
Micropropagation of Swertia chirayita -...
66
Micropropagation TERI University-Ph.D. Thesis, 2006 Micropropagation of Swertia chirayita 4.1 Introduction In vitro culture is an efficient method for ex situ conservation of plant biodiversity and multiplication of the endangered species. It enables the propagation of the endangered species from minimum plant material that is available for propagation. S. chirayita, though a widely used medicinal plant, is now a rare and endangered species, a consequence of its ruthless and destructive harvesting. Further, low germination percentage and poor viability of the seeds, long gestation periods and delicate field handling are some of the factors that discourage commercial cultivation and agro-technology development of S. chirayita. Considering the problems beset with conventional propagation of the species and its endangered status, efforts have been made to propagate the plant by in vitro techniques. Wawrosch et al. (1999) described adventitious shoot formation from root explants of in vitro germinated seedlings of S. chirata. The regenerated shoots were merely 3 mm in length, making handling of cultures a difficult task. Hence a two-step regeneration protocol was optimised in order to reduce the problems of callus formation, hyperhydricity and small shoot size. Ahuja et al. (2003) filed patent for medium composition for faster propagation of S. chirayita from mature explants. The patent report cited a shoot multiplication fold that ranged from 11.4 to 26.2, while the rooting frequency reportedly ranged between 50-80% in time duration of about 8 weeks, and a survival of 70% was reported. The present study describes axillary multiplication of S. chirayita from nodal explants derived from four-week-old aseptic seedlings and successful establishment of the plantlets in field. In this study, clusters obtained from the seedling-derived nodal explants gave a multiplication fold of 4.5 every four weeks. Hundred percent rooting was obtained within four weeks of inoculation on rooting medium. We also describe here a regeneration protocol from leaf explants procured from in vitro cultures. 4
Transcript of Micropropagation of Swertia chirayita -...

Micropropagation
TERI University-Ph.D. Thesis, 2006
Micropropagation of Swertia chirayita
4.1 Introduction
In vitro culture is an efficient method for ex situ conservation of plant
biodiversity and multiplication of the endangered species. It enables the
propagation of the endangered species from minimum plant material that is
available for propagation. S. chirayita, though a widely used medicinal plant, is
now a rare and endangered species, a consequence of its ruthless and destructive
harvesting. Further, low germination percentage and poor viability of the seeds,
long gestation periods and delicate field handling are some of the factors that
discourage commercial cultivation and agro-technology development of S.
chirayita.
Considering the problems beset with conventional propagation of the species
and its endangered status, efforts have been made to propagate the plant by in
vitro techniques. Wawrosch et al. (1999) described adventitious shoot formation
from root explants of in vitro germinated seedlings of S. chirata. The
regenerated shoots were merely 3 mm in length, making handling of cultures a
difficult task. Hence a two-step regeneration protocol was optimised in order to
reduce the problems of callus formation, hyperhydricity and small shoot size.
Ahuja et al. (2003) filed patent for medium composition for faster propagation
of S. chirayita from mature explants. The patent report cited a shoot
multiplication fold that ranged from 11.4 to 26.2, while the rooting frequency
reportedly ranged between 50-80% in time duration of about 8 weeks, and a
survival of 70% was reported.
The present study describes axillary multiplication of S. chirayita from nodal
explants derived from four-week-old aseptic seedlings and successful
establishment of the plantlets in field. In this study, clusters obtained from the
seedling-derived nodal explants gave a multiplication fold of 4.5 every four
weeks. Hundred percent rooting was obtained within four weeks of inoculation
on rooting medium. We also describe here a regeneration protocol from leaf
explants procured from in vitro cultures.
4

Micropropagation
TERI University - Ph.D. Thesis, 2006
4.2 Results
Micropropagation from seedling derived nodal explants
4.2.1. Seed germination and culture establishment
A protocol for the axillary multiplication of S. chirayita was established in this
study. In absence of mature explants, S. chirayita seedlings were used for
procuring initial nodal explants for culture initiation. The surface sterilised, GA3
pre-treated seeds showed 80% germination. In absence of pre-treatment with
GA3, the seeds failed to germinate. These seeds were allowed to grow in half
strength (MS major inorganic salts and iron reduced to half the original MS
strength) semi-solid MS medium supplemented with 3% sucrose, and later,
four-week-old seedlings were used for further in vitro studies.
4.2.2 Culture establishment
4.2.2.1 Effect of type and concentration of cytokinins on shoot bud
induction
Preliminary experiments were conducted to test the effect of equimolar
concentration of BAP, Kn and 2iP with MS basal as control on shoot bud
induction response of nodal explants (Figure 4.1). Sugar concentration was kept
constant at 3% and the medium was gelled with 0.8% agar. Data was recorded
after four weeks of culture period.
The nodal explants cultured on MS basal and on MS medium supplemented
with 3 µM each of BAP, Kn or 2iP separately, showed a significant variation in
terms of number of shoot buds induced per explant (Figure 4.2). A maximum
number of 5.9 and 5.8 shoot buds were induced on MS medium supplemented
with 3 M each of 2iP and BAP, respectively. On the other hand, only 4.9 shoot
buds per explant were induced on medium supplemented with an equimolar
concentration of Kn. The response from nodal explants was stabilised after the
fifth subculture and, by that time, the nodal cultures started forming shoot
clusters (Figure 4.1 c, d). The shoot bud induction response on basal medium
remained almost negligible with only 1.65 shoot buds being formed per explant.
Our results thus conclusively showed that cytokinins were essential for shoot
bud induction and multiplication. The effect of cytokinins on cluster elongation
was however not significant and remained more or less similar to that observed
on the basal medium used as control in the study (Figure 4.2). The nodal
explants initially inoculated on medium supplemented with BAP, Kn and 2iP
were further transferred to similar medium for multiplication. The shoot

Micropropagation
TERI University - Ph.D. Thesis, 2006
clusters obtained on BAP and Kn enriched medium were found to retain their
vigour and health. However, shoots multiplying on 2iP-enriched medium started
showing signs of hyper-hydricity by the end of the fifth passage. These shoots
were subsequently removed and excluded from further experimental studies.
Further experiments during the culture establishment phase were carried out
with MS medium supplemented with 3 M BAP as constant.
4.2.2.2 Effect of sugar concentration on the multiplication rate
The sugar concentration was varied from 1 to 5% to test its effect on
morphogenic response (Table 4.1). A gradual increase was observed in the
number of shoot buds induced per nodal explant as the sucrose concentration
was increased from 1 to 3%. However, an increase in sucrose concentration
beyond 3% resulted in decline in the number of shoot buds per explant. Hence,
maximum number of 4.27 shoots with an average height of 2.43 cm was
obtained from each nodal explant inoculated on medium with 3% sucrose.
Table 4.1 Effect of the varying sugar concentration on shoot multiplication
Sugar concentration (%) Shoot Buds Shoot length (cm)
1 1.5± 0.35c* 0.85±0.11b
2 2.8±0.46b 1.11±0.34b
3 4.27±0.22a 2.43±0.20a
4 2.96±0.23b 1.23±0.22b
5 2.64±0.26b 1.25±0.09b
LSD (0.05)** 0.58 0.38
On the other hand, multiplication fold reduced to a minimum of 1.5 when the
explants were cultured on medium supplemented with 1% sucrose and the
average cluster height also declined to 0.85 cm. Also, no significant variation in
shoot bud induction per explant was observed on medium supplemented with 2,
4 and 5% sucrose concentrations. Despite the quantitative variation in number
of shoot buds induced, the shoots remained healthy with dark green leaves
irrespective of sucrose concentrations.
* Means followed by the same letter within the column are not significantly different and the
values represent mean±standard deviation ** Least Significant Difference (LSD) helps in determining directly if two means differ
significantly. If the sample means differ more than the LSD, it is considered that the two are
significantly different.

Micropropagation
TERI University - Ph.D. Thesis, 2006
On the whole, varying sucrose concentration did have a significant effect on
the multiplication fold and shoot elongation as was evident from the highly
significant P values, both for cluster multiplication and elongation.
4.2.2.3 Effect of different gelling agents
The effect of different gelling agents viz. agar, agar gel and gelrite was
ascertained on explant multiplication and elongation (Table 4.2). Maximum
number of shoot buds (5.9/explant) were induced in medium gelled with 0.8%
agar, followed by 0.4% agar-gel and 0.2% gelrite, which resulted in 5 and 4.3
multiplication fold respectively.
However, completely contrasting results were observed for cluster elongation
in this experiment.
Table 4.2 Effect of the different gelling agents on multiplication fold and shoot elongation
Gelling agent Shoot buds Shoot length (cm)
Agar 5.9±0.69a 2.1±0.14b
Agar gel 5±0.10b 2.5±0.28b
Gelrite 4.3±0.06b 3.29±0.17a
LSD (0.05) 0.81 0.42
Shoot elongation obtained on gelrite medium was significantly higher as
compared to that obtained on medium gelled with agar-gel and agar (Figure
4.3). However, vitrification of shoots was observed on gelrite medium. The
variations observed for explant multiplication and shoot elongation on agar and
agar gel medium, were however, found to be statistically non significant, as
evaluated by Duncan’s Multiple Range Test (DMRT). Considering the higher
multiplication obtained on agar gelled medium, agar was employed as the
gelling agent in all subsequent studies.
4.2.3 Culture multiplication
4.2.3.1 Effect of different cytokinins on multiplication
The effect of BAP, 2iP and Kn at concentrations varying between 0.5 to 10.0 M
was studied on multiplication and growth response of shoot clusters (Figure 4.4)
Since, good cluster growth was obtained after the first five to six sub-cultures,
data for multiplication fold was, henceforth, recorded as the number of clusters,
instead of shoot buds (also referred as explants or propagules).
Among the eighteen combinations tested, a maximum of 3.6 multiplication
fold was obtained for the clusters inoculated on medium supplemented with 4

Micropropagation
TERI University - Ph.D. Thesis, 2006
M BAP (Figure 4.4). It was observed that the type and concentration of
cytokinin had a significant effect on the multiplication fold obtained on each
tested media combination. The lowest multiplication rate of 1.2 was obtained on
MS basal medium preceded by 2-fold multiplication on medium supplemented
with 0.5 M 2iP. For each type of cytokinin tested, the multiplication fold
showed a consistent increase upto a certain level (Figure 4.4), beyond which the
multiplication rate declined.
Also, the type of cytokinin and their concentration had a definite effect on
the cluster height. Among the three cytokinins tested, 2iP showed the most
optimal effect on cluster elongation. The elongation obtained for six
concentrations of 2iP ranged from a minimal of 2.13±0.09 cm to a maximum of
2.73±0.57. In case of BAP, these values ranged between 0.9±0.11 and 2.6±0.44
cm. Thus, the six different concentrations of BAP showed the highest variance in
terms of shoot elongation. At the highest concentration of 10 M BAP, the shoot
clusters showed stunted shoot growth, though, an average multiplication fold of
2.5 was obtained from each cluster. In case of explants inoculated in media with
different concentrations of Kn no variation on shoot elongation was observed.
A prolonged incubation of six weeks in culture medium, instead of the usual
4-week culture period, resulted in significant increase in cluster elongation.
However, this increase in cluster height was attributed to increase in internodal
distance. Hence, even after six weeks of incubation period, there was no increase
in the number of explants or the multiplication fold. Since, 4 M BAP produced
the most desirable results, both in terms of multiplication fold (3.6) and
cluster elongation (2.6 cm), further experiments were conducted with MS
medium gelled with 0.8% agar and supplemented with 3% sucrose
and 4 M BAP as constant. The media composition was employed as control
media while conducting further experiments for fine-tuning the media
composition.
4.2.3.2. Synergistic effect of BAP along with 2iP and Kn on cluster
multiplication and elongation
Synergistic effect of 4 M BAP with varying concentration of 2iP and Kn was
studied for cluster multiplication and elongation. The average multiplication
fold varied significantly (P= 0.0071) with the type and concentration of
cytokinin that was added to the control medium (MS + 4 M BAP + 3% sucrose).
For instance, maximum multiplication fold of 4.5 was obtained on MS + 4 M
BAP + 1.5 M 2iP medium, which was followed by 3.9-fold multiplication on
medium MS + 4 M BAP + 0.5 M 2iP (Table 4.3). A further increase in

Micropropagation
TERI University - Ph.D. Thesis, 2006
concentration of 2iP to 3 and 4 M, did not increase the multiplication fold,
which remained static at 3.7 for both the concentrations tested. It was observed
that the addition of Kn to control medium (MS + 4 M BAP) did not have any
significant effect on the regeneration response (Table 4.3). At 0.5 and 1.5 M
Kn, multiplication fold of 3.6 and 3.5 were obtained, which decreased to 3 and
2.9 folds when the concentration of kinetin was further increased to 3 and 4 M,
respectively.
The increase in shoot length for the above-discussed medium combinations,
was found to be statistically non- significant with a P value of 0.524.
Table 4.3 Synergistic effects of BAP (benzyl amino purine) with varying concentration of 2iP (6-[ , di-methylallylamino] purine) and Kn (kinetin), on multiplication fold and cluster height after 4 weeks of culture period
Cytokinins ( M) Number of propagules Cluster height (cm)
BAP 2iP Kn
4 0.5 - 3.9±0.43ab 2.2±0.10d
4 1.5 - 4.5±0.40a 2.8±0.3a
4 3 - 3.7±0.30bc 2.7±0.05abc
4 4 - 3.7±0.20bc 2.5±0.05abcd
4 - 0.5 3.0±0.23cd 2.2±0.16cd
4 - 1.5 3.6±0.49bcd 2.7±0.32ab
4 - 3 3.5±0.41bcd 2.4±0.32abcd
4 - 4 2.9±0.2d 2.4±0.32abcd
4 (control) - - 3.4±0.72bcd 2.3±0.21bcd
LSD 0.704 0.40
Maximum elongation of 2.8 and 2.7 cm was obtained for shoot clusters cultured
on control medium supplemented with 1.5 iP and 1.5 M Kn as well as 3 M
2iP separately.
4.2.3.3. Synergistic effect of 4 M BAP along with auxins on growth
of Swertia chirayita
Auxins, namely, IBA, IAA and NAA were tested along with 4 M of BAP to study
their effect on cluster multiplication and elongation. ANOVA for data recorded
for cluster multiplication and elongation showed highly significant P values for
the two variables. Nevertheless, addition of auxins to the control medium did
not have a positive effect on multiplication fold and cluster elongation (Table
4.4).

