
Long-Term Effects of Spinal Cord Transection on Fast and...
14
EXPERIMENTAL NEUROLOGY 91,435-448 (1986) Long-Term Effects of Spinal Cord Transection on Fast and Slow Rat Skeletal Muscle II. Morphometric Properties RICHARD L. LIEBER, JAN 0. FRID~N, ALAN R. HARGENS, AND EARL R. FBRINGA’ Division of Orthopaedics and Rehabilitation, Departments of Surgery and Neurology, Veterans Administration Medical Center and University of California, San Diego, California 92161 Received June 26, 1985; revision received October 14. I985 Morphometric properties of rat soleus and extensor digitorum longus muscles were studied 1 year following complete thoracic spinal cord transection (spinal cord level T9). Both muscles demonstrated almost complete type 1 to type 2 muscle fiber type conversion after 1 year. Muscle fiber atrophy was observed in both muscles. Type 2 fiber atrophy occurred to about the same extent in both muscles. Atrophy was most severe for the soleus type 1 fibers (50% decrease in size). Calculations based on the fiber type and size changes observed indicate that the percentage of the muscle cross- sectional area occupied by each fiber type was almost the same for both muscles 1 year after transection. Discriminant analysis of the data indicated that the percentage of type 2 fibers present in the muscle was the best discriminator between the various groups. These morphometric data provided a basis for understanding the contractile results presented in the previous study as well as insights into the mechanism of trans- formation in skeletal muscle. Furthermore, inherent differences between type 1 and type 2 fibers were demonstrated between predominantly slow and predominantly fast muscles. Thus, after almost one-half a lifetime of transection, rat muscles are almost completely transformed to fast muscle, and, regardless of initial conditions, have nearly identical properties. Q 1986 Academic Ress Inc. Abbreviations: SOL-soleus, EDL-extensor digitorum longus, SDH-succinate dehydroge- nase, MC-medial gastrocnemius. ’ The authors thank Dr. Roland R. Roy for his helpful suggestion regarding data presentation and Carina B. Johansson for her careful technical work. Please address reprint requests to Dr. R. L. Lieber, Division of Orthopedics and Rehabilitation, V- I5 1, VAMC, San Diego, CA 92 16 1. Supported by the Veterans Administration. 435 00 14-4886/86 $3.00 Copyri@t 0 1986 by Academic Press, Inc. All rights of reproduction in any form resewed.
Transcript of Long-Term Effects of Spinal Cord Transection on Fast and...

EXPERIMENTAL NEUROLOGY 91,435-448 (1986)
Long-Term Effects of Spinal Cord Transection on Fast and Slow Rat Skeletal Muscle
II. Morphometric Properties
RICHARD L. LIEBER, JAN 0. FRID~N, ALAN R. HARGENS, AND EARL R. FBRINGA’
Division of Orthopaedics and Rehabilitation, Departments of Surgery and Neurology, Veterans Administration Medical Center and University of California, San Diego, California 92161
Received June 26, 1985; revision received October 14. I985
Morphometric properties of rat soleus and extensor digitorum longus muscles were studied 1 year following complete thoracic spinal cord transection (spinal cord level T9). Both muscles demonstrated almost complete type 1 to type 2 muscle fiber type conversion after 1 year. Muscle fiber atrophy was observed in both muscles. Type 2 fiber atrophy occurred to about the same extent in both muscles. Atrophy was most severe for the soleus type 1 fibers (50% decrease in size). Calculations based on the fiber type and size changes observed indicate that the percentage of the muscle cross- sectional area occupied by each fiber type was almost the same for both muscles 1 year after transection. Discriminant analysis of the data indicated that the percentage of type 2 fibers present in the muscle was the best discriminator between the various groups. These morphometric data provided a basis for understanding the contractile results presented in the previous study as well as insights into the mechanism of trans- formation in skeletal muscle. Furthermore, inherent differences between type 1 and type 2 fibers were demonstrated between predominantly slow and predominantly fast muscles. Thus, after almost one-half a lifetime of transection, rat muscles are almost completely transformed to fast muscle, and, regardless of initial conditions, have nearly identical properties. Q 1986 Academic Ress Inc.
