
Mapping long-term cortical maturation of the auditory ... · Salima Jiwani Doctor of Philosophy...
304
Mapping long-term cortical maturation of the auditory system in adolescents who are deaf and have used a unilateral cochlear implant to hear by Salima Jiwani A Thesis submitted in conformity with the requirements for the degree of Doctor of Philosophy Institute of Medical Sciences University of Toronto © Copyright by Salima Jiwani 2015
-
Upload
doankhuong -
Category
Documents
-
view
213 -
download
0
Transcript of Mapping long-term cortical maturation of the auditory ... · Salima Jiwani Doctor of Philosophy...

Mapping long-term cortical maturation of the auditory system in
adolescents who are deaf and have used a unilateral cochlear implant
to hear
by
Salima Jiwani
A Thesis submitted in conformity with the requirements
for the degree of Doctor of Philosophy
Institute of Medical Sciences
University of Toronto
© Copyright by Salima Jiwani 2015

ii
Mapping long-term cortical maturation of the auditory system in
adolescents who are deaf and have used a unilateral cochlear implant
to hear
Salima Jiwani
Doctor of Philosophy
Institute of Medical Sciences
University of Toronto
2015
Abstract
In the present Thesis, we used novel imaging tools to study maturation of the auditory brain
in adolescents who are deaf and have use a unilateral cochlear implant (CI) to hear for most of their
lives. A CI is a surgically implanted auditory prosthesis that establishes hearing in children who are
deaf. The aims of implantation are to halt any effects of deafness on the brain and promote normal
auditory development. Unfortunately, CIs do not restore normal hearing as they provide only a crude
representation of sounds and eliminate important cochlear processing. It has recently been shown
that unilaterally stimulating the auditory system with a CI and leaving the opposite pathways deprived
of input for longer than 1.5 years compromises bilateral auditory development. We are now exploring
the cortical consequences of missing this sensitive period and driving maturation of the auditory cortex
with unilateral implant stimulation.
We measured electrically-evoked cortical responses in adolescents who had over a decade of
unilateral CI experience before receiving a second implant in the opposite deprived ear. This provided

iii
an unparalleled opportunity to assess the effects of long-term unilateral stimulation/deprivation on
the auditory pathways in the adolescent brain. We tracked the development of cortical responses with
CI experience and localized underlying areas of cortical activity in the brain using our beamformer
imaging methods. Neural synchrony of these responses was calculated to assess the co-ordination of
activity across brain regions in response to sound.
Our results indicate that long-term unilateral implant stimulation promotes normal-like
maturation of the auditory cortex with good speech perception outcomes, providing a general
impression that some degree of auditory development proceeds normally. However, abnormally
strengthened activity from the hearing ear to the contralateral cortex and increased synchrony in
networks known to be involved in cognitive processing suggests that cortical abnormalities persist
into maturation. In the opposite deprived ear, cortical responses were atypical, had abnormally large
dipoles and abnormal neural synchrony, perhaps reflecting cortical un-coupling/dis-connectivity in
response to sound. We suggest that unilateral maturation of the auditory cortex drives lasting auditory
asymmetries and leaves the deprived pathways unprotected from deafness-induced abnormalities.

iv
Acknowledgements
The pages of this Thesis hold much more than a culmination of years of studies. This Thesis
is built on the shoulders of more people than I can count, who have had a hand in shaping my personal
and intellectual growth. You have all thought me to push the boundaries of innovative research and
clinical practice to help people hear better and you have shown me how fun this can be! I came to
Archie’s Cochlear Implant Lab as an Audiologist. I now prepare to leave as an Audiologist-Scientist.
For that, I have many people to thank…
First and foremost, I would like to thank all of the children and adolescents who participated in my
studies. Thank you to you and your families for lending us your ears/implants and brains. This work
could not have been done without your generosity, contribution and time.
To my supervisor, Dr. Karen Gordon: Thank you for welcoming me as part of the CI family.
Working with you over the last few years has been an inspiration. You have thought me to think
smarter, write smarter, present smarter, and generally be smarter. Thank you. Thank you for your
incredible mind, your mentorship and your kind heart. Thank you also for always taking the time to
talk through ideas and thoughts with me, for opening doors of opportunity for me, and for becoming
a life-long friend. You truly are a one-of-a-kind Audiologist-Scientist, and I thank you for showing
me the path to success.
To my committee members, Dr. Robert Harrison, Dr. Margot Taylor and Dr. Sam Doesburg: Thank
you for always providing guidance and advice. Bob, you were the first to show me the wonderful
world of research at SickKids and you inspired me to follow a path of research with Karen. Thank
you for always having faith in me, for always asking the right questions, and for pushing me to be
better. Dr. Taylor, Thank you for always being so positive about my work and my abilities, for always
leaving revisions with me, and being so supportive. Sam, Thank you for always having the right ideas
to make the work better and more objective, for always making the time to explain methods to me,
and for always being so encouraging.
To the CI surgeons, Dr. Blake Papsin, Dr. Adrian James and Dr. Sharon Cushing: Thank you for
always sharing your brilliant minds with me, and always welcoming me into your ORs. Dr. Papsin,
Thank you for being an incredible leader, for always sharing your wisdom and for the many life/career
chats. Thank you also for always thinking about our data in terms of the big picture and all the many

v
things we can do with it to make a difference in a child’s life. I think the Nobel is on its way! Dr.
James and Dr. Cushing, Thank you for always smiling, for your constant drive to promote better
clinical care, and for teaching me how important it is to ask unique research questions.
To the CI Audiology Team, Vicky Papaioannou, Gina Goulding, Laurie MacDonald, Valérie Simard,
Mary-Lynn Feness, Pat Di Santo, Susan Druker, Rebecca Malcolmson and Cléo Audet-Halde: Thank
you all for your willingness to help and for sharing your knowledge, expertise, and sometimes even
your offices with me. You are an amazing group of people and I am lucky to have had the chance to
work with you all. To Patt Fuller and Debbie Andrade, Thank you for the laughs and all the help in
the clinic and the lab.
To the CI team at Archie’s Cochlear Implant Lab: You became my family and I am so fortunate to
have you in my life. Thank you for your hard work, your high spirits, your inquisitive nature, your
ability to think outside the box and making research so fun! Stephanie Jewell, you and I have shared
way too many laughs and adventures together! Thank you for your good nature and friendship. I am
lucky to have found you. Nikolaus Wolter, Thank you for sharing the Tower with me! Thank you
also for teaching me an abundance of interesting and random facts about science, otolaryngology,
history and animals. Thank you for your constant willingness to help others, for always finding the
good in all situations, for all of your funny jokes and for your friendship. Melissa Polonenko, Thank
you for making everyday more interesting, for always asking the difficult questions, and for pushing
me to do better. Parvaneh Abbasalipour, Thank you for always motivating me to work even harder
and helping me push my limits, as you always do. Thank you also for all of the time spent collecting
data and troubleshooting equipment with me. Patrick Yoo and Daniel Wong, Thank for spending all
of those days and weeks and months working through the Beamformer with me and the fun times in
the Satellite lab. Jerome Valero, Thank you for taking me under your wing early on, showing me the
ropes, and all the delicious lunch dates. Talar Hopyan, Sara Giannatonio, and Shazia Peer, Thank you
for always being so funny, so crazy and so light hearted! Claire Salloum, Sho Tanaka, Michael
Deighton, Morrison Steel, Désirée DeVreede, Lauren Schofield, Hailey Ainley, Catharine McCann,
Cullen Allemang, Bridget Allemang, Gurvinder Toor, Luis Vilchez Madrigal, Tulika Shingal, Heather
Osborn, Tony Eskander, William Parkes, Carmen Knight, Vijayalakshmi Easwar, Joshua
Gnanasegaram, Michael Chaikoff, Patricia Ungureanu, Mikaeel Valli, Eden Amber, Thank you for
making me feel at home in the lab.

vi
Finally, I would like to thank my family and friends. I am everything that I am because of you. Thank
you. Merci.
To my silent supporters, Dr. Steve Aiken and Philippe Fournier: Thank you for being the best of
cheerleaders and for inspiring me to make a difference in our field of Audiology.
To Akbar and Nasim Dharssi, Karim, Farrah and the boys: Thank you for your constant
encouragement and for always believing in me.
A mon père, ma mère, mes sœurs (Soraya et Sabrina), mes grands-parents (Papaji, Mama, Mama), et
les p’tit chouchous: Merci pour tout. Merci d’avoir toujours cru en moi, de m’avoir toujours
encouragé, et de m’avoir toujours supporté, en particulier dans les moments délicats. Mille Mercis
pour votre patience, générosité, bonne humeur, splendide énergie et bonté de cœur. Je vous dois
absolument tout.
To my dear husband, Salim Dharssi: Thank you for being my true better half over the last few years.
Thank you for always listening to me prep for talks, reading all my applications, and the countless
hours you spent helping me with the beamformer. Thank you for all of your support, all of your
laughs, all of your jokes, all of your kindness, and all of your heart. You inspire me to reach for the
stars and to be better. You have made my life richer than I ever could have imagined.
Contributions
Dr. Karen Gordon and Dr. Blake Papsin were the vision behind the studies presented in this Thesis.
They contributed to all aspects of the work. My Thesis advisory committee, Dr. Robert Harrison, Dr.
Margot Taylor and Dr. Sam Doesburg provided continuing guidance with the data analyses and results
for all studies, particularly the ones in Chapters Four and Five.
Stephanie Jewell and Parvaneh Abbasalipour helped with the EEG recordings for the data presented
in Chapters Three, Four and Five.
Daniel Wong developed the TRACS beamformer imaging method used in Chapter Four. Salim
Dharssi helped me develop an objective method to analyze the beamformer data and visualize dipole

vii
activity in 63,646 voxels in the brain, shown in Figures 4-2 and 4-4. Alexander Andrews provided
some assistance with this as well. Dr. Sam Doesburg and Marc Lalancette provided assistance with
the permutation analyses used in Chapter Four and shown in Figures 4-3, 4-5 and 4-6. Dr. Sam
Doesburg also provided the Matlab and Mathematica codes for the connectivity analyses in Figures
5-1, 5-2, 5-3. Catharine McCann helped me validate the suppression of the implant artifact using
independent component analyses, shown in Supplementary Figure 5-4.
Funding and Studentships
Studentship funding for my Doctoral work has been provided by an Entrance Award and Open
Fellowship from the Institute of Medical Sciences at the University of Toronto, a Post-Graduate
Medical Award (PGME) from the Faculty of Medicine at the University of Toronto, the Hilda &
William Courtney Clayton Paediatric Research Fund Fellowship, the Margaret & Howard Gamble
Research Grant, several Ontario Graduate Scholarships (OGS) from the Ontario Ministry of Training,
Colleges and Universities, a Sick Kid’s Research Training Competition Studentship (RESTRACOMP)
and a Sick Kid’s Clinician-Scientist Training Program Studentship (CSTP) from the Hospital for Sick
Children Foundation Student Scholarship Program, and a Medical Research Grant from The Hearing
Foundation of Canada. I was also granted a Conference Award from the School of Graduate Studies
at the University of Toronto, two Trainee Travel Awards from The Hospital for Sick Children
Foundation, and numerous Travel Awards from the National Institute of Health – National Institute
of Deafness and Other Communication Disorders (NIH NIDCD).

viii
Table of Contents
Abstract ..................................................................................................................................................... ii
Acknowledgements ................................................................................................................................ iv
Contributions .......................................................................................................................................... vi
Funding and Studentships.................................................................................................................... vii
Table of Contents................................................................................................................................. viii
List of Tables ......................................................................................................................................... xv
List of Figures ....................................................................................................................................... xvi
List of Abbreviations ........................................................................................................................... xix
Thesis Roadmap .................................................................................................................................... xx
1. Chapter One – Research rationale, questions and hypotheses ................................................... 1
2. Chapter Two – General introduction and background................................................................ 4
2.1 Cochlear implants establish hearing in children who are deaf ................................... 4
2.2 Normal hearing requires intact auditory structures and functions ............................ 9
2.3 Milestones in auditory development ............................................................................ 15
2.4 Auditory experience shapes auditory development ................................................... 19
2.5 Bilateral deafness in childhood drives abnormal reorganization in the brain ........ 22
2.6 Unilateral deafness promotes abnormal changes in the auditory brain .................. 26
2.7 Multiple effects of childhood deafness predict outcomes after cochlear
implantation ..................................................................................................................... 28
2.8 Electrophysiological measures assess auditory cortical development and map
underlying auditory activity in the brain ...................................................................... 33
2.9 Unilateral cochlear implantation restores hearing and promotes auditory
development in the brainstem and midbrain, but the trajectory of cortical auditory
maturation remains unclear ........................................................................................... 37

ix
2.10 Differences from normal persist in auditory processing despite long durations of
unilateral cochlear implant use...................................................................................... 41
2.11 Binaural processing is not available with unilateral hearing ..................................... 43
2.12 Evidence of a short sensitive period for bilateral input in human auditory
development .................................................................................................................... 45
2.13 Does long-term unilateral cochlear implant use have abnormal consequences for
cortical auditory development? ..................................................................................... 51
3. Chapter Three – Central auditory development after long-term cochlear implant use ........ 54
3.1 Abstract .................................................................................................................................... 54
3.2 Introduction ............................................................................................................................. 55
3.3.1 Deafness prior to cochlear implantation alters normal brain development .......... 55
3.3.2 Early auditory cortical development in cochlear implant users follows a normal-
like trajectory ................................................................................................................... 56
3.3.3 Auditory cortical maturation may be altered in cochlear implant users ................. 57
3.4 Methods .................................................................................................................................... 58
3.4.1 Participants ...................................................................................................................... 58
3.4.2 Evoked potential recordings ......................................................................................... 61
3.4.3 Analysis of the electrically-evoked cortical responses ............................................... 62
3.5 Results ....................................................................................................................................... 64
3.5.1 Cortical responses continue to mature with auditory experience in normal hearing
children and in users of cochlear implants.................................................................. 64
3.5.2 Cortical development in users of cochlear implants follows a normal trajectory with
time-in-sound with differences emerging in latencies greater than 150ms ............ 66
3.5.3 Normal-like cortical maturation in the 50 to 150ms latency range is time and
experience dependent..................................................................................................... 69
3.5.4 Cortical abnormalities in the 150 to 300ms latency range is experience dependent .
........................................................................................................................................... 71
3.6 Discussion ................................................................................................................................ 73

x
3.6.1 Cortical activity early in development is similar between cochlear implant users and
their normal hearing peers ............................................................................................. 73
3.6.2 Long-term cortical development follows a normal-like trajectory with time-in-
sound ................................................................................................................................ 76
3.6.3 Differences from normal in the later cortical peaks may reflect increased cortical
activity from non-auditory modalities ......................................................................... 78
3.7 Conclusion ............................................................................................................................... 80
4. Chapter Four – Early unilateral cochlear implantation promotes mature cortical asymmetries
in adolescents who are deaf ............................................................................................................ 82
4.1 Abstract .................................................................................................................................... 82
4.2 Introduction ............................................................................................................................. 83
4.3 Materials and Methods ........................................................................................................... 86
4.3.1 Participants ...................................................................................................................... 86
4.3.2 Recording cortical responses ........................................................................................ 87
4.3.3 Localization of cortical evoked peaks .......................................................................... 89
4.3.4 Speech perception tests to assess outcomes with CIs ............................................... 90
4.4 Results ....................................................................................................................................... 91
4.4.1 Tone-bursts preferentially stimulate the right auditory cortex in adolescents with
normal hearing ................................................................................................................ 91
4.4.2 Long periods of unilateral CI use drive abnormal patterns of auditory activity ... 95
4.4.3 Additional cortical areas are recruited by cochlear implant stimulation relative to
normal ........................................................................................................................... 101
4.4.4 Abnormal activity evoked by the naïve side predicts poor speech perception
outcomes ....................................................................................................................... 103
4.5 Discussion .............................................................................................................................. 104
4.5.1 Hemispheric specialization requires normal bilateral hearing ............................... 105
4.5.2 Long periods of unilateral CI use strengthens pathways from the stimulated ear ....
........................................................................................................................................ 107

xi
4.5.3 Activity evoked by stimulation of the newly implanted ear is abnormal ............. 110
4.6 Conclusion ............................................................................................................................. 111
4.7 Supplementary Information on Methods .......................................................................... 113
5. Chapter Five – Temporally coordinated activity in the brain is promoted by long-term
cochlear implant use in children ................................................................................................. 117
5.1 Abstract .................................................................................................................................. 117
5.2 Introduction ........................................................................................................................... 118
5.3 Methods .................................................................................................................................. 122
5.3.1 Participants and evoked potentials recordings ........................................................ 122
5.3.2 Phase synchronization analysis .................................................................................. 124
5.4 Results ..................................................................................................................................... 126
5.4.1 Less cortical synchrony is evoked from tone-bursts in normal right than left ears ..
........................................................................................................................................ 126
5.4.2 Right cochlear implants promote atypical cortical synchrony and leave deprived
pathways abnormally desynchronized ...................................................................... 130
5.5 Discussion .............................................................................................................................. 135
5.5.1 A specialized cortical hearing network normally matures by adolescence .......... 135
5.5.2 Increased connectivity in long-term unilateral CI users reflects greater processing
demands ........................................................................................................................ 137
5.5.3 Desynchronized activity evoked by the naïve-left CI suggests disorganization in the
deprived pathways ....................................................................................................... 139
5.6 Conclusion ............................................................................................................................. 142
5.7 Supplementary Information for Methods ......................................................................... 143
5.7.1 Independent component analysis to reject cochlear implant artifact .................. 143
5.7.2 Scalp current density to reduce spurious synchronization cause from volume
conduction .................................................................................................................... 146

xii
6. Chapter Six – General discussion ............................................................................................... 148
6.1 Normal maturation of the auditory system requires hearing in both ears to be
normal ..................................................................................................................................... 150
6.2 Unilateral implant stimulation promotes cortical maturation but leaves the brain with
abnormal organization.......................................................................................................... 154
6.3 Long-term unilateral deprivation drives abnormally altered and disorganized activity in
the unstimulated pathways................................................................................................... 161
7. Chapter Seven – Current and future directions ................................................................... 168
7.1 Does bilateral cochlear implant experience promote auditory development in pathways
from the newly implanted side? .......................................................................................... 168
7.2 Does the presence of residual hearing in the un-implanted ear protect these pathways
from abnormal effects of unilateral deprivation? ............................................................. 169
7.3 Can auditory development in the second implanted ear be promoted by using an aural
patching method? .................................................................................................................. 170
7.4 Is auditory activity evoked by cochlear implants stimulation mediated by mechanisms
of attention or multi-sensory stimulation? ........................................................................ 171
7.5 Can attention-driven and/or multi-modal auditory therapy drive improvements in
auditory processing and ease of listening? ......................................................................... 172
7.6 Can holistic therapies that incorporate music and/or exercise promote improvements
in auditory processing and accelerate auditory development after cochlear implantation?
................................................................................................................................................. 174
8. Chapter Eight – Conclusion ........................................................................................................ 176
References ........................................................................................................................................... 178
Appendices .......................................................................................................................................... 230

xiii
What is the optimal timing for bilateral cochlear implantation in children? ............................. 230
Abstract ............................................................................................................................................ 230
Introduction .................................................................................................................................... 231
Groups of Study Participants ........................................................................................................ 233
Mismatches in bilateral activity following a period of unilateral cochlear implant use ........ 234
Auditory brainstem responses ................................................................................................. 234
Cortical responses ..................................................................................................................... 237
Perception of speech and inter-implant timing and level cues ................................................ 239
Conclusion ....................................................................................................................................... 241
References ........................................................................................................................................ 242
Benefits and detriments of unilateral cochlear implant use on bilateral auditory development in
children who are deaf ................................................................................................................... 246
Abstract ............................................................................................................................................ 246
Introduction .................................................................................................................................... 246
The auditory system reorganizes when bilaterally deprived ..................................................... 247
Unilateral cochlear implantation restores hearing and promotes auditory development .... 249
Differences from normal persist in auditory processing despite long durations of unilateral
cochlear implant use.................................................................................................................................. 255
Binaural hearing is not available to traditional unilateral cochlear implant users .................. 256
Evidence of a short sensitive period for bilateral input in human auditory development .. 258
Long-term unilateral implant use in older children causes lasting asymmetry in the bilateral
auditory pathways ...................................................................................................................................... 264

xiv
Bilateral implantation within a sensitive period improves perception of binaural timing
cues 266
Conclusions ..................................................................................................................................... 268
References ........................................................................................................................................ 269
Copyright Acknowledgements ......................................................................................................... 282
Peer-Reviewed Publications .......................................................................................................... 282
Peer-Reviewed Presentations ........................................................................................................ 283

xv
List of Tables
Table 2-1: Milestones of structural, electrophysiological and behavioural maturation of activity in the
central auditory pathway. Reproduced with permission from Eggermont and Moore (2012) ........... 19
Table 3-1: Mean (X) ± standard deviations (SD) of demographic information of the CI users. ....... 60
Table 5-1: Demographic Information ...................................................................................................... 123

xvi
List of Figures
Figure 2-1: Illustration of the external and internal components of a CI device in a diagram of the
ear. Figure reproduced with permission from Papsin and Gordon (2007). ............................................. 7
Figure 2-2: Diagram of the ascending auditory pathway. Figure reproduced with permission from
(Netter 2010). ................................................................................................................................................... 11
Figure 2-3: Conceptual diagram of ascending projections of the primary auditory pathway (red boxes)
and the non-primary auditory pathways (green boxes). ............................................................................. 15
Figure 2-4: Widespread effects of bilateral deafness in all levels of the auditory pathways. Figure
reproduced with permission from Kral and Sharma (2012). ..................................................................... 24
Figure 2-5: Heterogeneity in the cause and onset of pediatric deafness. Figure reproduced with
permission from Morton and Nance (2006). ............................................................................................... 29
Figure 2-6: P1 latency as a function of age at implantation. Figure adapted then reproduced with
permission from Sharma, Dorman et al. (2002). ......................................................................................... 31
Figure 2-7: Speech perception outcomes as a function of age at implantation. Figure reproduced
with permission from Harrison, Gordon et al. (2005). .............................................................................. 32
Figure 2-8: Development of auditory brainstem responses after cochlear implantation. Figure
reproduced with permission from Gordon, Jiwani et al. (2013). .............................................................. 39
Figure 2-9: Development of auditory middle latency responses after cochlear implantation. Figure
reproduced with permission from Gordon, Jiwani et al. (2013). .............................................................. 41
Figure 2-10: Accuracy and reaction time for CI processing of auditory input with and without visual
input. .................................................................................................................................................................. 43
Figure 2-11: Cortical dipole activity evoked by auditory input in children with different durations of
unilateral implant use. Reproduced with permission from Gordon, Wong et al. (2013). .................... 51
Figure 3-1: Normal hearing mature cortical response. .............................................................................. 62

xvii
Figure 3-2: Development of the cortical response over time. ................................................................. 65
Figure 3-3: Cortical response difference as a function of time after cochlear implantation compared
to the normal hearing mature waveform. ..................................................................................................... 68
Figure 3-4: Difference in latency and amplitude of the P1-N1 complex as a function of time-in-sound
compared to normal hearing peers. ............................................................................................................... 70
Figure 3-5: Difference in latency and amplitude of the P2-N2 complex as a function of time-in-sound
compared to normal hearing peers. ............................................................................................................... 72
Figure 4-1: Balanced stimulus levels between the experienced and newly implanted ears determined
by matching peak eV amplitude of the brainstem response. ..................................................................... 88
Figure 4-2: Cortical dipole activity evoked by auditory stimulation of the right and left ears of normal
hearing adolescents. ......................................................................................................................................... 93
Figure 4-3: Aural preference evoked by auditory stimulation of the right and left ears of normal
hearing adolescents. ......................................................................................................................................... 94
Figure 4-4: Cortical dipole activity evoked by auditory stimulation of the experienced-right and naïve-
left ears of CI users. ......................................................................................................................................... 98
Figure 4-5: Cortical lateralization evoked by stimulation of the right/experienced and left/naïve
ear/implant in normal hearing adolescents and CI users. ...................................................................... 101
Figure 4-6: Group differences in cortical dipole activity between normal hearing adolescents and CI
users. ............................................................................................................................................................... 103
Figure 4-7: Speech perception performance on the experienced-right and naïve-left sides in CI users.
......................................................................................................................................................................... 104
Supplementary Figure 4-8: Example from one child with 15.95 years of CI experience in the right
ear indicates activity underlying the mature peak P2. ............................................................................... 116
Figure 5-1: Cortical oscillatory synchronization evoked by auditory stimulation of the right and left
ears of normal hearing adolescents. ........................................................................................................... 129

xviii
Figure 5-2: Cortical oscillatory synchronization evoked by auditory stimulation of the experienced-
right and naïve-left ears in CI users. ........................................................................................................... 133
Figure 5-3: Difference in cortical oscillatory synchronization evoked by auditory stimulation of the
right/experienced and left/naive ears/implant of normal hearing adolescents and CI users. .......... 134
Supplementary Figure 5-4: Independent component analysis to remove contamination of CI
artefact on the cortical response. ................................................................................................................ 146

xix
List of Abbreviations
AEP = Auditory Evoked Potentials
A-V Therapy = Auditory-Verbal Therapy
AVCN = Anteroventral Cochlear Nucleus
CAST = Computer-Assisted Speech Training
CI = Cochlear Implant
GJB-2 protein = Gap Junction Beta-2 protein
EEG = Electro-encephalography
FFW = Fast ForWord
ICA = Independent Component Analysis
LSO = Lateral Superior Olive
MEG = Magneto-encephalography
MNI = Montreal Neurological Institute
MSO = Medial Superior Olive
MRI = Magnetic Resonance Imaging
PBK words = Phonetically Balanced Kindergarden words
PET = Positron Emission Tomography
SOC = Superior Olivary Complex
TRACS Beamformer = Time Restricted Artifact and Coherent Suppression Beamformer

xx
Thesis Roadmap
This Thesis is organized into eight main chapters and one appendix chapter. Chapter One
and Two describe a general introduction to the main research questions explored in Chapters Three,
Four and Five. Background literature in this chapter consists of a description of auditory system
physiology and development both in children with normal hearing and known results from children
using cochlear implants. Chapter Three explores the long-term cortical auditory development in users
of cochlear implants, from infancy to adolescence. The trajectory of cortical auditory development in
this population is compared to that of normal hearing peers who are matched for chronological age
and hearing-age. This chapter has been published in Clinical Neurophysiology. In Chapters Four and
Five, we unwrap the cortical response and use a newly developed beamformer imaging method
(Chapter Four) and analyses of neural synchronization (Chapter Five) to map the activity underlying
the cortical response in both CI users and their peers with normal hearing. The beamformer imaging
method used in Chapter Four is an objective method that allows us to localize the cortical generators
underlying the recorded cortical activity in the temporal, frontal, parietal and occipital lobes in each
hemisphere when the first and second implants are stimulated separately. This manuscript has been
submitted to Human Brain Mapping and is currently under revision. In Chapter Five, we assess
whether different brain regions respond to sound in a synchronized, co-ordinated manner to
determine how different areas of the brain communicate with each other in response to sound.
Interactions across brain areas are believed to reflect cognitive processes that cochlear implant users
recruit to process auditory input. We compare this to normal hearing individuals to map the auditory
cortical network evoked by sound stimulation of the first and second implants. This manuscript has
been submitted to Cerebral Cortex and is currently under revision. Chapter Six draws the three studies
together into a general discussion. Chapter Seven provides a brief outline of relevant research that is
currently being conducted in our laboratory and suggests new directions for future research. Here, I
stress the importance of developing appropriate, effective and targeted auditory rehabilitation
therapies. Finally, I conclude this work in Chapter Eight with a brief recapitulation of the findings
shared in the present Thesis. The Appendix section of this Thesis includes two related review papers
published in Cochlear Implants International and Frontiers in Psychology. These papers are related
to this work but were written with a specific focus to address effects of unilateral CI stimulation on
binaural hearing after bilateral implantation. In these papers, we suggested that cochlear implants
should be provided to children early and in both ears.

1
1. Chapter One – Research rationale, questions and hypotheses
Children with severe to profound sensorineural hearing loss can now hear and develop oral
speech and language abilities with the use of an auditory prosthesis that is surgically placed in the
cochlea (inner ear), called a cochlear implant (CI). A CI is a small and complex electronic device that
sends electrical pulses from a series of electrodes in the cochlea to stimulate the auditory nerve with
auditory input. This input is carried by the auditory pathways to various regions of the cortex. The
user ultimately perceives this signal as sounds. The aims of implantation in children are to halt/reverse
any effects of deafness on the brain and to promote normal auditory development. Unfortunately,
CIs do not restore normal hearing, only provide a crude representation of acoustic sounds and may
not be able to completely reverse the effects of deafness in early life, which is likely to compromise
both of these objectives. It is clear that providing CIs early in life limits deafness-induced
reorganization of the auditory brain, as CI users show increasing activity in the auditory cortex (Suarez
et al., 1999, Pantev et al., 2006) and improvements in speech and language outcomes (Harrison,
Gordon et al. 2005) as they adapt to the artificial electrical stimulation from this unique device.
Unfortunately however, implants were traditionally provided to children in only one ear (i.e.,
unilaterally). Thus, the coupling of atypical electrical input delivered by this device to the stimulated
pathways with the long-term unilateral absence of auditory input to the opposite and deprived side
might drive abnormal maturation of auditory thalamo-cortical and cortico-cortical connections
(Ponton and Eggermont 2001; Eggermont and Ponton 2003) and/or maladaptive reorganization in
the auditory brain (Lee and Winer 2005; Lee, Giraud et al. 2007). These abnormalities could in turn
affect cognitive and perceptual functions such as memory processes, attention, and executive
processes. At the same time, it has recently been shown that leaving the opposite ear deprived of
auditory input beyond an early sensitive period of 1.5 years (Gordon, Wong et al. 2013), which
corresponds to the time-course of brainstem maturation, leaves those pathways unprotected from
abnormal deafness-induced reorganization and compromises bilateral auditory development (Gordon,
Jiwani et al. 2013; Gordon, Wong et al. 2013).
In the experiments of the present Thesis, we explore whether missing this early sensitive period
and driving maturation of the auditory cortex with over a decade of unilateral CI stimulation causes
permanent abnormalities in cortical activity and reorganization in the deprived pathways. In general,
we are asking whether long-term unilateral implant use drives development of pathways from this ear

2
at the expense of pathways from the other ear. Recently, many adolescents that we follow in our CI
Program who had over a decade of unilateral hearing experience with their implant have received a
second device in their opposite-deprived ear. This gave us a unique opportunity to explore the effects
of long-term unilateral implant stimulation/deprivation in the adolescent brain and ask whether the
development of cortical activity in the deprived auditory pathways is compromised by long-term
unilateral implant stimulation/deprivation. Through the course of my Doctorate work, we have
developed the necessary tools to image cortical function in this population using measures of electric
current evoked in the auditory cortex by the implant. We are now in a unique position to answer
important questions regarding the impact of early unilateral cochlear implantation on the development
of the auditory brain network and assess the extent to which our goals of preserving normal auditory
development in children who hear with a unilateral CI have been realized by asking the following
questions in chapters Three, Four and Five:
Chapter Three:
1) Does the auditory system mature in children who are deaf with long-term unilateral CI use?
Chapter Four:
2) Does long-term unilateral CI stimulation promote activity in cortical areas which normally
respond to sound?
3) Is this activity compromised in the deprived contralateral pathways?
Chapter Five:
4) Does long-term unilateral CI stimulation promote coordinated cortical activity (i.e., neural
networks), which is normally activated in response to sound?
5) Are the pathways from the deprived ear segregated from these cortical hearing networks?
We hypothesize that long-term unilateral CI stimulation in children will promote development
of the auditory cortex at normal rates, but will not eliminate reorganization of the brain caused by
bilateral deafness prior to implantation and unilateral deprivation afterward. We expect to find that
CI users will recruit more cortical resources to process sound from the ear they listened to for most
of their lives, reflecting a profound reorganization of the cortical network for CI listening. At the same

3
time, we hypothesize that cortical activity underlying the pathways from the opposite and deprived ear
will be abnormal and segregated from the experienced side, in turn placing this ear at a disadvantage
for auditory processing. This will translate into poor functional outcomes, thereby highlighting the
deleterious effects of unilateral deprivation on the brain. Understanding the effects of deafness and
abnormal hearing during development is important to improve clinical care for children who are deaf
and to guide novel targeted therapies to capitalize on parts of the auditory network that are activated
in CI users to promote optimal CI hearing for children/adolescents who are deaf.

4
2. Chapter Two – General introduction and background
2.1 Cochlear implants establish hearing in children who are deaf
Approximately 1.2 to 5.7 per 1,000 children in developed countries suffer from a permanent
hearing loss (Yoshinaga-Itano, Sedey et al. 1998; Mohr, Feldman et al. 2000; Fortnum, Summerfield et
al. 2001; Smith, Bale Jr et al. 2005; Morton and Nance 2006; Mehra, Eavey et al. 2009; Qi and Mitchell
2012). An estimated 1.33 per 1,000 children are born with a hearing loss that is both bilateral and at
least moderate in severity (thresholds >40dB HL) (Morton and Nance 2006; Mehra, Eavey et al. 2009)
with 4 per 10,000 babies born profoundly deaf each year (Smith, Bale Jr et al. 2005). The prevalence
of hearing impairments increases with age as progressive and acquired hearing losses develop during
childhood and adolescence (Mehra, Eavey et al. 2009; Shargorodsky, Curhan et al. 2010), and reaches
a rate of 2.7 per 1,000 children before the age of five years and 3.5 per 1,000 during adolescence
(Morton and Nance 2006). Such hearing impairments in childhood have detrimental consequences
for the development of hearing and spoken language acquisition (Yoshinaga-Itano, Sedey et al. 1998;
Yoshinaga-Itano, Coulter et al. 2000), educational and professional opportunities (Yoshinaga-Itano,
Coulter et al. 2001; Yoshinaga-Itano 2003; Yoshinaga‐Itano 2003; Qi and Mitchell 2012), and
psychosocial challenges related to low self-esteem and decreased quality of life – all of which lead to
difficulties with social and cultural integration (Bess, Dodd-Murphy et al. 1998) with a high cost to
society (Mohr, Feldman et al. 2000). Children who were identified with a hearing loss early (i.e., by 2
to 3 months) and received appropriate auditory intervention by 6 months of age had significantly better
receptive and expressive language skills (Yoshinaga-Itano, Sedey et al. 1996; Yoshinaga-Itano, Sedey
et al. 1998; Moeller 2000) as well as improved social and emotional development (Morton and Nance
2006) compared to those children who were diagnosed later in life, regardless of the degree of hearing
loss (Mah-rya and Yoshinaga-Itano 1995; Yoshinaga-Itano, Sedey et al. 1996).
For children with severe to profound sensorineural hearing loss who benefit little or not at all
from their hearing aids, hearing can only be established with the use of CIs. The CI is surgically
implanted into the cochlea and allows children who are deaf to develop oral speech and language. The
CI was made available to children in North America in the early 1990s and works by stimulating the
auditory pathways with electrical pulses. An illustration of the external and internal components of
the CI device in Figure 2.1 shows an array of 20 active electrodes (and two external ground electrodes),
which is surgically placed in the scala tympani of the cochlea. These electrodes deliver electrical pulses

5
to stimulate the auditory nerve. This electrical input is then carried by the auditory pathways to regions
of the auditory cortex and allows the user to perceive these signals as sounds. External equipment,
the behind-the-ear speech processor, takes in sounds through the microphone, converts them into a
digital signal, extracts frequency and intensity information from the temporal envelope of the acoustic
signal and sends instructions to an internal device through a transcutaneous radio frequency
transmitting coil.
The speech processor is a key component of the CI system (Zeng 2004; Zeng, Rebscher et al.
2008). It processes and encodes the incoming signal by analyzing its frequency and intensity, and
band-pass filters the sound into 22 independent channels, each corresponding to one implanted
electrode. It also contains a directional microphone that adapts to the sounds in the environment,
wireless capabilities to stream sounds from external commercial devices (i.e., TV, phone, MP3 player,
etc…) and memory which stores patient-specific information including dynamic range settings. The
dynamic range, which is the difference between the lower threshold (T) (minimum) and upper comfort
(C) (maximum loudness) level that a CI delivers to the auditory system is programmed for a number
of electrodes by an audiologist using proprietary programming software from the CI manufacturer
(Vaerenberg, Smits et al. 2014). Instructions regarding the current level and timing of stimulation are
delivered by an external transmitter to an internal receiver-stimulator through a radio frequency signal.
These components are held in place on either side of the scalp by a magnet that is placed underneath
the skin during surgery. The internal receiver-stimulator then sends this information to the electrodes
which are organized to mimic the normal cochlea; high frequency sounds are allocated to basal
electrodes with lower frequencies being allocated to progressively more apical electrodes. The
electrodes stimulate surviving spiral ganglion nerve fibers, which in turn, carry auditory activity through
the auditory pathways to various parts of the cortex. In this way, the child receives an electrical
representation of the acoustic world and learns to understand sounds including speech and language.

6

7
Unfortunately, the current CI only codes the temporal envelope of the acoustic signal. This
means that the fine-structure of sound that is important for discriminating between pitches, is lost in
CI listening (Zeng 2004; Zeng, Rebscher et al. 2008). CIs also have a much narrower bandwidth than
the normal hearing cochlea (i.e., 20 channels in the CI compared to activity provided by over 3,000
inner hair cells in the normal cochlea which synapse to ~30,000 afferent auditory nerve fibers). The
frequency resolution of CI hearing is therefore significantly poorer than normal (Rubinstein 2004)
particularly given that electrical stimulation excites a much broader/unfocused population of neurons
than acoustic stimulation (Drennan and Rubinstein 2008; Zeng, Rebscher et al. 2008). The limited
spectral selectivity of CI stimulation makes understanding tonal languages and music perception
particularly difficult. Indeed, since musical melodies are composed of complex tones, CI users typically
do not enjoy music, as musical appreciation relies on the ability to extract fine-structure information
from the signal (Drennan and Rubinstein 2008).
Figure 2-1: Illustration of the external and internal components of a CI device in a diagram of the ear.
Figure reproduced with permission from Papsin and Gordon (2007).
The external component houses a microphone, which picks up sounds from the environment and
sends it to the speech processor. The processor converts this acoustic input into an electric signal, and
analyzes the frequency and intensity information of the sound. The digitized signal is then sent by the
transmitting coil to the receiver-stimulator in the internal component via a radio frequency signal,
which in turn sends this information to an array of electrodes that is implanted into the cochlea. The
external transmitter coil and internal receiver-stimulator are connected over the skin flap by a pair of
magnets. The implanted electrodes are organized to mimic the tonotopic arrangement of the cochlea;
high frequencies are allocated to basal electrodes (i.e., electrode 3 in the Cochlear Nucleus device is
roughly equivalent to 1,500Hz) while low frequencies are allocated to more basal electrodes (i.e.,
electrode 20 in the same device corresponds to ~500Hz). The cross section of the cochlea on the top
right corner shows the array of electrodes that are surgically implanted into the scala tympani.
Electrical pulses, which are delivered to the electrode array in the form of biphasic pulses, stimulate
surviving spiral ganglion nerve fibers. This input, in turn, is carried along the ascending auditory system
to various regions of the cortex and allows the listener to perceive sounds.

8
In addition, current devices use a monopolar mode of stimulation, which further hinders the
spatial representation of the implant signal. Monopolar stimulation result in larger spread of electrical
activation/splattering and neural excitation. This occurs because the electric current from activated
electrodes in the cochlea is spread over a larger distance to the return ground electrode, which is placed
outside the cochlea (in bipolar stimulation which is only used in older devices such as the N22 implant,
each active electrode is paired with its own closely situated ground electrode) (Boëx, de Balthasar et al.
2003). A single electrode cannot provide the level of tuning and timing that resembles the normal
pattern of neural activity (Zeng, Rebscher et al. 2008). Thus, activation of a reduced number of
auditory neurons to a potentially abnormal auditory pathway with abnormal auditory input imposes
additional limitations to CI listening. This means that although CIs establish hearing to individuals
who are deaf, listening to sounds with a CI is far from normal.
Despite these limitations, children amaze us by achieving excellent listening and oral
communication abilities, learn and integrate in mainstream environments and communicate effectively.
Chronic auditory stimulation with a unilateral implant promotes development in the auditory
brainstem (Gordon, Papsin et al. 2003; Gordon, Papsin et al. 2006; Gordon, Papsin et al. 2007;
Gordon, Valero et al. 2007; Gordon, Valero et al. 2008; Gordon, Salloum et al. 2012) and thalamo-
cortex (Ponton, Don et al. 1996; Eggermont, Ponton et al. 1997; Ponton and Eggermont 2001; Sharma,
Dorman et al. 2002; Sharma, Dorman et al. 2002; Eggermont and Ponton 2003; Gordon, Papsin et al.
2005; Sharma, Dorman et al. 2005; Sharma and Dorman 2006; Gilley, Sharma et al. 2008; Gordon,
Tanaka et al. 2008; Sharma, Nash et al. 2009; Gordon, Wong et al. 2010; Gordon, Tanaka et al. 2011;
Kral and Sharma 2012; Gordon, Wong et al. 2013) in children who are deaf from infancy. Nonetheless,
performance on auditory processing tasks remains below that of normal hearing listeners even after
years of CI experience, especially in challenging listening situations (Gordon and Papsin 2009).
Children using CIs to hear still require extensive therapy to achieve optimal communication outcomes
and their hearing deteriorates significantly in noise and reverberant environments (Basura et al., 2009;
Gordon and Papsin, 2009a, 2009b; Papsin and Gordon, 2008). Furthermore, we are also finding that
children with CIs use compensatory multi-sensory strategies more effectively than their peers with
normal hearing to facilitate spoken language comprehension and complex auditory processing, such
as understanding subtle emotional cues in speech (Hopyan-Misakyan, Gordon et al. 2009). These
functional differences could be underpinned by strengthening of sensory input (most notably, vision)
into auditory cortices in deaf individuals (Giraud, Price et al. 2001; Dehmel, Cui et al. 2008) due to

9
bilateral deafness prior to cochlear implantation, unilateral deprivation afterwards, and the altered
representation of sound delivered by this unique device.
2.2 Normal hearing requires intact auditory structures and functions
In order to understand the effects of deafness and cochlear implantation on the auditory
system, we must first understand how normal hearing works and develops. Normal hearing requires
that structure and function in all parts of the auditory pathways be intact. Hearing loss of any kind,
whether it be temporary or permanent, conductive or sensorineural, congenital or acquired, stable or
progressive will disrupt the structure and/or function of the auditory system and may ultimately affect
the trajectory of auditory development in children.
Figure 2.2 shows the ascending pathways of the normal auditory system from cochlea to
cortex. Acoustic sound waves are collected by the pinna and travel into the ear canal to cause
vibrations of the tympanic membrane and ossicular chain in the middle ear cavity. Mechanical
vibrations of the stapes footplate against the oval window of the cochlea cause fluid
displacement/waves in the cochlea. The cochlea is a snail-shaped organ which consists of 2.5 turns
and is approximately 35 mm long in humans (Von Békésy and Wever 1960; Pujol and Hilding 1973;
Nadol Jr 1988; Pujol, Lavigne-rebillard et al. 1991). The spiral turns of the cochlea are divided
transversely into three fluid-filled spaces: the scala vestibuli which is anatomically connected to the
oval window, the scala media which houses the organ of corti, and the scala tympani, which is
connected to the round window. The scala media is separated from the scala vestibuli by Reissner’s
membrane and from the scala tympani by the Basilar membrane. The Basilar membrane is tonopically
organized. Its geometry, which is narrower and stiffer at the base (near the oval window) and wider
and flexible at the apical end, allows receptor cells arranged at different places along the basilar
membrane to respond to different frequencies (Von Békésy and Wever 1960; Lim 1980; Lim 1986;
Nadol Jr 1988; Gopen, Rosowski et al. 1997).
Vibrations of the ossicles (i.e., stapes against the oval window) in the middle ear causes waves
of perylimphatic fluid in the scala vestibuli, which in turn drives movement of endolymph in the
cochlear duct (i.e., scala media) and displacement of the basilar membrane into a sheering motion.
Displacement of the basilar membrane stimulates the hair cell receptors in the organ of corti.
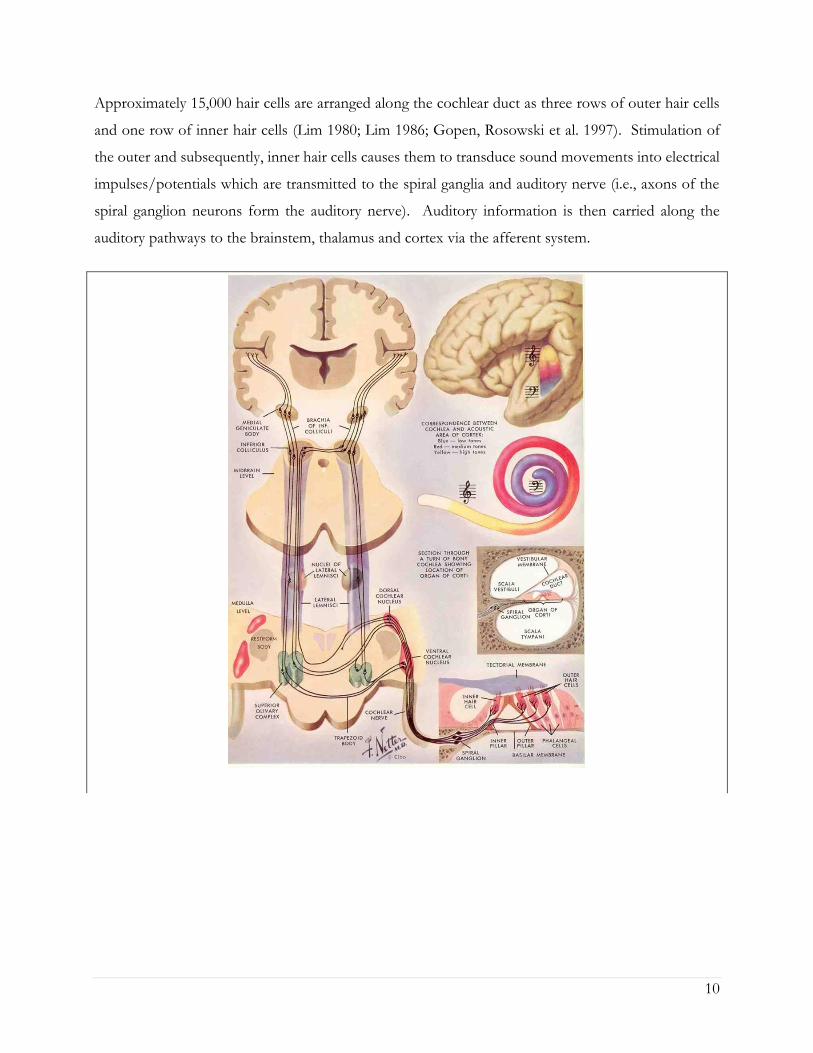
10
Approximately 15,000 hair cells are arranged along the cochlear duct as three rows of outer hair cells
and one row of inner hair cells (Lim 1980; Lim 1986; Gopen, Rosowski et al. 1997). Stimulation of
the outer and subsequently, inner hair cells causes them to transduce sound movements into electrical
impulses/potentials which are transmitted to the spiral ganglia and auditory nerve (i.e., axons of the
spiral ganglion neurons form the auditory nerve). Auditory information is then carried along the
auditory pathways to the brainstem, thalamus and cortex via the afferent system.

11
Axons from spiral ganglion neurons enter the brainstem ipsilaterally from the endbulb of Held
in the cochlear nucleus. The cochlear nucleus, whose regions retain the tonotopic organization of the
cochlea, is divided into the dorsal cochlear nucleus, the anteroventral cochlear nucleus (AVCN), and
the posteroventral cochlear nucleus. Signals travel from the cochlear nucleus to both the ipsilateral
and contralateral superior olivary complex (SOC), which form the first relay for binaural processing.
Binaural processing refers to the ability of the auditory system to process and integrate auditory
information by comparing subtle differences in level and timing of sounds reaching the two ears. Inter-
aural level differences are mediated by the head shadow effect, which refers to a mechanism of the
head to partially attenuate sounds coming from one side of head while at the same time improving the
signal-to-ratio on the listening side. As a rule of thumb, only sounds with wavelengths that are shorter
than the size of the head can be reflected. In humans, this typically corresponds to frequencies higher
Figure 2-2: Diagram of the ascending auditory pathway. Figure reproduced with permission from
(Netter 2010).
Diagram of the cochlea and cortex in the top right indicate the tonotopic distribution of activity. High
frequencies are located towards the base of the cochlea and medially in the primary auditory cortex,
whereas low frequency information is distributed on the apical end and towards the lateral end of
Heschl’s gyrus. Transverse cross-section of the cochlea below shows the three fluid spaces of the
cochlea: the scala vestibuli which is connected to the oval window at the top, the scala media which
houses the organ of corti in the middle and the scala tympani which is connected to the round window
at the bottom. The organ of corti is shown in the bottom right. Sound vibrations which are picked
up by the outer ear and causes vibrations of the middle ear ossicles against the oval window create
fluid waves in the cochlea. These movements causes the basilar membrane to move in a shearing
motion and results in the stimulation of outer hair cells and subsequent depolarization of inner hair
cells which in turn release neurotransmitters into a spiral ganglion nerve terminal. Auditory
information is then carried to the ipsilateral auditory nerve, travels through the medial nucleus of the
trapezoid body, project onto the contralateral superior olivary complex, and ascends in this way
through the brainstem, midbrain, medial geniculate body in the thalamus to the auditory cortex in
Hesch’s gyrus in the temporal lobe.

12
than ~1,500Hz. The lateral superior olive (LSO) neurons are the primary site for processing such level
cues between both ears. The LSO is innervated by glutamatergic excitatory input from spherical bushy
cells of the ipsilateral AVCN and inhibitory input from the ipsilateral medial nucleus of the trapezoid
body which receive excitatory input from globular bushy cells of the contralateral AVCN (Grothe,
Pecka et al. 2010). On the other hand, the medial superior olive (MSO) codes for differences in timing
between the ears. Inter-aural timing differences are also mediated in part by the head shadow effect
and relies on differences in phase coding of low frequency sounds (~<1,500Hz) arriving at each ear.
The MSO is innervated by excitatory input from spherical busy cells in both the ipsilateral and
contralateral AVCN as well as binaural inhibitory input from the medial nucleus of the trapezoid body
(Grothe, Pecka et al. 2010).
It is at this stage of the auditory system that the primary (also known as lemniscal) and non-
primary (extra-lemniscal) auditory pathways diverge. A conceptual diagram of afferent projections
from each system is shown in Figure 2.3. In the primary or lemnical pathway, shown by the red boxes
in Figure 2.3, auditory signals from the SOC project to the central nucleus of the inferior colliculus in
the midbrain through the lateral lemniscal fibre tract. Input then ascends to the ventral medial
geniculate body in the thalamus and terminates on middle layers of the primary auditory cortex in
Heshl’s gyrus in the temporal lobe. Signals are then relayed to belt and parabelt association auditory
regions for further processing (Hu, Senatorov et al. 1994). Primary pathway responses to sound are
specific to auditory input, fast, sharply tuned and exhibit great fidelity for temporal and fine-frequency
tuning (Kraus, Smith et al. 1988; McGee, Kraus et al. 1991; Kraus, McGee et al. 1992; LeDoux 1992;
McGee, Kraus et al. 1992; Hu, Senatorov et al. 1994; Kraus, McGee et al. 1994; Moller and Rollins
2002; Hu 2003).
On the other hand, the non-primary or extra-lemniscal auditory pathway receives ascending
activity directly from the SOC to the central nucleus of the inferior colliculus, bypassing the lateral
lemnicus. The ascending trajectory of this pathway is shown by the green boxes in Figure 2.3. Input
then projects to the external nucleus and dorsal cortex of the inferior colliculus. Neurons in these
areas of the inferior colliculus have been shown to respond to both auditory and somatosensory input
(Moller and Rollins 2002). This activity innervates a group of nuclei in the posterior thalamus (Jones
1985) including the medial and caudal portions of the medial geniculate body, the posterior
intralaminar nucleus, the suprageniculate nucleus and the lateral posterior nucleus (Hu 2003). These
neurons all respond to multi-sensory input including auditory, visual and somatosensory information

13
with the latter two, the suprageniculate nucleus and the lateral posterior nucleus, being primarily
devoted to visual processing (Hu 2003). Of note, neurons of this pathway have direct and reciprocal
connections to the mesencephalic reticular formation. This means that activity in this pathway is
particularly sensitive to effects of arousal/sleep (Kraus, Smith et al. 1988; McGee, Kraus et al. 1991;
Kraus, McGee et al. 1992; LeDoux 1992; McGee, Kraus et al. 1992; Hu, Senatorov et al. 1994; Moller
and Rollins 2002; Hu 2003). At the same time, they also send efferent projections to several limbic
structures (i.e, amygdala, insular temporal lobe and striatum), the frontal cortex, the parieto-temporal
regions and the cerebellum (LeDoux 1992; Hu 2003) as they ascend to both the primary and
association areas of the auditory cortex. Because of these connections, neurons of the non-primary
pathway are influenced by changes in arousal/sleep and attention (Kraus, Smith et al. 1988; Kraus,
McGee et al. 1989; McGee, Kraus et al. 1991; Kraus, McGee et al. 1992; McGee, Kraus et al. 1992;
Kraus and McGee 1993; McGee, Kraus et al. 1993; Kraus, McGee et al. 1994), are particularly sensitive
to multi-modal input (Moller and Rollins 2002), are involved in emotional learning and memory
formation of behaviourally relevant sensory input (LeDoux 1992; Hu, Senatorov et al. 1994; Hu 2003),
and are more likely to be affected by plasticity-related changes (i.e., to demonstrate plasticity) (Kraus,
McGee et al. 1994). Of note however, despite these differences in auditory function between the
primary and non-primary auditory pathways, the distribution of inhibitory-excitatory potentials and
the sensory receptive fields of neurons in both pathways are regulated in a similar way (Hu 2003).

14

15
2.3 Milestones in auditory development
Early exposure to sound shapes auditory development. Normal human auditory development
begins in utero. The ability to hear begins as early as 25 to 29 weeks gestational age. Responses to
sounds, voices (particularly the mother’s voice) and music has been observed by as early as 32 weeks
gestational age (Pujol, Lavigne-rebillard et al. 1991). Exposure to these sounds in the last 10 to 12
weeks of fetal life are necessary for fine tuning of cochlear hair cells and their connections to spiral
ganglion neurons and cochlear nuclei (Graven and Browne 2008). During this time, listening to
meaningful sounds allows babies to form memory circuits in non-primary auditory and language areas
of the cortex which are connected to the limbic system, and create emotional memories that are
associated with speech and music (Graven and Browne 2008). Indeed, newborns show preference for
their mother’s voice at birth (DeCasper and Fifer 1980; DeCasper and Spence 1986), are capable of
discriminating sounds of their native language by 2 days of age (Bosch and Sebastián-Gallés 1997;
Bosch and Sebastián-Gallés 2001), and interestingly, infants younger than 6 months of age are able to
discriminate phonemic contrasts in nearly all languages (Trehub, 1976).
Early auditory discrimination is associated with early maturation of the cochlea, auditory nerve
Figure 2-3: Conceptual diagram of ascending projections of the primary auditory pathway (red boxes)
and the non-primary auditory pathways (green boxes).
Peripherals projections from the cochlea to the contralateral superior olivary complex are similar
between the two pathways. From there, activity of the primary pathway ascends through the lateral
lemniscus tract to the central nucleus of the inferior colliculus, ventral medial geniculate body in the
thalamus and terminates in the primary auditory cortex. The primary pathway serves as the most direct
route to the primary auditory cortex. On the other hand, afferent projections in the non-primary
pathway ascend from the superior olivary complex to both the external nucleus and dorsal portions of
the inferior colliculus and project to the caudal and medial regions of the medial geniculate body.
Activity then innervates both areas of the primary and association auditory cortex. The non-primary
pathway shares direct connections with the limbic system and reticular activating system, and sends
efferent input to the frontal cortex, parietotemporal regions and the cerebellum.

16
fibers, brainstem pathways, reticular activating system and cortical layer I (Moore and Guan 2001;
Eggermont and Moore 2012). Maturation of these processes is largely complete at birth but continues
to be refined until the second to third year of life (Moore and Guan 2001) as developmental increases
in myelin density, dendritic arborization and synaptic modifications occur (Moore and Guan 2001;
Moore 2002). These changes allow for more efficient synapses, increased synchrony and rapid
transmission/conduction velocity in the developing pathways. Functionally, refinement of these
processes has been associated with the ability to attribute meaning to sounds (Eggermont and Moore
2012). At this young age (i.e., 2 to 3 years), the auditory brainstem response (which reflects activity in
the auditory nerve and brainstem pathways) (Salamy and McKean 1976; Starr, Amlie et al. 1977; Jerger
and Hall 1980; Salamy 1984), the middle latency response (which reflects subcortical auditory activity
generated in the thalamus and the primary auditory cortex) (Fifer and Sierra-Irizarry 1988; Frizzo,
Funayama et al. 2007) and the late component P2 of the cortical evoked response (which reflects
auditory activity driven from association auditory areas and the reticular activating system of the non-
primary pathways) (Wunderlich and Cone-Wesson 2006; Wunderlich, Cone-Wesson et al. 2006) are
fully mature. Components of these various responses continue to change with age and maturation,
especially for the cortical response. Table 2.1, reproduced from Eggermont and Moore (2012), shows
a beautiful summary of the structural, electrophysiological and behavioural/functional correlates of
these responses in different stages of development from infancy to adolescence (>12 years of age).
The auditory cortex has the longest developmental time-course. Myelination of thalamo-
cortical projections in deep cortical layers begins around 1 year of age and continues until 4 years
(Moore and Guan 2001; Moore 2002). Superficial layers however, which represent cortico-cortical
connections, take much longer to mature. Dendritic cortical neurofilament proteins which are
necessary for axonal development (i.e., formation of axonal cytoskeleton) only begin to be expressed
in superficial cortical layers around 5 years of age and continue to change until 11 to 12 years when
they are considered mature (Moore and Guan 2001). The maturational time-course of these cortical
structures coincides with the emergence of adult-like polyphasic cortical evoked response waveforms
(peaks P1, N1 and P2) (Albrecht, Suchodoletz et al. 2000; Ponton, Eggermont et al. 2000; Ponton,
Eggermont et al. 2002) and with adult-like development of complex auditory processing skills (such as
processing masked or degraded speech, and listening to sounds in noise or reverberant environments)
(Ponton, Eggermont et al. 2000; Ponton and Eggermont 2001; Ponton, Eggermont et al. 2002;
Eggermont and Ponton 2003; Eggermont and Moore 2012). This indicates that maturation of

17
structure and function in the auditory brain continues well into adolescence and only becomes mature
after the second decade of life (Albrecht, Suchodoletz et al. 2000). Of interest, the human visual
auditory cortex also requires a decade to mature, with the superficial layers only becoming adult-like
around 15 years of age (Moore and Guan 2001), suggesting that adolescence marks an important
maturational period in brain development.
While the primary and non-primary auditory pathways follow a parallel maturational time-
course, each system seems to dominate auditory processing during different times in development.
This has been examined by assessing the contribution of the reticular activating system (Kraus, McGee
et al. 1989; Kraus, McGee et al. 1992; McGee, Kraus et al. 1992; McGee, Kraus et al. 1993) and the
influence of multi-sensory stimulation (Moller and Rollins 2002) on auditory processing in children
and adults. Kraus and colleagues indicated that the middle latency response was clearly detectable
during wakefulness and light stages of sleep in sleeping children but became suppressed as they
transitioned into deeper stages of sleep. Sleep did not affect these responses in adults however, and
their detectability increased with age (Kraus, McGee et al. 1989; Kraus, McGee et al. 1992; McGee,
Kraus et al. 1993). This indicates that the reticular activating system (which regulates states of arousal
and controls the sleep/wake cycles of sleep), which shares reciprocal connections with the non-primary
pathways, mediates auditory activity in children but not in adults. This is consistent with results from
Moller and Rollins who showed that children under 9 years of age, but not adults, perceived sounds as
louder when they received simultaneous auditory and somatosensory stimulation (Moller and Rollins
2002). These findings suggest that the non-primary pathways dominate auditory processing in the
immature brain. Over time however, as the primary pathways develop into maturation around 9 to 12
years of age (Albrecht, Suchodoletz et al. 2000; Ponton, Eggermont et al. 2000; Ponton, Eggermont et
al. 2002), involvement of the non-primary pathways for auditory processing decreases and
contributions from the primary system increases (Kraus, McGee et al. 1989; Kraus, McGee et al. 1992;
McGee, Kraus et al. 1993; Moller and Rollins 2002).
As cortical pathways in the brain mature, so does cortical specialization of each hemisphere.
Adolescents and adults develop brains that are structurally and functionally highly specialized
(Davidson 1984; Zatorre and Belin 2001; Le Grand, Mondloch et al. 2003; Toga and Thompson 2003;
Rivera, Reiss et al. 2005; Gotts, Jo et al. 2013). In the auditory system, a relative specialization for
temporal resolution relevant to speech processing has been observed in the left auditory cortex,
whereas association auditory areas of the right hemisphere are biased for processing spectral patterns

18
of sounds such as tones, melodies and music (Zatorre and Belin 2001; Zatorre, Belin et al. 2002;
Schönwiesner, Rübsamen et al. 2005; Jamison, Watkins et al. 2006). Much of this specialization in the
brain is shaped by sensory experience and perceptual skills that are acquired during development
(Penhune, Zatorre et al. 1996; Johnson 2001; Johnson 2011) and is related to normal neuroanatomical
maturation of the cortex which begin in early life and is mostly complete by adolescence (Giedd, Snell
et al. 1996; Giedd, Blumenthal et al. 1999; Johnson 2001; Blakemore and Choudhury 2006; Lenroot
and Giedd 2006; Johnson 2011; Lebel and Beaulieu 2011; Blakemore 2012; Lohmann and Kessels
2014).
Normal exposure to sounds in utero, during infancy and during childhood is thus important
to the normal development and maturation of the auditory pathways. Indeed, certain auditory
functions such as learning to speak different languages (Johnson and Newport 1989; Flege, Yeni-
Komshian et al. 1999; Birdsong and Molis 2001) or play musical instruments (Hyde, Lerch et al. 2009;
Steele, Bailey et al. 2013) are optimal when learned early in life, when the brain is maximally plastic and
best able to adapt to the environment (Johnson and Newport 1989; Flege, Yeni-Komshian et al. 1999;
Birdsong and Molis 2001; Munte, Altenmuller et al. 2002; Trainor, Shahin et al. 2003; Fujioka, Ross et
al. 2006; Hyde, Lerch et al. 2009; Kraus and Chandrasekaran 2010; Steele, Bailey et al. 2013).
Deprivation of auditory input during initials stages of development may arrest or delay/desynchronize
maturational processes of myelination, dendritic arborization, synaptogenesis, conduction velocities,
etc… – all of which are necessary for normal organization and function in the auditory system. In
addition, since mechanisms of plasticity decrease with increasing age (Lohmann and Kessels 2014),
absence of auditory input during critical periods in development may hinder thalamo-cortical and
cortico-cortical connections (Ponton and Eggermont 2001; Eggermont and Ponton 2003). This may
in turn drive long-lasting abnormalities and reorganization in the central auditory system (Finney, Fine
et al. 2001; Lee, Lee et al. 2001; Bavelier and Neville 2002).

19
2.4 Auditory experience shapes auditory development
The development of the auditory system is shaped by both intrinsic (experience-independent)
and extrinsic (experience-dependent) processes. Activity independent processes such as the growth
of nerve cells and formation of rudimentary tonotopic projections from spiral ganglion neurons (Heid,
Hartmann et al. 1998) to the cochlear nucleus, superior olivary complex, lateral lemniscus, inferior
colliculus (Heid, Jähn-Siebert et al. 1997) and primary auditory cortex (Hartmann, Shepherd et al. 1997)
Table 2-1: Milestones of structural, electrophysiological and behavioural maturation of activity in the
central auditory pathway. Reproduced with permission from Eggermont and Moore (2012)

20
were all preserved in the congenitally deaf auditory pathways of adult cats, despite complete and long-
term absence of auditory input. Rudimentary binaural sensitivity of primary auditory neurons in the
cortex was also maintained (Kral 2007; Kral and Eggermont 2007). This suggests that rudimentary
response properties and feature detection in the auditory cortex are genetically pre-determined and not
influenced by sensory experience (Friauf and Lohmann 1999). Nonetheless, maturation of neuronal
circuits in the brain are shaped and influenced by sensory stimulation/experience during critical
periods in development. “Environmental sounds (e.g., speech) will modulate and increase auditory
nerve activity, conductive hearing loss (e.g., otosclerosis, otitis media) will decrease and desynchronize
nerve activity, and sensorineural loss will reduce and broaden, or abolish nerve activity from damaged
parts of the cochlea” (p.147, Moore 2002).
Hebbian theory of activity-dependent development suggests that successful and repeated
communication/synapses between pre- and post-synaptic neurons strengthens synaptic efficiency
(Hebb 1949) and underlies mechanisms of synaptogenesis, learning, memory and development in the
brain (Abbott and Nelson 2000; Song, Miller et al. 2000). In early development, longer post-synaptic
excitatory potentials (increased NMDA-mediated glutamatergic receptors) support higher levels of
plasticity. With development, the duration of synaptic transmissions decreases (due to changing
proportions of NMDA and AMPA (strength of AMPA-mediated transmission increases and
composition of NMDA changes)) (van Zundert, Yoshii et al. 2004; Elias, Apostolides et al. 2008) and
inhibition (GABA receptors) increases (Gao, Wormington et al. 2000). These developmental changes
in receptor currents promote dendritic elaboration (Cline 2001), are involved with mechanisms of
synaptogenesis in the primary auditory cortex (Huttenlocher and Dabholkar 1997) and support axonal
myelination – all of which serve to generate faster and more efficient synaptic transmission to promote
plasticity during sensitive periods in development (Abbott and Nelson 2000; Song, Miller et al. 2000).
Without stimulation, these receptors do not develop normally. Essential synapses do not get formed,
inappropriate ones may be pruned and normal growth of dendritic arbors may be interrupted (Kral
2007; Kral and Eggermont 2007). Normal auditory activity is thus important to promote activity-
dependent plasticity in the auditory pathways to promote development and maturation of circuits in
the auditory brain, fine-tune the auditory system and allow the brain to adapt to the environment
(Rauschecker 1999).
Interestingly, providing abnormal input to the developing system will still promote plasticity in
the auditory pathways but will impair the structure and function of auditory brain circuits. This has

21
been shown both in cases of sensorineural hearing loss as well as conductive impairments. For
example, sensorineural hearing loss caused from a cochlear lesion induced with ototoxic
aminoglycoside poisoning indicated reduced frequency tuning and frequency selectivity of cortical
neurons in kittens one year after the loss. This frequency deterioration was observed even in areas of
the cochlea which were still intact (Harrison, Stanton et al. 1995). Conductive hearing losses similarly
impaired cortical development. Children suffering from persistent and recurrent otitis media with
effusion, a common intermittent middle ear condition which attenuates and distorts sounds, had
impaired abilities for using binaural cues of timing and level differences between both ears (which in
turn compromised binaural hearing). This deficit was particularly pronounced when the hearing loss
was asymmetric (Moore 2002; Moore, Hartley et al. 2003). Long-term consequences of conductive
hearing loss on auditory processing were also shown in animal models with prolonged bilateral ear
plugging, and similarly indicated that impoverished auditory stimulation resulted in impaired bilateral
auditory function, in particular, poor sound localization and binaural unmasking abilities (Moore 2002;
Moore, Hartley et al. 2003). Abnormal stimulation of the auditory system with a distorted signal thus
has deleterious effects on auditory development and function.
This is a particular concern with CI listening because the implanted electrode array cannot
mimic stimulation provided by a normal cochlea. Specifically, CI processing will extract the envelope
of a speech signal and will transmit temporal information with high fidelity; but because the electrode
array consists of only 22 electrodes, spectral coding is not well represented. This is particularly true
with monopolar stimulation modes, as the CI will stimulate a broader cochlear region. In turn, this
results in broader spread of current excitation, which further limits spectral resolution in CI listeners.
This is unfortunate, given that tonotopic organization is one of the activity-independent mechanisms
that remains preserved in the deaf auditory system (Hartmann, Shepherd et al. 1997; Heid, Jähn-Siebert
et al. 1997; Heid, Hartmann et al. 1998). In addition, electrical pulses delivered by a CI will stimulate
a smaller number of surviving spiral ganglion neurons (because all do not survive cochlear damage
imposed by the hearing loss), and will do so with higher synchrony than normal acoustic hearing
(Rubinstein and Hong 2003). Moreover, the dynamic range of electric hearing is also reduced
compared to acoustic hearing, which means that CI listeners may not have a normal representation of
loudness growth (Zeng, Rebscher et al. 2008). Hearing with a CI is thus not normal. The distorted
signal delivered by this unique device might thus promote maladaptive plasticity in the developing
auditory system, which in turn could hinder expressive and receptive language skills, lead to decreased

22
performance in school and inflict social difficulties.
2.5 Bilateral deafness in childhood drives abnormal reorganization in the
brain
Prior to cochlear implantation, the absence of auditory input from bilateral deafness in early
life drives widespread physiological deficiencies and degeneration in the auditory system. Figure 2.4
summarizes some cortical effects of deafness observed from congenitally deaf adults and animal
models of deafness (i.e., cats) (Kral and Sharma 2012). As shown in A the number of non-responsive
units in the auditory cortex increased from 10% in normal hearing cats (blue bar) to 45% in the
congenitally deaf animal (red bar). The maximum firing rate (B) and dynamic range of responsive
units in the cortex (C) were reduced. Panel D shows that while rudimentary cochleotopy was preserved
in the deaf animal, cochleotopic resolution was weaker compared to the hearing cat. Current
thresholds for generating electrical potential in the auditory system were significantly lower in the
cortex (E), suggesting hypersensitivity to auditory input in the cortical pathways. By contrast,
sensitivity to interaural timing differences in the inferior colliculus and the cortex was decreased (F).
Bilateral deafness also altered the normal organization of activity in the cortex, as shown by the
development of symmetrical aural preference in the congenitally deaf animal in G, indicating a loss of
contralaterality in the auditory system, compared to normal hearing animals who showed preference
for the contralateral ear.
23

24
In addition to these changes, altered synaptic structures in auditory nerve endings particularly
at spherical and globular bushy cells in the endbulb of Held (Ryugo, Rosenbaum et al. 1998; O'Neil,
Limb et al. 2010), lower spiral ganglion cell count (Nadol Jr, Young et al. 1989; Nadol 1997), smaller
cochlear, vestibular and eight cranial nerve trunk diameter (Nadol Jr and Xu 1992; Nadol 1997),
shrinkage of dendrites and cell bodies (Drennan and Rubinstein 2008), asynchronous glutamate release
from hair cells to spiral ganglion neurons resulting in decreased and sometimes absent spontaneous
activity (Kral 2007) and delayed synaptic pruning (Kral et al., 2005) all also occur with bilateral deafness.
The number of synapses from the lateral lemniscus to the inferior colliculus becomes reduced (Hardie,
Martsi-McClintock et al. 1998; Nishiyama, Hardie et al. 2000) and the density of primary neurons
decreases (Ryugo, Rosenbaum et al. 1998). Bilateral deafness also leads to altered synaptic currents in
the auditory cortex. Activation of auditory cortical columns becomes desynchronized (Kral, et al.
2005), maturation of excitatory post-synaptic potentials becomes abnormal (Kral and Eggermont
2007) and development of inhibitory mechanisms becomes altered (Kral and Eggermont 2007).
Moreover, reduced synaptic activity in loop thalamo-cortical and cortico-cortical connections (Kral,
Hartmann et al. 2001) cause delayed development of supragranular layers and decreased activity in
Figure 2-4: Widespread effects of bilateral deafness in all levels of the auditory pathways. Figure
reproduced with permission from Kral and Sharma (2012).
A number of functional deficits were observed in cats with bilateral deafness compared to normal
hearing control cats. A. Bilateral deafness drives an increased proportion of non-responsive units in
the cortex shown by the red bars compared to normal hearing controls. B. Neural firing rate is
impaired and occurs over a significantly reduced dynamic range as shown in C. (blue). D.
Rudimentary cochleotopy is preserved in the deaf animal, but the resolution is much weaker and much
simpler than in the hearing animal. E. Thresholds of evoked responses are reduced in the cortex of
the deaf animal compared to the hearing animal, providing evidence of cortical hypersensitivity. Of
interest however, sensitivity to inter-aural timing cues is much reduced as shown in F. G. Analyses
of aural preference indicate a contralateral preference in the normal hearing control cat. By contrast,
aural preference is lost in the congenitally deaf animal as shown by similar responsiveness of the
auditory cortices to stimulation of the ipsilateral and contralateral ears.

