
Mechanisms of anion and fluid secretion by airway...
180
Mechanisms of anion and fluid secretion by airway epithelial cells Jiajie Shan Department of Physiology McGill University Montreal, Quebec, Canada December 2013 A thesis submitted to the faculty of Graduate Studies and Research in partial fulfillment of the degree of Doctorate in Philosophy Copyright © Jiajie Shan 2013
Transcript of Mechanisms of anion and fluid secretion by airway...

Mechanisms of anion and fluid secretion by airway epithelial cells
Jiajie Shan
Department of Physiology
McGill University Montreal, Quebec, Canada
December 2013
A thesis submitted to the faculty of Graduate Studies and Research in partial fulfillment of the degree of
Doctorate in Philosophy
Copyright © Jiajie Shan 2013

I
Table of contents
Abstract ………………………………………………………………………………………………………… VII
Résumé .............................................................................................................. IX
Acknowledgements ……………………………………………………………………………………... XI
Contributions of authors ..…………………………………………………………………………….. XII
Abbreviation …………………………………………………………………………………………………. XIV
Chapter 1 Introduction
1.1 The structure and function of airway epithelium …………………………………… 2
1.1.1 Ciliated cells …………………………………………………………………………….. 3
1.1.2 Basal cells ………………………………………………………………………………… 3
1.1.3 Mucous cells ……………………………………………………………………………. 3
1.1.4 Clara cells …………………………………………………………………………………. 4
1.1.5 Serous cells ………………………………………………………………………………. 4
1.1.6 Type I cells ………………………………………………………………………………… 5
1.1.7 Type II cells ……………………………………………………………………………….. 5
1.2 Junctional complexes in airways ……………………………………………………………… 5
1.3 The primary functions of airway epithelium: mucus production and
mucociliary clearance …………………………………………………………………………….. 6
1.4 Regulation of ASL ……………………………………………………………………………………. 8
1.4.1 Na+/K+-ATPase ……………………………………………………………………………. 9
1.4.2 V-ATPase ……………………………………………………………………………………. 10

II
1.4.3 NKCC ………………………………………………………………………………………….. 11
1.4.4 NBC …………………………………………………………………………………………… 11
1.4.5 The Na+-independent Cl-/HCO3- exchanger (AE) ………………………… 12
1.4.6 Pendrin (SLC26A4) …………………………………………………………………….. 12
1.4.7 The Na+/H+ exchanger (NHE) ……………………………………………………… 13
1.4.8 The H+ channel …………………………………………………………………………… 14
1.4.9 The chloride channels ………………………………………………………………… 14
1.4.10 ENaC ………………………………………………………………………………………….. 16
1.4.11 Carbonic anhydrase ……………………………………………………………………. 17
1.4.12 Regulation of ASL hydration ………………………………………………………. 18
1.4.13 pH regulation of ASL ………………………………………………………………….. 19
1.5 Models of anion transport in submucosal glands ……………………………………. 22
Chapter 2 anion transport in airway epithelial cells
2.1 Abstract …………………………………………………………………………………………………… 31
2.2 Introduction …………………………………………………………………………………………….. 32
2.3 Methods ………………………………………………………………………………………………….. 35
2.3.1 Cell culture …………………………………………………………………………………. 35
2.3.2 Solutions ……………………………………………………………………………………. 36
2.3.3 Immunoblotting …………………………………………………………………………. 37
2.3.4 Measurements of equivalent Isc and HCO3- secretion …………………. 37
2.3.5 Tracer fluxes ……………………………………………………………………………….. 38

III
2.3.6 Data analysis ………………………………………………………………………………. 39
2.4 Results ……………………………………………………………………………………………………… 39
2.4.1 HCO3- secretion accounts for the short-circuit current (Isc) …………. 39
2.4.2 HCO3- secretion is CFTR dependent ……………………………………………. 40
2.4.3 Proton secretion neutralizes some of the transported bicarbonate . 48
2.4.4 CFTR mediates apical HCO3- conductance …………………………………… 50
2.4.5 Electrogenic anion transport (Ieq) requires Na+ and CO2/HCO3- ……. 54
2.4.6 Some basolateral Cl- entry is independent of NKCC …………………….. 56
2.4.7 Evidence for basolateral Cl- loading by anion exchange ………………. 58
2.4.8 Electrogenic HCO3- secretion requires carbonic anhydrase …………. 62
2.4.9 Origin of the basal Ieq and HCO3- secretion …………………………………. 62
2.5 Discussion ………………………………………………………………………………………………… 69
2.5.1 Isc is mediated by electrogenic HCO3- transport …………………………… 69
2.5.2 Ieq depends on Na+ and CO2/HCO3- ……………………………………………… 70
2.5.3 Cl- loading depends on basolateral anion exchange mostly rather
than NKCC …………..…………………………………………………………………….. 71
2.5.4 Forskolin-stimulated HCO3- secretion requires carbonic anhydrase .. 72
2.5.5 Further evidence that CFTR mediates apical membrane HCO3-
and Cl- conductance …………………………………………………………………… 73
2.5.6 A revised anion transport model of Calu-3 cells …………………………. 75
Chapter 3 Fluid secretion by an airway epithelial cell line

IV
3.1 Abstract ………………………………………………………………………………………………….. 80
3.2 Introduction ……………………………………………………………………………………………. 81
3.3 Methods …………………………………………………………………………………………………. 84
3.3.1 Cell culture ………………………………………………………………………………… 84
3.3.2 Media ………………………………………………………………………………………… 84
3.3.3 Volume and composition of the secreted fluid ………………………….. 85
3.3.4 Data analysis ……………………………………………………………………………… 86
3.4 Results …………………………………………………………………………………………………….. 87
3.4.1 Fluid secretion is CFTR dependent ……………………………………………… 87
3.4.2 Cl- is the predominant anion in Calu-3 secretion ………………………… 87
3.4.3 HCO3-, Na+ and Cl- are all required for forskolin-stimulated
fluid secretion ……………………………………………………………………………. 89
3.4.4 Most of the fluid secretioin is independent of NKCC ………………….. 92
3.4.5 Basolateral anion exchange is important to fluid secretion ………… 92
3.4.6 Carbonic anhydrase has little effect on fluid secretion ……………….. 94
3.4.7 The time course of fluid secretion rate during 24 h stimulation …. 97
3.4.8 Evaporation does not affect the rate of fluid accumulation ………… 97
3.4.9 Osmolality determined the fluid secretion rate ………………………….. 99
3.5 Disscussion ………………………………………………………………………………………………. 101
3.5.1 Most fluid is driven by the net flux of Cl- ……………………………………… 103
3.5.2 Calu-3 secretions are only weakly alkaline ………………………………….. 104
3.5.3 A very low rate of net Cl- secretion is sufficient to

V
drive fluid secretion ……………………………………………………………………. 106
3.5.4 The osmolality determines the fluid secretion rate ……………………. 106
3.5.5 Carbonic anhydrase participates little in fluid secretion ……………… 108
Chapter 4 The role of carbonic anhydrase in anion and fluid secretion
4.1 Abstract ……………………………………………………………………………………………………. 112
4.2 Introduction ……………………………………………………………………………………………… 113
4.3 Methods …………………………………………………………………………………………………… 115
4.3.1 Cell culture …………………………………………………………………………………. 115
4.3.2 Solutions …………………………………………………………………………………….. 116
4.3.3 Measurements of equivalent Isc and HCO3- secretion ………………….. 117
4.3.4 RT-PCR ………………………………………………………………………………………… 117
4.3.5 Immunoblotting ………………………………………………………………………….. 118
4.3.6 Immunoprecipitation ………………………………………………………………….. 119
4.3.7 Fluid secretion assay …………………………………………………………………… 119
4.3.8 Data analysis ………………………………………………………………………………. 120
4.4 Results ……………………………………………………………………………………………………… 120
4.4.1 Carbonic anhydrase contributes partially to anion transport ……… 120
4.4.2 The expression of different isoforms of carbonic anhydrase ………. 121
4.4.3 CAIX is not involved in anion and fluid secretion by Calu-3 cells …. 123
4.4.4 CAII and CAXII are responsible for the forskolin-stimulated anion
secretion by Calu-3 cells …………………………………………………………….. 125

VI
4.4.5 CAII does not form a complex with CFTR ……………………………………… 128
4.5 Discussion ………………………………………………………………………………………………… 129
4.5.1 Carbonic anhydrase II is important for anion and fluid secretion .. 129
4.5.2 Further evidence that HCO3- is required for Cl- secretion ……………. 131
4.5.3 The bicarbonate transport metabolon ………………………………………… 132
4.5.4 The contributions of CAII and NBC to anion and fluid secretion …. 133
Chapter 5 General discussion
5.1 A novel model of anion transport of airway epithelial cells ………………………. 136
5.1.1 V-clamp vs. I-clamp …………………………………………………………………….. 136
5.1.2 pH-stat conditions ………………………………………………………………………. 138
5.1.3 The Calu-3 cell line ……………………………………………………………………… 138
5.1.4 Modification to existing models ………………………………………………….. 140
5.2 The determinants of fluid secretion ………………………………………………………….. 142
5.3 The role of carbonic anhydrases ……………………………………………………………….. 143
5.4 Conclusions and future direction ………………………………………………………………. 144
References ………………………………………………………………………………………………….. 146

VII
Abstract
Anion transport drives fluid into the airways and is essential for humidifying inspired
air and supplying surface liquid for mucociliary transport. Despite the importance of
airway secretion in diseases such as cystic fibrosis, the cellular mechanisms of anion
and fluid secretion remain poorly understood. To clarify these mechanisms, I studied
the Calu-3 cell line, a widely used model for serous cells in airway submucosal glands.
First, I studied anion transport using a technique which combines electrical
measurement of ion transport and pH-stat. Although a small fraction of the Cl-
secretion required basolateral entry through Na+, K+, 2Cl- cotransporter NKCC, I found
that most Cl- uptake was mediated by basolateral Cl-/HCO3- exchange. The HCO3
-
required for this exchange process was supplied by the basolateral Na+, HCO3-
cotransporter NBC and also by the hydration of CO2, which was catalyzed by carbonic
anhydrase. The data were consistent with most HCO3- and Cl- secretion occurring via
apical CFTR channels. Based on these findings, I propose a new model for
transepithelial anion secretion, which may explain early studies in the 1980’s which
indicated that most tracheal submucosal gland secretion is surprisingly insensitive to
bumetanide.
This new scheme for anion transport was further tested using fluid secretion assays
and was found to explain the ion dependencies and pharmacology of fluid secretion.
Moreover, fluid was actively secreted in the first 6 hours during the 24-h fluid

VIII
secretion assay, and secretions collected during that period had higher osmolality
than the culture medium. These results suggest that airway surface fluid is regulated
by osmolality and the water flux simply depends on the total amount of osmotically
active solute on the apical surface.
Finally, I explored the role of carbonic anhydrases because inhibitor studies indicated
that it is critically important for anion transport by Calu-3 cells. I found that several
isoforms of carbonic anhydrases are expressed in Calu-3 cells, namely CAII, CAIX and
CAXII. Inhibitor studies revealed that CAIX is not required for anion transport. CAII
is probably the isoform involved in bicarbonate secretion, although further studies
are needed to exclude the involvement of CAXII.
Taken together, these present findings support a novel cellular mechanism for anion
and fluid secretion by Calu-3 cells. This scheme may be applicable to airway
submucosal glands which control the pH, volume, and composition of airway surface
liquid and are critical for airway host defense.

IX
Résumé
Le transport d’anions entraîne le fluide dans les voies aériennes et est essentiel pour
l’humidification de l’air inspiré et pour apporter du liquide de surface nécessaire au
transport mucoscilliaire. Malgré l’importance de la sécrétion au niveau des voies
respiratoires dans le cas de maladies telles que la fibrose kystique, les mécanismes
cellulaires de la sécrétion d’anions et de fluide ne sont pas tout à fait élucidés. Afin de
clarifier les mécanismes, j’ai utilisé la lignée cellulaire Calu-3, un modèle connu de
cellules séreuses dans les glandes submucosales des voies aériennes.
En premier lieu, j’ai étudié le transport d’anions en utilisant une technique
combinant la mesure électrique de transport d’ions et le pH-stat. Bien qu’une petite
fraction de sécrétion de Cl- requiert l’entrée par le co-transporteur basolatéral Na+, K+,
2Cl- NKCC, j’ai démontré que la plupart de l’apport de Cl- est médié par l’échange
Cl-/HCO3-basolatéral. Le HCO3
- requis pour cet échange est apporté par le
co-transporteur basolatéral Na+, HCO3- NBC et également par l’hydratation de CO2,
qui est catalysée par l’anhydrase carbonique. Les données corrèlent avec la plupart
de la sécrétion de HCO3- et Cl- par les canaux CFTR apicaux. D’après ces résultats, je
propose un nouveau modèle de sécrétion transépithéliale d’anions, qui pourrait
expliquer des études précédentes datant des années 1980, qui indiquaient que la
sécrétion des glandes submucosales de trachée est majoritairement insensible au
bumétanide.

X
Ce nouveau modèle de transport d’anions a ensuite été testé par la méthode de
sécrétion de fluide et a permis d’expliquer les dépendances en ions et la
pharmacologie de la sécrétion de fluide. Aussi, le fluide est activement secrété
pendant les 6 premières heures au cours d’un essai de sécrétion de fluide sur 24
heures, et les secrétions collectées pendant cette période ont une osmolarité plus
importante que le milieu de culture. Ces résultats suggèrent que le fluide de surface
des voies aériennes est régulé par l’osmolarité et que le flux d’eau dépend
simplement de la quantité totale de soluté osmoticallement actif à la surface apicale.
Enfin, j’ai étudié le rôle des anhydrases carboniques car mes expériences d’inhibition
démontrent son importance dans le transport d’anions par les cellules Calu-3. J’ai mis
en évidence qu’il existe trois isoformes d’anhydrases carboniques exprimées dans les
cellules Calu-3 : CAII, CAIX et CAXII. Des expériences d’inhibition ont révélé que CAIX
n’est pas impliqué dans le transport d’anions. CAII est probablement l’isoforme
impliqué dans la sécrétion de bicarbonate, bien que des expériences
complémentaires soient nécessaires pour exclure l’implication de CAXII.
Pour conclure, ces résultats appuient un nouveau mécanisme cellulaire de sécrétion
d’anions et de fluide par les cellules Calu-3. Ce modèle pourrait être applicable aux
glandes submucosales des voies respiratoires qui contrôlent le pH, le volume et la
composition du liquide de surface des voies aériennes et qui sont essentielles à leur
défense.

XI
Acknowledgements
I would like to thank my supervisor Dr. John Hanrahan for his guidance, patience and
support throughout my study. I am grateful for his dedication and drive, without
which this work would not have been possible. I am also particularly thankful that I
learned a lot from his enormous knowledge throughout these years, which has
become my precious assets.
I would also like to thank my lab members, past and present. In particular, Dr. Jie Liao,
Mr. Junwei Huang, Ms. Adeline Wohlhuter, Ms. Alexandra Evagelidis and Ms. Julie
Goepp for their invaluable help throughout my study. The daily pleasant and joyous
company of my lab members eases my mind when I am stuck in the boring lab work.
I am very thankful to my committee members, Dr. John Orlowski, Dr. Gergely Lukacs
and Dr. Jean-Yves Lapointe. Their expertise provide me with insight, useful advice and
feedback for my work. I must thank them for spending their precious time in meeting
with me and discussing my work.
Finally, I would like to thank my parents for their constant support since I was born,
without them no accomplishment would be achieved.

XII
Contribution of authors
Chapter 2 contains part of a published manuscript: Shan J. et al.
Bicarbonate-dependent chloride transport drives fluid secretion by the human
airway epithelial cell line Calu-3. J Physiol 590(Pt 21): 5273-5297, 2012. For this study,
I performed Ieq and HCO3- secretion measurements in the Ussing chamber, analyzed
the data, and prepared the draft of the manuscript. Dr. Jie Liao performed Western
Blot to verify the CFTR-knockdown Calu-3 cell line, which is shown in Fig. 2.2A. Dr.
Renaud Robert performed Isc measurements shown in Fig. 2.4.
Chapter 3 also contains part of a published manuscript: Shan J. et al.
Bicarbonate-dependent chloride transport drives fluid secretion by the human
airway epithelial cell line Calu-3. J Physiol 590(Pt 21): 5273-5297, 2012. For this study,
I performed all the fluid secretion assay, analyzed the data, and prepared the draft of
the manuscript.
Chapter 4 contains a manuscript in preparation. For this study, I performed Ieq and
HCO3- secretion measurements, RT-PCR, Western Blot and co-immunoprecipitation. I
collected the data and analyzed.
Chapter 5 contains part of a published manuscript: Shan J. et al. Anion secretion by a
model epithelium: more lessons from Calu-3. Acta Physiol (Oxf) 202(3): 523-531,
2011. I mainly provided data, figure and discussion for this manuscript.

XIII
Throughout different stages of the work of this thesis, my supervisor Dr. John
Hanrahan provided guidance, instruction and discussion, including experimental
design, data analysis and the preparation of the final manuscripts.

XIV
Abbreviations
AE anion exchanger
ASL airway surface liquid
CA carbonic anhydrase
CF cystic fibrosis
CFTR cystic fibrosis transmembrane conductance regulator
Ieq equivalent short-circuit current
Isc short-circuit current
NBC sodium bicarbonate cotransporter
NKCC sodium potassium chloride cotransporter
pHi intracellular pH

1
Chapter 1 Introduction

2
1.1 The structure and function of airway epithelium
The human airways have 23 generations of dichotomous branches, which can be
divided into two parts according to their main functions, the conducting airways and
the respiratory airways. From generation 0 (trachea) to between 8 and 11, the
airways termed bronchi contain both cartilage and mucous glands. From around
generation 10 to 16, the airways lack cartilage, glands and alveoli, and are termed
conducting bronchioles. These two sections comprise the conducting airways, which
are responsible for transport of the inhaled air to the respiratory airway surfaces or
the exhaled air out of the lung. From generation 17 on, the airways become
respiratory bronchioles and have increasing numbers of alveoli. By generation 20, the
airway is full of alveoli and it ends in an alveolar sac at generation 23. These
respiratory airways are the site of gas exchange (Fischer and Widdicombe 2006,
Hollenhorst, Richter et al. 2011).
All parts of the airways are lined with an epithelium, which contains various
epithelial cell types having different morphologies and functions. Additionally, these
cell types are expressed in different proportions along the airway (Knight and Holgate
2003, Fischer and Widdicombe 2006, Hollenhorst, Richter et al. 2011, Tam,
Wadsworth et al. 2011). In the trachea, the surface epithelium is pseudostratified
and about 50 μm in height. The three main cell types are ciliated, basal and mucous
cells. With increasing airway generations, the height of the epithelium shortens
progressively. In the respiratory bronchioles, the height reaches ~10 μm and the

3
morphology of the epithelium becomes columnar.
1.1.1 Ciliated cells
Ciliated cells are the predominant cell type in the airways, accounting for ~50% of the
total cell number in the surface epithelium of all airway generations. Typically, each
cell possess up to 300 cilia on the apical surface, with numerous mitochondria
beneath them, indicating that the principal metabolic function of ciliated cells is to
provide ATP for ciliary beating and mucociliary clearance of inhaled particles out of
the lung (Knight and Holgate 2003, Fischer and Widdicombe 2006, Tam, Wadsworth
et al. 2011).
1.1.2 Basal cells
Basal cells are ubiquitously expressed along the conducting airways, though their
number decreases distally and are completely lost before the respiratory bronchioles
(Evans and Plopper 1988, Boers, Ambergen et al. 1998). These cells attach firmly to
the basement membrane and thus play a role in anchoring other superficial epithelial
cells (Evans and Plopper 1988, Evans, Cox et al. 1989, Evans, Cox et al. 1990). In
addition, basal cells act as stem cells in the lung and give rise to other epithelial cell
types, for example the ciliated cells (Hong, Reynolds et al. 2004, Hong, Reynolds et al.
2004).
1.1.3 Mucous cells

4
Mucous cells contain acidic-mucin granules and secret mucus into the airway to trap
foreign particles, which can be cleared away together with mucus by ciliated cells
(Knight and Holgate 2003, Tam, Wadsworth et al. 2011). In normal human trachea,
the estimated number of mucous cells is up to 6,800 cells/mm2 of surface epithelium
(Lumsden, McLean et al. 1984). Similar to basal cells, the number of mucous cells
declines progressively distally along the respiratory tree, and they are replaced by
Clara cells in the respiratory bronchioles (Boers, Ambergen et al. 1998). This switch
from mucous cells to Clara cells may reflect the fact that mucus is mainly a defense
against inhaled particles, which are deposited in the larger airways. Goblet cells
develop from basal cells, however in mice and perhaps humans, Clara cells can also
become mucus secreting during the mucus cell metaplasia induced by antigen
challenge (Evans, Williams et al. 2004).
1.1.4 Clara cells
Clara cells secret bronchiolar surfactants and antiproteases (Clara cell secretory
proteins, CCSPs) to protect the airway against toxic chemicals (Knight and Holgate
2003, Fischer and Widdicombe 2006, Tam, Wadsworth et al. 2011). Besides their
secretory function, they can also serve as stem cells when the number of basal cells
decreases (Hong, Reynolds et al. 2001).
1.1.5 Serous cells
Serous cells are present in the bronchioles. The morphology of serous cells resembles

5
that of goblet-shaped mucous cells however, their cellular contents differ. They are
specialized for fluid secretion into the airway to form the airway surface liquid and
also release antimicrobial factors (Rogers, Dewar et al. 1993, Fischer and
Widdicombe 2006).
1.1.6 Type I cells
Type I cells exist in the respiratory airway. Compared to other airway epithelial cells,
type I cells are flat and have very thin extensions, which cover over 98% of the
internal surface area of the lung. Thus, these cells form a thin barrier between the air
and the blood (Dobbs and Johnson 2007).
1.1.7 Type II cells
Unlike type I cells, type II cells are small cuboidal cells. These cells contain secretory
granules called lamellar bodies, and secret surfactant from these intracellular
organelles. Thus, the main functions of type II cells are the synthesis, secretion and
reuptake of the surfactant (Dobbs and Johnson 2007).
1.2 Junctional complexes in airways
Since there are various cells in the intact airway epithelium, cell-cell communication
is necessary between cells of the same cell type and between different cell types.
Therefore, paracrine purinergic signaling and junctional complexes at the contact
sites play important roles in cell-cell communication (Tam, Wadsworth et al. 2011).

6
Anchoring junctions, including desmosomes, hemidesmosomes and adherens
junctions, are vital in maintaining the structural integrity of the airway epithelium, as
they mediate direct or indirect cell-cell adhesion (Tam, Wadsworth et al. 2011).Tight
junctions, which include occludin, claudins and junctional adhesion molecule (JAM)
form zonula occludens, and are important in regulating the flow of solutes across the
epithelium by being selective for solutes of different sizes and charges (Harhaj and
Antonetti 2004).
Gap junctions consist of hexamers of connexin protein and form a pore that
transports molecules smaller than about 1 kDa between cells. They are especially
important in the bronchial and alveolar epithelium, since type I and type II cells
utilize these ‘channels’ to transit antioxidants, cytoplasmic metabolites and second
messenger signals between neighboring cells (Koval 2002, Boitano, Safdar et al.
2004).
1.3 The primary functions of airway epithelium: mucus
production and mucociliary clearance
The respiratory epithelial cells of the alveoli allow gas exchange while forming a
barrier that separates the external environment from the inner tissues and
vasculature of the lung. The conducting airways also form a barrier lined by epithelial
cells, but they are more complex and provide protection for the lung against harmful
foreign particles. Mucus-secreting goblet cells in the airway surface epithelium and

7
mucus tubules of the submucosal glands produce and secrete mucus that traps
pathogens and other particles so they can be cleared. Two important mucins in
human airways are MUC5AC and MUC5B. MUC5AC is produced mainly in the surface
epithelium and provide an acute response to direct contact with environmental
challenges, whereas MUC5B is most prominent in the submucosal glands and
involved in responses to chronic infection and inflammation (Hovenberg, Davies et al.
1996, Wickstrom, Davies et al. 1998, Thornton, Rousseau et al. 2008).
However, secreting mucus is not sufficient to protect against infection, mucus must
also be cleared by beating cilia which line the airways. To enable mucociliary
clearance, the epithelial cells secrete fluid in order to maintain a thin layer of liquid
6-7 μm in height on the epithelial surface (Tarran, Grubb et al. 2001). Most of the
secreted gel-forming mucins float on the surface of this liquid. Thus, ASL (airway
surface liquid) contains two layers, an outer mucous layer and a watery periciliary
layer between the epithelium and the mucus. The periciliary layer has lower viscosity
and enables the cilia to beat in a highly coordinated fashion that propels the mucus.
The tips of cilia penetrate into the mucous layer during the power stroke, propelling
it up the airways and out of the lung together with entrapped particles (Fischer and
Widdicombe 2006, Tam, Wadsworth et al. 2011).
In normal airways there is a fine balance between mucus production and clearance
(Evans and Koo 2009). This equilibrium is disrupted in cystic fibrosis, leading to

8
mucus plugging of the airways, infection, inflammation, epithelial damage, and
fibrosis (Boucher 2007, Boucher 2007, Chambers, Rollins et al. 2007, Mall 2008).
1.4 Regulation of ASL
The regulation of ASL hydration is a critical factor affecting mucociliary clearance, and
understanding its production is a focus of this study. Normal ASL is usually of 6-7 μm
in height, which approximates the length of the outstretched cilia and enables
efficient ciliary beating (Tarran, Grubb et al. 2001). The volume of the ASL and mucus
hydration are maintained by ion transport processes of airway epithelial cells, which
control the mass of salt (i.e. NaCl) on the airway surface, with water following
passively by osmosis (Matsui, Grubb et al. 1998). Ion transport by the surface
epithelium requires the coordinated activity of many transporters and channels,
including basolateral Na+,K+-ATPase, basolateral Na+,K+,Cl--cotransporter and apical
CFTR, apical ENaC (epithelial Na+ channel). In CF, the loss of CFTR activity reduces Cl-
secretion and may also lead to increased Na+ absorption by ENaC which prevent CF
epithelia from maintaining normal ASL volume (Boucher 2007, Boucher 2007,
Chambers, Rollins et al. 2007, Mall 2008). Consequently the height of the ASL
collapses to ~3 μm, which is not sufficient for ciliary mucociliary clearance (Tarran,
Button et al. 2005, Boucher 2007).
The pH of ASL is also important for airways defense although it is less well
understood. Firstly, pH can influence ENaC, and thus modulate sodium absorption

9
and the volume of the ASL (Awayda, Boudreaux et al. 2000). Secondly, several
proteins that actively participate in airway defense are pH-sensitive. Mucus
rheological properties are pH-dependent and the activity of some antibacterial
proteins and cationic antimicrobial peptides depend on pH (Ganz 2002). Regulation
of ASL pH is mediated by transporters, channels and enzymes, including the vacuolar
H+-ATPase, the H+ channels, and CFTR in the apical membrane, intracellular carbonic
anhydrase, and sodium bicarbonate cotransporter and anion exchangers in the
basolateral membrane etc. (Fischer and Widdicombe 2006).
Before considering current models for ion transport processes that regulate the
volume and pH of ASL, we first need to understand the structure and function of
these proteins.
1.4.1 Na+/K+-ATPase
Na+/K+-ATPase, also known as the sodium pump, is expressed ubiquitously in the
basolateral membrane of epithelia (with the exception of the choroid plexus and the
retinal pigment epithelium) (Ernst, Palacios et al. 1986, Gundersen, Orlowski et al.
1991, Crump, Askew et al. 1995). The enzyme belongs to the P-type ATPase family
because it becomes phosphorylated during the pump cycle. Na+/K+-ATPase is the
most important primary active transporter in the airway epithelium (Morth,
Pedersen et al. 2011). For each ATP hydrolysed, Na+/K+-ATPase transports 3 Na+ out
of the cell, and 2 K+ into the cell. Thus the pump generates both electrical and

10
chemical gradients that in turn drive ion movements through channels and
secondary active transporters.
Besides acting as a primary active transporter, Na+/K+-ATPase can also function as a
signaling molecule which regulates various enzymes and the formation and
maintenance of tight junctions between epithelial cells (Rajasekaran and Rajasekaran
2009).
1.4.2 V-ATPase
The vacuolar H+-ATPase (V-ATPase) is a proton pump responsible for acidifying
intracellular compartments such as endosomes and lysosomes. It also mediates
proton transport across the plasma membrane in some epithelial cells. For example,
V-ATPase in the apical membrane of intercalated cells of the renal distal tubule and
collecting duct secretes protons into the urine whereas V-ATPase in clear cells of the
epididymis acidify the epididymal fluid (Wagner, Finberg et al. 2004, Zuo, Huang et al.
2010). There is evidence for V-ATPase in airway epithelial cells, where it may be
required for sustained NADPH oxidase activity during infection (Fischer, 2011).
Bafilomycin A1 and concanamycin A are two potent macrolide inhibitors of V-ATPase.
They mainly bind to the c subunit of the V0 domain, thus preventing proton
translocation (Huss, Ingenhorst et al. 2002, Bowman, Graham et al. 2004).

