
Microbial Oxidation of Gaseous Hydrocarbons: Epoxidation of C2 to
8
APPLIED AND ENVIRONMENTAL MICROBIOLOGY, July 1979, p. 127-134 Vol. 38, No. 1 0099-2240/79/07-0127/08$02.00/0 Microbial Oxidation of Gaseous Hydrocarbons: Epoxidation of C2 to C4 n-Alkenes by Methylotrophic Bacteria CHING T. HOU,* RAMESH PATEL, ALLEN I. LASKIN, AND NANCY BARNABE Corporate Pioneering Research Laboratory, Exxon Research and Engineering Company, Linden, New Jersey 07036 Received for publication 23 April 1979 Over 20 new cultures of methane-utilizing microbes, including obligate (types I and II) and facultative methylotrophic bacteria were isolated. In addition to their ability to oxidize methane to methanol, resting cell-suspensions of three distinct types of methane-grown bacteria (Methylosinus trichosporium OB3b [type II, obligate]; Methylococcus capsulatus CRL Ml NRRL B-11219 [type I, obligate]; and Methylobacterium organophilum CRL-26 NRRL B-11222 [facul- tative]) oxidize C2 to C4 n-alkenes to their corresponding 1,2-epoxides. The product 1,2-epoxides are not further metabolized and accumulate extracellularly. Methanol-grown cells do not have either the epoxidation or the hydroxylation activities. Among the substrate gaseous alkenes, propylene is oxidized at the highest rate. Methane inhibits the epoxidation of propylene. The stoichiometry of the consumption of propylene and oxygen and the production of propylene oxide is 1:1:1. The optimal conditions for in vivo epoxidation are described. Results from inhibition studies indicate that the same monooxygenase system catalyzes both the hydroxylation and the epoxidation reactions. Both the hydrox- ylation and epoxidation activities are located in the cell-free particulate fraction precipitated between 10,000 and 40,000 x g centrifugation. On the basis of 180 incorporation from 1802 into the cellular constituents of Pseudomonas methanica, Leadbetter and Foster (9) suggested that the initial oxidative attack on methane in- volves an oxygenase. Higgins and Quayle (8) isolated CH318OH as the product of methane oxidation when suspensions of P. methanica or Methanomonas methanooxidans were allowed to oxidize methane in "802-enriched atmos- pheres. The subsequent observation of methane- stimulated reduced nicotinamide adenine dinu- cleotide (NADH) oxidation catalyzed by ex- tracts of Methylococcus capsulatus (14, 15) or Methylomonas capsulatus (6) suggested that the enzyme responsible for this oxygenation is a monooxygenase. These workers relied on indi- rect enzyme assays, measuring methane-stimu- lated NADH disappearance spectrophotometri- cally or methane-stimulated 02 disappearance polarographically. Recently, methane monoox- ygenase systems were partially purified from Methylosinus trichosporium OB3b (16, 17) and Methylococcus capsulatus (Bath) (3, 4). The epoxidation of 1-alkenes was first dem- onstrated by Van der Linden (18), who detected the formation of 1,2-epoxyoctane from 1-octene by heptane-grown resting cells ofP. aeruginosa. Cardini and Jurtshuk (2) found that a cell ex- tract of a Corynebacterium sp. oxidized I-octene to 1,2-epoxyoctane in addition to hydroxylating octane to octanol. Coon and his co-workers (5) isolated an enzyme system from P. oleovorans that catalyzed the hydroxylation of alkanes and fatty acids. We have also demonstrated the ex- poxidation of 1-octene by whole cells and a pu- rified monooxygenase system of P. oleovorans grown on n-octane (1, 10, 11). The epoxidation system of the octane-grown P. oleovorans was found not to be active on gaseous alkenes. In the course of our studies of gaseous hydro- carbon-utilizing microbes, we discovered that resting cell suspensions of methylotrophic bac- teria oxidize gaseous 1-alkenes to their corre- sponding 1,2-epoxyalkanes. Independently from our studies, Colby et al. reported the ability of a nonspecific, soluble methane monooxygenase from Methylococcus capsulatus (Bath) to oxy- genate n-alkanes, n-alkenes, ethers, and ali- cyclic, aromatic, and heterocyclic compounds (4). The present paper deals with the epoxida- tion of gaseous alkenes by resting cell suspen- sions of representatives of three distinct groups of methylotrophic bacteria. These are: M. trich- osporium OB3b (an obligate methylotroph with type II membrane structure); Methylococcus capsulatus CRL Ml (an obligate methylotroph with type I membrane structure); and Methylo- bacterium organophilum CRL 26 (a facultative 127 Downloaded from https://journals.asm.org/journal/aem on 30 January 2022 by 70.177.157.166.