25
deep infragranular cortical layers (Kral, Hartmann et al. 2001; Kral, Schroder et al. 2003; Kral 2007;
Kral and Eggermont 2007). This reduces connectivity of the primary auditory cortex with higher
cortical centers (Kral 2007; Kral and Eggermont 2007) and alters neural projections to association
auditory areas, leaving the immature and deaf auditory brain vulnerable to cross-modal recruitment by
non-auditory systems (Finney, Fine et al. 2001; Lee, Lee et al. 2001; Bavelier and Neville 2002; Bavelier,
Dye et al. 2006; Lomber, Meredith et al. 2010; Meredith and Lomber 2011).
The primary auditory cortex is spared while association areas of the auditory system become
the targets of other sensory systems (Lomber, Meredith et al. 2010; Meredith and Lomber 2011).
Recent evidence from a cat model of congenital deafness indicated that secondary and association
auditory areas, including parts of the planum temporale, all of which respond to multi-sensory input
including hearing, vision and touch (Pandya and Yeterian 1985; Giard and Peronnet 1999; Calvert,
Hansen et al. 2001; Calvert and Thesen 2004), become recruited by the visual (Finney, Fine et al. 2001;
Lee, Lee et al. 2001; Lee, Truy et al. 2007; Lomber, Meredith et al. 2010; Meredith and Lomber 2011)
and somatosensory (Levänen, Jousmäki et al. 1998; Levänen and Hamdorf 2001; Auer Jr, Bernstein et
al. 2007; Meredith and Lomber 2011) systems to perform non-auditory functions. As a consequence
of early auditory deprivation, processing of visual peripheral localization by the Posterior Auditory
Field (Lomber, Meredith et al. 2010), visual motion detection by the Dorsal Zone of the auditory
cortex (Lomber, Meredith et al. 2010) and somatosensory sensation by the Anterior Auditory Field
(Meredith and Lomber 2011) become enhanced in individuals who are deaf. These changes appear to
result from a direct competition for resources in areas which receive multi-sensory input. If governed
by principals of Hebbian processing (Hebb 1949; Abbott and Nelson 2000; Song, Miller et al. 2000),
neurons in these areas could preferentially form viable connections with non-auditory inputs to the
detriment of inputs carrying auditory information. Depending on how quickly these processes occur,
they may be impossible to reverse and could impair outcomes after cochlear implantation if auditory
input is not restored within sensitive periods in development.
Such cortical reorganization may also be attributed to anatomical changes of the auditory
cortex, occurring as a result of bilateral auditory deprivation in early life (Wong, Chabot et al. 2013;
Kok, Chabot et al. 2014). Indeed, recent findings from deaf cats have indicated that the auditory cortex
in early- or congenitally- deaf brains differs both in size and position from the hearing brain. The
volume of the auditory brain is significantly smaller in deaf cats compared to normal hearing cats and
the borders between the dorsal auditory areas and adjacent somatosensory and visual areas shifts

26
ventrally to allow for a greater representation of non-auditory input (Wong, Chabot et al. 2013). In
humans however, such volumetric changes were not found. While individuals with long-term bilateral
deafness had decreased white matter in the left superior temporal gyrus, the volume of the auditory
brain in either hemisphere was similar between individuals with deafness and normal hearing
(Emmorey, Allen et al. 2003; Shibata 2007). These differences in the cortical effects of early-onset
deafness between animal models and humans may be due to differences in the aetiology and/or onset
of deafness. Unlike children who may have had some hearing in utero, the onset of hearing in cats
occurs post-natally (Heid, Hartmann et al. 1998; Ryugo, Rosenbaum et al. 1998; Ryugo, Cahill et al.
2003). This means that congenitally deaf white cats never had access to sound, which may cause both
anatomical and physiological changes to the auditory pathways that are different from humans.
2.6 Unilateral deafness promotes abnormal changes in the auditory brain
While the effects of early-onset bilateral hearing loss on the auditory brain are well known,
fewer studies have explored the consequences of unilateral deafness on auditory development.
However, individuals with unilateral deafness are not a small population. The prevalence of unilateral
hearing (thresholds >45dB HL) is estimated to be 3 per 1,000 in school-aged children. This rate
increases to 13 per 1,000 when children with milder losses are included (Bess and Tharpe 1984; Bess
and Tharpe 1986; Bess, Dodd-Murphy et al. 1998). Bess and colleagues reported that approximately
one-third of children with unilateral hearing loss failed at least one grade in school, with almost 50%
of them needing access to resource assistance programs. Unilateral hearing impairments also affect
language development, with 38% of children suffering from reading problems, 31% have challenges
with spelling and 23% struggle with arithmetic. In addition to these education and language difficulties,
behavioural problems, low self-esteem and reduced intelligent quotient scores have all been reported
from children with unilateral hearing loss compared to those who have normal hearing in both ears
(Bess and Tharpe 1984; Bess and Tharpe 1986). These deficits might be attributed to functional
changes in the auditory system occurring a result of unilateral deprivation.
Several studies have demonstrated that unilateral hearing loss modifies tonotopic maps in the
auditory cortex (Reale, Brugge et al. 1987; Popescu and Polley 2010), alters the balance of excitation-
inhibition (Nordeen, Killackey et al. 1983; Moore and Kowalchuk 1988; Hashisaki and Rubel 1989;

27
Mossop, Wilson et al. 2000; Vale, Juíz et al. 2004; Kotak, Fujisawa et al. 2005), and drives abnormal
cortical lateralization in the auditory brain (Vasama, Mäkelä et al. 1994; Vasama and Mäkelä 1995;
Vasama, Mäkelä et al. 1995; Vasama and Mäkelä 1997; Scheffler, Bilecen et al. 1998; Ponton, Vasama
et al. 2001; Khosla, Ponton et al. 2003; Langers, van Dijk et al. 2005; Firszt, Ulmer et al. 2006; Hine,
Thornton et al. 2008; Popescu and Polley 2010; Kral, Heid et al. 2013; Kral, Hubka et al. 2013).
Importantly, studies have shown that profound unilateral deafness shifts cortical activation patterns
towards one that is more symmetrical between both the left and right hemispheres (Vasama, Mäkelä
et al. 1994; Vasama and Mäkelä 1995; Vasama, Mäkelä et al. 1995; Vasama and Mäkelä 1997; Scheffler,
Bilecen et al. 1998; Ponton, Vasama et al. 2001; Khosla, Ponton et al. 2003; Langers, van Dijk et al.
2005; Firszt, Ulmer et al. 2006; Hine, Thornton et al. 2008). This occurs as a decrease in normal
contralateral cortical activation and an increase in ipsilateral activation relative to the hearing ear (Firszt,
Ulmer et al. 2006), with earlier ipsilateral latencies (Ponton, Vasama et al. 2001; Khosla, Ponton et al.
2003). This is particularly true when the loss is in the left ear (Scheffler, Bilecen et al. 1998; Khosla,
Ponton et al. 2003), perhaps reflecting hemispheric specialization in the auditory brain (Zatorre and
Belin 2001; Zatorre, Belin et al. 2002). In adults with post-lingual deafness, this loss of contralaterality
in the cortex occurs almost immediately following the onset of hearing loss (Vasama and Mäkelä 1995;
Vasama, Mäkelä et al. 1995). Of interest however, the middle latency response (MLR) and peak N1 of
the cortical response are the least affected in both children and adults (Vasama, Mäkelä et al. 1994;
Vasama and Mäkelä 1997; Hine, Thornton et al. 2008), which suggests that association auditory areas
are most vulnerable to effects of deafness.
Changes in the hemispheric organization of auditory activity in the cortex might be attributed
to a disruption in excitatory-inhibitory activity when input to both ears is unbalanced (Nordeen,
Killackey et al. 1983; Kitzes 1984; Reale, Brugge et al. 1987; Moore and Kowalchuk 1988; Hashisaki
and Rubel 1989; Mossop, Wilson et al. 2000; Moore and Kitzes 2004; Nordeen, Killackey et al. 2004;
Vale, Juíz et al. 2004; Takesian, Kotak et al. 2009; O'Neil, Limb et al. 2010; Popescu and Polley 2010;
Kral, Heid et al. 2013; Kral, Hubka et al. 2013). There is evidence from animal models indicating
better acoustic thresholds and increased excitatory responses in the ventral cochlear nucleus (Kil,
Hkageyama et al. 1995; Kitzes, Kageyama et al. 1995; Moore and Kitzes 2004; Illing, Kraus et al. 2005),
superior olivary complex (Kil, Hkageyama et al. 1995; Kitzes, Kageyama et al. 1995; Moore and Kitzes
2004), inferior colliculus (Nordeen, Killackey et al. 1983; Nordeen, Killackey et al. 1983; Kitzes 1984;
Moore and Kitzes 2004; Nordeen, Killackey et al. 2004), and auditory cortex (Reale, Brugge et al. 1987;

28
Kral, Heid et al. 2013; Kral, Hubka et al. 2013) of the stimulated pathways. This has been attributed
to a loss of inhibitory processes from the opposite-deaf side (Mossop, Wilson et al. 2000; Vale, Juíz et
al. 2004; Takesian, Kotak et al. 2009) that would have normally developed with bilateral hearing (Sanes
and Takács 1993; Gao, Wormington et al. 2000; Grothe 2003; Dorrn, Yuan et al. 2010; King 2010;
Sanes and Kotak 2011). Of note, this strengthening of inputs in the ascending pathways is age-
dependent, occurring only when the loss is present in early life (Nordeen, Killackey et al. 1983; Reale,
Brugge et al. 1987; Moore and Kowalchuk 1988; Nordeen, Killackey et al. 2004) and when it is
unilateral (Moore 1990). These findings thus highlight the deleterious effects of unilateral hearing loss
on the auditory brain.
2.7 Multiple effects of childhood deafness predict outcomes after cochlear
implantation
It is becoming clear that abnormal reorganization of the auditory brain does not occur
uniformly in children who are deaf and may be related to the heterogeneity in the onset and cause of
pediatric deafness (Gordon, Wong et al. 2011; Gordon, Tanaka et al. 2011). This heterogeneity means
that there can be no single animal model to predict the effects and auditory pathway consequences of
childhood deafness (Gordon, Tanaka et al. 2011). More than half of all hearing losses are caused from
a genetic aetiology and approximately one-third have an environmental cause (Morton and Nance
2006; Papsin and Gordon 2007). As shown in Figure 2.5, the most common genetic cause of deafness
is a mutation in the Gap Junction Beta-2 (GJB-2) gene, which accounts for approximately 20% of all
deafness at birth and during childhood (Morton and Nance 2006; Propst, Papsin et al. 2006). Another
30 genes have been associated with syndromic forms of hearing losses, while 50 genes are non-
syndromic (Gordon, Tanaka et al. 2011). In addition, 35% of hearing losses at birth are related to
environmental factors and infections, although this number decreases slightly by 4 years of age. This
is perhaps because newborns have not yet developed immunity to many organisms (i.e., bacteria,
viruses, fungi, parasites) and are more susceptible to infectious agents than toddlers (Gao, Wormington
et al. 2000; Mackay, Rosen et al. 2001; Bach 2002; Openshaw, Yamaguchi et al. 2004). Common
environmental causes of deafness include prematurity, head trauma, infections such a cytomegalovirus
and meningitis, and pharmacologically-induced ototoxicity including aminoglucoside antibiotics and
chemotherapy drugs (Morton and Nance 2006). The heterogeneity of childhood deafness means that

29
there could be multiple effects of hearing loss on the auditory brain, each with its own developmental
trajectory and consequences to the cochlea, brainstem, thalamo-cortical pathways and cortico-cortical
pathways. The various effects of hearing loss on the auditory system could therefore disrupt structure
and function of activity along the auditory pathways and in turn affect how an auditory prosthesis,
such as a CI, stimulates the auditory system and promotes auditory development.
Limiting the period of bilateral deafness in early life is essential to drive maturation in the
auditory pathways (Eggermont, Ponton et al. 1997; O’Donoghue 1999; Kral, Hartmann et al. 2001;
Ponton and Eggermont 2001; Sharma, Dorman et al. 2002; Sharma, Dorman et al. 2002; Sharma,
Dorman et al. 2005; Papsin and Gordon 2007; Gordon, Tanaka et al. 2008; Papsin and Gordon 2008;
Figure 2-5: Heterogeneity in the cause and onset of pediatric deafness. Figure reproduced with
permission from Morton and Nance (2006).
Incidence at birth and prevalence at 4 years of age is shown for different etiologies of deafness. Genetic
mutations are the most common causes of deafness, with GJB-2 associated deafness being the most
common etiology both at birth and during childhood. Environmental factors and infections account
for 35% of deafness at birth, but this number decreases slightly during childhood, as children develop
immunity to infectious agents.

30
Nikolopoulos, O'Donoghue et al. 2009; Gordon, Wong et al. 2011; Gordon, Jiwani et al. 2011), and
promote optimal hearing and speech and language development in children (Beadle, McKinley et al.
2005; Harrison, Gordon et al. 2005; Nicholas and Geers 2007; Geers and Sedey 2011). As shown in
Figure 2.6A and 2.6C, children who were congenitally deaf in both ears and implanted within a critical
period of 3.5 years, when the brain was maximally plastic, followed age-appropriate developmental
changes in the cortical evoked response, compared to children who were implanted at older ages
(Sharma, Dorman et al. 2002). The latency of the P1 component of the cortical evoked response
(considered a biomarker of cortical development) of early implanted children (red circles) decreased
rapidly at initial stages of CI stimulation and reached values which were similar to their normal hearing
peers. By contrast, children who received a CI after 7 years of age (shown in B and by the green
triangles in C) showed abnormal cortical waveform morphologies with persistently delayed peak
latencies despite many of years of CI experience (Sharma, Dorman et al. 2002; Sharma, Dorman et al.
2002; Sharma, Dorman et al. 2005; Sharma, Gilley et al. 2007).

31
These age cut-offs are in line with findings from Harrison and colleagues shown in Figure 2.7
who similarly indicated that children who were implanted at younger ages (<5 years) and within limited
durations of bilateral deafness, shown by the black symbols, achieved higher scores on the open-set
Phonetically Balanced Kindergarden (PBK) words test and at faster rates compared to their peers who
were implanted at older ages and after longer durations of deafness, as shown by the white symbols
(Harrison, Gordon et al. 2005).
Figure 2-6: P1 latency as a function of age at implantation. Figure adapted then reproduced with
permission from Sharma, Dorman et al. (2002).
A. Grand mean cortical response from 18 early implanted children who received a CI by 3.5 years of
age is shown at the bottom and characterized by a large amplitude peak P1, similar to their age-matched
normal hearing peers (top). B. Grand mean cortical response from 13 late implanted children
(bottom) who were implanted later than 7 years of age after long-durations of bilateral deafness has an
abnormal waveform morphology and delayed positive peak latency compared to their age-matched
normal hearing peers (top). C. The latency of peak P1 was used as an index of auditory development
in children who were congenitally deaf and used a CI to hear for an average of 3 years. The solid lines
indicate the lower and upper bound 95% confidence interval of P1 latencies recorded from age-
matched normal hearing peers. Children who were congenitally deaf and received a CI by 3.5 years
had age-appropriate cortical responses as shown by the red circles, but those who were implanted late
after 7 years of age had abnormally delayed latencies (green triangles). Children who received a CI
between 3.5 and 7 years of age had variable responses (blue crosses)

32
Many studies investigating auditory development after cochlear implantation focus on children
who are deaf in infancy, but do not examine the larger heterogeneity in etiology, onset and/or degree
of deafness. These factors may each have unique effects on auditory activity in the brain prior to
implantation. For example, bilallelic mutations of the GJB-2 gene causes deficits in the cochlea at
likely very early stages of development with possible consequences for auditory function after
implantation (Propst, Papsin et al. 2006). The GJB-2 gene normally codes for the connexin-26 protein
which creates gap junctions in the cochlea necessary for the appropriate release and maintenance of
electrochemical gradients. This in turn, generates action potentials and stimulates the auditory nerve
(Kelley, Harris et al. 1998; Cohn and Kelley 1999; Gualandi, Ravani et al. 2002). Electrophysiological
Figure 2-7: Speech perception outcomes as a function of age at implantation. Figure reproduced with
permission from Harrison, Gordon et al. (2005).
Speech perception performance of pre-lingually deaf children who received a CI at ages ranging from
1 to 15 years of age was assessed prior to cochlear implantation and at regular intervals up to 96 months
post implantation using the open-set Phonetically Balanced Kindergarden (PBK) words speech test.
Children implanted prior to 5 years of age shown by the black symbols performed better and showed
faster rates of speech perception improvements compared to their later implanted peers who had
longer durations of bilateral deafness prior to implantation (white symbols).

33
recordings of auditory evoked cortical activity at initial CI activation in children with severe GJB-2
mutations revealed that responses from the cortex were more homogenous in this cohort compared
to those children who did not have such a mutation. Auditory evoked cortical responses in children
with GJB-2 mutations were characteristic of earlier stages of cortical development, perhaps reflecting
restricted spontaneous activity in the auditory system and more limited access to sound prior to
implantation compared to their peers who did not have a GJB-2 related deafness (Gordon, Tanaka et
al. 2011). This was further supported by poorer hearing sensitivity in the low frequencies in the GJB-
2 group (Propst, Papsin et al. 2006).
The degree of residual hearing is another important predictive factor for CI outcomes.
Traditional candidacy criteria for cochlear implantation in children include a diagnosis of permanent
severe-to-profound hearing loss bilaterally with little or limited access to acoustic input through hearing
aids (Osberger, Zimmerman-Phillips et al. 2002). It has recently been reported that children who had
better hearing at 250Hz used their hearing aids for longer durations prior to receiving a CI (Hopyan,
Peretz et al. 2012). Of interest, these children performed significantly better on tests of music
perception with their implants, particularly when detecting differences in rhythm, compared to children
who did not have acoustical access to these low frequencies prior to implantation (Hopyan, Peretz et
al. 2012). Thus, there are advantages of acoustical input for auditory development which can be
capitalized upon after cochlear implantation. In general, we are learning that the cause, onset and
degree of deafness in any one child will be important to understand in order to ensure that he/she
makes the best possible use of his/her device.
2.8 Electrophysiological measures assess auditory cortical development and
map underlying auditory activity in the brain
Auditory activity and development in the brain can be measured with a variety of neuroimaging
techniques, including Positron Emission Tomography (PET), Magnetic Resonance Imaging (MRI),
Magneto-encephalography (MEG) and electroencephalography (EEG), among others. In CI users
unfortunately, only PET and EEG can be used because the CI magnet in the receiver-stimulator
component of the subcutaneous CI component renders it incompatible with the magnetic field of
other imaging modalities (Portnoy and Mattucci 1991; Teissl, Kremser et al. 1998; Teissl, Kremser et

34
al. 1999). EEG imaging provides a non-invasive method to measure changes in the brain as a function
of brain states, as well as during sensory and cognitive processing.
In brief, the brain is composed of a large number of interconnected neurons, each containing
a cell body (soma), an axon and dendritic trees. Neurons are electrically excitable cells, which
communicate with each other via synapses to transmit messages to various parts of the brain through
chemical neurotransmitters. Activation of a cell results in a change in the membrane potential (i.e.,
voltage difference between the inside and outside of the cell). Release of an excitatory
neurotransmitter, reflecting depolarization, will result in an increase of the local intracellular potential.
By contrast, release of an inhibitory neurotransmitter, reflecting hyperpolarization, will cause the cell
interior to become more negative than its resting state. Membrane voltage changes result in small
potential differences between the inside and outside of the cell. Fast voltage changes, reflecting action
potentials, propagate along the axon of pre-synaptic nerve cells. On the other hand, slow voltage
changes are produced in cell dendrites of post-synaptic cells. The flow of ions through the dendrites
forms a primary current. In order to balance the current charge across the membrane, a return current
flows in the extracellular space and forms a secondary current. It is the electric field from this
secondary current that is reflected in EEG recordings.
The electric potential produced by a single neuron is too small to be detected by EEG
electrodes at the surface of the scalp however. Recorded EEG activity thus reflects a summation of
the resulting electrical fields produced by large populations of neurons, which fire in synchrony and
have a parallel spatial orientation. When sound stimulation is delivered to the auditory system, electric
responses in the brain undergo typical fluctuations, but are related in a time-locked fashion to changes
occurring in the auditory signal. These changes, known as auditory evoked potential (AEP) responses,
can be recorded from the brainstem, midbrain or cortex using surface electrodes (i.e., an EEG set-up),
as a waveform whose morphology, peak latencies and peak amplitudes reflect
developmental/maturational changes in the brain and has been used as an index of auditory
development and plasticity in the auditory system (Picton and Hillyard 1974; Eggermont 1985;
Eggermont 1988; Eggermont 2000; Ponton, Eggermont et al. 2000; Ponton and Eggermont 2001;
Eggermont and Ponton 2002; Ponton, Eggermont et al. 2002; Eggermont 2007; Picton and Taylor
2007). Figure 2.8 shows an auditory evoked response recorded from the brainstem in CI users. A
middle latency response to auditory input, which reflects auditory activity in the midbrain (i.e., thalamo-
cortical pathways) is shown in Figure 2.9. The data in Chapter Three describes the cortical response

35
in more depth. Because EEG signals measure the primary effects of neuro-electric activity, their
temporal resolution is very high (i.e., in the order of milliseconds) and can detect brief neuronal events.
Unfortunately, their spatial resolution is particularly poor. In order to record EEG or AEP activity
from distant scalp electrodes, the electric field must travel through tissues of varying conductivities
(i.e., brain, cerebral spinal fluid, skull bone and scalp) (van den Broek, Reinders et al. 1998). This
results in a smeared representation of electric potential at the scalp.
An important caveat with recording cortical evoked responses in CI users, is that the CI device
itself produces a large electrical artifact that is recorded by scalp electrodes (Gilley, Sharma et al. 2006;
Martin 2007; Wong and Gordon 2009; Castaneda-Villa, Manuel Cornejo-Cruz et al. 2010; Friesen and
Picton 2010; Castañeda-Villa and James 2011; Viola, De Vos et al. 2012; Mc Laughlin, Lopez Valdes
et al. 2013). This artifact is generated by the radio frequency transmission of the signal from the
processor to the receiver. It is large in amplitude, time-locked to the stimulus, lasts for at least the
duration of the stimulus, and interferes with recorded response. Methods have been developed to
remove this artifact by suppressing it (used in Chapter Four) or separating it from the cortical response
using independent component analyses (ICA) (used in Chapter Five) to protect the cortical response
from contamination.
Using these artifact suppression/separation methods and cortical responses recorded at 64-
cephalic surface electrodes, we can now objectively spatially localize areas of cortical activity using
beamformer imaging techniques (Dalal, Sekihara et al. 2006; Wong and Gordon 2009) and identify the
interaction of these brain areas with measures of neural oscillatory synchronization. Spatial smearing
of EEG activity recorded at the scalp still occurs with these methods, but using appropriate head
models to account for tissue conductivities and objective normalization techniques to account for
volume conduction and noise, these analyses are now possible. We have developed our unique and
validated ‘Time Restricted Artifact and Coherent Suppression’ (TRACS) beamformer method (Wong
and Gordon 2009; Jiwani, Papsin et al. Submitted) to localize auditory evoked cortical activity in the
hemispheres ipsilateral and contralateral to stimulation. Like many imaging methods, the beamformer
technique divides the brain into thousands of 3-dimensional coordinate spaces known as voxels. The
contribution of the dipole centered in each voxel to the measured field can be assessed by the adaptive
spatial filter of the TRACS beamformer. The signal at each voxel is normalized relative to the baseline
brain activity into a Pseudo-Z statistic (Van Veen, Van Drongelen et al. 1997). Dipole moment activity
can then be visualized topographically on age-appropriate head model templates at different latencies

36
for 63,646 voxels in brain space. In this way, the beamformer allows us to create an image of sound-
induced activity in the brain and assess the strength of activity in each hemisphere and in different
cortical regions, and ask questions regarding the lateralization and aural preference of cortical activity
evoked by sound in users of CIs. This is explored in Chapter Four.
More recently, investigators have indicated that co-ordinated oscillations of the various
frequency components of the cortical response underlie communication of various neural groups
between brain regions. Populations of neurons that are activated by sensory stimuli synchronize their
discharges with high precision, allowing them to oscillate in synchrony (Singer 1999; Uhlhaas, Pipa et
al. 2009). Oscillatory activity is generated by voltage changes in the cell membrane which occur with
de-polarization or hyper-polarization. As previously described, voltage changes can be fast (action
potentials) or slow (post-synaptic potentials). In this way, synchronously oscillating neurons,
particularly pyramidal cells in the cortex, exchange information effectively because their input-output
phases occur at the same time (Fries 2005).
The co-ordinated interaction of large numbers of oscillating neurons in different frequency
bands within and across brain areas is believed to underlie cognitive/perceptual dynamics in the brain,
such as attention, memory, learning, etc…. Different cognitive processes are represented by distinct
oscillatory signatures (Lachaux, Rodriguez et al. 1999; Rodriguez, George et al. 1999; Singer 1999;
Varela, Lachaux et al. 2001; Ward 2003; Doesburg, Kitajo et al. 2005; Uhlhaas and Singer 2006;
Doesburg, Emberson et al. 2008; Doesburg, Roggeveen et al. 2008; Doesburg, Green et al. 2009;
Doesburg and Ward 2009; Uhlhaas, Pipa et al. 2009; Doesburg, Herdman et al. 2010; Uhlhaas, Roux
et al. 2010; Green, Doesburg et al. 2011; Palva and Palva 2012). For example, increased synchrony in
the gamma-band activity (30 to 60Hz) is believed to reflect attention and memory (Doesburg,
Roggeveen et al. 2008; Fougnie 2008; Palva, Monto et al. 2010; Green, Doesburg et al. 2011; Doesburg,
Green et al. 2012). This activity occurs with an increase in theta activity (4 to 7Hz) during active
processing and an increase in alpha activity (10 to 13Hz) during integration and feature binding across
long-distances in the brain (Von Stein and Sarnthein 2000). Assessments of EEG frequency analyses
to auditory input can now be done to identify the regions of the brain which are responding in
synchrony and map interactions/connectivity in the cortex. This is explored in more depth in Chapter
Five.

37
2.9 Unilateral cochlear implantation restores hearing and promotes auditory
development in the brainstem and midbrain, but the trajectory of cortical
auditory maturation remains unclear
We have measured changes in evoked activity in the auditory brainstem and thalamo-cortical
pathways in children who were bilaterally deaf from early life and received a CI to hear within limited
durations of auditory deprivation. Auditory brainstem development, measured by decreasing latencies
of evoked potential peaks, is largely complete by the first year of CI use in children with early onset
deafness (Gordon, Papsin et al. 2003; Gordon, Papsin et al. 2006), indicating increasing efficiency of
neural conduction and improved neural synchrony with exposure to sound (Gordon, Papsin et al.
2003). Similar changes have been reported from the auditory brainstems of normal hearing children
over a similar time-course (Salamy and McKean 1976; Starr, Amlie et al. 1977; Jerger and Hall 1980;
Salamy 1984; Hecox and Burkard 2006). Data from Gordon et al (2006) is shown in Figure 2.8A; on
the left is an example of an electrically evoked auditory brainstem response. The stimulus artifact is
shown at time 0ms followed by waves eII, eIII and eV, and on the right, the latency values of wave eV
are plotted at initial device activation and over the first year following CI use in 44 children who had
early onset deafness and were implanted unilaterally (Gordon, Papsin et al. 2006). Recently, we
recorded these same responses in children who were in the original study once they had over a decade
of unilateral CI experience. Examples of these responses from 2 children are shown in B and C. In
both cases, wave eV latency clearly decreases over the first year of CI use, with no further changes
thereafter. This suggests that activity in auditory brainstem is largely complete by the first year
(Gordon, Papsin et al. 2006).

38

39
Further studies concentrated on thalamo-cortical activity as measured by the electrically evoked
middle latency response in a cohort of 10 adolescents who had early onset deafness, were implanted
by 3.18 ± 0.52 years of age, and had who had long-term experience with a unilateral CI (13.0 ± 1.65
years). These adolescents were 16.08 ± 1.87 years of age at the time of the test. Etiology of deafness
was heterogenous. Four of the children had genetic mutations associated with hearing loss: 2 had an
abnormal cochlea (1 confirmed mondini malformation), and deafness in 4 children was due to
unknown causes. The grand mean response recorded from the long-term CI group is shown by the
red waveform in Figure 2.9A along with a grand mean response (black waveform) from a group of 8
normal hearing peers matched for duration of hearing (15.1 ± 2.4 years). Upper and lower bound 95%
confidence intervals of the normal hearing acoustically evoked middle latency response are indicated
by the grey dotted lines. This response is characterized by 3 dominant peaks, Na, Pa and Nb, reflecting
auditory projections from the thalamus to the primary auditory cortex (Fifer and Sierra-Irizarry 1988;
Burton, Miller et al. 1989; McGee, Kraus et al. 1991; Kraus and McGee 1993; Frizzo, Funayama et al.
2007). As shown in Figure 2.9A, the middle latency response evoked by long-term CI users is similar
to that of the grand mean normal hearing waveform, with peak amplitudes reaching normal values
across latency. Analyses of peaks latencies and amplitudes are shown in B. No significant differences
Figure 2-8: Development of auditory brainstem responses after cochlear implantation. Figure
reproduced with permission from Gordon, Jiwani et al. (2013).
A. Example of an electrically evoked auditory brainstem response waveform is shown on the left. The
onset of the CI artifact is shown at time 0ms, followed by peaks eII, eIII and eV. Data from Gordon
et al. (2006) are plotted on the right and show the mean wave eV latency values of 44 children recorded
at initial activation of the implant, and at months 2, 6 and 12 following unilateral cochlear implantation.
B. and C. on the right show the changes in the brainstem responses of 2 children who were in the
original study (Gordon et al., 2006), recorded from initial activation of the device to different intervals
over the first year of cochlear implantation use. New responses recorded after 10 years of unilateral
CI experience are also shown further confirming that little change in the eV latency occurs beyond the
first year of implant use. The wave eV latencies at each time-point are represented on the right for
each child.

40
between the groups were found for any peak (Latencies: Na: t(16)=-2.76, p>0.05, Pa: t(16)=-1, p>0.05);
Nb: t(16)=0.12, p>0.05; Amplitudes: Na: t(16)=-2.72, p>0.05, Pa: t(16)=0.70, p>0.05, Nb: t(16)=-1.61,
p>0.05). The development of normal-like middle latency responses with implant use is consistent
with previous reports (Gordon, Papsin et al. 2005). Whilst the children represented in Figure 2.9
were young at the time of implantation, implant driven changes to the middle latency response can be
restricted in children who have long periods of auditory deprivation in early life prior to implantation
(Gordon, Papsin et al. 2005).
These changes suggest that some degree of auditory development proceeds normally at the
level of the brainstem and midbrain in children who are congenitally deaf when CIs are provided within
early durations of bilateral deafness. Of concern however, questions remain regarding the cortical
activity underlying auditory processing in CI users. Little continues to be known about the long-term
development of the auditory system and the neural networks that are activated by auditory input in
adolescents/young adults who have used a CI to hear for most of their lives. Investigators raised
concerns that maturation of axons in superficial layers of the auditory cortex may never occur in CI
users (Eggermont and Ponton 2003; Eggermont 2008). Unfortunately, these studies reported on only
2 children who had over ten years of implant experience (Eggermont and Ponton 2003). In Chapter
Three of the present Thesis, we address these concerns and investigate the long-term developmental
changes of the auditory cortical response with time and experience after unilateral cochlear
implantation in a larger cohort of children and adolescents.
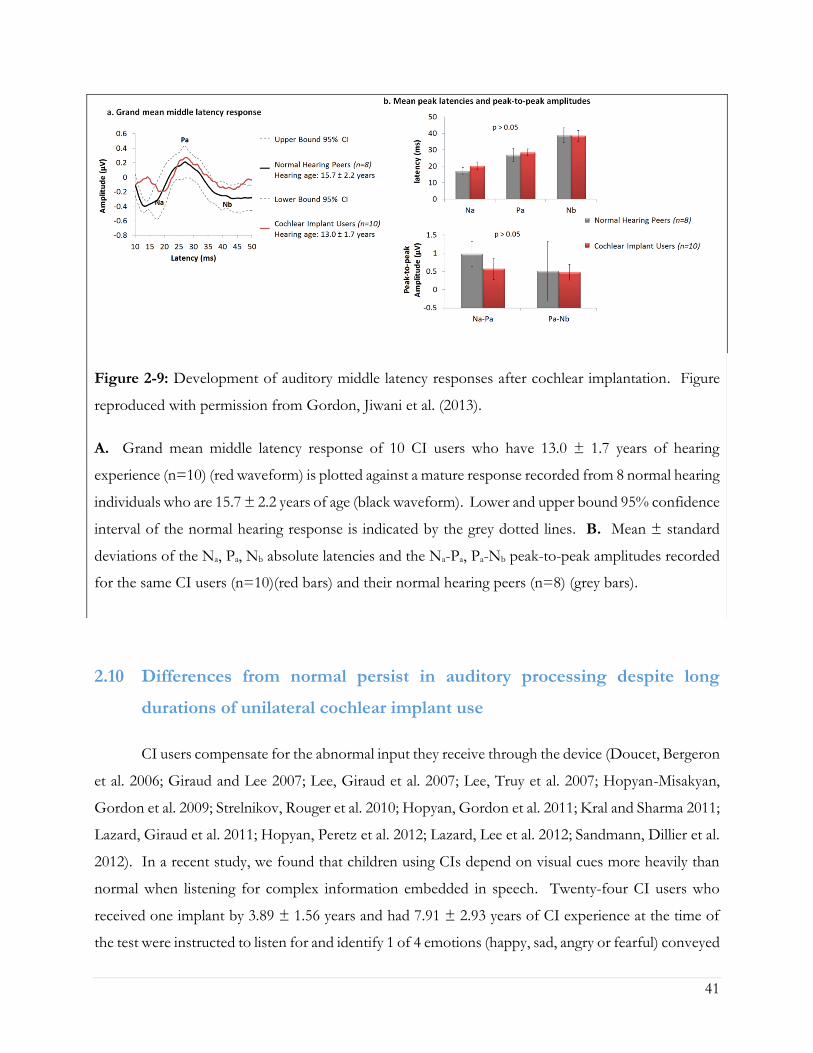
41
2.10 Differences from normal persist in auditory processing despite long
durations of unilateral cochlear implant use
CI users compensate for the abnormal input they receive through the device (Doucet, Bergeron
et al. 2006; Giraud and Lee 2007; Lee, Giraud et al. 2007; Lee, Truy et al. 2007; Hopyan-Misakyan,
Gordon et al. 2009; Strelnikov, Rouger et al. 2010; Hopyan, Gordon et al. 2011; Kral and Sharma 2011;
Lazard, Giraud et al. 2011; Hopyan, Peretz et al. 2012; Lazard, Lee et al. 2012; Sandmann, Dillier et al.
2012). In a recent study, we found that children using CIs depend on visual cues more heavily than
normal when listening for complex information embedded in speech. Twenty-four CI users who
received one implant by 3.89 ± 1.56 years and had 7.91 ± 2.93 years of CI experience at the time of
the test were instructed to listen for and identify 1 of 4 emotions (happy, sad, angry or fearful) conveyed
Figure 2-9: Development of auditory middle latency responses after cochlear implantation. Figure
reproduced with permission from Gordon, Jiwani et al. (2013).
A. Grand mean middle latency response of 10 CI users who have 13.0 ± 1.7 years of hearing
experience (n=10) (red waveform) is plotted against a mature response recorded from 8 normal hearing
individuals who are 15.7 ± 2.2 years of age (black waveform). Lower and upper bound 95% confidence
interval of the normal hearing response is indicated by the grey dotted lines. B. Mean ± standard
deviations of the Na, Pa, Nb absolute latencies and the Na-Pa, Pa-Nb peak-to-peak amplitudes recorded
for the same CI users (n=10)(red bars) and their normal hearing peers (n=8) (grey bars).

42
in the sentence: “I’m going out of the room now and I’ll be back later”. This sentence was presented
via direct auditory input to the child’s CI device by one of four actors (a girl, a boy, a man, a woman)
in an auditory-only or auditory-visual condition. Each emotion was presented twice by each actor, in
each condition, for a total of 64 trials. The listeners were asked to enter their decisions as quickly as
possible using an electronic response pad. Performance accuracy (Figure 2.10A) and reaction time
for accurate responses (Figure 2.10B) were both assessed for each condition and compared to 23
normal hearing controls who were matched for age (9.36 ± 1.01 years of age).
Although children using CIs showed significantly poorer than normal performance on this task
in either condition (Hopyan-Misakyan et al., 2009), they experienced a greater than normal
improvement in accuracy with the addition of visual information as shown in Figure 2.10A. This
means that they relied more on visual cues than their normal hearing peers to identify emotions carried
in speech (Hopyan-Misakyan, Gordon et al. 2009), but did so with longer reaction times, as shown in
Figure 2.10B. Even the accurate decisions took more time for children using CIs to make than their
hearing peers; this was true even when visual cues were available. We thus suggest that children using
unilateral CIs used greater than normal cognitive resources to perform this complex listening task.
The increased reliance on visual cues to process auditory input is consistent with functional
neuroimaging studies, which find abnormally increased activation of the visual cortex in CI users when
listening to meaningful speech (Giraud, Price et al. 2001; Lee, Giraud et al. 2007; Lee, Truy et al. 2007),
perhaps reflecting abnormal development of the auditory and visual systems without normal hearing
(Nishimura, Hashikawa et al. 1999; Lee, Lee et al. 2001; Doucet, Bergeron et al. 2006; Lee, Truy et al.
2007; Meredith and Lomber 2011; Sandmann 2012; Sandmann, Dillier et al. 2012).
These data suggest that unilateral cochlear implantation promotes the development of activity
in the auditory pathways over the long-term, but functional abnormalities persist. Increased
dependence on visual cues in addition to auditory cues when listening with a CI could reflect
compensation for: 1) deleterious or irreversible changes to neural reorganization, which occurred
during the period of auditory deprivation in early life, 2) abnormal representation of sound through
electrical pulses stimulation of the auditory system, and/or 3) absence of auditory input to the deprived
pathways from the opposite un-implanted ear.

43
2.11 Binaural processing is not available with unilateral hearing
Hearing through only one ear or one CI eliminates access binaural hearing. Binaural hearing
is the ability of the auditory system to process and integrate auditory input from both ears to allow
listeners to identify the location of sound sources in space (Basura et al., 2009; Brown and Balkany,
2007; Ching et al., 2007; Litovsky, 2008a, 2008b), increased loudness perception and ease of listening
(Ching et al., 2007; van Hoesel and Tyler, 2003; Steffens et al., 2008) and enhanced speech intelligibility
in the presence of competing noise and in reverberant environments (Basura et al., 2009; Brown and
Balkany, 2007; Ching et al., 2007; van Hoesel and Tyler, 2003). Binaural hearing is especially important
for children because they are rarely in one place and listening to a single speaker at a time. Children
need to attend to and discriminate between several sound sources when playing and learning. The
noise, reverberation and distance, predominant in most learning situations including typical
classrooms, make it challenging for children to listen and learn when binaural cues are not accessible.
Children who use only one CI do not have access to important binaural cues to aid listening in such
Figure 2-10: Accuracy and reaction time for CI processing of auditory input with and without visual
input.
Mean ± 1 standard error of the A. improvement in accuracy when visual input was added and B.
mean reaction time difference of identifying emotions in speech in an auditory only (black bars) and
auditory-visual (grey bars) condition. Responses of 24 CI users (7.9 ± 2.9 years hearing experience)
are compared to those of 23 normal hearing peers matched for hearing age (9.4 ± 1.0 years).

44
challenging conditions. It follows then that we should always strive to provide binaural hearing to
children with hearing loss. For children who are deaf in both ears, binaural hearing might only be
achieved with bilateral cochlear implantation (i.e., CIs in both ears) (van Hoesel and Tyler 2003;
Litovsky, Parkinson et al. 2004; Litovsky, Johnstone et al. 2006; Brown and Balkany 2007; Litovsky
2008; Steffens, Lesinski-Schiedat et al. 2008; Basura, Eapen et al. 2009; Eapen and Buchman 2009;
Gordon, Wong et al. 2010; Salloum, Valero et al. 2010; Chadha, Papsin et al. 2011; Gordon, Jiwani et
al. 2011). Bilateral cochlear implantation is now being increasingly provided to children either in the
same surgery (simultaneously) or in two different surgeries following a period of unilateral implant use
(sequentially).
Bilateral CIs attempt to restore binaural hearing by providing information to both ears.
Normally, the auditory system compares, processes and integrates subtle differences between level and
timing of sounds reaching each ear. In this way, binaural hearing allows: 1) the
identification/localization of sound sources in space (Batteau 1967; Lorenzi, Gatehouse et al. 1999;
Van Deun, Van Wieringen et al. 2009; Grothe, Pecka et al. 2010); 2) increased perception of loudness
and ease of listening through binaural summation (Bocca 1955; Blegvad 1975); and 3) improved
hearing in quiet, noisy and reverberant environments through the head shadow and squelch effects
(Hawley, Litovsky et al. 2004; Van Wanrooij and Van Opstal 2004). Moreover, binaural hearing
reduces the risk of auditory deprivation in the unaided ear (Gordon and Papsin, 2009a, 2009b) and
also makes communication less tiring which enables listening and communication to be a more
pleasant experience. Although restoring binaural hearing is the goal of bilateral implantation, this has
not been completely realized in either adults or children (van Hoesel and Tyler 2003; Seeber and Fastl
2008; Grieco-Calub and Litovsky 2010; Salloum, Valero et al. 2010).
Children who are deaf in both ears hear speech better with bilateral CIs than unilateral implants
(Litovsky, Parkinson et al. 2004; Brown and Balkany 2007; Ching, van Wanrooy et al. 2007; Galvin,
Mok et al. 2007; Peters, Litovsky et al. 2007; Litovsky 2008; Seeber and Fastl 2008; Steffens, Lesinski-
Schiedat et al. 2008; Basura, Eapen et al. 2009; Eapen and Buchman 2009; Gordon and Papsin 2009;
Van Deun, Van Wieringen et al. 2009; Salloum, Valero et al. 2010; Chadha, Papsin et al. 2011), but do
not hear binaural cues normally (Grieco-Calub and Litovsky 2010; Salloum, Valero et al. 2010).
Outcomes improve when both implants are provided with limited delays and at young ages (van Hoesel
and Tyler 2003; Gordon and Papsin 2009; Van Deun, Van Wieringen et al. 2009; Gordon, Wong et al.
2010; Chadha, Papsin et al. 2011). As the duration of inter-implant delay decreases, the two ears

45
develop more symmetric speech perception abilities and children show increasing advantages of
bilateral over unilateral implantation (Gordon and Papsin 2009). Significant improvements on
standardized speech perception tests are seen as early as 6 months following bilateral CI stimulation
in children who receive their second implant simultaneously or within short delays (Gordon and Papsin
2009). Furthermore, children implanted with both CIs simultaneously derive significantly more benefit
from spatial separation of noise compared to children who have longer delays between implants
(Chadha, Papsin et al. 2011). Sound localization improves in children who are provided access to
sound early and in both ears (Van Deun, Van Wieringen et al. 2009). By contrast, children who receive
both CIs sequentially after long inter-implant delays (>2 years) have persistent asymmetries in auditory
function and compromised bilateral benefits for speech perception, even after 36 months of bilateral
CI use (Gordon and Papsin 2009). Sequentially implanted children also seem to depend more on their
first implanted ear than their second for speech perception, and show less bilateral improvement
(relative to unilateral implant use) on speech outcomes than children implanted simultaneously or with
limited delay (Gordon and Papsin 2009). These children localize sound inaccurately and rely heavily
on level cues to do so (Grieco-Calub and Litovsky 2010). The negative effect of inter-implant delay
might be explained by underlying changes to the developing auditory pathways before and after
unilateral and bilateral implantation.
2.12 Evidence of a short sensitive period for bilateral input in human auditory
development
Data presented in Figures 2.8 and 2.9 show that unilateral stimulation promotes development
of the auditory pathways, thus limiting effects of deafness. At the same time, this development might
occur at the expense of pathways from the opposite and deprived ear. This might be explained by the
absence of inhibition, which would normally have come from input from the opposite ear during
binaural hearing (Grothe, Pecka et al. 2010). Without this inhibition, ascending projections from the
stimulated ear may be abnormally strengthened in children who are deaf and use unilateral CIs.
Gordon and colleagues suggested that the stage of unilaterally driven brainstem development
would be an important factor to consider when studying bilateral auditory function in children with
different durations of unilateral implant exposure (Gordon, Salloum et al. 2012; Gordon, Wong et al.

46
2013). Perhaps changes occurring in the brainstem at earlier stages of unilaterally driven development
would have less long lasting consequences on the bilateral pathways than after the unilaterally
stimulated brainstem reached maturity. As shown in Figure 2.8, development in the auditory
brainstem is largely complete by ~1 year of unilateral implant use. Thus, children with >2 years of
unilateral CI experience would be considered to have mature auditory brainstem function and long-
term unilateral use. On the other hand, children who have had <1 year of unilateral experience before
receiving a second implant on the opposite side would be considered to have short-term use with
continuing auditory brainstem development. Gordon and colleagues showed that all children receiving
bilateral implants sequentially produced brainstem responses which were faster when evoked by the
experienced ear compared to the newly implanted ear at initial bilateral implant use (Gordon, Valero
et al. 2008). This was expected and confirmed earlier findings that the first implant promoted
improved neural conduction through the brainstem. Repeated tests completed after (mean ± standard
deviation (SD)) 1.7 ± 1.65 year of bilateral implant use indicated that mismatches in response latencies
persisted in a group of children receiving the second implant after a long delay (>2 years) (Gordon,
Salloum et al. 2012). Increased response latencies in response to sound from the second implanted
side could reflect decreased axonal myelination, longer neural conduction times, slower or weaker
synapses or more asynchronous neural activity – all signs of more limited brainstem development.
Abnormal mismatches between brainstem response latencies were never present in children receiving
bilateral implants simultaneously and resolved with bilateral implant use in children who received both
implants after a short inter-implant delay (<1 year) (Gordon, Valero et al. 2007; Gordon, Valero et al.
2008; Gordon, Jiwani et al. 2011; Gordon, Salloum et al. 2012). Thus, allowing the brainstem to
develop unilaterally for >2 years compromises the later promotion of symmetrically functioning
bilateral auditory brainstem pathways.
Mismatched bilateral auditory development in sequentially implanted children was not
restricted to the brainstem, however. Effects of asymmetric activity in the pathways from the first
stimulated ear were also found in the auditory cortex. Consistent with the brainstem findings, cortical
abnormalities were not resolved by chronic bilateral implant use (3.57 ± 0.74 years) when unilateral
experience exceeded 1.5 years in children who were implanted early (1.87 ± 1.25 years of age). These
findings were recently reported by Gordon, Wong et al. (2013) and are shown in Figure 2.11. Using
a beamformer imaging method (Wong and Gordon 2009) to suppress the electrical artifact from the
CI device and spatially localize areas of cortical activity in the brain (method briefly described in Section

47
1.2.6)., dipole moments for a given voxel in both the left and right auditory cortices were calculated
across latency (virtual sensor) and peak values were used for analyses.
Cortical responses were evoked by unilateral electrical pulse trains delivered from one implant
electrode in 7 children with normal hearing, 8 children who were implanted unilaterally in the right ear
(2.32 ± 1.61 years) and had 7.21 ± 2.48 years of hearing experience and 26 children who used bilateral
CIs for 3.42 ± 0.59 years. Of the bilateral implant users, 10 children received both CIs simultaneously
and 16 were sequentially implanted (right ear implanted first with no hearing aid in the left ear).
Bilateral deafness prior to implantation was limited (1.74 ± 0.90 years) in all children. The children in
this study had less than 12 years of hearing experience, and therefore all produced a cortical evoked
response which was dominated by an immature large amplitude positive peak. The differences
between the dipoles from the left and right auditory cortices were normalized as a percent lateralization
[% lateralization = (dipole right – dipole left) / (dipole right + dipole left) x 100].
A larger than normal variability in the lateralization of cortical dipoles was found in children
receiving bilateral CIs sequentially. A factor analysis of multiple demographic variables identified the
duration of unilateral implant use as the factor, which best accounted for the spread of cortical
responses. Cortical lateralization data was further analyzed for effects of duration of unilateral implant
use occurring prior to bilateral implantation. When responses were evoked by the first (i.e., right)
implant, there was an increase in lateralization of activity to the contralateral left auditory cortex with
unilateral implant use. This became significantly larger than the percent of cortical lateralization in the
simultaneously implanted group at 1.48 years of unilateral implant use. Consistent results were
obtained in data evoked by the second (i.e., left) implant but in this case, cortical lateralization changed
from the normally expected contralateral direction to ipsilateral lateralization with unilateral implant
use. This abnormal switch to larger activity in the ipsilateral auditory cortex became significantly
different from responses in the simultaneously implanted group by 1.37 years of unilateral implant use.
These analyses indicated that children with longer than approximately 1.5 years of unilateral implant
use had experienced an abnormal strengthening of pathways from their first implanted right ear
through the auditory brainstem (Gordon, Valero et al. 2008; Gordon, Salloum et al. 2012) to their left
contralateral cortex. This was not resolved by several years of bilateral implant use and was associated
with poorer speech perception in the second than first implanted ear (Gordon, Wong et al. 2013).

48
The importance of restricting unilateral implant use to less than 1.5 years is further evident in
Figure 2.11. Here, the grand mean lateralization of cortical activity are shown (A), as well as the grand
mean dipole moments identified from the virtual sensors in each hemisphere (B). The group of 16
sequentially implanted children was divided into two groups based on the cut off of 1.5 years of
unilateral implant use. The Short Delay group included 7 children who had 0.86 ± 0.1 years of
unilateral implant experience at the time of testing. The other 9 children, the Long Delay group, had
more than 2 years of unilateral implant use (3.44 ± 1.27 years). The single positive peaked response is
clear in all of the group averaged waveforms shown in Figure 2.11B. The maximum dipoles in the
left and right auditory cortices were marked and analyzed in each child. The left plot of Figure 2.11C
shows that dipoles evoked by stimulation from the first/right implanted ear resulted in significantly
higher dipoles in the left auditory cortex (blue bars) of children who had >1.5 years of unilateral
implant use (Unilateral and Long Delay groups) than other groups of children. The similar findings
for these two groups confirm that unilaterally driven strengthening of projections to the contralateral
left auditory cortex was not reversed by the addition of a second CI. This was true despite the children
in the Long Delay group having had several years of bilateral implant experience at the time of the test.
The right plot in Figure 2.11C shows mean dipoles for each auditory cortex in response to left/second
CI stimulation. The Long Delay group shows significantly higher dipole moments in the left auditory
cortex than the other groups of children. Thus, regardless of which ear was stimulated, the left auditory
cortex (contralateral to the first/right implanted ear) was the more active side of the brain in children
who had used one implant for >1.5 years. One explanation for this finding is that the specialized
processing of language in left auditory cortex (Zatorre and Belin 2001; Zatorre, Belin et al. 2002;
Tervaniemi and Hugdahl 2003; Firszt, Ulmer et al. 2006) may be abnormally increased in unilateral CI
users. An alternate explanation is that unilateral stimulation allowed abnormal strengthening of
pathways from that ear.
Further evidence that the cortical changes were due to unilaterally driven strengthening was
found by assessing which ear preferentially activated the hemisphere contralateral to the ear deprived
during the period of unilateral implant use (i.e., the right auditory cortex). The right auditory cortex
was expected to respond more strongly to input from the left than right ear because the majority of
neurons from one ear normally cross to the contralateral brainstem and ascend ipsilaterally from there.
This was confirmed in the group of children with normal hearing and children with limited unilateral
implant use prior to bilateral implantation (Short Delay and Simultaneous). By contrast, this pattern

49
was reversed in children in the Long Delay group. This means that this group of children had
experienced a strengthening of pathways from their hearing ear to both the ipsilateral right auditory
cortex, as shown by the reversal of aural preference, as well as the contralateral left auditory cortex in
response to long-term stimulation of the experienced right implant. The same reversal of aural
preference in the cortex ipsilateral to the hearing ear has recently been reported in congenitally deaf
white cats (Kral, Hubka et al. 2013).
The abnormal strengthening of pathways from the unilaterally hearing ear to the immature
brain seems to initially occur at the level of the brainstem. This is supported by evidence of
mismatched brainstem latencies observed from children with long (>2 years) unilateral hearing
experience (Gordon, Salloum et al. 2012). The shorter wave eV latencies evoked from the more
experienced ear suggest an increasing efficiency of activity from this side and a weakening of pathways
from the opposite ear, as reflected by slower peak latencies on the second implanted side. This could
result from a lack of inhibitory processes in the brainstem which are normally present during binaural
hearing (Grothe, Pecka et al. 2010). Listening from one side would allow auditory input from the first
right implanted side to be projected to the cortex with abnormally high excitation during development
thus strengthening pathways to the contralateral cortex. It appears that if this is allowed to occur until
the brainstem is largely developed (i.e., >1 year of unilateral implant use), it establishes asymmetric
activity in the auditory pathways, which is not easily reversed by providing a second implant in the
deprived ear. Limiting the period of unilateral hearing in children by providing bilateral CIs with little
or no delay appears to protect the bilateral pathways from this abnormal development. These findings
thus suggest that there is a sensitive period of 1.5 years for binaural auditory development in children
(Gordon, Wong et al. 2013).

50

51
2.13 Does long-term unilateral cochlear implant use have abnormal
consequences for cortical auditory development?
We have reviewed evidence showing that access to sound within limited durations of bilateral
deafness in early life promotes development of activity along the auditory pathways in children who
have many years of hearing experience with a unilateral CI. At the same time however, unilaterally
driven stimulation of the auditory system leaves the opposite pathways deprived of input and
susceptible to reorganization. Thus, both bilateral and unilateral deprivation should be limited to
promote optimal hearing in children who use CIs and enable them to function better and more
naturally in challenging listening situations such as the playground or classroom environments.
Unfortunately, limiting the period of unilateral deprivation has not always been possible, particularly
in the early years of cochlear implantation when the device was provided in only one ear. There are
several reasons why CIs were traditionally provided only unilaterally. These include: 1) benefits of
Figure 2-11: Cortical dipole activity evoked by auditory input in children with different durations of
unilateral implant use. Reproduced with permission from Gordon, Wong et al. (2013).
“A. Per cent cortical lateralization (mean ± 1 standard error) is plotted for each participant group.
Greater than normal contralateral lateralization to right/CI-1 stimuli was found in long delay and
unilateral CI users (P<0.05 and <0.0001, respectively) but not in short delay and simultaneous groups
(P>0.05). The long delay group showed a decrease in contralateral lateralization/increase in ipsilateral
lateralization relative to those with normal hearing in response to left/CI-2 stimulation. This did not
occur in the short delay and simultaneous groups. B. Grand mean virtual sensor data for left and
right hemispheric sources of P1 (normal hearing) and P1ci (CI users for stimulation from right/CI-1
and left/CI-2). Large peaks in responses to CI-1 (right) stimulation can be seen in the long delay and
unilateral group data. C. Left and right hemispheric dipole moments (mean ± 1 SE) for P1/P1ci in
each group in response to right/CI-1 and left/CI-2 stimulation. In response to CI-1 (right)
stimulation, there is a marked increase in left hemispheric dipole moments in participant groups with
>2 years of unilateral hearing experience (long delay and unilateral; P<0.05).” (Gordon et al., 2013,
Brain, Figure 7, p.11).

52
implantation associated to linguistic, psychological, and social development were undetermined in the
early days of cochlear implantation (Lane and Bahan 1998; Svirsky, Robbins et al. 2000), 2) risks related
to the surgical procedure were unclear (Lane and Bahan 1998), 3) since the cost of cochlear
implantation is high, cost-utility analyses were not available at the time, and from a public policy
perspective, net savings to society were unknown (Cheng, Rubin et al. 2000; Mohr, Feldman et al.
2000; O'Neill, O'Donoghue et al. 2000), 4) some parents chose to ‘save’ one cochlea in the hope that
a cure or alternative treatments to deafness (i.e., stem cell therapy or gene therapy) may become
available in the future (Brown and Balkany 2007). Over the past decade, research has addressed many
of these questions related to the safety, efficacy and cost-utility of bilateral over unilateral cochlear
implantation. Positive outcomes for development of the auditory pathways, speech and language
performance, social/cultural integration and improved quality of life with two implants rather than
one all indicate that CIs should be provided to children early and in both ears within very limited or
no delays between implants.
Unilateral CI use exceeding a sensitive period of 1.5 years drives abnormal mismatches in
activity at the level of the brainstem (Gordon, Salloum et al. 2012) and cortex (Gordon, Wong et al.
2013) which are associated with abnormal asymmetries and integration of bilateral input (Hawley,
Litovsky et al. 2004; Peters, Litovsky et al. 2007; Litovsky 2008; Gordon and Papsin 2009; Van Deun,
Van Wieringen et al. 2009; Van Deun, Van Wieringen et al. 2009; Grieco-Calub and Litovsky 2010;
Salloum, Valero et al. 2010; Chadha, Papsin et al. 2011). Given these findings, in the present Thesis,
we asked whether there are cortical consequences of missing this early sensitive period and driving
maturation of the auditory cortex with unilateral implant stimulation. We tested this with 3
experiments. First, we used well established methods of auditory evoked potential recordings to
measure electric currents generated in the auditory pathways in response to sound to determine
whether auditory activity in the cortex undergoes normally expected maturational changes after 10
years of unilateral CI use. Second, we further developed the TRACS beamformer and used this
objective imaging method to determine which areas of the brain respond to sound in adolescents who
grow up using one CI. Recently, many of these adolescents/young adults who were implanted as
babies and have already had many years of unilateral hearing experience, received a second CI in their
opposite ear in hopes of deriving benefits of bilateral implantation. This provided us with a unique
opportunity to stimulate the unilaterally deaf ear for the first time and locate the cortical sources of
activity generated from the deprived auditory pathways to assess the plastic effects of long-term

53
unilateral stimulation/deprivation in the adolescent brain. Third, we explored how activity in these
areas of the brain is coordinated to respond to sound. We used a mathematical filtering algorithm to
assess the brain regions that respond to sound in a synchronized coordinated manner and determine
how this interaction across brain areas is involved in hearing after long durations of unilateral implant
use. Responses from the newly implanted side were examined to determine whether this ear is
segregated from the cortical hearing network.

54
3. Chapter Three – Central auditory development after long-term
cochlear implant use
This chapter has been published with required journal formatting:
Jiwani, S., Papsin, B.C., Gordon, K.A., 2013. Central auditory development after long-term cochlear
implant use. Clinical Neurophysiology, 124, 1868-80.
3.1 Abstract
Objective: We determined whether long-term cortical auditory development is altered or delayed in
children using cochlear implants (CIs) relative to their normal hearing peers. We hypothesized that
cortical development in children using unilateral CIs follows a normal trajectory with long-term
auditory input when the duration of bilateral auditory deprivation in childhood is limited.
Methods: Electrically-evoked cortical responses were recorded in 79 children who received one CI
within 2.03 ± 1.36 years of bilateral deafness and had up to ~16 years of time-in-sound experience,
and in 58 peers with normal hearing. Amplitude differences between the responses from children
using CIs and with normal hearing were calculated between 0 and 300ms.
Results: Responses from CI users remain different from those of their normal hearing peers. These
differences decreased over time, but were not eliminated even after 10 years of time-in-sound.
Specifically, the P1-N1-P2-N2 complex, typical of a normally mature response, began to emerge by 10
years of time-in-sound experience, but the amplitudes of P2 and N2 became abnormally large.
Conclusion: Mature-like cortical responses emerge in children after long-term unilateral CI use,
however, differences from normal persist.
Significance: Maturation of cortical responses with long-term CI use potentially underlies functional
improvements in hearing. Persistent differences from normal could reflect an increase in attention or
multi-sensory processing during listening.

55
3.2 Introduction
Hearing can be restored to children who are deaf by providing cochlear implants (CIs) which
electrically stimulate the auditory pathways. Although the CI promotes short term changes in the
auditory system (Ponton and Eggermont 2001; Sharma, Dorman et al. 2002; Sharma, Dorman et al.
2002; Gordon, Tanaka et al. 2005; Sharma and Dorman 2006; Gordon, Tanaka et al. 2008; Gordon,
Wong et al. 2010), there is little known about whether the children using these devices develop a mature
auditory system after long-term use. Normal maturation could be impeded by several factors including:
1) the abnormal auditory input provided by the implant; 2) stimulation in only one ear; and 3) bilateral
auditory deprivation during potentially sensitive developmental periods in early life. In the present
study, we took an optimistic approach and hypothesized that cortical development, measured by
assessing changes in the cortical evoked potential response, follows a normal trajectory with time-in-
sound over long-term unilateral CI use when the duration of bilateral auditory deprivation in childhood
is limited.
3.3.1 Deafness prior to cochlear implantation alters normal brain development
Prior to cochlear implantation, the immature and deaf auditory brain is vulnerable to take-over
by non-auditory neuronal networks (i.e., cross-modal plasticity) (Lee, Lee et al. 2001; Bavelier and
Neville 2002; Sharma, Dorman et al. 2002; Harrison, Gordon et al. 2005; Gordon, Papsin et al. 2007;
Moore and Shannon 2009; Sharma, Nash et al. 2009). Recent evidence of cross-modal reorganization
in a cat model of congenital deafness indicates that while the primary auditory cortex is spared (Kral,
Schroder et al. 2003), association areas of the auditory system become the targets of other sensory
systems (Lomber, Meredith et al. 2010; Meredith and Lomber 2011). Specifically, the Posterior
Auditory Field and the Dorsal Zone of the association auditory cortex become recruited by the visual
system to perform visual functions (Lomber, Meredith et al. 2010). These changes appear to result
from a direct competition for resources in areas that receive multi-sensory input and likely reflect the
brain’s attempt to compensate for auditory deprivation with enhanced visual processing.
Electrical stimulation of the auditory nerve with a CI could provide sufficient input to the brain
to slow or halt cross-modal changes (Kral, Hartmann et al. 2001; Harrison, Gordon et al. 2005; Papsin

56
and Gordon 2007; Gordon, Jiwani et al. 2011). Providing a CI at young ages drives cortical auditory
development within 3 to 6 months of implant use (Gordon, Papsin et al. 2005; Sharma and Dorman
2006). Yet, normal maturation of the auditory cortex requires much longer periods and is known to
continue throughout adolescence until ~20 years of age (Albrecht, Suchodoletz et al. 2000). This
suggests that long durations of hearing might be needed to achieve a mature auditory cortex in children
who are deaf and use CIs. Failure to reach maturity could result from delivering abnormal stimulation
to one ear alone with an implant (Ponton and Eggermont 2001) and any effects of bilateral deafness
in early development (Kral, Hartmann et al. 2001; Bavelier and Neville 2002; Eggermont and Ponton
2003). In the present investigation, we asked whether normal patterns of cortical maturation occur in
children who have used their CIs to hear for many years.
3.3.2 Early auditory cortical development in cochlear implant users follows a
normal-like trajectory
The development of the central auditory pathways over time can be studied using cortical
auditory evoked potential responses. Recent investigations indicate that electrically-evoked auditory
cortical responses of deaf children who have limited durations of bilateral deafness and have good
speech perception outcomes with their implants resemble the large amplitude broad positive peak
response recorded from young children with normal hearing who are under 10 years of age (Ponton
and Eggermont 2001; Sharma, Dorman et al. 2002; Sharma, Dorman et al. 2002; Gordon, Tanaka et
al. 2008). Many groups have suggested that this immature positive peaked response is homologuous
to the normal P1 cortical peak (Ponton, Eggermont et al. 2000; Ponton and Eggermont 2001; Ponton,
Eggermont et al. 2002; Sharma, Dorman et al. 2002; Gilley, Sharma et al. 2006; Sharma, Nash et al.
2009), which is generated by recurrent excitatory inputs from the thalamus to neurons in deep layers
(III to VI) of the auditory cortex (specifically the lateral portion of Heschl’s gyrus) (Liegeois-Chauvel,
Musolino et al. 1994). It may also reflect temporally overlapping auditory output from the reticular
activating system, which is known to integrate auditory and non-auditory multi-sensory input (Kraus,
McGee et al. 1992; Liegeois-Chauvel, Musolino et al. 1994; Ponton, Eggermont et al. 2000; Eggermont
and Ponton 2003).

57
Sharma and colleagues (2002; 2006) reported that when children with congenital deafness
receive a CI by 3.5 years, the positive P1-like peak in the cortical response decreases to age-appropriate
P1 latencies by 6 months post-implantation. They concluded that the central auditory system remains
highly plastic and minimally degenerate for a period of about 3 years following auditory deprivation in
these children (Sharma, Nash et al. 2009). These findings were consistent with reports by Eggermont,
Ponton and colleages (1997; 2003) who also found that the rate of cortical auditory maturation in
congenitally deaf CI users, measured by the latency of the positive peak, is delayed relative to normal
only by the amount of time that the child spent without significant hearing.
3.3.3 Auditory cortical maturation may be altered in cochlear implant users
Previous investigators raised concerns that the superficial layers of the auditory cortex may
never mature in CI users if the implant is not provided before 3.5 years of age (Eggermont and Ponton
2003; Sharma, Gilley et al. 2007), while others suggested that this might not occur even if the period
of auditory deprivation was further limited (Eggermont and Ponton 2003). The superficial layers of
the auditory cortex (III and II), which only mature around 9 to 12 years of age in typically developing
children (Ponton, Eggermont et al. 2000; Moore and Guan 2001), might require auditory input in early
life to develop (Eggermont and Ponton 2003). In normal hearing individuals, the cortical auditory
evoked potential response reflects this maturational process. Of importance, the large positive peaked
response seen early in development is thought to reflect activity in deeper auditory cortical layers (IV,
V and VI) (Kraus, McGee et al. 1992; Ponton and Eggermont 2001; Gordon, Tanaka et al. 2008),
which are known to develop first (Ponton, Eggermont et al. 2000; Moore and Guan 2001). However,
as primary auditory thalamo-cortical and cortico-cortical connections in superficial cortical layers
mature in adolescence, the development of a small negative amplitude peak, labelled N1, causes this
positive peak to bifurcate into two separate adult P1 and P2 components (Ponton, Eggermont et al.
2000; Ponton and Eggermont 2001; Eggermont and Ponton 2003; Wunderlich and Cone-Wesson
2006; Gordon, Tanaka et al. 2008). N1 is believed to be generated by late maturing auditory thalamo-
cortical and cortico-cortical activity in the superficial layers (III and II) of the superior temporal cortex
(Ponton, Eggermont et al. 2000; Ponton and Eggermont 2001; Ponton, Eggermont et al. 2002). The
development of these neural connections have been linked to the emergence of more complex
auditory-specific perceptual skills, such as hearing in noise, understanding degraded speech, sound

58
localization, and gap detection, as well as other complex temporal processing skills (Ponton,
Eggermont et al. 2000).
Eggermont and Ponton (2003) questioned whether children using CIs would ever develop
cortical responses containing this small negativity (Eggermont, Ponton et al. 1997; Ponton, Eggermont
et al. 2000; Ponton and Eggermont 2001; Eggermont and Ponton 2003). If this did not occur, it would
suggest that development of neuronal connections in the superficial layers of the auditory cortex of
children using CIs is persistently delayed or proceeds in an aberrant fashion. Effects of deafness
and/or unilateral CI stimulation could disrupt interhemispheric connections and auditory thalamo-
cortical and cortico-cortical pathways that are normally present by adolescence (Ponton, Eggermont
et al. 2000).
Although it is clear that chronic auditory stimulation with a CI promotes activity in the central
auditory system of children who are deaf from infancy (Kral, Hartmann et al. 2001; Harrison, Gordon
et al. 2005; Gordon, Valero et al. 2007; Papsin and Gordon 2007; Gordon, Jiwani et al. 2011), the long-
term development of the auditory cortex in these children has not been specifically evaluated. In the
present study, we used auditory evoked potential recordings to assess changes in the cortical evoked
response in a cohort of children who had up to 16 years of time-in-sound experience. Responses from
this group were compared with those recorded from more inexperienced CI users and age matched
normal hearing controls. Although normal maturation was not fully achieved, we found evidence of
continued change in cortical activity over long-term CI use.
3.4 Methods
3.4.1 Participants
Table 3.1 provides demographic details for the 79 children using CIs who participated in this
study. Thirty-nine of the children were female and 40 were male. This group of children had (mean
± standard deviation (SD)) 2.03 ± 1.36 years of bilateral deafness and between 0.69 to 15.95 years of
time-in-sound experience at the time of the test. Most of the children (n=56) were pre-lingually deaf,
10 children had a peri-lingual onset of severe-to-profound hearing loss, and 13 lost their hearing post-
lingually. Aetiology of deafness varied. Most were congenital in nature due to unknown causes (n=38);

59
37 of the hearing losses had a genetic aetiology, including 2 with a confirmed diagnosis of Pendred
syndrome, 4 with Ushers, 1 with KID syndrome, 3 with an abnormal cochlea and 1 with a
developmental delay. Two of the children acquired a severe-to-profound hearing loss due to
meningitis and 2 for other causes, including a progressive hearing loss resulting from a Cholesteatoma.
Of all the 79 children who participated in this study, 9 children had 5.68 ± 1.77 years of useable residual
hearing prior to cochlear implantation and combined with duration of CI experience, had, on average,
over 10 years of total time-in-sound experience (i.e., 13.22 ± 1.41 years) at the time of the test. The
other 70 children were deaf from infancy and received their CI by 2.35 ± 1.27 years of age. Of these
children, 17 had used an implant for over 10 years (i.e., 12.59 ± 1.44 years) and 50 had less experience
(i.e., 4.43 ± 2.67 years). All children were implanted with a CI device from Cochlear Inc. with full
insertion. Eight children used a Nucleus CI22 device, 26 used a CI24R(CS) contour implant, 15
children had a CI24R(CA) contour advance, 18 children received a CI24M device, and 12 used a
CI24RE Freedom Contour Advance implant. The type of CI device used by each child is indicated in
Table 1. To compare results with normal hearing responses, acoustically-evoked cortical responses
were recorded from 58 normal hearing controls who were between 7 and 19 years of age (31 female :
27 male). Children in the normal hearing control group were matched for chronological age and
duration of hearing experience (time-in-sound).
As expected, many variables changed together over time. Strong positive correlations were
found in the implant group between total time-in-sound experience and age at test (R=0.97, p<0.01),
duration of CI use (R=0.90, p<0.01), age at cochlear implantation (R=0.68, p<0.01), duration of
residual hearing (R=0.45, p<0.01) and duration of bilateral deafness (R=0.37, p<0.05). Thus, it was
impossible to isolate the duration of total time-in-sound experience from these other variables.
Accordingly, the analyses of the present study focused on experience-dependent changes in the cortical
response (i.e., total time-in-sound experience).

60
Table 3-1: Mean (X) ± standard deviations (SD) of demographic information of the CI users.

61
3.4.2 Evoked potential recordings
Electrically-evoked cortical responses were recorded using the NeuroScan 4.3 system with a
Synamps I amplifier. Responses were recorded from all participants while they sat in a soundproof
booth and watched a silent movie with closed captioning. Responses were recorded from a surface
electrode at a midline cephalic location (Cz) and referenced to each earlobe in a two channel recording.
The frontopolar point (Fpz) was used for the ground electrode. Responses were sampled at a rate of
500Hz and an online band-pass filter from 0.05Hz to 100Hz was applied. Averaged responses were
further filtered off-line when necessary, using a 30Hz low-pass digital filter with a 12dB/octave rolloff.
A minimum of 200 sweeps were recorded and at least 2 visually replicable responses were obtained for
all test conditions. All sweeps containing greater than ± 100µV were rejected from the average.
Biphasic electrical pulse trains of 250 pulses per second (i.e., 9 pulses per train) lasting 36ms
were delivered by a single electrode on the apical end of the electrode array (#20, #18, or #16,
depending on the stimulation mode). Pulse trains were presented at a rate of 1Hz to generate cortical
responses. Given the variability in the types of device used and the different MAPS, the stimuli were
delivered from a single electrode at intensities which were at maximum comfortably loud levels for
each child. To determine current presentation levels, auditory brainstem responses were electrically-
evoked by biphasic electrical pulses at a stimulus repetition rate of 11Hz. Current level was increased
to maximize the amplitude growth of the auditory brainstem response within the range of comfort for
the child. Once these levels were set according to the auditory brainstem response test, the current
levels were decreased by 10 current units for the pulse train stimuli, to account for the increased
perception of loudness with greater number of pulses presented in short periods. Stimulus intensity
of biphasic pulse trains used to record the electrically-evoked cortical responses in CI users ranged
from 180 to 240 current units. The stimulation intensity used for each child in our study is indicated
in Table 1. Auditory evoked cortical responses recorded from normal hearing individuals were
measured using 500Hz tone burst stimuli, lasting 36ms. A Tukey window was applied over the first
and last eighths of each tone, to minimize high frequency onset and offset effects. Stimuli were
presented at a rate of 1Hz through ER3-14A insert earphones to one ear at a time. The behavioural
threshold of hearing for this tone was measured in each of the children using the modified Hughson-
Westlake bracketing method (Carhart and Jerger 1959), and the tone was then presented at an intensity
level of 40dB above the threshold for that ear.

62
3.4.3 Analysis of the electrically-evoked cortical responses
Changes in the morphology of the cortical auditory evoked waveform and in the amplitude
and latency of the cortical responses have been used as indices of auditory maturation in typically
developing children (Albrecht, Suchodoletz et al. 2000; Wunderlich, Cone-Wesson et al. 2006) and in
deaf children who use a CI to hear (Gordon, Tanaka et al. 2005; Harrison, Gordon et al. 2005; Sharma,
Dorman et al. 2005; Gordon, Tanaka et al. 2008). Amplitude peaks in the response were visually
identified at latencies corresponding to those identified in normal responses (Gordon, Tanaka et al.
2008). The observer was blinded to child, age at implant, age at test, and duration of CI use. To
compare the differences in the cortical responses of long-term CI users relative to a normal hearing
mature auditory cortical response, a grand mean normal hearing mature cortical waveform was created
using acoustically-evoked auditory responses from 15 typically developing, normal hearing children
who were between 13 to 18 years of age at the time of the test (mean ± SD time-in-sound/age = 15.54
± 2.11 years). Figure 3.1 shows the grand mean normal hearing mature cortical waveform with upper
and lower bound 95% confidence intervals.
Figure 3-1: Normal hearing mature cortical response.
Grand mean normal hearing mature cortical waveform from 15 typically developing, normal hearing
children who were between 12 to 18 years of age at the time of testing. Mean age/time-in-sound is
15.54 ± 2.11 years.

