
Lung Tumor-Associated Dendritic Cell-Derived Amphiregulin
13
of April 10, 2019. This information is current as Progression Cancer Cell-Derived Amphiregulin Increased Lung Tumor-Associated Dendritic Hung, Chih-Jen Yang, Shah-Hwa Chou and Po-Lin Kuo Ya-Ling Hsu, Ming-Shyan Huang, Da-En Cheng, Jen-Yu http://www.jimmunol.org/content/187/4/1733 doi: 10.4049/jimmunol.1100996 2011; 2011; 187:1733-1744; Prepublished online 8 July J Immunol Material Supplementary 6.DC1 http://www.jimmunol.org/content/suppl/2011/07/08/jimmunol.110099 References http://www.jimmunol.org/content/187/4/1733.full#ref-list-1 , 20 of which you can access for free at: cites 33 articles This article average * 4 weeks from acceptance to publication Fast Publication! • Every submission reviewed by practicing scientists No Triage! • from submission to initial decision Rapid Reviews! 30 days* • Submit online. ? The JI Why Subscription http://jimmunol.org/subscription is online at: The Journal of Immunology Information about subscribing to Permissions http://www.aai.org/About/Publications/JI/copyright.html Submit copyright permission requests at: Email Alerts http://jimmunol.org/alerts Receive free email-alerts when new articles cite this article. Sign up at: Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists, Inc. All rights reserved. Copyright © 2011 by The American Association of 1451 Rockville Pike, Suite 650, Rockville, MD 20852 The American Association of Immunologists, Inc., is published twice each month by The Journal of Immunology by guest on April 10, 2019 http://www.jimmunol.org/ Downloaded from by guest on April 10, 2019 http://www.jimmunol.org/ Downloaded from
Transcript of Lung Tumor-Associated Dendritic Cell-Derived Amphiregulin

of April 10, 2019.This information is current as
ProgressionCancerCell-Derived Amphiregulin Increased
Lung Tumor-Associated Dendritic
Hung, Chih-Jen Yang, Shah-Hwa Chou and Po-Lin KuoYa-Ling Hsu, Ming-Shyan Huang, Da-En Cheng, Jen-Yu
http://www.jimmunol.org/content/187/4/1733doi: 10.4049/jimmunol.11009962011;
2011; 187:1733-1744; Prepublished online 8 JulyJ Immunol
MaterialSupplementary
6.DC1http://www.jimmunol.org/content/suppl/2011/07/08/jimmunol.110099
Referenceshttp://www.jimmunol.org/content/187/4/1733.full#ref-list-1
, 20 of which you can access for free at: cites 33 articlesThis article
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists, Inc. All rights reserved.Copyright © 2011 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on April 10, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from
by guest on April 10, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from

The Journal of Immunology
Lung Tumor-Associated Dendritic Cell-Derived AmphiregulinIncreased Cancer Progression
Ya-Ling Hsu,* Ming-Shyan Huang,†,‡ Da-En Cheng,* Jen-Yu Hung,*,†,‡ Chih-Jen Yang,*,†,‡
Shah-Hwa Chou,x and Po-Lin Kuo‡,{
The interaction of cancer within a microenvironment is an important factor determining cancer development. This study analyzed
the soluble factors secreted by tumor-associated dendritic cells (TADCs), which are responsible for increasing lung cancer growth,
migration, invasion, and epithelial-to-mesenchymal transition. Addition of amphiregulin, present in large amounts in TADC-
conditioned medium (CM), mimicked the inductive effect of TADC-CM on lung cancer progression, supported by the enhancement
of cell proliferation, migration, and invasion as well as osteolytic bone metastases phenotypes. In contrast, neutralization of amphir-
egulin fromTADC-CMdecreased the advancedmalignancy-inductive properties of TADC-CM. Significant upregulation of amphir-
egulin has been seen in tumor-infiltrating CD11c+ DCs in human lung cancer samples and patients’ sera. The enhancement of
amphiregulin in TADCs has also been noted in mice transplanted with lung cancer cells. Induction of lung cancer progression by
TADC-derived amphiregulin is associated with increased STAT3 and AKTactivation, which subsequently increases the expression
of cyclin D, Twist, and Snail. Blocking AKT significantly decreases TADC-CM and amphiregulin-mediated migration by de-
creasing the upregulation of Snail, whereas inhibition of STAT3 reduced the modulation of TADC-derived amphiregulin on Twist
and cyclin D expression, suggesting that cooperation of STAT3 and AKT plays a critical role in TADC-mediated cancer pro-
gression. Moreover, mice treated with anti-amphiregulin Abs showed decreased incidence of cancer development and increased
survival rates. Our study suggests that inhibition of amphiregulin or amphiregulin-related signaling is an attractive therapeutic
target in lung cancer patients. The Journal of Immunology, 2011, 187: 1733–1744.
Lung cancer, one of the leading causes of cancer mortalityin the world, is associated with very poor prognoses evenafter surgical resection (1). Cancer microenvironments
have been demonstrated to play an important role in modulatingtumor progression and chemoresistance to anticancer drugs.Cancer microenvironments consist of fibroblasts, immune cells(lymphocytes, dendritic cells [DCs], and macrophages), extra-cellular matrix components, growth factors, and metabolites. Agrowing body of studies indicates that cancer development is alsoregulated by an orchestrated series of bidirectional interactionsbetween nonneoplastic cells and cancer cells in the tumor mi-croenvironment (2, 3). The lungs are notably complex in terms ofthe different types of DCs to be found. DC subsets of the lung can
be divided into CD11chigh conventional DCs and CD11cdim plas-macytoid DCs (4). Different DC subsets may carry out differentfunctions. We have suggested that lung cancer has the ability topromote immune tolerance and escape the host’s immune sur-veillance by inhibiting the differentiation, maturation, and acti-vation of CD11chigh DCs (5). A CD11chigh myeloid DC, which ispresent in lung cancer, displays a “semimature” and immunno-suppression phenotype (6). In contrast, a variety of immune cells,such as macrophages and myeloid-derived suppressor cells, havebeen reported to contribute to tumor metastasis (7). However, it isstill not fully understood whether tumor-associated DCs (TADCs)are involved in lung cancer development or precisely whichmolecules are involved in the cross-talk of cancer and TADCs inthe tumor microenvironment. Strategies that harness the immunesystem to react against tumors can be integrated with existingforms of therapy for optimal responses in treating this devastatingdisease.Amphiregulin, an epidermal growth factor family growth factor,
binds and activates the epidermal growth factor receptor (EGFR).Amphiregulin is produced as a transmembrane precursor that isreleased from the cell membrane by a disintegrin and metallo-proteinases (ADAM17) (8). Elevated levels of amphiregulin havebeen characterized in various types of tumors, and amphiregulin isassumed to play a role in promoting cancer progression (9, 10).Amphiregulin also elevates angiogenic activity, promoting tumorgrowth and metastasis in pancreatic, colorectal, liver, and lungcancers (9–12). A recent study has indicated that cancer-derivedATP increased the expression of amphiregulin in DCs in a mouselung cancer cell line (13). Patients with high levels of amphir-egulin have poorer prognoses and higher resistance than patientswith low levels of amphiregulin (14). After activation of EGFR,amphiregulin signaling promotes activation of the PI3K, therebypromoting the activation of AKT, protein kinase C, calcium
*Graduate Institute of Medicine, Kaohsiung Medical University, Kaohsiung, Taiwan;†Division of Pulmonary and Critical Care Medicine, Kaohsiung Medical UniversityHospital, Kaohsiung, Taiwan; ‡Cancer Center, Kaohsiung Medical University Hos-pital, Kaohsiung, Taiwan; xDepartment of Chest Surgery, Kaohsiung Medical Uni-versity Hospital, Kaohsiung, Taiwan; and {Institute of Clinical Medicine, KaohsiungMedical University, Kaohsiung, Taiwan
Received for publication April 6, 2011. Accepted for publication June 6, 2011.