Abbreviations: SOL-soleus, EDL-extensor digitorum longus, SDH-succinate dehydroge- nase, MC-medial gastrocnemius.
’ The authors thank Dr. Roland R. Roy for his helpful suggestion regarding data presentation and Carina B. Johansson for her careful technical work. Please address reprint requests to Dr. R. L. Lieber, Division of Orthopedics and Rehabilitation, V- I5 1, VAMC, San Diego, CA 92 16 1. Supported by the Veterans Administration.
435
00 14-4886/86 $3.00 Copyri@t 0 1986 by Academic Press, Inc. All rights of reproduction in any form resewed.

436 LIEBER ET AL.
INTRODUCTION
As discussed in the previous paper (13), skeletal muscle demonstrates the ability to adapt to a changing environment. Changes in contractile properties are manifestations of structural changes within the muscle. The correlation between skeletal muscle structural and functional properties soon after cor- dotomy has been studied in single cat motor units ( 15), whole cat muscle (2, 18), and whole rat muscle (5). Type 1 to type 2 muscle fiber transformation has been observed in cat single motor units ( 15), cat whole muscle (2), whole guinea pig muscle (1 l), and whole rat muscle (5). The extent of fiber type transformation observed following cordotomy varies between studies. Several reports suggest that muscle fiber atrophy affects type 1 and type 2 fibers to the same extent (3, 11, 15) whereas others suggest that type 2 fibers are affected to a greater extent than type 1 fibers (7, 12). Based on these conflicting reports, the extent and type of adaptation which occurs after spinal cord transection is unclear. In this paper, we present a model which results in almost complete type 1 to type 2 fiber type transformation permitting unambiguous charac- terization of type 1 and type 2 fiber properties from a predominantly slow muscle (soleus, SOL) and a predominantly fast muscle (extensor digitorum longus, EDL). Our purpose was thus to determine the morphometric properties of the rat soleus and extensor digitorum longus 1 year (one-half to one-third the life span of the rat) after spinal cord transection.
METHODS
Experimental Preparation. Experimental animals, surgical cordotomy, and postoperative care were as described in the preceding report ( 13).
Histologic and Histochemical Preparation. After measurement of contractile properties, muscles were excised, blotted dry, stretched to & (the length which produced maximum twitch tension), and frozen in isopentane cooled by liquid nitrogen (-159°C). Muscles were stored at -80°C prior to sectioning. Eight- micrometer sections were cut at -20°C on a Tissue-Tek II cryostat (Lab-Tek, Naperville, Ill.). Sections were cut from the midportion of the muscle belly, air-dried, and stained for general morphologic features using hematoxylin and eosin. Further sections were stained for myofibrillar ATPase activity using a modification of the methods of Guth and Samaha (10). Muscle sections were preincubated at pH 4.20 and 10.00 with a short-term accuracy of kO.003 pH units as determined by an Altek model pHi 71 pH meter (Beckman In- struments, Irvine, Calif.). Long-term accuracy (during a period of hours) was kO.02 pH units. Relative succinate dehydrogenase (SDH) activity was deter- mined using the methods of Nachlas et al. (17). Inspection of SDH and ATPase serial sections permitted classification of fibers into type 1, type 2A, and type 2B.

MUSCLE MORPHOLOGY FOLLOWING CORLIOTOMY 437
Terminology. In this report, we claim that fast muscle fibers, as measured in the previous study (13) are identical to the type 2 fibers demonstrated histologically in this study. Similarly, slow fibers are assumed to be identical to type 1 fibers. The fibers are referred to by physiologic type (fast or slow) when discussing contractile properties and by their histochemical type (type 2 or type 1) when discussing morphologic properties.
For the sake of brevity, muscles from cordotomized rats will be referred to as “transected muscles,“e.g., the transected EDL.
Morphometric Measurements. Morphometric properties were determined by projecting the microscope slides onto a calibrated loo-intersection test grid using a Nikon Micropan Microslide projector (Nikon, Inc., Garden City, N.Y.) at magnifications between 100X and 500X. Six regions per slide were sampled using a modification of the systematic sampling procedures described by Weibel (23). The total number of fibers sampled was approximately 400 per muscle. Fields that contained cracks, obliquely sectioned fibers, or outer borders of the section were ignored. Measurements were carried out by a single investigator during a 2-day period.