11
1.4.3 NKCC
Sodium-potassium-chloride cotransporter (NKCC) is a member of the cation-chloride
cotransporter (CCC) superfamily (Haas and Forbush 2000, Russell 2000). It is
activated by cell shrinkage and participates in the regulation of cell volume. Besides
regulating the cell volume, NKCC maintains intracellular Cl- concentration ([Cl-]i)
above electrochemical equilibrium. The high [Cl-]i is used to drive net salt transport
in many epithelia. The cotransporter is expressed in the basolateral membrane of
polarized epithelial cells. Isoform 1 (NKCC1) is found in most epithelial organs except
the kidney, which express NKCC2 at the apical membrane (Gamba, Miyanoshita et al.
1994, Kaplan, Plotkin et al. 1996).
1.4.4 NBC
The sodium bicarbonate cotransporter (NBC) belongs to a superfamily of solute
carriers called the SLC4A family. NBC is mainly expressed in the basolateral
membrane of epithelial cells, and the dominant isoform in airway epithelial cells is
NBC1. The cotransporter helps regulate intracellular pH by transporting HCO3- into
the cell. The isoform in airway epithelial cells NBCe1 carries 1 Na+ and 3 HCO3- in the
same direction and therefore is electrogenic (Bernardo, Bernardo et al. 2006).
NBCe1 is inhibited by disulfonic stilbenes such as
4,4’-diisothiocyanostilbene-2,2’-disulfonic acid (DIDS) (Alpern 1985, Grassl and
Aronson 1986). The inhibition by disulfonic stilbenes occurs at the HCO3- interaction

12
site, and the amino acid motif that binds DIDS is KMIK and KLKK (Gross and Kurtz
2002). In addition to disulfonic stilbenes, NBCe1 is also inhibited competitively by
harmaline, which binds to the Na+ site (Soleimani and Aronson 1989).
1.4.5 The Na+-independent Cl-/HCO3- exchanger (AE)
Sodium independent Cl-/HCO3- exchangers also belong to the SLC4A family, and are
usually referred to simply as AEs (anion exchangers). AE isoform 2 (AE2) is widely
expressed, and in polarized epithelial cells it is usually located in the basolateral
membrane. AE2 carries out electroneutral exchange of Cl- and HCO3- in airway
epithelial cells (Dudeja, Hafez et al. 1999, Al-Bazzaz, Hafez et al. 2001), and the
directions of the fluxes depends on the chemical gradients of the two anions (Alper,
Chernova et al. 2002, Bonar and Casey 2008).
AE2 harbors covalent binding sites for disulfonic stilbenes, consequently these agents
inhibit AE2 and NBC1 unspecifically (Alper, Chernova et al. 2002).
1.4.6 Pendrin (SLC26A4)
Although pendrin, or SLC26A4, also functions as a Na+-independent Cl-/HCO3-
exchanger, it belongs to a different gene family called SLC26A, and is thus distinct
from SLC4A anion exchangers. Genetic defects in pendrin cause Pendred syndrome,
after which the exchanger was named (Wangemann 2011). Pendrin is expressed in
many different tissues, including the airways, where it localizes to the apical

13
membrane and exchanges HCO3- for other monovalent anions such as Cl-, I- or
formate (Scott, Wang et al. 1999, Scott and Karniski 2000, Pedemonte, Caci et al.
2007, Nakao, Kanaji et al. 2008).
Pendrin is predicted to have 12-15 transmembrane domains, and intracellular N- and
C- terminal domains (Mount and Romero 2004, Dossena, Rodighiero et al. 2009).
Motifs which are critical for anion transport are found within the hydrophobic core of
the transmembrane domains (Mount and Romero 2004). There is a STAS (sulfate
transporter and anti-sigma) domain at the C-terminus, which is believed to be
important for functional interactions with other proteins, including CFTR (Aravind
and Koonin 2000, Ko, Shcheynikov et al. 2002, Ko, Zeng et al. 2004).
Pendrin is inhibited by the chloride channel blocker NPPB
(5-Nitro-2-(3-phenylpropylamino) benzoic acid) and by the non-steroidal
anti-inflammatory drug niflumic acid (Dossena, Vezzoli et al. 2006, Pedemonte, Caci
et al. 2007).
1.4.7 The Na+/H+ exchanger (NHE)
NHE regulates intracellular pH and cell volume by exchanging 1 Na+ for 1 H+ and
normally transports Na+ in and H+ out. To date, six isoforms have been identified
(Fliegel 2005, Slepkov, Rainey et al. 2007). NHE1 is ubiquitously distributed and is the
only NHE isoform expressed basolaterally in airway epithelial cells (Dudeja, Hafez et

14
al. 1999, Al-Bazzaz, Hafez et al. 2001).
NHE1 is moderately sensitive to amiloride, a diuretic compound which acts mainly in
the distal nephron. To increase the potency and selectivity of this inhibitor for NHEs,
many derivatives have been synthesized such as DMA, EIPA and HMA. Some of these
novel inhibitors, notably benzoylguanidine and its derivatives, are more potent and
selective for NHE1 (Masereel, Pochet et al. 2003).
1.4.8 The H+ channel
An H+ channel in the apical membrane functions mainly in acid extrusion and is
activated by external alkaline pH and membrane depolarization (Cherny, Markin et al.
1995). So far, only one type of plasma membrane proton channel has been identified,
namely HVCN1. It has been found in many different cell types including airway
epithelial cells (DeCoursey 1991, DeCoursey and Cherny 1995, Fischer, Widdicombe
et al. 2002, Schwarzer, Machen et al. 2004, Iovannisci, Illek et al. 2010).
The most potent inhibitor of HVCN1 is Zn2+. It competitively binds to the external
surface of the proton channel, slowing the opening rate and shifting the voltage
dependence to more positive membrane potentials (Cherny and DeCoursey 1999).
1.4.9 The chloride channels
Different types of chloride channels have been found in airway epithelial cells

15
(Toczylowska-Maminska and Dolowy 2012). Among these channels, cystic fibrosis
transmembrane conductance regulator (CFTR) has been most intensively studied
because it is mutated in the common genetic disease cystic fibrosis (CF).
CFTR is apically localized in epithelia. It belongs to the ATPase-binding cassette (ABC)
transporter family, and is the only member of the family that functions as an ion
channel. It conducts both chloride and bicarbonate and has a Cl-:HCO3-
permeability ratio of 4:1 (Kim and Steward 2009). Two sets of six transmembrane
helices form transmembrane domains that contain the pore for anion flow. Each of
the membrane domains is followed by a nucleotide-binding domain (NBD), which
interacts with ATP. A central regulatory (R) domain links the N- and C-terminal
half-molecules. The R domain contains many sites for phosphorylation by protein
kinase A (PKA) and protein kinase C (PKC), and together with the two NBD s controls
the opening and closing rates of CFTR. Channel opening requires the binding of 2 ATP
molecules to the NBDs, which form a “nucleotide sandwich”, and phosphorylation of
R domain. Hydrolysis of ATP at NBD2 closes the channel gate, and dephosphorylation
of the R domain causes deactivation (Riordan 2008, Hwang and Sheppard 2009,
Lubamba, Dhooghe et al. 2012, Toczylowska-Maminska and Dolowy 2012). Besides
functioning as a chloride channel, CFTR may also exert regulatory effects on other
proteins, for example the epithelial sodium channel (ENaC) (Riordan 2008, Lubamba,
Dhooghe et al. 2012). The most potent and selective inhibitor of CFTR is the glycine
hydrazide GlyH-101

16
[N-(2-naphthalenyl)-((3,5-dibromo-2,4-dihydroxyphenyl)methylene)glycine hydrazide]
(Muanprasat, Sonawane et al. 2004, Sheppard 2004, Norimatsu, Ivetac et al. 2012). It
is voltage-dependent and therefore probably blocks the open CFTR pore. By contrast
the thiazolidinone CFTR(inh)-172
[3-[(3-trifluoromethyl)phenyl]-5-[(4-carboxyphenyl)methylene]-2-thioxo-4-thiazolidin
one] appears to inhibit within the bilayer by increasing the mean closed time and is
not voltage dependent (Kopeikin, Sohma et al. 2010).
The calcium activated chloride channel (CaCC) is another important chloride channel
which coexists with CFTR in the apical membrane of airway epithelial cells (Gabriel,
Makhlina et al. 2000). Its molecular identity is still under debate, but is probably
TMEM16A (Caputo, Caci et al. 2008, Schroeder, Cheng et al. 2008, Yang, Cho et al.
2008). Other chloride channels present in airway epithelial cells include volume
sensitive outwardly rectifying chloride channel (VSOR), bestrophins, ClC-2 channel
and outwardly rectifying chloride channel (ORCC) (Toczylowska-Maminska and
Dolowy 2012).
1.4.10 ENaC
The epithelial sodium channel ENaC is a member of the ENaC/Degenerin gene family,
which is comprised of channels with diverse functions including sodium transport
and mechanosensitivity (Kellenberger and Schild 2002). ENaC is located in the apical
membrane of epithelial cells and mediates Na+ transport across epithelia.

17
The functional unit of ENaC is probably a heterotrimer formed by α, β and γ subunits
(Kashlan and Kleyman 2011). It is interesting that in adult humans, rats and mice, all
the three ENaC subunits are highly expressed in small and medium-sized airways.
However, in the distal lung, only the α and γ subunits are expressed while the β
subunit is not found (Burch, Talbot et al. 1995, Farman, Talbot et al. 1997, Talbot,
Bosworth et al. 1999). The β- or γ-ENaC knockout mice died slightly later than the
α-knockout from severe hyperkalemia due to deficient renal K+ secretion. This
suggests that, in contrast to the kidney, Na+ transport in the lung can be maintained
efficiently by only two ENaC subunits, namely the α and β or the α and γ (Barker,
Nguyen et al. 1998, McDonald, Yang et al. 1999).
The channel blocker amiloride binds to residues in the TM2 domains of the subunits
(Kellenberger and Schild 2002, Kashlan and Kleyman 2011). These residues lie in the
outer vestibule of the channel pore, which has a diameter of ~ 5 Å, which
accommodates amiloride well (4-5 Å)
1.4.11 Carbonic anhydrase
Carbonic anhydrase is a zinc metalloenzyme which catalyzes the reversible hydration
of CO2. It is expressed ubiquitously in all living organisms, and 15 isoforms of
carbonic anhydrase have been identified in humans (Esbaugh and Tufts 2006,
Purkerson and Schwartz 2007). These isozymes can be divided into three groups, the

18
cytoplasmic CAs, the membrane-bound CAs, and CA-related proteins which have lost
their catalytic activity.
CAII is an important CA isoform, which has a high turnover rate of 106s-1 and
accounts for the majority of CA activity in different tissues (Khalifah 1971, Esbaugh
and Tufts 2006, Purkerson and Schwartz 2007). CAII is reported to bind to the
intracellular C-terminal region of AE1, AE2, NBC and NHE1 via its acidic motif in the
N-terminal region, forming a bicarbonate transport metabolon as discussed above
(McMurtrie, Cleary et al. 2004). The membrane-bound form of CA also has a high
turnover rate and may be of importance (Baird, Waheed et al. 1997, Wingo, Tu et al.
2001).
1.4.12 Regulation of ASL hydration
The hydration state of ASL is control by the mass of salt (NaCl) on the surface of
airways, with water flowing passively due to osmosis. Cl- secretion and Na+
absorption are important in regulating passive transepithelial water flow and thus
the height of the ASL. In this way, these ion transport processes are mainly
responsible for providing an optimal environment for ciliary beating (Hollenhorst,
Richter et al. 2011).
A large part of the Cl- secretion in the airways is mediated by apical CFTR (Riordan
2008, Lubamba, Dhooghe et al. 2012). In addition to CFTR, CaCC are also present on

19
the apical side and contribute to inflammatory secretion (Toczylowska-Maminska and
Dolowy 2012). NKCC and AE2 have been detected in the basolateral membrane of
airway epithelial cells, and supply Cl- from the basolateral side (Tessier, Traynor et al.
1990, Al-Bazzaz, Hafez et al. 2001). A wide variety of K+ channels are expressed in
airway epithelial cells, and several have been identified in the basolateral membrane
(Bardou, Trinh et al. 2009). Though these K+ channels are not directly involved in Cl-
secretion, they regulate the membrane potential and help to maintain the
electrochemical gradient favoring apical Cl- secretion (Bardou, Trinh et al. 2009,
Hollenhorst, Richter et al. 2011).
In terms of Na+ transport, the amiloride-sensitive ENaC contributes to apical Na+
absorption in the airways (Burch, Talbot et al. 1995), whereas on the basolateral side,
the Na+/K+ ATPase moves Na+ out of the cells and provides the driving force for apical
Na+ absorption (Hollenhorst, Richter et al. 2011). Thus, the Na+/K+ ATPase and ENaC
work in concert to mediate Na+ absorption in the airways. Besides acting as a
chloride channel, CFTR also regulates ENaC. In normal airways, CFTR inhibits ENaC
thus preventing Na+ absorption; however, in CF airways, ENaC becomes hyperactive
because of the defective CFTR (Matsui, Grubb et al. 1998, Boucher 2007, Boucher
2007). Therefore, ASL dehydrates under Na+ hyperabsorption.
1.4.13 pH regulation of ASL
Normal ASL pH is slightly acidic, averaging around 6.6-6.9 (Fischer and Widdicombe

20
2006). The ASL pH of CF patients becomes even more acidic, suggesting a breakdown
in the ASL pH regulation (Coakley, Grubb et al. 2003, Song, Salinas et al. 2006).
HCO3- is considered a weak base and may be the main determinant of the pH and
buffer capacity of the ASL (Song, Salinas et al. 2006). Under unstimulated conditions,
HCO3- has an outward net driving force across the apical membrane of airway
epithelial cells due to the negative membrane potential, and contributes to the anion
efflux through CFTR despite having lower permeability and conductance ratios
relative to Cl- (Fischer and Widdicombe 2006). The idea that CFTR alkalinizes
submucosal gland secretions is suggested by the reduced pH of CF gland secretions
(Coakley, Grubb et al. 2003, Song, Salinas et al. 2006).
NBCe1 and carbonic anhydrase are presumed to supply the HCO3- for secretion into
the lumen via CFTR (Smith and Welsh 1992, Devor, Singh et al. 1999, Devor, Bridges
et al. 2000, Krouse, Talbott et al. 2004, Ballard, Trout et al. 2006). NBCe1 supplies
HCO3- from the basolateral side whereas carbonic anhydrase catalyzes the
generation of endogenous HCO3-.
As discussed above, AE2 is also found basolaterally in airway epithelial cells (Dudeja,
Hafez et al. 1999, Al-Bazzaz, Hafez et al. 2001) but may move HCO3- out of the cell in
exchange of Cl-. Thus it would reduce the electrochemical driving force for apical
HCO3- exit via CFTR and tend to acidify the ASL (Fischer and Widdicombe 2006). It

21
should be noted however that this scheme is controversial (Garnett et al.)
Although HCO3- secretion via CFTR would cause alkalinization of the ASL, the fact
that normal ASL pH is consistently acidic (Fischer and Widdicombe 2006) indicates
that airway surface epithelial cells also secrete acid.
Other apical transporters also impact ASL pH, and some of these have been
discussed above. For example, The voltage-gated H+ channel was demonstrated in
the apical membrane of airway epithelial cells (DeCoursey 1991, DeCoursey and
Cherny 1995, Fischer, Widdicombe et al. 2002, Schwarzer, Machen et al. 2004,
Iovannisci, Illek et al. 2010). In some studies, blocking this channel by Zn2+ showed
50-70% inhibition on H+ secretion by airway epithelial cells (Fischer, Widdicombe et
al. 2002, Schwarzer, Machen et al. 2004), suggesting that in these studies the primary
acid secretion mechanism was mediated by the H+ channel. An H+,K+-ATPase is
present in the apical membrane of airway epithelial cells (Coakley, Grubb et al. 2003)
and blocking it for several hours by apical exposure to ouabain inhibits ASL
acidification, although the acute effects are small (Fischer, Widdicombe et al. 2002,
Coakley, Grubb et al. 2003, Krouse, Talbott et al. 2004, Schwarzer, Machen et al. 2004,
Song, Salinas et al. 2006). Another acid secreting transporter is the V-ATPase (Inglis,
Wilson et al. 2003). Its inhibitor bafilomycin A1 reduces ASL acidification to a variable
extent (0-60%) (Poulsen and Machen 1996, Fischer, Widdicombe et al. 2002, Inglis,
Wilson et al. 2003, Schwarzer, Machen et al. 2004, Song, Salinas et al. 2006). Though

22
NHE1 is expressed in the basolateral membrane and mainly participates in regulating
intracellular pH, it has been proposed that NHE1 could affect the apical H+ channel by
intracellular pH regulation, because the voltage-gated H+ channel can be activated by
intracellular acidification (Fischer and Widdicombe 2006). It remains unclear why
different acid transporters and channels have been identified as the primary H+
secretion pathway in airways by different investigators.
1.5 Models of anion transport in submucosal glands
Submucosal glands are important to airway defense, producing mucus and fluid in
response to neural signals (Maggi, Giachetti et al. 1995). The density of glands is
about 1 per mm2 in the trachea and becomes more abundant as the generation
grows. Each gland consists of multiple tubules that merge into a collecting duct, and
continue through a ciliated duct which eventually leads to the airway surface (Tos
1966, Meyrick, Sturgess et al. 1969). The abundant serous cells secrete water and
electrolytes, and a rich mixture of antimicrobial, anti-inflammatory, and antioxidant
compounds. CFTR is expressed in these cells according to most studies (Basbaum,
Jany et al. 1990, Engelhardt, Yankaskas et al. 1992).
To understand how serous cells regulate ASL, several groups have used submucosal
glands from human or animals and the model serous cell line Calu-3 to study anion
transport. Using Calu-3 cells, a scheme for anion transport of serous cells was
proposed which is widely accepted (Devor, Singh et al. 1999). In this cellular model,

23
cells secrete mostly HCO3- in response to elevated [cAMPi] by forskolin. HCO3
- is
taken up by cells via basolateral NBC, and then exits to the lumen through CFTR.
However, according to the model, secretion by serous cells can shift from HCO3- to Cl-
under certain conditions, for example during cell hyperpolarization, which favors the
electroneutral basolateral NKCC and thus causes the cells to secret Cl- (Fig. 1.1).
Another group made some modifications based on this model (Cuthbert and
MacVinish 2003). Though they confirmed the shift from HCO3- to Cl- secretion under
hyperpolarization, they emphasized that Cl- secretion was HCO3- dependent and the
role of carbonic anhydrase in generating HCO3-. They also suggested that carbonic
anhydrase (isoform 2) might form complexes with baolateral NBC, NHE and anion
exchanger to facilitate the transport of both HCO3- and Cl- (Fig. 1.2).

24
Figure 1.1 Model for anion transport by Calu-3 cells proposed by Devor
et al.. HCO3
- or Cl- is secreted through cAMP-stimulated apical CFTR channels according to the basolateral membrane potential (ψbl) and reversal potential of the sodium-bicarbonate cotransporter (ErevNaHCO3). Basolateral Cl- entry mediated by the sodium–potassium chloride cotransporter NKCC1 is favoured when the membrane potential is larger (i.e. more negative) than ErevNaHCO3. Basolateral HCO3
- entry via sodium-bicarbonate cotransporter NBC1 is favoured when the basolateral membrane potential is smaller (i.e. more positive) than ErevNaHCO3. 1-EBIO hyperpolarizes the basolateral membrane by activating charybdotoxin (CTX)-sensitive K+ channels. Bumetanide, ouabain and DNDS reduce secretion by inhibiting NKCC1, Na+/K+ ATPase and NBC1 respectively. Reproduced from Devor et al. 1999.

25
Figure 1.2 Model for anion transport by Calu-3 cells proposed by
Cuthbert et al.. This model is similar to the one proposed by Devor et al., except that besides NKCC1, carbonic anhydrase II (CAII) is also important to Cl- tranport. When the basolateral membrane is hyperpolarized by activating K+ channel, CAII converts CO2 to HCO3
-, which exchanges for Cl- via AE2. AE2 together with NKCC1 provides Cl- for secretion through apical CFTR. The generation of H+ by CAII conversion is extruded by NHE1 to maintain pHi. HCO3
- may activate soluble adenylyl cyclase (sAC), thus increaseing [cAMPi] and activating CFTR. Reproduced from Cuthbert et al. 2003.

26
When the above scheme was tested on submucosal glands of human and animal
models, some contradictions were found. When native glands were stimulated by
elevating [cAMPi], 1) pH of secretions from the gland was not as high as expected; 2)
the major anion in the secreted fluid was chloride rather than bicarbonate
(Jayaraman, Joo et al. 2001, Joo, Saenz et al. 2002). Bumetanide, an inhibitor of NKCC,
reduced gland secretion though not as well as did removal of HCO3-. These finding
led to the conclusion that both HCO3- and Cl- were important to gland secretion
when [cAMPi] was increased and CFTR was activated (Wine and Joo 2004) (Fig. 1.3).
All of the above studies were consistent with CFTR being the main apical exit
pathway for anion secretion, however a recent study suggested a different view
(Garnett, Hickman et al. 2011). Based on pHi measurements and fluid secretion
assays, Garnett et al. (2011) proposed that when [cAMPi] is increased, CFTR secretes
chloride while bicarbonate exits via apical pendrin in exchange for luminal chloride
(Fig. 1.4).
It has been almost 25 years since the cftr gene was cloned (Kerem, Rommens et al.
1989, Riordan, Rommens et al. 1989, Rommens, Iannuzzi et al. 1989). Nevertheless,
how anions are transported by serous cells and the role of CFTR in such process
remain controversial. The development of effective therapies for cystic fibrosis may
require an understanding of the mechanisms of anion and fluid secretion.

27
Figure 1.3 Model for anion and fluid secretion by submucosal glands
proposed by Joo et al.. Pathways that elevate intracellular Ca2+ level, such as acetylcholine (ACh), are hypothesized to activate fluid and macromolecular secretion from both serous and mucous cells. Pathways that elevate intracellular cyclic AMP level, such as VIP, are hypothesized to stimulate serous cells and mucin but not fluid secretion from mucous cells. The serous cells secrete mostly Na+, Cl- and water, and a small amount of HCO3
- comparied to Cl-. The CFTR-dependent fluid-secreting pathway is defective in CF glands. Reproduce from Joo et al., 2002.

28
Figure 1.4 Model for anion transport by Calu-3 cells proposed by
Garnett et al.. A, under non-stimulated conditions, Cl− is accumulated across the basolateral membrane by the combined action of the basolateral Na+-K+-2Cl− cotransporter, NKCC1, together with parallel activity of the NBC and AE2. Under these conditions, a basal level of CFTR activity drives a small amount of a Cl−-rich secretion with a pH of ~7.4 (25 mm HCO3
−) via electrogenic Cl− efflux through CFTR. B, elevation of cAMP inhibits basolateral AE2 and stimulates NBC. At the apical membrane, cAMP/PKA further increases CFTR activity as well as activates pendrin (SLC26A4). Stimulation of CFTR activity leads to a marked rise in net transepithelial fluid secretion driven by electrogenic Cl− efflux through CFTR, whereas the co-activation of pendrin increases the HCO3
− content of this secreted fluid to ~75 mm (pH 7.9), through coupled Cl−/HCO3
− exchange, with Cl− cycling across the apical membrane via CFTR and SLC26A4. Reproduced from Garnett et al., 2011.

29
Chapter 2 Anion transport in
airway epithelial cells

30
Previous studies by others showed that the activator of adenylyl cyclase, forskolin,
increases intracellular cAMP, stimulates PKA-phosphorylation, and activates CFTR in
Calu-3 cells. Under short-circuit current conditions, CFTR was proposed to secrete
mainly bicarbonate. However, our preliminary data revealed that the fluid secreted
by forskolin stimulated Calu-3 cell monolayers contained only ~25 mM bicarbonate.
In this chapter we investigated this and other discrepancies between our data and
previous studies. The result have led us to proposed a revised model for anion
transport by Calu-3 cells which may be applicable to airway submucosal gland serous
cells.

31
2.1 Abstract
Anion transport drives fluid onto the airway surface to supply airway surface liquid
for humidifying inspired air and also for mucociliary clearance. This anion transport is
defective in airway diseases such as cystic fibrosis. Despite the importance of anion
transport in airway epithelium, its cellular mechanisms remain poorly understood
and controversial. We studied Cl- and HCO3- fluxes across the human airway cell line
Calu-3 and a genetically matched CFTR-deficient cell line. Forskolin stimulated the
short-circuit current (Isc) across voltage-clamped monolayers, and the equivalent
short-circuit current (Ieq) calculated from the transepithelial voltage and resistance of
monolayers kept under open-circuit conditions. Isc was equivalent to the HCO3- flux
measured using the pH-stat technique. Ieq equaled the sum of net 36Cl- and HCO3-
fluxes, although Ieq and HCO3- were underestimated since both were increased by
bafilomycin and ZnCl2, inhibitors of electrogenic H+ secretion. Ieq was dependent on
the presence of Na+ and HCO3-. Forskolin-stimulated Ieq and HCO3
- secretion were
both rapidly abolished by the carbonic anhydrase inhibitor acetazolamide, indicating
that Na+,HCO3- cotransport is secondary to carbonic acid synthesis. Anion exchange is
the main mechanism of basolateral Cl- loading in these cells, consistent with the
weak (~20%) inhibition by bumetanide. Apical anion efflux was probably mediated by
CFTR channels because 1) a forskolin-stimulated current appeared after imposing a
transepithelial HCO3- gradient across basolaterally permeabilized monolayers, 2) the
HCO3- current was sensitive to CFTR inhibitors and drastically reduced in
CFTR-deficient cells, and 3) the net secretory HCO3- efflux was increased 25% in

32
bilateral Cl--free solutions, excluding a dependence on apical anion exchange. The
results suggest a model in which HCO3- recycles across the basolateral membrane in
exchange for Cl-, and the resulting HCO3--dependent supply of Cl- is secreted via CFTR
together with remaining intracellular HCO3-.
2.2 Introduction
The airways are protected by a microscopic layer of airway surface liquid (ASL), which
humidifies the air during inspiration and participates in host defense against inhaled
pathogens. Anion transport in airway epithelium is responsible for driving fluid into
the airway surface to form the ASL, and transport defects in CF impair the depth of
normal ASL (Widdicombe 2002, Tarran, Button et al. 2006, Boucher 2007), leading to
reduced mucociliary clearance and recurring infection and inflammation. Despite its
importance in innate defense and pathophysiology, the mechanisms of airway anion
transport remain poorly understood.
The Cystic Fibrosis Transmembrane conductance Regulator (CFTR) is the product of
the CF gene and plays a critical role in airway secretion. It is a
phosphorylation-regulated, non-rectifying anion channel of ~7 pS conductance which
determines the rate of secretion by many epithelia (Klyce and Wong 1977) including
those in the airways. The channel pore is permeable to Cl- and HCO3- (Gray, Pollard et
al. 1990, Poulsen, Fischer et al. 1994, Linsdell, Tabcharani et al. 1997), and secretion
of both anions is defective in CF (Widdicombe, Welsh et al. 1985, Smith and Welsh

33
1992). However, the relative contributions to apical HCO3- efflux of CFTR, or of
CFTR-regulated anion exchangers, remains controversial (Lee, Choi et al. 1999,
Ishiguro, Steward et al. 2009, Kim and Steward 2009, Garnett, Hickman et al. 2011).
Anion secretion has been studied extensively in the human airway cell line Calu-3,
which differentiates spontaneously into a predominant population of serous-like cells
which express CFTR and antimicrobial proteins, and a smaller population of
mucous-like cells which contain mucin granules. When cultured on porous supports
at the air-liquid interface, Calu-3 monolayers generate robust basal short-circuit
current (Isc) that is not explained by net 36Cl- or 22Na+ fluxes (Haws, Finkbeiner et al.
1994, Shen, Finkbeiner et al. 1994, Shan, Huang et al. 2011) and was therefore
suggested to be generated by active HCO3- secretion (Lee, Penland et al. 1998).
Forskolin stimulated both unidirectional 36Cl- fluxes across short-circuited Calu-3 cells
without inducing measureable net Cl- secretion, and the Isc was insensitive to the
NKCC1 inhibitor bumetanide (Devor, Singh et al. 1999). Based on these findings it was
suggested that the forskolin-stimulated Isc was due to electrogenic HCO3- transport
(Devor, Singh et al. 1999), and this was subsequently confirmed using the pH stat
technique (Krouse, Talbott et al. 2004).
Several cellular models have been proposed to explain transport by Calu-3
monolayers. According to one scheme, HCO3- transport occurs by Na+ coupled entry
at the basolateral membrane followed by diffusional exit through apical CFTR

34
channels (Devor, Singh et al. 1999). This scheme predicts the secretion of HCO3--rich
fluid during cAMP stimulation, and perhaps Cl- rich fluid during stimulation by
secretagogues that hyperpolarize the cell (based on unidirectional 36Cl- fluxes
measured during stimulation with the potassium channel activator 1-EBIO). Another
hyperpolarizing agonist, 7,8-benzoquinoline, was also found to stimulate Cl- secretion,
but in that study Cl- secretion was dependent on carbonic anhydrase activity and was
proposed to involve anion exchange (AE2)-mediated Cl- entry at the basolateral
membrane in parallel with NKCC (Cuthbert and MacVinish 2003). The relationship
between fluid secretion and anion transport remains uncertain. Elevated [HCO3-] was
observed in Calu-3 secretions (Irokawa, Krouse et al. 2004), however
cAMP-stimulated fluid secretion driven by a flow of bicarbonate is difficult to
reconcile with the pH of airway secretions, which are near neutrality or even slightly
acidic (Kyle, Ward et al. 1990, Coakley, Grubb et al. 2003, Fischer and Widdicombe
2006, Song, Salinas et al. 2006).
We studied HCO3- transport under open- and short-circuit conditions using an
automated pH stat apparatus and tracer fluxes. Under pH stat conditions we found
that Isc was stimulated by forskolin and was identical to the net HCO3- flux measured
simultaneously, and that the HCO3- current was probably mediated by CFTR as
previously suggested (Ballard, Trout et al. 1999, Devor, Singh et al. 1999). Unlike Isc,
the Ieq under open-circuit conditions was the sum of net 36Cl- and HCO3- fluxes, and
was dependent on both Na+ and HCO3-. Forskolin-stimulated Ieq and HCO3
- secretion

35
were both abolished by the carbonic anhydrase inhibitor acetazolamide, indicating
an important role of carbonic anhydrase in anion transport. We found that anion
exchange but not NKCC is the main mechanism of basolateral Cl- loading, which
probably explains the weak (~20%) inhibition by bumetanide. Under pH-stat
conditions, we found evidence that apical anion efflux is mediated by CFTR channels.
The results suggest a revised model for Calu-3 cell anion transport in which Cl-
transport is dependent on HCO3- via basolateral anion exchange, and most of the
HCO3- is synthesized intracellularly by carbonic anhydrase or enters the cell
basolaterally via NBCe1.
2.3 Methods
2.3.1 Cell Culture
We used two modified Calu-3 cell lines for this study, a CFTR knock-down cell line
which stably expresses a 21-mer shRNA specifically targeting CFTR mRNA, and a
control cell line expressing shRNA with four mutations that reduce its ability to
silence cftr (Sizt and Alter cell lines, respectively) (Palmer, Lee et al. 2006). Both cell
lines were cultured in EMEM containing 7% FBS. Some control experiments were also
performed using parental Calu-3 cells (HTB-55, American Type Culture Collection,
Manassas, VA) cultured in Eagle’s minimum essential medium (EMEM) containing 15%
fetal bovine serum (FBS) to allow comparison with previous studies.
Cells were seeded at 5 x 105 cells per cm2 on SnapwellsTM (1.12 cm2; Costar, Corning

36
Life Sciences, Lowell, MA) for measuring short-circuit current (Isc) and HCO3-
secretion. Fresh medium was placed on the basolateral side one day after plating,
and the apical medium was removed to establish air-liquid interface (ALI) conditions.
Any fluid that appeared spontaneously on the apical surface was removed after 3
days and cultures were maintained in a humidified, 5% CO2 incubator at 37℃ for
21-25 days. Polarization of the monolayers with respect to CFTR was confirmed by
imposing a transepithelial Cl- gradient and measuring current stimulated by forskolin
after permeabilization of the basolateral or apical membrane with nystatin (100
μg·ml-1).
2.3.2 Solutions
To measure HCO3- secretion, monolayers were mounted in modified Ussing
chambers. The apical surface was bathed with unbuffered solution containing
(mmol·l-1): 120 NaCl, 5 KCl, 1.2 MgCl2, 1.2 CaCl2, and the basolateral side was bathed
with 120 NaCl, 25 NaHCO3, 3.3 KH2PO4, 0.8 K2HPO4, 1.2 MgCl2, 1.2 CaCl2, and 10
glucose. Nominally Na+-free solution was prepared by replacing NaHCO3 and choline
chloride, and NaCl with N-methyl-D-glucamine chloride. Nominally Cl--free solution
was prepared by replacing Cl- with gluconate and increasing the [Ca2+] from 1.2 to 4
mmol·l-1 to compensate for gluconate binding. Nominally HCO3--free solution was
prepared by replacing NaHCO3 with NaHEPES. All solutions were adjusted to pH 7.4.
The apical side was stirred with 100% O2 while the basolateral side was bubbled with
95% O2/5% CO2. Both sides were gassed with 100% O2 during experiments with

