
MODELADO CINÉTICO DEL MICOPARASITISMO … · MODELADO CINÉTICO DEL MICOPARASITISMO POR...
14
32 MODELADO CINÉTICO DEL MICOPARASITISMO POR Trichoderma harzianum CONTRA Cladosporium cladosporioides AISLADO DE FRUTOS DE CACAO (Theobroma cacao) KINETIC MODELING OF MYCOPARASITISM BY Trichoderma harzianum AGAINST Cladosporium cladosporioides ISOLATED FROM COCOA FRUITS (Theobroma cacao) Teresa Romero-Cortes 1 , Pablo Antonio López-Pérez 1 , Mario Ramírez-Lepe 2 y Jaime Alioscha Cuervo-Parra 1* 1 Escuela Superior de Apan, Carretera Apan-Calpulalpan, Km. 8, Chimalpa Tlalayote s/n, Colonia Chimalpa, Apan, Hgo., México. C.P. 43900. 2 Unidad de Investigación y Desarrollo en Alimentos,2Instituto Tecnológico de Veracruz, Av. Miguel Angel de Quevedo 2779, Col Formando Hogar. C.P. 91897. Veracruz, Ver. México. * Autor para correspondencia E-mail: [email protected] RESUMEN Cepas de Cladosporium aisladas de frutos de Theobroma cacao e identificadas como C. cladosporioides se confrontaron contra Trichoderma harzianum VSL291 por medio de experimentos de confrontación. El Índice de Biocontrol (IBC) de T. harzianum VSL291 sobre C. cladosporioides observado a los siete días fue de 76,29%. Las mayores actividades proteolíticas detectadas en la cepa antagonista con paredes celulares de C. cladosporioides, Colletotrichum gloeosporioides y Corynespora cassicola se asociaron con un aumento de las actividades de las enzimas β-1,3 glucanasas, quitinasas, lipasas, proteasas y xilanasas. Estos resultados sugieren que las enzimas proteolíticas de acuerdo a su grado de inducción podrían participar en el efecto antagonista de T. harzianum VSL291 contra los hongos confrontados. Por otra parte, los valores más altos del IBC sugieren que las cepas de Trichoderma podrían tener la capacidad para antagonizar de manera más eficiente contra los patógenos fúngicos de este cultivo. Finalmente, se desarrolló un modelo cinético no estructurado bajo un sistema de ecuaciones diferenciales acopladas en función del tiempo basado en el mecanismo del micoparasitismo. Este modelo predice las concentraciones de las actividades enzimáticas de Trichoderma (β-1,3 glucanasas, quitinasas, lipasas, proteasas, xilanasas) con las paredes celulares de C. cladosporioides. Los parámetros cinéticos se ajustaron a los datos experimentales y se validaron por un coeficiente de correlación. Palabras clave: micoparasitismo simulado, Theobroma cacao, caracterización molecular, parámetros cinéticos. ABSTRACT Cladosporium strains isolated from Theobroma cacao pods identified as C. cladosporioides were confronted against Trichoderma harzianum VSL291 strain by confrontation experiments. Biocontrol index (IBC) of T. harzianum VSL 291 on C. cladosporioides observed at seven days was 76.29%. The highest proteolytic activities detected in the antagonistic strain with C. cladosporioides, Colletotrichum gloeosporioides and Corynespora cassicola cell walls were associated with increased enzymatic activities of β-1,3 glucanases, chitinases, lipases and proteases. These results suggest that proteolytic enzymes according to their degree of induction could participate in the antagonistic effect of T. harzianum VSL291 against the fungi tested. Moreover, the highest IBC values obtained suggest that the Trichoderma strains could be capable of antagonizing more efficiently against fungal ISSN 0719-3882 print ISSN 0719-3890 online Chilean J. Agric. Anim. Sci., ex Agro-Ciencia (2015) 31(3): 32-45. Recibido: 01 septiembre 2015. Aceptado: 23 noviembre 2015.
Transcript of MODELADO CINÉTICO DEL MICOPARASITISMO … · MODELADO CINÉTICO DEL MICOPARASITISMO POR...

Chilean J. Agric. Anim. Sci., ex Agro-Ciencia (2016) 32(1):32
MODELADO CINÉTICO DEL MICOPARASITISMO POR Trichoderma harzianum CONTRA Cladosporium cladosporioides AISLADO DE FRUTOS
DE CACAO (Theobroma cacao)
KINETIC MODELING OF MYCOPARASITISM BY Trichoderma harzianum AGAINST Cladosporium cladosporioides ISOLATED FROM COCOA FRUITS
(Theobroma cacao)
Teresa Romero-Cortes1, Pablo Antonio López-Pérez1, Mario Ramírez-Lepe2 y Jaime Alioscha Cuervo-Parra1*
1 Escuela Superior de Apan, Carretera Apan-Calpulalpan, Km. 8, Chimalpa Tlalayote s/n, Colonia Chimalpa, Apan, Hgo., México. C.P. 43900.
2 Unidad de Investigación y Desarrollo en Alimentos,2Instituto Tecnológico de Veracruz, Av. Miguel Angel de Quevedo 2779, Col Formando Hogar. C.P. 91897. Veracruz, Ver. México.
* Autor para correspondencia E-mail: [email protected]
RESUMEN
Cepas de Cladosporium aisladas de frutos de Theobroma cacao e identificadas como C. cladosporioides se confrontaron contra Trichoderma harzianum VSL291 por medio de experimentos de confrontación. El Índice de Biocontrol (IBC) de T. harzianum VSL291 sobre C. cladosporioides observado a los siete días fue de 76,29%. Las mayores actividades proteolíticas detectadas en la cepa antagonista con paredes celulares de C. cladosporioides, Colletotrichum gloeosporioides y Corynespora cassicola se asociaron con un aumento de las actividades de las enzimas β-1,3 glucanasas, quitinasas, lipasas, proteasas y xilanasas. Estos resultados sugieren que las enzimas proteolíticas de acuerdo a su grado de inducción podrían participar en el efecto antagonista de T. harzianum VSL291 contra los hongos confrontados. Por otra parte, los valores más altos del IBC sugieren que las cepas de Trichoderma podrían tener la capacidad para antagonizar de manera más eficiente contra los patógenos fúngicos de este cultivo. Finalmente, se desarrolló un modelo cinético no estructurado bajo un sistema de ecuaciones diferenciales acopladas en función del tiempo basado en el mecanismo del micoparasitismo. Este modelo predice las concentraciones de las actividades enzimáticas de Trichoderma (β-1,3 glucanasas, quitinasas, lipasas, proteasas, xilanasas) con las paredes celulares de C. cladosporioides. Los parámetros cinéticos se ajustaron a los datos experimentales y se validaron por un coeficiente de correlación.
Palabras clave: micoparasitismo simulado, Theobroma cacao, caracterización molecular, parámetros cinéticos.
ABSTRACT
Cladosporium strains isolated from Theobroma cacao pods identified as C. cladosporioides were confronted against Trichoderma harzianum VSL291 strain by confrontation experiments. Biocontrol index (IBC) of T. harzianum VSL 291 on C. cladosporioides observed at seven days was 76.29%. The highest proteolytic activities detected in the antagonistic strain with C. cladosporioides, Colletotrichum gloeosporioides and Corynespora cassicola cell walls were associated with increased enzymatic activities of β-1,3 glucanases, chitinases, lipases and proteases. These results suggest that proteolytic enzymes according to their degree of induction could participate in the antagonistic effect of T. harzianum VSL291 against the fungi tested. Moreover, the highest IBC values obtained suggest that the Trichoderma strains could be capable of antagonizing more efficiently against fungal
177-187.
ISSN 0719-3882 printISSN 0719-3890 online
Chilean J. Agric. Anim. Sci., ex Agro-Ciencia (2015) 31(3): 32-45.
Recibido: 01 septiembre 2015. Aceptado: 23 noviembre 2015.

33Romero-Cortes, T. et al. Micoparasitismo por Trichoderma contra Cladosporium aislado de cacao
pathogens of these crops. Finally, a kinetic model based on unstructured mycoparasitism mechanism was developed; this model was designed under a system of coupled differential equations with time. This model can predict the concentrations of the enzyme activities of Trichoderma (β-1, 3 glucanases, chitinases, lipases, proteases, xylanases) in the cell walls of C. cladosporioides. Kinetic parameters were fitted to the experimental data and validated by a correlation coefficient.
