
Microbiome Assembly across Multiple Body Sites in Low...
12
Microbiome Assembly across Multiple Body Sites in Low-Birthweight Infants Elizabeth K. Costello, a Erica M. Carlisle, b Elisabeth M. Bik, a Michael J. Morowitz, b * David A. Relman a,c,d Department of Microbiology & Immunology, Stanford University School of Medicine, Stanford, California, USA a ; Department of Surgery, University of Chicago Pritzker School of Medicine, Chicago, Illinois, USA b ; Department of Medicine, Stanford University School of Medicine, Stanford, California, USA c ; Veterans Affairs Palo Alto Health Care System, Palo Alto, California, USA d * Present address: Michael J. Morowitz, Department of Surgery, University of Pittsburgh School of Medicine, Pittsburgh, Pennsylvania, USA. ABSTRACT The purpose of this study was to evaluate the composition and richness of bacterial communities associated with low- birthweight (LBW) infants in relation to host body site, individual, and age. Bacterial 16S rRNA genes from saliva samples, skin swabs, and stool samples collected on postnatal days 8, 10, 12, 15, 18, and 21 from six LBW (five premature) infants were ampli- fied, pyrosequenced, and analyzed within a comparative framework that included analogous data from normal-birthweight (NBW) infants and healthy adults. We found that body site was the primary determinant of bacterial community composition in the LBW infants. However, site specificity depended on postnatal age: saliva and stool compositions diverged over time but were not significantly different until the babies were 15 days old. This divergence was primarily driven by progressive temporal turn- over in the distal gut, which proceeded at a rate similar to that of age-matched NBW infants. Neonatal skin was the most adult- like in microbiota composition, while saliva and stool remained the least so. Compositional variation among infants was marked and depended on body site and age. Only the smallest, most premature infant received antibiotics during the study period; this heralded a coexpansion of Pseudomonas aeruginosa and a novel Mycoplasma sp. in the oral cavity of this vaginally delivered, intubated patient. We conclude that concurrent molecular surveillance of multiple body sites in LBW neonates reveals a delayed compositional differentiation of the oral cavity and distal gut microbiota and, in the case of one infant, an abundant, unculti- vated oral Mycoplasma sp., recently detected in human vaginal samples. IMPORTANCE Complications of premature birth are the most common cause of neonatal mortality. Colonization by the indige- nous microbiota, which begins at delivery, may predispose some high-risk newborns to invasive infection or necrotizing entero- colitis (NEC), and protect others, yet neonatal microbiome dynamics are poorly understood. Here, we present the first cultivation-independent time series tracking microbiota assembly across multiple body sites in a synchronous cohort of hospi- talized low-birthweight (LBW) neonates. We take advantage of archived samples and publically available sequence data and compare our LBW infant findings to those from normal-birthweight (NBW) infants and healthy adults. Our results suggest po- tential windows of opportunity for the dispersal of microbes within and between hosts and support recent findings of substantial baseline spatiotemporal variation in microbiota composition among high-risk newborns. Received 13 September 2013 Accepted 25 September 2013 Published 29 October 2013 Citation Costello EK, Carlisle EM, Bik EM, Morowitz MJ, Relman DA. 2013. Microbiome assembly across multiple body sites in low-birthweight infants. mBio 4(6):e00782-13. doi: 10.1128/mBio.00782-13. Editor Martin Blaser, New York University Copyright © 2013 Costello et al. This is an open-access article distributed under the terms of the Creative Commons Attribution-Noncommercial-ShareAlike 3.0 Unported license, which permits unrestricted noncommercial use, distribution, and reproduction in any medium, provided the original author and source are credited. Address correspondence to Elizabeth K. Costello, [email protected]. T he composition of the human microbiota is body site specific in healthy adults (1–3), yet this is not the case in newborns shortly after delivery (4). While the postnatal assembly of an adult-like distal gut microbiota has been studied in healthy infants (5–7), relatively little is known about the development of the mi- crobiota at extraintestinal sites (8, 9) or about the compositional differentiation of the microbiota across multiple sites during the neonatal period. Knowledge of these spatiotemporal dynamics is particularly lacking for low-birthweight (LBW) infants, who are at high risk of invasive infection and other serious perinatal compli- cations, including necrotizing enterocolitis (NEC), a disease linked in part to microbial colonization (10, 11). LBW infants are often premature, and often receive antibiotics, experience delays in the initiation of enteral feedings, and/or require prolonged hos- pital stays—all of which can influence, and be influenced by, in- teractions with microbes. Certain complications, such as sepsis and NEC, are characterized by onset timing (12, 13); for example, the postnatal age at the onset of NEC is inversely correlated with the gestational age at delivery (14). These patterns underscore a need to understand better the temporal dynamics of microbiome development in high-risk neonates. Postnatal microbial colonization prompts the terminal matu- ration of host intestinal structures, mediates the development of the immune system, and induces resistance to invasion by would-be pathogens (15–17). Furthermore, early life colonization deficiencies have been associated with alterations in host metabo- RESEARCH ARTICLE November/December 2013 Volume 4 Issue 6 e00782-13 ® mbio.asm.org 1 on July 17, 2018 by guest http://mbio.asm.org/ Downloaded from
Transcript of Microbiome Assembly across Multiple Body Sites in Low...

Microbiome Assembly across Multiple Body Sites in Low-BirthweightInfants
Elizabeth K. Costello,a Erica M. Carlisle,b Elisabeth M. Bik,a Michael J. Morowitz,b* David A. Relmana,c,d
Department of Microbiology & Immunology, Stanford University School of Medicine, Stanford, California, USAa; Department of Surgery, University of Chicago PritzkerSchool of Medicine, Chicago, Illinois, USAb; Department of Medicine, Stanford University School of Medicine, Stanford, California, USAc; Veterans Affairs Palo Alto HealthCare System, Palo Alto, California, USAd
* Present address: Michael J. Morowitz, Department of Surgery, University of Pittsburgh School of Medicine, Pittsburgh, Pennsylvania, USA.
ABSTRACT The purpose of this study was to evaluate the composition and richness of bacterial communities associated with low-birthweight (LBW) infants in relation to host body site, individual, and age. Bacterial 16S rRNA genes from saliva samples, skinswabs, and stool samples collected on postnatal days 8, 10, 12, 15, 18, and 21 from six LBW (five premature) infants were ampli-fied, pyrosequenced, and analyzed within a comparative framework that included analogous data from normal-birthweight(NBW) infants and healthy adults. We found that body site was the primary determinant of bacterial community composition inthe LBW infants. However, site specificity depended on postnatal age: saliva and stool compositions diverged over time but werenot significantly different until the babies were 15 days old. This divergence was primarily driven by progressive temporal turn-over in the distal gut, which proceeded at a rate similar to that of age-matched NBW infants. Neonatal skin was the most adult-like in microbiota composition, while saliva and stool remained the least so. Compositional variation among infants was markedand depended on body site and age. Only the smallest, most premature infant received antibiotics during the study period; thisheralded a coexpansion of Pseudomonas aeruginosa and a novel Mycoplasma sp. in the oral cavity of this vaginally delivered,intubated patient. We conclude that concurrent molecular surveillance of multiple body sites in LBW neonates reveals a delayedcompositional differentiation of the oral cavity and distal gut microbiota and, in the case of one infant, an abundant, unculti-vated oral Mycoplasma sp., recently detected in human vaginal samples.
IMPORTANCE Complications of premature birth are the most common cause of neonatal mortality. Colonization by the indige-nous microbiota, which begins at delivery, may predispose some high-risk newborns to invasive infection or necrotizing entero-colitis (NEC), and protect others, yet neonatal microbiome dynamics are poorly understood. Here, we present the firstcultivation-independent time series tracking microbiota assembly across multiple body sites in a synchronous cohort of hospi-talized low-birthweight (LBW) neonates. We take advantage of archived samples and publically available sequence data andcompare our LBW infant findings to those from normal-birthweight (NBW) infants and healthy adults. Our results suggest po-tential windows of opportunity for the dispersal of microbes within and between hosts and support recent findings of substantialbaseline spatiotemporal variation in microbiota composition among high-risk newborns.
Received 13 September 2013 Accepted 25 September 2013 Published 29 October 2013
Citation Costello EK, Carlisle EM, Bik EM, Morowitz MJ, Relman DA. 2013. Microbiome assembly across multiple body sites in low-birthweight infants. mBio 4(6):e00782-13. doi:10.1128/mBio.00782-13.
Editor Martin Blaser, New York University
Copyright © 2013 Costello et al. This is an open-access article distributed under the terms of the Creative Commons Attribution-Noncommercial-ShareAlike 3.0 Unportedlicense, which permits unrestricted noncommercial use, distribution, and reproduction in any medium, provided the original author and source are credited.
Address correspondence to Elizabeth K. Costello, [email protected].
The composition of the human microbiota is body site specificin healthy adults (1–3), yet this is not the case in newborns
shortly after delivery (4). While the postnatal assembly of anadult-like distal gut microbiota has been studied in healthy infants(5–7), relatively little is known about the development of the mi-crobiota at extraintestinal sites (8, 9) or about the compositionaldifferentiation of the microbiota across multiple sites during theneonatal period. Knowledge of these spatiotemporal dynamics isparticularly lacking for low-birthweight (LBW) infants, who are athigh risk of invasive infection and other serious perinatal compli-cations, including necrotizing enterocolitis (NEC), a diseaselinked in part to microbial colonization (10, 11). LBW infants areoften premature, and often receive antibiotics, experience delays
in the initiation of enteral feedings, and/or require prolonged hos-pital stays—all of which can influence, and be influenced by, in-teractions with microbes. Certain complications, such as sepsisand NEC, are characterized by onset timing (12, 13); for example,the postnatal age at the onset of NEC is inversely correlated withthe gestational age at delivery (14). These patterns underscore aneed to understand better the temporal dynamics of microbiomedevelopment in high-risk neonates.