Transcript of Microbial Oxidation of Gaseous Hydrocarbons: Epoxidation of C2 to

APPLIED AND ENVIRONMENTAL MICROBIOLOGY, July 1979, p. 127-134 Vol. 38, No. 10099-2240/79/07-0127/08$02.00/0
Microbial Oxidation of Gaseous Hydrocarbons: Epoxidation ofC2 to C4 n-Alkenes by Methylotrophic Bacteria
CHING T. HOU,* RAMESH PATEL, ALLEN I. LASKIN, AND NANCY BARNABE
Corporate Pioneering Research Laboratory, Exxon Research and Engineering Company, Linden, NewJersey 07036
Received for publication 23 April 1979
Over 20 new cultures of methane-utilizing microbes, including obligate (typesI and II) and facultative methylotrophic bacteria were isolated. In addition totheir ability to oxidize methane to methanol, resting cell-suspensions of threedistinct types of methane-grown bacteria (Methylosinus trichosporium OB3b[type II, obligate]; Methylococcus capsulatus CRL Ml NRRL B-11219 [type I,obligate]; and Methylobacterium organophilum CRL-26 NRRL B-11222 [facul-tative]) oxidize C2 to C4 n-alkenes to their corresponding 1,2-epoxides. Theproduct 1,2-epoxides are not further metabolized and accumulate extracellularly.Methanol-grown cells do not have either the epoxidation or the hydroxylationactivities. Among the substrate gaseous alkenes, propylene is oxidized at thehighest rate. Methane inhibits the epoxidation of propylene. The stoichiometryof the consumption of propylene and oxygen and the production of propyleneoxide is 1:1:1. The optimal conditions for in vivo epoxidation are described.Results from inhibition studies indicate that the same monooxygenase systemcatalyzes both the hydroxylation and the epoxidation reactions. Both the hydrox-ylation and epoxidation activities are located in the cell-free particulate fractionprecipitated between 10,000 and 40,000 x g centrifugation.
On the basis of 180 incorporation from 1802into the cellular constituents of Pseudomonasmethanica, Leadbetter and Foster (9) suggestedthat the initial oxidative attack on methane in-volves an oxygenase. Higgins and Quayle (8)isolated CH318OH as the product of methaneoxidation when suspensions of P. methanica orMethanomonas methanooxidans were allowedto oxidize methane in "802-enriched atmos-pheres. The subsequent observation of methane-stimulated reduced nicotinamide adenine dinu-cleotide (NADH) oxidation catalyzed by ex-tracts of Methylococcus capsulatus (14, 15) orMethylomonas capsulatus (6) suggested thatthe enzyme responsible for this oxygenation is amonooxygenase. These workers relied on indi-rect enzyme assays, measuring methane-stimu-lated NADH disappearance spectrophotometri-cally or methane-stimulated 02 disappearancepolarographically. Recently, methane monoox-ygenase systems were partially purified fromMethylosinus trichosporium OB3b (16, 17) andMethylococcus capsulatus (Bath) (3, 4).The epoxidation of 1-alkenes was first dem-
onstrated by Van der Linden (18), who detectedthe formation of 1,2-epoxyoctane from 1-octeneby heptane-grown resting cells of P. aeruginosa.Cardini and Jurtshuk (2) found that a cell ex-tract of a Corynebacterium sp. oxidized I-octene
to 1,2-epoxyoctane in addition to hydroxylatingoctane to octanol. Coon and his co-workers (5)isolated an enzyme system from P. oleovoransthat catalyzed the hydroxylation of alkanes andfatty acids. We have also demonstrated the ex-poxidation of 1-octene by whole cells and a pu-rified monooxygenase system of P. oleovoransgrown on n-octane (1, 10, 11). The epoxidationsystem of the octane-grown P. oleovorans wasfound not to be active on gaseous alkenes.
In the course of our studies of gaseous hydro-carbon-utilizing microbes, we discovered thatresting cell suspensions of methylotrophic bac-teria oxidize gaseous 1-alkenes to their corre-sponding 1,2-epoxyalkanes. Independently fromour studies, Colby et al. reported the ability ofa nonspecific, soluble methane monooxygenasefrom Methylococcus capsulatus (Bath) to oxy-genate n-alkanes, n-alkenes, ethers, and ali-cyclic, aromatic, and heterocyclic compounds(4). The present paper deals with the epoxida-tion of gaseous alkenes by resting cell suspen-sions of representatives of three distinct groupsof methylotrophic bacteria. These are: M. trich-osporium OB3b (an obligate methylotroph withtype II membrane structure); Methylococcuscapsulatus CRL Ml (an obligate methylotrophwith type I membrane structure); and Methylo-bacterium organophilum CRL 26 (a facultative
127
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 30
Jan
uary
202
2 by
70.
177.
157.
166.
128 HOU ET AL.
methylotroph). Because of the instability of themonooxygenase system, the optimal conditionsfor the epoxidation were studied in cell suspen-sions of methylotrophs with particular referenceto the epoxidation of propylene oxide. Subse-quent studies revealed that the activity for boththe epoxidation of propylene and the hydroxyl-ation of methane was associated with the cell-free particulate fraction in the methylotrophstested. Evidence obtained indicated that a singleenzyme system catalyzed both the hydroxyl-ation of methane and the epoxidation of gaseous1-alkenes.