This work was supported by Grant NSC 98-2320-B-037-007-MY3 from the NationalScience Council of Taiwan, a grant from the Excellence for Cancer Research Center,and Grant DOH100-TD-C-111-002 from the Department of Health, Executive Yuan(Taipei, Taiwan).
Address correspondence and reprint requests to Dr. Po-Lin Kuo, Graduate Instituteof Medicine, Kaohsiung Medical University, Kaohsiung, Taiwan. E-mail address:[email protected]
The online version of this article contains supplemental material.
Abbreviations used in this article: CM, conditioned medium; DC, dendritic cell;EGFR, epidermal growth factor receptor; EMT, epithelial-to-mesenchymal transi-tion; LLC, Lewis lung carcinoma; mdDC, monocyte-derived dendritic cell; PTHrP,parathyroid hormone-related protein; Q-PCR, quantitative real-time PCR; siRNA,small interfering RNA; TADC, tumor-associated dendritic cell.
Copyright� 2011 by The American Association of Immunologists, Inc. 0022-1767/11/$16.00
www.jimmunol.org/cgi/doi/10.4049/jimmunol.1100996
by guest on April 10, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from

mobilization MAPK, or STAT3 signaling cascades (12, 14–16).AKT and STAT3 are known to be critical mediators in the controlof cancer growth and progression. Therefore, we hypothesized thatamphiregulin may play a role in mediating the regulatory effectsof lung cancer development via the DCs–lung cancer cell in-teraction.In this study, we discovered that TADCs secrete high amounts
of amphiregulin and play a role in promoting cancer progression.Our data show that TADCs-derived amphiregulin increases A549and H460 proliferation, migration, and epithelial-to-mesenchymaltransition (EMT) by the paracrine effect. These findings alsorevealed that DCs infiltrating tumor tissues of mice and humanpatients express high levels of amiphregulin, suggesting a novelrole of amphiregulin in the interaction of cancer and DCs. Fur-thermore, treatment of mice with anti-amphiregulin Abs decreasedthe incidence of lung tumors and increased survival rates. Inves-tigating the mechanisms by which TADCs contributes to lungcancer progression may facilitate the discovery of a potential targetfor developing therapeutic strategies targeting immune cells in thetumor microenvironment.
Materials and MethodsLung cancer cells and conditioned media
Human lung cancer cells A549 and H460 and mouse Lewis lung carcinoma(LLC) cells were obtained from the Bioresource Collection and ResearchCenter (Hsinchu City, Taiwan). To obtain the A549 and H460-conditionedmedium (CM), cells were seeded at 2 3 106 cells/100 mm dish and cul-tivated for 24 h. The medium was replaced, and the supernatants wereharvested after 48 h of incubation.
Serum samples from lung cancer patients
Preoperative blood samples were obtained from 62 lung cancer patients and19 healthy donors admitted to the Division of Pulmonary and Critical CareMedicine, Kaohsiung Medical University Hospital (Kaohsiung, Taiwan).Serum was separated by centrifugation and frozen at 280˚C. Approval forthese studies was obtained from the Institutional Review Board of Kaoh-siung Medical University Hospital. Informed consent was obtained fromall patients in accordance with the Declaration of Helsinki.
Measurement of secreted factors
Supernatants from TADC, monocyte-derived DCs (mdDCs), and A549and H460 cells were collected. IL-6 was assessed by IL-6 ELISA kit.Amphiregulin and IL-8 levels were quantified using the DuoSet ELISA.Parathyroid hormone-related protein (PTHrP) levels were determined byenzyme immunoassay kit (Peninsula Laboratories, San Carlos, CA).
Isolation of CD14+ monocytes and differentiation of mdDCs
Monocytes were purified from PBMCs obtained from healthy consentingdonors. Mononuclear cells were isolated from blood by Ficoll-Hypaquegradient (GE Healthcare Bio-Sciences, Little Chalfont, U.K.). CD14+
monocytes were purified using CD14+ mAb-conjugated magnetic beads(MACS MicroBeads; Miltenyi Biotec), according to the manufacturer’sprotocol. mdDCs were generated by culturing CD14+ monocytes in RPMI1640 medium containing 10% FBS (Invitrogen, Carlsbad, CA) and 20 ng/ml GM-CSF and 10 ng/ml IL-4 (R&D Systems, Minneapolis, MN) for 5 d.The medium was replaced with fresh medium containing GM-CSF and IL-4 on day 3. For maturation of DCs, immature mdDCs were stimulated withLPS (100 ng/ml) after priming with IFN-g for 3 h. A549 tumor-associatedmdDCs (A549-TADCs) and H460-TADCs were generated by culturingCD14+ monocytes in RPMI 1640 medium containing FBS, GM-CSF, andIL-4 presenting in A549-CM and H460-CM and then stimulated as de-scribed above. After washing, the supernatants were collected and iden-tified as A549-TADC-CM and H460-TADC-CM (Supplemental Fig. 1A).
Analysis of cell proliferation, migration, and invasion
Cell proliferation was assessed by Premixed WST-1 Cell ProliferationReagent (BD Clontech, Mountain View, CA), according to the manu-facturer’s instructions. Cell migration and invasion assays were conductedusing wound-healing, QCM 24-well Cell Migration Assay and InvasionSystem, as described previously (17). mdDC-CM and TADC-CM wereadded to the bottom wells for 24 h as a chemoattractant, and the fluores-
cence of the invading cells was read using a fluorescence plate reader atexcitation/emission wavelengths of 485/530 nm. The migration of A549and H460 cells was also assessed by a Scratch wound-healing assay.
Quantitative real-time PCR
RNA isolation was performed using the TRIzol reagent (Invitrogen). cDNAwas prepared using an oligo(dT) primer and reverse transcriptase (Takara,Shiga, Japan) following standard protocols. Real-time PCR was performedusing SYBR Green on the ABI 7500 Real-Time PCR System (AppliedBiosystems, Foster City, CA). Each PCRmixture contained 200 nM of eachprimer, 10 ml 23 SYBR Green PCR Master Mix (Applied Biosystems),and 5 ml cDNA and RNase-free water, with a total volume of 20 ml. ThePCR was carried out with a denaturation step at 95˚C for 10 min and thenfor 40 cycles at 95˚C for 15 s and at 60˚C for 1 min. All PCRs wereperformed in triplicate and normalized to internal control GAPDH mRNA.Relative expression was presented using the 2244CT method.
