Modulation de la santé digestive des poulets alimentés ...
231
© Amal Rouissi, 2020 Modulation de la santé digestive des poulets alimentés sans antibiotiques Thèse Amal Rouissi Doctorat en sciences animales Philosophiæ doctor (Ph. D.) Québec, Canada
Transcript of Modulation de la santé digestive des poulets alimentés ...

© Amal Rouissi, 2020
Modulation de la santé digestive des poulets alimentés sans antibiotiques
Thèse
Amal Rouissi
Doctorat en sciences animales
Philosophiæ doctor (Ph. D.)
Québec, Canada

Modulation de la santé digestive des poulets
alimentés sans antibiotiques
Thèse
Amal Rouissi
Sous la direction de :
Marie-Pierre Létourneau Montminy, directrice de recherche
Martine Boulianne, codirectrice de recherche

ii
Résumé
Bien que les principales indications pour l’utilisation des antibiotiques en alimentation
animale sont d’ordre curative et préventive, il est reconnu que leur utilisation permet
également l’amélioration des performances de croissance, ce qui leur a valu leur appellation
de « facteurs de croissance ». Les antibiotiques facteurs de croissance (AFC) permettent de
maintenir une concentration constante d’antibiotiques chez l’animal afin de pouvoir contrôler
la flore intestinale et donc prévenir ou minimiser certains problèmes de santé. Plusieurs
pratiques reliées à l’utilisation des antibiotiques en élevage pour améliorer les performances
zootechniques ont par contre été remises en question face à l’émergence de
l’antibiorésistance et ce, notamment du point de vue de la santé publique et de la sécurité
sanitaire des aliments. Depuis 1999, le retrait des antibiotiques dans les élevages de poulets
de chair en Europe a été associé à la résurgence d’entérite nécrotique, une maladie intestinale
causée par la bactérie Clostridium perfringens. Alors que la prévention de cette maladie
semblait bien maîtrisée, le retrait des antibiotiques a montré que peu de solutions de
remplacement sont disponibles pour prévenir cette maladie. C’est donc avec l’objectif
principal d’identifier des alternatives efficaces qu’a été tout d’abord réalisé une méta-analyse
pour distinguer les alternatives aux AFC les plus fréquemment étudiés et évaluer
numériquement leur effet dans l’améliorations des performances de croissance. Cette méta-
analyse fut suivie par des essais pratiques. Ces derniers, ont été testé l’impact du
remplacement des AFCen station expérimentale sur les performances de croissance, et la
santé digestive des poulets grâce à divers indicateurs (tels que le dosage de pH et humidité
de litière, l’histomorphométrie des cryptes et villosités intestinales, les mesures des
concentrations des acides gras à chaine courte dans les caeca et des indicateurs sériques). Les
alternatives étudiées étaient des acides organiques, des prébiotiques synthétique et naturels
(Avoine nue), des probiotiques et des produits phylogéniques et des huiles essentielles. La
présente étude a permis de quantifier l'effet des alternatives aux AFC sur les performances
de croissance à travers les divers modèles actuels établis par notre méta-analyse. En plus, elle
nous a permis de comparer ces alternatives aux AFC et de caractériser leur effet sur les
performances de croissance et la santé digestive des poulets de chair élevés sans antibiotiques
au Québec.

iii
Abstract
Although the principal use of antibiotics in animal feed is therapeutic and prophylaxic, it has
been known for a long time that their use also improves growth performance, which has
earned them the designation of "growth promoters ". Antibiotics (AGP) help maintain a
constant concentration of antibiotics in the gut in order to control the intestinal flora and
therefore prevent or minimize certain health problems. An important global problem in recent
years is the development of antibiotic resistance. This made questioned several practices
related to antibiotic use in animal production to improve zootechnical performance,
particularly from the point of view of food safety and public health. Since 2006, antibiotic
withdrawal from broiler chickens in Europe has been associated with the resurgence of
necrotic enteritis, an intestinal disease caused by the bacteria Clostridium perfringens. While
the pathogenesis of the disease seemed to be well understood, the absence of preventive in-
feed antibiotics has underlined that few alternatives are available to prevent this disease. In
this context, this thesis goal was to study the alternatives to AGP first with a meta-analysis,
to distinguish which AGP alternatives are the most studied and likely to improve growth
performance. This meta-analysis was then followed by field trials wherethe impact of the
AGP replacement were tested in animal husbandry in Quebec. Growth performances, as well
as chickens digestive health of were evaluated usingvarious markers (such as litter pH and
moisture, intestinal crypts and villi histomorphometry, cecal short chain fatty acids
concentrations and measurements of seric blood indicators). Tested alternatives included
organic acids, synthetic and natural prebiotics (naked oats), probiotics, phytogenic herbal
products and essential oils. The current study made it possible to quantify the effect of
alternatives to AGP on growth performance through the various current models established
by our meta-analysis. In addition, it has enabled us to better document, from a scientific point
of view, to compare these alternatives and to characterize their effect on the digestive health
of broilers raised without antibiotics in Quebec.

iv
Table des matières
Résumé ................................................................................................................................................. ii Abstract ............................................................................................................................................... iii Table des matières ............................................................................................................................... iv Liste des figures .................................................................................................................................. vi Liste des tableaux ................................................................................................................................ ix Liste des abréviations ........................................................................................................................... x Remerciements .................................................................................................................................... xi Avant-propos ..................................................................................................................................... xiii Introduction ......................................................................................................................................... 1 Chapitre 1 Revue de la littérature ................................................................................................. 4
1.1. Physiologie digestive chez le poulet ................................................................................... 5
1.1.1. Anatomie ..................................................................................................................... 5
1.1.2. pH ................................................................................................................................ 7
1.1.3. Le temps de rétention moyen ...................................................................................... 9
1.2. Devenir des nutriments dans le tube digestif des poulets .................................................. 13
1.2.1. Protéines .................................................................................................................... 13
1.2.2. Lipides ....................................................................................................................... 15
1.2.3. Fibres ......................................................................................................................... 17
1.3. Santé digestive des poulets ................................................................................................ 21
1.3.1. Pathologie digestive .................................................................................................. 22
1.3.2. Biomarqueurs de la santé digestive ........................................................................... 24
1.4. Alternatives aux antibiotiques facteurs de croissance ....................................................... 32
1.4.1. Prébiotiques ............................................................................................................... 33
1.4.2. Les probiotiques ........................................................................................................ 46
1.4.3. Les acides organiques ................................................................................................ 47
1.4.4. Les huiles essentielles ............................................................................................... 49
1.5. L’approche de la méta-analyse .......................................................................................... 51
1.5.1. Définition .................................................................................................................. 51
1.5.2. Étapes d’une méta-analyse ........................................................................................ 52
1.5.3. Les limites d’une méta-analyse ................................................................................. 60
Chapitre 2 Assessing growth promoter effect and modulating factors of dietary feed additive
commonly used in broilers: insights from meta-analysis approach .................................................. 62 2.1. Résumé .............................................................................................................................. 64
2.2. Abstract ............................................................................................................................. 65
2.3. Introduction ....................................................................................................................... 66
2.4. Materials and methods ...................................................................................................... 67
2.5. Results and discussion ....................................................................................................... 70

v
2.6. Conclusion ......................................................................................................................... 84
2.7. References ......................................................................................................................... 85
Chapitre 3 Effects of Bacillus subtilis, butyric acid, mannan-oligosaccharide and ß-glucans from
naked oat on growth performance, serum parameters and gut health of broiler chickens ................ 93 3.1. Résumé .............................................................................................................................. 95
3.2. Abstract ............................................................................................................................. 96
3.3. Introduction ....................................................................................................................... 97
3.4. Materials and methods ...................................................................................................... 99
3.5. Results ............................................................................................................................. 106
3.6. Discussion ....................................................................................................................... 119
3.7. Conclusions ..................................................................................................................... 123
3.8. References ....................................................................................................................... 124
Chapitre 4 Effect of different phytogenic feed additives on growth performance and intestinal
health 128 4.1. Résumé ............................................................................................................................ 130
4.2. Abstract ........................................................................................................................... 131
4.3. Introduction ..................................................................................................................... 132
4.4. Materials and methods .................................................................................................... 134
4.5. Results ............................................................................................................................. 140
4.6. Discussion ....................................................................................................................... 149
4.7. Conclusions ..................................................................................................................... 153
4.8. References ....................................................................................................................... 154
Chapitre 5 Discussion générale et conclusions ......................................................................... 157 5.1. Contexte de la thèse......................................................................................................... 158
5.2. Intérêt, résultats et limite de l’outil de méta-analyse ....................................................... 159
5.2.1. Intérêt de cette méta-analyse ................................................................................... 159
5.2.2. Résultats de cette méta-analyse ............................................................................... 161
5.2.3. Limites de cette méta-analyse ................................................................................. 164
5.3. Résultats pertinents des deux essais pratiques du projet ................................................. 166
5.3.1. Effet des traitements sur les indicateurs mesurés dans les deux essais ................... 166
5.3.2. Comparaison des effets des alternatives aux antibiotiques facteur de croissance sur les
performances de croissance entre les essaies pratique et la méta-analyse .............................. 177
5.3.3. Intérêts des indicateurs étudiés, limites et voies d’évolution .................................. 178
5.4. Contribution de la thèse à la diminution de l’utilisation des AFC .................................. 186
Conclusions ..................................................................................................................................... 191 Bibliographie ................................................................................................................................... 193

vi
Liste des figures
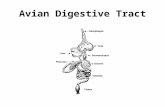
Figure 1-1 Rappel sur l’appareil digestif du poulet (en noir nom du segment et en orange les
fonctions principales) (Dusart, 2015). .................................................................................... 7
Figure 1-2 Variation du temps de transit moyen total avec l’âge des animaux (d’après Golian
et Maurice, 1992). ................................................................................................................. 10
Figure 1-3 Apports protéiques de différentes matières premières relatifs à ceux du tourteau
de soja (d'après Sauveur et al., 2002 Cité dans Bouvarel et al., 2014). ................................ 14
Figure 1-4 Répartition des différents types de « fibres » en fonction de leur solubilité et de la
méthode d’analyse (TDF, total dietary fibre; NSP, non-starch polysaccharides; NDF, neutral
detergent fibre; ADF, acid detergent fibre ; ADL, acid detergent lignine). ......................... 19
Figure 1-5 Le cycle biologique d'Eimeria (adaptée de Le Douarin, 2002). ......................... 22
Figure 1-6 Plage de croissance du pH de certains pathogènes bactériens de la volaille
(Mitscherlich et Marth, 1984). .............................................................................................. 25
Figure 1-7: A Structure de la paroi intestinale (Sarah, 2011) ; B : Villosité et Cryptes chez
les volailles (Richard;
http://environnement.wallonie.be/cgi/dgrne/aerw/ied/docs/20141126/r-ducatelle-sant-
intestinale-des-vollaillesgembloux.pdf). .............................................................................. 26
Figure 1-8 Composition du microbiote dans l’iléon et les Caeca du poulet (Lu et al., 2003).
.............................................................................................................................................. 31
Figure 1-9 Effet de la concentration du seigle et de la xylanase sur la viscosité de l’intestin
grêle proximal chez les poulets (tiré de Bedford et Classen, 1992). .................................... 45
Figure 1-10 Principales étapes de la méta-analyse (tiré de Sauvant et al., 2005). ............... 53
Figure 1-11 Les corrélations inter et intra étude (tiré de Loncke, 2009). ............................. 55
Figure 1-12 Exemple de la représentation graphique de la dispersion des résidus autour de la
droite de Henry pour le paramètre de gain de poids quotidien (ADG). ............................... 59
Figure 2-1 Prisma flow diagram. Number of scientific papers (trials) identified, screened and
selected for meta-analysis. .................................................................................................... 68
Figure 2-2 This forest plot shows the results of the meta-analysis with MOS as fixed effect.
For the average daily weight gain (ADG, in g) the mean difference estimate (black square
symbol) with corresponding 95% confidence interval (95% CI; black bar) between the
treatment group with MOS and the negative control group is shown. ................................. 76
Figure 2-3 This forest plot shows the results of the meta-analysis with MOS as fixed effect.
For the feed conversion ratio (FCR) the mean difference estimate (black square symbol) with
corresponding 95% confidence interval (95% CI; black bar) between the treatment group
with MOS and the negative control group is shown. ........................................................... 79
Figure 3-1 Concentrations of total SCFA (a), acetate (b), butyrate (c) and propionate (d) in
the caeca of growing broiler chickens fed alternative to antibiotic growth promoters (AGP);
Bs = Bacillus subtilis; absence of a letter in common indicates that the values. ............... 110
Figure 3-2 The pH of the crop (a), ileum (b), cecum (c) and litter (d) of growing broiler
chickens fed with alternative to antibiotic growth promoters (AGP); Bs = Bacillus subtilis;
absence of a letter in common indicates that the values differ significantly. ..................... 111
Figure 3-3 Litter moisture (%) variation overtime of broilers fed with alternatives to
antibiotics growth promoters; Bs = Bacillus subtilis. ......................................................... 118
Figure 4-1 Length (cm) of a) duodenum (Time: P<0.001, Trt: P=0.099, Time x Trt: P=0.092),
b) jejunum (Time: P<0.001, Trt: P=0.282, Time x Trt: P=0.029), and c) ileum (Time:

vii
P<0.001, Trt: P=0.057, Time x Trt: P=0.113) of broilers from the six different treatment
groups (n/treatment = 450). ................................................................................................ 142
Figure 4-2 Cecal concentration of a) total SCFA (mmol; Time: P<0.0001, Trt: P=0.0009,
Time x Trt: P= 0.0036), b) acetate (%; Time: P<0.0001, Trt: P=0.1791, Time x Trt:
P<0.0001); c) propionate (%; Time: P<0.0001, Trt: P=0.4077, Time x Trt: P=0.7648), and d)
butyrate (%; Time: P=0.0003, Trt: P=0.0269, Time x Trt: P=0.0031) of broilers from the six
different treatment groups (n/treatment =450). .................................................................. 144
Figure 4-3 Litter pH from 0- 21 day of broiler pens from the six different treatment groups
(n/treatment = 450) (Time: P<0.001, Trt: P=0.124, Time x Trt: P=0.0003). ..................... 147
Figure 4-4 Litter pH from 21- 34 day of broiler pens from the six different treatment groups
(n/treatment = 450). (Time: P<0.001, Trt: P=0.124, Time x Trt: P=0.0003). .................... 148
Figure 4-5 Litter moisture (%) of broiler pens from the six different treatment groups
(n/treatment = 450) (Time: P<0.001, Trt: P<0.001, Time x Trt: P<0.001). ....................... 149
Figure 5-1 Effet d’amélioration (%) des alternatives aux AFC étudié sur l’indice de
conversion alimentaire. ....................................................................................................... 162
Figure 5-2 Effet du pourcentage de l’énergie métabolisable sur l’indice de la conversion
alimentaire. ......................................................................................................................... 163
Figure 5-3 Effet des antibiotiques facteurs de croissance (AFC) (C- = traitement contrôle
sans AFC et C+ : traitement avec AFC) sur l’indice de la conversion alimentaire. ........... 164
Figure 5-4 Distribution du pourcentage de la protéine brute par rapport au besoin de la race,
de la race et du challenge sanitaire intrapublication dans la base de données des Mannan-
oligosaccharides.................................................................................................................. 166
Figure 5-5 Évolution du poids vif des poulets pour les différents traitements sur la période
d’essai par phase de croissance (nbr/traitement = 450). ..................................................... 168
Figure 5-6 Gain moyen quotidien pour les différents traitements sur la période d’essai par
phase de croissance (nbr/traitement= 450). ........................................................................ 169
Figure 5-7 La consommation moyenne journalière pour les différents traitements sur la
période d’essai par phase de croissance (nbr/traitement = 450). ........................................ 170
Figure 5-8 L’indice de consommation pour les différents traitements sur la période d’essai
par phase de croissance (nbr/traitement = 450). ................................................................. 171
Figure 5-9 Taux de mortalité pour les différents traitements sur la période d’essai par phase
de croissance (nbr/traitement = 450). ................................................................................. 171
Figure 5-10 Valeur de pH de la litière pour les différents traitements sur la période d’essai
par phase de croissance (nbr/traitement = 450). ................................................................. 172
Figure 5-11Taux d’humidité de la litière pour les différents traitements sur la période d’essai
par phase de croissance (nbr/traitement = 450). ................................................................. 173
Figure 5-12 Corrélation entre la concentration des acides gras à chaîne courte et ramifiée
avec les genres de bactéries dans le caecum de poulets nourris (10 poulets/ traitement) avec
des régimes alimentaires contenant des niveaux croissants de désoxynivalénol (0, 2,5, 5 ou
10 mg / kg) et avec ou sans ajout des lipopolysaccharides (LPS), 1 jour avant l’abattage. Les
couleurs font référence au degré de corrélation (Lucke et al., 2018). ................................ 180
Figure 5-13 Relation entre la CMJ et le pH de la litière (r=0.9; R2= 78.9%; P< 0.0001). 181
Figure 5-14 Relation entre l’humidité et le pH de la litière (r=0.63; R2= 65.3%; P< 0.0001).
............................................................................................................................................ 182
Figure 5-15 Relation entre la concentration en butyrate et en acétate dans le caeca des poulets
de chair (r=0.89; R2= 73.9%; P< 0.0001). ......................................................................... 183

viii
Figure 5-16 Concentration des acides gras dans les différents segments de l’intestin au jour
21 (Gonzalez-Ortiz et al., 2019). ........................................................................................ 184
Figure 5-17 Consommation de viande et protéine animale (kg) par habitant au Canada
(aliments disponible par personne, par année; (Statistique Canada, 2020)). ...................... 187

ix
Liste des tableaux
Tableau 1-1 pH des différents compartiments du tube digestif des poulets en fonction de
l’alimentation. ......................................................................................................................... 8
Tableau 1-2 Temps de rétention moyen (TRM, min) de l’aliment observé chez le poulet
(Sarah, 2011). ....................................................................................................................... 11
Tableau 1-3 Prolifération de Clostridium perfringens chez poulets de chair en fonction de
l’alimentation (Annett et al., 2002). ..................................................................................... 20
Tableau 1-4 Les acides gras volatils le long du tube digestif du poulet (Zhang et al., 2003)1.
.............................................................................................................................................. 28
Tableau 1-5 Les habitants du tube digestif des oiseaux. ...................................................... 29
Tableau 1-6. Effet de l’incorporation de l’avoine nue sur les performances de croissance de
poulet et le porc. ................................................................................................................... 38
Table 2-1 Descriptive statistics of the databases1, 2. ............................................................. 71
Table 2-2 Prediction of the effect of alternatives on feed conversion ratio (FCR) in function
of metabolizable energy (ME) and doses. ............................................................................ 74
Table 2-3 Prediction of the effect of alternatives on average daily gain (ADG) in function of
metabolizable energy (ME) and doses.................................................................................. 78
Table 2-4 Average daily gain and feed conversion ratio observed in experimental diets and
predicted by the models. ....................................................................................................... 83
Table 3-1 Broiler chicken basal diet composition (ingredient %). ..................................... 104
Table 3-2 Growth performance of broiler chickens fed with different alternatives to
antibiotics growth promoters (AGP) at different time points and P-values for treatment
interactions. ........................................................................................................................ 107
Table 3-3 Broiler chicken intestinal segment development in response to alternatives to
antibiotics growth promoters (AGP) at different time points (day of age), per treatment group
and interaction effects. ........................................................................................................ 112
Table 3-4 Ileal histomorphological parameters of broiler chickens fed with different
alternatives to antibiotics growth promoters and P-value for treatment interactions after the
starter and the grower phases. ............................................................................................ 115
Table 3-5 Serum parameters of broiler chickens fed different alternatives to antibiotics
growth promoters and P-values for treatment interactions. ................................................ 117
Table 4-1 Basal diet composition for starter, grower and finisher phase. .......................... 138
Table 4-2 Average daily gain, average daily feed intake, and feed conversion ratio of broiler
chicken fed with the six treatments for the three feeding periods (n/treatment=450). ....... 141
Table 4-3 Ileal histomorphological parameters of 28 day-old broiler chicken from the six
different treatment groups (n/treatment=450). ................................................................... 146
Table 5-1 Concentration des acides gras à chaîne courte totaux (mmol); de l’acétate (%),
propionate (%), butyrate (%) dans les caeca de poulets de chair pendant les trois phases de
croissance et paramètres histomorphologiques iléaux au jour 28 (nbr/traitement = 450). . 175
Table 5-2 Comparaison des effets des alternatives aux AFC sur les performances de
croissance entre les essaies pratique et la méta-analyse. .................................................... 178
Table 5-3 Effet des différentes alternatives sur le gain de poids quotidien, la consommation
moyenne journalière et l’indice de conversion alimentaire dans notre étude..................... 189

x
Liste des abréviations
ADFI Average Daily Feed Intake
ADG Average Daily Gain
AFC Antibiotique facteur de croissance
AGV Acides Gras Volatiles
AGP Antibitic growth promoter
CJM Consommation Journalière
CP Crude protein
EMA Énergie Métabolisable Apparente
FCR Feed Convertion Ratio
GMQ Gain Moyen Quotidien
IC Indice de Consommation
INRA Institut National de la Recherche Agronomique
Lys Lysine
NRC National Research Council
NSP Non-Starch Polysaccharides
OMS Organisation Mondiale de la Santé
P Probabilité
PB Protéine brut
pH Potentiel Hydrogène
RMSE Root mean square error
RMSEP Root mean square error of prediction

xi
Remerciements
يومًا ما ستبدو فخورًا بكل الصّعاب التي واجهتها بحياتك، بكل لحظة خوف، توترّ، قلق، سهر؛ ستبدو فخورًا جداً
بعبورك
جبران خليل جبران
Cette thèse a été réalisée à la Faculté des sciences de l’agriculture et de l’alimentation «
Département des sciences animales » de l’Université Laval.
Je tiens tout d’abord à remercier Alexandre Thibodeau de la Faculté de médecine vétérinaire
de Montréal, Martin Lessard, chercheur au Centre de recherche et de développement de
Sherbrooke et Agnès Narcy d’INRAE, France d’avoir accepté d’évaluer cette thèse.
J’exprime toute ma reconnaissance à Marie-Pierre Létourneau-Montminy, professeur-
chercheur au Département des sciences animales de l’Université Laval, qui a assumé la
direction de ma thèse.
Un grand merci aussi pour ma co-directrice de thèse, Martine Boulianne, professeur-
chercheur à la Faculté de médecine vétérinaire de l’Université de Montréal.
Merci infiniment à vous deux pour votre encadrement, votre gentillesse, votre disponibilité
pour ce projet, pour la confiance que vous m’avez accordée, pour vos multiples conseils et
critiques et pour votre compréhension surtout. Vous avez contribué à cette thèse par vos
judicieux conseils, révisions et suggestions qui ont su bien me guider tout au long du projet.
Un immense merci à Fréderic Guay, professeur-chercheur au Département des sciences
animales de l’Université Laval, pour sa gentillesse durant ces quatre années, pour ses conseils
scientifiques, son implication dans ce projet, pour son aide tout le long des différentes étapes
et surtout son soutien moral.
J’aimerais aussi exprimer ma gratitude envers les partenaires financiers et techniques :
la Chaire de recherche sur les stratégies alternatives d’alimentation des porcs et des volailles

xii
: approche systémique pour un développement durable, Mitacs, Agri-Marché et le groupe
WESTCO, sans qui ce projet n’aurait pu avoir lieu. Je suis également reconnaissante envers
mon cher pays, la Tunisie, qui m'a accordé une bourse d’étude pour pouvoir profité de ces
années d’apprentissage à l’Université LAVAL.
Un remerciement spécial à l’équipe de l’animalerie de CRSAD spécialement du poulailler.
Merci d’avoir si bien veillé sur mes poules tout au long des expérimentations, votre aide et
votre présence ont vraiment fait la différence lors des phases animales. J’aimerais aussi
remercier l’équipe du laboratoire sans qui ma charge de travail aurait été décuplée. Donc
merci à vous, Nancy, Dominic, Micheline, Yolaine at Alexandre pour votre expertise, votre
disponibilité et votre aide précieuse.
Je tiens à remercier toutes personnes qui m’a entourée durant ces années qui sont ma chère
coloc, mes cher(e)s amis et collègues en Tunisie ainsi qu’au Québec, qui m’ont toujours
soutenu avec un grand cœur, un grand merci pour votre amitié et votre disponibilité en tout
temps. Vous avez fait preuve d’une amitié sincère et durable et dont je garde un souvenir
merveilleux. Vous m’avez aimablement soutenu quand j’en avais besoin. Qu’ils trouvent ici
tous mes vœux de bonheur et de réussite. Je vous souhaite une bonne chance pour votre vie.
Merci énormément à Maroua Zouaoui, pour tous les bons moments qui sont devenus
inoubliables, merci d’être là et de m’écouter surtout pendant les moments difficiles.
A mes chers parents Hamadi et Monia, pour leur soutien, leur aide, leur patience et leur
présence tout au long de mes études. Je n’y serais pas arrivée sans vous. C’était assez difficile
de vous quitter pour venir au Canada, mais c’est la vie ! Que Dieu vous exauce de santé et
vous garde pour moi.
A mes sœurs Aya, Farah et Sirine, je ne peux pas exprimer à travers ses lignes tous mes
sentiments d’amour et de tendresse envers vous. Puisse l’amour et la fraternité nous unir à
jamais.
Encore une fois, merci à vous tous pour votre appui!

xiii
Avant-propos
La thèse qui suit comporte cinq chapitres avec insertion d’articles. Le premier chapitre
comporte l’introduction et la revue bibliographique. Il nous précise le contexte du projet à
partir d’un tour sur le contexte de restriction des AFC, la description de système digestive
des poulets, les différentes alternatives aux AFC et les multiples mesures de la santé digestive
des poulets de chair. Ce chapitre présente également une présentation de la méta-analyse.
Le deuxième chapitre est un article scientifique qui est en révision dans la revue Poultry
Science. Il s’agit du premier rapport résumant des données publiées indépendantes à travers
des modèles mécanistes obtenus avec un outil de méta-analyse décrivant l'effet des
alternatives aux antibiotiques facteurs de croissance (AFC) chez les poulets de chair. Cette
étude a été réalisée en collaboration avec Marie-Pierre Létourneau-Montminy et Frédéric
Guay professeurs à l’Université Laval et Martine Boulianne professeurs à l’Université de
Montréal.
Le troisième chapitre est un article scientifique qui sera soumis dans la revue Animal Feed
Science and Technology. Dans cette partie, basé sur les résultats du premier article de ma
thèse, une évaluation des effets in vivo de l'acide butyrique, de la manne-oligosaccharide, de
Bacillus subtilis et de l'avoine nue sur les performances de croissance, les paramètres
plasmatiques et la morphologie intestinale des poulets de chair et la qualité de la litière ont
été mesurés. Cette étude a été réalisée en collaboration avec Marie-Pierre Létourneau-
Montminy et Frédéric Guay professeurs à l’Université Laval et Martine Boulianne
professeurs à l’Université de Montréal.
Le quatrième chapitre consistait à étudier les effets in vivo de certaines huiles essentielles
commerciaux sur les performances de croissance, la santé intestinale et la qualité de la litière
des poulets de chair. Aussi, cette étude a été réalisée en collaboration avec Marie-Pierre
Létourneau-Montminy et Frédéric Guay professeurs à l’Université Laval et Martine
Boulianne professeurs à l’Université de Montréal. Le quatrième chapitre sera soumis dans
la revue Journal of Animal Science. Le cinquième et dernier chapitre représente une

xiv
discussion générale où les principaux éléments de la thèse ont été discutés pour finir avec
une conclusion renfermant des critiques et des perspectives de cette thèse.

1
Introduction
L’aviculture est un secteur très important des productions animales compte tenu qu’il s’agit
de la seconde viande la plus consommée mondialement juste derrière le porc (Statistique
Canada, 2020) et ce, sans compter l’importante consommation d’œufs. Par arriver à produire
ces quantités importantes de produits animaux, les volailles sont souvent maintenues à une
densité d’élevage élevée augmentant de par ce fait le risque de transmission d’agents
infections d’un animal à l’autre et même d’un élevage à un autre. Les agents infectieux
affectant le système digestif sont possiblement parmi les plus problématiques en productions
animales en raison de leur capacité à se transmettre rapidement et à provoquer des effets
négatifs sur la croissance et l’efficacité alimentaire des animaux (Williams, 2005). En fait, le
tube digestif des animaux est l’organe qui est plus en contact avec l’environnement extérieur.
Il agit à titre de barrière physique, chimique, immunologique et microbiologique sélective
entre le lumen intestinal et le milieu extracellulaire. La santé digestive est donc cruciale et
repose sur l’équilibre entre toutes ces composantes et est facilement perturbée par des
composants alimentaires ou des agents infectieux (Yegani et Korver, 2008). De ce fait,
l’utilisation des antibiotiques comme « facteurs de croissance » (AFC) en aviculture n’est
pas l’élimination d’une bactérie ou d’un groupe de microorganismes, que le contrôle de la
flore intestinale et la préventionde divers pathogènes (FAO/OIE/WHO, 2003).
Cependant, il est aujourd’hui reconnu que l’utilisation des AFC présente un risque pour la
sécurité alimentaire et la santé publique en raison du développement de l’antibiorésistance
(Sorum et Sunde., 2001). Cette résistante est un phénomène naturel de défense des bactéries
vis-à-vis de l’action exercée par l’antibiotique. Donc l’utilisation de ces « facteurs de
croissance » est une pratique qui est actuellement remise en question dans plusieurs pays.
Elle est strictement interdite par l’Union Européenne depuis le 1er Janvier 2006 (Régulation
(EC) No 183/2003) (Puyalto et al.,2018). Le Canada semble aujourd’hui suivre la même
tendance. C’est pourquoi, le 15 mai 2014, les Producteurs de poulet du Canada ont interdit
l’utilisation préventive des antibiotiques de catégorie I, de très haute importance et réservés
en médecine humaine au traitement des infections graves qui n’ont pas d’autres solutions,

2
dont principalement le ceftiofur (Les Producteurs de poulets du Canada, 2020). Ceci semble
avoir porté fruit puisqu’une diminution des résistances d’Escherichia coli au ceftiofur chez
les poulets a été observé par le Programme Intégré Canadien de surveillance de la Résistance
aux Antimicrobiens (PICRA). Depuis le 1er janvier 2018, les Producteurs de poulet du
Canada ont aussi interdit l’utilisation des antibiotiques de catégorie II, de haute importance
et réservés en médecine humaine pour le traitement d’infections graves qui disposent d’autres
options de traitement, en prévention. Un objectif pour éliminer l'utilisation préventive des
antibiotiques de catégorie III d'ici la fin de 2020 a aussi été fixé (Producteurs de poulet du
Canada, 2020). En revanche, la limitation de l’utilisation de ces antibiotiques a engendré une
recrudescence de l’entérite nécrotique et une diminution des performances (Timbermont et
al., 2011). L’entérite est l’un des problèmes les plus importants en production de volaille et
est responsable de performances de croissance diminuées et de mortalités augmentées
(Timbermont et al., 2011) se traduisant en pertes économiques (Dahiya et al., 2005).
Différents pathogènes peuvent causer des entérites, la forme la plus courante étant associée
à une prolifération de souches pathogènes de Clostridium perfringens qui cause l’entérite
nécrotique. La présence de la coccidiose semble favoriser le développement de l’entérite
nécrotique chez la volaille (Williams, 2005). En effet, les lésions causées par la coccidiose
présenteraient un avantage favorisant la prolifération de Clostridium perfringens. L’entérite
nécrotique peut tout de même se manifester dans l’élevage sans qu’il n’y ait de cas de
coccidiose concomitant. La gestion de l'écosystème intestinal pour améliorer la santé
digestive constitue actuellement un défi pour la production de volailles et ce, mondialement.
Des stratégies alimentaires alternatives qui modulent la microflore intestinale et favorisent
également un développement optimal de la muqueuse intestinale font donc l’objet de
nombreuses recherches. Toutefois, malgré les études réalisées sur ces stratégies, le fait qu'ils
puissent vraiment remplacer les AFC demeure une question controversée. En outre, il existe
de nombreuses alternatives telles que les immunostimulants, les bactériophages, les peptides
antimicrobiens, les pro- et les pré- biotiques, les huiles essentielles, les extraits de plantes,
les acides organiques et les enzymes (Diarra and Malouin, 2014). Le mode d'action de ces
produits n'est pas bien connu et leurs impacts sur la croissance et la prévention des maladies

3
digestives restent très variables (Diarra and Malouin, 2014). Afin de mieux définir les effets
des différentes alternatives, il apparait nécessaire de faire le point sur les recherches qui ont
été réalisées, quantifier les effets et identifier les facteurs de succès par l’étude détaillée de la
littérature scientifique.
La méta-analyse est une méthode pertinente pour résumer et quantifier les connaissances
acquises dans le cadre de recherches publiées précédemment (Sauvant et al., 2005, 2008,
2020). Cet outil est également utile pour identifier les facteurs de modulation et les éléments
clés pour développer une stratégie de remplacement des AFC efficace. Compte tenu du grand
nombre de publications sur les alternatives aux AFC et de leurs résultats contrastés chez les
poulets de chair, cette méthode statistique a été choisie pour répondre à l’objectif de la thèse
dans un premier temps. Ceci a permis d’identifier les stratégies les plus pertinentes et de les
tester dans des essais chez le poulet en croissance.
Notre objectif principal était de déterminer des stratégies alimentaires permettant de
maintenir les performances de croissance dans un contexte de retrait des AFC.
Prémiéremeent de développer des indicateurs de santé digestive pour évaluer l’impact des
stratégies testées. Deuxièmement, identifier les alternatives les plus étudiées et prometteuses
de façon objective par une approche de méta-analyse. Finalement de tester l’effet de ces
alternatives sur les performances de croissance et la santé digestive des poulets dans deux
essais en conditions expérimentales.

4
Chapitre 1 Revue de la littérature

5
1.1. Physiologie digestive chez le poulet
1.1.1. Anatomie
Le tractus gastro-intestinal des monogastriques présente quelques particularités anatomiques
comparativement à celui des mammifères monogastriques (Figure 1-1). Contrairement à
l’estomac de ces mammifères constitué d’une seule partie, celui des oiseaux comprend deux
parties : d’abord le proventricule qui a un rôle chimique, puis le gésier qui a un rôle
mécanique. La digestion consiste en une dégradation mécanique et chimique de l’aliment
dans le tube digestif en composés absorbables se retrouvant dans le sang et assimilables par
les cellules. Les différents organes constituant l’appareil digestif ont des actions spécifiques
et interviennent successivement dans le processus de digestion à mesure que l’aliment
transite le long du tractus gastro-intestinal (Larbier and Leclercq., 1992.). L'appareil digestif
des volailles est formé de:
La cavité buccale qui ne comprend ni lèvres ni dents, mais un bec corné qui permet la
préhension et une certaine fragmentation des aliments. Les glandes salivaires sont peu
développées résultant en une faible production de salive. Dans la bouche, les aliments sont
grossièrement insalivés au niveau du bec (Larbier et Leclercq., 1992).
L'œsophage assure le transport des aliments de la cavité buccale à l’estomac. Il est tapissé
dans toute sa longueur d’une muqueuse (Alamagot,1982).
Le jabot est un élargissement de l’œsophage en forme de réservoir en bas du cou. Les
aliments subissent une humectation et se ramollissent. Il est le lieu d'une digestion
microbienne par des Lactobacilles (Lan et al, 2002).
Le proventricule, l'estomac chimique, est riche en glandes qui sécrètent notamment l’acide
chlorhydrique et les pepsinogènes précurseurs de la pepsine permettant la digestion
chimique. La protéolyse y débute à un pH de 3 à 4,5. Dans le gésier et le proventricule, le
faible pH fait chuter la population bactérienne (Farner, 1943).
Le gésier est l’estomac mécanique qui est caractérisé par une couche superficielle très dure
entourée de muscles puissants. Il peut contenir de petits graviers qui sont nécessaires aux
animaux consommant des grains intacts, lorsque ceux-ci ont accès à de telles particules. C'est

6
donc au niveau du gésier que se produit véritablement la protéolyse sous l'action de la pepsine
(Gabriel et al., 2005).
Le petit intestin est un tube d'environ 1,2 m de longueur chez l’oiseau adulte dont la paroi
est bien équipée en glandes sécrétrices. Il se compose du duodénum, du jéjunum et de l’iléon.
Le duodénum reçoit les sécrétions du pancréas et du foie (Almargot, 1982). L'environnement
est plus favorable à la croissance bactérienne en raison de la plus faible pression d'oxygène
et de la faible concentration en enzyme et en sels biliaires.
Le pancréas est très développé et occupe l'espace entre les deux branches de l'anse
duodénale. La sécrétion pancréatique peut augmenter ou diminuer en fonction des besoins et
de la ration alimentaire.
Le gros intestin est peu développé chez l’oiseau, se réduisant pratiquement aux caeca où ont
lieu les fermentations bactériennes (Farner, 1943).
Le caecum : ils sont au nombre de deux et ils sont accolés à la partie terminale de l'iléon par
un méso. Ils sont en rapport ventralement avec l'anse duodénale et dorsalement avec la
portion moyenne de l'iléon. (Villate, 2001; Alamargot, 1982). Il est le siège de la fermentation
microbienne (Almargot, 1982).
Le rectum : il fait suite à l'iléon et débouche dans le cloaque. Il présente des villosités
comparables à ceux des mammifères. Ces villosités absorbent le liquide rectal et déshydrate
les fientes (fèces et urines) (Almargot, 1982).
Le cloaque : est la partie terminale de l'intestin dans laquelle débouchent les conduits
urinaires et génitaux. Il est formé de trois régions séparées par deux plis transversaux plus ou
moins nets (le coprodéum, l'urodéum, et le proctodéum) (Alamargot, 1982; Villate, 2001).

7
Figure 1-1 Rappel sur l’appareil digestif du poulet (en noir nom du segment et en orange les
fonctions principales) (Dusart, 2015).
1.1.2. pH
Le pH dans les contenus digestifs diffère d’un compartiment à un autre commençant par un
pH acide dans la zone gastrique à un pH presque neutre dans l’intestin. Le pH dépend des
sécrétions digestives et en plus de dépendre de la composition et de la forme de l’aliment
(Tableau 1-1).

8
Tableau 1-1 pH des différents compartiments du tube digestif des poulets en fonction de
l’alimentation.
Compartiment Régime à base de maïs, avoine et
blé 1
À jeun2 Farine3 Granulé3
Gésier 2,46-2,79 2,47-2,53 3,68 4
Duodénum 5,68-6,07 6,39-6,41 6,21 5,84
Jéjunum 5,72-6 6,58-6,62 6,17 6,05
Iléon 6,18-6,5 7,18-6,50 7,18 6,73
Rectum 6,08-6,58 6,98-7,02 6,47 5,97
Caeca 5,6-5,83 6,88-6,92 6,7 6,13
1Farner, 1943; 2Herpol et Van, 1967; 3Engberg et al, 2002
Un aliment en farine provoque une stimulation plus importante de la sécrétion de HCl dans
le gésier, ceci en raison d’un temps de séjour plus long dans le gésier que les aliments
granulés. Certaines études ont également rapporté que l’ajout de NSP (polysaccharides non
amylacés) insolubles pourrait entraîner une baisse de pH dans le gésier et dans les autres
compartiments du tube digestif (Jiménez - Moreno et al., 2009). L’ajout des fibres grossières
augmente le temps de séjour dans le gésier, ce qui favorise l’activation de la sécrétion de
pepsine et la production de HCl (Duke, 1997). Les problèmes qu’on peut observer au niveau
du pH des parties du tractus digestif s’observent lorsqu’on utilise des aliments à haut pouvoir
tampon, qui mènent à un pH gastrique élevé, altérant la digestibilité des protéines. Les
protéines non digérées atteignent le bas tube digestif où des excès de protéines entraîne une
fermentation produisant des amines biogènes toxiques (Skoufos et al., 2016). En outre, on
peut assister aussi à la prolifération de bactéries non bénéfiques dans le tube digestif
(Giannenas et al.,2014a; 2014b). La capacité du tube digestif à digérer les aliments et
absorber les nutriments est fortement liée au temps de rétention du digesta dans les différents
segments du système digestif. En effet, plus le temps de transit est long, plus le contact entre

9
le substrat et les enzymes digestives est long et plus la digestion des composants et
l’absorption des nutriments sont efficaces (Metzler et Mosenthin, 2008).
1.1.3. Le temps de rétention moyen
Contrairement aux mammifères qui ont un intestin grêle particulièrement long (18 m chez un
porc adulte), celui des oiseaux est environ 10 fois plus court (1,6 m chez un poulet de 8
semaines) (Denis et al, 2004). Mais, il présente un fonctionnement unique associant des
ondes péristaltiques et rétropéristatiques qui permettent d’optimiser la digestion et
l’absorption des nutriments. Cette particularité représente probablement une adaptation
permettant de réduire le poids du tractus gastro-intestinal (Susan, 2014). Le temps de
rétention moyen (TRM) varie d’un compartiment à l’autre. Au sein d’un même
compartiment, il dépend de l’âge des animaux (Figure 1-2), la composition de leur régime
(Palander et al., 2010) et leur lignée génétique (Shires et al., 1987). Dans le jabot, le TRM
varie de quelques minutes à plusieurs heures (Tableau 1-2). Le temps de stockage de
l’aliment dans cet organe dépend du taux de remplissage du gésier, ainsi un animal dont le
gésier est vide ne stocke pas d’aliment dans son jabot, ce qui explique les TRM, parfois très
courts observés (Klasing, 1998). Dans le proventricule, le TRM est très court, de l’ordre de
quelques minutes, et l’aliment est rapidement transféré vers le gésier. Le chyme reste dans le
gésier jusqu’à ce que la taille des particules alimentaires soit suffisamment réduite pour
passer la barrière du pylore, soit entre 15 et 85 minutes (Tableau 1-2). Dans l’intestin grêle,
le TRM augmente en allant du duodénum vers l’iléon, il dure de 3 à 18 minutes dans le
duodénum, entre 20 et 110 minutes dans le jéjunum et entre 30 et 150 minutes dans l’iléon
(Tableau 1-2). Ainsi en utilisant les observations rapportées dans le Tableau 2, le TRM dans
l’ensemble du tractus digestif, à l’exception des caeca, peut être estimé durer de 4 à 7h.

10
Figure 1-2 Variation du temps de transit moyen total avec l’âge des animaux (d’après Golian
et Maurice, 1992).

11
Tableau 1-2 Temps de rétention moyen (TRM, min) de l’aliment observé chez le poulet (Sarah, 2011).
Age
1 Lignée MPP2
Marqueu
r Temps de rétention moyen Références
Jabot Estomac Intestin grêle Colon Total6
Prov3 Gésier Estomac4 Duo5 Jéjunum Iléon Intestin
grêle
3 Mais
soja TiO2 26 98
Ouhida et al.,
2000
Orge
blé TiO2 21 104
Ouhida et al.,
2000
Seigle
soja TiO2 60 58 18 110 140 268 36 422
Danicke et al.,
1999
Ross 208 Blé
orge TiO2 52
Palander et al.,
2010
Ross 208 Avoin
e TiO2 32
Palander et al.,
2010
4
Ross
(femelle
)
Blé Cr2O3 64 93 Weurding et
al., 2001
Ross
(femelle
)
orge Cr2O3 53 82 Weurding et
al., 2001
6
Rhode
Island
x
Leghorn
51Cr 166 17 4 20 38 61 27 272 Sklan et al.,
1975

12
Rhode
Island
x
Leghorn
91Y 220 50 3 21 45 69 27 366 Sklan et al.,
1975
Hybro Mais
soja Cr2O3 41 33 5 71 90 166 26 266
Van der Kils et
al., 1990
Hubbard
Mais
soja+
mais
colza
103Ru 7 4 50 54 7 60 86 153 44 258 Shires et al.,
1987
Shaver
Starcros
s 208
Mais
soja+
mais
colza
103Ru 25 3 84 87 7 59 73 139 40 291 Sklan et al.,
1975
Ross 209 Blé
orge TiO2 151
Palander et al.,
2010
Ross 209 Avoin
e TiO2 89
Palander et al.,
2010
1Age exprimé en semaines 2M P principales : matières premières principales 3Prov : Proventricule, 4Estomac : proventricule + gésier, 5Duo : duodénum 6Temps de rétention moyen sur l’ensemble du tractus digestif à l’exception des caeca

13
1.2. Devenir des nutriments dans le tube digestif des poulets
La digestion met en jeu des phénomènes mécaniques, chimiques et microbiens. Les
phénomènes mécaniques sont la préhension et la mastication des aliments ainsi que les
contractions musculaires du tube digestif. Les principaux phénomènes chimiques sont dus
soit à des enzymes secrétées par l'animal qui conduisent à des réactions d'hydrolyse ou au pH
qui conduit à des solubilisations ou des insolubilisassions. Les phénomènes microbiens, eux-
mêmes de nature enzymatique, sont dus principalement à l'action de bactéries et de
protozoaires (Drogoul et Gadoud, 2004). Chez les monogastriques, l'insalivation des aliments
au niveau du bec a une fonction lubrifiante. C'est par l'œsophage que les aliments atteignent
l'estomac où est sécrété le suc gastrique. Après le mélange des nutriments et de suc gastrique
passe à l'intestin grêle où est secrété le suc pancréatique, le suc entérique et le bicarbonate
phosphate qui neutralisent l'acide chlorhydrique du suc gastrique et de la bile. L’hydrolyse
enzymatique dans le duodénum permet la libération des nutriments. Dans le duodénum et le
jéjunum sont absorbés la plupart des nutriments: minéraux, vitamines, glucose, acide gras,
acides aminés, vitamines, minéraux et eau (Larbier et Leclercq, 1992). L’aliment non digéré
dans l'estomac et l’intestin grêle passe au niveau de caeca (Fuller, 1984) qui constitue un
milieu favorable pour la multiplication bactérienne (le milieu est anaérobie, très liquide,
stagnant partiellement (l'évacuation ne se fait que toutes les 6 à 8 heures en moyenne), et le
pH est de l'ordre de 6,5 à 7,5). Par ce fait, la flore microbienne fermente majoritairement les
nutriments qui arrivent aux caeca et au colon. C'est à ces niveaux que sont absorbés les
produits obtenus par la fermentation microbienne (acides gras volatiles, groupes aminés,
etc.). Finalement, l'aliment non digestible ressort sous forme d'excrément (Fernandez et Ruiz
Matas, 2003).
1.2.1. Protéines
1.2.1.1. Définition et types de protéines
Les volailles ont besoin d’aliments énergétiques mais aussi riches en protéines surtout pour
les poules pondeuses et les poulets en pleine croissance. La source la plus riche en protéines

14
est le tourteau de soja et l’apport protéique des différentes autres matières premières est
variable (Figure 1-4). Les protéines sont composées par l’enchaînement de vingt acides
aminés (AA), dont 11 sont essentiels. Les acides aminés essentiels ne peuvent pas être
synthétisés par l’animal, par conséquent, ils doivent être nécessairement apportés par
l’alimentation en quantité répondant aux besoins. En alimentation de la volaille, on peut
distinguer deux sortes de protéines : des protéines végétales, on dit qu’elles sont incomplètes,
car déficientes en certains acides aminés essentiels, et des protéines animales qui sont
complètes et sont également plus facilement digestibles que les protéines d’origine
végétale. La lysine, la thréonine et la méthionine sont les trois acides aminés essentiels qui
sont les plus limitants pour la croissance dans les aliments pour volaille (Ajinomoto, 2018).
Figure 1-3 Apports protéiques de différentes matières premières relatifs à ceux du tourteau
de soja (d'après Sauveur et al., 2002 Cité dans Bouvarel et al., 2014).
1.2.1.2. Devenir intestinal des protéines
Dans l’estomac, le proventricule des oiseaux, de l’acide chlorhydrique est sécrété. Le contenu
de cet organe a donc un pH faible d’environ 2,5 chez le poulet. Cette acidité entraîne une
dénaturation des protéines et permet l’activation des pepsinogènes des proenzymes inactives
qui sont stockée dans les vésicules enzymatiques et excrétées au moment de la digestion.
L’activation du pepsinogène en pepsine est le résultat d’une hydrolyse acide dans le milieu
favorable de l’estomac, soit entre 1,8 et 4,4. La pepsine est ensuite inactivée par les

15
bicarbonates alcalins du suc pancréatique au niveau du duodénum. Les temps moyens de
séjour des aliments dans le gésier des oiseaux est court (moins d’une heure) avec toutefois
un séjour plus long pour les particules grossières que pour les fines particules (Crevieu-
Gabriel, 1999). Le digesta arrivant dans l’intestin subit l’action du suc pancréatique qui
contient plusieurs protéases, soit des endopeptidases (trypsine, chymotrypsine et élastase ;
carboxypeptidases), des aminopeptidases et des dipeptidases (Le Floch et Sève, 2000; Gabriel
et al, 2003). Les différents peptides et AA qui sont absorbés dans l’intestin grêle, alors que
leur absorption dans le gros intestin est pratiquement nulle. L’absorption le plus importante
des AA se fait principalement au niveau des entérocytes de l’intestin grêle (Denbow, 2000).
L’absorption intestinale s’effectue par l’action de transporteurs actifs ATP dépendants
couplés à du Na+ (Denbow, 2000). D’une façon générale, la vitesse d’absorption est
fortement diminuée chez les poulets nourris avec un aliment hypo-énergétique. Elle est en
revanche augmentée lorsque l’aliment est sous-carencé en acides aminés (Larbier et
Leclercq, 1992).
1.2.2. Lipides
1.2.2.1. Définitions et types des lipides
Il existe différentes classes de lipides : les triglycérides, qui représentent 95 à 98 % des lipides
alimentaires ingérés, les phospholipides, les sphingolipides et minoritairement les esters de
cholestérol. Ces différentes formes sont elles-mêmes constituées en majeure partie d’acides
gras (Afssa, 2001). Les acides gras peuvent être classés de différentes manières selon leur
structure (Anses, 2011a) en fonction de la chaine carbonée qui varie de 4 à plus de 24
carbones et en fonction de leur degré d’instauration, c’est-à-dire du nombre de doubles
liaisons carbone-carbone dans la molécule. On distingue ainsi :
- Les acides gras saturés : Ils sont synthétisés par l’organisme animal, en particulier dans le
foie, le cerveau et le tissu adipeux, et sont donc dits « non indispensables » (Anses, 2011b).
En plus de cette origine endogène, les acides gras saturés sont apportés par l’alimentation
(Anses, 2011a).

16
- Les acides gras monoinsaturés (une double liaison) : Comme les acides gras saturés, ils
proviennent aussi d’une part de la synthèse endogène et d’autre part de l’alimentation (Anses,
2011a). Quantitativement, l’acide oléique représente l’élément majeur de ces acides (Afssa,
2001).
- Les acides gras polyinsaturés (plusieurs doubles liaisons) : Parmi les AGPI, deux familles
nommées n-6 (ou « oméga 6 ») et n-3 (ou « oméga 3 ») sont particulièrement importantes.
Elles sont issues respectivement de l’acide linoléique (LA) et de l’acide α-linolénique (ALA),
synthétisés chez les végétaux à partir de l’acide oléique. Ces acides gras sont
« indispensables » car requis pour la croissance normale et les fonctions physiologiques des
cellules, mais non synthétisables dans l’organisme animal (Anses, 2011b).
1.2.2.2. Devenir intestinal des lipides
Du fait du caractère hydrophobe des lipides présents dans le milieu intestinal, qui lui, est
aqueux, leur digestion et absorption présentent deux différences majeures avec la digestion
et l'absorption des glucides et des protéines : leur digestion implique une émulsion, et leur
absorption par les entérocytes peut s’effectuer par simple diffusion à travers la bicouche
lipidique. La digestion des lipides commence dans le gésier par leur émulsion sous l’effet de
ses contractions, est facilitée par la présence de peptides issus de l’hydrolyse des protéines
(Klasing, 1998). Cette étape se poursuit dans l’intestin grêle sous l’action des acides biliaires.
On sait que les sels biliaires servent à la formation des micelles, leur faible concentration
réduit la solubilisation des lipides et donc leur absorption, en particulier ceux contenant des
acides gras saturés à longue chaîne (Larbier et Leclercq, 1992). La flore digestive modifie
largement les sels biliaires : déconjuguaison, désulfatation et déhydroxylation. Donc, elle
participe à la saturation des acides gras polyinsaturés par hydrogénation (Larbier et Leclercq,
1992). La digestibilité des lipides est faible chez le poussin où seuls les lipides insaturés sont
utilisés; par la suite, la digestion des graisses très saturées devient possible (Bouzouaia,
2019). Après l’hydrolyse, les acides gras libérés, les phospholipides et les monoglycérides
entrent dans les micelles formées grâce aux acides biliaires (Krogdahl, 1985; Klasing, 1998).
Les phospholipides alimentaires sont hydrolysés par la phospholipase pancréatique puis
absorbés sous forme de lysophospholipides (Larbier et Leclercq, 1992). L’absorption des

17
acides gras a lieu dans la partie distale du jéjunum et dans l’iléon (Krogdahl, 1985; Denbow,
2000). Les mécanismes d’absorption sont mal connus chez les oiseaux. Au niveau de la
bordure en brosse des entérocytes, les acides gras peuvent passer la membrane cellulaire par
diffusion passive.
1.2.3. Fibres
1.2.3.1. Définition des fibres
Le terme fibres alimentaires a été utilisé par Hipsley en 1953 pour la première fois. La fibre
est la partie indigestible des hydrates de carbone retrouvés principalement dans les plantes.
Les aliments les plus riches en fibres sont les céréales et leurs dérivés (son de blé, d'avoine,
orge...). Le terme fibre désigne en général les constituants des parois cellulaires des plantes,
comprenant une grande variété de polysaccharides structuraux qui sont souvent liés à des
protéines et à des phénols, particulièrement la lignine. Les principaux polysaccharides des
parois cellulaires des plantes sont: la cellulose, différentes hémicelluloses (i.e.
arabinoxylanes, β-glucane, xyloglucanes, arabinogalactanes) et des polysaccharides
pectiques (Figure 1-3). Les fibres jouent différents rôles au niveau physiologique y compris
la régularisation de la fonction gastro-intestinale, la diminution des taux de cholestérol ainsi
que la gestion de la glycémie (taux de sucre dans le sang). Des recherches récentes montrent
qu'il existe de nombreuses formes de fibres, chacune ayant un effet unique sur la nutrition et
la santé. Généralement, les fibres alimentaires sont classées en fonction de leur solubilité.
Deux catégories importantes de fibres sont alors définies : les fibres solubles et insolubles.
1.2.3.2. Les types de fibres
Tel que mentionné dans le paragraphe précédent, il existe deux types de fibres: les fibres
insolubles et les fibres solubles. Chaque type de fibres joue un rôle différent dans l'organisme.