MATERIALS AND METHODSBacterial strains. Methane-utilizing cultures were
isolated from lake water of Warinanco Park, Linden,N.J., and lake and soil samples of Bayway Refinery,Linden, N.J. (Table 1). These methylotrophs wereidentified by the classification given by Whittenburyet al. (19) and Patt et al. (13). M. trichosporium OB3b,Methylosinus sporium 5, Methylocystisparvus OBBP,Methylomonas methanica Si, Methylomonas albusBG 8, and Methylobacter capsulatus Y were kindlyprovided by R. Wittenbury (School of Biological Sci-ences, University of Warwick, Coventry, the UnitedKindgom). M. organophilum XX ATCC 24886 waskindly provided by R. S. Hanson (University of Wis-consin, Madison). The organisms were maintained onmineral salt plates in a desiccator jar under an atmos-phere of methane and air (1:1, vol/vol) at 30°C. Or-ganisms were grown at 30°C in 300-ml flasks contain-ing 50 ml of mineral salt medium (7) with methaneand air (1:1, vol/vol) as the sole carbon and energysource. When methanol (at 0.3%) was used, the gas-eous phase in the flask was air.
Chemicals. Gaseous hydrocarbons (methane, eth-ylene, propylene, 1-butene, and butadiene) and ethyl-ene oxide were obtained from Matheson Gas Products(East Rutherford, N.J.). 1,2-Epoxypropane, 1,2-epox-ybutane, 1,2-epoxybutene, and 1-pentene were pur-chased from Matheson, Coleman, and Bell, Norwood,Ohio.
Preparation of cell-free particulate fraction.Organisms were harvested by centrifugation (5,000 xg, 30 min, 4C) and washed twice on 0.05 M sodiumphosphate buffer (pH 7.0) containing 5 mM MgC12.Cells were suspended in the same buffer and disruptedby passing twice through a French pressure cell(American Instrument Co., Silver Spring, Md.). Celldebris and unbroken organisms were removed by cen-trifugation (10,000 x g, 30 min, 4C). The resultingextract was further centrifuged (40,000 x g, 90 min,4°C), yielding particulate and supernatant fractions.The particulate material was resuspended in 0.05 Msodium phosphate buffer (pH 7.0) containing 5 mMMgCl2.
Activity assay. When whole cells were used, theharvested cells from shake-flask cultures were washedtwice with 0.05 M phosphate buffer (pH 7.0). The finalpellet was resuspended in fresh buffer to obtain anoptical density at 660 nm of 0.5. A 0.5-mil amount ofthis washed cell suspension (containing a given
APPL. ENVIRON. MICROBIOL.
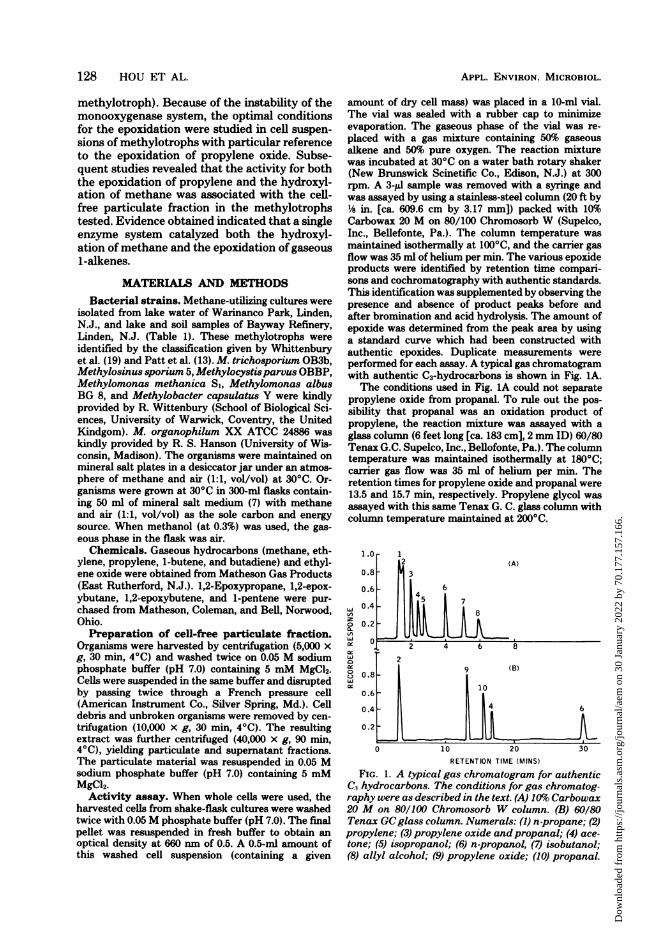
amount of dry cell mass) was placed in a 10-ml vial.The vial was sealed with a rubber cap to minimizeevaporation. The gaseous phase of the vial was re-placed with a gas mixture containing 50% gaseousalkene and 50% pure oxygen. The reaction mixturewas incubated at 30°C on a water bath rotary shaker(New Brunswick Scinetific Co., Edison, N.J.) at 300rpm. A 3-pl sample was removed with a syringe andwas assayed by using a stainless-steel column (20 ft by'h in. [ca. 609.6 cm by 3.17 mm]) packed with 10%Carbowax 20 M on 80/100 Chromosorb W (Supelco,Inc., Bellefonte, Pa.). The column temperature wasmaintained isothermally at 100°C, and the carrier gasflow was 35 ml of helium per min. The various epoxideproducts were identified by retention time compari-sons and cochromatography with authentic standards.This identification was supplemented by observing thepresence and absence of product peaks before andafter bromination and acid hydrolysis. The amount ofepoxide was determined from the peak area by usinga standard curve which had been constructed withauthentic epoxides. Duplicate measurements wereperformed for each assay. A typical gas chromatogramwith authentic C3-hydrocarbons is shown in Fig. 1A.The conditions used in Fig. 1A could not separate
propylene oxide from propanal. To rule out the pos-sibility that propanal was an oxidation product ofpropylene, the reaction mixture was assayed with aglass column (6 feet long [ca. 183 cm], 2 mm ID) 60/80Tenax G.C. Supelco, Inc., Bellofonte, Pa.). The columntemperature was maintained isothermally at 180°C;carrier gas flow was 35 ml of helium per min. Theretention times for propylene oxide and propanal were13.5 and 15.7 min, respectively. Propylene glycol wasassayed with this same Tenax G. C. glass column withcolumn temperature maintained at 200°C.
