
Mating prior to overwintering in the imported willow leaf beetle, Plagiodera versicolora...
6
Ecological Entomology (1989) 14,219-223 Mating prior to overwintering in the imported willow leaf beetle, PZugioderu versicolora (Coleoptera: C hrysomelidae) LORI STEVENS and DAVID E. McCAULEY* of Chicago, and *Department of General Biology, Vanderbilt University, Nashville Department of Biology, The University ABSTRACT. 1. We report on the mating status of overwintering adults of the imported leaf beetle, Plagioderu versicoloru Laicharting. This species overwinters as adults, under the bark of willow trees. 2. Individuals were collected both in the autumn, just after initiation of diapause, and in the spring, just before natural termination of diapause. From the autumn sample, we discovered that diapause can be terminated by husbanding the adults at 4°C for 4 weeks. 3. Most females lay fertile eggs upon the termination of diapause; how- ever, a significant number of females lay non-fertile eggs after overwinter- ing in the field. 4. Many females mate with more than one male before entering diapause. Progeny analysis using allozyme genetic markers shows that, on average, females who are fertile upon termination of diapause utilize sperm from 1.2 males to fertilize their eggs. Key words. Overwintering, multiple mating, diapause, Plugioderu versi- colora, Chrysomelidae. Introduction This study examines aspects of overwintering and diapause in the imported willow leaf beetle, Pfugioderu versicoloru (Laicharting), Chryso- melidae. We investigated: (1) whether females are inseminated when they enter diapause, and (2) whether females mate with more than one male before entering diapause. Adults of this species overwinter under the bark of willow trees (Hood, 1940). In Massachusetts it has been reported that the first and second generation adults feed, mate, and lay eggs. Adults of the third generation feed heavily and by the end of August practically all have entered diapause. A very few third generation adults mate and de- posit eggs that iiatch. Experimental attempts by Correspondence: Dr Lon Stevens, The University of Vermont, Department of Zoology, Marsh Life Sci- ence Building, Burlington, VT 05405-0086, U.S.A. Hood to induce diapause in first and second gen- eration adults were unsuccessful; however, third generation adults ‘voluntarily’ entered diapause (Hood, 1940). Although Hood’s observations imply that third generation adults either enter diapause or mate and lay eggs, this paper examines the possibility that adults mate before entering diapause. Previous studies from various geographic lo- cations have shown that females collected in the summer are often multiply mated (McCauley & O’Donnell, 1984; McCauley et ul., 1988). McCauley et ul. (1988) reported that the average degree of multiple paternity varies from locality to locality and also among generations across a season. It would be of interest, therefore, to de- termine whether the genetic composition of sib- ships born in the first generation of a given year is determined by the mating behaviour of the mother before entering or after terminating 219
-
Upload
lori-stevens -
Category
Documents
-
view
215 -
download
3
Transcript of Mating prior to overwintering in the imported willow leaf beetle, Plagiodera versicolora...

Ecological Entomology (1989) 14,219-223
Mating prior to overwintering in the imported willow leaf beetle, PZugioderu versicolora (Coleoptera: C hrysomelidae)
LORI STEVENS and DAVID E. McCAULEY* of Chicago, and *Department of General Biology, Vanderbilt University, Nashville
Department of Biology, The University
ABSTRACT. 1. We report on the mating status of overwintering adults of the imported leaf beetle, Plagioderu versicoloru Laicharting. This species overwinters as adults, under the bark of willow trees.
2. Individuals were collected both in the autumn, just after initiation of diapause, and in the spring, just before natural termination of diapause. From the autumn sample, we discovered that diapause can be terminated by husbanding the adults at 4°C for 4 weeks.
3. Most females lay fertile eggs upon the termination of diapause; how- ever, a significant number of females lay non-fertile eggs after overwinter- ing in the field. 4. Many females mate with more than one male before entering
diapause. Progeny analysis using allozyme genetic markers shows that, on average, females who are fertile upon termination of diapause utilize sperm from 1.2 males to fertilize their eggs.
Key words. Overwintering, multiple mating, diapause, Plugioderu versi- colora, Chrysomelidae.
