
Lymphoid organ virus of Penaeus monodon from Australia · DISEASES OF AQUATIC ORGANISMS Dis aquat...
8
DISEASES OF AQUATIC ORGANISMS Dis aquat Org Published October 26 Lymphoid organ virus of Penaeus monodon from Australia ' Department of Parasitology, Department of Microbiology, The University of Queensland, Brisbane. Queensland 4072, Australia ABSTRACT: Subadult Penaeus monodon shrimp were sampled from 4 farms in Queensland, Australia. Histological observations on lymphoid organs showed the formation of abnormal cell foci, which resembled tubules lacking a central hemolymph vessel. These formations contained hypertrophied nuclei with marginated chromatin, vacuolated cells and inclusion bodies which stained positive with Feulgen's reaction. Male and female F! monodon broodstock were sampled from 1 farm and showed the same abnormal histological features within the lymphoid organ as the subadults. Electron micro- scopy revealed tightly enveloped, cylindrical particles, 163-200 nm X 36-63 nm, packed into paracrys- talline arrays within the cytoplasm and infrequently within the nucleus. Arrays of virions were also observed within the cytoplasm of broodstock gill cells. Cross-sections of the particles revealed electron dense nucleic acid cores and envelopes. Nucleocapsids, 83-590 nm X 13-15 nm, were seen free and in vesicles within the cytoplasm. A negatively stained preparation of partially purified lymphoid organ revealed a nucleocapsid with one conical end. Lymphoid organ virus (LOV) resembles yellow-head virus (YHV) from Thailand in its morphology and cytopathology and rhabdovirus of penaeid shrimp (RPS) from the Americas in its cytopathology. KEY WORDS: Lymphoid organ virus (LOV) . Yellow-head virus (YHV) - Bacilliform . Asymptomatic INTRODUCTION At least 6 viruses have been described from the lym- phoid organ of penaeid shrimp. Infectious hypodermal and hematopoietic necrosis virus (IHHNV) was first described by Lightner et al. (1983) from Penaeus stylirostris, P. vannarnei and P. rnonodon and was recently reported from a hybrid shrimp raised in a lab- oratory in Queensland, Australia (Owens et al. 1992). Owens et al. (1991) described lymphoidal parvovirus- like virus (LPV), a virus which appears to be present exclusively in the lymphoid organ of F! monodon subadults. Lymphoid organ vacuolization virus (LOW), a togavirus, has been partially characterised from abnormal lymphoid organs of P vannamei (Bonami et al. 1992). Flegel et al. (1992) reported reovirus-like particles from the lymphoid organs of broodstock prawns in Thailand. The causative agent of yellow head disease in Penaeus monodon of Thailand has been identified as a bacilliform virus (YHV) which infects the lymphoid organ, gills and other organs (Boonyaratpalin et al. 1993).The genome is RNA and therefore YHV may be a rhabdovirus or a coronavirus (Wongteerasupaya et al. 1995). This virus is one of the most virulent disease agents so far reported for l? monodon, causing massive mortalities among subadult prawns within 3 d of first diagnosis (Chantanachookin et al. 1993). Rhabdovirus of penaeid shrimp (RPS)was described by Lu et al. (1991) for Penaeus stylirostris and F! vannamei infected with IHHNV. This virus appears to occur exclusively in the lymphoid organ. Prawns infected with RPS alone show no signs of clinical disease. The virus described in this paper, which we have called lymphoid organ virus (LOV), shares histopatho- logical and cytopathological features with YHV and RPS. O lnter-Research 1995 Resale of full artjcle not permitted
Transcript of Lymphoid organ virus of Penaeus monodon from Australia · DISEASES OF AQUATIC ORGANISMS Dis aquat...