1.0 12 (A)
0.8 3
0.6-6
LI0.4 45 8
o 0.2-
CZ 2 4 6 8°'6Xb.a 2°x |9 (B)*u 0.4 (
0 10 20 30RETENTION TIME (MINS)
FIG. 1. A typical gas chromatogram for authenticC, hydrocarbons. The conditions for gas chromatog-raphy were as described in the text. (A) 10%o Carbowax20 M on 80/100 Chromosorb W column. (B) 60/80Tenax GCglass column. Numerals: (1) n-propane; (2)propylene; (3) propylene oxide andpropanal; (4) ace-tone; (5) isopropanol; (6) n-propanol, (7) isobutanol;(8) allyl alcohol; (9) propylene oxide; (10) propanal.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 30
Jan
uary
202
2 by
70.
177.
157.
166.

EPOXIDATION OF GASEOUS 1-ALKENES 129
For the cell-free system, the methane hydroxylationand propylene epoxidation activities were measuredby determining methanol and propylene oxide forma-tion by gas-liquid chromatography (GLC). A 0.2-mlportion of the particulate fraction (1 mg of protein)was placed in a 5-ml vial. The vial was sealed with arubber cap to mininiize evaporation. The gaseousphase of the vial was replaced with a gas mixturecontaining 50% methane or propylene and 50% pureoxygen. In the case of methane hydroxylation, thebuffer concentration ofthe reaction mixture was raisedto 0.15 M sodium phosphate to inhibit further oxida-tion of methanol (17). The reaction mixture was in-cubated at 30°C on a water bath rotary shaker asdescribed above. A 3-pl sample was removed with asyringe and was assayed by GLC.
RESULTSResting cell suspensions of methane-grown
cells ofM. trichosporium OB3b, Methylococcuscus capsulatus CRL Ml, and M. organophilumCRL 26 oxidized propylene to 1,2-epoxypropane,which accumulated. Methanol-grown cellsshowed no activity for either epoxidation of pro-pyelene or hydroxylation of methane. After thereaction, the cell suspensions were centrifugedto remove the cells. The product propylene oxidewas found totally in the supernatant fraction inall the three strains tested, i.e., the productepoxide accumulated extracellularly. No prod-uct peak other than propylene oxide from pro-pylene was detected. The possibility of propanalas an oxidation product of propylene was ruledout on the basis of GLC analysis (Fig. 1B). Acomparison of the activity for epoxidation ofpropylene by methane-grown microbes is listedin Table 1. Epoxidation activity was found in allof the strains tested.The optimal conditions for the production of
propylene oxide were compared among threedistinct types of methylotrophic bacteria.Time course ofepoxide formation. Control
experiments with heat-killed cells indicated thatthe epoxide was produced enzymatically. Withthe standard assay system, the propylene oxideconcentration reached a maximum after 3 h ofincubation (Fig. 2). The rate of propylene oxideproduction was linear for the first 120 min forstrains CRL Ml and OB3b and for the first 60min for strain CRL 26. Therefore, epoxide pro-duction was mei,sured within 1 h whenever theeffect of a variable was tested.The possibilities that the slower reaction rate
after 1 to 2 h of incubation was due to enzymicor nonenzymic degradation of propylene oxideor due to product inhibition was examined. A4.8-,lmol amount of propylene oxide was addedto viable or heated-killed cell suspensions in thepresence or absence of substrate (propylene) andincubated under standard assay conditions. No
TABLE 1. Epoxidation ofpropylene to propyleneoxide by methane-grown microbes'
Epoxidation
Methylotrophs rate (Mmol/hper mg of pro-
tein)
Obligate, type IIMethylosinus sp. CRL 15Methylosinus tricho- OB3bsporium
Methylosinus sp. CRL 16Methylosinus spor- 5ium
Methylocystis sp. CRL 18Methylocystis parvus OBBP
Obligate, type IMethylomonas sp. CRL 4Methylomonas sp. CRL 21Methylomonas meth- Si
anicaMethylomonas sp. CRL 8Methylomonas albus BG 8Methylomonas sp. CRL 17Methylomonas sp. CRL 22Methylomonas sp. CRL M6PMethylomonas Sp. CRL 20Methylomonas sp. CRL 10Methylobacter sp. CRL M6Methylobacter sp. CRL 23Methylobacter sp. CRL M1YMethylobacter sp. CRL 19Methylobacter sp. CRL 5Methylococcus capsu- CRL MI
latusMethylococcus capsu- Y
latusMethylococcus sp. CRL 25Methylococcus sp. CRL 24
FacultativeMethylobacterium or- CRL 26ganophilum
Methylobacterium or- XXganophilum
2.21.8
1.61.0
0.70.8
1.01.21.3
0.60.70.50.80.51.00.70.61.20.71.10.82.5
0.7
0.60.9
1.3
0.9
" The reactions were conducted as described in thetext. Strains with CRL numbers are newly isolatedcultures in our lab.
