Lesion-Induced Alterations in Astrocyte Glutamate Transporter ...
17
Hindawi Publishing Corporation ISRN Neurology Volume 2013, Article ID 893605, 16 pages http://dx.doi.org/10.1155/2013/893605 Research Article Lesion-Induced Alterations in Astrocyte Glutamate Transporter Expression and Function in the Hippocampus Alexandra E. Schreiner, Eric Berlinger, Julia Langer, Karl W. Kafitz, and Christine R. Rose Institute of Neurobiology, Heinrich Heine University Duesseldorf, Universitaetsstraße 1, Building 26.02.00, 40225 Duesseldorf, Germany Correspondence should be addressed to Christine R. Rose; [email protected] Received 3 July 2013; Accepted 27 July 2013 Academic Editors: S. C. Barnett and J.-I. Satoh Copyright © 2013 Alexandra E. Schreiner et al. is is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. Astrocytes express the sodium-dependent glutamate transporters GLAST and GLT-1, which are critical to maintain low extracellular glutamate concentrations. Here, we analyzed changes in their expression and function following a mechanical lesion in the CA1 area of organotypic hippocampal slices. 6-7 days aſter lesion, a glial scar had formed along the injury site, containing strongly activated astrocytes with increased GFAP and S100 immunoreactivity, enlarged somata, and reduced capability for uptake of SR101. Astrocytes in the scar’s periphery were swollen as well, but showed only moderate upregulation of GFAP and S100 and efficiently took up SR101. In the scar, clusters of GLT-1 and GLAST immunoreactivity colocalized with GFAP-positive fibers. Apart from these, GLT-1 immunoreactivity declined with increasing distance from the scar, whereas GLAST expression appeared largely uniform. Sodium imaging in reactive astrocytes indicated that glutamate uptake was strongly reduced in the scar but maintained in the periphery. Our results thus show that moderately reactive astrocytes in the lesion periphery maintain overall glutamate transporter expression and function. Strongly reactive astrocytes in the scar, however, display clusters of GLAST and GLT-1 immunoreactivity together with reduced glutamate transport activity. is reduction might contribute to increased extracellular glutamate concentrations and promote excitotoxic cell damage at the lesion site. 1. Introduction Glutamate reuptake represents the principal mechanism for inactivation of synaptically released glutamate [1, 2]. In the rodent hippocampus, it is mainly accomplished by astrocytic glutamate transporters (EAATs: excitatory amino acid trans- porters), namely, GLAST (glutamate/aspartate transporter) and GLT-1 (glutamate-transporter-1; rodent analogues of EAAT1 and EAAT2, resp.; [3–7]). Glutamate uptake is ener- gized by the concomitant inward transport of three sodium ions and a proton, while one potassium ion is transported outward. Consequently, its activation is accompanied by an increase in the intracellular sodium concentration of astrocytes [8, 9]. Under pathological conditions, astrocytes undergo a complex reaction referred to as reactive astrogliosis, which is seen in diverse preparations and conditions ranging from primary cell culture to the intact brain [10, 11]. e hallmarks of reactive gliosis are a massive upregulation of the expression of the intermediate filament Glial Fibrillary Acidic Protein (GFAP) and a cellular hypertrophy [12, 13]. Reactive astro- cytes display several features of immature astrocytes (e.g., [14, 15]) and can partially reenter the cell cycle [16, 17]. e degree of astrogliosis can vary depending on the specific insult. Following traumatic brain injury, a dense glial scar forms at the lesion site due to strong astrocytic reorganization and proliferation [10, 11, 18]. At some distance to the lesion and scar, astrocytes usually do not divide, and their mor- phological reorganization and hypertrophy is less severe. e establishment of a glial scar around injured tissue is regarded as hindrance for the growth and regeneration of axons [19, 20]. Notwithstanding, reactive astrocytes might also exert a protective role and support regeneration [11, 13, 21]. ere is evidence that reactive gliosis is also accompanied by an alteration in the expression level of glial glutamate transporters [6]. Most studies performed in the rodent and
Transcript of Lesion-Induced Alterations in Astrocyte Glutamate Transporter ...