63
The amplitude differences between the individual cortical response waveforms of CI users and
the normal hearing mature waveform was then calculated in a recording window ranging from 0 to
300ms. The Trapezoid rule was used to calculate the amplitude difference between the 2 waveforms:
the grand mean normal hearing mature cortical waveform was subtracted from the cortical response
of each CI user at each 2ms amplitude point between 0 to 300 ms (i.e., 150 amplitude difference
points). The sum of these points was then multiplied by 2 (i.e., distance between each amplitude point)
to provide the total difference from normal amplitude for each child’s response. The amplitude
difference analysis was used to visually compare the morphological development of the cortical
waveform of CI users at different durations of device use, relative to a normal hearing mature response,
without making a priori decisions on the presence/absence of previously defined latency ranges of
amplitude peaks. In turn, this allowed for an objective quantification of any differences from normal
across latencies.
Amplitude differences in the 50 to 150ms and the 150ms to 300ms latency range were assessed
separately as dependent variables using multiple linear regression, with time-in-sound and implant type
as predictor variables. Duration of total time-in-sound was calculated as the duration of CI experience
plus the duration of any period of useable residual hearing prior to implantation. The duration of
useable residual hearing was defined as the number of years that thresholds of 40dB HL or better were
present with or without hearing aids at any two given frequencies. Given the large time-in-sound range
of the children in the CI group (i.e., 0.71 to 15.95 years) and the large age range in the normal hearing
group (i.e., 7 to 19 years), responses from children were separated into interval groups of 0 to 2 years,
2 to 4 years, 4 to 7 years, 7 to 10 years, 10 to 13 years, 13 to 16 years, and 16 to 19 years of time-in-
sound. This grouping by time-in-sound in the CI users and age in the normal hearing group allowed
us to analyze changes in the response as a function of time-in-sound experience. Data from normal
hearing participants and CI users in 3 groups (time-in-sound = 7 to 10 years, 10 to 13 years, and 13 to
16 years) were compared using t-tests with Bonferroni confidence interval adjustments. These analyses
were followed by more traditional peak picking methods to assess whether expected changes in specific
peaks were occurring over time. Differences between responses recorded from the CI users and the
children with normal hearing were evaluated using 2-tailed independent t-tests with Bonferroni
adjustments.

64
3.5 Results
3.5.1 Cortical responses continue to mature with auditory experience in normal
hearing children and in users of cochlear implants
Responses from CI users who had up to 16 years of auditory experience are shown in Figure
3.2 on the left column (Figure 3.2A) and responses from normal hearing children who were between
7 to 19 years of age at the time of the test are shown on the right (Figure 3.2B). Responses were
grouped by time-in-sound intervals into ranges of 0 to 2 years, 2 to 4 years, 4 to 7 years, 7 to 10 years,
10 to 13 years, 13 to 16 years, and 16 to 19 years of experience.
As indicated in Figure 3.2, the auditory cortical response recorded from CI users and normal
hearing children who had between 0.69 to 7 years of time-in-sound experience is dominated by a large
positive peak, followed by a large negative peak. This positive peaked response is largest in amplitude
in children who had less than two years of time-in-sound experience (mean ± SD: 13.05 ± 11.62µV),
and decreases as auditory experience extends to 7 years (5.44 ± 5.65µV). Between 7 and 10 years time-
in-sound, a small negative deflection begins to emerge around 100ms, causing the morphology of the
cortical response to change from one positive peaked response into a multi-peaked waveform. By 10
years of time-in-sound, the response in all cases is comprised of P1, N1, P2 and N2 peaks. The P1-N1
complex is not seen in children who had less than 10 years of time-in-sound experience. As shown in
Figure 3.2B on the right, morphological changes of the auditory evoked cortical response in normal
hearing children occur in a similar way and at similar durations of time-in-sound. Children with both
normal hearing and CIs required 10 years of auditory experience to develop responses with the
polyphasic P1-N1-P2-N2 complex, typical of a mature auditory cortical response. Indeed, cortical
waveforms produced by CI users indicated amplitude values similar to those of their normal hearing
peers across the latency range (i.e., 0 to 300ms) for children who had between 7 to 13 years of hearing
experience (p>0.05). However, differences from normal increased in children with more experience
(t(15.6) = 2.4, p<0.05).
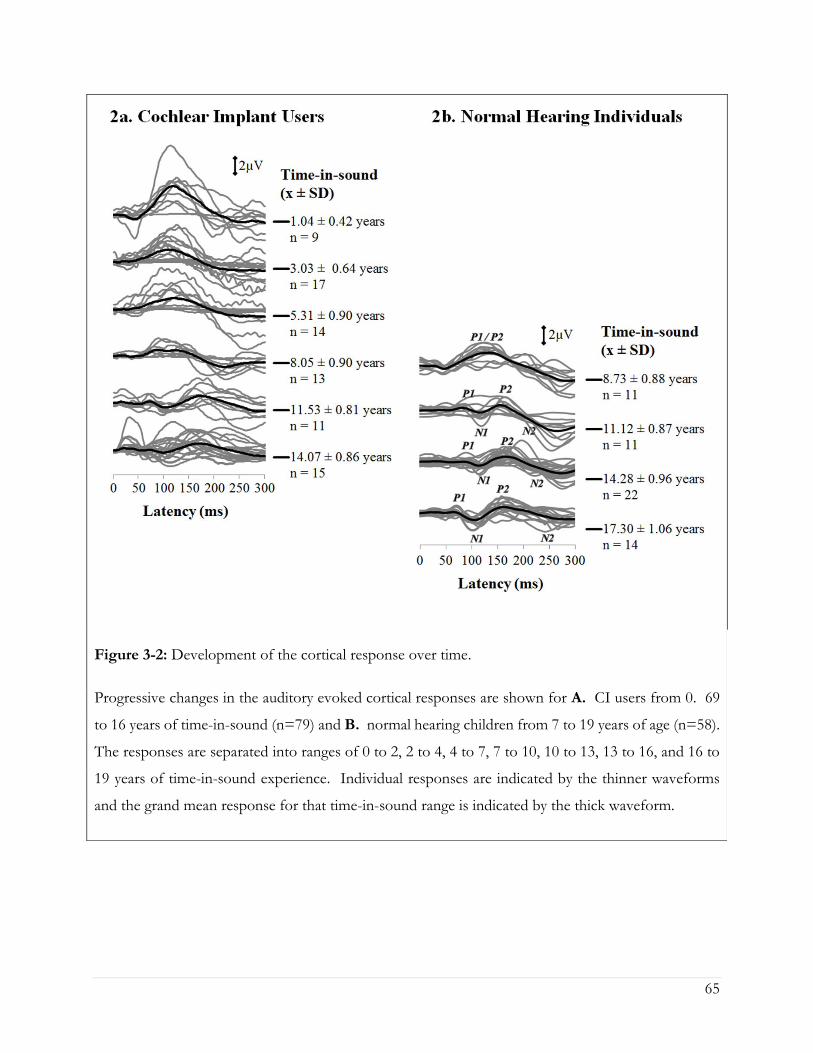
65
Figure 3-2: Development of the cortical response over time.
Progressive changes in the auditory evoked cortical responses are shown for A. CI users from 0. 69
to 16 years of time-in-sound (n=79) and B. normal hearing children from 7 to 19 years of age (n=58).
The responses are separated into ranges of 0 to 2, 2 to 4, 4 to 7, 7 to 10, 10 to 13, 13 to 16, and 16 to
19 years of time-in-sound experience. Individual responses are indicated by the thinner waveforms
and the grand mean response for that time-in-sound range is indicated by the thick waveform.

66
3.5.2 Cortical development in users of cochlear implants follows a normal
trajectory with time-in-sound with differences emerging in latencies
greater than 150ms
As shown in Figure 3.2, early cortical wave peaks appear to follow a normal maturational time
course with time-in-sound experience, but peaks in the later latency ranges remain different from
normal. Figure 3.3A shows the difference in amplitude (in grey shading) between the grand mean
cortical responses of CI users grouped by specific time-in-sound intervals (dark grey waveforms) and
a grand mean normal hearing mature waveform (black waveform) over a 0 to 300ms latency period.
The mean age at test of the responses from the normal hearing group used to create the mature cortical
waveform was 15.54 years. As seen from Figure 3.3A, the shaded areas of difference from normal
are largest in paediatric CI users who have less than 7 years of time-in-sound experience. However, as
their auditory experience approaches the mean time-in-sound of the normal hearing waveform (~15
years), differences between the CI and the normal cortical responses become minimal. Indeed,
responses from children with over 10 years of time-in-sound consistently contain a small biphasic peak
which occurs at the same latency range as the normal P1 and N1 peaks (i.e., between 50 to 150ms). The
P1-N1 complex in these more mature responses is followed by P2 and N2 peaks in the 150 to 300ms
latency range.
Figure 3.3B displays these amplitude differences between the CI responses and the grand
mean normal hearing cortical waveform in a 3-dimentional plot as a function of latency and against
time-in-sound. The colours are used to identify the magnitude of difference between the responses of
the children using CIs and the grand mean normal hearing mature waveform. The dark red colour
shows greater differences from normal and the dark blue represents the least differences from the
normal response. With time-in-sound experience, the cortical responses of long-term CI users
approach a mature normal hearing response. Of interest, amplitude differences from the normal
mature response in the 50 to 150ms latency range (i.e., P1-N1 latency range) are greatest (areas of dark
red) in children with less than 7 years of time-in-sound, compared to a mature normal hearing response.
However, differences from normal decrease with auditory experience. With over 10 years of time-in-
sound experience, the early cortical peaks in the responses recorded from CI users approach normal-
like latency and amplitude values compared to the normal hearing grand mean cortical waveform
(t(34.7)=0.6, p>0.05), as indicated by the dark blue areas. By contrast, abnormally large peak amplitudes

67
emerge with auditory experience in the 150 to 300ms latency range (i.e., P2-N2 latency range) in long-
term users of CIs who have over 10 years of time-in-sound experience (light blue colour) (t(37.8)=3.9,
p<0.01).

68
Figure 3-3: Cortical response difference as a function of time after cochlear implantation compared
to the normal hearing mature waveform.
A. Grand mean cortical waveforms of CI users are plotted for specific intervals of time-in-sound
experience. All mean responses are compared to a normal and mature waveform (mean age/time-in-
sound: 15.54 ± 2.11 years, n=15). B. The trajectory of cortical response waveforms of CI users
(n=79) from 1.04 ± 0.42 years to 14.07 ± 0.86 years of time-in-sound experience are plotted on a 3-
dimensional map: x-axis = Latency (ms); y-axis = Amplitude (µV); z-axis = Duration of time-in-sound
(years). Difference between the CI and normal responses is plotted by colour. The largest differences,
shown in dark red, occur at short time-in-sound. The dark blue colour represents minimum
differences.

69
3.5.3 Normal-like cortical maturation in the 50 to 150ms latency range is time
and experience dependent
To further assess changes in the cortical responses over time, we analysed early (50 to 150ms)
and later latency (150 to 300ms) ranges of the response separately. In Figure 3.4A., amplitude
differences in the 50 to 150ms latency range (i.e., P1-N1 latency range) between the cortical waveform
of the CI users and the grand mean normal hearing mature waveform are plotted against total time-in-
sound for all 79 CI users. Positive amplitude differences indicate larger amplitudes in CI users and
negative amplitudes indicate larger amplitudes in the group with normal hearing. Linear regression
analyses revealed that cortical responses of CI users approach normal values in the 50 to 150ms time
window with increasing time-in-sound experience (t(1, 77)=-3.7, p<0.01), regardless of implant type (t(2,
76)=-0.9, p>0.05).
Given that the differences from normal in the 50 to 150ms range decrease with time-in-sound,
we identified peak amplitude values for the P1-N1 complex in the 50 to 150ms latency range and
assessed how these change in latency and amplitude with time-in-sound for children with normal
hearing and their peers with CIs. Data summarized in Figure 3.4B illustrates the mean absolute
latencies and amplitudes of the P1 and N1 peaks once this complex emerges (i.e., after 10 years time-
in-sound experience). Cortical responses recorded from 11 children in each group who had between
10 to 13 years time-in-sound, and from 15 CI users and 22 normal hearing adolescents with 13 to 16
years time-in-sound experience, were compared. Independent 2-tailed t-test analyses revealed that
neither the latencies (P1: t(15.8)=-1.0, p>0.05; N1: t(16.8)=-0.7, p>0.05) nor the amplitudes (P1: t(16.6)=-0.1,
p>0.05; N1: t(16.6)=-0.6, p>0.05) of peaks P1 and N1 were significantly different from normal at this
long-term stage of CI use (p>0.05).

70
Figure 3-4: Difference in latency and amplitude of the P1-N1 complex as a function of time-in-sound
compared to normal hearing peers.
A. Amplitude differences between CI (n=79) and the normal mature grand mean response were
calculated between 50 and 150ms (amplitude x 50 latency measurements) (µV x ms) and plotted against
time-in-sound. B. Mean ± standard deviations of the P1 and N1 absolute latencies and amplitudes of
CI users (light gray squares) and normal hearing individuals (black diamonds) are plotted as a function
of time-in-sound ranging from 10 to 19 years.

71
3.5.4 Cortical abnormalities in the 150 to 300ms latency range is experience
dependent
In Figure 3.5A., the amplitude differences of all responses in the 150 to 300ms latency range
(i.e., P2-N2 latency range) are plotted against total time-in-sound for the responses recorded from all
79 CI users in this study. Linear regression analyses indicate that increases in amplitudes emerge in
the CI group, in this time window, with increasing time-in-sound experience (t(1, 77)=3.6, p<0.01). The
type of CI device used did not have an effect on these changes in amplitudes over time (t(2, 76)=-0.4,
p>0.05).
Changes in latency and amplitude of the peaks in this later latency range with time-in-sound
were assessed for 11 CI users and 11 normal hearing children who had between 10 to 13 years time-
in-sound, and for 15 CI users and 22 normal hearing children between 13 to 16 years. Figure 3.5B.
plots the absolute latencies and amplitudes of late latency peaks in responses from children who had
at least 10 years of time-in-sound experience. As shown, the latency of P2 is significantly increased
from normal at 10 to 13 years time-in-sound (when peaks P1 and N1 emerge) (t(6.7)=4.2, p<0.01), but
decreases with more auditory experience and is similar to normal values by 13 to 16 years (t(16.6)=1.2,
p>0.05). By contrast, the amplitude of peak P2 remains abnormally large in CI users even with 13 to
16 years of time-in-sound experience (t(15.9)=2.5, p<0.05).

72
Figure 3-5: Difference in latency and amplitude of the P2-N2 complex as a function of time-in-sound
compared to normal hearing peers.
A. Amplitude differences between CI (n=79) and the normal mature grand mean response were
calculated between 150 and 300ms (amplitude x 50 latency measurements) (µV x ms) and plotted
against time-in-sound. B. Mean ± standard deviations of the P2 and N2 absolute latencies and
amplitudes of CI users (light gray squares) and normal hearing individuals (black diamonds) are plotted
as a function of time-in-sound ranging from 10 to 19 years.

73
3.6 Discussion
In this study, we assessed whether cortical auditory development reaches maturation with long-
term unilateral CI use in children who had fairly short periods of bilateral auditory deprivation prior
to implantation. We found that auditory-evoked cortical waveforms approached normal-like
responses in children with at least 10 years of CI use, suggesting evidence of continuing cortical
development with auditory experience throughout adolescence. Importantly, a small normal-like P1-
N1 complex in the 50 to 150ms latency range, followed by a large and prominent P2-N2 complex
between 150 to 300ms was consistently observed in children who had over 10 years of time-in-sound
experience, but differences from normal persisted over time in latencies encompassing the P2
component.
3.6.1 Cortical activity early in development is similar between cochlear implant
users and their normal hearing peers
Analyses of cortical waveforms shown in Figures 3.2 and 3.3 indicate that early in
development, the cortical responses of CI users are dominated by a large and positive peak. This
positive peaked response (Ponton, Eggermont et al. 2000; Ponton and Eggermont 2001; Ponton,
Eggermont et al. 2002; Sharma, Dorman et al. 2002; Eggermont and Ponton 2003; Sharma and
Dorman 2006; Gordon, Valero et al. 2008; Sharma, Nash et al. 2009; Gordon, Jiwani et al. 2011;
Gordon, Tanaka et al. 2011) emerges as early as one month following CI activation (Sharma and
Dorman 2006) and persists with CI use up to ~10 years (Ponton, Eggermont et al. 2000; Ponton and
Eggermont 2001). These results are similar to previous reports indicating the dominance of this
response following early auditory experience (Ponton, Eggermont et al. 2000; Ponton and Eggermont
2001; Ponton, Eggermont et al. 2002; Sharma, Dorman et al. 2002; Eggermont and Ponton 2003;
Sharma and Dorman 2006; Gordon, Valero et al. 2008; Sharma, Nash et al. 2009; Gordon, Jiwani et
al. 2011; Gordon, Tanaka et al. 2011). This large positive peak is also seen in normal hearing children
early in development (Ponton and Eggermont 2001). It has been suggested that rapid decreases in the
latency of this large positive peaked response occur within the first 3 months following initial
stimulation with a CI in children who receive an implant by 3.5 years of age (Sharma and Dorman
2006; Sharma, Nash et al. 2009), reaching age-appropriate normal values within 6 to 9 months of

74
chronic CI use (Sharma, Dorman et al. 2002; Sharma, Dorman et al. 2002; Sharma and Dorman 2006;
Sharma, Nash et al. 2009). This suggests that when a CI is provided within a sensitive period, the
development of the auditory cortex in CI users who are younger than 10 years of age is similar to
normal (Eggermont and Ponton 2003; Sharma and Dorman 2006).
Investigators have suggested that the large amplitude positive peak in the cortical responses of
young children with normal hearing may either reflect excitatory activity in thalamo-cortical projections
to deep layers of the auditory cortex (Liegeois-Chauvel, Musolino et al. 1994), or may reflect a
convergence of temporally overlapping neural sources which generate both the adult peaks P1 and P2
(Ponton and Eggermont 2001). It is possible that this peak reflects immaturity in the auditory cortex,
since peak amplitudes are known to be largest earlier in development, and decrease with age as auditory
cortical sources mature (Ponton and Eggermont 2001). Alternatively, it is also possible that this large
positive peaked response may be synonymous to the adult peak P2. Wunderlich, Cone-Wesson and
Shepherd (2006) suggested that, in normal hearing newborns and young infants, the large amplitude
positive peak is more likely to be a P2 peak arising from deep cortical activity (Ponton and Eggermont
2001) in the non-lemniscal auditory pathways (Crowley and Colrain 2004). Since the generators of
peak P2 have an early maturational time course, it may indeed be that this large positive peaked
response reflects auditory activity driven from the association auditory cortex and the reticular
activating system, rather than from Heschl’s gyrus (i.e., P1 generator), which is known to have a slower
rate of maturation (Liegeois-Chauvel, Musolino et al. 1991; Kraus, McGee et al. 1992; Liegeois-
Chauvel, Musolino et al. 1994; Godey, Schwartz et al. 2001). Multi-channel recordings of electrically-
evoked cortical activity in children would be required to localize the sources of activity generating this
large response early in development. Nevertheless, data from the present study shown in Figure 3.2
indicates that the positive peak in CI users appears at similar latencies and amplitudes at very early
stages of implant use and thus likely has the same neural generators as normal. This is further
supported by our finding that the changes in this response over time are similar to normal.
As shown in the amplitude difference analyses of Figures 3.3A and 3.3B, the positive peak
decreased in amplitude over the first 7 years of time-in-sound in children using CIs, revealing
decreasing differences from the normal hearing mature response with auditory experience. The similar
peak in normally hearing children also decreases in amplitude and latency with age, as the auditory
system matures (Eggermont and Ponton 2003; Sharma and Dorman 2006). We know from normal
hearing data that maximum synaptic density in the auditory cortex is reached by ~3 months of age

75
(Huttenlocher and Dabholkar 1997). Thus, the large amplitude positive peaked cortical response
reported from normal hearing infants (Wunderlich, Cone-Wesson et al. 2006) and recorded from CI
users at initial stages of hearing experience may reflect the high synaptic density which is present in
early cortical development (Huttenlocher and Dabholkar 1997; Wunderlich and Cone-Wesson 2006;
Wunderlich, Cone-Wesson et al. 2006; Picton and Taylor 2007). Moreover, because the amplitude of
a peak is known to reflect synaptic density and efficiency (Eggermont 1988; Picton and Taylor 2007),
the reduction in amplitude in the early latency range (50 to 150ms) with time-in-sound (Figures 3.3
and 3.4) could reflect decreasing numbers of activated cells and synapses contributing to the response.
Synaptic pruning begins in late childhood (Huttenlocher and Dabholkar 1997; Ponton,
Eggermont et al. 2000) and is thought to be complete by ~12 years of age in the auditory cortex,
resulting in an abrupt decrease of synaptic density (Huttenlocher and Dabholkar 1997). Thus, it has
been suggested that the decrease in amplitude of the cortical response in children with normal hearing
up to 7 years of age likely reflects this reduction in synaptic density (Wunderlich, Cone-Wesson et al.
2006; Picton and Taylor 2007). Furthermore, neural maturation in childhood, characterized by changes
in the location and orientation of neural substrates in the auditory cortex may also influence the
amplitude of the cortical response (Wunderlich, Cone-Wesson et al. 2006). In addition, other
mechanisms may also affect the decrease in amplitude of cortical wavepeaks in early development.
Increased axon myelination and changes in synaptic mechanisms with development might contribute
to changes in the cortical waveform morphology occurring with time and auditory experience in
normal hearing children (Sharma, Kraus et al. 1997; Albrecht, Suchodoletz et al. 2000). Thus, as
neurotransmission becomes more efficient with auditory development, the observed decreases in
amplitude over the first 7 years of time-in-sound experience could reflect an ongoing process of
synaptogenesis in CI users that is similar to normal. These findings are consistent with previous reports
of decreasing amplitudes of the auditory evoked cortical response reported from normal hearing
children in this same time frame (Albrecht, Suchodoletz et al. 2000; Ponton, Eggermont et al. 2000;
Wunderlich and Cone-Wesson 2006; Wunderlich, Cone-Wesson et al. 2006).

76
3.6.2 Long-term cortical development follows a normal-like trajectory with
time-in-sound
The data presented in Figures 3.2 and 3.3A suggest that a developmental milestone is
occurring in the auditory cortex around 10 years of time-in-sound in children using CIs. Indeed, by
10 years, the auditory evoked cortical response recorded in all such children had developed into a
mature polyphasic P1-N1-P2-N2 waveform. This is in line with the time-course of development of the
mature polyphasic P1-N1-P2-N2 cortical waveform in normal hearing adolescents, illustrated in Figure
3.2 and previously reported by other groups as well (Ponton, Eggermont et al. 2000; Ponton and
Eggermont 2001; Wunderlich and Cone-Wesson 2006; Wunderlich, Cone-Wesson et al. 2006). Figure
3.3B shows that differences from the normal hearing mature response are largest in children who have
less than 7 years of time-in-sound experience. However, as peak N1 develops, the cortical response in
the 50 to 150ms latency range approaches the normal hearing mature waveform in all children who
had more than 10 years of hearing experience (areas of dark blue in Figure 3.3B), indicating minimal
differences from normal.
Linear regression analyses shown in Figures 3.4A and 3.4B indicate that the cortical responses
in the early latency range significantly change over time (R=0.39). Children who had limited duration
of bilateral deafness and had longer durations of auditory experience produced cortical waveforms that
approached the normal hearing response in the 50 to 150ms latency range (i.e., P1 and N1 latency
range). The moderate strength of correlation reflects inter-subject variability, which should be
expected, given that auditory development after implantation will depend on multiple factors (Holden,
Skinner et al. 2002; Skinner, Holden et al. 2002; Nicholas and Geers 2007; Geers and Sedey 2011).
The data in Figure 3.4B shows that both the latencies and amplitudes of peaks P1 and N1 reach age
appropriate values in children with more auditory experience. Since the development of activity-
dependent processes in the brainstem tracts do not mature without significant auditory input (Gordon,
Papsin et al. 2003; Gordon, Papsin et al. 2006; Gordon, Papsin et al. 2007; Gordon, Valero et al. 2007;
Gordon, Valero et al. 2008; Gordon, Jiwani et al. 2011), these cortical changes are very likely due to
the exposure to auditory input with CIs. Chronic auditory stimulation provided by a CI over the long-
term may contribute to the establishment and/or strengthening of activity-dependent synaptic
connections between neurons and perhaps increased myelination of nerve fibres throughout the

77
central auditory pathways. These mechanisms, known to increase neural synchronization (Gilley,
Sharma et al. 2005), may underlie the emergence of the P1 and N1 peaks in the evoked response.
It has been suggested that, although P1 generators are adult-like by 8 years of age and P2
generators are fully mature by 5 years of age (Albrecht, Suchodoletz et al. 2000; Ponton and Eggermont
2001), it is only at ~10 years of age that axons in superficial layers of the auditory cortex reach maturity
(Ponton, Eggermont et al. 2000; Moore and Guan 2001; Moore and Linthicum 2007). Interestingly,
it is also around this time (i.e., 9 to 12 years) that the cortical responses of normal hearing children and
CI users begin to develop into adult-like peaks (the P1-N1-P2-N2 complex) as shown in Figures 3.2,
3.3, and 3.4. This polyphasic waveform was identifiable only in children who had more than 10 years
of auditory experience and suggests that long-term CI users experience analogous cortical maturational
changes to those of their normal hearing peers at this stage of auditory processing. If the N1 reflects
re-entrant activity in thalamo-cortical and cortico-cortical pathways in the superficial layers of the
auditory cortex (Ponton and Eggermont 2001), then it is possible that these areas in the brain are
maturing in children who have used a CI for over 10 years, despite earlier concerns that this
development might never occur in children using implants (Ponton and Eggermont 2001; Eggermont
and Ponton 2003). We intend to explore this further using multi-channel recordings in future work to
assess the maturation of cortical sources in long-term CI users and their normal hearing peers.
Evidence of development of peak N1, perhaps reflecting maturation in superficial layers of the
cortex in children using CIs is encouraging, because these areas are essential for thalamo-cortical and
cortico-cortical maturation. Thalamo-cortical connections play a crucial role in the transfer of primary
(auditory specific) and non-primary (multi-sensory) auditory input from the medial geniculate body in
the thalamus (Razak, Zumsteg et al. 2009) to the ipsilateral and contralateral auditory cortices (Ponton,
Eggermont et al. 2000; Winer, Diehl et al. 2001; Winer, Miller et al. 2005; Razak, Zumsteg et al. 2009).
Cortico-cortical connections on the other hand, mediate the transfer of information between auditory
cortices in both hemispheres via commissural fibres and between the auditory cortex and other primary
sensory areas (Lee and Winer 2005; Winer, Miller et al. 2005; Klinge, Eippert et al. 2010). Thus, the
appropriate development of these connections in the brain has important implications for: 1) the relay
of auditory information from the outside world to the cortex; 2) communication between the two
hemispheres; and 3) the connectivity between different sensory areas. In addition, it has been
suggested that in normal hearing individuals, the maturation of these thalamo-cortical and cortico-
cortical auditory processes may be related to the development of complex auditory perceptual skills

78
which occur during adolescence, such as discriminating speech in the presence of noise and
understanding degraded speech input, among other functional auditory skills (Ponton, Eggermont et
al. 2000; Eggermont and Ponton 2003). Since the emergence of peak N1 coincides with the
maturational time course of these processes, it is very likely that similar underlying developmental
mechanisms are also taking place in long-term CI users who have over 10 years of auditory experience.
In line with the time course of development of these skills, behavioural studies have shown significant
and continued improvement on long-term language comprehension and verbal reasoning skills in early
implanted children who used a CI for over 10 years, compared with their performance on language
tests in early elementary grades (Geers and Sedey 2011). Thus, while auditory development of early
cortical wave peaks is likely dependent on both time-in-sound and age; it is possible that the duration
of auditory experience bears more weight, at least for the development of superficial cortical layers.
The findings of the present study indicate that the N1 peak, and in turn, mature adult-like auditory
cortical responses, do develop with long-term CI stimulation. These changes likely support the
remarkable improvements in auditory function seen over time in children using CIs (Beadle, McKinley
et al. 2005).
3.6.3 Differences from normal in the later cortical peaks may reflect increased
cortical activity from non-auditory modalities
While the mature polyphasic cortical waveform does emerge with chronic long-term CI
experience, differences in later latencies occur over time in children with CIs, relative to their normal
hearing peers. Specifically, responses in the later latency range, between 150 to 300ms, became
significantly larger than normal in children with more than 10 years of auditory experience, as shown
in Figures 3.2 and 3.3. These deviations from normal were confirmed in the regression analysis
(R=0.38) of Figure 3.5A and in the abnormally large amplitude values of the P2 component (p<0.05)
identified in Figure 3.5B. It is possible that the degree of residual hearing prior to implantation may
not have sufficiently promoted development in the neural generators of these later latency peaks,
leaving aberrant or altered development of activity in deep auditory cortical layers (generating peaks P2
and N2). It is also possible that the long-term absence of binaural hearing through CIs in this cohort
impeded the normal maturation of peak P2 generators.

79
Increases in the amplitude of peak P2 reflect more demand for attention and arousal while
hearing (Picton and Hillyard 1974; Rif, Hari et al. 1991; García-Larrea, Lukaszewicz et al. 1992; Novak,
Ritter et al. 1992; Kraus and McGee 1993; Ponton and Eggermont 2001; Tremblay, Kraus et al. 2001;
Moller and Rollins 2002; Tremblay and Kraus 2002; Eggermont and Ponton 2003; Alain, Snyder et al.
2007; Tremblay 2007; Tremblay, Shahin et al. 2009); increased recruitment of multi-sensory input,
including vision and touch, during complex decision-making processes (Hocherman, Benson et al.
1976; Hari 1990; Kraus, McGee et al. 1992; Kraus and McGee 1993; Grady, Van Meter et al. 1997;
Webster and Colrain 2000; Moller and Rollins 2002; Crowley and Colrain 2004; Busse, Roberts et al.
2005; Plailly, Howard et al. 2008; Kraus and Chandrasekaran 2010; Rinne 2010); and increased
responsiveness to the external environment during sleep (Weitzman and Kremen 1965; Ornitz, Ritvo
et al. 1967; Kraus, McGee et al. 1989; Kraus, McGee et al. 1992; Kraus and McGee 1993; McGee,
Kraus et al. 1993; Coull 1998; Colrain, Webster et al. 1999; Colrain and Campbell 2007). Enhanced P2
amplitudes observed following auditory training in normal hearing adults (Tremblay, Kraus et al. 2001;
Tremblay and Kraus 2002; Tremblay 2007; Tremblay, Shahin et al. 2009), suggests that focused
auditory learning increases the number of cortical neurons responding to the sound with improved
neural synchrony (Tremblay 2007; Tremblay, Shahin et al. 2009), and may reflect modulatory
influences from mechanisms of selective attention/arousal and other top-down processes (Alain,
Snyder et al. 2007; Tremblay, Shahin et al. 2009). These are the very processes which are known to
influence the amplitudes of peaks N1 and P2 (Picton and Hillyard 1974) and lead to improved auditory
perception (Shinn-Cunningham and Best 2008). Underlying generators could involve association
auditory areas and auditory activity driven from the reticular activating system, which includes frontal
and parietal areas (Hari 1990; Rif, Hari et al. 1991; Kraus and McGee 1993; Posner and Dehaene 1994;
Ponton and Eggermont 2001; Moller and Rollins 2002; Eggermont and Ponton 2003). It has been
suggested that these pathways supplement the primary auditory network for complex processing of
sound (Hocherman, Benson et al. 1976; Grady, Van Meter et al. 1997; Busse, Roberts et al. 2005). It
is therefore possible that the larger than normal amplitude of peak P2 observed in long-term CI users
reflects increased activity in the distributed cortical network involved in selective attention/arousal and
multi-sensory integration during auditory processing.
The larger than normal amplitude of peak P2 observed in the present study could reflect
increased recruitment/interaction between auditory and non-auditory multi-sensory processes (i.e.,
vision and/or touch) by children using CIs. Such children have been shown to use multi-sensory

80
information more effectively than their peers with normal hearing to facilitate the development of
complex auditory working memory skills (Brenner & Geers, 2011), spoken language comprehension
and complex auditory processing, such as understanding subtle emotional cues in speech (Hopyan-
Misakyan, Gordon et al. 2009; Hopyan, Gordon et al. 2011). Furthermore, functional neuroimaging
studies have also shown that activation of the visual cortex (Giraud, Price et al. 2001) and the posterior
portions of the auditory association areas (Nishimura, Hashikawa et al. 1999) increase during listening
tasks in long-term CI users, particularly when given auditory and visual input simultaneously
(Nishimura, Hashikawa et al. 1999). Despite receiving auditory information, users of CIs still rely on
contributions from non-auditory modalities when listening and processing speech (Nishimura,
Hashikawa et al. 1999; Finney, Fine et al. 2001; Giraud, Price et al. 2001; Doucet, Bergeron et al. 2006).
Together, these data suggest that CI users must recruit attentional resources and integrate multi-
sensory input to a greater than normal degree. This would help to compensate for: 1) the
reorganization of the developing brain when faced with sensory deprivation prior to cochlear
implantation; 2) the imperfect auditory signal provided by a CI to improve auditory perception; 3)
auditory input in only one ear, compromising binaural hearing and causing possible reorganization in
the pathways that were deprived of auditory input for many years.
3.7 Conclusion
It is hoped that early cochlear implantation will lead to normal maturation of the central
auditory system. In the present study, we assessed cortical activity in children and adolescents who
were deaf and received a unilateral CI during childhood after a limited duration of deafness. The CI
users studied had experienced up to sixteen years of time-in-sound. We found that, over the long-
term, their auditory responses matured into waveforms which were very similar to those of normal
hearing adolescents/young adults. This indicates that at least some degree of auditory development
proceeds normally with CI use. However, differences from the mature normal hearing cortical
response persist, even after sixteen years of auditory experience, suggesting slight alterations to normal
cortical processing. We question whether some of these differences reflect an increased dependence
on attentional resources or recruitment of multi-sensory input for complex auditory processing.

81
Nonetheless, the present findings provide encouraging evidence for long-term auditory development
in CIs users who were implanted as children after limited bilateral deafness.

82
4. Chapter Four – Early unilateral cochlear implantation promotes
mature cortical asymmetries in adolescents who are deaf
This paper was submitted to Human Brain Mapping using required journal formatting:
Jiwani, S., Papsin, B.C., Gordon, K.A. Eearly unilateral cochlear implantation promotes mature
cortical asymmetries in adolescents who are deaf.
4.1 Abstract
Unilateral cochlear implant (CI) stimulation establishes hearing to children who are deaf, but
compromises bilateral auditory development if a second implant is not provided within 1.5 years. In
the present study we are asking: 1) what are the cortical consequences of missing this early sensitive
period in adolescents who have developed a mature auditory cortex with unilateral implant
stimulation?, 2) what are the effects of unilateral deprivation on pathways from the opposite ear?
Cortical responses were recorded from 64-cephalic electrodes within the first week of bilateral
CI activation in adolescents who had over 10 years of unilateral implant experience and in normal
hearing peers. Cortical activation underlying the evoked peaks was localized to areas of the brain using
beamformer imaging.
Findings indicated stronger activity in the contralateral left hemisphere with stimulation from
the right CI. This could be driven by a reduction in inhibitory processes from the deprived side, and
indicate that cortical abnormalities remain in adolescents despite having developed a mature auditory
cortex with unilateral implant stimulation. Providing a second CI to the opposite and deprived ear
thereafter resulted in abnormal cortical responses with abnormally large and widespread dipole activity
across the cortex, indicating abnormal changes in the un-stimulated pathways. Thus, using a unilateral
CI to hear beyond the period of cortical maturation causes lasting asymmetries in the auditory system
and does little to protect the cortical pathways from effects of auditory deprivation, perhaps reflecting
a closing of a sensitive period for restoring auditory development on the deprived side.

83
4.2 Introduction
Cochlear implants (CI) are surgically implanted auditory prostheses which restore hearing to
individuals who are deaf. While CIs allow children to develop remarkable oral speech and language
abilities, their hearing is significantly poorer than normal because the CI provides only a crude
representation of acoustic sounds and eliminates important cochlear processing. Prior to cochlear
implantation, the absence of auditory input in early life leaves the auditory brain vulnerable to cross-
modal recruitment (Finney, Fine et al. 2001; Bavelier and Neville 2002; Bavelier, Dye et al. 2006) by
the visual (Lomber, Meredith et al. 2010) and somatosensory systems (Lomber, Meredith et al. 2010;
Meredith and Lomber 2011). If hearing is not restored during sensitive periods in development, this
reorganization will impair hearing with a CI (Lee, Lee et al. 2001). CIs have thus been provided to
children within limited durations of bilateral deafness with the aim to halt and hopefully reverse any
such effects of deafness on the brain.
Traditionally, CIs have been provided to children in only one ear, which put them at risk for
language delays and educational difficulties (Bess and Tharpe 1984; Bess and Tharpe 1986). While it
is clear that unilateral stimulation with an implant promotes auditory maturation in the brainstem
(Gordon, Papsin et al. 2006; Gordon, Jiwani et al. 2013), midbrain (Gordon, Papsin et al. 2005) and
cortex (Jiwani, Papsin et al. 2013), depriving the opposite pathways of auditory input might leave the
auditory system susceptible to deafness-induced reorganization. In turn, this could distort bilateral
auditory development as well as the ability to restore binaural hearing with a second implant at a later
time, perhaps permanently (Gordon, Jiwani et al. 2011; Gordon, Jiwani et al. 2013). We must therefore
determine whether there is an age at which it is too late to re-establish function in the opposite
pathways with a second implant.
Graham and colleagues suggested that the mid-teenage years could mark the end of a critical
period for implanting the non-implanted side in adolescents who are congenitally deaf and have used
a unilateral CI to hear for most of their lives (Graham, Vickers et al. 2009; Graham and Vickers 2011).
Adolescents who received a CI in the deprived ear after 15 years of age had significantly worse speech
perception abilities compared to their younger bilaterally implanted peers (Peters, Litovsky et al. 2007;
Graham, Vickers et al. 2009; Graham and Vickers 2011). Compromised benefit from spatial separation
of noise (Chadha, Papsin et al. 2011), and reduced sound localization (Van Deun, Van Wieringen et al.
2009; Grieco-Calub and Litovsky 2010) and lateralization (Salloum, Valero et al. 2010) abilities have

84
also been reported in children who received a second implant after long delays. Moreover, asymmetries
in auditory function increased as the duration of unilateral implant experience increased (Gordon and
Papsin 2009; Illg, Giourgas et al. 2013). This led to inconsistent use of the second implanted ear
(Fitzgerald, Green et al. 2013) and reduced binaural hearing abilities relative to children who received
bilateral implants within limited delays (Gordon and Papsin 2009; Van Deun, Van Wieringen et al.
2009; Grieco-Calub and Litovsky 2010; Salloum, Valero et al. 2010; Gordon, Jiwani et al. 2011). Poor
performance of older children with their second implant may be attributed to increased dependency
on the first and earlier implanted ear for hearing (Kral, Hubka et al. 2013), decreased motivation or
emotional resistance to change in teenagers (Fitzpatrick and Irannejad 2008), or decreased auditory
plasticity with maturation (Lohmann and Kessels 2014).
The latter issue, the remarkable capacity of the developing brain to change, has been studied
in animal models of unilateral deafness. Abnormally strengthened afferent projections from the
cochlear nucleus of the stimulated ear to the ipsilateral (Moore and Kowalchuk 1988; Kil, Hkageyama
et al. 1995; Kitzes, Kageyama et al. 1995; Nordeen, Killackey et al. 2004) and contralateral brainstem
(Kil, Hkageyama et al. 1995; Kitzes, Kageyama et al. 1995) were observed in neonate animals following
unilateral cochlear ablation. Cortical changes have also occurred as a result of unilateral hearing loss;
connections in the neural circuitry of the primary auditory cortex from the hearing ear increased,
whereas those of the opposite and deprived ear were weakened (Popescu and Polley 2010; Kral, Hubka
et al. 2013). These changes were not found when the loss was bilateral (Moore 1990) or induced later
in life (Nordeen, Killackey et al. 1983; Moore and Kowalchuk 1988; Nordeen, Killackey et al. 2004;
Popescu and Polley 2010; Kral, Hubka et al. 2013). Similar asymmetries were found in children using
unilateral CIs when the period of unilateral stimulation/deprivation exceeded 1.5 years. Faster
response latencies in the brainstem (Gordon, Salloum et al. 2012) reflected a strengthening of pathways
from the more experienced ear; and abnormally strong activity driven by the first CI to both auditory
cortices (Gordon, Wong et al. 2013; Kral, Hubka et al. 2013) indicated a strengthening of inputs in the
immature auditory pathways from the unilaterally implanted right ear. This has been explained by a
disruption in the normal organization of inhibitory-excitatory activity when input to both ears are
unbalanced (Mossop, Wilson et al. 2000; Nordeen, Killackey et al. 2004; Popescu and Polley 2010),
resulting in a loss of inhibitory processes (Vale, Juíz et al. 2004; Kotak, Fujisawa et al. 2005; Takesian,
Kotak et al. 2009) that would have normally occurred with bilateral hearing (Grothe, Pecka et al. 2010).
Because the cortical changes observed in the children using unilateral CIs occurred over approximately

85
the same time-course of CI-driven brainstem maturation (~1.5 year) (Gordon, Papsin et al. 2006), an
important human developmental period for bilateral input may have been missed (Gordon, Wong et
al. 2013).
Adolescence marks the maturation of the auditory cortex (Eggermont 1988; Ponton,
Eggermont et al. 2000; Moore and Linthicum 2007; Jiwani, Papsin et al. 2013) which could signal the
end of another important developmental period. Much of the brain is maturing during this time.
There is a slowing of white matter increase and a sharp decline in gray matter during adolescence
(Lebel and Beaulieu 2011) that likely reflects synaptic elimination (Giedd, Blumenthal et al. 1999;
Sowell, Thompson et al. 2001; Blakemore 2012), as existing connections in the mature brain are refined
(Blakemore and Choudhury 2006; Lohmann and Kessels 2014) and as each cortical hemisphere
becomes specialized (Davidson 1984; Zatorre and Belin 2001; Le Grand, Mondloch et al. 2003; Toga
and Thompson 2003; Rivera, Reiss et al. 2005; Gotts, Jo et al. 2013). In the auditory brain, the left
cortex becomes tuned to temporal processing, whereas association regions of the right auditory cortex
become biased to spectral variations of sound (Zatorre and Belin 2001; Schönwiesner, Rübsamen et
al. 2005; Jamison, Watkins et al. 2006).
Maturation driven by CI stimulation, particularly when only from one ear, could be very
different from normal. Whilst long-term speech and language outcomes in children with a single CI
show abilities which approach age-appropriate levels (Geers, Tobey et al. 2008; Geers and Sedey 2011;
Geers and Sedey 2011), they still face challenges as they mature including increased effort/attention
needed to hear (Lee, Giraud et al. 2007; Hopyan-Misakyan, Gordon et al. 2009; Pisoni, Conway et al.
2010; Sandmann, Dillier et al. 2012; Gordon, Jiwani et al. 2013; Kronenberger, Colson et al. 2014).
Having provided CIs to children for over two decades, we are now in a position to ask how their
auditory system has matured with the input provided from this unique device. Many such adolescents
have recently been seeking a second implant for their long-term deprived ear in the hopes of deriving
benefits of binaural hearing, decreasing listening effort and improving communication. This provided
a unique opportunity to stimulate the long-term deprived pathways for the first time and study the
plastic effects of long-term unilateral implant stimulation/deprivation in the adolescent brain. Given
the long durations of deprivation in the non-implanted ear and their older ages, we expect unique
cortical effects in this group of new bilateral implant users.

86
In the present study, we explored the underlying consequences of driving auditory maturation
in the brainstem and cortex with CI stimulation from only one ear. We asked: 1) how does missing an
early sensitive period for bilateral input (i.e., brainstem) and driving cortical maturation from one ear
compromise the normal development of the auditory cortex in adolescents who have used a unilateral
CI to hear for most of their lives?, and, 2) what are the effects of unilateral deprivation on pathways
from the opposite ear? Results indicate that allowing the auditory cortex to mature with over a decade
of unilateral CI use drives lasting asymmetries in the auditory system and does not protect the opposite
pathways from deafness-induced cortical changes. While the stimulated right ear strengthens activity
to the contralateral left hemisphere over the long-term, the deprived pathways from the left ear recruit
abnormally large cortical regions in the brain. Establishing binaural hearing may thus be challenging
after unilaterally-driven maturation of the auditory cortex.
4.3 Materials and Methods
4.3.1 Participants
Thirty-four adolescents who received one CI in their right ear by 3.2 ± 1.3 years of age after a
limited period of bilateral auditory deprivation (1.8 ± 1.3 years) participated in this study. They had
12.4 ± 1.7 years of unilateral CI experience at the time of the test. All were successful implant users
and did not ever receive auditory stimulation in their opposite ear. Twenty-one of these adolescents
later received a second implant in their opposite-left ear after 12.0 ± 2.1 years of unilateral deprivation
on that side. Age at implantation on this second implanted side was 15.9 ± 2.0 years. Responses were
recorded from the 21 bilateral CI recipients on the first day of activation of the second implant to
study the long-term cortical effects of single-sided deafness on the deprived pathways in the mature
auditory system. This provided us with a unique opportunity to stimulate the long-term deprived ear
for the first time and study the plastic effects of long-term unilateral implant stimulation/deprivation.
Etiology of deafness varied. Eight adolescents were diagnosed with a genetic anomaly; 2 had a 35delG
mutation on the GJB-2 gene; 3 lost their hearing after a meningitis infection; 2 were diagnosed with a
Mondini malformation (1 with hypoplasia of the vestibulocochlear nerve and 1 with KID syndrome)
and 3 had enlarged vestibular aqueducts. The other 16 adolescents had a congenital hearing loss of
unknown etiology. All adolescents were implanted with a Cochlear Nucleus device from Cochlear

87
Corporation Limited (4 N22 implants, 20 CI24M implants, 10 CI24R(CS) devices) in the right-
experienced ear. In the newly implanted side, 16 received a CI24RE(CA) implant and 6 were implanted
with a CI513. Of all 34 adolescents tested, 24 were boys and 10 were girls. All but 2 adolescents were
right handed. The control group was composed of sixteen adolescents (6 boys : 10 girls) who had
normal hearing in both ears. Adolescents in this group were 15.9 ± 6.4 years of age at the time of the
test and were matched for chronological age and duration of hearing experience.
4.3.2 Recording cortical responses
Cortical responses were evoked by each ear/implant separately using 64-cephalic electrodes
and referenced to the right earlobe. In the CI group, responses were generated using biphasic electrical
pulse trains of 250 pulses per second, lasting 36ms and delivered by a single electrode (#20) at the apex
of the electrode array. These electrical pulse trains were presented at a rate of 1Hz. Similar current
levels were presented from each CI at comfortably loud levels. These levels were determined as in
previous studies (Gordon, Wong et al. 2013) by recording auditory brainstem responses at increasing
intensities, as shown in the example in Figure 4.1. Electrically-evoked brainstem responses recorded
from the experienced and newly implanted sides separately are shown in response to electrical pulses
presented at 11Hz. Current levels were increased within a comfortable range of intensities for the
participant. The maximum levels at which wave eV amplitudes on each side were equal were used to
evoke cortical responses. Levels were reduced by 10 Clinical Units on each side so that stimulation
would be provided at a slightly reduced part of the dynamic range.

88
Cortical responses evoked in CI users were compared with the same responses recorded from
the 16 adolescents with normal hearing. Responses in normal hearing peers were evoked by 500Hz
tone-bursts. This stimulus was chosen because this frequency was allocated to the same apical
electrode (#20) of the CI device used to evoke responses in all CI participants. The same stimulus
duration (36ms) and rate of presentation (1Hz) was used as in the CI users. The tone was enveloped
Figure 4-1: Balanced stimulus levels between the experienced and newly implanted ears determined
by matching peak eV amplitude of the brainstem response.
A. Electrically evoked auditory brainstem responses recorded from one adolescent. Right ear had
13.35 years of CI experience. Left ear had 6 days of stimulation. Current levels on the experienced
side were increased to maximize the amplitude of the brainstem response within a comfortable
listening range. Current levels delivered to the naïve-left CI were increased until the wave eV amplitude
of the responses matched the maximum of the wave eV response from the experienced side. B. Peak-
to-peak amplitude of wave eV for responses recorded on the experienced-right and naïve-left sides for
different stimulus intensity levels. Matched wave eV responses are marked by the dashed line. The
corresponding current levels – 10 Clinical Units were used to stimulate each CI separately and record
the cortical evoked response.

89
with a Tukey window over the first and last eights to minimize effects of high frequency onset and
offset. Tone-bursts were delivered at 40dB above behavioural threshold and presented to the right
and left ears separately using ER3-14A insert earphones. Responses for all participants were recorded
using the NeuroScan-4.3 system with a Synamps-II amplifier. They were sampled at a rate of 1000Hz
and an online band-pass filter between 0.15 to 100Hz was applied. A minimum of 400 sweeps with at
least two visually replicable cortical responses were obtained. Epochs which were greater than ±100µV
between the 100 to 800ms latencies were rejected.
4.3.3 Localization of cortical evoked peaks
The Time Restricted, Artefact and Coherence Source suppression (TRACS) linearly
constrained minimum variance beamformer (Wong and Gordon 2009) was used to localize activity
underlying the evoked peaks in the cortex in response to stimulation from the experienced and naïve
CIs, and right and left ears in normal hearing peers. Suppression of the electrical artefact from the CI
device (Wong and Gordon 2009) and region suppression to eliminate coherent source interference
were applied (Dalal, Sekihara et al. 2006). See Supporting Information for additional details.
Covariance was estimated over latency windows encompassing each peak of the cortical response for
each individual (Van Veen, Van Drongelen et al. 1997). A pseudo-Z statistic was used to normalize
the signal-to-noise ratio of each 3 x 3 x 3 mm voxel. Once sources were localized using this lead
normalization process, the strength of source activity, measured in dipole moments (nAm), was
computed for the peaks of the cortical waveform. A one-tailed omnibus-noise T-test (Petersson,
Nichols et al. 1999) was used to calculate a statistical threshold pseudo-Z value (p≤0.0005) reflecting
baseline brain activity. Only voxels with pseudo-Z activity greater than this omnibus value were
accepted and used in the analyses.
Dipole activity was visualized topographically on age-appropriate head model templates
derived from the Montreal Neurological Institute (MNI) magnetic resonance imaging (MRI) library
for each adolescent in 63,646 voxels in brain space. Dipole strength was visualized as colours from
dark blue, reflecting weaker dipole activity, to dark red, indicating stronger dipoles. Mean beamformer
brain images were created to assess areas of cortical activity which were consistently evoked by sound
for each ear/implant. Two-sided uncorrected paired permutation tests were used to compare voxel-

90
by-voxel dipole activity in 2048 permutations (Blair and Karniski 1993; Chau, McIntosh et al. 2004) to
assess differences between the left and right hemispheres. Significant absolute differences (p<0.005)
were plotted on topographic brain maps for each cortical wavepeak. Un-paired permutation analyses
to assess differences in activity for each voxel-pair between ears and between each group was calculated
to measure aural preference and group differences for each cortical wavepeak. Significant absolute
differences at p<0.01 were considered significant for these analyses. The voxels reflecting the
strongest dipoles in the left and right auditory cortices were then extracted for each cortical wavepeak
in each individual. The location of the auditory cortex were identified within MNI coordinates (X =
<-55, Y = >-35 to <-5, Z = >-10 to <20) mm in the left hemisphere and (X = >55, Y = >-35 to <-
5, Z = >-10 to <20) mm in the right hemisphere in both the normal hearing and the CI group,
consistent with other reports (Wong and Gordon 2009; Gordon, Wong et al. 2010; Gordon, Wong et
al. 2013). Dipole locations are shown on glass brain images. Repeated measures ANOVA with
Bonferroni corrections were used to identify differences in peak dipole moment and latencies between
activity evoked in each auditory cortex and in each group. Percent cortical lateralization and aural
preference were calculated from the extracted peak dipoles using the following formulas:
% Cortical Lateralization =
(right hemisphere dipole – left hemisphere dipole) /
(right hemisphere dipole + left hemisphere dipole) x 100.
% Aural Preference =
(dipole from contralateral stimulus – dipole from ipsilateral stimulus) /
(dipole from contralateral stimulus + dipole from ipsilateral stimulus) x 100.
4.3.4 Speech perception tests to assess outcomes with CIs
Functional outcomes with CIs were assessed using the age-appropriate Phonemic Balanced
Kindergarten (PBK) monosyllabic words open-set speech perception test. This test assesses speech
recognition abilities using a list of twenty-five words. Words were presented at 0-degree azimuth in a
double-walled sound-proof booth, through a GSI-61 Grason-Stadler audiometer, using monitored-
live voice at 65dB sound pressure level. Words were presented in quiet to the first and second implants

91
separately after 8.0 ± 2.1 months (5.8 to 15.0 months) of bilateral CI use and scored as a percentage
of words that were repeated correctly (percent accuracy). Pearson correlations were used to assess
associations between speech perception scores, dipole moment and cortical lateralization.
4.4 Results
4.4.1 Tone-bursts preferentially stimulate the right auditory cortex in
adolescents with normal hearing
Cortical responses recorded at a midline-cephalic location (Cz) and mean global field power
with corresponding head topographies of potential distribution are shown in Figures 4.2A and 4.2B
for the normal hearing group. Adolescents with normal hearing produced cortical responses with
peaks P1, N1 and P2, characteristic of the mature response (Albrecht, Suchodoletz et al. 2000; Ponton,
Eggermont et al. 2000; Wunderlich, Cone-Wesson et al. 2006), when either the right (n=13) or left
ears (n=12) were stimulated by 500Hz tone-bursts. Mean latencies and amplitudes for all responses
appear in Table 4.1 in the Supplementary Information section. Responses were similarly distributed
over the scalp between the right and left ears. However, dipole moments underlying the mature
cortical peaks were stronger in the right hemisphere. This is shown by the dark red hotspots in the
right hemisphere mean topographic plots of Figure 4.2C and by the larger right (red bars) than left
cortex (blue bars) peak dipole magnitude in Figure 4.2E. Dipoles in auditory cortex showed effects
of side of stimulation (F(1,10)=5.976, p=0.035), hemisphere (F(1,10)=8.018, p=0.015) and an interaction
of these factors (F(1,10)=10.308, p=0.009) but no differences between the different cortical peaks
(F(2,20)=8.018, p>0.05). Post-Hoc t-tests with Bonferonni corrections revealed similar left hemisphere
activation when the right and left ears were stimulated separately (t(76)=-0.03, p>0.05), but confirm that
dipoles were significantly stronger in the right hemisphere for left ear stimulation (t(66.9)=-2.43, p=0.02).
Aural preference in each hemisphere, shown in Figure 4.3, was consistent with these results. Aural
preference in the left cortex was not evident in the mature auditory system (i.e., left auditory cortex
responded to both ears similarly), but a contralateral dominance remained present in the right auditory
cortex for the left ear (P1: t(10)=-4.5, p=0.001). This means that as mature cortical responses emerge,
the right hemisphere preferentially responds to tone-bursts, particularly when they are presented to
the left ear.

92

93
Figure 4-2: Cortical dipole activity evoked by auditory stimulation of the right and left ears of normal
hearing adolescents.
A. Auditory evoked cortical responses were stimulated from the right (n=13) and left (n=12) ears of
adolescents with normal hearing, aged 15.9 ± 6.4 years. Individual responses from a midline cephalic
electrode (Cz) (thin grey lines), and grand mean responses (thick black lines) show 3 peaks (P1, N1, P2),
characteristic of mature responses. B. Mean global field power is plotted for mean response data
from each ear. Topographic maps indicate similarly distributed scalp potentials (red = positive
potentials, blue = negative potentials) for the mature peaks in both ears. C. Mean peak dipole activity
evoked by each ear for each cortical wavepeak (P1, N1, P2) is plotted in each of 63,646 voxels in brain
space on age-appropriate topographic head models derived from the Montreal Neurological Institute
(MNI) MRI Library (MNI Axial orientation). Stronger activity, shown by hotspots of dark red, occurs
in the right hemisphere, particularly for left ear stimulation. D. Peaks dipoles underlying each cortical
wavepeak in the left (blue) and right (red) auditory cortices were chosen within the hotspots shown in
C. for each child. Peak dipole locations are shown on glass brain images. Axis are MNI coordinates
in millimeters. E. Mean (± 1 standard error (SE)) dipole moment strength is plotted for the left (blue
bars) and right (red bars) auditory cortices for all cortical peaks. Consistent with the topographic plots
in C., peak dipole activity is stronger in the right than left auditory cortex for stimulation of both ears,
particularly for left ear stimulation. F. Mean (± 1 SE) peak dipole latency are similar between the left
(blue) and right (red) auditory cortices and are in line with cortical wavepeak latencies shown in A. and
B.

94
Figure 4-3: Aural preference evoked by auditory stimulation of the right and left ears of normal
hearing adolescents.
A. Aural preference in each cortical hemisphere was assessed across 63,646 peak dipoles using
permutation testing in adolescents with normal hearing by comparing cortical activation with right
versus left ear stimulation. Significant absolute dipole moment differences (p<0.01) for right – left ear
stimulation are plotted for each cortical wavepeak on topographic brain plots. Voxels with dark red
colours indicate greater intra-hemispheric dipole differences between stimulation of each ear, and
those with dark blue colours reflect smaller dipole differences. P1 and N1 show large aural difference
for tone-burst stimuli, particularly in the right cortical hemisphere and frontal areas. B. Mean (± 1
SE) percent aural preference, [(dipole moment evoked from contralateral stimulus – dipole moment
evoked from ipsilateral stimulus) / (dipole moment evoked from contralateral stimulus + dipole
moment evoked from ipsilateral stimulus) x 100], from peak dipoles (from Figure 4.2D) are shown
for the mature cortical wavepeaks. Stimulation of the right and left ear evokes different activation in
the cortex. A contralateral aural preference in the right auditory cortex (red bars) was present for all
peaks (P1, N1, P2). Aural preference in the left auditory cortex (blue bars) was only present for P1.

95
4.4.2 Long periods of unilateral CI use drive abnormal patterns of auditory
activity
Auditory stimulation with a CI for over 10 years appears to promote normal-like maturation
of cortical responses. As shown in Figure 4.4A, the cortical waveform developed into a polyphasic
response with similar latencies to the normal hearing group (P1: t(10)=−0.88, p>0.05; N1: t(10.18)=−1.3,
p>0.05; P2: t(10.77)=1.43, p>0.05) and similar peak-to-peak amplitudes for the P1-N1 complex
(t(6.87)=1.75, p>0.05) and dipole moment magnitudes (Figures 4.4C and 4.4E), and latencies of
extracted peak dipoles in Figure 4.4F were also in line with those of the normal hearing group.
Nonetheless, differences from normal remained. The amplitude of peak P2 was significantly larger in
CI users (t(14.51)=2.49, p<0.05) and current distribution across the scalp, shown in Figure 4.4B, was
unusually similar for all three peaks of the mature response regardless of whether the wavepeak
recorded at Cz was of a positive or negative polarity. Furthermore, even though cortical activity evoked
by the experienced CI was generated predominantly in areas of the temporal cortex, auditory input in
these adolescents activated the left and right auditory cortices differently than their normal hearing
peers. Dipole activity underlying the mature cortical response was stronger in the left than right
auditory cortex as shown by the red hotspots in the topographic brain images of Figure 4.4C and
larger blue than red bars in Figure 4.4E reflecting magnitude of extracted peak dipoles from Figure
4.4D (F(2,24)=10.978, p<0.001; post hoc: P1: t(26)=2.4, p<0.05, P2: t(26)=3.9, p<0.01). This reflects
cortical activation in an opposite direction to the rightward hemispheric bias that was observed in
normal hearing peers in Figure 4.2 and indicates a contralateral dominance of the auditory pathways
in the left auditory cortex with long-term unilateral stimulation of the right-experienced implant. This
confirms that unlike their normal hearing peers, unilateral CI users did not develop right hemisphere
specialization to tone-bursts despite developing cortical responses with expected mature wavepeaks.
Activity evoked by stimulation of the naïve pathways on the first day of activation of that left
side was significantly different from the experienced side. As shown in Figures 4.4G and 4.4H, the
morphology of these cortical waveforms was atypical and different from the response evoked by the
experienced ear. These were characterized by a large negative amplitude peak, labelled N(ci), followed
by a large positive peak (P(ci)). This abnormal response was consistently evoked by the deprived
pathways in all CI users with long-term unilateral deafness. While the latency of these peaks, shown
in Figure 4.4L, were similar to that of N1 and P2 of the experienced side (Figure 4.4F) (N(ci)/N1:

96
t(11)=1.85, p>0.05; P(ci)/P2: t(11)=2.71, p>0.05), the absolute amplitudes (N(ci)/N1: t(11)=3.45, p<0.01;
P(ci)/P2: t(11)=3.31, p<0.01), peak-to-peak amplitudes (N(ci)-P(ci)/N1-P2: t(11)=-3.37, p<0.01), and global
field power (±5µV) were significantly larger than responses from the experienced and normal hearing
ears (F(3,70)=5.4, p<0.05). Underlying dipole activity shown by topographic plots in Figure 4.4I and
peak dipole strength in Figure 4.4K extracted from Figure 4.4J indicated further differences. Dipole
activity was more widespread and symmetric across both auditory cortices (N(ci): t(20)=-0.56, p>0.05;
P(ci): t(20)=-1.6, p>0.05), and dipole moments for both peaks were abnormally large in both hemisphere
(F(3,36)=5.481, p=0.03) compared to the experienced side and both the right and left normal hearing
ears.

97

98
Figure 4-4: Cortical dipole activity evoked by auditory stimulation of the experienced-right and naïve-
left ears of CI users.
Auditory evoked cortical responses were stimulated from the experienced-right (n=32) and naïve-left
(n=21) ears of adolescents with CIs, aged 15.9 ± 2.0 years. Cortical activity evoked by stimulation of
the right ear after 12.4 ± 1.7 years of unilateral CI experience is shown in A. – F. Left ear had 12.0 ±
2.1 years of unilateral deprivation. Responses recorded within the first week of CI activation on that
side are shown in G. – L. Individual responses from a midline cephalic electrode (Cz) (thin grey lines),
and grand mean responses (thick black lines) show 3 peaks (P1, N1, P2), characteristic of mature
responses, evoked by stimulation of the experienced-right CI in A., but an abnormal biphasic peaked
response (N(ci), P(ci)) of abnormally large amplitudes was evoked by the naïve-left side (n=21), shown
in G. Mean global field power is plotted for mean response data from each CI (B. indicates responses
from the experienced side and H. shows activity evoked by the naïve side). Topographic maps
indicate unusually similarly distributed scalp potentials (red = positive potentials, blue = negative
potentials) for the 3 peaks of the mature response on the experience side. These are different between
both peaks on the naïve-left side. Global field power is abnormally large on this side (±5µV) compared
to the experienced side (±2µV) and either ear of normal hearing peers (±1µV). C. Mean peak dipole
activity evoked by the experienced CI for each cortical wavepeak is plotted in each of 63,646 voxels in
brain space on age-appropriate topographic head models derived from the MNI MRI Library (MNI
Axial orientation). A strong contralateral bias in the left hemisphere, shown by hotspots of dark red,
occurs with stimulation of the experienced-right CI. By contrast, dipole activity evoked by stimulation
of the naïve-left ear in I. is symmetrically distributed in both hemispheres and is stronger compared
to right ear dipoles for both peaks of the response. D. & J. Peak dipoles underlying each cortical
wavepeak in the left (blue) and right (red) auditory cortices were chosen within the hotspots shown in
C. and I. for each child. Peak dipole locations are shown on glass brain images. Axis are MNI
coordinates in millimeters. E. Mean (± 1 SE) dipole moment strength is plotted for the left (blue
bars) and right (red bars) auditory cortices for all cortical peaks. Consistent with the topographic plots
in C., peak dipole activity is significantly stronger in the left than right auditory cortex for the mature
peaks P1 (p<0.05) and P2 (p<0.01) on the experienced side. K. The same responses are plotted for
the naïve side and shows dipole activity consistent with the data in I. F. & L. Mean (± 1 SE) peak
dipole latency are similar between the left (blue) and right (red) auditory cortices and are in line with
cortical wavepeak latencies shown in A. and B. for the experienced side and in G. and H. for the
naïve side.

99
Permutation analyses were used to identify significant differences in activity between left and
right auditory cortices in the normal hearing and CI groups. Figures 4.5A-D revealed significant
differences in activation between left and right temporo-frontal and temporo-parietal regions in both
groups with unique patterns in the CI users relative to their normal hearing peers. In the normal
auditory system, the emergence of mature cortical responses was expected to underlie the development
of hemispheric specialization (Zatorre and Belin 2001; Jamison, Watkins et al. 2006). We hypothesized
that this would be observed as a shift in lateralization toward the right auditory cortex in response to
tonal stimulation. As shown in Figure 4.5A, minimal differences in activity between the cortical
hemispheres were evident when tones bursts were presented to normal right ears using this analysis.
Significant differences were clearer in the auditory cortices as well as temporo-frontal and temporo-
parietal regions for normal left ear stimulation as shown in Figure 4.5B. These differences were larger
in the right than left cortical hemisphere. In the CI group, the left temporo-parietal junction was more
strongly activated than the right for all peaks, and the right temporo-frontal region was more strongly
activated than the left in the latency range of P2. As shown in Figure 4.5D, stimulation from the
newly implanted ear resulted in a greater activation of right than left parietal-temporal-occipital
association areas and left than right temporo-frontal cortex. Peak dipoles from the left and right
auditory cortex were compared. Normalized lateralization values shown in Figure 4.5E indicated that
while tonal stimuli lateralized to the right auditory cortex regardless of ear of stimulation in adolescents
with normal hearing, the opposite is true of adolescents with long term right CI use.
Despite developing mature-like cortical responses with over a decade of CI experience,
significantly strong left hemisphere lateralization was found for all peaks recorded in CI users (P1:
t(10)=-2.460, p=0.034, N1: t(14) =-2.260, p=0.04, P2: t(15) =-6.912, p<0.001). This pattern was significantly
different from the responses of either ear in normal hearing peers (F(4,92)=3.286, p=0.015, post-hoc:
right stimulation: P1: p=0.03, P2: p=0.003; left stimulation: P1: p=0.002, N1: p=0.001, P2: p<0.001).
Dipole activity evoked by stimulation in the newly implanted ear in both hemispheres was more
symmetrical as shown by a reduction in lateralization for peak N(ci) and P(ci) (N(ci): t(15) =-0.726, p=0.479
P(ci)=2.238: t(12), p=0.816). This was significantly different from the right lateralized responses
stimulated by tones in normal left ears (F(1,7)=6.674, p=0.036, post-hoc: N1: p=0.03, P2: p=0.006),
indicating that the cortex responds to input delivered by the naïve pathways very differently.
Symmetrical activation of both hemispheres indicated diffuse activation of the cortex and increased
cortical activity in response to sound when those pathways were stimulated for the first time.
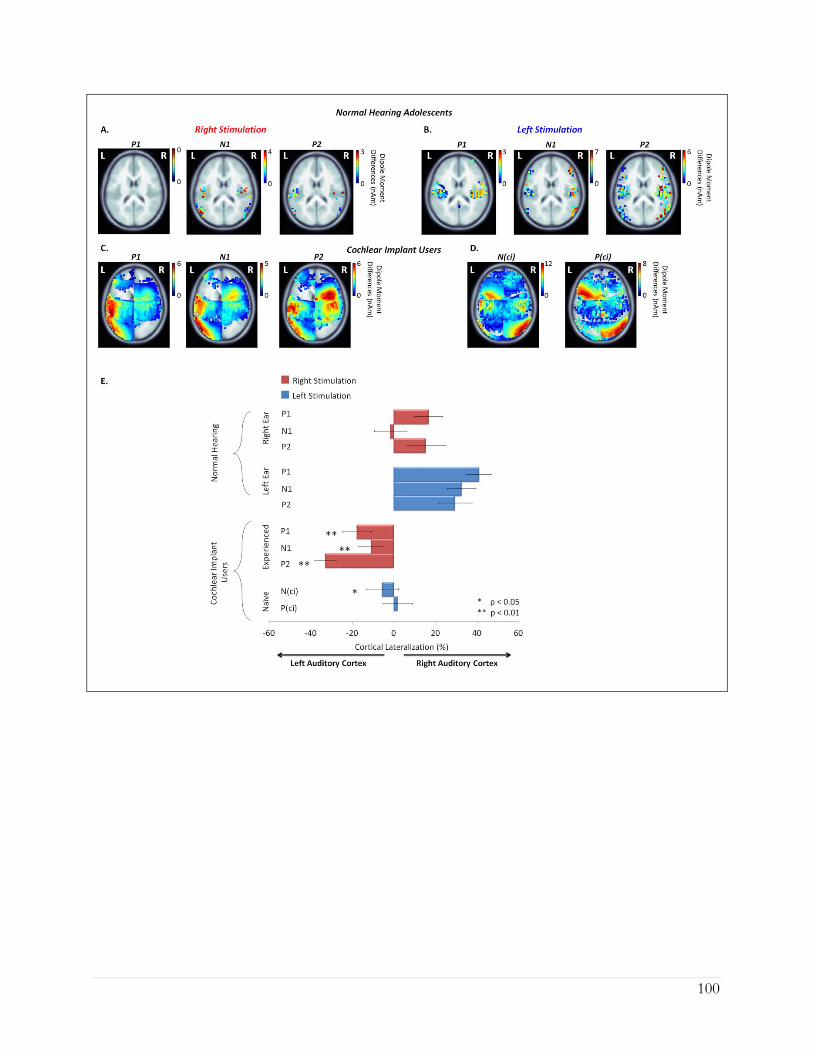
100

101
4.4.3 Additional cortical areas are recruited by cochlear implant stimulation
relative to normal
We hypothesized that long-term unilateral CI use would drive abnormal underlying cortical
activity in the stimulated pathways compared to normal and leave the opposite deprived pathways
vulnerable to significant change. To assess this, we compared responses between CI and normal
hearing groups using permutation analyses. As shown in Figure 4.6A, significantly larger dipoles were
identified in the frontal cortex underlying peak P1 and in the right precuneus region for peak P2 for the
Figure 4-5: Cortical lateralization evoked by stimulation of the right/experienced and left/naïve
ear/implant in normal hearing adolescents and CI users.
Cortical lateralization for each peak of the cortical waveform was assessed across 63,646 peak dipoles
using permutation testing to compare activation of the left and right hemispheres. Significant absolute
differences in dipole moments for right – left hemisphere (p<0.005) are plotted on topographic plots
for stimulation of the right ear in A. and the left ear in B. for adolescents with normal hearing; and
the right-experienced CI in C. and naïve-left side in D. in the implant cohort. Auditory activity
lateralized towards the right hemisphere in normal hearing individuals, particularly in response to left
ear stimulation, as shown by the dark red hotspots indicating greater inter-hemispheric dipole
differences. A significant shift in lateralization towards the contralateral left auditory cortex was
observed in CI users with stimulation of the experienced-right side. Cortical asymmetries towards the
auditory cortices in either hemisphere were not clear when stimulated from the naïve side.
Lateralization of secondary sources in the left temporo-parietal junction and right temporo-frontal
cortex was present for the experienced-right side, with stronger activity underlying P2. This activity
was reversed on the naïve-left side and occurred with increased dipole strength in the right parietal-
temporal-occipital association areas and left temporo-frontal cortex. E. Mean (± 1 SE) percent
cortical lateralization of peak dipoles is consistent with these results. Cortical lateralization from peak
dipoles was calculated using the formula: [(right hemisphere peak dipole moment – left hemisphere
peak dipole moment) / (right hemisphere peak dipole moment + left hemisphere peak dipole moment)
x 100]. Red bars indicate dipoles evoked from stimulation of the right ear/experienced CI and blue
bars indicate activity from the left ear/naïve CI.

102
experienced CI side compared to responses from the right ear in the normal hearing group, indicating
that CI users recruit additional cortical areas to process sound. At the same time, depriving auditory
input on the opposite side for over a decade led to significantly different activation in those pathways
relative to responses from the normal hearing left ear. Dipole activity in the naïve-left ear shown in
Figure 4.6B was significantly larger across most of the cortex, particularly for peak N(ci), compared to
normal left ear evoked activity. This might indicate aberrant cortical organization as a result of long-
term deafness, in line with our hypothesis.

103
4.4.4 Abnormal activity evoked by the naïve side predicts poor speech
perception outcomes
We asked whether there was a relationship between cortical activity in CI users and their speech
perception once they had used their bilateral CIs for at least 6 months. As shown in Figure 4.7A,
speech perception performance with the experienced CI was high, with a mean ± 1 standard-error
score of 77.54 ± 2.88%. By contrast, this was significantly worse (14 ± 4.92%) when adolescents
listened with their newly implanted ear alone (t(17)=5.70, p<0.01). All but two adolescents achieved
less than 50% accuracy. An analysis of speech perception and dipole activity between both ears, shown
in Figure 4.7B, indicated a significant correlation between speech perception outcomes and left
hemisphere lateralization for peak P2 (R=-0.55, p=0.02). We hypothesized that this relationship
reflects a supportive role of increased activity in the contralateral pathways towards the left cortex for
speech perception, rather than reduced activity in the right auditory cortex. In support, a significant
positive correlation between dipole strength and speech perception scores was found in the left
auditory cortex (P1: R=0.48, p=0.05, N1: R=0.51, p=0.03, P2: R=0.53, p=0.02) but not the right (P1:
R=-0.14, p=0.62, N1: R=0.43, p=0.06, P2: R=0.03, p=0.92) for the experienced-right CI. No
significant correlations with speech outcomes were found for cortical activity evoked by the naïve CI.
Figure 4-6: Group differences in cortical dipole activity between normal hearing adolescents and CI
users.
Permutation analyses were used to compare dipole activation between the group of CI users and the
group of normal hearing peers. Significant absolute dipole differences between the two groups
(p<0.01) are shown on topographic brain plots for experienced-right CI – normal hearing right ear
stimulation in A., and naïve-left CI – normal hearing left ear stimulation in B. Dipole magnitudes
underlying the mature cortical peaks were generally similar between adolescents with normal hearing
and experienced CI users, but CI stimulation of the experienced side activated cortical regions in the
left frontal cortex for peak P1 and the precuneous regions for P2 which were not present in normal
hearing peers, as shown by the dark red hotspots in A. By contrast, the naïve ear evoked abnormally
large and diffuse cortical activity across the brain, with significantly stronger dipole activity compared
to the left ear of the normal hearing cohort.