disappearance of propylene oxide or formationof propylene glycol was detected during 6 h ofincubation in either viable or heat-killed cellsuspensions. However, after a prolonged incu-bation (24 h), a slight disappearance of propyl-ene oxide and the formation of propylene glycolwas observed. The amount of the further oxi-dation of propylene oxide was identical in bothviable and heat-killed cell suspensions. Datashown in Fig. 3 indicate that there was no en-zymic degradation of propylene oxide, and thenonenzymic degradation of propylene oxide wasnegligible under our assay conditions. The viablecells incubated with both propylene and propyl-ene oxide showed further production of propyl-
VOL. 38, 1979
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 30
Jan
uary
202
2 by
70.
177.
157.
166.

APPL. ENVIRON. MICROBIOL.
6 -
Cl)
)(<
0~ZU~ ,, *
0 20 40 60 120 180 240TIME (MINS)
FIG. 2. Time course ofpropylene oxide productionby resting cell suspensions of methylotrophic bacte-ria. The reactions were conducted as described in thetext. Propylene oxide was estimated by gas chroma-tography. Three milligrams of cells (dry weight) wasused: Methylosinus trichosporium OB3b (0); Methy-lococcus capsulatus CRL MI (O); Methylobacteriumorganophilum CRL-26 (A).
8
> 6
0-
2X <:O _j
0o/-0k'~~~~~~~
0 2 4 6TIME (HRS)
FIG. 3. Degradation ofpropylene oxide by restingcell suspensions ofMethylosinus capsulatus CRL Ml.A 4.8-ttmol amount ofpropylene oxide was added toeach reaction mixture before incubation. Viable cellsuspension plus propylene oxide (0); heat-killed cellsuspension plus propylene oxide (A); viable cell sus-pension pluspropylene oxide andpropylene (0). Fourmilligrams of cells was used in each experiment.
ene oxide. However, the rate of epoxidation wasslower than in the standard assay system inwhich no external propylene oxide was added.Therefore, product inhibition might be countedas one of the reasons for the slower reaction rateafter 2 h of incubation.Optimum conditions for the production
of propylene oxide. (i) pH. The effect of pHon propylene oxide production by resting cellsof the three strains of methylotrophs was ex-amined. Sodium phosphate buffer (0.05 M) wasused for pH values from 5.5 to 8.0, andtris(hydroxymethyl)aminomethane buffer (0.05M) was used for values from 8.0 to 10.0. A pHbetween 6.0 and 7.0 appeared to be optimum forepoxide production for all three of the methy-lotrophs (Fig. 4). The initial and fmal pH read-ings in these experiments differed by less than0.5 pH unit, and similar amounts of epoxide were
produced in either buffer at pH 8.0. Authenticsamples of propylene oxide (final concentration,4 ltmol/ml) were added to heat-killed cell sus-pensions of Methylococcus capsulatus CRL Mlat pH 5.5, 7.0, and 10.0 to test for nonenzymaticdegradation of propylene oxide at these pH val-ues. The propylene oxide concentration in thesesuspensions did not decrease during 3 h of in-cubation, indicating that nonenzymatic oxida-tion or hydrolysis of propylene oxide was negli-gible under these assay conditions.
(ii) Temperature. The temperature opti-mum for the epoxidation of propylene by restingcell-suspensions of the three strains was about35°C (Fig. 5). Strain OB3b showed a broadertemperature optimum. At 40°C, there was anapparent decrease in the amount of epoxideaccumulated. This is possibly due to both theinstability of the monooxygenase system athigher temperature and the volatility of theproduct propylene oxide (bp 35°C).
(iii) Propylene concentration. Various con-centrations of propylene were used to examinethe production of propylene oxide by resting cellsuspensions of Methylococcus capsulatus CRLMl. The initial oxygen partial pressure in thegaseous phase was kept constant (50%, vol/vol).Helium gas was used to balance the rest of thegaseous phase. The amount of propylene oxideproduced was assayed after 30 min of incubation.A propylene concentration, in the gaseous phase,of ca. 15% (66 ,umol) supported maximum pro-pylene oxide production (Fig. 6). Higher propyl-ene concentration did not stimulate nor inhibitthe production of propylene oxide. The reactionrate appears to be dependent upon the solubilityof both substrates, propylene and oxygen.
x 4
Z e
2z .C
5.0 6.0 7.0 8.0 9.0 10.0
pH
FIG. 4. Effect ofpH on the epoxidation ofpropyl-ene by cell suspensions of methylotrophic bacteria.Reactions were conducted as described in the text.Propylene oxide was estimated after 1 h of incuba-tion. Sodium phosphate buffer (0.05 M) was used forpH values 5.0 to 8.0. Tris(hydroxymethyl)amino-methane-hydrochloride buffer (0.05 M) was used forvalues 8.0 to 10.0 (6mg ofcellsper vial). Methylosinustrichosporium OB3b (0); Methylococcus capsulatusCRL Ml (U); Methylobacterium organophilum CRL26 (A).