Key words: simulated mycoparasitism, Theobroma cacao, molecular characterization, kinetic parameters.
INTRODUCCIÓN
El cacao (Theobroma cacao L.) es un cultivo de gran valor comercial a nivel mundial, cuyo fruto se procesa para ser utilizado en diversos productos. Sin embargo, el gran volumen de residuos que generalmente permanecen disper-sos en la plantación y la acumulación de hume-dad favorecen el crecimiento y la propagación de hongos fitopatógenos. Las enfermedades de mayor impacto en el cultivo son la “moniliasis” causada por Moniliophthora roreri (Cuervo-Parra et al., 2011b), la “escoba de bruja” causada por Moniliophthora perniciosa (Kilarul et al., 2007) y la “mancha negra” causada por varias especies del género Phytophthora (Djocgoue et al., 2006). No obstante, otras enfermedades no específicas del cultivo del cacao como “la roña” o “podredum-bre de Cladosporium” causada por varias especies del género Cladosporium, pueden ocasionar pérdi-das económicas cuando no son controladas (Rias-cos et al., 2011). La podredumbre de Cladosporium puede infestar y matar plántulas y ramas, retrasar la producción y floración, y afectar la calidad de los frutos, y aunque el hongo no causa deterioro de la pulpa del grano, la apariencia y aceptación de los frutos en el mercado se ve reducido (Goes, 1998). De las 772 especies que comprende este gé-nero, Cladosporium herbarum y C. cladosporioides se encuentran entre las especies más predominantes de los ambientes internos y externos, prevalecien-do en materia orgánica en descomposición, por lo cual, comúnmente se encuentran como conta-minantes de alimentos (Schubert et al., 2007). En los últimos años, el género Cladosporium ha sido estudiado morfológica y molecularmente (Braun et al., 2008; Bensch et al., 2010).
Por otro lado, la capacidad antagonista de Tri-choderma es muy variable, el micoparasitismo que presenta es un proceso complejo que incluye una serie de eventos sucesivos. Cuando el antagonista hace contacto físico con el fitopatógeno, sus hifas se enrollan alrededor de éste o se le adhieren por medio de estructuras especializadas (Manocha y Sahai, 1993; Cuervo-Parra et al., 2011a). La inte-racción que presenta Trichoderma con su huésped es específica y está controlada por lectinas, pre-sentes en su pared celular. Las observaciones al microscopio electrónico muestran que Trichoder-
ma produce y secreta enzimas micolíticas respon-sables de la degradación parcial de la pared ce-lular de su huésped (Cuervo-Parra et al., 2011a).
Varios autores sugieren que el micoparasitis-mo es el mecanismo principal de acción de Tri-choderma por medio del cual penetra a las células del patógeno causándole daño extensivo, como la degradación de la pared celular, la retracción de la membrana plasmática y la desorganización del citoplasma. También actúa sobre la replicación celular al inhibir la germinación de esporas y la elongación del tubo germinativo (Harman et al., 1981; Agamez et al., 2009).
Las especies de T. polysporum, T. viride y T. harzianum han mostrado un eficiente biocontrol contra diferentes aislados de Cladosporium herba-rum (Barbosa et al., 2001). La confrontación direc-ta entre T. harzianum contra Cladosporium sp. fue estudiada por Mokhtar y Dehimat (2012), quienes reportaron una inhibición del 57% al cuarto día, y por la acción de los compuestos volátiles produ-cidos por T. harzianum a los 6 días. Por otra parte, Ahmad et al. (2012) reportaron la inhibición del crecimiento de Cladosporium spherospermun en un 72,2% en un periodo de 10 días.
En México la mayor parte de los trabajos se ha centrado en el estudio de los patógenos que cau-san enfermedades en el cultivo de cacao; es decir, Moniliophthora roreri y Phytophthora spp. (Phi-llips-Mora et al., 2006). En particular en la región Chontalpa, ubicada en el centro-oeste del estado de Tabasco, México, los hongos son los más agre-sivos con el cultivo. Desafortunadamente, existen pocos estudios enfocados en la identificación mo-lecular de otras especies de hongos fitopatógenos que afectan las plantaciones. Por otro lado, des-de el punto de vista biotecnológico, los sistemas de micoparasitismo son de gran importancia en los procesos ambientales. Sin embargo, la deter-minación de los parámetros cinéticos utilizando un modelo estructurado es un proceso complejo, debido a la interacción y medición de las con-centraciones de metabolitos intermediarios, bio-masa, ADN y enzimas involucradas (Villadsen et al., 2011). Por esta razón, los parámetros ci-néticos más comúnmente utilizados se estiman a través de un modelo cinético no estructurado (López-Pérez et al., 2013).
Por lo tanto, existe la necesidad de un cono-

Chilean J. Agric. Anim. Sci., ex Agro-Ciencia (2016) 32(1):34 32-45.
cimiento satisfactorio que describa el comporta-miento de la cinética del micoparasitismo basado en un modelo dinámico. Por lo cual, los objetivos de este estudio fueron (i) caracterizar molecular-mente las cepas de hongos aislados de frutos de cacao, (ii) probar la actividad antagonista in vi-tro de Trichoderma harzianum VSL291 frente a los patógenos aislados de las plantas de cacao, y (iii) proponer un modelo cinético no estructurado ba-sado en los datos experimentales del comporta-miento del micoparasitismo.
MATERIALES Y MÉTODOS
Aislamiento de las cepas de hongosCada fragmento de fruto con síntomas de
enfermedad se colocó en una cámara húmeda, posteriormente se incubó a 25°C durante 3 días en ausencia de luz. Las cepas fúngicas fueron ais-ladas de fragmentos de tejido enfermo, donde se observó crecimiento de micelio. Posteriormente, se tomó una parte del micelio crecido en cada uno de los tejidos enfermos con la ayuda de una aguja de disección y fueron resembrados en cajas Petri nuevas. Los cultivos puros se obtuvieron en me-dio PDA (Agar Papa Dextrosa) durante 5 días a temperatura ambiente. Las cepas se conservaron en una suspensión de esporas, en una solución de glicerol al 40% (v/v) y se almacenaron a una temperatura de -74ºC. Las cepas se identificaron como HT-ITV36, HT-ITV86, HT-ITV87, HT-ITV88 y HT-ITV89.
Caracterización molecular La extracción de ADN de las cepas se realizó
siguiendo la metodología propuesta por Kurza-tkowski et al. (1996). Las secuencias de ADN, que contenían la región ITS I/5,8s/ITS II, se amplifica-ron por PCR utilizando los primers ITS1 e ITS4 (Foster et al., 1993; Kendall y Rygiewicz, 2005). Las amplificaciones por PCR se realizaron en un volumen total de reacción de 50 µL, que contenía: 10 µL de buffer, 5x Colorless, 200 μM dNTPSs, 0,2 µM de cada primer, 2,5 unidades de Taq ADN polimerasa, y 10-50 ng de ADN templado. Las re-acciones de PCR se colocaron en un termociclador (TECHNE, modelo 3Prime, Bibby Scientific, Sta-ffordshire, United Kingdom) bajo los siguientes parámetros: 5 min de desnaturalización inicial a 95°C, seguido de 30 ciclos de 1 min de desnaturali-zación a 95°C, 1 min de hibridación de los primers a 57°C, 1 min de extensión a 72°C y un periodo final de extensión de 12 min a 72°C. Los productos resultantes se purificaron con el kit Geneclean® II (Bio101 Inc., California, EE.UU.) de acuerdo con el protocolo del fabricante. Las muestras de ADN fueron enviadas al Instituto de Biotecnología, Cuernavaca, Mor., México, para su secuenciación.
Análisis de las secuencias Las secuencias de ADN obtenidas de las ce-
pas amplificadas fueron ensambladas utilizan-do el programa Chromas 1,45 (School of Health Science, Griffith University, Gold Coast Cam-pus, Southport, Queensland, Australia) y se ali-nearon con el programa ClustalX (Larkin et al., 2007). Las secuencias seleccionadas se sometieron a una búsqueda local con el programa BLASTN (Altschul et al., 1997) contra secuencias dispo-nibles en el GenBank. El análisis filogenético de las secuencias alineadas se realizó por medio del análisis Neighbour-Joining y estuvo soportado por el método Bootstrap con 1000 repeticiones. El árbol filogenético sin raíz generado fue ensam-blado con la ayuda del programa PhyloDraw 0,8 (PhyloDraw, 1999).