37
bilateral HCO3--free solution.
Tracer fluxes were measured using the same solutions as for pH-stat experiments. In
some experiments the basolateral membrane was permeabilized by adding nystatin
from a 1,000 x stock solution in DMSO (final nystatin concentration 100 μg·ml-1). The
CFTR inhibitor GlyH-101 was added from a 1,000 x stock solution to give a final
concentration of 100 μmol·l-1 and 0.1% DMSO.
2.3.3 Immunoblotting
After SDS-PAGE on 8% gels, proteins were transferred to nitrocellulose membranes as
described previously (Luo, McDonald et al. 2009) and probed using monoclonal
antibodies: M3A7, which recognizes an epitope between amino acids 1365 and 1395
in the second nucleotide binding domain of CFTR (1:5000, gift from J.R. Riordan and
T.J. Jensen, UNC Chapel Hill, NC) (Kartner and Riordan 1998), TUB-1A2, which binds
the C-terminus of α-tubulin (1:5000, Sigma), and α5, which binds the α-subunit of
avian Na+/K+-ATPase (1:200, mAb gift from R.W. Mercer, Washington Univ., St. Louis
MO) (Takeyasu, Tamkun et al. 1988). Blots were washed, incubated with secondary
antibody conjugated to horseradish peroxidase (1:1000), visualized with enhanced
chemiluminescence (Amersham Biosciences), and analysed using ImageJ (Rasband,
2011).
2.3.4 Measurements of equivalent Isc and HCO3- secretion

38
Inserts were mounted in modified Ussing chambers (Physiologic Inst., San Diego, CA)
at 37℃ and initial studies were carried out under voltage clamp to allow comparison
of net HCO3- transport with Isc. In subsequent experiments HCO3
- secretion was
measured under open-circuit conditions, and was compared with the equivalent
short-circuit current (Ieq) calculated from Ohm’s law using the spontaneous
transepithelial potential (Vt) and transepithelial resistance (Rt). Rt was determined
from small deflections in Vt produced by bipolar current pulses (1 μA, 1 sec duration,
99.9 sec interval) delivered by voltage clamp (VCC200, Physiologic Instr.). Data were
digitized (Powerlab 8/30, AD Instruments, Montreal QC) and analyzed using Chart5
software.
HCO3- transport was measured using the pH-stat method under both open- and
short-circuit conditions. A mini-pH electrode (pHG200-8, Radiometer Analytical)
connected to the automated titration workstation (TitraLab 854, Radiometer)
delivered 1 μl aliquots of 10 mmol·l-1 HCl or 5 mmol·l-1 H2SO4 to maintain the pH
constant at 7.400 ± 0.002. The amount of acid required for this was used to calculate
the rate of HCO3- secretion. The volume of each half chamber was 4 ml. Solutions
containing 25 mmol·l-1 HCO3- were stirred vigorously with 95% O2/5% CO2.
Nominally HCO3--free solutions were bubbled with 100% O2.
2.3.5 Tracer fluxes
Calu-3 monolayers were mounted in modified Ussing chambers and equilibrated for

39
20 min. H36Cl (generous gift of W.S. Marshall, St. Francis-Xavier University, Antigonish
NS) was added to one side of the monolayer and neutralized, yielding a final Cl-
concentration of 125.3 mmol·l-1. After 20 min of mixing, duplicate 400 μl samples
were taken from the cold side at 15 min intervals and replaced with 800 μl saline.
Residual radioactivity was calculated, with correction for dilution, at the beginning of
each subsequent flux period. Samples were counted in 5 ml scintillation solution
(Tri-Carb 2810 TR, PerkinElmer, Woodbridge ON). Two 10 μl samples were taken from
the hot side at the beginning and end of the experiment and averaged to calculate
mean specific activity. Both unidirectional fluxes were measured in parallel
experiments performed at the same time using monolayers with similar Rt.
2.3.6 Data analysis
Isc or Ieq was determined at 100 sec intervals, and HCO3- net flux rate was calculated
every 5 min. Basal values were those obtained immediately before adding forskolin.
Steady-state fluxes during stimulation were calculated 30 - 60 min after forskolin
addition. Paired or unpaired student’s t-tests with p<0.05 were used for single
comparisons. One-way analysis of variance followed by the Bonferonni post-hoc test
was used for multiple comparisons
2.4 Results
2.4.1 HCO3- secretion accounts for the short-circuit current (Isc)
Electrogenic HCO3- secretion across control monolayers was measured under

40
voltage-clamp and compared with the Isc measured simultaneously. Unstimulated
HCO3- secretion and Isc were both low but increased ∼8-fold after forskolin (10
μmol·l-1) was added to the basolateral side (Fig. 2.1). The rate of HCO3- secretion
measured using pH-stat with the transepithelial potential clamped at 0 mV was equal
to the Isc within measurement error (net HCO3- secretion: 2.14 ± 0.36 μeq·cm-2·h-1; Isc:
2.30 ± 0.27 μeq·cm-2·h-1; Fig. 2.1), evidence that basal and forskolin-stimulated Isc
were both due to net HCO3- secretion. This confirms previous suggestions based on
the large discrepancy between Isc and the net 36Cl- and 22Na+ fluxes (Lee, Penland et
al. 1998, Devor, Singh et al. 1999), but differs from pH-stat results obtained during
1-EBIO stimulation, when HCO3− secretion accounted for 70% of the Isc and the
discrepancy was due to acid secretion by H+/K+-ATPase (Krouse, Talbott et al. 2004).
The effects of 1-EBIO were not examined in the present study. The pH-stat technique
required unbuffered solution on the apical side and therefore a transepithelial HCO3-
gradient; however, the basolateral-to-apical gradient of 25 mmol·l−1 HCO3- did not
cause overestimation of HCO3- secretion since the Isc was not noticeably affected
when 25 mol·l-1 HCO3-/5% CO2 was added acutely on the apical side (data not
shown), consistent with a previous report (Krouse, Talbott et al. 2004). The
stimulated Isc was similar whether symmetrical HCO3- solutions or pH-stat conditions
were used, further evidence that passive diffusion due to the HCO3- gradient
contributed little to Isc.
2.4.2 HCO3- secretion is CFTR dependent

41
To evaluate the role of CFTR during HCO3- transport, protein expression and anion
secretion were compared in control and CFTR knock-down monolayers. The
Figure 2.1 Relationship between short-circuit current (Isc) and HCO3-
secretion. Forskolin (10 μmol·l-1) and GlyH-101 (100 μmol·l-1) were added to control Calu-3 cells stably expressing shRNA that was mutated to inhibit targeting of CFTR mRNA transcripts. Note the close agreement between Isc and HCO3
- secretion and their sensitivity to the CFTR channel inhibitor GlyH-101 (n=4).

42
immunoblot in Fig. 2.2A shows CFTR expression. For comparison, Lanes 1 and 2 show
baby hamster kidney (BHK) cells stably transfected with wild-type or ΔF508-CFTR,
Lanes 3 and 4 show parental Calu-3 cells, and Lanes 5 and 6 show shRNA control and
CFTR knock-down Calu-3 cells, respectively.
BHK cells expressed mature (i.e. complex glycosylated or ‘band C' polypeptide) and
immature (core-glycosylated, band B) wild-type CFTR whereas only band B was
detected in cells expressing ΔF508 CFTR (Lanes 1 and 2, respectively). Mature and
immature CFTR glycoforms were also found in parental cells (Lanes 3 and 4), and in
cells expressing control (i.e. altered) shRNA (Lane 5). CFTR was usually not detectable
in cells that expressed shRNA targeting CFTR transcripts, indicating efficient
knock-down (Lane 6). Blots were also probed with anti-tubulin antibody to control
for variations in protein loading, and with anti-Na+/K+-ATPase antibody to assess
non-specific changes in membrane protein expression. Tubulin and Na+/K+-ATPase α
subunit levels were not affected by control or CFTR-specific shRNA. Densitometry of
CFTR expression revealed >95% reduction in band C compared with the control cell
line (Fig. 2.2A and inset), consistent with previous estimates (Palmer, Lee et al. 2006).
Band C was lower in shRNA control cells than in parental cells after normalization to
tubulin, despite the presence of four mutations in the shRNA which were intended to
reduce its binding to CFTR mRNA transcripts. Cells expressing control shRNA

43
(referred to previously as ‘alter' by (Palmer, Lee et al. 2006) were used as the control
cell line in subsequent transport experiments, bearing in mind that CFTR expression
Figure 2.2 CFTR expression and HCO3- secretion.
A, CFTR expression in stable cell lines. Lane 1, baby hamster kidney (BHK) cells transfected with wild-type CFTR expressed both mature and immature CFTR. Lane 2, BHK cells transfected with ΔF508 CFTR expressed only immature (band B) CFTR. Lanes 3,4 parental, and Lane 5 control transfected Calu-3 cells also expressed both band B and C. CFTR was not detectable in shRNA knock down cells. Comparison of μg band intensities and protein loaded indicates the following relative expression levels: BHK >> Calu-3 (Parental) > Calu-3 (control transfected) >> Calu-3 (shRNA transfected). B, The CFTR expression is compared to tubulin according to A. C, forskolin (10 μmol·l-1) stimulated Ieq and HCO3
- secretion across wild-type Calu-3

44
cells under open-circuited conditions (n=12). D, CFTR inhibitor GlyH-101 (100 μmol·l-1) nearly abolished the forskolin-stimulated Isc and HCO3
- secretion (n=6). E, the forskolin response was greatly reduced in CFTR-knockdown Calu-3 cells (n=6). F, Summary of forskolin-stimulated Ieq and HCO3
- secretion in B, C and D. ● Ieq; ○ HCO3
- secretion. Means±s.e.m.

45
is lower in these control cells than in parental Calu-3 cells that have been used in
previous studies published by others.
The effect of knocking down CFTR expression on HCO3- secretion was also
investigated under open-circuit conditions using the pH-stat technique (Fig. 2.2B-D).
In marked contrast to Isc, the Ieq calculated from transepithelial voltage and
resistance was higher than the simultaneously measured HCO3- flux; mean Ieq was
2.76 ± 0.13 μeq·cm-2·h-1 (n = 12) during forskolin stimulation whereas net HCO3-
transport was 1.14 ± 0.05 μeq·cm-2·h-1 (n = 12, Fig. 2.2B,C). This difference between
Ieq and net HCO3- flux (∼1.55 μeq·cm-2·h-1) was due to Cl- current because, as shown
below (Table 2.1), a net Cl- flux of 1.5 μeq·cm-2·h-1 in the secretory direction was
observed when unidirectional 36Cl fluxes were measured under these conditions (i.e.
open-circuited and with the same 25 mmol·l-1 HCO3- gradient that was present
during pH-stat experiments). Also, the discrepancy between Ieq and net HCO3- flux
was abolished in Cl--free solution. Ieq and net HCO3- secretion were both inhibited by
the CFTR channel blocker GlyH-101 (Fig. 2.2C); however, this inhibition developed
slowly (100 μmol·l-1; ∼20 min, see Fig. 1 and 2D in Muanprasat et al. 2004), perhaps
due to slow diffusion through the hydrophobic mucus gel on the apical surface.
Alternatively, GlyH-101 inhibition is voltage dependent therefore its affinity may
decline as CFTR channels become progressively blocked and the membrane potential

46
hyperpolarizes towards the potassium equilibrium potential EK. The stimulation of Ieq
and HCO3- secretion by forskolin was greatly reduced in CFTR knock-down cells
compared with
Table 2.1 Unidirectional 36Cl fluxes across Calu-3 cell monolayers.
Experiments were performed under open-circuit, pH-stat conditions (i.e. basolateral 25 mM HCO3
-, apical 0 mM HCO3-). The apical solution was stirred with 100% O2 and
buffered at pH 7.4 using 10 mmol·l-1 tricine.
Jsm Jms Jnet Ieq RT VT
control 1.10 ± 0.12 0.53 ± 0.11 0.58 ± 0.06 0.81 ± 0.06 375 ± 47 -8.14 ± 0.68
forskolin 3.13 ± 0.13 ***
1.63 ± 0.15 ***
1.51 ± 0.02 ***
2.75 ± 0.13 ***
289 ± 30 ***
-21.31 ± 0.97 ***
bumetanide 1.95 ± 0.08 +++
1.22 ± 0.06 +++
0.73 ± 0.05 +++
2.15 ± 0.10 +++
301 ± 32 +
-17.35 ± 0.64 +++
Jsm, Jms, Jnet , Ieq: μEquiv·cm-2·h-1, Rt: Ohms·cm2·h-1, n = 4, Mean±s.e.m. ***, P<0.001, forskolin vs. control. +++, P<0.001; +, P<0.05, bumetanide vs. forskolin

47
control cells (compare Fig. 2.2D and B), although the decrease in CFTR channel
function was somewhat less than the decrease in CFTR protein expression observed
on immunoblots (65% vs. 95% reduction). Figure 2E summarizes the mean Ieq (filled
bars) and HCO3- flux (open bars) across control vs. CFTR-deficient cells. HCO3
- was
secreted by CFTR knock-down cells during forskolin stimulation at a rate that was
similar to unstimulated control monolayers, and to stimulated control monolayers
treated with the CFTR inhibitor GlyH-101. Taken together the results confirm that
forskolin stimulates electrogenic HCO3− secretion and are consistent with CFTR
channels mediating a large part of this flux.
The stimulated Ieq under open-circuit, pH-stat conditions in Fig. 2C was 2.55
μeq·cm-2·h-1, higher than the HCO3- flux of 1.0 μeq·cm-2·h-1 measured simultaneously.
To examine if the discrepancy of ∼1.55 μeq·cm-2·h-1 is carried by Cl-,
unidirectional 36Cl- tracer fluxes were measured under the same conditions, i.e. open
circuited and with a basolateral → apical HCO3- gradient and apical O2 bubbling.
Apical pH was maintained by 25 mM HEPES during tracer experiments rather than by
titration with acid. As shown in Table 2.1, net Cl- secretion (Jnet) was observed under
basal conditions. Forskolin increased 36Cl- fluxes in both directions, but stimulation of
the secretory flow was larger and resulted in a 3-fold increase in the net flux. Thus,

48
Cl- was secreted under open circuit/pH-stat conditions (in marked contrast to Isc
conditions), and the net flux rate (1.51 ± 0.02 μeq·cm-2·h-1) accounted for the
discrepancy of 1.55 μeq·cm-2·h-1 between Ieq and net HCO3- flux measured using the
pH-stat.
2.4.3 Proton secretion neutralizes some of the transported bicarbonate
Acid secretion was also investigated because, if present, it could cause
underestimation of the HCO3- flux. Airway epithelial cells secrete H+ by multiple
mechanisms (Acevedo and Steele 1993, Fischer, Widdicombe et al. 2002, Coakley,
Grubb et al. 2003, Inglis, Wilson et al. 2003), two of which are electrically silent
(H+/K+-ATPase and Na+/H+ exchange) and therefore capable of increasing the
discrepancy between Ieq and HCO3− secretion. Two other mechanisms (electrogenic
vacuolar H+-ATPase and H+ channels) would be “invisible” in these experiments
because they would cause equivalent reductions in current and net HCO3- flux.
To assess the possible role of the non-gastric isoform of H+/K+-ATPase, the effect of
apical ouabain on Ieq and HCO3- secretion was investigated (Fig. 2.3A). Ouabain (100
μmol·l-1) did not alter basal or forskolin-stimulated Ieq or HCO3- secretion, suggesting
little role of ouabain-sensitive H+/K+-ATPase activity with an apical solution
containing 5 mmol·l-1 K+ and bubbled with 100% O2. Apical amiloride (1 mmol·l-1)
also had no effect (data not shown), evidence against significant apical proton
secretion via Na+/H+ exchange under these conditions. By contrast, pretreating cells

49
with the vacuolar H+-ATPase inhibitor bafilomycin for 30 min increased Ieq and HCO3-
secretion by ∼0.25 μeq·cm-2·h-1, and similar increases were obtained under basal and
forskolin-stimulated conditions (Fig. 2.3B and C). This implies that forskolin did not
Figure 2.3 Effect of proton transport inhibitors on Ieq and net HCO3-
secretion. A, ouabain (100 μmol·l-1) had no effect when added to the apical side to inhibit putative H+/K+-ATPase activity (n=3), B,C, pretreatment with the V-ATPase inhibitor bafilomycin (50 nmol·l-1), increased both basal and forskolin stimulated HCO3
- secretion. D, ZnCl2 (nominally 1 mmol·l-1) increased in Ieq and HCO3
- secretion slightly when added to the apical side to block putative H+ channels. ● Ieq; ○ HCO3
- secretion. Means±s.e.m.

50
cause insertion of additional proton pumps into the apical membrane (Paunescu,
Ljubojevic et al. 2010). Finally, acute exposure to ZnCl2 (1 mmol·l-1), which inhibits
acid secretion by blocking apical H+ (HVCN1) channels in airway epithelial cells
(Iovannisci, Illek et al. 2010), increased Isc and HCO3- secretion by ∼0.25 μeq·cm-2·h-1
(Fig. 2.3D), implicating proton channels in the H+ efflux. Taken together these
inhibitor studies indicate that HCO3- secretion is 30% higher across Calu-3
monolayers than the measured net flux and is partially neutralized by parallel H+
secretion. However these electrogenic H+ transport mechanisms would not increase
the discrepancy between Ieq and HCO3- net flux and are thus compatible with the
close agreement between the unidentified Ieq and 36Cl- net flux and the net HCO3-
flux and Isc under voltage clamp.
2.4.4 CFTR mediates apical HCO3- conductance
The next series of experiments used control and CFTR knock-down Calu-3 cells to
examine whether CFTR can directly mediate HCO3- flux. Monolayers were
voltage-clamped at 0 mV, the basolateral membrane was permeabilized using
nystatin (360 μg·ml-1), and an apical → basolateral gradient of 25 mM HCO3- was
imposed. Adding forskolin to control monolayers under these conditions caused a
rapid (i.e. negative) Isc, which was blocked by the inhibitor CFTRinh-172 (10 μmol·l-1)
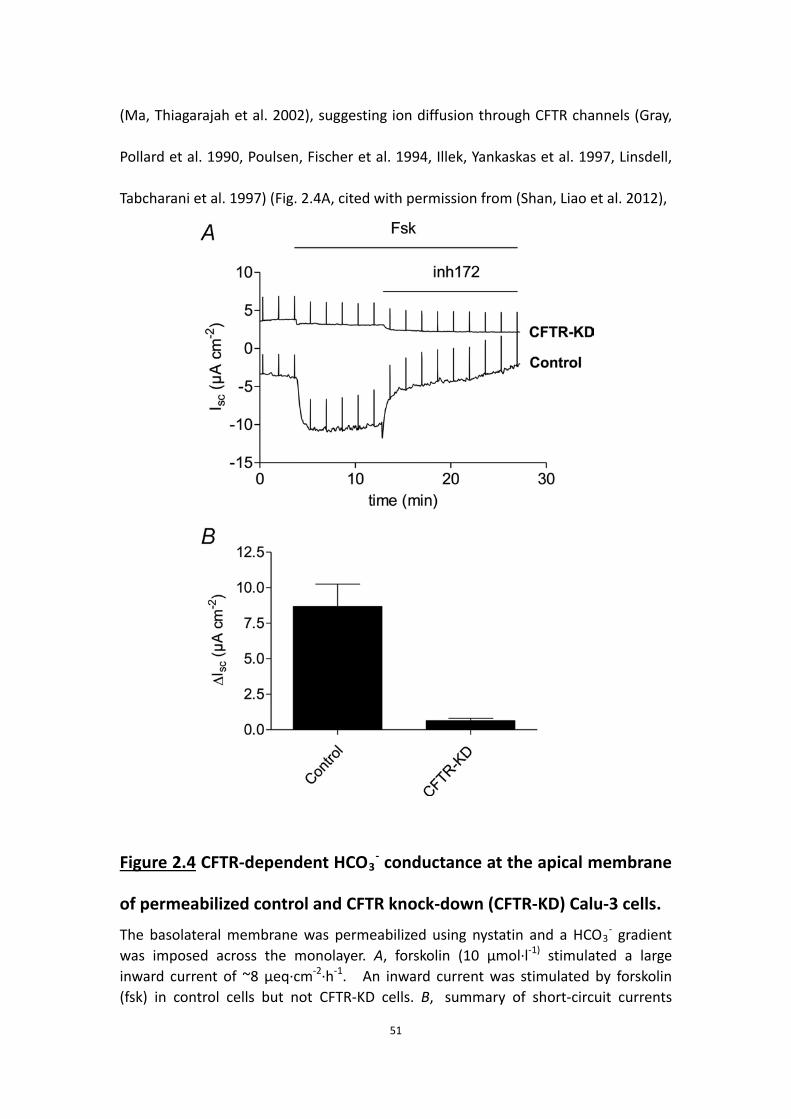
51
(Ma, Thiagarajah et al. 2002), suggesting ion diffusion through CFTR channels (Gray,
Pollard et al. 1990, Poulsen, Fischer et al. 1994, Illek, Yankaskas et al. 1997, Linsdell,
Tabcharani et al. 1997) (Fig. 2.4A, cited with permission from (Shan, Liao et al. 2012),
Figure 2.4 CFTR-dependent HCO3- conductance at the apical membrane
of permeabilized control and CFTR knock-down (CFTR-KD) Calu-3 cells. The basolateral membrane was permeabilized using nystatin and a HCO3
- gradient was imposed across the monolayer. A, forskolin (10 μmol·l-1) stimulated a large inward current of ~8 μeq·cm-2·h-1. An inward current was stimulated by forskolin (fsk) in control cells but not CFTR-KD cells. B, summary of short-circuit currents

52
measured across control cells and CFTR-KD cells in the presence of a bicarbonate gradient. Most of the forskolin stimulated secretion (ΔIsc) was abrogated in CFTR-KD cells. Means±s.e.m.
this experiment performed by Dr. R. Robert).
Forskolin did not stimulate Isc across CFTR knock-down cells under these conditions
(Fig. 2.4A). Figure 2.4B shows the mean Isc measured across basolaterally
permeabilized monolayers. Although these results do not exclude other apical exit
mechanisms, they show that forskolin-stimulated Isc is CFTRInh-172-sensitive and
reduced by 88% in CFTR knock-down cells, consistent with the >95% decrease in
CFTR protein expression. The simplest interpretation is that CFTR channels conduct
HCO3- and mediate electrogenic HCO3
- secretion at the apical membrane under
these conditions.
To test if apical anion exchangers carry HCO3- out of the cell according to the apical
Cl- gradient, we permeabilized the basolateral membrane using nystatin in
symmetrical Cl--free solutions while supplying the monolayers with basolateral HCO3-
and CO2 by gassing basolateral solutions with 95% O2/5% CO2. The apical solutions
were unbuffered so that the HCO3- flux could be measured by pH-stat. After
stabilization, we established Cl- gradients from the apical to the basolateral side of 0,
30 or 120 mM to favor any apical anion exchange. As shown in Fig. 2.5, the Cl-
gradient caused a reversed Ieq, except when the gradient is 0 or in the CFTR

53
knock-down cells, suggesting the reversed Cl- current was mediated by Cl- diffusion
through CFTR. With the Cl- gradient set at 30 mM to support anion exchange, no
HCO3- flux could be observed until forskolin was added to stimulate CFTR, although
some HCO3- flux
Figure 2.5 CFTR mediates HCO3- flux.
Under pH-stat and Cl--free conditions, the basolateral membrane of Calu-3 cell monolayers was permeabilized using nystatin (360 μg·ml-1). A, without Cl- gradients, forskolin (10 μM) alone elicited similar increases in Ieq and HCO3- flux. B, with a 30 mM Cl- gradient from the apical to the basolateral side, no HCO3
- flux could be observed until forskolin was added. A reversed Ieq was observed when the Cl- gradient was established, and this became larger in the presence of forskolin. C, a 120 mM Cl- gradient from the apical to the basolateral side was established after basolateral permeabilization. Unlike B, HCO3
- flux could be detected without forskolin under these conditions. ● Ieq; ○ HCO3
- flux. Means±s.e.m.

54
could be detected with extreme opposing gradients of 120 mM Cl- and 24 mM HCO3-.
The results showed that under pH-stat conditions, the HCO3- flux was mediated by
CFTR rather than apical anion exchangers, at least with a moderate Cl- gradient.
2.4.5 Electrogenic anion transport (Ieq) requires Na+ and CO2/HCO3-
To examine the ionic requirements of HCO3- transport under open-circuit conditions,
Ieq and the appearance of HCO3− on the apical side were measured after bilateral
removal of CO2/HCO3-, Na+ or Cl- (Fig. 2.6A–D). The basal and forskolin-stimulated Ieq
were both abolished in nominally CO2/HCO3--free solution (Fig. 2.6B).
This dependence of Ieq on CO2/HCO3- would be consistent with Na+-nHCO3
-
(NBC)-mediated cotransport at the basolateral membrane (e.g. (Devor, Singh et al.
1999)), but also with H2CO3 synthesis and subsequent Na+/H+ exchange (Cuthbert
and MacVinish 2003). Forskolin had little effect on Ieq under nominally Na+-free
conditions but did induce electrically silent HCO3- secretion at a rate of >0.5
μeq·cm-2·h-1 in the absence of sodium (Fig. 2.6C). Importantly, transepithelial HCO3-
transport during forskolin stimulation was 25% higher under bilateral Cl--free
conditions than when Cl- was present, despite a 50% reduction in Ieq (from 2.76 ±

55
0.13 to 1.35 ± 0.32 μeq·cm-2·h-1; compare mean values indicated by open circles in
Fig. 2.6A and E). This argues strongly against Cl-/HCO3- exchange as the mechanism of
apical HCO3- exit under these conditions. Ieq was identical to net HCO3
- secretion
under symmetrical Cl--free conditions (1.32 ± 0.31 vs. 1.32 ± 0.19 μeq·cm-2·h-1,
Figure 2.6 Effects of ion substitutions. A, the normal response of Calu-3 cells to 10 μmol·l-1 forskolin in control solutions

56
(n=3). B-D, effect of 10 µmol l-1 forskolin on Calu-3 cells bathed on both sides with solutions lacking HCO3
-/CO2, Na+, or Cl-, respectively (n=6 of each). Forskolin-stimulated HCO3
- secretion was abolished in the nominal absence of HCO3
- or Na+ but was increased by 25% in Cl--free conditions, and Ieq was identical to net HCO3
- secretion. E, monolayers were bathed with apical Cl--free solution, then stimulated with 10 μmol l-1 forskolin. Note the smaller and larger HCO3
- -independent Ieq compared to when cells were bathed with symmetrical Cl- solutions. ● Ieq; ○ HCO3
- secretion (n=6 each condition; means±s.e.m.).
respectively), providing further evidence that HCO3--independent Ieq measured in
control solutions is carried by Cl-. The large transient Ieq observed immediately after
forskolin addition under control conditions(e.g. Fig. 2.6A) was eliminated under
bilateral Cl--free conditions and therefore may reflect Cl- and K+ redistribution when
CFTR channels are activated. When Cl- was removed only from the apical side to
generate a favourable Cl- gradient, forskolin caused more robust stimulation of Ieq
and a larger discrepancy between Ieq and HCO3- flux consistent with an increased Cl-
flux (Fig. 2.6E).
2.4.6 Some basolateral Cl- entry is independent of NKCC
Previous studies of NKCC1's contribution to anion secretion by Calu-3 cells have
yielded varying results, perhaps due to different experimental conditions. In the
present work, the effect of bumetanide on Isc was measured and compared with its
effects on Ieq, net HCO3- flux, tracer fluxes, and fluid transport under open-circuit
conditions. Basolateral bumetanide (20–50 μmol·l-1) did not affect Isc during
stimulation by forskolin (data not shown), in good agreement with a previous study
(Devor, Singh et al. 1999). However, under open-circuit conditions bumetanide (20
μmol·l-1) inhibited ∼20% of the basal (Fig. 2.7A) and forskolin-stimulated (Fig. 2.7B)

57
Ieq, which monitors both Cl- and HCO3- transport. Bumetanide inhibition of the Ieq
component carried by Cl- was ∼35% in control-transfected Calu-3 cells (Fig. 2.7A and
B), similar to the results with parental cells (Huang et al. 2011) and with partial
inhibition of Isc noted previously during stimulation by the hyperpolarizing
Figure 2.7 Effect of bumetanide on forskolin-stimulated Ieq. A, pretreatment with the NKCC1inhibitor bumetanide (20 μmol·l-1) partially inhibited basal Ieq (by ~20%) but did not prevent forskolin-stimulation of Ieq or HCO3
- secretion (n=6). B, basolateral bumetanide (20 μmol·l-1) caused partial inhibition (~20%) of Ieq (●) without affecting HCO3
- secretion (○) (n=6).

58
secretagogues 1-EBIO and thapsigargin (Lee et al. 1998; Krouse et al. 2004).
Net 36Cl flux under these conditions confirmed the bumetanide-sensitive component
of Ieq was mediated by Cl- transport (Table 2.1) and confirmed that a fraction of the
transepithelial Cl- transport is mediated by NKCC1 when the basolateral membrane is
hyperpolarized, either by current flowing through paracellular and transcellular shunt
pathways under open-circuit conditions (i.e. loop current), or by activation of
Ca2+-activated potassium channels under short-circuit conditions. The partial
inhibitory effect of bumetanide on Cl--dependent Ieq in Ussing chambers indicates
that substantial Cl- transport is independent of NKCC1.
2.4.7 Evidence for basolateral Cl- loading by anion exchange
Since a large portion of Cl- and HCO3- transport was not carried by NKCC, we
examined whether other basolateral transporters mediate most of the anion
transport. We found that pretreatment with DIDS (100 μmol·l-1), an inhibitor of NBC
and anion exchangers, abolished the forskolin-stimulated responses in control
transfected Calu-3 cells (Fig. 2.8A). However DIDS is not a specific inhibitor and may
inhibit basolateral NBC activity and/or anion exchangers. To examine the involvement

59
of NBCe1, we substituted Cl- with gluconate in the solutions, which should abolish
any component due to anion exchange. Applying DIDS under these conditions caused
large decreases in both Ieq and HCO3- transport (Fig. 2.8B), clearly indicating the role
of NBC in anion transport of Calu-3 cells.