Immunoblot
Cells were lysed on ice for 15 min by M-PER lysis reagent (Pierce). Celllysate was centrifuged at 14,000 3 g for 15 min, and the supernatantfraction was collected for immunoblot. Equivalent amounts of protein wereresolved by SDS-PAGE (8–12%) and transferred to polyvinylidene dif-luoride membranes. After blocking for 1 h in 5% nonfat dry milk in TBS,the membrane was incubated with the desired primary Ab for 1–16 h. Themembrane was then treated with appropriate peroxidase-conjugated sec-ondary Ab, and the immunoreactive proteins were detected using an ECLkit (Millipore), according to the manufacturer’s instructions.
Immunofluorescence
Noncancerous lung tissue specimens obtained from human lung cancerpatients were embedded in OCTand frozen in liquid nitrogen. Sections (3–5mm) were fixed with acetone at 220˚C and then stained with amphiregulinAb (Abcam) or costained by anti-CD11c Ab. After a wash with PBS con-taining 0.1% Tween 20, slides were incubated with Dylight 488- or Dylight549-conjugated secondary Abs (Rockland, Gilbertsville, PA), with or withoutDAPI, for 1 h at room temperature. The data were analyzed with a confocallaser scanning microscope (Fluoview FV500; Olympus, Tokyo, Japan).
Osteoclast differentiation and activity assay
PBMCs were plated and incubated overnight at 37˚C. Nonadherent cellswere removed by washing with PBS, and the remaining adherent cellswere grown in culture medium containing mdDC-CM, A549-TADC-CM,H460-TADC-CM, or amphiregulin (10 ng/ml) presenting in 200 ng/mlhuman M-CSF and 100 ng/ml human RANKL for 7 d. The medium wasreplaced every 3 d. Osteoclast formation was measured by quantifyingcells positively stained by TRAP (Sigma-Aldrich, St. Louis, MO); osteo-clasts were determined to be TRAP-positive staining multinuclear (morethan three nuclei) cells by means of light microscopy. The TRAP-positivecells, and the number of nuclei per TRAP-positive cell in each well, werecounted. Osteoclasts’ bone resorption activity was assessed by BD Bio-Coat Osteologic Bone Resorption assay (BD Biosciences, Bedford MA), inthe same culture conditions as described above.
Gene knockdown by small interfering RNA
Monocytes were transfected with 1 mmol/l nontarget or amphiregulinAccell small interfering RNAs (siRNAs) pool (Dharmacon) in Accelldelivery media (B-005000), according to the manufacturer’s instructions.Positive controls Accell GAPDH siRNA and scrambled Accell siRNA poolwere used in the experiments. After 72 h transfection, the medium waschanged to RPMI 1640 medium containing 10% FBS, 20 ng/ml GM-CSFand 10 ng/ml IL-4 presenting in A549-CM and H460-CM, and thenstimulated by IFN-gand LPS for 24 h. Nontarget or amphiregulin AccellsiRNAs were added every 3 d to maintain the knockdown efficiency ofsiRNA. The supernatants of mdDCs were collected as CMs. The amphir-egulin changes of mdDCs were measured by real-time PCR as describedabove.
Animal models and isolation of CD11c+ cells from lungs
LLC cells were transplanted into C57BL/6 mice by tail vein injection. Lungtissuewas collected 24 d after injection, thenminced and incubated in RPMI1640 medium with collagenase type 1 (400 U/ml) (Worthington Bio-chemicals) for 1 h at 37˚C. The digested tissues were filtered through a 70-mm cell strainer and washed with RPMI 1640 medium. CD11c+ DCs wereisolated from the cell suspension by CD11c magnetic beads (MiltenyiBiotec). Mouse bone marrow cells were harvested from the long bones of
1734 AMPHIREGULIN INVOLVED IN CANCER–IMMUNE SYSTEM INTERACTION
by guest on April 10, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from

the limbs and then placed in RPMI 1640 containing murine GM-CSF (20ng/ml) and murine IL-4 (20 ng/ml, R&D Systems), with or without LLC-CM, for 24 h. The expression of various mRNAs was assayed by real-timePCR. Survival rates were evaluated in mice injected with LLC cells. Iso-type control Ig (IgG) and amphiregulin (50 mg/mice) Ab were i.p. injectedevery 7 d three times after administration of LLC. Animals were sacrificedon day 24 after LLC cell transplantation and the number of tumor nodulesrecorded for the analysis of lung cancer incidence. For survival assay, allanimals were closely monitored daily for signs of distress and were sac-rificed when they appeared lethargic or to have increased breathing rates.
Statistical analysis
Data were expressed as means 6 SD. Statistical comparisons of the resultswere made using ANOVA. Significant differences (p , 0.05) between themeans of the two test groups were analyzed by Student t test. Survivalstudies were assessed using Kaplan–Meier survival curves and analyzedwith the Mantel–Cox log-rank test.
ResultsTADCs increased lung cancer cell proliferation, migration andEMT
To investigate the influence of TADCs on lung cancer progression,we assessed the effect of TADC-CM on lung cancer cell pro-liferation, migration, and invasion. As shown in Fig. 1A, A549-TADC-CM (20%) and H460-TADC-CM (20%) increased theproliferation of A549 and H460 cells after 72 h of treatment. Inaddition, both A549-TADC-CM and H460-TADC-CM increasedthe migration and invasive abilities of A549 and H460 cells (Fig.1B, 1C). Furthermore, A549-TADC-CM and H460-TADC-CMalso caused A549 and H460 cells to undergo EMT, includingthe downregulation of epithelial markers (E-cadherin, claudin-3,and ZO-1), and upregulation of fibroblast markers (N-cadherin,vimentin, and fibronectin) (Fig. 1D).The effects of TADC on lung cancer migration and invasion
were also confirmed by coculture system. As shown in Supple-mental Fig.1B and 1C, coculture of A549 or H460 cells with A549-TADCs or H460-TADCs enhances the migratory and invasive ca-pability of A549 and H460 cells, respectively. These coculturesystem data coincide with that of TADC-CM treatment.