Micropropagation
TERI University - Ph.D. Thesis, 2006
Table 4.4 Synergistic effects of BAP and auxins on multiplication. All the concentrations are in M
BAP IBA IAA NAA Number of
propagules#
Cluster height
(cm)#
4 0.5 - - 2.43±0.15 bcd 2.35±0.36 ab
4 1 - - 2.5±0.2 bc 2.59±0.24 a
4 - 0.5 - 2.79±0.13 ab 2.53±0.28 ab
4 - 1 - 2.6±0.28 b 2.15±0.18b
4 - - 0.5 1.84±0.15 cde 1.41±0.17c
4 - - 1 1.75± 0.3 de 1.12±0.07 c
4 (control) - - - 3.32±0.9a 2.6±0.17 a
Basal medium - - - 1.65±0.21e 1.28±0.02 c
LSD (0.05) 0.65 0.38
Whereas 3.3 multiplication fold was obtained on the control medium (MS+ 4
M BAP), 2.79 fold multiplication was obtained on medium combination MS + 4
M BAP + 0.5 M IAA, followed by 2.6 multiplication fold on MS + 4 M + 1 M
IAA. The cluster height recorded for the two concentrations of IAA were 2.53
and 2.15 cm, respectively.
IBA when tested at 0.5 and 1 M concentration, along with 4 M BAP gave a
multiplication fold of 2.43 and 2.5, respectively. The cluster height recorded at
these two concentrations averaged to 2.35 and 2.59, respectively.
The effect of supplementing the control medium (MS + 4 M BAP) with 0.5
and 1.0 M NAA, were inhibitory both in terms of multiplication fold and cluster
elongation. Multiplication fold of 1.84 and 1.75 were obtained at 0.5 and 1.0 M
NAA and cluster height of 1.41 and 1.12 were recorded at these two
concentration, respectively. The cultures on NAA supplemented medium also
exhibited basal callusing.
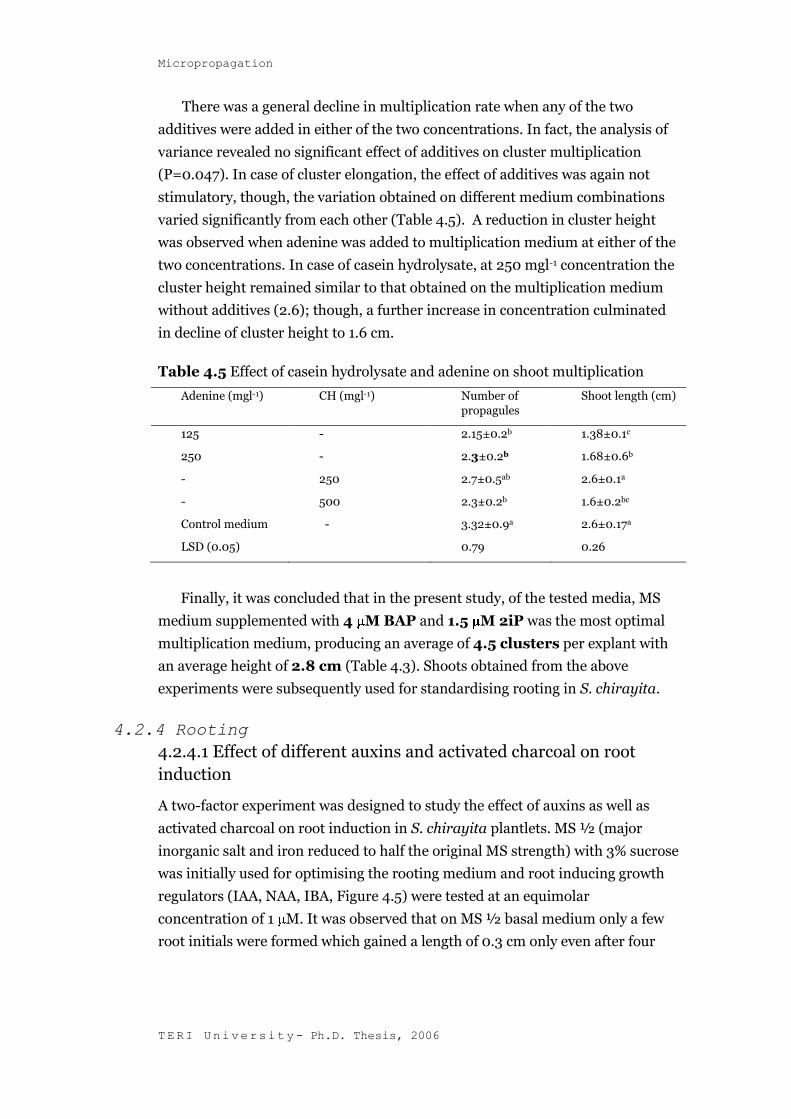
4.2.3.4 Effect of casein hydrolysate and adenine on cluster
multiplication and elongation
The addition of the two supplements, namely, adenine and casein hydrolysate
did not have a promotory effect on the overall regeneration response. Adenine
was added to the multiplication medium at 125 mgl-1 and 250 mgl-1
concentrations. Similarly, casein hydrolysate was added to the control medium
at 250 and 500 mgl-1 concentrations.
Micropropagation
TERI University - Ph.D. Thesis, 2006
There was a general decline in multiplication rate when any of the two
additives were added in either of the two concentrations. In fact, the analysis of
variance revealed no significant effect of additives on cluster multiplication
(P=0.047). In case of cluster elongation, the effect of additives was again not
stimulatory, though, the variation obtained on different medium combinations
varied significantly from each other (Table 4.5). A reduction in cluster height
was observed when adenine was added to multiplication medium at either of the
two concentrations. In case of casein hydrolysate, at 250 mgl-1 concentration the
cluster height remained similar to that obtained on the multiplication medium
without additives (2.6); though, a further increase in concentration culminated
in decline of cluster height to 1.6 cm.
Table 4.5 Effect of casein hydrolysate and adenine on shoot multiplication
Adenine (mgl-1) CH (mgl-1) Number of
propagules
Shoot length (cm)
125 - 2.15±0.2b 1.38±0.1c
250 - 2.3±0.2b 1.68±0.6b
- 250 2.7±0.5ab 2.6±0.1a
- 500 2.3±0.2b 1.6±0.2bc
Control medium - 3.32±0.9a 2.6±0.17a
LSD (0.05) 0.79 0.26
Finally, it was concluded that in the present study, of the tested media, MS
medium supplemented with 4 M BAP and 1.5 M 2iP was the most optimal
multiplication medium, producing an average of 4.5 clusters per explant with
an average height of 2.8 cm (Table 4.3). Shoots obtained from the above
experiments were subsequently used for standardising rooting in S. chirayita.
4.2.4 Rooting
4.2.4.1 Effect of different auxins and activated charcoal on root
induction
A two-factor experiment was designed to study the effect of auxins as well as
activated charcoal on root induction in S. chirayita plantlets. MS ½ (major
inorganic salt and iron reduced to half the original MS strength) with 3% sucrose
was initially used for optimising the rooting medium and root inducing growth
regulators (IAA, NAA, IBA, Figure 4.5) were tested at an equimolar
concentration of 1 M. It was observed that on MS ½ basal medium only a few
root initials were formed which gained a length of 0.3 cm only even after four

Micropropagation
TERI University - Ph.D. Thesis, 2006
weeks of incubation. On the contrary, 100% rooting was observed on MS ½
medium supplemented with 1 M of IAA, IBA or NAA separately.
In the present experiment MS ½ + 1 M NAA was found most optimal for
root induction and an average of 6.5 roots per shoot were induced, which gained
a length of 0.69 cm, after 4 weeks of incubation (Table 4.6). In case of medium
supplemented with auxins IBA and IAA, the root length remained more or less
consistent for the two media (0.49 and 0.50 respectively), however the number
of roots induced on IBA supplemented medium (11.5) far exceeded that obtained
on IAA or NAA supplemented media (7.8 and 6.5 respectively).
Table 4.6 Duncan’s multiple range tests for the effect of different auxins on root induction
Growth Regulators (1 M)
Shoot length (cm) Root length (cm) Root Number
Basal 2.33± 0.10 ab 0.30±0.19 c 2.83±1.9c
IBA 2.53±0.36a 0.49± 0.12b 11.5±2.4a
IAA 2.56± 0.32a 0.50±0.14b 7.8±0.75b
NAA 2.26± 0.18b 0.69± 0.23a 6.5±1.04b
LSD 0.05 0.22 0.14 1.73
The analysis of variance of the recorded experimental data revealed that the
influence of auxins was highly significant both for the number of roots induced
(P= 0.000) and the root length (P=0.0004) recorded on each media
combination.
The influence of auxins on shoot elongation was however not highly
significant as is evident from ANOVA which gave a P value of 0.029. The shoots
inoculated on IBA and IAA supplemented medium grew to a length of 2.5 cm
each, while those inoculated on NAA supplemented medium averaged to 2.2 cm,
which was approximately the same as recorded for shoots cultured on MS ½
basal medium (2.3 cm).
This experiment also highlighted the influence of activated charcoal on the
quality of roots. The shoots inoculated on medium without activated charcoal
developed basal callusing and the roots remained thick, small and tuberous
(Figure 4.6a). Addition of activated charcoal improved the quality of roots. The
roots formed were longer and thinner (Figure 4.6b). The basal callusing
observed in shoots inoculated on medium devoid of activated charcoal was
conspicuously absent in media combinations supplemented with activated
charcoal. Hence, in the subsequent rooting experiments activated charcoal

Micropropagation
TERI University - Ph.D. Thesis, 2006
supplemented medium was chosen for further experimentation to avoid basal
callusing.
Table 4.7 Duncan’s multiple range test for the effect of activated charcoal on root formation in Swertia chirayita
Medium Shoot length# (cm)
Root length# (cm) Root Number#
Without activated
charcoal
2.58±0.28a 0.37±0.17b 7.75±4.35a
With activated charcoal 2.26±0.15b 0.61±0.19a 6.5±2.53a
LSD 0.05 0.158 0.10 1.22
Analyses of variance for the recorded data showed that activated charcoal had a
highly significant effect on shoot length (P= 0.0005) and root length (P=0.0001,
Table 4.7). The average root length on medium containing activated charcoal
was 0.61 cm as compared to 0.37 cm for roots formed on medium without
activated charcoal. Similarly, shoots inoculated on medium without activated
charcoal gained an average length of 2.58 cm as compared to 2.26 cm of average
length of shoots inoculated on medium with activated charcoal. However, the
number of roots inoculated on medium with and without activated charcoal (7.7
and 6.5, respectively) remained statistically non-significant as is also deduced
from the ANOVA results. The interaction of growth regulators and activated
charcoal was found to be non significant for all the three variables (Table 4.8).
Further experiments were conducted with MS ½ medium supplemented with 1
M NAA, and 500 mgl-1 activated charcoal.