60
Figure 2.8 Effects of DIDS on Calu-3 cells. A, pretreatment of DIDS (100μmol·l-1) could abolish the plateau of forskolin-stimulated Ieq and HCO3
- secretion. B, in Cl--free solutions, forskolin elicited a reduced response of Ieq, which matched HCO3
- secretion, and it could be inhibited by 100 μmol·l-1 DIDS (basolateral), an inhibitor of NBC in Cl--free solutions. ●, Ieq; ○, HCO3
- secretion (n=6 each condition; means±s.e.m.).
Next, to examine if HCO3- efflux through basolateral anion exchangers could serve as
an alternative Cl- entry pathway during anion loading and fluid secretion,
experiments were designed to monitor the ‘trans' effect of anions on the flow of
HCO3- through the basolateral membrane (Fig. 2.9). The apical membrane was
permeabilized using nystatin (100 μg·ml-1) while monolayers were bathed with
symmetrical HCO3--free solution. After the current reached a plateau, 25 mmol·l-1
NaHCO3 was added on the apical side to generate an apical → basolateral HCO3-
gradient and pH-stat was used to monitor the appearance of HCO3- on the
basolateral side.
As shown in Fig. 2.9A, apical nystatin induced a large current that was apparently due
to electrogenic Na+ absorption since it was inhibited by basolateral ouabain in control
experiments (data not shown) as has been reported for other epithelia (Lewis, Wills
et al. 1978).
After Na+ current had stabilized, imposing a transepithelial HCO3- gradient with Cl-
solution bathing the basolateral (trans) side caused Ieq to decrease and HCO3- to
appear in the basolateral compartment (Fig. 2.9A). When the same experiment was

61
performed with gluconate solution on the basolateral (trans) side, no HCO3- flux to
the basolateral side was detected by pH-stat (Fig. 2.9B). However, robust HCO3- flux
was produced with basolateral (trans) NO3- solution (Fig. 2.9C). No osmotically
induced changes in Ieq or HCO3- flux were observed in control experiments when 50
Figure 2.9 Anion exchange mediates forskolin-stimulated anion and
fluid secretion by Calu-3 cells. A and B, monolayers were incubated in HCO3
-/CO2-free or Cl-&HCO3-/CO2-free
solutions, respectively, and gassed with 100% O2. The apical membrane was permeabilized using nystatin (100 μg/ml), and HCO3
- was added to the apical side with 95% O2/5% CO2. An increase of HCO3
- transport to the basolateral side could be detected in the presence of Cl- (A), but not if Cl- was replaced by gluconate (B) (n=4 of each). C, monolayers were incubated in HCO3
-/CO2-free solutions in which Cl- was replaced by NO3
-. The apical membrane was permeabilized and HCO3-
re-introduced as described in A and B. Basolateral HCO3- flux was observed under

62
these conditions, indicating anion exchange (n=4). ● Ieq; ○ HCO3- secretion.
mmol·l-1 mannitol was added instead of 25 mmol·l-1 NaHCO3 (data not shown). These
results indicate HCO3- flow through the basolateral membrane depends on the
nature of the trans-anion and are consistent with the activity of basolateral anion
exchangers which carry HCO3-, Cl- or NO3
- but not gluconate ions.
2.4.8 Electrogenic HCO3- secretion requires carbonic anhydrase
To study the role of HCO3- synthesis in anion and fluid secretion, the effects of
acetazolamide (100 μmol·l-1) on Ieq and HCO3- transport were assayed in Ussing
chambers under pH-stat conditions. Acetazolamide abolished forskolin-stimulated
HCO3- secretion and Ieq (Fig. 2.10 A,B), suggesting a large fraction of the secreted
HCO3- arises from the intracellular hydration of CO2, as suggested previously (Krouse,
Talbott et al. 2004).
Formation of the weak acid H2CO3 and efflux of HCO3- might be expected to
stimulate Na+/H+ exchangers (NHEs) and other pHi regulatory mechanisms. However,
the NHE inhibitor amiloride (1 mmol·l-1; IC50 = 24 μmol·l-1) and the NHE1
isoform-specific inhibitor HOE-694 (20 μmol·l-1; IC50 = 0.16 μmol·l-1) (Counillon,
Scholz et al. 1993) did not affect HCO3- transport or Ieq (Fig. 2.10C). This suggests pHi
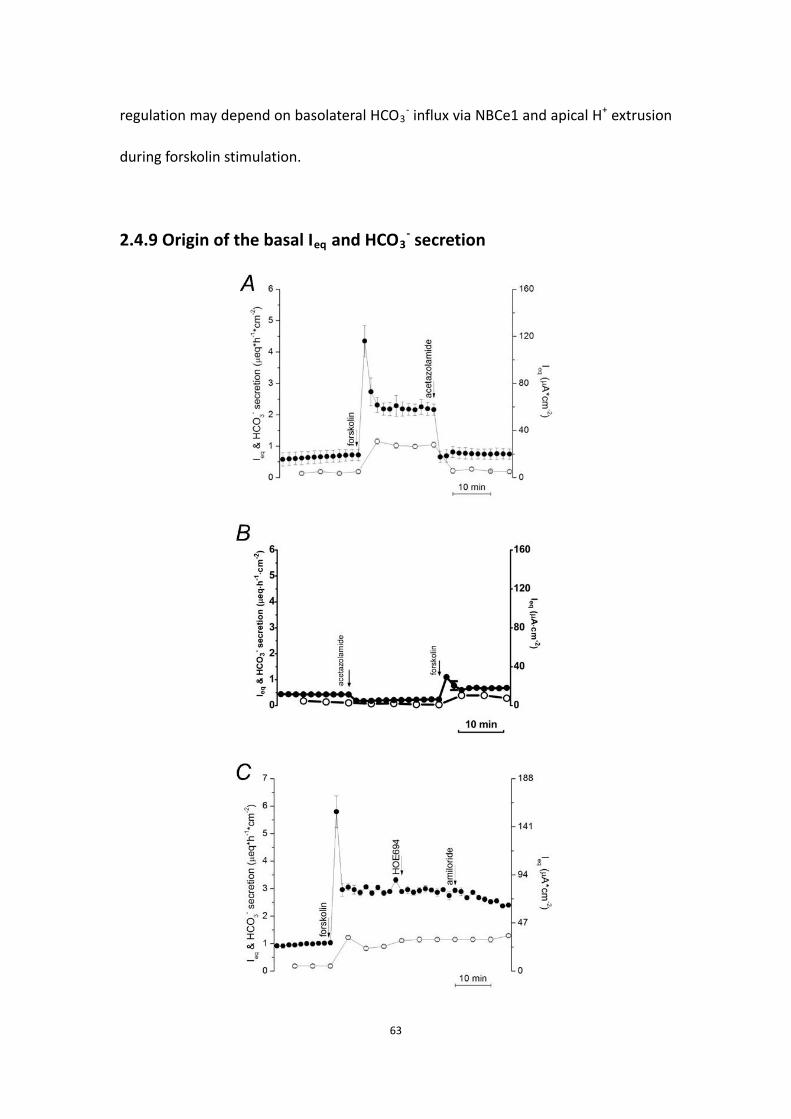
63
regulation may depend on basolateral HCO3- influx via NBCe1 and apical H+ extrusion
during forskolin stimulation.
2.4.9 Origin of the basal Ieq and HCO3- secretion

64
Figure 2.10 Effect of acetazolamide on anion by Calu-3 cells.
A, 100 µmol·l-1 acetazolamide, the inhibitor of carbonic anhydrase, greatly inhibited the forskolin-stimulated secretion (n=6). B, pretreatment with acetazolamide (100 µmol·l-1) also abolished the forskolin-stimulated secretion (n=6). C, sequential addition of the NHE inhibitors HOE694 (10 μmol·l-1) and amiloride (1 mmol·l-1) to the basolateral side did not inhibit HCO3
- secretion (n=3). ● Ieq; ○ HCO3- secretion.
Means±s.e.m.
Calu-3 monolayers have basal currents of 0.5–1.0 μeq·cm-2·h-1; however, the nature
of this constitutive transport remains uncertain. Since Calu-3 cells express purinergic
(Communi, Paindavoine et al. 1999) and adenosine receptors (Cobb, Ruiz et al. 2002)
and respond to prostaglandins (Palmer, Lee et al. 2006), we examined
pharmacologically whether these autocrine signals might be responsible for the basal
current. Ieq and HCO3- transport were unaffected by adding apyrase (10 units·ml-1) to
metabolize ATP released from the cells, 8-SPT (100 μmol·l-1) to prevent activation of
A2B adenosine receptors, or indomethicin (100 μmol·l-1) to inhibit prostaglandin
synthesis (Fig. 2.11A–C).
We then examined the role of CFTR and whether there is tonic adenylyl cyclase
activity. GlyH-101 (100 μmol·l-1), which blocks the CFTR channel with an IC50 ≈ 5
μmol·l-1 at −60 mV (Muanprasat, Sonawane et al. 2004), strongly inhibited both basal
Ieq and HCO3- transport (Fig. 2.12A).
These results suggest CFTR mediates basal secretion. As a further test for the
involvement of CFTR, the dependence of basal current on PKA was investigated.
Adding 100 μmol·l-1 Rp-cAMPS (Rp-adenosine-3’,5’-cyclic mono-phosphorothioate

65
triethylamine salt; IC50 for inhibition of PKA = 4.9 μmol·l-1), a membrane-permeant
competitive inhibitor of PKA, reduced basal Ieq and HCO3- secretion by 63% and 88%,
respectively (Fig. 2.12B), evidence that much of the basal secretion is mediated by
PKA in resting cells and can be rapidly down-regulated by a phosphatase when PKA is
Figure 2.11 Effects of autocrine signaling inhibitors on Ieq and HCO3-

66
secretion. A, the ATPase apyrase (10 units/ml) (n=3) and B, adenosine receptor antagonist 8-SPT (100 µmol·l-1) (n=3) added apically but had no effect on basal Ieq or HCO3
- secretion. C, the cyclo-oxygenase inhibitor indomethacin (100 μmol·l-1) did not inhibit Ieq or HCO3
- secretion when added bilaterally (n=3). Basal current and HCO3
- were CFTR-dependent however, since apical the CFTR channel inhibitor GlyH-101 (100 μM) abolished basal Ieq and HCO3
- secretion (n=4). ●, Ieq; ○, HCO3-
secretion.

67
Figure 2.12 Inhibitors suggest basal Ieq and HCO3- secretion depend on
CFTR and membrane-bound adenylyl cyclase activity, but not
HCO3- -stimulated adenylyl cyclase.
A, basal Ieq was abolished by the CFTR open channel blocker GlyH-101 (n=4). B, basal Ieq and net HCO3
- secretion were inhibited by 200 µmol·l-1 Rp-cAMPS, a competitive inhibitor of cAMP-dependent PKA (n=4). C, basal Ieq and HCO3
- secretion were reduced by the inhibitor of membrane-bound adenylyl cyclase, MDL-12330A (200 µmol·l-1) (n=4). D; inhibition of basal Ieq and HCO3
- secretion by 2-APB (100 μmol·l-1). E,F two chemically unrelated inhibitors of HCO3
--stimulated soluble adenylyl cyclase,

68
2-HE (20 µmol·l-1) and KH7 (50 µmol·l-1) had no effect on Ieq or HCO3- secretion (n=3
of each). G, summary of results in panels A - F. ●,■ Ieq; ○,□ HCO3- secretion.
inhibited.
The sensitivity of basal current to PKA inhibitors implies constitutive elevation of
cAMP, which may be localized near the apical membrane. There are nine
conventional membrane-bound adenylyl cyclases, and one soluble isoform which is
stimulated by HCO3- and is proposed to function as a HCO3
- sensor in various cells
(Chen, Cann et al. 2000), including Calu-3 (Wang et al. 2005). Inhibitors were used to
examine the role of these adenylyl cyclases. An antagonist of conventional
membrane-bound adenylyl cyclases, MDL-12330A (cis-N-(2-phenylcyclopentyl)
azacyclotridec-1-en-2-amine; Guellaen et al. 1977), reduced Ieq and HCO3- secretion
by 68% and 100%, respectively (200 μmol·l-1; IC50 < 10 μmol·l-1; Fig. 2.12C). Since
Ca2+-activated adenylyl cyclase might stimulate CFTR, 2-APB (100 μmol·l-1), a
non-specific inhibitor of store-operated Ca2+ entry, was also tested (Fig. 2.12D).
Remarkably, 2-APB was a potent inhibitor of basal anion secretion, reducing Ieq and
HCO3− secretion 65–90%. This suggests the membrane-bound adenylyl cyclase is
indeed activated by elevated Ca2+ near the membrane. By contrast, bilateral addition
of 2-hydroxyestradiol (2-HE; 20–50 μmol·l-1), an inhibitor of soluble adenylyl cyclase
(Fig. 2.12E), had no effect on basal Ieq or HCO3- transport. The same negative result

69
was obtained using a chemically unrelated inhibitor KH7 (50 μmol·l-1, IC50 = 3–10
μmol·l-1 in vivo; Fig. 2.12F). The effects of these inhibitors on Ieq and HCO3- transport
are summarized in Fig. 12G. Taken together they suggest the basal current in Calu-3
cells is mediated by CFTR channels, which are partially activated by PKA. This tonic
activity may be due to local production of cAMP by a membrane-bound adenylyl
cyclase in response to store-operated Ca2+ entry.
2.5 Discussion
In the current study, the human airway cell line Calu-3 was used because it forms
polarized monolayers, displays cAMP-stimulated bicarbonate and chloride secretion,
expresses the serous cell markers, and responds to gland secretagogues such as
vasoactive intestinal peptide and epinephrine (Shan, Huang et al. 2011). Some results
attained from Calu-3 cells are consistent with previous studies, while others suggest
a revised version of the current anion transport model for Calu-3 cells.
2.5.1 Isc is mediated by electrogenic HCO3- transport
The rate of HCO3- secretion measured in the present study by automated pH-stat was
equal to the simultaneously measured Isc, and both were stimulated by forskolin and
abolished by the CFTR inhibitor Inh-172. Close agreement between Isc and HCO3- flux
is consistent with previous studies of unstimulated (Lee, Penland et al. 1998) and
forskolin-stimulated Calu-3 monolayers (Devor, Singh et al. 1999). The 36Cl- net fluxes
measured under open-circuit conditions in Table 2.1 suggest that membrane

70
hyperpolarization leads to a discrepancy between Isc and HCO3- net flux, as
suggested previously during stimulation with 1-EBIO (Devor, Singh et al. 1999).
Although forskolin also induced some Cl- secretion in the present study, this is
expected since current flowing through shunt pathways would hyperpolarize the
basolateral membrane under open-circuit conditions in concert with apical
depolarization (Tamada, Hug et al. 2001). Thus, the present results extend earlier
findings to open-circuit conditions that would exist during fluid transport.
The present results with H+ transport inhibitors suggest HCO3- secretion is actually
∼30% higher than the net flux measured by pH-stat. Proton secretion by
H+/K+-ATPase has been demonstrated previously during 1-EBIO stimulation (Krouse,
Talbott et al. 2004) but was not observed in the present work. The different results
obtained using forskolin vs. 1-EBIO may be due to their opposite effects on the
membrane potential. Forskolin depolarizes the apical membrane (Tamada, Hug et al.
2001) and should favour electrogenic H+ efflux pathways whereas 1-EBIO would
cause membrane hyperpolarization and favour electroneutral acid secretion.
Electrogenic H+ secretion would be inconspicuous in the present study because it
would cause identical reductions in current and HCO3- net flux. H+/K+-ATPase activity
during 1-EBIO stimulation would consume HCO3- without affecting the current and
therefore would increase the unidentified component of the Isc, as was
demonstrated previously (Krouse, Talbott et al. 2004).

71
2.5.2 Ieq depends on Na+ and CO2/HCO3-
Basal and forskolin-stimulated Ieq were abolished in Na+ or HCO3−-free solution,
consistent with the ionic dependence of Isc reported previously (Devor, Singh et al.
1999). However, we noticed that ∼50% of the HCO3- net flux measured under
pH-stat conditions persisted despite the nominal absence of Na+, suggesting an
additional source of HCO3- besides NBCe1 (Krouse, Talbott et al. 2004). This flux may
reflect carbonic anhydrase-catalysed synthesis and apical efflux since
forskolin-stimulated HCO3- secretion was abolished by acetazolamide (Fig. 2.10A & B),
although we cannot exclude basolateral HCO3- loading through anion exchangers,
which may operate in reversed mode due to the favourable transepithelial HCO3-
gradient under pH-stat conditions.
2.5.3 Cl- loading depends on basolateral anion exchange mostly rather
than NKCC
Bumetanide (100 μM) had little effect on fluid secretion by submucosal glands in cat
trachea (Corrales, Nadel et al. 1984), which suggests the glands use mechanisms
other than NKCC1 unlike airway surface epithelial cells, which are mainly
bumetanide-sensitive. Cl- secretion by freshly isolated equine trachea declined ∼70%
when CO2 and HCO3- were removed under Isc conditions, suggesting basolateral
HCO3-/Cl- exchange (Tessier, Traynor et al. 1990). Further evidence for basolateral
anion exchange (and apical bicarbonate conductance) was obtained in human nasal
primary cells by measuring intracellular pH during extracellular Cl- replacement

72
(Paradiso, Coakley et al. 2003).
In this study, bumetanide (20-50 μM) inhibited only ~35% of the Cl- component of
the Ieq in control-transfected Calu-3 cells, indicating there is an alternative Cl- entry
mechanism besides NKCC1. We demonstrated the basolateral anion exchange by
permeabilizing the apical membrane, imposing an apical-to-basolateral HCO3-
gradient, and examining the effect of anions in the basolateral solution on the HCO3-
flux. At the presence of Cl- or NO3-, an apical to basolateral HCO3
- flux can be
detected. Though the anion exchangers mediating this basolateral Cl- loading were
not identified at the molecular level in the present study, the results suggest AE2 as
the most likely candidate since it is expressed at the basolateral membrane of Calu-3
cells (Loffing, Moyer et al. 2000). Recent studies of an AE2-deficient Calu-3 cell line
indicate that AE2 mediates essentially all Cl−/HCO3− exchange at the basolateral
membrane and plays an important role in fluid secretion; nevertheless, there is
another bumetanide-insensitive mechanism for Cl- loading at the basolateral
membrane in AE2 cells that remains to be identified (Huang, Shan et al. 2012).
2.5.4 Forskolin-stimulated HCO3- secretion requires carbonic anhydrase
Acetazolamide abolished forskolin-stimulated Isc and HCO3- secretion, as observed
previously during stimulation by other secretagogues (Krouse, Talbott et al. 2004).
The simplest explanation for this inhibition is that secreted HCO3- must be
synthesized in the epithelial cells by carbonic anhydrase to sustain electrogenic

73
secretion, even when NBCe1 is available to mediate basolateral HCO3- loading.
Acetazolamide inhibition was surprisingly rapid, perhaps because carbonic anhydrase
activity and the supply of intracellular bicarbonate are limiting when intracellular
pCO2 and [HCO3-] are low due to HCO3
-/CO2-free solution on the apical side.
Alternatively, during forskolin stimulation carbonic anhydrase may form a
bicarbonate transport metabolon with other transporters such as NBCe1 and AE2, so
that inhibiting the enzyme with acetazolamide also rapidly inhibits those transporters.
It is interesting to consider the role of carbonic anhydrase activity in intracellular pH
regulation. Forskolin-stimulated apical HCO3- efflux effectively converts the weak
intracellular acid H2CO3 to a strong acid H+. Cells must neutralize this acid load to
sustain a high rate of HCO3- secretion during steady-state forskolin stimulation.
Basolateral HCO3- entry via NBCe1 probably mediates most of this pHi regulation, as
it occurs during recovery from ammonium-induced acid loads (Inglis, Wilson et al.
2003). Thus, use of basolateral HCO3- entry to neutralize intracellular H+ may
generate CO2 which can be reused for carbonic anhydrase-catalysed HCO3- synthesis,
and this may be especially important when intracellular CO2 and HCO3- are low and
rate limiting. Future studies should examine whether the sensitivity to acetazolamide
is a universal feature of HCO3- secretion or a consequence of using pH-stat conditions
that are admittedly non-physiological.
2.5.5 Further evidence that CFTR mediates apical membrane HCO3− and
Cl− conductance

74
When the basolateral membrane was permeabilized using nystatin and an
apical-to-basolateral HCO3- gradient was imposed, subsequent addition of forskolin
produced a (reversed) Isc across control monolayers that was sensitive to the
inhibitor CFTRinh-172. Although GlyH-101 has also been shown to inhibit the Cl-
channel/exchanger Slc26a9 (Bertrand, Zhang et al. 2009), the anion exchangers
Slc26a3, -a6 and -a11 (Stewart, Shmukler et al. 2011), and mitochondrial function
(Kelly, Trudel et al. 2010), similar results were obtained using CFTR knock-down cells.
Together these results demonstrate the HCO3- conductance of CFTR under these
conditions, consistent with previous studies of Calu-3 monolayers (Illek, Yankaskas et
al. 1997, Tamada, Hug et al. 2001, Krouse, Talbott et al. 2004) and with the single
channel permeability ratio, which ranges between 0.13 and 0.26 (Gray, Pollard et al.
1990, Poulsen, Fischer et al. 1994, Linsdell, Tabcharani et al. 1997). Various factors
could influence permeability of the CFTR pore to Cl- and HCO3-. The pore is partially
blocked by intracellular anions and permeability is enhanced when extracellular Cl- is
replaced with [HCO3-] (Li, Holstead et al. 2011). It may also be modulated by the
WNK1-OSR1/SPAK kinase pathway during stimulated secretion (Park, Nam et al.
2010).
Forskolin-stimulated Isc was less affected by shRNA than CFTR protein expression,
consistent with studies using a different CFTR-deficient Calu-3 cell line (MacVinish,
Cope et al. 2007). Indeed, basal Ieq and HCO3− flux were similar in control and
CFTR-deficient monolayers even though pharmacological studies indicated that basal

75
anion secretion is mediated by CFTR. This result would be explained if some band C
protein that had been lost in knock down cells was not at the cell surface, or if the
decline in conductance caused by silencing CFTR was partly offset by an increase in
anion driving force as would be expected if CFTR was rate-limiting.
On the other hand, when the apical anion exchange is considered, our data suggest
that CFTR that mediates most HCO3- flux under pH-stat conditions. Nevertheless we
cannot exclude the possibility that apical anion exchangers participate in
physiological HCO3- secretion. The pH-stat constantly neutralizes any secreted HCO3
-
and prevents the accumulation of apical HCO3-, thus a basolateral-to-apical 25 mM
HCO3- gradient always exists under these conditions and HCO3
- may readily exit the
cell through CFTR. However, in the airways, HCO3- does accumulate in the thin film of
ASL. To overcome the less favorable transepithelial HCO3- gradient and drive more
HCO3- secretion, airway epithelial cells may then utilize the apical Cl- gradient and the
apical exchangers to transport HCO3-.
2.5.6 A revised anion transport model of Calu-3 cells
The present results suggest a revised model of anion secretion by Calu-3 cells (Fig.
2.13). 50–70% of the basolateral Cl- loading occurs by anion exchange, which is
increased during forskolin-stimulated secretion. This scheme differs from a model in
which NKCC mediates basolateral Cl- entry (Devor, Singh et al. 1999). We hypothesize
that basolateral Cl- loading via basolateral anion exchange increases during forskolin

76
stimulation, in contrast to another recent model for Calu-3 in which forskolin
stimulation was proposed to inhibit this basolateral anion exchange (Garnett,
Hickman et al. 2011).
Figure 2.13 Model for anion transport by Calu-3 cells proposed by Shan
et al.. In resting cells, an apical Ca2+ microdomain produced by store-operated Ca2+ entry causes partial activation of CFTR through stimulation of membrane-bound adenylyl cyclase and local elevation of [cAMP]. Secretagogues that further elevate [cAMPi] stimulate apical Cl− and HCO3
− efflux, creating an acid load that may further increase CFTR open probability (Chen et al. 2009). Regulation of pHi by HCO3
− that enters via NBCe1 provides CO2 for bicarbonate synthesis by carbonic anhydrase, which sustains HCO3
− secretion. Most bicarbonate entering via NBCe1 is recycled basolaterally by anion exchange during Cl− loading. Not shown in this scheme are other anion exchangers and Na+/H+ exchangers that are active under other conditions; i.e. Na+/H+ exchange is likely to mediate pHi regulation during stimulation by secretagogues that hyperpolarize the basolateral membrane.

77
Forskolin stimulates a large transient current which is abolished in Cl--free solution
and therefore probably mediated by apical Cl- and basolateral K+ efflux. While the
decline in intracellular Cl- activity would favour basolateral anion exchange, cell
shrinkage produced by the loss of these solutes may trigger a volume regulatory
increase and stimulation of Cl- entry through NKCC1 (Jiang, Chernova et al. 1997). In
this scheme for Calu-3 cells, most apical HCO3- efflux is sustained by intracellular
HCO3- synthesis, which is catalysed by carbonic anhydrase. The acid load produced by
forskolin-stimulated HCO3- efflux may be neutralized by HCO3
- entry through NBCe1,
whereas H+ extrusion via electroneutral NHE1 (Cuthbert and MacVinish 2003) and
H+/K+-ATPase (Krouse, Talbott et al. 2004) mediate pHi regulation during the
response to hyperpolarizing secretagogues.
Calu-3 monolayers had significant basal Ieq and HCO3- secretion. Basal activity was
sensitive to inhibitors of CFTR (GlyH-101), PKA catalytic subunit (Rp-cAMPS) and
membrane-bound adenylyl cyclase (MDL-12330A). Adenylyl cyclase may be
constitutively activated by Ca2+ entry through the store-operated Ca2+ entry channel
Orai1. A similar mechanism might explain the high basal activity of CFTR noted in
other tissues such as the sweat duct (Quinton 1983) and small airways (Wang, Lytle

78
et al. 2005). The present results do not allow discrimination of conductances that are
strictly dependent on CFTR activity (Bertrand, Zhang et al. 2009).
Chapter 3 Fluid secretion by an
airway epithelial cell line

79
In the previous chapter, we proposed a revised cellular scheme for anion transport by
Calu-3 cells and showed the importance of HCO3- for the short-circuit current and
transepithelial Cl- flux. However, preliminary studies of fluid secretion showed that
the concentration of HCO3- was surprisingly low, ~25 mM. We therefore proceeded
to study the relationships between different anions and fluid secretion, and
investigated how fluid secretion is regulated.

80
3.1 Abstract
Fluid secretion is reduced in cystic fibrosis, a disease caused by CFTR dysfuncion. In
normal airways, CFTR mediates the secretion of Cl- and HCO3- however the precise
roles of these anions in fluid transport by airway epithelial cells is not well
understood. In this study, we collected fluid produced by genetically matched
CFTR-deficient and CFTR-expressing cell lines derived from a human airway epithelial
cell line (Calu-3), and measured the volume, pH, anion composition and the
osmolality of the samples. Forskolin elicited CFTR-dependent fluid secretion.
Compared to the CFTR-expressing cell line, the volume of fluid secretion was greatly
reduced and the pH was more acidic in the CFTR-deficient cell line, suggesting less Cl-
and HCO3- transport, consistent with the results in the previous chapter. Fluid
secretion depended on the presence of all three ions (Na+, Cl- and HCO3-). Cl- was the
predominant anion in the secretions and therefore drove most of the fluid transport.
Consistent with the partial inhibition of Cl- transport by bumetanide (see Ieq
measurements in previous chapter), the volume of fluid produced was inhibited only
~20% by bumetanide, indicating a minimal role for NKCC1 and the presence of an
alternative Cl- transport mechanism at the basolateral membrane. Moreover adding
NO3-, which is not carried by NKCC1, restored most of the fluid secretion in nominally

81
Cl--free medium, suggesting an alternative mechanism, e.g. anion exchange, exists at
the basolateral membrane for Cl- loading. These results support a model proposed in
the previous chapter, in which most HCO3- taken up by NBCe1 is recycled
basolaterally by exchange for Cl-, and the resulting HCO3--dependent Cl- transport
generates most of the osmotic driving force for fluid secretion. Consistent with this
interpretation, additing acetazolamide in this fluid secretion assay, an inhibitor of
carbonic anhydrase that abolished Ieq and HCO3- secretion, caused only ~20%
inhibition of fluid transport. Time course studies revealed that most fluid was
produced during the first 6 h in this 24 h assay. On the other hand, we found that
evaporation did not affect the accuracy of the volume measurements. These data
prompted us to propose a novel but simple model for the regulation of ASL volume,
in which fluid secretion or absorption is mainly determined by osmolality created by
secreted ions.
3.2 Introduction
The airways are covered by a thin film of liquid which humidifies inspired air and
enables the cilia to beat and clear mucus containing pathogens and other inhaled
particles from the lungs. Most airway surface liquid (ASL) is produced by submucosal
glands, which secrete a complex mixture of ions, fluid, mucins and antimicrobial
factors when stimulated by agonists that mobilize intracellular Ca2+ and/or [cAMP]
(Wine and Joo 2004). The depth of ASL is 5–30 μm in healthy airways (Widdicombe
2002, Boucher 2003), but is reduced in cystic fibrosis (CF) due to abnormal

82
transepithelial ion and water transport (Tarran, Trout et al. 2006), and this diminishes
mucociliary clearance and leads to recurring infections and chronic inflammation.
Despite the important role of ASL in airway host defense and pathophysiology, the
relationship between anion and fluid secretion by airway epithelia remains poorly
understood.
Isosmotic fluid secretion by most epithelia is driven by the active transport of anions,
which are followed passively by cations and osmotically-obliged water (Ballard and
Inglis 2004). While NKCC1 is the main route for basolateral Cl- uptake during
secretion across many epithelia, in some tissues such as airway submucosal glands,
fluid secretion is insensitive to the NKCC1 inhibitor bumetanide (Corrales, Nadel et al.
1984). Basolateral anion exchange has been proposed as an alternative mechanism
for basolateral Cl- entry in salivary glands (Case, Hunter et al. 1984, Novak and Young
1986, Pirani, Evans et al. 1987, Turner and George 1988) and intact trachea, where
most secretion is from the glands (Tessier, Traynor et al. 1990). Removing exogenous
CO2 and HCO3- inhibits fluid secretion by individual airway submucosal glands ∼50%,
and similar results are obtained during stimulation by vasoactive intestinal peptide
(VIP) and/or acetylcholine (Joo, Saenz et al. 2002, Choi, Joo et al. 2007). This
dependence of fluid secretion on exogenous CO2/HCO3- would be greater if not for
the metabolic production of CO2 and HCO3− by the epithelium.
Previous studies have suggested various cellular models for anion and fluid secretion

83
in the airways. Although Cl- and HCO3- can permeate CFTR (Gray, Pollard et al. 1990,
Poulsen, Fischer et al. 1994, Linsdell, Tabcharani et al. 1997), apical anion exchange
has recently been proposed to mediate most HCO3− secretion by Calu-3 cells
(Garnett, Hickman et al. 2011). This model is similar to those that have been
proposed for duodenum (Simpson, Gawenis et al. 2005) and pancreatic duct (Lee,
Choi et al. 1999), which produce strongly alkaline secretions that are driven by HCO3−.
However, the pH of submucosal gland secretions is near neutrality in 5% CO2 (pH
6.9–7.1) (Song, Salinas et al. 2006) similar to the ASL pH, which ranges between 6.2
and 7.2 (Fischer and Widdicombe 2006). The low HCO3− concentration of airway
submucosal gland secretions and ASL (~20 mmol·l−1) is difficult to reconcile with the
high rates of HCO3− secretion in other tissues where HCO3
− drives fluid secretion.
Although HCO3− would contribute 20 mosmol·l−1 to the osmotic pressure of gland
secretions (i.e. < 7% of the total solute concentration of 300 mosmol·l−1), removing
exogenous CO2/HCO3− reduces the volume of secretions by ≥ 50% (Joo, Irokawa et al.
2002). These differences indicate that although HCO3- may not be at a high
concentration in the ASL, it is nevertheless vital for fluid secretion and the
maintenance of ASL volume.
In this study, we sampled fluid produced by polarized Calu-3 monolayers at different
time points and measured its volume, pH, Cl- concentration and osmolality. By
comparing CFTR-expressing and CFTR-deficient cell lines, we found that forskolin
elicits CFTR-dependent fluid secretion. Fluid transport by Calu-3 monolayers was