The following morphometric parameters were measured using the point- counting technique of Weibel(23) as implemented by Eisenberg (8): (i) average type 1 fiber area (pm*), (ii) average type 2 fiber area (pm*), (iii) percentage extracellular space, (iv) percentage type 1 fibers, and (v) percentage type 2 fibers.
Statistical Analysis. Morphometric properties of the SOL and EDL were analyzed separately using the BMDP program PlV (6). For each parameter listed above, a one-way ANOVA was used to compare means between the four groups (normal EDL, transected EDL, normal SOL, transected SOL). Following the preliminary ANOVA, multiple t tests were used to compare means between all possible pairs of four groups. Thus, the significance levels reported compensate for the number of comparisons made (21). Differences between groups were considered significant for P c 0.05. Following the AN- OVA, discriminant analysis was performed using BMDP program P7M with F-to-enter = 4.000 and F-to-remove = 3.996. This was done to determine which parameter(s) best differentiate(s) between the four muscle groups.
RESULTS
General inspection of muscle fibers from transected SOL and EDL muscle showed noticeable fiber atrophy, with increased endomysial and perimysial connective tissue. Nuclei were peripheral and of uniform staining intensity. No signs of degeneration, regeneration, or fiber type grouping were apparent.
Low magnification cross sections of SOL and EDL muscles stained for myofibrillar ATPase are presented in Figs. 1 and 2, respectively. Note the

438 LIEBER ET AL.
IWi. 1. Low-magnification micrograph of soleus (SOL) muscle myofibrillar ATPase activity after preincubation at pH 10.0. In this preparation, type 2 fibers appeared dark. A-normal SOL, B-transected SOL. Calibration bar = 0.5 mm. Note the dramatic type 1 to type 2 muscle fiber conversion that followed transection.
MUSCLE MORPHOLOGY FOLLOWING CORDOTOMY
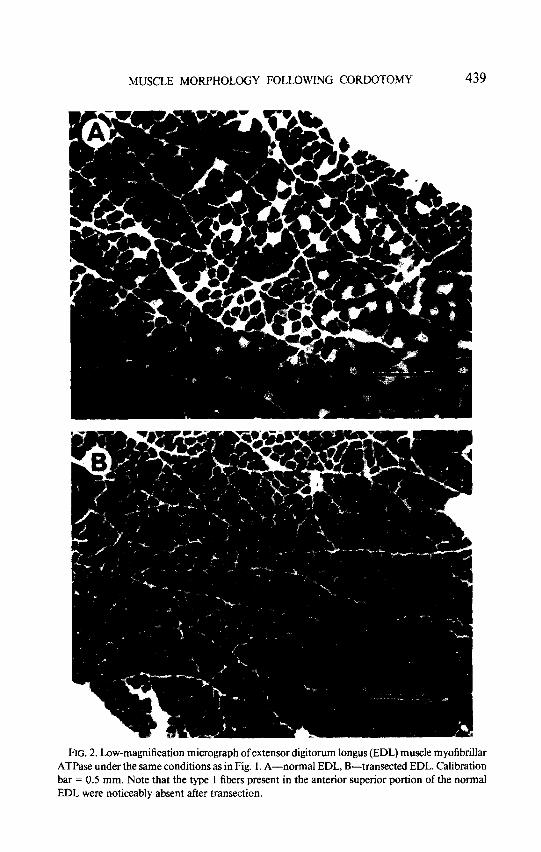
RG. 2. Low-magnification micrograph of extensor digitorum longus (EDL) muscle myofibrillar ATPase under the same conditions as in Fig. 1. A-normal EDL, B-transected EDL. Calibration bar = 0.5 mm. Note that the type 1 fibers present in the anterior superior portion of the normal EDL were noticeably absent after transection.

440 LIEBER ET AL.
scattered distribution of type 2 fibers in the normal SOL (Fig. 1A) contrasted with the transected SOL (Fig. 1B) where almost the entire muscle was com- posed of type 2 fibers. At higher magnification (Fig. 3), note the decrease in the size of both the type 1 and type 2 fibers. The changes in the EDL were much less dramatic but proceeded in the same direction. The normal EDL (Fig. 2A) contained scattered type 1 fibers (light staining) in the anterior su- perficial region of the muscle. However, after transection, few type 1 fibers were visible (Fig. 2B).