Amphiregulin plays an important role in TADCs-mediatedcancer progression
To investigate which factor is responsible for TADCs-mediatedlung cancer development, we used a microarray to assess the geneprofile of A549-TADCs. The data showed that several solublefactors were upregulated in A549-TADCs when compared withmdDCs. Among these upregulated genes, levels of amphiregulin,a lung cancer-related growth factor, increased 11.73-fold in A549-TADCs (Fig. 2A). In addition, ADAM17, an enzyme responsiblefor amphiregulin release, was also increased in A549-TADCs.Quantitative real-time PCR (Q-PCR) analysis further revealedthat A549-CM or H460-CM increased the expression of amphir-egulin in A549-TADCs and H460-TADCs (Fig. 2A). Furthermore,protein levels of amphiregulin were enhanced in A549-TADCsand H460-TADCs, as determined by ELISA analysis (Supple-mental Fig. 1D).We also assessed the effect of amphiregulin protein on the
proliferation, migration, invasion, and EMT of A549 and H460cells. Amphiregulin increased the proliferation of A549 and H460cells in a concentration-dependent manner (Supplemental Fig. 1E).Moreover, amphiregulin not only increased the migratory abilityof A549 and H460 (Fig. 2B) but also enhanced their invasiveability and EMT (Fig. 2C, 2D).To better understand the role of amphiregulin, we blocked the
effect of amphiregulin on A549 and H460 proliferation and mi-
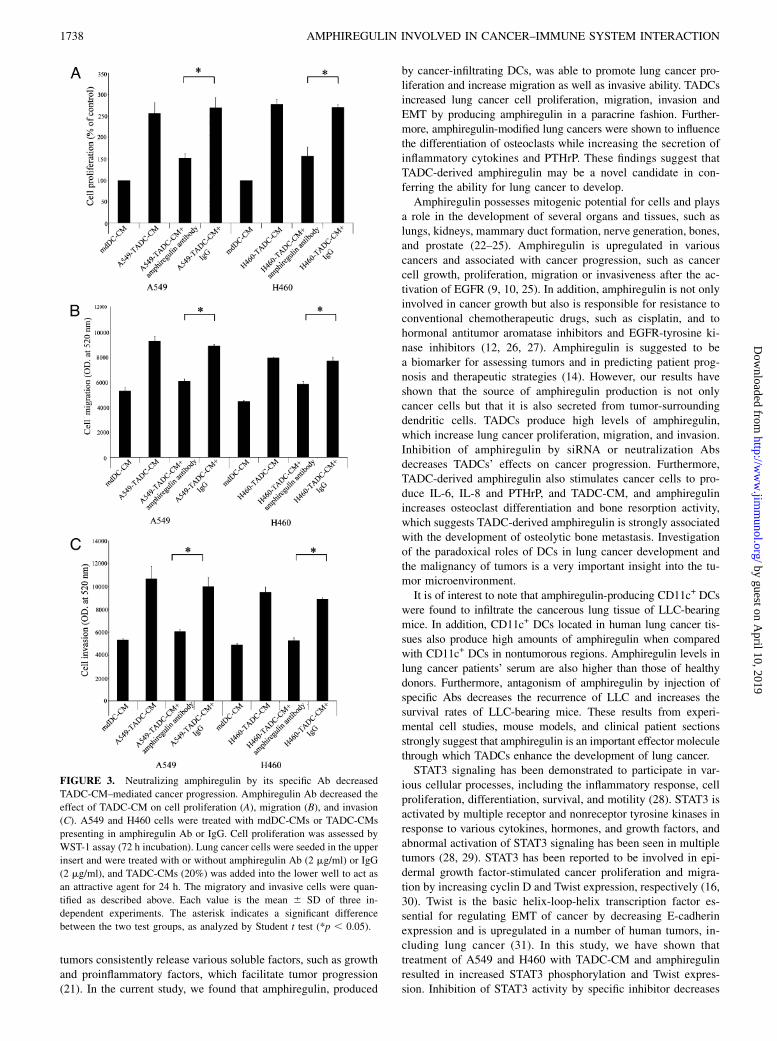
gration by using specific neutralization Ab and siRNA transfection.As shown in Fig. 3A, the effect of A549-TADC-CM and H460-TADC-CM on cell proliferation in A549 and H460 cells wasneutralized by treatment with anti-amphiregulin Ab but not byisotype control. Similarly, amphiregulin neutralization Ab alsodecreased the stimulatory effects of A549-TADC-CM and H460-TADC-CM on lung cancer cell migration and invasion (Fig. 3B,3C). The inhibitory effect of amphiregulin neutralization Ab onA549 migration and invasion was also confirmed by A549, H460,or mdDC coculture system (Supplemental Fig. 2A, 2B). Further-more, inhibition of amphiregulin in monocytes by amphiregulinsiRNA also reduced the effect of TADC-CM on proliferation, cellmigration, and invasive ability of A549 and H460 cells (Fig. 4B–D). These data suggest that tumor-infiltrated DCs may increaselung cancer progression by amphiregulin production.
TADC-derived amphiregulin stimulated lung cancer to producebone-metastasis factor (IL-6, IL-8 and parathyroidhormone-related protein) and osteoclastogenesis
Bone is a frequent target of lung cancer metastasis, and bonemetastasis is mediated by complex interactions between tumorcells and resident stromal cells in the cancer microenvironment(18). We therefore assessed the potential role of TADC-derivedamphiregulin in lung cancer metastasis. As shown in Fig. 5A and5B, A549-TADC-CM and H460-TADC-CM increased A549 orH460 cells’ expression of two inflammatory cytokines, IL-6 andIL-8, which have been implicated in bone metastasis (19, 20).Similar to TADC-CM, amphiregulin also increases the expressionof IL-6 and IL-8 in both A549 and H460 cells. PTHrP, a causativefactor that contributes to osteolytic metastases, is also regulatedin both A549-TADC-CM or amphiregulin-treated A549 andH460-TADC-CM or amphiregulin-treated H460 cells (Fig. 5C).Amphiregulin also increases the formation and activity of osteo-clasts from PBMCs presenting in M-CSF and RANKL (Fig. 5D).In addition, knockdown of amphiregulin in monocytes by siRNAalso decreases the upregulation of TADC-CMs on PTHrP ex-pression in both A549 and H460 cells (Fig. 5E).
STAT3 and AKT activation are involved in theamphiregulin-mediated effect on lung cancer
Because TADC-derived amphiregulin increased cell proliferation,migration, and invasion, we assessed the effect of TADC-CMs andamphiregulin on several oncogenic signaling pathways, includingAKT and STAT3 (15, 16). The results showed that exposure ofA549 and H460 cells to A549-TADC-CM or H460-TADC-CM ledto the phosphorylation of STAT3 and AKT. In contrast, the ex-pression of STAT3 and AKT (unphosphorylated form) was un-altered by TADC-CM treatment in either A549 or H460 cells (Fig.6A). Similar results were also found in amphiregulin-treatedA549 and H460 cells (Fig. 7A). In addition, both TADC-CMs andamphiregulin increased the expression of Snail and Twist, twoEMT-related transcriptional factors, and enhanced the levels ofcyclin D in both A549 and H460 cells (Figs. 6A, 7A).To investigate the functional impact of AKT and STAT3 acti-
vation on TADC-derived amphiregulin in lung cancer, we usedchemical inhibitors Ly294002 and AG490 to specifically inhibitAKT and STAT3 signaling, respectively. STAT3 inhibitor AG490decreased TADC-CM and amphiregulin-mediated cell prolifera-tion, migration, and invasion (Figs. 6B–D, 7B–D). In contrast, AKTinhibitor Ly294002 markedly inhibited two types of TADC-CMsand amphiregulin-induced cell migration in both A549 and H460cell lines, whereas it only slightly decreased TADC-CM andamphiregulin-driven cell proliferation. Furthermore, pretreatmentof A549 and H460 cells with STAT3 inhibitor completely
The Journal of Immunology 1735
by guest on April 10, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from

abrogated expression of Twist and cyclin D but only partiallydecreased Snail caused by A549-TADC-CM, H460-TADC-CM,
and amphiregulin (Figs. 6E, 7E). In contrast, AKT inhibitor
completely inhibited Snail expression, but not Twist and cyclin D.