18
Définition des fibres solubles
Les fibres solubles se dissocient facilement dans l’eau et formes des gels, que l’on définit
comme la viscosité. Les sources de fibres solubles comprennent le blé, l'avoine, l'orge, le
seigle et le triticale, ainsi que leurs coproduits (Choct, 2006). D’une façon générale, les fibres
solubles sont plus fermentées et mènent à une viscosité du digesta plus élevée que les fibres
insolubles (Bedford, 1995). Ce digesta visqueux atteignant l’intestin grêle, inhibe la digestion
de l'amidon, des lipides et des protéines chez la majorité des monogastriques (Beg et al.,
2001; Barrera et al., 2004) et augmente ainsi la sécrétion endogène dans l'intestin. Ceci
augmente également l'ingestion alimentaire, diminue l'efficacité alimentaire et réduit
l’absorption des nutriments (Wenk, 2001). Cette viscosité aussi causent l'apparition des
fientes collante (Bumett, 1966 cité par Sundberg et al, 1996).
Définition des fibres insolubles
Les fibres insolubles se dissocient difficilement dans l’eau. Les sources de fibres insolubles
sont en plus grande proportion dans les grains non visqueux et leurs dérivés tels le maïs, le
sorgho, le millet et le riz (Choct, 2006). Elles accélèrent le transit intestinal (Le Goff et al.,
2002). Malgré leur plus faible capacité de rétention d’eau, les fibres insolubles ont un effet
laxatif bénéfique en accentuant la masse des matières fécales, ce qui améliore leur circulation
dans le tube digestif, augmente les sécrétions de mucus et la fréquence de défécation
(Chutkan et al., 2012). Hetland et al (2003) ont montré que les fibres insolubles augmentent
les temps de rétention dans le gésier et amélioraient la digestibilité de l’amidon en
augmentant le reflux gastro-intestinal de sels biliaires.

19
Figure 1-4 Répartition des différents types de « fibres » en fonction de leur solubilité et de la
méthode d’analyse (TDF, total dietary fibre; NSP, non-starch polysaccharides; NDF, neutral
detergent fibre; ADF, acid detergent fibre ; ADL, acid detergent lignine).
1.2.3.3. Relation fibres et entérite nécrotique
La présence de polysaccharides non amylacés (NSP) dans l’aliment peut favoriser la maladie
en influençant les propriétés du contenu intestinal, comme la viscosité, le temps de transit
dans le tractus gastro-intestinal et le pH intestinal (Bedford, 1995; Timbermont et al., 2011;
Marie-Lou Gaucher, 2015). Plusieurs études dans la littérature qui ont étudié l’effet d’une
alimentation riche en fibres tels que l’avoine, l’orge, le seigle et le blé, présentent un risque
élevé au développement de l’entérite nécrotique (Timbermont, et al., 2011). Ces fibres
permettraient une diminution de la vitesse du transit intestinal, une augmentation de la
viscosité ou une modification du microbiote intestinal, favorisant la prolifération bactérienne
(Uzal, et al., 2015). Des études de Jia et al. (2009) et de Annett et al. (2002) (Tableau 1-3),
ont montré que l’orge et le blé diminuent la vitesse du transit intestinal et augmentant la

20
viscosité et permettant une plus grande prolifération de C. perfringens par rapport au maïs.
Donc il est bien important de faire un bon choix des composants de l’aliment afin de prévenir
l’entérite nécrotique.
Tableau 1-3 Prolifération de Clostridium perfringens chez poulets de chair en fonction de
l’alimentation (Annett et al., 2002).
Maïs Orge Blé
Médiane (x108 cfu/ml) 3.78 a 5.90 b 5.80 b
Premier quartile (x108 cfu/ml) 3.41 4.90 5.25
Troisième quartile (x108 cfu/ml) 4.06 7.95 6.90
L’inclusion de maïs dans l’aliment au lieu d’avoine, d’orge, de seigle ou de blé, est un moyen
de prévenir l’entérite nécrotique. Les mortalités associées aux céréales riches en ces
polysaccharides étaient aussi deux fois plus élevées lorsque comparées à celles des groupes
soumis à un aliment majoritairement composé de maïs (Shojadoost et al., 2012). Il a aussi
été démontré que certaines de ces céréales, telles que le seigle, augmentait l’adhésion de la
bactérie à la muqueuse intestinale (Kleessen et al., 2003; Shojadoost et al., 2012). Leur effet
s’explique par sa richesse en polysaccharides non amylasés solubles dans l’eau et faiblement
digestibles (ß-glucans et les arabinoxylans). Une fois rendus dans l’intestin, ces
polysaccharides peu digérés vont modifier l’environnement intestinal, en interagissant avec
des glycoprotéines de la surface épithéliale intestinale et provoqueraient une augmentation
de la production de mucine (Shojadoost et al., 2012). Puisque Clostridium perfringens
possède plus de 6 hydrolases capables de dégrader ces muco-oligosaccharides pour créer un
milieu favorable pour la bactérie (Shojadoost et al., 2012), en conséquence, une augmentation
du temps de transit intestinal et de la viscosité du digesta engendreront une prolifération
accrue de la bactérie (Williams ,2005). L’augmentation de la viscosité intestinale dépend de
la solubilité, de la taille moléculaire et de la conformation des ß -glucanes ainsi que de leur
vitesse de dégradation dans l'estomac et le petit intestin.

21
1.2.3.4. Devenir intestinal des fibres
Les glucides peuvent être distingués en deux types : ceux que l’oiseau peut digérer, soit
notamment l’amidon, la dextrine, les oligosaccharides et les monosaccharides, et ceux qui ne
peuvent être utilisés que par la microflore, qui regroupent notamment les polysaccharides
non amylacés tels la cellulose l’hémicellulose et les substances pectiques (Gabriel et al,
2003). Dans le cas des glucides utilisables par l’hôte, la microflore semble avoir un effet
limité. En effet, elle ne peut pas modifier l’activité des enzymes impliquées lors leur
digestion, telle que l’amylase pancréatique ou les disaccharides intestinaux, ni influencer
l’absorption du glucose. En ce qui concerne les glucides que l’oiseau ne peut utiliser, ils sont
fermentés par la microflore dans le jabot et principalement au niveau des caeca sans avoir un
rôle significatif.
1.3. Santé digestive des poulets
La santé digestive des volailles est un facteur très important en élevage, qui peut affecter
fortement la production et le bien-être animal. Elle repose sur trois piliers : l’état de santé du
tube digestif de l’animal, l’équilibre de la microflore et l’état du système immunitaire
(Sansonetti, 2015). Dans le secteur des volailles, les problèmes de la santé digestive sont
regroupés sous les rubriques de l'entérite nécrotique et de la dysbiose (≠ Eubiose) (Sansonetti,
2015). Selon Sokol et al. (2016), la dysbiose est un changement qualitatif ou quantitatif du
microbiote digestif, de son activité métabolique ou de sa distribution locale dans le tube
digestif conduisant à des effets néfastes chez l’animal. Une définition simplifiée donc de la
santé intestinale pourrait être : l'absence de paramètres décrivant la dysbiose (diarrhée,
constipation, ballonnement, inflammation, rougeur de la muqueuse intestinale). Dans les
parties qui suivent, les multiples indicateurs de la santé digestive chez les volailles seront
présentés.

22
1.3.1. Pathologie digestive
1.3.1.1. Coccidiose
La coccidiose chez le poulet est causée par un parasite, un protozoaire, le plus souvent en
élevage commercial, des espèces Eimeria tenella, Eimeria maxima ou Eimeria acervulina,
bien que l’on dénombre 9 espèces pouvant être en cause (Akçay et al., 2011). Il touche
directement les poulets en croissance, soient ceux âgés entre 3 et 6 semaines (Corrand et
Guérin, 2010). La coccidiose se présente sous forme de cycle dans les élevages avicoles
(Figure 1-5). Le processus infectieux est rapide (4 à 7 jours entre l’ingestion du parasite et
l’excrétion d’oocystes dans les fientes (Corrand et Guérin, 2010) et entraîne des dommages
intestinaux variables selon la charge parasitaire et la pathogénicité de la coccidie infectante.
1.3.1.2. L’entérite nécrotique
L’entérite nécrotique aviaire a été observée par Bennetts en 1930 en Australie, puis décrite
par Parish en 1961 en Angleterre (Martin et Smyth., 2009). Plus tard, la première
reproduction de la maladie a été faite par Bernier et al. (1977) chez des poulets de gril âgés
Figure 1-5 Le cycle biologique d'Eimeria (adaptée de Le Douarin, 2002).

23
de deux semaines, par l'administration orale et intraveineuse d'une culture pure de
Clostridium perfringens. L’entérite nécrotique se manifeste également chez de jeunes
volailles, généralement entre 2 et 5 à 6 semaines (Engstrom et al, 2003 ; Long, 1973). Elle
est causée par des souches pathogènes de Clostridium perfringens de type A ou C (Al-
Sheikhly et Truscott, 1977), bien que l’agent étiologique primaire semble être Clostridium
perfringens de type A, une bactérie à coloration Gram positive (Engstrom et al., 2003 ;
Gholamiandekhordi et al., 2006). Un nouveau toxinotype, type G qui est défini comme des
isolats qui produisent une toxine a et une toxine NetB (Rood et al.,2018). Il a été démontré
que les souches de C. perfringens de type G sont responsables pour l'entérite nécrotique chez
les poulets (Keyburn et al., 2008). Les preuves génétiques du rôle essentiel de la NetB dans
la virulence de l’entérite nécrotique sont très claires et étayé par des preuves
épidémiologiques solides (Rood et al., 2016; Prescott et al., 2016). De plus, plusieurs études
ont montré une relation entre la coccidiose et l’entérite nécrotique (Al-Sheikhly et Al-Saieg,
1980 ; Bedford, 2000). En effet, la coccidiose cause des lésions qui pourraient être un point
d’entrée pour le Clostridium perfringens. Ainsi, la coccidiose, sans être un facteur essentiel
au développement de l’entérite nécrotique, pourrait augmenter le risque pour les poulets de
développer une entérite nécrotique. La colonisation intestinale de C. perfringens chez les
poulets de chair cliniquement normaux est faible (environ 105 cfu / g de digesta), mais lorsque
la colonisation augmente à 107-109 cfu / g de digesta, la maladie clinique se développe si des
souches pathogènes sont présentes (Kondo, 1988). L’entérite nécrotique est une maladie
multifactorielle. Dahiya et al. (2005) ont signalé cinq facteurs prédisposants pour l'entérite
nécrotique: infection par Eimeria, élimination des coccidiostatiques ou des facteurs de
croissance antibiotiques des aliments, conditions environnementales et de gestion, stress
physiologique et immunosuppression et nature et forme de l'alimentation. Cette maladie peut
engendrer beaucoup de dégâts tels qu’une perte du gain de poids, une dépression, une
inappétence, une anorexie et des diarrhées et surtout une mortalité soudaine qui peut être
importante. L’entérite nécrotique est généralement traitée et prévenue par des antibiotiques.
La prévention de cette maladie passe également par la gestion des facteurs de risque,
principalement le contrôle de la coccidiose et par une bonne hygiène de l’environnement des
poulets. Au Canada, la prévention passe également par l’utilisation d’antibiotiques pour
contrôler les infections intestinales. En effet, la présence d’antibiotiques dans la nourriture

24
des poulets induit un changement dans le microbiote intestinal des volailles ce qui a une
influence sur leur immunité et leur santé (Lee et al, 2011).
1.3.2. Biomarqueurs de la santé digestive
1.3.2.1. Aspect des fientes et de la litière:
Les fientes sont un indicateur indirect d’un désordre de santé ou un déséquilibre nutritionnel.
Les fientes caecales en général sont d’un aspect crémeux et excrétées moins fréquemment
que les fientes intestinales, soit environ une fois par jour. Maria (2005) a indiqué que la
présence de fientes liquides peut-être due à un problème infectieux ou parasitaire, un
changement de régime alimentaire (composition, taille, procédé de fabrication…), une
température au sol insuffisante cumulée avec de l’humidité, ou encore une surconsommation
d’eau. Pour contrôler les diarrhées, Puterflam et al. (2007) ont calculé le ratio volumétrique
liquide/solide des fientes (Elanco-fluid). Un ratio supérieur à 0,5 (plus de 50% de fientes
humides deux jours consécutifs) est un indicateur de fientes liquides, donc de l’apparition de
diarrhées. Quand on parle de fientes, on doit directement penser à l’aspect de la litière. Les
facteurs affectant l’humidité de la litière sont multiples, comprenant l’alimentation, la saison,
la densité d’élevage, la ventilation, la conception du système d’abreuvement et la santé
digestive des animaux (Collett, 2006; 2012). Une augmentation considérable du taux
d’humidité de la litière est considérée comme le premier signe d’une entérite. Mortimer
(2002) a montré qu’en dessous de 20% de fientes humides, dans 95% des cas il n’y avait pas
de problème de litière dans l’élevage, alors qu’au-delà de 50% de fientes humides pendant
deux jours consécutifs l’élevage était en voie de développer de l’entérite dans 100% des cas.
Outre que l’humidité de la litière qui est un facteur important pour la santé des poulets, on
parle aussi du pH de la litière. La litière de volaille a un pH moyen de 6 à 9 (Jones-Hamilton,
2014). Parce que la litière de volaille a un pH neutre à tendance basique, elle peut favoriser
la croissance de Salmonella, Clostridium et d'autres bactéries pathogènes (Figure 1-6). Ainsi,
tout en abaissant le pH de la litière à 4,0 ou moins, cela réduira les bactéries pathogènes et
créera en fait un environnement favorable à la prolifération de bactéries saines (Jones-
Hamilton, 2014). La réduction du pH aussi conduit à une réduction de la production de

25
l'ammoniac (Burgess et al.,1998). Comme rapporté par Elliott et Collins (1982), les rejets
d'ammoniac de la litière sont liés au pH et à l'humidité de cette dernière. Les émissions
d'ammoniac ont été positivement corrélés avec le pH de la litière (Carr et al.1990) et
négativement corrélés avec la teneur en humidité de la litière (Ferguson et al. 1998). Cette
pratique d’acidification en général, est répandue et bien établie dans l'industrie américaine
des poulets de chair qui recycle la litière sur plusieurs lots d’élevage consécutifs (Jones-
Hamilton, 2014).
Figure 1-6 Plage de croissance du pH de certains pathogènes bactériens de la volaille
(Mitscherlich et Marth, 1984).
1.3.2.2. Cryptes et villosités
La muqueuse intestinale contient de nombreuses villosités qui sont des projections de la
muqueuse intestinale dans la lumière permettant d’augmenter la surface d’absorption
(Hodges, 1974). À la base des villosités se trouvent les cryptes de Lieberkühn qui sont
formées par une invagination de la muqueuse intestinale (Figure 1-7) et qui sont composées

26
de 3 types de cellules : les entérocytes ont une fonction d’absorption, les cellules de Paneth
ont une fonction d’immunité, les cellules progénitrices donnent des entérocytes et des cellules
caliciformes. La fonction d’immunité des cellules de Paneth consiste à libérer des enzymes
antimicrobiennes pour détruire les bactéries dans le tube digestif. Selon Lan et al. (2002), la
flore et la muqueuse intestinale ont des relations symbiotiques et compétitives, certaines
variations de la flore pouvant altérer les caractéristiques structurelles et fonctionnelles du
tube digestif. Les villosités intestinales sont plus hautes dans le jéjunum et l’iléon. Les cryptes
sont profondes tout le long de l’intestin grêle et le nombre de cellules en division est plus
élevé conduisant à un renouvellement cellulaire accéléré du duodénum distal à l’iléon. Les
nutriments (glucides, lipides, acides aminés), produits de la digestion et solubles dans la
lumière intestinale, sont absorbés au niveau des entérocytes, cellules qui tapissent les
villosités. Plus les villosités sont longues, plus la quantité d’entérocytes est importante.
Figure 1-7: A Structure de la paroi intestinale (Sarah, 2011) ; B : Villosité et Cryptes chez
les volailles (Richard;
http://environnement.wallonie.be/cgi/dgrne/aerw/ied/docs/20141126/r-ducatelle-sant-
intestinale-des-vollaillesgembloux.pdf).
Il a été montré que la hauteur des villosités et la profondeur des cryptes peuvent être utilisées
comme indicateurs de la santé de l'intestin et par conséquent, sont associées à l'état des
animaux et aux performances de croissance (Liu et al. 2007). Ainsi, des villosités plus
longues et plus larges indiqueraient une plus grande surface intestinale, conduisant à une plus

27
grande disponibilité des aliments et améliorant encore la santé intestinale (Baurhoo et al.,
2007). L'activité des enzymes digestives par masse d'intestin est étroitement associée au
nombre d'entérocytes par villosités chez les poulets. Par conséquent, une plus grande activité
de l'enzyme digestive a été notée dans les hautes villosités (Uni et al., 1995). Une diminution
de la hauteur et de la largeur des villosités et de la profondeur de la crypte peut conduire à
une réduction de l'absorption des nutriments, qui est associée à certaines pathologies et
intoxications (Awad et al., 2006). Il est bien connu que l’entérite nécrotique perturbe
l'intégrité intestinale et le rapport des villosités / cryptes, réduisant ainsi l'absorption des
nutriments et les performances des oiseaux. Golder et al (2010) ont montré que les poulets
du groupe témoin (sans challenge) présentaient des hauteurs de villosités et un rapport
hauteurs de villosités / profondeur des cryptes significativement plus élevées
comparativement aux poulets challengés avec Clostridium perfringens. Les cryptes plus
profondes suggèrent un renouvellement cellulaire (un turnover cellulaire) plus rapide,
produisant de nouvelles cellules épithéliales qui migrent vers le sommet des villosités,
compensant l'inflammation ou l'effondrement des cellules endommagées par des pathogènes
ou des toxines bactériennes (Awad et al., 2009 ; Xin et al, 2018).
1.3.2.3. Les acides gras volatils
Le microbiote intestinal produit des acides gras volatils (AGV) à chaîne courte et non
ramifiée tels que l’acétate, le propionate et le butyrate ou ramifiée tels que le valérate,
l’isovalérate et l’isobutyrate. Le type d’AGV et sa quantité dépendent de la nature de la flore
microbienne dans le tube digestif des poulets et des aliments disponibles. Dans le jabot,
l’acide acétique domine, et dans les caeca, en plus de l’acide acétique présent
majoritairement, on trouve des quantités non négligeables d’acide butyrique, de petites
quantités d’acide propionique ainsi que d’autres AVG en moins grande quantité (Zhang et
al., 2003, Tableau 1-4). Wielen et al (2000) suggèrent que les acides gras volatils (en
particulier l'acétate, le propionate et le butyrate) peuvent jouer un rôle clé dans le
développement de la microflore dans les caeca des poulets pendant la croissance. Une
expérience avec des poules de chair de race Ross et Cobb a montré que les AGV ont un effet

28
négatif sur la croissance de certaines bactéries, principalement les Enterobacteriacea (Gram
négatives) et, dans une moindre mesure, sur les entérocoques mais pas sur la croissance des
Lactobacilles (Wielen et al., 2000). Les concentrations en AGV dans les caeca surtout l’acide
propionique sont corrélés à la protection contre l’infection à Salmonella Typhimurium
(Kubena et al.,2001). L'acide butyrique entraîne une diminution de la colonisation caecale
peu après l'inoculation de poulets avec S. Enteritidis et réduit la colonisation des caeca par
Salmonella (Van Immerseel et al., 2004). Les acides formique, acétique, propionique et
butyrique sont des produits du métabolisme normal de la flore intestinale anaérobie (Mead,
2000). Les Lactobacilles produisent une grande quantité d’acide lactique, acide gras à chaîne
courte non volatil (Fuller 1984). C’est un produit intermédiaire de la fermentation des
glucides qui s’accumule par suite de l’inhibition de la production d’AGV à cause du pH
inférieur à 5,5 dans le jabot. La production de cet acide est favorable aux Lactobacilles mais
défavorable aux coliformes et à la plupart des autres bactéries (Fuller 1984). Cet acide gras
à chaîne courte est présent en quantité nettement plus faible dans l’intestin grêle et les caeca.
Tableau 1-4 Les acides gras volatils le long du tube digestif du poulet (Zhang et al., 2003)1.
Volatils Acid
Quantité (ug/ mL de surnageant; moyenne ± SE)
Jabot Gésier Jéjunum Iléon Caeca
Acétate 848 ± 144 389 ± 110 210 ± 27 594 ± 100 2856 ±
270
Propionate 22 ± 5 20 ± 4 21 ± 4 17 ± 6 154 ± 48
Butyrate 15 ± 5 10 ± 4 11 ± 5 39 ± 7 869 ±
247
Isobutyrate 20 ± 2 8 ± 2 8 ± 3 8 ± 2 69 ± 15
Valérate 19 ± 10 10 ± 5 11 ± 6 14 ± 6 91 ± 27
Isovalérate 9 ± 3 10 ± 2 18 ± 2 8 ± 3 81 ± 27
1 Poulets de chair (6 animaux de 50 à 55 j) à croissance rapide (Arbor Acres) élevés au sol en conditions
expérimentales (cage), consommant un régime composé de maïs, soja, farine de poisson et huile de soja, sans
antibiotique.

29
1.3.2.4. Microbiote
Le microbiote intestinal constitue un élément important pour l’hôte. Le microbiote permet à
l’hôte d’avoir une digestion plus efficace en convertissant les composants non digestibles en
composants utilisables par l’hôte. Le microbiote fait également office d’une puissante
barrière immunitaire pour l’organisme où l’équilibre de développement de la flore
commensale inhibe le développement de la flore bactérienne pathogène. On dit même que le
microbiote a une relation de type mutualiste voir symbiotique avec l’hôte car ils coévoluent
et interagissent dès leur premier contact (Irène Gabriel, 2014). Le tractus gastro-intestinal est
adapté à la présence de bactéries commensales et a une population bactérienne dans l'intestin
grêle dans les 24 heures suivant l'éclosion (Shapiro et Sarles, 1949). La connaissance de la
composition de ce microbiote est la première étape dans la compréhension des phénomènes
qui lui sont associés. La composition du microbiote est un mélange complexe qui est
influencé par un certain nombre de facteurs, notamment l'âge des animaux, leur régime
alimentaire et l'utilisation d'antibiotiques et de probiotiques (Gong et al., 2008 ; Shang et al.,
2018). C’est un écosystème constitué de plusieurs espèces de micro-organismes (Tableau 1-
5): bactéries, champignons, virus, archées, protozoaires, etc.
Tableau 1-5 Les habitants du tube digestif des oiseaux.
Espèces Intestin grêle (1) Caeca (2)
Bactéries 92 % 97,1-97,8 %
Archées 0,4 % 0,04-2,1 %
Eucaryotes 4,2 % 05-2,1 %
Virus 3,8 % 0.1- 0,3 %
(1) Kim et Mundt, 2011. (2) Danzeisen et al., 2011 et Sergeant et al.,2014.
Dans le jabot, on trouve principalement des Lactobacilles qui sont attachés à l'épithélium et
forment presque une couche continue. On trouve aussi des Streptocoques, des coliformes et
des levures. Dans le gésier et le proventricule, le faible pH fait chuter la population

30
bactérienne (Gabriel et al. 2005). En général, l'intestin grêle du poulet contient presque 108 à
109 cfu/g de bactéries, qui sont généralement dominées par les Lactobacilles, les
Entérocoques et les Clostridiums (van der Wielen et al., 2002; Rehman et al., 2007; Kohl,
2012; Pan et Yu, 2014; Stanleyet al., 2014; Waite et Taylor, 2014) avec certaines bactéries
de la famille des entérobactéries (Bjerrum et al., 2006). Parmi les segments de l'intestin grêle,
la densité bactérienne est la plus basse au niveau de duodénum en raison du court temps de
passage, de la présence de nombreuses enzymes, d’une forte pression en oxygène, d’une
présence de fortes concentrations de composés antimicrobiens tels que les sels biliaires et de
mouvements de reflux (Shapiro et Sarles., 1949; Gabriel et al. 2005). La population
bactérienne duodénale est principalement composée de Clostridies, Streptocoques,
Entérobactéries et Lactobacilles (Waite et Taylor., 2015). Plus loin dans l'intestin,
l'environnement devient plus favorable à la croissance bactérienne en raison de la plus faible
pression d'oxygène et de la faible concentration en enzymes et en sels biliaires (Gabriel et al.
2005). Concernant l’iléon, Lu et al. (2003) ont déterminé les bactéries iléales (Figure 1-8)
par séquences de gènes d’ARNr 16S et ont trouvé que les Lactobacilles représentaient la
majorité (70%) des bactéries, suivis des Clostridiaceae (11%), Streptococcus (6,5%) et
Enterococcus (6,5%). Plus récemment, un autre groupe a montré que les Lactobacilles étaient
le genre prédominant dans l’iléon (Kumar et al., 2018). Les caeca contiennent une
communauté plus diverse de bactéries (Figure 1-8), avec des genres de Bactéroïdes,
Bifidobacterium, Clostridium, Enterococcus, Escherichia, Fusobacterium, Lactobacillus,
Streptococcus et Campylobacter (Gong et al., 2002 ; Wise et Siragusa (2007); Pruesse et al.,
2007; Oakley et al., 2013; Wei et al.,2013). De façon similaire, Zhu et al. (2002) ont identifié
243 séquences différentes représentant 50 groupes ou sous-groupes phylogénétiques de
bactéries, avec 89% des séquences appartenant à 4 groupes phylogénétiques. Pour leur part,
Apajalahti et al. (2004) ont trouvé 640 espèces différentes et 140 genres bactériens différents
dans le tractus gastro-intestinal.

31
Figure 1-8 Composition du microbiote dans l’iléon et les Caeca du poulet (Lu et al., 2003).
1.3.2.5. Biomarqueurs sanguins
Le profil biochimique sanguin a été utilisé chez plusieurs espèces pour surveiller leur santé
et détecter les maladies subcliniques. Dans la plupart des espèces animales, un marqueur
précoce important d'un processus inflammatoire en cours est un niveau élevé de Protéine C-
Réactive (CRP) (Eckersall et Bell, 2010). La CRP est une protéine de phase aiguë dans les
réponses inflammatoires et elle est stimulée par la présence à la fois d'interleukine-1 (IL-1)
et d'interleukine-6 (IL-6) (Sproston et Ashworth, 2018). Une augmentation de la
concentration de CRP est liée aux dommages cellulaires causés par les bactéries, les virus,
les champignons et les parasites dans les 36 à 48 heures après l'exposition au stimulus nuisible
(Jarosz et al.2019). Une faible concentration de CRP dans la volaille est également observée
dans le sérum d'oiseaux apparemment en bonne santé (Cray et al. 2009). Cette faible
concentration de CRP est en partie liée à la présence de lésions chroniques dans les organes
et les tissus, indiquant que la CRP peut être utilisée comme marqueur de santé général. Les
endotoxines font partie de la membrane externe de la paroi cellulaire de toutes les bactéries
à Gram négatif, libérées par excrétion ou par lyse bactérienne (Rietschel et al., 1994).
L'endotoxine dans la lumière intestinale provoque une inflammation de l'épithélium intestinal
et augmente la perméabilité paracellulaire qui provoque une altération de l'intégrité
intestinale conduisant à une absorption et une utilisation altérées des nutriments qui affectent

32
les performances de croissance de l'oiseau (Ghareeb et al, 2016). Une diminution du niveau
d'endotoxine dans le sang était due à une réduction de la libération de lipopolysaccharides à
partir des parois cellulaires des bactéries Gram-négatives, des cellules bactériennes en
croissance ou des cellules qui sont lysées en raison d'une défense efficace de l'hôte ou des
activités de certains composés antimicrobiens. Dans l'intestin sain, Les lipopolysaccharides
(LPS) ne fuit pas par la voie paracellulaire. Il est internalisé dans les cellules épithéliales et
détoxifié par la phosphatase alcaline des cellules épithéliales (Guerville et Boudry, 2016).
Par conséquent, la détection et la quantification du LPS dans le sérum pourraient être un
indicateur élégant d'une augmentation de la perméabilité paracellulaire. Malheureusement,
la plupart des techniques de mesure du LPS ne sont pas très fiables chez le poulet (Ducatelle
et al., 2018).
1.4. Alternatives aux antibiotiques facteurs de croissance
À la suite du retrait maintenant mondial de l’utilisation des AFC ajoutés dans les aliments et
le retrait graduel de ceux utilisés à titre préventif, de nombreuses alternatives ont été
développées (Patel and Goyal, 2012, Alemu et al., 2018). Le but premier de ces alternatives
est de maintenir un faible taux de mortalité, de bonnes performances de croissance pour
l’élevage et ce, tout en préservant l'environnement. Parmi ces alternatives, les plus utilisées
pour améliorer la santé gastro-intestinale sont les probiotiques, les prébiotiques, les enzymes,
les acides organiques, les immunostimulants, les bactériocines, les bactériophages, les
additifs alimentaires phytogènes, les nanoparticules et les huiles essentielles (Medhi et al,
2018). Dans cette partie on s’intéressera aux probiotiques, aux prébiotiques, aux acides
organiques et aux huiles essentielles qui sont la base de cette thèse de recherche.

33
1.4.1. Prébiotiques
Le terme prébiotique désigne les fibres fermentescibles qui ont des effets bénéfiques pour la
santé stimulant la croissance et l’activité du microbiote intestinal de façon généralement
favorable (Teng and Kim, 2018). Les fibres solubles sont davantage fermentescibles et sont
donc davantage utilisées comme prébiotiques (Carlson et al. 2018). Tel que mentionné plus
tôt les fibres se caractérisent par différentes propriétés physiques telle la solubilité, la
viscosité et la fermentesciblé qui sont responsables de leurs effets physiologiques. Les fibres
solubles visqueuses ralentissent la vidange gastrique et prolongent le transit de l’intestin
grêle. Les fibres solubles, qui ont des effets hypocholestérolémiants, réduisent la glycémie et
l’insulinémie postprandiales. La production d’acides gras à chaîne courte lors de la
fermentation exerce plusieurs effets bénéfiques sur la santé. Par exemple, les prébiotiques
contribuent à rendre le microbiome intestinal plus sain, car ils stimulent la production de
bifidobactéries et de lactobacilles et inhibent la croissance des populations de bactéries
pathogènes en réduisant le pH fécal par la production d’acides gras à chaîne courte (Slavin,
2013; Voreades et al., 2014; Carlson et al., 2018) et favorisent également la maturation du
système immunitaire et renforcent la fonction de la barrière intestinale (Carlson et al., 2018).
Le butyrate, dont il sera question dans une section à venir, est particulièrement bénéfique, en
raison de ses propriétés anti-inflammatoires et anti-oxydantes (Slavin, 2013).
Les prébiotiques peuvent être synthétiser chimiquement ou se retrouver naturellement dans
les céréales. Les principaux prébiotiques utilisés en alimentation animale sont les mannan
oligosaccharides, les B-glucanes et les fructanes.
Mannan-oligosaccharides (MOS) sont dérivés des parois cellulaires de levure
(Saccharomyces cerevisiae) et sont riches en mannoprotéines (12,5%), mannane (30%) et
glucane (30%) (Baurhoo et al., 2009). Les oligosaccharides de mannane sont connus pour
leur capacité à se lier aux bactéries pathogènes, comme les E. coli et Salmonella (Spring et
al., 2000). Des études ont indiqué qu’une supplémentation en MOS de 0,08 à 0,5% pourrait
modifier la composition de la flore microbienne en augmentant le total des bactéries
anaérobiques tel que, Lactobacillus, Bifidobacterium, et en diminuant les bactéries

34
pathogènes tels que Salmonella, Escherichia coli, Clostridium perfringens et Campylobacter
(Spring et al., 2000; Baurhoo et al., 2009). Il est intéressant de noter que la MOS a augmenté
les Bacteroidetes caecaux chez des poulets de chair âgés de 7 et 35 jours (Lee et al., 2016)
qui est connus pour leur forte activité métabolique. Ils peuvent fermentent efficacement les
polysaccharides non digestibles en AGV et, par conséquent, améliorent l'absorption des
nutriments et protègent l'hôte d'une infection pathogène (Wexler, 2007).
Le β-glucane est un prébiotique dérivé de la levure ou des parois cellulaires des céréales, des
bactéries et des champignons. Ces polysaccharides à longue chaîne sont composés de
monomères de D-glucose avec des liaisons de liaisons β-glycosidiques 1-3, et ses chaînes
latérales sont liés par les liaisons 1 à 6. On suppose que les β-glucanes peuvent réguler le
système immunitaire en induisant la prolifération des cellules immunitaires pour attaquer les
cellules infectées par des agents pathogènes (Teng et Kim., 2018). Le β-glucane peut être
reconnu par récepteurs sur les cellules sentinelles, déclenchant la production de cytokines et
prolifération de lymphocytes (Swiatkiewicz et al., 2014). De plus, l'exposition au β-glucane
déclenche également la prolifération des macrophages et améliore la capacité phagocytaire
des macrophages (Cox et al., 2010; Guo et al., 2003).
Fructanes sont des extraits de différentes plantes, hydrolysés à partir de polysaccharides, ou
produits par des micro-organismes, ont été administrés récemment dans les régimes de
poulets de chair. Les fructanes sont classés en trois types distincts: le groupe inuline, le
groupe levan, et le groupe ramifié. Premièrement, le groupe de l'inuline, également connu
par les fructooligosaccharides (FOS) qui peuvent être divisés en différents catégories basées
sur les degrés de polymérisation (DP): Inuline, extraite de racines de chicorée (Cichorium
intybus L.), Oligofructose (OF), qui peut être généré par hydrolyse partielle de l'inuline et
conversion enzymatique de saccharose ou de lactose. Deuxièmement, le groupe levan est un
autre groupe de fructanes, qui sont principalement liés par des liaisons β-2,6
fructosylfructose. Enfin, les fructanes ramifiés contiennent à la fois des liaisons β-2,1
fructosyl-fructose et β-2,6 fructosylfructose en quantité équivalente (Zhao et al.,2013). C'est

35
la liaison β-glycosidique en fructanes qui résistent à leur dégradation par les enzymes
digestives chez les volailles et améliore la population de bactéries bénéfiques, comme les
bifidobactéries et les lactobacilles, et élimine les niveaux de bactéries pathogènes, telles que
Clostridium perfringens et E. coli, dans l'intestin des poulets de chair (Kim et al., 2011, Ricke,
2015).
Autres prébiotiques Outre que les trois principaux prébiotiques, MOS, β-glucane et
fructanes, d'autres oligosaccharides ont été pris en compte comme prébiotiques potentiels, y
compris les oligosaccharides de chitosane (COS) (Extrait de la chitine, le COS contient 2 à
10 unités de sucre de Nacétyl glucosamine avec 1 à 4 liaisons β), les galacto-oligosaccharides
(GOS) (prébiotiques synthétiques avec galactose avec 1–4 ou 1–6 liaisons β, sont
normalement produites à partir de lactose par l'enzyme lactase à haute activité
galactosyltransférase), le galactoglucomannane oligosaccharide (GGMO) (Oligosaccharides
de galactoglucomannane et galactoglucomannane, les oligosaccharides-arabinoxylane
(GGMO-AX) sont de nouveaux prébiotiques extraits et transformés à partir des copeaux de
bois des résineux (Capek et al., 2000). Ces oligosaccharides consistent en mannose, glucose
et monomères de galactose et xylo-oligosaccharides (XOS) (sont des oligosaccharides,
constitués de unités de sucre de xylose avec des liaisons β (De Maesschalck et al., 2015).
Xylan, le principal composant de fibre de céréales comme les épis de maïs, les pailles, les
coques et le son sont les ressources brutes pour la production XOS (Mussatto et Mancilha.,
2007). Xylan pourrait être dégradé en XOS par la xylanase de champignons, par la vapeur
ou dilué en solutions d'acide minéral (Mussatto et Mancilha., 2007).
1.4.1.1. Céréales à effet prébiotique
Avoine nue
Le Canada est un joueur important dans les domaines de l'amélioration génétique et de
l'utilisation de l'avoine nue ou sans écale (Agriculture Canada, 1993). Cependant, jusqu’à
présent son incorporation dans les aliments des animaux d’élevage et des volailles en

36
particulier n’est pas très répandue. L'avoine nue est un type d'avoine dont l'écale fibreuse
entourant l'amande du grain a été pratiquement éliminée par le biais de la sélection génétique
(Alain, 2001). Ainsi, les grains de l’avoine nue ont un profil nutritionnel très intéressant
comparativement à l'avoine traditionnelle par une concentration élevée en protéine et gras.
On remarque que son taux de protéine est de 30 % supérieur à l'avoine traditionnelle, alors
que sa teneur en ADF est 3 fois plus faible plutôt comparable au blé (NRC, 2001). De plus,
puisque la concentration en gras et en protéines de l'avoine nue est plus élevée et la teneur en
fibres est plus faible que l'avoine traditionnelle, l'énergie métabolisable volailles sera
supérieure (16.5 MJ/kg pour avoine nue vs 15 et 14.2 MJ/kg pour le blé et orge
respectivement ; Hoffman, 1995). Pour la composition minérale, il n'y a pas de différences
notables entre les deux avoines à l'exception du niveau de calcium qui est plus faible et du
niveau de phosphore dont la concentration est plus élevée pour l'avoine nue. De plus, après
l’orge elle est la deuxième céréale qui contient le plus de bêta-glucanes (β-glucanes) qui sont
des polysaccharides non-amylacés et qui sont connus pour avoir des effets prébiotiques donc
bénéfiques sur la composition de la flore bactérienne et l’inflammation dans l’iléon et le
colon chez le porc (Pieper et al., 2008). En plus du β-glucane, l’avoine nue contient d'autres
polysaccharides non amylacés tels que l’arabinoxylane et le xylane qui sont aussi connus
pour modifier la composition bactérienne et l'expression des facteurs inflammatoires dans
l'iléon et le colon des porcelets sevrés (Friend et al., 1988). L’avoine nue aurait donc une
activité antimicrobienne et pourrait modifier la composition bactérienne de l'intestin, puis
agir positivement sur le développement de la muqueuse intestinale. Un bénéfice pour la santé
digestive des volailles peut être noté grâce à l’effet prébiotique des fibres solubles de
l’avoine. Des chercheurs australiens (Farrell et al., 1991) ont démontré que les diètes? de
poulets de chair élevés avec de l’avoine nue doivent être granulés pour une performance
maximale. L’avoine nue a été ajoutée jusqu’à 84% de l’aliment des poussins et a entraîné
une performance de poulets de chair semblable à ce qui a été réalisé avec des régimes à base
de blé (Cave et al., 1989). Plusieurs recherches indiquent que jusqu'à 60% d'avoine nue
peuvent être incluses dans les régimes des volailles, en remplaçant le maïs et le tourteau de
soya (Maurice et al., 1985; Cave et al., 1989; Burrows, 2005). Torrallardona et al. (2000) ont
également montré qu’une consommation alimentaire post-sevrage plus élevée chez les
porcelets recevant des régimes de pré-sevrage contenant du riz ou de l'avoine nue, qui sont

37
connus pour avoir un amidon moins résistant et une meilleure digestibilité de l'amidon
(Tableau 1-6). Dans des études plus anciennes, l'inclusion de l'avoine nue dans les régimes
post-sevrage des porcelets a augmenté la consommation de porcelets sevrés nourris avec des
aliments à base de maïs (Brand et van der Merwe, 1996). De nouvelles variétés d'avoine nue
offrent la possibilité d'ajouter au porcelet sevré une source d'amidon hautement digestible qui
pourrait affecter positivement la croissance mais aussi le microbiote intestinal pendant la
période pré-sevrage.

38
Tableau 1-6. Effet de l’incorporation de l’avoine nue sur les performances de croissance de poulet et le porc.
Références Espèces Taux
d’incorporation
avoine nue (%)
Régime
alimentaire
Effet sur les performances de
croissance
Mahan and Newton, 1993 Porc - Maïs- Blé consommation
Brand et van der Merwe, 1996 Porc - Maïs- Blé consommation
Torrallardona et al. (2000) Porc - Maïs- Blé gain de poids quotidien
Flis et al.,2007 Porc 45 Triticale NS
Farell et al.,1991 Poulet 84 Blé NS
Hetland et al., 2003 Poulet 40 Maïs- Blé consommation
Svihus et al,2002 Poulet 18 Blé gain de poids quotidien
Burrows, 2005 Poulet 60 Maïs- Blé gain de poids quotidien
Szymczyk et al.,2005 Poulet 30 soja, maïs, blé gain de poids quotidien
Szymczyk et al., 2007 Poulet 50 soja, maïs, blé gain de poids quotidien
NS : effet non significatif

39
Son de blé
Le son de blé peut être une alternative économique et nutritionnelle pour l'alimentation des
animaux (Mateos et al., 2012). Il est le sous-produit obtenu après séparation de la farine de
blé par tamisage. Il a une teneur en protéines adéquate pour les volailles et des niveaux élevés
de fibres brutes (NRC, 1994), mais moins d'énergie métabolisable que de nombreux autres
ingrédients comme le maïs, le sorgho et l'orge (NRC, 1994). Souci et Kirchhoff (2008) ont
rapporté qu’il contient une quantité élevée de fibres alimentaires totales (451g / kg), de
protéines (160g / kg), de gras (47g / kg), de glucides (177g / kg) et de minéraux (61,5 g / kg).
Des études ont montré les effets positifs de l'utilisation de son de blé combiné ou non avec
des enzymes de type xylanases, sur les performances de croissance, la microflore intestinale,
la production d'œufs et la digestibilité de certains nutriments chez les volailles (Ali et al.,
2008 ; Courtin et al., 2008). De plus, Van Craeyveld et al. (2008) ont constaté que la
supplémentation alimentaire de son de blé riche en arabinoxylooligosaccharides (AXOS)
avait des effets prébiotiques et compétitifs en augmentant la colonisation intestinale par
Lactobacillus et Bifidobacterium et la production d’AGV. Les AXOS, sont le résultat de la
dégradation d’arabinoxylan (AX) par la xylanase qui agissent comme des prébiotiques dans
l'écosystème de l'intestin, en stimulant sélectivement les organismes bénéfiques et en
supprimant la croissance des organismes pathogènes. Les effets prébiotiques potentiels des
AXOS comprennent l'optimisation de la fonction du côlon, l'augmentation ou la modification
de la composition des AVG et la stimulation du système immunitaire. Courtin et al. (2008),
ont mené deux essais d'alimentation réalisés sur des poulets nourris avec du blé ou du maïs
comme composant principal. La supplémentation du son de blé à 0,5% pour le régime à base
de blé ou à 0,25% pour les régimes alimentaires à base de maïs améliorait significativement
le taux de conversion alimentaire sans augmenter le poids corporel des animaux, indiquant
ainsi une amélioration de l'efficacité de l'utilisation des nutriments. Les AXOS ont augmenté
significativement le taux de Bifidobactéries mais pas de bactéries totales dans les caeca des
poulets. Ces données suggèrent que le son de blé a des propriétés nutritionnelles intéressantes
et peut agir comme prébiotique. Dans le même concept, Ebersbach et al. (2012) ont rapporté
que XOS inhibait l'adhésion des pathogènes aux entérocytes in vitro. Keerqin et al. (2017)
ont examiné l'effet des AXOS sur les performances des poulets, le pH gastro-intestinal et la

40
production des AVG dans les caeca avec ou sans challenge à l’entérite nécrotique. Les
résultats de cette étude suggèrent que l’AXOS semblait être un prébiotique efficace en
améliorant le taux de conversion alimentaire et en augmentant la production des AVG dans
les caeca. Teng et al. (2017) ont constaté que le traitement avec 10% de son de blé fermenté
joue un rôle dans l’amélioration de la santé digestive et pourrait augmenter de façon
significative la hauteur des villosités iléales. D'autre part, le son de blé a des propriétés
bénéfiques qui peuvent maximiser la performance de croissance des poulets. D'après
Jørgensen et al. (1996), lorsqu'il est inclus à des niveaux modérés dans le régime alimentaire,
le son de blé améliore les performances de croissance, la digestibilité des nutriments et
l'activité microbienne chez les poulets de chair. Eeckhau et al. (2008) et Vermeulen et al.
(2017) ont signalé une réduction de la colonisation par Salmonella enteritidis chez les poulets
de chair supplémentés avec du son de blé et Akhtar et al. (2012) ont montré une meilleure
réponse immunitaire lorsque le son de blé a été inclus dans le régime alimentaire.
Martinez et al. (2015) ont montré que l'inclusion de 150 g de son de blé / kg en remplacement
partiellement le maïs dans le régime des poulettes pondeuses nourries pendant la phase de 10
à 16 semaines, favorisait une meilleure croissance par rapport aux 100 et 200 g de son de blé
/ kg. Zeisel et al. (2003) ont trouvé que le son de blé contient une grande quantité de bétaïne
(1505,6 mg / 100 g). La bétaïne est omniprésente chez les plantes, les animaux, les micro-
organismes et les sources alimentaires riches, y compris les fruits de mer, les épinards et le
son de blé. Elle protègerait les cellules intestinales des poussins contre les infections
coccidiennes, soulage les signes cliniques et améliore les performances (Fetterer et al., 2003,
Kettunen et al., 2001a). Le principal rôle physiologique de la bétaïne est de fonctionner
comme un donneur de méthyle et un osmolyte, pour maintenir l'équilibre cellulaire
osmotique et de balancer les ions afin d'améliorer la capacité des poulets à résister au stress
thermique en empêchant la déshydratation et l'inactivation osmotique (Kettunen et al.,
2001b). La bétaïne peut favoriser divers microbes intestinaux contre les variations
osmotiques et ainsi améliorer l'activité de fermentation microbienne (Saeed et al., 2017). Des
études antérieures ont montré que la supplémentation alimentaire de bétaïne dans les régimes
de volaille pourrait affecter positivement la digestibilité des nutriments, réduire le gras

41
abdominal et augmenter le rendement de la viande de poitrine. De plus, la bétaïne protège les
organes internes et améliore les performances des oiseaux. Son inclusion dans le régime
alimentaire de la volaille favorise les AA essentiels comme la méthionine. En outre, il peut
jouer un rôle important dans la production de viande maigre en affectant positivement le
métabolisme des lipides avec un catabolisme accru des acides gras et en réduisant ainsi le
dépôt de graisse des carcasses (Saeed et al., 2017).
Orge
L'orge peut être incluse dans les aliments pour volailles en tant que source d'énergie. Les
hydrates de carbone dans l'orge ne sont pas aussi faciles à digérer en raison de la présence de
polysaccharides non amylacés (NSP) en particulier les β-glucanes. La graine d'orge contient
généralement entre 52 et 60% d'amidon et 22% de fibres totales. Les β-glucanes sont des
polysaccharides non-amylacés et qui sont connus pour avoir des effets prébiotiques
bénéfiques. En revanche, il existe dans la littérature plusieurs résultats de recherche
controversés sur le β-glucanes. White et al. (1981) ont confirmé l'effet antinutritionnel des β-
glucanes, en isolant et en incluant les β-glucanes dans les régimes alimentaires des poulets
de chair. Il a été montré que les β-glucanes affectent l'activité des enzymes endogènes et
modifient la morphologie intestinale et la microflore. Par conséquent, non seulement la
digestion et l'absorption des nutriments sont fortement réduites, mais le taux de passage du
digesta est également affecté (Jacob et Pescatore, 2014). Plus récemment, Arena et al (2014)
ont trouvé que les β-glucanes de l'orge étaient capables d'améliorer le taux de croissance des
bactéries probiotiques, avec une influence sélective sur les espèces de Lactobacillus
plantarum. Notamment, la stimulation du métabolisme des microorganismes bénéfiques,
ainsi que la sélectivité, sont parmi les principales caractéristiques revendiquées pour les
produits alimentaires prébiotiques. L'alimentation des volailles en grains entiers est une
pratique de gestion courante en Europe, en Australie et au Canada (Biggs et Parsons, 2009).
Biggs et Parsons (2009) ont noté que l'alimentation de l'orge entière aux poussins (New
Hampshire × Columbian) améliorait la digestibilité des nutriments, en particulier pour
l’énergie métabolisable et les AA. Taylor et Jones (2004a) ont rapporté que l'inclusion de

42
20% d'orge à la ration a entraîné des altérations du développement de l'intestin, comme un
proventricule plus petit, un gésier plus gros et une meilleure performance globale. Avec
l'inclusion de 20% d'orge, des changements de la viscosité intestinale n'ont pas été observés
au-delà du duodénum (Taylor et Jones, 2004b). De même, Nahas et Lefrançois (2001) ont
rapporté que la croissance des poulets nourris avec des régimes contenant jusqu'à 20% d'orge
étaient similaires à ceux des poulets nourris au maïs et au soja avec une teneur similaire en
énergie et en protéines. A noté aussi qu’il y a une variété d’orge nue (Synergie, 2015-2016).
L’orge nue se dépouillant de ses glumes au champ, la teneur en toxines est ainsi
significativement diminuée. Les recherches sur l’effet de l’orge nue dans l’alimentation de
poulet de chair ne sont pas encore bien développées.
Seigle
Les principaux NSP solubles dans le seigle sont les arabinoxylanases et β-glucanes (Bach
Knudsen, 1997). Ces NSP solubles présents dans les parois des cellules de l'endosperme du
seigle ont été identifiés comme une cause d'un faible taux de croissance et d'une faible
digestibilité des nutriments chez les poulets de chair (Ward et Marquardt, 1988). Les résultats
de Mathlouthi et al. (2002) montrent que l'inclusion du seigle au lieu du maïs dans le régime
alimentaire a considérablement réduit les performances de croissance. Les performances
réduites chez les oiseaux nourris au seigle étaient au moins dues à la réduction de la
digestibilité des nutriments. Néanmoins, le seigle est la céréale menant à la plus haute
visqueuse du digesta (Danicke et al., 1999). L'augmentation de la viscosité digestive est donc
le principal mécanisme par lequel les NSP solubles exercent des propriétés antinutritives
(Bedford et Classen,1992). De plus, le régime à base de seigle augmente la consommation
d'eau et le rapport eau / alimentation (Langhout et al., 1997). Un autre effet de l’incorporation
du seigle dans l'alimentation des poulets est la forte stimulation de la déconjugaison précoce
des acides taurochénodésoxycholique et taurocholique, qui sont les acides biliaires
prédominants dans la bile de poulet (Elkin et al.,1990) Cette effet serait toutefois contrecarré
par une supplémentation enzymatique par des enzymes endogènes. De plus, Mathlouthi et al.
(2002) ont indiqué que le seigle a réduit non seulement la digestibilité des graisses, mais aussi

43
réduit la digestibilité des protéines. Cela pourrait s'expliquer par la prolifération de bactéries
dans l'intestin grêle, qui augmente la perte d'azote endogène (Smits et al.,1997) en
incorporant des acides aminés dans des protéines microbiennes (Salter et Coates, 1971) et
réduit ainsi la digestibilité apparente des protéines (Angkanaporn et al., 1994).
1.4.1.2. Les enzymes exogènes à effet prébiotique
Les enzymes sécrétées par le tube digestif et le pancréas sont indispensables pour la digestion
des aliments chez tous les animaux. Ces enzymes sont donc principalement produites par
l’animal ou peuvent provenir de microbes qui sont naturellement présents dans le tractus
digestif des animaux. La présence de protéines indigestibles et d’éléments antinutritionnels
dans les régimes alimentaires du porc et du poulet peut diminuer l’efficacité des enzymes
endogènes, ce qui explique que 15% à 25% des aliments ingérés par le porc et le poulet ne
sont pas digérés et ainsi les nutriments associés ne sont pas absorbés (Bedford et Partridge,
2010). L’incorporation dans les régimes de ces enzymes (apport exogène) est devenue une
voie de choix afin d’améliorer la digestibilité des aliments ou encore éliminer les facteurs
antinutritionnels. Les principales enzymes utilisées dans l'alimentation animale sont les
xylanases, les β-glucanases et les phytases (Aehle 2004, Kirk et al. 2002). Les xylanases et
les β-glucanases aident à la digestion des polysaccharides chez les monogastriques, qui,
contrairement aux ruminants, ont une capacité limitée à digérer complètement les aliments
d'origine végétale contenant de grandes quantités de cellulose et d'hémicellulose. La plupart
des enzymes alimentaires commerciales sont produites par des bactéries et des champignons
filamenteux (Aehle 2004, Kaur et al. 2007). Cependant, les enzymes alimentaires sont de
plus en plus produites de manière hétérologue dans les levures avec l'avènement de systèmes
d'expression hautement efficaces (Aehle 2004).
Les xylanases appartiennent aux familles des glycosides hydrolases GH10 et GH11, mais
aussi aux familles GH5, 7, 8 et 43. La famille GH10 à laquelle appartient l’endo-1,4-β-
xylanases et l’endo-1,3-β-xylanases comprend des xylanases d’origine végétale, bactérienne