A summary of all morphometric parameters measured is presented in Table 1 and selected parameters in Fig. 4. The morphometric changes observed to follow transection were similar for both muscles although the magnitude of the changes was greater for the SOL. As can be appreciated from the micro- graphs, the percentage of type 1 muscle fibers decreased significantly for both muscles. The average type 1 fiber area decreased significantly by about 50% for the SOL (Fig. 4B) but not for the EDL (Fig. 4B). The percentage of type 2 fibers increased significantly for both muscles (Fig. 4C). Again, the magnitude of the increase was greater for the SOL. Type 2 fiber area decreased to about the same extent for both muscles (Fig. 4D). The percentage of extracellular area [area fraction (23)] increased significantly for both muscles by about the same amount. Thus, both muscles demonstrated fiber atrophy, a significant type 1 to type 2 fiber transformation, and a significant increase in extracellular (both perimysial and epimysial) connective tissue.
A summary of the discriminant analysis on the morphometric parameters is presented in Table 2. Note that for most comparisons, the percentage of type 2 fibers was the best discriminator between groups. The most difficult groups to discriminate between were the transected EDL and transected SOL. Interestingly, the best discriminator between these groups was the percentage of extracellular space. A graphic representation of the discriminant analysis is presented in Fig. 5. The normal SOL was quite different than any of the other three groups, mainly due to the low percentage of type 2 muscle fibers (main component of horizontal axis canonical variable). The normal EDL, transected EDL, and transected SOL contained nearly the same percentage of type 2 fibers but differed in percent extracellular area (main component of vertical axis canonical variable). Thus, by considering both percent type 2 fibers and percent extracellular area, fair discrimination between groups was possible (56 to 100% reliability).
DISCUSSION
Because spinal cord transection results in reduced muscle use (1, 18), it is expected that skeletal muscle properties would change following transection. This study revealed that the morphometric properties of the SOL and EDL

MUSCLE MORPHOLOGY FOLLOWING CORDOTOMY 441
FIG. 3. Micrograph of SOL muscle myofibrillar ATPase activity under same conditions as in Fig. 1. A-normal SOL, B-transected SOL. Calibration bar = IOOrm. Note the decrease in size of the type 1 fibers in the transected muscle. Type 2 fibers decreased but to a lesser extent.

TABLE 1
Summary of Morphometry Data from Four Groups of Bat Muscles”
Normal (SE, N) Transected (SE, N)
Parameter EDL SOL EDL SOL
No. type 1 fibers measured
No. type 2 fibers measured
Type 1 area (pm’)
Type 2 area (pm’)
Ext. cell. area (96 of total)
%Typelfibers
IType2fibers
% Type 1 in cross section
% Type. 2 in cross section
19.3 *- * 268 I;-* 2.1 o- 0 12.3 (2.4, 13) (14.2, 13) (1.6, 9) (4.4, 7)
* * * *
* *
328 *- 40.0 *-* 435 t- * 501 (14, 13) (4.4, 13) (18,9) (20,7)
* * * l
* *
3733 v-s 6731 *- z 36420 -0 4604 (236, 13) (345, 13) (42,2) (80496)
0 0 * *
0 0
5899 +-* 4263 0 -0 4340 -* 3408 (245, 13) (241, 13) (212,9) (18597)
* * -*
* *
(l.:,813) *- 12.0 *-* 14.7 -* 20.0 (0.89, 13) (0.78, 9) (1.3,7)
* * * *
* *
(0.6;,513) *-* 86.8 *-* 0.43 0 -0 2.5 (1.5, 13) (0.3 1, 9) (0.84, 7)
* * * *
* *
94.5 a----s 13.2 -* 99.6 O- 0 97.5 (0.65, 13) (1.5, 13) (0.31,9) (0.84, 7)
* * * *
* *
3.51 e---e 91.25 * -* 0.413 o-----o 3.12 (0.46, 13) (0.90, 13) (0.31, 9) (1.2, 7)
(; *
0
96.5 *-a 8.74 *--* 99.6 O- 0 96.8 (0.46, 13) (0.90, 13) (0.31, 9) (1.237)
;, *
0
’ Values are connected by an asterisk if the means differed significantly (P -c 0.05) or by an open circle if the means were not significantly different.