Hindawi Publishing CorporationISRN NeurologyVolume 2013 Article ID 893605 16 pageshttpdxdoiorg1011552013893605
Research ArticleLesion-Induced Alterations in Astrocyte Glutamate TransporterExpression and Function in the Hippocampus
Alexandra E Schreiner Eric Berlinger Julia Langer Karl W Kafitz and Christine R Rose
Institute of Neurobiology Heinrich Heine University Duesseldorf Universitaetsstraszlige 1 Building 26020040225 Duesseldorf Germany
Correspondence should be addressed to Christine R Rose rosehhude
Received 3 July 2013 Accepted 27 July 2013
Academic Editors S C Barnett and J-I Satoh
Copyright copy 2013 Alexandra E Schreiner et al This is an open access article distributed under the Creative Commons AttributionLicense which permits unrestricted use distribution and reproduction in any medium provided the original work is properlycited
Astrocytes express the sodium-dependent glutamate transportersGLAST andGLT-1 which are critical tomaintain low extracellularglutamate concentrations Here we analyzed changes in their expression and function following a mechanical lesion in the CA1area of organotypic hippocampal slices 6-7 days after lesion a glial scar had formed along the injury site containing stronglyactivated astrocytes with increased GFAP and S100120573 immunoreactivity enlarged somata and reduced capability for uptake ofSR101 Astrocytes in the scarrsquos periphery were swollen as well but showed only moderate upregulation of GFAP and S100120573 andefficiently took up SR101 In the scar clusters of GLT-1 and GLAST immunoreactivity colocalized with GFAP-positive fibers Apartfrom these GLT-1 immunoreactivity declined with increasing distance from the scar whereas GLAST expression appeared largelyuniform Sodium imaging in reactive astrocytes indicated that glutamate uptake was strongly reduced in the scar but maintainedin the periphery Our results thus show that moderately reactive astrocytes in the lesion periphery maintain overall glutamatetransporter expression and function Strongly reactive astrocytes in the scar however display clusters of GLAST and GLT-1immunoreactivity together with reduced glutamate transport activity This reduction might contribute to increased extracellularglutamate concentrations and promote excitotoxic cell damage at the lesion site
1 Introduction
Glutamate reuptake represents the principal mechanism forinactivation of synaptically released glutamate [1 2] In therodent hippocampus it is mainly accomplished by astrocyticglutamate transporters (EAATs excitatory amino acid trans-porters) namely GLAST (glutamateaspartate transporter)and GLT-1 (glutamate-transporter-1 rodent analogues ofEAAT1 and EAAT2 resp [3ndash7]) Glutamate uptake is ener-gized by the concomitant inward transport of three sodiumions and a proton while one potassium ion is transportedoutward Consequently its activation is accompanied byan increase in the intracellular sodium concentration ofastrocytes [8 9]
Under pathological conditions astrocytes undergo acomplex reaction referred to as reactive astrogliosis whichis seen in diverse preparations and conditions ranging fromprimary cell culture to the intact brain [10 11] The hallmarks
of reactive gliosis are amassive upregulation of the expressionof the intermediate filament Glial Fibrillary Acidic Protein(GFAP) and a cellular hypertrophy [12 13] Reactive astro-cytes display several features of immature astrocytes (eg[14 15]) and can partially reenter the cell cycle [16 17] Thedegree of astrogliosis can vary depending on the specificinsult Following traumatic brain injury a dense glial scarforms at the lesion site due to strong astrocytic reorganizationand proliferation [10 11 18] At some distance to the lesionand scar astrocytes usually do not divide and their mor-phological reorganization and hypertrophy is less severeTheestablishment of a glial scar around injured tissue is regardedas hindrance for the growth and regeneration of axons [1920] Notwithstanding reactive astrocytes might also exert aprotective role and support regeneration [11 13 21]
There is evidence that reactive gliosis is also accompaniedby an alteration in the expression level of glial glutamatetransporters [6] Most studies performed in the rodent and
2 ISRN Neurology