DISEASES OF AQUATIC ORGANISMS Dis aquat Org Published October 26
Lymphoid organ virus of Penaeus monodon from Australia
' Department of Parasitology, Department of Microbiology, The University of Queensland, Brisbane. Queensland 4072, Australia
ABSTRACT: Subadult Penaeus monodon shrimp were sampled from 4 farms in Queensland, Australia. Histological observations on lymphoid organs showed the formation of abnormal cell foci, which resembled tubules lacking a central hemolymph vessel. These formations contained hypertrophied nuclei with marginated chromatin, vacuolated cells and inclusion bodies which stained positive with Feulgen's reaction. Male and female F! monodon broodstock were sampled from 1 farm and showed the same abnormal histological features within the lymphoid organ as the subadults. Electron micro- scopy revealed tightly enveloped, cylindrical particles, 163-200 nm X 36-63 nm, packed into paracrys- talline arrays within the cytoplasm and infrequently within the nucleus. Arrays of virions were also observed within the cytoplasm of broodstock gill cells. Cross-sections of the particles revealed electron dense nucleic acid cores and envelopes. Nucleocapsids, 83-590 nm X 13-15 nm, were seen free and in vesicles within the cytoplasm. A negatively stained preparation of partially purified lymphoid organ revealed a nucleocapsid with one conical end. Lymphoid organ virus (LOV) resembles yellow-head virus (YHV) from Thailand in its morphology and cytopathology and rhabdovirus of penaeid shrimp (RPS) from the Americas in its cytopathology.
KEY WORDS: Lymphoid organ virus (LOV) . Yellow-head virus (YHV) - Bacilliform . Asymptomatic
INTRODUCTION
At least 6 viruses have been described from the lym- phoid organ of penaeid shrimp. Infectious hypodermal and hematopoietic necrosis virus (IHHNV) was first described by Lightner et al. (1983) from Penaeus stylirostris, P. vannarnei and P. rnonodon and was recently reported from a hybrid shrimp raised in a lab- oratory in Queensland, Australia (Owens et al. 1992). Owens et al. (1991) described lymphoidal parvovirus- like virus (LPV), a virus which appears to be present exclusively in the lymphoid organ of F! monodon subadults. Lymphoid organ vacuolization virus (LOW), a togavirus, has been partially characterised from abnormal lymphoid organs of P vannamei (Bonami et al. 1992). Flegel et al. (1992) reported reovirus-like particles from the lymphoid organs of broodstock prawns in Thailand.
The causative agent of yellow head disease in Penaeus monodon of Thailand has been identified as a bacilliform virus (YHV) which infects the lymphoid organ, gills and other organs (Boonyaratpalin et al. 1993). The genome is RNA and therefore YHV may be a rhabdovirus or a coronavirus (Wongteerasupaya et al. 1995). This virus is one of the most virulent disease agents so far reported for l? monodon, causing massive mortalities among subadult prawns within 3 d of first diagnosis (Chantanachookin et al. 1993).
Rhabdovirus of penaeid shrimp (RPS) was described by Lu et al. (1991) for Penaeus stylirostris and F! vannamei infected with IHHNV. This virus appears to occur exclusively in the lymphoid organ. Prawns infected with RPS alone show no signs of clinical disease.
The virus described in this paper, which we have called lymphoid organ virus (LOV), shares histopatho- logical and cytopathological features with YHV and RPS.
O lnter-Research 1995 Resale of full artjcle not permitted

128 Dis aquat Org 23: 127-134. 1995
MATERIALS AND METHODS
Source and preparation of specimens. Forty sub- adult Penaeus monodon prawns, weighing between 10 and 30 g, were collected from 4 farms in Queens- land, Australia, 10 from each farm. Two male and eight female broodstock prawns, weighing between 96 and 130 g , were sampled from one of the farms. The cephalothorax of the prawns was separated from the abdomen, and split longitudinally. One longitudinal section was fixed in Davidson's fixative and processed for histology. Sections were stained using Feulgen's DNA reaction (Humason 1968).
Electron microscopy. The remaining portions of the lymphoid organ, gills, heart, nerve cord, hepato- pancreas and hematopoietic tissue were diced and fixed in 2.5% gluteraldehyde/2% paraformaldehyde in cacodylate buffer and postfixed in 1 % osmium tetroxide. The tissue was dehydrated through a graded series of alcohol and mounted in Spurr's resin (Spurr 1969). Sections were cut at 50 nm, stained with uranyl acetate/?O% methanol and Reynold's lead cit- rate and viewed at 80 kV under Phillips 400T and Hitachi H-800 transmission electron microscopes (TEM) .