Postnatal microbial colonization prompts the terminal matu-ration of host intestinal structures, mediates the development ofthe immune system, and induces resistance to invasion bywould-be pathogens (15–17). Furthermore, early life colonizationdeficiencies have been associated with alterations in host metabo-
RESEARCH ARTICLE
November/December 2013 Volume 4 Issue 6 e00782-13 ® mbio.asm.org 1
on July 17, 2018 by guesthttp://m
bio.asm.org/
Dow
nloaded from

lism and immune function (18, 19). In the neonatal intensive careunit (NICU), however, the promotion of potentially beneficialhost-microbe interactions must be carefully balanced against thecontrol of pathogen spread among a highly vulnerable patientpopulation (20, 21). This is distinctively challenging with regardto the prevention and treatment of NEC, a disease in which theinterrelated roles of antibiotic exposure, enteral feedings, andchanges in the intestinal microbiota are imprecisely defined (10,11). Recent studies of the fecal microbiota of premature infantsusing cultivation-independent approaches have revealed a lowlevel of diversity, high interindividual variability, and a capacityfor abrupt temporal shifts in species- and strain-level composition(22–32). However, most of these studies have been limited to arelatively small number of samples and to a single body site, thedistal gut.
In the present study, we simultaneously tracked the distal gut,oral cavity, and skin surface microbiota of six hospitalized LBWinfants, including 2 sets of twins, over the 2nd and 3rd weeks oflife. Our analysis focused on factors underpinning compositionalvariation during this critical time span. For the distal gut micro-biota, we also made comparisons to age-matched normal-birthweight (NBW) infants using archived samples from a prior
study (5); and for all sites, we made comparisons to adults usingpublically available sequence data (1, 2). Although the infantssampled here were unaffected by sepsis or NEC, their age rangerepresents an important window of vulnerability for both of theseconditions.
RESULTSLBW infant cohort characteristics. Five of the six infants (all butbaby 6) were premature; these five had completed �32 weeks ofgestation at the time of delivery. Among the premature infants,three were born weighing �1.5 kg, placing them in the category of“very LBW” (VLBW) and at highest risk for complications of pre-term birth. These three infants were born at Comer Children’sHospital, whereas the others were born at outside hospitals andthen transferred to Comer’s NICU prior to enrollment. The co-hort included two sets of premature twins, both delivered via Ce-sarean section. All infants received antibiotics in the first week oflife (Table 1). None of their mothers received antepartum antibi-otics.
Baby 3, the smallest, most premature infant in the study, wasintubated and mechanically ventilated throughout the samplingperiod, whereas the others either had no history of endotracheal
TABLE 1 LBW infant characteristics and clinical information
Babya Sexb
Deliverymodec
Birthwt (kg)
Gestationalage atdelivery Birth locatione
Postdeliveryantibiotics andlength of treatmentf Medical conditionsg
Complication(s) duringpregnancy and/or deliveryh
1 F Cs 1.82 311
7d Not at a UC
hospitalAp � Gm 48 hi Premature,
respiratorydistress,hyperbilirubinemia
Loss of fetal heart tones
2 M Cs 1.74 311
7Not at a UC
hospitalAp � Gm 48 hi Premature,
respiratorydistress,hyperbilirubinemia
Loss of fetal heart tones
3 M V 0.75 244
7UC hospital Ap � Gm 7 days Premature,
respiratorydistress,hyperbilirubinemia
PPROM
4 M Cs 1.38 300
7UC hospital Ap � Gm 48 h Premature,
hyperbilirubinemia,GE reflux, AOP
Preeclampsia, pretermlabor
5 M Cs 1.05 300
7UC hospital Ap � Gm 48 h Premature,
respiratorydistress,hyperbilirubinemia,GE reflux, AOP
Preeclampsia, pretermlabor
6 M V 1.72 381
7Not at a UC
hospitalAp � Gm �Cx
48-72 hj
IUGR, chromosome4p deletionsyndromek
None noted
a Babies 1 and 2 are dizygotic (DZ) twins; babies 4 and 5 are monozygotic (MZ) twins (monochorionic diamniotic).b F, female; M, male.c Cs, Cesarean section; V, vaginal.d 311
7, 31 weeks and 1 day.
e UC, University of Chicago.f Ap, ampicillin; Cx, cefotaxime; Gm, gentamicin.g GE, gastroesophageal; AOP, anemia of prematurity; IUGR, intrauterine growth restriction.h PPROM, preterm premature rupture of membranes.i A third antibiotic may have been given (non-UC chart unclear/unavailable).j The most likely treatment duration (non-UC chart unclear/unavailable).k Including various syndrome-associated medical problems.
Costello et al.
2 ® mbio.asm.org November/December 2013 Volume 4 Issue 6 e00782-13
on July 17, 2018 by guesthttp://m
bio.asm.org/
Dow
nloaded from

intubation (babies 4 and 5) or had been extubated by the timesampling commenced (babies 1, 2, and 6). Baby 3 was also treatedwith antibiotics on days 13 to 19 for a suspected case of sepsis(Table 2), but all cultures (blood, urine, and cerebrospinal fluid[CSF] samples) were negative; no respiratory tract samples werecultured. Baby 3 was the only subject to receive antibiotics duringthe sampling period. Finally, in some cases, modifications to theinfants’ feeding regimens and/or hospital locations were madeduring the sampling period (Table 2). Most feedings were deliv-ered via nasogastric or orogastric tube.
Baby 3 received antibiotics for a suspected case of NEC aroundday of life (DOL) 40, but his clinical signs resolved quickly withoutfurther intervention. To our knowledge, none of the other infantswent on to have invasive infections or NEC after DOL 21.
Overview of bacterial taxonomic representation. Of the 108samples collected for the study, 106 yielded sufficient quantities of16S rRNA gene sequences to warrant subsequent analysis (range,
219 to 1,914 sequences/sample; median, 1,066 sequences/sample).Due to low sequencing yield, two samples were dropped.
Overall, nine bacterial phyla were represented (Fig. 1). On av-erage, the most abundant were the Firmicutes (71.6%), Proteobac-teria (21.4%), Bacteroidetes (5.4%), Tenericutes (1.0%), and Acti-nobacteria (0.5%). Rare phyla (those with average abundances of�0.01%) included the Cyanobacteria, Deinococcus-Thermus,Chloroflexi, and Fusobacteria. In total, 119 bacterial genera weredetected, the most abundant of which are displayed in Fig. 1.Dominant genera were as follows: from the phylum Firmicutes,Staphylococcus, Streptococcus, Enterococcus, and Gemella; from theclass Gammaproteobacteria, Klebsiella/Enterobacter (genera indis-tinguishable using the available gene fragment), Haemophilus,Citrobacter, Proteus, and Pseudomonas; and from the phylum Bac-teroidetes, the genus Bacteroides.
The identities of the abundant taxa found here are generallyconsistent with those observed in prior studies of LBW infants (22,
TABLE 2 LBW infant age-related events and informationa
Baby Feature
Postnatal age in daysb
8 10 12 15 18 21
1 Feeding BMtr BMfull BMfull BMtr, Ftr BMfull BMfull
Wt (kg) 1.72 1.79 1.725 1.8 1.91 1.974Antibiotic(s) None noted None noted None noted None noted None noted None notedLocation NICU NICU NICU step-down NICU step-down NICU step-down NICU step-down
2 Feeding BMtr BMtr BMtr BMtr, Ftr BMtr Ffull
Wt (kg) 1.71 1.805 1.785 1.86 1.86 1.96Antibiotic(s) None noted None noted None noted None noted None noted None notedLocation NICU NICU NICU step-down NICU step-down NICU step-down NICU step-down
3 Feeding BMtr BMtr, Ftr BMtr, Ftr Ftr Ftr Ffull
Wt (kg) 0.84 0.86 0.92 0.94 0.9 0.97Antibiotic(s) None noted None noted None noted Vm � Gm � Cxc Gm � Cxc None notedLocation NICU NICU NICU NICU NICU NICU
4 Feeding Ftr BMtr BMtr Ffull Ffull Ffull
Wt (kg) 1.3 1.35 1.53 1.62 1.6 1.67Antibiotic(s) None noted None noted None noted None noted None noted None notedLocation NICU NICU NICU NICU NICU NICU
5 Feeding Ftr BMtr BMtr Ffull Ffull Ffull
Wt (kg) 1.05 1.115 1.205 1.245 1.234 1.26Antibiotic(s) None noted None noted None noted None noted None noted None notedLocation NICU NICU NICU NICU NICU NICU
6 Feeding Ffull Ffull Ffull Ffull Ffull Ffull
Wt (kg) 1.725 1.65 1.675 1.735 1.77 1.765Antibiotic(s) None noted None noted None noted None noted None noted None notedLocation NICU NICU NICU Transitional floor Transitional floor Transitional floor
a Respiratory support, baby 3 was intubated from day of life (DOL) ~1 to 44. Babies 1, 2, and 6 were intubated DOL 1 and 2, 1 and 2, and 4 to 6, respectively. Babies 4 and 5 werenot intubated. Most babies received oxygen via nasal cannulae throughout the study. Feeding support, most feedings were delivered via a naso- or orogastric tube.b BM, pumped or stored maternal breast milk; F, formula; tr, trophic (i.e., minimal); Vm, vancomycin; Gm, gentamicin; Cx, cefotaxime.c Sepsis ruleout. Vm from DOL 13 to 15, Gm from DOL 13 to 19, and Cx from DOL 14 to 19.
Gut, Oral, and Skin Microbiome Dynamics in LBW Neonates
November/December 2013 Volume 4 Issue 6 e00782-13 ® mbio.asm.org 3
on July 17, 2018 by guesthttp://m
bio.asm.org/
Dow
nloaded from

24–26, 28, 33), including studies of premature infants recruitedfrom the same NICU as that which served as the setting for thecurrent study (23, 27, 29).