Experimento de confrontación de Trichoderma vs C. cladosporioides
Para el experimento de confrontación se uti-lizó como antagonista la cepa de T. harzianum VSL291 contra las cepas de C. cladosporioides, de acuerdo a la técnica propuesta por Szekeres et al. (2006). Discos de 5 mm de diámetro de micelio de T. harzianum y de C. cladosporioides incubados durante 3 días se sembraron en medio PDA en puntos equidistantes. Las cajas se incubaron a 25°C durante 7 días en ausencia de luz. Cuando se observó la actividad antagonista se tomaron las imágenes digitales a la misma distancia (18 cm) con una cámara (Sony Cyber-shot DSC-P72). El porcentaje de inhibición se calculó utilizando el Índice de Biocontrol (IBC) de acuerdo con la fórmula descrita por Szekeres et al. (2006):
IBC = [A/B] x 100,
dónde A es el área de la colonia de T. harzianum y B es el área total ocupada por las colonias de T. harzianum y la cepa del hongo patógeno. El área del IBC se calculó con el software ImageJ (http://rsbweb.nih.gov/).
Micoparasitismo simulado T. harzianum - hongos fitopatógenos
Para determinar el micoparasitismo simulado de T. harzianum sobre los hongos fitopatógenos se utilizó paredes celulares de C. cladosporioides, Colletotrichum gloeosporioides y Corynespora cassicola como única fuente de carbono. En matraces de 250 mL se colocó 50 mL de medio mínimo (conteniendo: MgSO4·7H2O, 0,24 g; KCl, 0,24 g; NH4NO3, 1,2 g; ZnSO4·7H2O, 0,0024 g; MgCl2·7H2O, 0,0024; K2HPO4, 1,08 g; FeSO4·7H2O, 0,0024 g, pH 5,5; para 1200 mL) y 10 g L-1 de pared celular del micelio molido de C. cladosporioides, C. gloeosporioides y C. cassicola. Cada matraz

35Romero-Cortes, T. et al. Micoparasitismo por Trichoderma contra Cladosporium aislado de cacao
se sembró con una azada de micelio de la cepa VSL291 de T. harzianum y se fermentó durante 3 días. Se tomó muestra cada 24 h y se determinó la actividad enzimática para β-1, 3-glucanasa (Nobe et al., 2003), xilanasa (Rawashdeh et al., 2005), quitinasa (Monreal y Reese, 1969), lipasa (Nawani et al., 1998) y proteasa (Kunitz, 1946). La cuantificación de azúcares reductores se realizó por el método descrito por Miller (1959). Cada experimento se realizó por triplicado. Para evaluar si existió diferencias entre cada uno de los tratamientos (actividades enzimáticas), el análisis estadístico de los datos se hizo mediante un diseño completamente aleatorio con tres repeticiones, usando un análisis de varianza (ANOVA) con el software Statistix9. La comparación de medias se realizó con la prueba de Tukey HSD All-Parwise a una diferencia significativa de P ≤ 0,05.
RESULTADOS Y DISCUSIÓN
Aislamiento de cepasDe un total de ciento veintiocho cepas de hon-
gos aisladas de frutos de cacao con síntomas de moniliasis se identificaron cinco cepas como C. cladosporioides en base a sus características morfo-lógicas (datos no mostrados). Los especímenes del género Cladosporium, ya han sido reportados por varios autores como patógenos de plantas como el cártamo (Carthamus tinctorius; Quintana-Obre-gón et al., 2011), tabaco (Nicotiana tabacum; Wang et al., 2014), remolacha azucarera (Beta vulgaris L. subsp. vulgaris var. altissima Döll; Robeson y Jalal, 1992), maíz (Zea mays; Orole y Adejumo, 2009), trigo (Triticum spp.; Jacyno et al., 1993) y cacao (Theobroma cacao; Amegbedzi, 2013). Esos hongos son los responsables de causar algunas enferme-dades como las manchas foliares (Al-Sadi et al., 2014) y la enfermedad de la roña. Ésta última provoca la muerte de semillas y ramas, la pérdida de flores y disminuye la calidad del fruto (Goes, 1998) causando así pérdidas económicas en las plantaciones de cacao.
La enfermedad causada por estos hongos se diferencia de acuerdo a la planta que infectan y del tipo de síntomas. Por ejemplo, en los cereales, el hongo Cladosporium es responsable de causar la enfermedad llamada el punto negro de los ce-reales; en el género Cucurbita la roña de las cucur-bitáceas; en la papaya (Carica papaya) la roña de la papaya; y en el tomate (Solanum lycopersicum) causa manchas marrones en las hojas (Kryczyński y Weber, 2011).
Caracterización molecular Las cepas HT-ITV86 y HT-ITV88 mostraron
una homología de 99% con la cepa Cc_101 de C. cladosporioides (JX867558) en base a la región ITS
I/5,8s/ITS II. La secuencia de C. cladosporioides (KJ596320) aislada en la India, mostró una homo-logía de 99% con la cepa HT-ITV87. La secuencia de la cepa HT-ITV89 reveló su homología máxi-ma (99%) con la cepa NRRL 66016 de C. cladospo-rioides (KM030316), aislada en Estados Unidos de Norteamérica de plantas de maíz. Por último, la secuencia de la cepa HT-ITV36 reveló su homo-logía máxima (99%) con la cepa NRRL 52418 de C. cladosporioides (GQ221853), aislada de semillas de sorgo (Sorghum vulgare) en los Estados Unidos de Norteamérica. Los resultados obtenidos con el programa BLASTN confirman que las cepas HT-ITV36, HT-ITV86, HT-ITV87, HT-ITV88 y HT-ITV89 depositadas en las bases de datos del GenBank del National Center for Biotechnology Information (NCBI) con los números de acce-so KF040480, KP963942, KP963943, KP963944 y KP9633945, corresponden a C. cladosporioides.
Análisis filogenéticoSe realizó un análisis múltiple de secuencias
de las cepas de C. cladosporioides obteniéndose la secuencia completa de la región ITS I/5,8s/ITS II, junto con los extremos de las subunidades de los genes 18S y 28S. Las secuencias examinadas para la región ITS I/5,8s/ITS II, para las cepas de C. cladosporioides y otras secuencias de Cladosporium relacionadas del GenBank tuvieron una longitud de entre 830 a 1511 pb y la longitud de la región consenso para el alineamiento de todas las se-cuencias fue de 1518 pb.
Para el análisis de la subunidad 5.8S se ob-servaron variaciones muy limitadas al cambio de unas pocas bases en las secuencias de C. clados-porioides aisladas del cacao, lo cual indicó que es una región muy conservada entre las secuencias de Cladosporium estudiadas. Por otro lado, en las subunidades 18S y 28S, se observó una gran va-riabilidad, limitada a mutaciones puntuales, in-serciones o delesiones, predominantemente entre las secuencias de los aislamientos de C. cladospo-rioides.
Dentro de las regiones ITS se observó va-riación en término de delesiones, inserciones y substituciones de bases, especialmente en la re-gión ITS II, aunque muchas regiones conservadas estuvieron presentes en las regiones ITS I e ITS II. En general, la longitud de las secuencias para las regiones ITS I e ITS II, estuvo altamente con-servada en todas las secuencias de las especies de Cladosporium, con variaciones limitadas al cambio de unas pocas bases.
El análisis filogenético de las secuencias de las cepas de C. cladosporioides aisladas del cacao y otras 15 secuencias de Cladosporium relacionadas procedentes del GenBank, generó un árbol por el método Neighbour Joining. El análisis filoge-

Chilean J. Agric. Anim. Sci., ex Agro-Ciencia (2016) 32(1):36 32-45.
nético de las 5 secuencias representativas de C. cladosporioides aisladas de frutos de cacao y otras secuencias de especies relacionadas de las bases de datos del GenBank formaron tres grupos ma-yores (I, II y III) soportados por valores estadís-ticos bootstrap altos de entre 700 a 1000 (Fig. 1).