84
dependent on Na+, Cl- and HCO3- and was abolished by removal of any of these ions.
Cl- was the predominant anion in the secreted fluid, whereas the concentration of
HCO3- was relatively low. These and other unexpected results prompted us to
investigate the role of anions in fluid transport and propose a novel, but simple,
mechanism for the regulation of ASL volume.
3.3 Methods
3.3.1 Cell Culture
The CFTR-deficient cell line and the control CFTR-expressing cell lines called Sizt and
Alter, respectively, were obtained from Dr. Scott O’Grady at the Univ. of Minnesota
(Palmer, Lee et al. 2006). Both cell lines were cultured in Eagle's minimum essential
medium (EMEM; Wisent Bioproducts, St. Bruno, QC) containing 7% fetal bovine
serum (FBS). Cells were seeded at 5 × 105 cells per cm2 on Transwells (4.67 cm2,
Corning) for fluid secretion assays. Fresh medium was placed on the basolateral side
one day after plating and the apical medium was removed to establish an air–liquid
interface (ALI). Any fluid that appeared spontaneously on the apical surface was
removed after 3 days. Cultures were maintained in a humidified 5% CO2 incubator at
37°C and studied after 21–25 days.
3.3.2 Media
To maintain viability during long fluid secretion experiments, EMEM medium
containing essential amino acids was placed on the basolateral side. The salts of

85
amino acids (l-histidine·HCl, l-lysine·HCl) and vitamins (choline chloride,
pyridoxine·HCl, thiamine·HCl) contributed 0.6 mmol·l−1 Cl− to the nominally Cl−-free
medium. Cl− contamination probably did not affect the results however, since adding
this concentration of Cl− during pH-stat measurements had no detectable effect on
Ieq, which is a measure of the net Cl− + HCO3− flux under open-circuit conditions.
Monolayers bathed with basolateral 25 mmol·l−1 HCO3− medium were kept in 5%
CO2/95% air, which was nominally saturated with H2O during fluid secretion
measurements unless otherwise noted. Control experiments performed with
Transwells mounted on an orbital shaker in a 5% CO2/95% O2 atmosphere to provide
better oxygenation yielded only slightly higher fluid secretion rates (data not shown).
Humidified air (0.035% CO2) was used when measuring fluid secretion with
nominally HCO3−-free basolateral medium.
3.3.3 Volume and composition of the secreted fluid
Fluid was aspirated from the apical surface and fresh media containing replacement
ions, activators or inhibitors were added on the basolateral side to begin fluid
secretion assays. In some experiments, agents were added to the apical side in a
small volume of Krebs–Henseleit solution (mmol·l−1: 120 NaCl, 25 NaHCO3, 3.3
KH2PO4, 0.8 K2HPO4, 1.2 MgCl2, 1.2 CaCl2 and 10 mannitol), which was subtracted
from the volume measured at the end of the experiment when calculating secretion
rates. Apical fluid was collected at 24 h intervals using a pipettor and the
time-averaged fluid secretion rate was calculated after subtracting the volume of

86
vehicle if added on the apical side. Although cumulative volume increased linearly for
at least 3 days, the instantaneous secretion rate declined over the course of each 24
h collection period, therefore total volume secreted under these conditions is
sub-maximal. A low rate of evaporation was observed from siliconized Transwells in
control measurements (∼0.27 μl·cm−2·h−1) despite being kept in a humidified
incubator. However, when cultures were coated with a film of water-saturated
hexadecane (a solvent having low vapor pressure, which eliminates evaporation), the
volume of secretions that could be retrieved from cell monolayers was not altered.
High osmotic permeability apparently enables sufficient H2O diffusion through the
monolayer to compensate for evaporative H2O loss from the surface of uncoated
cultures so that the steady-state apical fluid volume is determined simply by the
quantity of solute on the surface. Correcting for evaporation would cause
overestimation of active fluid transport, therefore no correction was applied. The Cl−
concentrations of secretions and media were measured using an ADVIA 1650
biochemistry system (Bayer) after 10-fold dilution in distilled H2O. pH was measured
using a micro-electrode (9826BN, Orion) which was either kept inside the 5% CO2
incubator to reduce equilibration time or immersed in the sample while bubbling
with 5% CO2. The Henderson–Hasselbalch equation was used to calculate the
equilibrium HCO3− concentration:
where pK = 6.09 and PCO2 = %CO2/100·(760-47). No corrections were made for small
errors in water vapor pressure (slightly less than 47 mmHg), laboratory elevation

87
above sea level (∼31 m) or daily fluctuations in barometric pressure. The incubator
was set using the built-in infrared controller and confirmed independently within
±0.01% using an external, non-dispersive infrared (NDIR) sensor (GMM221, Vaisala,
Lake Villa, IL, USA).
3.3.4 Data analysis
Paired or unpaired Student's t tests with P < 0.05 were used for single comparisons,
whereas one-way analysis of variance followed by the Bonferonni post hoc test was
used for multiple comparisons.
3.4 Results
3.4.1 Fluid secretion is CFTR dependent
Dehydration is a hallmark of CF airway secretions, and suggests a role of CFTR in fluid
secretion. To test if fluid transport by Calu-3 cells is CFTR dependent, apical fluid was
removed at time 0 h and secretions were collected from control and CFTR
knock-down cultures at 24 h intervals under control conditions and during forskolin
stimulation (Fig. 3.1A). Little fluid was produced by control monolayers that were left
untreated or exposed only to vehicle (0.1% basolateral DMSO). However, fluid
secretion increased to > 40 μl·day−1 during stimulation with forskolin (10 μmol·l−1),
and a similar volume was produced over the course of several days, leading to the
cumulative secretion of ∼130 μl.

88
Figure 3.1B shows the time-averaged fluid secretion rate measured daily for 3 days
under each condition. GlyH-101 (100 μmol·l−1) nearly abolished forskolin-stimulated
fluid secretion when added to the apical side in 400 μl PBS, suggesting CFTR channel
activity is required for fluid transport.
3.4.2 Cl− is the predominant anion in Calu-3 secretions

89
Figure 3.1 CFTR-dependent fluid secretion by control and CFTR knock
down Calu-3 cells. A, Fluid secretion under different conditions: ■ control Calu-3 cells stimulated with 10 μmol·l-1 forskolin; ▲ untreated, ▼ 0.1 % DMSO, or ● forskolin + GlyH-101 (100 μmol·l-1) treated control Calu-3 cells, and ◆ CFTR knock down Calu-3 cells treated with forskolin (n=6 each condition). B, summary of results in A. Means±s.e.m.

90
To investigate if Cl- and/or HCO3- is essential to fluid secretion, the pH and the [Cl-] of
apical fluid on control and CFTR knock-down monolayers was measured. Surprisingly,
the pH of the fluid was in the range 7.2–7.6 when equilibrated with 5% CO2 (Table
3.1). The Cl− concentration in fluid secreted by control shRNA transfected, CFTR
knock-down and parental Calu-3 cell lines was ∼ 120 mmol·l−1. Thus, Cl− was the
predominant anion and was 2- to 4-fold higher than the [HCO3−] calculated using the
Henderson–Hasselbalch equation. [HCO3−] was similar in secretions collected after
several hours or several days of forskolin stimulation (data not shown), and the
osmotic pressure of secretions at the end of collection periods was equal to that of
the basolateral medium within measurement error (Table 3.1), suggesting most fluid
is osmotically driven by transepithelial Cl− rather than by HCO3− secretion despite the
close correspondence between Isc and HCO3− secretion (Fig. 2.1), and the absence of
detectable 36Cl− net flux under Isc conditions (Devor, Singh et al. 1999).
3.4.3 HCO3−, Na+ and Cl− are all required for forskolin-stimulated fluid
secretion
The low [HCO3−] and high [Cl−] of Calu-3 secretions prompted us to study the ionic
requirements for fluid secretion and Ieq. Fluid transport was reduced from 45.1 ± 7.9
μl·cm−2·day−1 to 0.8 ± 0.4 μl·cm−2·day−1 when monolayers were bathed with
nominally HCO3−-free medium (Fig. 3.2), and a similar decrease to 1.0 ± 0.8
μl·cm−2·day−1 was observed in nominally Na+-free medium. These effects of removing
HCO3− and Na+ were anticipated since Ieq also depended on these ions (e.g. Fig. 2.6B
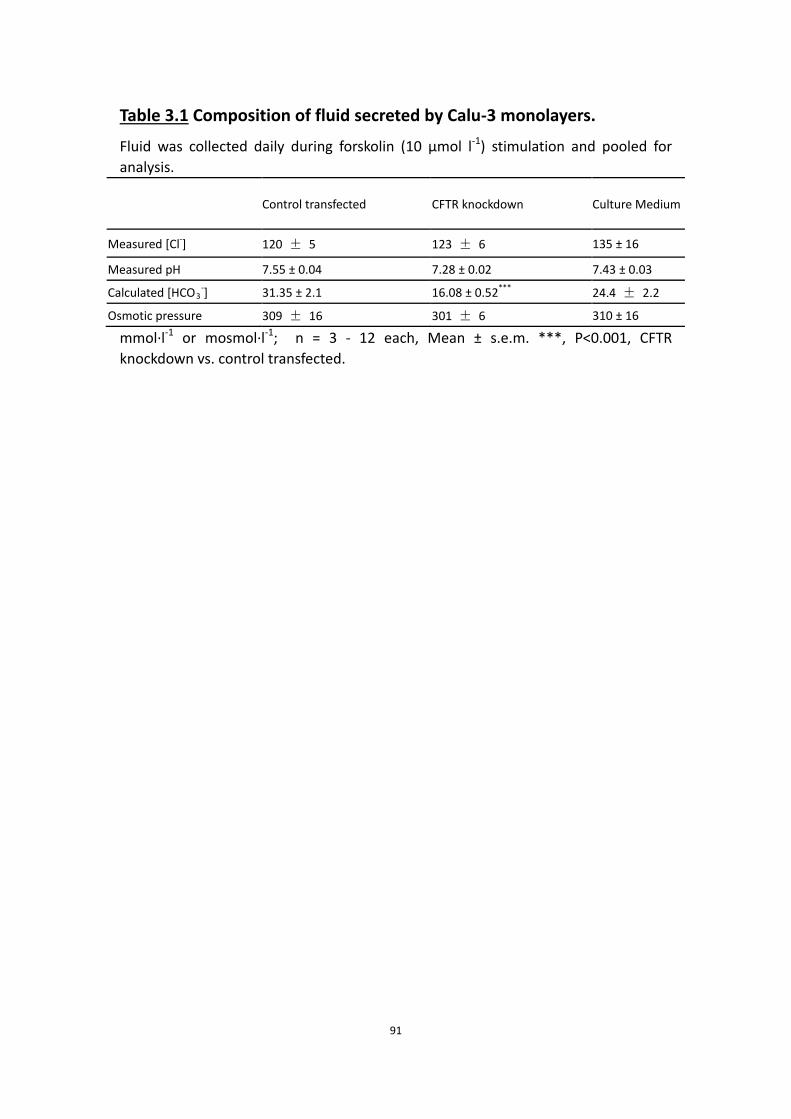
91
Table 3.1 Composition of fluid secreted by Calu-3 monolayers.
Fluid was collected daily during forskolin (10 μmol l-1) stimulation and pooled for analysis.
Control transfected CFTR knockdown Culture Medium
Measured [Cl-] 120 ± 5 123 ± 6 135 ± 16
Measured pH 7.55 ± 0.04 7.28 ± 0.02 7.43 ± 0.03
Calculated [HCO3-] 31.35 ± 2.1 16.08 ± 0.52*** 24.4 ± 2.2
Osmotic pressure 309 ± 16 301 ± 6 310 ± 16
mmol·l-1 or mosmol·l-1; n = 3 - 12 each, Mean ± s.e.m. ***, P<0.001, CFTR knockdown vs. control transfected.
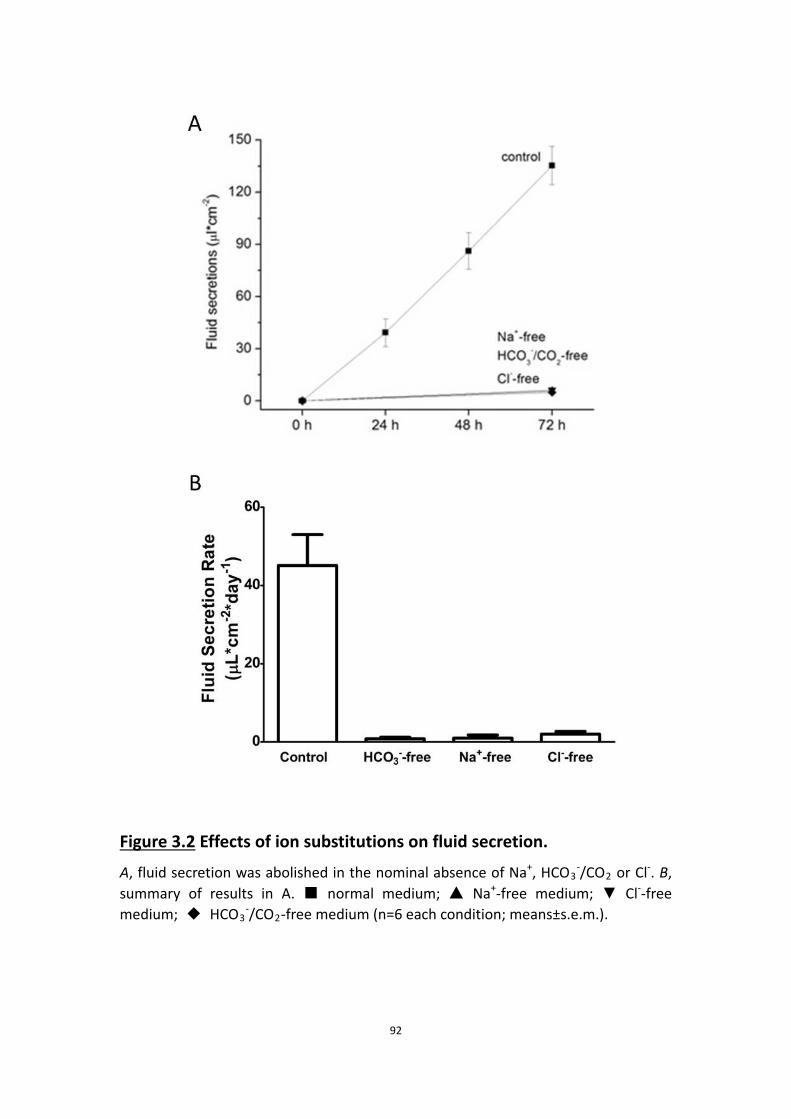
92
Figure 3.2 Effects of ion substitutions on fluid secretion. A, fluid secretion was abolished in the nominal absence of Na+, HCO3
-/CO2 or Cl-. B, summary of results in A. ■ normal medium; ▲ Na+-free medium; ▼ Cl--free medium; ◆ HCO3
-/CO2-free medium (n=6 each condition; means±s.e.m.).

93
and C). More surprising was the effect of replacing Cl− with gluconate, which nearly
abolished fluid secretion (from 45.1 ± 7.9 μl·cm−2·day−1 to 2.0 ± 0.7 μl·cm−2·day−1; Fig.
3.2). This contrasts with the effect on Ieq, 60% of which persisted in nominally
Cl−-free solution (Fig. 2.6D). Thus, forskolin-stimulated HCO3− secretion depends on
the simultaneous presence of Na+ and HCO3− but not Cl−, whereas fluid secretion
requires all three ions (Fig. 3.2).
Ion substitution effects and the high [Cl−] of secretions suggested that most fluid is
osmotically driven by transepithelial Cl− rather than HCO3− transport.
3.4.4 Most of the fluid secretion is independent of NKCC
Bumetanide (20 μmol·l−1) reduced fluid secretion from 33.5 ± 6.98 to 26.6 ± 1.39
μl·cm−2·day−1; however, the remaining 70–80% of the fluid secretion was insensitive
to this inhibitor despite the high [Cl−] of the secreted fluid (Fig. 3.3). Inhibition of fluid
transport was not further increased by raising bumetanide concentration to 50
μmol·l−1 (data not shown). Partial inhibition of fluid transport by bumetanide
paralleled its effect on Cl−-dependent Ieq in Ussing chambers, reinforcing the
conclusion that substantial Cl− and fluid transport is independent of NKCC1.
3.4.5 Basolateral anion exchange is important to fluid secretion
In the previous chapter, we have shown that basolateral anion exchange is important
to Cl- loading and AE2 is a candidate to be the mediator of the anion exchange. Since

94
Figure 3.3 Effects of bumetanide on forskolin-stimulated fluid
secretion. forskolin-stimulated fluid secretion was inhibited ~15% by bumetanide (▲) when compared to forskolin alone (■) (n=6 each condition).

95
NO3− substituted for Cl− during basolateral anion exchange, we wondered if it could
also support fluid transport. Forskolin stimulated robust fluid secretion when
monolayers were bathed with nominally Cl−-free NO3− medium on the basolateral
side (Fig. 3.4). The rate of fluid transport from basolateral NO3− solution was > 60% of
that measured with Cl− solution during the first 24 h stimulation, then gradually
declined, presumably because Cl− is needed for other cellular processes in order to
maintain long-term viability. Nevertheless, NO3− did sustain fluid secretion for many
hours, evidence that its basolateral entry is supported by HCO3− efflux in apically
permeabilized monolayers. HCO3− recycling through basolateral anion exchangers
can apparently load Calu-3 cells with either Cl− or NO3−, and both these anions
support fluid secretion because both are permeant through apical CFTR channels
(Linsdell, Tabcharani et al. 1997).
3.4.6 Carbonic anhydrase has little effect on fluid secretion
In chapter 2, we found that carbonic anhydrase is vital to anion transport by Calu-3
cells, since inhibiting it with acetazolamide abolished forskolin-stimulated
transepithelial anion transport as measured by Ieq. Thus, we speculated that carbonic
anhydrase may also play an important role in fluid transport, and that treatment with
acetazolamide should greatly reduce fluid secretion by stimulated Calu-3 cells.
However, application of acetazolamide (100 μM) only reduced fluid secretion by 27%
(Fig. 3.5), regardless of whether cells were pretreated with acetazolamide or if it was
added during forskolin stimulation.

96
Figure 3.4 Anion exchange mediates forskolin-stimulated fluid secretion
by Calu-3 cells. Fluid secretion was largely maintained for the first 24 h when Cl- was replaced with NO3
- but was nearly abolished after gluconate replacement. ■ normal medium; ▲ low-Cl- gluconate medium; ▼ low-Cl- nitrate medium.

97
Figure 3.5 Effects of acetazolamide on fluid secretion by Calu-3 cells. ■, without acetazolamide; ▲, with acetazolamide (n=6 of each). Acetazolamide caused only a small reduction in fluid secretion by Calu-3 cells.

98
3.4.7 The time course of fluid secretion rate during 24 h stimulation
To see if the fluid was secreted constantly during 24 h stimulation, we collected fluid
and measured the volume every hour, and then put the fluid back onto the apical
surface of Calu-3 monolayers for the next measurement to simulate a normal
secretion assay.
Fig. 3.6 shows that after forskolin stimulation, the control CFTR-expressing Calu-3
monolayers actively secreted during the first 6 h at an average rate of ~5 μl·cm-2·h-1,
and nearly 75% of the total daily amount was secreted during this initial period. After
the first 6 h, fluid secretion rate declined by ~90% to 0.5 μl·cm-2·h-1and fluid
accumulated slowly on the monolayers.
3.4.8 Evaporation does not affect the rate of fluid accumulation
It is believed that evaporation reduces the fluid volume under these conditions and
affects the accuracy of the volume measurements, consequently others have
corrected for evaporation (Garnett et al., 2011). We determined the evaporation rate
by sealing the Transwell with silicone polymer, and adding 200 μl medium onto the
apical compartment of the well. We observed a low rate of evaporation at ~0.3
μl·cm-2·h-1 despite keeing the Transwells in an atmosphere that was > 90% saturated
with H2O.
To test the impact of this evaporation on measurements of fluid secretion, we

99
Figure 3.6 Time course of fluid secretion during 24 h fluid secretion
assay. Calu-3 cell monolayers were stimulated by forskolin (10 μM). Most fluid was secreted during the first 6 h after stimulation. After the rapid secretion phase, there was a slow secretion phase, in which the secretion rate was almost one tenth of the one in the rapid phase (n=6 of each).

100
utilized water-saturated hexadecane, a solvent which has a very low vapor pressure
that should prevent evaporation. It was shown to be effective when the medium on
the siliconized Transwells was covered by a thin film of water-saturated hexadecane
(Fig.3.7A). However, when Calu-3 monolayers were coated with hexadecane, the
volume of secreted fluid that could be retrieved from the monolayers was not altered
(Fig. 3.7B). This rather surprising result suggests that the high osmotic permeability
and the osmotic driving force created by secreted ions is sufficient for H2O diffusion
through the monolayer to fully compensate for evaporative H2O losses. Therefore,
the apical fluid volume is determined simply by the quantity of solute on the surface.
The common practice of correcting for evaporation, which ignores this passive water
flux, is expected to cause overestimation of active fluid transport, therefore no
correction was applied to data from the fluid secretion assays.
3.4.9 Osmolality determined the fluid secretion rate
To explore the hypothesis that total solute content determines fluid flux, we applied
apical solutions with different osmolalities to unstimulated and stimulated Calu-3
monolayers.
Without forskolin stimulation, monolayers at the air-liquid interface (control) and in
solutions with 290 mOsm showed similar fluid secretion rates, ~10 μl·cm-2·h-1.
Solutions at 500 mOsm caused an increase in water flux at a rate of 60 μl·cm-2·h-1,
whereas exposure to 55 mOsm solution led to absorption at 12 μl·cm-2·h-1 rather
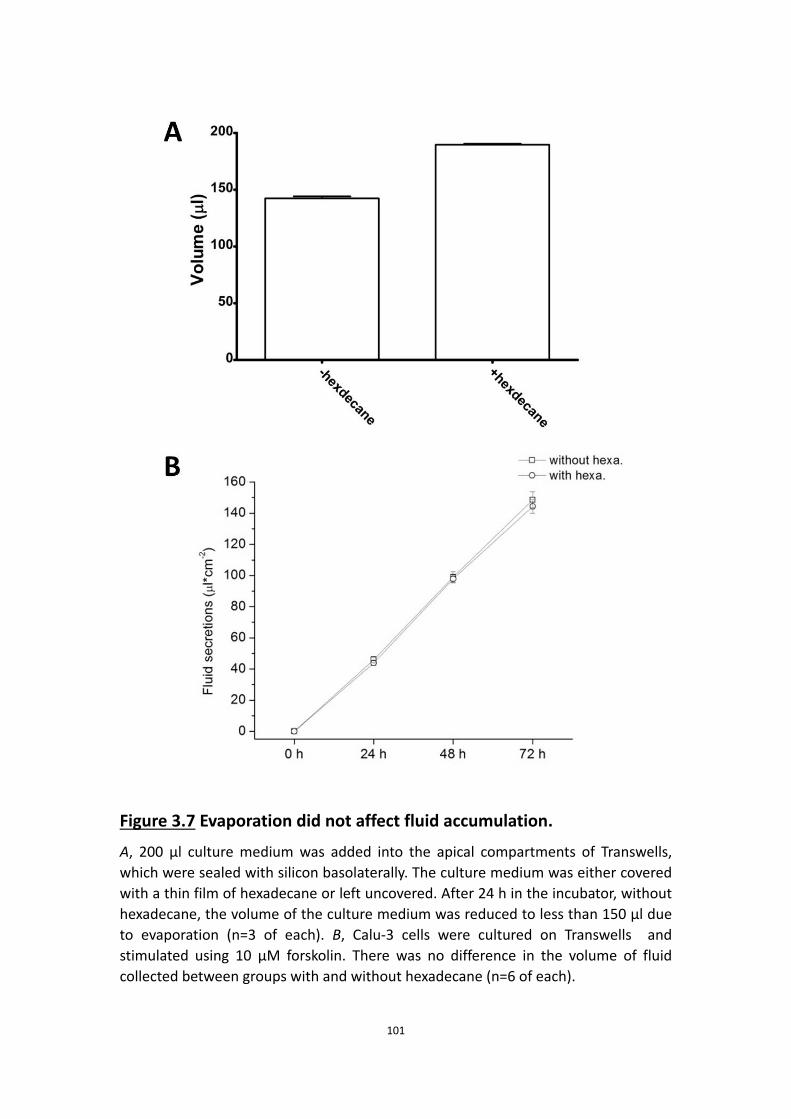
101
Figure 3.7 Evaporation did not affect fluid accumulation. A, 200 μl culture medium was added into the apical compartments of Transwells, which were sealed with silicon basolaterally. The culture medium was either covered with a thin film of hexadecane or left uncovered. After 24 h in the incubator, without hexadecane, the volume of the culture medium was reduced to less than 150 μl due to evaporation (n=3 of each). B, Calu-3 cells were cultured on Transwells and stimulated using 10 μM forskolin. There was no difference in the volume of fluid collected between groups with and without hexadecane (n=6 of each).

102
than secretion (Fig. 3.8A). During forskolin stimulation, addition of 290 mOsm
solution apically caused a significant reduction in the fluid secretion rate from ~40 to
~28 μl·cm-2·h-1, compared to control (Fig. 3.8B), presumably because adding solution
on the apical side caused dilution of the osmolytes secreted by the monolayer and
reduced the driving force that was generated initially by forskolin stimulated of salt
secretion.
To further investigate how osmolality regulates the fluid secretion rate, we studied
the time course of osmolality changes in secretions. We collected secreted fluid and
basolateral medium every hour for the first 5 h during a 24 h fluid secretion assay
and measured the osmolality. We found that the osmolality of apical fluid was always
higher than in the basolateral medium during the first 5 h of stimulated fluid
secretion by Calu-3 monolayers (Fig. 3.8C). After the initial active secretion phase,
the osmolality of the fluid and the medium became similar, consistent with the
observed slowing of the secretion rate in Fig. 3.6.
These results confirmed that we could elicit net fluid flux without stimulating the
monolayers, by imposing an osmotic gradient, and that the rate of active fluid
secretion was correlated with the transepithelial osmotic gradient generated by salt
transport.
3.5 Discussion

103
Figure 3.8 Osmolality determine the fluid secretion rate.
A, without stimulation by forskolin, Calu-3 cell monolayers were bathed apically with solutions at different osmolality values. Different fluid secretion (absorption) rates were observed accordingly to the osmolality (n=6 of each). B, with stimulation by forskolin, apical solution at 290 mOsm greatly reduced the fluid secretion rate, when compared to control (without apical solution)(n=6 of each). C, a time course of the fluid and the culture medium osmolality. Fluid secretion and the culture medium were collected every hour during the first 5 h and one final time at 24 h. the osmolality was measured. During the rapid secretion phase, the osmolality of the fluid was generally higher than the one of the medium. However, the osmolality of the fluid and the medium eventually became the same at the end of 24 h fluid secretion assay (n=6 of each).

104
In the present study, we compared fluid secretion by CFTR-expressing and
CFTR-deficient cell lines, investigated the ionic requirements for fluid secretion, and
studied aspects of fluid transport regulation. The results are consistent with those in
the previous chapter, and with the anion transport model we have proposed for
Calu-3 cells.
3.5.1 Most fluid is driven by the net flux of Cl−
The [Cl-] of secretions was ∼120 mmol·l−1, or about 4-fold higher than the [HCO3−].
This implies that most fluid secretion is driven by transepithelial Cl− rather than
HCO3− flux during forskolin stimulation despite the fact that active Cl− transport is not
detected under Isc conditions (Devor, Singh et al. 1999) or open circuit conditions
(this thesis), and that Isc can be fully explained by the net HCO3− flux. Using the [Cl-]
of secretions, we calculate that during a 24 h fluid secretion assay, the Cl- secretion
rate is ~0.2 μeq·cm−2·h−1, a number much smaller than the Cl- component of the Ieq
(1.5 μeq·cm−2·h−1). This calculation clearly shows that a very low rate of Cl− transport
would be sufficient to sustain fluid secretion at the observed rate. Such low rates are
difficult to detect using radiotracer fluxes because they are the difference between
two large unidirectional fluxes, and because there is only one useful radioisotope of
Cl- thus unidirectional fluxes must be measured across different monolayers, which
introduces variability larger than the net flux.
A related finding was that much of the Cl− transport which drives fluid secretion is

105
independent of NKCC1. This conclusion was based on anion selectivity and
pharmacological data. Robust fluid secretion was observed for 24 h when the
basolateral side was bathed with nominally Cl−-free solution containing NO3−, which
does not bind to the highly selective anion site on NKCC transporters and therefore is
not transported (Kinne, Kinne-Saffran et al. 1986). Further evidence for
NKCC1-independent Cl− entry is the relative insensitivity of fluid secretion to
bumetanide. These results suggest other basolateral Cl− entry pathways such as
anion exchangers are important for fluid secretion. NO3− is carried by many anion
exchangers including AE2 (Humphreys, Jiang et al. 1994).
Although HCO3− is secreted by Calu-3 cells under open-circuit conditions, its
concentration in the secreted fluid is low relative to Cl−. Thus HCO3- seems to be
involved in Cl- loading via basolateral anion exchange rather than driving the fluid
secretion. The high concentration of Cl− in secretions suggests that it together with
electrically coupled Na+ flux provides most of the osmotic driving force for fluid
secretion.
3.5.2 Calu-3 secretions are only weakly alkaline
The normal pH of ASL and gland secretions is in the range 6.2–7.2 (Fischer and
Widdicombe 2006). Weakly buffered solutions placed on cultures of surface
epithelium become acidic, and pH falls more rapidly on CF than non-CF monolayers,
consistent with a defect in parallel HCO3− secretion (Coakley, Grubb et al. 2003).