Micropropagation
TERI University-Ph.D. Thesis, 2006
Table 4.8 Analysis of variance (ANOVA) for the effect of auxins (growth regulators) and activated charcoal on the theree variables namely, shoot length, root length and root number
Source Shoot length Root length Root numbers
Degree
of
freedom
Mean
square
Probability Degree
of
freedom
Mean
square
Probability Degree
of
freedom
Mean
square
Probability
Individual
factor
Growth
regulator
(GR)
3 0.13 0.029* 3 0.14 0.0004*** 3 76.8 0.000***
Activated
charcoal
(AC)
1 0.63 0.0005*** 1 0.34 0.0001*** 1 8.1 0.604ns
Interaction
(GR X AC) 3 0.075 0.12ns
3 0.029 0.132ns
3 5.5 0.076ns
*** Very highly significant P<0.0001; ** highly significant P < 0.001, * less significant P<0.01; ns non significant

Micropropagation
TERI University-Ph.D. Thesis, 2006
4.2.4.2 Effect of different medium strengths on rooting
With the above mentioned factors (500 mgl-1 activated charcoal + 1 M NAA)
as constant, MS strengths were varied to study the effect of salt strengths on
root induction. The salt strengths were varied as shown in Table 4.9. The
roots induced on MS full strength medium were found to be small, tuberous
and were formed as dense clusters when compared to those induced on
medium with reduced MS salt strength. It was observed that shoot length
remained more or less same as the salt strengths were reduced from full to
half and quarter strengths (2.9, 2.4 and 2.7 cm, respectively); however, a
further decrease in salt strengths to one-eighth and one-tenth of the original
had a bolting effect on shoot length which shot to 4.76 and 4.38 cm,
respectively. This induced lankiness in the shoots, thus, adversely affecting
the plant vigour. These shoots were not suitable for further hardening due to
their lankiness.
Table 4.9 Effect of the different MS strengths on root induction and plant growth
Variation in MS
major and Iron
Medium
code
Shoot length
(cm)
Root length
(cm)
Root
number
Full SSR1 2.93±0.36b 0.59±0.07b 9.16±1.5a
Half SSR2 2.49±0.13b 1.18±0.32a 9.6±0.89a
Quarter SSR3 2.74±0.47b 1.10±0.35a 11.16±1.24a
One-eighth SSR4 4.76±0.73a 0.84±0.52ab 10.2±2.5a
One-tenth SSR5 4.38±0.55a 0.53±0.27b 10.93±2.7a
MS organics only SSR6 2.78±0.39b 0c 0b
No salt (water agar) SSR7 1.76±0.07c 0c 0b
LSD (0.05) - 0.51 0.476 1.92
The number of roots induced on these media combinations was however more
or less the same, with a lowest of 9 and a highest of 11 roots. However, the
variations in the salt strengths did induce a variable response in terms of root
length. The length of roots increased on MS ½ and ¼ medium (1.18 and 1.1,
respectively) when compared to root length on MS full (0.59). A further
reduction in salt strength did not result in further increase in root length,
instead the root length steadily declined from 0.84 to 0.53 cm as salt strength
was decreased from MS 1/8 to MS 1/10. In medium combination coded SSR6,
MS inorganic salts were completely eliminated and only MS organic salts were
added. On the other hand in medium coded SSR7 both, MS inorganic and
organic salts were completely eliminated (water agar medium). However,

Micropropagation
TERI University - Ph.D. Thesis, 2006
sucrose + 1 M NAA was supplemented in all the medium combinations tested.
In the later two medium combinations (lacking MS salts) a conspicuous absence
of root induction was observed. On the basis of these observations, MS ½ was
considered most optimal for root induction in S. chirayita.
4.2.4.3 Effect of sugar concentration on root induction
The effect of sucrose concentration (1-3%) was tested. It was observed that on
increasing the sugar concentration from 1 to 3% in the rooting medium, a
distinct increase in shoot length from 1.59 to 2.4 was observed. The number (5,
5.9 and 6.9) of induced roots, however, remained statistically similar at 1, 1.5
and 2% sugar concentration, respectively (Table 4.10). Only at 3% sucrose
concentration a statistically significant increase was observed in the number of
roots formed. However, root length was significantly affected by sucrose
concentration as compared to the root numbers. It was observed that root length
increased from 0.34 to 1 and 1.5 on medium supplemented with 1, 1.5 and 2%
sucrose, respectively. A further increase to 3% did not enhance the root length
and an average of 0.96 cm long roots were recorded. 2% sucrose concentration
was thus considered to be the most optimal for root induction with respect to
root number and root length.
Table 4.10 Effect of the varying sugar concentration on root induction and plant growth
Sucrose
concentration (%)
Root length (cm) Root number Shoot length (cm)
1 0.34±0.2 b 5.28±1.23b 1.59±0.31b
1.5 1.04± 0.4 ab 5.94±1.08b 1.97±0.41ab
2 1.49± 0.6a 6.96±0.6b 2.43±0.6a
3 0.96± 0.61 ab 9.10±1.71a 2.44±0.1a
LSD (0.05) 0.751 1.85 0.62
4.2.4.4 Effect of the medium content in culture vessel on root
induction
An attempt was made to evaluate the effect of the medium volume in the culture
vessels on root induction as well as on plant vigour. The volume of medium in
the culture vessel was varied from 20 ml to 60 ml and 15 shoots were inoculated
in each culture vessel. It was observed that the amount of medium in the culture
vessel does not affect root induction in S. chirayita (Table 4.11). The number of
roots induced and the length gained by them remained statistically non
significant for 20, 30 and 40 ml of medium pouring. However, 20 and 30 ml of

Micropropagation
TERI University - Ph.D. Thesis, 2006
the medium in 400 ml culture jars dried very soon due to exhaustion of media
and this in turn adversely affected the growth of the plant. The leaves dried and
the plants wilted within the culture period of 4 weeks itself. In case of 40, 50 and
60 ml of medium, the plant remained healthy. In fact, a significant increase in
length was observed in shoots (3.1 and 3.05 respectively) inoculated in culture
vessels with 50 and 60 ml of medium as compared to an average of 2.7 cm long
plantlets in 40 ml medium. However, this increase in shoot length resulted in
lanky plants, which adversely affects the hardening process. Therefore, it was
concluded that 40 ml of medium volume was optimal for 15 shoots in each
culture vessel.
Table 4.11 Effect of the different volumes of medium poured in the culture vessel on percentage rooting
Medium Volume
(ml)
Shoot length
(cm)
Root length (cm) Root number
20 2±0.63b 1.21±0.6a 4.6±0.02b
30 2.23±0.12b 1.45±0.32a 6.48±0.85b
40 2.75±0.39b 0.954±0.23a 6.84±2.1b
50 3.10±0.29a 1.23±0.2a 10.27±1.79a
60 3.05±0.3a 1.18±0.26a 12.05±1.2S
LSD (0.05) 0.51 0.476 2.20
4.2.5 Acclimatisation and field transplantations
4.2.5.1 Ex vitro hardening and effect of different months on the
survival rate
At a commercial scale hardening is carried out in the green house so as to
harden plants in large numbers. The green house experiments were conducted at
Gual Pahari (Gurgaon, Haryana), where the temperatures vary throughout the
year. The ambient temperature extremes recorded for the region varied from a
maximum of 40 0C to a minimum of 3 0C round the year. The temperature
conditions were maintained within a range of 11 to 27 0C throughout the year in
the green house. The rooted plants were, therefore, taken out during different
months of the year to record percentage survival of the plants. A minimal
survival rate of 20-35% was recorded during the months of May, June, July and
August (Table 4.12). However, the plants taken out after September showed a
substantial increase in survival percentage.

Micropropagation
TERI University-Ph.D. Thesis, 2006
Table 4.12 Acclimatisation success rate of the micropropagated S. chirayita plantlets in green house at Gual pahari, Haryana, India, during different months of the year
Month Temperature (oC) Relative humidity (%)
In Green House
Survival
percentage
Ambient Green house Minimum Maximum
Minimum Maximum Minimum Maximum
Dec 5.2 26 11.25 16.1 77.25 80 93.96
Jan 3 22.3 11.75 18.5 63.75 78.5 95.4
Feb 8.3 28.2 14.8 19.8 63 78.8 96.4
March 16.4 32.3 17.25 20 68.5 80.5 97
April 21.2 38.4 18.4 21.6 65.2 80.4 90
May 24.7 39.3 24.3 27.0 80.0 81.3 22
June 25.5 37 25 27 76.75 80.3 20
July 26 34.6 24.75 26.8 77.75 80 26
August 25.3 35.8 24.25 27.2 80 81.5 35
Sept 22 38.1 21.25 24.8 78 80.5 80
Oct-Nov 13.8 17.9 20 23 77.25 80 86

Micropropagation
TERI University-Ph.D. Thesis, 2006
In fact, a marked increase in percent survival from 80% (September) to 97%
(March) was observed. Hence, based on these observations it was concluded that
months of December to April are most conducive for successful hardening of S.
chirayita plantlets in green house conditions. Figure 4.7a shows tissue-cultured
plantlets freshly transferred to polybags and kept in green house for
acclimatisation. Figure 4.7b represents the fully hardened plants in nursery
ready for transplantation to the field. A total of 3596 plants were subjected to
hardening during the months of December to March, and high survival
percentage of 94.5 was recorded. Finally, a total of 3400 S. chirayita plantlets
were transferred to the shade area. These plants were taken to Mukteshwar
(Uttaranchal state) at an altitude of 2286 m above sea level for final
transplantation in March and April.
4.2.5.2 In vitro hardening
During the hardening trials in green house lowest survival percentage was
observed from May to August, when temperatures ranged between maximum of
27 0C to a minimum of 24 0C. Hence, in vitro hardening trials were conducted in
growth room with controlled conditions of temperature (22 2 0C) during the
months of May and August. A successful hardening response of 95.5% and
92.7% was obtained in May and August, respectively. These plants were
transplanted at Mandal (1568 m above sea level) and Mukteshwar (2286 m
above sea level) in the months of June and September. Figure 4.8 shows the
plantlets during in vitro hardening and Figure 4.9 represents the different stages
from rooting to successful hardening of S. chirayita plantlets.
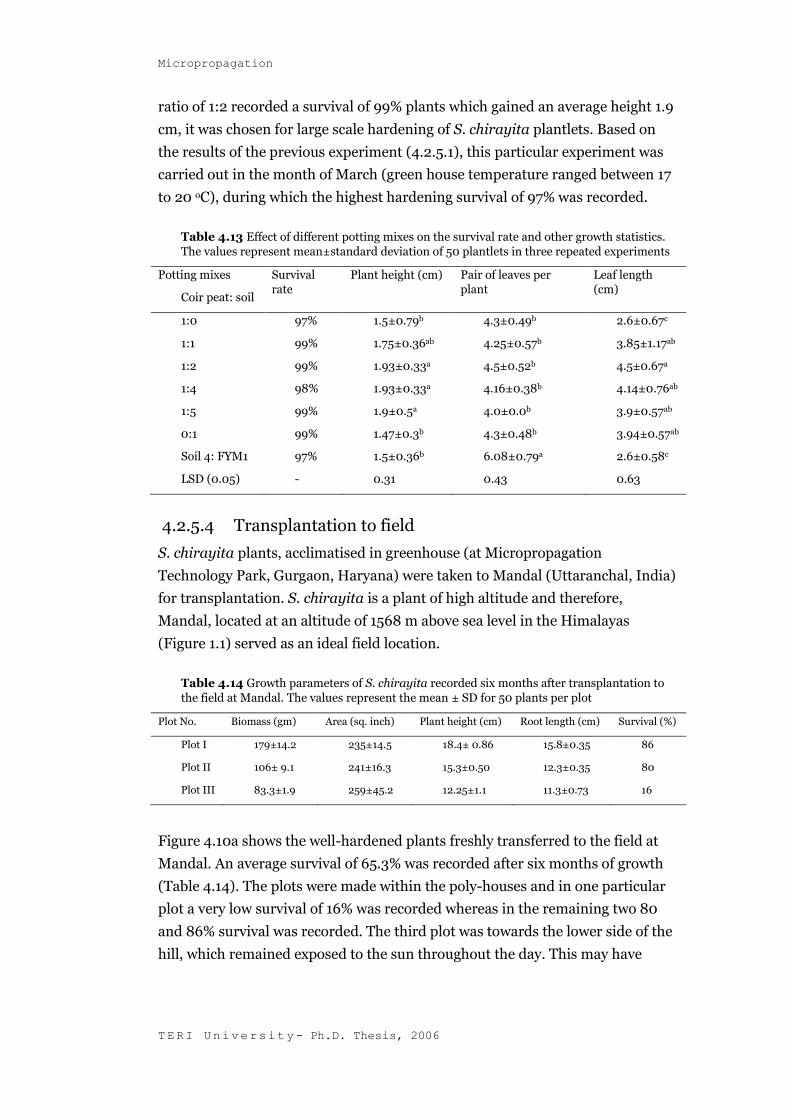
4.2.5.3 Effect of different potting mixes on initial survival and plant
growth
Different combination of coir-peat and soil were tested during transplantation
trials. However, no significant variation was found with respect to percent
survival in the different potting mixes tested and a high survival percentage of
97-99 was recorded. However, the growth of the plant was found to vary
depending on the potting mixes. Highest plant elongation (1.9 cm) was achieved
in potting mixes of coir peat and soil in varying ratios of 1:2, 1:4 and 1:5. When
soil and coir peat were used solely as potting mixes, the plant height did not
increase beyond 1.5 cm. In case of potting mix containing soil and FYM in the
ratio of 4:1, there was a substantial increase in the number of leaves produced
per plant (12) as compared to an average of 8 leaves per plant in other potting
mixes (Table 4.13). Since the potting mix containing coir peat and soil in the
Micropropagation
TERI University - Ph.D. Thesis, 2006
ratio of 1:2 recorded a survival of 99% plants which gained an average height 1.9
cm, it was chosen for large scale hardening of S. chirayita plantlets. Based on
the results of the previous experiment (4.2.5.1), this particular experiment was
carried out in the month of March (green house temperature ranged between 17
to 20 oC), during which the highest hardening survival of 97% was recorded.
Table 4.13 Effect of different potting mixes on the survival rate and other growth statistics.
The values represent mean±standard deviation of 50 plantlets in three repeated experiments
Potting mixes
Coir peat: soil
Survival rate
Plant height (cm) Pair of leaves per plant
Leaf length (cm)
1:0 97% 1.5±0.79b 4.3±0.49b 2.6±0.67c
1:1 99% 1.75±0.36ab 4.25±0.57b 3.85±1.17ab
1:2 99% 1.93±0.33a 4.5±0.52b 4.5±0.67a
1:4 98% 1.93±0.33a 4.16±0.38b 4.14±0.76ab
1:5 99% 1.9±0.5a 4.0±0.0b 3.9±0.57ab
0:1 99% 1.47±0.3b 4.3±0.48b 3.94±0.57ab
Soil 4: FYM1 97% 1.5±0.36b 6.08±0.79a 2.6±0.58c
LSD (0.05) - 0.31 0.43 0.63
4.2.5.4 Transplantation to field
S. chirayita plants, acclimatised in greenhouse (at Micropropagation
Technology Park, Gurgaon, Haryana) were taken to Mandal (Uttaranchal, India)
for transplantation. S. chirayita is a plant of high altitude and therefore,
Mandal, located at an altitude of 1568 m above sea level in the Himalayas
(Figure 1.1) served as an ideal field location.
Table 4.14 Growth parameters of S. chirayita recorded six months after transplantation to
the field at Mandal. The values represent the mean ± SD for 50 plants per plot
Plot No. Biomass (gm) Area (sq. inch) Plant height (cm) Root length (cm) Survival (%)
Plot I 179±14.2 235±14.5 18.4± 0.86 15.8±0.35 86
Plot II 106± 9.1 241±16.3 15.3±0.50 12.3±0.35 80
Plot III 83.3±1.9 259±45.2 12.25±1.1 11.3±0.73 16
Figure 4.10a shows the well-hardened plants freshly transferred to the field at
Mandal. An average survival of 65.3% was recorded after six months of growth
(Table 4.14). The plots were made within the poly-houses and in one particular
plot a very low survival of 16% was recorded whereas in the remaining two 80
and 86% survival was recorded. The third plot was towards the lower side of the
hill, which remained exposed to the sun throughout the day. This may have