44
et fongique, tandis que la famille GH11 ne comprend que les xylanases d’origine fongique
(Chakdar et al., 2016). Les xylanases ont été récupérées dans des Bacillus, des Streptomyces
ainsi que dans d’autres bactéries sans aucun effet pathogène sur les végétaux (Motta et al.,
2013). Ces bactéries ont attiré l’attention grâce à la production de xylanases thermostables et
alcalines avec un pH optimal supérieur à celui des xylanases fongiques. En plus des bactéries,
les xylanases peuvent être produites à partir des champignons filamenteux dont les
principaux sont Trichoderma, Aspergillus, Fusarium, et Pichia. Ces derniers produisent des
quantités d’enzymes plus importantes que les bactéries et les levures (Motta et al., 2013).
Les β-glucanases hydrolysent les liaisons 1,4 ou 1,3 trouvées dans les glucanes mixtes,
perturbant l'intégrité de la paroi cellulaire endospermique. Les β-glucanes à liaison mixte
sont également appelés (1,3 ou 1,4) -β-d-glucanes ou β-glucanes de céréales. Ce sont des
polysaccharides linéaires non ramifiés dans lesquels les monomères β-d-glucopyranosyle
sont polymérisés à la fois par des liaisons (1,4) et (1,3) (Saraol et al., 2011). Il a été rapporté
que dans l'orge, les β-glucanes contiennent environ 70% (1,4) de liaisons et 30% (1,3) de
liaison, dans lesquels des segments de deux ou trois (1,4) liaisons sont séparés par un seul
(1,3) -liens. Les structures résultantes des β-glucanes dans les céréales contiennent 58% à
72% d'unités liées β- (1,3) et 20% à 34% d'unités liées β- (1,4) (Parrish et al., 1960). Les β-
glucanases fongiques sont également fabriquées par des champignons du groupe Aspergillus.
Ils sont souvent formés comme activité secondaire dans la préparation de la pectinase.
(Villettaz et al., 1984; Dubourdieu et al., 1985).
Annison (1992) a démontré que la xylanase réduit la viscosité du contenu intestinal grâce à
la réduction du poids moléculaire du xylane soluble provoquant ainsi un temps de contact
plus long entre les enzymes et leurs substrats pour une meilleure digestibilité des nutriments.
En fait, le milieu visqueux peut constituer une barrière qui ralentit et empêche l’accès des
enzymes à leurs substrats (White et al., 1981). Ainsi, la diminution de la couche d’eau
augmente la vitesse de passage des nutriments au niveau de l’intestin des animaux (White et
al., 1981). Bedford et Classen (1992) ont montré que la xylanase engendre une diminution

45
de la viscosité du contenu intestinal à toutes les concentrations de seigle dans l’aliment : 0,
200, 400, et 600 g/kg (Figure 1-9). En revanche, la concentration d’enzyme optimale pour la
diminution de la viscosité augmente avec la concentration du seigle dans la ration. De ce fait,
il se pourrait que les facteurs à l’origine de cette viscosité soient surtout présents à des
concentrations élevées de seigle ou bien que les xylanes du seigle soient plus résistants à
l’hydrolyse comparée à ceux présents dans le blé. En fait, ceci est conforme aux résultats
cités ci-dessus concernant la forte viscosité trouvée dans le seigle comparé aux autres céréales
Figure 1-9 Effet de la concentration du seigle et de la xylanase sur la viscosité de l’intestin
grêle proximal chez les poulets (tiré de Bedford et Classen, 1992).
Kosieradzka et al. (2003) ont montré que l'addition de β-glucanase a augmenté la
digestibilité, a entraîné une digestibilité accrue de la fraction de fibres et de la valeur
énergétique de l'avoine. L'addition de ß-glucanase produit une augmentation d'environ 4%
de l'énergie métabolisable apparente de l'avoine nue pour les poulets de chair. Cet effet était
associé à une diminution de 70% de la viscosité jéjunale chez les poulets de chair recevant
un aliment contenant 500 g/kg d'avoine nue (Macleod et al., 2008). L'ajout de xylanase et de
β-glucanase au régime à base de seigle a amélioré la digestibilité des nutriments et les

46
performances des poulets de chair. Cet effet bénéfique des enzymes exogènes a déjà été
rapporté dans de nombreuses études (Choct et al., 1995; Scott et al.,1998). De plus, la
consommation d'eau était réduite lorsque les poulets de chair ont été nourris avec un régime
à base de seigle supplémenté de xylanase et de β-glucanase. Cela est en accord avec le résultat
obtenu par Yasar et Forbes (2000), et pourrait s'expliquer par l'hydrolyse des NSP dans
l'alimentation à base de seigle par la xylanase et la β-glucanase, qui diminuent leur capacité
de rétention d'eau. De plus, la supplémentation enzymatique restaure la capacité d'absorption
de la muqueuse intestinale en augmentant la taille des villosités intestinales, et il a supprimé
la déconjugaison précoce des acides biliaires. Ceci est probablement dû à une diminution des
bactéries qui hydrolysent la bile conjuguée acides (Smits et Annison, 1996).
1.4.2. Les probiotiques
L’utilisation des probiotiques a débuté au début des années 1900 (Patterson et Burkholder,
2003). Ils s’agissent de préparations microbiennes vivantes (WHO, 2001) utilisées comme
additifs alimentaires. Les principaux modes d’action sont la stimulation du système
immunitaire par l’exclusion compétitive des agents pathogènes (Hassanein et Soliman,
2010 ; Li et al. 2016), la production d'anticorps (Pan et Yu, 2014), la diminution de l'apoptose
cellulaire (Khan et al., 2016) et l’amélioration de la fonction barrière de l’intestin grâce à la
fortification des bactéries bénéfiques de la microflore intestinale (Bai et al., 2013). Les
probiotiques les plus utilisés dans les produits développés pour l’alimentation animale sont
les Lactobacillus acidophilus, Enterococcus faecium, les espèces de Bacillus,
Bifidobacterium bifidum et la levure Saccharomyces cerevisiae (Jacela et al., 2010). Dans le
même cadre, la supplémentation par d’autres types de probiotiques (tel que Lactobacillus
casei, Lactobacillus acidophilus, Bifidobacterium bifidum, Enterococcus faecium) améliore
aussi la croissance, l'efficacité alimentaire et la santé intestinale (Samli et al., 2007 ;
Giannenas et al., 2012 ; Ghasemi et al., 2014). Cette amélioration est obtenue par une
réduction du pH intestinal, un changement de la composition bactérienne intestinale et une
meilleure activité digestive. L'administration d'Enterococcus faecium dans l'alimentation du

47
poulet a eu un effet antibactérien sur la microflore bactérienne dans l'intestin grêle (Levkut
et al., 2012). Des résultats similaires ont été rapportés avec Streptomyces sp. (Latha et al.,
2016) et Bacillus subtilis (Zhang et al., 2013). D’autres travaux de recherche ont montré que
l’incorporation de levures entraîne une augmentation dans les digestibilités du calcium et du
phosphore ainsi que dans les digestibilités de la protéine brute, de l’énergie brute et de
certains acides aminés chez les poulets pendant la période de démarrage (Bai et al., 2013).
Les probiotiques augmentent le ratio longueur des villosités sur profondeur des cryptes dans
l’iléon (Afsharmanesh et Sadagh 2013). De plus, ils augmentent la quantité de Lactobacilles
dans l’iléon et diminuent la population de Clostridium perfringens dans le caecum, mais
n’affectent ni la morphologie intestinale, ni le pH, ni les concentrations d’AGV dans l’iléon
(Chen et al., 2015). Les probiotiques peuvent aussi jouer un rôle anticoccidien Giannenas et
al. (2012) suggèrent que le traitement avec des probiotiques a atténué l'impact de coccidiose
sur les poulets par un effet coccidiostatiques sur Eimeria tenella minimisant le risque et la
propagation de la coccidiose.
1.4.3. Les acides organiques
Les acides organiques sont des composés ayant un rôle de correction d’acidité qui sont
utilisés dans l’alimentation des poulets depuis très longtemps. Les acides organiques sont des
agents de conservation utilisés pour protéger contre la prolifération microbienne et fongique
(kum et al., 2010). L'action antimicrobienne de l'acide organique est due au fait que les acides
non dissociés peuvent diffuser à travers le système lipophile (Cherrington et al., 1991) Les
acides organiques seraient ainsi efficaces contre E. coli et Campylobacter, mais n’auraient
pas d’effet démontrés sur C. perfringens (M’Sadeq et al., 2015). Chez le poulet, ces derniers
sont encapsulés pour s’assurer qu’ils ne soient pas rapidement digérés ou (Bolton and Dewar
(1965). Les acides propionique, lactique et sorbique ont notamment été rapportés pour
diminuer la colonisation de bactéries pathogènes et la production de métabolites toxiques
pour améliorer la digestibilité des protéines et des minéraux (Ca, P, Mg, Zn) (Richards et al.,
2005). Les acides organiques peuvent être une bonne alternative aux antibiotiques en

48
réduisant la production de composés toxiques bactériens qui viennent altérer la morphologie
de l’intestin par la réduction de la colonisation de l’épithélium intestinale par des pathogènes
(Sultan et al. 2015). L'activité antibactérienne des acides organiques sur le Campylobacter
spp, E. coli, Salmonella et Listeria monocytogenes a été rapportée (Chaveerach et al., 2004 ;
Skrivanova et al., 2006 ; Over et al., 2009). Dans l'alimentation des volailles, des acides
organiques à chaîne courte tels que l'acide formique (C1), l'acide acétique (C2), l'acide
propionique (C3) et l'acide butyrique (C4) ont été particulièrement étudiés. D'autres acides
carboxyliques à chaîne courte utilisés comprennent les acides citrique, lactique, fumarique et
malique (Dibner et Buttin, 2002). Des études ont montré que l'addition d'acides organiques
aux aliments pour poulets de chair favorise la croissance, le taux de conversion des aliments
et leur utilisation (Hassan et al., 2010). L'ajout d'acides organiques dans l'eau de boisson
confèrerait aux jeunes poussins une protection contre l'infection à Campylobacter
(Chaveerach et al., 2004). Ces acides ont également une action protectrice contre E. coli (Izat
et al., 1990). Ainsi, Mohammadagheri et al. (2016) ont montré qu'une supplémentation en
acide citrique (2%) peut améliorer la prolifération épithéliale de cellules et la hauteur des
villosités. L’administration dans l’eau des acides formique et propionique a amené des
populations bactériennes plus stables et augmente surtout la population des Lactobacillus
dans l'iléon des poulets. Ces résultats montrent que les acides organiques peuvent être utilisés
comme alternative aux AFC pour réduire les bactéries pathogènes dans le tractus gastro-
intestinal (Nava et al., 2009).
Parmi les acides organiques, le butyrate augmente la production de protéines de jonction dans
les entérocytes, ce qui réduit la perméabilité de l'épithélium intestinal à l'invasion par des
agents pathogènes (Adil et al., 2010 ; Andreopoulou et al., 2014). Sunkara et al. (2011) ont
rapporté que le butyrate peut induire la synthèse de peptides de défense de l'hôte chez les
poulets induisant ainsi la réduction de Salmonella dans les caeca, ainsi que l'augmentation de
l'activité des monocytes contre l’entérite. L'administration de l'acide butyrique a également
montré une diminution de l'excrétion des bactéries à Gram négatif, et des infections
bactériennes intestinales, y compris la salmonelle (Fernandez-Rubio et al.,2009), a amélioré
les performances, a augmenté la résistance à l’entérite nécrotique (Van Immerseel et al.,

49
2005) et a changé la morphologie des villosités chez les poulets (Hu and Guo, 2007). Le
butyrate est une source d'énergie préférée pour les entérocytes. Cela se traduira par un
meilleur développement des villosités intestinales, mais aussi par une muqueuse intestinale
plus solide chez plusieurs espèces. Chez le poulet, Leeson et al (2005) ont trouvé des
villosités plus longues dans le duodénum des poulets recevant de l'acide butyrique dans
l'alimentation que les poulets témoins.
1.4.4. Les huiles essentielles
Les huiles essentielles sont des huiles volatiles extraites des plantes obtenues par la
distillation à la vapeur d'eau ou par l'extraction à l’aide d’un solvant organique (Basmacioğlu-
Malayoğlu et al., 2016). Les huiles essentielles peuvent être soit naturelles soit synthétiques
(Mehdi et al., 2018). Les plus utilisés sont le thymol, le trans-cinnamaldéhyde, le carvacrol
et l'eugénol. Leur mode d'action réside dans leur interférence avec le système enzymatique
des bactéries et la modulation des réponses immunitaires et de l'inflammation (Miguel, 2010).
Les phénols (thymol et carvacrol) constituent environ 80% des huiles essentielles d'origan,
l'espèce la plus répandue de la famille des Lamiacées, et sont principalement responsables de
ses activités antibactériennes et antioxydantes. Outre le thymol et le carvacrol, le ρ-cymène
a été trouvé comme un autre composant dominant de l'origan (Bouhaddouda et al., 2016).
Bien que le ρ-cymène ne soit pas un agent antimicrobien efficace en soi, il pourrait faciliter
le transport du carvacrol à travers la membrane cytoplasmique (Oke et al., 2009). La
littérature sur l’utilisation des huiles essentielles dans l’alimentation est riche. En revanche,
il est difficile de tirer des conclusions générales compte tenu de la complexité des mélanges
de molécules dans les produits. Des taux d’incorporation élevés sont à risques d’induire des
interactions avec d’autres composés de la ration (M’Sadeq et al., 2015). Bien que ces
composés soient reconnus comme ayant des propriétés antimicrobiennes, les modes d’action
ne sont pas encore bien définis. Le principal mode d'action reconnu est leur interférence avec
le système enzymatique de la bactérie et la modulation des réponses immunitaires et
inflammatoires (Mehdi et al., 2018). Plusieurs études ont montré que la supplémentation de

50
l’aliment avec des huiles essentielles améliore les performances de croissance. En
comparaison avec l’aliment témoin, de nombreuses études ont montré que les aliments
supplémentés à différents niveaux d’huile essentielle montraient soit des performances
améliorées ou équivalentes à l’aliment témoin en termes de gain de poids et de d’indice de
consommation. (ex. : Ghazanfari et al., 2015; Peng et al., 2016; Basmacioğlu-Malayoğlu et
al., 2016). De plus, les huiles essentielles peuvent jouer un rôle préventif et curatif dans le
traitement de l'entérite nécrotique chez les poulets de chair (Jerzsele et al., 2012). Saini et al.
(2003a et 2003b) ont montré des lésions relatives à l’entérite nécrotique (Saini et al., 2003a)
et à la coccidiose (Saini et al., 2003b) avec l’ajout d’huile essentielle d’origan dans l’aliment
des poulets de chair non significativement différentes de celles engendrées dans les rations
contenant un antibiotique. Tiihonen et al. (2010) ont quantifié la bactérie de Clostridium
perfringens, la bactérie associée à l’entérite nécrotique et il n’y avait cependant aucune
différence significative entre le traitement témoin et celui avec des huiles essentielles. Ces
résultats concernant la quantification de C. perfringens de cette dernière étude sont différents
de ceux obtenus par Mitsch et al. (2004) qui ont constaté que des mélanges avec des huiles
essentielles peuvent contrôler la colonisation intestinale et la prolifération de Clostridium
perfringens, et peuvent ainsi aider à prévenir l'entérite nécrotique chez les poulets de chair.
Un autre effet très intéressant des huiles essentielles est qu’elles entraînent un changement
morphologique bénéfique de l’intestin chez les poulets. En effet, les oiseaux qui ont reçu un
supplément alimentaire à base d’huiles essentielles avaient des villosités plus longues que les
oiseaux témoins ce qui peut être due à des meilleures capacités d’absorption (Alaeldein et
Alyemni, 2013). L’huile essentielle de thym (thymol) peut être utilisée dans l’alimentation
des poulets de chair, particulièrement pendant la phase de croissance, car elle améliore la
morphologie intestinale et augmente l’activité enzymatique digestive de l’intestin (Xin et al,
2018). La supplémentation avec le thymol entraine une profondeur de crypte plus importante
que le traitement témoin dans l’iléon, une augmentation de la taille des villosités et de la
couche musculaire du duodénum et augmente le ratio longueur des villosités sur profondeur
des cryptes dans le jéjunum, ce qui améliore la digestibilité et l’efficacité d’absorption (Xin
et al, 2018).

51
1.5. L’approche de la méta-analyse
Les avancées technologiques des dernières années ont permis de rendre disponible plus de
données disponibles sur un sujet d’intérêt. Il est donc de plus en plus intéressant, voir
nécessaire de pouvoir extraire des données quantitatives à partir des publications de la
littérature (Sauvant al., 2007, 2020). L’ensemble des études réalisées sur les effets des
alternatives aux antibiotiques facteurs de croissances (AFC) sur les performances de
croissance et la santé digestive des poulets de chair forme un ensemble de données montrant
une grande variabilité dans l’amplitude des réponses. Bien que l’analyse traditionnelle par
une revue de littérature systématique puisse nous donner une réponse satisfaisante des
réponses globales, la quantification des effets des alternatives reste difficile avec ce type
d’analyse (Sauvant et al., 2008). C’est pourquoi, on fait recours à l’outil de l’analyse des
données par méta-analyse, qui permet de quantifier ces effets.
1.5.1. Définition
Le terme « méta-analyse » a été utilisé pour la première fois par le statisticien américain Gene
V Glass, en 1976 qu’il a défini comme l’analyse statistique d’un large ensemble de résultats
provenant d’études individuelles. En effet pour une problématique bien ciblée, elle aide à
évaluer la cohérence entre les différents résultats et obtenir des conclusions générales
(Hedges, 1984). On la définit comme « une analyse globale faisant la synthèse des résultats
de différentes études, au moyen de méthodes statistiques appropriées » (Beaucage et al.,
1998). La méta-analyse comporte un volet qualitatif comme par exemple l’évaluation de la
qualité d’une étude par l’absence de biais ou la force du plan d’étude et un volet quantitatif
qui consiste à intégrer les données numériques de plusieurs études pour augmenter la
puissance statistique (Beaucage et al. 1998). En effet la méta-analyse permet d’analyser et
comprendre des phénomènes en prenant en compte plusieurs études (Sauvant et al. 2008) et
de synthétiser les données de manière objective dans un cadre quantitatif. Elle apporte ainsi
une valeur ajoutée par rapport à des synthèses qualitatives sur un sujet donné. Au cours des

52
dernières décennies, la multiplication des recherches menées sur les alternatives alimentaires
en nutrition animale dans le contexte de retrait des AFC a produit un bassin considérable de
connaissances. Bien qu’elles revêtent un important potentiel, ces connaissances restent
souvent inutilisées ou sous‑utilisées en raison du manque de synthèses permettant de les
intégrer efficacement. En effet, tous ces essais n’ont pas été conduits sous des conditions
similaires, ce qui rend difficile l’obtention d’une conclusion générale à partir des résultats
des expérimentations individuelles (Faridi et al. 2015). La méta-analyse se positionne donc
comme un outil pertinent pour compiler les résultats de différentes études. Elles sont un outil
très utilisé en médecine où elles ont pris leur essor. En science animales et agronomiques, les
bonnes pratiques de méta-analyse comprennent plusieurs étapes (Figure 1-10) qui ont été
bien décrites par Sauvant et al., (2005, 2008), peaufinées et récemment revues et discutées
par Sauvant et al., 2020. Les données extraites des publications permettent généralement de
couvrir un ensemble très large de conditions expérimentales différentes, améliorant ainsi la
qualité de la prédiction et la précision des modèles (Sauvant et al. 2008).
1.5.2. Étapes d’une méta-analyse
Au moins deux conditions sont tout d’abord nécessaires pour réaliser une méta-analyse de
qualité: l’exhaustivité et la rigueur dans le choix des publications et des tests statistiques. En
premier lieu l’objectif et les hypothèses de la méta-analyse doivent être clairement définis.
Ensuite, les étapes doivent être suivies avec rigueur et nécessitent souvent plusieurs
itérations.

53
Figure 1-10 Principales étapes de la méta-analyse (tiré de Sauvant et al., 2005).
1.5.2.1. Détermination de l’objectif
La première étape d’une méta-analyse consiste à définir la question de recherche qui
détermine par la suite les objectifs plus ou moins ciblés. Dans une méta-analyse, la
problématique pourra évoluer à la suite des premiers résultats trouvés et en fonction des
données disponibles.
1.5.2.2. Collectes des données et conception de la base
La seconde étape de la méta-analyse consiste à rassembler d’une façon exhaustive toutes les
études publiées cohérentes avec les objectifs du travail (Sauvant et al., 2005). La recherche
bibliographique s’effectue à partir de bases de données informatisées au moins deux pour
être certain d’être exhaustif (ex : Web of Science, résumés CAB, Prod INRA et Science

54
Direct). La sélection des données se fait par la suite sur des critères d’inclusion et exclusion
très clairs. Dans le présent travail, la constitution de la base consiste à rassembler le maximum
de données disponibles sur l’utilisation des additives alimentaires à la place des antibiotiques.
Ensuite, les données sont extraites et transcrites avec précision dans la base de données après
codage. Ce codage consiste à dissocier des ensembles de données pour lesquels plusieurs
objectifs expérimentaux sont confondus (Sauvant et al., 2005). Dans un premier temps, les
publications reçoivent chacune un code unique. Ensuite, s’il y a différente expérience dans
une publication elles reçoivent chacune un code (Sauvant et al., 2005). Plusieurs autres codes
(ex. : sujet à l’étude, type d’aliment, stade physiologique) pourront être réalisés pour créer
des sous-groupes de traitements (Loncke, 2009).
1.5.2.3. Étude de la méta-dispositif
Les données compilées suite à la sélection sont ce qu’on appelle le méta-dispositif qu’il
convient d'explorer minutieusement à la fois graphiquement et statistiquement afin de savoir
ce qui est possible de faire ou non avec les données.
Étude graphique
C’est une étape importante pour bien comprendre les données. Elle consiste à étudier les
variables 2 à 2 graphiquement afin d’avoir une vue générale de l’hétérogénéité et de la
cohérence des données ainsi que de la nature des relations entre les variables. Ceci doit se
faire tant au intra, i.e. au sein d’une même étude et inter-études (Figure 1-11). Cette
visualisation graphique permet aussi de repérer les valeurs aberrantes ou influentes (Sauvant
et al., 2005).

55
Figure 1-11 Les corrélations inter et intra étude (tiré de Loncke, 2009).
Statistiques descriptives
Les variables quantitatives doivent être décrites à l’aide des statistiques descriptives :
moyenne, écart-type, minimum, maximum, nombre de données renseignées (Loncke, 2009).
En ce qui est des variables qualitatives tels le type d’alternatives, l’âge ou encore la race il
est important de regarder les effectifs de chacune d’entre elles, afin de bien s’assurer de
l’hétérogénéité des classes représentées et de déterminer s’il est possible ou non de les
étudier.
Corrélation entre les variables

56
Il existe deux types de corrélation : inter-étude et intra-étude (Figure 1-11). La première
corrélation inter-étude dite la corrélation classique ou corrélation de Pearson qui consiste à
tester la liaison de 2 variables (X et Y) sans tenir compte du fait que certains traitements sont
issus de la même étude. En autres termes, cette corrélation est générale pour toutes les
données. En revanche, la corrélation intra-étude est testée en incluant l’effet de la publication
en fixe tout en sachant que certains résultats proviennent de la même étude (Sauvant et al.,
2005).
1.5.2.4. Choix des modèles
Un critère primordial dans le choix du modèle statistique se base sur le mode de prise en
compte de l'effet étude qui peut être considéré comme : fixe ou aléatoire. Ceci fait
énormément débat dans toutes les sphères où sont utilisées les méta-analyses (Sauvant et al.,
2020).
Effet fixe de l’expérience
L’effet étude est considéré comme fixe si chaque étude peut être vue comme étant issue d’une
population particulière (Sauvant et al,. 2005). C’est le cas lorsque, par exemple, les méthodes
d’études (méthodes de mesures, d’analyses biochimiques) varient entre publications et que
l’on suppose que ces méthodes conditionnent les résultats obtenus. En effet le modèle fixe
est utilisé quand on analyse les données d’une expérience. Il permet de dégager les effets des
facteurs de variation sur les variables d’intérêt (Liais, 2006). Il permet de prendre en compte
les variations dont on ne maîtrise pas l’effet. Il permet ainsi de tenir compte des conditions
expérimentales différentes. On peut utiliser plusieurs expérimentations dans le même modèle
et ainsi déterminer l’évolution générale indépendamment de l’effet de l’expérimentation
(Liais, 2006).

57
Effet aléatoire de l’expérience
L’autre modèle est le modèle aléatoire ou (random effect model) (St-Pierre, 2001), où l’effet
étude est traité comme aléatoire. L’effet étude sera pris comme aléatoire si chaque étude peut
être considérée comme un échantillon pris au hasard dans une population plus large. Dans ce
modèle, on considère que chaque étude constitue un échantillon pris au hasard d’une
population unique (Sauvant et al., 2005). En fait, une grande diversité au niveau des
méthodologies existe entre les publications scientifiques, ce qui a une influence sur les
données : effet lié aux publications. Comme il existe plusieurs facteurs de variation entre
expérience (type d’alternative, niveau énergétique, dose d’alternative, sexe des animaux).
Dans notre analyse, l’effet étude a été considéré comme étant aléatoire. En fait une grande
diversité au niveau des méthodologies existe entre les publications scientifiques, ce qui influe
sur les données : effet lié aux publications. Le modèle statistique de base est donc le modèle
général de variance covariance sous la forme suivante (Sauvant et al,. 2005):
Yij = μ + μi + βXij + βiXij + eij
Avec :
- i l’étude i - j le numéro du traitement alimentaire
- Yij la variable expliquée
- Xij la variable explicative quantitative
- eij erreur résiduelle aléatoire
- μ l’ordonnée à l’origine générale
- μi effet de l’étude i sur l’ordonnée à l’origine du modèle
- β coefficient général fixe de régression
- βi effet fixe de l’étude i sur le coefficient général du modèle
Notons qu’il peut y avoir plusieurs variables X et des interactions entre elles et que des
relations autres que linéaires peuvent exister entre les variables X et Y. De plus, différents
types de modèles tels des modèles linéaires, non-linéaires, exponentiels, etc., peuvent aussi
être utilisés.

58
1.5.2.5. Identification des facteurs interférents
C’est une étape importante de vérification. Elle a pour objectif la vérification de la stabilité
du modèle par la détection de facteurs secondaires qui sont susceptibles de moduler la
réponse de la variable expliquée à un traitement. L'influence de ces facteurs sera testée sur
chacun des paramètres du modèle (pentes individuelles, moyennes des moindres carrés et
résidus; Sauvant et al., 2005) et l'introduction dans le modèle de ces facteurs de variations
additionnels sera testée à condition qu'ils ne soient pas corrélés à la variable X identifiée
(Loncke, 2009). Finalement, un point majeur après l’analyse statistique est que le critère de
prédiction retenu soit cohérent quantitativement avec les connaissances zootechniques.
1.5.2.6. Post-analyses
Après établissement du modèle statistique, des démarches post-analytiques doivent être
appliquées, pour évaluer la qualité du modèle et détecter les valeurs aberrantes ou influentes
(Sauvant et al., 2005). Cette dernière étape de la méta-analyse est nécessaire pour connaitre
les limites de l’analyse effectuée et pour savoir s’il est nécessaire d’effectuer d’autres
analyses complémentaires. Cela implique l’analyse de la normalité des résidus par le test de
Shapiro-Wilk testant l'hypothèse nulle de normalité des résidus. Elle peut être examinée
graphiquement soit en comparant les quantiles (QQ-plot) des résidus estimés à l’espérance
des mêmes quantiles sous hypothèse de normalité ou à l’aide de la droite de Henry. Le calcul
des résidus « studentisés » permet de repérer les données aberrantes (Figure 1-12). En effet,
les résidus trop grands qui sont exclus de l’intervalle [-2 ; 2] peuvent identifier une donnée
influente sur la variable dépendante Y. Pour éliminer une donnée aberrante, il faut bien relire
la publication, vérifier toutes les données et faire attention aux conséquences sur le jeu de
données et sur les paramètres du modèle, et il faut absolument posséder une explication
logique concernant la présence de telle valeur aberrante (Sauvant et al., 2005). C’est souvent
à cette étape de l’analyse que l’on va réaliser que certains points ne sont pas bien prédits par
le modèle et donc sortent aberrants nous permettant d’identifier de nouveaux X qui devraient
être inclus dans le modèle. D’autres statistiques tel le calcul des effets de levier et des

59
distances de Cook permettent de déterminer le poids des divers traitements dans la
détermination des paramètres du modèle statistique en détectant la présence des valeurs
aberrantes sur la variable explicative X. Finalement, les traitements qui sont trop influents
peuvent être retirés du modèle, mais il faut tester l’impact de ce retrait sur le modèle (Sauvant
et al., 2005).
Figure 1-12 Exemple de la représentation graphique de la dispersion des résidus autour de la
droite de Henry pour le paramètre de gain de poids quotidien (ADG).
1.5.2.7. Évaluation des modèles
Lorsque la méta-analyse est utilisée à des fins prédictives, une étape de validation est
recommandée pour s’assurer de la pertinence et d’un risque d’erreur minimum dans la
prédiction des valeurs observées. Cependant, le fait que la méta-analyse soit un processus
exhaustif complique cette étape compte tenu que toutes les données disponibles ont été
utilisées.
En théorie, une validation du modèle nécessite des données externes (Steyerberg et al., 2001).
Il est possible de faire une validation interne (cross validation; Garcia et Agabriel, 2008) Ce

60
type de validation consiste à segmenter l’échantillon de base en deux ou plusieurs
échantillons (p) selon la méthode choisie (tests et validation ou holdout method, k-fold, cross-
validation et leave-one-out cross-validation; Shaikh, 2018). Pour résumer, un des k
échantillons de base est utilisé pour l’établissement du modèle et les (p-1) autres échantillons
seront utilisés pour la validation de l’échantillon testé (Messad, 2016). L’opération se répète
ainsi p fois pour qu’en fin de compte chaque sous-échantillon ait été utilisé exactement une
fois comme ensemble de validation. La moyenne des erreurs est calculée pour estimer
l’erreur moyenne de prédiction du modèle (RMSPE (Root Mean Square Prediction Error)
que l’on cherche faible. Ensuite, la vérification de l’absence de biais (Vaillant, 2010) est
établie si la valeur de l’ordonnée à l’origine de cette analyse GLM était non différente de 0,
si la pente était non significativement différente de 1 (Loncke, 2009). Un R2 élevé est
également signe d’une bonne prédiction.
1.5.3. Les limites d’une méta-analyse
La méta-analyse comportent une série d’avantages. En combinant les résultats d’études
multiples, la méta‑analyse augmente la puissance statistique, c’est‑à‑dire la probabilité de
détecter les relations ou les différences significatives entre variables (St-Amand et Saint-
Jacques, 2013) et de supprimer l’absence d’effet dans certaines études étant donné le pouvoir
statistique élevé (Sauvant et al., 2008). Elle génère en quelque sorte de nouvelles
connaissances. La méta‑analyse permet de minimiser les erreurs et les biais des études
individuelles Les techniques méta‑analytiques réduisent par le fait même les erreurs de type
II qui consistent à conclure qu’il n’y a pas de relation/différence alors qu’il y en a une en
réalité (St-Amand et Saint-Jacques, 2013). Elle permet donc de porter un regard sur les
éléments les plus fortement associés à la réussite des interventions nutritionnels dans le cas
présent. Malgré les avantages déjà cités, cette méthode possède également des contraintes
qui limitent l’analyse des données. Les critères qui composent la méthodologie d’une méta-
analyse découlent en grande partie de l’évaluation que certains auteurs ont fait de méta-
analyses publiées dans diverses sphères d’activités scientifiques ou encore de consensus entre
professionnels, en vue d’améliorer la qualité des méta-analyses. Ces critères portent sur le

61
processus de recherche, le contrôle des biais, l’analyse statistique rigoureuse, l’analyse de
sensibilité et l’application des résultats à la population. Ces critères méthodologiques ne sont
pas toujours respectés dans les méta-analyses publiées, ce qui a ouvert la voie à la critique
envers ce type de recherche (Gauthier and Pharm, 2002). Parmi les critères faisant l’objet
d’une critique, notons :
-L’inclusion ou non de publications en langues étrangères (Gauthier and Pharm, 2002).
-Les données sont l’une des principales limites. En effet, dans de nombreux cas, les données
sont déséquilibrées et les effets de facteurs non orthogonaux (indépendants), ce qui pose des
problèmes d’estimation statistique tels que les points de levier, la quasi-colinéarité et la
déconnexion des facteurs qui interdit de tester les effets complètement confondus avec
d’autres (Sauvant et al., 2008).
-La qualité des études pourrait affecter la méta-analyse. En fait, Wolf (1986) (cité par St-
Amand et Saint-Jacques, 2013), rappelle que bien que des méta‑analyses aient démontré
qu’une recherche s’appuyant sur une solide méthodologie résulte en des grandeurs d’effet
plus faibles qu’une recherche de qualité moindre, d’autres ont démontré l’absence de relation
entre la qualité d’une étude et les grandeurs d’effet rappelle qu’il y a des études qui montrent
que plus la méthodologie. Pour Littell et ses collaborateurs (2008), une façon efficace de
résoudre ce problème consiste à formuler un critère d’inclusion permettant d’identifier quels
types de protocoles sont justifiés compte tenu des questions à l’étude et à examiner
attentivement la qualité des études incluses afin d’en tenir compte dans l’interprétation des
résultats.
-Les études utilisées dans les méta-analyses sont majoritairement publiées lesquelles sont
plus susceptibles de présenter des résultats significatifs que les études non publiées (Glass et
al., 1981) induisant un biais. Il est cependant impossible de de retracer les études non publiées
(Gauthier and Pharm, 2002).
-Le manque de variabilité intra étude des données imposées en raison des designs
expérimentaux constitue une autre contrainte de la méta-analyse (Sauvant et al., 2008),
limitant l’étude des interactions entre les variables dans la base de données.
En somme, l’immense mérite de la démarche méta-analytique est alors de proposer des règles
de synthèse rigoureuse qui peuvent être partagées par tous les chercheurs, dans le domaine
souvent bien subjectif de la revue de la littérature.

62
Chapitre 2 Assessing growth promoter effect and
modulating factors of dietary feed additive
commonly used in broilers: insights from meta-
analysis approach
Ce chapitre est présenté sous forme d’article et il est en révision dans la revue Poultry
Science.

63
Assessing growth promoter effect and modulating factors of dietary feed additive
commonly used in broilers: insights from meta-analysis approach
A. Rouissi., * F. Guay*, M. Boulianne †, and M. P. Létourneau-Montminy *
* Department of Animal Science, Laval University, Québec City, QC, Canada. † Department
of Clinical Science, Faculty of Veterinary Medicine, University of Montreal, Saint-
Hyacinthe, QC, Canada.
Corresponding author : Marie-Pierre Létourneau-Montminy
E-mail : [email protected]
Running title: Growth promoter effect in broiler
Abbreviations: ADG, average daily gain; ADFI, average daily feed intake; FCR, feed
conversion ratio; AME, apparent metabolizable energy; RMSEP, root mean square error of
prediction, RMSE, root-mean-square error; CI, confidence interval; BA. Butyric acid; MOS,
mannaoligosacharides; Bs, Bacillus subtilus; OEO, oregano essential oil.

64
2.1. Résumé
L’objectif de cette étude était d'évaluer l'impact des alternatives aux AFC (acide butyrique,
MOS, Bacillus subtilis et huile essentielle d'origan) sur les performances des poulets de chair
par méta-analyse de 79 articles scientifiques publiés de 2000 à 2017. Aucun effet sur la CJM
n'a été trouvé, alors que le GMQ augmentait de manière linéaire et quadratique (P <0,05)
avec la dose d'acide butyrique, de MOS et d'huile d’origan et linéairement avec Bacillus
subtilis de 7%, 8%, 7% et 9% respectivement. La teneur en EM avait un effet linéaire (P
<0,05) sur le taux l’IC avec l'acide butyrique et le MOS. L'amélioration de IC est de 2,4%,
8%, 7% et 3% pour l'acide butyrique, le MOS, l'huile d'origan et le Bacillus subtilis. En
quantifiant l'effet des alternatives aux AFC, les modèles trouvés proposeront une base
rationnelle pour leur utilisation dans l'alimentation des poulets de chair.

65
2.2. Abstract
The impact of non-antibiotic growth promoters on broiler chicken performance was
evaluated by meta-analysis of 79 scientific articles published from 2000 to 2017. Four
alternatives were examined: butyric acid, mannanoligosaccharide (MOS), Bacillus subtilis
and oregano essential oil. ADG, average daily feed intake and feed conversion ratio were
modeled as dependent variables. No effect on feed intake was found, whereas ADG increased
linearly and quadratically (P < 0.05) with dose of butyric acid, MOS and essential oil and
linearly with Bacillus by up to respectively 7%, 8%, 7% and 9%. Metabolizable energy
content had a linear effect (P < 0.05) on feed conversion ratio in conjunction with butyric
acid and MOS. Improvement in conversion ratio reached 2.4%, 8%, 7% and 3% respectively
for butyric acid, MOS, oregano essential oil and Bacillus subtilis. The relative mean square
error of prediction varied from 3.04% to 7.71% for ADG and from 0.05% to 0.09% for FCR.
R2 varied from 0.88 to 0.97 for ADG and varied from 0.84 to 0.97 for FCR. By quantifying
the effect of non-antibiotic growth promoters, the model proposes a rational basis for their
use in broiler diets.
Key words: broiler, antibiotic, alternatives, growth performance, meta-analysis

66
2.3. Introduction
Almost since their discovery began in the 1920s, antibiotics have played an important role in
the production of livestock performance in swine, cattle, and poultry (National Research
Council, 1980). Fed in sub-therapeutic doses, they protect animals against infections that
would otherwise reduce appetite, feed efficiency and growth rate (Castanon, 2007). They are
therefore regarded as growth promotors (AGP). However, since bacteria tend to develop
resistance to AGP (Tang et al., 2017) and serious consequences for human health have
resulted (Public Health Agency of Canada, 2017), such usage in animal feed was banned in
the European Union in 2006 (Regulation EC N° 1831/2003). In 2009, the USA Food and
Drug Administration (FDA) testified that the use of AGP should be eliminated (FDA, 2009).
Chicken Farmers of Canada banned the preventive use of category 1 (medical purpose)
antibiotics in 2014 and Canada banned AGP in January 2018 (Reference: CFC website
address detailing these bans).The overall impact of such bans on growth performance
(Jackson et al., 2004; Yang et al., 2012), and intestinal health (Ducatelle et al., 2018) is likely,
but has not been measured.
Finding alternatives to AGP to maintain gut health and growth performance in broiler
chickens is a priority worldwide (Timbermont et al., 2011; Abudabos et al., 2017). Many
substances have been proposed on the basis of in vitro studies of modes of action (Guillot,
2001; Mroz, 2005) and some have been tested in broilers (Lan et al., 2005; Jeong and Kim,
2014; Mookiah et al., 2014). They act in various ways and each alternative has its own
specific mechanisms, but usually by modifying intestinal pH, improving absorption (effects
on crypts and villus), stimulating the immune system and maintaining or improving the
intestinal microbiota. However, the effectiveness of these and other modulators of intestinal
health as growth promoters remains poorly defined and quantified. Several reviews have tried
to highlight the effects of alternatives to AGP, all will little success given the wide responses
variation (Biggs et al., 2007; Pirgozliev et al., 2008; Nosrati et al., 2017). A meta-analysis
providing an overview of the alternatives that have been evaluated in vivo and quantify their
impacts in terms of feed conversion ratio and average daily feed intake with negative controls

67
would be helpful at this time. The objective of this study was to identify the most selected
alternatives to AGP for broilers and to quantify their effects on growth performance relative
to an antibiotic-free control diet and to shed light on the effective modulating factors.
2.4. Materials and methods
Data collection and coding
Information was extracted from peer-reviewed papers published from 2000 to 2017 and
retrieved from various public databases Web of Science, CAB abstracts and Ariane. This
selection of databases contains articles of enough diversity to make sampling bias unlikely.
The keywords used in the search were: ‘chicks’ or ‘poultry’ or ‘broiler’ + ‘prebiotic’,
‘essential oil’, ‘probiotic’, ‘organic acid’ and ‘performance’, ‘alternatives’, ‘necrotic
enteritis’. Documents retrieved from the different searches were pooled and duplicates were
removed. Articles retained for the meta-analysis met the following criteria: broiler trial of
feed containing a non-antibiotic growth promoter, inclusion of an antibiotic-free control diet,
and reporting of feed conversion ratio, diet composition and broiler genetic line. The
systematic review procedure is illustrated in Figure 2-1. The most studied non-antibiotic
growth promoters were grouped into four sub-database categories, namely organic acids,
prebiotics, probiotics and essential oils. General information (e.g. author name, date of
publication, name of journal, objective of study), qualitative data (e.g. sex, genetics, infection
challenge, ration composition, ration presentation, use of vaccines or other medication),
quantitative data (e.g. dietary crude protein, ME, stocking density in kg/m2, group size (n),
number of experimental replicates or units to which a treatment was applied, number and
duration of feeding phases) were compiled in a spreadsheet. A code was assigned to each
article and each experiment reported (Sauvant et al., 2008).
Data investigation and statistical analysis

68
Feed composition was used to recalculate the dietary nutrient profile, especially apparent
metabolizable energy (AME), crude protein and digestible amino acids, from INRA tables
of feedstuffs (Sauvant et al., 2004). The AME requirement, standardized ileal digestibility of
lysine and crude protein values were then compiled based on bird genetic line (ROSS 308
Broiler Nutrition Specifications, 2014; COBB 500 Performances et Recommendations
Nutritionnelles, 2015) and the dietary concentration in each experiment was expressed in %
of the requirement.
Figure 2-1 Prisma flow diagram. Number of scientific papers (trials) identified, screened and
selected for meta-analysis.
The continuous independent variables tested were dose of non-antibiotic alternative and
dietary provision relative to requirement and the fixed independent variables were genetic
line, presence of challenge (pathogenic bacteria administered orally) and bird age (starter,

69
grower, finisher). Interactions were tested. The meta-design was scrutinized (Sauvant et al.,
2008) first by examining the graphical relationships between the independent variables taken
two by two to assess the degree of variation in each experiment and to identify outliers. The
within-experiment and between-experiment responses of the dependent variables Y (average
daily gain, average daily feed intake and feed conversion ration) to covariates X (% ME
provided relative to required, % crude protein provided relative to required, dose and code
of non-antibiotic alternative) were then examined. The study was considered random since
each study is conceptually a random outcome among a large population of studies from which
inference is to be drawn and also because the aim of the meta-analysis is hypothesis testing
of the overall impact of non-antibiotic alternatives on growth performance (St-Pierre, 2001;
Sauvant et al., 2008).
The final model for estimation of the main effect on growth performance was:
Yij = μ + μi + β1Xij + β2 [Xij] 2 + eij
Where Yij is the dependent variable Y in experiment i with non-antibiotic alternative type j,
Xij is the independent variable, μ is the overall intercept across all studies, μi is the effect of
experiment i on intercept μ with the condition that Σμi = 0, β1 and β2 are respectively the
linear and quadratic coefficients of the relationship and eij is the residual error. All statistical
analyses were carried out using the MIXED procedure of SAS version 9.4. All variables
found significant (P < 0.05) were retained and the interactions between these variables were
tested.
Model evaluation
The entire dataset was used to determine the model. Model performance was cross-validated
by subtracting the observed values from the predicted values (X – Y). The model was
considered unbiased when the intercept was not significantly different from 0 and the slope
was not significantly different from 1. Leave-one-out cross-validation was used to obtain the

70
root mean square error of prediction (RMSEP) from the value of the predicted residual sum
of squares statistic (Causeur, 2003). The root-mean-square error (RMSE) and the RMSEP
were used to test the accuracy of equations at predicting the effect of non-antibiotic growth
promoters on ADG, average daily feed intake and feed conversion ratio.
2.5. Results and discussion
Of the 130 broiler chicken studies retrieved, 79 met the inclusion criteria and provided the
data considered in the meta-analysis. These reported a total of 112 trials and described 282
experimental treatments. Four categories of non-antibiotic growth promoters were identified
(Table 2-1).

71
Table 2-1 Descriptive statistics of the databases1, 2.
N Mean SD Min Max Median
Parameters Butyric Acid
Crude protein
(g/kg diet) 50 20.35 1.004 18.5 21.85 20.33
Metabolizable
energy (Kcal
/kg DM)
50 3031.6 55.8 2910 3148 3045
Dose (%) 50 0.14 0.159 0.007 0.5 0.085
ADG (g/d) 50 52.7 20.34 20.07 88.45 52.64
ADFI (g/d) 44 97.33 46.82 26.21 170.14 101.01
Mortality (%) 26 5.66 2.814 1.3 10.5 4.8
FCR 44 1.754 0.327 1.198 2.5715 1.7662
Parameters MOS
Crude protein
(g/kg) 94 21.17 1.227 18.75 23.6 21.4
Metabolizable
energy (Kcal
/kg DM)
94 3064.2 101.2 2800 3224 3100
Dose (g/Kg) 88 0.883 1.559 0.25 7.5 0.3
ADG (g/d) 94 58.41 32.59 16.98 169.1 50.34
ADFI (g/d) 94 101.93 56.98 26.56 258.87 100.74
Mortality (%) 18 1.46 1.434 0 4.51 1.32
FCR 94 1.7163 0.233 1.132 2.233 1.66
Parameters Bacillus sp.
Crude protein
(g/kg) 60 21.48 0.945 20 23 21.5

72
1 ADG= Average daily gain; ADFI= Average daily feed intake; FCR= feed conversion ratio; 2 Publications= (Botsoglou et al., 2002; Alçiçek et al., 2003;
Krkpnar and Acikgoz., 2004; Herna, 2004; Alcicek et al., 2004; Ertas et al., 2005; Leeson et al., 2005; Teo and Tan., 2007; Midilli et al., 2008; Bozkurt et
al., 2009; Ashayerizadeh et al., 2009; Navidshad et al., 2010; Knap et al., 2010; Traesel et al., 2011; Kim et al., 2011; Tsinas et al., 2011; Aghazadah et al.,
2012; Yitbarek et al., 2012; Bozkurt et al., 2012; Mathlouthi et al., 2012; Sen et al., 2012; Abudabos et al., 2013; Jayaraman et al., 2013; Ghahri et al., 2013;
Pouraziz et al., 2013; Chamba et al., 2014; Shahir et al., 2014; Murarolli et al., 2014; Afrouziyeh et al., 2014; Saiyed et al., 2015; Murshed and Abudabos.,
2015; Park and Kim., 2015; Peng et al., 2016; Li et al., 2016; Ginnenas et al., 2016; Yang et al., 2016; Wang et al., 2016; Chand et al., 2016; Méndez Zamora
et al., 2017; Masouri et al., 2017; Alaeldein et al., 2017; Jameel, 2017; Al-Sagan and Abudabos. 2017; Nosrati et al., 2017).
Metabolizable
energy (Kcal
/kg DM)
60 3105 90 2960 3200 3100.2
Dose (log) 60 3.821 3.217 4 9.2 5
ADG (g/d) 60 47.25 21.56 14.3 99.80 46.67
ADFI (g/d) 46 79.89 43.1 18.5 164.85 77.38
Mortality (%) 21 13.02 14.94 1.7 48.4 5
FCR 51 1.628 0.251 1.03 2.313 1.606
Parameters Oregano Essential Oil
Crude protein
(g/kg) 78 21.7 1.278 18.99 24.33 20.75
Metabolizable
energy (Kcal
/kg DM)
78 3036.6 106 2868 3200 3097
Dose (mg/kg) 78 187.5 24.1 24 750 100
ADG (g/d) 78 56.3 19.44 24.87 100.7 58.8
ADFI (g/d) 73 100.47 41.03 38.24 187.8 96.8
Mortality (%) 10 2.72 0.373 1.1 2.33 6.04
FCR 78 1.765 0.23 1.0394 1.808 2.26

73
Butyric Acid (BA)
For 8 trials of broiler chicken feeds containing added BA, the mean average daily weight
gain was on average 48 g (95% CI 47.20, 48.80; Forest plot not shown) higher when
compared to the negative control group, while no significant difference from the negative
control was observed for 21 trials (95% CI 39.82, 40.78; Forest plot not shown). Regarding
FCR, on average 74 g less feed was needed in the treatment with BA group when compared
to the negative control to gain 1 kg of body weight (Forest plot not shown). In summary,
results for BA (Tables 2-2, 2-3) showed no effect on ADFI while ADG increased linearly
(P= 0.01) and quadratically (P= 0.06) leading to a linear decrease (P=0.003) of FCR with a
positive effect of ME (P= 0.002). The effects relative to negative control diets reached 7%
for ADG and 2.4% for FCR at the same dose. Percent ME relative to the requirement had an
effect on the intercept, indicating that a higher energy content improves FCR and ADG (P
≤ 0.05), but no impact of energy on the effect of butyric acid was found. Based on a review
of published findings, it has been concluded that BA improves ADG and FCR, on average
by 3.12% and 2.92% respectively (Kim et al., 2015). In the present review, BA was again the
most intensively studied organic acid. Its effects on broiler gut health and growth
performance appear to be multiple and beneficial (Awad et al., 2009; Khodambashi et al.,
2013). It is a selective bactericidal agent, decreasing gastrointestinal pH (Panda et al., 2009),
inhibiting pathogens such as Salmonella, Escherichia coli and Campylobacter jejuni (Van
Deun et al., 2008) and promoting the growth of lactic acid bacteria such as lactobacilli and
Bifidobacteria (Vogt et al., 1982). It passes into bacterial cells by diffusion (Clark and
Cronan, 2005), where it may be toxic (Warnecke and Gill, 2005) and kill by direct
mechanisms (Dibner and Buttin, 2002). It appears to improve the digestion of protein and
other nutrients requiring low gastric pH and high pepsin activity (Wang et al., 2011). In the
intestine, it is absorbed by epithelial cells as a direct source of energy (Pryde et al., 2002;
Mahdavi and Torki, 2009) and accelerates enterocyte growth, leading to villus elongation
and increased crypt depth (Chamba et al., 2014). These effects depend on the chemical form
of the acid (which have different pKa values, Ahsan et al., 2016) and depend on the
presentation of the molecule (e.g. encapsulation, Khan and Iqbal, 2016). Unfortunately, this
information was not available in all publications and could not be factored into the models.

74
Table 2-2 Prediction of the effect of alternatives on feed conversion ratio (FCR) in function of metabolizable energy (ME) and
doses.
Alternatives N Model equations Intercept Dose Dose*dose ME R2 RMSE
BA, g/kg 44 Y= 15.472 - 0.138× a - 0.144× b 0.0014 0.003 NS 0.002 0.97 0.05
MOS, g/kg 94 Y= 3.479- 0.0174 ×a - 0.0638 ×b + 0.007×
b2 0.0007 <.0001 0.0003 0.05 0.91 0.07
Bs, log 51 Y= 1.6508 - 0.0156 ×b <.0001 <.0001 NS NS 0.90 0.07
OEO, mg/kg 78 Y= 1,814- 0,000679×b + 0,000001*×b2 <.0001 <.0001 <.0001 NS 0.84 0.091
BA: Butyric acid; MOS: Mannanoligosaccharide; Bs: Bacillus; OEO: Oregano Essential oil.
N = number of studies.
Y = feed conversion ratio (FCR).
a = Metabolizable Energy (ME). b = dose of alternatives.
NS = effect not statistically significant.
RMSE = root-mean-square error

75
Mannanoligosaccharide (MOS)
In 21 trials with MOS, ADG was on average 94.3 g (95% CI 94.3, 94.32; Figure 2-2) higher
than the negative control group, while in 25 trials no significant difference with the negative
control was observed (95% CI 43.95, 44.55; Figure 2-2). Three studies reported a negative
effect of MOS on ADG (95% CI 85.92, 85.99; Figure 2-2). Regarding FCR, on average 158
g less feed was needed in the treatment with MOS group when compared to the negative
control to gain 1 kg of body weight (Figure 2-3). ADG increased as a linear and quadratic
function of MOS dose (P < 0.001) whereas FCR decreased (P < 0.001 for linear and P =
0.0003 for quadratic). The percent ME shifted the intercept (Tables 2-2 and 2-3), showing
that additional energy improved ADG (P = 0.02) and FCR (P = 0.05). Both improvements
reached 8% relative to the control group with 99% of the ME requirement (%supplied vs
required). These results confirm those obtained in s previous meta-analysis of 44 trials
conducted with a commercially available dietary MOS (Bio-MOS, Alltech Inc.) from 1993
to 2003 and showing ADG improved by +1.61% and FCR lowered by 1.99% relative to the
negative control (Hooge, 2004). Similar effects were noted in a holo-analysis of 82 Bio-
Mos® supplementation trials conducted from 1997 to 2003 (Rosen, 2007). Body weight was
increased by 1.48% and FCR was reduced by 2.11% in conjunction with butyric-acid-
supplemented diet. A review of 15 trials (2009–2011) of the MOS Actigen® concluded that
ADG was increased by 0.129 kg or 5.41% in the treatment group (Hooge, 2011) and the FCR
was reduced by 2.54%. The present study suggests an optimal concentration of MOS of 2.5
g per kg of broiler feed, which is close to previous estimates of about 2 g/kg (Rosen, 2007;
Yang et al., 2007).

76
Figure 2-2 This forest plot shows the results of the meta-analysis with MOS as fixed effect.
For the average daily weight gain (ADG, in g) the mean difference estimate (black square
symbol) with corresponding 95% confidence interval (95% CI; black bar) between the
treatment group with MOS and the negative control group is shown.

