
FDSS/μCELLアプリケーションカタログ iPS細胞由来心筋細胞のCa2… · 2波長蛍光発光の同時測定も可能(オプション) iPS細胞由来心筋細胞内Ca2+測定
Upload
maniiiiiiiiCategory
view
32download
3description

1
1
Cell CycleRegulation and Checkpoints
Chi-Wu Chiang, Ph.D.IMM, NCKU
基醫所

2
2
-----Accompanied with cell division, cell growth and cell death
Cell mass=cell number+cell size
cell mass
From cell division to a full grown body

3
3
Cell division, cell growth, and cell death areindependently regulated but linked
Intrinsic programs
Extracellular signal molecules
Regulated cell cycle progression
Programmed cell death
Mitogens
Growth factors
Survival factors
Apoptosis-inducing factorsProliferation inhibitors

4
4
Introduction to cell cycle and cell cyclecheckpoints

5
5
Cell cycle
A cell reproduces by performing an orderly sequence of events in whichit duplicates its contents and then divides in two

6
6
The phases of cell cycle

7
7
M
S
M
G2 G1
S
Early embryonic cell cycle
Somatic cell cycle
Embryonic cell cycle and somatic cell cycle
12h
6h
6h
30 min

8
8
Yeast provides a good genetic system for studyon cell cycle
Mostly proliferating in a haploid state
Easy for genetic manipulation
Cdc genes and cdc mutants ----cell-division-cycle genes

9
9
Cdc mutants selected by temperaturesensitive mutations
Temperature-sensitive mutantsFunctioning in permissive condition (low temp.) andnonfunctioning in restrictive condition (high temp.)
A mutant that cannot complete the cell cycle, cannot bepropagated

10
10
Xenopus oocyte: a giant cell for study cell cyclebiochemically
1 mm in diameterCarrying 100,000 times more cytoplasm than an average cell in thehuman body
After fertilization occurs, the first division takes about 90 minutes,and the next 11 divisions occur at 30-mim intervals, producingabout 4096 cells within 7 hours
Each cycle is divided into S and M phase without detectable G1 or G2phases

11
11
Studying the cell cycle in a cell-free system
Observation of repeated nuclei decondense and DNA replication and mitosisIn vitro

12
12
Measurements of cell cycle progression
Propidium iodide (PI) staining
EMBO 2003, 22:5459
Oncogene 2001, 20:4507
BrdUstaining(bromo-deoxyuridine)
PIstaining

13
13
The cell-cycle control system

14
14
Operates like a timer thattriggers the events in a setsequence
The control of cell cycle
The system of switches is binary(ON/OFF) and launches events ina complete, irreversible fashion
The control system isindependent of the events itcontrols
The system is highly adaptable andcan be modified to suit specific celltypes

15
15
The Cyclin-Cdk complexes: the majorcomponents of the cell cycle control system

16
16
Cyclically activated protein kinases control cellcycle progression
Cyclin-dependent kinases (Cdks)
Kinase’s activity oscillates in the cell cycle
Cyclical changes in Cdk activity are controlled byan array of enzymes and other proteins
Among these, cyclins are the majormolecules
The “go”system (or the “engine”)
Expression is constant through the cellcycle

17
17
The cyclins
Four classes of cyclins:1. G1-cyclins—help to promote passage through“Start”or the restriction point in late G1
2. G1/S-cyclins—bind Cdks at the end of G1 andcommit the cell to DNA replication
3. S-cyclins–binds Cdks during S phase and arerequired for the initiation of DNA replication
4. M-cyclins—promote the events of mitosis

18
18
The major Cyclin-CDK complexes

19
19
Regulation of cell cycle

20
20
Regulation of the cyclin-Cdk complex
Post-translational modification
Transcriptional regulation
Cyclical proteolysis
Inhibitors of Cdks

21
21
Activation of the cyclin-Cdk complex
Partial activation of Cdks by cyclin associationFull activation by Cdk-activating kinase (CAK) phosphorylation
Active sitehindered byinhibitory T-loop
Exposure of T-loop by cyclinbinding
Phosphorylation byCAK causesconformationchange
Study on crystal structure of CDK2 revealed:

22
22
Regulation of the cyclin-Cdk complex:post translational modification
Inhibition of Cdks by inhibitory phosphorylation
Inhibition: Phosphorylation by Wee1 kinase at Tyr15Cdc25 phosphatase dephosphorylates the p-Tyr15 andactivates the Cdk
Human Cdk1 and also Cdc2 in yeast:
PP2A

23
23
The inhibition of a cyclin-Cdk complex by Cdk inhibitorproteins (CKI)
CKI (Cdk inhibitor proteins):
The INK family, P15, p16, p18, and p19 target to CDK4, 6
The CIP/KIP family, p21, p27, and p57 target to mostCDKs
Binding of CKI renders Cdk inactive by conformation changeand rearranged structure of active site
(during G1)
(during all phases)
24
24
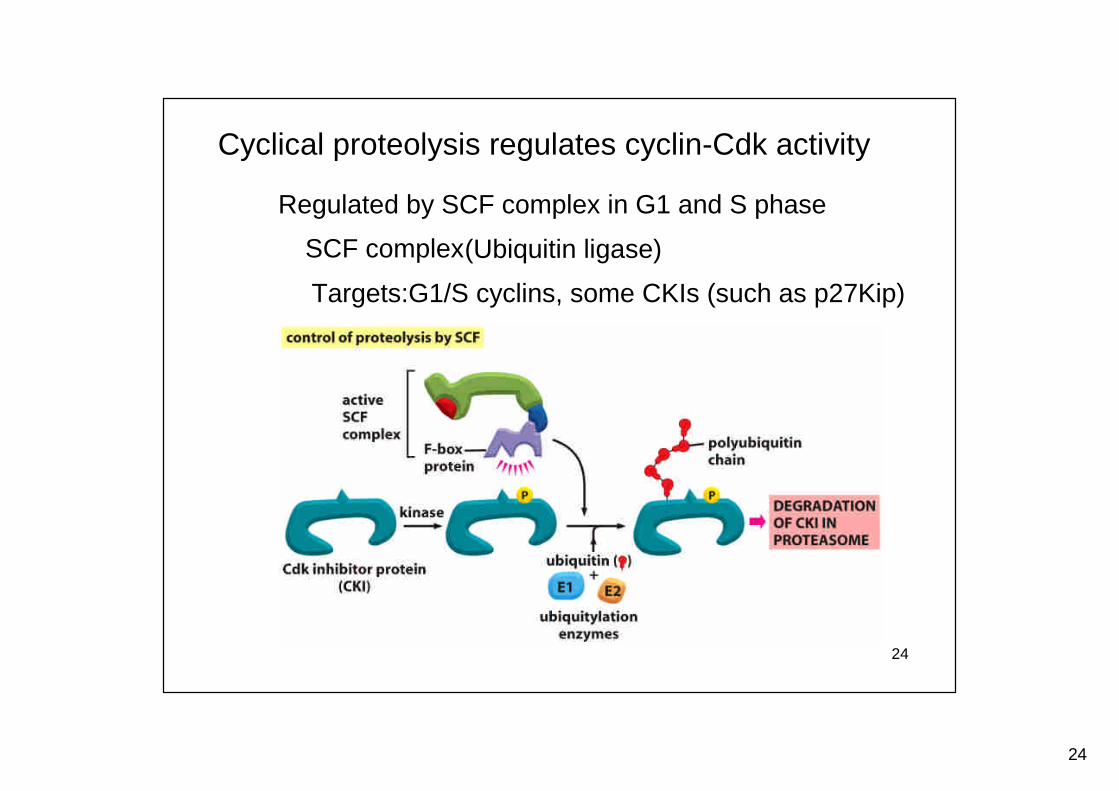
Cyclical proteolysis regulates cyclin-Cdk activity
SCF complex(Ubiquitin ligase)
Regulated by SCF complex in G1 and S phase
Targets:G1/S cyclins, some CKIs (such as p27Kip)