442

MUSCLE MORPHOLOGY FOLLOWING CORDOTOMY 443
125 A
2
100
::
ii 75 !?I
- F 50
k 25
0 :
125 .c
2 loo-
::
i 75 Lt. c-4
F
50:
I- 25:
0 PERCENT TYPE 1 FIBERS 7 TYPE 1 FIBER AREA
B6
I--
S -
N‘ 5
E 4 3 a 3
; 2
1
0
SOLEUS EDL p<O.OOl P’O.001
PERCENT TYPE 2 FIBERS
SOLEUS EDL p-co.02 p>O.8
ED 7 c
TYPE 2 FIBER AREA
sf3 8
5 5
E 4 -3
,Q
3
2 2
1
0 0
SOLEUS EDL SOLEUS EDL p<O.OOl pio.001 pxo.03 P<O.OOl
FIG. 4. Bar graphs of type 1 and type 2 fiber characteristics. For all graphs, the open bars represent normal muscle and the hatched bats transected muscle. Left sets of bars represent SOL muscles, right sets represent EDL muscles. Level of significance for a difference between means is listed beneath each muscle group and was determined using the multiple t test. A-percentage of type 1 fibers, B-average type 1 fiber diameter, C-percentage of type 2 fibers, D-average type 2 fiber diameter.
became almost identical after transection. Both muscles demonstrated almost complete type 1 to type 2 muscle fiber transformation with accompanying type 2 fiber atrophy. Such a complete change has not before been demontrated in animals and probably reflects the fact that the study period (1 year) com- prises almost one-half the life span of the normal rat. Near-complete trans- formations have been observed in humans to follow traumatic cord lesion (9). In spite of the similar changes observed in both muscles, several changes were observed which differ between muscles and fiber types.
Fiber Type and Size Changes. The similar degree of type 2 fiber atrophy (about 25%, Fig. 4D) in both SOL and EDL muscles may imply that the same driving force is acting on the two muscles after cordotomy. This similarity is especially interesting in light of the fact that in the transected SOL, most type 2 fibers present are transformed type 1 fibers, whereas in the EDL most type 2 fibers present are atrophied type 2 fibers. The transformation of type 1 SOL fibers was to the type 2A subtype, indicating that oxidative capacity did not change substantially after transformation. A greater degree of type 2 fiber atrophy (about 40%) has been observed in plantar flexors (medial gastroc-

444 LIEBER ET AL.
TABLE 2
Summary of Morphometry Discriminant Analysis’
Groups compared Best discriminators Percentage correct
(in order) decisions (%)
N-EDL vs. N-SOL
N-EDL vs. X-EDL
N-SOL vs. X-SOL
X-EDL vs. X-SOL
N-EDL vs. N-SOL vs. X-EDL vs. X-SOL
% Type 2, Type 1 area % Extracellular area
% Type 2, % Extracellular area
%Type2
% Extracellular area, % Type 2 fibers
%Type2,#Type2fibers, 96 Extracellular area
100
100
100
86-100
56-100
‘F-to-enter = 4.000, F-to-remove = 3.996. The scatter plot of the data indicated the most difficulty discriminating between transected EDL (X-EDL) and transected SOL (X-SOL); N, normal. When examining all four groups, incorrect classifications are: 1 N-EDL classified as X- EDL, 3 X-EDL classified as X-SOL, 1 X-EDL classified as N-EDL, and 1 X-SOL classified as X- EDL. Correct classifications were 92% of N-EDL (12/l 3), 100% of N-SOL ( 13/ 13), 56% of X-EDL (5/9), and 86% of X-SOL (6/7).
nemius) of the cat ( 15), guinea pig ( 1 1 ), and rat ( 12) to follow shorter periods of cordotomy. The atrophy and transformation observed is most severe be- ginning the first few months after cordotomy [when degenerative changes have also been reported, ( 1 1 )].