77
MOS are believed to improve broiler chicken growth primarily through three modes of action
(Teng and Kim, 2018). One is blocking attachment to and colonisation of the intestinal wall
by clostridia and Gram-negative pathogenic bacteria such as salmonella and E. coli (Spring
et al., 2000; Dimitroglou et al., 2009; Biggs et al., 2007). Another mode is modifying the
intestinal wall. Energy absorption is improved, villi are lengthened, and crypts are shallower
(Iji et al., 2001; Yang et al., 2008; Baurhoo et al., 2009). The epithelium is better protected
because of increased mucin production by goblet cells (Yang et al., 2008). The third mode
involves modulation of the immune response. At 2 g/kg in broiler feed, MOS appears to
stimulate expression of toll-like receptors TLR2 and TLR4 in ileal or cecal ganglions
(Yitbarek et al., 2012). In mammals, TLR4 recognizes bacterial lipopolysaccharide, whereas
responses to peptidoglycan and lipoprotein are mediated through TLR2. This is not yet clear
in poultry.

78
Table 2-3 Prediction of the effect of alternatives on average daily gain (ADG) in function of metabolizable energy (ME) and doses.
Alternatives N General regression equation (Model) Intercept Dose Dose×dose Energy R2 RMSE Dose max
BA, % 50 Y= -160.39 + 2.139 × a + 26.538 × b - 50.870 ×
b2 NS 0.01 0.01 0.05 0.97 3.52 0.261
MOS, g/kg 94 Y= 158 - 1,06 × a + 13,44 × b - 0,351 × b2 NS <.0001 <.0001 0.02 0.91 3.04 4.1
Bs, log 60 Y= 44.256 + 0.477 × b <.0001 <0.05 NS NS 0.88 7.64 †
OEO, mg/kg 78 Y= 54.54 + 0.024 × b - 0.00002 × b2 <.0001 0.0003 0.03 NS 0.95 4.48 300
BA: Butyric acid; MOS: Mannanoligosaccharide; Bs: Bacillus; OEO: Oregano Essential oil.N = number of studies.
Y = average daily gain (g/d). a = Metabolizable energy (ME). b = dose of alternatives.
NS = effect not statistically significant.
†= The response is linear.
RMSE = root-mean-square error.

79
Figure 2-3 This forest plot shows the results of the meta-analysis with MOS as fixed effect.
For the feed conversion ratio (FCR) the mean difference estimate (black square symbol) with
corresponding 95% confidence interval (95% CI; black bar) between the treatment group
with MOS and the negative control group is shown.

80
Oregano Essential Oil (OEO)
For 27 trials of broiler chicken feed supplemented with OEO, the mean ADG was on average
56.46 g (95% CI 56.22, 56.70; Forest plot not shown) higher compared to the negative control
group, while in 23 trials no significant difference with the negative control was observed
(95% CI 52.24, 53.28, Forest plot not shown). A negative effect on ADG with OEO dietary
inclusion was reported in two trials (95% CI 85.92, 85.99; Forest plot not shown). Regarding
FCR, on average 99.7 g less feed was needed in the treatment with OEO group when
compared to the negative control group. Results for OEO (Table2-2, 2-3) showed no effect
on ADFI while ADG was increased linearly (P<0.001) and quadratically (P=0.03).
Similarly, FCR also decreased linearly and quadratically (P<0.001) with increasing OEO
doses. When considering the quadratic effect, maximum effects for both ADG and FCR were
observed at 7% OEO inclusion rate. The optimal dose of OEO for broiler chicken production
was estimated at 300 mg/kg of diet in our study, which is close to the 500 mg/kg proposed
previously (Alagawany et al., 2018). OEO contains 78–85% bioactive compounds, the most
widely used being thymol and carvacrol (Basmacioglu et al., 2010). Based on the available
literature, its effect could be to increase crypt depth and villus height in the ileum (Gilani et
al., 2018). Increased villus height in the duodenum has also been reported (Skoufos et al.,
2016; Mohiti and Ghanaatparast, 2017; Fonseca-García et al., 2017). This effect was found
significant notably in a study involving challenge with Clostridium perfringens (Abudabos
et al., 2018). In vitro assays show that OEO is a strong inhibitor of E. coli (Roofchaee et al.,
2011), Salmonella spp., Listeria monocytogenes (Hulánková and Bořilová, 2012), Listeria
innocua and Staphylococcus aureus (Mathlouthi et al., 2012) and Mycobacterium avium
subsp. Paratuberculosis (Nowotarska et al., 2017). The positive effects of essential oils on
digestive enzyme secretion by the pancreas and intestinal mucosal have been reported in
many broiler studies (Jamroz et al., 2003, Jang et al., 2007; Basmacioğlu et al., 2010) and are
confirmed by increased digestibility of nutrients (Hernandez et al., 2004; García et al., 2007;
Amad et al., 2011).

81
Bacillus (Bs)
In 11 trials with Bs, ADG was on average 53.7 g (95% CI 53.2, 54.2; Forrest plot not shown)
higher when compared to the negative control group, while in 24 trials no significant
difference with the negative control group was observed (95% CI 56.2, 57.4; Forrest plot not
shown). A negative effect of Bs dietary inclusion was observed in 2 trials (95% CI 48.7, 50.6;
Forest plot not shown). Regarding FCR, on average 98.3 g less feed was needed in the Bs
treatment group when compared to the negative control group for a 1 kg gain in body weight.
Results for Bs (Table 2-2, 2-3) showed no effect on ADFI while ADG linearly increased
(P<0.05) and FCR linearly decreased (P<0.001) with increasing doses of this probiotic. At
doses from 106 to 108 cfu/g, feed conversion ratio fit a linear model with an effect reaching
3% whereas the effect on ADG reached 9%. Studies have demonstrated that strains of
Bacillus subtilis and Bacillus spp. may be effective as growth stimulators by competitive
exclusion of pathogenic bacteria (La Ragione and Woodward, 2003). Bacillus subtilis
modifies the intestinal microbiota and favors the growth of lactic acid bacteria, which have
recognized health-promoting properties (Knarreborg et al., 2008). The possible modes of
action of probiotics have been reviewed (Edens, 2003). The mechanisms proposed include
competition for nutrients and production of antibacterial compounds (organic acids,
hydrogen peroxide, and bacteriocins), interference with the ability of pathogens to colonise
and infect the mucosa and modulation of local and systemic immune responses (Fuller, 1989;
Rolfe, 2000; Frizzo et al., 2011). Amylolytic, proteolytic and lipolytic activities of probiotic
bacteria could accelerate digestion and thereby enhance growth performance (Rahman et al.,
2009). In the present meta-analysis, the duration of the experiment was not a significant
predictor of the effect of Bs on broiler growth, although it has been reported that the effect
of probiotics is strongest during the starter phase, when a stable gut microflora has not yet
been established and the birds are more vulnerable to environmental pathogens (Gaggìa et
al., 2010).

82
Prediction of growth performance
The cross validation (Table 2-4) showed that the intercept was not significantly different
from 0 and the slope was not significantly different from 1 for the four alternative’s database
indicating the absence of bias in the prediction. The current model presented also low
RMSPE with variation from 7.70% for Bs to 3.04% for MOS for ADG and from 0.093% for
OEO and 0.076% for Bs for FCR.

83
Table 2-4 Average daily gain and feed conversion ratio observed in experimental diets and predicted by the models.
BA: Butyric acid; MOS: Mannanoligosaccharide; Bs: Bacillus; OEO: Oregano essential oil. a = Mean deviation from the bisector expressed. b = the equation is: Yobs = a + bYpred. c = P-value that a and b differ from 0 and 1 respectively. d = R2, coefficient of determination. e = RMSEP, root mean square error of prediction.
BA, % MOS, g/kg Bs, log OEO, mg/kg
Parameters ADG FCR ADG FCR ADG FCR ADG FCR
Experimental
mean 53.7 1.76 59.1 1.72 47.5 1.62 56.7 1.77
Predicted
mean 53.7 1.76 59.4 1.72 47.6 1.64 56.6 1.76
da 0.0188 0.000260 -0.0490 -0.000127 -0.0490 0.00310 0.0514 0.000375
Regression equation b
a 0.273 0.003904 0.622 -0.00682 -0.617 0.0749 0.135 -0.0149
P-value c 0.853 0.934 0.929 0.9081 0.815 0.339 0.934 0.8704
b 0.995 0.998 0.716 1.00 1.01 0.956 0.999 1.01
P-value c 0.853 0.937 0.001 0.9083 0.814 0.349 0.957 0.866
R2d 0.97 0.97 0.91 0.91 0.88 0.9 0.95 0.84
RMSEP e 3.52168 0.055511 3.04197 0.0719 7.71 0.076 4.489249 0.093

84
2.6. Conclusion
Research on non-antibiotic alternative growth promoters for poultry production is
effervescent. Most of the articles published have dealt with growth performance. Between
2000 and 2017, 112 broiler chicken feed trials (282 experimental treatments described) with
negative (no antibiotic) controls were published. Meta-analysis shows clearly that non-
antibiotic growth promoters can improve performance indicators, including daily growth and
feed conversion ratio. Possible symbiotic combinations of this type of product remained to
be explored in depth.
Acknowledgement
The authors thank Agri-Marché Canada, Mitacs, Université Laval and the Tunisian
government national scholarship program for financial support.

85
2.7. References
Abudabos, A. M., Alyemni, A. H., Dafalla, Y. M., Khan, R.U., 2018. The effect of
phytogenics on growth traits, blood biochemical and intestinal histology in broiler
chickens exposed to Clostridium perfringens challengeJournal of Applied Animal
Research. 46, 691-695.
Abudabos, A. M., Alyemni, A. H., Dafalla, Y. M., Al-Owaimer, A. N., 2017. Effect of
organic acid blend and Bacillus subtilis on growth, blood metabolites and antioxidant
status in finishing broilers challenged with Clostridium perfringens. Journal of
Animal and Plant Sciences. 27, 1001-1007.
Afrouziyeh, M., Hanifian, S. H., Taghinejad, M., 2014. Effects of mannan oligosaccharides
on ileal digestibility of nutrients and microbial populations in the ceca of broiler
chickens. International Journal of Biosciences. 5, 373-380.
Aghazadeh, A. M., TahaYazdi, M., 2012. Effect of butyric acid supplementation and whole-
wheat inclusion on the performance and carcass traits of broilers. The South African
Journal of Animal Science. 42, 241-248.
Ahsan, U., Cengiz, Ö., Raza, I., Kuter, E., Chacher, M. F. A., Iqbal, Z., Çakir, S., 2016.
Sodium butyrate in chicken nutrition: the dynamics of performance, gut microbiota,
gut morphology, and immunity. Worlds Poultry Sciences. 72, 265-275.
Alagawany, M., Abd El-Hack, M. E., Farag, M. R., Shaheen, H. M., Abd El-Latif, M. A.,
Noreldin, A. E., Patra, A. K., 2018. The usefulness of oregano and its derivatives in
poultry nutrition. Worlds. Poultry Sciences. 74, 463- 474.
Al-Sagan, A. A., Abudabos, A. M., 2017. Effect of a prebiotic, probiotic and symbiotic on
performance of broilers under Clostridium perfringens challenge. The Thai Journal
of Veterinary Medicine. 47, 257-264.
Alcicek, A. H. M. E. T., Bozkurt, M., Çabuk, M., 2004. The effect of a mixture of herbal
essential oils, an organic acid or a probiotic on broiler performance. South African
Journal of Animal Science. 34, 217-222.
Amad, A.A., Männer, K., Wendler, K. R., Neumann, K., Zentek, J., 2011. Effects of a
phytogenic feed additive on growth performance and ileal nutrient digestibility in
broiler chickens. Poultry Sciences. 90, 2811–2816.
Ashayerizadeh, A., Dabiri, N., Ashayerizadeh, O., Mirzadeh, K.H., Roshanfekr, H.,
Mamooee, M., 2009. Effect of Dietary Antibiotic, Probiotic and Prebiotic as Growth
Promoters, on Growth Performance, Carcass Characteristics and Hematological
Indices of Broiler Chickens. Pakistan Journal of Biological Sciences. 12, 52-57.
Awad, W.A., Ghareeb, K., Abdel-Raheem, S., Böhm, J., 2009. Effects of dietary inclusion
of probiotic and symbiotic on growth performance, organ weights, and intestinal
histomorphology of broiler chickens. Poultry Sciences. 88, 49-55.
Basmacioglu, M. H., Baysal, S., Misiriloglu, Z., Polat, M., Yilmaz, H., Turan, M., 2010.
Effects of oregano essential oil with or without feed enzymes on growth performance,
digestive enzyme, nutrient digestibility, lipid metabolism and immune response of
broilers fed on wheat–soybean meal diets. British Poultry Science. 51, 67-80.
Baurhoo, B., Ferket, P. R., Zhao, X., 2009. Effects of diets containing different concentrations
of mannanoligosaccharide or antibiotics on growth performance, intestinal
development, cecal and litter microbial populations, and carcass parameters of
broilers. Poultry Sciences. 88, 62–72.

86
Biggs, P., Parsons, C. M., Fahey, G. C., 2007. Effects of several oligosaccharides on growth
performance, nutrient digestibilities, and caecal microbial populations in young
chicks. Poultry Sciences. 86, 2327-2336.
Botsoglou, N. A., Florou-Paneri, P., Christaki, E., Fletouris, D. J., Spais, A. B., 2002. Effect
of dietary oregano essential oil on performance of chickens and on iron-induced lipid
oxidation of breast, thigh and abdominal fat tissues. British Poultry Science. 43, 223-
230.
Bozkurt, M., Küçükyılmaz, K., Çatlı, A. U., Çınar, M., 2009. Effect of dietary mannan
oligosaccharide with or without oregano essential oil and hop extract supplementation
on the performance and slaughter characteristics of male broilers. South African
Journal of Animal Science. 39, 223-229.
Bozkurt, M., Küçükyılmaz, K., Çatlı, A. U., Çınar, M., Çabuk, M., Alçiçek, A. H. M. E. T.,
2012. Effects of administering an essential oil mixture and an organic acid blend
separately and combined to diets on broiler performance. Archiv fur Geflugelkunde.
76, 81-87.
Castanon, J.I.R., 2007. History of the use of antibiotic as growth promoters in European
poultry feeds. Poultry Sciences. 86, 2466–2471.
Causeur, D., G. Daumas, T. Dhorne, B. Engel, M. Fonti furnols, and S. Højsgaard. 2003.
Statistical Handbook for Assessing Pig Classification Methods: Recommendations
from the “EUPIGCLASS” Project Group, pp.132.
https://ec.europa.eu/info/sites/info/files/food-farming-
fisheries/animals_and_animal_products/documents/statistical-handbook-for-
assessing-pig-classification-methods_en.pdf. (Page consultée le 18 février 2018).
Chand, N., Faheem, H., Khan, R. U., Qureshi, M. S., Alhidary, I. A., Abudabos, A. M., 2016.
Anticoccidial effect of mannanoligosaccharide against experimentally induced
coccidiosis in broiler. Environmental Science and Pollution Research. 23, 14414-
14421.
Chamba, F., Puyalto, M., Ortiz, A., Torrealba, H., Mallo, J. J., Riboty, R., 2014. Effect of
partially protected sodium butyrate on performance, digestive organs, intestinal villi
and E. coli development in broilers chickens. International Journal of Poultry
Science.13, 390-396.
Clark, D.P., Cronan, J. E., 2005. Two-carbon compounds and fatty acids as carbon sources.
In: Neidhardt, F.C., Escherichia coli and Salmonella. Ecosal. Plus. 1(2).
COBB 500 Performances et Recommandations Nutritionnelles. 2015.
https://cobbstorage.blob.core.windows.net/guides/27811ef0-5d15-11e8-9602-
256ac3ce03b1. (Page consultée le 18 février 2017).
Dibner, J. J., Buttin, R. J., 2002 .Use of organic acids as a model to study the impact of Gut
microflora in nutrition and metabolism. Journal of Applied Poultry Research. 11, 453-
463.
Dimitroglou, A., Merrifield, D. L., Moate, R., Davies, S. J., Spring, P., Sweetman, J.,
Bradley, G., 2009. Dietary mannan oligosaccharide supplementation modulates
intestinal microbial ecology and improves gut morphology of rainbow
trout, Oncorhynchus mykiss (Walbaum). Animal Science Journal. 87, 3226–3234.
Ducatelle, R., Goossens, E., Meyer, F. D., Eeckhaut, V., Antonissen, G., Haesebrouck, F.,
Immerseel, V. F., 2018. Biomarkers for monitoring intestinal health in poultry:
present status and future perspectives. Veterinary Research. 49, 43.

87
Edens, F. W., 2003. An alternative for antibiotic use in poultry: Probiotics. Brazilian Journal
of Poultry Science. 5,75-97.
Ertas, O. N., Guler, T., Çiftçi, M., DalkIlIç, B., Simsek, U. G., 2005. The effect of an essential
oil mix derived from oregano, clove and anise on broiler performance. International
Journal of Poultry Science. 4, 879-884.
FDA, 2009. Food and Drug Administration. Summary report on antimicrobials sold or
distributed for use in food producing animals.
https://www.fda.gov/downloads/ForIndustry/UserFees/AnimalDrugUserFeeActAD
UFA/UCM231851.pdf. (Page consultée le 18 aout 2018).
Fonseca- Garcia, I., Escalera- Valente, F., Martinez-Gonzalez, S., Carmonagasca, C. A.,
Gutierrez-Arenas, D. A., Ávila-Ramos, F., 2017. Effect of oregano oil dietary
supplementation on production parameters, height of intestinal villi and the
antioxidant capacity in the breast of broiler. Australian Veterinary Journal. 49, 83-89.
Frizzo, L.S., Zbrun, M.V., Soto, L.P., Signorini, M. L., 2011. Effects of probiotics on growth
performance in young calves: a meta-analysis of randomized controlled trials. Animal
Feed Science and Technology. 169, 147-156.
Fuller, R., 1989. Probiotics in man and animalsJournal of Applied Microbiology. 66, 365-
378.
Gaggìa, F., Mattarelli, P., Biavati, B., 2010. Probiotics and prebiotics in animal feeding for
safe food production. International Journal of Food Microbiology. 141, 15–28.
García, V., Catalá-Gregori, P., Hernández, F., Megías, M. D., Madrid, J., 2007. Effect of
formic acid and plant extracts on growth, nutrient digestibility, intestine mucosa
morphology, and meat yield of broilers. British Poultry Science. 16, 555-562.
Ghahri, H., Toloei, T., Soleimani, B., 2013. Efficacy of antibiotic, probiotic, prebiotic and
symbiotic on growth performance, organ weights, intestinal histomorphology and
immune response in broiler chickens. Global Journal of Animal Scientific Research.
1, 25-41.
Gilani, S., Howarth, G., Nattrass, G., Kitessa, S., Barekatain, R., Forder, R., Tran, C. R.,
Hughes, R., 2018. Gene expression and morphological changes in the intestinal
mucosa associated with increased permeability induced by short-term fasting in
chickens. Animal Physiology and Animal Nutrition. 102, 653-661.
Guillot, J.F., 2001. Consequences of probiotics release in the intestine of animal. Pages 17-
21 in J. Brufeau (Ed.) Feed Manufacturing in the Mediterranean Region Improving
Safety: from feed to food. Zaragoza CIHEAM-IAMZ.
Hernandez, F., Madrid, J., Garcia, V., Orengo, J., Megias, M. D., 2004. Influence of two
plant extracts on broilers performance, digestibility, and digestive organ size. Poultry
Sciences. 83,169-174.
Hooge, D.M., 2011. Meta-Analysis Summary of Broiler Chicken Trials with Dietary
Actigen® (2009–2011). International Journal of Poultry Science. 10, 819-824.
Hooge, D.M., 2004. Meta-analysis of broiler chicken pen trials evaluating dietary
oligosaccharide, 1993–2003. International Journal of Poultry Science. 3, 163-174.
Iji, P.A., Saki, A. A., Tivey, D. R., 2001. Intestinal structure and function of broiler chickens
on diets supplemented with a mannanoligosaccharide. Journal of the Science of Food
and Agriculture. 81, 1186-1192.
Jackson, M.E., Geronian, K., Knox, A., McNab, J., McCartney, E., 2004. A dose-response
study with the feed enzyme β-mannanase in broilers provided with corn-soybean meal

88
based diets in the absence of antibiotic growth promoters. Poultry Sciences. 83, 1992-
1996.
Jameel, Y. J., 2017. Evaluation of probiotic, prebiotic and symbiotic on starter broilers
performance subjected to bursa vaccine and clostridium perfringens
challenge. International Journal of Advanced Biological and Biomedical Research. 7,
373-377.
Jamroz, D., Orda, I., Kamel, C., Wiliczkiewicz, A., Wertelecki, T., Skorupinska, J., 2003.
The influence of phytogenetic extracts on performance, nutrient digestibility, carcass
characteristics, and gut microbial status in broiler chickens. Journal of Animal and
Feed Sciences. 12, 583-596.
Jang, I.S., KO, Y. H., Kang, S. Y., Lee, C. Y., 2007. Effect of a commercial essential oil on
growth performance, digestive enzyme activity and intestinal microflora population
in broiler chickens. Animal Feed Science and Technology. 134, 304-315.
Jayaraman, S., Thangavel, G., Kurian, H., Mani, R., Mukkalil, R., Chirakkal, H., 2013.
Bacillus subtilis PB6 improves intestinal health of broiler chickens challenged with
Clostridium perfringens-induced necrotic enteritis. Poultry Sciences. 92, 370-374.
Jeong, J. S., Kim, I. H., 2014. Effect of Bacillus subtilis C-3102 spores as a probiotic feed
supplement on growth performance, noxious gas emission, and intestinal microflora
in broilers. Poultry Sciences. 93, 3097-3103.
Khan, S.H., Iqbal, J., 2016. Recent advances in the role of organic acids in poultry nutrition.
Journal of Applied Animal Research. 44, 359-369.
Khodambashi E. N., Zafari, N. S., Ruiz-Feria, C. A., 2013. Growth performance,
digestibility, immune response and intestinal morphology of male broilers fed
phosphorus deficient diets supplemented with microbial phytase and organic acids.
Livestock Science. 157, 506–513.
Kim, J.W., Kim, J. H., Kil, D. Y., 2015. Dietary organic acids for broiler chickens: a review.
Revista Colombiana de Ciencias Pecuarias. 28, 109-123.
Knap, I., B., Lund, A. B., Kehlet, C., Hofacre, G., Mathis, G., 2010. Bacillus licheniformis
prevents necrotic enteritis in broiler chickens. Avian Diseases. 54, 931-935.
Knarreborg, A., Brockmann, E., Høybye, K., Knap, I., Lund, B., Milora, N., Leser, T. D.,
2008. Bacillus subtilis (DSM17299) modulates the ileal microbial communities and
improves growth performance in broilers. International journal of Probiotics and
Prebiotics. 3, 83-88.
Lan Y., Verstegen, S., Tamminga, S., Williams, B. A., 2005. The role of the commensal gut
microbial community in broiler chickens. World's Poultry Science. 6, 95-104.
La Ragione, R. M., Woodward, M. J., 2003. Competitive exclusion by Bacillus subtilis
spores of Salmonella enterica serotype Enteritidis and Clostridium perfringens in
young chickens. Veterinary Microbiology. 94, 245-256.
Leeson, S., Namkung, H., Antongiovanni, M., Lee, E. H., 2005. Effect of butyric acid on the
performance and carcass yield of broiler chickens. Poultry Science. 84, 1418-1422.
Li, Y., Xu, Q., Huang, Z., Lv, L., Liu, X., Yin, C., Yan, H., Yuan, J., 2016. Effect of Bacillus
subtilis CGMCC 1.1086 on the growth performance and intestinal microbiota of
broilers. Journal of Applied Microbiology. 120, 195-204.
Masouri, L., Salari, S., Sari, M., Tabatabaei, S., Masouri, b., 2017. Effect of feed
supplementation with Satureja khuzistanica essential oil on performance and
physiological parameters of broilers fed on wheat-or maize-based diets. British
Poultry Science. 58, 425-434.

89
Mathlouthi, N., Bouzaienne, T., Oueslati, I., Recoquillay, F., Hamdi, M., Urdaci, M.,
Bergaoui, R., 2012. Use of rosemary, oregano, and a commercial blend of essential
oils in broiler chickens: in vitro antimicrobial activities and effects on growth
performance. Journal of Animal Science. 90, 813-823.
Mahdavi, R., Torki, M., 2009. Study on Usage Period of Dietary Protected Butyric Acid on
Performance, Carcass Characteristics, Serum Metabolite Levels and Humoral
Immune Response of Broiler Chickens. Journal of Animal and Veterinary Advances.
8, 1702-1709.
Méndez Zamora, G., Durán Meléndez, L.A., Hume, M. E., Vázquez, R. S., 2017.
Performance, blood parameters, and carcass yield of broiler chickens supplemented
with Mexican oregano oil. Revista Brasileira de Zootecnia. 46, 515-520.
Midilli, M., Alp, M., Kocabach, N., Muglah, O. H., Turan, N., Yilmaz, h., Cakir, S., 2008.
Effects of dietary probiotic and prebiotic supplementation on growth performance
and serum IgG concentration of broilers. South African Journal of Animal Science.
38, 21-27.
Mohiti- Asli, M., Ghanaatparast- Rashti, M., 2017. Comparing the effects of a combined
phytogenic feed additive with an individual essential oil of oregano on intestinal
morphology and microflora in broilers. Journal of Applied Animal Research. 46, 184-
189.
Mookiah, S., Sieo, C. C., Ramasamy, K., Abdullah, N., Ho, Y. W., 2014. Effects of dietary
prebiotics, probiotic and symbiotic on performance, caecal bacterial populations and
caecal fermentation concentrations of broiler chickens. Journal of the Science of Food
and Agriculture. 94, 341-348.
Mroz, Z., 2005. Organic acids as potential alternatives to antibiotic growth promoters for
pigs.
Advances in Pork Production. 16, 269-282.
Murshed, M. A., Abudabos, A. M., 2015. Effects of the dietary inclusion of a probiotic, a
prebiotic or their combinations on the growth performance of broiler
chickens. Brazilian Journal of Poultry Science. 17, 99-103.
Murarolli, V. D. A., Burbarelli, M. F. D. C., Polycarpo, G. V., Ribeiro, P. D. A. P., Moro, M.
E. G., Albuquerque, R. D., 2014. Prebiotic, probiotic and symbiotic as alternative to
Antibiotics on the Performance and Immune Response of Broiler Chickens. Brazilian
Journal of Poultry Science. 16, 279-284.
National Research Council, 1980. The Effects on Human Health of Subtherapeutic Use of
Antimicrobials in Animal Feeds. Washington, DC: The National Academies Press.
https://www.nap.edu/catalog/21/the-effects-on-human-health-of-subtherapeutic-use-
of-antimicrobials-in-animal-feeds. (Page consulté le 02 juin 2018).
Navidshad, B., Adibmoradi, M., Pirsaraei, Z. A., 2010. Effects of dietary supplementation of
Aspergillus originated prebiotic (Fermacto) on performance and small intestinal
morphology of broiler chickens fed diluted diets. Italian Journal of Animal Science.
9, 55-60.
Nosrati, M., Javandel, F., Camacho, L. M., Khusro, A., Cipriano, M., Seidavi, A., Salem, A.
Z. M., 2017. The effects of antibiotic, probiotic, organic acid, vitamin C, and
Echinacea purpurea extract on performance, carcass characteristics, blood chemistry,
microbiota, and immunity of broiler chickens. Journal of Applied Poultry Research.
26, 295-306.

90
Nowotarska, S.W., Nowotarski, K., Grant, I. R., Elliott, C. T., Friendmanf, M., SIitu, C.,
2017. Mechanisms of Antimicrobial Action of Cinnamon and Oregano Oils,
Cinnamaldehyde, Carvacrol, 2, 5-Dihydroxybenzaldehyde, and 2-Hydroxy-5-
Methoxybenzaldehyde against Mycobacterium avium subsp. Paratuberculosis (Map).
Foods. 6, 72-88.
Park, J. H., Kim, I. H., 2015. The effects of the supplementation of Bacillus subtilis RX 7
and B2A strains on the performance, blood profiles, intestinal Salmonella
concentration, noxious gas emission, and organ weight and breast meat quality of
broiler challenged with Salmonella typhimurium. Journal of Animal Physiology and
Animal Nutrition. 99, 326-334.
Panda, A.K., Rama, S.V.R., Raju, M. V. L. N., Sunder, G. S., 2009. Effect of Butyric Acid
on Performance, Gastrointestinal Tract Health and Carcass Characteristics in Broiler
Chickens. Asian-Australasian Journal of Animal Sciences. 22, 1026-1031.
Peng, Q. Y., Li, J. D., Li, Z., Duan, Z. Y., Wu, Y. P., 2016. Effects of dietary supplementation
with oregano essential oil on growth performance, carcass traits and jejunal
morphology in broiler chickens. Animal Feed Science and Technology. 214, 148-
153.
Pirgozliev, V., Murphy, C. T., Owens, B., George, J., McCann, M. E. E., 2008. Fumaric and
sorbic acid as additives in broiler feed. Research in Veterinary Science. 84, 387-394.
Pouraziz, S., Shahryar, H. A., Chekani-Azar, S., 2013. Effects of dietary saccharomyces
cerevisiae and butyric acid glycerides on performance and serum lipid level of broiler
chickens. Kafkas Universitesi Veteriner Fakultesi Dergisi. 19, 903-907.
Pryde, S. E., Duncan, S. H., Hold, G. L., Stewart, C. S., Flint, H. J., 2002. The microbiology
of butyrate formation in the human colon. FEMS Microbiology Letters. 217, 133–
139.
Public Health Agency of Canada, 2017. CANADIAN ANTIMICROBIAL RESISTANCE
SURVEILLANCE SYSTEM: Rapport 2017.
https://www.canada.ca/content/dam/phac-
aspc/documents/services/publications/drugs-health-products/canadian-
antimicrobial-resistance-surveillance-system-2017-report-executive-
summary/CARSS-Report-2017-En.pdf. (Page consultée le 12 juin 2018).
Rahman, M.M., Islam, M. R., Rahman, A., Rahman, M. S., 2009. Characterization of clinical
bacteria and their control by a medicinal plant (Cassia sophera L.) extracts. Journal
of Characterization and Development of Novel Materials. 1, 159-168.
ROSS 308 BROILER: Nutrition Specifications. 2014.
http://tmea.staging.aviagen.com/assets/Tech_Center/Ross_Broiler/Ross-308-
Broiler-Nutrition-Specs-2014r17-EN.pdf. (Page consultée le 08 fevrier 2017).
Roofchaee, A., Irani, M., Ebrahimzadeh, M. A., Akbari, M. R., 2011. Effect of dietary
oregano (Origanum vulgare L.) essential oil on growth performance, cecal microflora
and serum antioxidant activity of broiler chickens. African Journal of Biotechnology.
10, 6177-6183.
Rosen, G.D., 2007. Holo-analysis of the efficacy of Bio-Mos® in broiler nutrition. British
Poultry Science. 48, 21–26.
Rolfe, R. D., 2000. The role of probiotic cultures in the control of gastrointestinal
health. Journal of Nutrition 130, 396-402.
Saiyed, M. A., Joshi, R. S., Savaliya, F. P., Patel, A. B., Mishra, R. K., Bhagora, N. J., 2015.
Study on inclusion of probiotic, prebiotic and its combination in broiler diet and their

91
effect on carcass characteristics and economics of commercial broilers. Veterinary
World. 8, 225-231.
Sauvant, D., Perez, J. M., Tran, G., 2004. Tables of composition and nutritional value of feed
materials: Pig, poultry, sheep, goats, rabbits, horses, and fish. Wageningen Academic
Publishers; Wageningen, the Netherlands. INRA ed. Paris, France.
Sauvant, D., Schmidely, P., Daudin, J. J., St-Pierre, N. R., 2008. Meta-analyses of
experimental data in animal nutrition. Animal. 2, 203-1214.
SAS Version 9.4, 2014. Cary, NC: SAS Institue Inc.
Sen, S., Ingale, S. L., Kim, Y. W., Kim, J. S., Kim, K. H., Lohakare, J. D., Chae, B. J., 2012.
Effect of supplementation of Bacillus subtilis LS 1-2 to broiler diets on growth
performance, nutrient retention, caecal microbiology and small intestinal
morphology. Research in Veterinary Science. 93, 264-268.
Shahir, M. H., Afsarian, O., Ghasemi, S., Tellez, G., 2014. Effects of dietary inclusion of
probiotic or prebiotic on growth performance, organ weight, blood parameters and
antibody titers against influenza and Newcastle in broiler chickens. International
Journal of Poultry Science. 13, 70-75.
Skoufos, I., Giannenas, I., Tontis, D., Bartzanas, T., Kittas, C., Panagakis, P., Tzora, A.,
2016. Effects of oregano essential oil and attapulgite on growth performance,
intestinal microbiota and morphometry in broilers. The South African Journal of
Animal Science. 46, 77-88.
Spring, P., Wenk, C., Dawson, K. A., Newman, k. E., 2000. The effects of dietary
mannanoligosaccharide on cecal parameters and the concentrations of enteric bacteria
in the ceca of Salmonella-challenged broiler chicks. Poultry Sciences. 79, 205-211.
St-Pierre, N., 2001. Invited review: Integrating quantitative findings from multiple studies
using mixed model methodology1. Journal of Dairy Sciences. 84, 741-755.
Tang, K. L., Caffrey, N. P., Nóbrega, D. B., Cork, S. C., Ronksley, P. E., Barkema, H. W.,
Ghali, W. A., 2017. Restricting the use of antibiotics in food-producing animals and
its associations with antibiotic resistance in food-producing animals and human
beings: a systematic review and meta-analysis. The Lancet Planetary Health. 1, 316-
327.
Teng, P.Y., Kim, W. K., 2018. Review: Roles of Prebiotics in Intestinal Ecosystem of
Broilers. Frontiers in Veterinary Science. 5, 245.
Teo, A. Y., Tan, H. M., 2007. Evaluation of the performance and intestinal gut microflora of
broilers fed on corn-soy diets supplemented with Bacillus subtilis PB6 (CloSTAT).
Journal of Applied Poultry Research. 16, 296-303.
Timbermont, L., Haesebrouck, F., Ducatelle, R., Immerseel, V. F., 2011. Necrotic enteritis
in broilers: an updated review on the pathogenesis. Avian Pathology. 40, 341-347.
Traesel, C. K., Wolkmer, P., Schmidt, C., Silva, C. B., Paim, F. C., Rosa, A. P., Lopes, S. T.,
2011. Serum biochemical profile and performance of broiler chickens fed diets
containing essential oils and pepper. Comparative Clinical Pathology. 20, 453-460.
Tsinas, A., Giannenas, I., Voidarou, C., Tzora, A., Skoufos, J., 2011. Effects of an oregano
based dietary supplement on performance of broiler chickens experimentally infected
with Eimeria acervulina and Eimeria maxima. Poultry Science. 48, 194-200.
Van Deun, K., Haesebrouck, F., Immerseel, V. F., Ducatelle, R., Pasmans, F., 2008. Short-
chain fatty acids and l-lactate as feed additives to control Campylobacter jejuni
infections in broilers. Avian Pathology. 37, 379–383.

92
Vogt, H., Matthes, S., Harnisch, S., 1982. The effect of organic acids on productivity of
broilers. Archives Geflügelkunde. 46, 223-227.
Wang, X., Farnell, Y. Z., Peebles, E. D., Kiess, A. S., Wamsley, K. G. S., Zhai, W., 2016.
Effects of prebiotics, probiotics, and their combination on growth performance, small
intestine morphology, and resident Lactobacillus of male broilers. Poultry Science.
95,1332-1340.
Wang, Y., Manow, R., Finan, C., Wang, J., Garza, E., Zhou, S., 2011. Adaptive evolution of
no transgenic Escherichia coli KC01 for improved ethanol tolerance and homoethanol
fermentation from xylose. Journal of Industrial Microbiology and Biotechnology. 38,
1371-1377.
Warnecke, T., Gill, R. T., 2005. Organic acid toxicity, tolerance, and production in
Escherichia coli biorefining applications. Microbial Cell Factories. 4, 25.
Yang, G. Q., Yin, Y., Liu, H. Y., Liu., G. H., 2016. Effects of dietary oligosaccharide
supplementation on growth performance, concentrations of the major odor-causing
compounds in excreta, and the cecal microflora of broilers. Poultry Science. 95, 2342-
2351.
Yang, C. M., Cao, G. T., Ferket, P. R., Liu, T. T., Zhou, L., Zhang, L., Xiao, Y. P. Chen,
A. G., 2012. Effects of probiotic, Clostridium butyricum, on growth performance,
immune function, and cecal microflora in broiler chickens. Poultry Science. 91, 2121-
2131.
Yang, Y., Iji, P. A., Kocher, A., Mikkelsen, L. L., Choct, M., 2008. Effects of
mannanoligosaccharide and fructo-oligosaccharide on the response of broilers to
pathogenic Escherichia coli challenge. British Poultry Science. 49,550-559.
Yang, Y., Iji, P. A., Kocher, A., Mikkelsen, L. L., Choct, M., 2007. Effects of
mannanoligosaccharide on growth performance, the development of gut microflora
and gut function of broiler chickens raised on new litter. Journal of Applied Poultry
Research. 16, 280-288.
Yitbarek, A., Echeverry, H., Brady, J., Hernandez, J., Camelo-Jaimes, D. G., Sharif, S.,
Rodriguez-Lecompte, J. C., 2012. Innate immune response to yeast-derived
carbohydrates in broiler chickens fed organic diets and challenged with Clostridium
perfringens. Poultry Science. 91, 1105-1112.

93
Chapitre 3 Effects of Bacillus subtilis, butyric acid,
mannan-oligosaccharide and ß-glucans from naked
oat on growth performance, serum parameters and
gut health of broiler chickens
Ce chapitre est présenté sous forme d’article et sera soumis dans la revue Animal Feed
Sciences and Technology.

94
Effects of Bacillus subtilis, butyric acid, mannan-oligosaccharide and ß-glucans from
naked oat on growth performance, serum parameters and gut health of broiler
chickens
A. Rouissi., * F. Guay*, M. Boulianne †, and M. P. Létourneau-Montminy *
* Department of Animal Science, Laval University, Québec City, QC, Canada. † Department
of Clinical Science, Faculty of Veterinary Medicine, University of Montreal, Saint-
Hyacinthe, QC, Canada.
Corresponding author : Marie-Pierre Létourneau-Montminy
E-mail : [email protected]
Abbreviations: AGP, antibiotic growth promoters; ADFI, average daily feed Intake; ADG,
average daily Gain; BA, butyric acid; Bs, Bacillus Subtilis; BWG, body weight gain; Ca,
calcium; CD, crypt depth; CRP, C-reactive protein; DM, dry mater; FCR, feed conversion
ratio; FI, feed intake; FRAP, the ferric reducing antioxidant power; MDA, malondialdehyde;
MOS, mannan-oligosaccharide; Na, sodium; P, phosphor; GMQ, gain de poids quotidian;
SCFA, short-chain fatty acid; TP, total protein; VFA, Volatile fatty acid; VL, villus length;
VL/CD, villus length/ crypt depth.

95
3.1. Résumé
L’objectif était d'évaluer l’effet de Bacillus subtilis, de l'acide butyrique, de MOS et d’avoine
nue sur les performances de croissance et la santé digestive des poulets de chair. Le régime
d’avoine nue a amélioré le GMQ pendant la phase de démarrage, le MOS l’a amélioré durant
la phase de croissance et l'acide butyrique vers la fin. La meilleure amélioration de l’IC était
de 5% avec l’acide butyrique. Le régime à l'avoine nue a réduit le pH du caecum et de la
litière au jour 34. Le MOS a diminué la masse jéjunale au jour 34 et augmenté la longueur
des villosités (34%) et le rapport hauteur des villosités / profondeur de la crypte (32%) mesuré
au jour 10. Au jour 21, une augmentation significative (52%) de la protéine C-réactive avec
Bacillus subtilis. Les effets des alternatives aux AFC proposés sont mesurables mais leurs
modes d'action exacts restaient à définir.

96
3.2. Abstract
Four non-antibiotic alternative growth promoters for broiler chickens were evaluated. Ross
308 chicks were fed a control diet (mainly corn and soybean meal) or a diet supplemented
with a probiotic (Bacillus subtilis Gallipro DSM 17299), encapsulated butyric acid (Novyrate
C), mannan-oligosaccharide (Actigen MOS) or formulated with 20% naked oat (starter diet)
or 30% naked oat (grower and finisher). The study was carried out as a complete random
blocked design with 10 pens for each diet, 45 birds per pen. Compared to the control, the
naked oat diet improved the average daily gain by 16% during the starter phase (up to day
10). The probiotic did so during the grower phase as did butyric acid in the finisher phase
(up to day 34). For the experiment overall, the probiotic decreased ADG slightly. The best
improvement in feed conversion ratio was obtained in the butyrate group (5%). No significant
treatment effect on crop pH or on mortality was observed. The naked oat diet gave a slightly
lower cecum pH on day 34. The MOS supplement decreased jejunal mass on day 34 and
increased villus length (34%) and villus height/crypt depth ratio (32%) measured on day 10.
Naked oat, butyric acid and MOS diets all reduced serum endotoxin levels. The probiotic
increased serum C-reactive protein. All non-control diets reduced serum malondialdehyde.
The naked oat diet reduced day 34 litter pH by about 0.3. The effects of the proposed non-
antibiotic growth promoters are measurable and could contribute to livestock performance.
Their exact modes of action remained to be defined.
Keywords: Broilers, antibiotic, alternative to antibiotic growth promoters, growth
performance, gut health.

97
3.3. Introduction
For least 50 years, antibiotic growth promoters (AGP) have been commonly used to enhance
weight gain in poultry production (Engberg et al., 2000). This practice has participated to the
development of antibiotic resistance which has driven many countries to reduce their use,
and led to the withdrawal of antibiotic growth promoters in the European Union in January
1, 2006 (Castanon, 2007). In Canada, as of May 15, 2014, Canadian Chicken farmers are no
longer permitted to use preventatively Category I antibiotics, while the preventive use of
Category II antibiotics has been stopped in January 2019. The goal is to eliminate the
preventive use of Category III antibiotics by the end of 2020 (Chicken Farmers of Canada,
2020). Antibiotic resistance is defined as the ability of microorganisms to proliferate in the
presence of an antibiotic that generally inhibits or kills microorganisms of the same species
(Ruma, 2016). The resulting growth rate reduction with AGP removal impacts production
efficiency and may influence food safety and broiler health (Zhao et al., 2001). Consequently,
research efforts have been done to find alternatives to maintain feed efficiency and broiler
health in the absence of AGP (Diarra and Malouin, 2014). The efficacy of an alternative to
AGP is characterized by FCR maintenance, low mortality rate and a good gut health status
(Egani and Korver, 2008). Many alternatives to AGP have been proposed to improve animal
health, growth performance and immune response. A meta-analysis of 79 scientific articles
published from 2000 to 2017 (Rouissi et al., 2020) showed that the most frequently studied
alternatives to AGP in broilers were probiotics (76% Bacillus subtilis), prebiotics (82%
Mannan-oligosaccharides), organic acids (67% butyrate), and essential oils (52% oregano
based). Significant positive effects on average daily gain (7 to 9%) and FCR (2.4 to 8%) have
been observed for these AGP. Organic acid supplementation of broiler diets, particularly
butyric acid, has drawn attention because of its antimicrobial properties, effects on
gastrointestinal mucosa growth (Khan and Iqbal, 2016; Mehdi et al., 2018). Another category
of great interest are probiotics i.e., viable microorganisms, such as Bacillus, Bifidobacterium,
Enterococcus, Escherichia, Lactobacillus, Lactococcus, and Streptococcus, used as feed
additives. The modes of action of probiotics were extensively reviewed (Simon et al., 2001;
Ghadban, 2002; Edens, 2003) and consist in; 1) competitive exclusion including competition
for substrates, production of antimicrobial metabolites that inhibit pathogens, and

98
competition for attachment sites and, 2) immune modulation. Another category of alternative
to AGP are prebiotics, which are non-digestible food ingredients that beneficially affect the
host by selectively stimulating the growth and/or activity of intestinal bacteria and
fermentation (Choct, 2009; Huyghebaert et al., 2011). There is a plethora of commercially
available prebiotics such as fructo-oligosaccharides (FOS), oligofructose, inulin, mannan-
oligosaccharides (MOS), arabinoxylo oligosaccharides (AXOS), xylo-oligosaccharides
(XOS), and neoagaro-oligosaccharides (NAOS), (Femia et al., 2010; Patel and Goyal, 2012).
However, there are also natural prebiotics such as naked oats, which are too often
underlooked. For example, naked oats contain beta-glucans (β-glucans), non-starchy
polysaccharides with known prebiotic effects (Svihus and Gullord, 2002; Osek et al., 2003).
Based on the results of previous studies which were reviewed in a meta-analysis (Rouissi et
al., 2020), the objectives of this study were to evaluate the in vivo effects of butyric acid,
mannan-oligosaccharide, Bacillus subtilis and naked oat on growth performance, selected
serum parameters and gut morphology of broilers, as well as litter quality characteristics that
provide proxy evidence of bird wellbeing.

99
3.4. Materials and methods
The trial was conducted at the Deschambault Animal Science Research Center (120 Roy
street, Deschambault, QC G0A 1S0, Canada) from January 2019 to February 2019 and
approved by Animal Care Committee of Laval University (CPAUL) that is accredited by the
Canadian Council of Animal Care (CCAC) (Guidelines of the Canadian Council of Animal
Care, 2009).
Birds and housing
Day-old (male Ross 308) chicks (n=2.250) obtained from a local commercial hatchery were
weighed individually and assigned randomly to one of 50 floor pens (45 birds each). Ambient
temperature was maintained at 33°C during the first week and reduced gradually to 22°C by
the end of the third week and maintained at 22°C until the end of the trial. The lighting
program met Ross 308 guidelines, as did the starter phase diet (Ross 308 Broiler Nutrition
Specifications, 2014). During the grower and finisher phases, metabolizable energy and
standardized ileal digestible (SID) lysine were set at 96% of the requirements (Table 3-1) to
ensure that any growth promotion effect would be observable.
Experimental design
All birds were fed three basal diets: starter (0-10d), grower (11-21d) and finisher (22-34d)
diets. For each diet, seven treatments were manufactured as follows: 1) a Control diet (basal
diet; Table 1), 2) Control + butyric acid (an encapsulated butyric acid (SSFA, NUTRI-AD,
Belgium) with 30% of C4:0) incorporated at a rate of 1 kg/t in the starter phase, 0.50 kg/t in
the grower phase and 0.25 kg/ t in the finisher phase, 3) Control + Actigen prebiotic, (Alltech
Inc, St-Hyacinthe, Qc, Canada), a second generation MOS,) was added at a rate of 0.8 kg/t
in the starter phase, 0.4 kg/t in the grower phase and 0.2 kg/t in the finisher phase, 4) Control
+ probiotic, Gallipro®, (Chr. Hansen, Denmark, distributed by DCL, St-Hyacinthe Qc,

100
Canada) a Bacillus subtilis-based probiotic (strain DSM17299) was added at 0.125 kg/t for
all phases, and 5) naked oat as a natural ß-glucans source (SEMICAN inc. Plessisville, Qc,
Canada), was added at a rate of 20% in starter, and 30% in grower and finisher diets.
Data Collection
Litter moisture and pH
A 100g litter sample was collected from all 50 pens on day 9, 20 and 30 for moisture content
and pH measurement. Litter samples were oven-dried at 100°C for two days and percent
moisture content was calculated with the following formula: [100 - (dry weight-content
weight) / (wet weight -content weight)] * 100. For pH measurement (Rajkovich et al.2011),
1 g of ground litter sample (<1 mm) was added to 20 mL of deionized water. The suspension
was put on a shaker (Boekel Scientific, Orbitron II Laboratory Mixer, Model 260250,
Pennsylvania Blvd, USA) for an hour and then left to stand for one hour for the liquid and
solid phases to separate. The pH was measured in the liquid phase using a portable pH meter
(Ross, Orion Star A221, Thermo Scientific, Beverly, CA, USA).
Feed analysis
Feed was sampled during the batch bagging process and at the beginning of each dietary
phase. Dry matter, crude protein, calcium, phosphorus, sodium, fat and energy (Table 3-1)
were analyzed according to AOAC (2000) methods.
Performance data
Birds and feed were weighed (on days 0, 10, 21, 28 and 34) to evaluate average daily feed
intake (ADFI), average daily gain (ADG), and feed conversion ratio (FCR) for each growing

101
diets. Mortality was recorded daily. All dead birds and those euthanized for tissue sampling
(10, 21 and 34 d) were individually weighted and growth performance data were corrected
for mortality.
Intestinal morphology and histomorphometry parameters
At 10, 28, and 35 d of age, one bird per pen (10 per treatment) were euthanized by cervical
dislocation. The pH of the crop, distal ileum and cecum was measure using a portable pH
meter (Orion Star A221, Thermo Scientific™, Canada). The duodenum, jejunum, ileum, and
caeca were weighted. Once emptied of its contents, a section of the distal ileum was collected
for intestinal villi and crypts measurements. A first cross-section was made 1 cm from
Meckel's diverticulum and the second 1.5 cm distally. Each tissues were individually fixed
in a formalin solution and stored at room temperature (Marković et al., 2009). Multiple 5µm
sections of paraffined tissues were obtained from each sample with a microtome and stained
with hematoxylin and eosin (Scheuer et al., 1986). Villus and crypt measurements were based
on 18 villi and crypt per bird using a microscope (Microscope Central, Nikon Eclipse E600
Fluorescence, Feasterville, PA, USA) (objective lens: X4) and a PC-based image analysis
system and software analysis (Q-Capture, Fiji).
Cecal short-chain fatty acid profile in the cecum
One gram of cecum content was extracted upon euthanasia with an alcohol-cleaned spatula,
placed in a 15 mL conical tube, acidified with 2 ml of H2SO4 (1.5%), vortexed then frozen at
-20 ° C. The frozen sample was homogenized and centrifuged at 10,000 RPM for 15 min at
4oC. The supernatant was analysed on a Hewlett Packard 6890N gas chromatograph (Agilent
Technologies, Wilmington, DE, USA) equipped with a flame ionization detector and an
autosampler (Hewlett Packard, Avondale, PA, USA) for volatile fatty acid.

102
Fecal analysis
On day 34, 150 g of fresh droppings was collected, placed in a tube and stored at -20°C. DNA
was extracted from thawed samples using the Bioline Fecal DNA Extraction kit according to
the manufacturer's protocol (Clostridium perfringens was quantified in triplicate) by PCR
(Wu et al., 2011). Results for each sample were reported as number of copies per g of feces.
Biochemical analysis
Blood collected (in red cap tube, 6.0 mL with coagulation activator) from the brachial vein
prior to euthanasia (10, 28, and 34 day of age) at the end of each phase (n=10 birds/treatment).
Blood samples were centrifuged at 1,500 RPM at 4°C for 10 min and the serum was stored
at −20°C until analysis. Total protein was analyzed with commercially available diagnostic
kits (Thermo Scientific™ Pierce™ BCA Protein Assay Kit, 23227, USA). The serum
endotoxin levels was analyzed by a quantitative Chromogenic End-point Tachypleus
Amebocyte Lysate Endotoxin Detection Kit as per the manufacturer’s instructions
(" PyroGene™ Recombinant Factor C Endotoxin Detection Assay», LONZA, USA). C-
reactive protein (CRP) was analyzed using an assay kit, Chicken C-reactive protein Elisa kit
(Elabscience Biotechnology Inc. Canada). Uric Acid was measured with a commercially
available diagnostic kit (QuantiChrom TM Uric Acid Assay Kit, BioAssay Systems, USA).
Malondialdehyde (MDA) a lipid peroxidation marker that is used as an indicator of oxidative
stress, was determinated according to Ermis et al. (2005).
Statistical analyses
The study was carried out as a randomized complete block design. For repeated measures,
time and treatment effects, as well as their interactions, were tested with the MIXED
procedure (SAS 9.4, SAS, NC, USA). For the other measurements, feed additives were
compared to the control using the MIXED procedure and the Dunnett test (SAS 9.4, SAS,

103
NC, USA). A difference was considered significant at P <0.05 and P <0.10 was considered
to indicate a tendency.

104
Table 3-1 Broiler chicken basal diet composition (ingredient %).
Ingredient Diet
Starter Grower Finisher
Corn meal 55.018 39.4 56.66 31.82 61.4 36.56
Naked oat - 20 - 30 - 30
Soybean meal1 26.1 22.1 17.6 15 12.2 9.5
Durum wheat 0 0 10 10 10 10
Soybean meal2 10 10 9.3 7 10.5 8.3
Meat/bone meal 5 5 4.6 4.1 4.1 3.6
Calcium carbonate 0.66 0.69 0.49 0.61 0.55 0.67
Sodium chloride 0.22 0.21 0.21 0.19 0.22 0.19
Sodium bicarbonate 0.21 0.22 0.21 0.25 0.21 0.26
L-lysine 0.25 0.33 0.25 0.34 0.22 0.31
DL-methionine 0.37 0.34 0.29 0.25 0.24 0.2
Threonine 0.09 0.12 0.07 0.1 0.06 0.08
Micronutrient premix3 0.2 0.2 0.2 0.2 0.2 0.2
Others4 0.15 0.15 0.12 0.14 0.1 0.13
Calculated content
Dry matter 88.04 88.07 87.54 87.56 87.45 87.48
Fat 4.48 5.27 3.03 4.6 3.16 4.74

105
Metabolizable kcal/kg 3026.45 3026.78 3025.85 3025.31 3075.11 3075.11
Crude protein (N×6.25) 23.27 23.23 20.17 20.3 18.2 18.33
Fiber 2.79 2.86 2.74 2.81 8.71 2.78
Calcium 0.94 0.94 0.79 0.79 0.56 0.57
Phosphorus 0.69 0.69 0.61 0.62 0.56 0.57
Sodium 0.18 0.18 0.18 0.18 0.18 0.18
148% crude protein; 244% crude protein and 6% fat; 3vitamins and trace elements; 4phytase, choline, vitamin E, HyD.