Microbiota composition is primarily shaped by body site.Patterns of bacterial community-wide compositional variation
were evaluated using the unweighted UniFrac metric. Pairs ofsamples containing similar (i.e., closely related) lineages have rel-atively small UniFrac distances, whereas those containing diver-gent (i.e., distantly related) lineages have relatively large ones (34).The unweighted UniFrac metric is incidence based (i.e., presence/
Proportion of sequences
Baby 1
Baby 2
Cs
Baby 3
VB
aby 4B
aby 5C
sB
aby 6V
a. Saliva b. Skin c. Stool
Age
in d
ays
8
10
12
15
1821
8
10
12
15
1821
8
10
12
15
1821
8
10
12
15
1821
8
10
12
15
1821
8
10
12
15
1821
All other taxa (0.86%)
Mycoplasmataceae;Mycoplasma (0.98%)
Pseudomonadaceae;Pseudomonas (2.04%) Moraxellaceae;Acinetobacter (0.69%)
Pasteurellaceae;Haemophilus (2.11%)
Enterobacteriaceae;Serratia (0.09%) Enterobacteriaceae;Proteus (1.70%) Enterobacteriaceae;Klebsiella/Enterobacter (10.50%) Enterobacteriaceae;Citrobacter (3.31%)
Comamonadaceae;Acidovorax (0.14%) Burkholderiales Insertae Sedis;Aquabacterium (0.08%)
Sphingomonadales;Sphingomonadaceae;Sphingomonas (0.12%) Rhodobacterales;Rhodobacteraceae;Paracoccus (0.14%) Rhizobiales;Bradyrhizobiaceae;Bradyrhizobium (0.07%) Caulobacterales;Caulobacteraceae;Brevundimonas (0.05%)
Veillonellaceae;Veillonella (0.99%) Peptostreptococcaceae;Clostridium (0.14%) Incertae Sedis XI;Peptoniphilus (0.09%) Clostridiaceae;Clostridium (0.87%)
Streptococcaceae;Streptococcus (15.46%) Enterococcaceae;Enterococcus (10.08%) Carnobacteriaceae;Granulicatella (0.27%)
Staphylococcaceae;Staphylococcus (40.77%) Staphylococcaceae;Gemella (2.70%)
Streptophyta;chloroplast (0.11%)
Flavobacteriaceae;Cloacibacterium (0.09%)
Porphyromonadaceae;Dysgonomonas (0.16%) Bacteroidaceae;Bacteroides (5.02%)
Propionibacteriaceae;Propionibacterium (0.16%) Micrococcaceae;Micrococcus (0.05%) Corynebacteriaceae;Corynebacterium (0.12%)
Actinobacteria;Actinobacteria;Actinomycetales;
Bacteroidetes;Flavobacteria;Flavobacteriales;
Bacteroidetes;Bacteroidia;Bacteroidales;
Cyanobacteria;Cyanobacteria;Chloroplast;
Firmicutes;Bacilli;Bacillales;
Firmicutes;Bacilli;Lactobacillales;
Firmicutes;Clostridia;Clostridiales;
Proteobacteria;Gammaproteobacteria;Enterobacteriales;
Proteobacteria;Gammaproteobacteria;Pasteurellales;
Proteobacteria;Gammaproteobacteria;Pseudomonadales;
Tenericutes;Mollicutes;Mycoplasmatales;
Proteobacteria;Betaproteobacteria;Burkholderiales;
Proteobacteria;Alphaproteobacteria;
No data
No data
DZ tw
insM
Z twins
0.25 0.5
0.7
5 1 0.2
5 0.5
0.75 1 0
0.25 0.5
0.7
5 1 0 0
FIG 1 Stacked bar plots depicting the relative abundances of the 30 most abundant genus-level taxa in the LBW infants. Taxa were ranked according to theirmean abundance across all samples (percentages at right). Ten taxa had mean abundances of �1.00% (percentages in bold type within parentheses). Cs, Cesareansection delivery; V, vaginal delivery.
Costello et al.
4 ® mbio.asm.org November/December 2013 Volume 4 Issue 6 e00782-13
on July 17, 2018 by guesthttp://m
bio.asm.org/
Dow
nloaded from

absence based); thus, branch lengths associated with high- andlow-abundance taxa count equally.
Exploratory analysis using UniFrac-based principal coordinateanalysis (PCoA) revealed that, as in healthy adults (1, 2), bodysite—i.e., whether the community was from a saliva, skin, or stoolsample—was the primary determinant of bacterial communitycomposition in the LBW infants (Fig. 2). Indeed, microbiota com-position differed significantly across the three sites (permuta-tional multivariate analysis of variance [PERMANOVA] maintest, P � 0.001). This factor (“body site”) remained significantwhen hierarchically nested within “individuals” (i.e., when exam-ining within-infant distances only; PERMANOVA main test, P �0.001); however, in pairwise a posteriori tests, baby 4’s saliva andstool communities were undifferentiated overall (P � 0.323).
The relative abundance of seven genera differed significantlyacross the three body sites (ANOVA adjusted for Bonferroni’scorrection, P � 0.001). Among those with an average abundanceof �1.0%, Klebsiella/Enterobacter (genera indistinguishable usingthe available gene fragment), Enterococcus, and Citrobacter wereparticularly abundant in stool, as was Staphylococcus (largelyStaphylococcus epidermidis) on skin, and Streptococcus in saliva(Fig. 1). Controlling for sequencing effort, the number of opera-tional taxonomic units (OTUs) on skin was significantly higherthan the number in saliva or stool (see Fig. S2 in the supplementalmaterial).
Notably, Staphylococcus and Streptococcus, which are charac-teristically found on skin and in saliva, respectively, were surpris-ingly abundant at other sites in the LBW infants (Fig. 1), and thelevel of body site-driven compositional differentiation in the LBWinfants (as shown in Fig. 2) seemed lower than that reported forhealthy adults (1–3). Indeed, when we compared these groups
directly (see Fig. S1 in the supplemental material), we found thatthe effect of “body site” was smaller in LBW infants (PER-MANOVA �2 � 0.21) than in healthy adults (�2 � 0.34). Thisdirect comparison also revealed that, among the three sites exam-ined, LBW infant skin was the most adult-like in terms of micro-biota composition (Fig. 3).
Neonatal personalization of microbiota composition. Com-positional variation existed among the LBW infants (PER-MANOVA main test, P � 0.001), but the effect of “individual”(PERMANOVA �2 � 0.13) was smaller than the effect of “bodysite” (�2 � 0.21) (Fig. 2). It was also not the case that every babyharbored a highly personalized microbiota: in pairwise a posterioritests, the microbiota of babies 1 and 2 (the dizygotic [DZ] twins)were compositionally similar to each other and to the microbiotaof baby 5 (P values of �0.05). By day 21, the genus-level profilesfor the fecal bacterial communities of co-twins were remarkablysimilar (Fig. 1c); as follows, overall interindividual variability forthe distal gut decreased modestly as the cohort grew older (seeFig. S3 in the supplemental material). Throughout their hospital-ization, co-twins were generally colocated; however, specific as-pects of their care may have varied. For example, on DOL 21,babies 1 and 2 (the DZ twins) received different diets (Table 2).
The relative abundance of three genera differed significantlyamong the six infants (ANOVA adjusted for Bonferroni’s correc-tion, P � 0.001). Bacteroides (B. caccae) was particularly abundantin baby 6’s stool samples (at all ages), as was Proteus (P. mirabilis)in baby 3’s saliva and stool samples (early ages), and Haemophilus(H. parainfluenzae) in baby 4’s saliva and stool samples (early ages;also present at low abundance in monozygotic [MZ] co-twin)(Fig. 1). Baby 6 was the only term infant in the study; he was alsodelivered vaginally. Numerous studies link vaginal delivery toearly colonization by Bacteroides (35, 36).
A high degree of interindividual variation in fecal microbiotacomposition has been observed in preterm (23, 24, 28) and term(5, 7) infants. Our data suggest that this pattern extends to theneonatal skin and oral microbiota. The ultimate cause of interin-dividual variation may be difficult to ascertain— e.g., despite re-ceiving remarkably similar medical treatment (Tables 1 and 2),
-0.6 -0.4 -0.2 0.0 0.2 0.4-0.4
-0.2
0.0
0.2
0.4
0.6
PCo1 (18.8% of total variation)
PC
o2 (1
0.3%
of t
otal
var
iatio
n)
123456
SalivaSkinStool
Baby:
FIG 2 Unweighted UniFrac-based principal coordinate analysis (PCoA) ofLBW infant-associated bacterial communities. Each symbol represents thevalue for a sample, with the shape of the symbol indicating the infant (infants1 to 6) and the color indicating the body site. The percentages of variationexplained by the plotted principal coordinates (PCo1 and PCo2) are indicatedon the axes.
LBW babies versus healthy adults
Oral SkinStoo
l0.6
0.7
0.8
0.9
1.0
Uni
Frac
dis
tanc
e **** ********
FIG 3 Average unweighted UniFrac distances between LBW infants (pres-ent study) and healthy adults (references 1, 2, and 5; see Materials and Meth-ods) for oral, skin surface, and stool microbiota (250 sequences per sample).Values that are significantly different by Tukey’s posthoc tests are indicated bybars and 4 asterisks (P � 0.0001). Error bars represent 95% confidence inter-vals.
Gut, Oral, and Skin Microbiome Dynamics in LBW Neonates
November/December 2013 Volume 4 Issue 6 e00782-13 ® mbio.asm.org 5
on July 17, 2018 by guesthttp://m
bio.asm.org/
Dow
nloaded from
clear differences existed between the microbiomes of the MZ co-twins (Fig. 1).
Delayed compositional divergence of gut and oral communi-ties. As a categorical predictor, infant age was not associated withdifferences in bacterial community composition (PERMANOVAmain test, P � 0.935), and the relative abundance of only onegenus (Staphylococcus) changed consistently as the cohort grewolder (modest decline in stool and saliva; linear correlation, r ��0.485 and �0.387, respectively; adjusted for Bonferroni’s cor-rection, P � 0.011 and 0.064, respectively). Thus, microbiotacomposition was more stable over time (here, DOL 8, 10, 12, 15,18, and 21) than across body sites and host individuals.