The significant type 1 fiber atrophy observed in the SOL (Fig. 4A) but not the EDL (Fig. 4A) is further evidence that type 1 fibers in predominantly slow muscles differ from those in mixed muscles. In this regard they differ from the type 2 fibers which seem to respond similarly in both muscles. Observations of both type 1 fiber atrophy ( 11,15) and hypertrophy ( 12) have been reported for a number of species. The magnitude and direction of these changes also varies relative to the type 2 fiber changes presented in the same studies. It is clear from these studies that the behavior of the type 1 fibers is less well understood. Some of these difficulties may arise from sampling problems resulting from making measurements of the least prevalent fiber type in the muscle (only three of nine transected EDL muscles contained any type 1 fibers at all).
The functional consequences of the fiber type and size changes reported in this study are best appreciated by calculating the percentage of the total muscle area occupied by each fiber type before and after transection. Based on the average fiber area and percentage of each fiber type present, the percent of the muscle cross-sectional occupied by each type was calculated for each mus-

MUSCLE MORPHOLOGY FOLLOWING CORDOTOMY 445
N
-6L."'i"'.""""""""..""""""""~ -25 -20 -15 -10 -5 0 5 10 15
Cononicol Variable 1 0
FIG. 5. Graphic representation of the discriminant analysis presented in Table 2 for all four groups. The horizontal axis canonical variable reflects percentage of type 2 fibers. The vertical axis canonical variable reflects the percentage of extracellular space. Cl, normal SOL: 0 transected SOL: A, normal EDL: and 6, transected EDL. Note the normal SOL was very different from either of the other groups which were very similar based on fast fiber percentage alone. Differences in transected groups and normal EDL were manifested by differences in the percentage of extra- cellular space.
cle studied. The results, presented in Fig. 6, show that although the normal EDL and SOL initially contain very different fiber type and size distributions (Fig. 6A), 1 year after transection the two muscles are, in terms of fiber type, very similar (Fig. 6B). Interestingly, the two most similar muscles, 1 year after transection, are the normal EDL and transected SOL.
The 50% type 1 fiber atrophy observed in the SOL (but not in the EDL) apparently has little functional consequence as so few type 1 fibers remain
s $ 100
BEfORE TRANSECTION AFTER TRANSECTION
d s 80
% 2 60 I R 8 40
E 20 E
t 0 Sol*ur EDL EDL
FIG. 6. Bar graphs representing the percentage of muscle cross-sectional area occupied by each fiber type, calculated algebraically for each muscle using the average fiber area and percentage of each fiber type.. A-percentages of normal SOL and normal EDL cross sections occupied by type 1 fibers (open bars) and type 2 fibers (hatched bars). B-percentages of transected SOL and transected EDL cross sections occupied by type 1 fibers (open bars) and type 2 fibers (hatched bars). Note that although the normal distributions of the two muscles differed greatly, after tran- section they became nearly identical.

446 LIEBER ET AL.
after transection. Similarly, it is not surprising that the type 1 to type 2 trans- formation observed in the EDL would have little effect on its contractile properties, as very few type 1 fibers (~5%) are initially present in the muscle. One year after transection, almost no change is observed in the relative amount of muscle cross section occupied by type 1 fibers in the SOL and type 2 fibers in the EDL (Fig. 6B).
SpeciJic Tension. Although in the EDL we observed a 25% decrease in fast fiber area and 50% increase in extracellular space, resulting in a net decrease in cross-sectional area of 2% (Table l), no decrease in maximum tetanic tension (PO) was seen [Table 1 of (13)]. That such a decrease was not observed may be explained several ways. First, cordotomy may lead to an increase in specific tension of the individual fast muscle fibers. Second, muscle architecture of the EDL may change after cordotomy resulting in an increase of the true physiologic cross-sectional area which is not reflected in measurement of geo- metric cross-sectional area from muscle sections [architectural changes can account for apparent specific tension changes following immobilization at different muscle lengths (20, 22) but apparently do not occur in the cat fol- lowing transection (1 S)]. The only comparable studies of both morphometric and physiologic properties were on whole cat medial gastrocnemius (MG) and soleus (2, 18), and single motor units of the cat MG (15). A decrease in specific tension was observed for the cat fast fatigue-resistant and slow motor units (4, 15) and whole MG ( 18) in spite of the type 1 to type 2 transformation reported. This may not conflict with the present study based on the evidence that the specific tension of cat slow and fast fibers may not differ. The high specific tension reported in this study (13) for the whole rat transected SOL is the largest value reported in the literature. This value is even more surprising in light of the 20% area fraction of extracellular space present (Table 1) which would result in an underestimation of the specific tension of the muscle fibers themselves. It is difficult to obtain accurate estimates of specific tension from whole muscle studies, and thus, it would be interesting to monitor the con- tractile properties of isolated SOL muscle fibers before and after transection.