106
Calculations based on the pH of gland fluid collected in the presence vs. absence of
HCO3− (Song, Salinas et al. 2006) suggests that these secretions normally contain
about 14 mmol·l−1 HCO3−. Moreover, secretions from pig glands are 0.2–0.4 units
more acidic when secreted in the presence of CFTR inhibitors, mimicking the low pH
seen with CF glands (Song, Salinas et al. 2006).
HCO3−-driven fluid transport was expected to generate strongly alkaline secretions
having pH > 8; however, the maximal pH during forskolin stimulation in the present
experiments was ∼7.6. The calculated [HCO3−] in secretions was slightly higher than
in the basolateral medium (33 vs. 25 mmol·l−1), and higher apical [HCO3−] was
achieved by parental Calu-3 monolayers, which have significantly higher CFTR
expression (∼50 mm [HCO3−]; D. Kim, unpublished observations, Fig. 2A);
nevertheless, secretions were always much less alkaline than expected if the fluid
were driven significantly by HCO3− transport. The values obtained with parental cells
were generally consistent with previous studies in a virtual gland preparation (74 mM
HCO3− (Irokawa, Krouse et al. 2004)), and in cyclophilin B shRNA control Calu-3 cells
cultured on transwells (60 mM (Garnett, Hickman et al. 2011)). Silencing CFTR
reduced the [HCO3−] of secretions by ∼50% in the present study, but had no effect
on the pH and HCO3− concentration in a previous study (Garnett et al. 2011), perhaps
because the knock-down cells used in that study had higher residual CFTR expression
(28% vs. < 5% in the present work). The present results are consistent with the
decreased [HCO3−] reported for CF gland secretions (Song, Salinas et al. 2006),

107
although the starting value in glands was lower (i.e. 3.5 mmol·l−1 vs. 13.8 mmol·l−1
[HCO3−] in CF vs. non-CF glands, respectively).
In the present work, the total volume secreted and Cl− content were both reduced
10-fold by silencing CFTR expression and the [HCO3−] was reduced by 2-fold. Similar
changes in CF airways may contribute to disease symptoms by altering mucus release
or rheology, innate immunity or ciliary beating (Shan, Huang et al. 2011).
3.5.3 A very low rate of net Cl− secretion is sufficient to drive fluid
secretion
HCO3− secretion accounted for ∼40% of the Ieq under pH-stat conditions. The
remainder (1.5 μeq·cm−2·h−1) was carried by Cl−, as revealed by measuring 36Cl−
fluxes under the same conditions. Nevertheless, Cl− net flux was negligible during
previous short-circuit current experiments (Devor et al. 1999) and under open-circuit
conditions in the present study (Table 1), making it difficult to understand how Calu-3
monolayers could produce a Cl−-rich fluid. However, it is important to note that a net
Cl− flux of only 0.2 μeq·cm−2·h−1 would be sufficient to account for the Cl− content of
secretions in the present, and such low rates cannot be determined reliably in Ussing
chambers.
3.5.4 The osmolality determines the fluid secretion rate
The optimal height of ASL is ~7 μm, which is also the height of outstretched cilia. The

108
height of ASL is fine tuned in the airways, but it is not clear how airway epithelia
sense the volume and adjust the fluid secretion rate accordingly. Some studies
hypothesized that cilia of airway epithelial cells might act in a mechanosensory
capacity to signal ASL volume status to the underlying epithelia (Tarran, Grubb et al.
2001, Chambers, Rollins et al. 2007). However, a role for cilia in mechanically sensing
ASL volume has not been demonstrated. Other studies have suggested that soluble
stimulatory molecules such as adenosine and channel-activating proteases or sodium
channel regulators such as SPLUNC1 in the ASL may function as chemosensors that
regulate the balance between secretion and absorption (Garcia-Caballero,
Rasmussen et al. 2009).
Although Calu-3 does not maintain a constant ASL volume equivalent to surface
airway cells, our studies suggest a simple mechanism for regulating this balance in
which osmotic driving force created by secreted solutes determines fluid secretion
rate and steady-state volume. Firstly, our data from the evaporation test suggest that
the osmotic driving force created by forskolin stimulated ion secretion drives a
passive flux of solute-free water, and hypertonicity caused by evaporation has a
similar effect, drawing water from the basolateral side into the apical compartment
to compensate for that lost by evaporation.
Secondly, incubating Calu-3 monolayers with solutions having high or low osmolality
caused water flows without forskolin stimulation and rapidly generated absorption or

109
secretion. However adding isotonic solution on the apical side reduced the active
fluid secretion rate, presumably by diluting the local osmotic driving force created by
forskolin stimulated salt secretion. Thirdly, during the initial fast rate of secretion, the
osmolality of secretions was always slightly higher than the basolateral medium,
again suggesting that the osmotic driving force due to secretion drives fluid transport.
Later during the slow secretion phase, when some fluid had accumulated in the
apical compartment, the osmotic driving force was reduced and fluid was secreted at
a slower rate.
All of the present data suggest that osmolality determines fluid secretion rate and
thus regulates the ASL volume. In this simple view, osmotic pressure is both the
sensor and driving force for water flux, and luminal dessication caused by inhaled air
and sodium absorption by ENaC ensures that there is never too much ASL.
3.5.5 Carbonic anhydrase participates little in fluid secretion
In the previous chapter, we found that acetazolamide abolishes forskolin-stimulated
anion transport, indicating an important role of carbonic anhydrase in anion
transport. We therefore anticipated that carbonic anhydrase would also participate
actively in fluid secretion during forskolin stimulation. However, acetazolamide
caused only 27% inhibition of fluid transport when added before forskolin or acutely
during forskolin stimulation. We investigate the discrepancy between the impact of
carbonic anhydrase inhibitor on anion transport and fluid secretion in the next

110
chapter.

111
Chapter 4 The role of carbonic anhydrase in anion and fluid secretion

112
In previous chapters we found that pharmacological inhibition of carbonic anhydrase
dramatically reduced transepithelial anion transport but had only a modest effect on
the rate of fluid secretion. Therefore in this chapter we investigate further the role of
carbonic anhydrases in anion and fluid secretion by Calu-3 cells.

113
4.1 Abstract
Carbonic anhydrase is a ubiquitous metalloenzyme which catalyzes the hydration of
carbon dioxide to carbonic acid. The role of this enzyme in bicarbonate production
and transepithelial secretion has been demonstrated in many tissues, including the
airways. However, our results in previous chapters revealed a discrepancy between
the effect of a carbonic anhydrase inhibitor on anion transport and its effect on fluid
secretion, prompting us to study its role further. Without any contribution of NBC to
basolateral bicarbonate entry, forskolin still increased the equivalent short-circuit
current (Ieq) across Calu-3 monolayers although less than when NBC is operating.
Since we showed that bicarbonate is essential for Ieq this suggests that carbonic
anhydrase contributes significantly to transepithelial bicarbonate transport.
Expression of the carbonic anhydrase isozymes CAII, CAIX and CAXII was
demonstrated at the mRNA level by PCR and at the protein level by immunoblotting.
Attempts to co-immunoprecipitate CAII with CFTR were not successful, which argues
against a physical complex between these proteins (i.e. a metabolon), at least under
these conditions. Acetazolamide inhibited only the forskolin stimulated component
of Ieq attributable to bicarbonate, it did not affect the component mediated by
chloride flux through CFTR, therefore it did not act by blocking CFTR channels. CAIX is
often upregulated in cancer cells and was robustly expressed in Calu-3, which is a
tumor cell line, however the relatively CAIX-selective inhibitor
4-(2-aminoethyl)-benzenesulfonamide had no effect on forskolin-stimulated Ieq or
bicarbonate secretion, suggesting CAIX plays little role in anion and fluid secretion.

114
Taken together, the results suggest that bicarbonate production by the CAII isoform
may be important for anion and fluid secretion across Calu-3 cells.
4.2 Introduction
Carbonic anhydrase is a zinc metalloenzyme which catalyzes the reversible hydration
of CO2 to carbonic acid, which then dissociates spontaneously to protons and
bicarbonate ions. Carbonic anhydrase (CA) is expressed ubiquitously in all living
organisms, and 15 distinct isoforms have been identified in humans (Esbaugh and
Tufts 2006). The isozymes can be divided into three groups, the cytoplasmic CAs,
membrane-bound CAs, and the CA-related proteins which have lost their catalytic
activity. CAII is the most prevalent isoform and belongs to the cytoplasmic group. It
has a high enzymatic turnover rate of 106 s-1 and is thought to mediate much of the
CA activity in various tissues including airways (Khalifah 1971, Esbaugh and Tufts
2006, Purkerson and Schwartz 2007). On the other hand, some membrane-bound
CAs become over-expressed and may have roles in pathological states, for example
cancers. Examples of these isoforms include CAIX and CAXII, which are associated
with cancer progression and metastasis (McDonald, Winum et al. 2012).
Previous studies suggested that carbonic anhydrase is important for anion transport
by airway epithelial cells (Welsh and Smith 2001, Cuthbert, Supuran et al. 2003, Yue,
Lau et al. 2008). When CFTR was activated directly or indirectly, 35-50% of the anion
secretion was dependent on carbonic anhydrase based on the inhibition caused by

115
acetazolamide. The contribution of carbonic anhydrase is presumed to be due to its
production of endogenous bicarbonate. The CAII isoform may bind to the
intracellular C-terminal region of various transport proteins through an acidic motif
in its N-terminus, forming a functional complex that facilitates bicarbonate transport
called a metabolon (McMurtrie, Cleary et al. 2004). These partners include AE1, AE2,
NBC1, NBC3 and NHE1, some of which are essential for anion and fluid secretion by
Calu-3 cells as discussed in previous chapters. It should be noted however that the
idea of carbonic anhydrase forming metabolons remains controversial (Piermarini,
Kim et al. 2007).
CAIX and CAXII are over-expressed in tumors due to activation of a hypoxia
responsive element in their promoter, thus they have become important therapeutic
targets for the treatment of cancer (McDonald, Winum et al. 2012). Expression of
cancer-related carbonic anhydrases is restricted in normal tissues, and genetic
disruption of CAIX in mice results in a mild phenotype (Gut, Parkkila et al. 2002). It
seems that CAIX and CAXII play only minor, if any, role in bicarbonate generation in
normal cells when CAII is present. Interestingly, CAIX expression is upregulated
~2-fold in mice that are null for CAII (Pan, Leppilampi et al. 2006), suggesting that
CAIX compensates for the loss of functional CAII in this setting. Calu-3 is a widely
used model for airway epithelial cells and it is also an adenocarcinoma cell line,
therefore CAIX and CAXII could play important roles in the generation of endogenous
bicarbonate and the secretion of anions and/or fluid in this system.

116
CAII is the most widely expressed and usually the most abundant carbonic anhydrase
isoform, however carbonic anhydrases have not been characterized in airway
epithelial cells. In the present study we identified CA isoforms in Calu-3 cells and
studied their potential roles in anion and fluid secretion. We found that carbonic
anhydrase activity was able to partially sustain forskolin-stimulated Ieq
independently of basolateral NBC. Though CAII, CAIX and CAXII were the most
prominent isoforms, the CAIX-selective inhibitor 4-(2-aminoethyl)-
benzenesulfonamide had a negligible impact compared to the broad-spectrum CA
inhibitor acetazolamide. On the other hand, CAXII-specific inhibitor U-104
(1-(4-fluorophenyl)-3-(4-sulfamoylphenyl)urea) partially inhibited the
forskolin-stimulated Ieq and HCO3- secretion. These results suggest that CAII, but not
CAIX, is important for anion and fluid secretion across Calu-3 cells, presumably by
contributing some endogenous bicarbonate.
4.3 Methods
4.3.1 Cell Culture
Two Calu-3 cell lines provided by Dr. Scott O’Grady were used, a parental line with
normal CFTR expression and a CFTR-deficient cell line in which CFTR was knocked
down by stable expression of shRNA that targets CFTR mRNA transcripts. Both cell
lines were cultured in EMEM containing 7% FBS. Some control experiments were also
performed using parental Calu-3 cells (HTB-55, American Type Culture Collection,

117
Manassas, VA) cultured in Eagle’s minimum essential medium (EMEM) containing 15%
fetal bovine serum (FBS) to allow comparison with previous studies.
Cells were seeded at 5 x 105 cells per cm2 on SnapwellsTM (1.12 cm-2; Costar, Corning
Life Sciences, Lowell, MA) when measuring Ieq and HCO3- secretion, or on Transwells
(4.67 cm2, Corning) for experiments involving RT-PCR, immunoblotting,
co-immunoprecipitation and fluid secretion assays. Fresh medium was placed on the
basolateral side one day after plating and the apical medium was removed to
establish air-liquid interface (ALI) conditions. Any fluid that appeared spontaneously
on the apical surface after 3 days was removed. Cultures were maintained in a
humidified, 5% CO2 incubator at 37℃ for 21-25 days.
4.3.2 Solutions
To study the contribution of carbonic anhydrase to forskolin-stimulated Ieq and
HCO3- secretion, monolayers in the Ussing chamber were bathed apically with acidic
solution (pH was clamped at 6.0 by an automated titration workstation; TitraLab 854,
Radiometer) containing (mmol·l-1): 120 NaCl, 5 KCl, 25 NaHCO3, 1.2 MgCl2, 1.2 CaCl2.
The apical solution was constantly gassed with 95% O2/5% CO2. The basolateral side
was bathed with 120 NaCl, 3.3 KH2PO4, 0.8 K2HPO4, 1.2 MgCl2, 1.2 CaCl2, 10 HEPES
and 10 glucose, gassed with 100% O2. The nominally HCO3--free solution blocked
activity of basolateral NBCe1, while the apical solution provided CO2 to support the
activity of intracellular carbonic anhydrase. The pH of the apical solution was

118
clamped at 6 to minimize bicarbonate accumulation and avoid significant buffer
capacity that would preclude pH-stat measurements of HCO3- secretion.
4.3.3 Measurements of equivalent Isc and HCO3- secretion
Inserts were mounted in modified Ussing chambers (Physiologic Inst., San Diego, CA)
at 37℃. HCO3- secretion was measured under open-circuit conditions, and was
compared with the equivalent short-circuit current (Ieq) calculated from Ohm’s law
using the spontaneous transepithelial potential (Vt) and transepithelial resistance
(Rt). Rt was determined from the small deflections in Vt produced by bipolar current
pulses (1 μA, 1 sec duration, 99.9 sec interval) which were delivered by the voltage
clamp (VCC200, Physiologic Instr.). Data were digitized (Powerlab 8/30, AD
Instruments, Montreal QC) and analyzed using Chart5 software.
HCO3- transport was measured using the pH-stat method under open-circuit
conditions. A mini-pH electrode (pHG200-8, Radiometer Analytical) connected to an
automated titration workstation (TitraLab 854, Radiometer) delivered 1 μl aliquots of
10 mmol l-1 HCl to the apical solution to maintain the pH constant at 6.000 ± 0.002.
The amount of acid required for this was used to calculate the rate of HCO3-
secretion. The volume of each half chamber was 4 ml. Solutions containing 25 mmol
l-1 HCO3- were stirred vigorously with 95% O2/5% CO2. Nominally HCO3
--free
solutions were bubbled with 100% O2.

119
4.3.4 RT-PCR
Total RNA was isolated using the RNAqueous-4PCR extraction kit (Ambion) according
to the manufacturer's instructions. Samples were treated with DNase (Ambion) and
reverse transcription was performed using Superscript III (Invitrogen) and 2 μg of
RNA primed with 200 ng random hexamers and 50 μmol l−1 oligo(dT). mRNAs for
each CA isoform were detected as described previously (Tarun, Bryant et al. 2003).
Isoform-specific primers were used and RT-PCR was performed using a PTC-100
Thermal controller (MJ Research) with 30 ng of each primer in 20 μl PCR reaction
buffer. Amplification was carried out using the following thermal cycling conditions:
initial denaturation (2 min at 95°C), 15 cycles of touchdown PCR (95°C for 45 s, 75°C
(–1℃ for each subsequent cycle) for 1 min) and 30 cycles of two-step PCR (95°C for
45 s, 60°C for 1 min). For analysis of the PCR products, 10 µl of the PCR reactions
were loaded onto a 2% TAE (Tris acetate EDTA) agarose gel and subjected to
electrophoresis. After staining with ethidium bromide (0.5 µg/ml), the gel was
photographed. We used GAPDH as an endogenous control to normalize the results.
4.3.5 Immunoblotting
After SDS-PAGE on 12% gels, proteins were transferred to nitrocellulose membranes
and probed using antibodies against: CAII (1:50000, ab6621, Abcam), CAIX (1:2000,
ab107257, Abcam), Na+/K+-ATPase (1:200, mAb gift from R.W. Mercer, Washington
Univ., St. Louis MO; (Takeyasu, Tamkun et al. 1988)), β-actin (1:500, SC-1615, Santa
Cruz). Blots were washed, incubated with secondary antibody conjugated to

120
horseradish peroxidase (1:1000), and visualized with enhanced chemiluminescence
(Amersham Biosciences).
4.3.6 Immunoprecipitation
Calu-3 cell monolayers seeded on Transwells were treated with 10 μM forskolin or
DMSO for 24 h before protein crosslinking by Dithiobis(succinimidyl propionate)
(DSP). After crosslinking, cells were rinsed with PBS and lysed. CFTR was
immunoprecipitated on TrueBlot™ anti-mouse Ig IP beads (00-8811, eBioscience)
using the monoclonal antibody M3A7 (1:1000, gift from J.R. Riordan and T.J. Jensen,
UNC Chapel Hill, NC (Kartner and Riordan 1998)) by incubation for 1 h at 4℃. Beads
were then carefully washed to retrieve samples, whichwere subjected to 8%
SDS-PAGE. After transfer to nitrocellulose membrane, protein complexes were
probed using anti-CAII antibody (1:10000, ab6621, Abcam). Blots were washed,
incubated with secondary antibody conjugated to horseradish peroxidase (1:1000),
and visualized using enhanced chemiluminescence (Amersham Biosciences).
4.3.7 Fluid secretion assay
At the beginning of each assay, any fluid in the apical compartment of the Transwells
was removed. To eliminate the effects of serum and maintain viability during long
fluid secretion experiments, EMEM medium without serum but containing essential
amino acids was placed on the basolateral side. Calu-3 cell monolayers bathed with
basolateral 25 mmol·l−1 HCO3− medium were kept in 5% CO2–95% air, which was

121
nominally saturated with H2O during fluid secretion measurements. Chemicals were
added to the basolateral medium to avoid disrupting the air-liquid interface. After 24
h treatment, fluid secreted by cell monolayers was collected and the volume was
measured.
4.3.8 Data analysis
Isc or Ieq was determined at 100 sec intervals, and HCO3- net flux rate was calculated
every 5 min. Basal values were those obtained immediately before adding forskolin.
Paired or unpaired student’s t-tests with p<0.05 were used for single comparisons.
4.4 Results
4.4.1 Carbonic anhydrase contributes partially to anion transport
To investigate the effects of carbonic anhydrase on anion transport by airway
epithelial cells, we bathed the basolateral side of Calu-3 cell monolayers with
nominally HCO3-- (and CO2-) free solution gassed with 100% O2 to prevent
basolateral NBC from transporting bicarbonate into the cell. However, this should not
prevent the intracellular production of bicarbonate by carbonic anhydrase if CO2 is
supplied apically.
The apical solution was gassed with 95% O2/5% CO2 and clamped at pH 6 so that
pCO2 could be supplied to the cells while bicarbonate secretion is measured using
the pH-stat technique. Under these conditions, the apical solution should still

122
provide CO2 for carbonic anhydrase-catalyzed synthesis of HCO3- but would have
minimal buffer , enabling the measurement of HCO3- secretion by the monolayer.
Basal Ieq and HCO3- secretion under these conditions were similar to those measured
normal solutions (Fig. 4.1, compare with Fig. 2.2B). Applying forskolin again elicited a
transient peak Ieq followed by a plateau (Fig. 4.1), although the peak and plateau
were both reduced somewhat, probably due to the absence of basolateral HCO3- and
thus NBC activity. Based on comparison of the ΔIeq under these conditions with
control ΔIeq, we estimated the contribution of carbonic anhydrase towards
forskolin-stimulated anion current to be 35-40%, within the range reported
previously by other groups (Cuthbert, Supuran et al. 2003, Yue, Lau et al. 2008).
Interestingly, very little acid had to be added to the apical side to maintain pH
constant under these conditions, which implies that little of the forskolin-stimulated
current was carried by HCO3- secretion. The simplest interpretation is that
bicarbonate generated endogenously by carbonic anhydrase was exchanged for Cl- at
the basolateral membrane, and that Ieq was mainly due to net transepithelial Cl-
secretion under these conditions.
4.4.2 The expression of different isoforms of carbonic anhydrase
Two isoforms of carbonic anhydrase, CAIX and CAXII are commonly over-expressed in
cancer cells (McDonald, Winum et al. 2012). As Calu-3 is an adenocarcinoma cell line,

123
Figure 4.1 Carbonic anhydrase contributes partially to anion transport
by Calu-3 cells. Calu-3 cell monolayers were bathed basolaterally with nominally HCO3
-/CO2-free solution gassed with 100% O2. The apical unbuffered solution was gassed with 95% O2/5% CO2 and clamped at pH 6. 10 μM forskolin was added after basal Ieq and HCO3
- became stable (n=4). ●, Ieq; ○, HCO3- secretion.

124
it would not be surprising if it also had up-regulated CAIX and CAXII expression. This
raises the question “Is endogenous bicarbonate synthesis mediated by CAIX and/or
CAXII, in addition to other CA isoforms such as CAII?”
To answer this question, we began by determining the expression of the CA isoform
genes using the reverse transcriptase polymerase chain reaction (RT-PCR). The
RT-PCR revealed that CAII, CAIX and CAXII were highly expressed at the mRNA level
(Fig. 4.2A). Unfortunately, an antibody against CAXII is not available and we could not
confirm its expression at the protein level, however expression of CAII and CAIX
proteins was confirmed by immunoblotting (Fig. 4.2B, C). Both carbonic anhydrases
were exuberantly expressed in wild-type Calu-3 cells and CFTR-deficient cells stably
expressing shRNA against CFTR. Carbonic anhydrase expression was not altered by
raising intracellular cAMP with forskolin.
4.4.3 CAIX is not involved in anion and fluid secretion by Calu-3 cells
Our focus shifted to CAIX because of its up-regulation in cancer cells. We
hypothesized that CAIX might be a source of bicarbonate for chloride exchange and
that the 35-40% contribution carbonic anhydrase contribution to anion transport was
the combined efforts of CAIX and CAII.
To test this, we used 4-(2-aminoethyl)-benzenesulfonamide, a potent inhibitor which
is selective for CAIX over CAII (Svastova, Hulikova et al. 2004) in contrast to the

125
Figure 4.2 The expression of different isoforms of carbonic anhydrases
in Calu-3 cells.
A, mRNA expression of different carbonic anhydrases in Calu-3 cells. Lanes from the left to the right were ladder, CAI, CAII, CAIII, CAIX, CAXa, CAXb, CAXI, CAXII, CAIX, CAXII, CAXIII, CAXIX and GAPDH. The left graph was Calu-3 and the right one was negative control. B, CAII expression in Calu-3 cells. Lane 1 & 2, wild-type Calu-3 cells without forskolin stimulation; Lane 3 & 4, wild-type Calu-3 cells with 24 h forskolin stimulation; Lane 5 & 6, CFTR-deficient Calu-3 cells without forskolin stimulation; Lane 7 & 8, CFTR-deficient Calu-3 cells with 24 h forskolin stimulation. C, CAIX expression in Calu-3 cells. Lane 1 & 2, wild-type Calu-3 cells without forskolin stimulation; Lane 3 & 4, wild-type Calu-3 cells with 24 h forskolin stimulation; Lane 5 & 6, CFTR-deficient Calu-3 cells without forskolin stimulation; Lane 7 & 8, CFTR-deficient Calu-3 cells with 24 h forskolin stimulation.

126
non-specific CA inhibitor acetazolamide. However, during forskolin stimulation,
adding 4-(2-aminoethyl)- benzenesulfonamide at concentrations up to 100 µM did
not alter Ieq or HCO3- secretion (Fig. 4.3). Fluid secretion assays with
4-(2-aminoethyl)-benzenesulfonamide were also consistent with the findings from
Ussing chambers (data not shown). The volume of fluid secreted by Calu-3
monolayers was not affected by the CAIX inhibitor. These results suggest that CAIX
is expressed but plays little if any role in anion and fluid secretion by Calu-3 cells.
4.4.4 CAII and CAXII are responsible for the forskolin-stimulated anion
secretion in Calu-3 cells
As CAIX was excluded, only two candidates were left, one was CAII, the other was
CAXII. I used the CAXII-specific inhibitor U-104 to test if the enzyme was involved in
the forskolin-stimulated response in Calu-3 cells.
As Fig. 4.4 shows, applying U-104 (10 μM, a concentration to inhibit CAXII) after
forskolin partially inhibited the Ieq and HCO3- secretion, indicating CAXII contributed
to the anion transport.
Though I didn’t use any CAII-selective inhibitor to test the involvement of the enzyme,
it is clear that CAII also contributed to the anion transport in Calu-3 cell when
stimulated, because only two candidates were possibly responsible and CAXII
contributed partially, CAII must be the contributor to the remaining Ieq and HCO3-

127
Figure 4.3 CAIX is not involved in anion transport by Calu-3 cells.
The inhibitor of CAIX, 4-(2-aminoethyl)-benzenesulfonamide, did not affect forskolin-stimulated anion secretion by Calu-3 cells (n=3). ● , Ieq; ○ , HCO3
- secretion.

128
Figure 4.4 CAXII contributes partially to the anion transport by Calu-3
cells.
The inhibitor of CAXII U-104 (10 μM) partially inhibited the forskolin-stimulated anion secretion by Calu-3 cells (n=3). ●, Ieq; ○, HCO3
- secretion.

129
secretion.
4.4.5 CAII does not form a complex with CFTR
In the previous two chapters, we discovered a discrepancy between the role of CA in
anion transport and fluid secretion. Inhibiting CA with acetazolamide abolished
forskolin-stimulated Ieq and HCO3- secretion, but in the fluid secretion assay it caused
only ~30% inhibition. Enzyme inhibition by acetazolamide might be expected to
cause slow changes in bicarbonate transport and pH, however its effects were very
rapid, therefore we initially suspected that acetazolamide might abolish Ieq by
blocking CFTR. However the non-HCO3- component of Ieq was not inhibited by
acetazolamide, although it was presumably mediated by CFTR. This suggests that
acetazolamide is not a blocker of the CFTR pore.
Since carbonic anhydrase is thought to form a metabolon with acid/base transporters
which facilitates bicarbonate flux (McMurtrie, Cleary et al. 2004), we wondered if
acetazolamide might act by disrupting a complex between CFTR and CAII, the
predominant carbonic anhydrase in most cells.
To test this, we immunoprecipitated CFTR, separated the proteins by SDS-PAGE, and
probed the resulting blots with antibody against CAII to see if it was co-precipitated.
However no CAII was detected in the CFTR precipitates (Fig. 4.4). This suggests that
CAII and CFTR do not form a stable complex and makes it unlikely that acetazolamide

130
inhibits CFTR function by disrupting its association with CAII.
4.5 Discussion
Results in the current chapter support the involvement of carbonic anhydrase in
anion and fluid secretion by Calu-3 cells, with the former being more strongly
affected.
4.5.1 Carbonate anhydrase II is important for anion and fluid secretion
The role of CAII is suggested by several lines of evidence: 1) acetazolamide-sensitive
carbonic anhydrase maintained basal secretion when the basolateral HCO3- and NBC
was unable to supply HCO3- (Fig. 4.1), 2) CO2 was equally effective in supporting
basal secretion when supplied from the basolateral or apical side, compatible with its
hydration by a soluble carbonic anhydrase that is distributed throughout the cell, and
3) the data suggest that carbonic anhydrase contributes 30-40% of the
forskolin-stimulated anion and fluid secretion by Calu-3 cells through its ability to
generate endogenous bicarbonate.
Three carbonic anhydrases were identified in Calu-3 cells, the ubiquitously expressed
soluble CAII, and the membrane-bound forms CAIX and CAXII. Although all three
could potentially play a role in bicarbonate handling, the insensitivity of HCO3-
secretion to an inhibitor of CAIX suggested it plays little role in anion and fluid
secretion. Only the other hand, CAII and CAXII were showed to contribute to the

131
Figure 4.4 Co-immunoprecipitation between CAII and CFTR.
Wild-type Calu-3 cell protein (400 μg) was immunoprecipitated with CFTR antibody, and then blotted with CAII antibody. Lane 1, sample protein immunoprecipitated with CFTR antibody; Lane 2, sample protein without CFTR antibody; Lane 3, total lysate blotted against CFTR and CAII antibody.

132
anion secretion by Calu-3 cells. However the fact that CAIX and CAXII are normally
expressed at low levels and only become upregulated in adenocarcinomas suggests
that CAII may be the one that supports secretion in normal airway epithelial cells.
4.5.2 Further evidence that HCO3- is required for Cl- secretion
We showed in previous chapters that Cl- is the main anion secreted by Calu-3 cell
monolayers under physiological (open-circuit) conditions. This prominence is most
obvious when the activity of NBC is blocked by removing HCO3- from the basolateral
solution. Without functional NBC so that carbonic anhydrase supplied all the
bicarbonate to the cell, transepithelial bicarbonate secretion was reduced to 30-40%
that measured under normal conditions, and the bicarbonate generated by carbonic
anhydrase was insufficient to drive Cl- and HCO3- secretion via CFTR. Thus it seems
that airway epithelial cells have a mechanisms that preferably exchanges HCO3- for
Cl- at the basolateral membrane, so that Cl- becomes the main anion secreted under
these conditions (Fig. 4.1). The mechanisms were not investigated in detail in this
study, however we speculate that the selectivity of CFTR for Cl- over HCO3- and the
relative affinities of some transporters, e.g. AE2, for these anions may account for
this preferential exchange at the basolateral membrane. Moreover, as we found in
the previous chapters, bicarbonate supports chloride secretion but does not drive
fluid transport significantly in Calu-3 cells. Thus it is that airway epithelial cells would
humidify the airway lumen by secreting Cl- and fluid (rather than HCO3-) and recycle
HCO3- at the basolateral membrane when there is insufficient HCO3
- available to

133
support transepithelial flux of both anions.
4.5.3 The bicarbonate transport metabolon
CAII is thought to form complexes with other transport proteins that mediate
bicarbonate transport. These transporters include AE1, AE2, NBC1, NBC3 and NHE1
(McMurtrie, Cleary et al. 2004). Bicarbonate transport metabolons may exist in
Calu-3 cells since they express AE2 and NBC1, however our experiments did not
provide any evidence for such an association with CFTR.
Calu-3 cells need a supply of HCO3- to maintain HCO3
- secretion and also to support
basolateral Cl- entry by anion exchange during forskolin-stimulated Cl- secretion. CAII
may function in de novo HCO3- synthesis by generating HCO3
- from intracellular CO2.
We speculate that any HCO3- entering on NBCe1 which is not recycled by basolateral
anion exchange for Cl- becomes neutralized by the intracellular acid load that is
generated when HCO3- exits through CFTR. The resulting dehydration would produce
CO2 near CFTR which could then be rehydrated locally by carbonic anhydrase.
Alternatively, since CO2 diffuses rapidly, the basolateral supply of HCO3- that
supports Cl- entry via anion exchange may depend on a basolateral CAII-AE
metabolon that increases the efficiency of basolateral HCO3- extrusion.
We used a membrane-permeant crosslinking reagent in our attempt to capture
proteins complexed with CFTR, however we cannot exclude the possibility that CFTR

134
and CAII were weakly associated and the metabolon was disrupted during the
immunoprecipation, or during long-term experiments such as the fluid secretion
assay, which requires 24 h. Moreover, as shown in Chapter 3, active fluid secretion
was rapid initially but slowed dramatically after the first 6 h. The complex might be
disrupted at the beginning of the assay but remain undetected functionally because
CAII and NBC working independently can compensate during a 24 h assay.
Bicarbonate metabolons might also be inconspicuous if one of the interacting
proteins was inhibited before the supply of HCO3- became rate-limiting. This could
occur when Calu-3 monolayers are pre-treated with acetazolamide; i.e. if inhibiting
CAII beforehand prevented the formation of metabolons, this could potentially
explain why acetazolamide caused less dramatic inhibition of the forskolin-stimulated
secretion than during acute acetazolamide exposure.
4.5.4 The contributions of CAII and NBC to anion and fluid secretion
In our revised model for anion and fluid secretion by airway epithelial cells, CAII and
NBC both supply bicarbonate. When they are combined, some HCO3- is exchanged
for Cl- via basolateral AE and both anions are secreted through CFTR. Basolateral
NKCC also supplies Cl- which becomes predominant during stimulation by
secretagogues that hyperpolarize the cell (Devor, Singh et al. 1999), however during
forskolin (cAMP) stimulation it seems to play a minor role compared with AE2.