104
4.5 Discussion
Data from the present study suggest that hemispheric specialization emerges with normal
maturation of the auditory cortex. In response to tone-bursts, adolescents with normal hearing showed
Figure 4-7: Speech perception performance on the experienced-right and naïve-left sides in CI users.
A. Speech perception outcomes measured from the experienced side (n=28) (red) and naïve ear
(n=18) (blue) of CI users was assessed using the Phonemic Balanced Kindergarten (PBK) speech test
after 6 to 18 months of bilateral CI experience. Individual responses are indicated by the round
symbols for the experienced-right ear and diamonds for the naïve-left ear, with mean performance
shown by the bar graph. Performance on the experienced side was significantly better than the naïve
side (p<0.01). B. Analyses of cortical lateralization and performance on the PBK speech test indicated
significant correlation between left hemisphere lateralization for peak P2 (p<0.05) and better speech
scores on the experienced CI side. No significant correlations were found for dipole activity evoked
by stimulation of the naïve side and speech perception.

105
mature cortical responses, which lateralized to the right auditory hemisphere. Despite developing
normal-like cortical peaks, adolescents who had over a decade of unilateral CI experience developed
cortical responses which, in contrast to the normal hearing group, lateralized to the contralateral left
auditory cortex with stimulation of the right-experienced ear and bilateral cortical activity in response
to stimulation from the naïve-left ear. Increased response in the left auditory cortex reflects
strengthening from unilaterally stimulated pathways and may support speech perception in CI users.
Very poor performance in the newly implanted ear could reflect undefined and perhaps compromised
auditory processing. In support, additional cortical areas were stimulated in adolescents using CIs
relative to their normal hearing peers, particularly when evoked by the newly implanted ear, suggesting
alternate processing of auditory input. These findings indicate that long-term stimulation of the
auditory system with a unilateral CI promotes an asymmetric mature auditory cortex while, at the same
time, leaving pathways from the opposite ear unprotected from abnormal effects of deafness. Because
sensitive periods in development typically end once a period of maturation is reached (Lenroot and
Giedd 2006; Blakemore 2012; Lohmann and Kessels 2014), further changes from the newly implanted
side may be challenging to promote in these adolescents.
4.5.1 Hemispheric specialization requires normal bilateral hearing
Normal cortical activity was measured in adolescents with normal hearing in response to a non-
speech 500Hz tone to mimic the stimuli that CI users received through an apical electrode of their
device. Data in Figure 4.2 indicated a normally mature response to this tone-burst with more activity
in the right than left auditory cortex. Lateralization to the right hemisphere occurred for both
presentation to left and right ears as shown in Figure 4.5. This appears to have emerged in
development, as immature cortical responses to the same stimuli in younger children with normal
hearing showed contralateral lateralization of similar degrees for both left and right ear stimulation
(Gordon, Wong et al. 2013). Lateralization of immature responses to the contralateral cortex in the
normal hearing group was consistent with the anatomy of afferent auditory projections. The majority
of auditory fibers crosses to the contralateral side at the level of the brainstem and ascend to the
ipsilateral auditory cortex (Glendenning, Brusno‐Bechtold et al. 1981; Nordeen, Killackey et al. 1983;
Ponton, Vasama et al. 2001). Although it is possible for stimulation from one ear to reach the ipsilateral
cortex, the contralateral pathway will have the more direct route (Ponton, Vasama et al. 2001). Perhaps

106
it is for this reason that animals show an aural preference for the contralateral ear in each auditory
cortex (Kral 2013; Kral, Heid et al. 2013; Kral, Hubka et al. 2013).
The adult human auditory brain is more specialized; activity in the left auditory cortex is tuned
to temporal processing, whereas regions of the right auditory cortex respond to spectral processing
(Zatorre and Belin 2001; Zatorre, Belin et al. 2002; Schönwiesner, Rübsamen et al. 2005; Jamison,
Watkins et al. 2006). Much of this processing in the right hemisphere occurs in secondary association
areas (Schönwiesner, Rübsamen et al. 2005). Neurons in belt and parabelt regions of the superior
temporal gyrus, which are later to develop (Moore and Guan 2001; Moore and Linthicum 2007), are
involved in spectral analyses of complex sounds at early stages of processing (Rauschecker and Tian
2004; Schönwiesner, Rübsamen et al. 2005) and are more sensitive to spectral stimuli (Zatorre, Belin
et al. 2002; Schönwiesner, Rübsamen et al. 2005; Jamison, Watkins et al. 2006). Interactions with
frontal cortical regions have also been reported for processing patterns of tones, particularly when
demands on working memory are elicited by the task (Zatorre, Belin et al. 2002). Activation of
secondary sources in the right frontal and parietal regions underlying peaks N1 and P2, shown in Figure
4.5, indicate consistent findings in our cohort of adolescents with normal hearing. These cortical peaks
reflect responsiveness in association auditory regions (Ponton, Eggermont et al. 2000; Ponton,
Eggermont et al. 2002; Wunderlich and Cone-Wesson 2006; Wunderlich, Cone-Wesson et al. 2006)
which are involved in 1) complex auditory discrimination such as perceiving speech in the presence of
noise and understanding degraded speech (Eggermont and Ponton 2003), 2) increased
attention/arousal during listening (Picton and Hillyard 1974; Rif, Hari et al. 1991; Tremblay, Kraus et
al. 2001; Tremblay and Kraus 2002; Alain, Snyder et al. 2007; Tremblay 2007; Tremblay, Shahin et al.
2009), and 3) multi-sensory integration between auditory inputs with vision and touch (Grady, Van
Meter et al. 1997; Webster and Colrain 2000; Moller and Rollins 2002; Crowley and Colrain 2004;
Busse, Roberts et al. 2005; Rinne 2010). Thus, dipole activation underlying the wavepeaks across
secondary cortical regions in adolescents with normal hearing marks the establishment of appropriate
specialization once the auditory brain matures.
The brain undergoes significant structural and functional changes, which are more rapid in
earlier development and mostly complete by late adolescence (Giedd, Snell et al. 1996; Giedd,
Blumenthal et al. 1999; Blakemore and Choudhury 2006; Lenroot and Giedd 2006; Lebel and Beaulieu
2011; Blakemore 2012; Lohmann and Kessels 2014). Asymmetric function in the auditory cortices for
temporal and spectral processing of sound has been attributed to the development of structural

107
differences in each hemisphere. There are wider and more spaced cell columns coupled with heavier
axon myelination and increased inter-connectivity in left auditory regions compared to the right,
corresponding to rapid transmission and more precise temporal resolution in the left hemisphere
(Penhune, Zatorre et al. 1996; Zatorre and Belin 2001; Zatorre, Belin et al. 2002; Toga and Thompson
2003). Such inter-hemispheric structural specialization in the auditory system is mediated by auditory
experience (Penhune, Zatorre et al. 1996) and normal maturational changes occuring during
adolescence (Moore and Linthicum 2007; Kadis, Pang et al. 2011), in line with the maturational time-
course of the auditory evoked cortical response (Ponton, Eggermont et al. 2000; Ponton, Eggermont
et al. 2002; Eggermont and Ponton 2003; Moore and Linthicum 2007; Jiwani, Papsin et al. 2013).
Some of the same processes likely occurred in adolescents using unilateral CIs, given that
cortical peaks characteristic of mature responses were recorded in this group. The emergence of these
mature-like peaks is consistent with previous reports (Jiwani, Papsin et al. 2013) and suggests that the
neural generators, thalamo-cortical and cortical-cortical connections linked to the development of
superficial cortical layers (Ponton and Eggermont 2001), are maturing in adolescents who have used a
unilateral CI for most of their lives. The development of mature responses coupled with good speech
perception outcomes when using the implant in that ear (Figure 4.7) provides a general impression
that at least some degree of auditory development proceeds normally with long-term CI stimulation.
On the other hand, dipole activity in Figure 4.4 and lateralization measures in Figure 4.5 indicate
significantly stronger dipoles in the contralateral left auditory brain in response to stimulation of the
experienced-right implant. Thus, adolescents with right CIs did not develop the normal right brain
hemispheric bias associated with non-speech stimuli, despite having developed mature normal-like
cortical waveforms.
4.5.2 Long periods of unilateral CI use strengthens pathways from the
stimulated ear
There are several reasons why adolescents using CIs may not have developed specialized
auditory cortices similar to their normal hearing peers. They had been bilaterally deaf from early in life
(typically from birth) and listened, often unilaterally, to a representation of sound missing much of its
fine temporal information. The lack of temporal fine structure makes pitch discrimination difficult

108
(Drennan and Rubinstein 2008; Zeng, Rebscher et al. 2008). The asymmetric dipole activity shown in
Figures 4.5 and 4.7 could thus be explained by 1) persistent immaturity in the ascending pathways, 2)
disrupted excitatory-inhibitory balance arising from long periods of deprivation in the opposite ear
and/or 3) increased sensitivity of the left hemisphere for CI input, perhaps to support speech
processing (Zatorre and Belin 2001; Zatorre, Belin et al. 2002; Schönwiesner, Rübsamen et al. 2005;
Kadis, Pang et al. 2011; Chilosi, Comparini et al. 2014). This could occur to compensate for the
distorted access to acoustic input through the CI and/or the lack of binaural hearing.
It is possible that specialization has not yet emerged in these pathways, with contralateral
afferent projections still dominating auditory input as in earlier development (Gordon, Jiwani et al.
2013; Gordon, Wong et al. 2013). Effects of unilateral auditory deprivation/stimulation appear to
occur early and persist into maturation. In younger children with CIs, abnormal strengthening of
activity in the contralateral pathways from the experienced-right ear to the brainstem (Gordon, Salloum
et al. 2012) and cortex (Gordon, Wong et al. 2013) was found when unilateral implant use exceeded
1.5 years. The data in Figures 4.4 and 4.5 are consistent with these findings, indicating that increased
activity in the left auditory cortex from the right implanted ear continues into maturation with longer
periods of unilateral CI use. There was also strengthening of pathways to the ipsilateral cortex from
the unilaterally right stimulated ear in the immature cortex, as measured by a reversal of stimulus
preference from the contralateral ear to the ipsilateral first implanted ear (Gordon, Wong et al. 2013).
This could not be assessed in the present study because of the highly abnormal responses with very
large underlying dipoles evoked in the newly implanted side.
Asymmetric strengthening of the contralateral pathways from the stimulated ear might also be
explained by increased excitability of auditory neurons from this side. The normal organization of
inhibitory and excitatory activity breaks down when input to both ears are unbalanced. A significant
unilateral hearing loss during early stages of development not only leads to a loss of neurons in the
cochlear nucleus of the affected side (Hashisaki and Rubel 1989; Nordeen, Killackey et al. 2004), but
also results in reduced synaptic inhibition and enhanced excitatory glutamatergic conductance in the
superior olive (Kotak and Sanes 1996), the inferior colliculus (Kitzes 1984; Moore and Kitzes 2004;
Nordeen, Killackey et al. 2004; Vale, Juíz et al. 2004; Takesian, Kotak et al. 2009; Popescu and Polley
2010) and the primary auditory cortex of the stimulated pathways (Kotak, Fujisawa et al. 2005; Moore,
Devlin et al. 2005; Popescu and Polley 2010; Kral, Heid et al. 2013; Kral, Hubka et al. 2013).
Importantly, effects are found only when the loss is present in early life (Nordeen, Killackey et al. 1983;

109
Reale, Brugge et al. 1987; Moore and Kowalchuk 1988; Nordeen, Killackey et al. 2004) and when it is
unilateral (Moore 1990). Such deafness-induced disruptions to the delicate balance of excitatory-
inhibitory inputs influence and shape the development of binaural hearing in the brainstem (Grothe,
Pecka et al. 2010) and neuronal circuits in the auditory cortex (Turrigiano and Nelson 2004; King
2010). The coupling of excitatory-inhibitory synaptic inputs in the auditory system is influenced by
experience during development (King 2010). Maturation of inhibitory tuning occurs more slowly than
that of excitation (Dorrn, Yuan et al. 2010) and inhibitory synapses are more vulnerable to deafness-
induced perturbation (Sanes and Kotak 2011). Thus, the abnormal strengthening of pathways from
the unilaterally stimulated CI ear could reflect a disrupted balance of excitatory-inhibitory transmission
during early development.
An additional explanation for the abnormally increased dipoles in the left auditory cortex of
CI users is the possibility that this reflects compensation for inadequate input from the unilateral CI.
The left auditory cortex normally codes temporal information including speech. Although no speech
sounds were provided to evoke the cortical responses in this study, speech perception scores on the
PBK test in the experienced ear increased with stronger dipoles in the left auditory cortex, as shown
in Figure 4.7B. This is consistent with a previous report showing better grammar and expressive
language scores in children with right CIs who lateralized auditory input in the contralateral left
hemisphere (Chilosi, Comparini et al. 2014). In support, strong left cortical activity in individuals with
normal hearing has been associated with better language skills (Kadis, Pang et al. 2011), more precise
auditory processing (Zatorre and Belin 2001; Zatorre, Belin et al. 2002; Schönwiesner, Rübsamen et
al. 2005) and increased reading and spelling skills (Abrams, Nicol et al. 2006). Enhanced activity in the
left cortex of adolescents using CIs could thus mark plastic changes in the brain to compensate for the
abnormal electrical input provided by the implant device and/or stimulation of the auditory pathways
from only one side. Indeed, it is clear that individuals compensate for CI listening by using increased
multi-sensory information (Giraud, Price et al. 2001; Doucet, Bergeron et al. 2006; Giraud and Lee
2007; Lee, Giraud et al. 2007; Sandmann, Dillier et al. 2012) and increased effort/attention (Hopyan-
Misakyan, Gordon et al. 2009; Pisoni, Conway et al. 2010; Gordon, Jiwani et al. 2013; Kronenberger,
Colson et al. 2014). Increased cortical processing to support CI listening in good users has been shown
by increased activation of the left dorsolateral pre-frontal, frontal and parietal networks, including the
precuneus regions, which are involved with higher cognitive functions (Giraud, Price et al. 2001;
Giraud and Lee 2007), and larger amplitudes of the P2 peak of the electrophysiological response

110
(Jiwani, Papsin et al. 2013). In the present study, adolescents with CIs had abnormally increased activity
in the frontal cortex and precuneus regions, shown in Figure 4.6, even in a passive response evoked
by non-speech stimulation from the ear they had listened to for most of their lives. This suggests a
profound reorganization of the cortical network to support CI listening.
4.5.3 Activity evoked by stimulation of the newly implanted ear is abnormal
It is difficult to separate the effects of stimulation from one ear from the simultaneous effects
of deprivation in the other ear in adolescents using unilateral CIs. We suggest, however, that leaving
the opposite pathways deprived of input beyond the period of brainstem (Gordon, Papsin et al. 2006;
Gordon, Jiwani et al. 2013) and cortical (Jiwani, Papsin et al. 2013) maturation further disrupts
organization of activity in the auditory pathways. As shown in Figure 4.4, cortical responses at initial
stimulation of that side were atypical and dominated by an abnormally large biphasic waveform, despite
equal amplitudes of brainstem responses on each side. It may reflect cortical immaturity, as a similar
response has been observed in normal hearing pre-term infants (Wunderlich and Cone-Wesson 2006)
and children with little auditory input due to GJB-2 deafness (Gordon, Tanaka et al. 2011). It could
also reflect abnormal cortical activity, as the same response was recorded from children with congenital
deafness implanted late (Sharma, Dorman et al. 2002) and in children who had poor speech perception
with their implants (Gordon, Tanaka et al. 2008).
Cortical activity underlying the abnormal response in the newly implanted ear, shown in Figure
4.4, occurred with abnormally large dipole moments compared to the activity evoked by the opposite
experience side and the normal hearing group. Moreover, activity was more widespread as shown in
Figure 4.6, with a loss of cortical lateralization as plotted in Figure 4.5. Reduced cortical lateralization
has been associated with poor auditory processing skills, decreased academic performance and
increased difficulty with reading and spelling (Abrams, Nicol et al. 2006). Poor speech perception
outcomes when adolescents were listening with the new implant after ~8 months of experience, shown
in Figure 4.7, is consistent with these reports, suggesting altered cortical processing in the deprived
pathways.
The long period of deprivation in this ear could have left it vulnerable to many structural and
functional changes. Significant reorganization of subcortical projections (Kitzes, Kageyama et al. 1995;

111
Mossop, Wilson et al. 2000; Moore and Kitzes 2004; Moore and Kowalchuk 2004; Nordeen, Killackey
et al. 2004; Vale, Juíz et al. 2004) and cortical activation patterns have been reported with unilateral
auditory deprivation in early life, both in animals (Popescu and Polley 2010; Kral, Heid et al. 2013;
Kral, Hubka et al. 2013) and congenitally deaf children receiving sequential CIs (Gordon, Jiwani et al.
2013; Gordon, Wong et al. 2013). This reorganization drove increased allocation of resources (Kral,
Hubka et al. 2013) and hemispheric preference towards the hearing ear (Gordon, Wong et al. 2013;
Kral, Hubka et al. 2013) while reducing efficiency and responsiveness of the deprived side to activate
the cortex but not eliminating it (Popescu and Polley 2010; Kral, Heid et al. 2013). Similar cortical
consequences occur in the visual system, with monocular deprivation. Findings of reduced visual
acuity from a deprived eye has been attributed to uneven competition between the two pathways
(Hubel and Wiesel 1970; Hubel, Wiesel et al. 1977; Le Vay, Wiesel et al. 1980; Blakemore 1988; Lewis
and Maurer 2005), resulting in jittered representation of visual input in the cortex (Jeffrey, Wang et al.
2004). The widespread dipole activity observed from the naïve CI could thus reflect undefined cortical
activity across both hemispheres, and suggests that similar aberrant processes may underlie auditory
cortical activity in these pathways. This might reflect an immaturity or dis-integration of auditory areas
in the cortex (Kral and Sharma 2012), and would explain the inferior auditory outcome observed from
this side.
Kral and colleagues indicated that unilateral deprivation in early life drives a separation of the
experienced and naïve pathways, which places the deprived ear at a disadvantage for competition of
cortical resources (Kral, Heid et al. 2013). Our results of generally abnormal and undefined cortical
organization in response to novel auditory input evoked by the naïve CI compared to the experienced
side support this and highlight the deleterious effects of unilateral deprivation on the brain.
4.6 Conclusion
Maturation of the auditory brain is an experience-dependent process which underlies
hemispheric specialization in adolescents with normal hearing. Unfortunately, cortical organization in
the auditory pathways differs from normal in both ears in adolescents who have used a unilateral CI
to hear for most of their lives. Our findings indicate that driving maturation of the auditory cortex
with only one implant for over a decade leads to lasting asymmetries in the auditory system and leaves

112
the deprived pathways unprotected from effects of unilateral auditory deprivation. Because sensitive
periods typically end when maturation is reached, maturation of the cortex from the experienced CI
may mark the closing of an important developmental period in the adolescent brain for promoting
auditory development in the deprived pathways.

113
4.7 Supplementary Information on Methods
The Time-Restricted, Artefact and Coherent Source suppression (TRACS) beamformer (Wong
and Gordon 2009) imaging method was used to localize cortical activation underlying the evoked peaks
in the cortex of adolescents. This method was developed by our laboratory in linearly constrained
minimum variance beamformer analysis, and was designed specifically to suppress the electrical
artefact from the cochlear implant (CI) device and assess auditory activity in CI users (Wong and
Gordon 2009). Unfortunately, other imaging methods which offer better spatial resolution than
electroencephalography (EEG) (i.e., functional magnetic resonance imaging (fMRI), magneto-
encephalography (MEG)) are incompatible with the CI magnet in the receiver stimulator component
of the subcutaneous CI (Loizou 1999).
The beamformer is an adaptive spatial filter which uses age-appropriate head model templates
and lead potential matrices to estimate activity at each voxel in the brain (Van Veen, Van Drongelen
et al. 1997). Using these filters, the beamformer correlates brain electrical activity at a given source to
the electric potential distribution measured by sensors at the surface of the scalp while suppressing
activity originating at different locations (Van Veen, Van Drongelen et al. 1997). In this way, the
beamformer provides an estimate of neural power for each voxel (3 x 3 x 3mm) in the cortex (Van
Veen, Van Drongelen et al. 1997; Fuchs 2007) across 63,646 voxels in brain space. These
reconstructed sources provide information about the strength of activity at identified locations and
their respective latencies in response to auditory stimulation in both CI users and normal hearing peers.
A 3-layer boundary element mesh was constructed using age-appropriate head model templates
derived from the Montreal Neurological Institute (MNI) MRI library. This was used to represent age-
appropriate head geometry and tissue conductivities of the brain, meninges, skull and scalp to account
for spatial smearing of electric potentials at the surface of the head. Source activity in the left
hemisphere was assessed using region suppression of activity in the right hemisphere, and vice versa,
as bilaterally correlated activity is known to pose a particular problem for source localization in auditory
cortices (Dalal, Sekihara et al. 2006). In adolescents using CIs, a time restricted artefact suppression
algorithm was applied to suppress the implant-generated electrical artefact, corresponding to the 4
largest singular vector values over the -80 to 10ms latency range. This ensures that singular vectors
representing up to 97% of the CI artefact are suppressed (Wong and Gordon 2009). The artefact is
generated as the CI device processes sounds and transmits input from the external processor to the

114
internally implanted component/magnet. The CI artefact lasts for at least the duration of the stimulus
and likely reflects residual electrical energy created by the radio frequency transmission of the signal to
the receiver (Gilley, Sharma et al. 2006; Martin 2007; Debener, Hine et al. 2008; Friesen and Picton
2010). Without the suppression algorithm, source localization of cortical peaks become obscured and
distorted (Gilley, Sharma et al. 2006; Friesen and Picton 2010).
Peaks in the cortical response were visually identified at latencies corresponding to those
previously reported from normal hearing peers and CI users (Gordon, Tanaka et al. 2008; Jiwani,
Papsin et al. 2013). Peak latencies and amplitudes were marked for responses evoked by stimulation
of the right and left ears in the normal hearing group and the experienced-right and naïve-left sides in
the CI group (i.e., P1, N1 and P2 for normal hearing responses and experienced CI stimulation; and N(ci)
and P(ci) for responses evoked by stimulation of the naïve CI). Mean ± standard deviations (SD) are
shown in Supplementary Table 4.1 for each group.
P1
(Mean ± SD)
N1 / N(ci)
(Mean ± SD)
P2 / P(ci)
(Mean ± SD)
Latency (ms)
Normal Hearing Right
Normal Hearing Left
Experienced-Right CI
Naïve Left CI
78.87 ± 9.20
77.63 ± 4.46
79.59 ± 6.22
--
108.83 ± 9.53
111.64 ± 7.16
105.43 ± 9.57
114.10 ± 8.34
164.22 ± 17.94
163.87 ± 19.19
162.20 ± 18.24
184.58 ± 19.01
Amplitude (µV)
Normal Hearing Right
Normal Hearing Left
Experienced-Right CI
Naïve Left CI
0.92 ± 1.46
1.06 ± 1.41
0.63 ± 2.92
--
-1.56 ± 1.72
-2.27 ± 1.95
-1.92 ± 3.18
-12.27 ± 5.94
2.83 ± 1.96
2.61 ± 1.73
5.30 ± 3.61
8.77 ± 5.86
Supplementary Table 4.1: Latencies and amplitudes of cortical response peaks

115
The estimate of the covariance matrix for beamforming analysis was calculated over a latency
window >20ms encompassing each peak of the cortical response for each participant. A pseudo-Z
statistic was used to normalize the signal-to-noise ratio of each voxel to avoid an over-weighing of
source localization toward the center of the head (Vrba and Robinson 2001). The pseudo-Z value was
obtained by dividing the mean of the sample signal by the standard deviation of the sample noise.
Once sources were localized using this lead normalization process, the dipole moment of that source,
measured in nAm, was computed and were plotted on topographic maps. Supplementary Figure
4.8 is an example of a topographic image from one adolescent with long-term CI experience and shows
the activity underlying the mature peak P2 in 63,646 brain voxels. Source locations of the strongest
dipoles in the red hotspots are marked in Supplementary Figure 4.8 by the green cross hairs in the
left and right hemispheres separately. The waveforms to the left of the head maps show the
corresponding virtual channel of the voxel, reflecting the strongest peak dipole underlying the peak of
the cortical waveform. In this example, the strongest dipole activity occurs at ~150ms following
stimulus onset and is generated by areas of the left and right temporal lobes.
A statistical threshold of baseline brain activity was computed using a one-tailed omnibus noise
T-test (Petersson, Nichols et al. 1999). This was done to eliminate false detection of spurious sources.
To compute this, a minimum tolerance pseudo-Z statistic was calculated by creating a plus-minus
averaged dipole activity across the whole brain; the polarity of all odd-numbered epochs was inverted
and averaged with all even-numbered epochs. The beamformer analysis was then performed on this
resulting noise-only dataset. This analysis produced a global threshold pseudo-Z value reflecting
baseline brain activity which was significantly different from the evoked brain activity (p ≤ 0.0005).
All voxels with pseudo-Z activity that was equal to or less than this omnibus value were considered to
be baseline brain activity and not evoked by the auditory stimulus. Cortical activity was assessed and
visualized topographically in this way on age-appropriate head templates for each participant in 63,646
brain voxels, with only activity greater than the baseline omnibus threshold plotted.

116
Supplementary Figure 4-8: Example from one child with 15.95 years of CI experience in the right
ear indicates activity underlying the mature peak P2.
The virtual channel waveforms and topographic locations corresponding to the voxel with the
strongest activity (red hotspots) is shown for the left and right hemispheres separately. Source
locations are marked by the green cross hairs. As shown, the strongest dipole underlying peak P2
occurs in areas of the left and right auditory cortices at 150ms, in line with the latency of the cortical
wave P2.

117
5. Chapter Five – Temporally coordinated activity in the brain is
promoted by long-term cochlear implant use in children
This paper was submitted to Cerebral Cortex using required journal formatting:
Jiwani, S., Doesburg, S. M., Papsin, B.C., Gordon, K.A. Effects of long-term unilateral cochlear
implant use on cortical synchrony in adolescents.
5.1 Abstract
Unilateral cochlear implant (CI) use promotes maturational changes in the auditory system but
causes abnormally increased activity in the contralateral cortex if a second implant is not provided
within 1.5 years. This abnormal development might be attributed to a loss of inhibitory processes
from the deprived side which would have normally been present with bilateral hearing. At the same
time, leaving the opposite pathways deprived of auditory input beyond both the early period of
brainstem maturation and the later one of cortical maturation also disrupts activity in these pathways
and leaves this ear with relatively poorer auditory function. In the present study, we investigated the
effects of driving maturation of the auditory cortex with over a decade of unilateral implant use on the
neural networks that support CI listening in these adolescents. We hypothesized that unilaterally-
driven maturation of the auditory brain would: 1) promote abnormally increased coordinated activity
in brain areas that respond to sound, perhaps reflecting increased attention for auditory processing,
but 2) auditory activity in the deprived pathways would be abnormally desynchronized, perhaps
reflecting segregation of these pathways from the cortical hearing network.
Cortical responses were recorded from 64-cephalic electrodes in 34 adolescents who had over
a decade of unilateral CI experience in the right ear. Twenty-one of these adolescents received a second
implant in their left ear, allowing us to assess the effects of unilateral stimulation on pathways from
the opposite deprived ear. Responses were elicited by electrical pulses delivered from the right and
left CIs separately and compared to a group of 16 normal hearing peers. Neural synchrony was
calculated between electrodes in the theta, alpha, beta and gamma frequency-bands to assess

118
temporally coordinated responses to sound across brain regions. Time series of amplitude and phase-
locking values were calculated at 1Hz intervals between 4 and 60Hz for 171 pairs of 19 electrodes.
Our results indicated that stimulation of an experienced-right CI elicited abnormally increased
phase synchronization across the latency range in the theta and beta frequency bands with bursts of
activity in the gamma frequencies. This pattern of cortical synchronization suggests increased
recruitment of attention and higher-order perceptual processes to support CI listening, despite the
seemingly passive auditory task. By contrast, activity was globally desynchronized in response to initial
stimulation of the naïve-left ear. Findings indicate that unilaterally-driven cortical maturation causes
lasting asymmetries in the auditory pathways and altered functional neural networks. Mature auditory
responses could mark the closure of a sensitive period for establishing hearing on the deprived side.
5.2 Introduction
A cochlear implant (CI) is a surgically implanted auditory prosthesis, which stimulates the
auditory nerve with electrical pulses to allow children who are deaf to hear and develop oral
communication. Prior to cochlear implantation, the period of bilateral deafness leaves association
areas of the developing auditory brain vulnerable to recruitment by the visual (Lomber, Meredith et al.
2010) and somatosensory systems (Meredith and Lomber 2011). These cross-modal changes are
associated with enhanced function of the intact senses (Finney, Fine et al. 2001; Bavelier and Neville
2002; Bavelier, Dye et al. 2006) and reflect the brain’s remarkable capacity for plasticity during
development. Such changes may be impossible to reverse if hearing is not restored within sensitive
periods in development (Lee, Lee et al. 2001). Unilateral CIs have been provided to children in early
life to limit such deafness-induced reorganization (Lee, Lee et al. 2001) and promote auditory
development (Sharma, Dorman et al. 2002; Gordon, Papsin et al. 2005; Gordon, Papsin et al. 2006;
Jiwani, Papsin et al. 2013). Unilateral implant stimulation promotes maturational changes to the
auditory pathways (Gordon, Jiwani et al. 2013; Jiwani, Papsin et al. 2013), but drives abnormal
asymmetries in the brainstem (Gordon, Salloum et al. 2012) and cortex (Gordon, Wong et al. 2013;
Jiwani, Papsin et al. Submitted), which persist if a second implant is not provided within a sensitive
period of 1.5 years (Gordon, Wong et al. 2013). We have recently found that long-term unilateral
auditory stimulation/deprivation drove abnormally strengthened activity from the hearing ear to the

119
contralateral left auditory cortex (Gordon, Wong et al. 2013; Jiwani, Papsin et al. Submitted) and an
abnormal reversal of aural preference in the ipsilateral cortex (Gordon, Wong et al. 2013; Kral, Hubka
et al. 2013). These asymmetries might be explained by increased excitability of auditory neurons from
this side resulting from a loss of inhibitory processes in pathways from the opposite ear (Vale, Juíz et
al. 2004; Kotak, Fujisawa et al. 2005; Takesian, Kotak et al. 2009; Popescu and Polley 2010; Kral, Heid
et al. 2013; Kral, Hubka et al. 2013) that would have normally occurred with bilateral hearing (Grothe
2003; Grothe, Pecka et al. 2010). At the same time, leaving the opposite pathways deprived of auditory
input beyond the period of cortical maturation (Jiwani, Papsin et al. 2013) also led to significantly
abnormal activity in these pathways (Jiwani, Papsin et al. Submitted), suggesting that aberrant patterns
of activity develop in the absence of auditory input.
Long-term unilateral stimulation/deprivation thus seems to drive a profound reorganization
in the central auditory system. Yet some children amaze us by achieving excellent listening and oral
communication abilities (Geers, Tobey et al. 2008; Geers and Sedey 2011; Geers and Sedey 2011). We
suggest that these children are using compensatory strategies to facilitate listening and auditory
development with the reduced cues delivered by a CI in only one ear. We are already finding that
children with unilateral implants use multi-sensory information more effectively than their peers with
normal hearing to understand subtle emotional cues in speech (Hopyan-Misakyan, Gordon et al. 2009)
but do so with longer reaction times (Hopyan et al., Submitted; Steel et al., Submitted). This might
indicate more effort (Hughes and Galvin 2013; Pals, Sarampalis et al. 2013) and thus reduced efficiency
for auditory processing. More effortful listening in turn, could mean that unilateral CI users expend
more mental resources than their normal hearing peers to aid CI listening (Cleary, Pisoni et al. 2001;
Pisoni, Conway et al. 2010; Pisoni, Kronenberger et al. 2011; Geers, Pisoni et al. 2012; Hughes and
Galvin 2013; Kronenberger, Colson et al. 2014). In support, children who perform well with their
implants activate areas of the dorsolateral prefrontal cortex that are involved in higher cognitive
functions such as attention and working memory (Jeong Lee, Kang et al. 2005; Lee, Giraud et al. 2007),
and the precuneus region, which participates in visual imagery and episodic memory (Giraud, Price et
al. 2001; Jeong Lee, Kang et al. 2005). Activation of these areas indicate altered allocation of cortical
resources for CI listening (Kral and Sharma 2012; Kral 2013; Kral, Heid et al. 2013) and may underlie
differences in cognitive and perceptual processes that individuals with long-term unilateral CI
experience use compared to normal hearing peers.

120
We can now assess the task-dependent recruitment of distributed neural networks, which have
been associated with perceptual, motor and cognitive processing, including the engagement of higher
order cognitive processes such as selective attention and working memory in auditory processing,
through the measurement of inter-regional oscillatory synchronization (Lachaux, Rodriguez et al. 1999;
Varela, Lachaux et al. 2001; Fries 2005; Uhlhaas, Pipa et al. 2009; Uhlhaas, Roux et al. 2010). Cognitive
and perceptual functions in the brain are shaped by coordinated interactions/communication,
mediated by the synchronization of neural oscillations, across populations of neurons that are
distributed within and across brain regions (Uhlhaas, Roux et al. 2010). This coordinated activity
mediates the development and maturation of dynamic connections that are fundamental for functional
integration and processing of distributed information across large-scale networks (Varela, Lachaux et
al. 2001; Velazquez, Erra et al. 2009). In brief, neuronal communication is constrained by anatomical
connections between neurons (Fries 2005). A stimulated neuron synapses with all other neurons to
which it is anatomically connected. Activated neural groups then oscillate and those neurons that
oscillate in synchrony exchange bursts of action potentials, which are received by the input pyramidal
neurons during their depolarized phase, when they are most excitable (Fries 2005). Synchronization
between such assemblies of neurons thus allows them to exchange information effectively (Nunez
2000; Doesburg and Ward 2009). This neural encoding, which can be measured from inter-electrode
oscillatory synchronization in electroencephalography (EEG) and magnetoencephalography (MEG),
underlies neuronal communication and represent cognitive dynamics throughout the brain including
mechanisms of attention, memory, learning, anticipation, decision-making and consciousness (Ward
2003; Palva and Palva 2012).
Spectral power (i.e., amplitude) and phase locking of activity in different frequency bands are
measures of local and long-range neural synchronization respectively, which have been implicated in
information processing (Müller, Gruber et al. 2009). Increased long-range neural synchrony in the
gamma and theta frequencies, together with a reduction in local alpha activity has been linked to
increased cognitive processing during tasks of selective attention (Jensen, Kaiser et al. 2007; Doesburg,
Roggeveen et al. 2008; Green, Doesburg et al. 2011), auditory attention control (Doesburg, Green et
al. 2012), short-term and working memory (Doesburg, Herdman et al. 2010; Palva, Monto et al. 2010),
and multisensory integration (Doesburg, Emberson et al. 2008). Increased gamma-band
synchronization has been suggested to reflect local processing of cortical input, but feature binding
and information processing mechanisms are mediated by long-range synchronization in the theta and

121
alpha bands (Von Stein and Sarnthein 2000). Appropriate coupling of these frequencies are thus
crucial for maintenance of cortical organization in neural networks and support cognition/perception
and information processing. However, deviations from normal patterns of neural synchronization
have been associated with clinical impairments (Uhlhaas and Singer 2006) including epilepsy (Brugge,
Volkov et al. 2003; Matsumoto, Nair et al. 2004; Uhlhaas and Singer 2006), schizophrenia (Uhlhaas
and Singer 2006; Uhlhaas and Singer 2010; Uhlhaas and Singer 2011), Alzheimer’s disease (Uhlhaas
and Singer 2006), Parkinson’s disease (Schnitzler and Gross 2005; Uhlhaas and Singer 2006; Stoffers,
Bosboom et al. 2007), autism (Uhlhaas and Singer 2006; Vidal, Nicolson et al. 2006; Hughes 2007;
Wilson, Rojas et al. 2007; O’Connor 2012; Khan, Gramfort et al. 2013), attention deficit (Konrad and
Eickhoff 2010), and cognitive difficulties prevalent in children born very preterm (Doesburg, Ribary
et al. 2011; Doesburg, Chau et al. 2013; Doesburg, Moiseev et al. 2013). These conditions have been
associated with a general reduction in both local and long-range oscillatory synchronization.
Desynchronization of cortical activity might reflect functional disconnection between brain regions.
This might also be related to cortical slowing, which is characterized as a higher ratio of synchrony in
slow EEG activity (i.e., theta, alpha) relative to fast oscillations (i.e., higher gamma frequency)
(Monastra, Lubar et al. 1999; Monastra, Lubar et al. 2001; Englot, Yang et al. 2010). Cortical slowing
has been associated with impaired perception and has also been reported in many clinical populations
including attention deficits (Monastra, Lubar et al. 1999; Monastra, Lubar et al. 2001), epilepsy (Englot,
Yang et al. 2010), Alzheimer’s (Petit, Montplaisir et al. 1992) and Parkinson’s disease (Stoffers,
Bosboom et al. 2007) among others. These abnormalities underlie functional impairments in
cognition/perception (Stoffers, Bosboom et al. 2007; Konrad and Eickhoff 2010; Uhlhaas and Singer
2010; Uhlhaas and Singer 2011) and atypical cortical maturation (Konrad and Eickhoff 2010;
Doesburg, Ribary et al. 2011; Doesburg, Moiseev et al. 2013).
The role of synchrony in the impaired auditory system has not yet been defined however.
Given that the auditory pathways change during the period of bilateral and unilateral deafness and that
CIs do not restore normal hearing, we hypothesized that cortical network connectivity is altered in
adolescents who are deaf and have developed a mature auditory system with long-term unilateral CI
stimulation. In the present study, we asked how cortical networks underlying auditory processing are
affected by long-term unilateral CI use in adolescents who were congenitally deaf in both ears and
received a CI in only their right ear as babies. Children with over a decade of hearing experience with
this unique device have developed a mature auditory cortex as a result (Jiwani, Papsin et al. 2013).

122
They have subsequently received a second implant in their opposite deprived left ear, allowing us to
assess whether the long-term absence of auditory input in these naïve pathways compromises the
neural networks that support hearing in these adolescents. In the present study, we aimed to 1) map
the cortical areas that are working together to support CI listening and 2) define how the cortical
hearing network compensates for reorganization of the brain caused by long periods of unilateral
stimulation/deprivation.
5.3 Methods
5.3.1 Participants and evoked potentials recordings
Electrically-elicited cortical responses were recorded from thirty-four adolescents who received
a unilateral CI in their right ear at young ages (3.2 ± 1.3 years) and within limited durations of bilateral
deafness (1.8 ± 1.3 years). These adolescents had over a decade (12.0 ± 2.1 years) of experience with
their unilateral CI at the time of the test. Twenty-one of them then received a second implant in the
opposite-left ear at 15.9 ± 1.7 years of age, after 11.5 ± 1.7 years of unilateral deprivation on this side.
Responses were recorded from the bilaterally implanted adolescents within the first week of activation
of the new side. All participants used a CI device from Cochlear Limited in both ears. Etiology of
deafness varied. Ten adolescents had deafness associated with a genetic mutation, two had a Mondini
malformation, three had an enlarged vestibular aqueduct, and three lost their hearing after a meningitis
infection. Etiology of deafness was of unknown nature in the other sixteen adolescents. Table 1
provides the mean ± standard deviation (SD) demographic information.
Cortical responses were recorded from 64-cephalic electrodes and referenced to the right
earlobe. Responses were evoked using a 36ms pulse train of 250 pulses per second at a rate of 1Hz
and delivered to a single electrode at the apical end of the electrode array (#20). Stimulus intensity
levels for each ear were determined by recording auditory brainstem responses. Current levels were
increased to maximize the amplitude growth of the response within comfort levels for the child.
Stimulus levels were then balanced between both ears, as in previous reports (Salloum, Valero et al.
2010; Gordon, Wong et al. 2013; Jiwani, Papsin et al. Submitted), by matching the amplitudes of peak
eV when the experienced-right and naïve-left implants were stimulated separately.

123
The same responses were recorded from the right and left ears of sixteen normal hearing
adolescents who were 15.9 ± 6.4 years of age and matched for age and time-in-sound experience at
the time of the test. A 500Hz tone-burst with a 2-1-2 cycle (rise-plateau-fall) lasting 36ms was used to
stimulate the auditory system in these normal hearing peers. Tones were presented at 40dB sensation
level. This tone was chosen to mimic the stimulus delivered to the CI users, because 500Hz is the
frequency which has been assigned to electrode 20 in their internal array. Tone bursts were presented
at a rate of 1Hz and enveloped in a tapered cosine window, called a Tukey window, to minimize effects
of spectral leakage over the first and last eights of each tone. A minimum of 400 sweeps with two
visual replications of the cortical evoked waveforms were recorded. All sweeps with activity ±100µV
were rejected from the average. Responses were sampled at a rate of 1000Hz. An online band-pass
filter between 0.05 to 100Hz was applied with a 12dB/octave rolloff.
Table 5-1: Demographic Information
Normal Hearing
(mean ± SD) (years)
CI Users
(mean ± SD) (years)
Age of first implant - 3.2 ± 1.3
Age of second implant - 15.9 ± 2.0
Age at Test 15.9 ± 6.4 15.9 ± 1.7
Duration of Bilateral deafness - 1.8 ± 1.3
Duration of Unilateral hearing - 12.0 ± 2.1
Duration of Unilateral deafness - 11.5 ± 1.7
Duration of Bilateral Hearing 15.9 ± 6.4 0
Time-in-Sound 15.9 ± 6.4 12.4 ± 1.7

124
5.3.2 Phase synchronization analysis
CI-evoked cortical waveforms are contaminated by a large artefact from the CI device which
can obscure the response and render analyses of cortical activity impossible (Miller, Abbas et al. 2000;
Gilley, Sharma et al. 2006; Martin 2007; Friesen and Picton 2010; Castañeda-Villa and James 2011).
This artefact was removed from the recording with independent component analyses (Gilley, Sharma
et al. 2006; McMenamin, Shackman et al. 2010; Castañeda-Villa and James 2011; Viola, De Vos et al.
2012; Fatima, Quraan et al. 2013; Mc Laughlin, Lopez Valdes et al. 2013) using the extended-runica
Infomax ICA decomposition function in the EEGLab Matlab toolbox (Delorme and Makeig 2004).
Independent components representing the CI artifact were identified visually and removed from the
epoched recordings. Additional details on this method are provided in the Supplementary Materials
section. A scalp current density (SCD) analysis was then performed on the data using a MATLAB
script that implements a spherical spline operator algorithm (Perrin, Pernier et al. 1989) to reduce
effects of spurious synchronization caused from volume conduction as SCD more strongly
corresponds to superficial generators near the recording electrode, and accurately estimates the true
synchrony in the response. This is because SCD measures are much less influenced by the location of
the reference electrode. Additional details on this process are also provided in the Supplementary
Materials section.
Inter-electrode phase-locking values (PLV) were calculated at each time point for each 1Hz
frequency from 4Hz to 60Hz (passband = f ± 0.05f, where f represents the filter frequency) of the
SCD data across all 171 electrode pairs for 19 electrodes uniformly distributed on the scalp. The time-
frequency amplitude responses was also analyzed for the same 19-electrode montage This subset of
electrodes was selected from the 64-channel montage to avoid spurious synchronization originating
from common sources, which can often occur when measuring synchronization between nearby
electrodes. This was measured both for the individual data and for the group data to allow us to assess
temporally coordinated responses to sound across brain regions. PLV analysis takes the EEG
waveform and calculates the instantaneous phase of the signal for each trial, at each frequency and for
every latency point using a Hilbert transform, which also produces time series of instantaneous
amplitude values representing the signal envelope (similar to power). Inter-electrode PLVs were then
calculated by measuring the consistency of the phase angle between two electrodes, at a given
frequency and time point, across trials (Le Van Quyen, Foucher et al. 2001; Bruns 2004; Le Van Quyen
and Bragin 2007).

125
The advantage of using the Hilbert transform is that it can be used to calculate the phase over
relatively short epochs (Doesburg and Ward 2009). A disadvantage of this method however, is that it
introduces distortions to the beginning and end of the analyzed epochs, with stronger effects in the
lower frequencies. To account for these edge effects, epochs were extended in both directions relative
to stimulus onset from 1250ms before stimulus onset to 2000ms after stimulus. This allowed latencies
outside of the relevant time-window to be affected by the Hilbert transform analyses while protecting
the activity in relevant latencies from contamination. Amplitude and PLV analyses were normalized
relative to a 250ms pre-stimulus baseline window to eliminate synchronization of activity unrelated to
the auditory input. Normalization of PLVs was accomplished by subtracting the mean baseline PLV
from the PLV for every data point and dividing the difference by the standard deviation of baseline
PLV (Doesburg and Ward 2009). Baseline-corrected inter-electrode auditory-elicited synchronization
was then plotted on time-frequency plots for stimulation of the right and left ears in normal hearing
adolescents, and the experienced and naïve ears in CI users.
A surrogate distribution of neural synchrony, as measured by PLVs, was then calculated to
assess the statistical reliability in synchronization and desynchronization (Le Van Quyen, Foucher et
al. 2001; Le Van Quyen and Bragin 2007). This was done by comparing the changes in synchrony to
a null distribution derived from the data. To do this, epochs were randomly shuffled 200 times and
normalized PLVs were re-computed for each frequency and at every latency-point for the scrambled
data, thereby representing a null distribution of synchronization. Only changes in PLV exceeding the
97.5th percentile or below the 2.5th percentile were considered to reflect significant increases or
decreases in synchrony at p<0.05. Significant activity was plotted on topographic brain maps for the
analyzed frequencies at 25ms latency intervals, from 25ms to 800ms following stimulus onset (only
activity between 50 and 300ms is shown, as these were considered most relevant to the present
discussion). A difference analysis of the amplitude and PLVs, and a corresponding surrogate
distribution of phase-locking values, was then calculated to compare neural synchrony for right ear
stimulation between the normal hearing data and the experienced ear in the CI users, and left ear
stimulation between normal hearing adolescents and the CI naïve ear. This was done by subtracting
the activity evoked by CI stimulation from the normal hearing data.

126
5.4 Results
5.4.1 Less cortical synchrony is evoked from tone-bursts in normal right than
left ears
Cortical activity elicited by sound was assessed to map the underlying neural network that
supports hearing in typically developing adolescents. As shown from the 64-channel grand mean
average-referenced cortical response (i.e., butterfly plot) in Figures 5.1A and 5.1E, normal hearing
adolescents developed polyphasic cortical waveforms with peaks P1, N1 and P2 consistent with a mature
auditory cortex (Albrecht, Suchodoletz et al. 2000; Ponton, Eggermont et al. 2000; Ponton, Eggermont
et al. 2002; Wunderlich, Cone-Wesson et al. 2006). The distribution of topographic potentials is similar
between both ears; however, as shown by the frequency coherence plots, underlying temporally
coordinated activity in the brain differs for stimulation of the right (Figures 5.1B, 5.1C and 5.1D)
versus left ear (Figures 5.1F, 5.1G and 5.1H) in response to a tone-burst stimulus.
The amplitude time-frequency plot in Figure 5.1B indicates that bursts of increased gamma-
band amplitude centered at ~40Hz were observed, which were prominent over frontal regions (Figure
5.1D), following tones presented to the right ear. This coincided with an amplitude decrease in the
theta and alpha frequencies over parietal electrodes in both hemispheres, corresponding to local
changes in synchronized activity (Doesburg, Emberson et al. 2008; Doesburg, Roggeveen et al. 2008).
By contrast, this activity was reversed for the left ear. Gamma-band activity was reduced relative to
baseline (Figure 5.1F) and compared to the right ear, and occurred in concert with an amplitude
increase in theta and alpha activity across the temporo-parietal regions (Figure 5.1H). The pattern of
long-range oscillatory synchronization shown in Figures 5.1C and 5.1D for the right ear and Figures
5.1G and 5.1H for the left ear, reflective of coordinated activity between distant brain regions rather
than local changes in activity, indicated different long-range neural synchronization between
stimulation of both ears in our cohort of normal hearing adolescents. This is in line with the observed
differences in local EEG amplitude between both ears (Figures 5.1B and 5.1F). Auditory stimulation
of the right and left ears elicited bursts of long-range gamma oscillations, with the strongest
synchronization peaking between 50 and 60Hz. Most of this activity occurred in the latencies
underlying the cortical response and was identified primarily at electrodes over temporal and parietal
areas in both hemispheres and frontal regions in the right cortex. Right frontal activity was more

127
pronounced for left ear stimulation (Figure 5.1G). Of note, the left ear also elicited 30Hz-low
frequency gamma bursts between electrodes at left and right temporal cortex and frontal regions, as
shown in Figures 5.1F and 5.1G, indicating greater left ear-evoked activity in the brain compared to
the right ear. This is supported by observations of increased theta coupling in the left ear in Figure
5.1F, which has been suggested to regulate long-range communication within and across brain regions
and organize task-relevant information in the brain (Doesburg, Green et al. 2012).

128

129
Figure 5-1: Cortical oscillatory synchronization evoked by auditory stimulation of the right and left
ears of normal hearing adolescents.
Auditory cortical responses were recorded from the right and left ears separately in 13 normal hearing
adolescents aged 15.9 ± 6.4 years. Cortical activity elicited by stimulation of the right ear is shown in
A – D. Left ear evoked activity is shown in E – H. Grand mean average-referenced waveforms (i.e.,
butterfly plot) of the 64-channel cortical responses with the waveform recorded at a midline cephalic
electrode (Cz) highlighted in red for right ear stimulation (n=13) is shown in A. and in blue for left ear
stimulation (n=12) shown in E. Mean global field power (GFP) head topographies for each peak of
the cortical waveform are shown above the response for both ears (red = positive potentials, blue=
negative potentials, green = baseline). Normalized group mean amplitude activity, reflecting of local
changes in synchrony are shown in B. and F. for the right and left ears respectively for 4 to 60 Hz.
The right ear shows bursts of gamma band activity (red) with reduced low frequency amplitude (blue).
This activity is reversed in the left ear, where local changes in synchrony are more predominant in
oscillations in the theta, alpha and beta bands and reduced for gamma. C. & D. Normalized group
mean changes in long-range synchrony are plotted relative to baseline for frequencies from 4 to 60Hz
for latencies ranging from -200 to 800ms for the right ear in C. Topographic representation of this
activity is shown in D. for theta (4Hz), alpha (10Hz), low gamma (30Hz) and high gamma (60Hz)
frequencies for each 25ms latency interval from 50 to 300ms following stimulus onset. The black lines
indicate statistically significant increases in phase-locking activity between electrode pairs (>97.5th
percentile of the surrogate distribution), and the while lines show statistically significant decreased
activity (<2.5th percentile). These are superimposed over local changes in amplitude which are
represented by the colour gradients on the topographic brain plots. This activity is plotted for the left
ear in G. & H. Both ears show increased synchrony in the theta, beta and gamma frequency bands
but this activity is greater in the left ear, indicating that the left ear evokes greater cortical activity than
the right.

130
5.4.2 Right cochlear implants promote atypical cortical synchrony and leave
deprived pathways abnormally desynchronized
Long-term unilateral CI use appears to promote typical development of cortical responses. As
shown in Figure 5.2A, adolescents who had over a decade of CI stimulation in the right ear developed
cortical waveforms which were comprised of the mature peaks P1, N1 and P2. The latency (P1:
t(10)=−0.88, p>0.05; N1: t(10.18)=−1.3, p>0.05; P2: t(10.77)=1.43, p>0.05) and peak-to-peak amplitude (P1-
N1: t(6.87)=1.75, p>0.05) of cortical wavepeaks were generally similar between both groups, except that
peak P2 was significantly larger (t(14.51)=2.49, p<0.05) and global field power scalp amplitudes was
stronger (±2µV) in the CI users compared to either ear of the normal hearing group. On the newly
implanted side, initial stimulation of the naïve pathways shown in Figure 5.2E evoked cortical
responses, which had a significantly different morphology than the responses recorded from normal
(Figures 5.1A and 5.1E) and the experienced side (Figure 5.2A). Here, the morphology of the
waveform was characterized by a large negative amplitude biphasic response, N(ci), followed by a large
positive peak, P(ci). We have described this response in a previous report and suggested that it might
index abnormal or immature underlying activity in the brain (Jiwani, Papsin et al. Submitted). Peak-
to-peak amplitude of this response was significantly larger than any of the mature wavepeaks recorded
from the normal hearing group (F(3,70)=5.4, p<0.05) and the experienced ear (t(11)=-3.37, p<0.01), with
a stronger mean global field power amplitude (±5µV).
Analyses of changes in amplitude, phase-locking values and inter-electrode synchrony further
highlighted differences in cortical processing of sound evoked by CI stimulation compared to normal
hearing adolescents. Neural synchronization elicited by the experienced-right CI after long-term
unilateral experience was increased compared to normal hearing peers in both ears, shown in Figures
5.2B, 5.2C and 5.2D, whereas this activity was significantly and globally reduced on the naïve side
(Figures 5.2F, 5.2G and 5.2H). A general reduction in amplitude of the signal across the different
frequency bands was observed with CI stimulation in both the experienced (Figure 5.2B) and naïve
ears (Figure 5.2F) in latencies greater than 150 to 200ms. Of interest, on the experienced side only,
this activity was preceded by an increase in theta, beta and gamma-band amplitude over left temporal
and frontal electrodes in a narrow latency range underlying the cortical response. Long-range
synchronized activity was also greater on the experienced side compared to the naïve ear and normal
hearing peers, as shown in Figures 5.2C and 5.2D. Auditory stimulation of the experienced-right

131
implant evoked a pattern of theta activity that was maintained across a greater latency range (100 to
700ms) and spread across a larger network compared to typically developing normal hearing controls.
A sustained increase in theta activity over left temporal and left parietal regions to frontal electrodes
in both hemispheres was observed. These connections became more distributed across brain regions
(in electrodes over the temporal, parietal and frontal regions) and linked to the right parietal and right
visual cortices in later latencies (Figure 5.2D). These effects are clearly visible in a direct comparison
of long-range synchronization in long-term CI users and normal hearing peers shown in Figures 5.3A
and 5.3B. Theta activity was accompanied by sustained bursts of gamma-band synchronization
between left temporal, parietal and occipital regions to other electrodes, suggestive of increased
perceptual processing and perhaps increased alertness/attention in response to sound (Doesburg,
Green et al. 2012). Of interest, however, gamma activity exceeding 35Hz was reduced in the CI group
compared to normal, as shown in Figures 5.3A and 5.3B, and occurred with a greater-than-normal
ratio of alpha-band synchronization in later latencies (underlying complex perceptual processing)
(Figure 5.3A) across a distributed cortical network (Figure 5.3B), indicative of cortical slowing in
response to sound.
By contrast, long-range synchronization evoked by the naïve-left ear after over a decade of
unilateral deprivation on that side indicated generally desynchronized activity. Theta, beta and gamma-
band oscillations shown in Figure 5.2G were reduced across the cortex relative to baseline and
compared to the experienced side and normal hearing peers. A pattern of robust and sustained long-
range alpha synchrony was observed over widespread electrode locations in posterior, medial, temporal
and frontal regions (Figure 5.2H). A difference analysis of long-range synchronization with normal
hearing left ear stimulation in Figures 5.3C and 5.3D was consistent with these patterns of
oscillations, and indicated reduced cortical connectivity in all but the alpha-band frequencies at initial
stimulation of the naïve-left CI. This is in contrast to the synchronization observed from normal
hearing adolescents and the opposite experienced-right CI side, where theta and gamma activity,
believed to reflect perception and attention (Jensen, Kaiser et al. 2007; Doesburg, Emberson et al.
2008; Doesburg, Roggeveen et al. 2008; Green, Doesburg et al. 2011; Doesburg, Green et al. 2012),
was increased. General patterns of local (Figure 5.2F) and long-range desynchronization (Figures
5.2G, 5.2H, 5.3C and 5.3D) elicited by the newly implanted ear indicate abnormal network
connectivity in the brain after long-term unilateral auditory deprivation.

132

133
Figure 5-2: Cortical oscillatory synchronization evoked by auditory stimulation of the experienced-
right and naïve-left ears in CI users.
Auditory cortical responses were recorded from the experienced-right ear (n=34) and at initial
activation of the naïve-left (n=21) implant separately in adolescents who received a unilateral right CI
within limited durations of bilateral deafness, and received a second one on the naïve-left side after
over a decade of unilateral deprivation. Cortical activity elicited by stimulation of the experienced-
right after 12.0 ± 2.1 years of unilateral CI use is shown in A. – D. Left-naive ear elicited activity is
shown in E. – H. A. Grand mean average-referenced waveforms of the 64-channel cortical responses
evoked by the experienced-right CI show 3 peaks, P1, N1, P2, characteristic of a mature response, but
this responses evoked at initial stimulation of the naïve-left side shown in E. has an abnormal
morphology with a biphasic peaked waveform (N(ci), P(ci)) of abnormally large amplitudes. Mean
global field power (GFP) head topographies for each peak of the cortical waveform are shown above
the response for both ears (red = positive potentials, blue= negative potentials, green = baseline).
Normalized group mean amplitude activity reflective of local changes in synchrony is shown in B. and
F. for the experienced-right and naïve-left ears respectively for 4 to 60 Hz. An increase in alpha, beta
and gamma amplitude occurs in a narrow latency range underlying the cortical waveform on the
experienced side, shown by the red. Amplitude activity is generally reduced thereafter for both ears,
shown in blue. C. & D. Normalized group mean changes in long-range synchrony are plotted relative
to baseline for frequencies from 4 to 60Hz for latencies ranging from -200 to 800ms for the
experienced right ear in C. Topographic representation of this activity is shown in D. for theta (4Hz),
alpha (10Hz), low gamma (30Hz) and high gamma (60Hz) frequencies for 25ms latency interval from
50 to 300ms following stimulus onset. The black lines indicate statistically significant increases in
phase-locking activity between electrode pairs (>97.5th percentile of the surrogate distribution), and
the while lines show statistically significant decreased activity (<2.5th percentile). These are
superimposed over local changes in amplitude which are represented by the colour gradients on the
topographic brain plots. Neural synchrony is increased across the frequency bands for the
experienced-right ear. Increased synchrony is particularly evident for theta and gamma activity
indicating increased cortical processing to sound. Long-range neural synchrony for the naïve-left ear
is plotted in G. & H. As shown, an increase in neural synchronization occurs for the alpha frequency
bands. This occurs with a general widespread de-synchronization of activity in the other frequency
bands.

134
Figure 5-3: Difference in cortical oscillatory synchronization evoked by auditory stimulation of the
right/experienced and left/naive ears/implant of normal hearing adolescents and CI users.
A. Difference PLV time-frequency plots between normal hearing peers and CI users for each ear (red
= > synchronization in CI users, blue = > synchronization in normal hearing peers). B. Topographic
maps showing significantly differences in long range phase synchronization between normal hearing
peers and experienced CI users for theta, alpha, and gamma frequencies from 50 to 300ms (black lines
= increased phase-locking in CI users, white lines = increase phase-locking in normal hearing).
Gamma frequencies are reduced in the implant users, particularly at higher frequencies, but long-term
CI stimulation evokes greater than normal theta-band activity across a widespread network. This might
serve to maintain attention across distant brain regions for auditory processing. Altered inter-regional
synchronization elicited by the naïve-left side at initial stimulation of the newly implanted side
compared to normal hearing left ear activity is shown in C. and D. The dominance of blue in C. and
white lines in D. indicates that activity is significantly reduced and de-synchronized on the naïve-left
side compared to normal.

135
5.5 Discussion
5.5.1 A specialized cortical hearing network normally matures by adolescence
Results indicated that patterns of neural synchronization depend on the ear of stimulation in
normal hearing adolescents. Gamma and theta synchrony in the temporo-parietal regions of both
hemispheres as well as the right frontal cortex were increased for left compared with right ear
stimulation as shown in Figure 5.1. This suggests increased network connectivity in response to a
500Hz tone-burst in line with findings of specialization to this stimulus in the mature brain of normal
hearing adolescents. Regardless of ear of presentation, the same cohort showed strong right
hemisphere activity (Jiwani, Papsin et al. Submitted) consistent with previous reports (Hine and
Debener 2007; Schönwiesner, Krumbholz et al. 2007). The previous findings together with strong
theta and gamma activation in the right cortex from left ear stimulation, observed in this study, might
underlie mechanisms of hemispheric specialization in response to tonal stimuli (Kimura 1973;
Penhune, Zatorre et al. 1996; Zatorre and Belin 2001; Zatorre, Belin et al. 2002; Schönwiesner,
Rübsamen et al. 2005; Jamison, Watkins et al. 2006; Schönwiesner, Krumbholz et al. 2007). Of note,
cortical asymmetries between both hemispheres were not present in younger peers with an immature
auditory cortex (Gordon, Wong et al. 2013). A relative specialization in the right auditory hemisphere
for spectral processing and a temporal processing bias in the left auditory cortex (Zatorre and Belin
2001; Zatorre, Belin et al. 2002), which is mediated by inter-hemispheric structural differences
(Penhune, Zatorre et al. 1996), thus appears to develop with experience and age (Penhune, Zatorre et
al. 1996; Kadis, Pang et al. 2011).
It has been argued that the different specialization of the left and right hemispheres in the brain
arose as an evolutionary strategy to increase processing efficiency (Rogers 2000; Dharmaretnam and
Rogers 2005; Vallortigara 2006; MacNeilage, Rogers et al. 2009). Cortical specialization reduces
redundant processing in one hemisphere and allows the other side of the brain to perform additional
functions, thereby increasing neural capacity in the cortex (Vallortigara 2006). Normal hemispheric
specialization also increases the brain’s ability to perform parallel processing in the two hemispheres
and attend to two tasks simultaneously (Dharmaretnam and Rogers 2005). Larger amplitude activity
in the right cortex evoked by stimulation of both ears shown in Figures 5.1B and 5.1F and stronger
long-range network connectivity of the gamma-band network across temporal, parietal and right
frontal regions with left ear stimulation in Figures 5.1G and 5.1H, could thus underlie the functional

136
organization of cortical specialization to a 500Hz tone-burst. Indeed, increased network
synchronization (Barry, Clarke et al. 2004) and spectral power (Clarke, Barry et al. 2001) in theta, beta
and gamma frequencies such as that observed in Figure 5.1 have been shown to mark maturity and
efficiency in brain networks (Marosi, Harmony et al. 1992; Marosi, Harmony et al. 1997; Uhlhaas, Roux
et al. 2010). The pattern of cortical synchronization observed from our group of normal hearing peers
is in line with reports indicating that spectral processing occurs in the belt and parabelt regions of the
right auditory hemisphere (Zatorre, Belin et al. 2002; Rauschecker and Tian 2004; Schönwiesner,
Rübsamen et al. 2005) and interact with frontal regions for memory encoding and retrieval (Zatorre
and Samson 1991; Zatorre, Evans et al. 1994; Zatorre, Belin et al. 2002).
In addition to cortical specialization, functional correlates of theory of mind and mentalizing
become established during adolescence, as the brain matures (Blakemore 2012). Development of these
cognitive processes coincides with the maturation of the pre-frontal cortex (Moriguchi, Ohnishi et al.
2007) and are related to improvements in executive function (Anderson, Anderson et al. 2001) and
learning (Johnson 2011) that occur with age. These cognitive processes involve cortical activation of
a network of regions similar to that observed in Figure 5.1 (Blakemore and Choudhury 2006; Burnett,
Bird et al. 2009; Burnett and Blakemore 2009; Blakemore 2012), with activity being particularly
stronger in anterior cortical regions (i.e., medial prefontal cortex and inferior frontal gyrus) during
earlier adolescence and moving to posterior and dorsal regions in adulthood (Moriguchi, Ohnishi et
al. 2007; Blakemore 2012). The dynamic patterns of oscillations observed in Figure 5.1 from our
cohort of adolescents with normal hearing might thus reflect underlying processes of mentalizing,
which occur during passive listening tasks (Brown, Martinez et al. 2004; Wilson, Saygin et al. 2004;
Buckner, Andrews‐Hanna et al. 2008). This is supported by our findings of increased theta and beta
modulated changes of the gamma-band network which have been reported to underlie mechanisms of
response anticipation, attention preparation and/or perceptual readiness (Doesburg, Roggeveen et al.
2008; Uhlhaas, Pipa et al. 2009). This very pattern of activity likely governs complex auditory skills
(Ponton, Eggermont et al. 2000) and cognitive/perceptual processes of multi-sensory
attention/interaction (Posner and Dehaene 1994; Busse, Roberts et al. 2005; Fan, McCandliss et al.
2005; Awh, Vogel et al. 2006) which only become adult-like during adolescence (Ponton, Eggermont
et al. 2000; Eggermont and Ponton 2003). This development is coincident with the maturational time-
course of the auditory brain (Albrecht, Suchodoletz et al. 2000; Ponton, Eggermont et al. 2000; Ponton
and Eggermont 2001; Ponton, Eggermont et al. 2002; Wunderlich and Cone-Wesson 2006;

137
Wunderlich, Cone-Wesson et al. 2006; Jiwani, Papsin et al. 2013) and hemispheric specialization
(Davidson 1984; Penhune, Zatorre et al. 1996; Rogers 2000; Zatorre and Belin 2001; Zatorre, Belin et
al. 2002; Rivera, Reiss et al. 2005; Jamison, Watkins et al. 2006; Zatorre, Chen et al. 2007; Johnson
2011). The dynamic pattern of synchronization observed from our cohort of adolescents with normal
hearing thus suggests appropriate development of cortico-cortical connections.
Similar perceptual processes were likely activated by sound in CI users as well, since they
developed a mature auditory cortex (as indicated by mature cortical wavepeaks, P1, N1, P2) (Figure
5.2A), which was underlied by patterns of gamma synchronization which also appeared to be
modulated by theta and beta oscillations, as shown in Figure 5.2C. However, abnormalities relative
to normal hearing peers remain. Atypical organization of the auditory network in CI users is indicated
by the abnormally large peak P2 amplitude, as previously reported (Jiwani, Papsin et al. 2013), unusually
strong contralateral lateralization to the left auditory cortex (Jiwani, Papsin et al. Submitted), and
greater-than-normal coordinated synchronization in the brain (Figures 5.2 and 5.3).
5.5.2 Increased connectivity in long-term unilateral CI users reflects greater
processing demands
As shown in Figures 5.2 and 5.3, cortical responses evoked by stimulation of the experienced-
right CI were underlied by increased gamma synchronization over the left temporal, left parietal and
frontal regions in both hemispheres which appeared to be modulated by greater-than-normal theta and
beta oscillations. Increased gamma-band synchronization is linked to selective attention, short and
long-term memory encoding/retrieval and multisensory integration (Jensen, Kaiser et al. 2007;
Uhlhaas, Pipa et al. 2009; Uhlhaas, Roux et al. 2010). This activity increases in inferior frontal, posterior
parietal and anterior temporal cortical regions (association auditory areas) during both passive and
attentive listening tasks (Kaiser 2000, Kaiser 2002, Kaiser 2002, Kaiser 2004; Kaiser and Lutzenberger,
2005). These mechanisms are mediated by an attention control network which is represented by
stronger theta activity over the superior temporal lobe (Green, Doesburg et al. 2011). The pattern of
synchronization observed in the present study might thus indicate an enhanced role for attention in
adolescents who have used a CI to hear for most of their lives compared to typically developing
adolescents, even in a passive listening task evoked by non-speech stimuli. This is further supported

138
by observations of decreased local alpha amplitude in Figure 5.2B and increased long-range alpha-
band synchronization in the latency range of peak P2 (Figure 5.2C and 5.2D), which reflect
maintenance of attention (Cooper, Croft et al. 2003; Ward 2003; Palva and Palva 2007; Doesburg,
Roggeveen et al. 2008; Palva and Palva 2011), in CI users.
Individuals with long-term CI experience have been shown to recruit cortical areas linked to
attention (Posner and Dehaene 1994; Pugh, Shaywitz et al. 1996; Grady, Van Meter et al. 1997;
Benedict, Lockwood et al. 1998; Fujiwara, Nagamine et al. 1998; Jäncke, Mirzazade et al. 1999; Fan,
McCandliss et al. 2002; Fan, McCandliss et al. 2005; Okamoto, Stracke et al. 2007). These areas may
be used differently than their peers with normal hearing to facilitate listening and auditory processing
(Jeong Lee, Kang et al. 2005; Lee, Giraud et al. 2007). Activation of the left fronto-parietal network
(including dorsolateral prefrontal and inferior parietal cortices) has been associated with better speech
perception outcomes in CI users (Lee, Giraud et al. 2007). This indicates that recruitment of cognitive
mechanisms related to attention and working memory increase communication successes after
cochlear implantation (Cleary, Pisoni et al. 2001; Jeong Lee, Kang et al. 2005; Giraud and Lee 2007;
Lee, Giraud et al. 2007; Pisoni, Conway et al. 2010; Pisoni, Kronenberger et al. 2011; Geers, Pisoni et
al. 2012; Kronenberger, Colson et al. 2014). Increased attention may contribute to the significant
improvements on speech and language comprehension and production (Geers, Nicholas et al. 2003;
Geers, Tobey et al. 2008; Geers and Sedey 2011) and improvements on measures of verbal memory
capacity (Cleary, Pisoni et al. 2001; Pisoni, Kronenberger et al. 2011; Geers, Pisoni et al. 2012) reported
from CI users over the long-term. In line with these reports, our present findings suggest that
adolescents using CIs to hear capitalize on general cognitive strategies to process sounds. It is possible
that CI hearing drives increased cognitive load on the stimulated side (Kral, Heid et al. 2013), perhaps
reflecting maladaptive (Sandmann, Dillier et al. 2012) or compensatory (Kral and Sharma 2012;
Sharma, Campbell et al. 2014) cortical plasticity after long-term unilateral CI stimulation/deprivation.
Connectivity of the theta and gamma-band network with occipital cortical regions (Figure 5.2) further
suggests that CI users also supplement hearing with vision. The addition of visual input likely serves
to facilitate listening to the degraded auditory cues delivered by a CI (Rubinstein 2004; Drennan and
Rubinstein 2008) and to compensate for auditory limitations associated with bilateral deafness prior to
implantation (Finney, Fine et al. 2001; Lee, Lee et al. 2001; Bavelier and Neville 2002; Bavelier, Dye et
al. 2006; Lomber, Meredith et al. 2010; Meredith and Lomber 2011) and unilateral deprivation

139
afterward (Popescu and Polley 2010; Gordon, Jiwani et al. 2013; Gordon, Wong et al. 2013; Kral, Heid
et al. 2013; Kral, Hubka et al. 2013; Jiwani, Papsin et al. Submitted).
More attentive listening with increased reliance on vision to process auditory input could mean
increased effort and perhaps reduced efficiency for CI listening. Reduced gamma-band synchrony in
frequencies above 35Hz recorded from stimulation of the experienced side in adolescents using CIs
compared to normal shown in Figure 5.3, suggests a slowing of cortical processing to sound with CI
stimulation. This is consistent with findings of increased reaction time in response to speech sounds
(Steel, Papsin et al. Submitted) indicating increased listening effort in adolescents with CIs compared
to peers with bilateral implants (Hughes and Galvin 2013) and with normal hearing (Hughes and
Galvin 2013; Pals, Sarampalis et al. 2013). Indeed, adolescents using CIs have listened to a degraded
auditory input missing much of its fine temporal cues from only a single CI for most of their lives.
Distorted auditory input and/or decreased speech intelligibility exerts increased mental
processing/effort and uncertainty (Zekveld, Kramer et al. 2010; Zekveld, Kramer et al. 2011;
Koelewijn, Zekveld et al. 2012), which could mean slower auditory processing and in turn reduced
efficiency for CI listening. Increased effort and attention of the auditory brain for processing sounds
may be attributed to a reduction in inhibitory input from the opposite pathways which were left
deprived of input for over a decade (Vale, Juíz et al. 2004; Takesian, Kotak et al. 2009; Sanes and Kotak
2011). Indeed, investigators have shown that leaving the auditory system unilaterally deprived during
early stages of development drives a strengthening of activity in favor of the stimulated ear from
brainstem (Moore and Kowalchuk 1988; Moore 1990; Kil, Hkageyama et al. 1995; Kitzes, Kageyama
et al. 1995; Nordeen, Killackey et al. 2004) to cortex (Gordon, Wong et al. 2013; Kral, Heid et al. 2013;
Kral, Hubka et al. 2013) and a weakening of ascending connections in the deprived pathways (Popescu
and Polley 2010; Kral, Heid et al. 2013).
5.5.3 Desynchronized activity evoked by the naïve-left CI suggests
disorganization in the deprived pathways
Long-term unilateral stimulation of a right-experienced CI means that pathways of the
opposite-left ear were left deprived of auditory input. We hypothesized that this would lead to a
profound reorganization in the unstimulated pathways with abnormal desynchronized cortical

140
oscillations, reflecting immature and inefficient cortical networks. In line with our hypothesis, cortical
responses evoked by initial stimulation of the newly implanted side after over a decade of deprivation
revealed abnormal underlying activity in the brain (Figure 5.2E). Waveform morphology was
characterized by a biphasic response with abnormally large amplitude. We have described this
response previously and showed that abnormally strong and widespread dipole activity underlie both
cortical peaks N(ci) and P(ci) (Jiwani, Papsin et al. Submitted). This atypical response reflects altered
cortical activity in the deprived pathways (Sharma, Dorman et al. 2002; Gordon, Tanaka et al. 2008)
and might index cortical immaturity (Wunderlich, Cone-Wesson et al. 2006; Gordon, Tanaka et al.
2011). In support, coordinated oscillations underlying this response in Figure 5.2 also indicated
abnormal cortical activity. Spectral power (Figures 5.2F) and phase-locking values (Figure 5.2G)
were desynchronized in the theta, beta and gamma frequencies. The reduced temporal coherence
observed from the deprived side could indicate a lack of familiarity or meaninglessness with the
presented stimulus, as such global decreases in neural synchrony are associated with perception of
meaninglessness (Rodriguez, George et al. 1999). Local desynchronization of neuronal assemblies
might also reflect global inhibition (Cooper, Croft et al. 2003; Ward 2003) and/or functional dis-
connectivity between brain regions during stimulus processing (Wilson, Rojas et al. 2007). Of interest,
reductions in gamma amplitudes (Casanova, Buxhoeveden et al. 2002; Casanova, Buxhoeveden et al.
2003; Wilson, Rojas et al. 2007; Khan, Gramfort et al. 2013) and global long-range phase-locking
connectivity patterns (Hughes 2007; Khan, Gramfort et al. 2013) similar to the oscillations observed
following stimulation of the naïve side (Figure 5.2), are consistent with that of other clinical
populations (Uhlhaas and Singer 2006), including autism (Hughes 2007; Wilson, Rojas et al. 2007;
Tessier and Broadie 2009; Khan, Gramfort et al. 2013). It is possible that abnormal structural and
functional changes (Kitzes, Kageyama et al. 1995; Mossop, Wilson et al. 2000; Moore and Kitzes 2004;
Moore and Kowalchuk 2004; Nordeen, Killackey et al. 2004; Vale, Juíz et al. 2004; Popescu and Polley
2010; Kral, Heid et al. 2013; Kral, Hubka et al. 2013) occurring in the naïve pathways as a result of
long periods of unilateral deprivation drove abnormal network connectivity in those pathways. This
would explain earlier findings of poorer speech perception outcomes on this side even after 8 to 12
months of hearing experience with the second CI (Jiwani, Papsin et al. Submitted).
Alternatively, this pattern of oscillations could reflect immaturity in the deprived pathways.
Synchronization of oscillatory activity undergo significant changes and refinement towards more
precise oscillations during adolescence (Uhlhaas, Roux et al. 2010) which are related to development

141
of anatomical and physiological processes (Giedd, Blumenthal et al. 1999; Paus 2005; Uhlhaas, Roux
et al. 2009) as well as maturation of cognitive and perceptual functions (Varela, Lachaux et al. 2001;
Ward 2003; Uhlhaas, Roux et al. 2009; Blakemore 2012). Both local and long-range neural synchrony
in the theta-band (which are significantly reduced during childhood) increase until early adolescence
over fronto-central regions during auditory processing (Müller, Gruber et al. 2009), and in the beta and
gamma-bands over occipital (Werkle-Bergner, Shing et al. 2009) and parietal electrodes (Uhlhaas, Roux
et al. 2009) during visual processing. By contrast, this activity remains abnormally desynchronized in
earlier stages of immaturity of the preterm brain. A slowing of cortical oscillations (Doesburg, Ribary
et al. 2011; Doesburg, Chau et al. 2013; Doesburg, Moiseev et al. 2013) and weaker resting-state
network connectivity (Damaraju, Phillips et al. 2010) has been reported in children born under <32
weeks gestational age compared to children born at term. This indicates altered thalamo-cortical and
cortical dynamics in the preterm brain. The generally reduced oscillatory activity evoked by stimulation
of the naïve CI compared to the experienced ear (Figure 5.2) and normal hearing peers (Figure 5.3)
could thus be characteristic of earlier stages of cortical immaturity. This would explain the atypical
negative peaked cortical waveform similar to that shown in Figure 5.2E recorded from normal hearing
preterm babies under 35 weeks gestation (Wunderlich, Cone-Wesson et al. 2006) and children with
limited auditory input due to GJB-2 deafness (Gordon, Tanaka et al. 2011).
Cortical immaturity has been associated with oscillatory slowing (i.e., a shift in activity towards
slower frequencies) and in turn, poorer cognitive outcomes in preterm children (Doesburg, Moiseev
et al. 2013) and individuals suffering from Parkinson’s disease (Stoffers, Bosboom et al. 2007). In the
present study, global frequency desynchronization with increased long-range alpha synchrony
(Figures 5.2E-H and 3B) similarly indicates a widespread slowing of activity in the brain, perhaps
reflecting a general weakening for auditory processing in the deprived pathways. This is in agreement
with data from animal models of unilateral deafness (Popescu and Polley 2010; Kral, Heid et al. 2013;
Kral, Hubka et al. 2013) and studies of monocular deprivation in the visual system (Hubel and Wiesel
1970; Hubel, Wiesel et al. 1977; Le Vay, Wiesel et al. 1980; Blakemore 1988; Jeffrey, Wang et al. 2004;
Lewis and Maurer 2005). These studies indicate that lack of input competition between two pathways
drives functional changes in the balance of inhibitory-excitatory circuits (Vale, Juíz et al. 2004; Kotak,
Fujisawa et al. 2005; Takesian, Kotak et al. 2009; Sanes and Kotak 2011) which lead to weakened
activity in the deprived side (Popescu and Polley 2010; Kral, Heid et al. 2013). Significantly poorer
auditory performance (Gordon and Papsin 2009; Illg, Giourgas et al. 2013; Jiwani, Papsin et al.