Micropropagation
TERI University - Ph.D. Thesis, 2006
likely resulted in wilting of the plantlets at an early stage, immediately after
transplantation, thus resulting in the high mortality in the initial stages itself.
The results emphasise the significance of initial care required while handling the
tissue culture raised plants.
Besides recording the shoot length and area occupied by the plant,
destructive harvesting was done to determine the root length and the average
biomass of the plants. The root length averaged from 11 to 15 cm, whereas,
biomass per plant (fresh weight) averaged to 122 gm. Figure 4.10b show the
transplanted S. chirayita plantlets after six months of growth. Figure 4.11 gives a
comparative picture of the micropropagated plants at three months, six months
and after one year of growth. After the first year of growth the plant gained an
average of two-feet height as is evident in the figure. After bolting, the plant
enters into flowering phase (Figure 4.12 a). The pictures represent the flowering
twig (Figure 4.12 a, c) and a closer view of the flower (Figure 4.12 d). Figure
4.12b represents the root (the most potent part of the plant) after the completion
of flowering season. These illustrations represent the success of transplantation
trial carried out at Mandal.

Micropropagation
TERI University - Ph.D. Thesis, 2006
A schematic presentation for mass propagation of Swertia chirayita
**94.5% hardening represents the average of acclimatisation success obtained
for the months of December to April, which were found most conducive for
hardening in greenhouse conditions
Seeds
Transfer of four week-old seedlings to MS + 3µM BAP + 3%sugar + 0.8% agar
Germination on MS ½ medium + 3%sugar + 0.8% agar
(80% germination)
Subculture on MS with 4 µM BAP + 1.5 µM 2iP+ 3%sucrose + 0.8% agar (every 4 wk @ 4.5 multiplication fold)
Transfer of rootable shoots to MS ½ + 2% sucrose + 1µM NAA + 500 mgl-1 activated charcoal + 0.8% agar (4 wk)
(100% rooting)
Hardening of rooted shoots in green house in poly-bags containing
potting mix of coir-peat and soil (1:2) (15 days, 94.5% hardening**)
success)
Maintenance of hardened plants in nursery before final field trials at higher altitudes (temperate conditions)

Micropropagation
TERI University - Ph.D. Thesis, 2006
Production plan for 100,000 plants of Swertia chirayita - a schematic presentation
A surplus of 7000 plants was produced, keeping margin for contamination and
mortality during hardening.
Multiplication cycle (4 culture passages of 4 wk each) with 4.5 fold multiplication
21,200 shoot clusters
Hardening in green house
Final passage before rooting
12 nodal cultures
Establishment phase (5 passages of 4 wk each)
52 shoot clusters
Transfer of single shoots to rooting medium (4 wk)
(21,200*5=1,06,000 shoots)
12 nodal explants derived from four-week-old in vitro germinated
seedlings
(Week 1)
(4 wk)
(24 wk)
(40 wk)
(44 wk)
(48 wk)
(52 wk)

Micropropagation
TERI University - Ph.D. Thesis, 2006
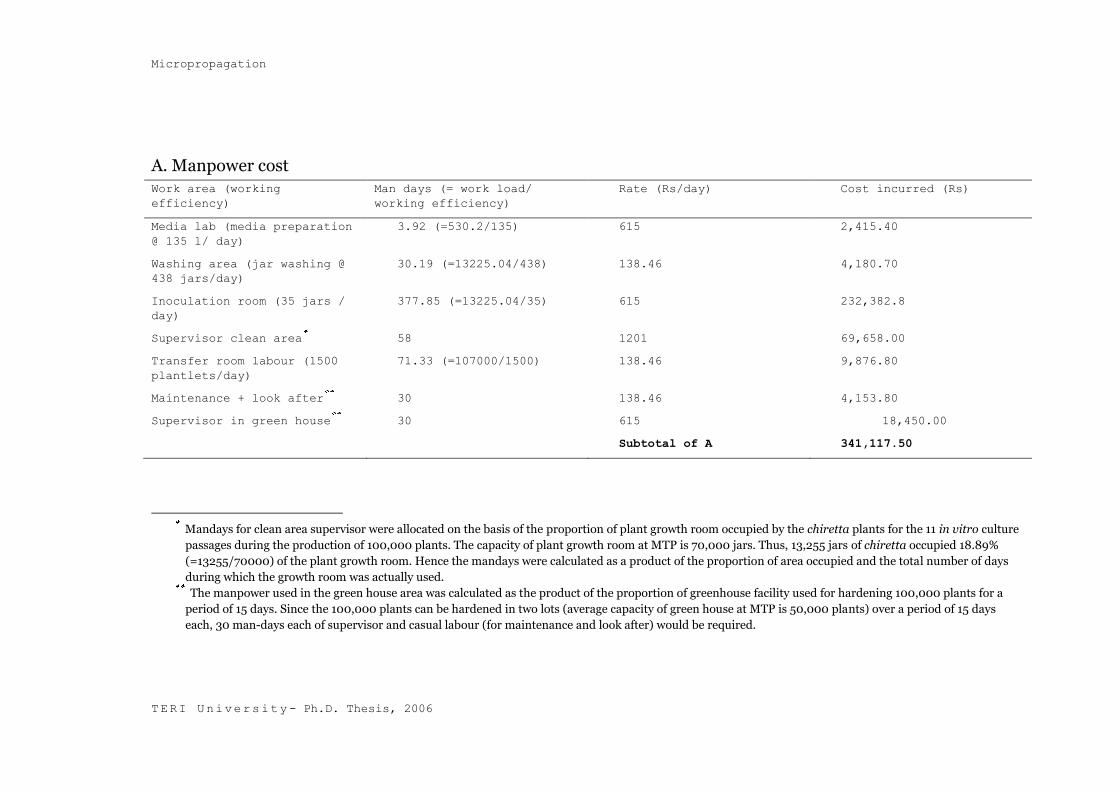
4.2.6 Cost Calculations
The cost of producing micropropagated plants at the Micropropagation
Technology Park (MTP) of The Energy and Resources Institute (TERI) have
been divided into two parts, namely, the direct cost and the indirect cost.
4.2.6.1 Direct Cost
Direct cost includes media cost, inoculation cost as well as the hardening cost,
at full occupancy and the costs incurred towards manpower and electricity
consumption. The parameters that decide the direct cost of the plant are
multiplication fold, rooting percentage and the hardening survival. The
parameters taken into consideration during production of 100,000 plants of S.
chirayita have been summarised below:
Parameters
Multiplication fold 4.5 in four weeks
Rooting percentage 100%
Hardening survival percentage 95%
Number of clusters in multiplication 5 shoot-clusters per culture
vessel
Number of shoots in rooting 15 shoots per culture vessel
Considering a 5% loss during the hardening stage and keeping a margin of 1%
for contamination, production of 1,07,00o plants was targeted in order to
produce a total of 1,00,000 successfully hardened plants of chiretta. It was
estimated from back calculations that a minimum of thirteen explants would be
required to eventually produce 1,07,000 rooted plants in eleven culture passages
(5 culture passages during establishment phase + 5 multiplication cycles + 1
rooting passage). Thus, during the establishment phase 13 nodal explants were
initially taken from 13 seedlings. The nodal explants established themselves into
clusters at the end of the fifth culture. In case of nodal explants 6 explants per
culture vessels were inoculated during the culture establishment stage. In case of
shoot clusters, five clusters were inoculated per culture vessel and a total of
fifteen rootable shoots were inoculated per culture vessel. Media pouring in each
jar was maintained at 40 ml throughout the production period. Table 4.15 gives
Inoculation cost includes the cost of consumables such as tissue paper, blades, rectified
spirit, sterile water, cling films, papers, sterilisation beads, savlon, garbage bag etc.
Culture vessel here refers to 400 ml jars used for commercial production of the chiretta
plants

Micropropagation
TERI University - Ph.D. Thesis, 2006
details regarding the media cosumption and number of culture vessels used
during production of 100,000 plants of S. chirayita.

Micropropagation
TERI University-Ph.D. Thesis, 2006
Table 4.15 Details of medium consumption and number of culture vessels used during production of 100,000 plants of Swertia chirayita
Stage Culture
passage
Number of
shoots/clusters
No. of culture vessels
(400 ml jars)
Medium consumption
(in litres, @ 40 ml/
culture vessel)
Establishment
phase
5 culture
passages
13 nodal explants 2*5 (6 nodal
explants/culture vessel)
0.4
Multiplication
phase
6th 58 clusters 12 (5 clusters/culture
vessel)
0.5
7th 261 clusters 52 2.08
8th 1174 clusters 235 9.4
9th 5284 clusters 1057 42.28
10th 23,778 clusters 4756 190.24
Rooting 11th 23,777*5=1,07,000
shoots
7133 (15 shoots/culture
vessel)
285.32
Grand total 13,255.04 530.2 litres
Micropropagation
TERI University - Ph.D. Thesis, 2006
A. Manpower cost Work area (working
efficiency)
Man days (= work load/
working efficiency)
Rate (Rs/day) Cost incurred (Rs)
Media lab (media preparation
@ 135 l/ day)
3.92 (=530.2/135) 615 2,415.40
Washing area (jar washing @
438 jars/day)
30.19 (=13225.04/438) 138.46 4,180.70
Inoculation room (35 jars /
day)
377.85 (=13225.04/35) 615 232,382.8
Supervisor clean area 58 1201 69,658.00
Transfer room labour (1500
plantlets/day)
71.33 (=107000/1500) 138.46 9,876.80
Maintenance + look after 30 138.46 4,153.80
Supervisor in green house 30 615 18,450.00
Subtotal of A 341,117.50
Mandays for clean area supervisor were allocated on the basis of the proportion of plant growth room occupied by the chiretta plants for the 11 in vitro culture
passages during the production of 100,000 plants. The capacity of plant growth room at MTP is 70,000 jars. Thus, 13,255 jars of chiretta occupied 18.89%
(=13255/70000) of the plant growth room. Hence the mandays were calculated as a product of the proportion of area occupied and the total number of days
during which the growth room was actually used.
The manpower used in the green house area was calculated as the product of the proportion of greenhouse facility used for hardening 100,000 plants for a
period of 15 days. Since the 100,000 plants can be hardened in two lots (average capacity of green house at MTP is 50,000 plants) over a period of 15 days
each, 30 man-days each of supervisor and casual labour (for maintenance and look after) would be required.