135
Chapter 5 General discussion

136
The first clear description of CF appeared in 1938 when Dorothy Anderson published
her paper entitled “Cystic fibrosis of the pancreas and its relation to celiac disease ”
(Lubamba, Dhooghe et al. 2012). Chronic respiratory infection was recognized as one
of the major symptoms, thus antibiotics were introduced for the treatment of CF in
the 1940s. As stated above, salty tasting skin is one of the hallmarks of CF and results
from increased secretion of Cl- and Na+ by the defective sweat glands, therefore the
diagnostic pilocarpine sweat test was brought into use in 1959 (Gibson and Cooke
1959). In the 1980s, the understanding of CF pathophysiology progressed rapidly
with reports of altered Na+ transport across CF respiratory epithelia (Knowles, Gatzy
et al. 1981), and impermeability of the CF sweat gland to Cl- (Quinton 1983, Quinton
and Bijman 1983). Another milestone in the CF field was reached in 1989 when the
CF gene was cloned (Kerem, Rommens et al. 1989, Riordan, Rommens et al. 1989,
Rommens, Iannuzzi et al. 1989). During the following decades, enormous progress
towards understanding of the molecular biology of the CF gene and its protein
product CFTR has firmly established that loss of CFTR function and/or expression is
the basic defect in CF. Nevertheless, the mechanisms of transepithelial ion transport
in the airways and the precise role of CFTR in CF pathogenesis has remained poorly
understood. The major goal of this thesis was to characterize the ion transport and
the related fluid secretion in airway epithelial cells.
In chapter 2, I focused on transepithelial anion transport by the polarized airway
epithelial cell line Calu-3. The airway epithelial monolayer forms a barrier with

137
transporters and channels on both sides (i.e. in the apical and basolateral
membranes). The results are consistent with the basolateral transporters NKCC, NBC
and AE mediating uptake of Cl- and HCO3- and are secreted mainly through apical
CFTR channels. In chapter 3, I investigated fluid secretion by airway epithelial cells,
which is driven by ions, or more accurately, by the osmolality gradient generated by
salt secretion. The results in chapter 3 confirmed the model for anion transport that I
proposed in chapter 2. In chapter 4 carbonic anhydrase was the main focus, and was
pursued due to the dramatic effect of inhibiting this enzyme on bicarbonate
transport and the important role of endogenous bicarbonate generation suggested in
chapter 2 and 3. I attempted to characterize its different isoforms and their
functions.
5.1 A novel model of anion transport of airway
epithelial cells
The results led to a novel model for the transepithelial anion transport in airway
epithelial cells. Before going into details of the model, I should discuss some of the
assumptions and limitations of the experimental design.
5.1.1 V-clamp vs. I-clamp
Short-circuit current (Isc) measurements are the most widely used method for
monitoring transepithelial ion transport (Ussing and Zerahn 1999). In this technique,
the transepithelial voltage is clamped at 0 mV by injecting current in the opposite

138
direction to that generated by the cells (V-clamp). There are no transepithelial
electrical or chemical gradients under classical short-circuit current conditions, thus
Isc provides a rigorous measure of net electrogenic ion transport across the
epithelium under these conditions.
Under I-clamp, voltage is measured and the current is clamped at 0 µA/cm2,
conditions that are referred to as open-circuit. Changes in transepithelial ion
transport under I-clamp are reflected by changes in transepithelial voltage. For
comparison with previous studies, the equivalent short-circuit current (Ieq) was also
calculated from the transepithelial voltage and resistance.
Ieq measures net electrogenic ion transport, but in the presence of a transepithelial
electrochemical gradient and therefore is not a measure of active transport.
Nevertheless, I-clamp has some advantages over V-clamp. I-clamp is more relevant to
physiological transport since it does not perturb the transepithelial and
transmembrane electrochemical gradients, which are important features of polarized
epithelia. Second, since no current is passed through the epithelium from an external
source, I-clamp does cause membrane dielectric breakdown or other damage to the
monolayer.
I used open-circuit conditions and measured V and R so that Ieq could be calculated.
This is appropriate for studying bicarbonate transport since rigorous short-circuit

139
conditions with symmetrical solutions are not possible under pH-stat conditions
anyway; CO2 and bicarbonate are present only on the basolateral side while the
apical solution must remain unbuffered to detect the secretion of base (see below).
Also, I found that the titration workstation worked best under open-circuit conditions,
presumably because no current flowed through reference electrode during pH
measurements under I-clamp.
5.1.2 pH-stat conditions
To directly measure HCO3- secretion, I used the pH-stat technique. A major
advantage of this method is that it provides an accurate measure of net bicarbonate
flux. One limitation is the possibility that the HCO3- gradient from the basolateral to
the apical side may lead to an artificially elevated rate of HCO3- secretion and
perhaps other artifacts caused by an electrochemical gradient for HCO3- which is
essentially infinite. For example the large outward gradient for HCO3- at the apical
membrane under these conditions might favor HCO3- apical exit via CFTR channels
rather than transporters or exchangers, although I obtained no evidence for this.
5.1.3 The Calu-3 cell line
In all the studies described in this thesis, I used the Calu-3 cell line as a model for
airway epithelial cells. Calu-3 is an adenocarcinoma cell line which was developed by
Jorgen Fogh (Fogh, Fogh et al. 1977). When Calu-3 cells were cultured on porous
supports at the air-liquid interface they formed tight junctions, had transepithelial

140
resistance >100 Ω·cm2, and generated short-circuit current (Isc) that was stimulated
by isoproterenol (Shen, Finkbeiner et al. 1994). Isc was also increased by
calcium-mediated secretagogues (bradykinin, histamine methacholine, Ca2+
ionophores). Those responses were abolished when the basolateral membrane was
permeabilized using the pore-forming antifungal agent nystatin, indicating that the
monolayers were polarized and that calcium mobilizing agonists acted primarily at
the basolateral membrane, presumably by activating K+ channels (Shen, Finkbeiner et
al. 1994).
Calu-3 cells express high levels of CFTR (Haws, Finkbeiner et al. 1994) and also
produce the serous cell markers lysozyme (Duszyk 2001) and lactoferrin (Dubin,
Robinson et al. 2004), however they lack the dense granules which characterize
serous cells in vivo. Moreover, secretagogues that normally stimulate lysozyme and
lactoferrin release by native glands (e.g methacholine, adrenaline, neutrophil
elastase, lipopolysaccharide) are not effective on Calu-3 monolayers (Dubin,
Robinson et al. 2004). Like most epithelial cell lines, Calu-3 cells also secrete some
mucus despite being a serous cell model. About 25% of the cells in a typical air-liquid
interface culture develop translucent mucin granules of 1–2 μm diameter (Shen,
Finkbeiner et al. 1994, Kreda, Okada et al. 2007) which contain the mucins MUC5AC
and MUC5B like those in submucosal glands (Kreda, Okada et al. 2007). Secretion of
antimicrobial factors and mucins suggests a mixed serous-mucous phenotype,
however this is not necessarily abnormal as serous cells in submucosal glands also

141
produce some mucin. Indeed, about one-third of the serous tubules in native glands
also express significant amounts of MUC7, which is colocalized with lysozyme
(Sharma, Dudus et al. 1998).
Another feature of Calu-3 cells which sometimes causes concern is their abnormal
karyotype. According to American Type Culture Collection (Manassas, VA, USA),
Calu-3 cells lack normal chromosomes 1, 13, 15 and 17, and are triploid for some
other (unidentified) chromosomes. Genes that are relevant for anion transport and
host defense would normally be situated on those missing chromosomes. For
example PDZD1/CAP70/NHERF3 is on chromosome 1 at 1q21, and the NADPH
oxidases DUOX1 and DUOX2 are on chromosome 15 at 15q21. However, it is clear
that at least some of the genes have been translocated to other chromosomes and
are still expressed. The loss of chromosome 17 should eliminate sodium-hydrogen
exchanger regulator factor 1 (NHERF1) at 17q25.1, yet NHERF1 is readily
immunoprecipitated from Calu-3 cells (Liedtke, Raghuram et al. 2004) and its role in
Calu-3 cells has been demonstrated using RNA interference. Thus although Calu-3
cells have chromosomal abnormalities, their consequences are less severe than
expected. In summary, the Calu-3 cell line is an imperfect, but still useful model for
airway epithelia.
5.1.4 Modifications to existing models
This section considers how existing models for Calu-3 transport might be revised to

142
accommodate the results from this study, which were obtained under conditions that
have not been used previously to study Calu-3 (i.e. open circuit, pH-stat).
The [Cl−] of fluid secreted by Calu-3 cells was ∼120 mmol·l−1, much higher than the
[HCO3−]. This implies that Calu-3 cells secrete mostly Cl− rather than HCO3
− during
forskolin stimulation despite the fact that active Cl− transport was not detected in a
previous study (Devor, Singh et al. 1999). Another interesting finding in this thesis is
that most of the Cl− transport was independent of NKCC1. This conclusion was based
on both anion selectivity and pharmacological data. Robust fluid secretion was
observed for 24 h when the basolateral side was bathed with nominally Cl−-free
solution containing NO3−, which does not bind to the highly selective anion site on
NKCC transporters and therefore is not transported (Kinne, Kinne-Saffran et al. 1986).
Further evidence for NKCC1-independent Cl− entry is the relative insensitivity of fluid
secretion to bumetanide. These results suggest that other basolateral Cl− entry
pathways such as anion exchangers are more important. The fact that NO3− is carried
by many anion exchangers including AE2 (Humphreys, Jiang et al. 1994) led to the
investigation of these exchangers in terms of their contribution to Cl- transport.
Basolateral anion exchange was demonstrated by permeabilizing the apical
membrane, imposing an apical-to-basolateral HCO3− gradient, and examining the
effect of anions in the basolateral solution (i.e. on the “trans” side) on the HCO3− flux
as measured by basolateral pH-stat. Our data strongly suggest that some HCO3− that

143
enters the cell via NBCe1 returns to the basolateral side by exchanging for Cl− (or
NO3−). The anion exchangers that mediate this basolateral Cl− loading were not
identified at the molecular level in the present study; however, AE2 is the most likely
candidate since it is expressed at the basolateral membrane of Calu-3 cells (Loffing,
Moyer et al. 2000). Recent studies of an AE2-deficient Calu-3 cell line generated in
this lab indicate that AE2 mediates essentially all Cl−/HCO3− exchange at the
basolateral membrane and plays an important role in fluid secretion (Huang, Shan et
al. 2012).
A recent study suggested that HCO3- is secreted apically via pendrin but not CFTR
(Garnett, Hickman et al. 2011). The results in this thesis provide no evidence for
significant bicarbonate transport via pendrin when functional CFTR is present.
However, as discussed above, our results do not preclude a role of pendrin under
other experimental conditions since the large bicarbonate gradient used here favors
bicarbonate secretion and might drive an abnormally large flux through CFTR,
masking the role of pendrin.
5.2 The determinants of fluid secretion
The [Cl-] of secretions was ∼120 mmol·l−1, or about 4-fold higher than the [HCO3−].
This implies that most fluid secretion is driven by transepithelial transport of Cl−
rather than HCO3−. However, HCO3
- plays a critical role in secretion as it is needed by
the basolateral anion exchanger for Cl- loading. The pharmacology results suggest

144
this is the predominant mechanism of Cl- uptake, with NKCC supplying a smaller
fraction of the Cl-.
Our further studies on fluid secretion indicate that osmolality provides the driving
force for fluid secretion as expected and that under physiological conditions, Cl-
secretion followed by passive cation transport establishes the osmotic gradient. Net
water flux can also be achieved artificially by adjusting osmolality with mannitol,
which supports the development of inhaled mannitol as a treatment for increasing
fluid secretion in CF airways.
5.3 The role of carbonic anhydrases
I found three carbonic anhydrase isoforms (II, IX, XII) in Calu-3 cells at the mRNA level,
and the expression of CAII and CAIX was further demonstrated at the protein level.
Expression of these isoforms is not surprising since CAII is ubiquitous while the other
two are cancer-associated, and Calu-3 is an adenocarcinoma cell line.
The fact that the broad inhibitor of carbonic anhydrases, acetazolamide, had a strong
inhibitory effect on anion transport by Calu-3 suggests carbonic anhydrase plays an
important role in this process. The lack of effect of
4-(2-aminoethyl)-benzenesulfonamide, a specific inhibitor of CAIX, suggests that this
isoform is not involved in anion transport.

145
Treating cells with acetazolamide had a strong inhibitory effect whether it was added
before or during forskolin stimulation, however the possibility of a metabolon with
NBC was not explored systematically. If a metabolon does exist in Calu-3 cells, CAII
would be the most likely candidate carbonic anhydrase.
Though CAXII also contributes to the anion secretion in Calu-3 cells, its effects are
probably due to the upregulation of CAXII in adenocarcinomas. In normal tissue
without such an upregulation, CAII may be the major player.
5.4 Conclusions and future direction
In my thesis, I successfully characterized transepithelial anion transport in airway
epithelial cells using the Calu-3 cell line as a model. I propose a revised model for
anion and fluid secretion by Calu-3 which is likely to be relevant to submucosal
glands and distinct from the mechanism present in the surface epithelium. This
model may explain the results in an early study by others showing that exposure to a
high concentration of bumetanide on the basolateral side (100 µM) has almost no
effect on fluid secretion by cat trachea during phenylephrine stimulation of the
submucosal glands (Corrales, Nadel et al. 1984). That convincing study is rarely cited
because the results could not be reconciled with the paradigm that NKCC drives fluid
transport in secretory epithelia.
There are still important questions which need to be addressed: Does pendrin

146
transport HCO3- under physiological and/or pathological conditions? Does the model
I have proposed for Calu-3 cells also apply to primary submucosal gland cultures?
What is the role of HCO3- in airway secretions beside supporting Cl- loading by
basolateral anion exchange? Answering these questions will provide a better
understanding of CF and may lead to better treatment of the disease.

147
References Acevedo, M. and L. W. Steele (1993). "Na(+)-H+ exchanger in isolated epithelial tracheal cells from sheep. Involvement in tracheal proton secretion." Exp Physiol 78(3): 383-394.
Al-Bazzaz, F. J., N. Hafez, S. Tyagi, C. A. Gailey, M. Toofanfard, W. A. Alrefai, T. M. Nazir, K. Ramaswamy and P. K. Dudeja (2001). "Detection of Cl--HCO3- and Na+-H+ exchangers in human airways epithelium." JOP 2(4 Suppl): 285-290.
Alper, S. L., M. N. Chernova and A. K. Stewart (2002). "How pH regulates a pH regulator: a regulatory hot spot in the N-terminal cytoplasmic domain of the AE2 anion exchanger." Cell Biochem Biophys 36(2-3): 123-136.
Alpern, R. J. (1985). "Mechanism of basolateral membrane H+/OH-/HCO-3 transport in the rat proximal convoluted tubule. A sodium-coupled electrogenic process." J Gen Physiol 86(5): 613-636.
Aravind, L. and E. V. Koonin (2000). "The STAS domain - a link between anion transporters and antisigma-factor antagonists." Curr Biol 10(2): R53-55.
Awayda, M. S., M. J. Boudreaux, R. L. Reger and L. L. Hamm (2000). "Regulation of the epithelial Na(+) channel by extracellular acidification." Am J Physiol Cell Physiol 279(6): C1896-1905.
Baird, T. T., Jr., A. Waheed, T. Okuyama, W. S. Sly and C. A. Fierke (1997). "Catalysis and inhibition of human carbonic anhydrase IV." Biochemistry 36(9): 2669-2678.
Ballard, S. T. and S. K. Inglis (2004). "Liquid secretion properties of airway submucosal glands." J Physiol 556(Pt 1): 1-10.
Ballard, S. T., L. Trout, Z. Bebok, E. J. Sorscher and A. Crews (1999). "CFTR involvement in chloride, bicarbonate, and liquid secretion by airway submucosal glands." Am J Physiol 277(4 Pt 1): L694-699.
Ballard, S. T., L. Trout, J. Garrison and S. K. Inglis (2006). "Ionic mechanism of forskolin-induced liquid secretion by porcine bronchi." Am J Physiol Lung Cell Mol Physiol 290(1): L97-104.
Bardou, O., N. T. Trinh and E. Brochiero (2009). "Molecular diversity and function of K+ channels in airway and alveolar epithelial cells." Am J Physiol Lung Cell Mol Physiol 296(2): L145-155.

148
Barker, P. M., M. S. Nguyen, J. T. Gatzy, B. Grubb, H. Norman, E. Hummler, B. Rossier, R. C. Boucher and B. Koller (1998). "Role of gammaENaC subunit in lung liquid clearance and electrolyte balance in newborn mice. Insights into perinatal adaptation and pseudohypoaldosteronism." J Clin Invest 102(8): 1634-1640.
Basbaum, C. B., B. Jany and W. E. Finkbeiner (1990). "The serous cell." Annu Rev Physiol 52: 97-113.
Bernardo, A. A., C. M. Bernardo, D. J. Espiritu and J. A. Arruda (2006). "The sodium bicarbonate cotransporter: structure, function, and regulation." Semin Nephrol 26(5): 352-360.
Bertrand, C. A., R. Zhang, J. M. Pilewski and R. A. Frizzell (2009). "SLC26A9 is a constitutively active, CFTR-regulated anion conductance in human bronchial epithelia." J Gen Physiol 133(4): 421-438.
Boers, J. E., A. W. Ambergen and F. B. Thunnissen (1998). "Number and proliferation of basal and parabasal cells in normal human airway epithelium." Am J Respir Crit Care Med 157(6 Pt 1): 2000-2006.
Boitano, S., Z. Safdar, D. G. Welsh, J. Bhattacharya and M. Koval (2004). "Cell-cell interactions in regulating lung function." Am J Physiol Lung Cell Mol Physiol 287(3): L455-459.
Bonar, P. T. and J. R. Casey (2008). "Plasma membrane Cl(-)/HCO(3)(-) exchangers: structure, mechanism and physiology." Channels (Austin) 2(5): 337-345.
Boucher, R. C. (2003). "Regulation of airway surface liquid volume by human airway epithelia." Pflugers Arch 445(4): 495-498.
Boucher, R. C. (2007). "Airway surface dehydration in cystic fibrosis: pathogenesis and therapy." Annu Rev Med 58: 157-170.
Boucher, R. C. (2007). "Evidence for airway surface dehydration as the initiating event in CF airway disease." J Intern Med 261(1): 5-16.
Bowman, E. J., L. A. Graham, T. H. Stevens and B. J. Bowman (2004). "The bafilomycin/concanamycin binding site in subunit c of the V-ATPases from Neurospora crassa and Saccharomyces cerevisiae." J Biol Chem 279(32): 33131-33138.
Burch, L. H., C. R. Talbot, M. R. Knowles, C. M. Canessa, B. C. Rossier and R. C. Boucher (1995). "Relative expression of the human epithelial Na+ channel subunits in

149
normal and cystic fibrosis airways." Am J Physiol 269(2 Pt 1): C511-518.
Caputo, A., E. Caci, L. Ferrera, N. Pedemonte, C. Barsanti, E. Sondo, U. Pfeffer, R. Ravazzolo, O. Zegarra-Moran and L. J. Galietta (2008). "TMEM16A, a membrane protein associated with calcium-dependent chloride channel activity." Science 322(5901): 590-594.
Case, R. M., M. Hunter, I. Novak and J. A. Young (1984). "The anionic basis of fluid secretion by the rabbit mandibular salivary gland." J Physiol 349: 619-630.
Chambers, L. A., B. M. Rollins and R. Tarran (2007). "Liquid movement across the surface epithelium of large airways." Respir Physiol Neurobiol 159(3): 256-270.
Chen, Y., M. J. Cann, T. N. Litvin, V. Iourgenko, M. L. Sinclair, L. R. Levin and J. Buck (2000). "Soluble adenylyl cyclase as an evolutionarily conserved bicarbonate sensor." Science 289(5479): 625-628.
Cherny, V. V. and T. E. DeCoursey (1999). "pH-dependent inhibition of voltage-gated H(+) currents in rat alveolar epithelial cells by Zn(2+) and other divalent cations." J Gen Physiol 114(6): 819-838.
Cherny, V. V., V. S. Markin and T. E. DeCoursey (1995). "The voltage-activated hydrogen ion conductance in rat alveolar epithelial cells is determined by the pH gradient." J Gen Physiol 105(6): 861-896.
Choi, J. Y., N. S. Joo, M. E. Krouse, J. V. Wu, R. C. Robbins, J. P. Ianowski, J. W. Hanrahan and J. J. Wine (2007). "Synergistic airway gland mucus secretion in response to vasoactive intestinal peptide and carbachol is lost in cystic fibrosis." J Clin Invest 117(10): 3118-3127.
Coakley, R. D., B. R. Grubb, A. M. Paradiso, J. T. Gatzy, L. G. Johnson, S. M. Kreda, W. K. O'Neal and R. C. Boucher (2003). "Abnormal surface liquid pH regulation by cultured cystic fibrosis bronchial epithelium." Proc Natl Acad Sci U S A 100(26): 16083-16088.
Cobb, B. R., F. Ruiz, C. M. King, J. Fortenberry, H. Greer, T. Kovacs, E. J. Sorscher and J. P. Clancy (2002). "A(2) adenosine receptors regulate CFTR through PKA and PLA(2)." Am J Physiol Lung Cell Mol Physiol 282(1): L12-25.
Communi, D., P. Paindavoine, G. A. Place, M. Parmentier and J. M. Boeynaems (1999). "Expression of P2Y receptors in cell lines derived from the human lung." Br J Pharmacol 127(2): 562-568.
Corrales, R. J., J. A. Nadel and J. H. Widdicombe (1984). "Source of the fluid

150
component of secretions from tracheal submucosal glands in cats." J Appl Physiol Respir Environ Exerc Physiol 56(4): 1076-1082.
Counillon, L., W. Scholz, H. J. Lang and J. Pouyssegur (1993). "Pharmacological characterization of stably transfected Na+/H+ antiporter isoforms using amiloride analogs and a new inhibitor exhibiting anti-ischemic properties." Mol Pharmacol 44(5): 1041-1045.
Crump, R. G., G. R. Askew, S. E. Wert, J. B. Lingrel and C. H. Joiner (1995). "In situ localization of sodium-potassium ATPase mRNA in developing mouse lung epithelium." Am J Physiol 269(3 Pt 1): L299-308.
Cuthbert, A. W. and L. J. MacVinish (2003). "Mechanisms of anion secretion in Calu-3 human airway epithelial cells by 7,8-benzoquinoline." Br J Pharmacol 140(1): 81-90.
Cuthbert, A. W., C. T. Supuran and L. J. MacVinish (2003). "Bicarbonate-dependent chloride secretion in Calu-3 epithelia in response to 7,8-benzoquinoline." J Physiol 551(Pt 1): 79-92.
DeCoursey, T. E. (1991). "Hydrogen ion currents in rat alveolar epithelial cells." Biophys J 60(5): 1243-1253.
DeCoursey, T. E. and V. V. Cherny (1995). "Voltage-activated proton currents in membrane patches of rat alveolar epithelial cells." J Physiol 489 ( Pt 2): 299-307.
Devor, D. C., R. J. Bridges and J. M. Pilewski (2000). "Pharmacological modulation of ion transport across wild-type and DeltaF508 CFTR-expressing human bronchial epithelia." Am J Physiol Cell Physiol 279(2): C461-479.
Devor, D. C., A. K. Singh, L. C. Lambert, A. DeLuca, R. A. Frizzell and R. J. Bridges (1999). "Bicarbonate and chloride secretion in Calu-3 human airway epithelial cells." J Gen Physiol 113(5): 743-760.
Dobbs, L. G. and M. D. Johnson (2007). "Alveolar epithelial transport in the adult lung." Respir Physiol Neurobiol 159(3): 283-300.
Dossena, S., S. Rodighiero, V. Vezzoli, C. Nofziger, E. Salvioni, M. Boccazzi, E. Grabmayer, G. Botta, G. Meyer, L. Fugazzola, P. Beck-Peccoz and M. Paulmichl (2009). "Functional characterization of wild-type and mutated pendrin (SLC26A4), the anion transporter involved in Pendred syndrome." J Mol Endocrinol 43(3): 93-103.
Dossena, S., V. Vezzoli, N. Cerutti, C. Bazzini, M. Tosco, C. Sironi, S. Rodighiero, G. Meyer, U. Fascio, J. Furst, M. Ritter, L. Fugazzola, L. Persani, P. Zorowka, C. Storelli, P.

151
Beck-Peccoz, G. Botta and M. Paulmichl (2006). "Functional characterization of wild-type and a mutated form of SLC26A4 identified in a patient with Pendred syndrome." Cell Physiol Biochem 17(5-6): 245-256.
Dubin, R. F., S. K. Robinson and J. H. Widdicombe (2004). "Secretion of lactoferrin and lysozyme by cultures of human airway epithelium." Am J Physiol Lung Cell Mol Physiol 286(4): L750-755.
Dudeja, P. K., N. Hafez, S. Tyagi, C. A. Gailey, M. Toofanfard, W. A. Alrefai, T. M. Nazir, K. Ramaswamy and F. J. Al-Bazzaz (1999). "Expression of the Na+/H+ and Cl-/HCO-3 exchanger isoforms in proximal and distal human airways." Am J Physiol 276(6 Pt 1): L971-978.
Duszyk, M. (2001). "CFTR and lysozyme secretion in human airway epithelial cells." Pflugers Arch 443 Suppl 1: S45-49.
Engelhardt, J. F., J. R. Yankaskas, S. A. Ernst, Y. Yang, C. R. Marino, R. C. Boucher, J. A. Cohn and J. M. Wilson (1992). "Submucosal glands are the predominant site of CFTR expression in the human bronchus." Nat Genet 2(3): 240-248.
Ernst, S. A., J. R. Palacios, 2nd and G. J. Siegel (1986). "Immunocytochemical localization of Na+,K+-ATPase catalytic polypeptide in mouse choroid plexus." J Histochem Cytochem 34(2): 189-195.
Esbaugh, A. J. and B. L. Tufts (2006). "The structure and function of carbonic anhydrase isozymes in the respiratory system of vertebrates." Respir Physiol Neurobiol 154(1-2): 185-198.
Evans, C. M. and J. S. Koo (2009). "Airway mucus: the good, the bad, the sticky." Pharmacol Ther 121(3): 332-348.
Evans, C. M., O. W. Williams, M. J. Tuvim, R. Nigam, G. P. Mixides, M. R. Blackburn, F. J. DeMayo, A. R. Burns, C. Smith, S. D. Reynolds, B. R. Stripp and B. F. Dickey (2004). "Mucin is produced by clara cells in the proximal airways of antigen-challenged mice." Am J Respir Cell Mol Biol 31(4): 382-394.
Evans, M. J., R. A. Cox, S. G. Shami and C. G. Plopper (1990). "Junctional adhesion mechanisms in airway basal cells." Am J Respir Cell Mol Biol 3(4): 341-347.
Evans, M. J., R. A. Cox, S. G. Shami, B. Wilson and C. G. Plopper (1989). "The role of basal cells in attachment of columnar cells to the basal lamina of the trachea." Am J Respir Cell Mol Biol 1(6): 463-469.

152
Evans, M. J. and C. G. Plopper (1988). "The role of basal cells in adhesion of columnar epithelium to airway basement membrane." Am Rev Respir Dis 138(2): 481-483.
Farman, N., C. R. Talbot, R. Boucher, M. Fay, C. Canessa, B. Rossier and J. P. Bonvalet (1997). "Noncoordinated expression of alpha-, beta-, and gamma-subunit mRNAs of epithelial Na+ channel along rat respiratory tract." Am J Physiol 272(1 Pt 1): C131-141.
Fischer, H. and J. H. Widdicombe (2006). "Mechanisms of acid and base secretion by the airway epithelium." J Membr Biol 211(3): 139-150.
Fischer, H., J. H. Widdicombe and B. Illek (2002). "Acid secretion and proton conductance in human airway epithelium." Am J Physiol Cell Physiol 282(4): C736-743.
Fliegel, L. (2005). "The Na+/H+ exchanger isoform 1." Int J Biochem Cell Biol 37(1): 33-37.
Fogh, J., J. M. Fogh and T. Orfeo (1977). "One hundred and twenty-seven cultured human tumor cell lines producing tumors in nude mice." J Natl Cancer Inst 59(1): 221-226.
Gabriel, S. E., M. Makhlina, E. Martsen, E. J. Thomas, M. I. Lethem and R. C. Boucher (2000). "Permeabilization via the P2X7 purinoreceptor reveals the presence of a Ca2+-activated Cl- conductance in the apical membrane of murine tracheal epithelial cells." J Biol Chem 275(45): 35028-35033.
Gamba, G., A. Miyanoshita, M. Lombardi, J. Lytton, W. S. Lee, M. A. Hediger and S. C. Hebert (1994). "Molecular cloning, primary structure, and characterization of two members of the mammalian electroneutral sodium-(potassium)-chloride cotransporter family expressed in kidney." J Biol Chem 269(26): 17713-17722.
Ganz, T. (2002). "Antimicrobial polypeptides in host defense of the respiratory tract." J Clin Invest 109(6): 693-697.
Garcia-Caballero, A., J. E. Rasmussen, E. Gaillard, M. J. Watson, J. C. Olsen, S. H. Donaldson, M. J. Stutts and R. Tarran (2009). "SPLUNC1 regulates airway surface liquid volume by protecting ENaC from proteolytic cleavage." Proc Natl Acad Sci U S A 106(27): 11412-11417.
Garnett, J. P., E. Hickman, R. Burrows, P. Hegyi, L. Tiszlavicz, A. W. Cuthbert, P. Fong and M. A. Gray (2011). "Novel role for pendrin in orchestrating bicarbonate secretion in cystic fibrosis transmembrane conductance regulator (CFTR)-expressing airway

153
serous cells." J Biol Chem 286(47): 41069-41082.
Gibson, L. E. and R. E. Cooke (1959). "A test for concentration of electrolytes in sweat in cystic fibrosis of the pancreas utilizing pilocarpine by iontophoresis." Pediatrics 23(3): 545-549.
Grassl, S. M. and P. S. Aronson (1986). "Na+/Hco3-Co-Transport in Basolateral Membrane-Vesicles Isolated from Rabbit Renal-Cortex." Journal of Biological Chemistry 261(19): 8778-8783.
Gray, M. A., C. E. Pollard, A. Harris, L. Coleman, J. R. Greenwell and B. E. Argent (1990). "Anion selectivity and block of the small-conductance chloride channel on pancreatic duct cells." Am J Physiol 259(5 Pt 1): C752-761.
Gross, E. and I. Kurtz (2002). "Structural determinants and significance of regulation of electrogenic Na(+)-HCO(3)(-) cotransporter stoichiometry." Am J Physiol Renal Physiol 283(5): F876-887.
Gundersen, D., J. Orlowski and E. Rodriguez-Boulan (1991). "Apical polarity of Na,K-ATPase in retinal pigment epithelium is linked to a reversal of the ankyrin-fodrin submembrane cytoskeleton." J Cell Biol 112(5): 863-872.
Gut, M. O., S. Parkkila, Z. Vernerova, E. Rohde, J. Zavada, M. Hocker, J. Pastorek, T. Karttunen, A. Gibadulinova, Z. Zavadova, K. P. Knobeloch, B. Wiedenmann, J. Svoboda, I. Horak and S. Pastorekova (2002). "Gastric hyperplasia in mice with targeted disruption of the carbonic anhydrase gene Car9." Gastroenterology 123(6): 1889-1903.
Haas, M. and B. Forbush, 3rd (2000). "The Na-K-Cl cotransporter of secretory epithelia." Annu Rev Physiol 62: 515-534.
Harhaj, N. S. and D. A. Antonetti (2004). "Regulation of tight junctions and loss of barrier function in pathophysiology." Int J Biochem Cell Biol 36(7): 1206-1237.
Haws, C., W. E. Finkbeiner, J. H. Widdicombe and J. J. Wine (1994). "CFTR in Calu-3 human airway cells: channel properties and role in cAMP-activated Cl- conductance." Am J Physiol 266(5 Pt 1): L502-512.
Hollenhorst, M. I., K. Richter and M. Fronius (2011). "Ion transport by pulmonary epithelia." J Biomed Biotechnol 2011: 174306.
Hong, K. U., S. D. Reynolds, A. Giangreco, C. M. Hurley and B. R. Stripp (2001). "Clara cell secretory protein-expressing cells of the airway neuroepithelial body

154
microenvironment include a label-retaining subset and are critical for epithelial renewal after progenitor cell depletion." Am J Respir Cell Mol Biol 24(6): 671-681.
Hong, K. U., S. D. Reynolds, S. Watkins, E. Fuchs and B. R. Stripp (2004). "Basal cells are a multipotent progenitor capable of renewing the bronchial epithelium." Am J Pathol 164(2): 577-588.
Hong, K. U., S. D. Reynolds, S. Watkins, E. Fuchs and B. R. Stripp (2004). "In vivo differentiation potential of tracheal basal cells: evidence for multipotent and unipotent subpopulations." Am J Physiol Lung Cell Mol Physiol 286(4): L643-649.
Hovenberg, H. W., J. R. Davies and I. Carlstedt (1996). "Different mucins are produced by the surface epithelium and the submucosa in human trachea: identification of MUC5AC as a major mucin from the goblet cells." Biochem J 318 ( Pt 1): 319-324.
Huang, J., J. Shan, D. Kim, J. Liao, A. Evagelidis, S. L. Alper and J. W. Hanrahan (2012). "Basolateral chloride loading by the anion exchanger type 2: role in fluid secretion by the human airway epithelial cell line Calu-3." J Physiol 590(Pt 21): 5299-5316.
Humphreys, B. D., L. Jiang, M. N. Chernova and S. L. Alper (1994). "Functional characterization and regulation by pH of murine AE2 anion exchanger expressed in Xenopus oocytes." Am J Physiol 267(5 Pt 1): C1295-1307.
Huss, M., G. Ingenhorst, S. Konig, M. Gassel, S. Drose, A. Zeeck, K. Altendorf and H. Wieczorek (2002). "Concanamycin A, the specific inhibitor of V-ATPases, binds to the V(o) subunit c." J Biol Chem 277(43): 40544-40548.
Hwang, T. C. and D. N. Sheppard (2009). "Gating of the CFTR Cl- channel by ATP-driven nucleotide-binding domain dimerisation." J Physiol 587(Pt 10): 2151-2161.
Illek, B., J. R. Yankaskas and T. E. Machen (1997). "cAMP and genistein stimulate HCO3- conductance through CFTR in human airway epithelia." Am J Physiol 272(4 Pt 1): L752-761.
Inglis, S. K., S. M. Wilson and R. E. Olver (2003). "Secretion of acid and base equivalents by intact distal airways." Am J Physiol Lung Cell Mol Physiol 284(5): L855-862.
Iovannisci, D., B. Illek and H. Fischer (2010). "Function of the HVCN1 proton channel in airway epithelia and a naturally occurring mutation, M91T." J Gen Physiol 136(1): 35-46.