human brain reported an overall downregulation of proteinlevels of both GLAST and GLT-1 andor a reduction infunctional glutamate uptake in response to brain injury andastrogliosis respectively (eg [22ndash27]) The reduction in theexpression of glial glutamate transporters has been suggestedto contribute to the elevation of extracellular glutamate con-centrations and to glutamate-mediated excitotoxicity whichis observed under many pathological conditions
Injury-induced changes in the expression of the gluta-mate transporter subtypes might however also depend onthe degree of astrogliosis This might be especially relevantand visible upon a mechanical lesion where reactivity ofastrocytes as judged for example based on their GFAPexpression and morphology ranges from severe astroglio-sis in the scar region to only moderate-mild astrogliosisdistant from the lesion [10 11] To study this questionwe analyzed expression of GLAST and GLT-1 following amechanical lesion in organotypic tissue slice cultures of themouse hippocampus Furthermore we employed ratiometricsodium imaging as functional assay for glutamate uptake inastrocytes Our results show that mechanical tissue injurygenerates subsets of reactive astrocytes depending on thedistance from the lesion site which differ in bothmorpholog-ical features and their ability to take up SR101 Furthermorethese subsets show discrete changes in glutamate transporterexpression and glutamate uptake capacity indicating that glu-tamate clearance might be largely functional in the peripheryof the lesion but strongly hampered in the scar region
2 Materials and Methods
21 Ethics Statement This study was carried out in strictaccordance with the institutional guidelines of the Hein-rich Heine University Duesseldorf Germany as well as theEuropean Community Council Directive (86609EEC) Allexperiments were communicated to and approved by theAnimal Welfare Office at the Animal Care and Use Facilityof the Heinrich Heine University Duesseldorf Germany(institutional act number O5205) In accordance with theGerman Animal Welfare Act (Tierschutzgesetz Articles 4and 7) no formal additional approval for the postmortemremoval of brain tissue was necessary For generation ofacute slices mice were quickly decapitated (following therecommendation of the European Commission published inEuthanasia of experimental animals Luxembourg Office forOfficial Publications of the European Communities 1997ISBN 92ndash827-9694-9)
22 Preparation of Tissue Slice Cultures Acute tissue slicesof the hippocampus were prepared from Balbc mice (Musmusculus) of both genders at postnatal days 7 to 8 (P7-8)using standard procedures For some experiments transgenicanimals (FVBN-Tg[GFAPGFP]14MesJ) expressing greenfluorescence protein (GFP) under the GFAP promoter wereused (obtained from Jackson Laboratory Harbor USA)After decapitation of the animals brains were quickly excisedand hemisected in ice-cold artificial cerebrospinal fluid(ACSF) composed of (in mM) 125 NaCl 25 KCl 2 CaCl
2
1 MgCl2 125 NaH
2PO4 26 NaHCO
3 and 20 glucose bub-
bled with 95 O2and 5 CO
2 and adjusted to a pH of 74
Hemisections were trimmed and transverse slices (200120583m)comprising the entorhinal cortex hippocampus fimbriaand thalamus were prepared using a vibratome (MicromHM650V Thermo Fischer Scientific Walldorf Germany)Slices were transferred to ACSF at 35∘C for 30 minutes
Organotypic hippocampal slice cultures were preparedand cultured according to the protocol introduced by Stop-pini et al [28] with minor modifications To this end acuteslices were transferred to a Millicell culture insert (PICMORG 50 hydrophilized PTFE pore size 04 120583m Merck Mil-lipore Darmstadt Germany) and maintained at the interfaceof a serum-based culture medium free of antibiotics in ahumidified incubator atmosphere of 5 CO
2at 37∘C The
culture medium was composed of 30 sterile filtered normalhorse serum (NHS GIBCOLife Technologies DarmstadtGermany) 30Dulbeccorsquos modified eagle medium (DMEMGIBCOLife Technologies Darmstadt Germany) and 40Hankrsquos balanced salt solution (HBSS GIBCOLife Tech-nologies Darmstadt Germany) supplemented with 38mMglucose (pH adjusted to 73-74) The medium was changedthree times a week and the insert surface was washed withmedium once a week