Virus purification. Lymphoid organs (LOS) (1.21 g) were dissected from 50 subadult prawns collected from one of the farms where infected prawns had been found. The LOS were immediately placed onto dry ice and stored at -70°C for 1 wk. Lymphoid organs were homogenized on ice in 40 m1 0.2 M NaC1/0.02 M Tris- HC1, pH 7.4 (NT), using a 7 m1 Corning sintered glass homogenizer. The homogenate was clarified at 2500 X g for 5 min at 4°C. The supernatant was then centrifuged in the same centrifuge at 48400 X g for 45 min at 4°C to pellet the virions. The pellet was resuspended in 1 m1 NT and centrifuged on a sucrose cushion, consisting of 1 m1 60% (w/w) sucrose in NT beneath 10 m1 20% (w/w) sucrose in NT at 103 000 X g for 2 h at 4°C in a Beckman L8-M ultracentrifuge. The bands that formed at the interface were collected, diluted 5-fold and then pelleted at 208 500 X g for 1 h at 4°C. Pellets were resuspended in 500 p1 NT, and 5 p1 pipetted onto a Formvar coated Cu-400 mesh grid, which was stained with 2 % sodium phosphotungstic acid (pH 6.8) and examined under a Hitachi H-800 electron microscope.
RESULTS
Histology
Normal lymphoid organs are composed of tubules with a central hemolymph vessel surrounded by a stromal matrix of cells (Bell & Lightner 1988). The lymphoid organs of all prawns from the 4 farms con- tained abnormal foci of cells, which resembled tubules lacking a central hemolymph vessel (Fig. 1). These areas counter-stained light green with Feulgen's reac- tion whereas normal tubules stained a darker green. The cells within these areas contained hypertrophied nuclei (diameter approximately 1.3 times that of nor- mal nuclei; Fig. 2) with marginated chromatin and large vacuoles. Evident in the Feulgen's preparations were globose, cytoplasmic inclusion bodies which stained positive (red), suggesting the presence of DNA (Fig. 2). It was difficult to distinguish these inclusions from pyknotic nuclei, except in cells where the inclu- sion was directly adjacent to the nucleus (Fig. 2) . These bodies occurred within the abnormal cell foci. All other organs showed normal histology. Pyknotic nuclei were occasionally seen in the gills, the wall of the oesopha- gus and the hepatopancreas of the subadults.
Electron microscopy
Cylindrical enveloped virus particles, 163 to 200 nm long and 36 to 63 nm wide, were found in the cytoplasm of some lyrnphoid organ cells, packed into paracrystalhe
The sampled prawns, subadults and broodstock, Fig. 1 Penaeus monodon. Lymphoid organ of a subadult
showed signs of 'linica1 disease and the p0pula- infected with LOV. Infected lymphoid organs contain foci of tions were not suffering mortalities at the time of cells (F) which resemble normal tubules (T) lacking a central sampling. hemolymph vessel (H). Scale bdr = 10 pm. Feulgen's reaction

Spann et al.: LOV of Penaeus monodon from Australia
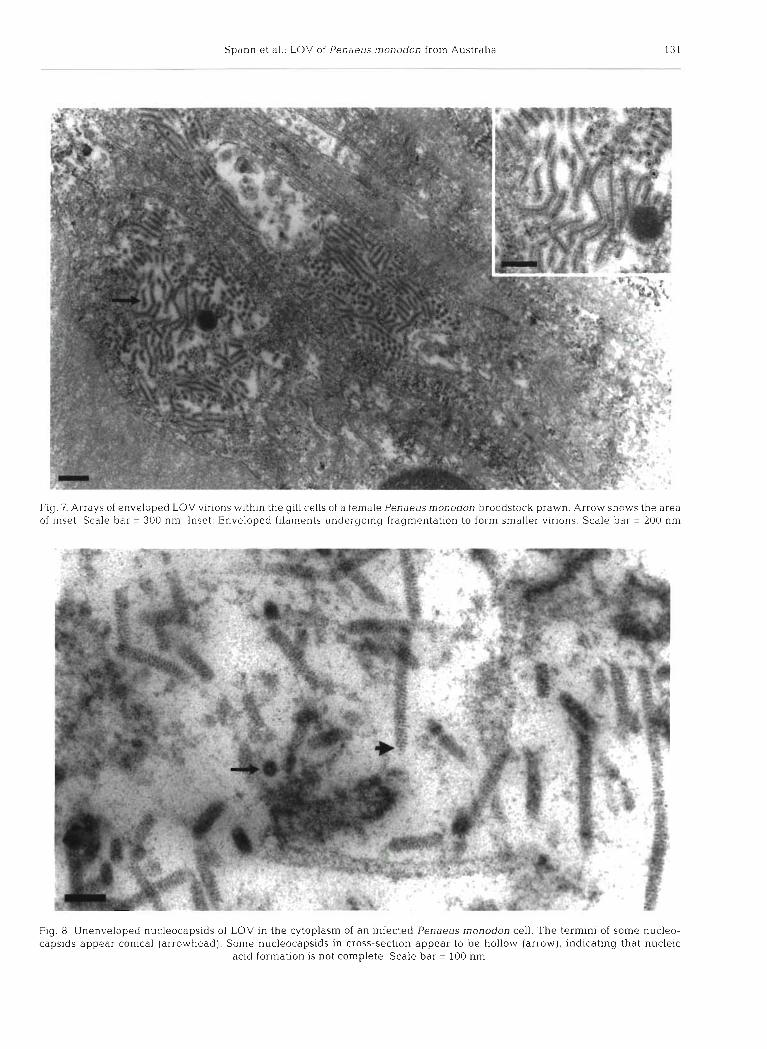
