
Metabolic pathway alternation, regulation and control (4)—Fatty acids Xiaochen Yu 01-31-2013.
37
Metabolic pathway alternation, regulation and control (4)—Fatty acids Xiaochen Yu 01-31-2013
-
Upload
primrose-clark -
Category
Documents
-
view
217 -
download
1
Transcript of Metabolic pathway alternation, regulation and control (4)—Fatty acids Xiaochen Yu 01-31-2013.

Metabolic pathway alternation, regulation and control (4)—Fatty acids
Xiaochen Yu01-31-2013

Fatty acids in Microorganism
• Highest energy content of the major nutrients (over 9 kcal/g)
• Crude fat is a mixture of triacylglycerols (TAG), phospholipids, fatty acids, sterols, waxes and pigments.
• TAG: the major storage form of fat in most of microorganisms
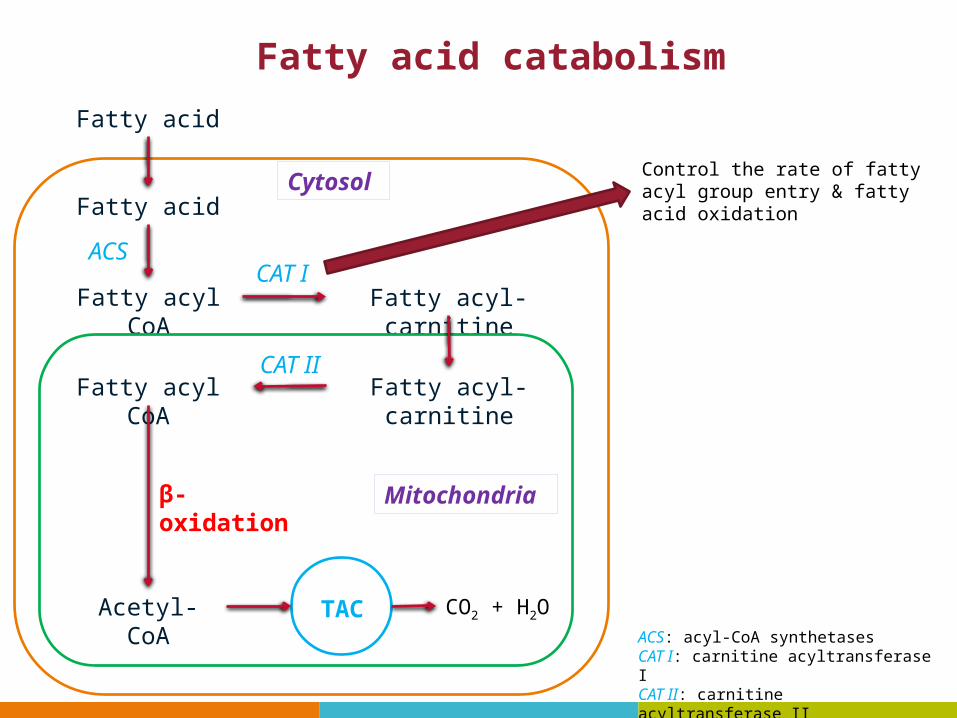
Fatty acid catabolism
Fatty acid
Fatty acid
Fatty acyl CoA Fatty acyl-carnitine
Fatty acyl-carnitineFatty acyl CoA
Acetyl-CoA
ACSCAT I
CAT II
β-oxidation
TAC CO2 + H2O
Control the rate of fatty acyl group entry & fatty acid oxidationCytosol
Mitochondria
ACS: acyl-CoA synthetasesCAT I: carnitine acyltransferase ICAT II: carnitine acyltransferase II