25
25
(Ubiquitin ligase)APC complex
Cyclical proteolysis regulates cyclin-Cdk activityRegulated by APC complex in M phase
APC: anaphase promoting complex
Target: M cyclin

26
26
The G1, G1-S, and S phase

27
27
The G1 phase is a state of stable Cdk inactivity
To reach a stable G1 for growth, Cdk reactivation is prevented
Decline in M-cyclin and in Cdc20-APC
Increase in Cdh1-APC* activitywhich targets to M-cyclins
*Cdh1 is a close relative of Cdc20
Sic1, a Cdk inhibitor ( CKI),inactivates M-Cdk
Decrease in M-cyclin transcription
*Cdh1-APC targets to M-cyclins to inactivate M-Cdk after mitosis

28
28
Initiation of S phase in budding yeast
Accumulation of G1-cyclin that is resistant to Sic-1 and Hct-1-APC
G1-Cdk stimulates G1/S-cyclin synthesis
G1/S-Cdk stimulates S-cyclin synthesis, and increase in S-Cdk activity
G1/S-Cdk phosphorylates Sic1 and Hct-1 and blocks their activity

29
29
Initiation of S phase in animal cellsIn G1, cells are in a state of Hct-1 activation, accumulation of aCKI, and inhibition of cyclin gene expression (M-cyclin)
In late G1, activation of G1-Cdk reverses the inhibitory state of G1
G1-Cdk and G1/S-Cdk phosphorylate Rb, an inhibitor of cellcycle progression, and results in release of E2F

30
30
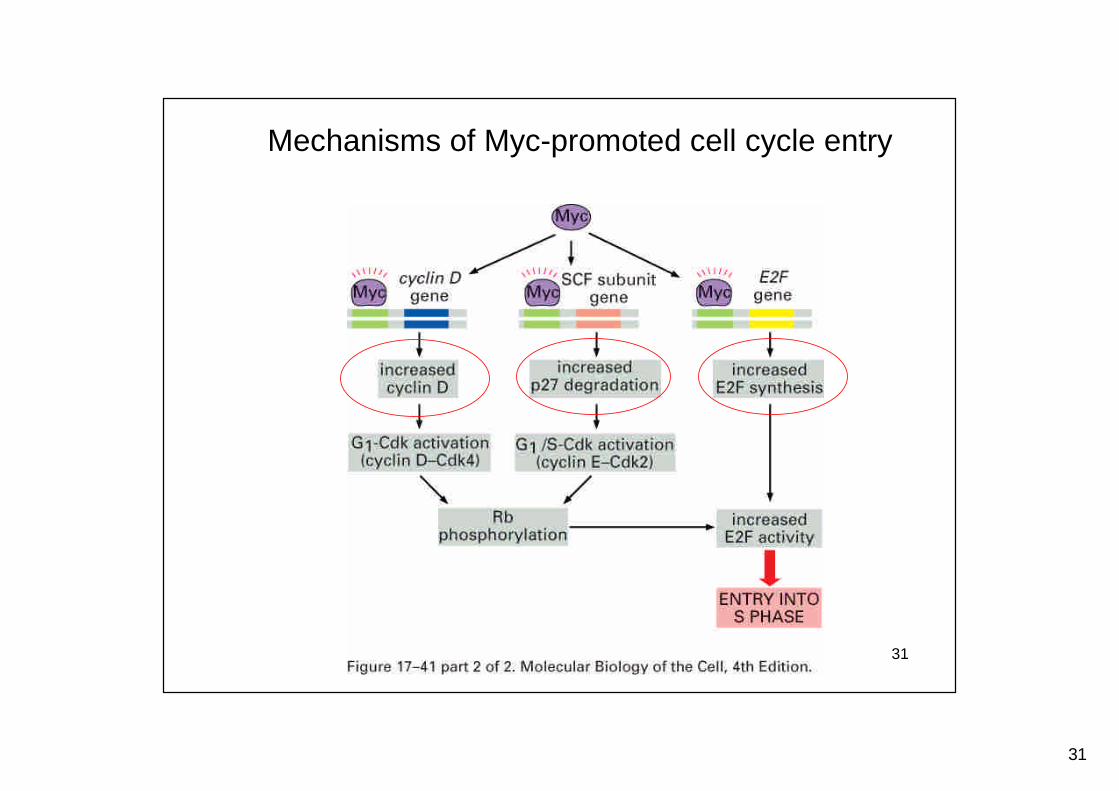
Mitogens stimulate activation of Ras and a MAPkinase cascade to trigger cell division
31
31
Mechanisms of Myc-promoted cell cycle entry