at the plasma membrane (Fig. 6). Virions had an electron dense nucleic acid core (12 to 22 nm in diameter) sur- rounded by a translucent layer (3 to 11 nm thick) and electron dense envelope (12 to 23 nnl thick) (Figs. 4 , 5 & 6). Enveloped virions were seen in the lyrnphoid organ and gills (Fig. 7) of both male and female broodstock. Long enveloped filaments which appeared to be under- going fragmentation were observed (Fig. 7 inset).
Unenveloped filaments, 83 to 290 nm long and 13 to 15 nm wide, occurred free in the cytoplasm (Fig. 8). The termini of some nucleocapsids appeared conical. Some of the nucleocapsid cores appeared hollow, sug- gesting that packing of nucleic acid into the capsids was not complete. Filaments were observed in associa- tion with endoplasmic reticulum (Fig. 9). Enveloped virions were occasionally seen along the inner surface of nuclear membranes (Fig. 10 and inset A) while un- enveloped nucleocapsids occurred outside the mem- branes (Fig. 10 and inset B).
Fig. 2. Penaeus monodon. Higher magnification of a focus of cells with hypertrophied nuclei (N) , vacuoles (V) and cyto- plasmic inclusion bodies (arrow). Scale bar = 4 pm Feulgen's
reaction
arrays (Fig. 3). These arrays were often associated with vesicles of viral material (Fig. 4). Virus particles were not observed in other organs of the subadult prawns. The nucleocapsids within the envelopes appeared striated (Fig. 5). The number of virions within an array varied from 3 to over 60 virions. Groups of virions were observed
Virus purification
Material collected at the sucrose cushion interface contained cell debris and a cylindrical, tightly enveloped particle (161 nm X 45 nm), with surface pro- jections up to 8 nm in length (Fig. 11). A cylindrical structure (145 nm X 25 nm), discernible within the envelope, possessed surface striations similar to those of unenveloped nucleocapsids (Fig. 8) and appeared to have one conical end.
Fig. 3. An LOV lntected Penaeus rnonodon cell showing paracrystalline arrays of enveloped virions w~thin the cytoplasm. Scale bar = 200 nm

130 Dis aquat Org 23 127-134, 1995
Fig. 4 . Enveloped virions of LOV (infecting Penaeus monotlvrll 111 paracrystalline arrays which are associated with vesicles (V) of viral mdterial and are surrounded by unenveloped virions free in the cytoplasm (arrow). Scale bar = 100 nm
Fig. 5. A small array of LOV virions contained within a vesicle in the cytoplasm of a n infected lymphoid organ cell of Fig. 6 . An array of LOV virions (infecting Penaeus rnonodon) Penaeus monodon. The striations of the nucleocapsid are which appears to be budding through the plasma membrane
evident (arrow). Scale bar = 50 nm (arrow) into an adjacent cell. Scale bar = 100 nm
Spann et al.: LOV of Penaeus xnonodon from Australia 131
- _ . . -7
" ' ' *--r=: - - L%. 1.6 . .
P
.:.:.R .'
-+if- ' $%%',g *.*G ,: .- 7 .
? , . . - . ..h. &g , . '9, c** - , '5 E."