Moreover, both STAT3 and AKT inhibitors reversed the inhibitory
effect of TADC-CMs on E-cadherin in A549 and H460 cells.
These data suggest that STAT3 and AKT play important roles in
lung cancer progression (Figs. 6E, 7E).
High amounts of amphiregulin in CD11c+ DCs are found in thetumor sections of lung cancer-bearing mice and human lungcancer patients
To confirm the role of amphiregulin on TADC-mediated cancerprogression in humans, we characterized the expression ofamphiregulin in tumor-infiltrated CD11c+ DCs. Immunofluores-cence staining of the marginal regions of a patient’s lung cancerreveals the presence of many CD11c+ DCs, which have infiltrated
FIGURE 1. TADCs increased lung cancer progression. TADC-CM increased lung cancer cell proliferation (A), migration (B), invasion (C), and EMT
(D). A549 cells were treated with A549-TADC-CM, and H460 cells were treated with H460-TADC-CM for 72 h. Cell proliferation was assessed by WST-1
assay. The invasiveness and migration ability of A549 and H460 cells were quantified by QCM 24-well Cell Migration and Invasion assay. Twenty percent
of A549-TADC-CM (for A549), H460-TADC-CM (for H460), or mdDC-CM (for control) acts as a chemoattractant of cancer migration and invasion. Each
value is the mean 6 SD of three independent experiments. The asterisk indicates a significant difference between the two test groups, as analyzed by
Student t test (*p , 0.05).
1736 AMPHIREGULIN INVOLVED IN CANCER–IMMUNE SYSTEM INTERACTION
by guest on April 10, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from

the area around the tumor. These areas express higher levels ofamphiregulin in comparison with tissue sections of non-tumorregions (Fig. 8A). Furthermore, a marked statistical differencehas also been found between lung cancer patients and healthydonor serum (Fig. 8B).We also used animal experiments to determine whether lung
cancer increased amphiregulin expression in DCs in vivo. Weinjected mouse lung cancer cell line LLC into mice and thenallowed the cells to develop for 14 d. LLC-CM increased theproduction of amphiregulin in mice bone marrow cells presentingin IL-4 and GM-SCF (Supplemental Fig. 3A). These data coincidewith human lung cancer data. Q-PCR and ELISA analysis havealso shown that lung tumor-infiltrated CD11c+ DCs produce ele-vated levels of amphiregulin in mRNA and protein levels (Fig. 8C,Supplemental Fig. 3B).
Amphiregulin Ab decreased cancer metastasis in vivo
Finally, we assessed whether inhibiting amphiregulin productionfrom tumor-infiltrated DCs could decrease lung cancer develop-ment. Treatment of mice by anti-amphiregulin mAb inhibited thegrowth of LLC in 25% (two of eight) of mice, in comparison with85% (six of seven) of rat IgG-treated control mice (Fig. 8D).Furthermore, treatment by anti-amphiregulin mAb also improvedlong-term survival of mice (66.6%) with LLC tumors (up to 90 d),whereas all mice treated by control IgG died of cancer within25 d (Fig. 8E).
DiscussionThe microenvironment of a tumor is widely known to be an im-portant factor regulating cancer development. Cells surrounding
FIGURE 2. Amphiregulin is involved in TADC-mediated cancer progression. A, The expression of amphiregulin and ADAM17 in A549-TADC. The
expression of A549-TADC and mdDC was assessed by microarray and Q-PCR. Amphiregulin increased cell migration (B) and invasion (C) as well as EMT
(D) in A549 and H460 cells. The invasiveness and migration ability of A549 and H460 cells were quantified as described above. Amphiregulin (10 ng/ml)
acts as a chemoattractant of cancer migration and invasion. Each value is the mean 6 SD of three independent experiments. The asterisk indicates
a significant difference with control, as analyzed by Student t test (*p , 0.05).
The Journal of Immunology 1737
by guest on April 10, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from
tumors consistently release various soluble factors, such as growthand proinflammatory factors, which facilitate tumor progression(21). In the current study, we found that amphiregulin, produced
by cancer-infiltrating DCs, was able to promote lung cancer pro-liferation and increase migration as well as invasive ability. TADCsincreased lung cancer cell proliferation, migration, invasion andEMT by producing amphiregulin in a paracrine fashion. Further-more, amphiregulin-modified lung cancers were shown to influencethe differentiation of osteoclasts while increasing the secretion ofinflammatory cytokines and PTHrP. These findings suggest thatTADC-derived amphiregulin may be a novel candidate in con-ferring the ability for lung cancer to develop.Amphiregulin possesses mitogenic potential for cells and plays
a role in the development of several organs and tissues, such aslungs, kidneys, mammary duct formation, nerve generation, bones,and prostate (22–25). Amphiregulin is upregulated in variouscancers and associated with cancer progression, such as cancercell growth, proliferation, migration or invasiveness after the ac-tivation of EGFR (9, 10, 25). In addition, amphiregulin is not onlyinvolved in cancer growth but also is responsible for resistance toconventional chemotherapeutic drugs, such as cisplatin, and tohormonal antitumor aromatase inhibitors and EGFR-tyrosine ki-nase inhibitors (12, 26, 27). Amphiregulin is suggested to bea biomarker for assessing tumors and in predicting patient prog-nosis and therapeutic strategies (14). However, our results haveshown that the source of amphiregulin production is not onlycancer cells but that it is also secreted from tumor-surroundingdendritic cells. TADCs produce high levels of amphiregulin,which increase lung cancer proliferation, migration, and invasion.Inhibition of amphiregulin by siRNA or neutralization Absdecreases TADCs’ effects on cancer progression. Furthermore,TADC-derived amphiregulin also stimulates cancer cells to pro-duce IL-6, IL-8 and PTHrP, and TADC-CM, and amphiregulinincreases osteoclast differentiation and bone resorption activity,which suggests TADC-derived amphiregulin is strongly associatedwith the development of osteolytic bone metastasis. Investigationof the paradoxical roles of DCs in lung cancer development andthe malignancy of tumors is a very important insight into the tu-mor microenvironment.It is of interest to note that amphiregulin-producing CD11c+ DCs