After at least 12 days in culture a mechanical lesionwas performed using a sterile scalpel blade The lesion waspositioned in the CA1 area perpendicular to the stratumpyramidale included the strata oriens pyramidale radiatumand lacunosum moleculare and spanned the entire depthof the slice (cf Figure 2) After lesioning slice cultureswere maintained for another 6-7 days (designated as 6-7days postlesion) Unlesioned control slices were cultured inparallel for a corresponding number of days
For visualization of cell death ACSF containing05 120583gmL propidium iodide (PI) was applied to the slicesurface and incubated for 3 hours at 37∘C and 5 CO
2
followed by a wash with ACSF Documentation was eitheraccomplished with an epifluorescence microscope (NikonEclipse 90i Nikon Instruments Dusseldorf Germany) orat an Olympus Fluoview300 laser scanning microscope(Olympus Hamburg Germany)
Astrocyte soma size in organotypic control and lesionedslices was calculated from images of GFP-fluorescencederived fromGFPGFAPmice which were obtained at a con-focal microscope (Olympus Fluoview300 Olympus Ham-burg Germany see also below) To this end a semiquantita-tive approach was used in which somata of single cells weremanually encircled (cf Figure 4(b)) and the resulting areawas calculated using ImageJ software (NIH Bethesda USA)
All chemicals were purchased from Sigma-AldrichChemical (Munich Germany) unless stated otherwise
23 Imaging Experiments For imaging experiments organ-otypic slices were excised from the Millicell inserts andincubated in ACSF containing 25120583M SR101 for 30minat 35∘C to stain astrocytes (see above) SR101 is a highlyspecific and widely used tool for the identification of matureastrocytes in many brain regions including the hippocampus
ISRN Neurology 3
[29 30] It has recently been shown to be taken up intomaturehippocampal astrocytes via an active transport mechanisminvolving organic anion transport polypeptides [31] Wide-field fluorescence imaging was performed using a variablescan digital imaging system (TILL Photonics MartinsriedGermany) attached to an upright microscope (BX51WiOlympus Europe Hamburg Germany) and a CCD camera(TILL Imago VGA Till Photonics Martinsried Germany)Imageswere collectedwith anAchroplan 40x objective (waterimmersion NA 08 Zeiss Gottingen Germany) SR101 wasexcited at 575 nm and emission was collected above 590 nmExcitation wavelength for detection of GFP was 488 nm andemission was collected above 510 nm
For intracellular sodium imaging slices were addition-ally incubated with the membrane permeable form of thesodium-sensitive fluorescent dye SBFI (SBFI-AM sodium-binding benzofuran isophthalate-acetoxymethyl ester Mol-ecular ProbesLife Technologies Darmstadt Germany) asdescribed earlier [32ndash34] Ratiometric sodium imaging wasperformed by alternate excitation of SBFI at 340 nm (weaklysodium-sensitive wavelength) and at 380 nm (sodium-sensitive wavelength) at 4Hz Emission (gt440 nm) wascollected in defined regions of interest (ROI) representingcell bodies Standard dynamic background correction wasperformed as described earlier [33 34] After backgroundcorrection the fluorescence ratio (F
340F380
) was calculatedfor the individual ROIs and analyzed offline using OriginPro8G Software (OriginLab Corporation Northampton MAUSA)
Changes in SBFI fluorescence ratio were expressed aschanges in sodium concentration based on in situ calibrationsas reported before [32 33 35] To equilibrate extra- andintracellular Na+-concentrations SBFI-loaded slices wereperfused with saline containing ionophores (3 120583M grami-cidin D 100 120583Mmonensin) and the Na+K+-ATPase blockerouabain (100 120583M) as well as different concentrations of Na+
The glutamate transporter agonist D-aspartate wasapplied by a pressure application device (PDES-02D NPIElectronic GmbH Tamm Germany) coupled to standardmicropipettes (Hilgenberg Waldkappel Germany) placed20ndash100120583m from cell bodies of selected cells
24 Antibodies Antibodies employed in this study are listedin Table 1 The following antisera were utilized whichrepresent well-established commercially available standardmarkers guinea-pig GLT-1 antiserum directed against theC-terminus of rat GLT-1 (Chemicon InternationalMilliporeCorp Carrigtwohill Ireland eg [36]) and guinea-pigGLAST antiserum directed against the C-terminus of ratGLAST (Chemicon InternationalMillipore Corp Carrigt-wohill Ireland eg [37]) Validation and a detailed charac-terization of both antibodies were provided before [38ndash40]For identification of astrocytes polyclonal rabbit antibodiesagainst GFAP (GFAP-pAb Dako Cytomation Denmark[40 41]) and S100120573 (Abcam United Kingdom [42]) wereemployedWhen double staining with the latter antibody wasperformed a monoclonal mouse antibody detecting GFAPwas used (GFAP-mAb Sigma Aldrich Germany [40 43])