142
Submitted) and inconsistent use of the CI on this later implanted side (Fitzgerald, Green et al. 2013)
compared to the experienced ear may thus be attributed to deleterious and/or immature development
of auditory pathways in the deprived ear. Given that the capacity for auditory plasticity decreases with
age (Lohmann and Kessels 2014) and sensitive periods in development typically close when maturation
is reached (Blakemore 2012; Lohmann and Kessels 2014), promoting development from this long-
term deprived side during adolescence may thus be challenging.
5.6 Conclusion
Cortical synchronization of neural activity between brain regions is crucial for maintaining
organization of information in cortical circuits and underlies a variety cognitive/perceptual brain
functions. As shown in the present study, normal development of oscillatory activity in the brain
reflects maturation of cortical specialization in adolescents with normal hearing. However, the cortical
auditory network is altered in adolescents who have used a unilateral CI to hear for most of their lives.
Our findings indicate for the first time that driving maturation of the auditory cortex from only one
ear with a CI alters neural dynamics in the auditory brain. Increased cortical synchronization evoked
by stimulation of the experienced-right CI reflects abnormally increased cognitive processing to sound,
perhaps reflecting increased recruitment of attention resources for CI listening. At the same time, this
maturation leaves the opposite deprived pathways with aberrant globally desynchronized connectivity.
This allows the unstimulated pathways to become segregated from the cortical hearing network,
thereby highlighting the deleterious effects of long-term unilateral deprivation.

143
5.7 Supplementary Information for Methods
5.7.1 Independent component analysis to reject cochlear implant artifact
Recording cortical evoked responses in cochlear implant (CI) users poses a particular problem
because the implant device itself produces a large electrical artifact that is recorded by scalp electrodes.
For the most part, this artifact is generated by the radio frequency transmission of the signal from the
processor to the receiver. It is large in amplitude, time-locked to the stimulus and will last for at least
the duration of the stimulus. The artifact thus interferes with the response and can completely obscure
the cortical waveform, in turn making any further analyses impossible. It is characterized by a biphasic
low-frequency pedestal of large amplitude which begins shortly after stimulus presentation and is
followed by a high frequency ringing (Gilley, Sharma et al. 2006). Devices which use monopolar
stimulation (i.e., CI24M, CI24R(CS), CI24RE(CA) and CI513) produce larger amplitude artifacts,
because the electric current from activated electrodes is spread over a larger distance to the return
ground electrode. This causes more distributed/dispersed electrical splattering/activity over the scalp,
compared to CIs that use bipolar stimulation, where each active electrode is paired with its own closely
situated ground electrode (Gilley, Sharma et al. 2006).
Several techniques have been proposed to suppress (Gilley, Sharma et al. 2006; Debener, Hine
et al. 2008; Wong and Gordon 2009; Friesen and Picton 2010; Viola, Thorne et al. 2011; Viola, De
Vos et al. 2012) or remove the electrical artifact from the CI response. One suggested method in single
channel recordings has been to place the reference electrode at a location on the scalp that would yield
the same artifact as the recording electrode, yet, be sufficiently far away from that electrode to ensure
that the recorded potential is neutral from the cortical evoked response (Gilley, Sharma et al. 2006).
This technique is based on the assumption that the CI artifact would be suppressed or cancelled out if
the artifact remains stable and if the reference electrode is placed at a location that would optimize the
differential recording and minimize its contribution to the recording. Another commonly used method
to remove the CI artifact is the subtraction technique. With this method, cortical responses are
recorded in different conditions or with different inter-stimulus intervals, and are then subtracted from
each other (Miller, Abbas et al. 2000; Friesen and Picton 2010). This method assumes that the artifact
remains the same in different recording conditions, even if the neural response is different (Friesen
and Picton 2010). The subtraction method would thus eliminate the artifact, and leave only the
difference waveform as a measure of the cortical response. However, this method requires that

144
additional conditions and additional averages of the cortical response be recorded, as the subtraction
technique introduces residual noise in the difference waveform (Friesen and Picton 2010) and may
distort the evoked response when the nerve is in only a partially refractory state (Miller, Abbas et al.
2000).
Most recently, investigators have used an independent component analysis (ICA) method to
separate artifacts from the neural response (Debener, Hine et al. 2008; Castaneda-Villa, Manuel
Cornejo-Cruz et al. 2010; McMenamin, Shackman et al. 2010; Castañeda-Villa and James 2011; Viola,
Thorne et al. 2011; Viola, De Vos et al. 2012; Fatima, Quraan et al. 2013). Scalp-recorded EEG signals
are a collection of signals from mixed sources which reflect both cortical activity and non-cortical
sources. ICA transforms this data by applying spatial filters over the scalp, using statistical techniques,
which decomposes a multivariate/mixed signal into its individual statistically independent components
(Hyvärinen and Oja 1997; Lee, Girolami et al. 1999; Delorme and Makeig 2004; Naik and Kumar
2011). ICA separation is based on two primary assumptions: 1) the sources of signal and artifact are
independent from each other, even though the mixed signals are not, and 2) source activity has a non-
gaussian distribution (Delorme and Makeig 2004). For the CI artifacts, both of these conditions are
met (Gilley, Sharma et al. 2006). In this way, ICA decomposition is able to spatially filter and identify
independent components produced by distinct sources from those of the cortical evoked activity.
In the present study, 64-channel cortical evoked responses were decomposed into 64
independent components using the extended-runica Infomax ICA decomposition function in the
EEGLab Matlab toolbox (Delorme and Makeig 2004). Independent components were identified
visually, and those representing the CI artifact were removed from the epoched recordings.
Supplementary Figure 5.4A shows an example of the component properties for four independent
components representing the CI artifact, and other common EEG artifacts, such as eye blinks and
60Hz noise, and brain related activity reflecting the cortical evoked response. Topographic distribution
of potentials is shown on the left with the individual trials and averaged activity on the right for each
component. As shown in Supplementary Figure 5.4B, the CI artifact, which occurs between -80
and 40ms, dominates the cortical evoked waveform and completely obscured/contaminated the
underlying distribution of frequency oscillations, as measured by the amplitude and phase-locking
synchrony plots, making this analysis impossible. After the components with the CI artifact were
removed using ICA decomposition, the cortical response and underlying frequency oscillation activity
could be seen, as shown in Supplementary Figure 5.4C.

145

146
5.7.2 Scalp current density to reduce spurious synchronization cause from
volume conduction
In EEG, since cortical evoked responses are recorded from surface sensors, far away from
their generators which are located deep within the scalp, the electric field becomes
distorted/distributed/smeared as it travels from current sources through tissues of varying
conductivities (i.e., brain, cerebral spinal fluid, skull bone and scalp) (van den Broek, Reinders et al.
1998). This imposes a problem of volume conduction, where the information recorded at two adjacent
electrode sites may reflect the integrated activity of shared neural populations (i.e., two electrodes
reflecting activity from the same neural sources/populations) (Lachaux, Rodriguez et al. 1999). This
is particularly exacerbated if the two electrodes are located at less than 3-4 cm from each other (Nunez,
Srinivasan et al. 1997). In turn, this creates spurious activity which can be ‘falsely’ misinterpreted as
synchronously oscillating activity. To reduce such effects of spurious synchronization (caused from
volume conduction) and estimate more accurately the true synchrony in the response, scalp current
density (SCD) was calculated, using a MATLAB script which implements a spherical spline operator
Supplementary Figure 5-4: Independent component analysis to remove contamination of CI artefact
on the cortical response.
A. Independent components representing the CI artifact, eye blinks, 60Hz electrical noise and the
cortical evoked response are shown. The scalp morphology of each component is shown by head
plots on the left. The voltage potential of the components in each trial and averaged across trials is
shown to the right of each head. B. 64-channel cortical evoked response is shown on the left with
corresponding amplitude (top) and phase-locking value (bottom) analyses of synchronization for
frequencies from 4 to 60Hz as a function of latency. The CI artefact dominates cortical activity in B.
as shown by the large red spot, rendering further cortical analyses impossible. C. Removal of CI
artifact contamination from the response using independent component analyses allows analyses of
cortical activity to be performed. The peaks of the cortical response in the left panel are clear and
dynamics in synchrony can be seen (red = increased synchrony, blue = decreased synchrony, green =
baseline)

147
algorithm (Perrin, Pernier et al. 1989). This is a reference-free measure of potential distribution over
the scalp, which reduces the overlap of current recorded from surface sensor electrodes. This method
has previously been used for similar analyses (Doesburg, Kitajo et al. 2005; Doesburg, Emberson et
al. 2008; Doesburg, Roggeveen et al. 2008). The scalp-distribution information that SCD yields is
independent of the location of the reference electrode used during EEG recordings, represents more
concentric current activity, and emphasizes dipoles with shallower generators (Pernier et al., 1988). In
addition, long-range neural synchrony was analyzed for 19 pairs of electrodes which were distributed
uniformly across the scalp (i.e., Fp1, Fp2, F7, F3, Fz, F4, F8, T7, C3, Cz, C4, T8, P7, P3, Pz, P4, P8,
O1, O2) and separated by ≥ 4cm, which further reduces the effects of volume conduction.

148
6. Chapter Six – General discussion
The aim of restoring auditory sensation through cochlear implants (CIs) is to drive
development of the auditory system, promote acquisition of receptive and expressive spoken language,
encourage integration of children who are deaf into mainstream educational, occupational and social
settings, and increase quality of life and emotional well-being. To achieve these goals, CIs have been
provided to children in early development to limit deafness-induced reorganization of the auditory
brain. They have also been increasingly provided in both ears with the intent to establish binaural
hearing and reduce the risks of auditory deprivation in the un-implanted ear. This is particularly
important for children, because they typically learn and play/interact in groups where they need to
attend to several sources, and are often in environments where noise, reverberation and distance pose
particular challenges for hearing and listening. Unfortunately, in the earlier days of cochlear
implantation, these devices were provided to children in only one ear. While this promoted
development of auditory pathways from the hearing ear, unilateral implant stimulation left the opposite
and deprived pathways unprotected from effects of deafness. At the time, the reasons for providing
children with only one implant (outlined in section 2.11) were all justified and reasonable.
However, the work from Bess, Tharpe and colleagues uncovered many difficulties that these
children were experiencing with hearing from only one side. Unilateral hearing in children has been
associated with language delays, educational/learning difficulties and lower intelligent quotient scores
(Bess and Tharpe 1984; Bess and Tharpe 1986; Culbertson and Gilbert 1986; Bess, Dodd-Murphy et
al. 1998). They also have greater challenges forming social relationships and integrating in peer groups.
Children with unilateral hearing also tend to have more withdrawn behaviours and generally poorer
emotional health compared to their bilaterally hearing peers (Culbertson and Gilbert 1986). In addition
to these difficulties, the atypical auditory input delivered by a unilateral CI in children who are
bilaterally deaf might further alter development of the auditory system over the long-term (Eggermont
and Ponton 2003). In turn, this would impose further difficulties for children and adolescents to hear
in mainstream listening environments, may affect their ability to make friends and secure employment,
and might have negative consequences for their overall psychosocial development (Hyde and Punch
2011; Hyde, Punch et al. 2011; Punch and Hyde 2011; Punch and Hyde 2011).
In the present Thesis we aimed to address concerns regarding auditory cortical maturation over
the long-term in unilateral CI users (Chapters Three, Four and Five). We also wondered about the

149
plastic effects of long-term unilateral hearing on the opposite pathways (Chapters Four and Five). Has
long-term unilateral cochlear implantation promoted development/maturation in the stimulated
auditory pathways? If so, has this occurred at the expense of pathways from the opposite side? The
auditory pathways ascending from both ears are not completely independent from each other (Gordon
et al. In Preparation; Jiwani et al., 2010 Abstract) and recent findings suggested that a sensitive period
of 1.5 years exists for binaural input in the auditory system (Gordon, Wong et al. 2013). Keeping this
in mind, we also wanted to explore whether hearing could be established to the opposite deprived
pathways with a second CI after this sensitive period had been missed and the auditory cortex reached
maturation with unilateral CI stimulation. To answer these general questions, in the present Thesis,
we asked 5 specific questions:
1) Does the cortical evoked potential response follows a normal trajectory with time-in-sound
over long-term unilateral CI use when the duration of bilateral auditory deprivation in
childhood is limited?
2) How does missing an early sensitive period for bilateral input (i.e., brainstem) and driving
cortical maturation from one ear compromise the normal development of the auditory cortex
in adolescents who have used a unilateral CI to hear for most of their lives?
3) What are the effects of long-term unilateral deprivation on pathways from the opposite ear?
4) How is the cortical network underlying auditory processing organized in adolescents who
were congenitally deaf in both ears and used a unilateral CI to hear for most of their lives?
5) Has long-term unilateral deprivation caused segregation of the unstimulated pathways from
the auditory network?
Our specific hypotheses for each question were: 1) auditory development in children using
unilateral CIs will follow a normal trajectory with long-term auditory input when the duration of
bilateral deafness in childhood is limited; 2) areas of the brain which normally respond to sound will
be activated by auditory input in children who grow up using one CI; by contrast, 3) activity from the
deprived ear will abnormally recruit more diffuse cortical activity in the brain; 4) long durations of
unilateral CI use will promote co-ordinated cortical activity (neural networks) between brain areas that
are normally activated by sound, but 5) the deprived pathways will be segregated from this network.

150
6.1 Normal maturation of the auditory system requires hearing in both ears to
be normal
As described in Chapter Two, normal hearing requires that all elements of the auditory
pathways have intact structure and function from early in life. Essential developmental steps in the
auditory system occur in utero (Pujol, Lavigne-rebillard et al. 1991; Graven and Browne 2008) and
during the first months to years after birth (Pujol and Hilding 1973; Pujol, Lavigne-rebillard et al. 1991;
Moore and Guan 2001; Eggermont and Moore 2012). However, activity-dependent maturational
changes in the auditory system continue well into adolescence (Albrecht, Suchodoletz et al. 2000;
Wunderlich and Cone-Wesson 2006; Wunderlich, Cone-Wesson et al. 2006). Importantly, distinct
developmental time-courses exist for different generators in the auditory system (Albrecht,
Suchodoletz et al. 2000). While maturation of the auditory brainstem pathways is mostly complete by
2 years of age, cortical maturation continues until approximately 20 years. Deep cortical layers and
association auditory regions mature first, but activity in superficial layers of the auditory cortex only
start to become adult-like after 9 years of age (Albrecht, Suchodoletz et al. 2000; Ponton, Eggermont
et al. 2000; Moore and Guan 2001; Ponton, Eggermont et al. 2002). Investigators have suggested that
while deeper cortical layers may mature in the absence of auditory input, maturation of superficial
layers require sound stimulation during critical periods in development (Eggermont and Ponton 2003).
Normal patterns of maturation and refinement in the auditory cortex are underlied by
development of excitatory and inhibitory synaptic transmissions, which are mediated by activity-
dependent mechanisms. Excitatory receptor cells promotes maturation of synaptic strength and
precise synaptic connections, while inbitory glycinergic transmissions allow specific synaptic
projections to become refined in many neuronal systems (Sanes and Takács 1993; Kotak and Sanes
1996). These mechanisms subserve the development of dentritic arbors (Cline 2001), axonal
myelination and support synaptogenesis (Huttenlocher and Dabholkar 1997; Cline 2001) – all of which
promote plastic changes in the brain during sensitive periods in development to fine tune the auditory
system, allow life-long learning and adaptation to the environment (Rauschecker 1999; Abbott and
Nelson 2000; Song, Miller et al. 2000; Eggermont 2008).
We tracked the development of the cortical response over time in normal hearing children and
adolescents in Chapter Three. As shown in Figure 3.2, the cortical waveform recorded from the
immature auditory system of children who were younger than 10 years of age was dominated by a large

151
positive amplitude peak. As auditory cortical pathways matured into adolescence, this response
developed into an adult-polyphasic waveform with peaks P1, N1 and P2, characteristic of a mature
auditory cortex. These findings were consistent with reports from other groups showing a similar
trajectory of auditory development in normal hearing individuals from childhood to adulthood
(Ponton, Don et al. 1996; Ponton, Eggermont et al. 2000; Ponton and Eggermont 2001; Ponton,
Eggermont et al. 2002; Wunderlich and Cone-Wesson 2006; Wunderlich, Cone-Wesson et al. 2006).
Presence of these cortical peaks reveals much about the development of underlying cortical
structure and function. The auditory cortex is known to receive excitatory afferent projections from
both the thalamus into deep and middle layers and input from other auditory cortical regions into
superficial layers II (Kral and Eggermont 2007). Cortical evoked response peaks recorded from
cephalic electrodes on the head reflect changes in extra-cellular potentials (i.e., the voltage charge
outside of the pyramidal cell membrane) in superficial cortical layers of the brain. Positive peaks
indicate that the extra-cellular voltage in superficial layers II is more positive. This occurs when
pyramidal cells in deep or middle cortical layers are activated by excitatory inputs. This causes the
dendritic membrane to depolarize (i.e., intracellular voltage becomes less negative than its resting
potential as an influx of Na+ flows into the dendrite via ion channels in the post-synaptic membrane)
and the extracellular current in these layers to become more negative in order to balance the current
change across the membrane. However, to balance the intracellular potential change from the
depolarized membrane in middle cortical layers, a return current flow at the other end of the dendrite
in superficial layers causes the charge in this intracellular space to become more negative and
subsequently the extracellular charge close to the scalp, more positive. This is recorded by scalp-
surface electrodes as a positive deflection (i.e., P1, P2, P(ci)) in the cortical response (Eggermont 2007;
Eggermont 2008). On the other hand, negative deflections such as N1 and presumably N(ci), reflect the
negative extra-cellular post-synaptic potential in superficial layers which results from depolarization of
dendritic cells when they are activated by excitatory cortico-cortical input (Eggermont 2007;
Eggermont 2008).
Late emergence of peak N1 suggests that axon neurofilaments in superficial cortical layers only
mature after ~9 years of age, at which time their conduction velocity and ability to synchronize
increases (Moore and Guan 2001). Of interest, development of this response coincides with
improvement in many complex and sophisticated auditory processing skills during adolescence such
as understanding degraded and masked speech and perceiving speech in the presence of noise

152
(Eggermont and Ponton 2003). Investigators have suggested that N1 may “reflect the conscious
detection of any discrete change in any subjective dimension of the auditory environment” (p. 281,
Hyde 1997) and may in addition reflect attention switching (Näätänen and Picton, 1987). Normal
maturation of this response might similarly represent hemispheric specialization in the cortex. Indeed,
a relative specialization of the left auditory cortex for processing rapidly changing sounds such as
speech, and of the right auditory regions for processing slower input such as tones and melodies exists
in the brain (Penhune, Zatorre et al. 1996; Zatorre and Belin 2001; Zatorre, Belin et al. 2002;
Schönwiesner, Rübsamen et al. 2005; Jamison, Watkins et al. 2006; Schönwiesner, Krumbholz et al.
2007). This cortical specialization is underlied by structural differences between the two hemispheres
(Penhune, Zatorre et al. 1996; Zatorre, Belin et al. 2002), which develop with auditory experience
(Penhune, Zatorre et al. 1996) and normal maturational changes that occur during adolescence (Moore
and Linthicum 2007; Kadis, Pang et al. 2011).
In Chapters Four and Five we investigated the activity underlying the mature cortical response
to map the areas of the brain that respond to sound and the cortical hearing network that is evoked by
a 500Hz tone-burst stimulus in normal hearing adolescents. This specific frequency was chosen to
mimic the stimulus that was delivered to users of CIs, as their responses were compared to that of the
normal hearing group. Localization of auditory activity measured in Chapter Four and neural
synchrony analyses in Chapter Five suggests that specialization of the cortical hemispheres emerged
with maturation of the cortical response in our cohort of normal hearing adolescents. As shown in
Figures 4.1, 4.5A, 4.5B and 4.5E dipole activity in response to a 500Hz tone was stronger in the right
than left auditory cortex for stimulation of both ears. This lateralization was particularly stronger for
stimulation of the left ear. Analyses of aural preference in Figure 4.3 confirm these results, further
indicating that the right auditory cortex preferentially responds to left ear stimulation. Of interest,
neural synchrony analyses in Figure 5.1 of Chapter Five also revealed differences in neural oscillations
when the right and left ears were stimulated separately. Increased coupling of theta and gamma-band
activity in the right hemisphere evoked by stimulation of the left ear compared to the right indicate
increased processing of cortical input and active feature binding analyses across long-range distance in
the brain between frontal, temporal and parietal regions. This suggests that in response to a tone-
burst, network connectivity in the cortex from the left ear is increased relative to the right ear. These
results are consistent with findings from other groups who similarly showed stronger auditory activity
in the right brain, with a greater degree of lateralization for left than right ear stimulation in response

153
to tones and white noise in normal hearing adults (Hine and Debener 2007). The present results
indicate that both the left and right auditory cortices are involved in processing spectral simuli, but
that the right hemisphere seems to serve a particularly important role for pitch processing in normal
hearing individuals.
This asymmetry likely emerged in maturation, as younger children with normal hearing who
had not yet developed mature cortical responses showed a more symmetrical contralateral lateralization
for stimulation of each ear in response to the same tone-burst stimulus (Gordon, Wong et al. 2013).
Lateralization of auditory activity in the brain is in line with the anatomy of ascending projections in
the auditory pathways. In the immature brain, the contralateral pathways have the more direct route
to the cortex because they contain fewer synapses and a greater number of afferent fibers compared
to the ipsilateral pathways (Glendenning, Brusno‐Bechtold et al. 1981; Nordeen, Killackey et al. 1983;
Ponton, Vasama et al. 2001). As the brain matures, structural asymmetries in white matter volume and
cellular organization of the left and right auditory cortex develop. The volume of white matter in the
left Heschl’s Gyrus and left Posterior Temporal lobe is larger compared to the right. Grey matter
volume between both hemispheres is similar, however. Pyramidal cells in layer III of the primary
auditory cortex are also larger on the left side and occur with wider and more widely spaced cell
columns. These cell columns receive thalamo-cortical input from a larger number of afferent fibers
that are heavily myelinated and have heavily branched axons relative to the complementary structure
in the right hemisphere (Penhune, Zatorre et al. 1996; Zatorre, Belin et al. 2002). These
interhemispheric structural differences refine connections in the brain and allow them to serve a
supportive role for specialized auditory processing of complex information contained in speech and
music. The larger number of heavily myelinated cells in the left hemisphere with thicker axonal
branching facilitates transmission and encoding of rapidly changing temporal events, such as speech
sounds (Penhune, Zatorre et al. 1996; Zatorre, Belin et al. 2002). The relatively smaller and slower cell
structure in the right cortex on the other hand seems to be more sensitive to tonal patterns and is
optimal for processing spectral content, such as music and melodies. Unfortunately, only a tone-burst
stimulus was used in ours and many other studies (Zatorre and Belin 2001; Langers, van Dijk et al.
2005; Schönwiesner, Rübsamen et al. 2005; Hine and Debener 2007; Schönwiesner, Krumbholz et al.
2007) investigating differences in cortical lateralization in the normal hearing adolescent/adult brain.
We suggest that future studies should investigate whether using 1) a fast broadband click stimulus and

154
2) a speech stimulus, might affect the direction of hemispheric lateralization after cortical maturation
is reached.
Specialization of the cortex allows for a division of labor between the two hemispheres which
becomes optimized with experience (Gotts, Jo et al. 2013). This allows the brain to reduce
redundancies, avoid competition, increase neural capacity and process input more efficiently and more
effectively (Rogers 2000; Toga and Thompson 2003; Dharmaretnam and Rogers 2005; Vallortigara
2006; MacNeilage, Rogers et al. 2009; Gotts, Jo et al. 2013). The degree of lateralization of brain
structure and function has been associated with improved cognitive abilities and functional task
performance (Abrams, Nicol et al. 2006; Gotts, Jo et al. 2013). Of interest, maturation of the pre-
frontal cortex, which is involved with complex cognitive and perceptual functions related to learning,
attention processing, decision making, problem solving, mentalizing, memory consolidation, etc… also
occurs around this time (Anderson, Anderson et al. 2001; Johnson 2001; Moriguchi, Ohnishi et al.
2007; Johnson 2011; Blakemore 2012). This likely contributes to the improvements in cognitive
behaviours and perceptual functions which are underpinned by brain maturation. Mechanisms of
plasticity decrease with increase age (Lohmann and Kessels 2014). Investigators have suggested that
once maturation is reached, the threshold for plasticity decreases, perhaps to protect the mature brain
from large-scale reorganization and allow fine-tuning of cortical connections (Lenroot and Giedd 2006;
Blakemore 2012; Lohmann and Kessels 2014). Adolescence thus highlights an important time in
development, which is characterized by much refinement and specialization in the cortex and could
mark the closing of an important period in brain plasticity.
6.2 Unilateral implant stimulation promotes cortical maturation but leaves the
brain with abnormal organization
Adolescents who were deaf in both ears from early life and used a CI to hear for over a decade
developed a similar cortical response to that of normal peers with polyphasic peaks P1, N1, P2 as shown
in Figures 3.2 and 3.3, characteristic of a mature auditory cortex. The cortical response has been
shown to decrease rapidly at initial stages of development in children who received a CI within limited
periods of bilateral deafness and reached age-appropriate peak latency and amplitude over 3 to 6
months of implant use (Sharma, Dorman et al. 2002). After this initial period, these responses changed

155
at a rate that is similar to normal (Eggermont, Ponton et al. 1997; Eggermont and Ponton 2003).
Eggermont and Ponton indicated that the latency of the cortical response was delayed in CI users
relative to age-matched normal hearing responses by an amount of time that was equivalent to the
period of deafness (Ponton and Eggermont 2001). This suggests that the duration of exposure to
sound (referred to as time-in-sound and calculated as chronological age – duration of bilateral deafness)
rather than chronological age, influences the rate of cortical maturation in children. These investigators
raised concerns that bilateral deprivation prior to cochlear implantation might alter the maturation of
thalamo-cortical and cortico-cortical loops in superficial layers, which develop around 9 to 12 years of
age and underlie the emergence of peak N1 in the cortical response. These pathways mediate the
transfer of primary auditory and multi-sensory input from the thalamus to various regions of the
ipsilateral and contralateral auditory cortices (Winer, Diehl et al. 2001; Winer, Miller et al. 2005; Razak,
Zumsteg et al. 2009), and the transmission of information from the auditory cortex to primary and
secondary sensory areas in both hemispheres (Read, Winer et al. 2002; Lee and Winer 2005; Klinge,
Eippert et al. 2010). Altered development of these pathways would hinder auditory processing and
may result in further struggles during listening for CI users.
The data from Chapter Three indicates that the developmental trajectory of the electrically
evoked cortical waveform (Figures 3.2 and 3.3) is similar between CI users and normal hearing
individual. Appropriate development of mature normal-like peaks P1, N1 and P2 in adolescents who
had over 10 years of time-in-sound experience suggests that long-term CI stimulation establishes
appropriate 1) relay of auditory input from the ear to the cortex, via the thalamus; 2) communication
between the two cortical hemispheres; and/or 3) connectivity between different sensory areas. The
data in Figure 3.4 suggests that this development is experience-dependent, with the cortical response
becoming more normal-like with time-in-sound experience. However, as shown in Figures 3.3 and
3.5, the waveform has at least one abnormality; the amplitude of peak P2 in CI users is larger than in
normal hearing peers. These results are consistent with previous studies indicating larger peak P2
amplitudes in a group of adults who were fitted with unilateral hearing aids (Bertoli, Probst et al. 2011)
and others who had a bilateral mild to moderate sensorineural hearing loss (Campbell and Sharma
2013), suggesting that hearing loss alters activity in the brain. Enhancements in the P2 amplitude,
particularly in the left hemisphere, have also been reported in normal hearing adults following focused
auditory training tasks (Tremblay and Kraus 2002; Tremblay 2007; Tremblay, Shahin et al. 2009) and
likely reflect increased listening effort and/or attention during listening. We suggest that the larger-

156
than-normal amplitude of peak P2 from our cohort of CI users might represent re-allocation of cortical
resources or recruitment of additional cortical areas to process sounds to support hearing from only
one side to the atypical electrical input delivered by the CI and/or compensatory cortical plasticity
resulting from early onset deafness. The importance of this recent finding is that it suggests that
deviations from normal cortical processing remain in these young people despite long-term unilateral
implant use.
Cortical localization measures and neural synchrony analyses in Chapters Four and Five further
indicate that the activity underlying the cortical response is abnormally organized in experienced CI
users compared to normal hearing peers. As shown in Figures 4.4 and 4.5, stimulation of the right
CI after over a decade of experience indicated strong lateralization towards the contralateral left
auditory cortex. This is opposite to the right hemisphere bias that was observed from adolescents with
normal hearing, shown in Figures 4.2 and 4.5. Consistent with these findings, neural synchronization
analyses indicating greater-than-normal coordinated synchronization in the theta, beta and gamma
bands from areas of the left temporo-parietal regions to the frontal cortex in Figure 5.2, further
suggests altered organization of the auditory network compared to normal hearing peers. The strong
asymmetric bias observed in the left cortex for CI listening could indicate that cortical specialization
to non-speech stimuli has either not yet emerged and that contralateral afferent input still dominates
auditory activity in these adolescents, as it does in the immature auditory cortex of younger implanted
peers (Gordon, Wong et al. 2013). Alternatively, it could also mean that the brain has re-allocated
cortical resources to support processing in the hemisphere that is specialized for speech. The latter
suggestion is supported by excellent speech perception scores on the PBK open-set speech test
(Figure 4.7A) and the significant positive correlation of these scores with lateralization and greater
dipole strength in the left hemisphere compared to the right (Figure 4.7B).
Differences in hemispheric asymmetries between CI users and normal hearing listeners might
be explained by differences in stimulus feature properties of the acoustic signal (in normal hearing
peers) versus electric signal delivered by the implant device to the auditory system (Sandmann, Eichele
et al. 2009). CIs were designed to establish and promote speech perception. Most digital signal
processing strategies encode the coarse features of auditory signals only (Zeng, Rebscher et al. 2008).
Current devices extract the temporal envelope of sound at a given rate of stimulation and band-pass
filter its frequency contents into a set number of channels. The number of channels with the largest
amplitude envelope are then selected and allowed to stimulate the specific electrodes corresponding

157
to the allocated frequency of the given filter (Rubinstein 2004; Zeng, Rebscher et al. 2008). In current
processing strategies this is usually limited to a maximum of ~8 channels (known as maxima),
regardless of the number of working electrodes (Henry and Turner 2003; Vaerenberg, Smits et al.
2014). The advantage of these envelope-based processing strategies is that they support high levels of
speech recognition in quiet. Unfortunately, they fare poorly in noise and with interfering speech
signals. In addition, acquisition and perception of tonal languages have proven to be particularly
challenging for CI listeners who use current digital processing strategies (Lee, Van Hasselt et al. 2002;
Wei, Cao et al. 2004; Xu and Zhou 2012). The same is true of music appreciation with an implant
(Drennan and Rubinstein 2008; Hopyan, Gordon et al. 2011; Hopyan, Peretz et al. 2012). It is
particularly difficult for CIs users to perceive pitch-related information (Rubinstein 2004; Drennan and
Rubinstein 2008), and perception of timbre and melody are impoverished (Hopyan, Gordon et al.
2011; Hopyan, Peretz et al. 2012). This is not surprising because electric stimulation with an implant
does not deliver fine structure and distorts detailed spectral information (i.e., CIs do not explicitly code
spectral information) that is normally used by the normal auditory system to recognize patterns of
tones (Xu and Zhou 2012) and extract complexities in music (Drennan and Rubinstein 2008).
Differences in hemispheric lateralization with right ear stimulation observed from normal hearing
adolescents and experienced CI users (Figure 4.5) might thus reflect differences in auditory experience
between the two groups. It is possible that absence of spectral and fine-structure auditory details with
CI stimulation over the long-term compromised mechanisms of cortical specialization in the right
hemisphere of the mature brain (which is known to be biased for spectral processing) and resulted in
an increased response bias in the left cortex. We are now investigating whether the same asymmetry
in the left cortex exists in adolescents who received an implant in their left ear as their first CI. Future
studies should work more closely with engineers and manufacturers of CIs to investigate whether
increasing the number of maxima and/or using higher stimulation rates to better represent fine-
structure information might normalize lateralization/specialization of auditory input with CI listening
while still preserving the gain in speech perception.
In addition to the reduced auditory cues delivered by the CI compared to normal hearing
listening, asymmetric strengthening of the contralateral pathways from the right-experienced ear could
be explained by increased excitability of postsynaptic currents in the stimulated ear and reduced
inhibition from the opposite ear, resulting from long-term unilateral deprivation (Sanes and Takács
1993; Vale, Juíz et al. 2004; Kotak, Fujisawa et al. 2005; Takesian, Kotak et al. 2009; Sanes and Kotak

158
2011). Unilateral auditory deprivation disrupts the delicate balance of excitatory and inhibitory inputs
in the afferent auditory system (Kitzes 1984; Moore and Kowalchuk 1988; Moore 1990; Kil,
Hkageyama et al. 1995; Kitzes, Kageyama et al. 1995; Mossop, Wilson et al. 2000; Nordeen, Killackey
et al. 2004; Popescu and Polley 2010; Kral, Heid et al. 2013; Kral, Hubka et al. 2013) that would have
normally occurred with bilateral hearing (Grothe, Pecka et al. 2010). A loss of inhibitory activity from
the contralateral ear (i.e., the deprived ear) would allow input from the stimulated side to be projected
to the contralateral cortex without suppression. This would in turn allow the left auditory cortex to
dominate auditory input in adolescents with pre- or peri-lingual deafness who were implanted in the
right ear as babies. Neural synchrony analyses in Figure 5.2 showing increased coordinated theta and
gamma band activity in pathways from the experienced right ear to the left temporo-parietal areas and
frontal regions in both hemispheres compared to normal might reflect such increased excitability from
this ear to the cortex.
Enhanced activity from this ear could also mark plastic changes in the brain occurring as a
compensatory mechanism for the abnormal electrical input provided by the implant device and/or
stimulation of the auditory pathways from only one side. The larger-than-normal P2 amplitude
observed in Figures 3.3 and 3.5 and additional source activation in the frontal and precuneus regions
(Figure 4.6) with increased long-range neural synchrony in the gamma and theta frequencies of these
cortical regions (Figure 5.2C and 5.2D) coupled with a reduction in local alpha activity (Figure 5.2B)
compared to normal all indicate recruitment of additional cortical processes to facilitate listening with
a unilateral CI. The observed pattern of activity suggests that adolescents with long-term implant
experience rely on brain networks linked to attention (Picton and Hillyard 1974; Hocherman, Benson
et al. 1976; Rif, Hari et al. 1991; García-Larrea, Lukaszewicz et al. 1992; Posner and Dehaene 1994;
Pugh, Shaywitz et al. 1996; Grady, Van Meter et al. 1997; Benedict, Lockwood et al. 1998; Fujiwara,
Nagamine et al. 1998; Jäncke, Mirzazade et al. 1999; Fan, McCandliss et al. 2002; Fan, McCandliss et
al. 2005; Okamoto, Stracke et al. 2007; Tremblay, Shahin et al. 2009) and multi-sensory integration
(Hari 1990; García-Larrea, Lukaszewicz et al. 1992; Levänen, Jousmäki et al. 1998; Webster and Colrain
2000; Moller and Rollins 2002; Crowley and Colrain 2004; Johnson and Zatorre 2005) for CI listening
of a non-speech auditory input.
Individuals using CIs have already been shown to use multi-perceptual (Jeong Lee, Kang et al.
2005; Lee, Giraud et al. 2007; Pisoni, Conway et al. 2010; Pisoni, Kronenberger et al. 2011; Geers,
Pisoni et al. 2012; Kronenberger, Colson et al. 2014) and multi-sensory (Nishimura, Hashikawa et al.

159
1999; Giraud, Price et al. 2001; Giraud, Price et al. 2001; Doucet, Bergeron et al. 2006; Giraud and Lee
2007; Hopyan-Misakyan, Gordon et al. 2009; Sandmann 2012; Sandmann, Dillier et al. 2012)
information more effectively than their peers with normal hearing to facilitate complex auditory
processing (Hopyan-Misakyan et al., 2009; Brenner & Geers, 2011). This is not surprising since
unilateral CI hearing only provide a crude representation of acoustic sounds and eliminates access to
binaural hearing. This in turn, makes listening and oral communication in general more difficult, more
tiring, more stressful, more unpleasant, and more effortful particularly in challenging listening
environments (Gordon, Jiwani et al. 2011; Gordon, Jiwani et al. 2013). Given that CIs do not restore
normal hearing and that bilateral and unilateral deafness drives changes in the normal organization of
the auditory system, it is reasonable that children who are deaf and have heard with a CI for most of
their lives would capitalize on intact senses and perceptual and cognitive strategies to supplement their
hearing.
Of note, imaging studies have shown increased recruitment of attentional cortical networks in
children who are deaf compared to normal hearing peers (Giraud, Price et al. 2001; Giraud, Price et al.
2001; Jeong Lee, Kang et al. 2005; Lee, Giraud et al. 2007). Consistent with our findings in Chapters
Four and Five, investigators have indicated that children who used their CIs successfully activated
cortical regions in the left posterior dorsolateral prefrontal gyrus which is involved with attention,
working memory, reasoning, and executive function (Jeong Lee, Kang et al. 2005; Lee, Giraud et al.
2007), and the right precuneous region which is involved with self-reflection, episodic memory,
visuospatial processing, and imageability (i.e., visual imagery) (Giraud, Price et al. 2001; Jeong Lee,
Kang et al. 2005). CI users might use these general cognitive strategies to form a visual representation
of the sound to aid sound recognition and comprehension (Giraud, Price et al. 2001). In addition,
activation of these cortical areas during listening might facilitate perceptual learning and speech
learning during rehabilitation (Jeong Lee, Kang et al. 2005; Alain, Snyder et al. 2007). In support,
Pisoni and colleagues suggested that successful CI users recruit increased attention demands and have
better memory capacity compared those children who are less successful with their implants (Pisoni,
Conway et al. 2010; Pisoni, Kronenberger et al. 2011; Kronenberger, Colson et al. 2014).
CI users also supplement their hearing with vision. Even normal hearing individuals use
multisensory input, notably vision, to support listening (Ross, Saint-Amour et al. 2007). However,
greater-than-normal cortical connectivity of theta and gamma oscillations in visual cortical areas in
experienced CI users shown in Figure 5.3 compared to normal hearing peers suggests a better ability

160
of these adolescents to utilize visual cues to compensate for the imperfect implant signal. In a recent
study (data shown in Figure 2.10) we found that adolescents who received a unilateral CI within
limited durations of bilateral deafness and had ~8 years of hearing experience with this device depend
on visual cues more heavily than their peers with normal hearing to identify emotions carried in speech
(Hopyan-Misakyan, Gordon et al. 2009; Gordon, Jiwani et al. 2013). The increased reliance on vision
to process auditory input is consistent with functional neuroimaging studies, which find abnormally
increased activation of the visual cortex in users of CIs when listening to meaningful speech (Giraud,
Price et al. 2001; Lee, Giraud et al. 2007; Lee, Truy et al. 2007). This may reflect maladaptive
(Nishimura, Hashikawa et al. 1999; Doucet, Bergeron et al. 2006; Lee, Truy et al. 2007; Sandmann
2012; Sandmann, Dillier et al. 2012) or compensatory (Kral and Sharma 2012; Campbell and Sharma
2013; Sharma, Campbell et al. 2014) plasticity of the auditory and visual systems without normal
hearing
Increased dependence on attention and other cognitive mechanisms with greater reliance on
vision might mean better functional hearing, but unfortunately, it could also mean increased effort and
perhaps reduced efficiency for listening. We suggest that the reduced gamma-band activity in
frequencies over 35Hz observed in Figure 5.3 from the experienced-right side of CI users and not
normal hearing peers reflects a general slowing of cortical processing that is likely attributed to more
effortful listening. As described earlier in this section, adolescents with long-term unilateral CI
experience activated regions of the frontal and precuneus cortices with greater than normal dipole
activity, suggesting that they require greater-than-normal cognitive resources during listening.
Unfortunately, this activity has been associated with increased listening effort (Pichora-Fuller 2003;
Collette, Hogge et al. 2006; Pichora-Fuller and Singh 2006; Hughes and Galvin 2013; Pals, Sarampalis
et al. 2013). Findings of increased reaction times in Figure 2.10B and changes in pupil diameter
reflective of increased mental resources (Steel, Papsin et al. Submitted) observed from CI listeners
compared to normal hearing peers provide evidence to support this. It is not surprising that listening
to a distorted auditory signal delivered by a single CI would be more effortful for CI users. Of concern,
however, increased expenditure of mental processing for children and adolescents who use CIs to hear
may negatively affect learning and perhaps even social interactions, as well as cause frustration and
exhaustion when listening. On the other hand, it is also possible that this might lend a supportive role
for CI listening. At the moment, it is unclear whether increased reliance on attention and vision for
listening with perhaps increased effort helps or hinders auditory processing. However, if the cognitive

161
load imposed on CI listening could be reduced, perhaps adolescents using CIs would be able to
respond to complex speech more quickly, fare better in challenging listening environments and better
keep up with their normal hearing peers in school, at work and in social situations.
In sum, the present data shows that while unilateral cochlear implantation promotes the
development of mature and normal-like activity in the auditory pathways over the long-term,
functional abnormalities persist. Long-term unilateral CI stimulation seems to drive a compensatory
plasticity of the auditory brain to re-allocate cortical resources across a larger functional network to
support hearing with the single implant. Auditory evoked activity in the contralateral left cortex (that
is normally specialized for speech) and occurring with increased oscillatory synchronization in cortical
areas linked to attention, general cognitive/perceptual processing and vision during listening could
reflect compensation for: 1) deleterious or irreversible changes to neural reorganization that occurred
during the period of auditory deprivation in early life, 2) abnormal representation of sound through
electrical pulses stimulation of the auditory system, and/or 3) absence of auditory input to the deprived
pathways from the opposite un-implanted ear.
6.3 Long-term unilateral deprivation drives abnormally altered and
disorganized activity in the unstimulated pathways
Many adolescents from this study who were implanted in their right ear as babies and have
already had over a decade of unilateral hearing experience, recently asked for a CI in their opposite
and deprived left ear in the hopes of deriving benefits of bilateral implantation. This provided us with
a unique opportunity to stimulate the deaf ear for the first time and study the effects of long-term
unilateral implant stimulation on the deprived pathways. We hypothesized that driving maturation of
the auditory cortex with over a decade of unilateral CI stimulation in the right ear would lead to
permanent abnormalities and reorganization of cortical activity in pathways from the deprived left ear.
Cortical responses were recorded from this second implanted left ear within a week of initial activation
on that side. Responses were measured at this early stage of activation to study cortical plasticity
associated with long-term unilateral deprivation before auditory input could be established in this ear
with CI stimulation.

162
As shown in Chapters Four and Five, despite long durations of inactivity to the auditory
pathways prior to implantation, the deprived ear was able to respond to sounds delivered by a CI, as
evidenced by the ability to record a responses from the brainstem (Figure 4.1) and cortex (Figure
4.4G and 5.2E) at initial device activation. However, the latency of wave eV of the brainstem response
was abnormally delayed compared to the experienced side. The lack of significant auditory input in
early life may have compromised or arrested the development of activity-dependent neural tracts in
these brainstem pathways. In normal hearing individuals, such activity-dependent development in
early life has been associated with the formation of more efficient synaptic transmission at gap
junctions, increased neural synchronization of spiral ganglion cells in response to auditory stimuli and
faster neural conduction in the auditory pathways from increased myelination of axonal tracts (Moore
and Goldberg 1963; Huttenlocher 1967; Huttenlocher 1970; Huttenlocher and Dabholkar 1997;
Moore and Guan 2001; Moore 2002; Moore and Linthicum 2007). It has also been shown that
spontaneous activity in the developing auditory system is required for neuronal survival, formation of
appropriate connectivity in the auditory pathways, and fine-tuning of tonotopic maps (Tritsch, Yi et
al. 2007). All of these mechanisms are reduced and/or hindered by bilateral deafness in early life prior
to cochlear implantation and may further be exacerbated by unilateral deprivation thereafter. In
support, the cortical response recorded from the naïve side also had an atypical waveform morphology
compared to the opposite experienced ear and both the right and left ears of age-matched normal
hearing peers. This indicated further disruptions/disorganization of activity in the unilaterally deprived
auditory pathways. The cortical waveform was characterized by a biphasic response of abnormally
large peak-to-peak amplitude (labelled N(ci)-P(ci)) and dominated by the negative peak, N(ci).
A similar cortical response has been observed from normal hearing pre-term infants younger
than 35 weeks gestational age (Weitzman and Graziani 1968; Wunderlich and Cone-Wesson 2006;
Wunderlich, Cone-Wesson et al. 2006) and was described as being characteristic of earlier stages of
cortical immaturity in the auditory pathways. This response type has also been recorded from
congenitally deaf children who were implanted after 7 years of age following long durations of bilateral
deafness (Sharma, Dorman et al. 2002; Jiwani, Tanaka et al. In Preparation) and in CI users who have
difficulty recognizing spoken language (Gordon, Tanaka et al. 2008). This abnormal response suggests
that a difference in cortical processing might exist in children who have difficulty recognizing spoken
language (Gordon, Tanaka et al. 2005; Gordon, Tanaka et al. 2008) compared to early implanted
children who have good auditory functions. Consistent with findings from congenitally deaf white

163
cats (Klinke, Kral et al. 1999; Kral, Hartmann et al. 2002), at initial CI activation, 49% of children with
bilateral deafness also produced a similar cortical response (Gordon, Tanaka et al. 2011). Of those,
79% were children with deafness related to bilallelic mutations on the GJB-2 gene (Gordon, Tanaka
et al. 2011). GJB-2 associated deafness results from a deficit in encoding the connexin 26 protein
which is necessary for cells to communicate with each other (Propst, Papsin et al. 2006). Such a deficit
would result in limited spontaneous activity in the auditory system and limited access to sound both in
utero and at birth prior to implantation (Propst, Papsin et al. 2006; Gordon, Tanaka et al. 2011). Thus,
the negative peaked cortical response type observed from the naïve pathways after initial stimulation
of this side might represent: 1) cortical immaturity of the auditory pathways, 2) restricted auditory
stimulation and disorganized activity in the pathways, 3) poor functional hearing outcomes.
We questioned whether this large negative peaked response might be homologous to the
negative peak N1 that emerges with maturation in normal hearing adults (Ponton, Eggermont et al.
2000) and adolescents with long-term CI experience (Jiwani, Papsin et al. 2013), since both of these
peaks have similar latencies. Of interest however, we noted several differences: 1) the amplitude of
peak N(ci) (Figure 4.4 and 5.2) is significantly larger than N1 (Figure 4.2 and 5.1); 2) dipoles underlying
peak N(ci) (Figure 4.4 and 4.6) are significantly larger than N1 (Figure 4.2); 3) they are also much more
widespread across the cortex (Figure 4.4 and 4.6), whereas this activity is more focused in the mature
response (Figure 4.2); 4) cortical lateralization is symmetrical between both hemispheres for peak N(ci)
(Figure 4.4 and 4.5). However, this activity shows asymmetric hemispheric lateralization for N1
(Figure 4.2 and 4.5), perhaps reflecting cortical specialization; 5) neural oscillations in the theta and
gamma frequencies are de-synchronized for the negative peaked waveform (Figure 5.2 and 5.3). On
the other hand this activity is increased for the mature cortical response (Figure 5.1 and 5.3); 6) the
latency and amplitude of peak N(ci) is not affected by changes in stimulus frequency, but both of these
peak properties change significantly with changes in frequency for N1, perhaps reflecting tonotopicity
in the auditory cortex (Gordon, Tanaka et al. 2008); 7) the negative peaked response N(ci) tends to be
associated with poor functional outcomes as shown in Figure 4.7 and in Gordon et al., 2005, 2008.
By contrast however, emergence of peak N1 tends to coincide with the development of complex
auditory perceptual skills such as hearing in noise and hearing degraded speech – these skills begin to
develop during adolescence (Ponton et al., 2000). In support, both normal hearing individuals and CI
users with long-term hearing experience who have developed a mature cortical response (with peak
N1) performed well on simple and complex listening tasks (average PBK speech score = 73.5 ± 3.2%

164
on the experienced CI side). However, these skills were significantly impaired in the newly implanted
ear, as shown in Figure 4.7 (simple open-set speech discrimination task had very poor performance
with only 26.8 ± 5.7% accuracy). Of interest, the amplitude of peak N(ci) has been correlated with poor
speech perception outcomes (Gordon, Tanaka et al. 2008). This negative-peaked response type might
thus represent an electrophysiological marker of immaturity in the auditory pathways and suggests that
different cortical trajectories and different cortical activity underlie peaks N(ci) and N1.
Nonetheless, it is possible that neural generators are still shared between the two responses.
As described in Section 5.1, a cortical peak with a negative deflection that is recorded from a midline-
cephalic location and referenced to the earlobe reflects activation in superficial layers of the cortex. N1
is believed to be generated by cortico-cortical input in these superficial layers and reflects maturation
of axon neurofilaments in layer II (Ponton, Eggermont et al. 2000; Moore and Guan 2001; Moore and
Linthicum 2007; Eggermont 2008). This developmental process only occurs around 9 to 12 years of
age (Ponton, Eggermont et al. 2000; Moore and Linthicum 2007) and is activity-dependent (Jiwani,
Papsin et al. 2013). In the same way, it is possible that the large peak N(ci) also reflects superficial
cortical activity. However, since this response type has been observed in pre-term normal hearing
babies (Weitzman and Graziani 1968; Wunderlich and Cone-Wesson 2006; Wunderlich, Cone-Wesson
et al. 2006) and at early device activation in implanted peers with little access to sound prior to
implantation due to GJB-2 deafness (Propst, Papsin et al. 2006; Gordon, Tanaka et al. 2011) who were
all younger than 9 years of age, N(ci) cannot reflect mature superficial layer axons. Rather, it is possible
that this response reflects immaturity of these axons in these cortical layers.
The large amplitude (Figures 4.4 and 5.2) and dipoles (Figure 4.4 and 4.6) underlying this
response may be a result of delayed or arrested mechanisms of synaptogenesis in the auditory cortex.
Indeed, early formation of synaptic contacts and age-dependent elimination of synapses have been
suggested to be an activity-dependent mechanism (Huttenlocher and Dabholkar 1997). Without
auditory input in the deprived pathways, it is possible that the areas that generate peak N(ci) have not
yet undergone synaptic pruning. In support, Ponton and colleagues similarly suggested that in the
normal hearing system, the fact that the amplitude of early developing peak P2 does not change with
maturation might indicate that its generators do not undergo synaptic pruning (Ponton, Eggermont et
al. 2000). If the same holds true for peak N(ci), the lack of cortical development in its generators would
be in contrast to the synaptic development that normally occurs in the maturing auditory cortex for
peak N1 (Huttenlocher and Dabholkar 1997), and rather more similar to the findings of delayed

165
synaptogenesis reported from cats with congenital bilateral deafness (Huttenlocher and Dabholkar
1997; Kral and O'Donoghue 2010; Kral and Sharma 2012).
Since the process of synapse elimination in the auditory cortex is normally complete by
adolescence (Huttenlocher and Dabholkar 1997), we are concerned that leaving the auditory pathways
unilaterally deprived beyond this important developmental period may lead to permanent
abnormalities. “In the visual cortex, unilateral deprivation of formed visual input during the period of
synapse elimination resulted in asymmetric synaptic input from the two eyes (Hubel et al., 1977;
Goodman and Shatz, 1993), and in permanent visual deficits (VonNoorden and Crawford, 1979; Assaf,
1982)” (p. 177, Huttenlocher and Dabholkar 1997). If long-term unilateral auditory deprivation drives
parallel changes in the auditory system, auditory development in the newly implanted side may be
challenging to promote. Of concern, unilateral auditory deprivation in the developing auditory system
has been associated with many subcortical and cortical structural and functional changes. Specifically,
long-term absence of hearing from one side alters the normal balance of excitatory-inhibitory activity
in the ascending auditory system (Vale, Juíz et al. 2004; Kotak, Fujisawa et al. 2005; Takesian, Kotak
et al. 2009) and results in suppressed input strength in the deprived pathways from brainstem (Moore
and Kowalchuk 1988; Kil, Hkageyama et al. 1995; Kitzes, Kageyama et al. 1995; Nordeen, Killackey
et al. 2004; Popescu and Polley 2010) to cortex (Popescu and Polley 2010; Kral, Heid et al. 2013; Kral,
Hubka et al. 2013). These abnormal changes persist even after hearing is restored to this side with a
CI (Gordon, Wong et al. 2013; Kral, Heid et al. 2013; Kral, Hubka et al. 2013). By contrast, this activity
becomes abnormally enhanced in pathways from the hearing side (Popescu and Polley 2010; Gordon,
Wong et al. 2013; Kral, Heid et al. 2013; Kral, Hubka et al. 2013).
Unfortunately, developmentally imbalanced auditory input between the two ears places the
deprived side at a disadvantage in competition for cortical resources compared to the hearing ear (Kral,
Heid et al. 2013) and leads to long-lasting and often irreversible perceptual deficits (Popescu and Polley
2010). Cortical consequences of unilateral deprivation was first shown in the visual system (Hubel and
Wiesel 1970; Blakemore and Van Sluyters 1974; Hubel, Wiesel et al. 1977; Cynader, Timney et al. 1980;
Le Vay, Wiesel et al. 1980; Cynader, Leporé et al. 1981; Cynader 1983; Blakemore 1988; Woodruff,
Hiscox et al. 1994; Holmes, Kraker et al. 2003; Williams, Northstone et al. 2003; Jeffrey, Wang et al.
2004) and later confirmed in the auditory system (Brookhouser, Worthington et al. 1991; Gordon,
Valero et al. 2008; Hine, Thornton et al. 2008; Gordon and Papsin 2009; Graham, Vickers et al. 2009;
Salloum, Valero et al. 2010; Chadha, Papsin et al. 2011; Gordon, Jiwani et al. 2011; Graham and Vickers

166
2011; Fitzgerald, Green et al. 2013; Gordon, Jiwani et al. 2013; Gordon, Wong et al. 2013; Hughes and
Galvin 2013; Illg, Giourgas et al. 2013; Kral, Heid et al. 2013; Kral, Hubka et al. 2013). In the visual
system, monocular deprivation resulted in permanently deficient visual acuity in the deprived eye,
impaired contrast sensitivity, reduced sensitivity to motion detection and deficient stereoscopic vision
(Hubel and Wiesel 1970; Hubel, Wiesel et al. 1977; Cynader, Timney et al. 1980; Le Vay, Wiesel et al.
1980; Cynader, Leporé et al. 1981; Cynader 1983; Blakemore 1988). These abnormalities persisted
even after years of treatment and re-training therapy of the poor eye. The same is true of the impaired
ear in children who have recurrent or long-lasting childhood conductive hearing losses (Popescu and
Polley 2010), asymmetric hearing losses (Bess and Tharpe 1984; Bess and Tharpe 1986; Tharpe 2008;
Hughes and Galvin 2013) and bilaterally deaf children with unilateral CIs (Gordon and Papsin 2009;
Salloum, Valero et al. 2010; Chadha, Papsin et al. 2011; Gordon, Jiwani et al. 2011; Gordon, Jiwani et
al. 2013; Gordon, Wong et al. 2013).
Perhaps the second implanted ear is left with a competitive disadvantage compared to the
experienced side for listening because activity in the deprived pathways has become slow, undefined
and disorganized. This would mean that these pathways may no longer be able to integrate and process
the incoming auditory input with clarity and accuracy. Representation of auditory input in the brain
might in turn become smeared or jittered, as it becomes in the visual cortex after long-term monocular
deprivation (Jeffrey, Wang et al. 2004). In support, the pattern of globally de-synchronized activity in
the theta and gamma frequency bands together with increased alpha oscillations shown in Figures 5.2
and 5.3 might reflect cortical slowing in the unstimulated pathways as a result of long-term unilateral
deafness (Monastra, Lubar et al. 1999; Monastra, Lubar et al. 2001; Uhlhaas and Singer 2006; Stoffers,
Bosboom et al. 2007; Englot, Yang et al. 2010; Uhlhaas and Singer 2010; Doesburg, Ribary et al. 2011;
Uhlhaas and Singer 2011; Doesburg, Moiseev et al. 2013). It might also reflect a general lack of
familiarity/meaninglessness with the novel stimulus or incongruency between the auditory input and
its representation in the brain (Rodriguez et al 1999, Doesburg 2008a). This would explain the
impaired speech perception performance observed from this side after months of hearing experience
with the implant (Figure 4.7). Poor functional hearing in the naïve ear might also be attributed to the
lack of asymmetry between the two hemispheres in the cortex, as shown in Figures 4.4 and 4.5.
Indeed, reduced cortical lateralization has been associated with poor auditory processing abilities,
reduced academic performance and impaired reading and writing skills (Abrams, Nicol et al. 2006).
Perhaps it is for this reason that early implanted adolescents who receive sequential CIs after years of

167
unilateral hearing are more likely to reject or inconsistently use their second implant (Fitzgerald, Green
et al. 2013). In line with our hypothesis, we suggest that the abnormal activity evoked by initial CI
stimulation of the deprived ear might reflect segregation of these pathways from the cortical hearing
network.

168
7. Chapter Seven – Current and future directions
7.1 Does bilateral cochlear implant experience promote auditory development
in pathways from the newly implanted side?
Having assessed the cortical effects of long-term unilateral stimulation/deprivation on the
auditory pathways in the adolescent brain, we are now asking about the longer term effects of single-
sided deafness in children after hearing is established in the deprived ear. To explore this, we have
been tracking cortical development in the deprived pathways in our cohort of bilaterally implanted
adolescents from chapters Four and Five as they gain more hearing experience with their second
implant. This will allow us to determine whether achieving maturation of the auditory cortex with
only one CI marks the closure of a sensitive period for restoring auditory function on the deprived
side. We have measured the cortical responses from this group of adolescents who have over a decade
of unilateral right implant experience after 1 month and 9 months of bilateral CI use. Cortical
waveform morphology from both the experienced and later implanted ears remained unchanged from
the first day of bilateral implant activation. As expected, on the experienced side, the cortical response
remained characterized by obligatory peaks P1-N1-P2-N2. Little changes in the response occurred in
the second implanted side over the first months of bilateral implant use. The response remained
dominated by the N(ci)-P(ci) complex, with little change in latency or amplitude to either peak despite
time after cochlear implantation. The lack of cortical development evoked by stimulation of the
second implanted side is in contrast to the rapid developmental change that occurs at early stages of
unilateral CI use in young children (Sharma, Dorman et al. 2002; Sharma and Dorman 2006) and
rather, more similar to the limited change reported in older children implanted after long durations of
bilateral deafness (Sharma, Dorman et al. 2002; Gordon, Tanaka et al. 2005; Gordon, Tanaka et al.
2008). Using beamformer and connectivity analyses, we will continue to explore the extent to which
9 months of CI experience on the second implanted side after a decade of deprivation promotes
auditory development on this side. We suggest that there are multiple sensitive periods in the maturing
auditory system. The first sensitive period closes with maturation of the brainstem. Missing this
sensitive period for restoring auditory input to both ears will compromise the normal lateralization of
auditory activity in both auditory cortices. We hypothesize that the results will indicate that a second
sensitive period coincides with maturation of the cortex, and that missing this second sensitive period
will compromise our ability to restore auditory development in the deprived pathways, perhaps

169
permanently. We speculate that this will be shown by similar underlying cortical activity at 9 months
following bilateral cochlear implantation as that of initial activation, shown in Chapters Four and Five.
7.2 Does the presence of residual hearing in the un-implanted ear protect these
pathways from abnormal effects of unilateral deprivation?
Recently in our lab, we have been asking whether the deprived pathways can be protected from
deleterious effects of deafness by promoting auditory stimulation in these pathways with the use of a
hearing aid. A recent paper by Hopyan and colleagues (2012) indicated that children who had better
hearing at 250Hz tended to use their hearing aids for longer durations prior to receiving a CI. Of
interest, these children performed significantly better on tests of music perception with their implants,
particularly for detecting differences in rhythm, compared to children who did not have acoustical
access to these low frequencies (Hopyan, Peretz et al. 2012). These findings suggest that capitalizing
on auditory development using acoustic input prior to cochlear implantation, even if only in the lower
frequencies, may be necessary to promote optimal implant hearing in children. The question of how
much residual hearing is needed in the un-implanted ear to provide a potential protective effect against
effects of unilateral stimulation/deprivation and whether receiving access to acoustic input in one ear
through the use of hearing aids and electric CI stimulation in the other (i.e., bimodal hearing) can be
used to restore binaural hearing is currently being examined in our lab.
Given that candidacy criteria for cochlear implantation were traditionally limited to individuals
with severe-to-profound hearing losses, surgical techniques for insertion of CIs were developed to
cause minimal damage to cochlear structure with little attention directed at preserving residual hearing
(Briggs, Tykocinski et al. 2005). More recently however, investigators indicated that individuals with
greater residual hearing prior to implantation had better functional outcomes with their implants
compared to their peers with no prior access to sounds (Turner, Gantz et al. 2004; Gantz, Turner et
al. 2005; Ching, van Wanrooy et al. 2007; Luntz, Yehudai et al. 2007; Campbell, Dillon et al. 2013).
Candidacy criteria for cochlear implantation have thus changed to include those with greater residual
hearing. Newer electrode designs, device technology and digital processing algorithms from CI
manufacturers (Campbell, Dillon et al. 2013) and advances in surgical techniques for cochlear
implantation now aim to preserve as much residual hearing in the implanted ear as possible to stimulate

170
the auditory pathway with both acoustic and electric hearing (with a hearing aid and a CI) in the same
ear (known as hybrid hearing), at the same time. This might restrict deafness-induced reorganization
in the auditory brain and promote development of the auditory cortex at normal rates. While positive
effects of stimulating the auditory pathways with both inputs in adults have been indicated (Turner,
Gantz et al. 2004; Briggs, Tykocinski et al. 2005; Gantz, Turner et al. 2005; Campbell, Dillon et al.
2013), this is still unclear in children. We suggest that this should be explored further in children as
preservation of residual hearing and the ability to use that hearing with CI stimulation is a tremendous
milestone in cochlear implantation. We suspect that hybrid hearing will provide a protective effect
against abnormal reorganization in the stimulated pathways and might support improved hearing in
noise, enjoyment of music and greater ease of listening in children. Given the large heterogeneity in
the cause and onset of deafness in children, as well as the degree of hearing loss across the frequency
range, understanding how much hearing was present prior implantation and how long the system was
left deprived of auditory input is essential to better predict the effects of deafness on auditory
development and outcomes after cochlear implantation.
7.3 Can auditory development in the second implanted ear be promoted by
using an aural patching method?
It is clear that providing CIs to children who are deaf within limited durations of bilateral
deafness and in both ears is essential to promote optimal hearing and auditory development of the
auditory pathways. In a recent study, Gordon and colleagues demonstrated that a sensitive period of
1.5 years exists for bilateral implantation to protect unilaterally deprived pathways from abnormal
development (Gordon, Wong et al. 2013). Unfortunately, providing a second CI to children within
this short time frame is not always possible. We have been exploring the benefits of different
rehabilitation strategies and have been asking whether the sensitive period for binaural development
can be re-opened in sequentially implanted children to promote hearing and auditory development
from the deprived ear. Recent work from our lab has been exploring whether this might be achieved
by using an aural patching method. Aural patching refers to a rehabilitation method which would
require children to remove the first implant and use the second implant alone for periods of time every
day. Successful outcomes of patching were first reported by Hubel and Weisel in kittens with
monocular deprivation (Hubel and Wiesel 1970) and was later confirmed in children with Amblyopia

171
who suffered from visual asymmetries following deprivation in one eye (Holmes, Kraker et al. 2003;
Williams, Northstone et al. 2003). Monocular patching allowed visual connections from the weaker
eye to become stronger, which in turn promoted development of binocular vision. Improvements
were found even when patching was introduced at older ages (Holmes, Kraker et al. 2003; Repka, Beck
et al. 2003; Group 2004; Menon, Shailesh et al. 2008). We wondered whether using a similar
intervention method in our sequentially implanted CI users might promote development in the
binaural pathways. We piloted this study in our lab by asking children/adolescents to use their second
implanted ear alone for 2, 8 or 24 hours per week. Unfortunately, it was difficult to encourage
children/teenagers to do this consistently. Perhaps if this therapy could be incorporated with clinical
rehabilitation techniques and tracked with the new data logging features of CIs, children may derive
benefits of aural patching.
7.4 Is auditory activity evoked by cochlear implants stimulation mediated by
mechanisms of attention or multi-sensory stimulation?
Behavioural experiments have shown that selectively directing attention to an auditory percept
is required for listeners to form an appropriate auditory image to extract meaning from words and
perceive the intended message (Hocherman, Benson et al. 1976; Rif, Hari et al. 1991; Pugh, Shaywitz
et al. 1996; Benedict, Lockwood et al. 1998; Fujiwara, Nagamine et al. 1998; Gomes, Molholm et al.
2000; Lavie 2005; Chun and Turk-Browne 2007; Okamoto, Stracke et al. 2007; Shinn-Cunningham
and Best 2008). Attention failures (i.e., disengaging attention) or alterations (i.e., over-focusing on a
stimulus), particularly for individuals with hearing impairments can significantly impact their ability to
perceive and understand speech, even when using an appropriate auditory prosthesis (Smith, Quittner
et al. 1998; Dye, Baril et al. 2007; Shinn-Cunningham and Best 2008). As suggested in the present
Thesis, children who use CIs to hear, depend on contributions from non-auditory systems when
listening to speech. We suggest that measuring the enhanced role for attention and degree of reliance
on multi-sensory input in children who are deaf and use CIs to hear is important to develop targeted
rehabilitation therapies. Busse and colleagues showed that an intimate relationship exists between
auditory and visual attention in normal hearing individuals (Busse, Roberts et al. 2005). Specifically,
visual attention has been shown to modulate auditory perception when attention is directed to a co-
occurring auditory-visual task (Oatman 1976; Alho, Woods et al. 1992; Woods, Alho et al. 1992; Alho,

172
Woods et al. 1994; Benson, Bandettini et al. 1996; Johansen-Berg, Christensen et al. 2000; Webster and
Colrain 2000; Eimer 2001; Hötting, Rösler et al. 2003; Shomstein and Yantis 2004; Sokolov, Pavlova
et al. 2004; Busse, Roberts et al. 2005; Johnson and Zatorre 2005; Rinne 2010).
Thus, given behavioural evidence for better hearing with selective attention and indications
that multi-modal attention is an important component of listening (Fujiwara, Nagamine et al. 1998;
Busse, Roberts et al. 2005; Lavie 2005; Obleser, Wise et al. 2007; Okamoto, Stracke et al. 2007; Rinne
2010), we suggest that it may be recruited differently in children who are trying to compensate for
limitations in auditory cues provided by a CI. Specifically, we wonder: Is auditory activity evoked by
CI stimulation in children modulated by multi-modal attention? Are these modulations over- or under-
expressed relative to normal? Using our beamformer and connectivity analyses, can we map the
cortical areas that mediate these changes in response to different sensory input and perceptual states?
To investigate this, we need to assess the influence of vision and touch on auditory perception by
recording cortical responses to auditory only, auditory-visual, auditory-somatosensory, and auditory-
visual-somatosensory stimuli in both a passive listening task and in a task of directed attention.
7.5 Can attention-driven and/or multi-modal auditory therapy drive
improvements in auditory processing and ease of listening?
If children using CIs are indeed subject to larger effects of attention for hearing, novel therapies
focused on training selective attention may be needed to help them capitalize on this unique way of
listening and promote optimal language and hearing skills for these children.
Current oral and aural rehabilitation therapies rely exclusively on auditory-verbal (A-V) input
to encourage acquisition of speech and language. These therapies (i.e., Lindamood Phoneme
Sequencing Program, Laureate Learning Systems and Earobics) use analytic bottom-up techniques to
rehabilitate the auditory system at a peripheral level. In general, these consist of ten 1-hour speech
perception training sessions with an A-V therapist. The focus of these interventions is on the elements
of speech to teach the listener to distinguish between sounds and to perceive simple auditory signals
provided by a CI to acquire oral speech and language. A-V therapy focuses on repetition of single
words, phrases or sentences, and it is generally administered in quiet listening environments. However,
speech is complex and conversation exchanges very seldomly take place in quiet environments.

173
Current therapies do not train the auditory system to pick out target auditory signals amidst a number
of sources, which makes it very challenging for children to transfer their skills to real-world situations.
In addition, these therapies do not target complex multi-sensory listening skills, and none have shown
any influence on selective attention processes (Fu et al., 2005; 2007; Stevens et al, 2009).
It seems that there is a gap in the generalization of rehabilitation from the laboratory to the
real-world. Rehabilitation strategies should teach CI users to use top-down processes to hear, listen
and process speech in difficult listening situations such a noise. This would enable them to learn to
suppress interfering auditory (i.e., multiple talkers – cocktail party effect) and/or non-auditory (i.e.,
integrating in a multi-sensory environment including visual, somatosensory, olfactory stimuli) inputs.
A more holistic approach to rehabilitation is needed to combine bottom-up and top-down approaches
for more effective generalization of the aural rehabilitation programs to real-world benefits. We
suggest that attention-driven auditory therapies may serve to reduce distractibility from non-auditory
inputs and teach CI users to focus auditory attentional resources to better encode, process and integrate
target auditory signals in challenging listening environments. In addition, A-V therapy is typically only
provided between 6 months to 2 years following initial cochlear implantation. However, basic
cognitive skills such as selective attention, strategic memory, planning and problem solving show rapid
development around 5 to 7 years of age in typically developing normal hearing children and around 8
to 9 years of age in CI users (Smith et al., 1998; Tharpe et al., 2002). I suggest that it may be necessary
to re-enrol these children in auditory rehabilitation therapy programs once these basic cognitive skills
have developed (Tharpe et al., 2002). More research is necessary to determine the age or duration of
CI experience needed to derive the most benefits from re-entering an auditory therapy program.
A proposed auditory rehabilitation program which targets neural mechanisms of attention is
called the Computer-Assisted Speech Training (CAST) program, developed at the House Ear Institute
(Fu et al., 2005; 2007). The CAST can be used at home and training is conducted for 1 hour per day,
5 days a week for a period of 1 month. The CAST was designed to accelerate CI users’ rehabilitation
process through intensive, repetitive and complex speech training. The CAST targets expressive and
receptive oral language skills. Over time, complexity of the tasks is increased by introducing multiple
speaker as well as background noise. A mean improvement in receptive language scores of up to 28%
in different conditions was reported and the improved performance was retained for a period of 8
weeks beyond the training period (Fu et al., 2005; 2007).