En el grupo I se agruparon dos subgrupos (C1 y C2). El subgrupo C1 incluyó los aislados KP963942, KP963944 y JX867558, de los cuales las dos primeras corresponden a secuencias de C. cladosporioides aisladas de frutos de cacao, proce-dentes de México y la tercera de la India. El va-lor estadístico bootstrap alto que se obtuvo entre el alineamiento de la cepa HT-ITV88 (GenBank KP963944) y la secuencia de la cepa Cc_101 de C. cladosporioides (JX867558) confirmó la identifica-ción de la cepa HT-ITV88. Por otro lado, aunque el valor estadístico bootstrap que se obtuvo para la cepa HT-ITV86 no fue muy alto (248) su alinea-miento entre secuencias de C. cladosporioides den-tro del subgrupo C1 muestra la probabilidad de pertenecer a esta especie.
Por su parte, en el subgrupo C2 se incluyó los aislados KP963943 y KJ596320, correspondientes a secuencias de C. cladosporioides de México y la
India, respectivamente. En este subgrupo la cepa HT-ITV87 se alineó junto a la secuencia de la cepa TSP1 de C. cladosporioides y se respaldó por un valor estadístico de soporte bootstrap de 523. Por otro lado, el grupo II se dividió en dos subgrupos (C3 y C4).
El subgrupo C3 incluyó los aislados KP963945, KM030316, FR837924 y GQ221852, todos de C. cla-dosporioides (2 Estados Unidos de Norteamérica, 1 México, 1 República Checa). En este subgrupo la cepa HT-ITV89 se alineó con la cepa NRRL 66016 de C. cladosporioides, con un valor de soporte de bootstrap de 1000. El subgrupo C4 incluyó los ais-lados EF679372, EF679390, EF679382 y EF679371, correspondientes a secuencias de C. macrocarpum, C. subtillissimum, C. ossifragi y C. macrocarpum (1 Turquía, 1 Estados Unidos de Norteamérica, 1 Noruega y 1 Rumania), cuyos alineamientos estu-vieron soportados por valores estadísticos boots-trap altos. Por último, el grupo III se dividió en tres subgrupos (C5, C6 y C7).
El subgrupo C5 incluyó los aislados KF040480, GQ221853 y HQ130717, con dos secuencias de C. cladosporioides y una secuencia de Cladosporium sp. procedentes de México, Estados Unidos de
Fig. 1. Árbol filogenético generado por el método Neighbour Joining derivado de los datos de las secuencias de los hongos para la región ITS I/5,8s/ITS II del ARN ribosomal. Los números adyacentes a los puntos de ramificación son los valores bootstrap para 1000 repeticiones.
Fig. 1. Phylogenetic tree generated by the Neighbour Joining method derived from the sequence data for fungal ITS I/5.8s/ITS II region of the RNAr. Numbers adjacent to branch points are bootstrap values from 1000 interactions.

37
América y África, respectivamente. En este sub-grupo la secuencia de la cepa HT-ITV36 se alineó con la secuencia de la cepa NRRL 52418 de C. cladosporioides, soportado por un valor estadísti-co bootstrap de 1000. Dicho resultado estadístico confirma que ésta cepa pertenece a la especie C. cladosporioides. El subgrupo C6 incluyó los ais-lados AY251070, AY251075 y GQ458030 de los cuales la mayoría son secuencias de cepas de C. cladosporioides procedentes de Países Bajos y Ca-nadá, soportados por valores estadísticos altos. Por último, el subgrupo C7 incluyó solo el aislado AY251074 procedente de Países Bajos.
Experimento de confrontación El experimento de confrontación mostró una
zona de inhibición progresiva producida por T. harzianum VSL291 en contra de C. cladosporioides (datos no mostrados), que se incrementó a través del tiempo, acompañada por el ataque del micelio del hongo patógeno (Tabla 1). T. harzianum mos-tró un efecto marcado de híperparasitismo, que fue observado por el crecimiento sobre la colonia del fitopatógeno y por la competencia por espacio y nutrientes (Ezziyyani et al., 2004).
La aplicación de cepas de T. harzianum en fru-tas de uvas y manzanas ha sido reportada redu-ciendo parcialmente la enfermedad causada por el hongo Botrytis cinerea y las protegió por más de dos meses, e igualmente mostró efecto sobre Penicillium expansum (Latorre et al., 1997). Así mismo, se observó que T. harzianum y T. pseudoko-ningi controlaron el desarrollo de Monilinia laxa en ciruela de hueso y B. cinerea en manzanas (Ve-lázquez-del Valle et al., 2008).
Simulación de micopararasitismo T. harzianum secretó en el medio de cultivo
algunas enzimas como la β-1,3-glucanasas (0.58
mU mg-1), quitinasas (0.08 mU mg-1), xilanasas (0.31 mU mg-1), proteasas (0.21 mU mg-1) y lipasas (0.15 mU mg-1), en presencia de paredes celula-res de las cepas HT-ITV36, HT-ITV86, HT-ITV87, HT-ITV88 y HT-ITV89 de C. cladosporioides, y de las cepas HT-ITV41 y HT-ITV32 de C. gloeospo-rioides y C. cassicola, respectivamente. La activi-dad micoparasítica de Trichoderma dependió de la secreción de una mezcla compleja de enzimas hidrolíticas capaces de degradar las paredes ce-lulares de los hongos patógenos (Suarez et al., 2005). En muchos miembros del género Trichoder-ma se ha demostrado su acción antagónica contra patógenos de plantas y la producción de enzimas líticas (Haggag et al., 2006).
Para la producción de quitinasas se observó que a las 12 y 24 h se produjo la actividad enzi-mática mayor (0,078 mU mg-1 ± 0,01 y 0,054 mU mg-1 ± 0,02 de proteína, respectivamente) y para C. gloeosporioides y C. cassicola (0,084 mU mg-1 ± 0,01 y 0,024 mU mg-1 ± 0,015). De la Cruz et al. (1992) obtuvieron valores de actividad específi-ca de entre 670 y 800 mU mg-1 de proteína para la actividad quitinasa con la cepa CECT 2413 de T. harzianum, crecida con paredes celulares de B. cinerea, Gibberella fujikuroi, Rhizoctonia solani, Phytophthtora citrophthora y T. harzianum, que son más altos a los valores obtenidos en este traba-jo. Kucuk y Kivanc (2008) produjeron quitinasas en medios de cultivo líquidos que contenían pa-redes celulares de Gibberella zeae y Aspergillus us-tus como única fuente de carbono con cepas de T. harzianum y reportaron valores similares a los encontrados en este trabajo.
La producción máxima de xilanasas por T. harzianum VSL291 se observó a las 12 h (0,3 mU mg-1 de proteína ± 0,003) y se mantuvo constan-te a una concentración de 0,25 hasta las 96 horas y para C. gloeosporioides y C. cassicola (0,083 mU
Tabla 1. Índice de Biocontrol (IBC) de la cepa VSL 291 contra C. cladosporioides Table 1. Biocontrol index (IBC) of VSL 291 strain against C. cladosporioides
Tiempo de incubación (h) Áreaa T cm-2 ± DE Áreab T + P cm-2 ± DE IBCc (%) 0 0 0 0 24 10,98 ± 0,58 20,78 ± 0,50 52,83 48 16,34 ± 1,13 27,84 ± 1,18 58,69 72 18,23 ± 2,21 29,23 ± 2,20 62,37 96 19,68 ± 1,29 31,18 ± 0,73 63,12 120 23,34 ± 0,63 35,29 ± 1,09 66,14 144 30,12 ± 0,78 42,31 ± 0,33 71,18 168 43,03 ± 1,21 56,22 ± 0,07 76,29
a Área T: área total de la colonia de Trichoderma.b Área T + P: área total ocupada por las colonias de Trichoderma y el hongo patógeno. c IBC del crecimiento del hongo patógeno.