32
32
Figure 17-23 Molecular Biology of the Cell (© Garland Science 2008)
Control of the initiation for DNA replication
(formed by Orc proteins)

33
33
The control for DNA replication
Regulation of pre-replicative complex (pre-RC) by S-CDK
ORC: Origin recognition complexBinds to the replication origin through the cell cycle
Cdc6, Cdt1 associate with ORC at early G1
help to recruit Mcm proteins, a DNA helicase
S-Cdk causes:
Control only one replication, by Cdc6 degradation andby Mcm export from nucleus
Start of DNA replication, by ORC phosphorylation,MCM activation, and replication firing

34
34
The M phase:Mitosis and cytokinesis

35
35
Regulation of entry into mitosis

36
36
• M-cyclin accumulates by reduced degradation before M phasein embryonic cell cycles
• M-cyclin increases in transcription in most cell types
• Inactivated M-cyclin-Cdk complex accumulates due toinhibitory phosphorylation
• Cdc25 phosphatase dephosphorylates and activates M-Cdk
• Polo kinase and active M-Cdk activate Cdc25
• Active M-Cdk inactivates Wee1
Entry into mitosis

37
37
Multiple roles of M-Cdk in mitosis
Induce the assembly of mitotic spindle
Ensure replicated chromosomes attach to the mitoticspindle
Chromosome condensation
Nuclear envelope breakdown
Reorganization of the Golgi apparatus and endoplasmicreticulum

38
38
Mitosis

39
39
Mitosis, continued
Exit

40
40
Sister chromatid separation is triggered by proteolysis
Separase is required forsister chromatids separation
Securin, can inactivateseparase; degradation ispromoted by APC
APC: anaphase-promotingcomplex, a highly regulatedubiquitin ligase; promote thedestruction of several mitoticregulatory proteins
Cdc20 activates APC

41
41
The spindle-attachment checkpoint
Ensure that all chromosomes are properly attached tothe spindle before sister-chromatid separation occurs
A sensor mechanism monitors the state of thekinetochore, the specialized region of thechromosome that attaches to microtubules of thespindle
Improper attachment of kinetochore to the spindlesends out a negative signal to the cell-cycle controlsystem, blocking Cdc20-APC activation and sisterchromatid separation
Several proteins, including Mad2, are recruited tounattached kinetochores. Mad2 binding results in Inhibitionof Cdc20-APC and blocking securin destruction

42
42
Exit from mitosis and start of G1
The mitotic spindle must be dissembled
Complex changes at the end of mitosis
Chromosomes decondensedThe nuclear envelope reformed
Inactivation of M-Cdk is required for exit from mitosis
Cdc20-APC complex mediated ubiquitin-dependentproteolysis of M-cyclin

43
43
Decline in M-cyclin and in Cdc20-APC
Increase in Cdh1-APC* activitywhich targets to M-cyclins
*Cdh1 is a close relative of Cdc20
Sic1, a Cdk inhibitor ( CKI),inactivates M-Cdk
Decrease in M-cyclin transcription
*Cdh1-APC targets to M-cyclins to inactivate M-Cdk after mitosis
Exit from the M phase and entry ofG1 phase in somatic cells