174
In the same way, Stevens and colleagues (2009) found that providing more focused high-
intensity language-training with a computerized intervention program led to increased activity in
auditory attentional networks and significant improvements in receptive language scores. This
intervention program, called Fast ForWord (FFW), targets oral language skills by training auditory
attention through intensive computer based sessions using speech and non-speech receptive and
expressive language subtests (Cohen et al., 2005; Stevens et al, 2009). The FFW training is conducted
for 100 minutes per day, 5 days a week for a period of 4 to 6 weeks, and requires the child to attend to
complex auditory stimuli. Neuroimaging evidence found increased activity in the anterior cingulate
gyrus and the lateral prefrontal cortex following FFW training (Temple et al., 2003). These areas have
strong connectivity with the inferior frontal gyrus, the dorso-lateral pre-frontal cortex and the parietal
cortex, which are crucial for attention control. Investigators reported that increased activity in this
area indicated an improvement in selective attention skills related to FFW training (Bush et al., 2000;
Stevens et al., 2009; Temple et al., 2003) and led to significant improvements on measures of executive
attention and non-verbal intelligence (Rueda et al., 2005). This suggests that attention training can
serve not only to enhance oral language skills, but also generalized intelligence and cognition (Rueda
et al., 2005; Stevens et al., 2009) in children. We suggest that developing task-specific multi-sensory
attention-driven training approaches may prove to be more appropriate for CI listening than simpler
auditory therapies. Incorporating attention-driven auditory rehabilitation programs such as FFW may
target auditory processing at a more central/integrative level, and would allow CI users to use and
manipulate the increased dependency that they have on multi-sensory/multi-perceptual input to
enhance encoding, processing and integration of auditory input in everyday, real-world challenging
listening environments.
7.6 Can holistic therapies that incorporate music and/or exercise promote
improvements in auditory processing and accelerate auditory
development after cochlear implantation?
The developing brain has a remarkable capacity to change in response to environmental
demands. It is therefore possible that alternative and holistic rehabilitation techniques which include
musical training and/or exercise therapy may also promote improvements in hearing after cochlear
implantation. Many studies have shown that musical training in early life (Moreno, Marques et al.

175
2009; Strait, Parbery-Clark et al. 2012) drives structural (Hyde, Lerch et al. 2009, Herholz and Zatorre
2012) and functional (Hyde, Lerch et al. 2009; Moreno, Marques et al. 2009; Strait, Kraus et al. 2009;
Kraus and Chandrasekaran 2010; Strait, Kraus et al. 2010; Strait and Kraus 2011; Herholz and Zatorre
2012; Strait, Parbery-Clark et al. 2012; Zendel and Alain 2012; Alain, Zendel et al. 2014) changes in the
brain which correlate with improved attention and memory capacity (Hyde, Lerch et al. 2009; Moreno,
Marques et al. 2009; Herholz and Zatorre 2012; Strait, Kraus et al. 2009; Strait, Kraus et al. 2010; Strait
and Kraus 2011), higher intelligent quotient (Moreno, Marques et al. 2009), increased reading abilities
(Moreno, Marques et al. 2009; Strait, Kraus et al. 2009; Strait, Kraus et al. 2010; Strait and Kraus 2011)
and better auditory skills in both quiet and noisy environments (Hyde, Lerch et al. 2009; Moreno,
Marques et al. 2009; Strait, Kraus et al. 2009; Kraus and Chandrasekaran 2010; Strait, Kraus et al. 2010;
Strait and Kraus 2011; Herholz and Zatorre 2012; Strait, Parbery-Clark et al. 2012). This has been
attributed to enhanced neural efficiency (i.e., activation of a larger population of neurons or increased
synchronized activity of these neurons) in response to sound and/or greater recruitment of attention
processes to discriminate auditory complexities embedded in music.
In addition to the tremendous benefits of music to auditory development and processing,
musical training in young adults with normal hearing showed significantly reduced age-related declines
in gap detection and speech-in-noise tests compared to non-musicians (Zendel and Alain 2012; Alain,
Zendel et al. 2014). This suggests that musical training may have a protective effect on general
cognitive and perceptual processes and may in turn serve to improve or accelerate auditory
development in children using CI to hear. In the same way, forms of exercise therapies have also been
shown to promote plasticity in individuals with autism (Lang, Koegel et al. 2010; Pan 2011; Sowa and
Meulenbroek 2012), influence brain recovery after a stroke in adults (Dickstein, Hocherman et al. 1986)
and lessen effects of Parkinson’s Disease (Murray, Sacheli et al. 2014). Regular, graded, customized
and integrated physical exercise improves mental health in general (Byrne and Byrne 1993; Blumenthal,
Babyak et al. 1999; Daley 2002; Colcombe and Kramer 2003) and slows down cognitive aging (Daley
2002; Heyn, Abreu et al. 2004). Thus, given the many reported positive effects of both music and
exercise therapy on brain development, plasticity and recovery, we suggest that incorporating a more
holistic approach to CI rehabilitation may help children capitalize on non-auditory functions and
develop enhanced abilities to utilize general cognitive strategies to support learning and promote
optimal hearing with a CI.

176
8. Chapter Eight – Conclusion
Data presented in this Thesis demonstrates the use of innovative methods/techniques to assess
the plasticity and development of the central auditory system in children with normal hearing and
children who are deaf and hear with a CI. We provide evidence that normal mechanisms of maturation
and specialization in the brain are experience-dependent and require hearing in both ears to be normal
with intact structure and function in the auditory pathways. Unfortunately, cortical organization in the
auditory system of adolescents who have been bilaterally deaf from early life and received a unilateral
CI as babies differs from normal in both ears. We have shown that while long-term unilateral CI
stimulation promotes maturation of auditory activity in the brainstem, midbrain and cortex, functional
abnormalities persist. Abnormally increased cortical activity in pathways from the hearing ear to the
contralateral cortex, coupled with increased recruitment of cortical areas beyond the auditory cortex
with increased neural synchrony in response to sound and compared to normal hearing peers, suggests
that CI users have developed a 1) processing bias in the left cortex, perhaps to support speech, and 2)
reliance on multi-perceptual and multi-sensory information for CI listening. These plastic changes
likely reflect cortical compensation for reorganization of the brain caused by bilateral deafness prior
to implantation, unilateral deprivation afterward, and the abnormal auditory input delivered by the CI
device. Whether this helps or hinders auditory processing remains to be understood. Increased
reliance on these non-auditory mechanisms however might mean more effortful listening and thus
reduced efficiency of CI hearing.
Despite these differences, children using CIs amaze us with their remarkable improvements in
auditory function over time and leave us hopeful that at least some degree of auditory development
proceeds normally with long-term unilateral CI use. Of concern however, some of this development
might have occurred at the expense of pathways from the opposite and deprived ear. Findings of
abnormal cortical waveform morphologies underlied by abnormally large, widespread and
disorganized/dis-connected cortical activity suggests that implanting the deprived side after the period
of brainstem and cortical maturation has passed leads to restricted and aberrant auditory development
in those pathways. It is unfortunate that abnormal changes in these pathways could not be prevented,
protected or reversed by unilateral stimulation with an implant in the opposite ear. Our current
findings highlight the deleterious effects of unilateral deprivation on the brain. We are now concerned
that these adolescents may not benefit from implantation on the second side. We must strive to

177
develop targeted therapies to exploit the parts of the auditory network that are activated by sound in
CI users to encourage optimal hearing for these children and promote their integration into the
mainstream with strong hearing and spoken language skills, educational and professional opportunities,
and social and cultural well-being with improved quality of life.

178
References
Abbott, L. F. and S. B. Nelson (2000). Synaptic plasticity: taming the beast. Nature Neuroscience 3:
1178-1183.
Abrams, D. A., T. Nicol, S. G. Zecker and N. Kraus (2006). Auditory brainstem timing predicts
cerebral asymmetry for speech. The Journal of Neuroscience 26(43): 11131-11137.
Alain, C., J. S. Snyder, Y. He and K. S. Reinke (2007). Changes in auditory cortex parallel rapid
perceptual learning. Cerebral Cortex 17(5): 1074-1084.
Albrecht, R., W. Suchodoletz and R. Uwer (2000). The development of auditory evoked dipole source
activity from childhood to adulthood. Clinical Neurophysiology 111(12): 2268-2276.
Alho, K., D. L. Woods and A. Algazi (1994). Processing of auditory stimuli during auditory and visual
attention as revealed by event‐related potentials. Psychophysiology 31(5): 469-479.
Alho, K., D. L. Woods, A. Algazi and R. Näätänen (1992). Intermodal selective attention. II. Effects
of attentional load on processing of auditory and visual stimuli in central space.
Electroencephalography and Clinical Neurophysiology 82(5): 356-368.
Anderson, V. A., P. Anderson, E. Northam, R. Jacobs and C. Catroppa (2001). Development of
executive functions through late childhood and adolescence in an Australian sample.
Developmental Neuropsychology 20(1): 385-406.
Auer Jr, E. T., L. E. Bernstein, W. Sungkarat and M. Singh (2007). Vibrotactile activation of the
auditory cortices in deaf versus hearing adults. Neuroreport 18(7): 645-648.
Awh, E., E. Vogel and S.-H. Oh (2006). Interactions between attention and working memory.
Neuroscience 139(1): 201-208.
Bach, J.-F. (2002). The effect of infections on susceptibility to autoimmune and allergic diseases. New
England Journal of Medicine 347(12): 911-920.

179
Barry, R. J., A. R. Clarke, R. McCarthy, M. Selikowitz, S. J. Johnstone and J. A. Rushby (2004). Age
and gender effects in EEG coherence: I. Developmental trends in normal children. Clinical
Neurophysiology 115(10): 2252-2258.
Basura, G. J., R. Eapen and C. A. Buchman (2009). Bilateral cochlear implantation: current concepts,
indications, and results. Laryngoscope 119(12): 2395-2401.
Batteau, D. W. (1967). The role of the pinna in human localization. Proceedings of the Royal Society of
London. Series B. Biological Sciences 168(1011): 158-180.
Bavelier, D., M. W. G. Dye and P. C. Hauser (2006). Do deaf individuals see better? Trends in Cognitive
Sciences 10(11): 512-518.
Bavelier, D. and H. J. Neville (2002). Cross-modal plasticity: where and how? Nature Reviews in
Neuroscience 3(6): 443-452.
Beadle, E. A. R., D. J. McKinley, T. P. Nikolopoulos, J. Brough, G. M. O'Donoghue and S. M.
Archbold (2005). Long-term functional outcomes and academic-occupational status in
implanted children after 10 to 14 years of cochlear implant use. Otology & Neurotology 26(6):
1152-1160.
Benedict, R. H., A. H. Lockwood, J. L. Shucard, D. W. Shucard, D. Wack and B. W. Murphy (1998).
Functional neuroimaging of attention in the auditory modality. Neuroreport 9(1): 121-126.
Benson, R. R., P. A. Bandettini, K. K. Kwong, R. J. Howard, T. Talavage, J. Belliveau and B. R. Rosen
(1996). Modulation of auditory and visual cortex by selective attention is modality-dependent.
Neuroreport 7: 1909-1913.
Bertoli, S., R. Probst and D. Bodmer (2011). Late auditory evoked potentials in elderly long-term
hearing-aid users with unilateral or bilateral fittings. Hearing Research 280(1): 58-69.
Bess, F. H., J. Dodd-Murphy and R. A. Parker (1998). Children with minimal sensorineural hearing
loss: prevalence, educational performance, and functional status. Ear and Hearing 19(5): 339-
354.

180
Bess, F. H. and A. M. Tharpe (1984). Unilateral hearing impairment in children. Pediatrics 74(2): 206-
216.
Bess, F. H. and A. M. Tharpe (1986). An introduction to unilateral sensorineural hearing loss in
children. Ear and Hearing 7(1): 3-13.
Birdsong, D. and M. Molis (2001). On the evidence for maturational constraints in second-language
acquisition. Journal of Memory and Language 44(2): 235-249.
Blair, R. C. and W. Karniski (1993). An alternative method for significance testing of waveform
difference potentials. Psychophysiology 30(5): 518-524.
Blakemore, C. (1988). The sensitive periods of the monkey visual cortex. Strabismus and Amblyopia:
Experimental Basis for Advances in Clinical Management: 219-234.
Blakemore, C. and R. C. Van Sluyters (1974). Reversal of the physiological effects of monocular
deprivation in kittens: further evidence for a sensitive period. The Journal of Physiology 237(1):
195-216.
Blakemore, S.-J. (2012). Imaging brain development: the adolescent brain. Neuroimage 61(2): 397-
406.
Blakemore, S. J. and S. Choudhury (2006). Development of the adolescent brain: implications for
executive function and social cognition. Journal of child psychology and psychiatry 47(3‐4): 296-312.
Blegvad, B. (1975). Binaural Summation of Surface-Recorded Electrocochleographic Responses
Normal-hearing Subjects. Scandinavian Audiology 4(4): 233-238.
Blumenthal, J. A., M. A. Babyak, et al. (1999). Effects of exercise training on older patients with major
depression. Archives of Internal Medicine 159(19): 2349-2356.
Bocca, E. (1955). Binaural hearing: Another approach. The Laryngoscope 65(12): 1164-1171.
Boëx, C., C. de Balthasar, M.-I. Kós and M. Pelizzone (2003). Electrical field interactions in different
cochlear implant systems. The Journal of the Acoustical Society of America 114(4): 2049-2057.

181
Bosch, L. and N. Sebastián-Gallés (1997). Native-language recognition abilities in 4-month-old infants
from monolingual and bilingual environments. Cognition 65(1): 33-69.
Bosch, L. and N. Sebastián-Gallés (2001). Evidence of early language discrimination abilities in infants
from bilingual environments. Infancy 2(1): 29-49.
Briggs, R. J., M. Tykocinski, K. Stidham and J. B. Roberson (2005). Cochleostomy site: implications
for electrode placement and hearing preservation. Acta Oto-Laryngologica 125(8): 870-876.
Brookhouser, P. E., D. W. Worthington and W. J. Kelly (1991). Unilateral hearing loss in children.
Laryngoscope 101(12): 1264-1272.
Brown, K. D. and T. J. Balkany (2007). Benefits of bilateral cochlear implantation: a review. Current
Opinion in Otolaryngology Head Neck Surgery 15(5): 315-318.
Brown, S., M. J. Martinez and L. M. Parsons (2004). Passive music listening spontaneously engages
limbic and paralimbic systems. Neuroreport 15(13): 2033-2037.
Brugge, J. F., I. O. Volkov, P. C. Garell, R. A. Reale and M. A. Howard, 3rd (2003). Functional
connections between auditory cortex on Heschl's gyrus and on the lateral superior temporal
gyrus in humans. Journal of Neurophysiology 90(6): 3750-3763.
Bruns, A. (2004). Fourier-, Hilbert-and wavelet-based signal analysis: are they really different
approaches? Journal of Neuroscience Methods 137(2): 321-332.
Buckner, R. L., J. R. Andrews‐Hanna and D. L. Schacter (2008). The brain's default network. Annals
of the New York Academy of Sciences 1124(1): 1-38.
Burnett, S., G. Bird, J. Moll, C. Frith and S.-J. Blakemore (2009). Development during adolescence
of the neural processing of social emotion. Journal of Cognitive Neuroscience 21(9): 1736-1750.
Burnett, S. and S. J. Blakemore (2009). Functional connectivity during a social emotion task in
adolescents and in adults. European Journal of Neuroscience 29(6): 1294-1301.
Burton, M. J., J. M. Miller and P. R. Kileny (1989). Middle-Latency Responses: I. Electrical and
Acoustic Excitation. Archives of Otolaryngology - Head and Neck Surgery 115(1): 59-62.

182
Busse, L., K. C. Roberts, R. E. Crist, D. H. Weissman and M. G. Woldorff (2005). The spread of
attention across modalities and space in a multisensory object. Proceedings of the National Academy
of Siences of the United States of America 102(51): 18751-18756.
Byrne, A. and D. Byrne (1993). The effect of exercise on depression, anxiety and other mood states:
a review. Journal of Psychosomatic Research 37(6): 565-574.
Calvert, G. A., P. C. Hansen, S. D. Iversen and M. J. Brammer (2001). Detection of audio-visual
integration sites in humans by application of electrophysiological criteria to the BOLD effect.
Neuroimage 14(2): 427-438.
Calvert, G. A. and T. Thesen (2004). Multisensory integration: methodological approaches and
emerging principles in the human brain. Journal of Physiology - Paris 98(1): 191-205.
Campbell, A. P., M. T. Dillon, C. A. Buchman and O. F. Adunka (2013). Hearing Preservation
Cochlear Implantation. Current Otorhinolaryngology Reports 1(2): 69-79.
Campbell, J. and A. Sharma (2013). Compensatory changes in cortical resource allocation in adults
with hearing loss. Frontiers in Systems Neuroscience 7(71): 1-9
Carhart, R. and J. F. Jerger (1959). Preferred method for clinical determination of pure-tone
thresholds. Journal of Speech and Hearing Disorders 24(4): 330-345.
Casanova, M. F., D. Buxhoeveden and J. Gomez (2003). Disruption in the inhibitory architecture of
the cell minicolumn: implications for autisim. The Neuroscientist 9(6): 496-507.
Casanova, M. F., D. P. Buxhoeveden, A. E. Switala and E. Roy (2002). Minicolumnar pathology in
autism. Neurology 58(3): 428-432.
Castañeda-Villa, N. and C. J. James (2011). Independent component analysis for auditory evoked
potentials and cochlear implant artifact estimation. Biomedical Engineering, IEEE Transactions on
58(2): 348-354.
Castaneda-Villa, N., J. Manuel Cornejo-Cruz and C. J. James (2010). Independent component analysis
for robust assessment of auditory system maturation in children with cochlear implants.
Cochlear Implants International 11(2): 71-83.

183
Chadha, N. K., B. C. Papsin, S. Jiwani and K. A. Gordon (2011). Speech detection in noise and spatial
unmasking in children with simultaneous versus sequential bilateral cochlear implants. Otology
& Neurotology 32(7): 1057-1064.
Chau, W., A. R. McIntosh, S. E. Robinson, M. Schulz and C. Pantev (2004). Improving permutation
test power for group analysis of spatially filtered MEG data. Neuroimage 23(3): 983-996.
Cheng, A. K., H. R. Rubin, N. R. Powe, N. K. Mellon, H. W. Francis and J. K. Niparko (2000). Cost-
utility analysis of the cochlear implant in children. JAMA 284(7): 850-856.
Chilosi, A. M., A. Comparini, P. Cristofani, M. Turi, S. Berrettini, F. Forli, G. Orlandi, A. Chiti, N.
Giannini and P. Cipriani (2014). Cerebral lateralization for language in deaf children with
cochlear implantation. Brain and Language 129: 1-6.
Ching, T. Y., E. van Wanrooy and H. Dillon (2007). Binaural-bimodal fitting or bilateral implantation
for managing severe to profound deafness: a review. Trends in Amplification 11(3): 161-192.
Chun, M. M. and N. B. Turk-Browne (2007). Interactions between attention and memory. Current
Opinion in Neurobiology 17(2): 177-184.
Clarke, A. R., R. J. Barry, R. McCarthy and M. Selikowitz (2001). Age and sex effects in the EEG:
development of the normal child. Clinical Neurophysiology 112(5): 806-814.
Cleary, M., D. B. Pisoni and A. E. Geers (2001). Some measures of verbal and spatial working memory
in eight-and nine-year-old hearing-impaired children with cochlear implants. Ear and Hearing
22(5): 395-411.
Cline, H. T. (2001). Dendritic arbor development and synaptogenesis. Current Opinion in Neurobiology
11(1): 118-126.
Cohn, E. S. and P. M. Kelley (1999). Clinical phenotype and mutations in connexin 26
(DFNB1/GJB2), the most common cause of childhood hearing loss. American Journal of Medical
Genetics 89(3): 130-136.
Colcombe, S. and A. F. Kramer (2003). Fitness effects on the cognitive function of older adults a
meta-analytic study. Psychological science 14(2): 125-130.

184
Collette, F., M. Hogge, E. Salmon and M. Van der Linden (2006). Exploration of the neural substrates
of executive functioning by functional neuroimaging. Neuroscience 139(1): 209-221.
Colrain, I. M. and K. B. Campbell (2007). The use of evoked potentials in sleep research. Sleep Medecine
Reviews 11(4): 277-293.
Colrain, I. M., K. E. Webster and G. Hirst (1999). The N550 component of the evoked K‐complex:
A modality non‐specific response? Journal of Sleep Research 8(4): 273-280.
Cooper, N. R., R. J. Croft, S. J. Dominey, A. P. Burgess and J. H. Gruzelier (2003). Paradox lost?
Exploring the role of alpha oscillations during externally vs. internally directed attention and
the implications for idling and inhibition hypotheses. International Journal of Psychophysiology
47(1): 65-74.
Coull, J. T. (1998). Neural correlates of attention and arousal: insights from electrophysiology,
functional neuroimaging and psychopharmacology. Progress in Neurobiology 55(4): 343-361.
Crowley, K. E. and I. M. Colrain (2004). A review of the evidence for P2 being an independent
component process: age, sleep and modality. Clinical Neurophysiology 115(4): 732-744.
Culbertson, J. L. and L. E. Gilbert (1986). Children with unilateral sensorineural hearing loss:
cognitive, academic, and social development. Ear and Hearing 7(1): 38.
Cynader, M. (1983). Prolonged sensitivity to monocular deprivation in dark-reared cats: effects of age
and visual exposure. Developmental Brain Research 8(2): 155-164.
Cynader, M., F. Leporé and J.-P. Guillemot (1981). Inter-hemispheric competition during postnatal
development Nature 290: 139-140.
Cynader, M., B. Timney and D. E. Mitchell (1980). Period of susceptibility of kitten visual cortex to
the effects of monocular deprivation extends beyond six months of age. Brain Research 191(2):
545-550.
Dalal, S. S., K. Sekihara and S. S. Nagarajan (2006). Modified beamformers for coherent source region
suppression. Biomedical Engineering, IEEE Transactions on 53(7): 1357-1363.

185
Daley, A. J. (2002). Exercise therapy and mental health in clinical populations: is exercise therapy a
worthwhile intervention? Advances in Psychiatric Treatment 8(4): 262-270.
Damaraju, E., J. R. Phillips, J. R. Lowe, R. Ohls, V. D. Calhoun and A. Caprihan (2010). Resting-state
functional connectivity differences in premature children. Frontiers in Systems Neuroscience 4(23):
1-13.
Davidson, R. (1984). Affect, cognition, and hemispheric specialization. Emotions, cognition, and
behavior, Springer: 320.
Debener, S., J. Hine, S. Bleeck and J. Eyles (2008). Source localization of auditory evoked potentials
after cochlear implantation. Psychophysiology 45(1): 20-24.
DeCasper, A. J. and W. P. Fifer (1980). Of human bonding: Newborns prefer their mothers' voices.
Science 208(4448): 1174-1176.
DeCasper, A. J. and M. J. Spence (1986). Prenatal maternal speech influences newborns' perception
of speech sounds. Infant Behavior and Development 9(2): 133-150.
Dehmel, S., Y. Cui and S. Shore (2008). Cross-modal interactions of auditory and somatic inputs in
the brainstem and midbrain and their imbalance in tinnitus and deafness. American Journal of
Audiology 17(2): S193-S209.
Delorme, A. and S. Makeig (2004). EEGLAB: an open source toolbox for analysis of single-trial
EEG dynamics including independent component analysis. Journal of Neuroscience Methods
134(1): 9-21.
Dharmaretnam, M. and L. Rogers (2005). Hemispheric specialization and dual processing in strongly
versus weakly lateralized chicks. Behavioural Brain Research 162(1): 62-70.
Dickstein, R., S. Hocherman, T. Pillar, and R. Shaham (1986). Stroke rehabilitation three exercise
therapy approaches. Physical Therapy 66(8): 1233-1238.

186
Doesburg, S. M., C. M. Chau, T. P. Cheung, A. Moiseev, U. Ribary, A. T. Herdman, S. P. Miller, I. L.
Cepeda, A. Synnes and R. E. Grunau (2013). Neonatal pain-related stress, functional cortical
activity and visual-perceptual abilities in school-age children born at extremely low gestational
age. Pain 154(10): 1946-1952.
Doesburg, S. M., L. L. Emberson, A. Rahi, D. Cameron and L. M. Ward (2008). Asynchrony from
synchrony: long-range gamma-band neural synchrony accompanies perception of audiovisual
speech asynchrony. Experimental Brain Research 185(1): 11-20.
Doesburg, S. M., J. J. Green, J. J. McDonald and L. M. Ward (2009). Rhythms of consciousness:
binocular rivalry reveals large-scale oscillatory network dynamics mediating visual perception.
PloS One 4(7) - e6142: 1-14.
Doesburg, S. M., J. J. Green, J. J. McDonald and L. M. Ward (2012). Theta modulation of inter-
regional gamma synchronization during auditory attention control. Brain Research 1431: 77-85.
Doesburg, S. M., A. T. Herdman, U. Ribary, T. Cheung, A. Moiseev, H. Weinberg, M. Liotti, D. Weeks
and R. E. Grunau (2010). Long-range synchronization and local desynchronization of alpha
oscillations during visual short-term memory retention in children. Experimental Brain Research
201(4): 719-727.
Doesburg, S. M., K. Kitajo and L. M. Ward (2005). Increased gamma-band synchrony precedes
switching of conscious perceptual objects in binocular rivalry. Neuroreport 16(11): 1139-1142.
Doesburg, S. M., A. Moiseev, A. T. Herdman, U. Ribary and R. E. Grunau (2013). Region-specific
slowing of alpha oscillations is associated with visual-perceptual abilities in children born very
preterm. Frontiers in Human Neuroscience 7(791): 1-9.
Doesburg, S. M., U. Ribary, A. T. Herdman, A. Moiseev, T. Cheung, S. P. Miller, K. J. Poskitt, H.
Weinberg, M. F. Whitfield and A. Synnes (2011). Magnetoencephalography reveals slowing
of resting peak oscillatory frequency in children born very preterm. Pediatric Research 70(2):
171-175.
Doesburg, S. M., A. B. Roggeveen, K. Kitajo and L. M. Ward (2008). Large-scale gamma-band phase
synchronization and selective attention. Cerebral Cortex 18(2): 386-396.

187
Doesburg, S. M. and L. M. Ward (2009). Synchronization between sources: emerging methods for
understanding large-scale functional networks in the human brain. Coordinated Activity in the
Brain, Springer: 25-42.
Dorrn, A. L., K. Yuan, A. J. Barker, C. E. Schreiner and R. C. Froemke (2010). Developmental
sensory experience balances cortical excitation and inhibition. Nature 465(7300): 932-936.
Doucet, M., F. Bergeron, M. Lassonde, P. Ferron and F. Lepore (2006). Cross-modal reorganization
and speech perception in cochlear implant users. Brain 129(12): 3376-3383.
Drennan, W. R. and J. T. Rubinstein (2008). Music perception in cochlear implant users and its
relationship with psychophysical capabilities. Journal of Rehabilitation Research and Development
45(5): 779.
Dye, M. W., D. E. Baril and D. Bavelier (2007). Which aspects of visual attention are changed by
deafness? The case of the Attentional Network Test. Neuropsychologia 45(8): 1801-1811.
Eapen, R. J. and C. A. Buchman (2009). Bilateral cochlear implantation: current concepts. Current
Opinions in Otolaryngology - Head & Neck Surgery 17(5): 351-355.
Eggermont, J. (1985). Evoked potentials as indicators of auditory maturation. Acta Oto-laryngologica
99(S421): 41-47.
Eggermont, J. (2007). Electric and magnetic fields of synchronous neural activity. Auditory Evoked
Potentials: Basic Principles and Clinical Application, Springer: 2-21.
Eggermont, J. J. (1988). On the rate of maturation of sensory evoked potentials. Electroencephalography
and Clinical Neurophysiology 70(4): 293-305.
Eggermont, J. J. (2000). Sound-induced synchronization of neural activity between and within three
auditory cortical areas. Journal of Neurophysiology 83(5): 2708-2722.
Eggermont, J. J. (2008). The role of sound in adult and developmental auditory cortical plasticity.
Ear and Hearing 29(6): 819-829.

188
Eggermont, J. J. and J. K. Moore (2012). Morphological and functional development of the auditory
nervous system. Human auditory development, Springer: 61-105.
Eggermont, J. J. and C. W. Ponton (2002). The neurophysiology of auditory perception: from single
units to evoked potentials. Audiology and Neurotology 7(2): 71-99.
Eggermont, J. J. and C. W. Ponton (2003). Auditory-evoked Potential Studies of Cortical Maturation
in Normal Hearing and Implanted Children: Correlations with Changes in Structure and
Speech Perception. Acta Oto-laryngologica 123(2): 249-252.
Eggermont, J. J., C. W. Ponton, M. Don, M. D. Waring and B. Kwong (1997). Maturational delays in
cortical evoked potentials in cochlear implant users. Acta Oto-laryngologica 117(2): 161-163.
Eimer, M. (2001). Crossmodal links in spatial attention between vision, audition, and touch: evidence
from event-related brain potentials. Neuropsychologia 39(12): 1292-1303.
Elias, G., P. Apostolides, A. Kriegstein and R. Nicoll (2008). Differential trafficking of AMPA and
NMDA receptors by SAP102 and PSD-95 underlies synapse development. Proceedings of the
National Academy of Sciences 105(52): 20953-20958.
Emmorey, K., J. S. Allen, J. Bruss, N. Schenker and H. Damasio (2003). A morphometric analysis of
auditory brain regions in congenitally deaf adults. Proceedings of the National Academy of Sciences
100(17): 10049-10054.
Englot, D. J., L. Yang, H. Hamid, N. Danielson, X. Bai, A. Marfeo, L. Yu, A. Gordon, M. J. Purcaro
and J. E. Motelow (2010). Impaired consciousness in temporal lobe seizures: role of cortical
slow activity. Brain: 1-14.
Fan, J., B. D. McCandliss, J. Fossella, J. I. Flombaum and M. I. Posner (2005). The activation of
attentional networks. Neuroimage 26(2): 471-479.
Fan, J., B. D. McCandliss, T. Sommer, A. Raz and M. I. Posner (2002). Testing the efficiency and
independence of attentional networks. Journal of Cognitive Neuroscience 14(3): 340-347.
Fatima, Z., M. A. Quraan, N. Kovacevic and A. R. McIntosh (2013). ICA-based artifact correction
improves spatial localization of adaptive spatial filters in MEG. Neuroimage 78: 284-294.

189
Fifer, R. C. and B. Sierra-Irizarry (1988). Clinical applications of the auditory middle latency response.
Otology & Neurotology 9(5): 47-56.
Finney, E. M., I. Fine and K. R. Dobkins (2001). Visual stimuli activate auditory cortex in the deaf.
Nature Neuroscience 4(12): 1171-1173.
Firszt, J. B., J. L. Ulmer and W. Gaggl (2006). Differential representation of speech sounds in the
human cerebral hemispheres. The Anatomical Record Part A: Discoveries in Molecular, Cellular, and
Evolutionary Biology 288(4): 345-357.
Fitzgerald, M. B., J. E. Green, Y. Fang and S. B. Waltzman (2013). Factors influencing consistent
device use in pediatric recipients of bilateral cochlear implants. Cochlear Implants International
14(5): 257-265.
Fitzpatrick, M. R. and S. Irannejad (2008). Adolescent readiness for change and the working alliance
in counseling. Journal of Counseling & Development 86(4): 438-445.
Flege, J. E., G. H. Yeni-Komshian and S. Liu (1999). Age constraints on second-language acquisition.
Journal of Memory and Language 41(1): 78-104.
Fortnum, H. M., A. Q. Summerfield, D. H. Marshall, A. C. Davis, J. M. Bamford, A. Davis, C.
Yoshinaga-Itano and S. Hind (2001). Prevalence of permanent childhood hearing impairment
in the United Kingdom and implications for universal neonatal hearing screening:
questionnaire based ascertainment studyCommentary: Universal newborn hearing screening:
implications for coordinating and developing services for deaf and hearing impaired children.
BMJ 323(7312): 536.
Fougnie, D. (2008). The relationship between attention and working memory. New Research on Short-
Term Memory: 1-45.
Friauf, E. and C. Lohmann (1999). Development of auditory brainstem circuitry. Cell and Tissue
Research 297(2): 187-195.
Fries, P. (2005). A mechanism for cognitive dynamics: neuronal communication through neuronal
coherence. Trends in Cognitive Sciences 9(10): 474-480.

190
Friesen, L. M. and T. W. Picton (2010). A method for removing cochlear implant artifact. Hearing
Research 259(1-2): 95-106.
Frizzo, A. C. F., C. A. R. Funayama, M. L. Isaac and J. F. Colafêmina (2007). Auditory Middle Latency
Responses: a study of healthy children. Revista Brasileira de Otorrinolaringologia 73(3): 398-403.
Fuchs, A. (2007). Beamforming and its applications to brain connectivity. Handbook of Brain
Connectivity: 357-378.
Fujioka, T., B. Ross, R. Kakigi, C. Pantev and L. J. Trainor (2006). One year of musical training affects
development of auditory cortical-evoked fields in young children. Brain 129(10): 2593-2608.
Fujiwara, N., T. Nagamine, M. Imai, T. Tanaka and H. Shibasaki (1998). Role of the primary auditory
cortex in auditory selective attention studied by whole-head neuromagnetometer. Cognitive
Brain Research 7(2): 99-109.
Galvin, K. L., M. Mok and R. C. Dowell (2007). Perceptual benefit and functional outcomes for
children using sequential bilateral cochlear implants. Ear and Hearing 28(4): 470-482.
Gantz, B. J., C. Turner, K. E. Gfeller and M. W. Lowder (2005). Preservation of hearing in cochlear
implant surgery: advantages of combined electrical and acoustical speech processing. The
Laryngoscope 115(5): 796-802.
Gao, W. J., A. B. Wormington, D. E. Newman and S. L. Pallas (2000). Development of inhibitory
circuitry in visual and auditory cortex of postnatal ferrets: Immunocytochemical localization of
calbindin‐and parvalbumin‐containing neurons. Journal of Comparative Neurology 422(1): 140-
157.
García-Larrea, L., A. C. Lukaszewicz and F. Mauguiére (1992). Revisiting the oddball paradigm. Non-
target vs neutral stimuli and the evaluation of ERP attentional effects. Neuropsychologia 30(8):
723-741.
Geers, A., E. Tobey, J. Moog and C. Brenner (2008). Long-term outcomes of cochlear implantation
in the preschool years: From elementary grades to high school. International Journal of Audiology
47(S2): S21-S30.

191
Geers, A. E., J. G. Nicholas and A. L. Sedey (2003). Language skills of children with early cochlear
implantation. Ear and Hearing 24(1): 46S-58S.
Geers, A. E., D. B. Pisoni and C. Brenner (2012). Complex Working Memory Span in Cochlear
Implanted and Normal Hearing Teenagers. Otology & Neurotology 34(3): 396-401.
Geers, A. E. and A. L. Sedey (2011). Language and verbal reasoning skills in adolescents with 10 or
more years of cochlear implant experience. Ear and Hearing 32(1 Suppl): 39S-48S.
Giard, M. and F. Peronnet (1999). Auditory-visual integration during multimodal object recognition
in humans: a behavioral and electrophysiological study. Journal of Cognitive Neuroscience 11(5):
473-490.
Giedd, J. N., J. Blumenthal, N. O. Jeffries, F. X. Castellanos, H. Liu, A. Zijdenbos, T. Paus, A. C.
Evans and J. L. Rapoport (1999). Brain development during childhood and adolescence: a
longitudinal MRI study. Nature Neuroscience 2(10): 861-863.
Giedd, J. N., J. W. Snell, N. Lange, J. C. Rajapakse, B. Casey, P. L. Kozuch, A. C. Vaituzis, Y. C. Vauss,
S. D. Hamburger and D. Kaysen (1996). Quantitative magnetic resonance imaging of human
brain development: ages 4–18. Cerebral Cortex 6(4): 551-559.
Gilley, P. M., A. Sharma, M. Dorman, C. C. Finley, A. S. Panch and K. Martin (2006). Minimization
of cochlear implant stimulus artifact in cortical auditory evoked potentials. Clinical
Neurophysiology 117(8): 1772-1782.
Gilley, P. M., A. Sharma, M. Dorman and K. Martin (2005). Developmental changes in refractoriness
of the cortical auditory evoked potential. Clinical Neurophysiology 116(3): 648-657.
Gilley, P. M., A. Sharma and M. F. Dorman (2008). Cortical reorganization in children with cochlear
implants. Brain Research 1239: 56-65.
Giraud, A. L. and H. J. Lee (2007). Predicting cochlear implant outcome from brain organisation in
the deaf. Restorative Neurology and Neuroscience 25(3): 381-390.
Giraud, A. L., C. J. Price, J. M. Graham and R. S. Frackowiak (2001). Functional plasticity of language-
related brain areas after cochlear implantation. Brain 124(7): 1307-1316.

192
Giraud, A. L., C. J. Price, J. M. Graham, E. Truy and R. S. J. Frackowiak (2001). Cross-modal plasticity
underpins language recovery after cochlear implantation. Neuron 30(3): 657-664.
Glendenning, K., J. Brusno‐Bechtold, G. Thompson and R. Masterton (1981). Ascending auditory
afferents to the nuclei of the lateral leminscus. Journal of Comparative Neurology 197(4): 673-703.
Godey, B., D. Schwartz, J. De Graaf, P. Chauvel and C. Liegeois-Chauvel (2001). Neuromagnetic
source localization of auditory evoked fields and intracerebral evoked potentials: a comparison
of data in the same patients. Clinical Neurophysiology 112(10): 1850-1859.
Gomes, H., S. Molholm, C. Christodoulou, W. Ritter and N. Cowan (2000). The development of
auditory attention in children. Frontiers in Bioscience 5(1): D108-120.
Gopen, Q., J. J. Rosowski and S. N. Merchant (1997). Anatomy of the normal human cochlear
aqueduct with functional implications. Hearing Research 107(1): 9-22.
Gordon, K., D. D. Wong, J. Valero, S. Jewell, P. Yoo and B. Papsin (2011). Use it or lose it? Lessons
learned from the developing brains of children who are deaf and use cochlear implants to hear.
Brain Topography 24(3): 204-219.
Gordon, K. A., S. Jiwani and B. C. Papsin (2011). What is the optimal timing for bilateral cochlear
implantation in children? Cochlear Implants International 12 Suppl 2: S8-14.
Gordon, K. A., S. Jiwani and B. C. Papsin (2013). Benefits and detriments of unilateral cochlear
implant use on bilateral auditory development in children who are deaf. Frontiers in Psychology
4(719): 1-14.
Gordon, K. A. and B. C. Papsin (2009). Benefits of short interimplant delays in children receiving
bilateral cochlear implants. Otology & Neurotology 30(3): 319-331.
Gordon, K. A., B. C. Papsin and R. V. Harrison (2003). Activity-dependent developmental plasticity
of the auditory brain stem in children who use cochlear implants. Ear and Hearing 24(6): 485-
500.
Gordon, K. A., B. C. Papsin and R. V. Harrison (2005). Effects of cochlear implant use on the
electrically evoked middle latency response in children. Hearing Research 204(1-2): 78-89.

193
Gordon, K. A., B. C. Papsin and R. V. Harrison (2006). An evoked potential study of the
developmental time course of the auditory nerve and brainstem in children using cochlear
implants. Audiology and Neurotology 11(1): 7-23.
Gordon, K. A., B. C. Papsin and R. V. Harrison (2007). Auditory brainstem activity and development
evoked by apical versus basal cochlear implant electrode stimulation in children. Clinical
Neurophysiology 118(8): 1671-1684.
Gordon, K. A., C. Salloum, G. S. Toor, R. van Hoesel and B. C. Papsin (2012). Binaural Interactions
Develop in the Auditory Brainstem of Children Who Are Deaf: Effects of Place and Level of
Bilateral Electrical Stimulation. The Journal of Neuroscience 32(12): 4212-4223.
Gordon, K. A., S. Tanaka and B. C. Papsin (2005). Atypical cortical responses underlie poor speech
perception in children using cochlear implants. Neuroreport 16(18): 2041-2045.
Gordon, K. A., S. Tanaka, D. D. Wong and B. C. Papsin (2008). Characterizing responses from
auditory cortex in young people with several years of cochlear implant experience. Clinical
Neurophysiology 119(10): 2347-2362.
Gordon, K. A., S. Tanaka, D. D. Wong, T. Stockley, J. D. Ramsden, T. Brown, S. Jewell and B. C.
Papsin (2011). Multiple effects of childhood deafness on cortical activity in children receiving
bilateral cochlear implants simultaneously. Clinical Neurophysiology 122(4): 823-833.
Gordon, K. A., J. Valero and B. C. Papsin (2007). Auditory brainstem activity in children with 9-30
months of bilateral cochlear implant use. Hearing Research 233(1-2): 97-107.
Gordon, K. A., J. Valero, R. van Hoesel and B. C. Papsin (2008). Abnormal timing delays in auditory
brainstem responses evoked by bilateral cochlear implant use in children. Otology & Neurotology
29(2): 193-198.
Gordon, K. A., D. D. Wong and B. C. Papsin (2013). Bilateral input protects the cortex from
unilaterally-driven reorganization in children who are deaf. Brain 136(5): 1609-1625.
Gordon, K. A., D. D. E. Wong and B. C. Papsin (2010). Cortical function in children receiving
bilateral cochlear implants simultaneously or after a period of interimplant delay. Otology &
Neurotology 31(8): 1293-1299.

194
Gotts, S. J., H. J. Jo, G. L. Wallace, Z. S. Saad, R. W. Cox and A. Martin (2013). Two distinct forms
of functional lateralization in the human brain. Proceedings of the National Academy of Sciences
110(36): E3435-3444.
Grady, C. L., J. W. Van Meter, J. M. Maisog, P. Pietrini, J. Krasuski and J. P. Rauschecker (1997).
Attention-related modulation of activity in primary and secondary auditory cortex. Neuroreport
8(11): 2511-2516.
Graham, J. and D. Vickers (2011). Evidence of a ‘critical age’for sequential implantation of the second
ear in congenitally deaf children. Cochlear Implants International 12(s1): S121-S123.
Graham, J., D. Vickers, J. Eyles, J. Brinton, G. A. Malky, W. Aleksy, J. Martin, L. Henderson, D.
Mawman and P. Robinson (2009). Bilateral sequential cochlear implantation in the
congenitally deaf child: evidence to support the concept of a ‘critical age’after which the second
ear is less likely to provide an adequate level of speech perception on its own. Cochlear Implants
International 10(3): 119-141.
Graven, S. N. and J. V. Browne (2008). Auditory development in the fetus and infant. Newborn and
Infant Nursing Reviews 8(4): 187-193.
Green, J. J., S. M. Doesburg, L. M. Ward and J. J. McDonald (2011). Electrical neuroimaging of
voluntary audiospatial attention: evidence for a supramodal attention control network. Journal
of Neuroscience 31(10): 3560-3564.
Grieco-Calub, T. M. and R. Y. Litovsky (2010). Sound localization skills in children who use bilateral
cochlear implants and in children with normal acoustic hearing. Ear and Hearing 31(5): 645.
Grothe, B. (2003). New roles for synaptic inhibition in sound localization. Nature Reviews Neuroscience
4(7): 540-550.
Grothe, B., M. Pecka and D. McAlpine (2010). Mechanisms of sound localization in mammals.
Physiological Reviews 90(3): 983-1012.
Group, P. E. D. I. (2004). A prospective, pilot study of treatment of amblyopia in children 10 to< 18
years old. American Journal of Ophthalmology 137(3): 581.

195
Gualandi, F., A. Ravani, A. Berto, A. Sensi, C. Trabanelli, F. Falciano, P. Trevisi, M. Mazzoli, M.
Tibiletti and E. Cristofari (2002). Exploring the clinical and epidemiological complexity of
GJB2‐linked deafness. American Journal of Medical Genetics 112(1): 38-45.
Hardie, N. A., A. Martsi-McClintock, L. M. Aitkin and R. K. Shepherd (1998). Neonatal sensorineural
hearing loss affects synaptic density in the auditory midbrain. Neuroreport 9(9): 2019-2022.
Hari, R. (1990). The neuromagnetic method in the study of the human auditory cortex. Advances in
Audiology 6: 222-282.
Harrison, R. V., K. A. Gordon and R. J. Mount (2005). Is there a critical period for cochlear
implantation in congenitally deaf children? Analyses of hearing and speech perception
performance after implantation. Developmental Psychobiology 46(3): 252-261.
Harrison, R. V., S. G. Stanton and R. J. Mount (1995). Effects of chronic cochlear damage on
threshold and frequency tuning of neurons in AI auditory cortex. Acta Oto-Laryngologica
115(S519): 30-35.
Hartmann, R., R. Shepherd, S. Heid and R. Klinke (1997). Response of the primary auditory cortex
to electrical stimulation of the auditory nerve in the congenitally deaf white cat. Hearing Research
112(1): 115-133.
Hashisaki, G. T. and E. W. Rubel (1989). Effects of unilateral cochlea removal on anteroventral
cochlear nucleus neurons in developing gerbils. Journal of Comparative Neurology 283(4): 465-
473.
Hawley, M. L., R. Y. Litovsky and J. F. Culling (2004). The benefit of binaural hearing in a cocktail
party: Effect of location and type of interferer. The Journal of the Acoustical Society of America 115:
833-843.
Hebb, D. O. (1949). The Organization of Behavior, Wiley, New York.
Hecox, K. and R. Burkard (2006). Developmental dependencies of the human brainstem auditory
evoked response. Annals of the New York Academy of Sciences 338(1): 538-556.

196
Heid, S., R. Hartmann and R. Klinke (1998). A model for prelingual deafness, the congenitally deaf
white cat–population statistics and degenerative changes. Hearing Research 115(1): 101-112.
Heid, S., T. Jähn-Siebert, R. Klinke, R. Hartmann and G. Langner (1997). Afferent projection patterns
in the auditory brainstem in normal and congenitally deaf white cats. Hearing Research 110(1-
2): 191-199.
Henry, B. A. and C. W. Turner (2003). The resolution of complex spectral patterns by cochlear
implant and normal-hearing listeners. The Journal of the Acoustical Society of America 113(5): 2861-
2873.
Herholz, S. C. and R. J. Zatorre (2012). Musical training as a framework for brain plasticity: behavior,
function, and structure. Neuron 76(3): 486-502.
Heyn, P., B. C. Abreu and K. J. Ottenbacher (2004). The effects of exercise training on elderly persons
with cognitive impairment and dementia: a meta-analysis. Archives of Physical Medicine and
Rehabilitation 85(10): 1694-1704.
Hine, J. and S. Debener (2007). Late auditory evoked potentials asymmetry revisited. Clinical
Neurophysiology 118(6): 1274-1285.
Hine, J., R. Thornton, A. Davis and S. Debener (2008). Does long-term unilateral deafness change
auditory evoked potential asymmetries? Clinical Neurophysiology 119(3): 576-586.
Hocherman, S., D. Benson, M. Goldstein, H. Heffner and R. Hienz (1976). Evoked unit activity in
auditory cortex of monkeys performing a selective attention task. Brain Research 117(1): 51-68.
Holden, L. K., M. W. Skinner, T. A. Holden and M. E. Demorest (2002). Effects of stimulation rate
with the Nucleus 24 ACE speech coding strategy. Ear and Hearing 23(5): 463-476.
Holmes, J., R. Kraker, R. Beck, E. Birch, S. Cotter, D. Everett, R. Hertle, G. Quinn, M. Repka and M.
Scheiman (2003). A randomized trial of prescribed patching regimens for treatment of severe
amblyopia in children. Ophthalmology 110(11): 2075-2087.

197
Hopyan-Misakyan, T. M., K. A. Gordon, M. Dennis and B. C. Papsin (2009). Recognition of affective
speech prosody and facial affect in deaf children with unilateral right cochlear implants. Child
Neuropsychology 15(2): 136-146.
Hopyan, T., K. Gordon and B. Papsin (2011). Identifying emotions in music through electrical hearing
in deaf children using cochlear implants. Cochlear Implants International 12(1): 21-26.
Hopyan, T., I. Peretz, L. P. Chan, B. C. Papsin and K. A. Gordon (2012). Children using cochlear
implants capitalize on acoustical hearing for music perception. Frontiers in psychology 3(125): 1-
9.
Hötting, K., F. Rösler and B. Röder (2003). Crossmodal and intermodal attention modulate event-
related brain potentials to tactile and auditory stimuli. Experimental Brain Research 148(1): 26-
37.
Hu, B. (2003). Functional organization of lemniscal and nonlemniscal auditory thalamus. Experimental
Brain Research 153(4): 543-549.
Hu, B., V. Senatorov and D. Mooney (1994). Lemniscal and non-lemniscal synaptic transmission in
rat auditory thalamus. The Journal of Physiology 479(Pt 2): 217-231.
Hubel, D. H. and T. N. Wiesel (1970). The period of susceptibility to the physiological effects of
unilateral eye closure in kittens. The Journal of Physiology 206(2): 419-436.
Hubel, D. H., T. N. Wiesel and S. LeVay (1977). Plasticity of ocular dominance columns in monkey
striate cortex. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences: 377-
409.
Hughes, J. R. (2007). Autism: the first firm finding= underconnectivity? Epilepsy & Behavior 11(1):
20-24.
Hughes, K. C. and K. L. Galvin (2013). Measuring listening effort expended by adolescents and young
adults with unilateral or bilateral cochlear implants or normal hearing. Cochlear Implants
International 14(3): 121-129.

198
Huttenlocher, P. R. (1967). Development of cortical neuronal activity in the neonatal cat. Experimental
Neurology 17(3): 247-262.
Huttenlocher, P. R. (1970). Myelination and the development of function in immature pyramidal
tract. Experimental Neurology 29(3): 405-415.
Huttenlocher, P. R. and A. S. Dabholkar (1997). Regional differences in synaptogenesis in human
cerebral cortex. Journal of Computational Neurology 387(2): 167-178.
Hyde, K. L., J. Lerch, A. Norton, M. Forgeard, E. Winner, A. C. Evans and G. Schlaug (2009). Musical
training shapes structural brain development. The Journal of Neuroscience 29(10): 3019-3025.
Hyde, K. L., J. Lerch, A. Norton, M. Forgeard, E. Winner, A. C. Evans and G. Schlaug (2009). Musical
training shapes structural brain development. The Journal of Neuroscience 29(10): 3019-3025.
Hyde, M. and R. Punch (2011). The modes of communication used by children with cochlear implants
and role of sign in their lives. American Annals of the Deaf 155(5): 535-549.
Hyde, M., R. Punch and P. Grimbeek (2011). Factors predicting functional outcomes of cochlear
implants in children. Cochlear Implants International 12(2): 94-104.
Hyvärinen, A. and E. Oja (1997). A fast fixed-point algorithm for independent component analysis.
Neural Computation 9(7): 1483-1492.
Illg, A., A. Giourgas, A. Kral, A. Büchner, A. Lesinski-Schiedat and T. Lenarz (2013). Speech
comprehension in children and adolescents after sequential bilateral cochlear implantation with
long interimplant interval. Otology & Neurotology 34(4): 682-689.
Illing, R.-B., K. S. Kraus and M. A. Meidinger (2005). Reconnecting neuronal networks in the auditory
brainstem following unilateral deafening. Hearing Research 206(1): 185-199.
Jamison, H. L., K. E. Watkins, D. V. Bishop and P. M. Matthews (2006). Hemispheric specialization
for processing auditory nonspeech stimuli. Cerebral Cortex 16(9): 1266-1275.

199
Jäncke, L., S. Mirzazade and N. Joni Shah (1999). Attention modulates activity in the primary and the
secondary auditory cortex: a functional magnetic resonance imaging study in human subjects.
Neuroscience Letters 266(2): 125-128.
Jeffrey, B. G., Y.-Z. Wang and E. E. Birch (2004). Altered global shape discrimination in deprivation
amblyopia. Vision Research 44(2): 167-177.
Jensen, O., J. Kaiser and J. P. Lachaux (2007). Human gamma-frequency oscillations associated with
attention and memory. Trends in Neuroscience 30(7): 317-324.
Jeong Lee, H., E. Kang, S.-H. Oh, H. Kang, D. Soo Lee, M. Chul Lee and C.-S. Kim (2005).
Preoperative differences of cerebral metabolism relate to the outcome of cochlear implants in
congenitally deaf children. Hearing Research 203(1): 2-9.
Jerger, J. and J. Hall (1980). Effects of age and sex on auditory brainstem response. Archives of
Otolaryngology - Head & Neck Surgery 106(7): 387-391.
Jiwani, S., B. C. Papsin and K. Gordon (Submitted). Early unilateral cochlear implantation promotes
mature cortical asymmetries in adolescents who are deaf. Human Brain Mapping.
Jiwani, S., B. C. Papsin and K. A. Gordon (2013). Central auditory development after long-term
cochlear implant use. Clinical Neurophysiology 124(9): 1868-1880.
Jiwani, S., S. Tanaka, B. C. Papsin and K. Gordon (In Preparation). Auditory development after
cochlear implantation in children who use sign language as their primary mode of
communication.
Johansen-Berg, H., V. Christensen, M. Woolrich and P. M. Matthews (2000). Attention to touch
modulates activity in both primary and secondary somatosensory areas. Neuroreport 11(6):
1237-1241.
Johnson, J. A. and R. J. Zatorre (2005). Attention to simultaneous unrelated auditory and visual
events: behavioral and neural correlates. Cerebral Cortex 15(10): 1609-1620.

200
Johnson, J. S. and E. L. Newport (1989). Critical period effects in second language learning: The
influence of maturational state on the acquisition of English as a second language. Cognitive
Psychology 21(1): 60-99.
Johnson, M. H. (2001). Functional brain development in humans. Nature Reviews Neuroscience 2(7):
475-483.
Johnson, M. H. (2011). Interactive specialization: a domain-general framework for human functional
brain development? Developmental Cognitive Neuroscience 1(1): 7-21.
Kadis, D. S., E. W. Pang, T. Mills, M. J. Taylor, M. P. McAndrews and M. L. Smith (2011).
Characterizing the normal developmental trajectory of expressive language lateralization using
magnetoencephalography. Journal of the International Neuropsychological Society 17(5): 896-904.
Kelley, P., D. Harris, B. Comer, J. Askew, T. Fowler, S. Smith and W. Kimberling (1998). Novel
mutations in the connexin 26 gene (GJB2) that cause autosomal recessive (DFNB1) hearing
loss. The American Journal of Human Genetics 62(4): 792-799.
Khan, S., A. Gramfort, N. R. Shetty, M. G. Kitzbichler, S. Ganesan, J. M. Moran, S. M. Lee, J. D.
Gabrieli, H. B. Tager-Flusberg and R. M. Joseph (2013). Local and long-range functional
connectivity is reduced in concert in autism spectrum disorders. Proceedings of the National
Academy of Sciences 110(8): 3107-3112.
Khosla, D., C. W. Ponton, J. J. Eggermont, B. Kwong, M. Dort and J.-P. Vasama (2003). Differential
ear effects of profound unilateral deafness on the adult human central auditory system. JARO-
Journal of the Association for Research in Otolaryngology 4(2): 235-249.
Kil, J., G. Hkageyama, M. N. Semple and L. M. Kitzes (1995). Development of ventral cochlear
nucleus projections to the superior olivary complex in gerbil. Journal of Comparative Neurology
353(3): 317-340.
Kimura, D. (1973). The asymmetry of the human brain. Scientific American.
King, A. J. (2010). Auditory neuroscience: balancing excitation and inhibition during development.
Current Biology 20(18): R808-810.

201
Kitzes, L. (1984). Some physiological consequences of neonatal cochlear destruction in the inferior
colliculus of the gerbil,< i> Meriones unguiculatus</i>. Brain Research 306(1): 171-178.
Kitzes, L. M., G. H. Kageyama, M. N. Semple and J. Kil (1995). Development of ectopic projections
from the ventral cochlear nucleus to the superior olivary complex induced by neonatal ablation
of the contralateral cochlea. Journal of Comparative Neurology 353(3): 341-363.
Klinge, C., F. Eippert, B. Roder and C. Buchel (2010). Corticocortical connections mediate primary
visual cortex responses to auditory stimulation in the blind. Journal of Neurosci 30(38): 12798-
12805.
Klinke, R., A. Kral, S. Heid, J. Tillein and R. Hartmann (1999). Recruitment of the auditory cortex in
congenitally deaf cats by long-term cochlear electrostimulation. Science 285(5434): 1729-1733.
Koelewijn, T., A. A. Zekveld, J. M. Festen and S. E. Kramer (2012). Pupil dilation uncovers extra
listening effort in the presence of a single-talker masker. Ear and Hearing 33(2): 291-300.
Kok, M. A., N. Chabot and S. G. Lomber (2014). Cross‐modal reorganization of cortical afferents to
dorsal auditory cortex following early‐and late‐onset deafness. Journal of Comparative Neurology
522(3): 654-675.
Konrad, K. and S. B. Eickhoff (2010). Is the ADHD brain wired differently? A review on structural
and functional connectivity in attention deficit hyperactivity disorder. Human Brain Mapping
31(6): 904-916.
Kotak, V. C., S. Fujisawa, F. A. Lee, O. Karthikeyan, C. Aoki and D. H. Sanes (2005). Hearing loss
raises excitability in the auditory cortex. The Journal of Neuroscience 25(15): 3908-3918.
Kotak, V. C. and D. H. Sanes (1996). Developmental influence of glycinergic transmission: regulation
of NMDA receptor-mediated EPSPs. The Journal of Neuroscience 16(5): 1836-1843.
Kral, A. (2007). Unimodal and cross-modal plasticity in the 'deaf' auditory cortex. International Journal
of Audiology 46(9): 479-493.
Kral, A. (2013). Auditory critical periods: a review from system’s perspective. Neuroscience 247: 117-
133.

202
Kral, A. and J. J. Eggermont (2007). What's to lose and what's to learn: development under auditory
deprivation, cochlear implants and limits of cortical plasticity. Brain Research Reviews 56(1): 259-
269.
Kral, A., R. Hartmann, J. Tillein, S. Heid and R. Klinke (2001). Delayed maturation and sensitive
periods in the auditory cortex. Audiology and Neurotology 6(6): 346-362.
Kral, A., R. Hartmann, J. Tillein, S. Heid and R. Klinke (2002). Hearing after congenital deafness:
central auditory plasticity and sensory deprivation. Cerebral Cortex 12(8): 797-807.
Kral, A., S. Heid, P. Hubka and J. Tillein (2013). Unilateral hearing during development: hemispheric
specificity in plastic reorganizations. Frontiers in Systems Neuroscience 7(93): 1-13.
Kral, A., P. Hubka, S. Heid and J. Tillein (2013). Single-sided deafness leads to unilateral aural
preference within an early sensitive period. Brain 136(1): 180-193.
Kral, A. and G. M. O'Donoghue (2010). Profound deafness in childhood. New England Journal of
Medicine 363(15): 1438-1450.
Kral, A., J. H. Schroder, R. Klinke and A. K. Engel (2003). Absence of cross-modal reorganization
in the primary auditory cortex of congenitally deaf cats. Experimental Brain Research 153(4):
605-613.
Kral, A. and A. Sharma (2012). Developmental neuroplasticity after cochlear implantation. Trends in
Neurosciences 35(2): 111-122.
Kraus, N. and B. Chandrasekaran (2010). Music training for the development of auditory skills. Nature
Reviews Neuroscience 11(8): 599-605.
Kraus, N. and T. McGee (1993). Clinical implications of primary and nonprimary pathway
contributions to the middle latency response generating system. Ear and Hearing 14(1): 36-48.
Kraus, N., T. McGee and C. Cornperatore (1989). MLRs in children are consistently present during
wakefulness, stage 1, and REM sleep. Ear and Hearing 10(6): 339-345.

203
Kraus, N., T. McGee, T. Littman and T. Nicol (1992). Reticular formation influences on primary and
non-primary auditory pathways as reflected by the middle latency response. Brain Research
587(2): 186-194.
Kraus, N., T. McGee, T. Littman, T. Nicol and C. King (1994). Nonprimary auditory thalamic
representation of acoustic change. Journal of Neurophysiology 72(3): 1270-1277.
Kraus, N., D. Smith and T. McGee (1988). Midline and temporal lobe MLRs in the guinea pig
originate from different generator systems: a conceptual framework for new and existing data.
Electroencephalography and Clinical Neurophysiology 70(6): 541-558.
Kronenberger, W. G., B. G. Colson, S. C. Henning and D. B. Pisoni (2014). Executive Functioning
and Speech-Language Skills Following Long-Term Use of Cochlear Implants. Journal of Deaf
Studies and Deaf Education: 1-15.
Lachaux, J. P., E. Rodriguez, J. Martinerie and F. J. Varela (1999). Measuring phase synchrony in
brain signals. Human brain mapping 8(4): 194-208.
Lane, H. and B. Bahan (1998). Article Commentary: Ethics of cochlear implantation in young
children: A review and reply from a Deaf-World perspective. Otolaryngology - Head and Neck
Surgery 119(4): 297-313.
Lang, R., L. K. Koegel, K. Ashbaugh, A. Regester, W. Ence, and W. Smith (2010). Physical exercise
and individuals with autism spectrum disorders: A systematic review. Research in Autism
Spectrum Disorders 4(4): 565-576.
Langers, D. R., P. van Dijk and W. H. Backes (2005). Lateralization, connectivity and plasticity in the
human central auditory system. Neuroimage 28(2): 490-499.
Lavie, N. (2005). Distracted and confused?: selective attention under load. Trends in Cognitive Science
9(2): 75-82.
Lazard, D., A.-L. Giraud, D. Gnansia, B. Meyer and O. Sterkers (2011). Understanding the deafened
brain: Implications for cochlear implant rehabilitation. European Annals of Otorhinolaryngology,
Head and Neck Diseases.

204
Lazard, D. S., H. J. Lee, E. Truy and A. L. Giraud (2012). Bilateral reorganization of posterior
temporal cortices in post‐lingual deafness and its relation to cochlear implant outcome. Human
Brain Mapping.
Le Grand, R., C. J. Mondloch, D. Maurer and H. P. Brent (2003). Expert face processing requires
visual input to the right hemisphere during infancy. Nature Neuroscience 6(10): 1108-1112.
Le Van Quyen, M. and A. Bragin (2007). Analysis of dynamic brain oscillations: methodological
advances. Trends in Neurosciences 30(7): 365-373.
Le Van Quyen, M., J. Foucher, J.-P. Lachaux, E. Rodriguez, A. Lutz, J. Martinerie and F. J. Varela
(2001). Comparison of Hilbert transform and wavelet methods for the analysis of neuronal
synchrony. Journal of Neuroscience Methods 111(2): 83-98.
Le Vay, S., T. N. Wiesel and D. H. Hubel (1980). The development of ocular dominance columns in
normal and visually deprived monkeys. Journal of Comparative Neurology 191(1): 1-51.
Lebel, C. and C. Beaulieu (2011). Longitudinal development of human brain wiring continues from
childhood into adulthood. The Journal of Neuroscience 31(30): 10937-10947.
LeDoux, J. E. (1992). Brain mechanisms of emotion and emotional learning. Current Opinions in
Neurobiology 2(2): 191-197.
Lee, C. C. and J. A. Winer (2005). Principles governing auditory cortex connections. Cerebral Cortex
15(11): 1804-1814.
Lee, D. S., J. S. Lee, S. H. Oh, S. K. Kim, J. W. Kim, J. K. Chung, M. C. Lee and C. S. Kim (2001).
Deafness: cross-modal plasticity and cochlear implants. Nature 409(6817): 149-150.
Lee, H. J., A. L. Giraud, E. Kang, S. H. Oh, H. Kang, C. S. Kim and D. S. Lee (2007). Cortical activity
at rest predicts cochlear implantation outcome. Cerebral Cortex 17(4): 909-917.
Lee, H. J., E. Truy, G. Mamou, D. Sappey-Marinier and A. L. Giraud (2007). Visual speech circuits
in profound acquired deafness: a possible role for latent multimodal connectivity. Brain
130(11): 2929-2941.

205
Lee, K., C. Van Hasselt, S. Chiu and D. Cheung (2002). Cantonese tone perception ability of cochlear
implant children in comparison with normal-hearing children. International Journal of Pediatric
Otorhinolaryngology 63(2): 137-147.
Lee, T.-W., M. Girolami and T. J. Sejnowski (1999). Independent component analysis using an
extended infomax algorithm for mixed subgaussian and supergaussian sources. Neural
Computation 11(2): 417-441.
Lenroot, R. K. and J. N. Giedd (2006). Brain development in children and adolescents: insights from
anatomical magnetic resonance imaging. Neuroscience and Biobehavioral Reviews 30(6): 718-729.
Levänen, S. and D. Hamdorf (2001). Feeling vibrations: enhanced tactile sensitivity in congenitally
deaf humans. Neuroscience Letters 301(1): 75-77.
Levänen, S., V. Jousmäki and R. Hari (1998). Vibration-induced auditory-cortex activation in a
congenitally deaf adult. Current Biology 8(15): 869-872.
Lewis, T. L. and D. Maurer (2005). Multiple sensitive periods in human visual development: evidence
from visually deprived children. Developmental Psychobiology 46(3): 163-183.
Liegeois-Chauvel, C., A. Musolino, J. Badier, P. Marquis and P. Chauvel (1994). Evoked potentials
recorded from the auditory cortex in man: evaluation and topography of the middle latency
components. Electroencephalography and Clinical Neurophysiology/Evoked Potentials Section 92(3):
204-214.
Liegeois-Chauvel, C., A. Musolino and P. Chauvel (1991). Localization of the primary auditory area
in man. Brain 114(1): 139-153.
Lim, D. J. (1980). Cochlear anatomy related to cochlear micromechanics. A review. The Journal of the
Acoustical Society of America 67(5): 1686-1695.
Lim, D. J. (1986). Functional structure of the organ of Corti: a review. Hearing Research 22(1): 117-
146.
Litovsky, R. (2008). Potential Advantages from Bilateral Cochlear Implants. White Paper Series.

206
Litovsky, R. Y., P. M. Johnstone and S. P. Godar (2006). Benefits of bilateral cochlear implants and/or
hearing aids in children. International Journal of Audiology 45(S1): 78-91.
Litovsky, R. Y., A. Parkinson, J. Arcaroli, R. Peters, J. Lake, P. Johnstone and G. Yu (2004). Bilateral
cochlear implants in adults and children. Archives of Otolaryngology - Head and Neck Surgery 130(5):
648-655.
Lohmann, C. and H. W. Kessels (2014). The developmental stages of synaptic plasticity. The Journal
of Physiology 592(1): 13-31.
Loizou, P. C. (1999). Introduction to cochlear implants. Engineering in Medicine and Biology Magazine,
IEEE 18(1): 32-42.
Lomber, S. G., M. A. Meredith and A. Kral (2010). Cross-modal plasticity in specific auditory cortices
underlies visual compensations in the deaf. Nature Neuroscience 13(11): 1421-1427.
Lorenzi, C., S. Gatehouse and C. Lever (1999). Sound localization in noise in normal-hearing listeners.
The Journal of the Acoustical Society of America 105: 1810-1820.
Luntz, M., N. Yehudai and T. Shpak (2007). Hearing progress and fluctuations in bimodal-binaural
hearing users (unilateral cochlear implants and contralateral hearing aid). Acta Otolaryngol
127(10): 1045-1050.
Mackay, I. R., F. S. Rosen and R. M. Zinkernagel (2001). Maternal antibodies, childhood infections,
and autoimmune diseases. New England Journal of Medicine 345(18): 1331-1335.
MacNeilage, P. F., L. J. Rogers and G. Vallortigara (2009). Origins of the left & right brain. Scientific
American 301(1): 60-67.
Mah-rya, L. A. and C. Yoshinaga-Itano (1995). Early identification of infants with significant hearing
loss and the Minnesota Child Development Inventory. Seminars in Hearing, Copyright© 1995
by Thieme Medical Publishers, Inc.
Marosi, E., T. Harmony, A. Reyes, J. Bernal, T. Fernandez, V. Guerrero, M. Rodri'guez, J. Silva, G.
Yáñez and H. Rodri'guez (1997). A follow-up study of EEG coherences in children with
different pedagogical evaluations. International Journal of Psychophysiology 25(3): 227-235.

207
Marosi, E., T. Harmony, L. Sánchez, J. Becker, J. Bernal, A. Reyes, A. E. D. de León, M. Rodríguez
and T. Fernández (1992). Maturation of the coherence of EEG activity in normal and
learning-disabled children. Electroencephalography and Clinical Neurophysiology 83(6): 350-357.
Martin, B. A. (2007). Can the acoustic change complex be recorded in an individual with a cochlear
implant? Separating neural responses from cochlear implant artifact. Journal of the American
Academy of Audiology 18(2): 126-140.
Matsumoto, R., D. R. Nair, E. LaPresto, I. Najm, W. Bingaman, H. Shibasaki and H. O. Luders (2004).
Functional connectivity in the human language system: a cortico-cortical evoked potential
study. Brain 127(Pt 10): 2316-2330.
Mc Laughlin, M., A. Lopez Valdes, R. B. Reilly and F.-G. Zeng (2013). Cochlear implant artifact
attenuation in late auditory evoked potentials: A single channel approach. Hearing Research 302:
84-95.
McGee, T., N. Kraus, C. Comperatore and T. Nicol (1991). Subcortical and cortical components of
the MLR generating system. Brain Research 544(2): 211-220.
McGee, T., N. Kraus, M. Killion, R. Rosenberg and C. King (1993). Improving the reliability of the
auditory middle latency response by monitoring EEG delta activity. Ear and Hearing 14: 76-
76.
McGee, T., N. Kraus, T. Littman and T. Nicol (1992). Contributions of medial geniculate body
subdivisions to the middle latency response. Hearing Research 61(1-2): 147-154.
McMenamin, B. W., A. J. Shackman, J. S. Maxwell, D. R. Bachhuber, A. M. Koppenhaver, L. L.
Greischar and R. J. Davidson (2010). Validation of ICA-based myogenic artifact correction
for scalp and source-localized EEG. Neuroimage 49(3): 2416-2432.
Mehra, S., R. D. Eavey and D. G. Keamy Jr (2009). The epidemiology of hearing impairment in the
United States: newborns, children, and adolescents. Otolaryngology - Head and Neck Surgery
140(4): 461-472.

208
Menon, V., G. Shailesh, P. Sharma and R. Saxena (2008). Clinical trial of patching versus atropine
penalization for the treatment of anisometropic amblyopia in older children. Journal of American
Association for Pediatric Ophthalmology and Strabismus 12(5): 493-497.
Meredith, M. A. and S. G. Lomber (2011). Somatosensory and visual crossmodal plasticity in the
anterior auditory field of early-deaf cats. Hearing Research 280(1-2): 38-47.
Miller, C. A., P. J. Abbas and C. J. Brown (2000). An improved method of reducing stimulus artifact
in the electrically evoked whole-nerve potential. Ear and Hearing 21(4): 280-290.
Moeller, M. P. (2000). Early intervention and language development in children who are deaf and
hard of hearing. Pediatrics 106(3): e43-43.
Mohr, P. E., J. J. Feldman, J. L. Dunbar, A. McConkey-Robbins, J. K. Niparko, R. K. Rittenhouse and
M. W. Skinner (2000). The societal costs of severe to profound hearing loss in the United
States. International Journal of Technology Assessment in Health Care 16(4): 1120-1135.
Moller, A. R. and P. R. Rollins (2002). The non-classical auditory pathways are involved in hearing in
children but not in adults. Neuroscience Letters 319(1): 41-44.
Monastra, V. J., J. F. Lubar and M. Linden (2001). The development of a quantitative
electroencephalographic scanning process for attention deficit–hyperactivity disorder:
Reliability and validity studies. Neuropsychology 15(1): 136-144.
Monastra, V. J., J. F. Lubar, M. Linden, P. VanDeusen, G. Green, W. Wing, A. Phillips and T. N.
Fenger (1999). Assessing attention deficit hyperactivity disorder via quantitative
electroencephalography: an initial validation study. Neuropsychology 13(3): 424-433.
Moore, D. R. (1990). Auditory brainstem of the ferret: bilateral cochlear lesions in infancy do not
affect the number of neurons projecting from the cochlear nucleus to the inferior colliculus.
Developmental Brain Research 54(1): 125-130.
Moore, D. R. (2002). Auditory development and the role of experience. British Medical Bulletin 63(1):
171-181.