155
Irokawa, T., M. E. Krouse, N. S. Joo, J. V. Wu and J. J. Wine (2004). "A "virtual gland" method for quantifying epithelial fluid secretion." Am J Physiol Lung Cell Mol Physiol 287(4): L784-793.
Ishiguro, H., M. C. Steward, S. Naruse, S. B. Ko, H. Goto, R. M. Case, T. Kondo and A. Yamamoto (2009). "CFTR functions as a bicarbonate channel in pancreatic duct cells." J Gen Physiol 133(3): 315-326.
Jayaraman, S., N. S. Joo, B. Reitz, J. J. Wine and A. S. Verkman (2001). "Submucosal gland secretions in airways from cystic fibrosis patients have normal [Na(+)] and pH but elevated viscosity." Proc Natl Acad Sci U S A 98(14): 8119-8123.
Jiang, L., M. N. Chernova and S. L. Alper (1997). "Secondary regulatory volume increase conferred on Xenopus oocytes by expression of AE2 anion exchanger." Am J Physiol 272(1 Pt 1): C191-202.
Joo, N. S., T. Irokawa, J. V. Wu, R. C. Robbins, R. I. Whyte and J. J. Wine (2002). "Absent secretion to vasoactive intestinal peptide in cystic fibrosis airway glands." J Biol Chem 277(52): 50710-50715.
Joo, N. S., Y. Saenz, M. E. Krouse and J. J. Wine (2002). "Mucus secretion from single submucosal glands of pig. Stimulation by carbachol and vasoactive intestinal peptide." J Biol Chem 277(31): 28167-28175.
Kaplan, M. R., M. D. Plotkin, W. S. Lee, Z. C. Xu, J. Lytton and S. C. Hebert (1996). "Apical localization of the Na-K-Cl cotransporter, rBSC1, on rat thick ascending limbs." Kidney Int 49(1): 40-47.
Kartner, N. and J. R. Riordan (1998). "Characterization of polyclonal and monoclonal antibodies to cystic fibrosis transmembrane conductance regulator." Methods Enzymol 292: 629-652.
Kashlan, O. B. and T. R. Kleyman (2011). "ENaC structure and function in the wake of a resolved structure of a family member." Am J Physiol Renal Physiol 301(4): F684-696.
Kellenberger, S. and L. Schild (2002). "Epithelial sodium channel/degenerin family of ion channels: a variety of functions for a shared structure." Physiol Rev 82(3): 735-767.
Kelly, M., S. Trudel, F. Brouillard, F. Bouillaud, J. Colas, T. Nguyen-Khoa, M. Ollero, A. Edelman and J. Fritsch (2010). "Cystic fibrosis transmembrane regulator inhibitors CFTR(inh)-172 and GlyH-101 target mitochondrial functions, independently of

156
chloride channel inhibition." J Pharmacol Exp Ther 333(1): 60-69.
Kerem, B., J. M. Rommens, J. A. Buchanan, D. Markiewicz, T. K. Cox, A. Chakravarti, M. Buchwald and L. C. Tsui (1989). "Identification of the cystic fibrosis gene: genetic analysis." Science 245(4922): 1073-1080.
Khalifah, R. G. (1971). "The carbon dioxide hydration activity of carbonic anhydrase. I. Stop-flow kinetic studies on the native human isoenzymes B and C." J Biol Chem 246(8): 2561-2573.
Kim, D. and M. C. Steward (2009). "The role of CFTR in bicarbonate secretion by pancreatic duct and airway epithelia." J Med Invest 56 Suppl: 336-342.
Kinne, R., E. Kinne-Saffran, B. Scholermann and H. Schutz (1986). "The anion specificity of the sodium-potassium-chloride cotransporter in rabbit kidney outer medulla: studies on medullary plasma membranes." Pflugers Arch 407 Suppl 2: S168-173.
Klyce, S. D. and R. K. Wong (1977). "Site and mode of adrenaline action on chloride transport across the rabbit corneal epithelium." J Physiol 266(3): 777-799.
Knight, D. A. and S. T. Holgate (2003). "The airway epithelium: structural and functional properties in health and disease." Respirology 8(4): 432-446.
Knowles, M., J. Gatzy and R. Boucher (1981). "Increased bioelectric potential difference across respiratory epithelia in cystic fibrosis." N Engl J Med 305(25): 1489-1495.
Ko, S. B., N. Shcheynikov, J. Y. Choi, X. Luo, K. Ishibashi, P. J. Thomas, J. Y. Kim, K. H. Kim, M. G. Lee, S. Naruse and S. Muallem (2002). "A molecular mechanism for aberrant CFTR-dependent HCO(3)(-) transport in cystic fibrosis." EMBO J 21(21): 5662-5672.
Ko, S. B., W. Zeng, M. R. Dorwart, X. Luo, K. H. Kim, L. Millen, H. Goto, S. Naruse, A. Soyombo, P. J. Thomas and S. Muallem (2004). "Gating of CFTR by the STAS domain of SLC26 transporters." Nat Cell Biol 6(4): 343-350.
Kopeikin, Z., Y. Sohma, M. Li and T. C. Hwang (2010). "On the mechanism of CFTR inhibition by a thiazolidinone derivative." J Gen Physiol 136(6): 659-671.
Koval, M. (2002). "Sharing signals: connecting lung epithelial cells with gap junction channels." Am J Physiol Lung Cell Mol Physiol 283(5): L875-893.

157
Kreda, S. M., S. F. Okada, C. A. van Heusden, W. O'Neal, S. Gabriel, L. Abdullah, C. W. Davis, R. C. Boucher and E. R. Lazarowski (2007). "Coordinated release of nucleotides and mucin from human airway epithelial Calu-3 cells." J Physiol 584(Pt 1): 245-259.
Krouse, M. E., J. F. Talbott, M. M. Lee, N. S. Joo and J. J. Wine (2004). "Acid and base secretion in the Calu-3 model of human serous cells." Am J Physiol Lung Cell Mol Physiol 287(6): L1274-1283.
Kyle, H., J. P. Ward and J. G. Widdicombe (1990). "Control of pH of airway surface liquid of the ferret trachea in vitro." J Appl Physiol (1985) 68(1): 135-140.
Lee, M. C., C. M. Penland, J. H. Widdicombe and J. J. Wine (1998). "Evidence that Calu-3 human airway cells secrete bicarbonate." Am J Physiol 274(3 Pt 1): L450-453.
Lee, M. G., J. Y. Choi, X. Luo, E. Strickland, P. J. Thomas and S. Muallem (1999). "Cystic fibrosis transmembrane conductance regulator regulates luminal Cl-/HCO3- exchange in mouse submandibular and pancreatic ducts." J Biol Chem 274(21): 14670-14677.
Lewis, S. A., N. K. Wills and D. C. Eaton (1978). "Basolateral membrane potential of a tight epithelium: ionic diffusion and electrogenic pumps." J Membr Biol 41(2): 117-148.
Li, M. S., R. G. Holstead, W. Wang and P. Linsdell (2011). "Regulation of CFTR chloride channel macroscopic conductance by extracellular bicarbonate." Am J Physiol Cell Physiol 300(1): C65-74.
Liedtke, C. M., V. Raghuram, C. C. Yun and X. Wang (2004). "Role of a PDZ1 domain of NHERF1 in the binding of airway epithelial RACK1 to NHERF1." Am J Physiol Cell Physiol 286(5): C1037-1044.
Linsdell, P., J. A. Tabcharani, J. M. Rommens, Y. X. Hou, X. B. Chang, L. C. Tsui, J. R. Riordan and J. W. Hanrahan (1997). "Permeability of wild-type and mutant cystic fibrosis transmembrane conductance regulator chloride channels to polyatomic anions." J Gen Physiol 110(4): 355-364.
Loffing, J., B. D. Moyer, D. Reynolds, B. E. Shmukler, S. L. Alper and B. A. Stanton (2000). "Functional and molecular characterization of an anion exchanger in airway serous epithelial cells." Am J Physiol Cell Physiol 279(4): C1016-1023.
Lubamba, B., B. Dhooghe, S. Noel and T. Leal (2012). "Cystic fibrosis: insight into CFTR pathophysiology and pharmacotherapy." Clin Biochem 45(15): 1132-1144.
Lumsden, A. B., A. McLean and D. Lamb (1984). "Goblet and Clara cells of human

158
distal airways: evidence for smoking induced changes in their numbers." Thorax 39(11): 844-849.
Luo, Y., K. McDonald and J. W. Hanrahan (2009). "Trafficking of immature DeltaF508-CFTR to the plasma membrane and its detection by biotinylation." Biochem J 419(1): 211-219, 212 p following 219.
Ma, T., J. R. Thiagarajah, H. Yang, N. D. Sonawane, C. Folli, L. J. Galietta and A. S. Verkman (2002). "Thiazolidinone CFTR inhibitor identified by high-throughput screening blocks cholera toxin-induced intestinal fluid secretion." J Clin Invest 110(11): 1651-1658.
MacVinish, L. J., G. Cope, A. Ropenga and A. W. Cuthbert (2007). "Chloride transporting capability of Calu-3 epithelia following persistent knockdown of the cystic fibrosis transmembrane conductance regulator, CFTR." Br J Pharmacol 150(8): 1055-1065.
Maggi, C. A., A. Giachetti, R. D. Dey and S. I. Said (1995). "Neuropeptides as regulators of airway function: vasoactive intestinal peptide and the tachykinins." Physiol Rev 75(2): 277-322.
Mall, M. A. (2008). "Role of cilia, mucus, and airway surface liquid in mucociliary dysfunction: lessons from mouse models." J Aerosol Med Pulm Drug Deliv 21(1): 13-24.
Masereel, B., L. Pochet and D. Laeckmann (2003). "An overview of inhibitors of Na(+)/H(+) exchanger." Eur J Med Chem 38(6): 547-554.
Matsui, H., B. R. Grubb, R. Tarran, S. H. Randell, J. T. Gatzy, C. W. Davis and R. C. Boucher (1998). "Evidence for periciliary liquid layer depletion, not abnormal ion composition, in the pathogenesis of cystic fibrosis airways disease." Cell 95(7): 1005-1015.
McDonald, F. J., B. Yang, R. F. Hrstka, H. A. Drummond, D. E. Tarr, P. B. McCray, Jr., J. B. Stokes, M. J. Welsh and R. A. Williamson (1999). "Disruption of the beta subunit of the epithelial Na+ channel in mice: hyperkalemia and neonatal death associated with a pseudohypoaldosteronism phenotype." Proc Natl Acad Sci U S A 96(4): 1727-1731.
McDonald, P. C., J. Y. Winum, C. T. Supuran and S. Dedhar (2012). "Recent developments in targeting carbonic anhydrase IX for cancer therapeutics." Oncotarget 3(1): 84-97.
McMurtrie, H. L., H. J. Cleary, B. V. Alvarez, F. B. Loiselle, D. Sterling, P. E. Morgan, D. E.

159
Johnson and J. R. Casey (2004). "The bicarbonate transport metabolon." J Enzyme Inhib Med Chem 19(3): 231-236.
Meyrick, B., J. M. Sturgess and L. Reid (1969). "A reconstruction of the duct system and secretory tubules of the human bronchial submucosal gland." Thorax 24(6): 729-736.
Morth, J. P., B. P. Pedersen, M. J. Buch-Pedersen, J. P. Andersen, B. Vilsen, M. G. Palmgren and P. Nissen (2011). "A structural overview of the plasma membrane Na+,K+-ATPase and H+-ATPase ion pumps." Nat Rev Mol Cell Biol 12(1): 60-70.
Mount, D. B. and M. F. Romero (2004). "The SLC26 gene family of multifunctional anion exchangers." Pflugers Arch 447(5): 710-721.
Muanprasat, C., N. D. Sonawane, D. Salinas, A. Taddei, L. J. Galietta and A. S. Verkman (2004). "Discovery of glycine hydrazide pore-occluding CFTR inhibitors: mechanism, structure-activity analysis, and in vivo efficacy." J Gen Physiol 124(2): 125-137.
Nakao, I., S. Kanaji, S. Ohta, H. Matsushita, K. Arima, N. Yuyama, M. Yamaya, K. Nakayama, H. Kubo, M. Watanabe, H. Sagara, K. Sugiyama, H. Tanaka, S. Toda, H. Hayashi, H. Inoue, T. Hoshino, A. Shiraki, M. Inoue, K. Suzuki, H. Aizawa, S. Okinami, H. Nagai, M. Hasegawa, T. Fukuda, E. D. Green and K. Izuhara (2008). "Identification of pendrin as a common mediator for mucus production in bronchial asthma and chronic obstructive pulmonary disease." J Immunol 180(9): 6262-6269.
Norimatsu, Y., A. Ivetac, C. Alexander, N. O'Donnell, L. Frye, M. S. Sansom and D. C. Dawson (2012). "Locating a plausible binding site for an open-channel blocker, GlyH-101, in the pore of the cystic fibrosis transmembrane conductance regulator." Mol Pharmacol 82(6): 1042-1055.
Novak, I. and J. A. Young (1986). "Two independent anion transport systems in rabbit mandibular salivary glands." Pflugers Arch 407(6): 649-656.
Palmer, M. L., S. Y. Lee, D. Carlson, S. Fahrenkrug and S. M. O'Grady (2006). "Stable knockdown of CFTR establishes a role for the channel in P2Y receptor-stimulated anion secretion." J Cell Physiol 206(3): 759-770.
Pan, P., M. Leppilampi, S. Pastorekova, J. Pastorek, A. Waheed, W. S. Sly and S. Parkkila (2006). "Carbonic anhydrase gene expression in CA II-deficient (Car2-/-) and CA IX-deficient (Car9-/-) mice." J Physiol 571(Pt 2): 319-327.
Paradiso, A. M., R. D. Coakley and R. C. Boucher (2003). "Polarized distribution of HCO3- transport in human normal and cystic fibrosis nasal epithelia." J Physiol 548(Pt

160
1): 203-218.
Park, H. W., J. H. Nam, J. Y. Kim, W. Namkung, J. S. Yoon, J. S. Lee, K. S. Kim, V. Venglovecz, M. A. Gray, K. H. Kim and M. G. Lee (2010). "Dynamic regulation of CFTR bicarbonate permeability by [Cl-]i and its role in pancreatic bicarbonate secretion." Gastroenterology 139(2): 620-631.
Paunescu, T. G., M. Ljubojevic, L. M. Russo, C. Winter, M. M. McLaughlin, C. A. Wagner, S. Breton and D. Brown (2010). "cAMP stimulates apical V-ATPase accumulation, microvillar elongation, and proton extrusion in kidney collecting duct A-intercalated cells." Am J Physiol Renal Physiol 298(3): F643-654.
Pedemonte, N., E. Caci, E. Sondo, A. Caputo, K. Rhoden, U. Pfeffer, M. Di Candia, R. Bandettini, R. Ravazzolo, O. Zegarra-Moran and L. J. Galietta (2007). "Thiocyanate transport in resting and IL-4-stimulated human bronchial epithelial cells: role of pendrin and anion channels." J Immunol 178(8): 5144-5153.
Piermarini, P. M., E. Y. Kim and W. F. Boron (2007). "Evidence against a direct interaction between intracellular carbonic anhydrase II and pure C-terminal domains of SLC4 bicarbonate transporters." J Biol Chem 282(2): 1409-1421.
Pirani, D., L. A. Evans, D. I. Cook and J. A. Young (1987). "Intracellular pH in the rat mandibular salivary gland: the role of Na-H and Cl-HCO3 antiports in secretion." Pflugers Arch 408(2): 178-184.
Poulsen, J. H., H. Fischer, B. Illek and T. E. Machen (1994). "Bicarbonate conductance and pH regulatory capability of cystic fibrosis transmembrane conductance regulator." Proc Natl Acad Sci U S A 91(12): 5340-5344.
Poulsen, J. H. and T. E. Machen (1996). "HCO3-dependent pHi regulation in tracheal epithelial cells." Pflugers Arch 432(3): 546-554.
Purkerson, J. M. and G. J. Schwartz (2007). "The role of carbonic anhydrases in renal physiology." Kidney Int 71(2): 103-115.
Quinton, P. M. (1983). "Chloride impermeability in cystic fibrosis." Nature 301(5899): 421-422.
Quinton, P. M. and J. Bijman (1983). "Higher bioelectric potentials due to decreased chloride absorption in the sweat glands of patients with cystic fibrosis." N Engl J Med 308(20): 1185-1189.
Rajasekaran, S. A. and A. K. Rajasekaran (2009). "Na,K-ATPase and epithelial tight

161
junctions." Front Biosci 14: 2130-2148.
Riordan, J. R. (2008). "CFTR function and prospects for therapy." Annu Rev Biochem 77: 701-726.
Riordan, J. R., J. M. Rommens, B. Kerem, N. Alon, R. Rozmahel, Z. Grzelczak, J. Zielenski, S. Lok, N. Plavsic, J. L. Chou and et al. (1989). "Identification of the cystic fibrosis gene: cloning and characterization of complementary DNA." Science 245(4922): 1066-1073.
Rogers, A. V., A. Dewar, B. Corrin and P. K. Jeffery (1993). "Identification of serous-like cells in the surface epithelium of human bronchioles." Eur Respir J 6(4): 498-504.
Rommens, J. M., M. C. Iannuzzi, B. Kerem, M. L. Drumm, G. Melmer, M. Dean, R. Rozmahel, J. L. Cole, D. Kennedy, N. Hidaka and et al. (1989). "Identification of the cystic fibrosis gene: chromosome walking and jumping." Science 245(4922): 1059-1065.
Russell, J. M. (2000). "Sodium-potassium-chloride cotransport." Physiol Rev 80(1): 211-276.
Schroeder, B. C., T. Cheng, Y. N. Jan and L. Y. Jan (2008). "Expression cloning of TMEM16A as a calcium-activated chloride channel subunit." Cell 134(6): 1019-1029.
Schwarzer, C., T. E. Machen, B. Illek and H. Fischer (2004). "NADPH oxidase-dependent acid production in airway epithelial cells." J Biol Chem 279(35): 36454-36461.
Scott, D. A. and L. P. Karniski (2000). "Human pendrin expressed in Xenopus laevis oocytes mediates chloride/formate exchange." Am J Physiol Cell Physiol 278(1): C207-211.
Scott, D. A., R. Wang, T. M. Kreman, V. C. Sheffield and L. P. Karniski (1999). "The Pendred syndrome gene encodes a chloride-iodide transport protein." Nat Genet 21(4): 440-443.
Shan, J., J. Huang, J. Liao, R. Robert and J. W. Hanrahan (2011). "Anion secretion by a model epithelium: more lessons from Calu-3." Acta Physiol (Oxf) 202(3): 523-531.
Shan, J., J. Liao, J. Huang, R. Robert, M. L. Palmer, S. C. Fahrenkrug, S. M. O'Grady and J. W. Hanrahan (2012). "Bicarbonate-dependent chloride transport drives fluid secretion by the human airway epithelial cell line Calu-3." J Physiol 590(Pt 21): 5273-5297.

162
Sharma, P., L. Dudus, P. A. Nielsen, H. Clausen, J. R. Yankaskas, M. A. Hollingsworth and J. F. Engelhardt (1998). "MUC5B and MUC7 are differentially expressed in mucous and serous cells of submucosal glands in human bronchial airways." Am J Respir Cell Mol Biol 19(1): 30-37.
Shen, B. Q., W. E. Finkbeiner, J. J. Wine, R. J. Mrsny and J. H. Widdicombe (1994). "Calu-3: a human airway epithelial cell line that shows cAMP-dependent Cl- secretion." Am J Physiol 266(5 Pt 1): L493-501.
Sheppard, D. N. (2004). "CFTR channel pharmacology: novel pore blockers identified by high-throughput screening." J Gen Physiol 124(2): 109-113.
Simpson, J. E., L. R. Gawenis, N. M. Walker, K. T. Boyle and L. L. Clarke (2005). "Chloride conductance of CFTR facilitates basal Cl-/HCO3- exchange in the villous epithelium of intact murine duodenum." Am J Physiol Gastrointest Liver Physiol 288(6): G1241-1251.
Slepkov, E. R., J. K. Rainey, B. D. Sykes and L. Fliegel (2007). "Structural and functional analysis of the Na+/H+ exchanger." Biochem J 401(3): 623-633.
Smith, J. J. and M. J. Welsh (1992). "cAMP stimulates bicarbonate secretion across normal, but not cystic fibrosis airway epithelia." J Clin Invest 89(4): 1148-1153.
Soleimani, M. and P. S. Aronson (1989). "Ionic mechanism of Na+-HCO3- cotransport in rabbit renal basolateral membrane vesicles." J Biol Chem 264(31): 18302-18308.
Song, Y., D. Salinas, D. W. Nielson and A. S. Verkman (2006). "Hyperacidity of secreted fluid from submucosal glands in early cystic fibrosis." Am J Physiol Cell Physiol 290(3): C741-749.
Stewart, A. K., B. E. Shmukler, D. H. Vandorpe, F. Reimold, J. F. Heneghan, M. Nakakuki, A. Akhavein, S. Ko, H. Ishiguro and S. L. Alper (2011). "SLC26 anion exchangers of guinea pig pancreatic duct: molecular cloning and functional characterization." Am J Physiol Cell Physiol 301(2): C289-303.
Svastova, E., A. Hulikova, M. Rafajova, M. Zat'ovicova, A. Gibadulinova, A. Casini, A. Cecchi, A. Scozzafava, C. T. Supuran, J. Pastorek and S. Pastorekova (2004). "Hypoxia activates the capacity of tumor-associated carbonic anhydrase IX to acidify extracellular pH." FEBS Lett 577(3): 439-445.
Takeyasu, K., M. M. Tamkun, K. J. Renaud and D. M. Fambrough (1988). "Ouabain-sensitive (Na+ + K+)-ATPase activity expressed in mouse L cells by transfection with DNA encoding the alpha-subunit of an avian sodium pump." J Biol

163
Chem 263(9): 4347-4354.
Talbot, C. L., D. G. Bosworth, E. L. Briley, D. A. Fenstermacher, R. C. Boucher, S. E. Gabriel and P. M. Barker (1999). "Quantitation and localization of ENaC subunit expression in fetal, newborn, and adult mouse lung." Am J Respir Cell Mol Biol 20(3): 398-406.
Tam, A., S. Wadsworth, D. Dorscheid, S. F. Man and D. D. Sin (2011). "The airway epithelium: more than just a structural barrier." Ther Adv Respir Dis 5(4): 255-273.
Tamada, T., M. J. Hug, R. A. Frizzell and R. J. Bridges (2001). "Microelectrode and impedance analysis of anion secretion in Calu-3 cells." JOP 2(4 Suppl): 219-228.
Tarran, R., B. Button and R. C. Boucher (2006). "Regulation of normal and cystic fibrosis airway surface liquid volume by phasic shear stress." Annu Rev Physiol 68: 543-561.
Tarran, R., B. Button, M. Picher, A. M. Paradiso, C. M. Ribeiro, E. R. Lazarowski, L. Zhang, P. L. Collins, R. J. Pickles, J. J. Fredberg and R. C. Boucher (2005). "Normal and cystic fibrosis airway surface liquid homeostasis. The effects of phasic shear stress and viral infections." J Biol Chem 280(42): 35751-35759.
Tarran, R., B. R. Grubb, J. T. Gatzy, C. W. Davis and R. C. Boucher (2001). "The relative roles of passive surface forces and active ion transport in the modulation of airway surface liquid volume and composition." J Gen Physiol 118(2): 223-236.
Tarran, R., L. Trout, S. H. Donaldson and R. C. Boucher (2006). "Soluble mediators, not cilia, determine airway surface liquid volume in normal and cystic fibrosis superficial airway epithelia." J Gen Physiol 127(5): 591-604.
Tarun, A. S., B. Bryant, W. Zhai, C. Solomon and D. Shusterman (2003). "Gene expression for carbonic anhydrase isoenzymes in human nasal mucosa." Chem Senses 28(7): 621-629.
Tessier, G. J., T. R. Traynor, M. S. Kannan and S. M. O'Grady (1990). "Mechanisms of sodium and chloride transport across equine tracheal epithelium." Am J Physiol 259(6 Pt 1): L459-467.
Thornton, D. J., K. Rousseau and M. A. McGuckin (2008). "Structure and function of the polymeric mucins in airways mucus." Annu Rev Physiol 70: 459-486.
Toczylowska-Maminska, R. and K. Dolowy (2012). "Ion transporting proteins of human bronchial epithelium." J Cell Biochem 113(2): 426-432.

164
Tos, M. (1966). "Development of the tracheal glands in man. Number, density, structure, shape, and distribution of mucous glands elucidated by quantitative studies of whole mounts." Acta Pathol Microbiol Scand 68: Suppl 185:183+.
Turner, R. J. and J. N. George (1988). "Cl(-)-HCO3- exchange is present with Na+-K+-Cl- cotransport in rabbit parotid acinar basolateral membranes." Am J Physiol 254(3 Pt 1): C391-396.
Ussing, H. H. and K. Zerahn (1999). "Active transport of sodium as the source of electric current in the short-circuited isolated frog skin. Reprinted from Acta. Physiol. Scand. 23: 110-127, 1951." J Am Soc Nephrol 10(9): 2056-2065.
Wagner, C. A., K. E. Finberg, S. Breton, V. Marshansky, D. Brown and J. P. Geibel (2004). "Renal vacuolar H+-ATPase." Physiol Rev 84(4): 1263-1314.
Wang, X., C. Lytle and P. M. Quinton (2005). "Predominant constitutive CFTR conductance in small airways." Respir Res 6: 7.
Wangemann, P. (2011). "The role of pendrin in the development of the murine inner ear." Cell Physiol Biochem 28(3): 527-534.
Welsh, M. J. and J. J. Smith (2001). "cAMP stimulation of HCO3- secretion across airway epithelia." JOP 2(4 Suppl): 291-293.
Wickstrom, C., J. R. Davies, G. V. Eriksen, E. C. Veerman and I. Carlstedt (1998). "MUC5B is a major gel-forming, oligomeric mucin from human salivary gland, respiratory tract and endocervix: identification of glycoforms and C-terminal cleavage." Biochem J 334 ( Pt 3): 685-693.
Widdicombe, J. H. (2002). "Regulation of the depth and composition of airway surface liquid." J Anat 201(4): 313-318.
Widdicombe, J. H., M. J. Welsh and W. E. Finkbeiner (1985). "Cystic fibrosis decreases the apical membrane chloride permeability of monolayers cultured from cells of tracheal epithelium." Proc Natl Acad Sci U S A 82(18): 6167-6171.
Wine, J. J. and N. S. Joo (2004). "Submucosal glands and airway defense." Proc Am Thorac Soc 1(1): 47-53.
Wingo, T., C. Tu, P. J. Laipis and D. N. Silverman (2001). "The catalytic properties of human carbonic anhydrase IX." Biochem Biophys Res Commun 288(3): 666-669.

165
Yang, Y. D., H. Cho, J. Y. Koo, M. H. Tak, Y. Cho, W. S. Shim, S. P. Park, J. Lee, B. Lee, B. M. Kim, R. Raouf, Y. K. Shin and U. Oh (2008). "TMEM16A confers receptor-activated calcium-dependent chloride conductance." Nature 455(7217): 1210-1215.
Yue, G. G., C. B. Lau, K. P. Fung, P. C. Leung and W. H. Ko (2008). "Effects of Cordyceps sinensis, Cordyceps militaris and their isolated compounds on ion transport in Calu-3 human airway epithelial cells." J Ethnopharmacol 117(1): 92-101.
Zuo, W. L., J. H. Huang, J. J. Shan, S. Li, P. Y. Wong and W. L. Zhou (2010). "Functional studies of acid transporter in cultured rat epididymal cell." Fertil Steril 93(8): 2744-2749.

















![A Cation-Chloride Cotransporter Gene Is … Cation-Chloride Cotransporter Gene Is Required for Cell Elongation and Osmoregulation in Rice1[OPEN] Zhi Chang Chen, Naoki Yamaji, Miho](https://static.fdocuments.net/doc/165x107/5d0f99e188c99345078b4fbd/a-cation-chloride-cotransporter-gene-is-cation-chloride-cotransporter-gene-is-required.jpg)