130 HOU ET AL.
0-.. 0
.. a4,
A
----711
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 30
Jan
uary
202
2 by
70.
177.
157.
166.

EPOXIDATION OF GASEOUS 1-ALKENES 131
0.51
5 Stoichiometry of propylene oxidation.The stoichiometry of epoxidation of propyleneby a cell suspension of M. trichosporium OB3b
4 was examined. The amount of oxygen consumedduring the reaction was determined polaro-graphically with a Clark oxygen electrode. The
3 - propylene consumed and the propylene oxideformed were estimated by gas chromatography.
2//^ The stoichiometry of consumption of propylene
2 -/ / / and oxygen and the production of propyleneoxide was found to be approximately 1:1:1.
I/ Substrate specificity. The substrate speci-ficity for the epoxidation of 1-alkenes by thethree strains of methylotrophs was determined.
15 20 25 30 35 40 Epoxides were produced from gaseous alkenesTEMPERATURE (°C) only (Table 2). The highest production of epox-
5. Effect oftemperature on the epoxidation Of ide was for propylene in all three strains.!ne by cell suspensions of methylotrophic bac- Inhibition studies. The epoxidation of pro-leactions were conducted as described in the pylene to propylene oxide and the hydroxylationropylene oxide was estimated after I h of of methane to methanol by the three types ofeion (6mg ofcellsper vial). Methylosinus trich- methane-utilizing bacteria were inhibited byCm OB3b (0); Methylococcus capsulatus CRL metal-binding and metal-chelating agents suchMethylobacterium organophilum CRL 26 as potassium cyanide, 1,10-phenanthroline, a,a-
bipyridyl, thiourea, and imidazol (Table 3). Thissuggests the involvement of metal ion(s) in boththe epoxidation of propylene and the hydroxyl-ation of methane.
___ *- s-- The effect of methane on the oxidation ofpropylene to propylene oxide was also studied.The production of propylene oxide from propyl-ene by a cell suspension of M. trichosporiumOB3b was assayed in the presence of a givenamount of methane. The initial partial pressureof propylene in the gaseous phase was keptconstant in all of the experiments. Methane in-hibits the propylene oxidation reaction by the
I)5 110 2'0 3|0 ;H0 whole cell system of strain OB3b (Table 4).PROPYLENE (e/, IN GASEOUS PHASE)
FIG. 6. Effect ofpropylene partial pressure on theproduction ofpropylene oxide by cell suspensions ofmethylotrophic bacteria. Reactions were conductedas described in the text. Oxygen partial pressure waskept constant at 50%, vol/vol, of the gaseous phase.Propylene was diluted with helium to obtain variouspartial pressures ofpropylene. Propylene oxide wasestimated after 30 min ofincubation (3 mg of cellspervial). Methylococcus capsulatus CRL Ml (M.
(iv) Cell concentrations. The cell concen-tration also influences the rate of propylene ox-ide production. The amount of propylene oxideaccumulated after 1 h of incubation increased asthe cell concentration was increased up to about9 mg/ml (Fig. 7). No further increase in propyl-ene oxide production was observed at higher cellconcentrations. This indicated that at cell con-centrations higher than 9 mg/ml, the solubilityof the gaseous substrates becomes the rate-lim-iting factor in the production of propylene oxide.
o-woi <
I0
6
2
0/
0 2 4 6 8 10 12
CELL CONCENTRATION (mg)
FIG. 7. Effect of cell concentration on the produc-tion ofpropylene oxide by cell suspensions of methy-lotrophic bacteria. Reactions were conducted as de-scribed in the text. Propylene oxide was estimatedafter I h of incubation. Methylosinus trichosporiumOB3b (0); Methylococcus capsulatus CRL Ml (U);Methylobacterium organophilum CRL 26 (A).
<0)
z E
woJ °
O -JCL
FIG.propyle,teria. Rtext. Piincubatosporiu,Ml (-)(A).
2 .0
1.01w tn'm Ln
0 -gwz LC!LAJ 0-i 1->- cn0. W0 -jIX C>CL 2M.
14
VOL. 38, 1979
c
°or
.
0
a
0
w
aI
IA
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 30
Jan
uary
202
2 by
70.
177.
157.
166.

TABLE 2. Oxidation ofgaseous alkenes and methane by resting cell suspensions ofmethane-grown bacteriaRate of oxidation (pmol/h per assya)
Propl 1ua-Pen-Strains Meth- Ethyl- eneto; 1-Butene diene to teneStrains ~~~~~~aneto ene to ene
opl to 1,2- 12 o12metha- ethylene P PY- epoxy bu- 2 epoytaneos epoxynol oxide ide tane butene pen-
tane
Obligate, type II membrane structureMethylosinus trichosporium OB3b 1.6 1.9 3.6 0.45 2.5 0
Obligate, type I membrane structureMethylococcus capsulatus CRL Ml NRRL 2.5 5.5 5.5 1.3 4.4 0B11219
FacultativeMethylobacterium organophilum CRL 26 NRRL 0.7 0.9 2.5 0.9 2.8 0
B-11222
aTwo milligrams of cell (dry weight) per assay was used.