209
Moore, D. R., J. T. Devlin, J. Raley, E. Tunbridge, K. Lanary, A. Floyer-Lea, C. Narain, I. Cohen, P.
Jezzard and M. J. Burton (2005). Effects of long term unilateral hearing loss on the
lateralization of fMRI measured activation in human auditory cortex. Plasticity and Signal
Representation in the Auditory System, Springer: 335-346.
Moore, D. R., D. E. Hartley and S. Hogan (2003). Effects of otitis media with effusion (OME) on
central auditory function. International Journal of Pediatric Otorhinolaryngology 67: S63-S67.
Moore, D. R. and L. M. Kitzes (2004). Projections from the cochlear nucleus to the inferior colliculus
in normal and neonatally cochlea‐ablated gerbils. The Journal of Comparative Neurology 240(2):
180-195.
Moore, D. R. and N. E. Kowalchuk (1988). Auditory brainstem of the ferret: Effects of unilateral
cochlear lesions on cochlear nucleus volume and projections to the inferior colliculus. The
Journal of Comparative Neurology 272(4): 503-515.
Moore, D. R. and N. E. Kowalchuk (2004). Auditory brainstem of the ferret: Effects of unilateral
cochlear lesions on cochlear nucleus volume and projections to the inferior colliculus. The
Journal of Comparative Neurology 272(4): 503-515.
Moore, D. R. and R. V. Shannon (2009). Beyond cochlear implants: awakening the deafened brain.
Nature Neuroscience 12(6): 686-691.
Moore, J. K. and Y.-L. Guan (2001). Cytoarchitectural and Axonal Maturation in Human Auditory
Cortex. JARO 2(4): 297-311.
Moore, J. K. and F. H. Linthicum, Jr. (2007). The human auditory system: a timeline of development.
International Journal of Audiology 46(9): 460-478.
Moore, R. and J. Goldberg (1963). Ascending projections of the inferior colliculus in the cat. Journal
of Comparative Neurology 121(1): 109-135.
Moreno, S., C. Marques, A. Santos, M. Santos, S. L. Castro and M. Besson (2009). Musical training
influences linguistic abilities in 8-year-old children: more evidence for brain plasticity. Cerebral
Cortex 19(3): 712-723.

210
Moriguchi, Y., T. Ohnishi, T. Mori, H. Matsuda and G. Komaki (2007). Changes of brain activity in
the neural substrates for theory of mind during childhood and adolescence. Psychiatry and
Clinical Neurosciences 61(4): 355-363.
Morton, C. C. and W. E. Nance (2006). Newborn hearing screening—a silent revolution. New England
Journal of Medicine 354(20): 2151-2164.
Mossop, J., M. Wilson, D. Caspary and D. Moore (2000). Down-regulation of inhibition following
unilateral deafening. Hearing Research 147(1): 183-187.
Müller, V., W. Gruber, W. Klimesch and U. Lindenberger (2009). Lifespan differences in cortical
dynamics of auditory perception. Developmental Science 12(6): 839-853.
Munte, T. F., E. Altenmuller and L. Jancke (2002). The musician's brain as a model of neuroplasticity.
Nature Reviews Neuroscience 3(6): 473-477.
Murray, D. K., M. A. Sacheli, J. J. Eng and A. J. Stoessl (2014). The effects of exercise on cognition
in Parkinson’s disease: a systematic review. Translational Neurodegeneration 3(1): 5.
Nadol, J. B. (1997). Patterns of neural degeneration in the human cochlea and auditory nerve:
implications for cochlear implantation. Otolaryngology - Head and Neck Surgery 117(3): 220-228.
Nadol Jr, J. and W. Xu (1992). Diameter of the cochlear nerve in deaf humans: implications for
cochlear implantation. The Annals of Otology, Rhinology, and Laryngology 101(12): 988-993.
Nadol Jr, J., Y. Young and R. Glynn (1989). Survival of spiral ganglion cells in profound sensorineural
hearing loss: implications for cochlear implantation. The Annals of Otology, Rhinology, and
Laryngology 98(6): 411-416.
Nadol Jr, J. B. (1988). Comparative anatomy of the cochlea and auditory nerve in mammals. Hearing
Research 34(3): 253-266.
Naik, G. R. and D. K. Kumar (2011). An overview of independent component analysis and its
applications. Informatica: An International Journal of Computing and Informatics 35(1): 63-81.
Netter, F. H. (2010). Atlas of human anatomy, Elsevier Health Sciences.

211
Nicholas, J. G. and A. E. Geers (2007). Will they catch up? The role of age at cochlear implantation
in the spoken language development of children with severe to profound hearing loss. Journal
of Speech, Language, and Hearing Research 50(4): 1048-1062.
Nikolopoulos, T. P., G. M. O'Donoghue and S. Archbold (2009). Age at implantation: its importance
in pediatric cochlear implantation. The Laryngoscope 109(4): 595-599.
Nishimura, H., K. Hashikawa, K. Doi, T. Iwaki, Y. Watanabe, H. Kusuoka, T. Nishimura and T. Kubo
(1999). Sign language ‘heard’in the auditory cortex. Nature 397(6715): 116-116.
Nishiyama, N., N. Hardie and R. Shepherd (2000). Neonatal sensorineural hearing loss affects
neurone size in cat auditory midbrain. Hearing Research 140(1): 18-22.
Nordeen, K., H. Killackey and L. Kitzes (1983). Ascending auditory projections to the inferior
colliculus in the adult gerbil, Meriones unguiculatus. Journal of Comparative Neurology 214(2):
144-153.
Nordeen, K., H. Killackey and L. Kitzes (1983). Ascending projections to the inferior colliculus
following unilateral cochlear ablation in the neonatal gerbil, Meriones unguiculatus. Journal of
Comparative Neurology 214(2): 144-153.
Nordeen, K., H. Killackey and L. Kitzes (2004). Ascending projections to the inferior colliculus
following unilateral cochlear ablation in the neonatal gerbil, Meriones unguiculatus. The Journal
of comparative neurology 214(2): 144-153.
Novak, G., W. Ritter and H. G. Vaughan Jr (1992). Mismatch detection and the latency of temporal
judgments. Psychophysiology 29(4): 398-411.
Nunez, P. L. (2000). Toward a quantitative description of large-scale neocortical dynamic function
and EEG. Behavioral and Brain Sciences 23(3): 371-398.
Nunez, P. L., R. Srinivasan, A. F. Westdorp, R. S. Wijesinghe, D. M. Tucker, R. B. Silberstein and P.
J. Cadusch (1997). EEG coherency: I: statistics, reference electrode, volume conduction,
Laplacians, cortical imaging, and interpretation at multiple scales. Electroencephalography and
Clinical Neurophysiology 103(5): 499-515.

212
O'Neil, J. N., C. J. Limb, C. A. Baker and D. K. Ryugo (2010). Bilateral effects of unilateral cochlear
implantation in congenitally deaf cats. The Journal of Comparative Neurology 518(12): 2382-2404.
O'Neill, C., G. M. O'Donoghue, S. M. Archbold and C. Normand (2000). A Cost‐Utility Analysis of
Pediatric Cochlear Implantation. The Laryngoscope 110(1): 156-160.
O’Connor, K. (2012). Auditory processing in autism spectrum disorder: a review. Neuroscience and
Biobehavioral Reviews 36(2): 836-854.
O’Donoghue, G. M. (1999). Hearing without ears: do cochlear implants work in children?: Yes, so
long as they are given to the right children early enough. BMJ 318(7176): 72.
Oatman, L. C. (1976). Effects of visual attention on the intensity of auditory evoked potentials.
Experimental Neurology 51(1): 41-53.
Obleser, J., R. J. Wise, M. A. Dresner and S. K. Scott (2007). Functional integration across brain
regions improves speech perception under adverse listening conditions. The Journal of
Neuroscience 27(9): 2283-2289.
Okamoto, H., H. Stracke, C. H. Wolters, F. Schmael and C. Pantev (2007). Attention improves
population-level frequency tuning in human auditory cortex. The Journal of Neuroscience 27(39):
10383-10390.
Openshaw, P. J., Y. Yamaguchi and J. S. Tregoning (2004). Childhood infections, the developing
immune system, and the origins of asthma. Journal of Allergy and Clinical Immunology 114(6):
1275-1277.
Ornitz, E. M., E. R. Ritvo, E. M. Carr, S. L. Franchi and R. D. Walter (1967). The effect of sleep
onset on the auditory averaged evoked response. Electroencephalography and Clinical
Neurophysiology 23(4): 335-341.
Osberger, M. J., S. Zimmerman-Phillips and D. B. Koch (2002). Cochlear implant candidacy and
performance trends in children. The Annals of Otology, Rhinology & Laryngology. 189(5): S62-65.
Pals, C., A. Sarampalis and D. Başkent (2013). Listening effort with cochlear implant simulations.
Journal of Speech, Language, and Hearing Research 56(4): 1075-1084.

213
Palva, J. M., S. Monto, S. Kulashekhar and S. Palva (2010). Neuronal synchrony reveals working
memory networks and predicts individual memory capacity. Proceedings of the National Academy
of Science of the United States of America 107(16): 7580-7585.
Palva, S. and J. M. Palva (2007). New vistas for α-frequency band oscillations. Trends in Neurosciences
30(4): 150-158.
Palva, S. and J. M. Palva (2011). Functional roles of alpha-band phase synchronization in local and
large-scale cortical networks. Frontiers in Psychology 2(204): 1-15.
Palva, S. and J. M. Palva (2012). Discovering oscillatory interaction networks with M/EEG:
challenges and breakthroughs. Trends in Cognitive Sciences 16(4): 219-230.
Pan, C.-Y. (2011). The efficacy of an aquatic program on physical fitness and aquatic skills in children
with and without autism spectrum disorders. Research in Autism Spectrum Disorders 5(1): 657-
665.
Pandya, D. N. and E. H. Yeterian (1985). Architecture and connections of cortical association areas.
Cerebral Cortex 4: 3-61.
Papsin, B. C. and K. A. Gordon (2007). Cochlear implants for children with severe-to-profound
hearing loss. New England Journal of Medicine 357(23): 2380-2387.
Papsin, B. C. and K. A. Gordon (2008). Bilateral cochlear implants should be the standard for children
with bilateral sensorineural deafness. Current Opinions in Otolaryngology - Head Neck Surgery 16(1):
69-74.
Paus, T. (2005). Mapping brain maturation and cognitive development during adolescence. Trends in
Cognitive Sciences 9(2): 60-68.
Penhune, V., R. Zatorre, J. MacDonald and A. Evans (1996). Interhemispheric anatomical differences
in human primary auditory cortex: probabilistic mapping and volume measurement from
magnetic resonance scans. Cerebral Cortex 6(5): 661-672.
Perrin, F., J. Pernier, O. Bertrand and J. Echallier (1989). Spherical splines for scalp potential and
current density mapping. Electroencephalography and Clinical Neurophysiology 72(2): 184-187.

214
Peters, B. R., R. Litovsky, A. Parkinson and J. Lake (2007). Importance of age and postimplantation
experience on speech perception measures in children with sequential bilateral cochlear
implants. Otology & Neurotology 28(5): 649-657.
Petersson, K. M., T. E. Nichols, J.-B. Poline and A. P. Holmes (1999). Statistical limitations in
functional neuroimaging II. Signal detection and statistical inference. Philosophical Transactions
of the Royal Society of London. Series B: Biological Sciences 354(1387): 1261-1281.
Petit, D., J. Montplaisir, D. Lorrain and S. Gauthier (1992). Spectral analysis of the rapid eye
movement sleep electroencephalogram in right and left temporal regions: a biological marker
of Alzheimer's disease. Annals of Neurology 32(2): 172-176.
Pichora-Fuller, M. K. (2003). Cognitive aging and auditory information processing. International Journal
of Audiology 42(2): S26-S32.
Pichora-Fuller, M. K. and G. Singh (2006). Effects of age on auditory and cognitive processing:
implications for hearing aid fitting and audiologic rehabilitation. Trends in Amplification 10(1):
29-59.
Picton, T. and S. Hillyard (1974). Human auditory evoked potentials. II: Effects of attention.
Electroencephalography and Clinical Neurophysiology 36: 191-200.
Picton, T. W. and M. J. Taylor (2007). Electrophysiological evaluation of human brain development.
Developmental Neuropsychology 31(3): 249-278.
Pisoni, D., W. Kronenberger, A. Roman and A. Geers (2011). Measures of digit span and verbal
rehearsal speed in deaf children following more than 10 years of cochlear implantation. Ear
and Hearing 32(S1): 60-70.
Pisoni, D. B., C. M. Conway, W. Kronenberger, S. Henning and E. Anaya (2010). Executive Function,
Cognitive Control, and Sequence Learning in Deaf Children with Cochlear Implants. The
Oxford Handbook of Deaf Studies, Language, and Education.
Plailly, J., J. D. Howard, D. R. Gitelman and J. A. Gottfried (2008). Attention to odor modulates
thalamocortical connectivity in the human brain. The Journal of Neuroscience 28(20): 5257-5267.

215
Ponton, C., J. J. Eggermont, D. Khosla, B. Kwong and M. Don (2002). Maturation of human central
auditory system activity: separating auditory evoked potentials by dipole source modeling.
Clinical Neurophysiology 113(3): 407-420.
Ponton, C. W., M. Don, J. J. Eggermont, M. D. Waring and A. Masuda (1996). Maturation of human
cortical auditory function: differences between normal-hearing children and children with
cochlear implants. Ear and Hearing 17(5): 430.
Ponton, C. W. and J. J. Eggermont (2001). Of kittens and kids: altered cortical maturation following
profound deafness and cochlear implant use. Audiology and Neurotology 6(6): 363-380.
Ponton, C. W., J. J. Eggermont, B. Kwong and M. Don (2000). Maturation of human central auditory
system activity: evidence from multi-channel evoked potentials. Clinical Neurophysiology 111(2):
220-236.
Ponton, C. W., J.-P. Vasama, K. Tremblay, D. Khosla, B. Kwong and M. Don (2001). Plasticity in
the adult human central auditory system: evidence from late-onset profound unilateral
deafness. Hearing Research 154(1): 32-44.
Popescu, M. V. and D. B. Polley (2010). Monaural deprivation disrupts development of binaural
selectivity in auditory midbrain and cortex. Neuron 65(5): 718-731.
Portnoy, W. M. and K. Mattucci (1991). Cochlear implants as a contraindication to magnetic
resonance imaging. The Annals of Otology, Rhinology, and Laryngology 100(3): 195-197.
Posner, M. I. and S. Dehaene (1994). Attentional networks. Trends in Neuroscience 17(2): 75-79.
Propst, E. J., B. C. Papsin, T. L. Stockley, R. V. Harrison and K. A. Gordon (2006). Auditory
responses in cochlear implant users with and without GJB2 deafness. The Laryngoscope 116(2):
317-327.
Pugh, K. R., B. A. Shaywitz, S. E. Shaywitz, R. K. Fulbright, D. Byrd, P. Skudlarski, D. P. Shankweiler,
L. Katz, R. T. Constable and J. Fletcher (1996). Auditory selective attention: an fMRI
investigation. Neuroimage 4(3): 159-173.

216
Pujol, R. and D. Hilding (1973). Anatomy and physiology of the onset of auditory function. Acta
Oto-Laryngologica 76(1-6): 1-10.
Pujol, R., M. Lavigne-rebillard and A. Uziel (1991). Development of the human cochlea. Acta Oto-
Laryngologica 111(S482): 7-13.
Punch, R. and M. Hyde (2011). Social participation of children and adolescents with cochlear
implants: A qualitative analysis of parent, teacher, and child interviews. Journal of Deaf Studies
and Deaf Education 16(4): 474-493.
Punch, R. and M. B. Hyde (2011). Communication, psychosocial, and educational outcomes of
children with cochlear implants and challenges remaining for professionals and parents.
International Journal of Otolaryngology: 1-11.
Qi, S. and R. E. Mitchell (2012). Large-scale academic achievement testing of deaf and hard-of-hearing
students: Past, present, and future. Journal of Deaf Studies and Deaf Education 17(1): 1-18.
Rauschecker, J. P. (1999). Auditory cortical plasticity: a comparison with other sensory systems. Trends
in Neurosciences 22(2): 74-80.
Rauschecker, J. P. and B. Tian (2004). Processing of band-passed noise in the lateral auditory belt
cortex of the rhesus monkey. Journal of Neurophysiology 91(6): 2578-2589.
Razak, K. A., T. Zumsteg and Z. M. Fuzessery (2009). Development of auditory thalamocortical
connections in the pallid bat, Antrozous pallidus. Journal of Computational Neurology 515(2): 231-
242.
Read, H. L., J. A. Winer and C. E. Schreiner (2002). Functional architecture of auditory cortex. Current
Opinion in Neurobiology 12(4): 433-440.
Reale, R. A., J. F. Brugge and J. C. Chan (1987). Maps of auditory cortex in cats reared after unilateral
cochlear ablation in the neonatal period. Developmental Brain Research 34(2): 281-290.
Repka, M., R. Beck, J. Holmes, E. Birch, D. Chandler, S. Cotter, R. Hertle, R. Kraker, P. Moke and G.
Quinn (2003). A randomized trial of patching regimens for treatment of moderate amblyopia
in children. Archives of Ophthalmology 121(5): 603-611.

217
Rif, J., R. Hari, M. S. Hämäläinen and M. Sams (1991). Auditory attention affects two different areas
in the human supratemporal cortex. Electroencephalography and Clinical Neurophysiology 79(6): 464-
472.
Rinne, T. (2010). Activations of human auditory cortex during visual and auditory selective attention
tasks with varying difficulty. The Open Neuroimaging Journal 4: 187-193.
Rivera, S., A. Reiss, M. Eckert and V. Menon (2005). Developmental changes in mental arithmetic:
evidence for increased functional specialization in the left inferior parietal cortex. Cerebral
Cortex 15(11): 1779-1790.
Rodriguez, E., N. George, J.-P. Lachaux, J. Martinerie, B. Renault and F. J. Varela (1999). Perception's
shadow: long-distance synchronization of human brain activity. Nature 397(6718): 430-433.
Rogers, L. J. (2000). Evolution of hemispheric specialization: advantages and disadvantages. Brain
and Language 73(2): 236-253.
Ross, L. A., D. Saint-Amour, V. M. Leavitt, D. C. Javitt and J. J. Foxe (2007). Do you see what I am
saying? Exploring visual enhancement of speech comprehension in noisy environments.
Cerebral Cortex 17(5): 1147-1153.
Rubinstein, J. T. (2004). How cochlear implants encode speech. Current Opinion in Otolaryngology &
Head and Neck Surgery 12(5): 444-448.
Rubinstein, J. T. and R. Hong (2003). Signal coding in cochlear implants: exploiting stochastic effects
of electrical stimulation. Annals of Otology Rhinology and Laryngology 191: 14-19.
Ryugo, D., B. Rosenbaum, P. Kim, J. Niparko and A. Saada (1998). Single unit recordings in the
auditory nerve of congenitally deaf white cats: morphological correlates in the cochlea and
cochlear nucleus. The Journal of Comparative Neurology 397(4): 532-548.
Ryugo, D. K., H. B. Cahill, L. S. Rose, B. T. Rosenbaum, M. E. Schroeder and A. L. Wright (2003).
Separate forms of pathology in the cochlea of congenitally deaf white cats. Hearing Research
181(1): 73-84.

218
Salamy, A. (1984). Maturation of the auditory brainstem response from birth through early childhood.
Journal of Clinical Neurophysiology 1(3): 293-329.
Salamy, A. and C. M. McKean (1976). Postnatal development of human brainstem potentials during
the first year of life. Electroencephalography & Clinical Neurophysiology; Electroencephalography &
Clinical Neurophysiology 40(4): 418-426.
Salloum, C. A. M., J. Valero, D. D. E. Wong, B. C. Papsin, R. van Hoesel and K. A. Gordon (2010).
Lateralization of interimplant timing and level differences in children who use bilateral cochlear
implants. Ear Hear 31(4): 441-456.
Sandmann, P. (2012). User Research Visual processing in the auditory cortex of cochlear-implant
users. Brain Products Press Release 42.
Sandmann, P., N. Dillier, T. Eichele, M. Meyer, A. Kegel, R. D. Pascual-Marqui, V. L. Marcar, L.
Jäncke and S. Debener (2012). Visual activation of auditory cortex reflects maladaptive
plasticity in cochlear implant users. Brain 135(2): 555-568.
Sandmann, P., T. Eichele, M. Buechler, S. Debener, L. Jäncke, N. Dillier, K. Hugdahl and M. Meyer
(2009). Evaluation of evoked potentials to dyadic tones after cochlear implantation. Brain
132(7): 1967-1979.
Sanes, D. H. and V. C. Kotak (2011). Developmental plasticity of auditory cortical inhibitory synapses.
Hearing Research 279(1): 140-148.
Sanes, D. H. and C. Takács (1993). Activity‐dependent Refinement of Inhibitory Connections.
European Journal of Neuroscience 5(6): 570-574.
Scheffler, K., D. Bilecen, N. Schmid, K. Tschopp and J. Seelig (1998). Auditory cortical responses in
hearing subjects and unilateral deaf patients as detected by functional magnetic resonance
imaging. Cerebral Cortex 8(2): 156-163.
Schnitzler, A. and J. Gross (2005). Normal and pathological oscillatory communication in the brain.
Nature Reviews Neuroscience 6(4): 285-296.

219
Schönwiesner, M., K. Krumbholz, R. Rübsamen, G. R. Fink and D. Y. Von Cramon (2007).
Hemispheric asymmetry for auditory processing in the human auditory brain stem, thalamus,
and cortex. Cerebral Cortex 17(2): 492-499.
Schönwiesner, M., R. Rübsamen and D. Y. Von Cramon (2005). Hemispheric asymmetry for spectral
and temporal processing in the human antero‐lateral auditory belt cortex. European Journal of
Neuroscience 22(6): 1521-1528.
Seeber, B. U. and H. Fastl (2008). Localization cues with bilateral cochlear implants. The Journal of the
Acoustical Society of America 123: 1030.
Shargorodsky, J., S. G. Curhan, G. C. Curhan and R. Eavey (2010). Change in prevalence of hearing
loss in US adolescents. JAMA 304(7): 772-778.
Sharma, A., J. Campbell and G. Cardon (2014). Developmental and cross-modal plasticity in deafness:
Evidence from the P1 and N1 event related potentials in cochlear implanted children.
International Journal of Psychophysiology (In Press).
Sharma, A. and M. Dorman (2006). Central auditory development in children with cochlear implants:
clinical implications. Advances in Oto Rhino Laryngology 64: 66-88.
Sharma, A., M. F. Dorman and A. Kral (2005). The influence of a sensitive period on central auditory
development in children with unilateral and bilateral cochlear implants. Hearing Research 203(1):
134-143.
Sharma, A., M. F. Dorman and A. J. Spahr (2002). Rapid development of cortical auditory evoked
potentials after early cochlear implantation. Neuroreport 13(10): 1365.
Sharma, A., M. F. Dorman and A. J. Spahr (2002). A sensitive period for the development of the
central auditory system in children with cochlear implants: implications for age of implantation.
Ear and Hearing 23(6): 532-539.
Sharma, A., P. M. Gilley, M. F. Dorman and R. Baldwin (2007). Deprivation-induced cortical
reorganization in children with cochlear implants. International Journal of Audiology 46(9): 494-
499.

220
Sharma, A., N. Kraus, T. J McGee and T. G. Nicol (1997). Developmental changes in P1 and N1
central auditory responses elicited by consonant-vowel syllables. Electroencephalography and
Clinical Neurophysiology/Evoked Potentials Section 104(6): 540-545.
Sharma, A., A. A. Nash and M. Dorman (2009). Cortical development, plasticity and re-organization
in children with cochlear implants. Journal of Communication Disorders 42(4): 272-279.
Shibata, D. (2007). Differences in brain structure in deaf persons on MR imaging studied with voxel-
based morphometry. American Journal of Neuroradiology 28(2): 243-249.
Shinn-Cunningham, B. G. and V. Best (2008). Selective attention in normal and impaired hearing.
Trends in Amplification 12(4): 283-299.
Shomstein, S. and S. Yantis (2004). Control of attention shifts between vision and audition in human
cortex. The Journal of Neuroscience 24(47): 10702-10706.
Singer, W. (1999). Neuronal synchrony: a versatile code for the definition of relations? Neuron 24(1):
49-65.
Skinner, M. W., L. K. Holden, L. A. Whitford, K. L. Plant, C. Psarros and T. A. Holden (2002).
Speech recognition with the nucleus 24 SPEAK, ACE, and CIS speech coding strategies in
newly implanted adults. Ear and Hearing 23(3): 207-223.
Smith, L. B., A. L. Quittner, M. J. Osberger and R. Miyamoto (1998). Audition and visual attention:
the developmental trajectory in deaf and hearing populations. Developmental Psychology 34(5):
840-850.
Smith, R. J., J. F. Bale Jr and K. R. White (2005). Sensorineural hearing loss in children. The Lancet
365(9462): 879-890.
Sokolov, A., M. Pavlova, W. Lutzenberger and N. Birbaumer (2004). Reciprocal modulation of
neuromagnetic induced gamma activity by attention in the human visual and auditory cortex.
Neuroimage 22(2): 521-529.
Song, S., K. D. Miller and L. F. Abbott (2000). Competitive Hebbian learning through spike-timing-
dependent synaptic plasticity. Nature Neuroscience 3: 919-926.

221
Sowa, M. and R. Meulenbroek (2012). Effects of physical exercise on autism spectrum disorders: a
meta-analysis. Research in Autism Spectrum Disorders 6(1): 46-57.
Sowell, E. R., P. M. Thompson, K. D. Tessner and A. W. Toga (2001). Mapping continued brain
growth and gray matter density reduction in dorsal frontal cortex: inverse relationships during
postadolescent brain maturation. The Journal of Neuroscience 21(22): 8819-8829.
Starr, A., R. Amlie, W. Martin and S. Sanders (1977). Development of auditory function in newborn
infants revealed by auditory brainstem potentials. Pediatrics 60(6): 831-839.
Steel, M. M., B. C. Papsin and K. A. Gordon (Submitted). Binaural fusion and listening effort in
children who use bilateral cochlear implants: A psychoacoustic and pupillometric study. PloS
one.
Steele, C. J., J. A. Bailey, R. J. Zatorre and V. B. Penhune (2013). Early musical training and white-
matter plasticity in the corpus callosum: evidence for a sensitive period. The Journal of
Neuroscience 33(3): 1282-1290.
Steffens, T., A. Lesinski-Schiedat, J. Strutz, A. Aschendorff, T. Klenzner, S. Rühl, B. Voss, T. Wesarg,
R. Laszig and T. Lenarz (2008). The benefits of sequential bilateral cochlear implantation for
hearing-impaired children. Acta Oto-laryngologica 128(2): 164-176.
Stoffers, D., J. Bosboom, J. Deijen, E. C. Wolters, H. Berendse and C. Stam (2007). Slowing of
oscillatory brain activity is a stable characteristic of Parkinson's disease without dementia. Brain
130(7): 1847-1860.
Strait, D. L. and N. Kraus (2011). Can you hear me now? Musical training shapes functional brain
networks for selective auditory attention and hearing speech in noise. Frontiers in Psychology
2(113): 1-10.
Strait, D. L., N. Kraus, A. Parbery-Clark and R. Ashley (2010). Musical experience shapes top-down
auditory mechanisms: evidence from masking and auditory attention performance. Hearing
Research 261(1): 22-29.

222
Strait, D. L., N. Kraus, E. Skoe, and R. Ashley (2009). Musical experience and neural efficiency–
effects of training on subcortical processing of vocal expressions of emotion. European Journal
of Neuroscience 29(3): 661-668.
Strait, D. L., A. Parbery-Clark, E. Hittner and N. Kraus (2012). Musical training during early
childhood enhances the neural encoding of speech in noise. Brain and Language 123(3): 191-
201.
Strelnikov, K., J. Rouger, J.-F. Demonet, S. Lagleyre, B. Fraysse, O. Deguine and P. Barone (2010).
Does brain activity at rest reflect adaptive strategies? Evidence from speech processing after
cochlear implantation. Cerebral Cortex 20(5): 1217-1222.
Svirsky, M. A., A. M. Robbins, K. I. Kirk, D. B. Pisoni and R. T. Miyamoto (2000). Language
development in profoundly deaf children with cochlear implants. Psychological Science 11(2):
153-158.
Takesian, A. E., V. C. Kotak and D. H. Sanes (2009). Developmental hearing loss disrupts synaptic
inhibition: implications for auditory processing. Future neurology 4(3): 331-349.
Teissl, C., C. Kremser, E. S. Hochmair and I. J. Hochmair-Desoyer (1998). Cochlear implants: in
vitro investigation of electromagnetic interference at MR imaging - compatibility and safety
aspects. Radiology 208(3): 700-708.
Teissl, C., C. Kremser, E. S. Hochmair and I. J. Hochmair-Desoyer (1999). Magnetic resonance
imaging and cochlear implants: compatibility and safety aspects. Journal of Magnetic Resonance
Imaging 9(1): 26-38.
Tervaniemi, M. and K. Hugdahl (2003). Lateralization of auditory-cortex functions. Brain Research
Reviews 43(3): 231-246.
Tessier, C. R. and K. Broadie (2009). Activity-dependent modulation of neural circuit synaptic
connectivity. Frontiers in Molecular Neuroscience 2(8): 1-13.
Thai-Van, H., S. Cozma, F. Boutitie, F. Disant, E. Truy and L. Collet (2007). The pattern of auditory
brainstem response wave V maturation in cochlear-implanted children. Clinical Neurophysiology
118(3): 676-689.

223
Tharpe, A. M. (2008). Unilateral and mild bilateral hearing loss in children: past and current
perspectives. Trends in Amplification 12(1): 7-15.
Toga, A. W. and P. M. Thompson (2003). Mapping brain asymmetry. Nature Reviews Neuroscience 4(1):
37-48.
Trainor, L. J., A. Shahin and L. E. Roberts (2003). Effects of musical training on the auditory cortex
in children. Annals of the New York Academy of Sciences 999(1): 506-513.
Tremblay, K., N. Kraus, T. McGee, C. Ponton and B. Otis (2001). Central auditory plasticity: changes
in the N1-P2 complex after speech-sound training. Ear and Hearing 22(2): 79-90.
Tremblay, K. L. (2007). Training-related changes in the brain: evidence from human auditory-evoked
potentials. Seminars in Hearing, Thieme Medical Publishers Inc.
Tremblay, K. L. and N. Kraus (2002). Auditory training induces asymmetrical changes in cortical
neural activity. Journal of Speech, Language and Hearing Research 45(3): 564-572.
Tremblay, K. L., A. J. Shahin, T. Picton and B. Ross (2009). Auditory training alters the physiological
detection of stimulus-specific cues in humans. Clinical Neurophysiology: Official Journal of the
International Federation of Clinical Neurophysiology 120(1): 128-135.
Tritsch, N. X., E. Yi, J. E. Gale, E. Glowatzki and D. E. Bergles (2007). The origin of spontaneous
activity in the developing auditory system. Nature 450(7166): 50-55.
Turner, C. W., B. J. Gantz, C. Vidal, A. Behrens and B. A. Henry (2004). Speech recognition in noise
for cochlear implant listeners: benefits of residual acoustic hearing. The Journal of the Acoustical
Society of America 115(4): 1729-1735.
Turrigiano, G. G. and S. B. Nelson (2004). Homeostatic plasticity in the developing nervous system.
Nature Reviews Neuroscience 5(2): 97-107.
Uhlhaas, P., G. Pipa, B. Lima, L. Melloni, S. Neuenschwander, D. Nikolić and W. Singer (2009).
Neural synchrony in cortical networks: history, concept and current status. Frontiers in Integrative
Neuroscience 3(17): 1-19.

224
Uhlhaas, P. J., F. Roux, E. Rodriguez, A. Rotarska-Jagiela and W. Singer (2010). Neural synchrony
and the development of cortical networks. Trends in Cognitive Sciences 14(2): 72-80.
Uhlhaas, P. J., F. Roux, W. Singer, C. Haenschel, R. Sireteanu and E. Rodriguez (2009). The
development of neural synchrony reflects late maturation and restructuring of functional
networks in humans. Proceedings of the National Academy of Sciences 106(24): 9866-9871.
Uhlhaas, P. J. and W. Singer (2006). Neural synchrony in brain disorders: relevance for cognitive
dysfunctions and pathophysiology. Neuron 52(1): 155-168.
Uhlhaas, P. J. and W. Singer (2010). Abnormal neural oscillations and synchrony in schizophrenia.
Nature Reviews Neuroscience 11(2): 100-113.
Uhlhaas, P. J. and W. Singer (2011). The development of neural synchrony and large-scale cortical
networks during adolescence: relevance for the pathophysiology of schizophrenia and
neurodevelopmental hypothesis. Schizophrenia Bulletin 37(3): 514-523.
Vaerenberg, B., C. Smits, G. De Ceulaer, E. Zir, S. Harman, N. Jaspers, Y. Tam, M. Dillon, T. Wesarg
and D. Martin-Bonniot (2014). Cochlear implant programming: a global survey on the state
of the art. The Scientific World Journal.
Vale, C., J. M. Juíz, D. R. Moore and D. H. Sanes (2004). Unilateral cochlear ablation produces greater
loss of inhibition in the contralateral inferior colliculus. European Journal of Neuroscience 20(8):
2133-2140.
Vallortigara, G. (2006). The evolutionary psychology of left and right: costs and benefits of
lateralization. Developmental Psychobiology 48(6): 418-427.
van den Broek, S. P., F. Reinders, M. Donderwinkel and M. Peters (1998). Volume conduction effects
in EEG and MEG. Electroencephalography and Clinical Neurophysiology 106(6): 522-534.
Van Deun, L., A. Van Wieringen, F. Scherf, N. Deggouj, C. Desloovere, F. E. Offeciers, P. H. Van de
Heyning, I. J. Dhooge and J. Wouters (2009). Earlier intervention leads to better sound
localization in children with bilateral cochlear implants. Audiology and Neurotology 15(1): 7-17.

225
Van Deun, L., A. Van Wieringen, T. Van den Bogaert, F. Scherf, F. E. Offeciers, P. H. Van de Heyning,
C. Desloovere, I. J. Dhooge, N. Deggouj and L. De Raeve (2009). Sound localization, sound
lateralization, and binaural masking level differences in young children with normal hearing.
Ear and Hearing 30(2): 178-190.
van Hoesel, R. J. and R. S. Tyler (2003). Speech perception, localization, and lateralization with
bilateral cochlear implants. The Journal of the Acoustical Society of America 113: 1617.
Van Veen, B. D., W. Van Drongelen, M. Yuchtman and A. Suzuki (1997). Localization of brain
electrical activity via linearly constrained minimum variance spatial filtering. Biomedical
Engineering, IEEE Transactions on 44(9): 867-880.
Van Wanrooij, M. M. and A. J. Van Opstal (2004). Contribution of head shadow and pinna cues to
chronic monaural sound localization. The Journal of neuroscience 24(17): 4163-4171.
van Zundert, B., A. Yoshii and M. Constantine-Paton (2004). Receptor compartmentalization and
trafficking at glutamate synapses: a developmental proposal. Trends in Neurosciences 27(7): 428-
437.
Varela, F., J.-P. Lachaux, E. Rodriguez and J. Martinerie (2001). The brainweb: phase synchronization
and large-scale integration. Nature Reviews Neuroscience 2(4): 229-239.
Vasama, J.-P. and J. P. Mäkelä (1995). Auditory pathway plasticity in adult humans after unilateral
idiopathic sudden sensorineural hearing loss. Hearing Research 87(1): 132-140.
Vasama, J.-P. and J. P. Mäkelä (1997). Auditory cortical responses in humans with profound unilateral
sensorineural hearing loss from early childhood. Hearing Research 104(1): 183-190.
Vasama, J.-P., J. P. Mäkelä, L. Parkkonen and R. Hari (1994). Auditory cortical responses in humans
with congenital unilateral conductive hearing loss. Hearing Research 78(1): 91-97.
Vasama, J.-P., J. P. Mäkelä, I. Pyykkö and R. Hari (1995). Abrupt unilateral deafness modifies function
of human auditory pathways. Neuroreport 6(7): 961-964.

226
Velazquez, J. L. P., R. G. Erra, R. Wennberg and L. G. Dominguez (2009). Correlations of cellular
activities in the nervous system: physiological and methodological considerations. Coordinated
Activity in the Brain, Springer: 1-24.
Vidal, C. N., R. Nicolson, T. J. DeVito, K. M. Hayashi, J. A. Geaga, D. J. Drost, P. C. Williamson, N.
Rajakumar, Y. Sui and R. A. Dutton (2006). Mapping corpus callosum deficits in autism: an
index of aberrant cortical connectivity. Biological Psychiatry 60(3): 218-225.
Viola, F. C., M. De Vos, J. Hine, P. Sandmann, S. Bleeck, J. Eyles and S. Debener (2012). Semi-
automatic attenuation of cochlear implant artifacts for the evaluation of late auditory evoked
potentials. Hearing Research 284(1): 6-15.
Viola, F. C., J. D. Thorne, S. Bleeck, J. Eyles and S. Debener (2011). Uncovering auditory evoked
potentials from cochlear implant users with independent component analysis. Psychophysiology
48(11): 1470-1480.
Von Békésy, G. and E. G. Wever (1960). Experiments in hearing, McGraw-Hill New York.
Von Stein, A. and J. Sarnthein (2000). Different frequencies for different scales of cortical integration:
from local gamma to long range alpha/theta synchronization. International Journal of
Psychophysiology 38(3): 301-313.
Vrba, J. and S. E. Robinson (2001). Signal processing in magnetoencephalography. Methods 25(2):
249-271.
Ward, L. M. (2003). Synchronous neural oscillations and cognitive processes. Trends in Cognitive Sciences
7(12): 553-559.
Webster, K. E. and I. M. Colrain (2000). The respiratory-related evoked potential: effects of attention
and occlusion duration. Psychophysiology 37(03): 310-318.
Wei, C.-G., K. Cao and F.-G. Zeng (2004). Mandarin tone recognition in cochlear-implant subjects.
Hearing Research 197(1): 87-95.
Weitzman, E. D. and L. J. Graziani (1968). Maturation and topography of the auditory evoked
response of the prematurely born infant. Developmental Psychobiology 1(2): 79-89.

227
Weitzman, E. D. and H. Kremen (1965). Auditory evoked responses during different stages of sleep
in man. Electroencephalography and Clinical Neurophysiology 18(1): 65-70.
Werkle-Bergner, M., Y. L. Shing, V. Müller, S.-C. Li and U. Lindenberger (2009). EEG gamma-band
synchronization in visual coding from childhood to old age: evidence from evoked power and
inter-trial phase locking. Clinical Neurophysiology 120(7): 1291-1302.
Williams, C., K. Northstone, R. Harrad, J. Sparrow and I. Harvey (2003). Amblyopia treatment
outcomes after preschool screening v school entry screening: observational data from a
prospective cohort study. British Journal of Ophthalmology 87(8): 988-993.
Wilson, S. M., A. P. Saygin, M. I. Sereno and M. Iacoboni (2004). Listening to speech activates motor
areas involved in speech production. Nature Neuroscience 7(7): 701-702.
Wilson, T. W., D. C. Rojas, M. L. Reite, P. D. Teale and S. J. Rogers (2007). Children and adolescents
with autism exhibit reduced MEG steady-state gamma responses. Biological Psychiatry 62(3):
192-197.
Winer, J. A., J. J. Diehl and D. T. Larue (2001). Projections of auditory cortex to the medial geniculate
body of the cat. Journal of Comparative Neurology 430(1): 27-55.
Winer, J. A., L. M. Miller, C. C. Lee and C. E. Schreiner (2005). Auditory thalamocortical
transformation: structure and function. Trends in Neuroscience 28(5): 255-263.
Wong, C., N. Chabot, M. A. Kok and S. G. Lomber (2013). Modified areal cartography in auditory
cortex following early-and late-onset deafness. Cerebral Cortex: 1-15.
Wong, D. D. E. and K. A. Gordon (2009). Beamformer suppression of cochlear implant artifacts in
an electroencephalography dataset. Biomedical Engineering, IEEE Transactions on 56(12): 2851-
2857.
Woodruff, G., F. Hiscox, J. Thompson and L. K. Smith (1994). Factors affecting the outcome of
children treated for amblyopia. Eye 8(6): 627-631.

228
Woods, D. L., K. Alho and A. Algazi (1992). Intermodal selective attention. I. Effects on event-
related potentials to lateralized auditory and visual stimuli. Electroencephalography and Clinical
Neurophysiology 82(5): 341-355.
Wunderlich, J. L. and B. K. Cone-Wesson (2006). Maturation of CAEP in infants and children: a
review. Hearing Research 212(1-2): 212-223.
Wunderlich, J. L., B. K. Cone-Wesson and R. Shepherd (2006). Maturation of the cortical auditory
evoked potential in infants and young children. Hearing Research 212(1-2): 185-202.
Xu, L. and N. Zhou (2012). Tonal languages and cochlear implants. Auditory Prostheses, Springer:
341-364.
Yoshinaga-Itano, C. (2003). Universal newborn hearing screening programs and developmental
outcomes. Audiological Medicine 1(3): 199-206.
Yoshinaga-Itano, C., D. Coulter and V. Thomson (2000). The Colorado Newborn Hearing Screening
Project: effects on speech and language development for children with hearing loss. Journal of
Perinatology: Official Journal of the California Perinatal Association 20(8 Pt 2): S132-137.
Yoshinaga-Itano, C., D. Coulter and V. Thomson (2001). Developmental outcomes of children with
hearing loss born in Colorado hospitals with and without universal newborn hearing screening
programs. Seminars in Neonatology, Elsevier.
Yoshinaga-Itano, C., A. Sedey, M. Apuzzo, A. Carey, D. Day and D. Coulter (1996). The effect of early
identification on the development of deaf and hard-of-hearing infants and toddlers. American
Academy of Audiology Annual Conference.
Yoshinaga-Itano, C., A. L. Sedey, D. K. Coulter and A. L. Mehl (1998). Language of early-and later-
identified children with hearing loss. Pediatrics 102(5): 1161-1171.
Yoshinaga‐Itano, C. (2003). Early intervention after universal neonatal hearing screening: impact on
outcomes. Mental Retardation and Developmental Disabilities Research Reviews 9(4): 252-266.
Zatorre, R. J. and P. Belin (2001). Spectral and temporal processing in human auditory cortex. Cerebral
Cortex 11(10): 946-953.

229
Zatorre, R. J., P. Belin and V. B. Penhune (2002). Structure and function of auditory cortex: music
and speech. Trends in Cognitive Sciences 6(1): 37-46.
Zatorre, R. J., J. L. Chen and V. B. Penhune (2007). When the brain plays music: auditory–motor
interactions in music perception and production. Nature Reviews Neuroscience 8(7): 547-558.
Zatorre, R. J., A. C. Evans and E. Meyer (1994). Neural mechanisms underlying melodic perception
and memory for pitch. The Journal of Neuroscience 14(4): 1908-1919.
Zatorre, R. J. and S. Samson (1991). Role of the right temporal neocortex in retention of pitch in
auditory short-term memory. Brain 114(6): 2403-2417.
Zekveld, A. A., S. E. Kramer and J. M. Festen (2010). Pupil response as an indication of effortful
listening: The influence of sentence intelligibility. Ear and Hearing 31(4): 480-490.
Zekveld, A. A., S. E. Kramer and J. M. Festen (2011). Cognitive load during speech perception in
noise: The influence of age, hearing loss, and cognition on the pupil response. Ear and Hearing
32(4): 498-510.
Zeng, F.-G. (2004). Trends in cochlear implants. Trends in Amplification 8(1): 1-34.
Zeng, F.-G., S. Rebscher, W. Harrison, X. Sun and H. Feng (2008). Cochlear implants: system design,
integration, and evaluation. Biomedical Engineering, IEEE Reviews in 1: 115-142.

230
Appendices
What is the optimal timing for bilateral cochlear implantation in
children?
This chapter has been published with required journal formatting:
Gordon, K. A., Jiwani, S., & Papsin, B. C. (2011). What is the optimal timing for bilateral cochlear
implantation in children? Cochlear Implants Int, 12 Suppl 2, S8-14.
Abstract
Bilateral cochlear implants have been provided to children who are deaf in both ears with intent
to promote binaural hearing. If it is possible to establish binaural hearing with two cochlear implants,
these children would be able to make use of interaural level and timing differences to localize sound
and to distinguish between sounds separated in space. These skills are central to the ability to attend
to one particular sound amidst a number of sound sources. This may be particularly important for
children because they are typically learning and interacting in groups. However, the development of
binaural processing could be disrupted by effects of bilateral deafness, effects of unilateral cochlear
implant use, or issues related to the child’s age at onset of deafness and age at the time of the first and
second cochlear implantation. This research aims to determine whether binaural auditory processing
is affected by these variables in an effort to determine the optimal timing for bilateral cochlear
implantation in children. It is now clear that the duration of bilateral deafness should be limited in
children in order to restrict reorganization in the auditory thalamo-cortical pathways. It has also been
shown that unilateral cochlear implant use can halt such reorganization to some extent and promote
auditory development. At the same time however, unilateral input might compromise the development
of binaural processing if cochlear implants are provided sequentially. Mismatches in responses from
the auditory brainstem and cortex evoked by the first and second cochlear implant after a long period
of unilateral cochlear implant use suggest asymmetry in the bilateral auditory pathways which is
significantly more pronounced than in children receiving bilateral implants simultaneously. Moreover,
behavioral responses to level and timing differences between implants suggest that these important

231
binaural cues are not being processed normally by children who received a second cochlear implant
after a long period of unilateral cochlear implant use and at older ages. In sum, there may be multiple
sensitive periods in the developing auditory system which must be considered when determining the
optimal timing for bilateral cochlear implantation.
Introduction
The benefits of bilateral cochlear implantation have been examined in our laboratory by studying
auditory activity in the developing auditory system using both electrophysiological recordings and
behavioural measures of speech perception. We are asking the following questions: (1) Is the auditory
system in children who are deaf and receive cochlear implants able to integrate electrical information
provided by the two independent implants? (2) Does a period of unilateral cochlear implant use change
the bilateral auditory pathways? (3) Does a period of unilateral cochlear implant use disrupt binaural
processing once bilateral cochlear implants are provided? (4) Should bilateral cochlear implants be
provided simultaneously or sequentially?
Children who are deaf and benefit little or not at all from their hearing aids are now able to hear,
communicate and develop oral speech and language skills through the use of cochlear implants.
Indeed, research has shown that providing chronic auditory stimulation with a cochlear implant
promotes development in the auditory brainstem (Gordon, Papsin et al. 2003; Gordon, Papsin et al.
2006; Gordon, Papsin et al. 2007; Thai-Van, Cozma et al. 2007) and thalamo-cortex (Gordon et al.,
2005a, 2005b, 2008a, 2010b; Sharma et al., 2002a, 2002b, 2005, 2009) in children who are deaf from
infancy. Children who use a unilateral cochlear implant show significant improvements, such that
many are able to develop speech and language, learn in mainstream environments and communicate
effectively. Nonetheless, these children still require extensive therapy to achieve optimal
communication outcomes and their hearing deteriorates significantly in noise (Basura et al., 2009;
Gordon and Papsin, 2009a, 2009b; Papsin and Gordon, 2008). Bilateral cochlear implants are now
being provided to children who are deaf with an aim to improve hearing in both quiet and noisy
environments. The two cochlear implants have been provided in the same surgery (simultaneously)
or in two different surgeries with an inter-implantation delay (sequentially) (Kühn-Inackera et al.,
2004).

232
It is clear that cochlear implants should be provided in a timely manner to minimize abnormal
changes in the auditory system caused by bilateral auditory deprivation 2010b; Gilley et al., 2005, 2008;
; Gordon, Tanaka et al. 2005; 2008; 2010b; Gilley et al., 2005, 2008; Lee, Lee et al. 2001; Lomber et
al., 2010; Sharma et al., 2002a, 2009). We are now studying whether using a cochlear implant on one
side and leaving the other side deprived from auditory stimulation also causes abnormal re-
organization in the auditory system. If such changes occur during a sensitive period in development,
they could disrupt binaural processing permanently. We therefore hypothesize that minimizing both
bilateral and unilateral auditory deprivation is important to promote optimal auditory development and
maturation of the auditory system. It is important to test this hypothesis in order to determine the
optimal timing for bilateral cochlear implantation in children.
The auditory system is able to process and integrate auditory information by comparing subtle
differences in level and timing of sounds reaching the two ears. Research has shown that advantages
of binaural hearing, compared to monaural hearing, come from the improved ability to identify the
location of sound sources in space (sound localization) (Basura et al., 2009; Brown and Balkany, 2007;
Ching et al., 2007; Litovsky, 2008a, 2008b) as well as from binaural summation, which allows for
increased loudness perception and ease of listening (Ching et al., 2007; van Hoesel and Tyler, 2003;
Steffens et al., 2008). Binaural hearing also enhances speech intelligibility in the presence of competing
noise and in reverberant environments using the head shadow and squelch effects (Basura et al., 2009;
Brown and Balkany, 2007; Ching et al., 2007; van Hoesel and Tyler, 2003). Moreover, binaural hearing
reduces the risk of auditory deprivation in the unaided ear (Gordon and Papsin, 2009a, 2009b).
Binaural processing is particularly important for children because they are rarely in one location
listening to a single talker in a quiet environment. Indeed, children need to be able to attend to several
sound sources when interacting and playing in groups (i.e., in playgrounds). In addition, learning often
takes place in environments where noise, reverberation and distance pose particular challenges for
hearing and listening (i.e., classroom situations). Children who use only one cochlear implant do not
have access to important binaural cues to aid listening in such challenging conditions. It follows then
that we should always strive to provide binaural hearing to children with hearing loss.
For children who are affected by profound hearing loss and limited residual hearing in both ears,
binaural hearing might only be achieved through bilateral cochlear implantation at the present time
(Basura et al., 2009; Eapen and Buchman, 2009; Papsin and Gordon, 2008). Functional outcomes of

233
bilateral cochlear implantation in children are becoming available; however, it remains unclear to what
extent the developing auditory system is able to process electrical input delivered bilaterally from two
independent cochlear implants.
This paper will review data from a large group of children who were prospectively recruited to
participate in our study of bilateral cochlear implantation in children. The duration of bilateral deafness
was limited to less than 3 years in 3 of 4 groups and the duration of unilateral cochlear implant use
ranged from nil to > 2 years. Current findings show that: 1) unilateral cochlear implant use sets up a
mismatch in responses evoked by the right and left cochlear implants, and 2) improvements in speech
perception when using bilateral versus unilateral cochlear implants are clearest when the duration of
bilateral deafness and unilateral cochlear implant use are limited.
Groups of Study Participants
As of April 2010, 156 children had been recruited into the bilateral cochlear implant study at the
Hospital for Sick Children in Toronto, Canada. Details of these children are shown in Table 1. Of
these, 146 had a short duration of bilateral deafness (< 3 years) and 10 had longer periods of bilateral
auditory deprivation. Of the children with limited bilateral deafness, 72 children received both
implants in the same surgery (Simultaneous group), 19 had a short inter-implant delay of less than 1
year (Short Delay group) and 55 of the children had an average of 5 years of unilateral cochlear implant
use before they received their second implant (Long Delay group). The group with longer durations
of bilateral deafness also had long periods (> 2 years) of unilateral cochlear implant use (Long Deaf-
Long Delay group). The majority of the children had early onset severe-to-profound sensorineural
hearing loss and the etiologies for deafness were often unknown. Any period of usable hearing
(defined by audiometric thresholds with or without hearing aids of ≤ 40 dBHL at any test frequency)
was subtracted from the period of bilateral deafness. This meant that several children had access to
some sounds in normal conversational speech prior to cochlear implantation and were therefore older
than 3 years of age when they received their first implant.

234
Table 1: Demographic variables of bilateral cochlear implant (CI) groups
Mismatches in bilateral activity following a period of unilateral
cochlear implant use
Auditory brainstem responses
We were interested in the ability of the developing auditory system to compare electrical
information delivered by two cochlear implants. Because unilateral cochlear implant use has already
been shown to promote latency decreases in responses from the auditory brainstem (Gordon et al.,
2003, 2006, 2007b, 2007c, 2008b) and cortex (Sharma et al., 2002a, 2002b, 2005), we hypothesized that
evoked responses from the newly implanted side would be prolonged relative to the more experienced
side in children receiving cochlear implants sequentially. If so, this could set up abnormal differences
along the bilateral auditory pathways and potentially compromise binaural processing.
Figure 1 shows two examples of auditory brainstem responses measured on the first day of
bilateral cochlear implant use in a child from the Simultaneous group and a child from the Long Delay
Sequential group. Waves eII, eIII, and eV are clear in waveforms evoked by the right and left cochlear
implants in both children. Wave latencies are similar for right and left evoked responses in the child
who received bilateral cochlear implants simultaneously. Latencies from this child are highlighted in
gray and reproduced in the plot of responses from the child receiving bilateral cochlear implants
sequentially. Responses from that child, who was in the Long Delay group, show mismatches in wave
latencies in line with our hypothesis. Because electrically evoked auditory brainstem responses were
found not to change after the first year of life in children with bilateral deafness (Gordon et al., 2010a),

235
these findings likely reflect the effect of unilateral cochlear implant use rather than increasing age. In
addition, latencies of both responses from the child in the Long Delay group are decreased relative to
the child in the Simultaneous group (shown by the gray highlighted lines).
Figure 1: Brainstem response waveforms of a child receiving bilateral cochlear implants simultaneously
and another child receiving bilateral cochlear implants sequentially with a long inter-implant delay.
Responses from the latter child show a mismatch in timing of brainstem activity evoked by the right
versus left sides. The two traces on each plot represent responses evoked by the right and left sides.
Mean wave eV latency differences between groups are plotted in Figure 2 and confirm that there
was no significant latency differences between sides in the Simultaneous group but that significant
differences were found in both the Short Delay and Long Delay groups. Data from this test time
(initial bilateral cochlear implant use) (Gordon et al., 2007b) were compared with results measured
after bilateral cochlear implant experience (Gordon et al., 2007c, 2008b). Data from the groups studied
at that time showed that wave eV differences in the Short Delay group resolved by 9 months of bilateral
cochlear implant use (Gordon et al., 2007c, 2008b) but did not resolve even after 1 year of bilateral
cochlear implant use in the Long Delay group (Gordon et al., 2007c). Given previous findings that
the time course for maturation of electrically evoked auditory brainstem latencies was approximately
12 months (for wave eV) (Gordon et al., 2006), we would expect wave eV latency differences to resolve
within a similar time period of bilateral cochlear implant use. The persistent delay between wave eV
evoked by the second implanted side relative to the first suggests that auditory brainstem development
promoted after long periods of unilateral cochlear implant use will be different from that which occurs

236
after bilateral deafness. We have also been interested in the decrease of wave latencies from both sides
in the Sequential group compared to the Simultaneous group at initial bilateral implant use. This
finding was expected in the response from the experienced side, as latencies are known to decrease
with unilateral cochlear implant use (Gordon et al., 2003, 2006), but was unexpected for responses
from the newly implanted side. We are presently examining whether this effect can be explained by
the difference in age at testing between the groups or whether this reflects a cross-over effect of
unilateral cochlear implant use to the pathways from the opposite ear. We also need to consider
whether this change has positive implications for bilateral auditory development as suggested by others
(Bauer et al., 2006) or whether this reflects re-organization along the pathways which may be
detrimental to binaural processing.
Figure 2: Brainstem response latencies for waves eV of the naïve ear relative to the experienced ear on
the first day of bilateral device activation. Responses from 13 children in the Simultaneous group, 15
in the Short Delay group and 16 children in the Long Delay group were included in this analysis.
Children with short and long inter-implant delays showed significant mismatches in brainstem activity
evoked from the naïve ear, relative to the experienced ear. This was not seen in the children who
received both cochlear implants simultaneously.
The mismatched timing found between auditory brainstem responses evoked by the right and
left cochlear implants in children implanted sequentially was also found in the binaural difference

237
response (Gordon et al., 2007c, 2008b). This response is calculated by summing responses evoked by
the right and left cochlear implants and then subtracting the response evoked by the coordinated
stimulation from both implants. The resulting amplitude difference, found at (mean ± SD) 0.09 ±
0.00 ms for children in the Simultaneous group, 0.07 ± 0.22 ms for the Short Delay group, 0.15 ± 0.07
ms for children in the Long Delay group and 0.35 ± 0.35 ms for the Older Long Delay group (Gordon
et al., 2007c), is thought to reflect an inhibitory interaction in the auditory brainstem in response to
bilateral input and indicates that the brainstem detects the presence of bilateral input. Because binaural
processing first occurs in the auditory brainstem, the presence of binaural difference responses in most
children receiving bilateral cochlear implants suggests that the auditory pathways at this level of the
system retain the ability to detect bilateral electrical input (Gordon et al., 2007b, 2007c, 2008b). The
consequence of abnormal timing of this response in children who receive bilateral cochlear implants
sequentially rather than simultaneously remains unclear. Interestingly, a similar prolongation in timing
of the binaural difference response has been reported in response to interaural timing cues in normal
hearing individuals (Furst et al., 1985; Riedel and Kollmeier, 2006). Thus, it may be possible that the
mismatch in timing of auditory brainstem activity in some children using bilateral cochlear implants
creates abnormal increases in timing differences between the two ears. The functional consequences
for sound localization and other binaural processing should be explored. Our own efforts to measure
perception of inter-implant level and timing cues are reviewed below (see Section, Perception of speech
and inter-implant timing and level cues).
Cortical responses
Along with measures from the auditory brainstem, we have been exploring effects of bilateral
cochlear implantation with different inter-implant delays on the auditory cortex. Figure 3, shows
examples of cortical responses from three children in the Simultaneous group and one child in the
Long Delay group. Children in the Simultaneous group show a variety of response waveform types as
recently reported (Gordon, Tanaka et al. in press) but each child shows a similar response to right as
to left cochlear implant stimulation. On the other hand, responses in the child who had used a
unilateral cochlear implant for > 2 years are very different from one another (Gordon et al., 2005b,
2008a, 2010b). Given the variability in cortical responses found at the initial stage of bilateral cochlear
implant use in ears with no prior cochlear implant stimulation, we have been unable to determine

238
which of these peaks, if any, are similar to the P1 peak found in children with normal hearing or with
unilateral cochlear implant experience (Sharma et al., 2002a, 2002b, 2005, 2009). Group data, shown
in Figure 4, confirm that most children receiving bilateral cochlear implants simultaneously have similar
cortical responses to right and left cochlear implant stimulation (62 of 72 children – i.e., 86%), whereas
the majority of children receiving a second cochlear implant at older ages and after unilateral implant
use showed differences in cortical response waveforms. Why cortical responses should show this
degree of variability upon acute cochlear implant stimulation in children with both bilateral and
unilateral auditory deprivation remains an interesting question which will need to be addressed by
defining the cortical generators of these responses. We have begun to do this work using our own
beamforming analysis (Wong and Gordon, 2009; Gordon et al., 2010b) which has allowed us to focus
specifically on activity generated in the auditory cortex in response to cochlear implant stimulation. In
addition, we need to determine whether there are functional implications for the mismatches in cortical
responses evoked by right and left cochlear implant stimulation in children receiving cochlear implants
sequentially.
Figure 3: Cortical response waveforms of 3 children receiving bilateral cochlear implants
simultaneously and 2 children receiving a second cochlear implant after a long inter-implant delay.
Several waveform types occur in the children in the Simultaneous group but responses were similar
within children. Cortical responses were very different when evoked by the newly implanted side
compared to the experienced side in the children with long inter-implant delays. The two traces on
each plot represent responses evoked by the right and left sides.

239
Figure 4: Cortical responses were similar in most children in the Simultaneous group and were largely
mismatched in children receiving bilateral cochlear implants sequentially.
Perception of speech and inter-implant timing and level cues
We have been exploring whether mismatches in timing of auditory brainstem activity and
mismatches in cortical responses between right and left evoked responses in children receiving bilateral
cochlear implants sequentially are associated with differences in auditory function in these children
compared to those receiving bilateral devices simultaneously. We are assessing both the ability of
children using bilateral cochlear implants to perceive differences in level and timing cues between two
cochlear implants and their ability to understand speech in quiet and noise.
Findings from these studies revealed that children who receive a second cochlear implant
following a long inter-implant delay (4.9 ± 2.8 years) are able to perceive changes in electrical pulses
when level cues are different between the two implants, but have more difficulty detecting changes in
inter-implant timing cues (Salloum et al., 2010). These findings are consistent with previous reports
showing that adults who use bilateral cochlear implants also interpret inter-implant level differences
more effectively than timing differences (van Hoesel & Tyler, 2003). In addition, findings also revealed
that children who have a long period of unilateral cochlear implant use before receiving their second
implant do not normally hear changes in lateralization of sounds with increasing difference in inter-
implant timing cues (Gordon et al., 2010b; Salloum et al., 2010). Using a behavioural lateralization

240
task, we showed that children often heard bilateral input as coming from their second (less
experienced) implanted side when electrical pulses of the same current level were presented to both
implants simultaneously. This likely occurs when two different cochlear implant devices are used. In
addition, unlike their normal hearing peers, children receiving bilateral cochlear implants after a long
inter-implant delay rarely perceived the sound as coming from the mid-line of the head and sometimes
reported that they heard bilateral input as coming from the left and right sides separately (Salloum et
al., 2010). It is still unclear whether these children are using unilateral or bilateral processing, but the
poor ability to lateralize sound based on timing cues and the absence of bilateral inputs perceived at
the mid-line of the head, suggest that binaural processing may be hindered in children receiving
bilateral cochlear implants following a long period of unilateral cochlear implant use and at older ages.
To examine the effects of bilateral and unilateral deafness on development and on speech
perception outcomes, we have been further investigating speech perception performance in quiet and
in noise, in all four groups indicated in Table 1 after 6 to 12 months of bilateral cochlear implant
experience. Speech perception has been measured using standardized tests which are appropriate for
the age and language level of each child. We have previously reported results in a smaller group
(Gordon and Papsin, 2009b). In order to control for differences in test and age, we calculated the
change in speech perception test scores in bilateral conditions (in quiet and noise) relative to the scores
obtained using the first cochlear implant in quiet. For children in the Simultaneous group, scores
obtained while using the right implant alone in quiet were used, because most children in the sequential
groups had received the first implant in the right ear. Speech perception when the children were
wearing one cochlear implant in the side opposite to the more experienced/right ear in quiet was also
examined. Findings revealed that all groups of children found it most challenging to recognize speech
in noise. However, children who received both of their cochlear implants with a limited duration of
bilateral deafness and with minimal inter-implant delays achieved the largest bilateral improvements in
speech perception scores in both quiet and noise (over unilateral implant use) compared to older
children who had inter-implant delays longer than 2 years (Gordon and Papsin, 2009b). Children with
longer inter-implant delays showed poorer abilities to perceive speech in quiet when using their new
implant compared with an implant on their more experienced side. In contrast, children with minimal
inter-implant delays showed similar performance between ears, when listening to speech with either
the first or second implant alone (Gordon and Papsin, 2009b).

241
Conclusion
We used electrophysiological measures to assess auditory activity at the level of the brainstem
and cortex in children who received cochlear implants bilaterally. We found that children who were
implanted at an early age with a short duration of bilateral deafness and inter-implant delays shorter
than 12 months derived the most benefits from bilateral implantation. Mismatches of timing in
auditory activity in the brainstem and cortex, evoked by the experienced and naïve sides at initial
bilateral device activation resolved by 9 months of consistent bilateral cochlear implant use, suggesting
maturation of the binaural pathways from the brainstem to the cortex within the first year of bilateral
implantation. These children also showed clear improvements in speech perception in quiet and in
noise when using bilateral rather than unilateral cochlear implants during the same period of time. By
contrast, children who received their cochlear implants with an inter-implant delay longer than 2 years
showed mismatched timing of auditory activity in the brainstem and in the cortex. Behavioural
responses show that these children have difficulties perceiving inter-implant timing differences and
have poorer speech perception when using the second cochlear implant compared to the first during
the first 6 to 12 months of bilateral cochlear implant use. At this stage of bilateral implant use, they
do not show a clear improvement in speech perception in noise with bilateral rather than unilateral
implant use. Our findings suggest that children receiving bilateral cochlear implants simultaneously
have the best chance for development of bilateral auditory pathways capable of processing binaural
cues.
Bilateral implants have demonstrated a clear advantage to unilateral stimulation and bilateral
cochlear implantation has been supported and applied in many clinical settings. Through universal
newborn hearing screening initiatives we are now able to identify hearing loss early and provide timely
intervention with the use of auditory prostheses, with the aim of limiting the duration of bilateral and
unilateral deafness in children. We are continuing to expand our research in this domain to better
understand the effects of childhood hearing loss on the developing auditory system and to promote
better hearing through binaural development for all children with hearing loss. Further research will
determine whether this is possible by providing bilateral cochlear implants at young ages and with a
minimal inter-implant delay.

242
References
Bauer, P.W., Sharma, A., Martin, K., Dorman, M. 2006. Central auditory development in children
with bilateral cochlear implants. Archives of Otolaryngology - Head & Neck Surgery, 132(10): 1133-
1136.
Basura, G.J., Eapen, R., Buchman, C.A. 2009. Bilateral cochlear implantation: current concepts,
indications, and results. The Laryngoscope, 119: 2395-2401.
Brown, K.D., Balkany, T.J. 2007. Benefits of bilateral cochlear implantation: A review. Current Opinion
in Otolaryngology & Head and Neck Surgery, 15: 315-318.
Ching, T.Y.C., van Wanrooy, E., Dillon, H. 2007. Binaural-bimodal fitting or bilateral implantation
for managing severe to profound deafness: A review. Trends in Amplification, 11(3): 161-192.
Eapen, R., Buchman, C.A. 2009. Bilateral cochlear implantation: Current concepts. Current Opinions
in Otolaryngology & Head and Neck Surgery, 17: 351-355.
Furst, M., Levine, R.A., McGaffigan, P.M. 1985. Click lateralization is related to the beta component
of the dichotic brainstem auditory evoked potentials of human subjects. Journal of the Acoustical
Society of America, 78: 1644-51.
Gilley, P., Sharma, A., Dorman, M. 2008. Cortical reorganization in children with cochlear implants.
Brain Research, 1239: 56-65.
Gilley, P.M., Sharma, A., Dorman, M., Martin K. 2005. Developmental changes in refractoriness of
the cortical auditory evoked potential. Clinical Neurophysiology, 116(3): 648-657.
Gordon, K.A., Papsin, B.C. 2009a. Understanding hearing loss in children: What is known and what
remains to be learned? Otorinolaringology, 59: 221-227.
Gordon, K.A., Papsin, B.C. 2009b. Benefits of short interimplant delays in children receiving bilateral
cochlear implants. Otology & NeuroOtology, 30(3): 319-331.
Gordon, K.A., Papsin, B.C., Harrison, R.V. 2003. Activity-dependent developmental plasticity of the
auditory brainstem in children who use cochlear implants. Ear & Hearing, 24(6): 485-500.

243
Gordon, K.A., Papsin, B.C., Harrison, R.V. 2005a. Effects of cochlear implant use on the electrically
evoked middle latency response in children. Hearing Research, 204: 78-89.
Gordon, K.A., Papsin, B.C., Harrison, R.V. 2006. An Evoked Potential Study of the Developmental
Time Course of the Auditory Nerve and Brainstem in Children Using Cochlear Implants.
Audiology & NeuroOtology, 11(1): 7-23.
Gordon, K.A., Papsin, B.C., Harrison, R.V. 2007a. Auditory brainstem activity and development
evoked by apical versus basal cochlear implant electrode stimulation in children. Clinical
Neurophysiology, 118(8): 1671-1684.
Gordon, K.A., Tanaka, S., Papsin, B.C. 2005b. Atypical cortical responses underlie poor speech
perception in children using cochlear implants, NeuroReport, 16(18): 2041-2045.
Gordon, K.A., Tanaka, S., Wong, D.D., Papsin, B.C. 2008a. Characterizing responses from auditory
cortex in young people with several years of cochlear implant experience, Clinical Neurophysiology,
119(10): 2347-2362.
Gordon, K.A., Tanaka, S., Wong, D.D., Stockley, T., Ramsden, J.D., Brown, T., Jewell, S., Papsin, B.C.
2010c. Multiple effects of childhood deafness on cortical activity in children receiving bilateral
cochlear implants simultaneously, Clinical Neurophysiology, 122(4): 823-833
Gordon, K.A., Valero, J., Jewell, S.F., Ahn, J., Papsin, B.C. 2010a. Auditory development in the
absence of hearing in infancy, NeuroReport, 21: 163–167.
Gordon, K.A., Valero, J., Papsin, B.C. 2007b. Binaural processing in children using bilateral cochlear
implants. NeuroReport, 18(6): 613-617.
Gordon, K.A., Valero, J., Papsin, B.C. 2007c. Auditory brainstem activity in children with 9-30
months of bilateral cochlear implant use. Hearing Research, 233: 97-107.
Gordon, K.A., Valero, J., van Hoesel, R., Papsin, B.C. 2008b. Abnormal timing delays in auditory
brainstem responses evoked by bilateral cochlear implant use in children. Otology & NeuroOtology,
29(2): 193-198.

244
Gordon, K.A., Wong, D.D., Papsin, B.C. 2010b. Cortical function in children receiving bilateral
cochlear implants simultaneously or after a period of interimplant delay. Otology & NeuroOtology,
31(8): 1293-1299.
Kühn-Inacker, H., Shehata-Dieler, W., Müller, J., Helms, J. 2004. Bilateral cochlear implants: A way
to optimize auditory perception abilities in deaf children? International Journal of Pediatric
Otorhinolaryngology, 68: 1257-1266.
Lee, D.S., Lee, J.S., Oh, S.H., Kim, S.K., Kim, J.W., Chung, J.K., Lee, M.C., Kim, C.S. 2001. Cross-
modal plasticity and cochlear implants. Nature, 409(6817): 149-150.
Lomber, S.G., Meredith, M.A., Kral, A. 2010. Cross-modal plasticity in specific auditory cortices
underlies visual compensations in the deaf. Nature Neuroscience, 13(11): 1421-1427
Litovsky, R.Y. 2008a. Binaural hearing. White paper series, published by Cochlear Americas, February.
Litovsky, R.Y. 2008b. Potential advantages for bilateral cochlear implants. White paper series, published
by Cochlear Americas, February.
Papsin, B.C., Gordon, K.A. 2008. Bilateral cochlear implants should be the standard for children with
bilateral sensorineural deafness. Current Opinions in Otolaryngology & Head and Neck Surgery, 16: 1-
6.
Riedel, H., Kollmeier, B. 2006. Interaural delay-dependent changes in the binaural difference potential
of the human auditory brain stem. Hearing Research, 218: 5-19.
Salloum, C.A., Valero, J., Wong, D.D., Papsin, B.C., van Hoesel, R., Gordon, K.A. 2010.
Lateralization of interimplant timing and level differences in children who use bilateral cochlear
implants. Ear & Hearing, 31(4): 441-56.
Sharma, A., Dorman, M. F., Kral, A. 2005. The influence of a sensitive period on central auditory
development in children with unilateral and bilateral cochlear implants, Hearing Research, 203(1-
2): 134-143.

245
Sharma, A., Dorman, M.F., Spahr, A.J. 2002a. A sensitive period for the development of the central
auditory system in children with cochlear implants: Implications for age of implantation. Ear &
Hearing, 23: 532-539.
Sharma, A., Dorman, M.F., Spahr, A.J. 2002b. Rapid development of cortical auditory evoked
potentials after early cochlear implantation. NeuroReport, 13(10): 1365–1368.
Sharma, A., Nash, A.A., Dorman, M. 2009. Cortical development, plasticity and re-organization in
children with cochlear implants. Journal of Communication Disorders, 42: 272–279.
Steffens, T., Lesinski-Schiedat, A., Strutz, J., Aschendorff, A., Klenzner, T., Rühl, S., Voss, B., Wesarg,
T., Laszig, R., Lenarz, T. 2008. The benefits of sequential bilateral cochlear implantation for
hearing-impaired children. Acta Oto-Laryngologica, 128(2): 164-176.
Thai-Van, H., Cozma, S., Boutitie, F., Disant, F., Truy, E., Collet, L. 2007. The pattern of auditory
brainstem response wave V maturation in cochlear-implanted children, Clinical Neurophysiology,
118(3): 676-689.
van Hoesel, R.J.M., Tyler, R.S. 2003. Speech perception, localization and lateralization with bilateral
cochlear implants. Journal of the Acoustical Society of America, 113(3): 1617-1630.
Wong, D.D., Gordon, K.A. 2009. Beamformer suppression of cochlear implant artifacts in an
electroencephalography dataset. IEEE Transactions on Biomedical Engineering, 56(12): 2851-2857.