106
3.5. Results
Growth performance
At the end of the starter phase, the ADFI and ADG were respectively 6% and 16% greater in
the naked oat diet group compared to the control diet and the FCR was 9% lower (P < 0.001).
No significant difference in growth performance was associated with any other diet at this
phase (Table 3-2). No diet had any significant effect on mortality. In the grower phase, the
ADG remained greater and the FCR lower in the naked oat group (P < 0.001) while the
Bacillus diet appeared to increase the FCR by 2.8% compared to the control diet (P = 0.002).
No significant effect on ADFI was observed between the treatments. Mortality tended to be
higher in the naked oat group than in the control group (P = 0.06). In the finisher phase, the
butyric acid supplement brought a 4% increase in ADG (P = 0.002) and a 5% decrease in
FCR (P = 0.005). The Bacillus supplement had the opposite effect, with an 8% decrease in
ADG (P < 0.001) and a 6% increase in FCR (P < 0.001). No diet had any effect on ADFI or
mortality. For the 34-day trial overall, ADG was lower and FCR was higher in the Bacillus
group (P < 0.001), the ADFI was increased by 2% in the naked oat group and no significant
effects were noted for the other diets (Table 3-2). The best (i.e. lowest) FCR was observed in
the butyrate group (P < 0.005) and no significant treatment effect on mortality was observed.

107
Table 3-2 Growth performance of broiler chickens fed with different alternatives to antibiotics growth promoters (AGP) at different
time points and P-values for treatment interactions.
Dietary enrichment P value1
Parameter Control butyrate MOS Bs oat SEM T2 T3 T4 T5
Starter (0–10d)
ADG 17.29 17.54 17.62 17.70 20.08 0.168 0.242 0.203 0.141 <0.001
ADFI 25.76 26.31 26.06 26.21 27.35 0.191 0.097 0.433 0.256 <0.001
FCR 1.495 1.496 1.501 1.466 1.369 0.012 0.758 0.639 0.114 <0.001
Mortality2 0.650 0.650 1.300 0.650 0.650 0.391 0.553 0.253 1.000 1.000
Grower (10–21d)
ADG 64.96 63.80 65.62 63.85 69.30 0.522 0.242 0.556 0.319 0.0003
ADFI 81.08 81.27 82.43 81.48 82.67 0.748 0.887 0.371 0.789 0.2973
FCR 1.239 1.268 1.248 1.274 1.194 0.006 0.009 0.429 0.002 <0.001
Mortality2 0 0.990 0.470 0.310 1.090 0.307 0.126 0.343 0.478 0.0631
Finisher (21–34d)
ADG 105.98 110.66 108.01 98.26 105.60 0.931 0.002 0.182 <0.001 0.7835
ADFI 165.13 164.60 164.65 163.8 168.25 1.181 0.784 0.784 0.4655 0.0811
FCR 1.5570 1.4890 1.5250 1.658 1.5930 0.015 0.004 0.165 <0.001 0.1147
Mortality2 0.5100 0.5400 0.6400 0.320 0.7600 0.271 0.990 0.796 0.6766 0.5960

108
Overall (0–34d)
ADG 65.84 67.26 66.77 62.88 67.73 0.550 0.080 0.306 0.001 0.041
ADFI 95.59 95.09 96.00 94.72 97.87 0.700 0.542 0.619 0.299 0.007
FCR 1.451 1.422 1.437 1.52 1.441 0.008 0.035 0.311 0.0001 0.475
FBW 2.3223 2.3173 2.3508 2.385 2.3329 0.026 0.878 0.385 0.0594 0.7459
Mortality 1.73 2.39 2.82 2.17 3.261 1.20 0.537 0.305 0.680 0.153
ADG: average daily gain (g); ADFI: Average daily feed intake (g/kg); FCR: feed conversion ratio; FBW: final body weight (kg); 1Control versus butyric
acid (T2), mannan-oligosaccharide (T3), Bacillus subtilis (T4), or naked oat (T5); 2Percentage of birds at this stage, not cumulative.

109
Short chain fatty acid content of the cecum
The total short chain fatty acid (SCFA) content of the cecum decreased in all groups from
starter to grower (72.8 to 55.1 mmol/l, P<0.001; Figure 3-1a) and then rose sharply during
the finisher phase (55.1 to 102 mmol/l, P<0.001). Acetate proportion was influenced by age
and dietary treatment (P=0.02) showing that birds receiving naked oat had the highest acetate
proportion at day 10 and then the lowest at day 21 and 34 (Figure 3-1b). A similar tendency
(day x treatment, P = 0.07) was observed for butyrate, but with opposite results, showing an
increased butyrate in the naked oat group (Figure 3-1c) while the other treatments resulted in
a decreased butyrate proportion from day 10 to day 21.

110
Figure 3-1 Concentrations of total SCFA (a), acetate (b), butyrate (c) and propionate (d) in
the caeca of growing broiler chickens fed alternative to antibiotic growth promoters (AGP);
Bs = Bacillus subtilis; absence of a letter in common indicates that the values.
Weight and pH of the intestinal segments
Crop pH was not influenced by dietary treatments but decreased with age (P<0.001).
However, ileum pH increased with age and tended to decrease in birds fed with butyrate
treatment (P=0.06; Figure 3-2 b) when compared to controls (6.9 at d 10 vs 7.2 at d 34). A
time per treatment interaction was observed for cecal pH (P=0.02) with the highest pH
measured at day ten and the lowest at day 34 in the naked oat diet group. The opposite was
observed for the Bacillus-supplemented diet group (Figure 3-2c). As expected, weight of

111
digestive tract section increase with age (P<0.001) without any treatment effect except for a
decrease in jejunal weight at day 34 in the MOS group (Table 3-3) (Interaction day x
treatment, P = 0.027).
Figure 3-2 The pH of the crop (a), ileum (b), cecum (c) and litter (d) of growing broiler
chickens fed with alternative to antibiotic growth promoters (AGP); Bs = Bacillus subtilis;
absence of a letter in common indicates that the values differ significantly.

112
Table 3-3 Broiler chicken intestinal segment development in response to alternatives to antibiotics growth promoters (AGP) at
different time points (day of age), per treatment group and interaction effects.
Items Duodenum Jejunum Ileum Cecum
Time (Day)
10 4.87 c 6.69 c 4.91 c 1.49 c
21 11.8 b 14.7 b 10.3 b 2.51 b
34 19.4 a 25.5 a 20.0 a 6.60 a
Diet
Control 12.3 15.3 11.0 2.61
Butyrate 11.6 15.3 11.6 3.05
MOS 11.7 15.2 11.1 2.87
Bs 11.9 15.6 12.0 3.14
Naked oat 11.9 16 12.3 3.56
Probabilities
SEM 0.3732 0.4695 0.4088 0.1706
Treatments 0.2685 0.1189 0.0669 0.1222

113
Time. Day < 0.001 < 0.001 < 0.001 < 0.001
Treatments x Time 0.0841 0.0267 0.169 0.8238

114
Ileum histology
No treatment effect on ileum crypt depth was apparent on day 10 (Table 3-4). However, villus
length and villus height/crypt depth ratio were higher in birds receiving mannan-
oligosaccharides compared to the control birds (+34%; P<0.001 and +32%; P = 0.05)). At
day 21, villus length was also higher in this group (15%; P =0.03) but there were no
significant differences between the Control and the other treatment groups for crypt depth
and villus height/crypt depth ratio, for a tendency to a higher villus height/crypt depth ratio
(14%, P=0.08) in naked oat in comparison to Control birds.

115
Table 3-4 Ileal histomorphological parameters of broiler chickens fed with different alternatives to antibiotics growth promoters
and P-value for treatment interactions after the starter and the grower phases.
Parameter*
Dietary treatment P value1
Control Butyrate MOS Bs Oat SEM T2 T3 T4 T5
starter (0–10d)
Villus
length (μm) 165.3 188.2 221.2 180.9 193.6 13.975 0.15 0.0008 0.32 0.21
Crypt
depth (μm) 39.44 41.1 39.29 38.02 40.43 2.7535 0.63 0.96 0.68 0.77
length/dept
h 4.82 4.46 6.35 4.97 5.51 0.6146 0.56 0.05 0.1 0.23
Grower (10–21d)
Villus
length (μm) 287.9 298.9 330 318.8 310.7 15.093 0.56 0.03 0.1 0.23
Crypt
depth (μm) 27.5 27.82 27.65 27.75 24.35 1.8402 0.89 0.94 0.91 0.2
length/dept
h 11.22 11.46 12.67 12.58 12.75 0.8775 0.83 0.22 0.26 0.08
1Control versus enrichment with butyric acid (T2), mannan-oligosaccharide (T3), Bacillus subtilis (T4) or naked oat (T5); *: Villus lengths and depths in
μm.

116
Serum biochemistry
On day 10, no treatment effects on plasma C-reactive protein or malondialdehyde were
apparent, whereas the butyrate diet appeared to lower the endotoxin concentration (P = 0.02)
as did mannan-oligosaccharide (P = 0.05) and naked oat (P = 0.03) in comparison with the
control diet (Table 3-5). On day 21, a significant increase (52%, P = 0.036) in C-reactive
protein appeared in the Bacillus treatment group in comparison to the control group, and
malondialdehyde was lowered in all non-control groups (P < 0.05).

117
Table 3-5 Serum parameters of broiler chickens fed different alternatives to antibiotics growth promoters and P-values for
treatment interactions.
Parameter Dietary enrichment P value1
Contro
l Butyrate MOS BS Oat SEM T2 T3 T4 T5
starter (0–10d)
CRP 0.9341 1.0191 0.6646 0.8709 0.6741 0.1946 0.718 0.2547 0.7881 0.2574
MDA 7.0578 8.5677 9.3812 9.1394 9.5943 1.2401 0.302 0.1146 0.1566 0.0857
Endotoxin 4.5173 3.0669 3.4797 4.0346 3.2335 0.4122 0.016 0.0507 0.4113 0.0319
Grower (10–21d)
CRP 1.3809 1.8691 1.6061 2.0973 1.5876 0.2707 0.1473 0.4874 0.0355 0.5235
MDA 9.1568 5.7729 6.466 6.4349 6.9372 0.8789 0.0058 0.0263 0.0247 0.0857
Endotoxin 5.6090 7.3026 6.5695 5.6578 6.3579 0.6361 0.0500 0.2740 0.9554 0.3926
Finisher (22-34d)*
CRP 0.2002 0.2056 0.203
7 0.2854 0.1575 0.0463 0.159 0.6630 0.0008 0.4672
1Control versus enrichment with butyric acid (T2), mannan-oligosaccharide (T3), Bacillus subtilis (T4) or naked oat (T5); CRP = C-reactive protein (ng/mL);
MDA = malondialdehyde (µM); Endotoxin = EU per mL; *: we did not do the analysis for MDA and endotoxin at the finisher phase for lack of samples.

118
Litter Properties
As expected, litter moisture increased with age (P<0.001; Figure 3-3) and no treatment effect
was observed. Litter pH also increased with age (P<0.001) but at a different rate with the
naked oat treatment group the kinetic change depending of the dietary treatment with naked
oat that reduced pH at day 30 in comparison to Control (6.52 vs 6.82; Interaction Day x
Treatments, P = 0.011; Figure 3-2d).
Figure 3-3 Litter moisture (%) variation overtime of broilers fed with alternatives to
antibiotics growth promoters; Bs = Bacillus subtilis.

119
3.6. Discussion
There is a worldwide interest in finding alternatives to antibiotics growth promoters which
can both optimize growth performance and poultry health. The main objective of this study
was to validate the results of our meta-analysis we conducted (Rouissi et al., 2020) of 79
previous studies published from 2000 to 2017, on the impact of alternatives to AGP on
growth performance. The second objective was more specifically to measure directly the
effects of alternatives to AGP on growth and intestinal morphology and to evaluate easily
performed analyses of serum and pen litter as proxies of bird health. Results obtained from
this meta-analysis (Rouissi et al., 2020), showed no effect on feed intake, whereas average
daily gain increased linearly and quadratically (P<0.05) with incremental doses of butyric
acid and mannan-oligosaccharide, and linearly with Bacillus subtilis by up to respectively
7%, 8% and 9% at recommended doses varying from 106 to 108 cfu/g. Improvement in feed
conversion ratio reached 2.4%, 8% and 3% respectively for butyric acid, mannan-
oligosaccharide, and Bacillus subtilis. In the current study, energy and lysine supply were
reduced in the grower and finisher phases based on a strategy used in many publications of
the meta-analysis. Field experiments have shown that reducing energy and lysine contents to
96% of their requirements results in a measurable decrease in growth performance.
In the current study, butyrate treatment tended to increase ADFI during the starter phase,
followed by a negative effect on FCR during the grower phase, and then a significant
improvement of ADG and FCR during the finisher phase. Some studies showed that FCR
was increased in butyrate supplemented diets during the grower phase, (Moquet, 2018;
Gonzalez-Ortiz et al., 2019) and growth performance was improved in others (Abd El-Hakim
et al., 2009; Raza et al., 2017). In the present study, the higher growth performances in the
finisher phase could not be associated with any modification of the intestinal health
parameters except for ileal pH at day 21. Because fat-coated butyrate is slowly released
(Chamba et al., 2014; Sikandar et al., 2017), this could explain why the butyrate used in this
study was more effective in reducing ileal pH. Probiotics influence health through
modulation of immunological factors, intestinal permeability and bacterial translocation, and

120
secretion of various metabolites (de Vrese et al., 2008). Surprisingly, a negative effect of
Bacillus subtilis diet on ADG and FCR was observed during the grower (11-21d) and the
finisher (22-34d) phases. Previous publications have shown a positive effect of Bacillus
subtilis supplementation on growth performance (Park et al., 2018, Gadde et al., 2017; Gong
et al., 2018). Lee et al. (2015) have shown that Bacillus subtilis DFM fed from hatch
enhanced innate immunity and improved protection against avian coccidiosis. In the present
study, concomitantly with the reduced growth performance, the serum concentration of C-
reactive protein (CRP) was increased at day 21 and at day 34 (Table 5) in birds receiving
Bacillus subtilis in comparison to the Control group. Because CRP is an acute-phase protein
in inflammatory responses, and it is stimulated by the presence of both interleucine-1 (IL-1)
and interleucine-6 (IL-6), such increase might indicate the development of a local
inflammatory process (Eckersall and Bell, 2010). An increase in the concentration of CRP is
associated with cell damage caused by pathogens, that within 36-48 h post-exposure to such
stimulus (Jarosz et al. 2019). The negative effect of Bacillus subtilis coincides with the
reduction in AMEn and digestible Lys and other amino acids supplied in ratio with Lys in
grower and finisher diet. From a nutritional viewpoint, substrates such as amino acids and
energy are needed to support immune system, with amino acids and energy deficiency being
one of the major causes of immunodeficiency globally (Ruth et al., 2013; Field et al., 2002).
Moreover, both innate and adaptive immune systems are highly dependent upon an adequate
availability of amino acids (Kim et al., 2007). One of the hypotheses for the lower
performance in birds receiving the probiotic could thus be that the energy and / or protein
deficiency would have induced a global inflammatory state or targeted in the intestine or
both. This could lead to a probiotic-induced dysbiosis decreasing absorption due to alteration
of the gut microflora and intestinal mucosal integrity (Suez et al., 2018) or to a use of energy
for the increased immune response (Lei et al., 2015) both leading to a decrease in growth
performance.
The positive effect of the naked oat diet could be due to the reduced soybean meal and hence
antinutritional factor content. The higher pH and higher proportion of acetate in the cecum
on day 10 but no longer apparent on day 21 suggest that less substrate and probably different

121
substrates passed from the ileum to the cecum during these early growth phases in birds fed
the naked oat diet (Svihus et al., 2002) and that digesta arrived in the cecum with relatively
little unabsorbed nutrient. In fact, although the naked oat starter diet was formulated to meet
all requirements, absorption of energy or other nutrients may have been underestimated,
leading to higher gain and lower FCR compared to the corn-based diet. It was supposed to
provide 17.5 MJ/kg of crude energy and contain about half as much neutral detergent fiber
as the corn diet and mostly (i.e. 62%) the soluble NSP type (Sauvant et al., 2004; Bach
Knudsen, 2014). The apparent metabolizable energy or AMEn value (zero-nitrogen
corrected) of naked oat is higher than that of wheat, due presumably to its high oil content
and absence of husks (MacLeod et al., 2008). However, since none of these possible
advantages were apparent at the end of the finishing phase, perhaps only young broilers
benefit from them. Although corn and wheat AMEn values for chickens appear to increase
linearly with bird age (Yang et al., 2020), this might not be the case for naked oat. Whereas
the mass of the small intestine more than doubled during the grower phase and almost
doubled during the finisher phase, nearly quadrupling overall, bird weight increased by a
factor of six over these two periods. The absorption capacity relative to body weight thus
may have decreased with age. Meanwhile, the cecum grew faster during the finisher phase,
by more than four-fold over the two phases. Although the cecum was 60% smaller on day 10
than on day 21, total SCFA inside it was 1.3 times higher, suggesting that it received more
unabsorbed nutrients from the ileum on day 10. These results are consistent with previous
observations (Van den Borne et al., 2015). The intestinal mucosa of broiler chickens
undergoes major structural development during the first 7 days post hatch, especially in terms
of villus height or relative length in the jejunum (Iji et al., 2001), which is known to be the
principal site of nutrient absorption (Collins et al., 2019). The disappearance of the positive
effects of the naked oat diet by the end of the finisher phase is reflected in the lower pH
measured in the cecum and the litter. Naked oat fiber is highly fermentable and yields larger
amounts of butyric acid that do other types of dietary fiber (Casterline et al., 1997). The
proportion of butyrate increased from 10 to 21 days of age and stay higher in comparison to
other diets. Proportion of SCFA also changed with age, with reduction in acetate and
propionate and increase of butyrate showing a switch over time to more butyrogenic
microflora in the caeca as previously observed (Gonzalez-Ortiz et al., 2017). It is worthy to

122
note that SCFA production is influenced by many factors (e.g. diet, feeding phase duration,
pen or cages) other than just age. Oat β-glucan might thus behave as a prebiotic: a no
digestible feed ingredient that affects the host by selectively stimulating the growth and/or
activity of one strain or a limited number of bacterial strains in the colon and thus improves
host health (Gibson and Roberfroid 1995). The modification of cecal and colon fermentation
can lead to change of excreta and then litter characteristics. Thus, the reduction of litter pH
with naked oat diet at day 30 (6.19 vs 6.23) is probably the result of higher fermentability of
oat β-glucan (Daou and Zhang, 2012). Low litter pH is a good indicator of bird health
condition. Hardin and Roney, (1989) showed that the litter pH reduction resulted in a decline
in microbial populations, including E. coli, Salmonella, and Clostridium, to below detectable
limits. Additionally, Payne et al. (2007) showed that achieving a low pH led to the fastest
reduction in Salmonella populations when compared with higher pH. The current results
showed another positive effect of naked oat on broiler health. In addition to modifying the
cecal SCFA content and the pH, naked oat tended to increase villus height/crypt depth ratio
(+14% vs control) during the grower phase that may be attributed again to its high level of
β-glucan that has been showed to increase this ratio (Teng and Kim, 2018). This higher ratio
could improve the nutrient absorption capacity of birds in this group and then growth
performances. These results indicate that naked oat can be incorporated in broiler diets at 20
to 30% without negatives effects. Previous studies have shown that naked oat can substitute
other cereals in broiler diets (Kamińska., 2003; Osek et al., 2003, Szymczyk et al., 2005).
The MOS treatment had no effect on growth performances during different phases of
experiment. These results are opposite to those obtained in several studies, including those
reported in the meta-analysis of Roussi et al. (2020). Although MOS had no effect on
performance, an increase of the villus height was observed during the starter and grower
phase (0-21 d) and villus height/crypt depth ratio during the starter phase (0-10d). These
results are consistent with previous studies showing that mannan-oligosaccharides increase
villus height and surface area, decrease crypt depth, induce numbers of sulphated-acidic
goblet cells, and upregulate gene expression of MUC, which is related to mucin secretion
(Chee et al., 2010; Baurhoo et al., 2009). Longer villi correspond to a larger intestinal surface

123
area and higher digestive enzymes activities, therefore, increased nutrient absorption and
improved digestibility (Gao et al., 2008). Additionally, it was proposed that a decreased crypt
depth indicates a lower epithelial cell turnover, therefore better efficient energy use by the
host for growth and enhanced productivity (Murugesan et al., 2015). However, this change
in the morphology of the mucosa did not improve the growth of the birds, suggesting that the
height of the villi was not a factor limiting absorption in this study. In fact, the absorption of
nutrients depends on the villi but also on the length of the intestine and the presence of
specific transporters.
3.7. Conclusions
As regulations and the rise of consumer demand for poultry “Raised without antibiotics”
increase pressure to abandon the use of antibiotics as growth promoters (AGP), the search
for alternative approaches to stimulating the growth of broiler chickens intensifies. Much of
the published body of research has focused on the development of feed additives that
maintain or improve poultry health as well as performance. This current study focuses on the
effects of common alternatives on broiler chicken growth performance and gut health. The
alternatives examined herein were limited to a probiotic, prebiotics and butyric acid.
Challenges remain to understanding the mechanisms underlying their efficacy, advantages
and drawbacks. Their modes of action need to be defined more precisely. Optimal
combinations of various alternatives may be the key to maximizing performance and
maintaining productivity.
Acknowledgements
The authors thank Agri-Marché Canada, Mitacs, Université Laval, The Chair in Poultry
Research at the Faculty of Veterinary Medicine, University of Montreal and the Tunisian
Government National Scholarship Program for financial support.

124
3.8. References
Abd El-Hakim, A.S., Cherian, G., Ali, M.N., 2009. Use of organic acids, herbs and their
combination to improve the utilization of commercial low protein broiler diets.
International. Poultry Science. 8, 14-20.
AOAC, 2000. Official Methods of Analysis. 17th Edition, the Association of Official
Analytical Chemists, Gaithersburg, MD, USA. Methods 925.10, 65.17, 974.24,
992.16.
Bach Knudsen, K. E ., 2014. Fiber and nonstarch polysaccharide content and variation in
common crops used in broiler diets. Poultry Science. 2380–2393.
Baurhoo, B., Ferket, P. R., Zhao, X., 2009. Effects of diets containing different
concentrations of mannanoligosaccharide or antibiotics on growth performance,
intestinal development, cecal and litter microbial populations, and carcass parameters
of broilers. Poultry Science. 88, 2262-2272.
Castanon, J.I.R., 2007. History of the Use of Antibiotic as Growth Promoters in European
Poultry Feeds. Poultry Science. 86, 2466-2471.
Casterline, J.L., Oles, C.J., Ku, Y., 1997. In vitro fermentation of various food fiber fractions.
Journal of Agricultural and Food Chemistry. 45, 2463–7.
Chamba, F., Puyalto, M., Ortiz, A., Torrealba, H., Mallo, J.J., Riboty, R., 2014. Effect of
Partially Protected Sodium Butyrate on Performance, Digestive Organs, Intestinal
Villi and E. coli Development in Broilers ChickensInternational Journal of Poultry
Science. 13, 390-396.
Chee, S. H., Iji, P. A., Choct, M., Mikkelsen, L. L., Kocher, A., 2010. Functional interactions
of manno-oligosaccharides with dietary threonine in chicken gastrointestinal tract. I.
Growth performance and mucin dynamics. British Poultry Science. 5, 658–666.
Chicken Farmers of Canada, 2020. The Antimicrobial Use Reduction Strategy.
https://www.chickenfarmers.ca/the-antimicrobial-use-reduction-strategy/. (Page
consultee le 18 juillet 2020).
Choct, M., 2009. Managing gut health through nutrition. British Poultry Science. 50, 9-15.
Collins, J.T., Badireddy, M., Anatomy, Abdomen, Pelvis., 2019. Small Intestine. In:
StatPearls. StatPearls Publishing, Treasure Island (FL).
Daou, C., Zhang, H., 2012. Oat beta-glucan: Its role in health promotion and prevention of
diseases. Comprehensive Reviews in Food Science and Food Safety.11, 355-365.
De Vrese, M, Schrezenmeir, J., 2008. Probiotics, prebiotics, and synbiotics. Advances in
Biochemical Engineering-Biotechnology. 111, 1-66.
Diarra, M.S., Malouin, F., 2014. Antibiotics in Canadian Poultry Productions and
Anticipated Alternatives. Frontiers in Microbiology. 5, 282.
Eckersall, P.D., Bell, R., 2010. Acute phase proteins: biomarkers of infection and
inflammation in veterinary medicine. Veterinary Journal. 185, 23-27.
Edens, F.W., 2003. An alternative for antibiotic use in poultry: probiotics. Revista Brasileira
de Ciência Avícola. 5, 75-97.

125
Engberg, R.M., Hedemann, M.S., Leser, T.D., Jensen, B.B., 2000. Effect of Zinc Bacitracin
and Salinomycin on Intestinal Microflora and Performance of Broilers. Poultry
Science. 79, 1311-1319.
Ermis, B., Yildirim, A., Örs, R., Tastekin, A., Ozkan, B., Akcay, F., 2005. Influence of
Smoking on Serum and Milk Malondialdehyde, Superoxide Dismutase, Glutathione
Peroxidase, and Antioxidant Potential Levels in Mothers at the Postpartum Seventh
Day. Biological Trace Element Research. 105, 27-37.
Femia, A.P., Salvadori, M., Broekaert, W.F., Francois, I.E.J.A., Delcour, J.A., Courtin, C.M.,
Caderni, G., 2010. Arabinoxylan-oligosaccharides (AXOS) reduce preneoplastic
lesions in the colon of rats treated with 1, 2-dimethylhydrazine (DMH). European
Journal of Nutrition. 49, 127-132.
Field, C., Johnson, J., I., R, Schley, P., D., 2002. Nutrients and their role in host resistance to
infection. Journal of Leukocyte Biology. 71, 6–32.
Gadde, U., Oh, S. T., Lee, Y. S., Davis, E., Zimmerman, N., Rehberger, T., 2017. The effects
of direct-fed microbial supplementation, as an alternative to antibiotics, on growth
performance, intestinal immune status, and epithelial barrier gene expression in
broiler chickens. Probiotics and Antimicrobial Proteins. 9, 397.405.
Gao, J., Zhang, H. J., Yu, S. H., Wu, S. G., Yoon, I., Quigley, J., et al., 2008. Effects of yeast
culture in broiler diets on performance and immunomodulatory functions. Poultry
Science. 87, 377–1384.
Ghadban, G. S., 2002. Probiotics in broiler nutrition – a review. Arch. Geflugelk. 66, 49-58.
Gibson, G. R., Roberfroid, M. B., 1995. Dietary modulation of the human colonic microflora
introducing the concept of probiotics. Journal of Nutrition. 125, 1401-1412.
Gong, L., Baikui, W., Xiaoqiang, M., Han, X., Yan, Q., Weifen, L., Yingshan, Z., 2018.
Effects of three probiotic Bacillus on growth performance, digestive enzyme
activities, antioxidative capacity, serum immunity, and biochemical parameters in
broilers. Animal Science. 89, 1561-1571.
Gonzalez-Ortiz, G., Dos Santos, T. T., Vienola, K., Vartiainen, S., Apajalahti, J., Bedford,
M.R., 2019. Response of broiler chickens to xylanase and butyrate supplementation.
Poultry Science. 98, 3914–3925.
Gonzalez-Ortiz, G., Sola-Oriol, D., Martinez-Mora, M., Perez, J. F., Bedford, M. R., 2017.
Response of broiler chickens fed wheat based diets to xylanase supplementation.
Poultry Science. 96, 2776– 2785.
Hardin, B. E., Roney, C.S., 1989. Effects of pH on selected bacteria. Alabama Department
of Agriculture and Industry Report.
Huyghebaert, G., Ducatelle, R., Van Immerseel, F., 2011. An update on alternatives to
antimicrobial growth promotors for broilers. Veterinary Journal. 187, 182-188.
Iji, P. A., Saki, A. A., Tivey, D. R., 2001. Intestinal structure and function of broiler chickens
on diets supplemented with a mannanoligosaccharide. Journal of the Science of Food
and Agriculture. 81, 1186–1192.
Jarosz, Ł., Marek, A., Grądzki, Z. et al., 2019. Effect of Zinc Sulfate and Zinc Glycine
Chelate on Concentrations of Acute Phase Proteins in Chicken Serum and Liver
Tissue. Biological Trace Element Research. 187, 258–272.
Kamińska B., 2003. The effect of various grains in grower diet on broiler performance and
dietetic value of carcass. Annals of Animal Science, Suppl. 2, 185-188.
Khan, S.H., Iqbal, J., 2016. Recent advances in the role of organic acids in poultry nutrition.
Journal of Applied Animal Research. 44, 359-369.

126
Kim, S.W., Mateo, R.D., Yin, Y.L., Wu, G., 2007. Functional amino acids and fatty acids for
enhancing production performance of sows and piglets. Asian-Australasian Journal
of Animal Sciences. 20, 295–306
Lee, K. W., Kim, D. K., Lillehoj, H. S., Jang, S. I., Lee, S. H., 2015. Immune modulation by
Bacillus subtilis -based direct-fed microbials in commercial broiler chickens. Animal
Feed Science and Technology. 200, 76–85.
Lei, X., Piao, X., Ru, Y., Zhang, H. and Peron, A., 201. Effect of Bacillus
amyloliquefaciens‐based direct‐fed microbial on performance, nutrient utilization,
intestinal morphology and cecal microflora in broiler chickens. Asian-Australasian
Journal of Animal Sciences. 28, 239-246.
MacLeod, M. G., Valentine, J., Cowan, A., Wade, A., McNeill, L., Bernard, K., 2008. Naked
oats: metabolisable energy yield from a range of varieties in broilers, cockerels and
turkeys. British Poultry Science. 49, 368-377.
Marković, R., Šefer, D., Krsticv, M., Petrujkicv, B., 2009. Effect of different growth
promoters on broiler performance and gut morphology. Archivos de Medicina
Veterinaria. 4, 163-169.
Mehdi, Y., Létourneau-Montminy, M. P., Gaucher, M. L., Chorfi, Y., Gayatri, S., Rouissi,
T., et al., 2018. Use of antibiotics in broiler production: global impacts and
alternatives. Animal Nutrition. 4, 170–178.
Moquet, P. C. A., Salami, S. A., Onrust, L., Hendriks, W. H., Kwakkel; R. P., 2018. Butyrate
presence in distinct gastrointestinal tract segments modifies differentially digestive
processes and amino acid bioavailability in young broiler chickens. Poultry Science.
97, 167-176.
Murugesan, G. R., Ledoux, D. R., Naehrer, K., Berthiller, F., Applegate, T. J., Grenier, B.,
Phillips, T. D., Schatzmayr, G., 2015. Prevalence and effects of mycotoxins on
poultry health and performance, and recent development in mycotoxin counteracting
strategies. Poultry Science. 94, 1298–1315.
Osek M., Janocha A., Klocek B., Wasiłowski Z., Milczarek A., 2003. The influence of
different content of naked oat in plant feed on performance and post-slaughter value
of broiler chicken. Annals of Animal Science, Suppl. 2, 205-208.
Patel, S., Goyal, A., 2012. The current trends and future perspectives of prebiotics research:
A review. 3 Biotech. 2, 115-125.
Park, J. H., Yun, H. M., Kim, I. H., 2018. The effect of dietary Bacillus subtilis
supplementation on the growth performance, blood profile, nutrient retention, and
caecal microflora in broiler chickens. Journal of Applied Animal Research. 46, 868-
872.
Payne, J. B., Osborne, J. A., Jenkins, P. K., Sheldon, B. W., 2007. Modeling the growth and
death kinetics of Salmonella in poultry litter as a function of pH and water activity.
Poultry Science. 86:191–201.
Rajkovich, S., Enders, A., Hanley, K., Hyland, C., Zimmerman, A.R., Lehmann, J., 2011.
Corn growth and nitrogen nutrition after additions of biocars with varying properties
to a temperate soil. Biology and Fertility of Soils. 48, 271-284.
Raza, M., Biswas, A., Mandal, A.B., Yadav, A.S., 2017. Effect of Dietary Supplementation
of Butyric Acid on Growth Performance and Intestinal Microbial Load in Broiler
Chickens. Animal Feed Science and Technology.17, 353-359.

127
Ruma., 2016. Responsible Use of Medicines in Agriculture Alliance (Ruma) Information on
Antibiotic Resistance. https://www.ruma.org.uk/2016/11/?cat=47. (Page consultee le
05 avril 2019).
Ruth, M., R, Field, C., J, 2013. The immune modifying effects of amino acids on gut-
associated lymphoid tissue. Journal of Animal Science and Biotechnology. 4, 1–10.
Sauvant, D., J. Perez, M., Tran, G., 2004. Tables of composition and nutritional value of feed
materials: Pig, poultry, sheep, goats, rabbits, horses, and fish. Wageningen Academic
Publisher.
SAS Version 9.4. 2014. Cary, NC: SAS Institue Inc.
Scheuer, J.P., Chalk, T.B., 1986. Clinical tests: Histology. Wolfe Medical Publ Ltd,
Netherlands. London, 1986. No. of pages: 128.
Sikandar, A., Zaneb, H., Younus, M., Masood, S., Aslam, A., Khattak, F., 2017. Effect of
sodium butyrate on performance, immune status, microarchitecture of small intestinal
mucosa and lymphoid organs in broiler chickens. Asian-Australasian Journal of
Animal Sciences. 30, 690–9.
Simon, O., Jadamus, A., Vahjen, W., 2001. Probiotic feed additives-effectiveness and
expected modes of action. Journal of Animal and Feed Sciences.10, 51-67.
Suez, J., Zmora, N., Zilberman-Schapira, G., Mor, U., Dori-Bachash, M., Bashiardes, S., et
al. 2018. Post-antibiotic gut mucosal microbiome reconstitution is impaired by
probiotics and improved by autologous FMT. Cell. 174, 1406–1423.
Svihus, B., Gullord, M., 2002. Effect of chemical content and physical characteristics on
nutritional value of wheat, barley and oats for poultry. Animal Feed Science and
Technology. 102, 71–92.
Szymczyk B., Hanczakowski P., Szczurek W., 2005. Performance and intestinal viscosity in
broilers fed diets containing dehulled or naked oats and enzymes. Journal of Animal
and Feed Sciences.14, 491-494.
Teng, P.Y.; Kim, W.K., 2018. Review: Roles of Prebiotics in Intestinal Ecosystem of
Broilers. Frontiers in Veterinary Science. 5, 245.
Van den Borne, J. J. G. C., Heetkamp, M. J. W., Buyse, J., Niewold, T.A., 2015. Fat coating
of Ca butyrate results in extended butyrate release in the gastrointestinal tract of
broilers. Livestock Science. 175, 96–100.
Wu, S. B., Rodgers, N., Choct, M., 2011. Real-time PCR assay for Clostridium perfringens
in broiler chickens in a challenge model of necrotic enteritis. Applied and
Environmental Microbiology. 77, 1135-1139.
Yang, Z., Qin, C., Weng, P., Zhang, X., Xia, Q., Wu, Z., Xiao, J., 2020. In vitro evaluation
of digestive enzyme inhibition and antioxidant effects of naked oat phenolic acid
compound (OPC). International Journal of Food Science Technology. 55, 2531-2540.
Yegani, M., Korver, D.R., 2008. Factors affecting intestinal health in poultry. Poultry
Sciences. 87, 2052-2063.
Zhao, C., Ge, B., De Villena, J., Sudler, R., Yeh, E., Zhao, S., 2001. Prevalence of
Campylobacter Spp., Escherichia Coli, and Salmonella Serovars in Retail Chicken,
Turkey, Pork, and Beef from the Greater Washington, Dc, Area. Applied and
Environmental Microbiology. 67, 5431-5436.

128
Chapitre 4 Effect of different phytogenic feed additives
on growth performance and intestinal health
Ce chapitre est présenté sous forme d’article et sera soumis dans la revue Journal of Animal
Science.

129
Effect of different phytogenic feed additives on broiler chicken growth performance
and intestinal health
A. Rouissi., * F. Guay*, M. Boulianne †, and M. P. Létourneau-Montminy *
* Department of Animal Science, Laval University, Québec City, QC, Canada. † Department
of Clinical Science, Faculty of Veterinary Medicine, University of Montreal, Saint-
Hyacinthe, QC, Canada.
Corresponding author : Marie-Pierre Létourneau-Montminy
E-mail : [email protected]
Abbreviations: AGP, antibiotic growth promoters; ADFI, average daily feed Intake; ADG,
average daily Gain; BWG, body weight gain; Ca, calcium; CD, crypt depth; DM, dry mater;
FCR, feed conversion ratio; FI, feed intake; oligosaccharide; Na, sodium; P, phosphor;
SCFA, short-chain fatty acid; VFA, Volatile fatty acid; VL, villus length; VL/CD, villus
length/ crypt depth.

130
4.1. Résumé
Le but était d'évaluer l’effet d’un antibiotique, d’additifs phytogènes; Phyto1, Phyto2, Phyto3
et Phyto4 sur les performances de croissance et la santé digestive des poulets de chair. Pour
la phase de finition, tous les phytogéniques ont augmenté la CJM et le GMQ (3 à 5%), à
l'exception du Phyto4 qui était le seul à augmentr l’IC (3%) par rapport au régime témoin
sans antibiotiques. Phyto4 augmente la longueur de duodénum et l’iléon au jour 35, tandis
que Phyto2 avaient le jéjunum et l'iléon plus courts au jour 35. Phyto4 avait la plus forte
concentration des AGCC aux jours 28 et 35 et la valeur la plus élevée de concentration en
butyrate dans le caeca. Phyto1, Phyto3 et Phyto4 réduisent la LV tandis que phyto2
l’augmente. Les effets des alternatives aux AFC proposés sont mesurables mais leurs modes
d'action exacts restent à définir car la composition exacte des produits phytogéniques n'est
pas connue.

131
4.2. Abstract
The use of phytogenic feed additives in poultry production as an alternative to antibiotic
growth promoters has gained recent interest. The aim of the present study was to investigate
the effect of four phytogenic feed additives on the growth performance and gut health of
broiler chickens. Five hundred Cobb 500 chicks were fed a control diet (C) or a diet
supplemented with antibiotic (PC) or phytogenic feed additives; Phyto1, Phyto2, Phyto3, and
Phyto4. The study was carried out as a complete random blocked design with 10 pens for
each diet, 45 birds per pen. PC diet improved AFG and FCR (by +6%; -3%, respectively)
during the starter phase (day 0 to day 10). In the grower phase, Phyto1 increased FCR
(P<0.05) diet while ADG and ADFI were similar. Phyto2 increased ADFI and ADG without
influencing FCR. Phyto3 did not modify growth performance. Phyto4 increased ADFI by
3% without affecting ADG, but increased FCR (2%). For the finisher phase, all phytogenics
increased ADFI and ADG (3 to 5%), excepted for Phyto4, This was the only phytogenic feed
additives with an increased FCR (3%) when compared to the control diet i.e., without
antibiotics. Birds fed Phyto4 had a longer duodenum at d14 and d35 and ileum at d35, while
birds fed Phyto2, had a shorter jejunum and ileum at d35. Phyto4 had the highest SCFA
concentration at day 28 and 35. At d28, butyrate percentage was highest in PC, while at d35
the highest butyrate concentration was observed in the Phyto4 group. At 28d of age, birds
fed Phyto1, Phyto3, and Phyto4 had lower villus length than control birds while Phyto2
showed higher villus length and crypt depth. Litter moisture was higher in PC and Phyto2.
At the end, Phyto1 had lowest litter moisture while C had highest moisture. Litter pH on d34
was lower in C and higher in Phyto1 and Phyto4. The effects of the proposed non-antibiotic
growth promoters are measurable and could contribute to livestock performance.
Nevertheless, their exact modes of action remained to be defined because their exact
composition is not publicized.
Keywords: Broilers, antibiotic, alternative to antibiotic growth promoters, growth
performance, gut health, Herbal extracts.

132
4.3. Introduction
Since their discovery, antibiotics have played a major role in the prevention, control, and
treatment of humans and animals bacterial diseases. It is also well known that antibiotic use
in animal feed promotes animal growth, enhances feed efficiency, and improves the quality
of animal products (Mehdi et al., 2018; Yang et al., 2019; Hamid et al., 2019). Antibiotics
added directly to the feed as growth promoter, namely antibiotic growth promoters (AGP)
have been an important component in the development of intensive and large-scale farming.
However, the wide use of antibiotics as feed additives have led to the development of
resistant bacteria to drugs used to treat infections hence posing a potential risk for humans
(Castanon, 2007) which led to their progressive ban worldwide. This has had important
impacts on animal production such as the increase in disease prevalence, increased mortality
and morbidity rates and decreased performance. To counter the withdrawal of antibiotics as
feed additives, several alternatives or replacements solutions have been proposed (Diarra and
Malouin, 2014; Mehdi et al., 2018) all aiming for a good feed conversion ratio, low mortality
and no intestinal dysbiosis by favoring good bacteria and healthy gut. Among these,
probiotics, prebiotics, enzymes, organic acids and considerable attention has been given to
phytogenic feed additives (Gurbuz and Ismael 2016; Yatoo et al. 2017).
Phytogenics are a group of natural feed additives derived from herbs, spices, or plants
(Hashemi and Davoodi, 2010). They have been exploited in animal nutrition, particularly for
their antimicrobial, antiinflammatory, antioxidative, and antiparasitic activity. Phytogenic
compounds can be divided in herbs (flowering, nonwoody, and nonpersistent plants), spices
(herbs with an intensive smell or taste commonly added to human food), essential oils
(volatile lipophilic compounds derived by cold expression or by steam or alcohol
distillation), or oleoresins (extracts derived by nonaqueous solvents) (Windisch et al., 2008)
and then into several categories, such as phenolics/polyphenols, terpenoids/essential oils
(EO), alkaloids, lectins/polypeptides (Windisch et al., 2008). The exact antimicrobial
mechanism is poorly understood since most studies investigate blends of various active
compounds and report the effects on production performance rather than the physiological

133
impacts. However, some in vivo observations support the assumption that most of the
antimicrobial effect of phytogenic feed additives resides in a reduction of intestinal pathogen
pressure (Windisch et al., 2008). The growth promoting effect is likely related to pH
stabilisation of the gut and role as organic acid. It has also been reported that essential oils
and oleoresins mixed products act on the activity of intestinal microbiota in broilers (Jamroz
et al., 2003, 2005; Mitsch et al., 2004). Morphological changes in gastrointestinal tissues and
their possible benefits to the gut are also reported for other AGP (Cheng et al., 2014).
Based on the results of previous studies and a meta-analysis by Rouissi et al. (2020), the
objective of this study was to evaluate the in vivo effects of phytogenic products based on
essential oil, oleoresin, and plant extracts already available and used in Canada for their
growth promoting, gut morphology, and litter quality effects.

134
4.4. Materials and methods
The research was approved by Animal Care Committee of Laval University (CPAUL)
(accredited by Canadian Council of Animal Care (CCAC), Guidelines of the Canadian
Council of Animal Care, 2009) and conducted from July 4 to August 8, 2017 at the
Deschambault Animal Science Research Center (120 street Roy, Deschambault, QC G0A
1S0, Quebec, Qc, Canada).
Birds and housing
A total of two thousand and seven hundred (2,700) day-old Cobb 500 chicks were obtained
from a commercial hatchery, weighed and randomly divided in 6 dietary treatment groups
(10 replicates) with 45 birds per pen. Lighting program was in accordance with Ross 308
guidelines. The temperature was maintained at 33 °C during the first week and gradually
reduced to 22 °C by the end of the third week and maintained until 35 days. Diet fulfilled
nutrient requirements of Cobb 500 broilers (Cobb 500 Broiler Performance and Nutrition
Supplement, 2017) (Table 4-1). All chicks received no antibiotics but were vaccinated
against Marek's disease and infectious bronchitis virus in ovo.
Experimental design
Experimental groups were divided in three phases: starter (0-10 d), grower (11-21 d) and
finisher (22-35 d). All treatment diets contained contained zoalene (Zoamix, concentration
250mg/kg) been used with the dose of integration into the feed for the period of 0-21 days,
were manufactured as follows: 1) a control with antibiotic (0.375 kg/tonne of feed of
Stafac©: virginiamycin) 2) a positive control without antibiotic (C), 3) positive control +
Phyto1 (Oleobiotec Poultry; Mixture of EO and oleoresins which contains mainly carvacrol,
thymol, cinnamaldehyde, eugenol, ginger, curcumin, and capsaicin.) incorporated at a rate of
50 mg/kg in the starter phase, 52.5 mg/kg in the grower phase and 117.5 mg/kg in the finisher

135
phase, 4) positive control + Phyto2 (Micro-Aid; Yucca schidigera extract, verxite granules,
and calcium carbonate) was added at a rate of 125 mg/kg in the starter phase, 131.25 mg/kg
in the grower phase and 293.75 mg/kg in the finisher phase, 5) positive control + Phyto3
(XTract Instant; Mixture of EO and oleoresin which contains mainly carvacrol,
cinnamaldehyde, and capsicum oleoresin) was added at a rate of 50 mg/kg in the starter
phase, 52.5 mg/kg in the grower phase and 117.5 mg/kg in the finisher phase and 6) positive
control + Phyto4 (Phyteck-P; Mixture of vitamin K3, oleoresin of curcuma and EO from
thyme, pepper mint, and cinnamon) was added at a rate of 250 mg/kg in the starter phase,
262.5 mg/kg in the grower phase and 587.5 mg/kg in the finisher phase.
Data Collection
Litter moisture and pH
A 100g litter sample was collected from all 50 pens on day 9, 20 and 30 for moisture content
and pH measurement. Litter samples were oven-dried at 100°C for two days and percent
moisture content was calculated with the following formula: [100 - (dry weight-content
weight) / (wet weight -content weight)] * 100. For pH measurement (Rajkovich et al 2011),
1 g of ground litter sample (<1 mm) was added to 20 mL of deionized water. The suspension
was put on a shaker (Boekel Scientific, Orbitron II Laboratory Mixer, Model 260250,
Pennsylvania Blvd, USA) for an hour and then left to stand for one hour for the liquid and
solid phases to separate. The pH was measured in the liquid phase using a portable pH meter
(Ross, Orion Star A221, Thermo Scientific, Beverly, CA, USA).
Feed analysis
Feed was sampled during the batch bagging process and at the beginning of each dietary
phase. Dry matter, crude protein, calcium, phosphorus, sodium, fat and energy (Table 1) were
analyzed according to AOAC (2000) methods.

136
Performance data
Birds and feed were weighed (on days 0, 10, 21, 29 and 35 to evaluate average daily feed
intake (ADFI), average daily gain (ADG), and feed conversion ratio (FCR) for each growing
diets. Mortality was recorded daily. All dead birds and those euthanized for tissue sampling
(10, 21 and 35 d) were individually weighted and growth performance data were corrected
for mortality.
Intestinal physiology and histomorphometry parameters
At 14, 28 and 35 d of age one bird per pen (10 per treatment) was euthanized by cervical
dislocation. Duodenum, jejunum and ileum lengths were measured in all euthanized birds.
Once emptied of its contents, a section of the distal ileum was retrieved for
histomorphometry. A first cross-section was made 1 cm from Meckel's diverticulum and a
second one 1.5 cm distally. Each ileal sample was individually fixed in a formalin solution
and stored at room temperature (Marković et al., 2009). Multiple 5µm sections of paraffined
tissues were obtained from each sample with a microtome and stained with hematoxylin and
eosin (Scheuer et al., 1986). Villus and crypt measurements were based on 18 villi and crypt
per bird using a microscope (Microscope Central, Nikon Eclipse E600 Fluorescence,
Feasterville, PA, USA) (objective lens: X4) and a PC-based image analysis system and
software analysis (Q-Capture, Fiji).
Caeca short-chain fatty acid (SCFA) concentrations
One gram of cecal content was extracted upon euthanasia with an alcohol-cleaned spatula,
placed in a 15 mL conical tube, acidified with 2 ml of H2SO4 (1.5%), vortexed then frozen
at -20 ° C. The frozen sample was homogenized and centrifuged at 10,000 RPM for 15 min
at 4oC. The supernatant was analysed on a Hewlett Packard 6890N gas chromatograph

137
(Agilent Technologies, Wilmington, DE, USA) equipped with a flame ionization detector
and an autosampler (Hewlett Packard, Avondale, PA, USA) for volatile fatty acid.
Statistical analyses
The study was carried out as a randomized complete block design. For repeated measures,
time and treatment effects, as well as their interactions, were tested with the MIXED
procedure (SAS 9.4, SAS, NC, USA). For the other measurements, feed additives were
compared to the control using the MIXED procedure and the Dunnett test (SAS 9.4, SAS,
NC, USA). A difference was considered significant at P <0.05 and P <0.10 was considered
to indicate a tendency.

138
Table 4-1 Basal diet composition for starter, grower and finisher phase.
Ingredient % as fed Starter (0-10 d) Grower (11-21 d) Finisher (22-35 d)
Maize meal 60.4 62.6 63.08
Soybean meal 30.3 26.4 24.5
meat and bone meal 5.0 5.0 5.0
Phosphate monocalcique 0.82 0.61 0.47
Limestone 0.55 0.39 0.62
Fat mixed, % 1.0 3.2 4.3
Biolys 54.6% 0.46 0.4 0.4
DL-methionine 0.385 0.34 0.265
Micropremix 0.2 0.2 0.2
Salt 0.12 0.15 0.19
Bicarbonate 0.5 0.19 0.16
Premix 0.15 0.15 0.15
Threonine 0.13 0.105 0.75
Choline chloride 60% 0.076 0.07 0.076
Phytase 0.15 0.015 0.055
Zoamix* 0.05 0.05 -
Nutritional content (calculated)
Fat, % 2.9 4.8 6.8
Crude protein (N × 6.25), % 21.4 19.8 18.8
Metabolisable energy, kcal/kg 3249 3118 3249
Na, % 0.18 0.18 0.18

139
Ca, % 0.78 0.82 0.78
P, % 0.57 0.58 0.57
** Stafac (Antibiotic) (0.375 kg / ton)
* All treatments will contain Zoamix (0.5kg / t) for the 0-21 day period.

140
4.5. Results
Growth performance
During the starter phase (0-10d), there was an increased of ADG (6%; P<0.05) and decrease
of FCR (3%; P=0.08) in antibiotics diet in comparison to C diet without antibiotics (Table
4-2). When comparing the C with the AGP, no effect was found. In the grower phase (11-
21d), birds receiving antibiotics ate more than C diet (4%, P=0.05). Phyto1 increased FCR
(P<0.05) compared to C diet while ADG and ADFI were similar. Phyto2 increased ADFI
(+7%, P<0.05) and ADG (+7%, P<0.001) without affecting FCR. Phyto3 did not modified
growth performance. In comparison to C diet, Phyto4 tended to increase ADFI (3%, P=0.09)
resulting in an increased FCR (2%, P=0.05). In the finisher phase (22-35d), birds receiving
antibiotics had a 2% higher ADG (0.04). In comparison to C diet, all the phytogenics
increased ADFI by 3 to 5% (P<0.05); ADG was increased for Phyto1 (5%, P<0.01), Phyto2
(4%, P<0.01), and Phyto3 (3% P<0.05), but not for Phyto4. The latter was the only AGP to
show an increased FCR (3%, P<0.05) in comparison to C diet. For the overall experience,
birds fed antibiotics had higher ADFI and ADG (4%, P<0.01) than C diet. As for the finisher
phase, all phytogenics increased ADFI (between 4 to 6%, P<0.01) and ADG (3 to 5%,
P<0.05) excepted for Phyto4 which showed an increased FCR (3%, P<0.01) in comparison
to the control diet without antibiotics.

141
Table 4-2 Average daily gain, average daily feed intake, and feed conversion ratio of broiler chicken fed with the six treatments
for the three feeding periods (n/treatment=450).
Dietary treatment P-value1
Items Antibiotic Control Phytogenic1 Phytogenic2 Phytogenic3 Phytogenic4 SEM T1 T3 T4 T5 T6
starter (0-10d)
ADG, g/d 18.17 17.08 17.59 17.31 17.48 17.62 0.351 0.021 0.270 0.597 0.379 0.264
ADFI, g/d 18.30 17.77 18.00 18.27 18.28 18.33 0.350 0.267 0.641 0.308 0.291 0.261
FCR 1.008 1.042 1.023 1.055 1.046 1.042 0.014 0.085 0.325 0.512 0.828 0.978
Mortality2,% 0.023 0.041 0.036 0.027 0.025 0.038 0.008 0.206 0.991 0.392 0.318 0.934
Grower (11-21d)
ADG, g/d 55.99 54.20 53.89 58.32 55.61 55.09 0.873 0.139 0.775 <0.001 0.239 0.475
ADFI, g/d 73.34 70.59 72.10 75.77 72.00 73.02 1.346 0.050 0.300 <0.001 0.321 0.093
FCR 1.311 1.300 1.339 1.298 1.295 1.327 0.020 0.580 0.041 0.914 0.759 0.050
Mortality2,% 0.016 0.024 0.018 0.030 0.032 0.028 0.006 0.492 0.732 0.567 0.361 0.870
Finisher (22-35d)
FBW, kg 2.226 2.208 2.258 2.315 2.263 2.242 0.952 0.075 0.080 <0.001 0.031 0.051
ADG, g/d 97.03 94.76 99.11 98.31 97.80 95.48 1.087 0.040 0.005 0.002 0.050 0.634
ADFI, g/d 157.22 154.27 159.49 161.39 159.14 160.18 1.610 0.201 0.026 0.002 0.038 0.012
FCR 1.620 1.628 1.610 1.643 1.627 1.678 0.017 0.756 0.477 0.566 0.961 0.048
Mortality2,% 0.021 0.031 0.030 0.030 0.021 0.031 0.008 0.466 0.924 0.738 0.478 0.846
Overall (0-35d)
ADG, g/d 60.75 58.58 60.33 61.32 60.42 59.18 0.571 0.006 0.025 <0.001 0.019 0.436
ADFI, g/d 89.64 86.05 89.35 91.26 89.39 89.77 0.871 0.003 0.006 <0.001 0.006 0.002
FCR 1.475 1.469 1.481 1.488 1.479 1.517 0.010 0.661 0.431 0.208 0.488 0.002
Mortality,% 0.041 0.074 0.065 0.067 0.058 0.076 0.014 0.508 0.999 0.999 0.991 0.998
ADG: average daily gain (g); ADFI: Average daily feed intake (g/kg); FCR: feed conversion ratio; FBW: final body weight (kg); 1Control versus antibiotic
(T1), Phytogenic1 (T3), Phytogenic2 (T4), Phytogenic3 (T5) or Phytogenic4 (T6); 2Percentage of birds at this stage, not cumulative.