Micropropagation
TERI University - Ph.D. Thesis, 2006
B. Electricity Electricity consumption
during
Cost (Rs) Cost incurred (Rs)
Autoclaving
Dry cycle
Media cycle
Unit cost (Rs)
9
9
Units consumed per
cycle
22.5
45
Total number of cycles
20
32
4,050 (=9*22.5*20)
12,960 (=9*45*32)
Laminar flow (two
seater)
Electricity cost / flow /
day
Flow required for - -
184.93 189 days - 34,951.77 (=184.93*189)
Plant growth room 0.17 / jar/ day 13225.04 jars - 2,248.25
(=0.17*13225.04)
Green house 1.5 / plant 107000 plants 160,500 (=1.5*107000)
Subtotal of B 214,710.02
C. Media cost
Break up Cost per litre (Rs) Total media (l) Cost incurred (Rs)
LPG cost towards media melting 0.6 530.2 318.12
Material cost of media 28.1 530.2 14,898.62
Subtotal of C 15,216.74
Calculated on the basis of the number of sterile petri plates required during inoculations of 13225.04 jars
Calculated on the basis of the total number of media jars to be autoclaved and the capacity of the autoclave

Micropropagation
TERI University-Ph.D. Thesis, 2006
D. Inoculation cost Cost / jar/ day (Rs) Jars/day Cost incurred (Rs)
37.85 377.85 (= total number
of jars / jars inoculated per day)
14301.93
Subtotal of D 14301.93
Direct cost of 100,000 chiretta plant = grand total of A + B + C + D
= Rs 283949 + Rs 214710.02 + Rs 15216.74 + Rs 341117.5
= Rs 585346.2
Direct cost of one chiretta plant = (585346.2/100000) = Rs 5.85
4.2.6.2 Indirect Cost
The indirect cost towards production of micropropagated plants at a commercial
scale is an important parameter to be considered. It includes the administrative
cost, depreciation of the infrastructure and equipment, maintenance cost,
professional cost associated with supervision as well as transport of the plants to
the transplantation site. The cost of following was taken into account while
calculating the indirect cost of the micropropagated chiretta plants
A. Annual administrative cost
B. Annual maintenance cost
C. Depreciation of building
D. Depreciation of equipment
E. Transport
F. Professional cost
A. Annual administrative cost
The annual administrative and maintenance cost on a yearly basis were
calculated separately from the expenses incurred round the year, during which
an average of 140, 000 plants (70% of the production capacity) were produced.
The administrative cost totalled to Rs. 500,000.
A. Annual administrative cost Yearly
Administartive
cost (Rs)
Cost per plant in Rs
(=total yearly cost / total
number of plants
produced round the year)
Total
plants
produced
Cost incurred in Rs
(= cost / plant *
total number of
plants produced)
500,000 0.357 (=500000/140000) 107,000 38,214.28

Micropropagation
TERI University - Ph.D. Thesis, 2006
B. Annual repair and maintenance cost Yearly
Maintenance
cost (Rs)
Cost per plant in Rs
(=total yearly cost
/ total number of
plants produced
round the year)
Total
plants
produced
Cost incurred
in Rs (= cost
/ plant *
total number
of plants
produced)
673,162 0.48
(=673162/140000)
107,000 38,246.88
C. Depreciation of building
The original cost of MTP is estimated at Rs. 1,50,51,000. A depreciation of 5%
flat was levied on this cost and was added accordingly towards production cost
of the plants.
Original
cost (Rs)
Yearly
depreciation
(@ 5% flat)
Cost per plant
(Rs)
Total Cost incurred (Rs)
1,50,51,000 752,550 0.537
(=722550/140000)
57,516.32
(=[722550/140000]/107000)
D. Depreciation of equipment
Total cost of equipments
Equipment Cost (Rs)
Hot air oven 36,575
Refrigerator 36,519
Refrigerator 38,000
Glass bead steriliser 19,855
pH meter 10,044
Laminar flow 248,240
Autoclave 500,590
Autoclave 280,732
Grand total 1,170,555
The depreciation cost on equipments at a rate of 10% flat comes out as Original
cost
(Rs)
Yearly
depreciation
(@ 10 %
flat)
Cost per plant
(Rs)
Total Cost incurred (Rs)
1,170,555 1,17,055.5 0.0836
(=117055.5/140000)
8,946.378
(=[117055.5/140000]/107000)

Micropropagation
TERI University - Ph.D. Thesis, 2006
E. Transport
The transport cost included the cost incurred towards loading of 100,000 plants
and transportation. This has been calculated as follows:
Loading cost Loading
efficiency
Man days
(=Total
plants/
working
efficiency)
Rate (Rs) Cost incurred (Rs)
1000
plants/man-
hour
13.3 man days 138.46 1846.1
The transport cost of 100,000 plants was calculated at Rs. 0.40 per plant.
Vehicle cost Cost per (Rs) Total
plants
transported
Cost incurred
(= cost /
plant * total
number of
plants
transported)
0.40 100,000 40,000
Total transport cost = loading cost + vehicle cost = Rs 1846.1 + Rs 40000 = Rs
41846.1
F. Professional cost Man days Rate per day (Rs) Cost incurred (Rs)
26 8457.20 219,887.20
Indirect cost of 100,000 chiretta plant = grand total of A + B + C + D + E + F
= Rs 38214.28 + Rs 11464.28 + Rs 57516.32 + Rs 8946.37 + Rs 41846.1 + Rs
219,887.20
= Indirect cost of one chiretta plant = (417909.70 / 100000) = Rs 4.17
4.2.6.3 Total Cost
Per plant cost of chiretta thus turns out to be = direct cost + indirect cost
= Rs 5.85 + Rs 4.17
Taken as 10% of the 52 weeks (260 man days) required for producing 100,000 chiretta
plants

Micropropagation
TERI University - Ph.D. Thesis, 2006
= Rs 10.02
Based on these calculations it can be inferred that the production of chiretta is
not a commercially feasible proposition. However, the present protocol can be
used for initial bulking up of the chiretta plantlets, which can later be
reintroduced in their natural habitat and established in herbal gardens as a
viable conservation measure. Once populations of elite chiretta germplasm has
been established, further propagation can be undertaken through seeds.

Micropropagation
TERI University - Ph.D. Thesis, 2006
Regeneration in Swertia chirayita from leaf explants
4.2.7.1 Response of different explants for regeneration
Leaf, internode and root explants of S. chirayita from in vitro grown plants were
tested for callus induction and regeneration response. The root explants and
internodal segments were cut into ca. 1 cm segments. The leaf explants were cut
into ca. 1 cm X 1 cm sections and were inoculated on medium slants. The MS
medium was modified for the regeneration experiments; with major and iron
salts reduced to half of the original MS salt strength. The rest of the components
were kept as the original MS medium. In a preliminary experiment, effect of
2,4,5-T (0.5 to 4 mg l-1) along with 0.5 mgl-1 BAP was tested for callus and shoot
bud induction response. The explants showing viable response were transferred
to fresh medium of similar composition after six weeks. Higher concentration of
2,4,5-T, ranging from 1-4 mgl-1 resulted in water uptake (visible as swelling of
explants) as well as cell proliferation (expansion of explants). The explants
remained green for 12 weeks (2 culture passages) and subsequently turned
brown and died, without showing any evidence of callus induction or
organogenesis. The root explants and internodal segments showed no response
at 0.5 mg l-1 2,4,5-T, though in case of leaf explants, organogenesis and callus
induction was observed.
Different combinations of auxins and cytokinins were also tested for the
regeneration response. On the medium combination supplemented with NAA
and IBA, each at 0.75 mgl-1 concentrations, shoot buds were induced in root
explants only (Figure 4.13). However, on repeating the experiments, it was
observed that this response was sporadic and not reproducible. These
preliminary experiments enabled selection of leaf explants for further
experimentation with MS half supplemented with BAP and 2,4,5-T (0.5 mgl-1
each).
4.2.6.2 Effect of varying concentration of BAP on callus induction
and shoot regeneration from leaf explants
The effect of varying concentrations of BAP
The BAP concentration was also varied to study its effect on shoot regeneration.
The regeneration response increased with increase in concentration of BAP from
0.25 to 2 mgl-1 (Figure 4.14). However, an increase in BAP concentration beyond
2 mgl-1 resulted in a decline in percentage regeneration. Callus formation was
conspicuous in most of the media combinations tested. On the other hand, in
absence of any growth regulator, the leaf explants exhibited minor curling and

Micropropagation
TERI University - Ph.D. Thesis, 2006
retained their green colour; however, no callus formation was noticed. The leaf
explants inoculated on medium supplemented with BAP at concentrations
ranging from 0.25 to 2.0 mgl-1 showed yellowish nodular growth, callus
induction and formation of root like outgrowths. Figure 4.15a illustrates the leaf
explant exhibiting initial swelling on regeneration medium whereas Figure 4.15b
show callus formed on leaf explant inoculated on MS 1/2 medium supplemented
with 2 mgl-1 BAP and 0.5 ml-1 2,4,5-T.
Figure 4.14 shows the graphical representation of the effect of varying
concentration of BAP on callus formation and shoot bud induction. At a
concentration range of 0.25 to 2.0 mgl-1, an increase in the percent regeneration
response from 16 to 76% was revealed. At 0.5, 1.0 and 2.0 mgl-1 concentrations of
BAP the leaf explants showed a regeneration response of 30, 36 and 76%,
respectively. The maximum numbers of 10 shoot buds per explant were induced
on medium supplemented with 1 and 2 mgl-1 BAP. A further increase in BAP
concentration to 3 mgl-1 led to a decline in regeneration response and only 16%
explants exhibited callus and shoot bud induction, whereas, the rest eventually
turned brown. A further increase of BAP to 4 mgl-1 led to browning of all the
explants inoculated on the medium.
The leaf explants inoculated on medium supplemented with 0.5 mgl-1, 1.0
mgl-1 and 2.0 mgl-1 of BAP when transferred to MS 1/2 basal medium (Figure
4.16), showed a substantial increase in shoots buds and shoot length. The
numbers of shoot buds increased from 8.3 (MS ½ + 0.5 mgl-1 2,4,5-T + 0.5 mgl-
1 BAP), 10.3 (MS ½ + 0.5 mgl-1 2,4,5-T + 1.0 mgl-1 BAP) and 10.7 (MS ½ + 0.5
mgl-1 2,4,5-T + 2 mgl-1 BAP) to 24, 34.8 and 43, respectively when transferred
from regeneration medium, as detailed in parenthesis, to MS 1/2 basal medium.
Figure 4.17 gives a comparison of the shoot bud induction in explants inoculated
on regeneration medium and then transferred to MS 1/2 basal. The shoot buds
elongated to an average of 3 cm after transfer to MS ½ basal medium (Figure
4.17 and 4.18).
The effect of additives on callus growth and shoot bud induction
The effect of three additives, namely casein hydrolysate (CH), glutamine and
adenine was analysed to study their effect on shoot bud induction from leaf
explants. Varying CH concentrations (125, 250 and 500 mgl-1) were tested
whereas glutamine and adenine were tested at 125 mgl-1 concentrations. It was
observed that the latter two, at the tested concentration did not have any
stimulatory effect on shoot bud induction. Though an initial callus formation
was observed on media supplemented with adenine and glutamine, the explants

Micropropagation
TERI University - Ph.D. Thesis, 2006
eventually turned brown (Figure 4.19). On the other hand, in case of CH, at
concentrations of 125 and 250 mgl-1, leaf explants showed expansion and
formation of nodular structures, without any shoot bud induction. As the
concentration of CH was increased to 500 mgl-1, the leaf explants exhibited
shoot bud regeneration. However, the addition of CH at any of the
concentrations did not improve the number of shoot buds induced on the
regeneration medium (MS ½ + 0.5 mgl-1 2,4,5-T + 2 mgl-1 BAP) as discussed in
the above section.
4.2.6.3 Effect of different gelling agents on explant growth and
callus induction
In order to study the influence of gelling agents on regeneration response, the
leaf explants were inoculated on medium gelled with 0.8% agar, 0.4% agar gel
and 0.2% gelrite. The medium composition was otherwise kept constant (MS ½
+ 0.5 mgl-1 2,4,5-T + 2 mgl-1 BAP). The initial expansion of the leaf explants was
observed for all the three gelling agents. However, gelrite and agar gel did not
favour any further growth and the explants turned brown by the end of the six-
week incubation period. In case of agar, the explants showed callus and shoot
bud induction. Hence, agar at 0.8% was consistently used in all further
experiments.
4.2.6.4 Effect of MS salt strength on the regeneration response of
leaf explants
The data recorded in percent values was arcsine transformed (section 2.6.2.1)
and the transformed data was subjected to analysis of variance. The data
expressed as percentage or proportions generally has a binomial distribution.
Since one of the assumptions of ANOVA is that the data should have a normal
distribution, therefore all the data recorded in percentage was arcsine
transformed prior to ANOVA.
The regeneration response of leaf explants varied significantly with
variations in MS salt strengths. Whereas on MS full strength medium only 24.7%
leaf explants exhibited shoot induction with an average of six shoots buds
induced per explant, on MS ½ and ¼ strength medium, an average of 78% and
44.7% explants showed regeneration response, respectively (Table 4.16). The
number of shoot buds per explant however did not vary much for these salt
strengths and an average of 11 and 10 shoots per explant were produced on MS
½ and MS ¼ media combinations, respectively.