were found to infiltrate the cancerous lung tissue of LLC-bearingmice. In addition, CD11c+ DCs located in human lung cancer tis-sues also produce high amounts of amphiregulin when comparedwith CD11c+ DCs in nontumorous regions. Amphiregulin levels inlung cancer patients’ serum are also higher than those of healthydonors. Furthermore, antagonism of amphiregulin by injection ofspecific Abs decreases the recurrence of LLC and increases thesurvival rates of LLC-bearing mice. These results from experi-mental cell studies, mouse models, and clinical patient sectionsstrongly suggest that amphiregulin is an important effector moleculethrough which TADCs enhance the development of lung cancer.STAT3 signaling has been demonstrated to participate in var-
ious cellular processes, including the inflammatory response, cellproliferation, differentiation, survival, and motility (28). STAT3 isactivated by multiple receptor and nonreceptor tyrosine kinases inresponse to various cytokines, hormones, and growth factors, andabnormal activation of STAT3 signaling has been seen in multipletumors (28, 29). STAT3 has been reported to be involved in epi-dermal growth factor-stimulated cancer proliferation and migra-tion by increasing cyclin D and Twist expression, respectively (16,30). Twist is the basic helix-loop-helix transcription factor es-sential for regulating EMT of cancer by decreasing E-cadherinexpression and is upregulated in a number of human tumors, in-cluding lung cancer (31). In this study, we have shown thattreatment of A549 and H460 with TADC-CM and amphiregulinresulted in increased STAT3 phosphorylation and Twist expres-sion. Inhibition of STAT3 activity by specific inhibitor decreases
FIGURE 3. Neutralizing amphiregulin by its specific Ab decreased
TADC-CM–mediated cancer progression. Amphiregulin Ab decreased the
effect of TADC-CM on cell proliferation (A), migration (B), and invasion
(C). A549 and H460 cells were treated with mdDC-CMs or TADC-CMs
presenting in amphiregulin Ab or IgG. Cell proliferation was assessed by
WST-1 assay (72 h incubation). Lung cancer cells were seeded in the upper
insert and were treated with or without amphiregulin Ab (2 mg/ml) or IgG
(2 mg/ml), and TADC-CMs (20%) was added into the lower well to act as
an attractive agent for 24 h. The migratory and invasive cells were quan-
tified as described above. Each value is the mean 6 SD of three in-
dependent experiments. The asterisk indicates a significant difference
between the two test groups, as analyzed by Student t test (*p , 0.05).
1738 AMPHIREGULIN INVOLVED IN CANCER–IMMUNE SYSTEM INTERACTION
by guest on April 10, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from

cyclin D and Twist expression, suggesting that STAT3 is an up-stream regulator of cyclin D and Twist. Moreover, STAT3 in-hibitor decreases the effects of TADC-CM and amphiregulin onbone metastasis factor PThrP production and cell migration inA549 and H460 cells, suggesting that the activation of STAT3signaling plays a crucial role in the development of lung cancergrowth and metastasis.
AKT, a downstream effector of PI3K has been shown to bea critical mediator of cell proliferation, survival, and metastasis ina variety of cell types. AKT has been reported to act as an im-portant factor in amphiregulin-mediated cell survival and mi-gration (32, 33). In our study, we found that treating A549 andH460 cells with TADC-CM or amphiregulin dramatically in-creases the activation of AKT. The inductive effect of TADC-
FIGURE 4. Inhibition of amphiregulin decreased the enhancement of TADCs on lung cancer progression. A, The effect of amphiregulin siRNA on
monocytes. Knockdown of amphiregulin reduced TADCs on cancer proliferation (B), migration (C), and invasion (D). Knockdown of amphiregulin in
monocytes was performed by Accell siRNA at 1 mM concentration of scramble or amphiregulin siRNA. The supernatants of mdDCs and TADCs were
collected. The expression of amphiregulin was assessed by Q-PCR. Cell proliferation was assessed by WST-1 assay. The invasiveness and migration ability
of A549 and H460 cells were quantified by QCM 24-well Cell Migration and Invasion assay. Each value is the mean 6 SD of three independent
experiments. The asterisk indicates a significant difference between the two test groups, as analyzed by ANOVAwith Student t test post hoc (*p , 0.05).
The Journal of Immunology 1739
by guest on April 10, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from

FIGURE 5. TADC-derived amphiregulin increased bone metastasis-related factors in lung cancer. A, TADCd and amphiregulin increased the production
of IL-6 (A), IL-8 (B), and PTHrP (C) in A549 and H460 cells. A549 and H460 cells were treated with A549-TADC-CM (20%) or H460-TADC-CM (20%)
and amphiregulin (10 ng/ml) for 24 h, and then, the supernatants were collected and assessed by ELISA kits. D, Amphiregulin increased osteoclast
differentiation and activity. PBMCs were treated with amphiregulin (10 ng/ml) presenting in RANKL (100 ng/ml) and M-CSF (200 ng/ml) for 7 d. Os-
teoclast cells were stained for TRAP activity, and bone resorption activity was determined by the Osteologic MultiTest Slide. E, Inhibition of amphiregulin
decreased the enhancement of TADCs on PTHrP expression. A549 and H460 cells were treated with various amphiregulin siRNA or scrambled siRNA-
transfected A549 or H460-associated DC-CMs (20%) for 24 h, and then, the cells’ supernatants were collected and assessed by PTHrP ELISA kits. Each
value is the mean 6 SD of three independent experiments. The asterisk indicates a significant difference with control, as analyzed by Student t test (*p ,0.05).
1740 AMPHIREGULIN INVOLVED IN CANCER–IMMUNE SYSTEM INTERACTION
by guest on April 10, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from

FIGURE 6. TADC-CM promote cancer progression by STAT3 and AKT activation. A, TADC-CM increased STAT3 and AKT activation and enhanced
Snail and Twist expression. Cells were treated with TADC-CM (20%) for the indicated times, and the expression of various proteins was assessed by
immunoblot assay. STAT3 and AKT inhibitors decreased TADC-mediated cell proliferation (B), migration (C), invasion (D), and EMT (E). Cells were
pretreated with or without AKT (5 mM) or STAT3 (1 mM) inhibitor and then cultured with TADC-CM (20%) for the indicated times (72 h for cell
proliferation, 24 h for E-cadherin, and 6 h for Snail, Twist, and cyclin D). Cell proliferation was assessed by WST-1 assay. The invasiveness and migration
ability of A549 and H460 cells were quantified by QCM 24-well Cell Migration and Invasion assay. Cells were seeded in the upper inserts were treated with
or without AKT and STAT3 inhibitors, with amphiregulin acting as a chemoattractant for cancer migration and invasion. Each value is the mean 6 SD of
three independent experiments. The asterisk indicates a significant difference between the two test groups, as analyzed by Student t test (*p , 0.05).