Introduction
This study examines aspects of overwintering and diapause in the imported willow leaf beetle, Pfugioderu versicoloru (Laicharting), Chryso- melidae. We investigated: (1) whether females are inseminated when they enter diapause, and (2) whether females mate with more than one male before entering diapause. Adults of this species overwinter under the bark of willow trees (Hood, 1940). In Massachusetts it has been reported that the first and second generation adults feed, mate, and lay eggs. Adults of the third generation feed heavily and by the end of August practically all have entered diapause. A very few third generation adults mate and de- posit eggs that iiatch. Experimental attempts by
Correspondence: Dr Lon Stevens, The University of Vermont, Department of Zoology, Marsh Life Sci- ence Building, Burlington, VT 05405-0086, U.S.A.
Hood to induce diapause in first and second gen- eration adults were unsuccessful; however, third generation adults ‘voluntarily’ entered diapause (Hood, 1940). Although Hood’s observations imply that third generation adults either enter diapause or mate and lay eggs, this paper examines the possibility that adults mate before entering diapause.
Previous studies from various geographic lo- cations have shown that females collected in the summer are often multiply mated (McCauley & O’Donnell, 1984; McCauley et ul., 1988). McCauley et ul. (1988) reported that the average degree of multiple paternity varies from locality to locality and also among generations across a season. It would be of interest, therefore, to de- termine whether the genetic composition of sib- ships born in the first generation of a given year is determined by the mating behaviour of the mother before entering or after terminating
219

220
diapause. It is also of interest to compare the average number of mates for overwintering females with those reported in earlier studies. For these reasons we examined whether females are multiply inseminated when they enter diapause.
Lori Stevens and David E. McCauley
Materials and Methods
Adults in diapause (autumn sample) or dor- mancy (spring sample) were collected from under willow bark (Salk sp.). Two samples were collected from Van Patton Forest Preserve, Lake County, Illinois, twenty-eight adults were collected in the autumn (September 1986) and eighteen adults in the spring (April 1987). One collection of sixty-seven adults was made at Kellogg Biological Station, Kalamazoo County, Michigan, in the spring (March 1987). The sam- ples from Illinois were pooled and contrasted with the Michigan sample to determine if there was variation between these two geographic populations in the average number of mates per female. In addition we determined if over- wintering in the field affected the proportion of females that laid non-fertile eggs, possibly owing to sperm becoming inviable during the winter.
The autumn collection was made while the weather was still relatively warm, prior to a hard freeze; however, all of the willow leaves had fallen and the beetles were in diapause as deter- mined by their presence under bark, general in- active state, and lack of feeding when brought into the laboratory. The spring collections were made while the weather was still relatively sold (March and April) and we did not observe beetles active in the field at this time. Egg laying has been reported to begin in May in Illinois (Wade & Breden, 1986).
The insects collected in the spring were stored at 4°C for 1 week. Upon removal, they became active and started laying eggs. The spring stor- age at 4°C was done for convenience, to prevent the beetles from becoming active before we were ready to start the experiment. We believe they would have become active, fed and laid eggs im- mediately if permitted to do so, i.e. they were dormant, not in diapause (Tauber et al., 1986). This was not true for the insects collected in the autumn. In order to break diapause, the autumn collections were placed in a 4°C incubator, re- moved 4 weeks later, maintained at room temp-
erature and light conditions, and fed willow leaves. They started eating about a week later and laying eggs about 12 days later (see Results).
We had originally planned to determine the length of cold temperature required to terminate diapause by placing beetles in the 4°C incubator and removing adults at 4-week intervals. The first sample of adults became active and laid eggs; therefore, we conclude that this is the maximum cold period at this temperature re- quired to terminate diapause.
Parental individuals collected from the field were husbanded individually in 90 ml snap cap jars I containing willow leaves. Holes were punched in the caps to allow for air circulation and a moistened cotton swab was suspended through the cap. Fresh leaves were provided every 2-3 days. Newly laid eggs were moved to a separate jar and progeny were raised to adults. Both parental females and their offspring were stored at -70°C prior to starch-gel electro- phoresis of the protein phosphoglucomutase (PGM) (for methods see McCauley & O'Donnell, 1984). We were unable to determine the genotype of one of the females from the Illinois sample and excluded her from the analysis. Three of the fifteen females from the Michigan sample produced fewer than five adult progeny (some offspring died in rearing) and were excluded from the genetic analysis.