Standard fluorochrome-conjugated antibodies (Alexa-Fluor InvitrogenLife Technologies Darmstadt Germany)were employed as secondary antibodies for immunohisto-chemistry
25 Immunohistochemistry Prior to immunohistochemicalprocessing organotypic slices were immersion fixed for30min at RT in 4 paraformaldehyde (PFA) in phosphate-buffered saline (PBS) following three washes every30min with PBS Cell membranes were permeabilizedand unspecific binding sites were blocked in PBS containing025 triton-X100 (TX) and 2 normal goat serum (NGSGIBCOLife Technologies Darmstadt Germany) for 90minat 4∘C followed by incubation with the primary antibodyGFAP-pAb (1 1000 Dako Cytomation Glostrup Denmark)diluted in the same solution over night at 4∘C In case ofS100120573GFAP double stainings slices were incubated witha mixture of the primary antibodies GFAP-mAb (1 1000Dako Cytomation Glostrup Denmark) and rabbit-S100120573(1 100 Abcam United Kingdom) After five washes in PBScontaining 025 TX and 2 NGS slices were incubatedeither with guinea pig-anti-GLAST or with guinea pig-anti-GLT-1 (both 1 1000 diluted in 025 TX2 NGSPBS) for4 hrs at RT Excess primary antibody was removed with fivewashes using 2 NGSPBS Antirabbit-AlexaFluor594 andantiguinea pig-AlexaFluor488 or antimouse-AlexaFluor488(1 100 in blocking solution) were used for visualizationof antibody binding and incubated for 2 hrs at RT Theslices were subjected to DAPI staining (410158406-diamidino-2-phenylindole 05120583m Invitrogen) washed three times andmounted on glass slides with mowiolDABCO (CalbiochemFluka distributed by Sigma-Aldrich Chemical MunichGermany)
Identical conditions were applied to all performed stain-ings regarding tissue processing and staining procedureNegative controls were run in parallel to each staining byeither omitting all or just one of the primary antibodiesControl stainings in which one of the primary antibodieswas omitted showed the identical labeling pattern for theremaining antibody as in the double stainings Omitting bothprimary antibodies never resulted in a staining
Documentation of immunofluorescence was either per-formed with an epifluorescence microscope (Nikon Eclipse90i Nikon Instruments Dusseldorf Germany) or a confocallaser scanningmicroscope (Olympus Fluoview300OlympusHamburg Germany) The epifluorescence microscope wasequipped with a standard DAPI (EX 340ndash380 DM 400BA 435ndash485) FITC (EX 465ndash495 DM 505 BA 515ndash555)and TRITC (EX 54025 DM 365 BA 60555) filter setIllumination was provided by an Intensilight fiber lamp(C-GHFI Nikon Instruments Dusseldorf Germany) andemission was detected with a monochrome digital cam-era (DS-Qi1Mc Nikon Instruments Dusseldorf Germany)Images were collected with either a 20x075 (PlanApoVCNikon Instruments Dusseldorf Germany) air objective ora 60x140 oil immersion objective (PlanApoVC NikonInstruments Dusseldorf Germany) NIS-Elements software(Nikon Instruments Dusseldorf Germany) was used forimage acquisition For confocal microscopy an Olympus
4 ISRN Neurology
Table 1 List of antibodies used
Antibody Structure of theimmunogen Company Order number Species Type Dilution
GLAST(EAAT1)
KPYQLIAQDNEPEKPVADSETKM ChemiconMillipore AB1782lowast Guinea
Pig IgGPolyclonal 1 1000
GLT-1(EAAT2)
AANGKSADCSVEEEPWKREK ChemiconMillipore AB1783 Guinea
Pig IgGPolyclonal 1 10001 2000
GFAP ns (full-length proteinfrom cow spinal cord) Dako Cytomation Z 0334 Rabbit
IgGPolyclonal 1 10001 2000
GFAP
clone G-A-5(LQSLTCDVESLRGTNESLERQMREQEERHAREAASYQEALTRLEEEGQSLKDEMARHLQEYQELLNVKLALDIEIATY)
Sigma-Aldrich G3893 MouseIgG
Monoclonal 1 10001 2000
S100szlig ns (recombinantfull-length cow protein) Abcam ab868 Rabbit
IgGPolyclonal 1 100
MouseIgG AF488 conjugated Invitrogen (Life Technologies) A11029 Goat mdash 1 100
RabbitIgG AF594 conjugated Invitrogen (Life Technologies) A11012 Goat mdash 1 100
Guineapig IgG AF488 conjugated Invitrogen (Life Technologies) A11073 Goat mdash 1 100