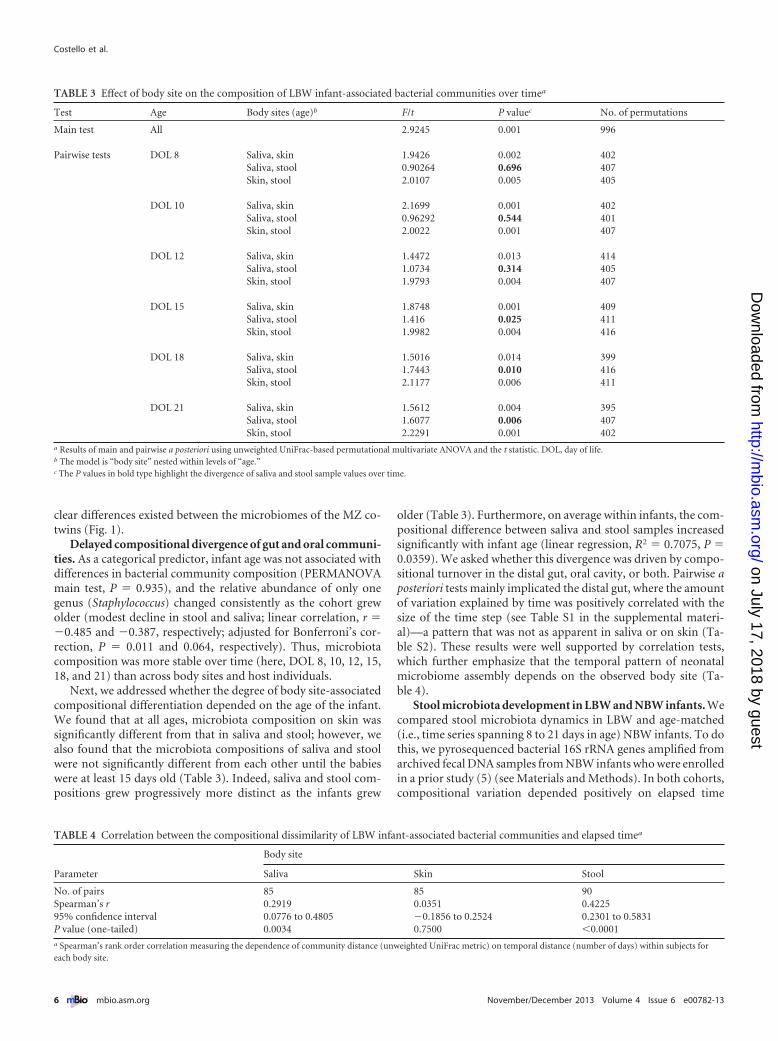
Next, we addressed whether the degree of body site-associatedcompositional differentiation depended on the age of the infant.We found that at all ages, microbiota composition on skin wassignificantly different from that in saliva and stool; however, wealso found that the microbiota compositions of saliva and stoolwere not significantly different from each other until the babieswere at least 15 days old (Table 3). Indeed, saliva and stool com-positions grew progressively more distinct as the infants grew
older (Table 3). Furthermore, on average within infants, the com-positional difference between saliva and stool samples increasedsignificantly with infant age (linear regression, R2 � 0.7075, P �0.0359). We asked whether this divergence was driven by compo-sitional turnover in the distal gut, oral cavity, or both. Pairwise aposteriori tests mainly implicated the distal gut, where the amountof variation explained by time was positively correlated with thesize of the time step (see Table S1 in the supplemental materi-al)—a pattern that was not as apparent in saliva or on skin (Ta-ble S2). These results were well supported by correlation tests,which further emphasize that the temporal pattern of neonatalmicrobiome assembly depends on the observed body site (Ta-ble 4).
Stool microbiota development in LBW and NBW infants. Wecompared stool microbiota dynamics in LBW and age-matched(i.e., time series spanning 8 to 21 days in age) NBW infants. To dothis, we pyrosequenced bacterial 16S rRNA genes amplified fromarchived fecal DNA samples from NBW infants who were enrolledin a prior study (5) (see Materials and Methods). In both cohorts,compositional variation depended positively on elapsed time
TABLE 3 Effect of body site on the composition of LBW infant-associated bacterial communities over timea
Test Age Body sites (age)b F/t P valuec No. of permutations
Main test All 2.9245 0.001 996
Pairwise tests DOL 8 Saliva, skin 1.9426 0.002 402Saliva, stool 0.90264 0.696 407Skin, stool 2.0107 0.005 405
DOL 10 Saliva, skin 2.1699 0.001 402Saliva, stool 0.96292 0.544 401Skin, stool 2.0022 0.001 407
DOL 12 Saliva, skin 1.4472 0.013 414Saliva, stool 1.0734 0.314 405Skin, stool 1.9793 0.004 407
DOL 15 Saliva, skin 1.8748 0.001 409Saliva, stool 1.416 0.025 411Skin, stool 1.9982 0.004 416
DOL 18 Saliva, skin 1.5016 0.014 399Saliva, stool 1.7443 0.010 416Skin, stool 2.1177 0.006 411
DOL 21 Saliva, skin 1.5612 0.004 395Saliva, stool 1.6077 0.006 407Skin, stool 2.2291 0.001 402
a Results of main and pairwise a posteriori using unweighted UniFrac-based permutational multivariate ANOVA and the t statistic. DOL, day of life.b The model is “body site” nested within levels of “age.”c The P values in bold type highlight the divergence of saliva and stool sample values over time.
TABLE 4 Correlation between the compositional dissimilarity of LBW infant-associated bacterial communities and elapsed timea
Parameter
Body site
Saliva Skin Stool
No. of pairs 85 85 90Spearman’s r 0.2919 0.0351 0.422595% confidence interval 0.0776 to 0.4805 �0.1856 to 0.2524 0.2301 to 0.5831P value (one-tailed) 0.0034 0.7500 �0.0001a Spearman’s rank order correlation measuring the dependence of community distance (unweighted UniFrac metric) on temporal distance (number of days) within subjects foreach body site.
Costello et al.
6 ® mbio.asm.org November/December 2013 Volume 4 Issue 6 e00782-13
on July 17, 2018 by guesthttp://m
bio.asm.org/
Dow
nloaded from

(Fig. 4a, P values of �0.05). We also found that there was nosignificant difference between the cohorts with respect to the rateof compositional turnover (Fig. 4a, P � 0.7911). On average, thestool microbiotas of LBW infants were slightly enriched in the
observed number of OTUs (controlling for sequencing effort; P �0.01) and significantly enriched in OTUs assigned to Enterobacter,the Enterobacteriaceae, Enterococcus, and Staphylococcus (adjustedfor Bonferroni’s correction, P values of �0.001). Escherichia wasabundant in the NBW infants and virtually absent from the LBWinfants (P � 0.001). Despite these differences, over time, thecommunity-wide composition of LBW infant stool grew moresimilar to that of 21-day-old NBW infant stool (i.e., to that of ahealthy reference group; Fig. 4b). These results suggest that whilegestational age at delivery, delivery mode, or other factors mayaffect gut microbiota makeup, its rate of development may de-pend more on intrinsic community-level factors, e.g., the amountof time the site has been available to colonists, microbe-microbeinteractions, microbe-host interactions (that are independent ofhost gestational age), or increasing hypoxia/anaerobiosis.
Dynamics of particular taxa in LBW infants, including anuncultivated Mycoplasma. Several noteworthy taxa were brieflyabundant in LBW infant stool samples (Fig. 1c). On day 18, Clos-tridium perfringens represented ~40% of sequences from baby 2,but it was below the detection level on all other days. On day 15,Dysgonomonas capnocytophagoides comprised ~8% of sequencesfrom baby 3; this fastidious organism (and opportunistic patho-gen) has not, to our knowledge, been reported in pediatric clinicalsamples. Finally, on day 15, a Peptoniphilus sp. represented ~7% ofsequences from baby 4, having been detected previously in his day12 skin swab (1%; Fig. 1b)—a possible bellwether for the taxon’sappearance in the distal gut.
However, the most striking example emerged from the oraldata set and involved taxa from baby 3’s saliva samples: specifi-cally, the genera Mycoplasma (several species) and Pseudomonas(P. aeruginosa), which became dominant on days 15, 18, and 21(Fig. 1a). Indeed, the sequences comprising one, highly abundantMycoplasma-related OTU appeared to be phylogenetically novel.This finding prompted an in-depth analysis of these and relatedsequences belonging to the phylum Tenericutes.
Among the OTUs detected in the LBW infants, three were as-signed to the phylum Tenericutes; together, they contained 788sequences. Representatives of the first and second OTUs were�99% identical to Mycoplasma hominis and Ureaplasma parvum,respectively. However, the representative of the third OTU, whichcontained 771 sequences, was only 88% identical to the closestnamed species in GenBank (e.g., Mycoplasma iowae, Myco-plasma microti, and Mycoplasma muris). This novel OTU was vir-tually exclusive to baby 3, the only extremely LBW (ELBW) infantin the study (ELBW is defined as �1.0 kg). Its expansion in baby3’s oral cavity, which peaked on DOL 18 at 47.2% of sequences,coincided with antibiotic treatment for suspected (but ultimatelyunconfirmed) sepsis (Fig. 5 and Table 2).