Romero-Cortes, T. et al. Micoparasitismo por Trichoderma contra Cladosporium aislado de cacao
Chilean J. Agric. Anim. Sci., ex Agro-Ciencia (2016) 32(1):38 32-45.
mg-1 ± 0,01 y 0,133 mU mg-1 ± 0,05). Stricker et al. (2006), encontraron que la cepa QM9414 de Hypo-crea jecorina produjo la actividad máxima de xi-lanasa (0,206 ± 0,015 U mg-1 de proteína) a las 48 h utilizando xilano como fuente de carbono. Sin embargo, Seiboth et al. (2003) evaluaron la pro-ducción de xilanasa con H. jecorina utilizando seis sustratos diferentes, encontrando que la mejor actividad específica (0,33 ± 0,05 U mg-1) se obtuvo cuando H. jecorina creció en el medio conteniendo xilitol como sustrato. Ujiie et al. (1991) reporta-ron una actividad específica de xilanasa de 24,4 U mg-1 de proteína para la enzima cruda producida por Trichoderma viride. Otros autores evaluaron las actividades específicas de xilanasa produci-das por cepas nativas y recombinantes (ATX2-12, ATX2-16 y TU-6) de Trichoderma reesei en medio suplementado con glucosa y xilano, y reportan que las actividades específicas de xilanasa a partir de xilano por las cepas ATX2-12, ATX2- 16 y TU-6 fueron 30, 27 y 24 U mg-1 de proteína, respecti-vamente (Kurzatkowski et al., 1996). Rezende et al. (2002) reportaron una actividad específica de xilanasa de 13 U mg-1 de proteína. Los resultados obtenidos en los trabajos anteriores son superio-res a los mostrados en este estudio, ya que en esos trabajos se utilizaron fuentes de carbono específi-cas para la producción de xilanasas.
Para el caso de las proteasas, la actividad máxima se detectó a las 24 h (0,220 mU Tirosina
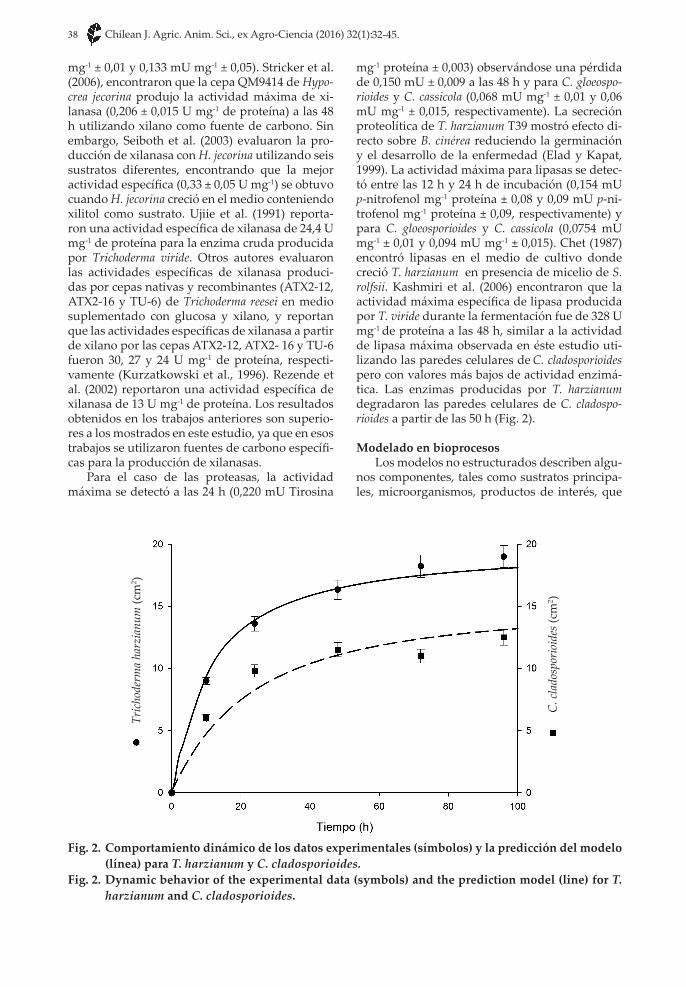
mg-1 proteína ± 0,003) observándose una pérdida de 0,150 mU ± 0,009 a las 48 h y para C. gloeospo-rioides y C. cassicola (0,068 mU mg-1 ± 0,01 y 0,06 mU mg-1 ± 0,015, respectivamente). La secreción proteolítica de T. harzianum T39 mostró efecto di-recto sobre B. cinérea reduciendo la germinación y el desarrollo de la enfermedad (Elad y Kapat, 1999). La actividad máxima para lipasas se detec-tó entre las 12 h y 24 h de incubación (0,154 mU p-nitrofenol mg-1 proteína ± 0,08 y 0,09 mU p-ni-trofenol mg-1 proteína ± 0,09, respectivamente) y para C. gloeosporioides y C. cassicola (0,0754 mU mg-1 ± 0,01 y 0,094 mU mg-1 ± 0,015). Chet (1987) encontró lipasas en el medio de cultivo donde creció T. harzianum en presencia de micelio de S. rolfsii. Kashmiri et al. (2006) encontraron que la actividad máxima específica de lipasa producida por T. viride durante la fermentación fue de 328 U mg-1 de proteína a las 48 h, similar a la actividad de lipasa máxima observada en éste estudio uti-lizando las paredes celulares de C. cladosporioides pero con valores más bajos de actividad enzimá-tica. Las enzimas producidas por T. harzianum degradaron las paredes celulares de C. cladospo-rioides a partir de las 50 h (Fig. 2).
Modelado en bioprocesosLos modelos no estructurados describen algu-
nos componentes, tales como sustratos principa-les, microorganismos, productos de interés, que
Fig. 2. Comportamiento dinámico de los datos experimentales (símbolos) y la predicción del modelo (línea) para T. harzianum y C. cladosporioides.
Fig. 2. Dynamic behavior of the experimental data (symbols) and the prediction model (line) for T. harzianum and C. cladosporioides.
Tric
hode
rma
harz
ianu
m (c
m2 )
C. cl
ados
porio
ides
(cm
2 )

39
permiten limitar el número de variables y pará-metros del sistema (Dochain, 2003). En la fermen-tación por lotes, la dinámica del sistema puede ser descrito por el siguiente modelo:
(1)
dónde nx
m x nK
es el vector de concentraciones;
nx
m x nK es la matriz de coeficientes estequiométricos (m < n);
ni
es el vector de términos cinéticos;
es el tiempo; y (2)
dónde t
es la tasa de crecimiento específico que es una función de diferentes concentraciones de acuerdo al sistema en estudio.
Tales velocidades de reacción varían con el tiempo y por lo general se ven influidas por mu-chos factores físico-químicos y biológicos am-bientales como el sustrato, los microorganismos y las concentraciones de productos, así como el pH, la temperatura, la concentración de oxígeno di-suelto o varios inhibidores del crecimiento micro-biano. Los cuales, para este trabajo representan las concentraciones de las actividades enzimáti-cas de lipasas, glucanasas, proteasas, quitinasas, y xilanasas, de T. harzianum con paredes celulares de C. cladosporioides.
Modelo cinético propuesto
El modelo cinético propuesto representa el mecanismo de control biológico mediante la velo-cidad de crecimiento de los hongos y la secreción de enzimas. El modelo se basó en el balance de materia vía cinéticas no estructuradas represen-
tadas por ecuaciones diferenciales en función del tiempo (modelo dinámico). Para la solución del modelo se diseñó una red de programación gráfi-ca (Model Maker®) representando el mecanismo de reacción del micoparasitismo en función de las enzimas involucradas. Para la solución numérica se utilizó el método de Runge-Kutta. Con la reso-lución de las ecuaciones (4) a la (10), se obtuvie-ron 34 parámetros que permitieron caracterizar el comportamiento del micoparasitismo simulado. Los parámetros se ajustaron a los datos experi-mentales y se validaron mediante el cálculo de un índice de desempeño global (ecuación 3) lo cual indicó un ajuste satisfactorio (Fig. 3 y Fig. 4).
(3)
dónde ∆(ti ) es el valor numérico simulado vari-able a tiempo ti; M(ti )el valor observado de la misma variable en el tiempo ti; M* es el valor me-dio de la variable observada, y N es el número de datos. ∆ varía entre (-∞, 1) en dónde un valor positivo de ∆ representa un simulación aceptable mientras que ∆ > 0,5 representa un buena simu-lación.
Modelo dinámicoBalance de T. harzianum (S):
(4)
txKdt
tdx
*t
tt-1 N
1ii
N
1iii
Xk
X *kS-1*
YdtdS
xs
max
G
B*L
L* P
*Q
* Xk
X *kS-1*
dtdX
32
2
1
1
xsmax
4
t xt tx
*t
tt-1 N
1ii
N
1iii
Xk
X *kS-1*
YdtdS
xs
max
G
B*L
L* P
*Q
* Xk
X *kS-1*
dtdX
32
2
1
1
xsmax
4
Fig. 3. Red de programación gráfica para el micoparasitismo. Fig. 3. Graphical network programming for mycoparasitism.