Mechanism of Fiber Transformation. It is interesting to compare the cor- dotomy model of transformation with other models reported. The only other model reported to cause as complete a type 1 to type 2 fiber transformation as that reported here is the denervated rat soleus subjected to chronic high- frequency stimulation (14). It is tempting to hypothesize a similar mechanism for the changes observed in two studies-namely that cordotomy results in a prolonged high-frequency input to the muscle via the peripheral nerves ( 19). This explanation is unlikely for two reasons. First, no change in muscle ox- idative capacity, as indicated by SDH staining intensity, was observed after cordotomy (data not shown), but a large increase in oxidative capacity and fatigue index was observed after prolonged electrical stimulation (14). These

MUSCLE MORPHOLOGY FOLLOWING CORDOTOMY 447
data are supported by the observations of Roy et al. (18) and Baldwin et al. (2) who reported no change in cat muscle oxidative capacity (as measured histochemically and biochemically) and fatigue index. Second, in cats, the EMG profile demonstrates decreased electrical activity soon after cordotomy (I), making it unlikely that electrical activity of the peripheral nerve induced the changes observed.
A second explanation for the observed transformation may be that the prolonged immobilization secondary to cordotomy causes type 1 to type 2 fiber type transformation, independent of the status of the central nervous system. This explanation is unlikely based on the wealth of data acquired from long-term immobilization studies on the rat (22), cat (16), and guinea pig (11). In general, immobilization causes muscle fiber atrophy (which may be fiber type-specific) with little or no change in fiber type percentage-even after extended periods.
The observed transformation does not reflect selective degeneration of slow alpha-motor neurons accompanying the denervation-reinnervation process. This conclusion is based on the observation that no fiber type grouping was observed in this preparation or others (11, 15, 18) and that the slow fibers present, although few, were randomly scattered throughout the muscle.
Skeletal muscle fiber trnsformation is thus not a function of only electrical activity, tension, or innervation. In addition, contractile speed and myofibrillar ATPase activity can change significantly without a corresponding change in muscle oxidative capacity. The actual stimulus to transformation is therefore c&cted by muscle activity, tension, and neuromuscular status but is not solely a function of one of those parameters.
REFERENCES
1. ALAIMO, M. A., J. L. SMITH, R. R. ROY, AND V. R. EDGERTON. 1984. EMG activity of slow and fast ankle extensors following spinal cord transection. J. Appl. Physiol.: Resp. Environ. Exercise Physiol. 56: 1608- 16 13.
2. BALDWIN, K. M., R. R. ROY, R. D. SACKS, C. BLANCO, AND V. R. EDGERTON. 1984. Relative independence of metabolic enzymes and neuromuscular activity. .I Appl. Physiol.: Respir. Environ. Exercise Physiol. 56 (6): 1602- 1607.
3. BROOK& M. H., AND W. K. ENGEL. 1969. The histographic analysis of human muscle biopsies with regard to fiber types. 2. Diseases of the upper and lower motor neurons. Neurology 19: 378-393.
4. BURKE, R. E., D. N. LEVINE P. TSAIRIS, AND F. E. ZAJAC. 1973. Physiological types and histochemical profiles in motor units of the cat gastrocnemius. .I. Physiol. (London) 234: 723-748.
5. DAVEY, D. F., C. DUNLOP, J. F. Y. HOH, AND S. Y. P. WONG. 198 1. Contractile properties and ultrastructure of extensor digitorum longus and soleus muscles in spinal cord transected rats. Amt. J. Exp. Biol. Med. Sci. 59: 383-404.