Phylogenetic analysis suggests that the novel OTU belongs to asingle, well-supported clade comprising uncultivated lineagesfrom cow rumen, which are among its closest relatives at 94.3 to94.8% sequence identity, and termite gut (see Fig. S4 in the sup-plemental material). Interestingly, a recently deposited GenBanksequence (uncultured Mycoplasma sp. clone Mnola; accession no.JX508800) is 99% identical to our infant-derived OTU (Fig. S4);this clone was isolated from a vaginal swab from a Trichomonasvaginalis-infected patient (37). Finally, we amplified and clonednear-full-length 16S rRNA gene sequences from baby 3’s DOL 18saliva (see Materials and Methods). This yielded sequences be-longing to the novel OTU that confirmed the phylogenetic place-
0 2 4 6 8 10 12 140.5
0.6
0.7
0.8
0.9
8 10 12 15 18 210.5
0.6
0.7
Elapsed time (days)
a
b
Uni
Frac
dis
tanc
eU
niFr
ac d
ista
nce
LBW baby age (days)
LBW babies (age 8-21 d; this study)NBW babies (age 8-21 d; reference 5)
Compared to 21-d-old NBW babies (ref. 5)
FIG 4 Relationship between neonatal stool microbiota composition andtime. (a) Average (95% CI) within-subject, unweighted UniFrac distance plot-ted against the age difference (lag) in days for LBW (R2 � 0.5) and NBW (R2 �0.5) infants. Lines indicate best fit linear regressions. The NBW infants werenot sampled on day 18; thus, for this analysis, the corresponding age wasexcluded from the LBW data set. (b) For LBW infants at various ages, average(95% CI) unweighted UniFrac distance compared to a healthy reference co-hort (the 21-day-old NBW infants).
Gut, Oral, and Skin Microbiome Dynamics in LBW Neonates
November/December 2013 Volume 4 Issue 6 e00782-13 ® mbio.asm.org 7
on July 17, 2018 by guesthttp://m
bio.asm.org/
Dow
nloaded from

ment of the shorter pyrosequences (Fig. S4). To our knowledge,this is the first report of infant-derived (and second report ofhuman-derived) sequences from this as-yet-uncultivatedMycoplasma-related clade.
DISCUSSION
In a small cohort of 8- to 21-day-old LBW infants, we found thatmicrobiota composition was shaped primarily by body site andhost individual; this is consistent with patterns observed inhealthy adults (1–3). Minutes after delivery, the composition ofthe newborn microbiota is undifferentiated across body sites (4).Our results suggest that site-specific bacterial communitiesemerge relatively early—indeed, within the neonatal period— de-spite an overall dearth of microbes characteristic of healthy adults(see Fig. S1 in the supplemental material). To our knowledge, thisis the first study to assess microbiota differentiation across multi-ple body sites in neonates; at the present time, there are no otherdata available from multiple body sites in the same baby, so wecannot directly evaluate whether similar patterns occur in, forexample, NBW infants.
Among the three sites examined, LBW infant skin was the mostadult-like in terms of microbiota composition (Fig. 3); this mayresult from infant skin being more selective for, and/or moreheavily exposed to, the skin microbiota of adult caretakers in theNICU compared to other body sites (33), although we did notquantify the amount of time each infant spent in direct contactwith mothers or other caregivers. (In the mouth and gut, the maindifference between neonates and adults seems to be a relative lackof strict anaerobes [38].) While developmental changes over thefirst year of life have been reported for the infant skin microbiome
(8), they were not apparent within the relatively short, neonataltime frame of the current study (Table 4).
Finally, delivery mode has been noted to exert a strong influ-ence on the composition of the newborn microbiota (4); while thiseffect was conceivably manifest in our study (e.g., Ureaplasma inbaby 3; Bacteroides in baby 6 [Fig. 1] [36, 39]), its pervasivenessand persistence will require examination in larger cohorts of high-risk infants.
We found that microbiota composition was relatively stableover time within LBW neonates. This small effect size for time,compared to those for body site and host individual, is also con-sistent with patterns observed in healthy adults (1–3, 40). None-theless, our comparative approach uncovered subtle yet impor-tant temporal changes that occurred over the 8- to 21-day agerange: in particular, a gradual (i.e., delayed) compositional diver-gence of the oral and fecal microbiota (Table 3), largely driven byprogressive temporal turnover in the distal gut (Table 4), the latterof which proceeded at a rate indistinguishable from that of age-matched NBW infants (Fig. 4a). Long recognized as a key processtaking place in early infancy (38, 41–43), our study draws intofocus the initiation phase of gut microbiome development, cap-turing, possibly, the time span over which the site begins to receiveand select for gut-specific microbes, which may then grow to out-number or outcompete transient or generalist immigrants fromthe oral cavity (or other sources shared by the two sites). However,given our small cohort of six infants for which there were a num-ber of uncontrolled variables (e.g., gestational age at delivery, mul-tiple gestation, medical treatment, delivery mode), we cautionthat our data are likely limited in terms of their generalizabilityand capacity to detect subtle effects. The biogeographic patternswe report warrant follow-up in larger, well-controlled, prospec-tive cohort studies.
We also detected a novel, uncultivated lineage of Mycoplasmaat high abundance in the oral cavity of ELBW baby 3. Mycoplasmaand Ureaplasma spp. colonize the human respiratory and urogen-ital tracts, and some play roles as perinatal pathogens (39).M. hominis and Ureaplasma spp. can cause chorioamnionitis (arisk factor for preterm premature rupture of membranes[PPROM]) and pass from mother to newborn, and the latterorganisms have been associated with preterm labor and low birth-weight (39, 44). In neonates, they cause respiratory, blood, andcentral nervous system (CNS) infections (39). Lacking cell walls,these organisms are innately resistant to beta-lactam (e.g., ampi-cillin, cefotaxime) and glycopeptide (e.g., vancomycin) antibiotics(45). Although not innately resistant, their susceptibility to ami-noglycosides (e.g., gentamicin) is variable (46).
Baby 3 was delivered vaginally after PPROM at ~24.5 weeks ofcompleted gestation and was treated intravenously with ampicil-lin and gentamicin for the first 7 days of life. Thus, carriage ofMycoplasma- and Ureaplasma-related OTUs at low abundance atthe start of the study, on DOL 8, may have been due to verticaltransmission at delivery, followed by resistance to the initialcourse of antibiotics, although alternative scenarios are possible(e.g., later exposure in the NICU). Baby 3 was again treated withantibiotics (vancomycin, gentamicin, cefotaxime) from DOL 13to 19 (Table 2), and this coincided with a marked increase in theproportional abundance of Pseudomonas aeruginosa (Fig. 1a) andOTU 15, a member of a novel, uncultivated clade belonging to theMycoplasmataceae, in baby 3’s oral samples (Fig. 5; see Fig. S4 inthe supplemental material). Intriguingly, a recent study found
8 10 12 14 16 18 20 220.0
0.5
10
20
30
40
50
Age in days
Perc
enta
ge o
f seq
uenc
es
+-
++
++
-+
-+
-+
Antibiotics:Breast milk:
Formula:
0
OTU 15 (uncultivated Mycoplasma sp.)OTU 53 (Ureaplasma parvum)OTU 144 (Mycoplasma hominis)
FIG 5 Relative abundances of three OTUs belonging to the Mycoplasmata-ceae from oral samples from extremely LBW baby 3. OTU 15, a novel, uncul-tivated Mycoplasma sp., is plotted against the upper y axis. OTUs 53 and 144,which are closely related to OTUs from cultivated strains, are plotted againstthe lower y axis. Expansion of OTU 15 coincided with antibiotic treatmentfrom DOL 13 to 19 (see Table 2 for details). Feedings were delivered via naso-or orogastric tube. For antibiotics, the date range is indicated. The baby’s diet(breast milk or formula) is given for each sample date.
Costello et al.
8 ® mbio.asm.org November/December 2013 Volume 4 Issue 6 e00782-13
on July 17, 2018 by guesthttp://m
bio.asm.org/
Dow
nloaded from

high abundances of this uncultivated Mycoplasma in the vaginalmicrobiota of Trichomonas vaginalis-infected women (detected in19/30 T. vaginalis-infected and 1/29 uninfected individuals) (37),again raising the possibility that this organism too was transferredfrom mother to infant at delivery. Further investigation into thediversity, distribution, and clinical significance of this novel, un-cultivated Mycoplasma in human hosts is warranted, particularlyin pregnant women and premature infants.
Although the LBW infants in this study were relatively free ofmajor medical problems, we found that their microbiomes weredominated at times by bacterial taxa that have been associatedwith neonatal infections and NEC, e.g., Staphylococcus, C. perfrin-gens, P. aeruginosa, and others (28, 32, 47, 48). Yet, despite theabundance of taxa with pathogenic potential, it appears that cer-tain normal processes were under way, including the developmentof body site-specific bacterial communities and progressive com-positional turnover in the distal gut, as observed in healthy hosts(2, 38). Our analysis was cohort based; however, it might be usefulto know whether individual infants vary in the precise timing ofbody site-associated compositional differentiation, and if so,whether such variation depends on gestational age at delivery orparticular NICU management protocols. Unfortunately, our co-hort was not well suited to this analysis because of its small size,but also because gestational age at delivery was confounded withdelivery location and the amount of time spent in the NICU (Ta-bles 1 and 2). This underscores a need for larger and distinct co-horts but also highlights a challenge: the smallest, most prematureinfants will almost always require the most intensive medical sup-port, thus entangling factors such as gut and immune immaturitywith, for example, the number of invasive procedures or days onantibiotics. Nevertheless, monitoring of oral and other potentialsource communities in the NICU might be particularly warrantedduring the time the gut microbiome remains “undifferentiated”and, possibly, more open to invasion.
MATERIALS AND METHODSPatients and sample collection. Six low-birthweight (LBW) infants wererecruited from a level III NICU at the University of Chicago Comer Chil-dren’s Hospital. The infants were born within 1 week of each other in thesummer of 2010. The cause of the low birthweight was preterm delivery infive of the infants (a singleton and two pairs of twins) and fetal growthrestriction in the sixth. Birth weights ranged from 0.75 to 1.82 kg (seeTable 1 for clinical details; �2.5 kg is considered low birthweight). Stooland saliva samples and skin swabs were obtained from each infant onpostnatal days 8, 10, 12, 15, 18, and 21. The age range of 8 to 21 days wasselected because it may represent a critical window for the colonization ofthe infant, and although it did not occur in the present cohort, for theonset of NEC. Stool sampling involved manual perineal stimulation witha lubricated cotton swab, which induced prompt defecation. Oral and skinsamples were collected by gently swabbing the dorsum of the tongue andthe anterior upper chest wall, respectively. For the oral samples, we simplycall the collected materials “saliva,” because it is likely that multiple siteswere contacted during the gentle swabbing. Samples were collected usingsterile nylon or cotton swabs, placed in 3 ml of universal transport me-dium (UTM; EMD Millipore, Billerica, MA), and promptly frozen at�80°C. A total of 108 samples were collected for the study. Data pertain-ing to the care and location of the infants during the sampling period arepresented in Table 2. All infants remained hospitalized throughout thestudy. The Institutional Review Board of the University of Chicago ap-proved the study protocol, and the infants’ parents provided written in-formed consent.