β oxidation
• Oxidation of saturated fatty acids with an even # of carbons
R–CH2–CH2–CH2–C–S–CoA
O
R–CH2–CH=C–C–S–CoA
O
H
H
R–CH2–CH–CH2–C–S–CoA
O
OH
H
R–CH2–C–CH2–C–S–CoA
OO
R–CH2–C–S–CoA
O
CH3–C–S–CoA
O
FAD
FADH2
H2O
NAD+ NADH + H+
+
CoA-SH
Palmitoyl-CoA
(C14) Acyl-CoA Acetyl-CoA
Acyl-CoA dehydrogenase
Enoyl-CoA hydratase
3-hydroxyacyl-CoA dehydrogenase
Acyl-CoA acetyltransferase
1st dehydrogenation
Hydration
2nd dehydrogenation

Fatty acid biosynthesis
ONLY exist in oleaginous
microorganism

Fatty acid biosynthesis

Nitrogen limitation
(Papanikolaou and Aggelis, 2011)
AMP

Fatty acid profiles
Docosahexaenoic acid (C22:6 n-3): Crypthecodinium cohnii & Schizochytrium sp.Gamma linoleic acid (C18:3 n-6): Mucor sp.

Metabolic pathway of xylose to fatty acid
Phosphoketolase pathwayPentose phosphate pathway (PPP)
(Tanaka et al., 2002)

Metabolic pathway of glycerol & Acetate
to fatty acid
Acetate
Acetyl-CoA synthetaseATP
ADPAcetyl-CoA

Lipid yield from carbon sources
100 g of glucose (~0.56 moles)~~1.1 moles of acetyl-CoA-------around 0.32 g lipids/g
100 g of xylose (~0.66 moles)-------around 0.34 g lipids/g
Assuming xylose is assimilated for the phosphoketolase pathway (the most efficient) in oleaginous microorganisms.
1. 1.2 moles of acetyl-CoA (phosphoketolase pathway) or2. 1.0 moles of acetyl-CoA (PPP)
100 g of glycerol--------0.30 g lipids/g
(Papanikolaou and Aggelis, 2011)

12
CodeD-Glucose:initCobraToolboxmodel=readCbModel('iNL895');model1=changeRxnBounds(model,'r_51_exchange',10,'u')solution1=optimizeCbModel(model1)
D-Xylosemodel1=changeRxnBounds(model,'r_51_exchange',0,'u')model2=changeRxnBounds(model1,'r_54_exchange',10,'u')
Acetate:model2=changeRxnBounds(model1,'r_30_exchange',10,'u’)
Glycerol:model2=changeRxnBounds(model1,'r_71_exchange',10,'u')

Example 1 Growth rate optimization
• Task Use FBA to predict flux distribution for optimal growth by yeast Yarrowia lipolytica model iNL895 using different carbon sources.
13
Uptake rate (10 mmol gDW-1 h-1) Biomass flux (h-1) Aerobic
D-Glucose 3.8879
D-Xylose 3.2862
Acetate 0.7432
Glycerol 2.0828
The maximal theoretical growth rate under different carbon sources
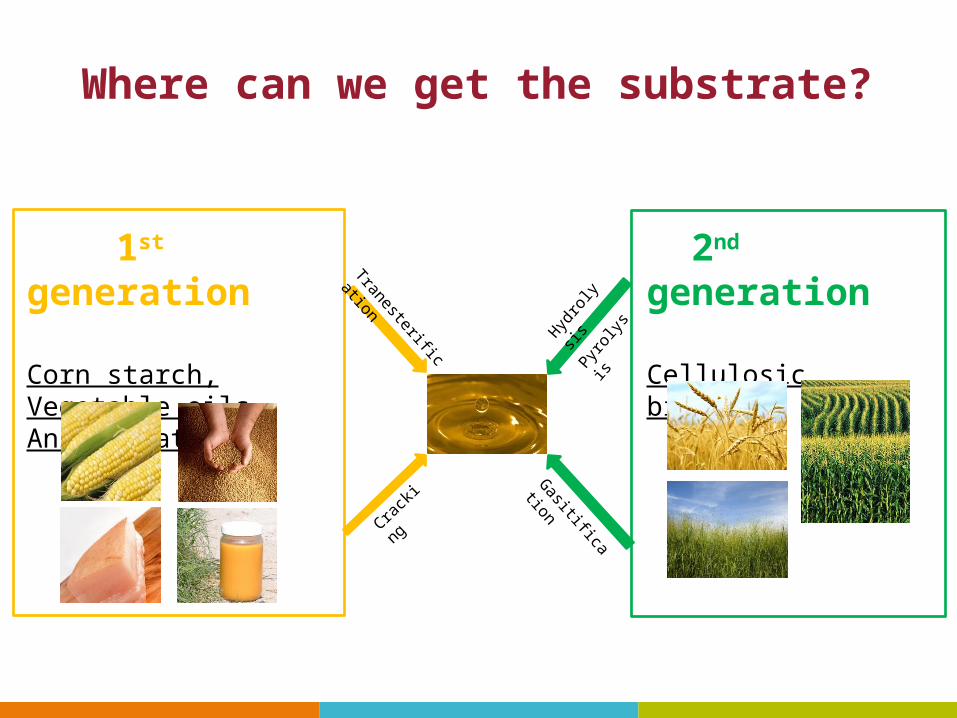
2nd generation
Cellulosic biomassHydro
lysis
Pyrol
ysis
Gasitification
1st generation
Corn starch, Vegetable oils, Animal fats
TranesterificationCra
cking
Where can we get the substrate?