GLAST glutamate aspartate transporter GLT-1 glutamate transporter 1 GFAP glial fibrillary protein AF AlexaFluor appl application IHC immunohisto-chemistry WB western blot ns not specifiedlowastThemanufacturer has lately stopped the sale and distribution of the GLAST antibody used here
BX51WI microscope coupled to a confocal laser scanningsystem (FV300) equipped with a multiline argon (488 nm)and a helium-neon laser (543 nm both Melles Griot Ben-sheim Germany) was used Images were collected witheither a 20x050 (UMPlanFl Olympus Hamburg Ger-many) a 40x080 water immersion (LUMPlan OlympusHamburg Germany) or a 60x140 oil immersion objective(PlanApoVC Nikon Instruments Dusseldorf Germany) AKalman filter 4 was employed at every scan Simultaneousor sequential scanning of both fluorophores revealed nodifference in their staining patterns indicating absence ofcross-excitation or spectral bleed through The thickness ofz-plane sections was 1120583m and the number of optical sectionsvaried depending on the preparation
Figures show extended focus images as specified inthe figure legends which were calculated from z-stacksof optical sections using ImageJ software (NIH BethesdaUSA) Images were overlaid employing Adobe PhotoshopCS2 (Adobe Systems Cologne Germany)
26 Data Presentation and Statistics Unless otherwise spec-ified data are expressed as means plusmn SEM Data were statis-tically analyzed by Studentrsquos 119905-test employing the proceduresimplemented in OriginPro 8G Software (OriginLab Corpo-ration Northampton MA USA) A 119875 value of lt005 wasconsidered significant If not stated otherwise 119899representsthe number of analyzed cells and119873 the number of slices Eachset of experimentswas performedon at least three tissue slicesobtained from different animals
3 Results
31 Identification of Astrocytes in Organotypic Slice CulturesThe aim of the present study was to analyze changes inglutamate transporter expression and function in astrocytesin response to a mechanical injury To this end we preparedorganotypic tissue slice cultures of themouse hippocampus awell-establishedmodel system inwhich the layering and basiccellular organization of the neural network are maintainedover time [28] After 19ndash25 days in culture organotypic tissueslices had flattened to a thickness of 40ndash50 120583m and werecomposed of 3 to 4 cell layers At this stage propidium iodideassays detected only few dead cells dispersed throughoutthe slices (3 plusmn 06 cells per CA1 subfield 18 plusmn 34 in entirepreparation 119873 = 6 data not shown) indicating that thepreparation was viable and in a stable condition
To visualize astrocytes in cultured slices we used hip-pocampi of transgenic mice in which green fluorescentprotein (GFP) is expressed under the control of the promoterof the astrocyte-specific intermediate filament glial fibrillaryacidic protein (GFAP FVBN-Tg[GFAPGFP] ldquoGFPGFAPmicerdquo) A visualization of theGFP fluorescence demonstratedthat the general distribution and organization of astrocyteswere preserved in the strata pyramidale and radiatum ofthe CA1 region of organotypic cultures confirming earlierreports (119873 = 3 Figure 1(a) [44 45]) The vast majority ofGFP-expressing cells (sim95) also labeled with the astrocyte-specific vital marker SR101 in organotypic slices (119873 = 3Figures 1(a) and 1(b)) Furthermore immunohistochemicalstainings for GFAP in slice cultures derived from wildtype
ISRN Neurology 5
Slic
e cul
ture
(20
dic
)
GFP-GFAP MergeSR101
lowast lowast lowast lowast
lowast
lowast
lowast lowast
StrP
StrR
(a)
StrR
StrP
StrLM
StrM
SR101
StrO
(b)
GFAP S100120573
StrP
StrR
StrP
StrR
StrP
StrR
StrP
StrR
DAPI Merge
(c)
Figure 1 Astrocytes in organotypic hippocampal slice cultures (a) Organotypic slice cultures at 20 days in culture (dic) Preparations werederived from animals in which GFP was expressed under control of the GFAP promoter Shown is the expression of GFP (left) the labelingpattern of SR101 (center) and the corresponding merged image (GFAP green SR101 magenta) Asterisks point out SR101-labeled cells thatdo not express GFP arrowheads point out GFP-positive but SR101-negative cells Images represent maximum intensity projections of 15optical sections taken at 1120583m intervals (b) SR101 fluorescence of an organotypic slice at 21 dic Image represents a stack of 20 optical sectionstaken at 