TABLE 3. Effect of inhibitors on the epoxidation ofpropylene and the hydroxylation of methane'% Inhibition
Methylosinus tricho- Methylococcus capsu- Methylobacterium or-Inhibitor sporium OB3b latus CRL Ml ganophilum CRL 26
Epoxida- Hydroxyl- Epoxida- Hydroxyl- Epoxida- Hydroxyl-tion ation tion ation tion ation
Thiourea 100 100 100 100 100 1001,10-Phenanthroline 90 92 95 95 90 90a,a-Bipyridyl 100 90 100 100 100 100Imidazole 95 90 95 95 100 100Potassium cyanide 100 100 100 100 95 95
" The reactions were conducted as described in the text. The products were estimated by gas chromatographyafter 1 h of incubation at 30°C. Each inhibitor was added at a final concentration of 1 mM.
Cell-free system. Cells of methane-grownMethylococcus capsulatus CRL Ml were usedfor cell-free studies on the epoxidation of pro-pylene and the hydroxylation of methane. Thecrude extract after 10,000 x g centrifugation ofsonically disrupted cells (3 x 50 s, Wave Energyultrasonic oscillator, model W 201) was found tohave no activity for either epoxidation or hy-droxylation. However, when the cells were dis-rupted by passing twice through a French pres-sure cell (1,000 kg of pressure), both activitieswere found in the crude extract after 10,000 x gcentrifugation. All of the activity in the crudeextract was collected as a particulate fraction byfurther centrifugation of the crude extract at40,000 x g for 90 min at 4°C. NADH stimulatedboth the epoxidation (20%) and the hydroxyl-ation (30%) reactions. Further information onepoxidation of propylene by cell-free systemswill be published (Patel et al., J. Bacteriol., inpress).
DISCUSSIONBoth the system of P. aeruginosa demon-
strated by Van der Linden (18) and the system
TABLE 4. Effect of methane on the epoxidation ofpropylenea
PropyleneCompositioof gaseousphas oxide % Inhibi-Composition of gaseous phase fomd tnformed tion
(,umol)
Propylene + helium + 02 1.6 0(25:25:50, vol/vol)
Propylene + methane + 02 0.8 50(25:25:50, vol/vol)
' Reactions were conducted as described in the textexcept that various gaseous compositions were used tomaintain a constant propylene partial pressure. Cellsuspensions of methane-grown M. trichosporiumOB3b (3.6 mg) were used. Propylene oxide was esti-mated by gas chromatography after 15 min of theincubation.
of P. oleovorans demonstrated in our laboratory(1, 10, 1 1) epoxidized liquid 1-alkenes from C6to C12, but not gaseous alkenes.
In this report, we describe the epoxidation ofethylene, propylene, 1-butene, and butadiene bycell suspensions of all three distinct groups ofmethane-utilizing bacteria. The epoxidation ofalkenes and the hydroxylation of methane were
132 HOU ET AL. APPL. ENVIRON. MICROBIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 30
Jan
uary
202
2 by
70.
177.
157.
166.

EPOXIDATION OF GASEOUS 1-ALKENES 133
not found under anaerobic condition or in meth-anol-grown cells, suggesting that the enzymesystem is inducible. The product 1,2-epoxidesaccumulated extracellularly. The nonenzymicdegradation of propylene oxide in our standardassay system was not significant even after aprolonged incubation time (Fig. 3). Van der Lin-den (18) demonstrated the production of 1,2-epoxyoctane from 1-octene by heptane-growncells of Pseudomonas sp. and also stated thatthe epoxide was not further oxidized enzymati-cally. However, May and Abbott (10, 11) re-ported that when 1-octene was supplied as asubstrate to the w-hydroxylation enzyme systemof P. oleovorans, both 8-hydroxy-1-octene and1,2-epoxyoctane were formed. In addition, Ab-bott and Hou (1) found that the methyl groupof the latter compound was also susceptible tohydroxylation. The present results obtainedfrom the studies of viable cell suspensions of themethane-utilizing bacteria, however, indicatedthat propylene oxide was not further metabo-lized enzymatically (Fig. 3).Epoxide accumulation from 1-octene by P.
aeruginosa was accompanied by the metabolismof a large quantity of 1-octene via methyl groupoxidation (18). In the epoxidation of propyleneby cell suspensions ofmethane-utilizing bacteria,however, no formation of 3-hydroxy propene-1was detected.Both the epoxidation of 1-alkene and the hy-
droxylation of methane were inhibited by var-ious metal-binding and metal-chelating agents,indicating the involvement of metal(s)-contain-ing enzyme system(s). The epoxidation of pro-pylene to propylene oxide by a cell suspensionof methane-grown strain CRL MI was inhibited(50%) in the presence of the hydroxylation sub-strate methane (Table 4).The optimum conditions for the in vivo epox-
idation of propylene by the three distinct groupsof methane-utilizing bacteria are quite similar.The pH optima were around 6 - 7, and thetemperature optimum was around 350C. Theapparent decrease in epoxidation above 40°Cmay be due to both the instability of the mono-oxygenase system and the volatility of the prod-uct propylene oxide (bp, 350C).Both the hydroxylation and epoxidation activ-
ities are located in the cell-free particulate frac-tion precipitated between 10,000 and 40,000 x gcentrifugation. Tonge et al. (16, 17) have re-ported the purification of a membrane-boundmethane monooxygenase from the particulatefraction (precipitated between 10,000 and150,000 x g centrifugation) of M. trichosporiumOB3b. Recently, Colby et al. (4) demonstrated aunique soluble methane monooxygenase from
Methylococcus capsulatus (Bath strain) whichcatalyzes the oxidation of n-alkanes, n-alkenes,ethers, and alicyclic, aromatic, and heterocycliccompounds. The strains from the three distinctgroups of methane-utilizing bacteria that wehave examined all catalyze the epoxidation ofgaseous alkenes (C2 to C4) and the hydroxylationof gaseous alkanes (Cl to C4). In contrast toMethylococcus capsulatus (Bath), these meth-ylotrophs did not oxidize liquid alkanes and al-kenes.