Our choice: Renewable Lipid-based Biofuel Production Strategy
Lignocellulosic biomass
Sugars
Pretreatment & hydrolysis
Fermentation
Characteristics of oleaginous microorganisms applied Similar fatty acid compositions with those of vegetable oils
A wide range of sugars can be utilized
High productivity
Tolerance to some inhibitors
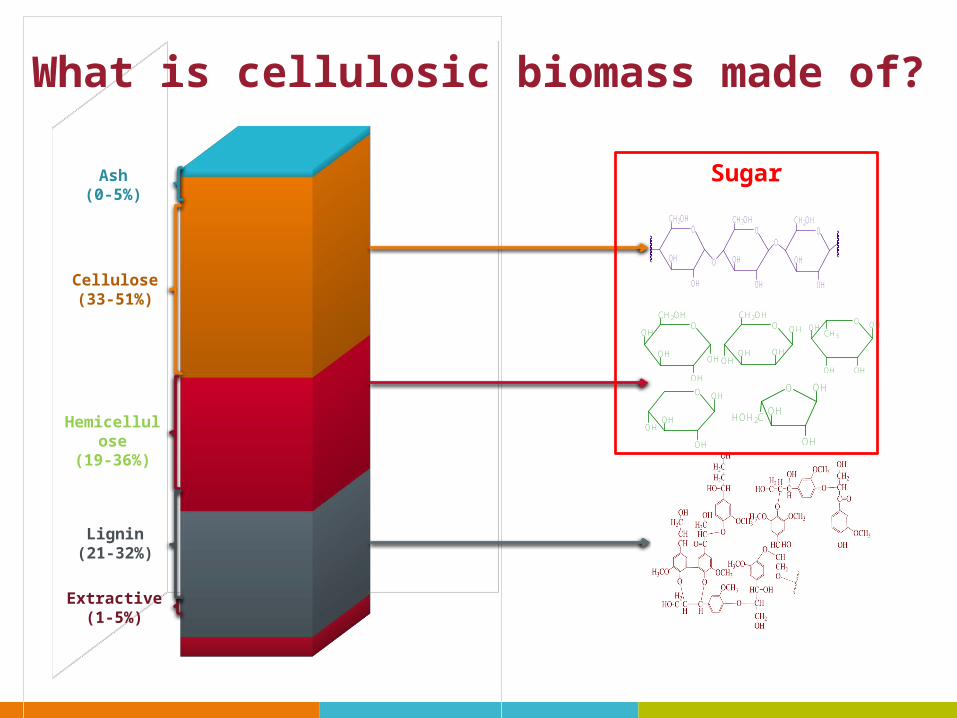
Ash(0-5%)
Cellulose(33-51%)
Hemicellulose(19-36%)
Lignin(21-32%)
Extractive(1-5%)
O
O
OH
OH
CH2OHO
O
OH
OH
CH2OHO
OH
OH
CH2OH
O
OH
OH
CH2OH
OH
OHO OH
OHOH
CH2OH
OH
O OHCH3
OH
OHOH
O OH
OHOH
OH
O OH
OH
OHHOH2C
Sugar
What is cellulosic biomass made of?