1 120583m intervals (c) Immunohistochemical double staining for GFAP and S100120573 as well as DAPI fluorescence and merged image ofthe fluorescence of GFAP (green) and S100120573 (magenta) of part of the CA1 region encompassing the strata pyramidale and radiatum of anorganotypic slice at 20 dic Note the radially oriented GFAP-positive fibers crossing the pyramidal cell layer Images were taken at a wide-field microscope ((a)ndash(c)) Dashed lines indicate the approximate boundaries of the stratum pyramidale StrO stratum oriens StrP stratumpyramidale StrR stratum radiatum StrLM stratum lacunosum moleculare and StrM stratum moleculare Scale bars (a) 50 120583m (b) 100120583mand (c) 50 120583m
animals showed that themajority of GFAP-positive cells werealso positive for S100120573 (119873 = 12 Figure 1(c)) a marker formature astrocytes [46]
Taken together the nearly complete overlap in the cellularstaining pattern for the vital dye SR101 with the expressionof GFP in GFPGFAP mice as well as the overlap in theimmunofluorescence for GFAP and S100120573 in organotypicslices indicates that these markers are well suited to identifyastrocytes in this preparation
32 Astrocyte Markers and Astrocyte Morphology followingMechanical Lesion To induce astrogliosis in organotypicslice cultures we used a scalpel blade and performed
a scratch under semisterile conditions through the entireCA1 region oriented perpendicular to the stratum pyramidale(Figure 2(a)) 6-7 days after the lesion a general enhance-ment of GFAP immunoreactivity was observed indicatingastrogliosis (Figure 2(b))The greatest enhancement of GFAPimmunoreactivity occurred within a distance of less than100 120583malong the lesion (119873 = 26 Figures 2(b) and 2(c)) Herea densemeshwork of thick GFAP-positive labels was presentindicative of the formation of a glial scar Furthermore theregion close to the lesion site exhibited a discernible increasein S100120573 immunoreactivity (119873 = 12 Figure 2(c)) Both theprominent expressions of GFAP and S100120573 are characteristicsof strongly reactive astrocytes which represent the major
6 ISRN Neurology
GFAP
DAPI
CA1 subregion
GFAP
DAPI
StrR
StrP StrP
StrR
StrOStrO
StrO
StrP
StrR
StrO
StrP
StrR
Control Lesioned
GFAP
Lesio
n sit
e
Hi
StrMStrG
StrP
StrR
StrLM
Scratch wound
StrO
Lesio
n sit
e
Lesioned
(a) (c)
(b)
350120583m350120583m
S100120573
Figure 2 Lesion-induced reactive gliosis in organotypic slice cultures (a) Schematic overview of the hippocampus indicating the positionof the scratch woundThe box delineates the area primarily analyzed (b) GFAP immunofluorescence and DAPI staining of the CA1 area in acontrol (left) and a lesioned (right) slice The boxes indicate the areas enlarged below The dashed line in the lower right indicates the lesion(c) Double staining for GFAP (top) and S100120573 (bottom) adjacent to the lesion site Note the gradual increase in GFAP and S100120573 expressiontowards the lesion (b) widefield microscopy (c) confocal microscopy (optical sections taken at 1120583m intervals) StrO stratum oriens StrPstratum pyramidale StrR stratum radiatum and StrLM stratum lacunosum moleculare Scale bars (b) 200 120583m (top) and 50 120583m (bottom) (c)10120583m
ISRN Neurology 7
StrR
StrP
StrLM
StrO
StrP StrRStrO StrLM
Lesio
n sit
e
SR101-staining pattern
(a)
SBFI
Lesio
n sit
e
SR101 GFAP-GFP
(b)
Figure 3 SR101 labeling in lesioned slices (a) SR101 fluorescence in the CA1 area in a slice 7 days after lesion The delineated area comprisesthe stratum pyramidale and the box indicates the area enlarged on the right side Right SR101 fluorescence at the lesion site illustrating thatSR101 labeling is reduced along the lesionThe dotted line indicates the transition between the scar area and the adjacent moderately reactivetissue The boxed area is further enlarged at the bottom (b) Image series obtained at a wide-field fluorescence microscope The slice wasprepared from a transgenic GFPGFAP mouse and the lesion site is indicated by the dotted line Left Image of SR101 fluorescence centerGFP fluorescence indicating GFAP expression and right SBFI fluorescence SR101 labeling is only weak in GFPSBFI-positive cells in the scarregion (see arrowheads) StrO stratum oriens StrP stratum pyramidale StrR stratum radiatum and StrLM stratum lacunosum-moleculareScale bars 100120583m ((a) upper left) and 20 120583m (other images)