Romero-Cortes, T. et al. Micoparasitismo por Trichoderma contra Cladosporium aislado de cacao
t
Chilean J. Agric. Anim. Sci., ex Agro-Ciencia (2016) 32(1):40 32-45.
Balance de C. cladosporioides (X):
(5)
dónde:max= 0,0156 Velocidad máxima de crecimiento [cm2 h-1]; ks = 0,25 término de inhibición para el modelo Luong [cm2]; kx = 15 Constante de afinidad de T. harzianum [cm2]; α= 1,6 Término exponencial para el modelo Luong [adimensional]; Y = 0,8 Rendimiento [adimensional]; φ= 2,3 constante de modelo de Bulton para quitinasas sobre C. clados-porioides [mU mg-1]; φ2= 3,9 constante de modelo de Bulton para proteasas sobre C. cladosporioides [mU mg-1]; φ3= 0,01 constante de modelo de Bul-ton para lipasas sobre C. cladosporioides [mU mg-1]; φ4= 0,01 constante de modelo de Bulton para β-1, 3 glucanasas sobre C. cladosporioides [mU mg-1]
Balance de xilanasas (B):
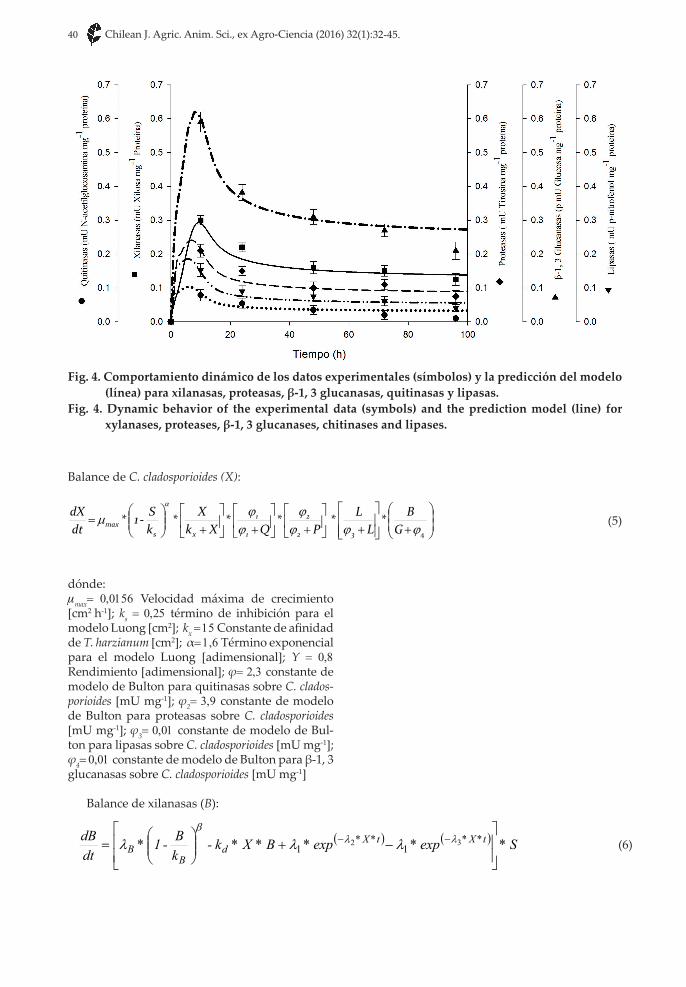
Fig. 4. Comportamiento dinámico de los datos experimentales (símbolos) y la predicción del modelo (línea) para xilanasas, proteasas, β-1, 3 glucanasas, quitinasas y lipasas.
Fig. 4. Dynamic behavior of the experimental data (symbols) and the prediction model (line) for xylanases, proteases, β-1, 3 glucanases, chitinases and lipases.
*t
tt-1 N
1ii
N
1iii
Xk
X *kS-1*
YdtdS
xs
max
G
B*L
L* P
*Q
* Xk
X *kS-1*
dtdX
32
2
1
1
xsmax
4
(6) S*exp*exp*B*X*k-kB-1*
dtdB t*X*t*X*
dB
B
32
11
S*exp*P*X*k-kP-1*
dtdP t*X*
dP
P
2
1
S*exp*Q*X*k-kQ-1*
dtdQ t*X*
dQ
Q
2
1

41
dónde:λB = 0,015 velocidad especifica de producto de xilanasas [mU mg-1 h-1 cm-2]; ks = 13 término de inhibición para el modelo Luong para xilanasas [mU mg-1]; β = 3,12 Término exponencial para el modelo Luong [adimensional]; kd = 0,01 constan-te de decaimiento referida a C. cladosporioides [h-1 cm-2]. También fue reportada por Cunniffe y Gilli-gan (2011) con un valor de 1 a 5, λ1 = 3,3 constante referida al metabolismo de xilanasas [mU mg-1 h-1 cm-2]; λ2 = 1,36 constante exponencial de la activi-dad de xilanasa (activas) [adimensional]; λ3 = 2,0 constante exponencial de la actividad de xilanasa (inactivas) [adimensional],
Balance de proteasas (P):
(7)
dónde:ρp= 0,024 velocidad especifica de producto de proteasas [mU mg-1 h-1 cm-2]; kp=16 término de inhibición para el modelo Luong para proteasas [mU mg-1]; X = 1,6 Término exponencial para el modelo Luong [adimensional]; ρ1 = 0,2 constante referida al metabolismo de proteasas [mU mg-1 h-1 cm-2]; ρ2 = 1,5 constante exponencial de la activi-dad de proteasas [adimensional].
Balance de quitinasas (Q):
S*exp*exp*B*X*k-kB-1*
dtdB t*X*t*X*
dB
B
32
11
S*exp*P*X*k-kP-1*
dtdP t*X*
dP
P
2
1
S*exp*Q*X*k-kQ-1*
dtdQ t*X*
dQ
Q
2
1
S*exp*exp*B*X*k-kB-1*
dtdB t*X*t*X*
dB
B
32
11
S*exp*P*X*k-kP-1*
dtdP t*X*
dP
P
2
1
S*exp*Q*X*k-kQ-1*
dtdQ t*X*
dQ
Q
2
1
(8)
dónde:KQ = 0,0056 velocidad especifica de producto de quitinasas [mU mg-1 h-1 cm-2]; KQ = 15 término de inhibición para el modelo Luong para quitinasas [mU mg-1]; δ = 1,6 Término exponencial para el modelo Luong [adimensional]; κ1 = 1,2 constante referida al metabolismo de quitinasas [mU mg-1 h-1 cm-2]; κ2 = 3,5 constante exponencial de la acti-vidad de quitinasas [adimensional].
Balance β-1, 3 glucanasas (G):
(9) S*exp*G*X*k-
kG-1*
dtdG t*X*
dG
G
2
1
S*exp*L*X*k-kL-1*
dtdL t*X*
dL
L
2
1
Romero-Cortes, T. et al. Micoparasitismo por Trichoderma contra Cladosporium aislado de cacao

Chilean J. Agric. Anim. Sci., ex Agro-Ciencia (2016) 32(1):42 32-45.
dónde:VG = 0,048 velocidad especifica de producto de glucanasas [mU mg-1 h-1 cm-2]; kG = 15 término de inhibición para el modelo Luong para glucanasas [mU mg-1]; ε = 1,6 Término exponencial para el modelo Luong [adimensional]; V1 = 1,25 constante referida la metabolismo de glucanasas [mU mg-1 h-1 cm-2]; V2 = 1,4 constante exponencial de la acti-vidad de glucanasas [adimensional].
Balance de lipasas (L):
(10)
S*exp*G*X*k-kG-1*
dtdG t*X*
dG
G
2
1
S*exp*L*X*k-kL-1*
dtdL t*X*
dL
L
2
1
dónde:σL = 0,001 velocidad especifica de producto de lipasas [mU mg-1 h-1 cm-2]; kL = 15 término de inhibición para el modelo Luong para lipasas [mU mg-1]; ε = 1,6 Término exponencial para el modelo Luong [adimensional]; σ1 = 0,63 constan-te referida al metabolismo de lipasas [mU mg-1 h-1 cm-2]; σ2 = 1,4 constante exponencial de la activi-dad de lipasas [adimensional].