Fermentation of glucose & xylose
Glucose and xylose are the major monomeric sugars in the lignocellulosic biomass hydrolysates
A major challenge in efficient utilization of mixed sugars for biofuel production
Many oleaginous yeasts can utilize glucose and xylose but lack the capability of simultaneous fermentation of both sugars, resulting in xylose accumulation

Glucose repression on non-glucose sugar utilization
Microorganisms generally consume the mixed sugars sequentially
Glucose first and then xylose
Allosteric competition for sugar transporters (Kawaguchi et al., 2006)
Solutions
1. Express xylose transporters
2. Introduce cellodextrin transporters
(Kim et al., 2012; Kawaguchi et al., 2006; Zaldivar and Olsson, 2001; Hu et al., 2011; Kastner et al., 1998)

(Ha et al., 2011)
Xylose-specific transporter

(Ha et al., 2011)
Cellodextrin transporters

Sugar transport study
MethodologyDetermine uptake rates of the target sugar by measuring the radio activities of C14 labeled sugar entering into the cells.
Major equipment: Liquid scintillation counter

• Model I, a Michaelis-Menten function: representing a single, saturable carrier
• Model II, a Michaelis-Menten function plus a linear term: representing a single carrier and a non-saturable diffusion process
• Model III, a sum of two Michaelis-Menten functions: representing two saturable carriers in the membrane
Kinetic Model of sugar transport

Example 2 Kinetics of xylose transport
S (mM) V(umol/min/g DCW)2.00 2.024.00 3.646.00 3.878.00 5.5910.00 4.5012.00 6.7414.00 5.5516.00 8.4320.00 6.6150.00 7.87
150.00 10.49200.00 10.74500.00 14.25
Experiment
Measure the uptake rates in C. curvatus at different concentrations of xylose
Task Determine the model of xylose transport in Cryptococcus curvatus

Data analysis: Non-linear regression
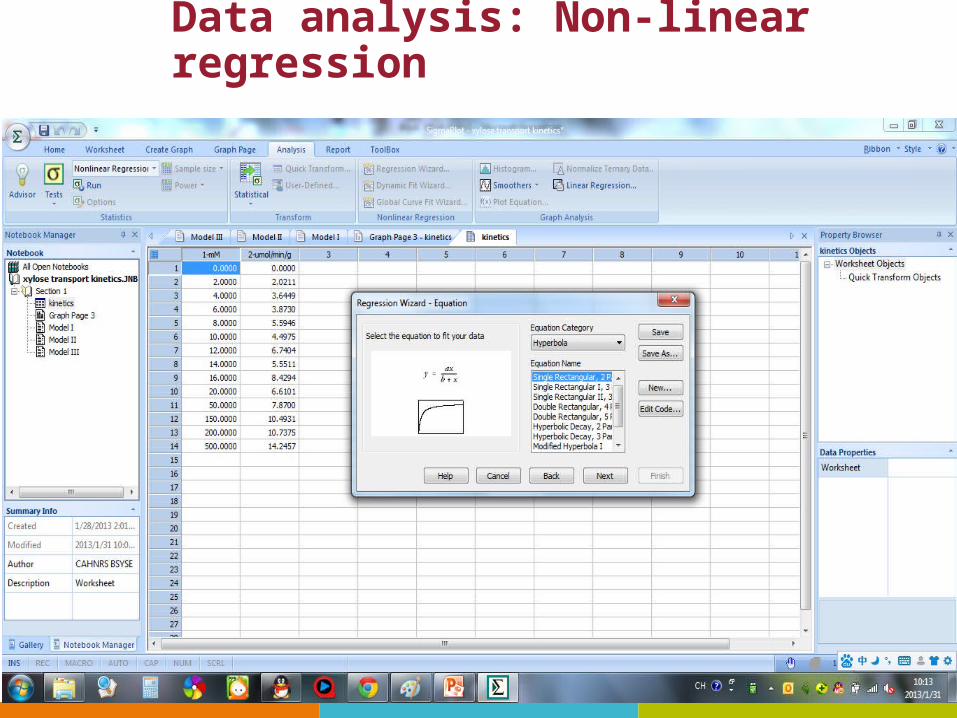
Data analysis: Non-linear regression