The Journal of Immunology 1741
by guest on April 10, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from

CM or amphiregulin on AKT is responsible for the increaseof Snail expression, a repressor of epithelial type marker E-cadherin. Furthermore, selective inhibition of AKT by chemical
inhibitor also decreases the effects of TADC-CM or amphiregulinon cell migration and invasion, suggesting that phenotypic tran-sition of lung cancer by TADC involves amphiregulin-mediated
FIGURE 7. Amphiregulin promotes cancer progression by STAT3 and AKT activation. A, Amphiregulin increased STAT3 and AKT activation and
enhanced Snail and Twist expression. Cells were treated with amphiregulin (10 ng/ml) for the indicated times, and the expression of various proteins was
assessed by immunoblot assay. STAT3 and AKT inhibitors decreased amphiregulin-mediated cell proliferation (B), migration (C), invasion (D), and EMT
(E). Cells were pretreated with or without AKT or STAT3 inhibitor and then cultured with amphiregulin (10 ng/ml) for the indicated times (72 h for cell
proliferation, 24 h for E-cadherin, and 6 h for Snail, Twist, and cyclin D). Cell proliferation, invasiveness, and migration ability of A549 and H460 cells
were quantified by as described above. Each value is the mean 6 SD of three independent experiments. The asterisk indicates a significant difference
between the two test groups, as analyzed by Student t test (*p , 0.05).
1742 AMPHIREGULIN INVOLVED IN CANCER–IMMUNE SYSTEM INTERACTION
by guest on April 10, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from

FIGURE 8. Elevated amounts of amphiregulin on CD11c+ DCs in the lung cancer-bearing patient and mice. A, AmphiregulinhighD11c+ DCs infiltrated
cancer sections. Nontumor and tumor regions (n = 15) were cut and stained and then samples were analyzed by confocal microscopy (original magnification
340). Arrows indicate the CD11c+/amphiregulatin+ cells. B, High levels of amphiregulin were found in the serum of lung cancer patients. Amphiregulin
levels of lung cancer patients’ (n = 62) and healthy donors’ sera (n = 19) were assessed by ELISA. C, Elevated levels of amphiregulin were found in
CD11c+ DCs isolated from the lungs of lung-cancer bearing mice. Culture media and LLC were injected into mice via the tail vein (control group, n = 10;
LLC group, n = 8). After 24 d, nontumorous and tumorous regions of the lungs were harvested. CD11c+ DCs were isolated from fresh lung or tumor tissue,
and amphiregulin levels were assessed by Q-PCR. Neutralization of amphiregulin decreased the occurrence of lung cancer (D) and improved survival rates
(E) in mice. LLCs were injected into mice via the tail vein. Mice were dosed every 7 d with i.p. injections IgG (n = 7) or amphiregulin Abs (50 mg/ml) (n =
8). Arrows indicate the time of administration of amphiregulin Ab. The asterisk indicates a significant difference with the control, as analyzed by ANOVA
with Student t test post hoc (*p , 0.05).
The Journal of Immunology 1743
by guest on April 10, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from

activation of AKT and subsequent AKT-activated Snail gene ex-pression.Taken together, our findings suggest that a new mechanism of
TADCs promotes lung cancer progression, in which TADC-derivedamphiregulin causes phenotypic transition and reinforces cancerprogression. Amphiregulin enhances the cooperation of STAT3 andAKT signaling and subsequently increases the expression of Twistand Snail, resulting in the promotion of lung cancer development.Neutralization of amphiregulin by amphiregulin Ab significantlydecreases the incidence of cancer development in mice. Our find-ings provide important insights into the biology of tumors withvicious interactions of DCs and cancer as well as establish newrationales for using anti-amphiregulin strategies to target aggres-sive invasive lung cancer.
DisclosuresThe authors have no financial conflicts of interest.
References1. Jemal, A., R. Siegel, E. Ward, Y. Hao, J. Xu, and M. J. Thun. 2009. Cancer
statistics, 2009. CA Cancer J. Clin. 59: 225–249.2. Doedens, A. L., C. Stockmann, M. P. Rubinstein, D. Liao, N. Zhang,
D. G. DeNardo, L. M. Coussens, M. Karin, A. W. Goldrath, and R. S. Johnson.2010. Macrophage expression of hypoxia-inducible factor-1a suppresses T-cellfunction and promotes tumor progression. Cancer Res. 70: 7465–7475.
3. Barnas, J. L., M. R. Simpson-Abelson, S. P. Brooks, R. J. Kelleher, Jr., andR. B. Bankert. 2010. Reciprocal functional modulation of the activation ofT lymphocytes and fibroblasts derived from human solid tumors. J. Immunol.185: 2681–2692.
4. Plantinga, M., H. Hammad, and B. N. Lambrecht. 2010. Origin and functionalspecializations of DC subsets in the lung. Eur. J. Immunol. 40: 2112–2118.
5. Kuo, P. L., J. Y. Hung, S. K. Huang, S. H. Chou, D. E. Cheng, Y. J. Jong,C. H. Hung, C. J. Yang, Y. M. Tsai, Y. L. Hsu, and M. S. Huang. 2011. Lungcancer-derived galectin-1 mediates dendritic cell anergy through inhibitor ofDNA binding 3/IL-10 signaling pathway. J. Immunol. 186: 1521–1530.
6. Perrot, I., D. Blanchard, N. Freymond, S. Isaac, B. Guibert, Y. Pacheco, andS. Lebecque. 2007. Dendritic cells infiltrating human non-small cell lung cancerare blocked at immature stage. J. Immunol. 178: 2763–2769.
7. Robinson-Smith, T. M., I. Isaacsohn, C. A. Mercer, M. Zhou, N. Van Rooijen,N. Husseinzadeh, M. M. McFarland-Mancini, and A. F. Drew. 2007. Macro-phages mediate inflammation-enhanced metastasis of ovarian tumors in mice.Cancer Res. 67: 5708–5716.
8. Lu, X., Q. Wang, G. Hu, C. Van Poznak, M. Fleisher, M. Reiss, J. Massague, andY. Kang. 2009. ADAMTS1 and MMP1 proteolytically engage EGF-like ligandsin an osteolytic signaling cascade for bone metastasis. Genes Dev. 23: 1882–1894.
9. Yamada, M., Y. Ichikawa, S. Yamagishi, N. Momiyama, M. Ota, S. Fujii,K. Tanaka, S. Togo, S. Ohki, and H. Shimada. 2008. Amphiregulin is a prom-ising prognostic marker for liver metastases of colorectal cancer. Clin. CancerRes. 14: 2351–2356.
10. Gilmore, J. L., R. M. Gonterman, K. Menon, G. Lorch, D. J. Riese, II,A. Robling, and J. Foley. 2009. Reconstitution of amphiregulin-epidermalgrowth factor receptor signaling in lung squamous cell carcinomas activatesPTHrP gene expression and contributes to cancer-mediated diseases of the bone.Mol. Cancer Res. 7: 1714–1728.
11. Li, X. D., S. Y. Miao, G. L. Wang, L. Yang, Y. Q. Shu, and Y. M. Yin. 2010.Amphiregulin and epiregulin expression in colorectal carcinoma and the corre-lation with clinicopathological characteristics. Onkologie 33: 353–358.