DNA extraction, PCR amplification, and pyrosequencing. GenomicDNA was isolated from each sample (1.5 ml UTM) using a QIAamp DNAstool minikit (Qiagen, Valencia, CA) with modifications, including beadbeating (49). A fragment of the 16S rRNA gene spanning the V3-V5 hy-pervariable regions was amplified. The forward primer (5= CGT ATCGCC TCC CTC GCG CCA TCA GNN NNN NNN NNN NGC ACT CCTACG GGA GGC AGC A 3=) contained the 454 Life Sciences primer Asequence, a unique 12-nucleotide (nt) error-correcting Golay barcodeused to label each amplicon (designated by the N’s) (50), the broad-rangebacterial primer 338F (F stands for forward), and a two-base linker lo-cated between the bar code and the rRNA primer (GC). The reverseprimer (5= CTA TGC GCC TTG CCA GCC CGC TCA GAA CCG TCAATT CCT TTG AGT TT 3=) contained the 454 Life Sciences primer Bsequence, a two-base linker (AA), and the broad-range bacterial primer906R (R stands for reverse). Amplifications were carried out in triplicate25-�l reactions using 0.4 �M forward and reverse primers, 3-�l templateDNA, and 1� HotMasterMix (5 PRIME, Gaithersburg, MD). Bovine se-rum albumin (BSA) was added at a final concentration of 0.1 �g/�l toreaction mixtures containing fecal DNA. Thermal cycling was carried outat 94°C for 2 min, followed by 35 cycles, with 1 cycle consisting of 94°C for45 s, 50°C for 30 s, and 72°C for 90 s, with a final extension step of 10 minat 72°C. Replicate reactions were pooled and then purified using an Ultra-Clean-htp 96-well PCR clean-up kit according to the manufacturer’s in-structions (MO BIO, Carlsbad, CA).
DNA concentrations were determined using a high-sensitivityQuant-iT double-stranded DNA (dsDNA) kit according to the manufac-turer’s instructions (Invitrogen, Carlsbad, CA). Purified amplicons werecombined in equimolar ratios into a single tube, ethanol precipitated, andresuspended in 100 �l of nuclease-free water. The pooled DNA was gelpurified and recovered using a QIAquick gel extraction kit (Qiagen). Uni-directional amplicon sequencing was performed by the W. M. Keck Cen-ter for Comparative and Functional Genomics at the University of Illinois,Urbana-Champaign using a 454 Life Sciences genome sequencer FLX in-strument, titanium (Ti) series reagents, primer A, and 6 regions of a 16-region gasket (Roche, Branford, CT). Sequencing generated 186,428 rawreads.
Sequence analysis. Raw reads were filtered using the QIIME softwarepackage (51). Reads were removed from the analysis if they were �200 or�600 nt in length, contained an ambiguous base, had a mean quality scoreof �25 across the entire read, contained a homopolymer run �6 nt inlength, did not contain the forward primer sequence, or contained anuncorrectable barcode. Remaining reads were truncated at the first base ofthe first 50-nt sliding window with a mean quality score of �25 (if found),and retained unless �200 nt in length after truncation. Filtered reads wereassigned to samples by examining the 12-nt barcode. A total of 119,191filtered reads were associated with samples at this step (mean read length,535 nt).
Error correction, chimera detection (using UCHIME), and clusteringof filtered reads into de novo operational taxonomic units (OTUs) at 97%sequence identity were performed in USEARCH using otupipe-likescripts enabled in QIIME (http://www.drive5.com/usearch/manual/otu_clustering.html) (52, 53). A representative sequence was chosen fromeach OTU by selecting the “first” sequence (i.e., the UCLUST clusterseed). Representative sequences were aligned against the Greengenes coreset (54) using PyNAST (55) with a minimum alignment length of 150 ntand a minimum identity of 80%. Fifteen OTU representative sequencesfailed to align; BLASTn searches against GenBank’s nr/nt database re-vealed 13 human OTUs, 1 Candida albicans OTU (representing 276 readsfrom baby 6, day 8 stool), and 1 poor-quality OTU, all of which wereexcluded from further analysis. Taxonomic assignments were made usingthe Ribosomal Database Project (RDP) classifier version 2.2 with a mini-mum support threshold of 80% and the RDP taxonomic nomenclature(56). For the most abundant OTUs study-wide (here, those with �0.05%average abundance across all samples), RDP assignments were manuallyconfirmed and, when possible, annotated with species-level information
Gut, Oral, and Skin Microbiome Dynamics in LBW Neonates
November/December 2013 Volume 4 Issue 6 e00782-13 ® mbio.asm.org 9
on July 17, 2018 by guesthttp://m
bio.asm.org/
Dow
nloaded from

using BLASTn searches against the nr/nt database. A table of sequencecounts per classified OTU � sample was generated in which the criteriafor an OTU’s inclusion were that it contained at least 2 sequences and wasassigned at least to the genus level. The final OTU table consisted of 321OTUs containing a total of 105,462 sequences.
Sequences representing OTUs that did not make it into the final tablewere removed from the alignment. Hypervariable (i.e., uninformative)positions were then excluded using the PH Lane mask (57). A phylogenywas inferred using FastTree version 2.1.3 (58) with the Jukes-Cantor plusCAT model. The final OTU table and phylogeny served as inputs to sub-sequent analyses, including rarefaction, � and � diversity calculations,unweighted UniFrac-based principal coordinate analysis (PCoA), andphylum- and genus-level taxonomic summaries implemented in QIIME.Unweighted UniFrac-based permutational multivariate analysis of vari-ance (PERMANOVA) was performed in PRIMER-E version 6 (59). Otherstatistical tests were performed in QIIME or Prism (GraphPad Software,Inc.).
Sequence analysis focused on a novel, uncultivated oral Myco-plasma. Phylogenetic relationships among sequences belonging to OTUsassigned to the phylum Tenericutes (3 OTUs) were investigated in detail.This analysis was prompted by the identification of an OTU assigned tothe genus Mycoplasma containing 771 reads (99% of which were frombaby 3’s saliva) and exhibiting low sequence identity (~88%) to the mostclosely related cultivated strains represented in GenBank (http://www.ncbi.nlm.nih.gov/genbank/). Sequences were aligned against the Green-genes core set using the NAST algorithm (60) (http://greengenes.lbl.gov)and imported into ARB (version 08.08.27) (61). In ARB, the alignmentwas manually improved using secondary structure information and align-ment to nearest neighbors in the context of an expanded, in-house data-base founded upon the Greengenes alignment. Phylogenetic relationshipsamong the 3 OTUs found in the present study, their closest relatives (un-cultivated mycoplasmas), and selected representatives of cultivated Te-nericutes were inferred using bootstrapped maximum likelihood infer-ence methods in RAxML (version 7.2.8) (62). In order to confirm andfurther explore the phylogenetic placement of the novel Mycoplasma-related OTU, a small number of near-full-length 16S rRNA gene se-quences were recovered from baby 3’s day 18 saliva sample via amplifica-tion (with primers 8F/1391R), cloning, and Sanger sequencing usingmethods described elsewhere (63). Fifteen high-quality sequences wereassembled (4 uncultivated Mycoplasma sequences and 11 Pseudomonasaeruginosa sequences). The near-full-length Mycoplasma sequences wereanalyzed using NAST, ARB, and RAxML as described above.
Comparison to microbiota of NBW infants via pyrosequencing ofarchived stool DNA. Archived stool DNA samples from healthy,age-matched (i.e., time series spanning 8 to 21 days in age), normal birth-weight (NBW) (�2.5 kg) infants enrolled in a prior study (5) were ampli-fied, sequenced, and analyzed using the pyrosequencing and bioinformat-ics approaches described herein. The archived DNA had been isolatedusing the QIAamp stool DNA minikit (Qiagen) and stored at �80°C. TheStanford University Administrative Panel on Human Subjects in MedicalResearch approved this work, and the infants’ parents provided writteninformed consent.
Comparison to microbiota of healthy adults using publically avail-able sequence data. Sequence data from the LBW and NBW infants werecompared to publically available sequence data from the correspondingbody sites of healthy adults. Adult data were selected from two publishedstudies that used pyrosequencing approaches similar to those used here(1, 2). From the first study (1), we selected samples from 7 adults (3female), “days 1 and 2” (of 4 sampling dates), including dorsal tongueswabs, skin swabs (forehead and right forearm), and stool samples (56samples in total). These 16S rRNA gene sequences were V2 region FLXreads originating from the distal primer (338R). From the second study(2), we selected samples from 6 adults (a subset chosen at random butmatched for gender to the LBW infants), “visit 2” (of 2 sampling visits),including saliva samples, skin swabs (right retroauricular crease), and
stool samples (18 samples in total). These were V3-V5 region Ti readsoriginating from the distal primer (926R). By comparison, the infant-derived sequences generated for the present study were V3-V5 region Tireads originating from the proximal primer (338F). Thus, given the dif-ferences in sequence length and sequenced region among the data sets, thepooled sequences were trimmed to a length of not more than 300 nt andOTUs were picked against a set of reference sequences. This was accom-plished in QIIME using uclust_ref-based OTU picking against the Green-genes gg_97_otus_4feb2011.fasta reference set at an identity threshold of95% (relaxed from 97% to allow for greater recruitment), with reversestrand matching enabled and no new clusters allowed. A total of 3,158reference OTUs were detected; these encompassed 96% of the 475,080total sequences. Rarefied and unrarefied OTU tables, along with a refer-ence tree (gg_97_otus_4feb2011.tre), were used to calculate unweightedUniFrac distance matrices, which served as inputs for PCoA in QIIME.