Cabe mencionar que muchas de estas constan-tes están en rango con algunos experimentos rela-cionados con el micoparasitismo (Beg et al., 2001; Cai et al., 2004; Lu et al., 2008; Knob y Carmona, 2010). En la literatura solo se tienen reportes de cinéticas referidas a los hongos antagonistas y/o patógenos sin incluir el efecto de las proteínas en el modelado cinético (Beg et al., 2001; Cunniffe y Gilligan, 2011).
CONCLUSIONES
Las técnicas de caracterización molecular permitieron identificar a las secuencias de las cepas HT-ITV36, HT-ITV86, HT-ITV87, HT-ITV88 y HT-ITV89 como especies de C. cladosporioides sensu lato. Por otro lado, la actividad enzimática de T. harzianum VSL291 contra C. cladosporioides, C. gloeosporioides y C. cassicola se relacionó con el aumento de las actividades enzimáticas. Además, los valores del IBC obtenidos para la cepa de T. harzianum VSL291, sugieren que las cepas de Trichoderma aisladas de T. cacao tiene la capacidad para antagonizar de manera más eficiente a los patógenos fúngicos de este cultivo. Finalmente, el modelo cinético propuesto predijo las concentraciones de las enzimas β-1,3 glucanasas, quitinasas, lipasas, proteasas, xilanasas producidas por Trichoderma en paredes celulares de C. cladosporioides, C. gloeosporioides y C. cassicola. Además, este modelo puede utilizarse
como una herramienta para la estimación de actividades enzimáticas con un enfoque de instrumentación virtual.
LITERATURA CITADA
Agamez, R.E., V.J. Barrera, y L.Z. Oviedo. 2009. Evaluación del antagonismo y multiplicación de Trichoderma sp. en sustrato de plátano en medio líquido estático. Acta Biol. Colomb. 14:61-70.
Al-Sadi, A.M., Z.A. Al-Alawi, M.L. Deadman, and P. Annette. 2014. Etiology of four foliar and root diseases of wild plants in Oman. Can. J. Plant Pathol. 36:514-522.
Altschul, S.F., T.L. Madden, A.A. Schaffe, J. Zhang, Z. Zhang, W. Milleer, and D.J. Lip-man. 1997. Grapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucl. Acids Res. 25(17):3389-3402.
Amegbedzi, M.A. 2013. Effects of two fungicides, Caocobre and Ridomil on the rhizosphere microflora of cocoa (Theobroma cacao L) seed-lings. Int. J. Pure Appl. Sci. Technol. 15(1):31-42.
Ahmad, M.L., W.M. Rafiq, S.A. Sheikh, S. Sahay, and D.M. Suliman. 2012. Antagonistic po-tenciality of Trichoderma harzianum against Cladosporium spherospermum, Aspergillus niger and Fusarium oxysporum. J. Biol. Agric. and Healthc. 2(8):72-76.
Barbosa, M.A.G., K.G. Rehn, M. Menezes, and R.L. Mariano. 2001. Antagonism of Tricho-derma species on Cladosporium herbarum and their enzimatic characterization. Braz. J. Mi-crobiol. 32:98-104.
Beg, Q.K., M. Kapoor, L. Mahajan, and G.S. Hoondal. 2001. Microbial xylanases and their industrial applications: A review. Appl. Mi-crobiol. Biotechnol. 56:326-338.

43
Bensch, K., J.Z. Groenewald, J. Dijksterhuis, M. Starink-Willemse, B. Andersen, B.A. Sum-merell, et al. 2010. Species and ecological diversity within the Cladosporium cladospo-rioides complex (Davidiellaceae, Capnodiales). Stud. Mycol. 67:1-94.
Braun, U., P.W. Crous, and K. Schubert. 2008. Taxonomic revision of the genus Cladospori-um s. lat. 8. Reintroduction of Graphiopsis (= Dichocladosporium) with further reassess-ments of Cladosporioid hyphomycetes. Myco-taxon. 103:207-216.
Cai, Q., X. Yue, and T. Niu. 2004. The screening of culture condition and properties of xilanasas by white-rot fungus Pleurotus ostreatus. Pro-cess Biochem. 39:1561-1566.
Chet, I. 1987. Trichoderma - Aplication, mode of action and potential as a biocontrol agent of soil borne plant pathogenic fungi. p. 137-160. In I. Chet (ed.) Innovative Approaches to Plant Disease Control. Wiley, New York, USA.
Cuervo-Parra, J.A., M. Ramírez-Suero, V. Sán-chez-López, and M. Ramírez-Lepe. 2011a. Antagonistic effect of Trichoderma harzianum VSL 291 on phytopathogenic fungi isolated from cocoa (Theobroma cacao L.) fruits. Afr. J. Biotechnol. 10(52):10657-10663.
Cuervo-Parra, J.A. V. Sánchez-López, M. Ra-mirez-Suero, and M. Ramírez-Lepe. 2011b. Morphological and molecular characteriza-tion of Moniliophthora roreri causal agent of frosty pod rot of cocoa tree in Tabasco, Mexi-co. Plant Pathol. 10(3):122-127.
Cunniffe, N.J., and C.A. Gilligan. 2011. A theoret-ical framework for biological control of soil-borne plant pathogens: Identifying effective strategies. J. Theor. Biol. 278:32-43.
De la Cruz J., A. Hidalgo-Gallego, J.M. Lora, T. Benitez, J.A. Pintor-Toro, and A. Llobell. 1992. Isolation and characterization of three chitinases from Trichoderma harzianum. Eur. J. Biochem. 206:859-867.
Djocgoue, P.F., T. Boudjeko, D.J. Nankeu, M.I.B. Efombagn, S. Nyasse, and D.N. Omokolo. 2006. Comparative assessment of the resis-tance of cocoa (Theobroma cacao L.) progenies from SNK10 x SNK413; ICS84 x ICS95 to Phy-tophthora megakarya in Cameroon by measur-ing size of necrotic lesion along the midrib. Plant Pathol. 5:329-335.
Dochain, D. 2003. State and parameter estimation in chemical and biochemical processes: a tu-torial. J. Process Contr.13:801-818.
Elad, Y., and A. Kappat. 1999. Role of Trichoder-ma harzianum protease in the biocontrol of Botrystis cinerea. Eur. J. Plant Pathol. 105:177-189.
Ezziyyani, M., S.C. Pérez, A.A. Sid, M.E. Re-quena, y M.E. Candela. 2004. Trichoderma har-zianum como biofungicida para el biocontrol de Phytophthora capsici en plantas de pimien-to (Capsicum annuum L.). Anales de Biología 26:35-45.
Foster, D.M., K.R. Kozak, M.G. Loftus, J.J. Ste-vens, and I.K. Ross. 1993. The polymerase chain reaction and its application to filamen-tous fungi. Mycol. Res. 97:769-781.
Goes, A. 1998. Donças fúngicas. p. 208-216. In Simposio Brasileiro sobre a Cultura do Mara-cujazeiro. 10-13 fevreiro 1998. Faculdade de Ciencias Agrárias e Veternárias, Universida-de Estadual Paulista, Jaboticabal, Brasil.
Haggag, M.W., L.A. Kansohy, and M.A. Aly. 2006. Characterization and antifungal activ-ity against brown spot disease on faba bean. Plant Pathology Bulletin 15:231-239.
Harman, G., I. Chet, and R. Baker. 1981. Fac-tors affecting Trichoderma hamatum applied to seed as a biocontrol agent. Phytopathol. 71:569-572.
Jacyno, J.M., J.S. Harwood, and M.K. Lee. 1993. Isocladosporin, a biologically active isomer of cladosporin from Cladosporium cladospori-oides. J. Nat. Prod. 56(8):1397-1401.
Kashmiri, M.A., A. Adnan, and B.W. Butt. 2006. Production, purification and partial charac-terization of lipase from Trichoderma viride. Afr. J. Biotechnol. (10):878-882.
Kendall, J.M., and P.T. Rygiewicz. 2005. Fun-gal-specific PCR primers developed for anal-ysis of the ITS region of environmental DNA extracts. BMC Microbiol. 5:1-11.