Micropropagation
TERI University - Ph.D. Thesis, 2006
Table 4.16 Effect of MS salt strength on shoot buds induction in leaf explants
MS strength % Response Shoot buds/explant
MS full 24.7c (29.9)
# 6.3
b
MS half 78.4b (62.3) 11.3
a
MS quarter 44.7a (42) 10.3
a
LSD 7.7 3.05
# Values in the bracket ( ) represent the arcsine-transformed values
4.2.6.5 Effect of different sugar concentration on regeneration
frequency
Sucrose concentrations were varied from 1 to 5% to study its effect on shoot bud
induction in leaf explants. The leaf explants showed significant differences in
response to the five different sucrose concentrations tested. At 1% sucrose
concentration the leaf explants turned brown, whereas at 4% and 5%
concentration, though the explants remained green and an outgrowth of white,
root-like structures was observed, no shoot bud induction was recorded. On the
other hand, at 3% sucrose concentration, 78% explants responded to shoot
regeneration with an average of 7.4 shoot buds per explant. On medium
supplemented with 2% sucrose, only 47% explants responded producing an
average of 5.5 shoots per explant. Thus, a significant difference was observed on
medium supplemented with 2 and 3% sucrose with respect to the percentage of
explants responding to regeneration. However, no significant differences were
observed in the number of shoot buds induced per explant at these two sucrose
concentrations (Table 4.17).
Table 4.17 Effect of sugar concentration on regeneration response
Sucrose concentration
(%)
Response (%) Shoot buds/explant
1 0c (0)
# 0
b
2 47b (43.3) 5.5
a
3 78a (62) 7.4
a
4 0c (0) 0
b
5 0c (0) 0
b
LSD 12.52 2.3
# Values in the bracket ( ) represent the arcsine-transformed values
Eventually it was observed that MS 1/2 supplemented with 3% sucrose +
0.5 mgl-1 2,4,5-T + 2.0 mgl-1 BAP and solidified with 0.8% agar produced

Micropropagation
TERI University - Ph.D. Thesis, 2006
the most optimal results for shoot bud induction. On transfer of the explants to
the MS ½ medium an increase in number of shoot buds was observed. The
shoots developed roots on the MS ½ basal medium itself and were successfully
hardened. The clonal fidelity analysis (detailed in Chapter 5) of the regenerants
proved that no variations were developed during the regeneration process.

Micropropagation
TERI University - Ph.D. Thesis, 2006
4.3 Discussions
In spite of the immense demand for Swertia chirayita and the reported
scarcity in its supply, very few reports dealing with large-scale propagation of S.
chirayita or even its allied species are available in the literature. This situation
has remained practically unchanged since the seventies when in vitro culture of
S. japonica, another species of the genus Swertia (Miura et al. 1978 a, b) was
reported for the first time. Ahuja et al. (2003) patented micropropagation
protocol of S. chirayita, reporting a shoot multiplication rate ranging between
11.4-26.2 using single nodal explants and a 50-80% rooting at the end of 8 wk
inoculation period. In the present study an attempt has been made to develop a
micropropagation protocol for S. chirayita from seedling derived nodal
explants.
4.3.1. Seed germination
In absence of mature explants, S. chirayita seedlings were used for culture
initiation. Very few studies on germination and viability of S. chirayita seeds
have been reported. Raina et al. (1994) reported 91% germination in S. chirayita
seeds after a chilling treatment at –3 0C in comparison to 3.4% germination of
control seeds. In a more recent study, Basnet (2001) reported a maximum of
81% and a minimum of 76% germination in S. chirayita in the nursery
conditions. The high variations in germination percentage reported in these
different studies can be best explained by the variability in the genotypes of the
germplasm. Besides, factors such as storage conditions (ambient temperature
and humidity) and maturity of seeds also influence the germination proportion
(Colbach and Durr 2003, Wei et al. 2003). In the present study, seeds were pre-
treated with 400 ppm GA3 that resulted in 80% germination. Seeds without GA3
pre-treatment did not germinate at all. Wawrosch et al. (1999) also found the
treatment of seeds with GA3 essential for germination. The seeds lost their
viability and failed to germinate in the subsequent germination experiments
conducted after two months.
4.3.2 Culture establishment
4.3.2.1 Effect of different cytokinins on initial shoot bud induction
The role of cytokinins in axillary bud induction has been repeatedly illustrated in
the literature. Preliminary experiments were conducted to test the effect of
equimolar concentration (3 M) of BAP, Kn and 2iP with MS basal as control, on
shoot bud induction in nodal explants. The nodal explants inoculated on MS
basal and those inoculated on MS medium supplemented with 3 µM BAP, Kn

Micropropagation
TERI University - Ph.D. Thesis, 2006
and 2iP separately, showed a significant variation in terms of number of shoot
buds induced per explant. A maximum number of 5.9 shoot buds were induced
on 2iP supplemented MS medium, followed by 5.7 shoot buds / explant on MS
medium supplemented with BAP.
Shoot bud induction on basal medium remained almost negligible with only
1.6 shoot buds being formed per explant. The nodal explant formed shoot
clusters by the end of the third subculture on cytokinin containing medium. Our
results thus conclusively showed that cytokinins were essential for shoot bud
induction and multiplication.
The requirement of exogenous cytokinins for shoot bud induction during
culture establishment has also been illustrated in many other studies on
medicinal plants. For instance, shoot burst during culture establishment was
cytokinin dependent in Campotheca acuminata (Jain and Nessler 1996) as was
also the case in Nothapodites foetida (Satheeskumar and Seeni 2000). In fact,
the type of cytokinin also has a marked effect on shoot regeneration from nodal
explants. BAP was also found most effective for shoot bud induction in
Chlorophytum borivilianum (Purohit et al. 1994). In Gymnema elegans, BAP
showed better response over Kn in shoot bud induction (Komalavalli and Rao
1997). In Vitex negundo too, Sahoo and Chand (1998) reported BAP as most
effective for shoot bud induction and multiple shoot formation. In our study,
both 2iP and BAP were found to be equally conducive for initial culture
establishment and shoot bud induction.
However, in the present study, effect of cytokinins on shoot length did not
vary much and remained more or less similar to that observed on basal medium.
Hence, during the initial stages of culture establishment stunted shoot growth
was observed. Over a period of time the shoot length attained an average length
of 2 to 2.5 cm in four weeks time, as is evident in the experiments discussed
next. This can be attributed to the fact that explants get acclimatised to in vitro
conditions and hence respond more favourably. We can also attribute the lack of
elongation in in vitro cultures partially to the fact that S. chirayita is a slow
growing plant. In its natural conditions, the plant remains in a rosette form
during the vegetative phase of growth and elongates to 1-2 m length only prior to
flowering.
4.3.2.2 Effect of different sugar concentration on multiplication rate
and elongation
Sucrose, a disaccharide also synthesized and transported naturally by the plant,
is most commonly used as the carbon source in in vitro cultures. Sucrose

Micropropagation
TERI University - Ph.D. Thesis, 2006
present in the culture medium is rapidly broken down to fructose and glucose by
the extracellular enzymes. Since the culture conditions are deficient in light
energy and CO2 concentration, sucrose is indispensable for in vitro growth and
development (Tombolata and Costa, 1998). Sucrose concentration of 20 gl-1 and
30 gl-1 are most commonly used for plant tissue culture studies (Arditti 1974).
In the present study, the treatments containing different sucrose
concentration produced significant differences in plant height and
multiplication rate after the stipulated culture period of four weeks. Medium
supplemented with 3% sucrose proved most optimal for shoot bud induction as
well as shoot elongation and an average of 4.27 shoots with 2.43 cm length were
produced per explant.
4.3.2.3 Effect of different gelling agents on plant growth
Most cultures require a gelling agent to act as a support for the plant tissue.
Gelling agents influence the gel strength, water potential and contribute to
regulation of humidity, which in turn affects the availability of water (Debergh
1983) because the water content in semisolid medium exists in bound form.
Further, determining the better gelling agent for a particular species is
worthwhile as the effect of gelling agent is mostly considered to be genotype
dependent (Makunga et al. 2005).
Agar is the most commonly used gelling agent and has been used in many
protocols defining micropropagation of medicinal plants (Joshi and Dhar 2003,
Chaturvedi et al. 2004, Makunga et al. 2005). In comparison to agar, which is
derived from seaweed (red algae Gelidium amansii, Araki and Arai 1967),
gelrite, derived from bacterium Pseudomonas elodea, is a natural anionic
heteropolysaccharide, consisting of glucuronic rhamnose, glucose and O-acetil
molecules that form rigid, brittle agar like gel in the presence of soluble salts
(Kang et al. 1982). Agargel on the other hand is a blend of agar and gelrite and
was developed to help control vitrification in plant tissue cultures. It is superior
to gelrite where vitrification is a problem and is as an economical alternative to
agar.
In our study, response of explants in medium gelled with agar was found
superior as compared to medium gelled with agar gel and gelrite. However, it
was on gelrite medium that explants showed increase in shoot length. A few
scattered cases of hyperhydricity were also observed on gelrite medium. In fact,
gelrite is known to encourage hyperhydricity in some plant species (Debergh
1983, Thomas et al. 2000, Makunga et al. 2005). Hyperhydricity was also
evident in S. chirayita cultures as studied by Wawrosch et al. (1999), however,

Micropropagation
TERI University - Ph.D. Thesis, 2006
they attributed this behaviour to the influence of cytokinins. Eventually, we
opted for agar as the gelling agent, which gave a higher multiplication fold (5.9
shoot buds/explant).
4.3.3 Culture multiplication
The organogenic potential of an explant under in vitro conditions is closely
associated with content of natural and exogenously applied plant growth
regulators (PGRs). Hence, it becomes mandatory to work out the type and
concentration of different PGRs for deriving optimal multiplication results.
4.3.3.1 Effect of different cytokinins on culture multiplication
The regulatory action of cytokinin and apical dominance in in vitro shoot
induction and multiplication has been well documented (Wickson and Thiman
1958). Among the three cytokinins that were tested at varying concentration,
BAP at 4 M was most effective in promoting axillary shoot proliferation and
culture multiplication. In the presence of 4 M BAP the multiplication rate
increased to 3.6 (in terms of shoot clusters) as compared to 1.3 fold
multiplication observed on MS basal medium. Thus, the superiority of BAP over
other cytokinins was established in this experiment. Benzyladenine has often
been found to be more effective at stimulating axillary shoot development in
many medicinal plants such as Cholorophytum borivilianum (Dave et al. 2003),
Rotula aquatica (Martin 2003), S. chirata (Ahuja et al. 2003) and Orthosiphon
stamineus (Leng and Lai-Keng 2004).
It was observed that the optimal concentration of each of the cytokinin was
considerably low and an increase in concentration beyond a certain threshold
resulted in decrease in the multiplication rate for each of the cytokinin tested.
The reduction in number of shoots generated from each explant at cytokinin
concentration higher than the optimal level was also reported in Withania
somnifera (Sen and Sharma 1991), Kaempferia galanga (Vincent et al. 1992)
and Vitex negundo (Sahoo and Chand 1998). Decrease in shoot multiplication at
higher concentration has been variously attributed to callus induction and
adventitious bud formation (Zimmerman et al. 1980), inhibition of shoot
initiation (Sen and Sharma 1991) and hyperhydricity (Wawrosch et al. 1999).
An associated advantage of low concentration of cytokinin in culture medium
is that the occurrence of adventitious bud formation is reduced and this
eliminates the consequent risk of somaclonal variations (George 1996) thus
increasing probability of propagation of clonally uniform plants as was achieved
in our study (detailed in Chapter 5).