The DNA extraction method varied among the studies compared:studies of adults used a MO BIO kit, while studies of infants used a Qiagenkit. To investigate potential kit-associated bias, we pyrosequenced 16SrRNA genes amplified from archived adult stool DNA that had been iso-lated using a Qiagen kit (from the NBW infants’ fathers [5]; mothers wereexcluded due to possible pregnancy-associated shifts in microbiota com-position [64]). These new adult sequences were filtered as describedherein and trimmed to a length of not more than 300 nt, pooled with theother sequences, and analyzed as described in the preceding paragraph.Because the Qiagen-extracted adult stool samples (5) clustered with theMO BIO-extracted ones (1, 2) (see Fig. S1 in the supplemental material),we concluded that DNA extraction kit did not grossly bias the results ofthe unweighted UniFrac-based PCoA.
Nucleotide sequence accession numbers. The sequence data gener-ated for this study were deposited in the QIIME database (study identifi-cation numbers 2042 and 2046).
SUPPLEMENTAL MATERIALSupplemental material for this article may be found at http://mbio.asm.org/lookup/suppl/doi:10.1128/mBio.00782-13/-/DCSupplemental.
Figure S1, PDF file, 0.4 MB.Figure S2, PDF file, 0.2 MB.Figure S3, PDF file, 0.2 MB.Figure S4, PDF file, 0.4 MB.Table S1, DOCX file, 0.1 MB.Table S2, DOCX file, 0.1 MB.
ACKNOWLEDGMENTS
We thank the subjects and their families for their participation and theNICU staff for their support. We also thank members of the Morowitz andRelman laboratories, and in particular Valeriy Poroyko (University ofChicago) for technical assistance and Diana Proctor (Stanford University)for critical review of the manuscript.
This work was supported in part by NIH grant 1R01AI092531-01(M.J.M. and D.A.R.; Jill Banfield, principal investigator [PI]), a Walter V.and Idun Berry postdoctoral fellowship (E.K.C.), NIH Pioneer awardDP1OD000964 (D.A.R.), March of Dimes Foundation research grant5-FY10-103 (M.J.M.), the March of Dimes Prematurity Research Centerat Stanford University School of Medicine (D.A.R.), and by the Thomas C.and Joan M. Merigan Endowment at Stanford University (D.A.R.).
REFERENCES1. Costello EK, Lauber CL, Hamady M, Fierer N, Gordon JI, Knight R.
2009. Bacterial community variation in human body habitats across spaceand time. Science 326:1694 –1697.
2. Microbiome, Human Project Consortium. 2012. Structure, function anddiversity of the healthy human microbiome. Nature 486:207–214.
3. Caporaso JG, Lauber CL, Costello EK, Berg-Lyons D, Gonzalez A,Stombaugh J, Knights D, Gajer P, Ravel J, Fierer N, Gordon JI, KnightR. 2011. Moving pictures of the human microbiome. Genome Biol. 12:R50. doi:10.1186/gb-2011-12-5-r50.
4. Dominguez-Bello MG, Costello EK, Contreras M, Magris M, Hidalgo
Costello et al.
10 ® mbio.asm.org November/December 2013 Volume 4 Issue 6 e00782-13
on July 17, 2018 by guesthttp://m
bio.asm.org/
Dow
nloaded from

G, Fierer N, Knight R. 2010. Delivery mode shapes the acquisition andstructure of the initial microbiota across multiple body habitats in new-borns. Proc. Natl. Acad. Sci. U. S. A. 107:11971–11975.
5. Palmer C, Bik EM, DiGiulio DB, Relman DA, Brown PO. 2007. Devel-opment of the human infant intestinal microbiota. PLoS Biol. 5:e177.doi:10.1371/journal.pbio.0050177.
6. Koenig JE, Spor A, Scalfone N, Fricker AD, Stombaugh J, Knight R,Angenent LT, Ley RE. 2011. Succession of microbial consortia in thedeveloping infant gut microbiome. Proc. Natl. Acad. Sci. U. S. A.108(Suppl 1):4578 – 4585.
7. Yatsunenko T, Rey FE, Manary MJ, Trehan I, Dominguez-Bello MG,Contreras M, Magris M, Hidalgo G, Baldassano RN, Anokhin AP,Heath AC, Warner B, Reeder J, Kuczynski J, Caporaso JG, LozuponeCA, Lauber C, Clemente JC, Knights D, Knight R, Gordon JI. 2012.Human gut microbiome viewed across age and geography. Nature 486:222–227.
8. Capone KA, Dowd SE, Stamatas GN, Nikolovski J. 2011. Diversity of thehuman skin microbiome early in life. J. Invest. Dermatol. 131:2026 –2032.
9. Crielaard W, Zaura E, Schuller AA, Huse SM, Montijn RC, Keijser BJ.2011. Exploring the oral microbiota of children at various developmentalstages of their dentition in the relation to their oral health. BMC Med.Genomics 4:22. doi:10.1186/1755-8794-4-22.
10. Morowitz MJ, Poroyko V, Caplan M, Alverdy J, Liu DC. 2010. Rede-fining the role of intestinal microbes in the pathogenesis of necrotizingenterocolitis. Pediatrics 125:777–785.
11. Neu J, Walker WA. 2011. Necrotizing enterocolitis. N. Engl. J. Med.364:255–264.
12. Stoll BJ, Hansen N, Fanaroff AA, Wright LL, Carlo WA, EhrenkranzRA, Lemons JA, Donovan EF, Stark AR, Tyson JE, Oh W, Bauer CR,Korones SB, Shankaran S, Laptook AR, Stevenson DK, Papile LA, PooleWK. 2002. Late-onset sepsis in very low birth weight neonates: the expe-rience of the NICHD Neonatal Research Network. Pediatrics 110:285–291.
13. Yee WH, Soraisham AS, Shah VS, Aziz K, Yoon W, Lee SK, CanadianNeonatal Network. 2012. Incidence and timing of presentation of necro-tizing enterocolitis in preterm infants. Pediatrics 129:e298 – e304.
14. González-Rivera R, Culverhouse RC, Hamvas A, Tarr PI, Warner BB.2011. The age of necrotizing enterocolitis onset: an application ofSartwell’s incubation period model. J. Perinatol. 31:519 –523.
15. Levy O. 2007. Innate immunity of the newborn: basic mechanisms andclinical correlates. Nat. Rev. Immunol. 7:379 –390.
16. Hooper LV, Gordon JI. 2001. Commensal host-bacterial relationships inthe gut. Science 292:1115–1118.
17. Sekirov I, Finlay BB. 2009. The role of the intestinal microbiota in entericinfection. J. Physiol. (Lond.) 587:4159 – 4167.
18. Cho I, Yamanishi S, Cox L, Methé BA, Zavadil J, Li K, Gao Z, MahanaD, Raju K, Teitler I, Li H, Alekseyenko AV, Blaser MJ. 2012. Antibioticsin early life alter the murine colonic microbiome and adiposity. Nature488:621– 626.
19. Olszak T, An D, Zeissig S, Vera MP, Richter J, Franke A, Glickman JN,Siebert R, Baron RM, Kasper DL, Blumberg RS. 2012. Microbial expo-sure during early life has persistent effects on natural killer T cell function.Science 336:489 – 493.
20. Carlisle EM, Morowitz MJ. 2011. Pediatric surgery and the human mi-crobiome. J. Pediatr. Surg. 46:577–584.
21. Manzoni P, De Luca D, Stronati M, Jacqz-Aigrain E, Ruffinazzi G,Luparia M, Tavella E, Boano E, Castagnola E, Mostert M, Farina D.2013. Prevention of nosocomial infections in neonatal intensive careunits. Am. J. Perinatol. 30:81– 88.
22. Millar MR, Linton CJ, Cade A, Glancy D, Hall M, Jalal H. 1996.Application of 16S rRNA gene PCR to study bowel flora of preterm infantswith and without necrotizing enterocolitis. J. Clin. Microbiol. 34:2506 –2510.
23. Wang Y, Hoenig JD, Malin KJ, Qamar S, Petrof EO, Sun J, Antono-poulos DA, Chang EB, Claud EC. 2009. 16S rRNA gene-based analysis offecal microbiota from preterm infants with and without necrotizing en-terocolitis. ISME J. 3:944 –954.
24. Mshvildadze M, Neu J, Shuster J, Theriaque D, Li N, Mai V. 2010.Intestinal microbial ecology in premature infants assessed with non-culture-based techniques. J. Pediatr. 156:20 –25.
25. Chang JY, Shin SM, Chun J, Lee JH, Seo JK. 2011. Pyrosequencing-based molecular monitoring of the intestinal bacterial colonization in pre-term infants. J. Pediatr. Gastroenterol. Nutr. 53:512–519.
26. Mai V, Young CM, Ukhanova M, Wang X, Sun Y, Casella G, TheriaqueD, Li N, Sharma R, Hudak M, Neu J. 2011. Fecal microbiota in prema-ture infants prior to necrotizing enterocolitis. PLoS One 6:e20647. doi:10.1371/journal.pone.0020647.
27. Morowitz MJ, Denef VJ, Costello EK, Thomas BC, Poroyko V, RelmanDA, Banfield JF. 2011. Strain-resolved community genomic analysis ofgut microbial colonization in a premature infant. Proc. Natl. Acad. Sci.U. S. A. 108:1128 –1133.
28. Madan JC, Salari RC, Saxena D, Davidson L, O’Toole GA, Moore JH,Sogin ML, Foster JA, Edwards WH, Palumbo P, Hibberd PL. 2012. Gutmicrobial colonisation in premature neonates predicts neonatal sepsis.Arch. Dis. Child. (Fetal Neonatal Ed.) 97:F456 –F462.