The genetic marker PGM has four alleles and has been used in several studies of paternity and group relatedness in the imported willow leaf beetle (McCauley & O'Donnell, 1984; McCauley et al., 1988). When the genotypes of the mother and a group of her offspring are known, the presence of at least three different paternally derived alleles among the offspring proves multiple paternity for that family. The number of cases of multiple paternity was esti- mated by counting the number of paternally derived alleles for each family. This underesti- mates the actual number of cases by about 50% (McCauley & O'Donnell, 1984) because it does not detect instances where a female uses the sperm of several males with identical PGM alleles. In addition, the paternal contribution cannot be identified when the mother and off- spring are identical heterozygotes. An estimate of the average number of mates per female was obtained using a statistical method described by Wilson (1981;seealsovanderHaveetaf., 1988). This method uses a one-locus two-allele model
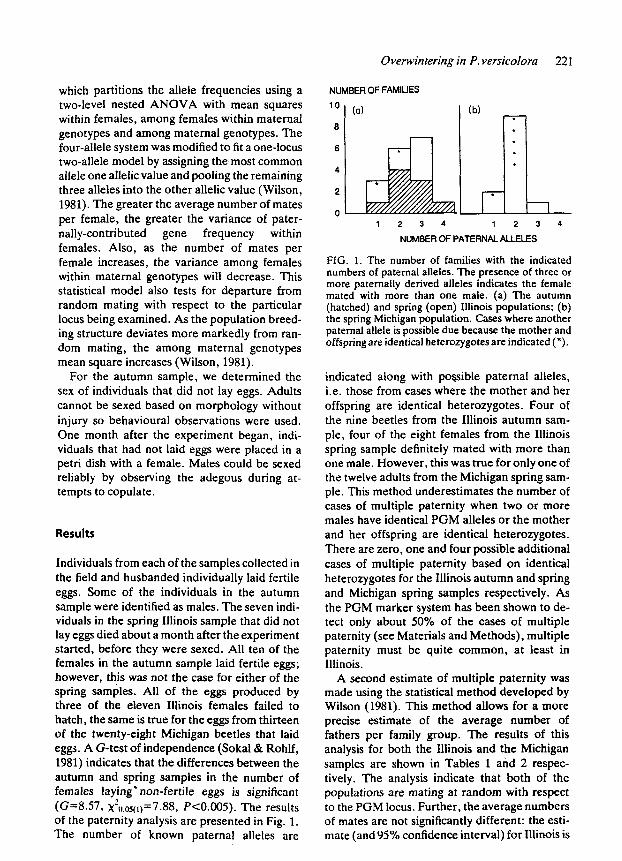
Overwintering in P. versicolora 221
which partitions the allele frequencies using a two-level nested ANOVA with mean squares within females, among females within maternal genotypes and among maternal genotypes. The four-allele system was modified to fit a one-locus two-allele model by assigning the most common aIlele one allelic value and pooling the remaining three alleles into the other allelic value (Wilson, 1981). The greater the average number of mates per female, the greater the variance of pater- nally-contributed gene frequency within females. Also, as the number of mates per female increases, the variance among females within maternal genotypes will decrease. This statistical model also tests for departure from random mating with respect to the particular locus being examined. As the population breed- ing structure deviates more markedly from ran- dom mating, the among maternal genotypes mean square increases (Wilson, 1981).
For the autumn sample, we determined the sex of individuals that did not lay eggs. Adults cannot be sexed based on morphology without injury so behavioural observations were used, One month after the experiment began, indi- viduals that had not laid eggs were placed in a petri dish with a female. Males could be sexed reliably by observing the adegous during at- tempts to copulate.