Data analysis: Non-linear regression
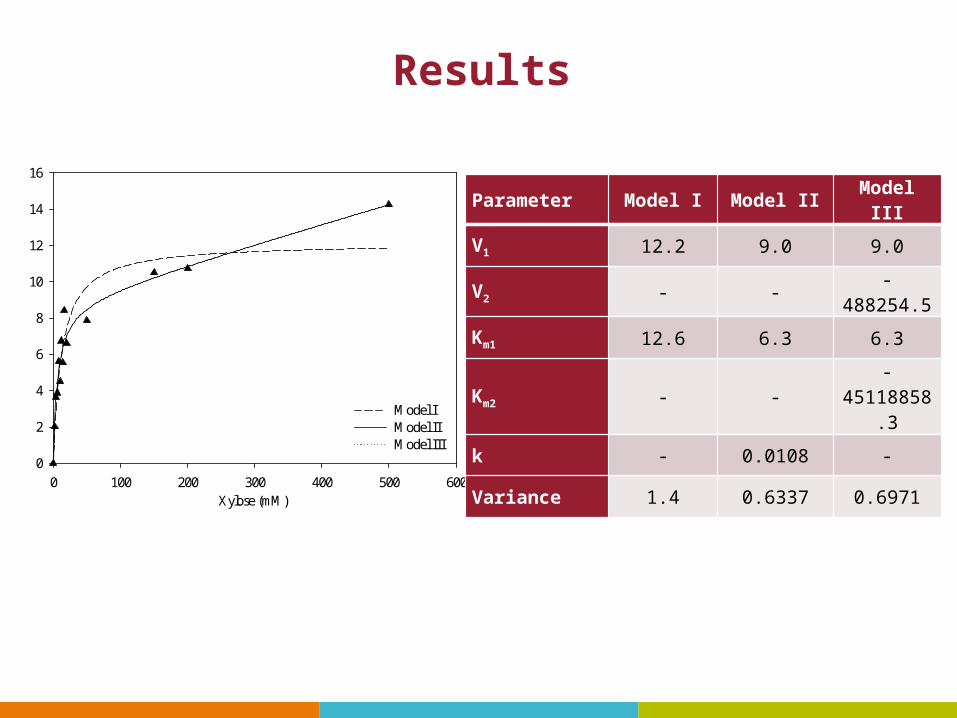
Results
Xylose (mM)0 100 200 300 400 500 600
V (nm
ol/min/m
g DC
W)
0
2
4
6
8
10
12
14
16
Model IModel IIModel III
Parameter Model I Model II Model III
V1 12.2 9.0 9.0
V2 - - -488254.5
Km1 12.6 6.3 6.3
Km2 - --
45118858.3
k - 0.0108 -
Variance 1.4 0.6337 0.6971

Other application of Michaelis-Menten kinetics
Enzyme behavior ----- competitive, non-competitive and uncompetitive inhibition
Hill reaction---- the laboratory equivalent of the light reaction of photosynthesis
The reaction center of photosystem II can be saturated with photons of red light just like the active site of an enzyme can be saturated with substrate molecules.