'-l , i,. .ei L '
Fig. 7. Arrays of enveloped LOV virions within the gill cells of a female Penaeusrnonodon broodstock prawn. Arrow shows the area of inset. Scale bar = 300 nm. Inset: Enveloped filaments undergoing fragmentation to form smaller virions. Scale bar = 200 nm
Fig. 8. Unenveloped nucleocapsids of LOV in the cytoplasm of an infected Penaeus rnonodon cell. The termini of some nucleo- capsids appear conical (arrowhead). Some nucleocapsids in cross-section appear to be hollow (arrow), indicating that nucleic
acid formation is not complete. Scale bar = 100 nm

Dis aquat Org 23: 127-134, 1995
DISCUSSION
Fig. 9. Unenveloped filaments associated with endoplasmic reticulum (E) within an infected lymphoid organ cell of
Penaeus monodon. Scale bar = 100 nm
The results indicate the presence of a previously undescribed bacilliform virus which occurs primarily in the lymphoid organs of Penaeus monodon prawns farmed in Queensland. The histopathological features associated with this virus (cell foci resembling tubules lacking a central hemolymph vessel, hypertrophied nuclei with marginated chromatin, large cytoplasmic vacuoles and Feulgen-positive, cytoplasmic inclusion bodies) are also associated with other lymphoid organ viruses, LOVV (Bonami et al. 1992), LPV (Owens et al. 1991), a reovirus and YHV (Flegel et al. 1992) and RPS (Lu et al. 1991).
The presence of cytoplasmic inclusion bodies, which stain positive with Feulgen's reaction, suggests the presence of a DNA-containing virus. However, repli- cation and virion accumulation appear to occur in the cytoplasm, which is typical of RNA viruses and few DNA viruses (Franki et al. 1991). The genome of this virus has not yet been characterised.
The virus described here is reminiscent of rhabdo- viruses in size and morphology. The virion supports short surface projections and may be bullet shaped as there is a suggestion that the nucleocapsid has a
Fig. 10. Nucleus of an infected lyrnphoid organ cell of Penaeus monodon. Scale bar = 600 nm. Arrows show the areas of insets. Inset A: Enveloped virions (arrow) within the nuclear membrane. Scale bar = 400 nm. Inset B: Unenveloped vitions (arrow)
outside the nuclear membrane. Scale bar = 200 nm

Spann et a1 LOV of Penae ! L I S monodon from Austral~a 133
Fig. 11. Cylindrical, enveloped particle (purified from Penaeus monodon lymphoid organs) with apparent projections, striated nucleocapsid and one conical end (arrow). Scale bar = 50 nm
conical end. The nucleocapsid is of helical symmetry (Franki et al. 1991). Rhabdovirus particles may be as long as those seen in Fig. 8. The accumulation of the virus within vesicles in the cytoplasm is typical of slowly maturing rhabdoviruses (Brun 1991). Replica- tion appears to occur in the cytoplasm, however there is evidence of some nuclear involvement which is atypical of rhabdoviruses.
LOV is similar in its ultrastructural and cytopatho- logical features to YHV of Thailand and in its cytopathological features to RPS of the Americas. The virus described here and YHV both infect Penaeus monodon subadults and involve the lymphoid organ and gills. Paracrystalline arrays of virions within the cytoplasm are characteristic of both viruses, and the virions are similar in size [ l73 + 13 nm X 44 * 6 nm for enveloped YHV (Chantanachookin et al. 1993) and 163-200 nm X 36-62 nm for LOV). The abundant fila- mentous material, possibly nucleic acid cores, is char- acteristic of both viruses.
The partially purified enveloped particle is similar in appearance to purified YHV particles (Wongteera- supaya et al. 1995). Viral envelopes appear to be acquired by passage through the rough endoplasmic reticulum which then forms a vesicle around the viri- ons. This process of envelope acquisition has been sug- gested for YHV by Chantanachookin et al. (1993).
A notable difference between this virus and YHV is the lack of any obvious effect on the health of the shrimp, as YHV is one of the most highly pathogenic prawn viruses with mortalities reaching 100% (Chan- tanachookin et al. 1993). However, Flegel et al. (1992) observed cytoplasmic, enveloped, rod-shaped virions
identical to YHV in the lymphoid organs of outwardly healthy broodstock and Chantanachookin et al. (1993) have suggested that this infection is the latent asymp- tomatic form of YHV. Particles apparently identical to LOV were observed in Queensland broodstock. I t is possible that YHV may have arisen from an asympto- matic virus similar to LOV.