12. Yotsumoto, F., T. Fukami, H. Yagi, A. Funakoshi, T. Yoshizato, M. Kuroki, andS. Miyamoto. 2010. Amphiregulin regulates the activation of ERK and Aktthrough epidermal growth factor receptor and HER3 signals involved in theprogression of pancreatic cancer. Cancer Sci. 101: 2351–2360.
13. Bles, N., L. Di Pietrantonio, J. M. Boeynaems, and D. Communi. 2010. ATPconfers tumorigenic properties to dendritic cells by inducing amphiregulin se-cretion. Blood 116: 3219–3226.
14. Addison, C. L., K. Ding, H. Zhao, A. Le Maıtre, G. D. Goss, L. Seymour,M. S. Tsao, F. A. Shepherd, and P. A. Bradbury. 2010. Plasma transforminggrowth factor a and amphiregulin protein levels in NCIC Clinical Trials GroupBR.21. J. Clin. Oncol. 28: 5247–5256.
15. Berasain, C., E. R. Garcıa-Trevijano, J. Castillo, E. Erroba, M. Santamarıa,D. C. Lee, J. Prieto, and M. A. Avila. 2005. Novel role for amphiregulin inprotection from liver injury. J. Biol. Chem. 280: 19012–19020.
16. Lo, H. W., S. C. Hsu, W. Xia, X. Cao, J. Y. Shih, Y. Wei, J. L. Abbruzzese,G. N. Hortobagyi, and M. C. Hung. 2007. Epidermal growth factor receptorcooperates with signal transducer and activator of transcription 3 to induceepithelial-mesenchymal transition in cancer cells via up-regulation of TWISTgene expression. Cancer Res. 67: 9066–9076.
17. Kuo, P. L., Y. H. Chen, T. C. Chen, K. H. Shen, and Y. L. Hsu. 2011. CXCL5/ENA78 increased cell migration and epithelial-to-mesenchymal transition ofhormone-independent prostate cancer by early growth response-1/snail signalingpathway. J. Cell. Physiol. 226: 1224–1231.
18. Vicent, S., D. Luis-Ravelo, I. Anton, I. Garcıa-Tunon, F. Borras-Cuesta, J. Dotor,J. De Las Rivas, and F. Lecanda. 2008. A novel lung cancer signature mediatesmetastatic bone colonization by a dual mechanism. Cancer Res. 68: 2275–2285.
19. Hsu, Y. L., J. Y. Hung, Y. C. Ko, C. H. Hung, M. S. Huang, and P. L. Kuo. 2010.Phospholipase D signaling pathway is involved in lung cancer-derived IL-8 in-creased osteoclastogenesis. Carcinogenesis 31: 587–596.
20. Kim, M. Y., T. Oskarsson, S. Acharyya, D. X. Nguyen, X. H. Zhang, L. Norton,and J. Massague. 2009. Tumor self-seeding by circulating cancer cells. Cell 139:1315–1326.
21. McAllister, S. S., and R. A. Weinberg. 2010. Tumor-host interactions: a far-reaching relationship. J. Clin. Oncol. 28: 4022–4028.
22. Xian, C. J. 2007. Roles of epidermal growth factor family in the regulation ofpostnatal somatic growth. Endocr. Rev. 28: 284–296.
23. Schuger, L., G. R. Johnson, K. Gilbride, G. D. Plowman, and R. Mandel. 1996.Amphiregulin in lung branching morphogenesis: interaction with heparan sulfateproteoglycan modulates cell proliferation. Development 122: 1759–1767.
24. Berasain, C., J. Castillo, M. J. Perugorrıa, J. Prieto, and M. A. Avila. 2007.Amphiregulin: a new growth factor in hepatocarcinogenesis. Cancer Lett. 254:30–41.
25. Willmarth, N. E., and S. P. Ethier. 2008. Amphiregulin as a novel target forbreast cancer therapy. J. Mammary Gland Biol. Neoplasia 13: 171–179.
26. Hurbin, A., J. L. Coll, L. Dubrez-Daloz, B. Mari, P. Auberger, C. Brambilla, andM. C. Favrot. 2005. Cooperation of amphiregulin and insulin-like growth factor-1 inhibits Bax- and Bad-mediated apoptosis via a protein kinase C-dependentpathway in non-small cell lung cancer cells. J. Biol. Chem. 280: 19757–19767.
27. Eckstein, N., K. Servan, L. Girard, D. Cai, G. von Jonquieres, U. Jaehde,M. U. Kassack, A. F. Gazdar, J. D. Minna, and H. D. Royer. 2008. Epidermalgrowth factor receptor pathway analysis identifies amphiregulin as a key factorfor cisplatin resistance of human breast cancer cells. J. Biol. Chem. 283: 739–750.
28. Behera, R., V. Kumar, K. Lohite, S. Karnik, and G. C. Kundu. 2010. Activationof JAK2/STAT3 signaling by osteopontin promotes tumor growth in humanbreast cancer cells. Carcinogenesis 31: 192–200.
29. Barbieri, I., S. Pensa, T. Pannellini, E. Quaglino, D. Maritano, M. Demaria,A. Voster, J. Turkson, F. Cavallo, C. J. Watson, et al. 2010. Constitutively activeStat3 enhances neu-mediated migration and metastasis in mammary tumors viaupregulation of Cten. Cancer Res. 70: 2558–2567.
30. Catalano, S., C. Giordano, P. Rizza, G. Gu, I. Barone, D. Bonofiglio,F. Giordano, R. Malivindi, D. Gaccione, M. Lanzino, et al. 2009. Evidence thatleptin through STAT and CREB signaling enhances cyclin D1 expression andpromotes human endometrial cancer proliferation. J. Cell. Physiol. 218: 490–500.
31. Hung, J. J., M. H. Yang, H. S. Hsu, W. H. Hsu, J. S. Liu, and K. J. Wu. 2009.Prognostic significance of hypoxia-inducible factor-1a, TWIST1 and Snail ex-pression in resectable non-small cell lung cancer. Thorax 64: 1082–1089.
32. Fukumoto, J., C. Harada, T. Kawaguchi, S. Suetsugu, T. Maeyama, I. Inoshima,N. Hamada, K. Kuwano, and Y. Nakanishi. 2010. Amphiregulin attenuatesbleomycin-induced pneumopathy in mice. Am. J. Physiol. Lung Cell. Mol.Physiol. 298: L131–L138.
33. Qian, J., Y. Zou, J. S. Rahman, B. Lu, and P. P. Massion. 2009. Synergy betweenphosphatidylinositol 3-kinase/Akt pathway and Bcl-xL in the control of apo-ptosis in adenocarcinoma cells of the lung. Mol. Cancer Ther. 8: 101–109.
1744 AMPHIREGULIN INVOLVED IN CANCER–IMMUNE SYSTEM INTERACTION
by guest on April 10, 2019
http://ww
w.jim
munol.org/
Dow
nloaded from


