Micropropagation
TERI University - Ph.D. Thesis, 2006
4.3.3.2 Synergistic effect of BAP along with 2iP and Kn on cluster
multiplication and elongation
Synergistic effect of two or more cytokinins on promotion of shoot
multiplication is well documented for medicinal plants. Various synergistic
combinations have been reported such as BAP + Kn for Kaempferia galanga
(Vincent et al. 1992) and Feronia limonia (Hossain et al. 1994). Synergistic use
of BAP and Kn resulted in improvement in shoot elongation as well as
multiplication in Chlorophytum borivilianum (Purohit et al. 1994). Similarly, a
significant synergistic effect of cytokinins is well documented in a study on Piper
longum in which a combination of BAP and Kn produced significant increase in
culture multiplication (Philip et al. 2000). In the present study also it was
observed that BAP along with 1.5 M 2iP produced the highest multiplication
fold of 4.5 in S. chirayita explants followed by 3.9 multiplication fold obtained
on MS medium supplemented with 4 M BAP and 0.5 M 2iP.
4.3.3.3 Synergistic effect of 4 M BAP along with auxins on growth
of S. chirayita
Cytokinins, along with auxins also play important role in plant growth and shoot
regeneration. Sachs and Thimann (1964) illustrated that morphogenesis is
controlled by the interplay of auxins and cytokinins. Synergistic effect of
cytokinins with auxins has been reported to produce superior results for shoot
proliferation in many medicinal plants (Sudha and Seeni 1996, George and
Ravishankar 1997, Khan et al. 1997). Various successful combinations have been
reported, such as, BA + IAA for Thapsia garganica (Makunga et al. 2005); BAP
+IBA for Hagenia abyssinicia (Feyissa et al. 2005) and BA + NAA for
Phellodendron amurense (Azad et al. 2005).
Contrary to the above studies, no significant synergistic effect of any of the
tested auxins (namely IAA, NAA or IBA) along with 4 M BAP was established
in S. chirayita with respect to cluster multiplication. The multiplication rate
observed for all the treatments, was much lower than that recorded for the
medium with 4 M BAP alone. In fact, the multiplication fold reduced to values
equivalent to those observed on basal medium for the two concentrations of
NAA (0.5 and 1.0 M) tested. Cluster height was stunted and shoot clusters
obtained on these two medium combinations were the smallest among all the
medium combinations tested. Ahuja et al. (2003) on the other hand reported the
use of 1 mgl-1 IAA in their patented media composition for faster propagation of
S. chirayita. This variation in S. chirayita response might be due to the

Micropropagation
TERI University - Ph.D. Thesis, 2006
difference in the genotypes used in the two studies. Also, differnce in the
physiological state of the explants (mature and juvenile) used in the two studies
also explains the variations in their response.
Similar inhibitory influence of NAA on shoot multiplication was also
reported for Aegle marmelos (Varghese et al. 1993, Ajithkumar and Seeni 1998).
Inhibition of multiple shoot induction on auxin supplemented multiplication
medium was also reported for Gymnema elegans (Komalavalli and Rao 1997),
and is usually attributed to callus induction and in many cases to incidences of
rooting as well.
4.3.3.4. Effect of casein hydrolysate and adenine on cluster
multiplication and elongation
Additives such as casein hydrolysate (CH) and adenine sulphate provide an
additional source of nitrogen and have been frequently used as supplements for
enhancing multiplication and elongation in micropropagation protocols. Casein
hydrolysate contains a mixture of essential amino acids and is an excellent
source of reduced nitrogen for many plant tissue culture systems. However in
our study, neither the use of adenine nor that of CH produced any increase in
multiplication of S. chirayita. Similar results were also evident in Bixa orellana
(Sharon and D’Souza 2000), where addition of casein hydrolysate did not result
in increased number of shoots. In Peganum harmala (Saini and Jaiwal 2000)
addition of adenine sulphate increased the efficiency of regeneration; however,
there was a definite reduction in the number of shoots regenerated per explant.
Thus, contrary to the expected result of an increase in multiplication, use of
additives resulted in decline of cluster growth in S. chirayita.
4.3.4 Rooting
4.3.4.1 Effect of different auxins and activated charcoal on root
induction
Among the different auxins tested for root induction, most optimal response was
recorded for NAA at 1 M concentration. In an earlier study on S. chirata,
Wawrosch et al. (1999) used considerably higher concentration of auxins (10
M) for rooting and this resulted in induction of short, tuberous roots
accompanied with prolific callusing. They subsequently altered the rooting
procedure and rooting was obtained on basal medium after a short pulse
treatment of 2 seconds in 15 ppm NAA. Ahuja et al (2003) on the other hand
reported 60-65% rooting in time duration of 8 weeks, on MS media
supplemented with IAA ranging between 1-5 mgl-1.

Micropropagation
TERI University - Ph.D. Thesis, 2006
In the present study, 1 M auxin (IAA, NAA or IBA) concentration was found
sufficient to induce rooting. However, thick swollen tuberous growth of roots
was evident. This was circumvented by addition of activated charcoal to the
rooting medium.
Addition of activated charcoal is known to result in adsorption of various
organic compounds, such as excess hormones, vitamins, abscisic acid, phenolic
metabolites and ethylene (Van Winkle et al. 2003) leading to the improved
growth of the cultures. The carry over effect of PGRs used during the
multiplication cycles can also lead to a difficulty in root induction and can affect
the quality of roots as well. In the present study, thick and tuberous roots were
induced in rooting medium without activated charcoal. However, addition of
activated charcoal in the rooting medium, improved the quality of roots and
thin, long roots developed directly from the shoot base without an intervening
callus. This can most likely be attributed to activated charcoal, which is known to
eliminate the residual effects of cytokinins applied during the multiplication
stage (Maene and Debergh 1985). Effect of activated charcoal on root induction
has been previously reported in different plant systems such as, Decalepis
hamiltonii (Reddy et al. 2001), Rollina mucosa (Figueiredo et al. 2001) and
Curcuma zedory (Loc et al. 2005).
4.3.4.2 Effect of different medium strengths on rooting
The influence of mineral concentration of culture medium on rooting can be
attributed to the participation of inorganic ions in processes regulating
hormonal balance (Amzallag et al. 1992). The salt strength concentration has
also been known to affect auxin stability thus altering the auxin’s concentration
(Dunlap et al. 1986). Further, the favourable effect of diluted mineral solution on
rooting can be explained by the reduction of nitrogen concentration (Driver and
Suttle 1987). In fact, in some plant species, dilution of salt concentration
prepares the plants for better adaptation to the acclimatization regime.
Therefore, it becomes mandatory to study the effect of varying salt strengths on
root induction besides optimising auxin concentration for root induction.
In many plant species shoots are transferred from high strength medium to
less concentrated solutions as a routine practice. The practice has been used for
most of the plant species irrespective of their being herbaceous, for instance
Saussurea obvallata (Joshi and Dhar 2003); or woody medicinal plants such as
Vitex negundo (Sahoo and Chand 1998) and Azadirachta indica (Chaturvedi et
al. 2004). In our study, the number of roots remained unaffected by diluting the
salt concentration to as low as one-tenth. However, a definite variation was seen

Micropropagation
TERI University - Ph.D. Thesis, 2006
in the length of the roots induced on medium with different MS salt strengths. It
increased as salt strength was reduced from MS full to MS half and one fourth;
albeit, further reduction in salt strengths resulted in decrease of root length.
Root induction response remained more or less similar on MS half and MS one-
fourth medium. The salt strength also affected the quality of shoots and
therefore, while determining the optimal dilutions, it is important to consider
quality of roots as well as shoots, as these have significant effect on
acclimatisation of the plant.
4.3.4.3 Effect of sugar concentration on root induction
Besides serving as a carbohydrate source, sucrose regulates the osmolarity of the
culture medium and also plays a definite role during morphogenesis (Sopory
1979). Results of the experiment on different sucrose concentration show that S.
chirayita shoots rooted well on medium containing sucrose concentration
ranging from 10 to 30 gl-1. However, at 2% concentration, optimal number of
roots (6.9) were induced without compromising the plant growth.
While considering cost economics, the presence of sugar in the culture
medium is one of the factors that contribute to a significant increase in
production cost of micropropagated plantlets (Kozai 1991). Also, according to
Debergh (1991), the absence / reduction of sugar in the culture medium reduces
contamination problems. Further, a decrease in sugar concentration in the
subsequent stages of micropropagation is usually favoured as it leads to increase
in hardening success during acclimatisation of the plants (Muller 2004).
Therefore, the decrease in sucrose concentration during the rooting stage augurs
well for commercial production of plants through tissue culture.
4.3.5 Acclimatisation and field transplantations
The success of a tissue culture protocol largely depends on ex vitro survival and
growth performance of micropropagated plants. In the present experimental
study, a hardening success of 94.5% (average of hardening achieved from
December to April, the most conducive months of the year for hardening of
chiretta at MTP at Gual Pahari, Haryana) and a transplantation success of 65.3%
were recorded.
During the warmer months of May and August, when the initial hardening
trials were conducted, successful hardening was feasible only in growth room
under controlled conditions of temperature (22 2 0C) and humidity ranging
between 65-70%. Similar details have been reported for many other Himalayan
plants. Wawrosch et al. (1999) obtained hardening survival of 90-100% for

Micropropagation
TERI University - Ph.D. Thesis, 2006
Swertia chirata. Low temperatures (17-20 0C) were maintained stringently and
an increase in temperature even to 25 0C resulted in drastic mortality in their
study. In Picrorhiza kurroa, another medicinal plant of temperate Himalayas, a
high rate of plantlet survival (87.7%) was possible under the growth chamber
conditions of low temperature (15-20 0C) and constant relative humidity (60-
70%, Lal et al. 1988).
The potting mixes in which the plants are transferred also affect the survival
percentage of the plant. In Vitex negundo, of the four different potting mixes
examined, percentage survival of the plants was highest (93%) in vermin-
compost (Sahoo and Chand 1998). Similarly, in Melia azedarach, the plant
survival and growth were better in vermiculite, and it was reasoned that
lightweight and porous nature of the soil substitute prevents root damage and
thus allows for better root growth and hence healthier plants (Thakur et al.
1998). Contrary to these reports, survival percentage in S. chirayita plants
remained more or less similar (97-99%) for the different planting substrates that
were examined. Similarly, not much variation was noticed in the growth of the
plants on different planting substrates. The statistical variation in the data
recorded was not conspicuous when the plants were finally transferred to the
field.
The plants were eventually transferred to the field and 65.3% survival of the
micropropagated plants was recorded after the initial three months of growth.
The lower survival rate during field transplantations can be well explained by the
sudden change of climatic regime to which the plant is exposed during the
transportation of the plants from tropical conditions of Delhi to the temperate
environment of Himalayas. Despite the shock that the plants were exposed to, a
considerably high survival percentage (65.3%) was recorded and the plants
successfully completed their life cycles. In most in vitro studies conducted for
temperate plants, a low transplantation success has been recorded. For instance,
in Saussurea obvallata the highest percent survival recorded was 66.7 after the
initial 15 days of transfer to the field and this further reduced to 55.6% after the
first 30 days (Joshi and Dhar 2003). Wawrosch et al (1999) recorded 60% field
survival of micropropagated Swertia chirata after one year.
The post-transplantation growth of the micropropagated plants based on
visual observation was uniform. The uniformity of the plants raised via tissue
culture was also ascertained by ISSR assay as detailed in the next chapter. The
tissue culture protocol detailed above has been successfully utilised for large-
scale propagation of the endangered species and more than 100,000 plants have

Micropropagation
TERI University - Ph.D. Thesis, 2006
been produced and successfully transferred to the field at Mandal and
Mukteshwar.
4.3.6 Regeneration in Swertia chirayita from leaf explants
4.3.6.1 Differential regeneration response of leaf, root and shoot
explants
Morphogenetic potential of different explants has been regularly explored to
standardise regeneration protocols for different species. It is generally observed
that regeneration response of explants is genotype dependent and the number of
shoot buds regenerated also varies according to the species and type of explant.
For instance, Bhat et al. (1995) evaluated the morphogenetic potential of root,
leaf, node and internode explants of species belonging to genus Piperaceae.
Piper longum leaf explants formed shoot buds whereas those of P. belte and P.
nigrum did not. In the present study, most optimal regeneration response for S.
chirayita was obtained from leaf explants. Similarly, the leaves of Pothomorphe
umbellata showed greater organogenic potential for shoot induction (Pereira et
al. 2000). Soniya and Das (2002) also reported regeneration from leaf explant of
Piper longum using picloram instead of the regular auxins that are used for
regeneration. The use of leaf explants for shoot bud induction has also been
reported for medicinal plants, such as, Piper longum (Soniya and Das 2002),
Aconitum balfourii (Pandey et al. 2004), Phellodendron amurense (Azad et al.
2004) and Hypericum perforatum (Ayan et al. 2005).
In contrast to our results, Wawrosch et al. (1999) reported adventitious shoot
regeneration from root explants. Since the germplasm of S. chirayita for the two
studies was procured from different places, the variation in genotypes can likely
explain the difference in morphogenetic response. The root explants in our study
also induced shoot buds; however, this response was sporadic and non-
reproducible. The differential response of internodal segments and root and leaf
explants of S. chirayita on similar media might be related to the variations in
tissue and cell differentiation, as well as the endogenous amounts of
phytohormones in the explant.
During optimisation of regeneration protocol for most species, such as,
Panax notoginseng (Shoyama et al. 1997), Withania somnifera (Manickam et al.
2000) and Aconitum balfouri (Pandey et al. 2004) to name a few, auxins, such
as, IAA, IBA, NAA and 2,4-D are routinely used. In our study 2,4,5-T (0.5 mgl-1
concentration), a less frequently used auxin, produced the most optimal results
in terms of callus and shoot bud induction. However, at higher concentration,

Micropropagation
TERI University - Ph.D. Thesis, 2006
2,4,5-T resulted in browning of explants. In corroboration with our study, 2,4,5-
T has been successfully employed for organogenesis studies in Eleusine
coracana (Kumar et al. 2001), Sorghum bicolor (Nirwan and Kothari 2004) and
Cicer arietinum (Kiran et al. 2005).
4.3.6.2 Effect of varying concentration of BAP on callus induction
and shoot regeneration from leaf explants
The effect of varying concentration of BAP
Among the different cytokinins regularly used for inducing regeneration, BAP
was employed in the present study. In the preliminary experiments MS ½ + 0.5
mgl-1 BAP + 0.5 mgl-1 2,4,5-T was found to induce regeneration response from
leaf explants. BAP concentration was tested in the range of 0.25 mgl-1 to 4 mgl-1
and it was revealed that 2 mgl-1 of BAP with 0.5 mgl-1 of 2,4,5-T induced
regeneration in maximum number of explants (76%). However, browning of
explants was noticed as BAP concentration was increased beyond 2 mgl-1,
resulting in a decline in regeneration response.
Wawrosch et al. (1999) also observed that BAP at 5 M concentration was
most optimal for regeneration in S. chirata. In their study, the use of 2iP
resulted in 40% hyperhydrated shoots. The authors, therefore, opted for two-
step regeneration protocol, which consisted of, an initial 3-week cultivation on
modified MS medium supplemented with 3 M BAP followed by another period
of three weeks on hormone free medium. This two-step protocol helped in
reducing hyperhydricity as well as led to elongation of shoots. In our study, the
transfer of explants from regeneration medium on to MS PGR-free medium
displayed increased proliferation in number of shoot buds as well as shoot
length.
The effect of additives on callus growth and shoot bud induction
Addition of additives, such as, casein hydrolysate, adenine and glutamine is
usually effective in increasing the regeneration response. In the present study,
the addition of adenine or glutamine to the regeneration medium led to
browning of callus. On the other hand, the addition of CH was not inhibitory as
was the case with adenine and glutamine. However, response of explants on
medium with CH did not supersede that of explants inoculated on regeneration
medium, which lacked CH. Similar results were also evident for axillary
multiplication of S. chirayita from nodal explants as discussed in section
4.3.3.4.