142
Length of digestive segments
As expected, the length of all intestinal segment increased with age (P<0.001; Figure 4-1).
However, the growth of the intestinal tract was different depending on the dietary treatments
(Interaction diet x time P<0.05). These results showed that birds fed Phyto4 treatment had
the longest duodenum at d14 and d35 and ileum at d35 (Figure 4-1a, 4-1b, 4-1c), while birds
fed Phyto2 treatment, had the shortest jejunum and ileum at d35.
Figure 4-1 Length (cm) of a) duodenum (Time: P<0.001, Trt: P=0.099, Time x Trt: P=0.092),
b) jejunum (Time: P<0.001, Trt: P=0.282, Time x Trt: P=0.029), and c) ileum (Time:
P<0.001, Trt: P=0.057, Time x Trt: P=0.113) of broilers from the six different treatment
groups (n/treatment = 450).

143
Caeca short chain fatty acid content
Total short chain fatty acid concentration in the caeca (SCFA) was influenced by age
(P<0.0001) but this effect depended on dietary treatment (Interaction diet x time, P < 0.001).
Birds fed Phyto4 had the highest SCFA concentration at day 28 and 35 (Figure 4-2a) while
those fed Phyto1 had the lowest concentration. At d 14, birds receiving antibiotics and C diet
had the higher percentage of acetate while Phyto2 had the lowest percentage (Figure 4-2b).
At d 22 and d 28, Phyto3 had the highest acetate percentage of all groups. At d 35, acetate
percentage was highest in bird fed Phyto1 and lowest for Phyto4 while opposite effect was
observed for butyrate percentage (Figure 4-2c). No significant difference was observed for
propionate percentage between treatments at d 22, 28 and 35, but on d 14, birds fed Phyto1
had the highest percentage while those fed Phyto3 and C had the lowest percentage (Figure
2d). The butyrate percentage at d14 was lowest in antibiotics and Phyto1 treatments and
highest in birds fed Phyto3 (Figure 4-2d). At d 28, butyrate percentage was highest in PC
treatment while at d35 Phyto4 showed the highest butyrate concentration value.

144
Figure 4-2 Cecal concentration of a) total SCFA (mmol; Time: P<0.0001, Trt: P=0.0009,
Time x Trt: P= 0.0036), b) acetate (%; Time: P<0.0001, Trt: P=0.1791, Time x Trt:
P<0.0001); c) propionate (%; Time: P<0.0001, Trt: P=0.4077, Time x Trt: P=0.7648), and
d) butyrate (%; Time: P=0.0003, Trt: P=0.0269, Time x Trt: P=0.0031) of broilers from the
six different treatment groups (n/treatment =450).
Ileum histology
At 28d of age, birds fed Phyto1, Phyto3, and Phyto4 had lower villus lengths than C birds
(11%, 17% and 13% respectively; P<0.05; Table 4-3) while Phyto2 had higher villus lengths
than C birds (26%; P<0.05). For crypt depth, only birds fed Phyto2 had higher values than

145
those fed the C diet (P<0.05). Villus length /crypt depth ratios were not affected by the
dietary treatment.

146
Table 4-3 Ileal histomorphological parameters of 28 day-old broiler chicken from the six different treatment groups
(n/treatment=450).
1Control versus antibiotic (T1), Phytogenic1 (T3), Phytogenic2 (T4), Phytogenic3 (T5) or Phytogenic4 (T6); *: Villus lengths and depths in μm.
Dietary treatment P-value1
Items* Antibiotic Control Phytogenic1 Phytogenic2 Phytogenic3 Phytogenic4 SEM T1 T3 T4 T5 T6
Villus
length (μm)
391.10 351.70 315.12 443.96 292.77 308.63 23.66 0.245 0.028 0.034 0.005 0.017
Crypt
depth (μm)
44.50 43.18 43.66 47.24 43.48 42.44 7.11 0.876 0.921 0.047 0.904 0.808
VL/CD 8.94 11.00 9.25 8.68 9.27 10.36 1.34 0.238 0.858 0.882 0.846 0.413

147
Litter Properties
As expected, litter moisture increased with age (P<0.001; Figure 4-5) but the kinetic change
depended on the dietary treatment (Interaction diet x time, P <0.001). Litter moisture was
higher in antibiotic fed birds on d 7, 13 and 27 when Phyto2 reached the same level. On d
34, Phyto1 treatment had the lowest litter moisture while C treatment had the highest
moisture.
Figure 4-3 Litter pH from 0- 21 day of broiler pens from the six different treatment groups
(n/treatment = 450) (Time: P<0.001, Trt: P=0.124, Time x Trt: P=0.0003).

148
Figure 4-4 Litter pH from 21- 34 day of broiler pens from the six different treatment groups
(n/treatment = 450). (Time: P<0.001, Trt: P=0.124, Time x Trt: P=0.0003).
Litter pH also increased with age (P<0.001; Figure 4-3, 4-4) but rising was different
according to the dietary treatment (Interaction diet x time, P= 0.01). On d 7, litter pH was
clearly higher in antibiotics fed birds (5%) while on d13 Phyto3 and antibiotics treatments
had higher pH litter (5.6) while Phyto 1 and 4 litter samples showed decreased pH
(respectively, 5.3 and 5.2). Litter pH on d34 was lower on C and higher in Phyto1 and Phyto4
treatments (respectively, 6.9, 7.1 and 7.1).

149
Figure 4-5 Litter moisture (%) of broiler pens from the six different treatment groups
(n/treatment = 450) (Time: P<0.001, Trt: P<0.001, Time x Trt: P<0.001).
4.6. Discussion
The main objective of this study was to validate the results of our meta-analysis (Rouissi et
al., 2020), measure the effect of phytogenics product as alternatives to AGP and learning
about their mode of action into the gut. Our meta-analysis (Rouissi et al., 2020) closely
looked at 79 publications from 2000 to 2017, and verified the impact of alternatives to AGP
on growth performance. In total, 27% of the papers studied the effect of essential oils with
the most studied EO oregano-based (65% of publications). Results obtained from this meta-
analysis (Rouissi et al., 2020), showed no effect on average daily feed intake, whereas
average daily gain increased linearly (P<0.001) and quadratically (P<0.05). Similarly, FCR
also decreased linearly and quadratically (P<0.001) with increasing oregano essential oil
(OEO) doses. When considering the quadratic effect, maximum effects for both ADG and
FCR were 7% improvement. The optimal dose of OEO for broiler chicken production was
estimated at 300 mg/kg of diet in our study.

150
When comparing the diet with and without antibiotics, an increased growth performance was
observed in all studies, for an overall improvement of 4% ADG and ADFI. Birds raised
without antibiotic likely had room for AGP to positively affect growth performance. The
exact mode of action of the growth promoting effect of antibiotics is still not exactly known.
Some research showed that AGP modifies the number and diversity of the normal gut
bacterial, increasing the bioavailability of nutrients available to the host and/or reducing the
production of microbial metabolites deleterious to animal growth (Gaskins et al., 2002;
Knarreborg et al., 2004). Alternatively, antibiotics were suggested to improve growth
performance through an anti-inflammatory effect directed toward the intestinal epithelium
(Niewold, 2007). Some recent studies showed that antibiotics is inversely correlated with the
activity of bile salt hydrolase, an intestinal bacteria-produced enzyme that exerts negative
impact on host fat digestion and utilization, and the abundance of
corresponding Lactobacilli producers (Lin, 2014).
On the phytogenic tested, three of them (Phyto1, 3 and 4) claimed to contain EO and
oleoresin, but the exact composition is not known. In terms of growth performance, these
three phytogenics showed an enhanced feed consumption, which can partially explained their
growth promoting effect. Phyto1 and Phyto3 improved in similar magnitude ADG and ADFI
in the finisher phase, both without affecting the FCR. Phyto 4 improved ADFI in the grower
fed birds without modifying the ADG hence resulting in an increased FCR. This is not
consistent with many studies conducted by Windisch et al. (2008) were the main mechanism
of the growth promoting effect was reported to be a reduced feed intake for a largely
unchanged body weight gain or final body weight, leading to an improved feed to gain when
feeding phytogenic compounds. However, the conditions in which these compounds were
tested (e.g. diet type, presence of challenge, bird age) can largely change the results
(Windisch et al., 2008).
Phyto1 significantly increased ADFI (4%) and ADG (3%) and marginally improved final
body weight (2%). To our knowledge there has been no scientific publication testing this

151
product as AGP. In a recent meta-analysis reporting different research and commercial trials
(n=25) using Phyto1 (Létourneau-Montminy et al., 2019), ADG was improved by 2.8±0.70%
and FCR decreased by 3.6±1.2% (P=0.006), similar to the current results. Surprisingly, this
phytogenic reduced the cecal SCFA concentration production by about 2 times in comparison
to C diet during the finisher phase. One of the main active compound of this EO blend is the
phenolic substance carvacrol. This lipophilic substance was found in vitro to be a very
efficient antimicrobial compound (Ben Arfa et al., 2006). Given that SCFA are the result of
the microbial activity (Van der Wielen er al., 2000), this result is likely related to the
antimicrobial activity of EO. Similar results have been obtained in piglets were the lower
SCFA production was correlated with a decrease in bacterial colony counts (Kroismayr et
al., 2008). Such effect on SCFA production was not observed with Phyto3 and Phyto4 that
are also supposed to contain carvacrol. Large variations in the concentration of active
compounds in EO have been reported (e.g. Burt, 2004) and might explain these discrepancies.
Phyto1 was also the only one to reduce litter moisture, which is often associated with a better
balance of nutrient or reduced dysbacteriosis (Collett, 2012). In fact, this can be due to the
reduction in the contents of anaerobic and aerobic germs, volatile fatty acids, and ammonia
small and large intestine as well as contents of biogenic amines in caecal chyme (Kroismayr
et al., 2008), as showed in piglets that reduce the need for water to excrete these toxics
compounds (Collet, 2012).
Regarding Phyto3, it significantly increased ADFI (4%) and ADG (3%) and improved final
body weight (2%). More results are available in the scientific literature on this phytogenic.
These results have also been reviewed in a meta-analysis (Oguey and Bravo, 2015) reporting
20 trials. It showed no significant effect on ADFI, an improvement of 4.5±0.72% of ADG
and of 3.9±0.83% of FCR. The other components measured did not differed for this
phytogenic. Jamroz et al. (2003) found that the body weight gain in broilers fed diets
supplemented with this phytogenic was 5.4% (150 mg/kg) and 8.1% (300 mg/kg) higher in
comparison with the negative control group. That can be explained by a positive action on
metabolism and the digestive system by stimulating of enzymes secretion (Garcia et al.,
2007), improving apparent metabolizable energy (Oguey and Bravo, 2015) and their

152
antioxidant and anti-inflammatory effects hence protecting the gut epithelium (Jamroz et al.,
2005; Pirgozliev et al., 2019).
Phyto 4 increased ADFI by 4% from grower to the end of the experiment without affecting
ADG resulting in lower FCR. This phytogenic showed reduced jejunum and ileum lengths
(Sarica et al., 2005; Hashemipour et al., 2016), which can explain that the higher feed intake
was not translated in a better gain. This lower gut length might result in a lesser digestion
time, which can also be correlated with an increased total SCFA production, an indicator of
bacterial carbohydrate fermentation (Bauer et al., 2019). This increased of total SCFA seems
to be the result of more butyrate production, which is known to favoring gut health (van der
Beek et al., 2017).
Phyto2 that is based on Yucca schidigera extract, a source of saponin, is a completely
different type of phytogenic in comparison of the three other tested compounds. It however
showed similar improvement of ADG and ADFI (8% vs C) more importantly in the grower
phase, resulting in a 5% heavier final body weight than C fed birds. Yucca schidigera extract
has successfully been used as feed additives in the poultry industry (Sahoo et al., 2015). It
enhances growth and productivity in broiler production (Sun et al., 2017, 2019; Sahoo et al.,
2015). Its saponins are proposed to reduce intestinal ammonia formation, thus aerial poultry
house pollution, an important health stress for broilers (Francis et al., 2002). Broilers exposed
to high concentrations of ammonia has been shown to have lower villus height and crypt
depth (Wei et al., 2012). This can explain the fact that this phytogenic was the only one
increasing villus length (26% vs C), partly explaining the positive effect of this AGP on
growth performance. Finally, effects related to lipid metabolism with Yucca are well known
(Cheeke et al., 2006). Sun et al. (2019) demonstrated that in the grower period, Yucca had a
positive antioxidative function of small intestine in broilers. Further studies are needed to
fully understand the effects of Yucca. schidigera extract supplementation on the intestinal
mucosa of the animals.

153
4.7. Conclusions
This study focuses on the effects of common alternatives on broiler chicken growth
performance and gut health. The alternatives examined herein were limited to some
commercial phytogenics composant. We found diverse results on the mode of actions of
those alternatives. Some inconsistent found may be attributed to the complexity of the
chemical composition of essential oil and the active comounds, dietary inclusion levels, and
utilization form of the feed additives applied. Further studies are needed to fully understand
the effects of phytogenic extract supplementation on the intestinal health of the broilers.
Acknowledgements
The authors thank the Department of Animal Science of Université Laval, Faculty of
Veterinary Medicine, University of Montreal, Westco Group who is one of the largest
companies in the poultry sector in Canada and the Tunisian government national scholarship
program for financial support.

154
4.8. References
AOAC, 2000. Official Methods of Analysis. 17th Edition, the Association of Official
Analytical Chemists, Gaithersburg, MD, USA. Methods 925.10, 65.17, 974.24,
992.16.
Bauer, B.W., Gangadoo, S., Bajagai, Y.S., Van, T. T. H., Moore, R. J., Stanley, D., 2019.
Oreganopowder reduces Streptococcus and increases SCFA concentration in a mixed
bacterial culture assay. PLoS ONE. 14, 12:e0216853.
Ben Arfa, A., Combes, S., Preziosi-Belloy, L., Gontard, N., Chalier, P., 2006. Antimicrobial
activity of carvacrol related to its chemical structure. Letters in Applied
Microbiology. 43, 149-154.
Burt, S., 2004. Essential oils: Their antibacterial properties and potential applications in
foods - A review. International Journal of Food Microbiology. 94, 223-253.
Castanon, J. I., 2007. History of the use of antibiotic as growth promoters in European poultry
feeds. Poultry Science. 86, 2466–2471.
Cheeke, P. R., Piacente, S., Oleszek, W., 2006. Anti-inflammatory and anti-arthritic effects
of Yucca schidigera: a review. J. Inflame.3, 1-7.
Cheng, G., Hao, H., Xie, S., Wang, X., Dai, M., Huang, L., Yuan, Z., 2014. Antibiotic
alternatives: the substitution of antibiotics in animal husbandry?. Frontiers in
microbiology. 5, 217-232.
Collett, S. R., 2012. Nutrition and wet litter problems in poultry. Animal Feed Science and
Technology. 173, 65-75.
Diarra, S. M., Malouin, F., 2014. Antibiotics in Canadian poultry productions and
anticipated alternatives. Frontiers in Microbiology. 5, 282.
Francis, G., Kerem, Z., Makkar, H.P., Becker, K., 2002. The biological action of saponins in
animal systems: a review. British Journal of Nutrition. 88,587–605.
Garcia, V., Catala-Gregori, P., Hernandez, F., Megias, M.D., Madrid, J., 2007. Effect of
formic acid and plant extracts on growth, nutrient digestibility, intestine mucosa
morphology and meat yield of broilers. Journal of Applied Poultry Research. 16, 555-
562.
Gaskins, H. R., Collier, C. T. Anderson, D. B., 2002. Antibiotics as growth promotants:
mode of action. Anim. Biotechnology. 13, 29–42.
Gurbuz, Y. and Ismael, I., 2016. Effect of peppermint and basil as feed additive on broiler
performance and carcass characteristics. Iranian Journal of Applied Animal Science.
6, 149-156.
Hamid, H., Zhao, L. H., Ma, G. Y., Li, W. X., Shi, H. Q., Zhang, J. Y., Ma, Q. G., 2019.
Evaluation of the overall impact of antibiotics growth promoters on broiler health and
productivity during the medication and withdrawal period. Poulry Science. 98, 3685-
3694.
Hashemi, S. R., and Davoodi, H., 2010. Phytogenics as new class of feed additive in poultry
industry. Journal of Animal and Veterinary Advances. 9, 2295–2304.
Hashemipour, H., Khaksar, V., Rubio, L. A., Veldkamp, T., Van Krimpen, M. M., 2016.
Effect of feed supplementation with a thymol plus carvacrol mixture, in combination
or not with an NSP-degrading enzyme, on productive and physiological parameters

155
of broilers fed on wheat-based diets. Animal Feed Science and Technology. 211, 117-
131.
Jamroz, D., Orda, I., Kamel, C., Wiliczkiewicz, A., Wertelecki, T., Skorupinska, I., 2003.
The influence of phytogenic extracts on performance, nutrient digestibility, carcass
characteristics, and gut microbial status in broiler chickens Animal Feed Science and
Technology. 12, 583–596.
Jamroz, D., Wiliczkiewicz, A., Wertelecki, T., Orda, J., Skorupińska, J., 2005. Use of active
substances of plant origin in chicken diets based on maize and locally grown cereals.
British Poultry Science. 46, 485-493.
Knarreborg, A., Lauridsen, C., Engberg, R. M., Jensen, S. K., 2004. Dietary antibiotic growth
promoters enhance the bioavailability of α-tocopheryl acetate in broilers by altering
lipid absorption. Journal of Nutrition. 134, 1487–1492.
Létourneau-Montminy, M.P., Messad, F., Medina, B., Gabarrou, J. F., 2019. Effects of a
specific blend of oleoresins of spices and essential oils on growth performance of
broilers: Insight from a meta-analysis approach. Poult. Sci Vol. 98, E-Suppl. 1, 659
Lin, J., 2014. Antibiotic growth promoters enhance animal production by targeting
intestinal bile salt hydrolase and its producers. Frontiers in Microbiology. 5, 33.
Marković, R., Šefer, D., Krsticv, M., Petrujkicv, B., 2009. Effect of different growth
promoters on broiler performance and gut morphology. Archivos de Medicina
Veterinaria. 41, 163-169.
Mehdi, Y., Létourneau-Montminy, M. P., Gaucher, M. L., Chorfi, Y., Suresh, G., Rouissi,
T., Brar, S. K., Côté, C., Ramirez, A. A., Godbout, S., 2018. Use of antibiotics in
broiler production: Global impacts and alternatives. Animal Nutrition. 4, 170–178.
Mitsch, P., Zitterl-Eglseer, K., Köhler, B., Gabler, C., Losa, R., Zimpernik, I., 2004. The
Effect of Two Different Blends of Essential Oil Components on the Proliferation of
Clostridium perfringens in the Intestines of Broiler Chickens. Poultry Science. 83,
669–675.
Niewold, T. A., 2007. The nonantibiotic anti-inflammatory effect of antimicrobial growth
promoters, the real mode of action? A hypothesis. Poultry Science. 86, 605–609.
Oguey et Bravo. PSA 2015. http://alliednutrition.com.www99.jnb2.host-h.net/wp-
content/uploads/2016/02/A4_Xtract_leaflet_proof.pdf. (Page consulté le 21 juin
2018).
Pirgozliev, V., Mansbridge, S. C., Rose, S. P., Mackenzie, A. M., Beccaccia, A., Karadas,
F., Bravo, D., 2019. Dietary essential oils improve feed efficiency and hepatic
antioxidant content of broiler chickens. Animal. 13, 502-508.
Rajkovich, S., Enders, A., Hanley, K., Hyland, C., Zimmerman, A.R., Lehmann, J., 2011.
Corn growth and nitrogen nutrition after additions of biocars with varying properties
to a temperate soil. Biology and Fertility of Soils. 48, 271-284.
Sahoo, S. P., Kaur, D., Sethi, A. P. S., Sharma, A., Chandra, M., 2015. Evaluation of Yucca
schidigera extract as feed additive on performance of broiler chicks in winter season.
Veterinary World. 8, 556-560.
Sarica, S., Ciftci, A., Demir, E., Kilinc, K., Yildirim, Y., 2005. Use of an antibiotic growth
promoter and two herbal natural feed additives with and without exogenous enzymes
in wheat based broiler diets. South African Journal of Animal Science. 35, 61–72.
Scheuer, J.P., Chalk, T.B., 1986. Clinical tests: Histology. Wolfe Medical Publ Ltd,
Netherlands.

156
Sun, D., Jin, X., Camerlink, I., Tong, M., Su, J., Zhao, F., Shi, B., 2019. Effects of Yucca
schidigera extract on growth performance and antioxidative function of small
intestine in broilers. Journal of animal physiology and animal nutrition. 103, 738-746.
Sun, D., Jin, X., Shi, B., Su, J., Tong, M., Yan, S., 2017. Dietary Yucca schidigera extract
improved growth performance and liver antioxidative function in broilers. Italian
Journal of Animal Science. 16, 677-684.
Van Der Wielen, P.W., Biesterveld, S., Notermans, S., Hofstra, H., Urlings, B.A., van
Knapen, F., 2000. Role of Volatile Fatty Acids in Development of the Cecal
Microflora in Broiler Chickens during Growth. Applied and Environmental
Microbiology. 66, 2536–2540.
Van der Beek, C. M, Dejong, C. H., Troost, C F. J., Masclee, A. M., Lenaerts, k., 2017. Role
of short-chain fatty acids in colonic inflammation, carcinogenesis, and mucosal
protection and healing. Nutrition Reviews. 75, 286–305.
Wei, F.X., Xu, B., Hu, X.F., Li, S.Y., Liu, F.Z., Sun, Q.Y., 2012. The effect of ammonia and
humidity in poultry houses on intestinal morphology and function of broilers. Journal
of Animal and Veterinary Advances. 11, 3641–46.
Windisch, W., Schedle, K., Plitzner, C., Kroismayer, A., 2008. Use of phytogenetic products
as feed additives for swine and poultry. J.ournal of Animal Science. 86, E140-E148.
Yang, Y., Ashworth, A. J., Cook, K., Willett, C., Upadhyay, A., Owens, P., Moore, P. A.,
2019. Review of antibiotic resistance, ecology, dissemination, and mitigation in US
broiler poultry systems. Frontiers in Microbiology. 10, 2639-2649.
Yatoo, M. I., Dimri, U., Gopalakrishnan, A., Karthik, K., Gopi, M., Khandia, R., Saminathan,
M., Saxena, A., Alagawany, M., Farag, M.R., Munjal, A., Dhama, K., 2017.
Beneficial health applications and medicinal values of Pedicularis plants: a review.
Biomed Pharmacother. 95, 1301–1313.

157
Chapitre 5 Discussion générale et conclusions

158
5.1. Contexte de la thèse
L’utilisation des antibiotiques en élevage avicole, représente un atout puisqu’elle contribue
à l’amélioration des performances de croissance des oiseaux et à la prévention des maladies
(Les Producteurs de poulets du Canada, 2018). Il est aujourd’hui démontré que les
antibiotiques facteurs de croissance (AFC) présentent un risque pour la sécurité alimentaire
et la santé humaine en favorisant la résistance de certains micro-organismes à certaines
familles d’antibiotiques (WHO, 2014). Leur retrait est par contre associé à des conséquences
importantes pour la production (Les Producteurs de poulets du Canada, 2018). Au Canada,
les exigences rattachées à une appellation « sans antibiotiques » pour la viande de volaille
sont d’autant plus strictes (Santé Canada, 2017). C’est dans ce contexte que s’est déroulée
ma thèse en collaboration entre la Chaire de recherche sur les stratégies alternatives
d’alimentation des porcs et des volailles de l’Université Laval de Québec, la ferme
expérimentale du Centre de recherche en sciences animales de Deschambault, l’entreprise
Agri-Marché, l’entreprise Westco ainsi que la Faculté de Médecine vétérinaire de
l’Université de Montréal.
L'objectif principal du présent travail de thèse était de développer des stratégies alimentaires
permettant de maintenir les performances de croissance dans un contexte de retrait des
antibiotiques facteurs de croissance (AFC). Pour ce faire, des indicateurs de santé digestive
devaient être développés pour évaluer l’impact des stratégies testées. Ensuite, compte tenu
de la complexité du sujet avec une multitude d’alternatives aux antibiotiques facteurs de
croissance disponibles, l’approche par méta-analyse a été préconisée dans un premier temps.
Ceci a permis d’identifier les alternatives les plus étudiées et prometteuses de façon objective.
Enfin, en lien avec le travail de revue de littérature quantitative et les différentes stratégies
déjà utilisées sur le terrain au Québec, deux essais ont été réalisés pour développer les
indicateurs et tester des alternatives aux AFC.

159
Dans cette section de la thèse, les résultats obtenus sont discutés en trois étapes. La première
étape est basée sur la discussion des intérêts, des résultats et des limites de l’étude par méta-
analyse compte tenu de la multitude d’alternatives disponibles et de la variabilité des
réponses obtenues. La deuxième étape se décline en deux sous parties. Une première partie
pour comparer les différents traitements et leur effet sur les indicateurs des performances de
croissance et de santé digestive mesurés dans les deux essais et une deuxième partie pour
étudier les corrélations entre les indicateurs, discuter leurs utilités et limites. Enfin, la
contribution de la thèse à la diminution souhaitée dans l’utilisation des AFC sera discutée.
5.2. Intérêt, résultats et limite de l’outil de méta-analyse
5.2.1. Intérêt de cette méta-analyse
Pour tirer des conclusions pertinentes sur les effets des alternatives aux AFC sur les
performances de croissance et les considérées comme une connaissance utilisable et
généralisable, il n’est pas possible de se baser sur une seule expérimentation puisque d’une
part, elle est limitée par sa taille empêchant l’obtention de la précision sur les effets et les
relations observés (Sauvant, 1992) et d’autre part, sa dépendance au milieu expérimental est
très élevée (Sauvant et al., 2005). Compte tenu des multitudes alternatives disponibles et la
variabilité des réponses obtenues dans les travaux antérieurs, l’outil de méta-analyse
apparaissant tout indiqué. En effet, la méta-analyse est un outil qui permet de récolter des
données de façon exhaustive de plusieurs travaux déjà publiés (Sauvant et al., 2008) afin de
les étudier statistiquement. L’étude des facteurs de variations affectant différents paramètres
Y étudiés permet de tester les hypothèses avec plus de précision (puissance statistique).
La première phase de la thèse consistait donc à une étude par méta-analyse comme proposée
par Sauvant et al., (2005). Une première recherche bibliographique et une discussion avec le
groupe Agri-Marché a permis de diriger les recherches sur certains types d’alternatives,

160
soient les acides organiques, les prébiotiques, les probiotiques, et les huiles essentielles qui
ont été organisés en sous-bases. D’une façon exhaustive, toutes les études publiées sur ces
alternatives aux AFC entre 2000 et 2017 ont été recherchées dans une base de données.
Toutes les études contenaient des informations sur les effets de ces alternatives sur les
performances de croissance, la race de poulet, la teneur en protéine brute et l’énergie
métabolisable de régimes alimentaires et le challenge sanitaire. Ceci a permis de faire un état
des lieux, d’identifier les alternatives aux AFC les plus étudiées et prometteuses de façon
objective. Ainsi, une base de 79 articles scientifiques contenant 282 essais divisés en quatre
alternatives aux AFC les plus utilisés, à savoir l’acide butyrique, les MOS, les probiotiques
(Bacillus subtillis) et l’huile essentielle d’origan. Incluant des données de performance de
croissancede, la race de poulet, la teneur en protéine brute et l’énergie métabolisable de
régimes alimentaires et le challenge sanitaire Le regroupement de plusieurs expérimentations
a permis d’établir des lois générales de prédiction des performances de croissance (gain
moyen quotidien de poids (GMQ), consommation moyenne journalière (CMJ) et indice de
conversion alimentaire (IC)) à partir de la dose d’alternative, de % calculé de l’énergie
métabolisable de la ration de base par rapport aux besoins de la race de poulet et de leurs
interactions (Chapitre 1).
La distribution des données (uniformité, variations intra et interétudes), la relation entre les
variables dépendantes (GMQ, CMJ et IC) et les variables indépendantes, dont la dose des
alternatives, le % calculé de la protéine brute et de l’énergie métabolisable de la ration de
base par rapport aux besoins de la race de poulet, la race de poulet et le challenge sanitaire
(bactéries pathogènes administrées par voie orale) ont été étudiés graphiquement. Cette
visualisation graphique a permis la détection et la suppression des données aberrantes ainsi
que la définition du modèle statistique à savoir ; les variables susceptibles d'influencer les
performances de croissance ainsi que la définition de la nature des relations entre ces
dernières (linéaire/ quadratique).

161
Le choix des prédicteurs était basé sur l’étude des corrélations entre les variables testées en
interétudes (coefficient de Pearson) et en intra études par des modèles GLM avec inclusion
de l’effet publication. L’objectif principal de cette étude du méta-dispositif était
l’identification de prédicteurs pour établir la loi de réponse. Pour le choix du modèle, l’effet
étude a été considéré comme étant aléatoire (Sauvant et al., 2008; St-Pierre, 2001). La
dernière étape de cette étude était la vérification des modèles établis par l’intermédiaire d’une
validation interne (cross validation).
5.2.2. Résultats de cette méta-analyse
Les résultats de cette méta-analyse ont montré qu’il n’y aucun effet sur la CJM, alors que le
GMQ a augmenté de manière linéaire et quadratique (P <0,05) avec la dose d'acide
butyrique, de MOS et d'huile essentielle d’origan et linéairement avec Bacillus subtilis
jusqu'à respectivement 7%, 8%, 7% et 9%. L'amélioration de l’IC (Figure 5-1) était de 2,4
%, 8 %, 7 % et 3 % pour l'acide butyrique, le MOS, l'huile essentielle d'origan et Bacillus
subtilis respectivement. Les détails suivront pour chaque alternative aux AFC.

162
Figure 5-1 Effet d’amélioration (%) des alternatives aux AFC étudié sur l’indice de
conversion alimentaire.
Pour l’acide butyrique, une diminution linéaire (P = 0,003) de l’IC avec un effet positif du
% de l’énergie métabolisable par rapport aux besoins (P = 0,002). Ce dernier a eu un effet
sur l’ordonnée à l’origine, indiquant qu'un contenu énergétique plus élevé améliore l’IC (P
= 0,002) et le GMQ (P = 0,05). En comparant avec d’autres études récemment effectuées, il
a été démontré que l’acide butyrique améliore le GMQ et l’IC, en moyenne de 3,12 % et 2,92
% respectivement (Kim et al., 2015). Le MOS a exercé une amélioration de l’IC avec une
diminution linéaire (P <0,001) et quadratique (P = 0,0003) à 99 % de l’énergie métabolisable
par rapport aux besoins. Le pourcentage de l’énergie métabolisable (Figure 5-2) par rapport
aux besoins a eu un effet sur l’ordonnée à l'origine, montrant qu’une énergie plus élevée
améliorait l’IC (P = 0,05). Ces résultats confirment ceux obtenus dans des méta-analyses
récentes incluant 44 essais menés avec un MOS commercial (Bio-MOS, Alltech Inc.) publiés
entre 1993 et 2003 (Hooge, 2004) et incluant 82 essais entre 1997 et 2003 (Rosen, 2007) et
montrant une amélioration du GMQ de l’ordre de +1,61 % et 1,48 % respectivement et une
réduction de l’IC de l’ordre de 1,99 et 2,11 % respectivement. La présente étude suggère une

163
concentration optimale de MOS égale à 2,5 g/ kg d'aliments pour poulets de chair, ce qui est
proche des estimations précédentes d'environ 2 g / kg (Rosen, 2007; Yang et al., 2007). De
même, l’IC a également été diminué de manière linéaire (P <0,001) et quadratique (P
<0,001) avec l'augmentation des doses d’huile essentielle d’origan. La dose optimale d’huile
essentielle d’origan pour les poulets de chair a été estimée à 300 mg / kg du produit mélangé
dans le régime alimentaire dans notre étude, ce qui est proche des 500 mg / kg proposés
précédemment par Alagawany et al. (2018). L’effet de Bacillus subtillus sur le GMQ et l’IC
était juste linéaire, pas d’effet noté du % de l’énergie métabolisable par rapport aux besoins.
Aux doses de 106 à 108 cfu / g, l’IC a diminué de 3% alors que le GMQ a augmenté de 9%.
Figure 5-2 Effet du pourcentage de l’énergie métabolisable sur l’indice de la conversion
alimentaire.
En parallèle avec ce travail, dans certaines études des traitements avec et sans antibiotiques
facteurs de croissance étaient présents et ont été rassemblés dans une sou-base. L’étude de
cette dernière n’était pas simple avec différents antibiotiques, parfois interdits depuis
longtemps, non-utilisés au Canada ou avec des dosages très différents. Après discussion avec
l’équipe, une base de données a été conçue et étudiée par méta-analyse. Cette dernières a
montré que dans 64 % des cas il n’y avait pas d’effet des AFC, alors que dans le reste des cas
un effet positif sur l’IC était noté avec l’addition d’AFC (Figure 5-3). Dans le but de bien
comprendre l’effet marqué des AFC, qui demeure le traitement à remplacer, des antibiotiques
1,5
1,6
1,7
1,8
1,9
0 2 4
Ind
ice
de
co
nve
rsio
n
alim
enta
ire
Dose des MOS (g/kg)
Énergie 99%
Énergie 95%
Énergie 92%
-7,2%
-7,5%
-8,0%

164
ont été ajouté dans un des traitements du deuxième essai expérimental sur terrain dans le
cadre de cette thèse (Chapitre 5).
Figure 5-3 Effet des antibiotiques facteurs de croissance (AFC) (C- = traitement contrôle
sans AFC et C+ : traitement avec AFC) sur l’indice de la conversion alimentaire.
5.2.3. Limites de cette méta-analyse
Comme indiqué dans la figure 5-1, les doses des alternatives aux AFC étudiées sont variées
et spécifique au type d’alternative et on ne peut pas les uniformiser dans une seule unité sur
une même échelle. Ainsi, leurs effets en fonction de la dose restent toujours liés à chaque
alternative et on n’a pas pu les comparer pour déterminer la meilleure alternative aux AFC
et faire une conclusion générale pertinente sur le choix de l’alternative. En effet, il est
important de noter que l’outil de méta-analyse demande énormément de temps avec de
multiples itérations, certainement une limite de cette approche. Autre limite détectée dans
notre étude, était la base de données des huiles essentielles. Les huiles essentielles sont
généralement des mélanges avec d’autres composés comme des épices et des oléorésines.
Dans les publications, nous avions très rarement le taux d’incorporation de chacun des

165
composés et parfois que le nom commercial du produit, limitant notre capacité à différencier
les types de produits.
D'autres complications surviennent parce que les huiles essentielles peuvent varier
considérablement en ce qui concerne l'origine botanique de la plante et son mode de
traitement et de fabrication (Windisch et al., 2008). Ce fût un gros défi qui a pris beaucoup
de temps afin de tester différentes méthodes et hypothèses. Conséquemment, beaucoup de
travail reste à faire en ce qui concerne les huiles essentielles et est actuellement en cours avec
un partenariat avec un producteur de ce type de produit qui a beaucoup à nous apporter. Une
autre limite fut rencontrée dans la base de données des acides organiques. Il est déjà bien
rapporté dans la littérature que ses effets dépendent de la forme chimique de l'acide avec des
valeurs de pKa différentes (Ahsan et al., 2016) et de la présentation de la molécule (e.x. :
l'encapsulation; Khan et Iqbal, 2016). Malheureusement, ces informations n'étaient pas
disponibles dans toutes les études et n'ont pas pu être prises en compte dans les modèles
statistiques.
De plus, le sujet des alternatives aux AFC concernent des additifs nutritionnels probablement
souvent financés par les compagnies elle-même induisant un biais. En effet, il est fort
possible que les études utilisées dans la méta-analyse soit non représentatif de l’ensemble des
données utilisé dans la résolution d’une question de recherche. Dans ces bases de données
nous faisions également face à un manque de variabilité intra-étude en raison des designs
expérimentaux qui était souvent des doses réponses aux alternatives aux AFC dans une
condition donnée ce qui constitue une autre contrainte de la méta-analyse (Sauvant et al.,
2008). En effet, ceci limitait l’étude des effets de d’autres facteurs que la dose. En fait, une
grande diversité au niveau des méthodologies existe entre les publications scientifiques, ce
qui a une influence sur les données, tels que l’âge, la lignée génétique, la teneur en énergie

166
métabolisable et la teneur en protéines brutes qui n’étaient pas significative probablement en
raison des faibles variations dans une même expérience (figure 5-4). Néanmoins, la prise en
compte de l’effet étude a été pris en compte et corrigeait pour tous ces facteurs en même
temps.
Figure 5-4 Distribution du pourcentage de la protéine brute par rapport au besoin de la race,
de la race et du challenge sanitaire intrapublication dans la base de données des Mannan-
oligosaccharides.
5.3. Résultats pertinents des deux essais pratiques du projet
5.3.1. Effet des traitements sur les indicateurs mesurés dans les deux essais
Pour tenter d’avoir des conclusions générales sur les effets des alternatives aux AFC, une
base de données rassemblant les données communes des deux essais expérimentaux a été

167
créée en utilisant la procédure MIXED et le test Tukey Kramer (SAS 9.4, SAS, NC, USA).
L’effet essai (1 ou 2) a été inclus dans le modèle comme aléatoire pour tenter de prendre en
compte le fait que les conditions environnementale (pas au même moment) et les aliments
n’étaient pas les mêmes entre les deux essais. Une différence a été considérée comme
significative à P <0,05 et P <0,10 a été considérée comme une tendance.
Dans un premier temps, il est facile de regarder l’évolution du poids vif des poulets au cours
des deux essais (Figure 5-5). En effet, c’est le critère auquel les éleveurs de poulets font
référence pour envoyer leurs animaux à l’abattoir. Bien que les régimes et les nutriments
n’étaient pas les mêmes, l’avoine nue se démarque. Durant la phase de démarrage (Phase 1)
et la phase de finition (Phase 3) les poulets nourris avec l’avoine nue ont un poids
significativement plus élevé que les poulets de tous les autres traitements, témoin avec
antibiotique compris. Pendant la phase de croissance (Phase 2), l’incorporation de l’avoine
nue donne les poulets les plus lourds, mais ce résultat n’est plus significativement différent
de ceux nourris avec Bacillus Subtilis et ceux du traitement Témoin. Toutefois, ce résultat
seul n’est pas suffisant. Les calculs du gain moyen quotidien (GMQ), de la consommation
moyenne journalière (CMJ), de l’indice de conversion alimentaire (IC) et les mesures des
indicateurs de santé digestive (profondeur des cryptes et longueur des villosités, pH et
humidité de la litière et production des acides gras à chaînes courtes caecaux) peuvent éclairer
ces différences et donner de nouveaux objectifs pour choisir la meilleure alternative.

168
Figure 5-5 Évolution du poids vif des poulets pour les différents traitements sur la période
d’essai par phase de croissance (nbr/traitement = 450).
Les figures allant de 5-6 à 5-9, représentent les différents résultats liés aux performances de
croissance et à la mortalité en fonction du type de traitement sur les trois périodes de
croissance des deux expériences. Pour les trois périodes, le type de traitement a un effet
significatif sur le GMQ, la CJM, l’IC et la mortalité.

169
Figure 5-6 Gain moyen quotidien pour les différents traitements sur la période d’essai par
phase de croissance (nbr/traitement= 450).
L’avoine nue semble être l’alternative avec lequel on a le GMQ le plus élevé comparant aux
autres traitements durant les deux phases de croissance, ce qui a mené à un IC plus faible. En
revanche, une disparition des effets positifs du régime d'avoine nue à la fin de la phase de
finition contre la supplémentation avec l’acide butyrique sous forme protégée. Cet effet se
traduit par la baisse du pH de la litière (Figure 5-10). La réduction du pH de la litière (Figure
5-10) avec un régime d'avoine nue durant la phase de finition est probablement le résultat
d'une fermentation plus élevée du β-glucane d'avoine (Daou et Zhang, 2012). Les résultats
actuels ont montré un autre effet positif de l'avoine nue sur la santé des poulets de chair. La
proportion de butyrate a augmenté durant la phase de finition avec le régime d’avoine nue
montrant un passage au fil du temps à une microflore plus butyrogène dans les caeca comme
précédemment observé par Gonzalez-Ortiz et al. 2017. Le butyrate est connu pour favoriser
la santé intestinale (van der Beek et al., 2017).
b

170
Figure 5-7 La consommation moyenne journalière pour les différents traitements sur la
période d’essai par phase de croissance (nbr/traitement = 450).
Xtract-Instant aussi réduit le pH de la litière durant la phase de démarrage (Figure 5-10). Le
faible pH de la litière est un bon indicateur de l'état de santé des oiseaux. Hardin et Roney,
(1989) ont montré que la réduction du pH de la litière entraînait un déclin des populations
microbiennes, y compris E. coli, Salmonella et Clostridium. De plus, Payne et al. (2007) ont
montré qu’un pH bas entraînait la réduction la plus rapide dans les populations de Salmonella
par rapport à un pH plus élevé.
Micro-Aid était la seule alternative à augmenter la longueur des villosités et la profondeur
des cryptes réduisant ainsi leur rapport (Tableau 5-1). Les saponines qu'il contient sont
proposées pour augmenter la perméabilité des cellules de la muqueuse intestinale in vitro,
inhibant ainsi le transport actif et facilitant l'absorption intestinale des nutriment (Johnson et
al., 1986). Seeman et al. (1973) ont démontré qu’à des niveaux d’inclusion optimaux de
saponines, il avait une augmentation de l'absorption des nutriments par l'intestin grâce à
l’augmentation de diamètre des villosités de 40 à 50 Ao, ce qui rendrait celles-ci perméables

171
aux grosses molécules comme la ferritine; et peut être responsable d’un meilleur taux de
croissance des villosités (Seeman et al., 1973).
Figure 5-8 L’indice de consommation pour les différents traitements sur la période d’essai
par phase de croissance (nbr/traitement = 450).
Figure 5-9 Taux de mortalité pour les différents traitements sur la période d’essai par phase
de croissance (nbr/traitement = 450).

172
En revanche, le phytek-P est l’alternative qui agit négativement sur l’IC et le GMQ et
augmente le pH de la litière durant la phase de finition. Il augmente la production de AGCC
total durant la phase de croissance (Tableau 5-1), ce qui est un indicateur de la fermentation
des glucides par les bactéries (Bauer et al., 2019) et produit moins d’acide butyrique.
Figure 5-10 Valeur de pH de la litière pour les différents traitements sur la période d’essai
par phase de croissance (nbr/traitement = 450).
L'effet positif du régime à base d'avoine nue pourrait être dû à la réduction du tourteau de
soja et donc à la réduction de la teneur en facteurs antinutritionnels. Le β-glucane de l’avoine
nue se comporte comme un prébiotique naturel: un ingrédient alimentaire non digestible qui
affecte l'hôte en stimulant sélectivement la croissance et / ou l'activité d'une souche ou d'un
nombre limité de souches bactériennes dans le côlon et améliore ainsi la santé de l'hôte
(Gibson et Roberfroid 1995). Ces résultats indiquent que l'avoine nue pourrait être incorporée
dans les régimes de poulets de chair de l’ordre de 20 à 30% sans effets négatifs. Des études
antérieures ont montré que l'avoine nue pourrait remplacer d'autres céréales dans
l'alimentation des poulets de chair (Kamińska., 2003; Osek et al., 2003, Szymczyk et al.,
2005). Pour l’effet de l’acide butyrique, certaines études ont montré que l’IC était augmenté

173
dans les régimes supplémentés en butyrate pendant la phase de croissance (Moquet, 2018;
Gonzalez-Ortiz et al., 2019) et que les performances de croissance étaient améliorées dans
d'autres (Abd El-Hakim et al., 2009; Raza et al., 2017). Parce que le butyrate utilisé dans
notre étude était enrobé de graisse végétale, donc il est libéré lentement (Chamba et al., 2014;
Sikandar et al., 2017), cela pourrait expliquer pourquoi le butyrate utilisé dans cette étude
était plus efficace vers la fin de l’expérience. Pour le taux d’humidité de la litière (Figure 5-
11), pas de différences significatives entre les traitements durant la phase de démarrage.
Cependant, le régime avec le Bacillus subtillus était le seul à réduire l'humidité de la litière
(Figure 5-11), ce qui est souvent associé à un meilleur équilibre des nutriments ou à une
diminution de la dysbactériose (Collett, 2012). En effet, il a été démontré que des litières plus
humides renfermaient des charges microbiennes significativement plus élevées (Gaucher.,
2016).
Figure 5-11Taux d’humidité de la litière pour les différents traitements sur la période d’essai
par phase de croissance (nbr/traitement = 450).

174
Dans le même ordre d’idée, l’action bénéfique attribuable aux produits commerciaux à base
d’huiles essentielles (Oleo-Poultry, Micro-Aid, Xtract-Instant et phytek-P) évaluée durant
notre étude est difficile à évaluer vue leur composition complexe. Cela pourrait être dû à
plusieurs facteurs qui influencent leur effet sur les performances tels que: la teneur variable
des huiles essentielles dans les produits (Jang et al., 2004), l’âge de la plante à la récolte, la
zone géographique de la plante, la partie de la plante utilisée pour extraire le produit
(Windisch et al., 2006). De nombreuses études ont documenté le pouvoir antibactérien des
huiles essentielles (Ben Arfa et al., 2006; Brenes et Roura, 2010), mais plus spécifiquement,
il a été démontré que des mélanges d’huiles essentielles, telles que le thymol, et le carvacrol,
des composantes actives retrouvées dans les produits commerciaux utilisés dans notre étude,
agiraient de diverses façons pour exercer leur impact positif (Oguey and Bravo, 2015). Entre
autres, les huiles essentielles et surtout leurs composantes actives auraient pu stimuler la
libération de sécrétions digestives (Garcia et al., 2007) chez les poulets. Tout ceci aurait eu
comme effet l’optimisation du processus d’assimilation des nutriments chez les pouletschez
les poulets. Tout ceci aurait eu comme effet l’optimisation du processus d’assimilation des
nutriments chez les poulets.

175
Table 5-1 Concentration des acides gras à chaîne courte totaux (mmol); de l’acétate (%), propionate (%), butyrate (%) dans les
caeca de poulets de chair pendant les trois phases de croissance et paramètres histomorphologiques iléaux au jour 28 (nbr/traitement
= 450).
Items* T1 T2 T3 T4 T5 T6 T7 T8 T9 T10 T11 SEM P
Phase de démarrage
AGCCT 67,83
d
75
cd
71,81
cd
78,51
cd
80,94
c
111,7
ab
115,8
ab
110,2
b
125,7
a
120,1
a
113,7
ab
7,55 <.0001
Acétate 75,73 74,31 75,68 74,37 72,83 76,46 76,46 76,05 72,65 73,5 74,39 1,04 0.1242
Propionate 4,196 4,451 4,307 4,596 5,127 4,705 3,725 5,832 4,971 3,756 4,42 0,58 0.5081
Butyrate 17,95 18,97 17,82 18,48 19,53 16,11 17,58 15,84 19,89 20,6 19,14 1,09 0.0873
Phase de croissance
LV 294,6
c
305,5
bc
336,5
bc
325,3
bc
317,1
bc
397,7
ab
358,4
b
321,6
bc
450,5
a
299,2
c
315,2
bc
21,5 <.0001
PC 43,12
ab
43,1
ab
43,05 ab 42,89
ab
39,45
b
59,94
ab
58,81
ab
58,75
ab
62,56
a
58,39
ab
57,72
ab
5,14 0.0002
LV/PC 8,94
bc
9,25
bc
10,46 ab 10,38
b
11,16
a
6,71
bc
8,74
bc
7,06
bc
6,46
C
7,11
bc
8,15
bc
1,21 0.0182
AGCCT 42,87
c
55,89
bc
53,11 bc 56,21
bc
55,81
bc
65,28
ab
67,01
ab
42,65
c
76,66
ab
88,75
ab
99,76
a
9,06 <.0001
Acétate 75,61
ab
77,79
ab
76,13 ab 79,09
a
73,58
bc
69,65
c
73,08
bc
73,25
bc
75,75
ab
76,36
ab
74,84
ab
1,12 <.0001
Propionate 6,416 6,881 6,781 5,617 5,251 7,681 7,662 7,261 6,925
9
6,393
5
6,769 0,71 0.2594
Butyrate 13,11
bc
10,39
c
13,21 bc 11,24
bc
17,16
ab
18,06
a
14,84
ab
13,31
bc
12,97
bc
13,76
b
14,71
ab
1,52 0.0017

176
*AGCCT: acides gras à chaîne courte totaux (mmol); LV : longueur des villosités (μm); PC : profondeur des cryptes (μm); LV/PC : rapport de longueur des
villosités sur profondeur des cryptes; T1 : Controle1; T2 : T1+Butyrate; T3 : T1+Mos; T4 : T1+Bs; T5 : T1+avoine nue; T6 : antibiotique; T7 : Controle2;
T8 : T7+ Pleo-Poultry; T9 : T7+ Micro-Aid; T10 : T7+ Xtarct-Instant; T11 : T7+ Phytek-P.
Phase de finition
AGCCT 99,05
b
108,7 ab 127,7
a
99,63
b
83,53
bc
81,52
bc
87,91
bc
63,91
c
72,14
bc
82,52
bc
105,1
ab
11,1 0.0060
Acétate 75,54 74,25 75,78 74,37 72,95 72,31 72,97 73,73 74,21 72,67 73,05 1,08 0.3225
Propionate 4,085
c
4,579
c
4,541
c
4,596
c
4,593
c
7,171
ab
7,094
b
7,115
ab
7,086
ab
7,855
a
7,477
ab
0,61 <.0001
Butyrate 18,12
ab
18,58
ab
17,38
ab
18,42
ab
20,07
a
16,05
b
16,58
ab
15,09
bc
15,06
bc
15,39
bc
14,48
c
1,26 0.0318

177
5.3.2. Comparaison des effets des alternatives aux antibiotiques facteur de croissance sur
les performances de croissance entre les essaies pratique et la méta-analyse
Ces résultats sont prometteurs, car ils étudient les différents effets des alternatives aux AFC
disponibles présentement sur le marché sur les performances de croissances du poulet de
chair et ses caractéristiques morphologiques intestinales. Par comparaison à notre méta-
analyse (Tableau 5-2), les résultats des deux essais ont la même allure d’amélioration pour
l’acide butyrique. Pour les résultats de MOS, ils se concordent en termes de GMQ mais pas
d’effet sur l’IC. En revanche, l’effet de Bacillus subtillus était à l’inverse. Pour les produits
photogéniques testés, ils augmentaient le GMQ et la CMJ induisant une augmentation dans
l’IC. L’une des hypothèses des performances réduites chez les poulets recevant la
supplémentation avec Bacillus subtilis est le déficit énergétique et / ou protéique dans notre
essai (96 % des besoins). Ce déficit aurait pu induire un état inflammatoire chez les poulets.
Cela pourrait avoir conduit à une dysbiose se traduisant en une diminution de l'absorption en
raison d'une altération de la microflore intestinale et de l'intégrité de la muqueuse intestinale
(Suez et al., 2018) ou à une utilisation d'énergie pour l'augmentation de la réponse
immunitaire (Lei et al., 2015). D'un point de vue nutritionnel, des substrats tels que les acides
aminés et l'énergie sont nécessaires pour soutenir le système immunitaire, les acides aminés
et le déficit énergétique étant l'une des principales causes d'immunodéficience (Ruth et al.,
2013; Field et al., 2002). De plus, les systèmes immunitaires innés et adaptatifs dépendent
fortement d'une disponibilité adéquate d'acides aminés (Kim et al., 2007). Il est à notre que
dans la base de données des probiotiques de tels effets négatifs n’étaient pas rapportés bien
qu’un biais puisse être suspecté.

178
Table 5-2 Comparaison des effets des alternatives aux AFC sur les performances de
croissance entre les essaies pratique et la méta-analyse.
Effet sur Essaies pratiques (0-35j) Méta-analyse (Rouissi et al., 2020)
Acide butyrique
GMQ (g/j) +2.2 +7
CMJ (g/j) NS NS
IC -2.1 -2.4
MOS
GMQ (g/j) +3 +8
CMJ (g/j) +2 NS
IC NS -8
Bacillus Subtilis
GMQ (g/j) -4 +9
CMJ (g/j) NS NS
IC +4 -3
Huiles essentielles
GMQ (g/j) +3 +7
CMJ (g/j) +4.7 NS
IC +2.8 -4
5.3.3. Intérêts des indicateurs étudiés, limites et voies d’évolution
5.3.3.1. Performances de croissance
D’une manière générale, les variables liées aux performances de croissance, telles que le
GMQ, le poids vif final, la CMJ et l’IC semblent être corrélées entre elles (exp : pour la
totalité de la période expérimentale). Ceci semble logique puisque la CMJ affecte directement
le GMQ (r=0.88; P< 0.0001). De plus, le GMQ est corrélé négativement à l’IC (r= -0.70;
P< 0.0001), mais c’est un sens positif, rappelons qu’un indice de consommation bas est une
bonne chose : cela veut dire que les poulets prennent beaucoup de poids en mangeant peu.