LOV resembles RPS in that prawns infected with RPS show no overt signs of disease (Lu & Loh 1992). RPS appears to exclusively infect the lymphoid organ and has the previously mentioned characteristics typi- cal of rhabdoviruses. The primary site of budding of rhabdoviruses is the cellular membrane, however virions of LOV and RPS also bud from cytoplasmic vesicular sites.
Acknowledgements. Mr R. Webb of the Centre for Micro- scopy and Microanalysis. The University of Queensland, is thanked for his interpretation of photomicrographs. The research was supported by the Australian Research Council, AA8931446 and A092 105 1
LITERATURE CITED
Bell TA, Lightner DV (1988) A handbook of normal penaeid shrimp histology. The World Aquaculture Society, Baton Rouge
Bonami J R , Lightner DV, Redman KM, Poulos BT (1992) Par- tial characterization of a togavirus (LOVV) associated with histopathological changes of the lymphoid organ of penaeid shrimps. Dis aquat Org 14:145-152
Boonyaratpalin S, Supamattaya K, Kasornchandra J , Direk- busaracom S. Aekpanithanpong U, Chantanachooklin C (1993) Non-occluded baculo-like virus the causative agent of yellowhead disease in the black tiger shrimp. Gyobyo Kenkyu 28:103-109
Brun G (1991) Rhabdoviridae. In: Adams JR, Bonami J R (eds) Atlas of invertebrate viruses. CRC Press, Boca Raton, p 443-460
Chantanachookin C, Boonyaratpalin S, Kasornchandra J, Direkbusarakom, S, Ekpanithanpong U, Supamataya K, Sriurairatana S, Flegel TW (1993) Histology and ultra- structure reveal a new granulosis-like virus in Penaeus monodon affected by yellow-head dlsease. Dis aquat Org 17:145-157
Flegel TW, Fegan DF, Kongsom S, Vuthikornudomkit S, Snu- rairatana S, Boonyaratpalin S, Chantanachookin C, Vick- ers JE, Macdonald OD (1992) Occurrence, diagnosis and treatment of shrimp diseases in Thailand. In: Fulks W, Main KL (eds) Diseases of cultured penaeid shrimp in Asia and the United States. The Oceanic Institute, Honolulu, p 57-112
Franclu RIB, Farquet CM, Knudson DL, Brown F (eds) (1991) Classification and nomenclature of viruses. Fifth report of the International Committee on Taxonomy of Viruses. Springer-Verlag, New York, p 250-262
Humason GL (1968) Animal tissue techniques. WH Freeman and CO, San Francisco
Lightner DV, Redman RH, Bell TA (1983) Infectious hypoder- mal and hematopoietic necrosis vlrus, a newly recognised virus disease of penaeid shrimp. J Invertebr Path01 42:62-70

Dis aquat Org 23: 127-134, 1995
Lu Y, Loh PC (1992) Some biological properties of a rhab- dovirus isolated from penaeid shnmps. Arch Virol 127: 339-343
Lu Y, Nadala ECB Jr, Brock J A , Loh PC (1991) A new virus isolate from infectious hypodermal and hematopoietic necrosis virus (1HHNV)-infected penaeid shnmps. J virol Methods 31:189-196
Owens L, Andersen [G, Kenway M, Trott L, Benzie JAH (1992) Infectious hyperdermal and haematopoietic necro- sis virus (IHHNV) in a hybrid penaeid prawn from tropical Australia. Dis aquat Org 14:219-228
Responsible Subject Editoc J. E. Stewart, Dartmouth, Nova Scotia, Canada
Owens L, De Beer S, Smith J (1991) Lynlphoid parvo-like par- ticles in Australian penaeid prawns. Dis aquat Org 11: 129-134
Spurr AR (1969) A low viscosity epoxy resin embedding medium for electron microscopy. J Ultrastruct Res 26: 31-43
Wongteerasupaya C, Sriurairatana S, Vickers JE, Akrajamorn A, Boonsaeng V, Panyim S, Tassanakajon A, Withyachun- marnkul B, Flegel TLV (1995) Yellow-head virus of Penaeus monodon is an RNA virus. Dis aquat Org 22: 45-50
Manuscript first received: December 29, 1994 Revised version accepted: May l l , 1995


