Micropropagation
TERI University - Ph.D. Thesis, 2006
4.3.6.3 Effect of different gelling agents on explant growth and
callus induction
Gelling agents are considered to be inert components of a culture medium, with
a limited role of providing support to the growing cultures. However, these
gelling substrates have different origin and therefore differences exist in their
quantitative and qualitative organic fractions. In fact, a definite variation has
been observed in response of explants of different species with respect to explant
growth and callus induction on different gelling agent. For instance, in Siberian
elm, proliferation in gelrite medium was greater than that on agar-solidified
medium (Cheng and Shi 1995), whereas Corchete et al. (1993) had reported
success of Siberian elm regeneration on agar medium. The variations reported
can be explained by the sensitivity of the explants of different species and
genotypes to varying organic fractions of the gelling agent. In our study, gelrite
and agar gel completely inhibited the regeneration response from leaf explants,
whereas medium solidified with agar promoted regeneration. Wawrosch et al.
(1999) also used agar as gelling agent for shoot regeneration from root explants
in S. chirata.
4.3.6.4 Effect of MS salt strength on the regeneration response of
leaf explants
Basal medium are known to influence growth response of various plants in
different ways. This is considered to be a consequence of differences in their salt
components and proportions (Bhojwani and Razdan 1996). Since MS salt
composition is considered very rich in terms of the total nitrogen available (60
M of nitrogen ions), usually the MS composition is modified with reduction in
major salt to ½ or ¼ of the original strength for inducing morphogenetic
responses, such as, somatic embryogenesis and shoot and root induction. In our
study, the best regeneration response of leaf explants with 78% explant
responding to shoot bud induction, was obtained on modified MS salt with
major and iron reduced to half the original strength. Further reduction in the
MS salt composition, however, resulted in a decrease in regeneration response.
The lowest regeneration response (24.7%) was obtained on MS full medium,
thus corroborating the fact that lower MS salt concentration favoured
regeneration in S. chirayita.
4.3.6.5 Effect of different sugar concentration on regeneration
frequency

Micropropagation
TERI University - Ph.D. Thesis, 2006
Sucrose is the main carbohydrate source for in vitro culture and is necessary to
sustain growth and also serves as an osmoticum. Sucrose concentration has also
been known to regulate shoot bud induction in different plant species.
For instance, on increasing sugar concentration from 1 to 3%, an increase in
regeneration percent was observed in quince (Baker and Bhatia 1993). Chevreau
et al. (1989) found positive linear effect on shoot regeneration in pear as sucrose
concentration was increased from 1 to 5%. In case of S. chirayita, in the present
study, 3% sucrose concentration proved most optimal for shoot bud induction
inducing a 78% regeneration response from the leaf explants. However, further
increase in sucrose concentration to 4% and 5% led to browning of the leaf
explants.

Figure 4.1 a) Nodal explant of Swertia chirayita derived from 4-week-old in vitro germinated seedlings and (b-d ) nodal explant developing into clusters after the initial establishment stages on MS medium supplemented with 3 M BAP and 3% sucrose for a culture period of 4 weeks.
a
c
b
d

1.7
5.84.9
5.9
1.26 1.281.25
1.44
0.0
1.0
2.0
3.0
4.0
5.0
6.0
7.0
8.0
basal 2iP Kn BAP
cytokinins
shoot buds
0.00
0.50
1.00
1.50
2.00
shoot heig
ht (cm
)
sh oot bu ds shoot length
Figure 4.2 The effect of BAP, Kn and 2iP at equimolar concentration of 3 M on shoot bud induction and shoot elongation in Swertia chirayita after a culture period of 4 week each. A significant variation was visible in explants cultured on MS basal medium as compared to medium supplemented with 3
M each of the three cytokinins, namely 2iP, Kn and BAP with respect to shoot buds/explant.

Figure 4.3 The growth of S. chirayita shoot-clusters as obtained on MS medium supplemented with 3 M BAP + 3% sucrose and solidified with (a) agar (b) agarose and (c) gelrite. The difference in shoot length is visible in the three medium with gelrite showing the maximum elongation after 4 wk of culture period.
a b
c

0
0.5
1
1.5
2
2.5
3
3.5
4
0.5 1 4 6 8 10
con cen tr a t ion (m icr om ola r s)
mu
ltip
lica
tio
n fo
ld
0
0.5
1
1.5
2
2.5
3
3.5
sh
oo
t len
gth
(cm
)
BA P sh oot bu ds kin et in sh oot bu d 2 iP sh oot bu ds
BA P sh oot len g th Kin et in sh oot len g th 2 iP sh oot len g th
Figure 4.4 Pictorial representation of the variations in cluster multiplication and cluster height in S. chirayita as obtained for the varying concentrations of BAP, Kinetin and 2iP after 4 wk of culture period. Among the eighteen different combinations tested, 4
M BAP produced the most desirable results, both in terms of multiplication fold (3.6) and cluster elongation (2.6 cm).

Figure 4.5 Effect of different auxins on root induction in S. chirayita shoots when inoculated on (a) basal medium (MS half +3% sucrose) and that supplemented with (b) 1 M IAA (c) 1 M NAA and (d) 1 M IBA as observed after 4 wk of culture period.
a
b
d c

Fig4.6 (a) Roots induced on rooting medium (MS ½ + 3% sucrose + 1 M NAA) without activated charcoal remain thick and small along with basal callusing as compared to (b) thin slender roots induced on rooting medium supplemented with 500 mgl-1 of activated charcoal.
b
a

Figure 4.7 In vitro hardening of micropropagated S. chirayita plantlets in growth room. (a) Plantlets transferred to culture vessels containing coir peat with polypropylene caps which were gradually opened over a period of three weeks for successful hardening and (b) successfully hardened plantlets 3 weeks after hardening.
b
a

Figure 4.8 Ex vitro hardening of Swertia chirayita plantlets representing (a) ex-agar plants transferred to poly bags kept in green house for acclimatization for a period of two weeks and (b) 4 week-old hardened plants shifted to shade net after acclimatization.
a
b

Figure 4.9 Swertia chirayita plantlets from rooting stage to a well acclimatized plant. (a) micropropagated shoots in rooting medium (MS ½ + 2% sucrose + 1
M NAA) b) an ex-agar, well rooted plantlet before hardening c) few plantlets after hardening in green house d) a bare root micropropagated chiretta plant ready for transplantation.
a b
c d

Figure 4.10 (a) 2-month old hardened plants immediately after transplantation in field at Mandal, Dist. Chamoli, Uttaranchal, India (b) Micropropagated plants after six months of growth in the same field. An average height of 15 cm, growth area of 240 square inch and biomass of 106 gm was recorded for the plants six months after transplantation to the field.
a
b

Figure 4.11 In vitro multiplied plants of Swertia chirayita after ( a) 3-months, (b) 6-months and (c) one year of growth in field (Mandal, Uttaranchal). The plants show rosette growth with a maximum height averaging between 12-18 cm during the vegetative phase of growth (a and b). The plants bolt before flowering and attain an average height of 1-1.5 m ©.
a
c b

Figure 4.12 The chiretta plant during flowering stage after two years of vegetative growth. a) The shoot apex immediately before bearing flowers b) a well developed root after two years of growth c) A flowering twig d) flower of chiretta showing the four yellow-green petals, also visible are the two glands which are the identifying feature of Swertia chirayita.
a b
d c

Figure 4.13 Sporadic shoot regeneration from root explants (a) root explants after incubation period of six weeks on NAA + IBA (0.75 mgl-1 each) supplemented media (b) shoot elongation of regenerants on MS ½ basal medium after 4 weeks of culture period.
a
b

0
30
36
76
16.59
0
17
0.0 0.0
8.3
10.710.3
8.3
3.3
0
1 0
2 0
3 0
4 0
5 0
6 0
7 0
8 0
9 0
0 0.2 5 0.5 1 2 3 4
BAP (mg/l)
ex
pla
nt re
sp
on
se
(%
)
0.0
2.0
4.0
6.0
8.0
10.0
12.0
14.0
nu
mb
er o
f sh
oo
t b
ud
s p
er e
xp
la
nt
Figure 4.14 The effect of varying BAP concentrations (0-4 mgl-1 ) on the regeneration response of leaf explants and the number of shoot buds induced per explant. Maximum regeneration response of 76% was observed at BAP concentration of 2 mgl-1 with an average of 11 shoots buds/explant.

Figure 4.15 (a) Leaf explants on regeneration medium showing initial swelling and expansion due to water absorption and cell proliferation in the initial six weeks of culture followed by (b) profuse callus formation after another 6 weeks of culture period on regeneration medium (MS ½ supplemented with 2.0 mgl-1 BAP and 0.5 mg l-1 2,4,5-T with 3% sucrose).
a b

8.3
10.3 10.7
34.8
24.0
43.0
0.0
5.0
10.0
15.0
20.0
25.0
30.0
35.0
40.0
45.0
50.0
1 2 3
r eg en er a t ion m ediu m
nu
mb
er o
f s
ho
ot b
ud
s p
er e
xp
lan
t
r eg en er a t ion m edia ba sa l m edia
Figure 4.16 Comparative graphical illustration of shoot buds induced on the three best regeneration medium (among the different tested combinations) numbered as 1, 2 and 3 and the increase in their numbers after transfer to basal medium at the end of 4 week culture period. The regeneration medium numbered 1, 2 and 3 represent MS ½ supplemented with 0.5 mg/l 2,4,5-T each and BAP at 0.5, 1.0 and 2.0 mg/l, respectively.

4 3 .0
3 4 .8
2 4 .0
2 .9 5
3 .02
3 .1 5
0.0
5 .0
1 0.0
1 5 .0
2 0.0
2 5 .0
3 0.0
3 5 .0
4 0.0
4 5 .0
5 0.0
1 2 3
Or g a n og en esis in du ct ion m ediu m
nu
mb
er o
f s
ho
ot b
ud
s p
er e
xp
lan
t
2 .6 5
2 .7
2 .7 5
2 .8
2 .8 5
2 .9
2 .9 5
3
3 .05
3 .1
3 .1 5
3 .2
sh
oo
t l
en
gth
(cm
)
Sh oot bu ds sh oot len g th
Figure 4.17 The increase in number of shoots and elongation in shoot length after the transfer of explants from the regeneration medium, numbered as 1 (MS ½ + 0.5 mgl-1 2,4,5-T + 0.5 mgl-1 BAP ), 2 (MS ½ + 0.5 mgl-1 2,4,5-T + 1.0 mgl-1 BAP) and 3 (MS ½ + 0.5 mgl-1 2,4,5-T + 2 mgl-1 BAP) to MS1/2 basal medium after 4 weeks of culture period.

Figure 4.18 Shoot elongation and proliferation after transfer of explants from regeneration medium (MS ½ supplemented with 2.0 mgl-1 BAP and 0.5 mg l-1 2,4,5-T with 3% sucrose) to MS ½ basal medium after 4 weeks.

Figure 4.19 a) Callus growth and shoot buds induction on control medium (MS ½ medium supplemented with 2.0 mgl-1 BAP+ 0.5 mgl-1 2,4,5-T) (b) browning of callus browning in case of explants cultured on control medium supplemented with 500 mgl-1 CH (c) control + 125 mgl-1 adenine and (d) control + 125 mgl-1 glutamine.
a b
c d


