44
44
DNA damage checkpoints and p53 activation
Normally, P53 is targeted forubiquitin-dependentdegradation by Mdm2, aubiquitin ligase
p53 is phosphorylated byDNA damage-activatedkinase and is released frombinding to Mdm2

45
45
An overview of the cell-cycle control system
Accelerators (cyclin-CDK) and brakes (checkpoints)

46
46
From cell dividing to senescence
-gal-positiveFlat morphology
Abnormal nuclear morphology

47
47
In 1965, Leonard Hayflick first suggested thatthere was a “finite limit to the cultivation periodof diploid cell strains”and this was“attributable to intrinsic factors which areexpressed as senescence at the cellular level”.
Hayflick proposed that normal cells cannot divideindefinitely because they are programmed for aset proliferative lifespan.
Cell senescence observed by Hayflick
48
48
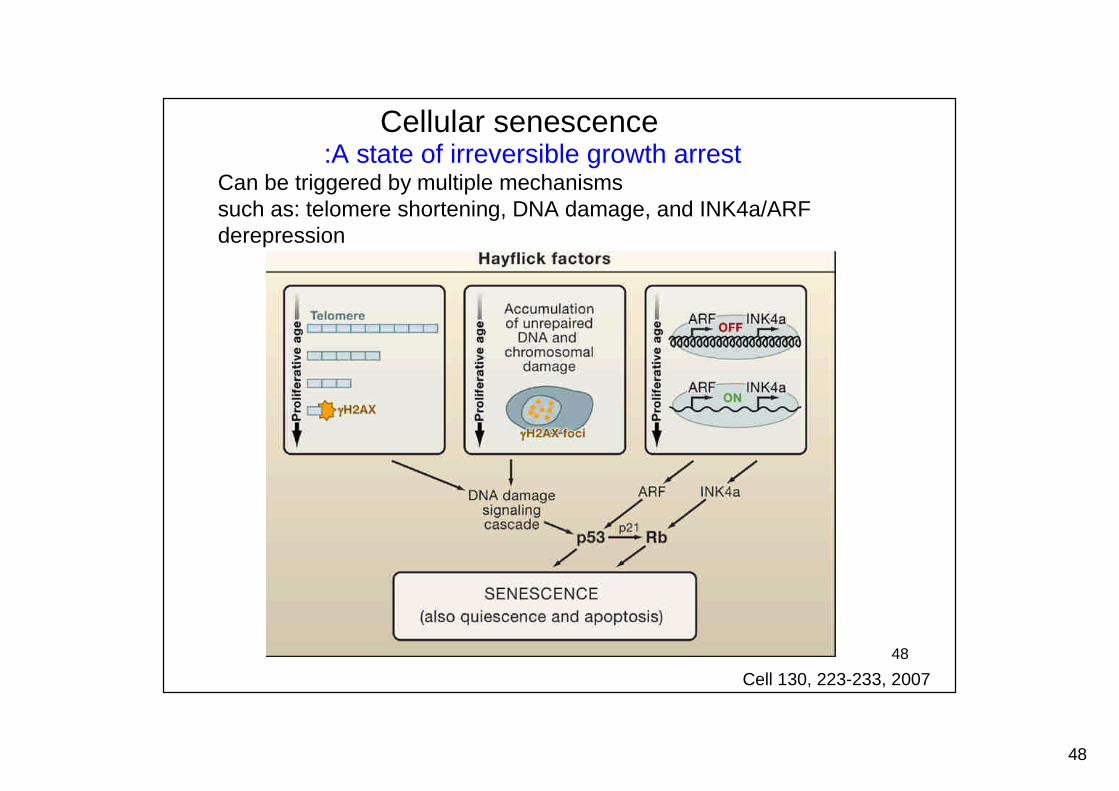
Cellular senescence:A state of irreversible growth arrest
Can be triggered by multiple mechanismssuch as: telomere shortening, DNA damage, and INK4a/ARFderepression
Cell 130, 223-233, 2007

49
49
Nature Reviews Cancer 1:203, 2001
Telomere
Molecular Cell Biology, 6th edition
GGGTTA repeats (near thousands) in human’s telomere