(Savage, 2011)
Final biofuel products

Comparison of chemical properties of transportation
fuels
Structural features influencing the physical and fuel properties of a fatty ester molecule
• Carbon chain length• degree of saturation• branching of the chain
(Li et al., 2010)
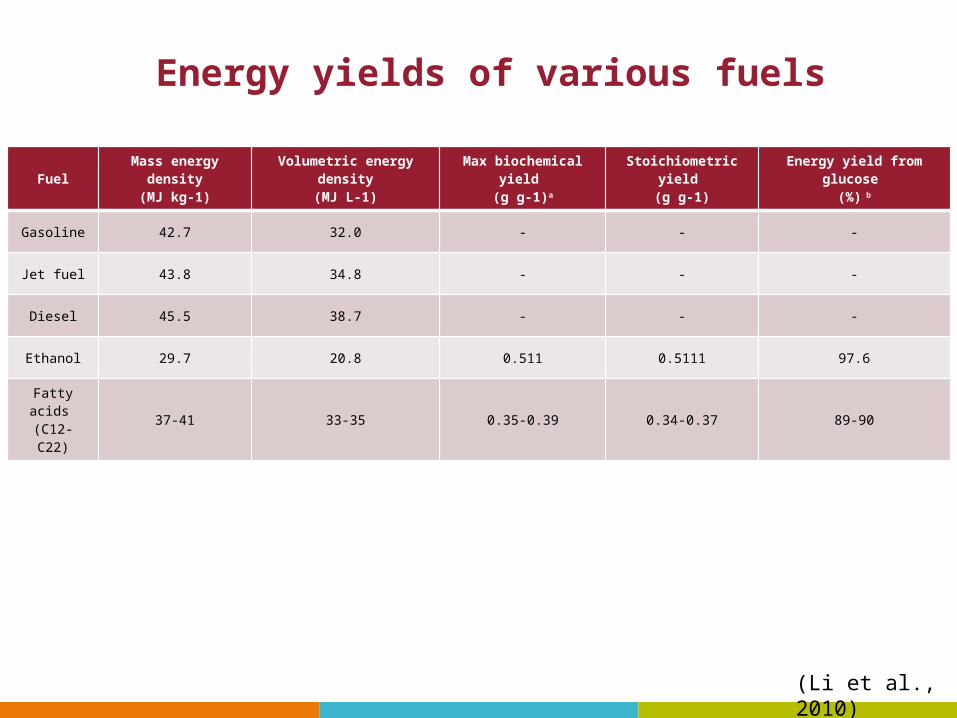
Energy yields of various fuels
FuelMass energy density
(MJ kg-1)Volumetric energy density
(MJ L-1)Max biochemical yield
(g g-1)a
Stoichiometric yield (g g-1)
Energy yield from glucose (%) b
Gasoline 42.7 32.0 - - -
Jet fuel 43.8 34.8 - - -
Diesel 45.5 38.7 - - -
Ethanol 29.7 20.8 0.511 0.5111 97.6
Fatty acids (C12-C22)
37-41 33-35 0.35-0.39 0.34-0.37 89-90
(Li et al., 2010)
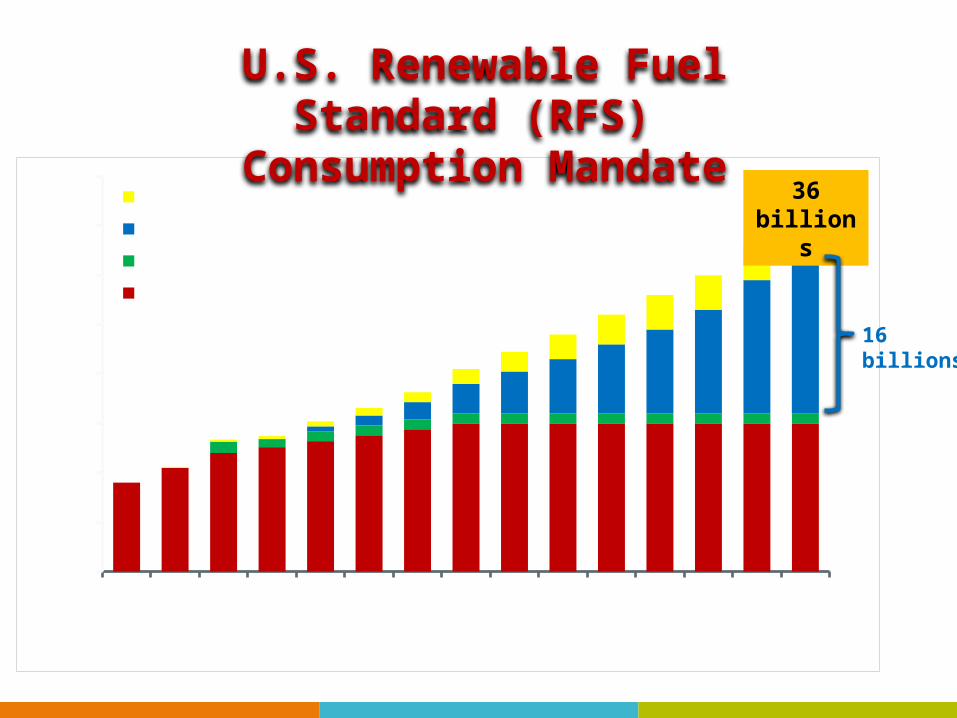
20082009
20102011
20122013
20142015
20162017
20182019
20202021
20220
5
10
15
20
25
30
35
40Other
Cellulosic biofuels
Biomass-based diesel
Conventional biofuels
Year
Bil
lion
s of
Gal
lon
sU.S. Renewable Fuel Standard (RFS)
Consumption Mandate
36 billions
16 billions