246
Benefits and detriments of unilateral cochlear implant use on bilateral
auditory development in children who are deaf
This chapter has been published with required journal formatting:
Gordon, K. A., S. Jiwani and B. C. Papsin (2013). Benefits and detriments of unilateral cochlear
implant use on bilateral auditory development in children who are deaf. Front Psychol 4(719):
1-14.
Abstract
We have explored both the benefits and detriments of providing electrical input through a
cochlear implant in one ear to the auditory system of young children. A cochlear implant delivers
electrical pulses to stimulate the auditory nerve, providing children who are deaf with access to sound.
The goals of implantation are to restrict reorganization of the deprived immature auditory brain and
promote development of hearing and spoken language. It is clear that limiting the duration of
deprivation is a key factor. Additional considerations are the onset, etiology, and use of residual hearing
as each of these can have unique effects on auditory development in the pre-implant period. New
findings show that many children receiving unilateral cochlear implants are developing mature-like
brainstem and thalamo-cortical responses to sound with long term use despite these sources of
variability; however, there remain considerable abnormalities in cortical function. The most apparent,
determined by implanting the other ear and measuring responses to acute stimulation, is a loss of
normal cortical response from the deprived ear. Recent data reveal that this can be avoided in children
by early implantation of both ears simultaneously or with limited delay. We conclude that auditory
development requires input early in development and from both ears.
Introduction
A cochlear implant is an auditory prosthesis which is surgically implanted into the cochlea (inner
ear), and allows children who are deaf to develop oral speech and language. Because the brain is most

247
susceptible to changes in early life, providing access to sound at a young age is essential to promote
auditory development (Papsin and Gordon, 2007;Kral and O'Donoghue, 2010). The implant cannot
restore normal hearing. It provides only a crude representation of acoustic sounds, eliminates
important cochlear processing, and may not be able to completely reverse the effects of deafness. In
addition, cochlear implants were traditionally provided unilaterally (i.e., in only one ear) in children,
leaving the opposite pathways deprived of input and susceptible to degeneration and reorganization
(O'Neil et al., 2010;Gordon et al., 2013;Kral et al., 2013). Yet, despite these disadvantages, many
children achieve excellent listening and oral communication abilities. In the present review, we share
findings from studies exploring whether cochlear implantation can limit reorganization of the deprived
immature auditory brain and promote appropriate and normal-like development along the auditory
pathways.
The auditory system reorganizes when bilaterally deprived
Prior to cochlear implantation, the absence of auditory input to the auditory system leaves the
brain vulnerable to reorganization (Nishimura et al., 1999;Bavelier et al., 2000;Finney et al., 2001;Lee
et al., 2001;Bavelier and Neville, 2002;Bavelier et al., 2006;Merabet and Pascual-Leone, 2010).
Secondary and association auditory areas, including parts of the planum temporale, all of which
respond to multi-sensory input including hearing, vision and touch (Pandya and Yeterian, 1985;Giard
and Peronnet, 1999;Calvert et al., 2001;Calvert and Thesen, 2004), become recruited by the visual
(Finney et al., 2001;Lee et al., 2001;Lee et al., 2007b;Lomber et al., 2010;Meredith and Lomber, 2011)
and somatosensory (Levänen et al., 1998;Levänen and Hamdorf, 2001;Auer Jr et al., 2007;Meredith
and Lomber, 2011) systems to perform non-auditory functions. As a consequence of early auditory
deprivation, processing of visual peripheral localization by the posterior auditory field (Lomber et al.,
2010), visual motion detection by the dorsal zone of the auditory cortex (Lomber et al., 2010), and
somatosensory sensation by the anterior auditory field (Meredith and Lomber, 2011) become enhanced
in individuals who are deaf. These changes appear to result from a direct competition for resources in
areas which receive multi-sensory input. If governed by principals of Hebbian processing (Hebb,
1949;Abbott and Nelson, 2000;Song et al., 2000), neurons in these areas might preferentially form
viable connections with non-auditory inputs to the detriment of inputs carrying auditory information.
We must be concerned by the reorganization of the deaf auditory cortex because, depending on how

248
quickly these processes occur, they may be impossible to reverse and could impair outcomes after
cochlear implantation. It is also becoming clear that these changes do not occur uniformly in children
who are deaf and may be related to the heterogeneity in the onset and cause of pediatric deafness
(Gordon et al., 2011a;Gordon et al., 2011c).
Limiting the period of bilateral deafness in early life is essential to drive maturation in the
auditory pathways (O’Donoghue, 1999;Kral et al., 2001;Ponton and Eggermont, 2001; Sharma et al.,
2005;Papsin and Gordon, 2007;Gordon et al., 2008;Nikolopoulos et al., 2009;Gordon et al., 2010), and
promote optimal hearing and speech and language development (Beadle et al., 2005;Harrison et al.,
2005;Nicholas and Geers, 2007;Geers and Sedey, 2011). Many studies investigating auditory
development after cochlear implantation focus on children who are deaf in infancy, but do not examine
the larger heterogeneity in etiology, onset and/or degree of deafness. These factors may each have
unique effects on auditory activity in the brain prior to implantation. For example, bilallelic mutations
of the Gap Junction Beta-2 (GJB-2) gene causes deficits in the cochlea at likely very early stages of
development with possible consequences for auditory function after implantation (Propst et al., 2006).
The GJB-2 gene normally codes for the connexin-26 protein, which creates gap junctions in the
cochlea necessary for the appropriate release and maintenance of electrochemical gradients. This in
turn, generates action potentials and stimulates the auditory nerve (Kelley et al., 1998;Cohn and Kelley,
1999;Gualandi et al., 2002). Electrophysiological recordings of auditory evoked cortical activity at
initial cochlear implant activation in children with severe GJB-2 mutations revealed that responses
from the cortex were more homogenous in this cohort compared to those children who did not have
such a mutation. Auditory evoked cortical responses in children with GJB-2 mutations were
characteristic of earlier stages of cortical development, perhaps reflecting restricted spontaneous
activity in the auditory system and more limited access to sound prior to implantation compared to
their peers who did not have a GJB-2 related deafness (Gordon et al., 2011c). This was further
supported by poorer hearing sensitivity in the low frequencies in the GJB-2 group (Propst et al., 2006).
The degree of residual hearing is another important predictive factor for cochlear implant
outcomes. Traditional candidacy criteria for cochlear implantation in children include a diagnosis of
permanent severe-to-profound hearing loss bilaterally with little or limited access to acoustic input
through hearing aids (Osberger et al., 2002). We recently reported that children who had better hearing
at 250Hz used their hearing aids for longer durations prior to receiving a cochlear implant (Hopyan et
al., 2012). Of interest, these children performed significantly better on tests of music perception with

249
their implants, particularly when detecting differences in rhythm, compared to children who did not
have acoustical access to these low frequencies prior to implantation (Hopyan et al., 2012). Thus, there
are advantages of acoustical input for auditory development which can be capitalized upon after
cochlear implantation. In general, we are learning that the cause, onset and degree of deafness in any
one child will be important to understand in order to ensure that he/she makes the best possible use
of his/her device.
Unilateral cochlear implantation restores hearing and promotes
auditory development
The cochlear implant was made available to children in North America in the early 1990s and
works by stimulating the auditory pathways with electrical pulses. The implant contains an array of
electrodes which is surgically placed in the scala tympani of the cochlea. These electrodes each deliver
electrical pulses to stimulate the auditory nerve. External equipment is worn which takes in acoustic
sound through the microphone, extracts frequency and intensity information in a speech processor
and sends instructions to an internal device through an FM transmitting coil. The internal receiver-
stimulator sends this information to the electrodes which are organized to mimic the normal cochlea;
high frequency sounds are allocated to basal electrodes with lower frequencies being allocated to
progressively more apical electrodes. In this way, the child receives an electrical representation of the
acoustic world and learns to understand sounds including speech.
Auditory brainstem development, measured by decreasing latencies of evoked potential peaks,
is largely complete by the first year of cochlear implant use in children with early onset deafness
(Gordon et al., 2003;Gordon et al., 2006), indicating increasing efficiency of neural conduction and
improved neural synchrony with exposure to sound (Gordon et al., 2003). Similar changes have been
reported from the auditory brainstems of normal hearing children over a similar time-course (Salamy
and McKean, 1976;Starr et al., 1977;Jerger and Hall, 1980;Salamy, 1984;Hecox and Burkard, 2006).
Data from Gordon et al (2006) is shown in Figure 1a; on the left is an example of an electrically evoked
auditory brainstem response. The stimulus artifact is shown at time 0ms followed by waves eII, eIII
and eV, and on the right, the latency values of wave eV are plotted at initial device activation and over
the first year following cochlear implant use in 44 children who had early onset deafness and were

250
implanted unilaterally (Gordon et al., 2006). Recently, we recorded these same responses in 2 children
who were in the original study once they had over a decade of unilateral cochlear implant experience.
Their responses are shown in Figures 1b and 1c. In both cases, wave eV latency clearly decreases over
the first year of cochlear implant use, with no further changes thereafter. This suggests that activity in
auditory brainstem is largely complete by the first year (Gordon et al., 2006).

251

252
Figure 1: (A) Example of an electrically evoked auditory brainstem response waveform is shown on
the left. The onset of the cochlear implant artifact is shown at time 0ms, followed by peaks eII, eIII
and eV. Data from Gordon et al. (2006) are plotted on the right and show the mean wave eV latency
values of 44 children recorded at initial activation of the implant, and at months 2, 6 and 12 following
unilateral cochlear implantation. Figures (B) and (C) on the right show the changes in the brainstem
responses of 2 children who were in the original study (Gordon et al., 2006), recorded from initial
activation of the device to different intervals over the first year of cochlear implantation use. New
responses recorded after 10 years of unilateral cochlear implant experience are also shown further
confirming that little change in the eV latency occurs beyond the first year of implant use. The wave
eV latencies at each time-point are represented on the right for each child.
Further studies concentrated on the development of cortical auditory activity in children with
time after cochlear implantation. Cochlear implants provided to children who are congenitally deaf
within 3.5 years of bilateral deafness promote age-appropriate cortical responses over the first 3-6
months of implant use (Sharma et al., 2002a). After this initial period, these responses change at a rate
which is similar to normal (Eggermont et al., 1997;Eggermont and Ponton, 2003). We recently
assessed changes in cortical responses after longer term unilateral cochlear implant use in children
implanted early (Jiwani et al., 2013a). Grand mean cortical evoked responses from 79 unilateral
cochlear implant users (red waveforms) are plotted in Figure 2 along with the grand mean responses
from 58 normal hearing peers (black waveforms) for different intervals of hearing experience. Figures
2a, 2b and 2c show grand mean cortical evoked waveforms from children who have between 0 to 7
years (40 cochlear implant users; 11 normal hearing), 7 to 12 years (21 cochlear implant users; 18
normal hearing) and over 12 years (18 cochlear implant users; 29 normal hearing) of hearing
experience, respectively. Cochlear implant users represented in these Figures had limited durations of
bilateral deafness prior to implantation (2.03 ± 1.36 years) with typical heterogeneity in their etiologies
of deafness.

253
Figure 2: Grand mean cortical evoked responses from 79 cochlear implant users (red waveform)
(Jiwani et al., 2013a) are plotted for children who have (A) under 7 years of hearing experience (4.3 ±
1.7 years; n=40), (B) between 7 to 12 years of hearing experience (9.4 ± 1.6 years; n=21), and (C) those
who have more than 12 years of hearing experience (13.8 ± 0.9 years; n=18). Mean responses for each
range of hearing experience are compared to a normal and mature cortical waveform (black waveform)
(Jiwani et al., 2013a), recorded from normal hearing peers who are matched for hearing age (n=58).
As shown in Figure 2a, responses from children with up to 7 years of hearing experience with
an implant or with normal bilateral hearing are dominated by a large and broad positive amplitude
peak, labelled P1/P2. Comparison of peak latencies (t(47.3)=-1.63; p>0.05) and amplitudes (t(42.1)=-0.64;
p>0.05) reveal no significant differences between the two groups. This positive-peaked response is
believed to reflect either excitatory auditory activity from the thalamus to deep layers of the auditory
cortex (Liegeois-Chauvel et al., 1994), or auditory driven activity from association auditory areas to the
reticular activating system in the non-lemniscal auditory pathways (Kraus et al., 1992;Ponton et al.,
2000;Ponton and Eggermont, 2001). As thalamo-cortical and cortico-cortical connections develop
around 9 to 12 years of age in superficial layers of the auditory cortex, a small negative amplitude peak,
labelled N1, develops in the cortical evoked response and bifurcates the large P1/P2 response into three
peaks: P1, N1 and P2. Similar developmental changes to the cortical response are observed in early
implanted cochlear implant users who have equal durations of hearing experience. Indeed, as shown

254
in Figure 2b, with 7 to 12 years of auditory experience (9.38 ± 1.57 years in cochlear implant users;
9.92 ± 1.57 years in normal hearing individuals), the cortical response in both groups begins to develop
into a polyphasic waveform. The grand mean response from all 21 unilaterally implanted children
begins to bifurcate into a 3-peaked cortical response at this stage of implant use (Figure 2b).
Differences in the wavepeak latencies (P1: t(10)=-0.88, p>0.05; N1: t(10.18)=-1.3, p>0.05; P2: t(10.77)=1.43,
p>0.05) and peak-to-peak amplitudes (P1-N1: t(6.87)=1.75, p>0.05; N1-P2: t(10.67)=2.2, p>0.05) between
both groups were not significant. This response continues to develop with time. As auditory pathways
mature in the auditory cortex, peaks P1-N1-P2-N2 become clearly present (Figure 2c) when auditory
experience exceeds 12 years in all 18 cochlear implant users (13.81 ± 0.92 years of unilateral implant
experience) and 29 normal hearing peers (15.30 ± 1.81 years of age and hearing) (Jiwani et al., 2013a).
The data from individuals with normal hearing shown in Figure 2 is consistent with findings by
Ponton, Eggermont and colleagues who suggested that peak N1 normally emerges around 9 to 12 years
of age reflecting maturation of thalamo-cortical and cortico-cortical loops in superficial layers of the
auditory cortex (Ponton et al., 2000;Eggermont and Ponton, 2003). These pathways mediate the
transfer of primary auditory and multi-sensory input from the thalamus to various regions of the
ipsilateral and contralateral auditory cortices (Winer et al., 2001;Winer et al., 2005;Razak et al., 2009),
and the transmission of information from the auditory cortex to primary and secondary sensory areas
in both hemispheres (Read et al., 2002;Lee and Winer, 2005;Klinge et al., 2010). The developmental
trajectory of the electrically evoked cortical waveform suggests that similar development is taking place
in children using cochlear implant (Jiwani et al., 2013a), perhaps establishing: 1) appropriate relay of
auditory input from the ear to the cortex, via the thalamus; 2) communication between the two cortical
hemispheres; and/or 3) connectivity between different sensory areas. These normal-like
developmental changes to the auditory cortex may underlie the impressive improvements in auditory
function observed with cochlear implant use, over time (Beadle et al., 2005;Nicholas and Geers,
2007;Geers and Sedey, 2011).

255
Differences from normal persist in auditory processing despite long
durations of unilateral cochlear implant use
Although early implantation of young children results in normal-like cortical response peaks, as
shown in Figure 2c, the waveform has at least one abnormality. Specifically, the amplitude of the P2
peak in cochlear implant users is larger than in normal hearing peers (t(14.51)=2.49, p<0.05) (Jiwani et
al., 2013a). The importance of this recent finding is that it suggests deviations from normal cortical
processing remain in these young people despite long-term unilateral implant use. Enhanced P2 peak
amplitudes in normal hearing adults are known to reflect increases in selective attention (Picton and
Hillyard, 1974;Hocherman et al., 1976;Rif et al., 1991;García-Larrea et al., 1992;Posner and Dehaene,
1994;Grady et al., 1997;Fujiwara et al., 1998;Tremblay et al., 2009) and increases in multi-sensory
integration during auditory processing (Hari, 1990;García-Larrea et al., 1992;Levänen et al.,
1998;Webster and Colrain, 2000;Moller and Rollins, 2002;Crowley and Colrain, 2004;Johnson and
Zatorre, 2005). These processes cause a reduction in the primary network which becomes
supplemented by the frontal and parietal areas through increased neural recruitment and synchrony
(Tremblay et al., 2001;Tremblay and Kraus, 2002;Tremblay, 2007;Tremblay et al., 2009) from the non-
primary and association auditory pathways (Hocherman et al., 1976;Kraus and McGee, 1993;Kraus et
al., 1994;Grady et al., 1997;Busse et al., 2005). It is therefore possible that the larger than normal
amplitude of peak P2 observed in children with long-term cochlear implant experience reflects
increased cognitive demands for attention and multi-sensory system integration during hearing. This
may reflect compensatory mechanisms to offset: 1) the reorganization in the auditory brain potentially
occurring during the period of deafness prior to implantation; 2) the abnormal auditory input provided
by the cochlear implant; and/or, 3) the absence of sound to the un-implanted ear which may lead to
reorganization in the deprived pathways.
Cochlear implant users compensate for the abnormal input they receive through the device
(Doucet et al., 2006;Giraud and Lee, 2007;Lee et al., 2007a;Lee et al., 2007b;Hopyan-Misakyan et al.,
2009;Strelnikov et al., 2010;Hopyan et al., 2011;Kral and Sharma, 2011;Lazard et al., 2011;Hopyan et
al., 2012;Lazard et al., 2012;Sandmann et al., 2012). We found that children using cochlear implants
depend on visual cues more heavily than normal to listen for complex information embedded in
speech. Emotion perception was tested using 2 subtests of the standardized Diagnostic Analysis of
Nonverbal Behavior-2 (DANVA-2) in 18 cochlear implant users who received one implant by 2.9 ±

256
0.9 years, had 7.2 ± 1.3 years of cochlear implant experience at the time of the test, and had good
speech perception skills. In the first test, children listened to the spoken sentence: “I’m going out of
the room now and I’ll be back later” (24 trials), and had to decide which 1 of 4 emotions (happy, sad,
angry or fearful) was conveyed by the voice. In the second test, children watched pictures of other
children’s faces, each depicting 1 of the same 4 emotions, and had to decide which emotion was
conveyed by the photographs. Performance accuracy was assessed for each task, and compared to 18
normal hearing controls who were matched for age (10.3 ± 1.5 years of age) (Hopyan-Misakyan et al.,
2009).
Children using cochlear implants showed significantly poorer than normal performance on the
emotion identification task in the auditory subtest (F(1,34) = 43.7, p>0.01). This deficit does not
reflect a general failure to identify emotions, however, since they performed as well as their peers with
normal hearing when the emotions were presented in the visual modality (F(1,34) = 0.1, p>0.05)
(Hopyan-Misakyan et al., 2009). The inability of these children to perceive emotions in speech might
reflect abnormal development of cortical representation of emotional prosody in speech without
normal hearing (Nishimura et al., 1999;Lee et al., 2001;Doucet et al., 2006;Lee et al., 2007b;Meredith
and Lomber, 2011;Sandmann, 2012;Sandmann et al., 2012).
In sum, unilateral cochlear implantation promotes the development of normal-like activity in the
auditory pathways over the long-term, but functional abnormalities persist. These could reflect: 1)
deleterious or irreversible changes to neural reorganization which occurred during the period of
auditory deprivation in early life, 2) abnormal representation of sound through electrical pulses
stimulation of the auditory system, and/or 3) abnormal cortical development driven by the absence of
auditory input to the deprived pathways from the opposite un-implanted ear. We have been studying
effects of the latter issue in children.
Binaural hearing is not available to traditional unilateral cochlear
implant users
Hearing through only one cochlear implant eliminates access to binaural hearing, which is the
ability of the auditory system to process and integrate auditory input from both ears. Binaural hearing

257
is especially important for children because they are rarely in one place and listening to a single speaker
at a time. Children need to attend to and discriminate between several sound sources when playing
and learning. The noise, reverberation and distance, predominant in most situations including typical
classrooms, make it challenging for children to listen and learn when binaural cues are not accessible.
For children who are deaf in both ears, binaural hearing might achieved with bilateral cochlear
implantation (i.e., cochlear implants in both ears) (van Hoesel and Tyler, 2003;Litovsky et al.,
2004;Litovsky et al., 2006;Brown and Balkany, 2007;Litovsky, 2008;Steffens et al., 2008b;Basura et al.,
2009;Eapen and Buchman, 2009;Gordon et al., 2010;Salloum et al., 2010;Chadha et al., 2011;Gordon
et al., 2011b). Bilateral cochlear implantation is now being increasingly provided to children either in
the same surgery (simultaneously) or in two different surgeries following a period of unilateral implant
use (sequentially).
Bilateral cochlear implants attempt to restore binaural hearing by providing information to both
ears. Normally, the auditory system compares, processes and integrates subtle differences between
level and timing of sounds reaching each ear. In this way, binaural hearing allows: 1) the
identification/localization of sound sources in space (Batteau, 1967;Lorenzi et al., 1999;Van Deun et
al., 2009b;Grothe et al., 2010); 2) increased perception of loudness through binaural summation
(Bocca, 1955;Blegvad, 1975); and 3) improved hearing in quiet and in noisy environments through the
head shadow and squelch effects (Hawley et al., 2004;Van Wanrooij and Van Opstal, 2004). Binaural
hearing also makes communication less tiring which enables listening and communication to be a more
pleasant experience. Although restoring binaural hearing is the goal of bilateral implantation, this has
not been completely realized in either adults or children (van Hoesel and Tyler, 2003;Seeber and Fastl,
2008;Grieco-Calub and Litovsky, 2010;Salloum et al., 2010).
Children who are deaf in both ears hear speech better with bilateral cochlear implants than
unilateral implants (Litovsky et al., 2004;Brown and Balkany, 2007;Ching et al., 2007;Galvin et al.,
2007;Peters et al., 2007;Litovsky, 2008;Seeber and Fastl, 2008;Steffens et al., 2008a;Basura et al.,
2009;Eapen and Buchman, 2009;Gordon and Papsin, 2009;Van Deun et al., 2009a;Salloum et al.,
2010;Chadha et al., 2011), but do not hear binaural cues normally (Grieco-Calub and Litovsky,
2010;Salloum et al., 2010). Outcomes improve when both implants are provided with limited delays
and at young ages (van Hoesel and Tyler, 2003;Gordon and Papsin, 2009;Van Deun et al.,
2009a;Gordon et al., 2010;Chadha et al., 2011). As the duration of inter-implant delay decreases, the
two ears develop more symmetric speech perception abilities and children show increasing advantages

258
of bilateral over unilateral implantation (Gordon and Papsin, 2009). Significant improvements on
standardized speech perception tests are seen as early as 6 months following bilateral cochlear implant
stimulation in children who receive their second implant simultaneously or within short delays
(Gordon and Papsin, 2009). Furthermore, children implanted with both cochlear implants
simultaneously derive significantly more benefit from spatial separation of noise compared to children
who have longer delays between implants (Chadha et al., 2011). Sound localization improves in
children who are provided access to sound early and in both ears (Van Deun et al., 2009a). By contrast,
children who receive both cochlear implants sequentially after long inter-implant delays (>2 years)
have persistent asymmetries in auditory function and compromised bilateral benefits for speech
perception, even after 36 months of bilateral cochlear implant use (Gordon and Papsin, 2009).
Sequentially implanted children also seem to depend more on their first implanted ear than their second
for speech perception, and show less bilateral improvement (relative to unilateral implant use) on
speech outcomes than children implanted simultaneously or with limited delay (Gordon and Papsin,
2009). These children localize sound inaccurately and rely heavily on level cues to do so (Grieco-Calub
and Litovsky, 2010). The negative effect of inter-implant delay might be explained by underlying
changes to the developing auditory pathways before and after unilateral and bilateral implantation.
Evidence of a short sensitive period for bilateral input in human
auditory development
Data presented in Figures 1 and 2 shows that unilateral stimulation promotes development of
the auditory pathways (Jiwani et al., 2013a), thus limiting effects of deafness. At the same time, this
development might occur at the expense of pathways from the opposite and deprived ear. This might
be explained by the absence of inhibition which would normally have come from input from the
opposite ear during binaural hearing (Grothe et al., 2010). Without this inhibition, ascending
projections from the stimulated ear may be abnormally strengthened in children who are deaf and use
unilateral cochlear implants.
We studied bilateral auditory function in children who had different durations of unilateral
exposure. We hypothesized that the stage of unilaterally driven brainstem development would be an
important factor to consider. Perhaps changes occurring in the brainstem at earlier stages of

259
unilaterally driven development would have less long lasting consequences on the bilateral pathways
than after the unilaterally stimulated brainstem reached maturity. Development in the auditory
brainstem is largely complete by 1 year of unilateral implant use (Gordon et al., 2006). Thus, children
with >2 years of unilateral experience were categorized as having mature auditory brainstem function
and long-term unilateral use. Children with <1 year of unilateral experience were considered to have
short-term use with continuing auditory brainstem development. Auditory development in these
children was compared to that of children who were deaf and had not yet used cochlear implants (i.e.,
limited to no auditory brainstem development). All children were implanted bilaterally, allowing us to
assess auditory brainstem function evoked by stimulation from each ear. All children receiving bilateral
implants sequentially showed brainstem responses which were faster when evoked by the experienced
ear compared to the newly implanted ear at initial bilateral implant use (Gordon et al., 2008b). This
was expected and confirmed earlier findings that the first implant promoted improved neural
conduction through the brainstem. Repeated tests completed after 1.7 ± 1.65 year of bilateral implant
use indicated mismatches in response latencies persisted in a group of children receiving the second
implant after a long delay (>2 years) (Gordon et al., 2012). Increased response latencies in response
to sound from the second implanted side could reflect decreased axonal myelination, longer neural
conduction times, slower or weaker synapses or more asynchronous neural activity – all signs of more
limited brainstem development. Abnormal mismatches between brainstem response latencies were
never present in children receiving bilateral implants simultaneously and resolved with bilateral implant
use in children who received both implants after a short inter-implant delay (<1 year) (Gordon et al.,
2007b;Gordon et al., 2008b;Gordon et al., 2011b;Gordon et al., 2012). Thus, allowing the brainstem
to develop unilaterally for >2 years compromises the later promotion of symmetrically functioning
bilateral auditory brainstem pathways.
Mismatched bilateral auditory development in sequentially implanted children was not restricted
to the brainstem. Effects of asymmetric activity in the pathways from the first stimulated ear were
also found in the auditory cortex. Consistent with the brainstem findings, cortical abnormalities were
not resolved by chronic bilateral implant use (3.57 ± 0.74 years) when unilateral experience exceeded
1.5 years in children who were implanted early (1.87 ± 1.25 years of age). These findings were recently
reported by Gordon et al. (2013) and are shown in Figure 3 (re-printed from that paper). We used a
unique and validated ‘Time Restricted Artifact and Coherent Suppression’ (TRACS) beamformer
method (Wong and Gordon, 2009) to suppress the electrical artifact from the cochlear implant device

260
and spatially localize areas of cortical activity in hemispheres ipsilateral and contralateral to stimulation.
Like many imaging methods, the brain was divided into thousands of 3-dimensional coordinate spaces
(voxels). Responses were recorded at 64-cephalic surface electrodes and the contribution of the dipole
centered in each voxel to the measured field was assessed by the adaptive spatial filter of the TRACS
beamformer. Dipole moments for a given voxel were calculated across latency (virtual sensor) and
peak values were used for analyses.
Cortical responses were evoked by unilateral electrical pulse trains delivered from one implant
electrode in 7 children with normal hearing, 8 children who were implanted unilaterally in the right ear
(2.32 ± 1.61 years) and had 7.21 ± 2.48 years of hearing experience and 26 children who used bilateral
cochlear implants for 3.42 ± 0.59 years. Of the bilateral implant users, 10 children received both
cochlear implants simultaneously and 16 were sequentially implanted (right ear implanted first with no
hearing aid in the left ear). Bilateral deafness prior to implantation was limited (1.74 ± 0.90 years) in
all children. The children in this study had less than 12 years of hearing experience, and therefore all
produced a cortical evoked response which was dominated by an immature large amplitude positive
peak, similar to the one shown in Figure 2a. The differences between the dipoles from the left and
right auditory cortices were normalized as a percent lateralization (% lateralization = (dipole right –
dipole left) / (dipole right + dipole left) x 100).
A larger than normal variability in the lateralization of cortical dipoles was found in children
receiving bilateral cochlear implants sequentially. A factor analysis of multiple demographic variables
identified the duration of unilateral implant use as the factor which best accounted for the spread of
cortical responses. We thus further analyzed the cortical lateralization data for effects of duration of
unilateral implant use occurring prior to bilateral implantation. When responses were evoked by the
first (i.e., right) implant, there was an increase in lateralization of activity to the contralateral left
auditory cortex with unilateral implant use. This became significantly larger than the percent of cortical
lateralization in the simultaneously implanted group at 1.48 years of unilateral implant use. Consistent
results were obtained in data evoked by the second (i.e., left) implant but, in this case, cortical
lateralization changed from the normally expected contralateral direction to ipsilateral lateralization
with unilateral implant use. This abnormal switch to larger activity in the ipsilateral auditory cortex
became significantly different from responses in the simultaneously implanted group by 1.37 years of
unilateral implant use. These analyses indicated that children with longer than approximately 1.5 years
of unilateral implant use had experienced an abnormal strengthening of pathways from their first

261
implanted right ear through the auditory brainstem (Gordon et al., 2008b;Gordon et al., 2012) to their
left contralateral cortex. This was not resolved by several years of bilateral implant use and was
associated with poorer speech perception in the second than first implanted ear (Gordon et al., 2013).
The importance of restricting unilateral implant use to less than 1.5 years is further evident in
Figure 3 (reprinted from Gordon et al., 2013). Here, the grand mean lateralization of cortical activity
are shown (Figure 3a), as well as the grand mean dipole moments identified from the virtual sensors
in each hemisphere (Figure 3b). The group of 16 sequentially implanted children have been divided
into two groups based the cut off of 1.5 years of unilateral implant use. The Short Delay group includes
7 children who had 0.86 ± 0.1 years of unilateral implant experience at the time of testing. The other
9 children, the Long Delay group, had more than 2 years of unilateral implant use (3.44 ± 1.27 years).
The single positive peaked response is clear in all of the group averaged waveforms shown in Figure
3b. The maximum dipoles were marked and analyzed in each child. The left plot of Figure 3c shows
that dipoles evoked by stimulation from the first/right implanted ear resulted in significantly higher
dipoles in the left auditory cortex (blue bars) of children who had >1.5 years of unilateral implant use
(Unilateral and Long Delay groups) than other groups of children (F(4,36)=3.52, p<0.05). The similar
findings for these two groups confirm that unilaterally driven strengthening of projections to the
contralateral left auditory cortex was not reversed by the addition of a second cochlear implant. This
was true despite the children in the Long Delay group having had several years of bilateral implant
experience at the time of the test. The right plot in Figure 3c shows mean dipoles for each auditory
cortex in response to left/second cochlear implant stimulation. The Long Delay group shows
significantly higher dipole moments in the left auditory cortex than the other groups of children
(F(3,29)=5.31, p<0.01). Thus, regardless of which ear was stimulated, the left auditory cortex
(contralateral to the first/right implanted ear) was the more active side of the brain in children who
had used one implant for >1.5 years. One explanation for this finding is that the specialized processing
of language in left auditory cortex (Zatorre and Belin, 2001;Zatorre et al., 2002;Tervaniemi and
Hugdahl, 2003;Firszt et al., 2006) is abnormally increased in unilateral cochlear implant users. It is not
clear, however, how such a network would have been recruited by the simple non-speech stimuli used
in the present experiment. An alternate explanation is that unilateral stimulation allowed abnormal
strengthening of pathways from that ear.

262

263
Figure 3: Re-printed with permission from Gordon et al. (2013). “(A) Per cent cortical lateralization
(mean ± 1 standard error) is plotted for each participant group. Greater than normal contralateral
lateralization to right/CI-1 stimuli was found in long delay and unilateral cochlear implant users
(P<0.05 and <0.0001, respectively) but not in short delay and simultaneous groups (P>0.05). The long
delay group showed a decrease in contralateral lateralization/increase in ipsilateral lateralization relative
to those with normal hearing in response to left/CI-2 stimulation. This did not occur in the short delay
and simultaneous groups. (B) Grand mean virtual sensor data for left and right hemispheric sources
of P1 (normal hearing) and P1ci (cochlear implant users for stimulation from right/CI-1 and left/CI-
2). Large peaks in responses to CI-1 (right) stimulation can be seen in the long delay and unilateral
group data. (C) Left and right hemispheric dipole moments (mean ± 1 SE) for P1/P1ci in each group
in response to right/CI-1 and left/CI-2 stimulation. In response to CI-1 (right) stimulation, there is a
marked increase in left hemispheric dipole moments in participant groups with >2 years of unilateral
hearing experience (long delay and unilateral; P<0.05).” (Gordon et al., 2013, Brain, Figure 7, p.11)
Further evidence that the cortical changes were due to unilaterally driven strengthening was
found by assessing activity in the ipsilateral/right auditory cortex. We assessed which ear preferentially
activated the hemisphere contralateral to the ear deprived during the period of unilateral implant use
(i.e., the right auditory cortex). The right auditory cortex was expected to respond more strongly to
input from the left than right ear because the majority of neurons from one ear normally cross to the
contralateral brainstem and ascend ipsilaterally from there. This was confirmed in the group of
children with normal hearing and children with limited unilateral implant use prior to bilateral
implantation (Short Delay and Simultaneous). By contrast, this pattern was reversed in children in the
Long Delay group. This meant that this group of children had experienced a strengthening of
pathways from their hearing ear to both the ipsilateral (right) cortex, as shown by the reversal of aural
preference, as well as the contralateral (left) cortex as shown by the data in Figure 3. The same reversal
of aural preference in the cortex ipsilateral to the hearing ear has recently been reported in congenitally
deaf white cats (Kral et al., 2013).
The abnormal strengthening of pathways from the unilaterally hearing ear to the immature brain
seems to initially occur at the level of the brainstem. This is supported by evidence of mismatched
brainstem latencies observed from children with long (>2 years) unilateral hearing experience (Gordon

264
et al., 2012). The shorter wave eV latencies evoked from the more experienced ear suggest an
increasing efficiency of activity from this side and a weakening of pathways from the opposite ear, as
reflected by slower peak latencies on the second implanted side. This could result from a lack of
inhibitory processes in the brainstem which are normally present during binaural hearing (Grothe et
al., 2010). Listening from one side would allow auditory input from the first right implanted side to
be projected to the cortex with abnormally high excitation during development thus strengthening
pathways to the contralateral cortex. It appears that if this is allowed to occur until the brainstem is
largely developed (i.e., >1 year of unilateral implant use), it establishes asymmetric activity in the
auditory pathways which is not easily reversed by providing a second implant in the deprived ear.
Limiting the period of unilateral hearing in children by providing bilateral cochlear implants with little
or no delay appears to protect the bilateral pathways from this abnormal development. These findings
thus suggest that there is a sensitive period of 1.5 years for binaural auditory development in children.
Long-term unilateral implant use in older children causes lasting
asymmetry in the bilateral auditory pathways
We make the case above that unilateral implant use in children who have been deaf since infancy
should be limited to less than 1.5 years to promote normal-like symmetrical development of the
auditory pathways from both ears. However, providing bilateral implants within this time frame may
not always be possible. For example, many adolescents/young adults who were implanted as babies
and have already had many years of unilateral hearing experience are now seeking a cochlear implant
for their opposite ear in hopes of deriving benefits of bilateral implantation. These children are
different in several ways from our previous research cohorts of sequentially implanted children. They
have had very long periods of unilateral cochlear implant use concurrently with long durations of
deprivation in their non-implanted ear, and they are no longer children. We thus expect unique cortical
development in this new group of bilateral implant users relative to our previous study groups.
Figure 4 shows the cortical responses recorded at a midline cephalic location on the head (Cz)
and evoked by cochlear implant stimulation from each ear on the first day of activation of the second
implant in a child who had 15.95 years of hearing experience on the right side and was deprived of
auditory input in the left ear. These measures were repeated after 1 month of bilateral implant use and

265
then again after 9 months. Responses from the latter two time points are shown in Figures 4b and 4c,
respectively. The red waveform shows the grand mean response recorded from the side with long-
term unilateral cochlear implant experience, and the blue is the cortical waveform evoked by
stimulation of the newly implanted side (naïve side). The two responses are very different from one
another at all time points. Consistent with previous findings, the cortical responses from the
experienced side (red waveform) in Figure 4 were dominated by a mature-like morphology, comprised
of the obligatory peaks P1-N1-P2-N2, similar to those expected in same aged peers with normal hearing
(Jiwani et al., 2013a). By contrast, responses recorded from the newly implanted ear (blue waveform)
were characterized by different peaks occurring with much larger amplitudes than the responses from
the side with long-term hearing experience; a large negative peak (N(ci), followed by a large positive
peak (P(ci)) can be seen (Jiwani et al., 2013b;Jiwani et al., 2013c). Little changes to either response
occurred over the first months of bilateral implant use. Slight decreases in the latencies and amplitudes
of the peaks evoked by the newly implanted ear were found after one month (Figure 4b), with almost
no change in latency, amplitude or waveform morphology thereafter. This is shown by the response
recorded at 9 months following activation of the second implant in Figure 4c.
Figure 4: Example of cortical evoked responses from an adolescent in the Jiwani et al. (2013b) and
Jiwani et al. (2013c) study cohorts. She received a right unilateral cochlear implant (red waveform)
within limited durations of bilateral deafness (3 years of age) and used it unilaterally to hear for 15.95
years. She then received a second implant in the opposite and deprived left side (naïve side) (blue
waveform). Cortical responses evoked from both implants are shown at: (A) the first day of activation
of the second implanted ear (Jiwani et al., 2013b), (B) one month after bilateral implantation (Jiwani et
al., 2013c) and (C) nine months following bilateral cochlear implant experience (Jiwani et al., 2013c).

266
The lack of cortical development evoked by stimulation of the second implanted side is in
contrast to the rapid developmental change expected to occur at early stages of unilateral cochlear
implant use in young children (Sharma et al., 2002a;Sharma and Dorman, 2006), and, rather, more
similar to the limited change reported in older children implanted after long durations of bilateral
deafness (Sharma et al., 2002b;Gordon et al., 2005;Gordon et al., 2008a). This might reflect immaturity
or abnormalities in auditory development from the second implanted side, driven by either long
duration of auditory deprivation or by maturation of the auditory cortex from unilateral cochlear
implant use. Providing a second implant to children after this period has passed may prevent the naïve
cortical pathways from developing after an important period in cortical auditory development has been
missed. The findings from our previous study (Gordon et al., 2013) (discussed above and shown in
Figure 3) suggest that there is an early sensitive period for bilateral brainstem development (exceeded
after 1.5 years of unilateral implant use) and a later cortical maturation promoted by unilateral use of
over 10 years (Jiwani et al., 2013a), as shown by the data in Figure 4 (Jiwani et al., 2013b;Jiwani et al.,
2013c). Together, these results suggest that there are multiple sensitive periods in the developing
auditory system.
Bilateral implantation within a sensitive period improves perception of
binaural timing cues
As reviewed above, several lines of investigation suggest that the potential for promoting
binaural hearing in children who are deaf will be best realized by limiting the period of bilateral deafness
and providing bilateral implants with little delay. We have been studying the perception of binaural
level and timing cues in children who received bilateral cochlear implants because these cues are
important for binaural hearing. Interaural level and timing cues arise because sounds coming from
one side of the head reach the closer ear at higher intensities and/or faster than the other ear. Level
and timing differences are coded in the auditory brainstem by the degree of inhibition (Grothe et al.,
2010).
We found that 19 children receiving one implant at 2.1 ± 1.1 years of age and the second after
4.9 ± 2.8 years of unilateral implant use can hear changes in interaural level differences but have
particularly poor abilities to detect interaural timing cues even after several years of bilateral cochlear

267
implant use (Salloum et al., 2010). Poor detection of binaural timing cues by sequentially implanted
children was surprising given evidence from a similar group showing that the auditory brainstem
integrates input from both implants as measured by the electrophysiological binaural interaction
component (Gordon et al., 2012). This measure is a calculated difference between the sum of the left
and right evoked auditory brainstem responses and the bilaterally evoked brainstem response. Peaks
in the difference response reflect inhibition occurring with binaural processing (Dobie and Berlin,
1979;Dobie and Norton, 1980;Brantberg et al., 1999). Using this difference measure, we found that
tonotopic organization is maintained in the bilateral brainstem of children who are deaf and that the
pathways continue to code interaural level cues despite development driven from one ear before the
other. There are consequences of the mismatches in development resulting from unilateral implant
use. Although the auditory brainstem codes interaural timing differences, this does not occur normally
(Gordon et al., 2008b). A miscalculation of binaural brainstem interactions results from the mismatch
in neural conduction (measured by shorter peak latencies responses from the more experienced ear).
More recent findings show that a sound arriving first to the more experienced ear by 1ms, for example,
reduces the binaural interaural component more than when it arrives first by the same amount to the
second implanted ear (Gordon, et al., in preparation). Nonetheless, coding of interaural timing remains
(albeit abnormally calibrated); thus abnormal brainstem processing cannot account for the profound
difficulties these children have detecting timing differences sent by their bilateral implants. This
suggests a deficit for interaural timing processing in more central areas of the auditory system which
likely occurred during the period before bilateral implantation. In support, the numbers of cortical
neurons specialized to respond to interaural timing cues are reduced in congenitally deaf white cats
(Tillein et al., 2010) as are numbers of neurons in auditory cortices responsible for sound localization
(Malhotra et al., 2008). In more recent work, we are asking whether binaural timing cues are better
heard by children who received bilateral cochlear implants simultaneously. Preliminary findings
suggest good potential for development of binaural hearing in children who have limited durations of
bilateral and unilateral deafness, but is compromised in children with long unilateral cochlear implants
experience (>1.5 years).

268
Conclusions
We have reviewed evidence showing that access to sound within limited durations of bilateral
deafness in early life promotes normal-like development of activity along the auditory pathways in
children who have many years of hearing experience with a unilateral cochlear implant. At the same
time, however, the unilaterally driven stimulation leaves the opposite pathways deprived of input and
susceptible to reorganization. We find that providing bilateral cochlear implants to children after a
period of unilateral deafness of longer than 1.5 years drives abnormal mismatches in activity at the
level of the brainstem and cortex. This is characterized by abnormal strengthening of activity to both
the contralateral and ipsilateral auditory cortices from the first implanted ear. These abnormalities in
auditory development are associated with more asymmetric speech perception, poorer hearing in noise,
abnormal sound localization, and an inability to identify inter-aural timing cues. These skills are
important for normal integration and processing of auditory input. We therefore suggest that binaural
hearing is compromised in children who receive bilateral cochlear implants after a period of unilateral
implant use exceeding 1.5 years. With that in mind, cochlear implants should be provided to children
early as well as bilaterally within very limited or no delays between implants (i.e., simultaneously). Our
current studies are now examining how much residual hearing is needed in the un-implanted ear to
provide a potential protective effect against unilaterally driven reorganization and whether bimodal
hearing (acoustic and electrical input) can be used to restore binaural hearing. Further, we are asking
whether the sensitive period for bilateral input can be “reopened” by attempting to strengthen
pathways from the second implanted ear to restore symmetric bilateral pathways and binaural hearing.
Our findings suggest that both bilateral and unilateral deprivation should be limited to promote
optimal binaural hearing in children who use cochlear implants, and enable them to function better
and more naturally in challenging listening situations such as the playground or classroom
environments.

269
References
Abbott, L.F., and Nelson, S.B. (2000). Synaptic plasticity: taming the beast. Nat Neurosci 3, 1178-
1183.
Auer Jr, E.T., Bernstein, L.E., Sungkarat, W., and Singh, M. (2007). Vibrotactile activation of the
auditory cortices in deaf versus hearing adults. Neuroreport 18, 645.
Basura, G.J., Eapen, R., and Buchman, C.A. (2009). Bilateral cochlear implantation: current concepts,
indications, and results. Laryngoscope 119, 2395-2401.
Batteau, D.W. (1967). The role of the pinna in human localization. Proceedings of the Royal Society of
London. Series B. Biological Sciences 168, 158-180.
Bavelier, D., Dye, M.W.G., and Hauser, P.C. (2006). Do deaf individuals see better? Trends in Cognitive
Sciences 10, 512-518.
Bavelier, D., and Neville, H.J. (2002). Cross-modal plasticity: where and how? Nat Rev Neurosci 3, 443-
452.
Bavelier, D., Tomann, A., Hutton, C., Mitchell, T., Corina, D., Liu, G., and Neville, H. (2000). Visual
attention to the periphery is enhanced in congenitally deaf individuals. Journal of Neuroscience
20, 1-6.
Beadle, E.a.R., Mckinley, D.J., Nikolopoulos, T.P., Brough, J., O'donoghue, G.M., and Archbold, S.M.
(2005). Long-term functional outcomes and academic-occupational status in implanted
children after 10 to 14 years of cochlear implant use. Otology & Neurotology 26, 1152.
Blegvad, B. (1975). Binaural Summation of Surface-Recorded Electrocochleographic Responses
Normal-hearing Subjects. Scandinavian Audiology 4, 233-238.
Bocca, E. (1955). Binaural hearing: Another approach. The Laryngoscope 65, 1164-1171.
Brantberg, K., Fransson, P.-A., Hansson, H., and Rosenhall, U. (1999). Measures of the binaural
interaction component in human auditory brainstem response using objective detection
criteria. Scandinavian Audiology 28, 15-26.

270
Brown, K.D., and Balkany, T.J. (2007). Benefits of bilateral cochlear implantation: a review. Curr
Opin Otolaryngol Head Neck Surg 15, 315.
Busse, L., Roberts, K.C., Crist, R.E., Weissman, D.H., and Woldorff, M.G. (2005). The spread of
attention across modalities and space in a multisensory object. Proc Natl Acad Sci U S A 102,
18751-18756.
Calvert, G.A., Hansen, P.C., Iversen, S.D., and Brammer, M.J. (2001). Detection of audio-visual
integration sites in humans by application of electrophysiological criteria to the BOLD effect.
Neuroimage 14, 427-438.
Calvert, G.A., and Thesen, T. (2004). Multisensory integration: methodological approaches and
emerging principles in the human brain. Journal of Physiology-Paris 98, 191-205.
Chadha, N.K., Papsin, B.C., Jiwani, S., and Gordon, K.A. (2011). Speech detection in noise and spatial
unmasking in children with simultaneous versus sequential bilateral cochlear implants. Otology
& Neurotology 32, 1057.
Ching, T.Y., Van Wanrooy, E., and Dillon, H. (2007). Binaural-bimodal fitting or bilateral
implantation for managing severe to profound deafness: a review. Trends Amplif 11, 161-192.
Cohn, E.S., and Kelley, P.M. (1999). Clinical phenotype and mutations in connexin 26
(DFNB1/GJB2), the most common cause of childhood hearing loss. American Journal of Medical
Genetics 89, 130-136.
Crowley, K.E., and Colrain, I.M. (2004). A review of the evidence for P2 being an independent
component process: age, sleep and modality. Clin Neurophysiol 115, 732-744.
Dobie, R., and Norton, S. (1980). Binaural interaction in human auditory evoked potentials.
Electroencephalography and Clinical Neurophysiology 49, 303-313.
Dobie, R.A., and Berlin, C.I. (1979). Binaural interaction in brainstem-evoked responses. Archives of
Otolaryngology—Head & Neck Surgery 105, 391.
Doucet, M., Bergeron, F., Lassonde, M., Ferron, P., and Lepore, F. (2006). Cross-modal
reorganization and speech perception in cochlear implant users. Brain 129, 3376-3383.

271
Eapen, R.J., and Buchman, C.A. (2009). Bilateral cochlear implantation: current concepts. Curr Opin
Otolaryngol Head Neck Surg 17, 351-355.
Eggermont, J.J., and Ponton, C.W. (2003). Auditory-evoked Potential Studies of Cortical Maturation
in Normal Hearing and Implanted Children: Correlations with Changes in Structure and
Speech Perception. Acta Oto-laryngologica 123, 249-252.
Eggermont, J.J., Ponton, C.W., Don, M., Waring, M.D., and Kwong, B. (1997). Maturational delays
in cortical evoked potentials in cochlear implant users. Acta Oto-laryngologica 117, 161-163.
Finney, E.M., Fine, I., and Dobkins, K.R. (2001). Visual stimuli activate auditory cortex in the deaf.
Nat Neurosci 4, 1171-1174.
Firszt, J.B., Ulmer, J.L., and Gaggl, W. (2006). Differential representation of speech sounds in the
human cerebral hemispheres. The Anatomical Record Part A: Discoveries in Molecular, Cellular, and
Evolutionary Biology 288, 345-357.
Fujiwara, N., Nagamine, T., Imai, M., Tanaka, T., and Shibasaki, H. (1998). Role of the primary
auditory cortex in auditory selective attention studied by whole-head neuromagnetometer.
Cognitive brain research 7, 99-109.
Galvin, K.L., Mok, M., and Dowell, R.C. (2007). Perceptual benefit and functional outcomes for
children using sequential bilateral cochlear implants. Ear Hear 28, 470.
García-Larrea, L., Lukaszewicz, A.C., and Mauguiére, F. (1992). Revisiting the oddball paradigm.
Non-target vs neutral stimuli and the evaluation of ERP attentional effects. Neuropsychologia
30, 723-741.
Geers, A.E., and Sedey, A.L. (2011). Language and verbal reasoning skills in adolescents with 10 or
more years of cochlear implant experience. Ear Hear 32, 39S.
Giard, M., and Peronnet, F. (1999). Auditory-visual integration during multimodal object recognition
in humans: a behavioral and electrophysiological study. Journal of Cognitive Neuroscience 11, 473-
490.

272
Giraud, A.L., and Lee, H.J. (2007). Predicting cochlear implant outcome from brain organisation in
the deaf. Restorative neurology and neuroscience 25, 381-390.
Gordon, K., Wong, D.D., Valero, J., Jewell, S., Yoo, P., and Papsin, B. (2011a). Use it or lose it?
Lessons learned from the developing brains of children who are deaf and use cochlear implants
to hear. Brain Topography 24, 204-219.
Gordon, K.A., Jiwani, S., and Papsin, B.C. (2011b). What is the optimal timing for bilateral cochlear
implantation in children? Cochlear Implants Int 12 Suppl 2, S8-14.
Gordon, K.A., and Papsin, B.C. (2009). Benefits of short interimplant delays in children receiving
bilateral cochlear implants. Otology & Neurotology 30, 319.
Gordon, K.A., Papsin, B.C., and Harrison, R.V. (2003). Activity-dependent developmental plasticity
of the auditory brain stem in children who use cochlear implants. Ear Hear 24, 485-500.
Gordon, K.A., Papsin, B.C., and Harrison, R.V. (2006). An evoked potential study of the
developmental time course of the auditory nerve and brainstem in children using cochlear
implants. Audiology and Neurotology 11, 7-23.
Gordon, K.A., Papsin, B.C., and Harrison, R.V. (2007a). Auditory brainstem activity and development
evoked by apical versus basal cochlear implant electrode stimulation in children. Clin
Neurophysiol 118, 1671-1684.
Gordon, K.A., Salloum, C., Toor, G.S., Van Hoesel, R., and Papsin, B.C. (2012). Binaural Interactions
Develop in the Auditory Brainstem of Children Who Are Deaf: Effects of Place and Level of
Bilateral Electrical Stimulation. The Journal of neuroscience 32, 4212-4223.
Gordon, K.A., Tanaka, S., and Papsin, B.C. (2005). Atypical cortical responses underlie poor speech
perception in children using cochlear implants. Neuroreport 16, 2041.
Gordon, K.A., Tanaka, S., Wong, D.D., and Papsin, B.C. (2008a). Characterizing responses from
auditory cortex in young people with several years of cochlear implant experience. Clin
Neurophysiol 119, 2347-2362.

273
Gordon, K.A., Tanaka, S., Wong, D.D., Stockley, T., Ramsden, J.D., Brown, T., Jewell, S., and Papsin,
B.C. (2011c). Multiple effects of childhood deafness on cortical activity in children receiving
bilateral cochlear implants simultaneously. Clin Neurophysiol 122, 823-833.
Gordon, K.A., Valero, J., and Papsin, B.C. (2007b). Auditory brainstem activity in children with 9-30
months of bilateral cochlear implant use. Hear Res 233, 97-107.
Gordon, K.A., Valero, J., Van Hoesel, R., and Papsin, B.C. (2008b). Abnormal timing delays in
auditory brainstem responses evoked by bilateral cochlear implant use in children. Otology &
Neurotology 29, 193.
Gordon, K.A., Wong, D.D., and Papsin, B.C. (2013). Bilateral input protects the cortex from
unilaterally-driven reorganization in children who are deaf. Brain 136, 1609-1625.
Gordon, K.A., Wong, D.D.E., and Papsin, B.C. (2010). Cortical function in children receiving
bilateral cochlear implants simultaneously or after a period of interimplant delay. Otology &
Neurotology 31, 1293.
Grady, C.L., Van Meter, J.W., Maisog, J.M., Pietrini, P., Krasuski, J., and Rauschecker, J.P. (1997).
Attention-related modulation of activity in primary and secondary auditory cortex. Neuroreport
8, 2511.
Grieco-Calub, T.M., and Litovsky, R.Y. (2010). Sound localization skills in children who use bilateral
cochlear implants and in children with normal acoustic hearing. Ear and Hearing 31, 645.
Grothe, B., Pecka, M., and Mcalpine, D. (2010). Mechanisms of sound localization in mammals.
Physiological Reviews 90, 983-1012.
Gualandi, F., Ravani, A., Berto, A., Sensi, A., Trabanelli, C., Falciano, F., Trevisi, P., Mazzoli, M.,
Tibiletti, M., and Cristofari, E. (2002). Exploring the clinical and epidemiological complexity
of GJB2‐linked deafness. American Journal of Medical Genetics 112, 38-45.
Hari, R. (1990). The neuromagnetic method in the study of the human auditory cortex. Advances in
audiology 6, 222-282.

274
Harrison, R.V., Gordon, K.A., and Mount, R.J. (2005). Is there a critical period for cochlear
implantation in congenitally deaf children? Analyses of hearing and speech perception
performance after implantation. Dev Psychobiol 46, 252-261.
Hawley, M.L., Litovsky, R.Y., and Culling, J.F. (2004). The benefit of binaural hearing in a cocktail
party: Effect of location and type of interferer. The Journal of the Acoustical Society of America 115,
833.
Hebb, D.O. (1949). "The Organization of Behavior". Wiley, New York).
Hecox, K., and Burkard, R. (2006). Developmental dependencies of the human brainstem auditory
evoked response. Annals of the New York Academy of Sciences 338, 538-556.
Hocherman, S., Benson, D., Goldstein, M., Heffner, H., and Hienz, R. (1976). Evoked unit activity
in auditory cortex of monkeys performing a selective attention task. Brain Res 117, 51-68.
Hopyan-Misakyan, T.M., Gordon, K.A., Dennis, M., and Papsin, B.C. (2009). Recognition of affective
speech prosody and facial affect in deaf children with unilateral right cochlear implants. Child
Neuropsychology 15, 136-146.
Hopyan, T., Gordon, K., and Papsin, B. (2011). Identifying emotions in music through electrical
hearing in deaf children using cochlear implants. Cochlear Implants Int 12, 21-26.
Hopyan, T., Peretz, I., Chan, L.P., Papsin, B.C., and Gordon, K.A. (2012). Children using cochlear
implants capitalize on acoustical hearing for music perception. Frontiers in psychology 3.
Jerger, J., and Hall, J. (1980). Effects of age and sex on auditory brainstem response. Archives of
Otolaryngology—Head & Neck Surgery 106, 387.
Jiwani, S., Papsin, B.C., and Gordon, K.A. (2013a). Central auditory development after long-term
cochlear implant use. Clinical Neurophysiology In Press.

275
Jiwani, S., Papsin, B.C., and Gordon, K.A. (2013b, February). Extensive areas of the cortex are evoked
by stimulation from the newly implanted ear in children who were long-term unilateral cochlear
implant users [Abstract]. 36th Annual MidWinter Meeting of the Association for Research in
Otolaryngology. Baltimore, Maryland, USA.
(http://www.aro.org/documents/ARO2013FinalAbstractBook_000.pdf).
Jiwani, S., Papsin, B.C., and Gordon, K.A. (2013c, July). Long durations of unilateral cochlear implant
use do not protect the un-stimulated pathays from effects of auditory deprivation [Abstract].
2013 Conference on Implantable Auditory Prosthesis, Lake Tahoe, California, USA.
(http://www.ciap2013.org/PDFs/CIAP2013%20Program%20Book%20draft.pdf).
Johnson, J.A., and Zatorre, R.J. (2005). Attention to simultaneous unrelated auditory and visual
events: behavioral and neural correlates. Cereb Cortex 15, 1609-1620.
Kelley, P., Harris, D., Comer, B., Askew, J., Fowler, T., Smith, S., and Kimberling, W. (1998). Novel
mutations in the connexin 26 gene (GJB2) that cause autosomal recessive (DFNB1) hearing
loss. The American Journal of Human Genetics 62, 792-799.
Klinge, C., Eippert, F., Roder, B., and Buchel, C. (2010). Corticocortical connections mediate primary
visual cortex responses to auditory stimulation in the blind. J Neurosci 30, 12798-12805.
Kral, A., Hubka, P., Heid, S., and Tillein, J. (2013). Single-sided deafness leads to unilateral aural
preference within an early sensitive period. Brain 136, 180-193.
Kral, A., and O'donoghue, G.M. (2010). Profound deafness in childhood. New England Journal of
Medicine 363, 1438-1450.
Kral, A., and Sharma, A. (2011). Developmental neuroplasticity after cochlear implantation. Trends
in Neurosciences.
Kraus, N., and Mcgee, T. (1993). Clinical implications of primary and nonprimary pathway
contributions to the middle latency response generating system. Ear Hear 14, 36.
Kraus, N., Mcgee, T., Littman, T., and Nicol, T. (1992). Reticular formation influences on primary
and non-primary auditory pathways as reflected by the middle latency response. Brain Res 587,
186-194.

276
Kraus, N., Mcgee, T., Littman, T., Nicol, T., and King, C. (1994). Nonprimary auditory thalamic
representation of acoustic change. J Neurophysiol 72, 1270-1277.
Lazard, D., Giraud, A.-L., Gnansia, D., Meyer, B., and Sterkers, O. (2011). Understanding the
deafened brain: Implications for cochlear implant rehabilitation. European Annals of
Otorhinolaryngology, Head and Neck Diseases.
Lazard, D.S., Lee, H.J., Truy, E., and Giraud, A.L. (2012). Bilateral reorganization of posterior
temporal cortices in post‐lingual deafness and its relation to cochlear implant outcome. Human
Brain Mapping.
Lee, C.C., and Winer, J.A. (2005). Principles governing auditory cortex connections. Cereb Cortex 15,
1804-1814.
Lee, D.S., Lee, J.S., Oh, S.H., Kim, S.K., Kim, J.W., Chung, J.K., Lee, M.C., and Kim, C.S. (2001).
Deafness: cross-modal plasticity and cochlear implants. Nature 409, 149-150.
Lee, H.J., Giraud, A.L., Kang, E., Oh, S.H., Kang, H., Kim, C.S., and Lee, D.S. (2007a). Cortical
activity at rest predicts cochlear implantation outcome. Cereb Cortex 17, 909-917.
Lee, H.J., Truy, E., Mamou, G., Sappey-Marinier, D., and Giraud, A.L. (2007b). Visual speech circuits
in profound acquired deafness: a possible role for latent multimodal connectivity. Brain 130,
2929-2941.
Levänen, S., and Hamdorf, D. (2001). Feeling vibrations: enhanced tactile sensitivity in congenitally
deaf humans. Neuroscience Letters 301, 75-77.
Levänen, S., Jousmäki, V., and Hari, R. (1998). Vibration-induced auditory-cortex activation in a
congenitally deaf adult. Current Biology 8, 869-872.
Liegeois-Chauvel, C., Musolino, A., Badier, J., Marquis, P., and Chauvel, P. (1994). Evoked potentials
recorded from the auditory cortex in man: evaluation and topography of the middle latency
components. Electroencephalography and Clinical Neurophysiology/Evoked Potentials Section 92, 204-
214.
Litovsky, R. (2008). Potential Advantages from Bilateral Cochlear Implants. White Paper Series.

277
Litovsky, R.Y., Johnstone, P.M., and Godar, S.P. (2006). Benefits of bilateral cochlear implants
and/or hearing aids in children. Int J Audiol 45, 78-91.
Litovsky, R.Y., Parkinson, A., Arcaroli, J., Peters, R., Lake, J., Johnstone, P., and Yu, G. (2004).
Bilateral cochlear implants in adults and children. Archives of Otolaryngology- Head and Neck
Surgery 130, 648.
Lomber, S.G., Meredith, M.A., and Kral, A. (2010). Cross-modal plasticity in specific auditory cortices
underlies visual compensations in the deaf. Nat Neurosci 13, 1421-1427.
Lorenzi, C., Gatehouse, S., and Lever, C. (1999). Sound localization in noise in normal-hearing
listeners. The Journal of the Acoustical Society of America 105, 1810.
Malhotra, S., Stecker, G.C., Middlebrooks, J.C., and Lomber, S.G. (2008). Sound localization deficits
during reversible deactivation of primary auditory cortex and/or the dorsal zone. Journal of
Neurophysiology 99, 1628-1642.
Merabet, L.B., and Pascual-Leone, A. (2010). Neural reorganization following sensory loss: the
opportunity of change. Nat Rev Neurosci 11, 44-52.
Meredith, M.A., and Lomber, S.G. (2011). Somatosensory and visual crossmodal plasticity in the
anterior auditory field of early-deaf cats. Hear Res 280, 38-47.
Moller, A.R., and Rollins, P.R. (2002). The non-classical auditory pathways are involved in hearing in
children but not in adults. Neuroscience letters 319, 41-44.
Nicholas, J.G., and Geers, A.E. (2007). Will they catch up? The role of age at cochlear implantation
in the spoken language development of children with severe to profound hearing loss. Journal
of Speech, Language, and Hearing Research 50, 1048.
Nishimura, H., Hashikawa, K., Doi, K., Iwaki, T., Watanabe, Y., Kusuoka, H., Nishimura, T., and
Kubo, T. (1999). Sign language ‘heard’in the auditory cortex. Nature 397, 116-116.
O'neil, J.N., Limb, C.J., Baker, C.A., and Ryugo, D.K. (2010). Bilateral effects of unilateral cochlear
implantation in congenitally deaf cats. The Journal of comparative neurology 518, 2382-2404.

278
Osberger, M.J., Zimmerman-Phillips, S., and Koch, D.B. (2002). Cochlear implant candidacy and
performance trends in children. The Annals of otology, rhinology & laryngology. Supplement 189, 62.
Pandya, D.N., and Yeterian, E.H. (1985). Architecture and connections of cortical association areas.
Cerebral Cortex 4, 3-61.
Papsin, B.C., and Gordon, K.A. (2007). Cochlear implants for children with severe-to-profound
hearing loss. New England Journal of Medicine 357, 2380-2387.
Peters, B.R., Litovsky, R., Parkinson, A., and Lake, J. (2007). Importance of age and postimplantation
experience on speech perception measures in children with sequential bilateral cochlear
implants. Otology & Neurotology 28, 649.
Picton, T., and Hillyard, S. (1974). Human auditory evoked potentials. II: Effects of attention.
Electroencephalography and clinical neurophysiology 36, 191-200.
Ponton, C.W., and Eggermont, J.J. (2001). Of kittens and kids: altered cortical maturation following
profound deafness and cochlear implant use. Audiology and Neurotology 6, 363-380.
Ponton, C.W., Eggermont, J.J., Kwong, B., and Don, M. (2000). Maturation of human central auditory
system activity: evidence from multi-channel evoked potentials. Clinical Neurophysiology 111,
220-236.
Posner, M.I., and Dehaene, S. (1994). Attentional networks. Trends Neurosci 17, 75-79.
Propst, E.J., Papsin, B.C., Stockley, T.L., Harrison, R.V., and Gordon, K.A. (2006). Auditory
responses in cochlear implant users with and without GJB2 deafness. The Laryngoscope 116,
317-327.
Razak, K.A., Zumsteg, T., and Fuzessery, Z.M. (2009). Development of auditory thalamocortical
connections in the pallid bat, Antrozous pallidus. J Comp Neurol 515, 231-242.
Read, H.L., Winer, J.A., and Schreiner, C.E. (2002). Functional architecture of auditory cortex. Curr
Opin Neurobiol 12, 433-440.

279
Rif, J., Hari, R., Hämäläinen, M.S., and Sams, M. (1991). Auditory attention affects two different areas
in the human supratemporal cortex. Electroencephalography and Clinical Neurophysiology 79, 464-
472.
Salamy, A. (1984). Maturation of the auditory brainstem response from birth through early childhood.
Journal of Clinical Neurophysiology 1, 293-329.
Salamy, A., and Mckean, C.M. (1976). Postnatal development of human brainstem potentials during
the first year of life. Electroencephalography & Clinical Neurophysiology; Electroencephalography &
Clinical Neurophysiology.
Salloum, C.a.M., Valero, J., Wong, D.D.E., Papsin, B.C., Van Hoesel, R., and Gordon, K.A. (2010).
Lateralization of interimplant timing and level differences in children who use bilateral cochlear
implants. Ear Hear 31, 441.
Sandmann, P. (2012). User Research Visual processing in the auditory cortex of cochlear-implant
users. Brain 42.
Sandmann, P., Dillier, N., Eichele, T., Meyer, M., Kegel, A., Pascual-Marqui, R.D., Marcar, V.L.,
Jäncke, L., and Debener, S. (2012). Visual activation of auditory cortex reflects maladaptive
plasticity in cochlear implant users. Brain 135, 555-568.
Seeber, B.U., and Fastl, H. (2008). Localization cues with bilateral cochlear implants. The Journal of
the Acoustical Society of America 123, 1030.
Sharma, A., and Dorman, M. (2006). Central auditory development in children with cochlear implants:
clinical implications. Advances in Oto Rhino Laryngology 64, 66-88.
Sharma, A., Dorman, M.F., and Spahr, A.J. (2002a). Rapid development of cortical auditory evoked
potentials after early cochlear implantation. Neuroreport 13, 1365.
Sharma, A., Dorman, M.F., and Spahr, A.J. (2002b). A sensitive period for the development of the
central auditory system in children with cochlear implants: implications for age of implantation.
Ear Hear 23, 532-539.

280
Song, S., Miller, K.D., and Abbott, L.F. (2000). Competitive Hebbian learning through spike-timing-
dependent synaptic plasticity. Nat Neurosci 3, 919-926.
Starr, A., Amlie, R., Martin, W., and Sanders, S. (1977). Development of auditory function in newborn
infants revealed by auditory brainstem potentials. Pediatrics 60, 831-839.
Steffens, T., Lesinski-Schiedat, A., Strutz, J., Aschendorff, A., Klenzner, T., Ruhl, S., Voss, B., Wesarg,
T., Laszig, R., and Lenarz, T. (2008a). The benefits of sequential bilateral cochlear
implantation for hearing-impaired children. Acta Otolaryngol 128, 164-176.
Steffens, T., Lesinski-Schiedat, A., Strutz, J., Aschendorff, A., Klenzner, T., Rühl, S., Voss, B., Wesarg,
T., Laszig, R., and Lenarz, T. (2008b). The benefits of sequential bilateral cochlear
implantation for hearing-impaired children. Acta Oto-laryngologica 128, 164-176.
Strelnikov, K., Rouger, J., Demonet, J.-F., Lagleyre, S., Fraysse, B., Deguine, O., and Barone, P. (2010).
Does brain activity at rest reflect adaptive strategies? Evidence from speech processing after
cochlear implantation. Cerebral Cortex 20, 1217-1222.
Tervaniemi, M., and Hugdahl, K. (2003). Lateralization of auditory-cortex functions. Brain Research
Reviews 43, 231-246.
Tillein, J., Hubka, P., Syed, E., Hartmann, R., Engel, A.K., and Kral, A. (2010). Cortical representation
of interaural time difference in congenital deafness. Cereb Cortex 20, 492-506.
Tremblay, K., Kraus, N., Mcgee, T., Ponton, C., and Otis, B. (2001). Central auditory plasticity:
changes in the N1-P2 complex after speech-sound training. Ear and Hearing 22, 79-90.
Tremblay, K.L. (Year). "Training-related changes in the brain: evidence from human auditory-evoked
potentials", in: Seminars in Hearing: Thieme Medical Publishers Inc., 120.
Tremblay, K.L., and Kraus, N. (2002). Auditory training induces asymmetrical changes in cortical
neural activity. Journal of Speech, Language and Hearing Research 45, 564.
Tremblay, K.L., Shahin, A.J., Picton, T., and Ross, B. (2009). Auditory training alters the physiological
detection of stimulus-specific cues in humans. Clinical neurophysiology: official journal of the
International Federation of Clinical Neurophysiology 120, 128.

281
Van Deun, L., Van Wieringen, A., Scherf, F., Deggouj, N., Desloovere, C., Offeciers, F.E., Van De
Heyning, P.H., Dhooge, I.J., and Wouters, J. (2009a). Earlier intervention leads to better
sound localization in children with bilateral cochlear implants. Audiology and Neurotology 15, 7-
17.
Van Deun, L., Van Wieringen, A., Van Den Bogaert, T., Scherf, F., Offeciers, F.E., Van De Heyning,
P.H., Desloovere, C., Dhooge, I.J., Deggouj, N., and De Raeve, L. (2009b). Sound
localization, sound lateralization, and binaural masking level differences in young children with
normal hearing. Ear and Hearing 30, 178-190.
Van Hoesel, R.J., and Tyler, R.S. (2003). Speech perception, localization, and lateralization with
bilateral cochlear implants. The Journal of the Acoustical Society of America 113, 1617.
Van Wanrooij, M.M., and Van Opstal, A.J. (2004). Contribution of head shadow and pinna cues to
chronic monaural sound localization. The Journal of neuroscience 24, 4163-4171.
Webster, K.E., and Colrain, I.M. (2000). The respiratory-related evoked potential: effects of attention
and occlusion duration. Psychophysiology 37, 310-318.
Winer, J.A., Diehl, J.J., and Larue, D.T. (2001). Projections of auditory cortex to the medial geniculate
body of the cat. J Comp Neurol 430, 27-55.
Winer, J.A., Miller, L.M., Lee, C.C., and Schreiner, C.E. (2005). Auditory thalamocortical
transformation: structure and function. Trends Neurosci 28, 255-263.
Wong, D.D.E., and Gordon, K.A. (2009). Beamformer suppression of cochlear implant artifacts in
an electroencephalography dataset. Biomedical Engineering, IEEE Transactions on 56, 2851-2857.
Zatorre, R.J., and Belin, P. (2001). Spectral and temporal processing in human auditory cortex.
Cerebral Cortex 11, 946-953.
Zatorre, R.J., Belin, P., and Penhune, V.B. (2002). Structure and function of auditory cortex: music
and speech. Trends in Cognitive Sciences 6, 37-46.

282
Copyright Acknowledgements
Peer-Reviewed Publications
Papers deriving from this thesis have been published in:
Gordon, K. A., Jiwani, S., & Papsin, B. C. (2013). Benefits and detriments of unilateral cochlear
implant use on bilateral auditory development in children who are deaf. Frontiers in
Psychology, 4(719): 1-14.
Jiwani, S., Papsin, B. C., & Gordon, K. A. (2013). Central auditory development after long-term
cochlear implant use. Clinical Neurophysiology, 124(9), 1868-1880.
Chapter 4 has been submitted for publication in Human Brain Mapping and is in review:
Jiwani, S., Papsin, B. C., & Gordon, K. A. Early unilateral cochlear implantation promotes mature
cortical asymmetries in adolescents who are deaf
Chapter 5 has been submitted for publication Cerebral Cortex and is in review:
Jiwani, S., Doesburg, S. M., Papsin, B. C., & Gordon, K. A. Temporally coordinated activity in the
brain is promoted by long-term unilateral cochlear implant use in adolescents
Figure 2.9 and Figure 3.2 have been published in:
Jiwani, S. & Gordon, K. A. (2012). “Figure 9-1”, Chapter 9 - p.134 in Objective Measures in Cochlear
Implants, Michelle L. Hughes, Editor. Plural Publishing Inc
Jiwani, S., & Gordon, K. A. (2012). “Figure 10-2”, Chapter 10 - p.142 in Objective Measures in
Cochlear Implants, Michelle L. Hughes, Editor. Plural Publishing Inc

283
Peer-Reviewed Presentations
Papers deriving from this thesis have been presented at the following conferences:
Jiwani, S., Doesburg, S. M., Papsin, B. C. and Gordon, K. A. (October 2014). Unilaterally driven
cortical maturation leads to lasting asymmetries in the bilateral auditory pathways in adolescents
who are deaf and use one cochlear implant. 8th International Symposium on Objective
Measures in Auditory Implants, Toronto-ON, Canada. Podium (Student Travel Award)
Jiwani, S., Doesburg, S. M., Papsin, B. C. and Gordon, K. A. (February 2014). Temporally
coordinated activity in the brain is promoted by long-term cochlear implant use in children.
Association for Research in otolaryngology 37th MidWinter Meeting, San Diego-CA,
USA (NIH NIDCD Audiologist Travel Award; The Hospital for Sick Children Travel Award)
Gordon, K. A., Wong, D., Jiwani, S., De Vreede, D. and Papsin, B. C. (February 2014).
Developmental consequences of unilateral stimulation/deprivation in children using one
cochlear implant. Association for Research in otolaryngology 37th MidWinter Meeting,
San Diego-CA, USA. Podium
Gordon, K. A., Jiwani, S., Wong, D. and Papsin, B. C. (November 2013). Does unilateral cochlear
implant use promote cortical development at the expense of pathways from the unstimulated
ear?. 1st Global Otology Research Forum, Antalya, Turkey. Podium
Jiwani, S. (October 2013). Understanding brain responses in adolescents who are new bilateral
cochlear implant users. 4th Annual MidWest Conference Miniconference on Cochlear
Implants, Madison-WI, USA. Podium
Jiwani, S., Papsin, B. C. and Gordon, K. A. (July 2013). Long durations of unilateral cochlear
implant use do not protect the un-stimulated pathways from effects of auditory deprivation.
Conference on Implantable Auditory Prosthesis, Lake Tahoe-CA, USA (NIH NIDCD
Podium Presenter Travel Award)

284
Jiwani, S. (May 2013). Long durations of deafness and use of sign language do not promote auditory
development in adolescents who are unilateral cochlear implant users. Percy Ireland Day,
University of Toronto, Department of Otolaryngology, Toronto – ON. Podium (Best
Presentation Award - Graduate Students Category)
Jiwani, S., Wong, D. D. E., Papsin, B. C. and Gordon, K. A. (April 2013). Does a sensitive period
exist for the development of the bilateral auditory pathways in cochlear implant users?
American Academy of Audiology – Academy Research Conference, Anaheim-CA, USA.
Podium (NIH NIDCD Podium Presenter Travel Award - Ranked 1st)
Jiwani, S., Papsin, B. C. and Gordon, K. A. (February 2013). Extensive areas of the cortex are
evoked by stimulation from the newly implanted ear in children who were long-term unilateral
cochlear implant users. Association for Research in otolaryngology 36th MidWinter
Meeting, Baltimore-MD, USA (The Hospital for Sick Children Travel Award; NIH NIDCD
Audiologist Travel Award; NIH NIDCD Graduate Student Travel Award)
Jiwani, S., Tanaka, S., Papsin, B. C. and Gordon, K. A. (October 2012). “Auditory development
after cochlear implantation in children who use sign language as their primary mode of
communication”. Canadian Academy of Audiology Conference, Ottawa – ON. Podium
(University of Toronto School of Graduate Studies Conference Grant)
Jiwani, S. (May 2012). Stimulation from the newly implanted ear in children who were long-term
unilateral cochlear implant users drives diffuse activity in the brain. Percy Ireland Day,
University of Toronto, Toronto – ON. Podium
Jiwani, S., Papsin, B. C. and Gordon, K. A. (July 2011). Central auditory development after long-
term cochlear implant use. 13th International Symposium on Cochlear Implants in
Children, Chicago – Illinois, USA (NIH NIDCD Mentored Doctoral Student Award)
Jiwani, S., Valero, J., Jewell, S. Papsin, B. C. and Gordon, K. A. (September 2010). Electrically
evoked middle latency responses at initial bilateral cochlear implant use. 6th International
Symposium on Objective Measures in Auditory Implants, Washington University Medical
Center, St. Louis – Missouri, USA (NIH NIDCD Student Poster Travel Award)


