Results
Individuals from each of the samples collected in the field and husbanded individually laid fertile eggs. Some of the individuals in the autumn sample were identified as males. The seven indi- viduals in the spring Illinois sample that did not lay eggs died about a month after the experiment started, before they were sexed. All ten of the females in the autumn sample laid fertile eggs; however, this was not the case for either of the spring samples. All of the eggs produced by three of the eleven Illinois females failed to hatch, the same is true for the eggs from thirteen of the twenty-eight Michigan beetles that laid eggs. A G-test of independence (Sokal& Rohlf, 1981) indicates that the differences between the autumn and spring samples in the number of females laying' non-fertile eggs is significant (Gz8.57, x20.~s(t)=7.88, P<0.005). The results of the paternity analysis are presented in Fig. 1. The number of known paternal alleles are
NUMBER OF FAMILIES
1 2 3 4 1 2 3 4
NUMBER OF PATERNAL AUELES
FIG. 1. The number of families with the indicated numbers of paternal alleles. The presence of three or more paternally derived alleles indicates the female mated with more than one male. (a) The autumn (hatched) and spring (open) Illinois populations; (b) the spring Michigan population. Cases where another paternal allele is possible due because the mother and offspring are identical heterozygotes are indicated (*).
indicated along with possible paternal alleles, i.e. those from cases where the mother and her offspring are identical heterozygotes. Four of the nine beetles from the Illinois autumn sam- ple, four of the eight females from the Illinois spring sample definitely mated with more than one male. However, this was true for only one of the twelve adults from the Michigan spring sam- ple. This method underestimates the number of cases of multiple paternity when two or more males have identical PGM alleles or the mother and her offspring are identical heterozygotes. There are zero, one and four possible additional cases of multiple paternity based on identical heterozygotes for the Illinois autumn and spring and Michigan spring samples respectively. As the PGM marker system has been shown to de- tect only about 50% of the cases of multiple paternity (see Materials and Methods), multiple paternity must be quite common, at least in Illinois.
A second estimate of multiple paternity was made using the statistical method developed by Wilson (1981). This method allows for a more precise estimate of the average number of fathers per family group. The results of this analysis for both the Illinois and the Michigan samples are shown in Tables 1 and 2 respec- tively. The analysis indicate that both of the populations are mating at random with respect to the PGM locus. Further, the average numbers of mates are not significantly different: the esti- mate (and 95% confidence interval) for Illinois is

222 Lori Stevens and David E. McCauley
TABLE 1. Results of statistical analysis for the Illinois sample. This analysis tests for random mating and estimates the average number of mates per female. See text for a description of the method and Wilson (1981) for its derivation.
Component Sum of squares d.f. Mean square (MS) ~ ~~ ~~
Among maternal genotypes 0.1803 2 0.0902 Among mothers within genotypes 18.4504 14 1.3179 Among progeny within mothers 27.9947 170 0.1647
Test for random mating: Fl9=O.07, P=0.9342. - Estimate of the average number of mates (95% confidence interval): L’=1.27
(0.80,2.15).
TABLE 2. Results of statistical analysis for the Michigan sample. This analysis tests for random mating and estimates the average number of mates per female. See text for a description of the method and Wilson (1981) for its derivation.
Component Sumof squares d.f. Meansquare (MS)
Among maternal genotypes 2.5789 2 1.2895 Among mothers within genotypes 8.5086 9 0.9454 Among progeny within mothers 10.2255 74 0.1382
Test for random mating: F2,9=1.36, P=0.3038. Estimate of the average number of mates (95% confidence interval): =1.12
(0.67,2.20).
1.27 (0.80, 2.15) and for Michigan is 1.12 (0.67, 2.20). Because this statistical analysis inciudes only those females that laid fertile eggs; the lower limit should be: 1 .O + number of observed cases of multiple paternity. When the averages are adjusted to include those females that laid eggs that did not hatch, the estimate of the aver- age number of mates is 1.08 for the Illinois sam- ple and 0.54 for the Michigan sample. Both the method of counting paternal alleles and the statistical analysis indicate that in the Illinois population most females matk before they enter diapause; however, not all females lay fertile eggs and some females use sperm from more than one male, indicating they were multiply mated before entering diapmse. In contrast, al- most half (13/28=46%) of the females from the Michigan population did not lay viable eggs. Among the females that laid viable eggs, similar numbers used sperm from multiple males in both populations.