How can we achieve the goal?
Progress in biofuel research, development and commercialization
Solazyme, Inc.


Algae-based biodiesel fuel now available at pump 11.14.2012 abc news

Reference• Liping Xiao, 2010. Evaluation of Extraction Methods for Recovery of Fatty Acids from Marine Products.
Retrieved from http://www.cursos.ualg.pt/emqal/documents/thesis/Liping_Xiao.pdf
• Tanaka, K. et al, 2002. Two different pathways for D-xylose metabolism and the effect of xylose concentration on the yield coefficient of L-lactate in mixed-acid fermentation by the lactic acid bacterium Lactococcus lactis IO-1. Appl Microbiol Biotechnol 60:160–167
• Papanikolaou, S. and Aggelis, G. 2011. Lipids of oleaginous yeasts. Part I: Biochemistry of single cell oil production. Eur. J. Lipid. Sci. Technol. 113: 1031-1051
• Kawaguchi, H., et al., Engineering of a Xylose Metabolic Pathway in Corynebacterium glutamicum. Applied and Environmental Microbiology, 2006. 72(5): p. 3418-3428.
• Kim, S.R., et al., Simultaneous co-fermentation of mixed sugars: a promising strategy for producing cellulosic ethanol. Trends in Biotechnology, 2012. 30(5): p. 274-282.
• Zaldivar, J., J. Nielsen, and L. Olsson, Fuel ethanol production from lignocellulose: a challenge for metabolic engineering and process integration. Applied Microbiology and Biotechnology, 2001. 56(1): p. 17-34.
• Hu, C., et al., Simultaneous utilization of glucose and xylose for lipid production by Trichosporon cutaneum. Biotechnology for Biofuels, 2011. 4: p. 25.
• Kastner, J.R., W.J. Jones, and R.S. Roberts, Simultaneous utilization of glucose and D-xylose by Candida shehatae in a chemostat. Journal of Industrial Microbiology & Biotechnology, 1998. 20(6): p. 339-343.
• Li, H., et al., Biofuels: Biomolecular Engineering Fundamentals and Advances. Annu. Rev. Chem. Biomol. Eng. 2010. 1:19–36


