Kilaru1, A., B.A. Bailey, and K.H. Hasenstein. 2007. Moniliophthora perniciosa produces hormones and alters endogenous auxin and salicylic acid in infected cocoa leaves. FEMS Microbiol. 274:238-244.
Knob, A., and E.C. Carmona. 2010. Purification and characterization of two extracellular xy-lanases from Penicillium sclerotiorum: A novel acidophilic xylanase. Appl. Biochem. Bio-technol. 162:429-443.
Kryczyński, S., and Z. Weber. 2011. Fitopatolo-gia. Tome 2. Choroby roślin uprawnych. Powszechne Wydawnictwo Rolnicze i Leśne. Oddział, cop. Poznań, Polska.
Kucuk, C., and M. Kivanc. 2008. Mycoparasitism in the biological control of Gibberella zeae and Aspergillus ustus by Trichoderma harzianum strains. J. Agri. Technol. 4:49-55.
Kunitz, M. 1946. Spectrophotometric method for the measurement of ribonuclease activity. J. Biol. Chem. 164:563-568.
Romero-Cortes, T. et al. Micoparasitismo por Trichoderma contra Cladosporium aislado de cacao

Chilean J. Agric. Anim. Sci., ex Agro-Ciencia (2016) 32(1):44 32-45.
Kurzatkowski, W., A. Törrönen, J. Filipek, R. Mach, P. Herzog, S. Sowka, and C.P. Kubi-cek. 1996. Glucose-induced secretion of Trichoderma reesei Xylanases. Appl. Environ. Microbiol. 62(8):2859–2865.
Larkin, M.A., G. Blackshields, N.P. Brown, R. Chenna, P.A. McGettigan, H. McWilliam, et al. 2007. ClustalW and ClustalX version 2. Bioinformatics 23(21):2947-2948.
Latorre, B.A., E. Agosin, R. San Martín, y G.S. Vasquez. 1997. Effectiveness of conidia of Trichoderma harzianum produced by liquid fermentation against Botrytis bunch rot table grape in Chile. Crop Protection 16:209-214.
López-Pérez, P.A., M.I. Neria-González, R. Agui-lar-López. 2013. Cadmium concentration sta-bilization in a continuous sulfate reducing bioreactor via sulfide concentration control. Chem. Pap. 67(3):326–335.
Lu, F., M. Lu, Z. Lu, X. Bie, H. Zhao, and Y. Wang. 2008. Purification and characterization of xy-lanase from Aspergillus ficuum AF-98. Biore-source Technol. 99:5938-5941.
Manocha, M.S., and A.S. Sahai. 1993. Mechanisms of recognition in necrotrophic and biotrophic mycoparasites. Can. J. Microbiol. 39:269-275.
Miller, G.L. 1959. Use of dinitrinosalicylic acid re-agente for determination of reducing sugar. Anal. Chem. 31:426-428.
Mokhtar, H., and A. Dehimat. 2012. Antagonism capability in vitro of Trichoderma harzianum against some pathogenic fungi. Agric. Biol. J.N. Am. 3(11):452-460.
Monreal, J., and E.T. Reese. 1969. The chitinase of Serratia marcescens. Can. J. Microbiol. 15:689-696.
Nawani, N., S.D. Nirpjit, and K. Jagdeep. 1998. A novel thermostable lipase from a thermo-philic Bacillus sp. Characterization and ester-ification studies. Biotechnol. Lett. 20(10):997-1000.
Nobe, R., Y. Sakakibara, N. Fukuda, N. Yoshida, K. Ogawa, and M. Suijo. 2003. Purification and characterization of laminaran hydro-lases from Trichoderma viride. Biosci. Biotech-nol. Biochem. 67:1349-1357.
Orole, O.O., and T.O. Adejumo. 2009. Activity of fungal endophytes against four maize wilt pathogens. Afr. J. Microbiol. Res. 3(12):969-973.
Phillips-Mora, W., A. Coutiño, C.F. Ortiz, A.P. López, J. Hernández, and M.C. Aime. 2006. First report of Moniliophthora roreri causing frosty pod rot (moniliasis disease) of cacao in Mexico. Plant Pathol. 55:584.
PhyloDraw. 1999. PhyloDraw versión 0.8. Graph-ics Application Lab, Pusan National Univer-sity. Available at http://search.wareseeker.com/phylodraw-version-0.8/ (Accessed 30 July 2008).
Quintana-Obregón, E.A., M. Plascencia-Jatomea, J. López-Cervantes, D.L.A. Cira-Chávez, D.I. Sánchez-Machado, and M.O. Cortez-Rocha. 2011. Antifungal activity of chitosan in Clado-sporium cladosporioides isolated from safflow-er. Rev. Mex. Micol. 34:93-96.
Rawashdeh, R., I. Saadoun, and A. Mahasneh. 2005. Effect of cultural conditions on xy-lanase production by Streptomyces sp. (strain Ib 24D) and its potential to utilize tomato pomace. Afr. J. Biotechnol. 4(3):251-255.
Rezende, M.I., A.M. Barbosa, A.F.D. Vasconcelos, and A.S. Endo. 2002. Xylanase production by Trichoderma harzianum rifai by solid state fermentation on sugarcane bagasse. Braz. J. Microbiol. 33:67-72.
Riascos, D., I. Quiroga, y L. Hoyos-Carvajal. 2011. Análisis de la sintomatología de la roña en gulupa (Pasiflora edulis f. edulis SIMS). Agron. 19(1):20-30.
Robeson, D.J., and M.A.F. Jalal. 1992. Formation of Ent-isophleichrome by Cladosporium herba-rum isolated from sugar beet. Biosci. Biotech. Biochem. 56(6):949-952.
Schubert, K., J.Z. Groenewald, U. Braun, J. Dijk-sterhuis, M.S. Starink, C.F. Hill, et al. 2007. Biodiversity in the Cladosporium herbarum complex (Davidiellaceae, Capnodiales), with standardisation of methods for Cladospo-rium taxonomy and diagnostics. Studies in Mycology 58:105-156.
Seiboth, B., L. Hartl, M. Pail, and P.C. Kubicek. 2003. D-Xylose metabolism in Hypocrea jeco-rina; loss of the xylitol dehydrogenase step can be partially compensated for by lad1-En-codad L-Arabinitol-4-Dehydrogenase. Eu-karyot. Cell. 5:867-875.
Stricker, R.A., K. Grosstessner-Hain, E. Wurleit-ner, and P.L. Mach. 2006. Xyr1 (Xylanase Regulator 1) Regulates both the hydrolytic enzyme system and D-Xylose metabolism in Hypocrea jecorina. Eukariot. Cell. 12:2128-2137.
Suarez, M.B., L. Sanz, M.L. Chamorro, M. Rey, F.J. Gonzalez, A. Llobell, and E. Monte. 2005. Proteomic analysis of secreted proteins from Trichoderma harzianum. Identification of a fungal cell wall-induced aspartic protease. Fungal Genet. Biol. 42:924-934.
Szekeres, A., B. Leitgeb, L. Kredics, L. Manczing-er, and C. Vágvölgyi. 2006. A novel, image analysis-based method for the evaluation of in vitro antagonism. J. Microbiol. Methods 65(3):619-622.

45
Ujiie, M., C. Roy, and M. Yaguch. 1991. Low-mo-lecular weight xylanase from Trichoderma viride. Appl. Environ. Microbiol. 57(6):1860-1862.
Velázquez-del Valle, M.G., S. Bautista-Baños, A.N. Hernández-Lauzardo, M.G. Guerra-Sánchez, and E. Amora-Lazcano. 2008. Estrategias de control de Rhizopus stolonifer Ehrenb. (EX FR) Lind, agente causal de pudriciones postcose-cha en productos agrícolas. Rev. Mex. Fito-patol. 26(1):49-55.
Villadsen, J., J. Nielsen, and G. Lidén. 2011. Biore-actor engineering principles. 3r. ed. Springer, USA.
Wang, H.C., J. Wang, W.H. Li, Y.F. Huang, H.Q. Xia, M.S. Wang, et al. 2014. Cladosporium cladosporioides identified in China on tobacco seeds. Plant Dis. 98(7):1002.
Romero-Cortes, T. et al. Micoparasitismo por Trichoderma contra Cladosporium aislado de cacao


