ACKNOWLEDGMENTSWe thank Andre Felix and I. Marczak for their excellent
technical assistance.
LITERATURE CITED1. Abbott, B. J., and C. T. Hou. 1973. Oxidation of 1-
alkenes to 1,2-epoxyalkanes by Pseudomonas oleovor-ans. Appl. Microbiol. 26:86-91.
2. Cardini, G., and P. Jurtshuk. 1970. The enzymatichydroxylation of n-octane by Corynebacterium sp.strain 7E1C. J. Biol. Chem. 245:2789-2796.
3. Colby, J., and H. Dalton. 1978. Resolution of the meth-ane mono-oxygenase of Methylococcus capsulatus(Bath) into three components. Biochem. J. 171:461-468.
4. Colby, J., D. I. Stirling, and H. Dalton. 1977. Thesoluble methane mono-oxygenase of Methylococcuscapsulatus (Bath). Its ability to oxygenate n-alkanes, n-alkenes, ethers, and alicyclic, aromatic and hetero-cycliccompounds. Biochem. J. 165:395-402.
5. Coon, M. J., H. W. Strabel, A. P. Autor, J. Heidema,and W. Duppel. 1972. In G. S. Boyd and R. M. Smellie(ed.), Biological hydroxylation mechanisms, p. 45. Ac-ademic Press Inc., New York.
6. Ferenci, T. 1974. Carbon monoxide-stimulated respira-tion in methane-utilizing bacteria. FEBS Lett. 41:94-98.
7. Foster, J. W., and R. H. Davis. 1966. A methane-de-pendent coccus, with notes on classification and nomen-clature of obligate, methane-utilizing bacteria. J. Bac-teriol. 91:1924-1931.
8. Higgins, I. J., and J. R. Quayle. 1970. Oxygenation ofmethane by methane-grown Pseudomonas methanicaand Methanomonas methanooxidans. Biochem. J. 118:201-208.
9. Leadbetter, E. R. L., and J. W. Foster. 1959. Incorpo-ration of molecular oxygen in bacterial cells utilizinghydrocarbons for growth. Nature (London) 184:1428-1429.
10. May, S. W., and B. J. Abbott. 1972. Enzymatic epoxi-dation. I. Alkene epoxidation by the w-hydroxylationsystem ofPseudomonas oleovorans. Biochem. Biophys.Res. Commun. 48:1230-1234.
11. May, S. W., and B. J. Abbott. 1973. Enzymatic epoxi-dation. II. Comparison between the epoxidation andhydroxylation reactions catalyzed by the w-hydroxyl-ation system of Pseudomonas oleovorans. J. Biol.Chem. 248:1725-1730.
12. Patel, R., C. T. Hou, and A. Felix. 1976. Inhibition ofdimethyl ether and methane oxidation in Methylococ-cus capsulatus and Methylosinus trichosporium. J.Bacteriol. 126:1017-1019.
13. Patt, T. E., G. C. Cole, and R. S. Hanson. 1976. Meth-ylobacterium, a new genus of facultatively methylo-trophic bacteria. Int. J. Syst. Bacteriol. 26:226-229.
14. Ribbons, D. W. 1975. Oxidation of C, compounds by
VOL. 38, 1979
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 30
Jan
uary
202
2 by
70.
177.
157.
166.

134 HOU ET AL. APPL. ENVIRON. MICROBIOL.
particulate fractions from Methylococcus capsulatus:distribution and properties of methane-dependent re-duced nicotinamide adenine dinucleotide oxidase(methane hydroxylase). J. Bacteriol. 122:1351-1363.
15. Ribbons, D. W., and J. L Michalover. 1970. Methaneoxidation by cell-free extracts of Methylococcus capsu-latus. FEBS Lett. 11:41-44.
16. Tonge, G. M., D. E. F. Harrison, and I. J. Higgins.1977. Purification and properties of the methane mono-oxygenase enzyme system from Methylosinus tricho-sporium OB3b. Biochem. J. 161:333-344.
17. Tonge, G. M., D. E. F. Harrison, C. J. Knowles, andI. J. Higgins. 1975. Properties and partial purificationof the methane-oxidizing enzyme system from Methy-losinus trichosporium. FEBS Lett. 58:293-299.
18. Van der Linden, A. C. 1963. Epoxidation of a-olefins byheptane-grown Pseudomonas cells. Biochem. Biophys.Acta 77:157-159.
19. Whittenbury, R., K. C. Phillips, and J. F. Wilkinson.1970. Enrichment, isolation and some properties ofmethane-utilizing bacteria. J. Gen. Microbiol. 61:205-218.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 30
Jan
uary
202
2 by
70.
177.
157.
166.


