6. DIXON, W. J. 1983. BMDP Statistical Software. Univ. of California Press, Los Angeles.

448 LIEBER ET AL.
7. EDSTROM, L. 1970. Selective changes in the size of red and white muscle fibers in upper motor neuron lesion and Parkinsonism. J. Net&. Sci. 11: 523-550.
8. EISENBERG, B. R. 1983. Quantitative ultrastructure of mammalian skeletal muscle. Handbook ofPhysiology, Chap. 3 Amer. Physiol. Sot., Bethesda, Maryland.
9. GRIMBY, G., C. BROBERG, I. KROTKIEWSKA, AND M. KROTKIEWSKI. 1976. Muscle fiber composition in patients with traumatic cord lesion. &and. J. Rehab. Med. 8: 37-42.
10. GUTH, L., AND J. SAMAHA. 1969. Qualitative differences between actomyosin ATPase of slow and fast mammalian muscle. Exp. Neurobiol. 25: 138 152.
1 I. KARPATI, G., AND W. K. ENGEL. 1968. Correlative histochemical study of skeletal muscle after suprasegmental denervation, peripheral nerve section, and skeletal fixation. Neurology 18: 681-692.
12. KINOSHITA, M., K. KAWASAKI, Y. SUZUKI, N. KAMEI, AND E. SATAYOSHI. 1976. Type 1 fiber hypertrophy in the “spastic muscle.” Clin. Neurol. (Tokyo) 16: 417-423.
13. LIEBER, R. L., C. B. JOHANSSON, H. L. VAHLSING, A. R. HARGENS, AND E. R. FERINGA. 1986. Long-term effects of spinal cord transection on fast and slow rat skeletal muscle. I. Contractile properties. Exp. Neurol. 91: 423-434.
14. I&MO, T., R. H. WESTGAARD, AND L. ENGELBRETSEN. 1980. Different stimulation patterns at%% contractile properties of denervated rat soleus muscles. Pages 297-309 in D. PETrE, Ed., Plasticity of Muscle. de Gruyter, New York.
15. MAYER, R. F., R. E. BURKE, J. TOOP, B. WALLMSLEY, AND J. A. HODGSON. 1981. The effect of spinal cord transection on the motor units in cat medial gastrocnemius muscles. Muscle Nerve 23-3 1.
16. MAYER, R. F., R. E. BURKE, J. TOOP, J. A. HODGSON, K. KANDA, AND B. WALLMSLEY. 1981. The effect of long-term immobilization on the motor unit population of the cat medial hastrocnemius muscle. Neuroscience 6(4): 725-739.
17. NACHLAS, M. M., K-C. Tsou, E. DESOUZA, C-S. CHENG, AND A. M. SELIGMAN. 1957. Cytochemical demonstration of succinic dehydrogenase by the use of a new pnitrophenyl substituted ditetrazole. .I. Histochem. Cytochem. 420-436.
18. ROY, R. R., R. D. SACKS, K. M. BALDWIN, M. SHORT, AND V. R. EDGERTON. 1984. Inter- relationships of contraction time, V,, and myosin ATPase after spinal transection. J. Appl. Physiol.: Respir. Environ. Exercise Physiol. 56(6): 1594- 160 1.
19. SALMONS, S., AND F. A. STRETER. 1976. Significance of impulse activity in the transformation of skeletal muscle type. Nature 263: 30-34.
20. SIMARD, C. P., S. A. SPECTOR, AND V. R. EDGERTON. 1982. Contractile properties of rat hind limb muscles immobilized at different lengths. Exp. Neural. 77: 467-482.
21. SOKOL, R. R., AND F. J. ROHLF. 1969. Biometry. Freeman, San Francisco. 22. SPECTOR, S. A., C. P. SIMARD, M. FOURIER, E. STERNLICHT, AND V. R. EDGERTON. 1982.
Architectural alterations of rat hind limbs skeletal muscles immobilized at different lengths. Exp. Neural. 76: 94-l 10.
23. WEIBEL, E. R. 1980. Stereological Methods, Vol. 1, Practical Methods for Biological Mor- phometry. Academic Press, New York.


