50
50
Telomere shortening and senescence
Human diploid fibroblasts (HDFs) undergo 60-80doublings in culture, after which they cease divisionand develop a senescent phenotype
Telomerase expression in HDFs is sufficient tobypass the cell senescence and crisis
Nature Reviews Cancer 1:203, 2001

51
51
The telomere problem of a linear DNAreplication
The “must be”5’to 3’polymerization, the lagging stand, and the telomere

52
52
Maintaining telomere length immortalizes cells
Nature Reviews Cancer 1:203, 2001
Exogenous expression of telomerase reverse transcriptase (TERT)at any stage in the replicative period allows immortalization of thesecells

53
53
Telomere hypothesis
• Most adult cells have limiting amounts of telomerase that arenot sufficient for preventing telomere loss, resulting in theshortening of telomeres with ages
• Telomerase-deficient mice with short telomeres show reducedfunction of various stem cell compartments including those inthe bone marrow and skin
• Telomerase-deficient mice show a decrease in both the medianand maximum lifespan, and these effects become morepronounced with each subsequent generation
• Terc transgenic mice show increased mortality due to cancer,but show evidence improved tissue regeneration as well as aslight increase in maximum life span
Nature 2007, 448:767-774

54
54
Telomere stabilization: an important step in tumordevelopment
Most human cells don’t express telomerase, whereas mosthuman tumors express telomerase
Expression of dominant-negative telomerase reversetranscriptase (TERT) results in apoptosis of tumor cells
Those tumors that don’t express telomerase maintain thetelomere by an mechanism of alternative lengthening oftelomeres (ALT)

55
55
Age-dependent increase in the number of senescent cellsin differentiated tissues

56
56
Genomic instability in aging cells
DNA damage and mutations accumulate with agein mammals
Cytogenetically visible lesions such as translocations,insertions, dicentrics, and acentric fragmentsaccumulate in aging mammalian cells

57
57
DNA damage
DNA repair
Genomic instability
Recovery and regenerate
Defect in DNA repair
Aging?
The role of DNA repair in aging

58
58
Genomic instability increased not only in agingcells but also in most cancer cells

59
59
Excessive oncogenic induction can result in cell-cyclearrest or apoptosis

60
60
OIS: Oncogene-induced senescence
ARF-p53 pathway
p16INK4a-RB pathway
Implication: in response to the activation of mitogeniconcogenes, checkpoint-mediated failsafe mechanismssuch as apoptosis or cellular senescence may berecruited to terminate a pre-malignant condition
Trends cell biol 11, s27-s31, 2001
Nature Rev Cancer 3, 286-295, 2003

61
61
Pathways to senescence
Nature Reviews Cancer 1:203, 2001

62
62
• Please introduce some key background information regarding thispaper, including the motivation or hypotheses for pursuing this study,cdk2, myc, relationship between cdk2 and myc, oncogene-inducedsenescence (such as Ras), etc.
• What has been found in this paper is different from the finding inprevious studies regarding myc activity in cell proliferation andapoptosis?
• Why authors think myc-induced senescence in the absence of cdk2 isnot due to blocking apoptosis? And myc-induced cell proliferation isindependent of cdk2?
• Why authors conclude that Arf-p53-p21 and p16-pRb pathways wereintegrally required for myc-induced senescence in cdk2-/- cells?
Assignments for next week’s class
Cdk2 suppresses cellular senescence induced by the c-myc oncogene,Nat Cell Biol. 2010 Jan;12(1):54-9

63
63
• Did authors conclude that cdk2 regulates myc-induced senescencethrough controlling myc-induced genotoxic stresses? Why?
• Why oxidative stress may contribute to myc-induced senescence?What is the role of cdk2 in this?
• Since CDK2 is dispensable in many cancer cell lines, why authorsstill think inhibition of CDK2 can be a therapeutic consideration fortreatment of cancer?
Assignments for next week’s class, continued,


