29. Sharon I, Morowitz MJ, Thomas BC, Costello EK, Relman DA, BanfieldJF. 2013. Time series community genomics analysis reveals rapid shifts inbacterial species, strains, and phage during infant gut colonization. Ge-nome Res. 23:111–120.
30. Normann E, Fahlén A, Engstrand L, Lilja HE. 2013. Intestinal microbialprofiles in extremely preterm infants with and without necrotizing entero-colitis. Acta Paediatr. 102:129 –136.
31. Mai V, Torrazza RM, Ukhanova M, Wang X, Sun Y, Li N, Shuster J,Sharma R, Hudak ML, Neu J. 2013. Distortions in development ofintestinal microbiota associated with late onset sepsis in preterm infants.PLoS One 8:e52876. doi:10.1371/journal.pone.0052876.
32. Morrow AL, Lagomarcino AJ, Schibler KR, Taft DH, Yu Z, Wang B,Altaye M, Wagner M, Gevers D, Ward DV, Kennedy MA, HuttenhowerC, Newburg DS. 2013. Early microbial and metabolomic signatures pre-dict later onset of necrotizing enterocolitis in preterm infants. Micro-biome 1:13. doi:10.1186/2049-2618-1-13.
33. Keyworth N, Millar MR, Holland KT. 1992. Development of cutaneousmicroflora in premature neonates. Arch. Dis. Child. 67:797– 801.
34. Lozupone C, Knight R. 2005. UniFrac: a new phylogenetic method forcomparing microbial communities. Appl. Environ. Microbiol. 71:8228 – 8235.
35. Bennet R, Nord CE. 1987. Development of the faecal anaerobic micro-flora after caesarean section and treatment with antibiotics in newborninfants. Infection 15:332–336.
36. Penders J, Thijs C, Vink C, Stelma FF, Snijders B, Kummeling I, vanden Brandt PA, Stobberingh EE. 2006. Factors influencing the compo-sition of the intestinal microbiota in early infancy. Pediatrics 118:511–521.
37. Martin DH, Zozaya M, Lillis RA, Myers L, Nsuami MJ, Ferris MJ. 2013.Unique vaginal microbiota which include an unknown Mycoplasma-likeorganism are associated with Trichomonas vaginalis infection. J. Infect.Dis. 207:1922–1931.
38. Mackie RI, Sghir A, Gaskins HR. 1999. Developmental microbial ecologyof the neonatal gastrointestinal tract. Am. J. Clin. Nutr. 69:1035S–1045S.
39. Waites KB, Katz B, Schelonka RL. 2005. Mycoplasmas and ureaplasmasas neonatal pathogens. Clin. Microbiol. Rev. 18:757–789.
40. Kuczynski J, Costello EK, Nemergut DR, Zaneveld J, Lauber CL,Knights D, Koren O, Fierer N, Kelley ST, Ley RE, Gordon JI, Knight R.2010. Direct sequencing of the human microbiome readily reveals com-munity differences. Genome Biol. 11:210. doi:10.1186/gb-2010-11-5-210.
41. Schaedler RW, Dubos R, Costello R. 1965. The development of thebacterial flora in the gastrointestinal tract of mice. J. Exp. Med. 122:59 – 66.
42. Savage DC. 1977. Microbial ecology of the gastrointestinal tract. Annu.Rev. Microbiol. 31:107–133.
43. Murgas Torrazza R, Neu J. 2011. The developing intestinal microbiomeand its relationship to health and disease in the neonate. J. Perinatol.31(Suppl 1):S29 –S34.
44. DiGiulio DB, Romero R, Amogan HP, Kusanovic JP, Bik EM, GotschF, Kim CJ, Erez O, Edwin S, Relman DA. 2008. Microbial prevalence,diversity and abundance in amniotic fluid during preterm labor: a molec-ular and culture-based investigation. PLoS One 3:e3056. doi:10.1371/journal.pone.0003056.
45. Taylor-Robinson D, Bébéar C. 1997. Antibiotic susceptibilities of myco-plasmas and treatment of mycoplasmal infections. J. Antimicrob. Che-mother. 40:622– 630.
46. Waites KB, Crouse DT, Cassell GH. 1992. Antibiotic susceptibilities andtherapeutic options for Ureaplasma urealyticum infections in neonates.Pediatr. Infect. Dis. J. 11:23–29.
47. de la Cochetiere MF, Piloquet H, des Robert C, Darmaun D, GalmicheJP, Rozé JC. 2004. Early intestinal bacterial colonization and necrotizingenterocolitis in premature infants: the putative role of Clostridium. Pedi-atr. Res. 56:366 –370.
Gut, Oral, and Skin Microbiome Dynamics in LBW Neonates
November/December 2013 Volume 4 Issue 6 e00782-13 ® mbio.asm.org 11
on July 17, 2018 by guesthttp://m
bio.asm.org/
Dow
nloaded from

48. Stewart CJ, Marrs EC, Magorrian S, Nelson A, Lanyon C, Perry JD,Embleton ND, Cummings SP, Berrington JE. 2012. The preterm gutmicrobiota: changes associated with necrotizing enterocolitis and infec-tion. Acta Paediatr. 101:1121–1127.
49. Zoetendal EG, Heilig HG, Klaassens ES, Booijink CC, Kleerebezem M,Smidt H, De Vos WM. 2006. Isolation of DNA from bacterial samples ofthe human gastrointestinal tract. Nat. Protoc. 1:870 – 873.
50. Hamady M, Walker JJ, Harris JK, Gold NJ, Knight R. 2008. Error-correcting barcoded primers for pyrosequencing hundreds of samples inmultiplex. Nat. Methods 5:235–237.
51. Caporaso JG, Kuczynski J, Stombaugh J, Bittinger K, Bushman FD,Costello EK, Fierer N, Peña AG, Goodrich JK, Gordon JI, Huttley GA,Kelley ST, Knights D, Koenig JE, Ley RE, Lozupone CA, McDonald D,Muegge BD, Pirrung M, Reeder J, Sevinsky JR, Turnbaugh PJ, WaltersWA, Widmann J, Yatsunenko T, Zaneveld J, Knight R. 2010. QIIMEallows analysis of high-throughput community sequencing data. Nat.Methods 7:335–336.
52. Edgar RC. 2010. Search and clustering orders of magnitude faster thanBLAST. Bioinformatics 26:2460 –2461.
53. Edgar RC, Haas BJ, Clemente JC, Quince C, Knight R. 2011. UCHIMEimproves sensitivity and speed of chimera detection. Bioinformatics 27:2194 –2200.
54. DeSantis TZ, Hugenholtz P, Larsen N, Rojas M, Brodie EL, Keller K,Huber T, Dalevi D, Hu P, Andersen GL. 2006. Greengenes, a chimera-checked 16S rRNA gene database and workbench compatible with ARB.Appl. Environ. Microbiol. 72:5069 –5072.
55. Caporaso JG, Bittinger K, Bushman FD, DeSantis TZ, Andersen GL,Knight R. 2010. PyNAST: a flexible tool for aligning sequences to a tem-plate alignment. Bioinformatics 26:266 –267.
56. Wang Q, Garrity GM, Tiedje JM, Cole JR. 2007. Naive Bayesian classifier
for rapid assignment of rRNA sequences into the new bacterial taxonomy.Appl. Environ. Microbiol. 73:5261–5267.
57. Lane DJ. 1991. 16S/23S rRNA sequencing, p 115–148. In Stackebrandt E,Goodfellow M (ed), Nucleic acid techniques in bacterial systematics. JohnWiley & Sons, Chichester, United Kingdom.
58. Price MN, Dehal PS, Arkin AP. 2010. FastTree 2—approximatelymaximum-likelihood trees for large alignments. PLoS One 5:e9490. doi:10.1371/journal.pone.0009490.
59. Clarke KR, Gorley RN. 2006. PRIMER v6: user manual/tutorial.PRIMER-E, Plymouth, United Kingdom.
60. DeSantis TZ, Hugenholtz P, Keller K, Brodie EL, Larsen N, Piceno YM,Phan R, Andersen GL. 2006. NAST: a multiple sequence alignment serverfor comparative analysis of 16S rRNA genes. Nucleic Acids Res. 34:W394 –W399.
61. Ludwig W, Strunk O, Westram R, Richter L, Meier H, Kumar Y,Buchner A, Lai T, Steppi S, Jobb G, Förster W, Brettske I, Gerber S,Ginhart AW, Gross O, Grumann S, Hermann S, Jost R, König A, LissT, Lüssmann R, May M, Nonhoff B, Reichel B, Strehlow R, StamatakisA, Stuckmann N, Vilbig A, Lenke M, Ludwig T, Bode A, Schleifer KH.2004. ARB: a software environment for sequence data. Nucleic Acids Res.32:1363–1371.
62. Stamatakis A, Hoover P, Rougemont J. 2008. A rapid bootstrap algo-rithm for the RAxML Web servers. Syst. Biol. 57:758 –771.
63. Bik EM, Long CD, Armitage GC, Loomer P, Emerson J, Mongodin EF,Nelson KE, Gill SR, Fraser-Liggett CM, Relman DA. 2010. Bacterialdiversity in the oral cavity of 10 healthy individuals. ISME J. 4:962–974.
64. Koren O, Goodrich JK, Cullender TC, Spor A, Laitinen K, Bäckhed HK,Gonzalez A, Werner JJ, Angenent LT, Knight R, Bäckhed F, Isolauri E,Salminen S, Ley RE. 2012. Host remodeling of the gut microbiome andmetabolic changes during pregnancy. Cell 150:470 – 480.
Costello et al.
12 ® mbio.asm.org November/December 2013 Volume 4 Issue 6 e00782-13
on July 17, 2018 by guesthttp://m
bio.asm.org/
Dow
nloaded from


