179
5.3.3.2. Indicateurs de la santé digestive
D’une façon générale, les performances de croissance (GMQ, CMJ et IC) qui sont les
indicateurs les plus importants sur le terrain ne corrèlent pas avec les paramètres
histologiques intestinaux (la longueur des villosités, les profondeurs des cryptes) et la
production des AGCC. Ce qui contredit des recherches antérieures, qui montrent des relations
entre les paramètres histologiques intestinaux, le type d’aliment consommé et la CMJ
(Mathlouti et al., 2002; Engberg et al., 2004; den Besten et al., 2013).
Le type d’aliment consommé et la CMJ auront un effet sur la production des AGCC
puisqu’ils sont les produits finaux de la fermentation des fibres alimentaires par la flore
microbienne intestinale. On sait que la flore bactérienne intestinale produit des acides à
chaîne courte et non ramifiée (acétate, propionate, butyrate) ou ramifiée (valérate,
isovalérate, isobutyrate). C’est pour ça qu’on compte faire l’analyse de profil bactérien dans
nos deux essaies expérimentaux afin de mieux comprendre les résultats des AGCC, puisque
ces indicateurs et bien signe d’une modification de la flore intestinale. Le type d’acide et leur
quantité dépendent de deux facteurs essentiels : a) la nature de la flore (Figure 12). Lucke et
al. (2018) ont montré 15 corrélations significatives entre la concentration des acides gras à
chaîne courte et ramifiée avec les genres de bactéries dans le caecum, et b) des substrats
disponibles (Gabriel et al., 2005).

180
Figure 5-12 Corrélation entre la concentration des acides gras à chaîne courte et ramifiée
avec les genres de bactéries dans le caecum de poulets nourris (10 poulets/ traitement) avec
des régimes alimentaires contenant des niveaux croissants de désoxynivalénol (0, 2,5, 5 ou
10 mg / kg) et avec ou sans ajout des lipopolysaccharides (LPS), 1 jour avant l’abattage. Les
couleurs font référence au degré de corrélation (Lucke et al., 2018).
Par ailleurs, la CMJ est corrélée positivement avec le pH de la litière (Figure 5-13). Les
poulets qui consomment plus semblent donc avoir rejeté davantage de fientes que les autres.
Ce résultat concorde avec l’IC obtenu sur ces poulets qui était le plus élevé. Ces poulets ont
donc davantage mangé et produit plus de fientes. Il est possible aussi que ces poulets
consomment plus d’eau. Les quantités importantes de fientes et d’eau pourraient expliquer
l’augmentation du pH dans ces litières (Olivère, 2010). On observe une corrélation positive
entre l’humidité de la litière et son pH (Figure 5-14). Ils augmentent dans le même sens. À
noter que la litière constitue un paramètre important à prendre en compte en ce qui concerne

181
la santé des poulets, puisqu’ ils sont toujours en contact avec cette matrice (Bilgili et al.,
2009) et son humidité est un gros enjeu. Cette corrélation paraît logique, puisqu’une litière
humide entraîne une prolifération bactérienne responsable de la dégradation des fientes et
donc d’une augmentation de la teneur en ammoniaque (Thomke et Elwinger 1998) qui a un
pH alcalin (Martland, 1986).
Figure 5-13 Relation entre la CMJ et le pH de la litière (r=0.9; R2= 78.9%; P< 0.0001).
Dans un souci d’exactitude de la réalité terrain, il serait intéressant de suivre le
développement des propriétés de la litière, par le contrôle de ces indicateurs au fur et à mesure
de la durée expérimentale dans chaque élevage de poulets de chair. L’étude d’autres
caractéristiques physico-chimiques, comme le dosage d’azote total et ammoniacal, la mesure
des températures des litières et la présence de champignons et de bactéries seraient
intéressants pour une meilleure explication des résultats.

182
Figure 5-14 Relation entre l’humidité et le pH de la litière (r=0.63; R2= 65.3%; P< 0.0001).
Il y a une tendance pour que le butyrate augmente la longueur des villosités (P=0.07). Sinon,
il n’y a pas d’autre corrélation entre les autres AGCC et les autres indicateurs. Le butyrate
semble jouer un rôle dans le développement de l'épithélium intestinal (Leeson et al., 2005)
et est censé être important pour le développement normal des cellules épithéliales (Pryde et
al., 2002). Les effets du butyrate sur la santé intestinale ont déjà été décrits en détail dans de
nombreuses recherches (Guilloteau et al., 2010; Canani et al., 2011). Le butyrate intestinal
augmente le nombre des Lactobacillus, qui renforce la barrière muqueuse en augmentant la
production de mucine, qui est un élément important de défense contre les agents pathogènes
intestinaux (Chang et al., 2019). La concentration en butyrate est corrélée positivement avec
celle de l’acétate dans le caecum des poulets (Figure 5-15). Il a été rapporté que l’acide
acétique et l’acide butyrique influencent positivement l’état énergétique de l’hôte en
fournissant une source de carbone pour le microbiote intestinal grâce à l’activation des
enzymes de la voie du glyoxylate (Clark, 1996; Donohoe et al., 2011).

183
Figure 5-15 Relation entre la concentration en butyrate et en acétate dans le caeca des poulets
de chair (r=0.89; R2= 73.9%; P< 0.0001).
Nos résultats sur les interactions entre les autres AGCC entre eux ne sortaient pas
significatifs. Rappelons qu’on a mesuré leur concentration uniquement au niveau des caeca
de poulet. Peut-être qu’il serait pertinent de mesurer aussi leurs concentrations aux niveaux
de jabot ou d’autres parties de l’intestin. En effet au niveau du jabot, les lactobacilles
produisent une grande quantité d’acide lactique et d’acides gras à chaîne courte (Fuller,
1984). Dans le jabot, l’acide acétique domine, et dans les caeca, en plus de l’acide acétique
présent majoritairement, on trouve des quantités non négligeables d’acide butyrique, de
petites quantités d’acide propionique ainsi que d’autres acides en moins grandes quantités
(Zhang et al., 2003). Gonzalez-Ortiz et al. (2019) ont montré que chez les poulets de 21 et 42
jours, la section intestinale et non le traitement était le facteur le plus important qui a affecté
les concentrations des acides organiques (Gonzalez-Ortiz et al., 2019). La proportion d'acide
acétique a augmenté progressivement du duodénum aux caeca. L'acide butyrique était le
troisième acide gras volatil le plus abondant (Figure 5-16).

184
Figure 5-16 Concentration des acides gras dans les différents segments de l’intestin au jour
21 (Gonzalez-Ortiz et al., 2019).
Les genres bactériens varient considérablement selon l'emplacement tout au long de l’intestin
grêle des poulets (Pourabedin et Zhao, 2015), ce qui explique les différences de profils des
acides organiques dans chaque segment. Le duodénum et le jéjunum sont principalement
dominés par le genre Lactobacillus (Gong et al., 2007). En revanche, la composition
microbienne de l'iléon est plus diversifiée et moins stable (Pourabedin et Zhao, 2015). L'iléon
est dominé par Lactobacillus, Candidatus Arthromitus, Enterococcus et Escherichia coli
(Mohd Shaufi et al., 2015). Par conséquent, l'acide lactique domine dans ces segments
(Rinttilä et Apajalahti, 2013). L'acide lactique est réduit en concentration au niveau de caeca
aux dépens de l'acide acétique et de l'acide butyrique. Étant donné qu’il est rapidement
absorbé par l'intestin ou utilisé comme substrat pour les bactéries utilisant du lactate, tel que
les Eubacterium, Anaerostipes, Veillonella et Megaspharea (Belenguer et al., 2007). Le
caecum est la partie clé pour la fermentation bactérienne des glucides non digestibles et des

185
protéines non digérées. Trois familles majeures colonisent le caecum : les Clostridiacées, les
Lachnospiracées et les Ruminococcacées (Danzeisen et al., 2011). Ces familles sont des
producteurs importants de butyrate. Pour résumer, avec toutes les données sur les AGCC
intestinaux mesurés dans notre projet ainsi que les données présentées et publiées dans la
littérature, il y a toujours plusieurs considérations à prendre en compte, car ils peuvent
affecter la qualité des résultats:
La motricité iléale est caractérisée à la fois par la présence de mouvements de
segmentation et de péristaltisme inversé chez les poulets peut entraîner une variabilité
de concentrations des AGCC (Godwin et Russell, 1997).
Le turnover de l'intestin vers le sang des AGCC est extrêmement rapide (Pouteau et
al., 2008).
La répétabilité et la reproductibilité des AGCC mesurés à un jour donné durant
l’élevage remettent en question leur pertinence. Ceux-ci nous laissent penser, aurait-
il une possibilité d’'utilisation d'appareils électroniques capables de mesurer les
AGCC en temps réel comme celle utilisée pour le pH (Lee et al., 2018)? Ça sera très
utile pour une meilleure compréhension de la fermentation intestinale.
En ce qui concerne la profondeur des cryptes et la longueur des villosités au niveau de l’iléon,
mesurées par une technique d’histologie par microdissection, on a eu peu de réponses
(section 5.3.1.), pas de corrélation avec les performances, ni les AGCC outre que la tendance
d’amélioration avec l’acide butyrique mentionnée précédemment. Comme il est déjà connu
que l'intestin grêle est le site principal de digestion et d'absorption des nutriments (Choct,
2009). Peut-être, comme pour le cas des AGCC, il est mieux de mesurer les paramètres
histologiques dans différents segments de l’intestin. A noter qu’une première coupe
transversale a été réalisée à 1 cm du diverticule de Meckel et la seconde à 1,5 cm distalement.
La hauteur des villosités est la plus élevée dans le duodénum et diminue en longueur vers
l'iléon (Choct, 2009). Une augmentation de la hauteur des villosités entraîne une
augmentation de la surface et donc une plus grande zone d'absorption des nutriments (Saeid
et al., 2013). Les cryptes sont responsables de la production des entérocytes et des cellules
caliciformes (Shen, 2009). Ces derniers sont responsables de la production de glycoprotéines
de mucine, qui est le composant principal de la couche de mucus qui recouvre l'ensemble du
tractus gastro-intestinal (Smirnov et al., 2006). Cette couche de mucus joue un rôle important

186
dans l'absorption des nutriments et la protection du tractus gastro-intestinal contre les agents
pathogènes (Smirnov et al., 2006). Les cryptes profondes indiquent un fort renouvellement
des cellules le long des villosités, et une forte demande pour produire de nouvelles cellules
pour rassurer la croissance des villosités (Xu et al., 2003). Comme les entérocytes migrent
vers le haut des villosités, où ils sont en contact avec la lumière intestinale, ils subissent un
changement fonctionnel, changement de la fonction sécrétoire dans les cryptes, à une
fonction absorbante (Uni et al., 2000). D’autres indicateurs pourraient être intéressants dans
notre cas, à savoir la détermination du nombre des entérocytes et de cellules caliciformes ou
l’analyse génétique de la mucine intestinale (Smirnov et al., 2006).
5.4. Contribution de la thèse à la diminution de l’utilisation des AFC
Dans le contexte actuel de production, il est important de faire la distinction entre les
antibiotiques utilisés dans le traitement et la prévention des maladies chez les animaux
d'élevage (utilisation thérapeutique et prophylactique prescrite), et de leur utilisation comme
additifs alimentaires pour améliorer les performances de croissance (Castanon, 2007). En
effet, l’effet bénéfique des antibiotiques sur les performances de croissance a été découvert
dans les années 1940, lorsqu'il a été observé que des animaux nourris avec des mycéliums
séchés de Streptomyces aureofaciens contenant des résidus de chlorotétracycline
amélioraient leur croissance. Depuis, plusieurs antibiotiques ont été utilisés comme facteurs
de croissance (AFC) chez les animaux d'élevage. L'introduction de ces agents a coïncidé avec
l'élevage intensif des animaux. Ces produits ont amélioré la conversion alimentaire et la
croissance des animaux et ont réduit la morbidité et la mortalité dues aux maladies (Ewing
et Cole, 1994). Le mécanisme d'action des AFC est en effet lié aux interactions avec la
population microbienne intestinale (Dibner et Richards, 2005; Niewold, 2007). La
supplémentation en AFC favoriserait un microbiote équilibré ce qui réduirait la réponse
inflammatoire et améliorait l’efficacité de l’utilisation de l'énergie à partir des nutriments
(Huyghebaert et al., 2011). Alternativement, ils ont un effet anti-inflammatoire et bénéfique
pour l'épithélium intestinal (Niewold, 2007). Au Canda, le secteur du poulet poursuit sa

187
croissance depuis 2006 avec une progression de 13 % (Figure 5-17; Statistique Canada,
2020). En 2019, elle a augmenté de 2,47 % par rapport à l’année 2018, pour atteindre 1 297
Mkg éviscérés de production totale annuelle (Les Éleveurs La production avicole au Québec,
2019). En raison des demandes accrues des consommateurs pour une viande plus saine (c'est-
à-dire sans résidus ou d’utilisation d’antibiotiques) et de l'augmentation de maladies chez
l'homme dues à la résistance aux antibiotiques (Centres pour le contrôle et la prévention des
maladies, 2013), l’utilisation des alternatives dans l’alimentation animales est devenue
primordiale pour réduireces problèmes de santé.
Figure 5-17 Consommation de viande et protéine animale (kg) par habitant au Canada
(aliments disponible par personne, par année; (Statistique Canada, 2020)).
C’est dans ce contexte que ce projet de doctorat présenté s’est déroulé. Il avait pour objectif
d’étudier les impacts des alternatives aux AFC : l’acide butyrique, MOS, Bacillus subtillus,
avoine nue et quatre produits photogéniques sur les performances de croissance et la santé
digestive des poulets de chair. Nos résultats sont discutés en détail dans la deuxième section
de ce chapitre. En résumé, les effets des alternatives sont rassemblés dans le Tableau 5-3.
Tous nos alternatives ont amélioré le GMQ entre 1,16 et 8 % sauf pour l’avoine nue durant
la phase de croissance qui a montré une plus grande efficacité (+16% du GMQ) et à l’opposé
pour le Bacillus subtilis qui a diminué le GMQ de 8 % durant la phase finition. L’IC été

188
améliorée de 2 à 9 % avec acide butyrique, MOS, avoine nue (sauf durant la phase de
finition). Les produits photogéniques (Micro Aid et Phyteck-P) ont augmenté l‘IC par
l’augmentation de la consommation. Les résultats des études antérieurs ont montré que l'ajout
d'AGP dans l'alimentation des poulets de chair améliorait le gain de poids corporel, la prise
alimentaire et le taux de conversion alimentaire (Corrêa et al., 2003; Ferket, 2004; Miles et
al., 2006; Yang et al., 2009). L'amélioration moyenne de GMQ a été estimée entre 4 et 8%,
et l’IC été améliorée de 2 à 5% (Ewing et Cole., 1994). Pour mieux évaluer leur potentiel,
des études futures devraient être menées sur les poulets élevés dans des fermes, car en
générale les conditions hygiéniques et de conduite d’élevage ne sont pas totalement
optimisés. Et à ne pas oublier qu’il faut toujours améliorer plusieurs aspects reliés à l’élevage
avicole, tels que la conception des bâtiments, l’équipement, la prévention des maladies
infectieuses, la génétique des oiseaux.

189
Table 5-3 Effet des différentes alternatives sur le gain de poids quotidien, la consommation moyenne journalière et l’indice de
conversion alimentaire dans notre étude.
Traitements GMQ (g/j) Effet (%) CMJ (g/j) Effet (%) IC Effet (%)
Phase de démarrage
Contrôle 17,3 +1,4 25,8 +2,1 1,50 Pas d'effet
Acide butyrique 17,5 26,3 1,50
Contrôle 17,3 +1,9 25,8 +1,1 1,50 Pas d'effet
MOS 17,6 26,1 1,50
Contrôle 17,3 +2,3 25,8 +1,7 1,50 -2
Bacillus subtilis 17,7 26,2 1,47
Contrôle 17,3 +16 25,8 +6,1 1,50 -9
Avoine nue 20,1 27,4 1,37
Contrôle 17,1 +2,9 17,8 +1,2 1,04 -2,0
Oleo Poultry 17,6 18,0 1,02
Contrôle 17,1 +1,3 17,8 +2,8 1,04 +1,2
Micro Aid 17,3 18,3 1,06
Contrôle 17,1 +2,3 17,8 +2,8 1,04 +0,4
Xtract Instant 17,5 18,3 1,05
Contrôle 17,1 +3,1 17,8 +3,1 1,04 Pas d'effet
Phyteck-P 17,6 18,3 1,04 Phase de croissance
Contrôle 65,0 -2 81 Pas d'effet 1,24 +2
Acide butyrique 63,8 81 1,27
Contrôle 65,0 +1,16 81 +1,6 1,24 Pas d'effet
MOS 65,6 82 1,24
Contrôle 65,0 -2 81 Pas d'effet 1,24 2
Bacillus subtilis 63,9 81 1,27
Contrôle 65,0 +6,6 81 +1,9 1,24 -4
Avoine nue 69,3 83 1,19
Contrôle 54,2 -0,1 71 +2,1 1,30 -3
Oleo Poultry 53,9 72 1,34
Contrôle 54,2 +7,6 71 +7,3 1,30 Pas d'effet
Micro Aid 58,3 76 1,30
Contrôle 54,2 +2,6 71 +1,9 1,30 Pas d'effet
Xtract Instant 55,6 72 1,30
Contrôle 54,2 +1,6 71 +3,4 1,30 +2
Phyteck-P 55,1 73 1,33 Phase de finition

190
Contrôle 106 +4,4 165 Pas d'effet 1,55 -5
Acide butyrique 111 165 1,48
Contrôle 106 +1,9 165 Pas d'effet 1,55 -2
MOS 108 165 1,52
Contrôle 106 -8 165 -0,7 1,55 +6
Bacillus subtilis 98 164 1,65
Contrôle 106 Pas d'effet 165 +1,8 1,55 +2,5
Avoine nue 106 168 1,59
Contrôle 94,8 +4,5 154 +3,3 1,63 -2
Oleo Poultry 99,1 159 1,61
Contrôle 94,8 +3,7 154 +4,6 1,63 +0,9
Micro Aid 98,3 161 1,64
Contrôle 94,8 +3,2 154 +3,1 1,63 Pas d'effet
Xtract Instant 97,8 159 1,63
Contrôle 94,8 +3,2 154 +3,8 1,63 +3
Phyteck-P 97,8 160 1,68

191
Conclusions
Cette thèse résume les connaissances expérimentales sur les effets, les modes d'action
possibles, et les aspects de l'application des alternatives aux AFC dans l’alimentation des
volailles. L’élevage de poulets sans AFC est un sujet d’actualité qui implique de nombreux
intervenants parmi la chaîne de production avicole, allant des producteurs jusqu’aux
consommateurs. Le projet de doctorat présenté ici avait pour objectif d’étudier les effets des
alternatives aux AFC chez le poulet de chair sur les performances de croissance et la santé
digestive.
Ce projet a permis de concevoir une base de données à partir de la littérature scientifique, qui
inclut des données expérimentales sur la composition chimique et les valeurs nutritives des
aliments ainsi que sur le type et les doses des alternatives. Cet outil est renouvelable et il sera
intéressant de maintenir à jour les bases de données permettant d’apporter des nouveautés
qui élargiront le champ de validité des modèles de prédiction. Ce projet aussi a permis de
concrétiser que l’élevage de poulets de chair sans antibiotiques est possible au Québec
lorsque les AFC sont remplacés par un protocole d’élevage incluant diverses stratégies
alternatives telles que des huiles essentielles, des acides organiques et des prébiotiques
naturels ou synthétiques. C’est vrai que les résultats obtenus et les observations faites pendant
cette thèse ont permis de répondre à certaines interrogations, mais ont aussi contribué à
soulever d’autres questionnements.
La présente étude ouvre donc sur une toute nouvelle vision de la production à grande échelle
de poulets de chair élevés sans AFC au Québec. À la lumière des résultats obtenus, ce projet
pourrait être complété par des modèles de prédiction sur les paramètres de la santé digestive
et les mesures sanguines afin de construire des modèles mécanistes permettant de bien
expliquer et de prévoir le comportement des poulets de chair nourries avec des alternatives

192
aux AFC. Il serait aussi pertinent de compléter le profil bactérien détaillé des poulets durant
les deux projets sur terrain afin de mieux comprendre l’effet spécifique de ces alternatives
aux AFC sur le microbiote intestinal et donc sur l’inhibition de certains microorganismes
indésirables qui pourraient compromettre la santé digestive des poulets de chair.

193
Bibliographie
Abd El-Hakim, A.S., Cherian, G., Ali, M.N., 2009. Use of organic acids, herbs and their
combination to improve the utilization of commercial low protein broiler diets.
International. Poultry Science. 8, 14-20.
Adil, S., Banday, T., Bhat, G.A., Mir, M.S., Rehman, M., 2010. Effect of Dietary
Supplementation of Organic Acids on Performance, Intestinal Histomorphology, and
Serum Biochemistry of Broiler Chicken. Veterinary Medicine International. 2010,
479-485.
Aehle, W., 2004. Industrial enzymes: overview of industrial enzyme applications. Pages
257–262 in Enzymes in Industry: Production and Applications. 2nd ed. Wiley-VCH
Verlag GmbH & Co. KGaA, Weinheim.
Afsharmanesh, M., Sadaghi, B., 2013. Effects of dietary alternatives (probiotic, green tea
powder, and kombucha tea) as antimicrobial growth promoters ongrowth, ileal
nutrient digestibility, blood parameters, and immune responseof broiler chickens.
Comparative Clinical Pathology. 23, 717–724.
Afssa, 2001., Apports nutritionnels conseillées pour la population française. 3e édition.
Coordonateur Ambroise Martin.
Agriculture Canada, 1993. http://biblio.uqar.ca/archives/30428359.pdf. (Page consultee le 20
Septembre 2017).
Ahsan, U., Cengiz, Ö., Raza, I., Kuter, E., Chacher, M. F. A., Iqbal, Z., Çakir, S., 2016.
Sodium butyrate in chicken nutrition: the dynamics of performance, gut microbiota,
gut morphology, and immunity. Worlds Poultry Sciences. 72, 265-275.
Ajinomoto, 2018. Ideal Amino Acid Profile and Low Crude Protein Diets for Broiler
Chickens.
Akçay, A., Ertuğrul. O., Gurcan, I. S., Karaer, Z., 2011. "Quantification of risk factors of
coccidiosis in broilers by using logistic regression analysis." Ankara Üniversitesi
Veteriner Fakültesi Dergisi. 58, 195-202.
Akhtar, M., Tariq, A. F., Awais, M. M., Iqbal, Z., Muhammad, F., Shahid, M., Hiszczynska-
Sawicka, E., 2012. Studies on wheat bran Arabinoxylan for its immunostimulatory
and protective effects against avian coccidiosis. Carbohydrate Polymers. 90, 333-339.
Alaeldein, M. A., Abdullah, H. A., 2013. Effects of the Essential Oil Blend CRINA Poultry
in Feed on Broiler Performance and Gut Microbiology.
Italian Journal of Animal Science. 12, 513-517.
Alagawany, M., Abd El-Hack, M. E., Farag, M. R., Shaheen, H. M., Abd El-Latif, M. A.,
Noreldin, A. E., Patra, A. K., 2018. The usefulness of oregano and its derivatives in
poultry nutrition. Worlds. Poultry Sciences. 74, 463- 474.
Alain, F., 2001. L'avoine nue pour les vaches laitières… oui c'est intéressant! Agri Réseau.
https://www.agrireseau.net/bovinslaitiers/Documents/Avoine%20nue.pdf (Page
consulté le 26 juin 2017).
Alemu, R., Nyachoti, C. M., 2018. Application of resistant starch in swine and poultry diets
with particular reference to gut health and function. Animal Nutrition. 4, 305-310.
Ali, M. N., Abou, M. S., El-kloub, M., 2008. Incorporation of wheat bran in broiler diets.
International Journal of Poulty Science.7, 6-13.
Almargot, J., 1982. L'appareil digestif et ses annexes, pages 15-32. In : Manuel d'anatomie
et d'autopsie aviaires. Edition : Le point vétérinaire.

194
Al-Sheikhly, F., Al-Saieg, A., 1980. Role of Coccidia in the occurrence of necrotic enteritis
of chickens. Avian Diseases. 24, 324-333
Al-Sheikhly, F., Truscott, R.B., 1977. The pathology of necrotic enteritis of chickens
following infusion of broth cultures of Clostridium perfringens into the duodenum.
Avian Diseases. 21, 230-240.
Andreopoulou, M., Tsiouris, V., Georgopoulou, I., 2014. Effects of organic acids on the gut
ecosystem and on the performance of broiler chickens. Journal of the Hellenic
Veterinary Medical Society. 65, 289-302.
Angkanaporn, K., Choct, M., Bryden, W.L., Annison, E.F., Annison, G., 1994. Effects of
wheat pentosans on endogenous amino acid losses in chickens. Journal of the Science
of Food and Agriculture. 66, 399-404
Annett, C. B., Viste, J. R., Chirino-Trejo, M., Classen, H. L., Middleton, D. M., Simko, E.,
2002. Necrotic enteritis: effect of barley, wheat and corn diets on proliferation of
Clostridium perfringens type A. Avian Pathology. 31, 599-602.
Annison, G., 1992. Commercial enzyme supplementation of wheat-based diets raises ileal
glycanase activities and improves apparent metabolisable energy, starch and pentosan
digestibilities in broiler chickens. Animal Feed Science and Technology. 38, 105-121.
Anses, 2001 a. Actualisation des apports nutritionnels conseillés pour les acides gras.
Rapport d’expertise collective. Edition scientifique. Mai 2011.
Anses, 2011b. Alimentation humaine. Lipides et recommandations nutritionnelles. Juillet
2011. Disponible sur le site : http://www.anses.fr/PNE001.htm. (Page consultée le12
février 2017).
Apajalahti, J., Kettunen, A., Graham, H., 2004. Characteristics of the gastrointestinal
microbial communities, with special reference to the chicken. World's Poultry
Science Journal. 60: 223-325.
Arena, M. P., Caggianiello, G., Fiocco, D., Russo, P., Torelli, M., Spano, G., Capozzi, V.,
2014. Barley β-glucans-containing food enhances probiotic performances of
beneficial bacteria. International journal of molecular sciences. 15, 3025-3039.
Awad, W. A., Böhm, J; Razzazi-Fazeli, E., Ghareeb, K., Zentek, J., 2006. Effect of addition
of a probiotic microorganism to broiler diets contaminated with deoxynivalenol on
performance and histological alterations of intestinal villi of broiler chickens. Poulty
Science. 85, 974-979.
Awad, W. A., Ghareeb, K., Abdel-Raheem, S., Bohm, J., 2009. Effects of dietary inclusion
of probiotic and synbiotic on growth performance, organ weights, and intestinal
histomorphology of broiler chickens. Poulty Science. 88, 49–56.
Bach-Knudsen, K. E., 1997. Carbohydrate and lignin contents of plant materials used in
animal feeding. Animal Feed Science and Technology. 67, 319–338.
Bai, S. P., Wu, A. M., Ding, X. M., Lei, Y., Bai, J., Zhang, K.Y., Chio, J. S., 2013. Effects
of probiotic supplemented diets on growth performance and intestinal immune
characteristics of broiler chickens. Poultry Science. 92, 663-670.
Barrera, M., Cervantes, M., Sauer, W. C., Araiza, A. B., Torrentera, N., Cervantes, M., 2004.
Ileal amino acid digestibility and performance of growing pigs fed wheat-based diets
supplemented with xylanase. Journal of Animal Sciences. 82, 1997‑2003.
Basmacioğlu-Malayoğlu, H., Ozdemir, P., Bağriyanik, H. A., 2016. Influence of an organic
acid blend and essential oil blend, individually or in combination, on growth
performance, carcass parameters, apparent digestibility, intestinal microflora and
intestinal morphology of broilers. British Poultry Science. 57, 227-234.

195
Bauer, B.W., Gangadoo, S., Bajagai, Y.S., Van, T. T. H., Moore, R. J., Stanley, D., 2019.
Oreganopowder reduces Streptococcus and increases SCFA concentration in a mixed
bacterial culture assay. PLoS ONE. 14, 12:e0216853.
Baurhoo, B., Ferket, P. R., Zhao, X., 2009. Effects of Diets Containing Different
Concentrations of Mannanoligosaccharide or Antibiotics on Growth Performance,
Intestinal Development, Cecal and Litter Microbial Populations, and Carcass
Parameters of Broilers. Poultry Science. 88, 2262-2272.
Baurhoo, B., Phillip, L., Ruiz-Feria, C. A., 2007. Effects of purified lignin and mannan
oligosaccharides on intestinal integrity and microbial populations in the ceca and litter
of broiler chickens. Poultry science. 86, 1070-1078.
Beaucage, C., Bonnier, V. Y., 1998. Épidémiologie appliquée. Une initiation à la lecture
critique de la littérature en sciences de la santé (Book Review). Population (French
Edition). 53, 660-661.
Bedford, M. R. et G. G. Partridge., 2010. Enzymes in farm animal nutrition. 2e édition. CAB
International, Wallingford, Royaume-Uni.
Bedford, M. R., Classen, H. L., 1992. Reduction of intestinal viscosity through manipulation
of dietary rye and pentosanase concentration is effected through changes in the
carbohydrate composition of the intestinal aqueous phase and results in improved
growth rate and food conversion efficiency of broiler chicks. Journal of Nutrition.
122, 560‑569.
Bedford, M., 2000. Removal of antibiotic growth promoters from poultry diets: implications
and strategies to minimise subsequent problems. World Poultry Science journal. 56,
347-365.
Bedford, M.R., 1995. Mechanism of action and potential environmental benefits from the
use of feed enzymes. Anim. Feed Sci. Tech. 53, 145-155.
Beg, Q. K., Kapoor, M., Mahajan, L., Hoondal, G. S., 2001. Microbial xylanases and their
industrial applications: A review. Applied Microbiology and Biotechnology. 56,
326‑338.
Belenguer, A., Duncan, S. H., Holtrop, G., Anderson, S. E., Lobley, G. E., Flint, H. J., 2007.
Impact of pH on lactate formation and utilization by human fecal microbial
communities. pplied and Environmental Microbiology. 73, 6526–6533.
Ben Arfa, A., Combes, S., Preziosi-Belloy, L., Gontard, N., Chalier, P., 2006. Antimicrobial
activity of carvacrol related to its chemical structure. Letters in Applied
Microbiology. 43, 149-154.
Bernier, G., Phaneuf, J.B., Filion, R., 1977. [Necrotic enteritis in broiler chickens. III. Study
of the factors favoring the multiplication of Clostridium perfringens and the
experimental transmission of the disease]. Canadian journal of comparative medicine.
41, 112-116.
Biggs, P., Parsons, C. M., 2009. The effects of whole grains on nutrient digestibilities, growth
performance and cecal short-chain fatty acid concentrations in young chicks fed
ground corn-soybean meal diets. Poultry Science. 88, 1893–1905.
Bilgili, S.F., Hess, J.B., Blake, J.P., Macklin, K.S., Saenmahayak, B., Sibley, J.L., 2009.
Influence of bedding material on footpad dermatitis in broiler chickens. Journal
Applied Poultry Research, 18, 583 – 589.
Bjerrum, L., Engberg, R.M., Leser, T.D., Jensen, B.B., Finster, K., Pedersen, K., 2006.
Microbial Community Composition of the Ileum and Cecum of Broiler Chickens as

196
Revealed by Molecular and Culture Based Techniques. Poultry Science. 85, 1151-
1164.
Bolton, W., Dewar, W.A., 1965. The digestibility of acetic, propionic and butyric acids by
the fowl. British Poultry Science. 6, 103-105.
Bouhaddouda, N., Aouadi, S., Labiod, R., 2016. Evaluation of Chemical Composition and
Biological Activities of Essential Oil and Methanolic Extract of Origanum vulgare L.
ssp. glandulosum (Desf.) Ietswaart from Algeria. International Journal of
Pharmacognosy and Phytochemical Research. 8, 104-112.
Bouvarel, I., Lessire, M., Narcy, A., Duval, E., Grasteau, S., Quinsac, A., Heuze, V., 2014.
Des sources de protéines locales pour l’alimentation des volailles: quelles voies de
progrès?. OCL, 21(4), D405.
Bouzouaia, M., 2019. Rappels sur les maladies nutritionnelles chez le poulet de chair. El
jadida, Hôtel Mazagan, 11 et 12 juillet
2019.https://www.novusint.com/Portals/0/NovusDocuments/1%20Physio%20Patho
%20Digestive%20BM%20NOVUS%20Mc%20VF.pdf. (Page consultée le 19 Juillet
2020).
Brand, T.S., van der Merwe, J.P., 1996. Naked oats (Avena nuda) as a substitute for maize
in diets for weanling and grower-finisher pigs. Animal Feed Science and Technology.
57, 139-147.
Brenes A., Roura E., 2010. Essential oils in poultry nutrition: Main effects and modes of
action. Animal Feed Science and Technology. 158, 1-14.
Burrows, V. D., 2005. Hulless oats: “Specialty grains for food and feed”. in E. S. M. Abdel-
Aal and P. Wood, eds.. American Association of Cereal Chemists, St. Paul, MN.
Pages 223–251.
Canani, R.B., Costanzo, M.D., Leone, L., Bedogni, G., Brambilla, P., Cianfarani, S., 2011.
Epigenetic mechanisms elicited by nutrition in early life. Nutrition Research
Reviews. 24, 198–205.
Capek, P., Kubacková, M., Alfoldi, J., Bilisics, L., Liskova, D., Kakoniova, D., 2000.
Galactoglucomannan from the secondary cell wall of Picea abies L. Karst.
Carbohydrate Research. 329, 635–645.
Carlson, J.L., Erickson, J.M., Lloyd, B.B., Slavin, J.L., 2018. Health effects and sources of
prebiotic dietary fiber. Current Developments in Nutrition. 2(3):nzy005.
Carr, L. E., Wheaton, F. W., Douglass, L. W., 1990. Empirical models to determine ammonia
concentrations from broiler chicken litter. American Society of Agricultural
Engineers. 33, 1337-1342
Castanon, J. I. R., 2007. History of the use of antibiotic as growth promoters in European
poultry feeds. Poultry Science. 86, 2466–2471.
Cave, N. A., Hamilton, R.M.G., Burrows, V. D., 1989. Evaluation of naked oats (Avena
nuda) as a feeding stuff for laying hens. Canadian Journal of Animal Science. 69,
789-799.
Chakdar, H., Kumar, M., Pandiyan, K., Singh, A., Nanjappan, K., Kashyap, P.L., Srivastava,
A.K., 2016. Bacterial xylanases: biology to biotechnology. 3 Biotech. 6, 150.
Chamba, F., Puyalto, M., Ortiz, A., Torrealba, H., Mallo, J. J., Riboty, R., 2014. Effect of
partially protected sodium butyrate on performance, digestive organs, intestinal villi
and E. coli development in broilers chickens. International Journal of Poultry
Science.13, 390-396.

197
Chang, C.H., Teng, P.Y., Lee, T.T., Yu, B., 2019. Effects of Multi-Strain Probiotics
Combined with Gardeniae fructus on Intestinal Microbiota, Metabolites, and
Morphology in Broilers. Poultry Science. 56, 32-43.
Chaveerach, P., Keuzenkamp, D. A., Lipman, L. J., Van Knapen, F., 2004. Effect of Organic
Acids in Drinking Water for Young Broilers on Campylobacter Infection, Volatile
Fatty Acid Production, Gut Microflora and Histological Cell Changes. Poultry
Science. 83, 330-334.
Chen, H., Liu, S., Xu, X. R., Zhou, G. L., Liu, S. S., Yue, W. Z., Kai-Feng, S., Guang, Y.,
2015. Antibiotics in the Coastal Environment of the Hailing Bay Region, South
China Sea: Spatial Distribution, Source Analysis and Ecological Risks. Marine
Pollution Bulletin. 95, 365-373.
Cherrington, C. A., Hinton, M., Mead, G. C., Chopra, I., 1991.Organic acids: Chemistry,
antibacterial activity and practicalapplications. Advances in Microbial Physiology.
32, 87–108.
Choct, M., 2006. Enzymes for the feed industry: past, present and future. World's Poultry
Science Journal. 62, 5‑15.
Choct, M., 2009. Managing gut health through nutrition. British Poultry Science. 50, 9-15.
Choct, M., Hughes, R. J., Wang, J., Bedford, M. R., Morgan, A. J., Annison, G., 1995. Feed
enzymes eliminate the anti-nutritive effect of non-starch polysaccharides and modify
the fermentation in broilers. In Proceedings of of the Australian Poultry Science
Symposium 1995. Poultry Research Foundation, University of Sydney.
Chutkan, R., Fahey, G., Wright, W. L., Mcrorie, J., 2012. Viscous versus nonviscous soluble
fiber supplements: Mechanisms and evidence for fiber-specific health benefits.
Journal of the American Association of Nurse Practitioners. 24, 476‑487.
Clark, D.P., Cronan, J.E., 1996. Two-carbon compounds and fatty acids as carbon
sources Escherichia coli and Salmonella: cellular and molecular biology, 2nd ed
ASM Press, Washington, DC: 343–357.
Collet, S. R., 2006. Wet litter: Its causes and prevention and the role of nutrition. In: G. C.
Perry, editor, Avian gut function in health and disease. CABI Publishing,
Wallingford, UK. p. 195–209.
Collett, S. R. 2012. Nutrition and wet litter problems in poultry. Anim. Feed Sci. Technol.
173:65–75.
Corrand, L., Guérin, J. L., 2010. Les coccidioses aviaires. Avi Campus.
http://www.avicampus.fr/PDF/PDFpathologie/coccidioses2.pdf (Page consulté le 9
juin 2017).
Corrêa G., Gomes, A., Corrêa A., Salles, A., Mattos, E., 2003. Efeito de antibiótico e
probióticos sobre o desempenho e rendimento de carcaça de frangos de corte Effect
of antibiotic and probiotic on the performance and carcass yield of broilers. Arquivo
Brasileiro de Medicina Veterinária e Zootecnia. 55, 467-473.
Courtin, C. M., Broekaert, W. F., Swennen, K., Lescroart, O., Onagbesan, O., Buyse, J.,
Decuypere, E., Wiele, T., Marzorati, M., Verstraete, W., Huyghebaert, G., Delcour,
J. A., 2008. Dietary inclusion of wheat bran arabinoxylooligosaccharides induces
beneficial nutritional effects in chickens. Cereal Chemistry .85, 607-613.
Cox, C. M., Dalloul, R. A., 2010. Beta-glucans as immunomodulators in poultry: use and
potential applications. Avian Biology Research. 3, 171-178.
Cray, C., Zaias, J., Altman, N. H., 2009. Acute phase response in animals: a review.
Comparative Medicine. 59, 517-526.

198
Crevieu-Gabriel, I., 1999. Digestion des protéines végétales chez les monogastriques.
Exemple des protéines de pois. Productions animales. 12, 147-161.
Dahiya, J.P., Hoehler, D., Wilkie, D.C., Van Kessel, A.G., Drew, M.D., 2005. Dietary
glycine concentration affects intestinal Clostridium perfringens and lactobacilli
populations in broiler chickens. Poultry Science.84, 1875-1885.
Danicke, S., Vahjen, W., Simon, O., Jeroch, H., 1999. Effects of dietary fat type and xylanase
supplementation to rye-based broiler diets on selected bacterial groups adhering to
the intestinal epithelium, on transit time of feed, and on nutrient digestibility. Poultry
Science. 78, 1292-1299.
Danzeisen, J. L., Kim, H. B., Isaacson, R.E., Tu, Z.J., Johnson, T.J., 2011. Modulations of
the chicken cecal microbiome and metagenome in response to anticoccidial and
growth promoter treatment. PLoS One. 6, 927-949.
Daou, C., Zhang, H., 2012. Oat beta-glucan: Its role in health promotion and prevention of
diseases. Comprehensive Reviews in Food Science and Food Safety.11, 355-365
De Maesschalck, C., Eeckhaut, V., Maertens, L., De Lange, L., Marchal, L., Nezer, C.,
Haesebrouck, F., 2015. The effects of xylo-oligosaccharides on performance and
microbiota in broiler chickens. Applied and environmental microbiology.
Den Besten, G., van Eunen, K., Groen, A. K., Venema, K., Reijngoud, D. J., Bakker, B. M.,
2013. The role of short-chain fatty acids in the interplay between diet, gut microbiota,
and host energy metabolismJournal of Lipid Research. 2013. 54, 2325–2340.
Denbow D., 2000. Gastrointestinal anatomy and physiology. Sturkie's avian physiology. 5,
299-325.
Denis, O. K., House, D. J., Nyachoti, M., 2004. Alternatives to antibiotics in swine diets: a
molecular approach. Department of Animal Science. University of Manitoba.
Canada.
Diarra, M. S., Malouin, F., 2014. Antibiotics in Canadian poultry productions and anticipated
alternatives. Frontiers in microbiology. 5, 282.
Dibner, J.J., Buttin, P., 2002. Use of organic acids as a model to study the impact of gut
microflora on nutrition and metabolism. Journal of Applied Poultry Research.11, 453-
463.
Dibner, J., Richards, J., 2005. Antibiotic growth promoters in agriculture: history and mode
of action. Poultry Science. 84, 634-643.
Donohoe, D.R., Garge, N., Zhang, X., Sun, W., O'Connell, T.M., 2011. The microbiome and
butyrate regulate energy metabolism and autophagy in the mammalian colon. Cell
Metabolism. 13, 517–526.
Drogoul, C., Gadoud, R., 2004. Nutrition et alimentation des animaux d’élevage.
L’alimentation des monogastriques. Digon. Educagri Editions.
Dubourdieu, D., Desplanques, C., Villettaz, J., Ribereau-Gayon, P., 1985. Investigations of
an industrial β-D-glucanase from Trichoderma harzianum. Carbohydrate Research.
144, 277-287.
Ducatelle, R., Goossens, E., De Meyer, F., Eeckhaut, V., Antonissen, G., Haesebrouck, F.,
Van Immerseel, F., 2018. Biomarkers for monitoring intestinal health in poultry:
present status and future perspectives. Veterinary research. 49, 1-9.
Duke, G. E., 1997. Gastrointestinal physiology and nutrition in wild birds. Proceedings of
the Nutrition Society. 56, 1049-1056.

199
Dusart, L., 2015. Quelques rappels sur les mécanismes
physiologiques.http://itab.asso.fr/downloads/cahier-
volailles/cahier_volaille_chapitre2_web.pdf. (Page consultée le 15 Janvier 2019)
Ebersbach, T., Andersen, J. B., Bergstrom, A., Hutkins, R. W., Licht, T. R., 2012. Xylo-
Oligosaccharides Inhibit Pathogen Adhesion to Enterocytes In Vitro. Research in
Microbiology. 163, 22–27.
Eckersall, P. D., Bell, R., 2010. Acute phase proteins: Biomarkers of infection and
inflammation in veterinary medicine. The veterinary journal. 185, 23-27.
Eeckhaut, V., Van Immerseel, F., Dewulf, J., Pasmans, F., Haesebrouck, F., Ducatelle, R.,
Broekaert, W. F., 2008. Arabinoxylooligosaccharides from wheat bran inhibit
Salmonella colonization in broiler chickens. Poultry science. 87, 2329-2334.
Elliott, H. A., Collins, N. E., 1982.Factors affecting ammonia release in broiler houses Trans.
American Society of Agricultural Engineers. 25, 413-418
Engberg, R. M., Hedemann, M. S., Jensen, B. B., 2002. The influence of grinding and
pelleting of feed on the microbial composition and activity in the digestive tract of
broiler chickens. British Poultry science. 43, 569-579.
Engberg, R.M., Hedemann M.S., Steenfeldt S., Jensen B.B., 2004. Influence of whole wheat
and xylanase on broiler performance and microbial composition and activity in the
digestive tract Poultry Science. 2004. 83, 925-938.
Engstrom, B.E., Fermer, C., Lindberg, A., Saarinen, E., Baverud, V., Gunnarsson, A., 2003.
Molecular typing of isolates of Clostridium perfringens from healthy and diseased
poultry. Veterinary Microbiology. 94, 225-235.
Ewing, W. N., Cole, D. J. A., 1994. The living gut. An introduction to microorganisms in
nutrition. Context, Dungannon, Ireland.
FAO/OIE/WHO. 2003. Joint FAO/OIE/WHO Expert Workshop on Non-Human
Antimicrobial Usage and Antimicrobial Resistance: Scientific Assessment. Food and
Agriculture Organization of the United Nations, World Organization for Animal
Health and World health Organization.
http://apps.who.int/iris/bitstream/10665/68883/1/WHO_CDS_CPE_ZFK_2004.7.pd
f. (Page consulté le 30 juillet 2018).
Faridi, A., Gitoee, A., France, J., 2015. A meta-analysis of the effects of non-phytate
phosphorus on broiler performance and tibia ash concentration. Poultry Science. 94,
2753–2762.
Farner, D.S, 1943. Gastric Hydrogen Ion Concentration and Acidity in the Domestic Fowl.
Poultry Science. 22, 79–82.
Farrell, D. J., Takhar, B. S., Barr, A. R., Pell, A. S., 1991. Naked oats: their potential as a
complete feed for poultry. Proceedings of Recent Advances in Animal Nutrition. 312-
25.
Ferguson, N. S., Gates, R. S., Taraba, J. L., Cantor, A. H., Pescatore, A. J., Straw, M. L.,
Burnham, D. J., 1998. The effect of dietary protein and phosphorus on ammonia
concentration and litter composition in broilers. Poultry Science. 77, 1085-1093.
Ferket, P. R., 2004. Alternatives to antibiotics in poultry production: responses, practical
experience and recommendations. Pages 56–67 in Nutritional Biotechnology in the
Feed and Food Industries. Lyons, T. P., Jacques, K. A., ed. Proceedings of Alltech’s
20th Annual Symposium. Lexington, KY: Nottingham University Press.
Fernandez, V., Ruiz, M., 2003. Técnico en ganadería, Volume 1. Técnico en ganadería,
Volume 1. Editeur : Culural. 556 pages.

200
Fernandez-Rubio, C., Ordonez, C., Abad-Gonzalez, J., Garcia-Gallego, A., Honrubia, M. P.,
Mallo, J. J., Balana-Fouce, R., 2009. Butyric acid-based additives help protect broiler
chickens from Salmonella enteritidis infection. Poultry science. 88, 943–948.
Fetterer, R. H., Augustine, P. C., Allen, P. C., Barfield, R. C., 2003. The effect of dietary
betaine on intestinal and plasma levels of betaine in uninfected and coccidia-infected
broiler chicks. Parasitology Research. 90, 343-348.
Field, C., Johnson, J., I., R, Schley, P., D., 2002. Nutrients and their role in host resistance to
infection. Journal of Leukocyte Biology. 71, 6–32.
Flis, M., Sobotka, W., Antoszkiewicz, Z., Lipiński, K., Zduńczyk, Z., 2007. Effect of husked
and naked oat used in the diets supplemented with linseed oil on the growth
performance of pigs, carcass and meat quality. Archiv fur Tierzucht, Dummerstorf
Special Issue. 50, 161-171.
Friend, D.W., Fortin, A., Poste, L.M., Butler, G., Kramer, J.K., Burrows, V.D., 1988. Feeding
and metabolism trials and assessment of carcass and meat quality for growing-
finishing pigs fed naked oats. Canadian Journal of Animal Science. 68, 511-522.
Fuller, R., 1984. Microbial activity in the alimentary tract of birds. Proceedings of the
Nutrition Society. 43, 55-61.
Gabriel, I., Mallet, S., Lessire, M., 2003. La microflore digestive : une composante oubliée
de la nutrition des volailles. Cinquièmes journées de la recherche avicole. Tours, 26
et 27 mars 2003.
Gabriel, I., Mallet, S., Sibille, P., 2005. La microflore digestive des volailles: facteurs de
variation et conséquences pour l’animal. INRA Production Animales. 18, 309-322.
Garcia, F., Agabriel, J., 2008. CompoCow: a predictive model to estimate variations in body
composition and the energy requirements of cull cows during finishing. The Journal
of Agricultural Science. 146, 251-265.
Garcia, V., Catala-Gregori, P., Hernandez, F., Megias, M.D., Madrid, J., 2007. Effect of
formic acid and plant extracts on growth, nutrient digestibility, intestine mucosa
morphology and meat yield of broilers. Journal of Applied Poultry Research. 16, 555-
562.
Gaucher, M. L., 2016. Étude de l'impact de deux traitements, dont un sans antibiotiques, sur
la santé digestive et les populations de Clostridium perfringens dans des élevages de
poulets de chair. Thése de Doctorat. Université de
Montreal.https://papyrus.bib.umontreal.ca/xmlui/bitstream/handle/1866/13372/Gauc
her_Marie-Lou_2015_these.pdf?sequence=4&isAllowed=y.com. (Page consultée le
20 Juin 2020)
Gauthier, L., Pharm, B., 2002. L’élaboration d’une méta-analyse : un processus complexe!
Pharmactuel. 35, 211-213.
Ghareeb, K., Awad, W. A., Böhm, J., Zebeli, Q., 2016. Impact of luminal and systemic
endotoxin exposure on gut function, immune response and performance of chickens.
World's Poultry Science Journal. 72, 367-380.
Ghasemi, H. A., Kasani, N., Taherpour,K., 2014. Effects of Black Cumin Seed (Nigella
Sativa L.), a Probiotic, a Prebiotic and a Synbiotic on Growth Performance, Immune
Response and Blood Characteristics of Male Broilers. Livestock Science. 164, 128-
134.
Ghazanfari, S., Mohammadi, Z., Adib Moradi, M., 2015. Effects of Coriander Essential Oil
on the Performance, Blood Characteristics, Intestinal Microbiota and Histological of
Broilers. Brazilian Journal of Poultry Science. 17, 419-426.

201
Gholamiandekhordi, A.R., Ducatelle, R., Heyndrickx, M., Haesebrouck, F., Van Immerseel,
F., 2006. Molecular and phenotypical characterization of Clostridium perfringens
isolates from poultry flocks with different disease status. Veterinary Microbiology.
113,143-152.
Giannenas, I., Papadopoulos, E., Tsalie, E., Triantafillou, E., Henikl, S., Teichmann, K.,
2012. Assessment of Dietary Supplementation with Probiotics on Performance,
Intestinal Morphology and 614 Microflora of Chickens Infected with Eimeria
Tenella. Veterinary Parasitology. 188, 31-40.
Giannenas, I., Papaneophytou, C., Tsalie, E., Mavridis, S., Triantafillou, E., Kontopidis, G.
and Tontis, D., 2014b. Dietary Supplementation of Benzoic Acid and Essential Oil
Compounds Affects Buffering Capacity of the Feeds, Performance of Turkey Poults
and Their Antioxidant Status, pH in the Digestive Tract, Intestinal Microbiota and
Morphology. Asian-Australasian Journal of Animal Sciences. 27, 225-236.
Giannenas, I.A., Papaneophytou, C.P., Tsalie, E., Triantafillou, E., Tontis, D., Kontopidis,
G.A., 2014a. The effects of benzoic acid and essential oil compounds in combination
with protease on the performance of chickens. Journal of Animal and Feed Sciences.
23, 73-81.
Glass, G.V., Mcgaw, B.M., Smith, M.L., 1981. Meta-analysis in social research. Beverly
Hills: Sage publications.
Glass, G: V., 1976. Primary, secondary, and meta-analysis of research. Educational
Researcher. 5, 3-8.
Godwin, I. R., Russell, W. J., 1997. Reverse peristalsis in the chicken digestive tract. Pages
229 in Proceedings of the Recent Advances in Animal Nutrition in Australia. J. L.
Corbett, M. Choct, J. V. Nolan, and J. B. Rowe eds., University of New England,
Armidale, Australia.
Golder, H.M., Geier, M.S., Hynd, P.I., Forder, R.E.A., Boulianne, M., Hughes, R.J., 2010.
Feed additives influence goblet cell distribution and villus-crypt architecture in
broilers after necrotic enteritis challenge. Annual Australian Poultry Science
Symposium (21st : 2010 : Sydney, Australia).211-214.
Golian, A., Maurice, D.V., 1992. Dietary poultry fat and gastrointestinal transit-time of feed
and fat utilization in broiler-chickens. Poultry Science. 71, 1357-1363.
Gong, J., Forster, R.J., Yu, H., Chambers, J.R., Wheatcroft, R., Sabour, P.M., Chen, S., 2002.
Molecular analysis of bacterial populations in the ileum of broiler chickens and
comparison with bacteria in the cecum. FEMS Microbiology Ecology. 41, 171-179.
Gong, J., Yu, H., Liu, T., Gill, J. J., Chambers, J. R., Wheatcroft, R., Sabour, P. M., 2008.
Effects of zinc bacitracin, bird age and access to range on bacterial microbiota in the
ileum and caeca of broiler chickens. Journal of Applied Microbiology. 104, 1372-
1382.
Gonzalez-Ortiz, G., Dos Santos, T. T., Vienola, K., Vartiainen, S., Apajalahti, J., Bedford,
M.R., 2019. Response of broiler chickens to xylanase and butyrate supplementation.
Poultry Science. 98, 3914–3925.
Guerville, M., Boudry, G., 2016. Gastrointestinal and hepatic mechanisms limiting entry and
dissemination of lipopolysaccharide into the systemic circulation. American Journal
of Physiology-Gastrointestinal and Liver Physiology. 311, G1-G15.
Guilloteau, P., Zabielski, R., David, J.C., Blum, J.W., Morisset, J.A., Biernat, M., 2009.
Sodium-butyrate as a growth promoter in milk replacer formula for young
calves. Journal of Dairy Sciences. 92:1038–1049.