Discussion
This study shows that females of the imported willow leaf beetle, P.versicolora, overwinter in-
seminated and often utilize the sperm from more than one male when laying eggs in the spring. The observation that females usually mate before entering diapause differs from previous reports (Hood, 1940) that suggest that late sum- mer females either enter diapause as virgins or mate and lay eggs. Females that mate prior to overwintering may have an advantage in that they do not need to find mates prior to laying eggs in the spring. Mating in this species has been observed in the field in the spring as the first eggs appear (Wade & Breden, 1986); thus, if selection acts during the winter season, females may increase the fitness of their offspring by re- mating after dormancy with males that survive the winter. In addition, there is a chance that sperm can, become inviable during the winter and this possibility gives another advantage to remating in the spring. Males that mate prior to overwintering risk mating with females that
-don’t survive the winter; however when the males themselves don’t survive the winter but their mates do, their genes may be passed on nonetheless.
Early instar larvae of P.versicolora feed in aggregations that are a mixture of full and half sibs. McCauley er al. (1988) found a difference in

Overwintering in P. versicolora 223
the average genetic composition of family groups depending on whether the larval groups were collected during the first or second gener- ation. They ascribed this to temporal variation in the frequency of multiple insemination. The re- sults from this study show that the genetic com- position of first generation larval groups is a function of both the mating behaviour of females prior to initiating diapause in the autumn and any copulations they experience immediately after terminating diapause in the spring. Hulburt (1987) found little evidence of sperm precedence but rather considerable sperm mixing among the offspring of females mated to successive males. Thus, first generation larval groups could con- tain a mixture of individuals fathered by mates encountered by the mother both before and after diapause. Such a pattern could maximize both male and female fitness, adults can mate before entering diapause and mate again in the spring with individuals that survive the winter.
In summary, we have shown that females of this species overwinter inseminated, eliminating the need to find mates before they lay eggs in the spring. Females often mate with more than one male before entering diapause and have been observed mating in the spring when the first eggs appear. Thus the relatedness of larval groups in the spring is a function of both the females mat- ing behaviour in the autumn prior to entering diapause and in the spring prior to egg laying.
Acknowledgments
We thank C. A. Heckman, J. Cronberg and M. C. Whitlock for technical assistance and C. J. Goodnight and M. J . Wade for comments on the
manuscript. This work was supported by NSF grant BSR86-05672 to D. B. Mertz and C. J. Goodnight, an award from the Vanderbilt Uni- versity Research Council to D. E. McCauley and NIH grant GM222523 to M. J. Wade.
References
Hood, C.E. (1940) Life history and control of the im- ported willow leaf beetle. U.S. Department of Ag- riculture Circular, 572, 1-9.
Hulburt, A.V. (1987) The effects of multiple mating and patterns of sperm precedence in Plagiodera versicolora (Coleoptera: Chrysomelidae). M.S. thesis, University of Virginia, Charlottesville. Va.
McCauley, D.E. & O’Donnell, R. (1984) The effect of multiple mating on genetic relatedness in larval aggregations of the imported willow leaf beetle (Plagiodera versicolora Coleoptera: Chryso- melidae). Behavioral Ecology and Sociobiology,
McCauley, D.E., Wade, M.J., Breden, F. & Wohltman, M. (1988) Spatial and temporal vari- ation in group relatedness: evidence from the im- ported willow leaf beetle. Evolufion, 42, 184-192.
Sokal, R.R. & Rohlf, F.J. (1981) Biometry, 2nd edn. W. H. Freeman, San Francisco.
Tauber, M.J., Tauber, C.A. & Masaki, S. (1986) Sear sonal Adaptations of Insects. Oxford University Press, New York.
van der Have, T.M., Boomsa, J.J. & Menken, S.B.J. (1988) Sex-investment ratios and relatedness in the monogynous ant Lasius niger (L.). Evolution, 42,
Wade, M.J. & Breden. F. (1986) Life history of natural populations of the imported willow leaf beetle, Plagiodera versicolora (Coleoptera: Chryso-rnelidae). Annals of the Entomological Society of America, 79,73-79.
Wilson, J. (1981) Estimating the degree of polyandry in natural populations. Evolution. 35,664-673.
15,287-29 I .
160-172.
Accepted 26 October 1988



