202
Guo, Y., Ali, R.A. Qureshi, M.A., 2003. The influence ofβ-glucan on immune responses in
broilerchicks.Immunopharmacology and Immunotoxicology. 25, 461-472.
Hardin, B. E., Roney, C.S., 1989. Effects of pH on selected bacteria. Alabama Department
of Agriculture and Industry Report.
Hassan, H.M.A., Mohamed, M.A., Youssef, A.W., Hassan, E.R., 2010. Effect of Using
Organic Acids to Substitute Antibiotic Growth Promoters on Performance and
Intestinal Microflora of Broilers. Asian-Australasian Journal of Animal Sciences. 23,
1348-1353.
Hassanein, S.M., Soliman, N.K., 2010. Effect of Probiotic (Saccharomyces Cerevisiae)
Adding to Diets on Intestinal Microflora and Performance of Hy-Line Layers Hens.
Journal of American Science. 6,159-169.
Hedges, L. V., 1984. Advances in statistical methods for meta-analysis. In P. Wortman, &
W. H. Yeaton (Eds.), Issues in Data Synthesis: New Directions in Program
Evaluation (pp. 25-42). Jossey-Bass.
Herpol, C., Van Grembergen, G., 1967. La signification du pH dans le tube digestif de gallus
domesticus. In Annales de Biologie Animale, Biochimie, Biophysique. 7, 33-38.
Hetland, H., Svihus, B., Krogdahl, A., 2003.Effects of oat hulls and woods havings on
digestion in broilers and layers fed diets based on whole or ground wheat. British
Poultry science. 44, 275-282.
Hipsley, E.H., 1953. Dietary “fiber” and pregnancy toxaemia. British Medical Journal. 2,
420-422.
Hodges, R.D., 1974. The digestive System. In: The hystology of the fowl (ed. by Press A).
35-108.
Hoffman, L. A., 1995. World production and use of oats. In: Welch, R. W. (Ed.), the oat
crop: production and utilization. Chapman & Hall.
Hooge, D.M., 2004. Meta-analysis of broiler chicken pen trials evaluating dietary
oligosaccharide, 1993–2003. International Journal of Poultry Science. 3, 163-174.
Hu, Z., Y. Guo., 2007. Effects of dietary sodium butyrate supplementation on the intestinal
morphological structure, absorptive function, and gut flora in chickens. Animal Feed
Science and Technology. 132, 240-249.
Huyghebaert, G., Ducatelle, R., Van Immerseel, F., 2011. An update on alternatives to
antimicrobial growth promoters for broilers. Veterinary Journal. 187, 182–188.
Irène Gabriel, 2014. Microbiote et santé digestive des volailles : État des lieux. INRA.
https://www.nobivet.fr/download/st-malo-2014-presentations-02-gabriel.pdf. (Page
consulté le 17 février 2019).
Izat, A.L., Tidwell, N.M., Thomas, R.A., Reiber, M.A., Adams, M.H., Colberg, M.,
Waldroup, P.W., 1990. Effects of a Buffered Propionic Acid in Diets on the
Performance of Broiler Chickens and on Microflora of the Intestine and Carcass.
Poultry Science.69, 818-826.
Jacela. J. Y., DeRouchey. J. M., Tokach. M. D., Goodband. R. D., Nelssen. J. L., Renter. D.
G., Dritz. S. S., 2010. Practice tip Peer reviewed feed additives for swine: Fact sheets–
prebiotics and probiotics, and phytogenics. Journal of Swine Health and Production.
18, 132-136.
Jacob, J.P., Pescatore, A.J., 2014. Barley beta-glucans in poultry diets. Annals of
Translational Medicine. 2, 20-26.
Jang, I. S., Ko, Y. H., Yang, H. Y., Ha, J. S., Kim, J. Y., Kang, S. Y., Yoo, D. H., Nam, D.
S., Kim, D. H., Lee, C. Y., 2004. Influence of essential oil components on growth

203
performance and the functional activity of the pancreas and small intestine in broiler
chickens. Asian-Australasian Journal of Animal Sciences. 17, 394–400.
Jarosz, Ł., Marek, A., Grądzki, Z., Laskowska, E., Kwiecień, M., 2019. Effect of zinc sulfate
and zinc glycine chelate on concentrations of acute phase proteins in chicken serum
and liver tissue. Biological trace element research. 187, 258-272.
Jerzsele, A., Szeker, K., Csizinszky, R., Gere, E., Jakab, C., Mallo, J., Galfi, P., 2012.
Efficacy of Protected Sodium Butyrate, a Protected Blend of Essential Oils, Their
Combination, and Bacillus Amyloliquefaciens Spore Suspension against Artificially
Induced Necrotic Enteritis in Broilers. Poultry Science. 91, 837-843.
Jia, W., Slominski, B.A., Bruce, H.L., Blank, G., Crow, G., Jones, O., 2009. Effects of diet
type and enzyme addition on growth performance and gut health of broiler chickens
during subclinical Clostridium perfringens challenge. Poultry Science. 88, 132-140.
Jiménez-Moreno, E., González-Alvarado, J. M., Lázaro, R., Mateos, G. G., 2009. Effects of
type of cereal, heat processing of the cereal, and fiber inclusion in the diet on gizzard
pH and nutrient utilization in broilers at different ages. Poultry Science. 88, 1925-
1933.
Johnson, I.T., Gee, J.M., Price, K., Curl, C., Fenwick, G.R., 1986. Influence of saponins on
gut permeability and active nutrient transport in vitro. Journal of Nutrition. 116, 2270-
2277.
Jones-Hamilton, 2014. Role of pH in Poultry Production. http://joneshamiltonag.com/wp-
content/uploads/Role-of-pH-in-Poultry-Production-FINAL-for-web-use.pdf. (Page
consultée le 20 Jsanvier 2018).
Jørgensen, H., Zhao, X.Q., Knudsen, K.E.B., Eggum, B.O., 1996. The influence of dietary
fiber source and level on the development of the gastrointestinal tract, digestibility
and energy metabolism in broiler chickens. British Journal of Nutrition. 75, 379–395.
Kamińska B., 2003. The effect of various grains in grower diet on broiler performance and
dietetic value of carcass. Annals of Animal Science, Suppl. 2, 185-188.
Kaur, J., Chadha, B.S., Kumar, B.A., Kaur, G.S., Saini, H.S., 2007. Purification and
characterization of βglucosidase from Melanocarpus sp. MTCC 3922. Electronic
Journal of Biotechnology. 10, 260 – 270.
Keerqin, C., Morgan, N. K., Wu, S. B., Swick, R. A., Choct, M., 2017. Dietary inclusion of
arabinoxylooligosaccharides in response to broilers challenged with subclinical
necrotic enteritis. British Poultry Science. 58, 418-424.
Kettunen, H., Peuranen, S., Tiihomen, K., 2001 b. Betaine aids in the osmoregulation of
duodenal epithelium of broiler chicks and affects the movement of water across the
small intestinal epithelium. Comparative Biochemistry and Physiology Part A:
Molecular & Integrative Physiology.129, 595–560.
Kettunen, H., Tiihonen, K., Peuranen, S., Saarinen, M.T., Remus, J.C., 2001 a. Dietary
betaine accumulates in the liver and intestinal tissues and stabilizes the intestinal
epithelial structure in healthy and coccidia infected broiler chicks. Comparative
Biochemistry and Physiology Part A: Molecular & Integrative Physiology.130, 750–
769.
Keyburn, A.L., Boyce, J.D., Vaz, P., Bannam, T.L., Ford, M.E., Parker, D., Di Rubbo, A.,
Rood, J.I., Moore, R.J., 2008. NetB, a new toxin that is associated with avian necrotic
enteritis caused by Clostridium perfringens. PLoS Pathogens. 4, e26.
Khan, R.U., Naz, S., Dhama, K., Karthik, K., Tiwari, R., Abdelrahman, M.M., Alhidary, I.A.,
Zahoor, A., 2016. Direct-fed microbial: beneficial applications, modes of action and

204
prospects as a safe tool for enhancing ruminant production and safeguarding health.
International Journal of Pharmacology.12, 220–231.
Khan, S.H., Iqbal, J., 2016. Recent advances in the role of organic acids in poultry nutrition.
Journal of Applied Animal Research. 44, 359-369.
Kim, J.W., Kim, J. H., Kil, D. Y., 2015. Dietary organic acids for broiler chickens: a review.
Revista Colombiana de Ciencias Pecuarias. 28, 109-123.
Kim, S.W., Mateo, R.D., Yin, Y.L., Wu, G., 2007. Functional amino acids and fatty acids
for enhancing production performance of sows and piglets. Asian-Australasian
Journal of Animal Sciences. 20, 295–306.
Kim, T., Mundt, E., 2011. Metagenomic analysis of intestinal microbiomes in chickens. In
High-throughput Next Generation Sequencing: Methods and Applications. Y. M. Min
Kwon, and S. C. Ricke, ed. Methods in Molecular Biology. 733, 185-194.
Kim,Y., Park, S.C., Wolf, B.W., Hertzler, S.R., 2011. Combination of erythritol and fructose
increases gastrointestinal symptoms in healthy adults. Nutr Res. 31, 836–41.
Kirk, O., Borchert, T.V., Fuglsang, C.C., 2002. Industrial enzyme applications. Current
Opinion in Biotechnology. 13, 345–351.
Klasing, K.C., 1998. Anatomy and physiology of the digestive system. In: Comparative avian
nutrition (ed. by Klasing KC).
Kleessen, B., Hartmann, L., Blaut, M., 2003. Fructans in the diet cause alterations of
intestinal mucosal architecture, released mucins and mucosa-associated
bifidobacteria in gnotobiotic rats. British Journal of Nutrition. 89, 597–606.
Kohl, K.D., 2012. Diversity and function of the avian gut microbiota. Journal of Comparative
Physiology B. 182, 591–602.
Kondo, F., 1988. In vitro lecithinase activity and sensitivity to 22 antimicrobial agents of
Clostridium perfringens isolated from necrotic enteritis of broiler chickens. Research
in Veterinary Science. 45, 337-340.
Kosieradzka, I., Fabijanska, M., Biuletyn, I., 2003. Application of naked oats in feeding pigs
and poultry. Part II. Naked oats in feeding broiler chickens. Institut Roslin d'élevage
et d'acclimatation Pologne. 229, 329-339.
Krogdahl, A., 1985. Digestion and absorption of lipids in poultry. The Journal of nutrition.
115, 675-685.
Kubena, L. F., Bailey, R. H., Byrd, J. A., Young, C. R., Corrier, D. E., Stanker, L. H.,
Rottinghaus, G. E., 2001. Cecal Volatile Fatty Acids and Broiler Chick Susceptibility
to Salmonella typhimurium Colonization as Affected by Aflatoxins and T-2 Toxin.
Poultry Science.80, 411–417.
Kum, S., Eren, U., Onol, A., Sandikci, M., 2010. Effects of dietary organic acid
supplementation on the intestinal mucosa in broilers. Revue de médecine vétérinaire.
10, 463-468.
Kumar, S., Chen, C., Indugu, N., Werlang, G.O., Singh, M., Kim, W.K., Thippareddi, H.,
2018. Effect of antibiotic withdrawal in feed on chicken gut microbial dynamics,
immunity, growth performance and prevalence of foodborne pathogens. PloS ONE.
13,1-23.
Lan, P.T., Hayashi, H., Sakamoto, M., Benno, Y., 2002. Phylogenetic analysis of cecal
microbiota in chicken by the use of 16S rDNA clone libraries. Microbiology and
Immunology. 46, 371-382.
Langhout, D. J., Schutte, J. B., Geerse, C., Kies, A. K., De Jong, J., Verstegen, M. W. A.,
1997. Effects on chick performance and nutrient digestibility of an endo‐xylanase

205
added to a wheat‐and rye‐based diet in relation to fat source. British poultry science.
38, 557-563.
Larbier, M., Leclercq, B., 1992.Nutrition et alimentation des volailles. Absorption des
nutriments. Nutrition et alimentation des volailles, édit. INRA. 38 - 47.
Latha, S., Vinothini, G., John Dickson Calvin, D., Dhanasekaran, D., 2016. In Vitro,
Probiotic Profile Based Selection of Indigenous Actinobacterial Probiont
Streptomyces Sp. Jd9 for Enhanced Broiler Production. Journal of Bioscience and
Bioengineering. 121, 124-131.
Le Douarin, P., 2002. Coccidioses en aviculture - Les coccidies sont bien adaptées aux
élevages modernes. Réussir aviculture.
http://aviculture.reussir.fr/actualites/coccidioses-en-aviculture-les-coccidies-sont-
bien-adaptees-aux-elevages-modernes:24352.html (Page consulté le 2 octobre 2018).
Le Floch, N., Sève, B., 2000. Le devenir des protéines et des acides aminés dans l'intestin du
porc: de la digestion à l'apparition dans la veine porte. INRA Productions Animales.
13, 303-314.
Le Goff, G., 2001. Étude des mécanismes impliqués dans l’évolution de la digestion et
l’utilisation métabolique des fibres alimentaires au cours de la vie du porc.
Conséquences pour l’estimation de la valeur énergétique des aliments. Thèse, École
nationale supérieure agronomique de Rennes, Rennes, France, pp270.
Le Goff, G., Van Milgen, J., Noblet, J., 2002. Influence of dietary fibre level on digestive
and metabolic utilisation of energy in growing and finishing pigs. Animal Research.
51, 245‑259.
Lee, K.W., Lillehoj, H.S., Jeong, W., Jeoung, H.Y., An, D.J., 2011. Avian necrotic enteritis:
experimental models, host immunity, pathogenesis, risk factors, and vaccine
development. Poultry Science. 90,1381–1390.
Lee, S. A., Dunne, J., Febery, E., Brearley, C. A., Mottram, T., Bedford, M. R., 2018.
Exogenous phytase and xylanase exhibit opposing effects on real-time gizzard pH in
broiler chickens. British Poultry Science. 59, 568–578.
Lee, S.I., Park, S.H., Ricke, S.C., 2016. Assessment of cecal microbiota, integron occurrence,
fermentation responses, and Salmonella frequency in conventionally raised broilers
fed a commercial yeast-based prebiotic compound. Poultry Science. 95, 144–53.
Leeson, S., Namkung, H., Antongiovanni, M., Lee, E. H., 2005. Effect of butyric acid on the
performance and carcass yield of broiler chickens. Poultry science. 84, 1418-1422.
Lei, X., Piao, X., Ru, Y., Zhang, H. and Peron, A., 2015. Effect of Bacillus
amyloliquefaciens‐based direct‐fed microbial on performance, nutrient utilization,
intestinal morphology and cecal microflora in broiler chickens. Asian-Australasian
Journal of Animal Sciences. 28, 239-246.
Les Éleveurs La production avicole au Québec, 2019, Rapport annuel 2019.
https://volaillesduquebec.qc.ca/a-propos/publications/rapports-annuels. (Page
consulté 17 Septembre 2020.)
Levkut, M., Revajova, V., Laukova, A., Sevcikova, Z., Spisakova, V., Faixova, Z., 2012.
Leukocytic Responses and Intestinal Mucin Dynamics of Broilers Protected with
Enterococcus Faecium Ef55 and Challenged with Salmonella Enteritidis. Research in
Veterinary Science. 93, 195-201.
Li, Y., Xu, Q., Huang, Z., Lv, L., Liu, X., Yin, C., Yan, H., Yuan, J., 2016. Effect of Bacillus
subtilis CGMCC 1.1086 on the growth performance and intestinal microbiota of
broilers. Journal of Applied Microbiology. 120, 195–204.

206
Liais, J.C., 2006. Méta-analyses. En vue de la modélisation de l’évolution des
Caractéristiques propriétés du muscle chez le bovin. Rapport de stage de fin d’étude
Licence professionnelle de Bio-informatique.
Littell, J. H., Corcoran, J., Pillai, V., 2008. Systematic Reviews and Meta-Analysis. Pocket
guides to social work research methods. New York, NY: Oxford University Press.
202 pp.
Liu, J.R., Lai, S.F., Yu, B., 2007. Evaluation of an intestinal Lactobacillus reuteri strain
expressing rumen fungal xylanase as a probiotic for broiler chickens fed on a wheat
based diet. British Poultry Science. 48, 507-514.
lkin, R. G., Wood, K. V., Hagey, L. R., 1990. Biliary bile acid profiles of domestic fowl as
determined by high performance liquid chromatography and fast atom bombardment
mass spectrometry. Comparative Biochemistry and Physiology B. 96,157-161.
Loncke, C., 2009. Modélisation des relations entre l’alimentation et les flux splanchniques
de nutriments énergétiques chez le ruminant. L’Institut des Sciences et Industries du
Vivant et de l’Environnement (Agro Paris Tech), 456.
Long, J.R., 1973. Necrotic enteritis in broiler chickens. I. A review of the literature and the
prevalence of the disease in Ontario. Canadian Journal of Comparative Medicine. 37,
302–308.
Lu, J., Idris, U., Harmon, B., Hofacre, C., Maurer, J.J., Lee, M.D., 2003. Diversity and
Succession of the Intestinal Bacterial Community of the Maturing Broiler Chicken.
Applied and Environmental Microbiology. 69, 6816-6824.
Lucke, A., Böhm, J., Zebeli, Q., Metzler-Zebeli, B.U., 2018. Dietary Deoxynivalenol
Contamination and Oral Lipopolysaccharide Challenge Alters the Cecal Microbiota
of Broiler Chickens. Frontiers in Microbiology. 9, 804.
M’Sadeq, S.A., Shubioa, W., Swick, R.A., Choct, M., 2015. Towards the control of necrotic
enteritis in broiler chickens with in-feed antibiotics phasing-out
worldwide. Animal Nutrition. 1, 1-11.
Macleod, M.G., Mcneill, L., Bernard, K., Valentine, J., Cowan, A., Wade, A., 2008. Naked
oats: Metabolizable energy yield from a range of varieties in broilers, cockerels and
turkeys. British Poultry Science. 49, 368-377.
Mahan, D.C., Newton, E.A., 1993. Evaluation of feed grains with dried skim milk and
added carbohydrate sources on weanling performance. Journal of Animal Sciences.
71, 3376-3382.
Maria, F., 2005. Sixièmes Journées de la Recherche Avicole, 30-31 mars 2005, Saint-Malo.
Martin, T.G., Smyth, J.A., 2009. Prevalence of netB among some clinical isolates of
Clostridium perfringens from animals in the United States. Veterinary Microbiology.
136, 202-5.
Martinez, Y., Carrión, Y., Rodríguez, R., Valdivié, M., Olmo, C., Betancur, C., Liu, G., Al-
Dhabi, N.A., Duraipandiyan, V., 2015. Growth Performance, Organ Weights and
Some Blood Parameters of Replacement Laying Pullets Fed with Increasing Levels
of Wheat Bran. Brazilian Journal of Poultry Science. 17, 347-354.
Martland, M.F., 1984. Wet litter as a cause of plantar pododermatitis, leading to foot
ulceration and lameness in fattening turkeys. Avian Pathology. 13, 241 – 252.
Mateos, G.G., Jiménez-Moreno, E., Serrano, M.P., Lázaro, R.P., 2012. Poultry response to
high levels of dietary fiber sources varying in physical and chemical characteristics.
Journal Applied Poultry Research. 21, 156-174.

207
Mathlouthi, N., Lallès, J. P., Lepercq, P., Juste, C., Larbier, M., 2002. Xylanase and β-
glucanase supplementation improve conjugated bile acid fraction in intestinal
contents and increase villus size of small intestine wall in broiler chickens fed a rye-
based diet. Journal of Animal Science. 80, 2773-2779.
Maurice, D. V., Jones, J. E., Hall, M. A., Castaldo, D. J., Whisenhunt, J. E., McConnell, J.
C., 1985. Chemical composition and nutritive value of naked oats (Avena nuda L.) in
broiler diets. Poultry Science. 64, 529-535.
Mehdi, Y., Létourneau-Montminy, M.P., Gaucher, M.L., Chorfi, Y., Gayatri, S., Rouissi, T.,
Brar, S.K., Côté, C., Ramirez, A.A., Godbout, S., 2018. Use of antibiotics in broiler
production: Global impacts and alternatives. Animal Nutrition Journal. 4, 170-178.
Messad, F., 2016. Prédiction de la digestibilité des acides aminés des ingrédients utilisés en
alimentation porcine : approche par méta-analyse. Thèse de Doctorat, Université
Laval, Canada.
Metzler, B. U., Mosenthin, R., 2008. A review of interactions between dietary fiber and the
gastrointestinal microbiota and their consequences on intestinal phosphorus
metabolism in growing pigs. Asian-Australasian Journal of Animal Sciences. 21,
603‑615.
Miguel, M. G., 2010. Antioxidant and anti-inflammatory activities of essential oils: a short
review. Molecules. 15, 9252-9287.
Miles, R., Butcher, G., Henry, P., Littell, R., 2006. Effect of antibiotic growth promoters on
broiler performance, intestinal growth parameters, and quantitative morphology.
Poultry Science. 85, 476-485.
Mitsch, P., Zitterl-Eglseer, K., Köhler, B., Gabler, C., Losa, R., Zimpernik, I., 2004. The
effect of two different blends of essential oil components on the proliferation of
Clostridium perfringens in the intestines of broiler chickens. Poultry Science. 83, 669-
675.
Mitscherlich, E., Marth, E.H., 1984.Microbial Survival in the Environment, E. Mitscherlich
and E.H. Marth (eds.). Springer-Verlag, Berlin and Heidelberg.
http://joneshamiltonag.com/wp-content/uploads/Role-of-pH-in-Poultry-Production-
FINAL-for-web-use.pdf. (Page consultée le 18 Mars 2020).
Mohammadagheri, N., Najafi, R., Najafi, G., 2016. Effects of Dietary Supplementation of
Organic Acids and Phytase on Performance and Intestinal Histomorphology of
Broilers. Veterinary Research Forum. 7, 189-195.
Mohd Shaufi, M. A., Sieo, C. C., Chong, C. W., Gan, H. M., Ho, Y. W., 2015. Deciphering
chicken gut microbial dynamics based on high-throughput 16S rRNA metagenomics
analyses. Gut Pathogens. 7, 4.
Moquet, P. C. A., Salami, S. A., Onrust, L., Hendriks, W. H., Kwakkel; R. P., 2018. Butyrate
presence in distinct gastrointestinal tract segments modifies differentially digestive
processes and amino acid bioavailability in young broiler chickens. Poultry Science.
97, 167-176.
Mortimer, I., 2002.The detection of dysbacteriosis. Proceedings of ELANCO Symposium,
Cambridge, UK.
Motta, F. L., Andrade, C. C. P., Santana, M. H. A., 2013. A Review of Xylanase Production
by the Fermentation of Xylan: Classification, Characterization and Applications.
Dans: A. K. C. and S. S. da Silva, éditeur. Intech. Vol. Sustainabl. p. 251‑275.
Mussatto, S.I., Mancilha, I.M., 2007. Non-Digestible Oligosaccharides: A Review.
Carbohydrate Polymers. 68, 587-597.

208
Nahas, J., Lefrançois, M. R., 2001. Effects of feeding locally grown whole barley with or
without enzyme addition and whole wheat on broiler performance and carcass traits.
Poultry Science. 80, 195–202.
National Research Council (NRC), 1994. Nutrient requirements of poultry. 9th ed.
Washington: Academy Press.
National Research Council (NRC), 2001. Nutrient Requirements of Dairy Cattle (7th rev.
ed.), Natl. Acad. Press, Washington, DC.
Nava, G.M., Attene-Ramos, M.S., Gaskins, H.R., Richards, J.D., 2009. Molecular Analysis
of Microbial Community Structure in the Chicken Ileum Following Organic Acid
Supplementation. Veterinary Microbiology. 137, 345-353.
Niewold, T. A., 2007. The nonantibiotic anti-inflammatory effect of antimicrobial growth
promoters, the real mode of action? A hypothesis. Poultry Science. 86, 605–609.
Oakley, B.B., Morales, C.A., Line, J., Berrang, M.E., Meinersmann, R.J., Tillman, G.E.,
Wise, M.G., Siragusa, G.R., Hiett, K.L., Seal, B.S., 2013. The poultry-associated
microbiome: network analysis and farm-to-fork characterizations. PLoS One. 8,
e57190.
Oguey et Bravo. PSA 2015. http://alliednutrition.com.www99.jnb2.host-h.net/wp-
content/uploads/2016/02/A4_Xtract_leaflet_proof.pdf. (Page consulté le 21 juin
2018).
Oke, F., Aslim, B., Ozturk, S., Altundag, S., 2009. Essential oil composition, antimicrobial
and antioxidant activities of Satureja cuneifolia Ten. Food Chemistry. 112, 874–879
Olivère, P., 2010. Bien-être du poulet de chair: détermination des conditions d’ambiance et
des caractéristiques physico-chimiques de la litière responsable de l’apparition de
dermatites de contact en Poulet de chair. Sciences de l’environnement.
Osek M., Janocha A., Klocek B., Wasiłowski Z., Milczarek A., 2003. The influence of
different content of naked oat in plant feed on performance and post-slaughter value
of broiler chicken. Annals of Animal Science, Suppl. 2, 205-208.
Ouhida, I., Pérez, J.F., Piedrafita, J., Gasa, J., 2000. The effects of sepiolite in broiler chicken
diets of high, medium and low viscosity. Productive performance and nutritive value.
Animal Feed Science and Technology. 85, 183-194.
Over, K., Hettiarachchy, N., Johnson, M., Davis, B., 2009. Effect of organic acids and plant
extracts on Escherichia coli O157: H7, Listeria monocytogenes, and Salmonella
typhimurium in broth culture model and chicken meat systems. Journal of Food
Sciences. 74, 515–521.
Palander, S., Nasi, M., Palander, P., 2010. Digestibility and energy value of cereal-based
diets in relation to digesta viscosity and retention time in turkeys and chickens at
different ages estimated with different markers. Archives of Animal Nutrition. 64,
238253
Pan, D., Yu, Z., 2014. Intestinal microbiome of poultry and its interaction with host and diet.
Gut Microbes. 5, 108–119.
Parrish, F.W., Perlin, A.S., Reese, E.T., 1960. Selective enzymolysis of poly-b-Dglucans and
the structure of the polymers. Canadian Journal of Chemistry.38, 2094–2104.
Patel, S., Goyal, A., 2012. The current trends and future perspectives of prebiotics research:
a review. 3 Biotech. 2, 115–125.
Patterson, J. A., Burkholder, K. M., 2003. Application of prebiotics and probiotics in poultry
production. Poultry Science. 82, 627– 631.

209
Payne, J. B., Osborne, J. A., Jenkins, P. K., Sheldon, B. W., 2007. Modeling the growth and
death kinetics of Salmonella in poultry litter as a function of pH and water activity.
Poultry Science. 86:191–201. Gibson, G. R., Roberfroid, M. B., 1995. Dietary
modulation of the human colonic microflora introducing the concept of probiotics.
Journal of Nutrition. 125, 1401-1412.
Peng, Q.Y., Li, J.D., Li, Z., Duan, D.Y., Wu, Y.P., 2016. Effects of Dietary Supplementation
with Oregano Essential Oil on Growth Performance, Carcass Traits and Jejunal
Morphology in Broiler Chickens. Animal Feed Science and Technology. 214, 148-
153.
Pieper, R., Jha, R., Rossnagel, B., Van Kessel, A. G., Souffrant, W. B., Leterme, P., 2008.
Effect of barley and oat cultivars with different carbohydrate compositions on the
intestinal bacterial communities in weaned piglets. FEMS Microbiology Ecology. 66,
556–566.
Pourabedin, M., Guan, L., Zhao, X., 2015. Xylo-oligosaccharides and virginiamycin
differentially modulate gut microbial composition in chickens. Microbiome. 3, 15.
Pouteau, E., Rochat, F., Jann, A., Meirim, I., Sanchez-Garcia, J. L., Ornstein, K., German,
B., Ballevre, O., 2008. Chicory increases acetate turnover, but not propionate and
butyrate peripheral turnovers in rats. British Journal of Nutrition. 99, 287–296.
Prescott, J.F., Valeria, R.P., Gohari, I.M., Lepp, D., Gong, J., 2016. The pathogenesis of
necrotic enteritis in chickens: what we know and what we need to know. Avian
Pathology. 45, 288–294.
Pruesse, E., Quast, C., Knittel, K., Fuchs, B.M., Ludwig, W., Peplies, J., Glockner, F.O.,
2007. SILVA: a comprehensive online resource for quality checked and aligned
ribosomal RNA sequence data compatible with ARB. Nucleic Acids Research. 35,
7188–7196.
Pryde, S. E., Duncan, S. H., Hold, G. L., Stewart, C. S., Flint, H. J., 2002. The microbiology
of butyrate formation in the human colon. FEMS Microbiology Letters. 217, 133–
139.
Puterflam, J., Grontec, E., Derel, R., Pinsard, J. L., Martin, D., Léorat, J., 2007. LES
ENTÉRITES NON SPÉCIFIQUES DU POULET DE CHAIR : INDICATEURS,
LEVIERS DE MAÎTRISE. Septièmes Journées de la Recherche Avicole, Tours, 28
et 29 mars 2007.http://www.journees-de-la-recherche-cunicole.org/PDF/P105-
PUTERFLAM-version%20def.pdf (Page consulté le 04 Avril 2019).
Puyalto, M., Sol, C., Mallo, J.J., 2018. Organic Acids in Today’s Livestock Production:
Usage actuel des acides organiques en productions animales. Animal Nutrition
Conference of Canada (ANAC), cutting edge nutritional strategies for improving
performance, profitability and sustainability, 2-3 May 2018, Edmonton, Canada.
http://toc.proceedings.com/39984webtoc.pdf (Page consulté le 01 Mai 2019).
Raza, M., Biswas, A., Mandal, A.B., Yadav, A.S., 2017. Effect of Dietary Supplementation
of Butyric Acid on Growth Performance and Intestinal Microbial Load in Broiler
Chickens. Animal Feed Science and Technology.17, 353-359.
Rehman, H.U., Vahjen, W., Awad, W.A., Zentek, J., 2007. Indigenous bacteria and bacterial
metabolic products in the gastrointestinal tract of broiler chickens. Archives of
Animal Nutrition. 61, 319–335.
Richard, D., La santé intestinale des vollailles et la lutte contre l’antibiorésistance. Faculty
of Veterinary Medicin. Ghent
University.http://environnement.wallonie.be/cgi/dgrne/aerw/ied/docs/20141126/r-

210
ducatelle-sant-intestinale-des-vollaillesgembloux.pdf. (Page consulté le 05 Avril
2018).
Richards, J.D., Gong, J., de Lange, C.F.M., 2005. The gastrointestinal microbiota and its role
in monogastric nutrition and health with an emphasis on pigs: current understanding,
possible modulations, and new technologies for ecological studies. Canadian Journal
of Animal Sciences. 85, 421–435.
Ricke, S. C., 2015. Potential of fructooligosaccharide prebiotics in alternative and
nonconventional poultry production systems. Poultry Science. 94, 1411–1418.
Rietschel, E. T., Kirikae, T., Schade, F. U., Mamat, U., Schmidt, G., Loppnow, H., Ulmer,
A. J., Zähringer, U., Seydel, U., Di Padova, F., 1994. Bacterial endotoxin: Molecular
relationships of structure to activity and function. Federation of American Societies
for Experimental Biology. 8, 217–225.
Rinttilä, T., Apajalahti, J., 2013. Intestinal microbiota and metabolites −implications for
broiler chicken health and performance. Journal of Applied Poultry Research. 22,
647-658.
Rood, J. I., Adams, V., Lacey, J., Lyras, D., McClane, B. A., Melville, S. B., Uzal, F. A.,
2018. Expansion of the Clostridium perfringens toxin-based typing scheme.
Anaerobe 53, 5-10.
Rood, J.I., Keyburn, A.L., Moore, R.J., 2016. NetB and necrotic enteritis: the hole movable
story. Avian Pathology. 45, 295-301.
Rosen, G.D., 2007. Holo-analysis of the efficacy of Bio-Mos® in broiler nutrition. British
Poultry Science. 48, 21–26.
Ruth, M., R, Field, C., J, 2013. The immune modifying effects of amino acids on gut-
associated lymphoid tissue. Journal of Animal Science and Biotechnology. 4, 1–10.
Saeed, M., Babazadeh, D., Naveed, M., Arain, M. A., Hassan, F. U., Chao, S., 2017.
Reconsidering betaine as a natural anti-heat stress agent in poultry industry: a review.
Tropical animal health and production. 49, 1329-1338.
Saeid, J. M., Mohamed, A. B., Al-baddy, M. A., 2013. Effect of adding Garlic Powder
(Allium sativum) and Black Seed (Nigella sativa) in Feed on Broiler Growth
Performance and Intestinal Wall Structure. 3, 35–42.
Saini, R., Davis, S., Dudley-Cash, W., 2003a. Oregano essential oil reduces the expression
of coccidiosis in broilers. Proc. 52nd West. Poult. Dis. Conf., Sacramento, CA. Vet.
Extension, Univ. Calif., Davis, pp: 97-98.
Saini, R., Davis, S., Dudley-Cash, W., 2003b. Oregano essential oil reduces necrotic enteritis
in broilers. Pages 95-97 dans: Proceedings of the fifty-second western poultry disease
conference. Frame, D. (éd.). Sacramento, Californie, États-Unis.
Salter, D. N., Coates, M. E., 197 l. The influence of the microflora of the alimentary tract on
protein digcstion in the chick. British Journal of Nutrition. 26, 55-69.
Samli, H.E., Senkoylu, N., Koc, F., Kanter, M., Agma, A., 2007. Effects of Enterococcus
Faecium and Dried Whey on Broiler Performance, Gut Histomorphology and
Intestinal Microbiota. Archives of Animal Nutrition. 61, 42-49.
Sansonetti, P., 2015. Microbiologie et maladies infectieuses. L’annuaire du Collège de
France. Cours et travaux, (114), 363-383.
Sarah, Guardia, 2011. Thèse : EFFETS DE PHYTOBIOTIQUES SUR LES
PERFORMANCES DE CROISSANCE ET L’EQUILIBRE DU MICROBIOTE
DIGESTIF DU POULET DE CHAIR. Université François – Rabelais. France.

211
http://www.applis.univ-tours.fr/theses/2011/sarah.guardia_3573.pdf. (Page consultée
le 04 Mai 2018).
Sarao, L. K., Arora, M., Sehgal, V. K., Bhatia, S., 2011. The use of fungal enzymes viz
protease, cellulase, and xylanase for polishing rice. International Journal of Food
Safety. 13, 26-37.
Sauvant D 1992. La modélisation systémique en nutrition. Reproduction Nutrition
Development. 32, 217-230.
Sauvant, D., P. Schmidely, J. J. Daudin, and N. R. St-Pierre. 2008. Meta-analyses of
experimental data in animal nutrition. Animal. 2, 203-1214.
Sauvant, D., Schmidely, P., Daudin, J.J., 2005. Les méta-analyses des données
expérimentales: applications en nutrition animale. INRA Productions Animales. 18,
63-73.
Scott, T. A., Silversides, F. G., Classen, H. L., Swift, M. L., Bedford, M. R., 1998. Effect of
cultivar and environment on the feeding value of Western Canadian wheat and barley
samples with and without enzyme supplementation. Canadian Journal of Animal
Sciences. 78, 649–656.
Seeman, P., Cheng, D., Iles, G.F., 1973. Structure of membrane holes in, osmotic and
saponins hemolysis. Journal of Cell Biology. 56, 519-527.
Sergeant, M.J., Constantinidou, C., Cogan, T.A., Bedford, M, R., Penn, C.W., Pallen, M.J.,
2014. Extensive microbial and functional diversity within the chicken cecal
microbiome. PLoS One. 9, 991-941.
Shang, Y., Kumar, S., Oakley, B., Kim, W.K., 2018. Chicken Gut Microbiota: Importance
and Detection Technology. Frontiers in Veterinary Science. 5,254.
Shapiro, S.K., Sarles, W.B., 1949. Microorganisms in the intestinal tract of normal chickens.
Journal of Bacteriology. 58, 531–544.
Shen, L., 2009. Functional Morphology of the Gastrointestinal Tract. In: Molecular
Mechanisms of Bacterial Infection via the Gut. Ed. Sasakawa, C., Springer,
Heidelberg. 337, 1-35.
Shires, A., Thompson, J.R., Turner, B.V., Kennedy, P.M., Goh, Y.K., 1987. Rate of passage
of corncanola meal and corn-soyabean meal diets through the gastrointestinal tract of
broiler and white legorn chickens. Poultry Science. 66, 289-298
Shojadoost, B., Vince, A.R., Prescott, J.F., 2012. The successful experimental induction of
necrotic enteritis in chickens by Clostridium perfringens: a critical review. Veterinary
Research. 43, 74.
Sikandar, A., Zaneb, H., Younus, M., Masood, S., Aslam, A., Khattak, F., 2017. Effect of
sodium butyrate on performance, immune status, microarchitecture of small intestinal
mucosa and lymphoid organs in broiler chickens. Asian-Australasian Journal of
Animal Sciences. 30, 690–9.
Sklan, D., D. Dubrov, et al., 1975. "51Cr-EDTA, 91Y and 141Ceas Nonabsorbed Reference
630 Substances in the Gastrointestinal Tract of the Chicken." The Journal of
Nutrition. 105, 1549-1552.
Skoufos, I., Giannenas, I., Tontis, D., Bartzanas, T., Kittas, C., Panagakis, P., 2016. Effects
of oregano essential oil and attapulgite on growth performance, intestinal microbiota
and morphometry in broilers. South Africa Journal of Animal Science. 46, 78-88.
Skrivanova, E., Marounek, M., Benda, V., Brezina, P., 2006. Susceptibility of Escherichia
coli, Salmonella sp. and Clostridium perfringens to organic acids and monolaurin.
Veterinární Medicína. 51, 81- 88.

212
Slavin, J., 2013. Fiber and prebiotics: Mechanisms and health benefits. Nutrients. 5, 1417-
35.
Smirnov, A., Tako, E., Ferket, P., Uni, Z., 2006. Mucin gene expression and mucin content
in the chicken intestinal goblet cells are affected by in ovo feeding of carbohydrates.
Journal of Poultry Science. 85, 669-673.
Smits, C. H. M., Annison, G., 1996. Non-starch plant polysaccharides in broiler nutrition–
towards a physiologically valid approach to their determination. World's Poultry
Science Journal. 52, 203-221.
Smits, C. H. M., Veldman, A., Verstegen, M. W. A., Beynen, A. C., 1997. Dietary
carboxymethylcellulose with high instead of low viscosity reduces macronutrient
digestion in broiler chickens. The Journal of Nutrition. 127, 483-487.
Sokol, H., Leducq, V., Aschard, H., Pham, H.P., Jegou, S., Landman, C., Cohen, D., Liguori,
G., Bourrier, A., Nion-Larmurier, I., Cosnes, J., Seksik, P., Langella, P., Skurnik, D.,
Richard, M.L., Beaugerie, L., 2016. Fungal microbiota dysbiosis in IBD. Gut.
66,1039-1048.
Sorum, H. Sunde, M., 2001. Resistance to antibiotics in the normal flora of
animals. Veterinary Research. 32, 227-241.
Souci, S.W., Kirchhoff, E., 2008. Food composition and nutrition tables. 7th ed. Stuttgart
Germany: Medpharm Scientific Publ.
Spring, P., Wenk, C., Dawson, K.A., Newman, K.E., 2000. The effects of dietary
mannaoligosaccharides on cecal parameters and the concentrations of enteric bacteria
in the ceca of salmonella-challenged broiler chicks. Poultry Science. 79, 205-211.
Sproston, N. R., Ashworth, J. J., 2018. Role of C-reactive protein at sites of inflammation
and infection. Frontiers in immunology. 9, 754.
St-Amand, A., Saint-Jacques, M. C., 2013. Comment faire? une méta-analyse, méthode
agrégative de synthèse des connaissances. Université Laval, Faculté des sciences
sociales, Centre de recherche sur l'adaptation des jeunes et des familles à risque
(JEFAR).
Stanley, D., Hughes, R.J., Moore, R.J., 2014. Microbiota of the chicken gastrointestinal tract:
influence on health, productivity and disease. Applied Microbiology and
Biotechnology. 98, 4301–4310.
Statistique Canada, 2020, Consommation de viande et protéine animale par habitant au
Canada(Aliments disponible par personne, par année).
https://www.agr.gc.ca/fra/production-animale/information-sur-le-marche-de-la-
volaille-et-des-oeufs/indicateurs-de-l-industrie/consommation-par-
habitant/?id=1384971854413. (Page consulté 17 Septembre 2020.)
Steyerberg, E. W., Harrell Jr, F. E., Borsboom, G. J., Eijkemans, M. J. C., Vergouwe, Y.,
Habbema, J. D. F., 2001. Internal validation of predictive models: efficiency of some
procedures for logistic regression analysis. Journal of Clinical Epidemiology. 54,
774- 781.
St-Pierre, N., 2001. Invited review: Integrating quantitative findings from multiple studies
using mixed model methodology1. Journal of dairy science. 84, 741-755.
Suez, J., Zmora, N., Zilberman-Schapira, G., Mor, U., Dori-Bachash, M., Bashiardes, S., et
al. 2018. Post-antibiotic gut mucosal microbiome reconstitution is impaired by
probiotics and improved by autologous FMT. Cell. 174, 1406–1423.

213
Sultan, A., Ullah, T., Khan, S., Khan, R.U., 2015. Effect of organic acid supplementation on
the performance and ileal microflora of broiler during finishing period. Pakistan
Journal of Zoology. 47, 635–639.
Sundburg, B., Petcersson, D., Aman, P., 1996. Nutritional properties of fibre-rich barley
products fed to broiler chickens. Journal of the Science of Food and Agriculture. 67,
469-476.
Sunkara, L.T., Achanta, M., Schreiber, N.B., Bommineni, Y.R., Dai, G., Jiang, W., Lamont,
S., Lillehoj, H.S., Beker, A., Teeter, R.G., 2011. Butyrate enhances disease resistance
of chickens by inducing antimicrobial host defense peptide gene expression. PLoS
One. 6, 227-225.
Susan, E. O., 2014. ANATOMIE ET PHYSIOLOGIE DU TRACTUS
GASTROINTESTINAL AVIAIRE. https://lafeber.com/vet/wp-
content/uploads/Note-de-Anatomie-et-Physiologie-du-Tractus-Gastro-Intestinal-
Aviaire.pdf. (Page consultée le 18 Mars 2020).
Svihus, B., Gullord, M., 2002. Effect of chemical content and physical characteristics on
nutritional value of wheat, barley and oats for poultry. Animal Feed Science and
Technology. 102, 71-92.
Świątkiewicz, S., Arczewska-Włosek, A., Józefiak, D., 2014. Immunomodulatory efficacy
of yeast cell products in poultry: a current review. World's Poultry Science Journal.
70, 57-68.
Szymczyk B., Hanczakowski P., Szczurek W., 2005. Performance and intestinal viscosity in
broilers fed diets containing dehulled or naked oats and enzymes. Journal of Animal
and Feed Sciences.14, 491-494.
Szymczyk, B., Hanczakowski, P., Szczurek, W., Frys-Zurek, M., 2007. Effect of naked oat
and enzymes in diets for broiler chickens on quality, fatty acid profile and oxidative
stability of breast muscle. Polish Journal of Food and Nutrition Sciences. 57, 541-
545.
Taylor, R. D., Jones, G. P. D., 2004a. The influence of whole grain inclusion in pelleted
broiler diets on proventricular dilatation and ascites mortality. British Poultry
Science. 45, 247–254.
Taylor, R. D., Jones, G. P. D., 2004b. The incorporation of whole grain into pelleted broiler
chicken diets. II. Gastrointestinal and digesta characteristics. British Poultry Science.
45, 237–246.
Teng, P. Y., Chang, C. L., Huang, C. M., Chang, S. C., Lee, T. T., 2017. Effects of solid-
state fermented wheat bran by Bacillus amyloliquefaciens and Saccharomyces
cerevisiae on growth performance and intestinal microbiota in broiler chickens.
Italian Journal of Animal Science. 16, 552-562.
Teng, P. Y., Kim, W. K., 2018. Review: Roles of Prebiotics in Intestinal Ecosystem of
Broilers. Frontiers in Veterinary Science. 5.
Thomke, S., Elwinger, K., 1998. Growth promotants in feeding pigs and poultry. I. Growth
513 and feed efficiency responses to antibiotic growth promotants. Annales de
zootechnie. 47, 85-97.
Tiihonen, K., Kettunen, H., Bento, M. H. L., Saarinen, M., Lahtinen, S., Ouwehand, A. C.,
Schulze, H., Rautonen, N., 2010. The effect of feeding essential oils on broiler
performance and gut microbiota. British Poultry Science.51, 381-392.
Timbermont, L., Haesebrouck, F., Ducatelle, R., Van Immerseel, F., 2011. Necrotic enteritis
in broilers: an updated review on the pathogenesis. Avian Pathology. 40, 341-347.

214
Torrallarodona, D., Llaurado, L. L., Roura, E., Matas, J., Fort, F., 2000. Enhancement of the
performance of 21 d old weanling pigs with the addition of feed flavors. In Abstracts
of the 51st Annual Meeting of the EAAP. (Ed. J. A. M. van Arendonk, A. Hofer, Y.
van der Honing, F. Mdec, K. Sejrsen, D. Pullar, L. Bodin, J. A. Fernandez and E. W.
Bruns). Wageningen Pers, Wageningen, p. 346.
Uni, Z., Geyra, A., Ben-Hur, H., Sklan, D., 2000. Small intestinal development in the young
chick: Crypt formation and enterocyte proliferation and migration. British Poultry
Science. 41, 544-551.
Uni, Z., Noy, Y., Sklan, D., 1995, Posthatch changes in morphology and function of the small
intestines in heavy and light strain chicks. Poultry Science. 74, 1622–1629.
Uzal, F.A., McClane, B.A., Cheung, J.K., Theoret, J., Garcia, J.P., Moore, R.J., Rood, J.I.,
2015. Animal models to study the pathogenesis of human and animal Clostridium
perfringens infections. Veterinary Microbiology. 179, 23-33.
Vaillant, R., 2010. Linear regression diagnostics for survey data. SSC Annual Meeting,
Proceedings of the Survey Methods Section, 1-
10.https://ssc.ca/sites/default/files/data/Members/public/about/Awards/Survey/2010
_Proceedings/SSC2010_RValliant.pdf. (Page consulté le 02 Mai 2019).
Van Craeyveld, V., Swennen, K., Dornez, E., Van de Wiele, T., Marzorati, M., Verstraete,
W., Delaedt, Y., Onagbesan, O., 2008. Structurally different wheat-derived
arabinoxylooligosaccharides have different prebiotic and fermentation properties in
rats. The Journal of Nutrition. 138, 2348–2355.
Van der Beek, C. M, Dejong, C. H., Troost, C F. J., Masclee, A. M., Lenaerts, k., 2017. Role
of short-chain fatty acids in colonic inflammation, carcinogenesis, and mucosal
protection and healing. Nutrition Reviews. 75, 286–305.
Van der Kils, J.D., Verstegen, M.W.A., De Wit, W., 1990. Absorption of minerals and
retention time of dry matter in the gastrointestinal tract of broilers. Poultry Science.
69, 2185-2194.
Van der Wielen, P.W., Keuzenkamp, D.A., Lipman, L.J., van Knapen, F., Biesterveld, S.,
2002. Spatial and temporal variation of the intestinal bacterial community in
commercially raised broiler chickens during growth. Microbial Ecology. 44, 286–
293.
Van Immerseel, F., Boyen, F., Gantois, I., Timbermont, L., Bohez, L., Pasmans, F.
Haesebrouck, F., Ducatelle, R., 2005. Supplementation of coated butyric acid in the
feed reduces colonization and shedding of Salmonella in poultry. Poultry Science. 84,
1851–1856.
Van Immerseel, F., Fievez, V., De Buck, J., Pasmans, F., Martel, A., Haesebrouck, F.,
Ducatelle, R., 2004. Microencapsulated short-chain fatty acids in feed modify
colonization and invasion early after infection with Salmonella Enteritidis in young
chickens. Poultry Science. 83, 69–74.
Vermeulen, K., Verspreet, J., Courtin, C.M., Haesebrouck, F., Ducatelle, R., Van Immerseel,
F., 2017. Reduced particle size wheat bran is butyrogenic and lowers Salmonella
colonization, when added to poultry feed. Veterinary Microbiology. 198, 64-71.
Villate, D., 2001, Les maladies des volailles. Edition : INRA. S. Beghou. Appareil digestif
dela poule: particularités anatomo-physiologiques. L'appareil digestif. pp : 27-38.
Villettaz, J. C., 1984. Les enzymes en oenologie. Bull. L’OIV. 635, 19-29.
Voreades, N., Kozil, A., Weir, T.L., 2014. Diet and the development of the human intestinal
microbiome. Frontiers in Microbiology. 5, 494.

215
Waite, D., Taylor, M., 2014. Characterising the avian gut microbiota: membership, driving
influences and potential function. Frontiers Microbiology. 5, 223.
Waite, D., Taylor, M., 2015. Exploring the avian gut microbiota: current trends and future
directions. Frontiers Microbiology. 6, 673.
Ward, A.T., Marquardt, R.R., 1988. Effect of various treatments on the nutritional value of
ray or ray fractions. Brit. Poultry Science. 29, 709–720.
Wei, S., Morrison, M., Yu, Z., 2013. Bacterial census of poultry intestinal microbiome.
Poultry Science. 92, 671–683.
Wenk, C., 2001. The role of dietary fibre in the digestive physiology of the pig. Animal Feed
Science and Technology. 90, 21‑33.
Weurding, R.E., Veldman, A., Veen, W.A.G., van der Aar, P.J., Verstegen, M.W.A., 2001.
Starch digestion rate in the small intestine of broiler chickens differs among
feedstuffs. The Journal of Nutrition. 131, 2329-2335.
Wexler, H.M., 2007. Bacteroides: the good, the bad, and the nitty-gritty. Clinical
Microbiology Reviews. 20, 593–621.
White, W.B., Bird, H.R., Sunde, M.L., Prentice, N., Burger, W.C., Marlett, J.A., 1981. The
viscosity interaction of barley beta-glucan with Trichoderma viride cellulase in the
chick intestine. Poultry Science. 60, 1043-8.
WHO, 2001. Food and Agriculture Organization of the United Nations/World Health
Organization (Fao/Who). Health and Nutritional Properties of Probiotics in Food
Including Powder Milk with Live Lactic Acid Bacteria [Internet]. Report of a Joint
Fao/Who Expert Consultation. http://www.fao.org/3/a-a0512e.pdf. (Page consulté le
08 janvier 2018).
Wielen, P.W.J.J.V.D., Biesterveld S., Notermans S., Hofstra H., Urlings B.A.P., Knapen
F.V., 2000. Role of volatile fatty acids in development of the cecal microflora in
broiler chickens during growth. Applied and Environmental Microbiology. 66, 2536-
2540.
Williams, R.B., 2005. Intercurrent coccidiosis and necrotic enteritis of chickens: rational,
integrated disease management by maintenance of gut integrity. Avian Pathology. 34,
159-80.
Windisch, W., Kroismayr, A., 2006. The effect of phytobiotics on performance and gut
function in monogastrics. World nutrition forum: The future of animal nutrition. 85-
90.
Windisch, W., Schedle, K., Plitzner, C., Kroismayer, A., 2008. Use of phytogenetic products
as feed additives for swine and poultry. J.ournal of Animal Science. 86, E140-E148.
Wise, M.G., Siragusa, G.R., 2007. Quantitative analysis of the intestinal bacterial community
in one- to three-week-old commercially reared broiler chickens fed conventional or
antibiotic-free vegetable-based diets. Journal of Applied Microbiology I. 102, 1138–
1149.
Wolf, F.M., 1986. Meta-analysis. Quantitative methods for research synthesis. Beverly Hills,
California, Sage Publications, 1986. 65 p. (Quantitative Applications in the Social
Sciences Number 07-059.)
Xin, Y., Fangyu, L., Hongliang, X., Chengbo, Y., Xiaojun, Y., 2018. Impact of essential oils
and organic acids on the growth performance, digestive functions and immunity of
broiler chickens. Animal Nutrition. 4, 388- 393.

216
Xu, Z., Hu, C., Xia, M., Zhan, Wang, M., 2003. Effects of dietary fructooligosaccharide on
digestive enzyme activities, intestinal microflora and morphology of male broilers.
Journal of Poultry Science. 82, 1030-1036.
Yang, Y., Iji, P. A., Kocher, A., Mikkelsen, L. L., Choct, M., 2007. Effects of
mannanoligosaccharide on growth performance, the development of gut microflora
and gut function of broiler chickens raised on new litter. Journal of Applied Poultry
Research. 16, 280-288.
Yang, Y., Iji, P., Choct, M., 2009.Dietary modulation of gut microflora in broiler chickens:
a review of the role of six kinds of alternatives to in-feed antibiotics. World's Poultry
Science. 65, 97-114.
Yasar, S., Forbes, J.M., 2000. Enzyme supplementation of dry and wet wheat-based feeds
for broiler chickens: performance and gut responses. British Journal of Nutrition. 84,
297–307.
Yegani, M., Korver, D. R., 2008. Factors affecting intestinal health in poultry. Poultry
science. 87, 2052-2063.
Zeisel, S.H., Mar, M.H., Howe, J.C., Holden, J.M., 2003. Concentrations of choline-
containing compounds and betaine in common foods. Journal of Nutrition.133, 1302–
1307.
Zhang, W.F., Li D.F., Lu W.Q., Yi G.F., 2003. Effects of isomalto-oligosaccharides on
broiler performance and intestinal microflora. Poultry Science.82, 657-663.
Zhang, Z.F., Cho, J.H., Kim, I.H., 2013. Effects of Bacillus Subtilis Ubt-Mo2 on Growth
Performance, Relative Immune Organ Weight, Gas Concentration in Excreta, and
Intestinal Microbial Shedding in Broiler Chickens. Livestock Science. 155, 343-347.
Zhao, M., Jiang, B., Hang, H., Fang, Y., Jiang, F., Phillips, G. O., 2013. Efficient induction
of inulin fructotransferase by inulin and by difructose anhydride III in Arthrobacter
aurescens SK 8.001. European Food Research and Technology. 236:991–998.
Zhu, X.Y., Zhong, T.Y., Pandya, Y., Joerger, R.D., 2002. 16S rRNA-based analysis of 399
microbiota from the cecum of broiler chickens. Applied and Environmental
Microbiology. 68, 124-137.