cellular component of the glial scar forming after mechanicaltissue injury [10] More distal to the lesion (gt100ndash350 120583m) aless dramatic rearrangement of GFAP immunoreactivity wasobserved
A clear difference in astrocyte properties between thescar region and its periphery was also seen following SR101labeling In the periphery of the scar (100ndash350 120583m distancefrom the lesion) SR101 resulted in a reliable and brightcellular staining similar to what had been observed incontrol unlesioned slices (119873 = 12 Figure 3(a) cf Figure 1)
In contrast to this cellular SR101 staining was detectable butonly faint within a distance of less than 100 120583m on both sidesof the lesion (Figure 3(a) meanwidth of SR101-free area 111plusmn3 120583m 119873 = 5) In slices from GFPGFAP transgenic animals(119873 = 5 Figure 3(b)) GFAP-positive weakly SR101-positivescar cells also stained with the sodium-sensitive fluorescenceindicator SBFI-AM (acetoxymethyl ester of sodium bindingbenzofuran isophthalate see below 119873 = 3) indicatingthat they were able to take up and de-esterify this dye(Figure 3(b))
8 ISRN Neurology
lowastlowastlowast
lowastlowastlowast
80
lowast
Control ScarPeriphery
120
160
0
40
200
Som
a siz
e (120583
m2)
(a)
Lesioned
StrP
StrR
StrP
Control
StrR
StrLM
Lesio
n sit
e
(b)
Figure 4 Lesion-induced changes in somatic area (a) Quantification of somatic area based on GFAPGFP fluorescence in control andlesioned slices The soma size increased both in astrocytes in the scar region (ldquoscarrdquo) and in astrocytes in the periphery of the lesion(ldquoperipheryrdquo lowastlowastlowast119875 lt 0001 lowast119875 lt 005) (b) Confocal images of GFP fluorescence in cultured slices obtained from GFPGFAP transgenicanimals under control conditions (left) and subjected to a lesion (right) Cells surrounded by boxes are shown at higher magnification in theinsets and the dashed lines around the soma illustrate the areas which served to determine their size StrP stratum pyramidale StrR stratumradiatum StrLM stratum lacunosum moleculare and scale bars 40 120583m and 10120583m (inset)
In addition to GFAP-positive cells the scar region hostedmicroglial cells identified by their vital staining with TexasRed-coupled lectin from lycopersicon esculentum (tomatolectin119873 = 4 not shown) Tomato lectin-positive microgliahowever did not take up SBFI-AM (119873 = 4 not shown) con-firming earlier reports thatmicroglial cells do not incorporateAM-ester dyes in situ [47]
To further characterize reactive gliosis in response to thescratch wound we determined the area of astrocyte somatain confocal images of the GFP fluorescence in control andlesioned slices obtained fromGFPGFAP transgenic animalsThesemeasurements revealed that compared to astrocytes in
control slices the soma area of astrocytes in the periphery ofa lesion was increased by 18 and that of astrocytes in thescar regionwas increased by 24 (control 163plusmn2 120583m2 lesionperiphery 193plusmn3 120583m2 scar tissue 201plusmn4 120583m2119875 lt 0001 119899 =945 496 and 439 in 7 control and 9 lesioned slices Figures4(a) and 4(b)) Thus astrocytes show a second hallmark ofreactive gliosis after lesion namely a hypertrophy of their cellbodies which is observed both in the scar tissue as well as inthe periphery of the lesion
Taken together these data show that 6-7 days after themechanical lesion a glial scar has formed along the lesionsite This scar encompasses strongly activated astrocytes
ISRN Neurology 9
characterized by a robust increase in GFAP and S100120573expressionThese proximal reactive astrocytes locatedwithina distance of less than 100120583m from the lesion exhibit longGFAP-positive processes have significantly swollen somataand only weakly stain with SR101 Astrocytes in the peripheryof the scar (100ndash350 120583m from the lesion) display swollen cellbodies as well but show onlymoderate upregulation of GFAPand maintain their ability to efficiently take up SR101
33 Expression Levels and Spatial Distribution of GLT-1 andGLAST To determine the spatial distribution of the gluta-mate transporters GLAST and GLT-1 in organotypic slicesand changes therein in response to the lesion immunohisto-chemistry was employed In control slices GLAST (119873 = 13Figure 5(a)) as well as GLT-1 immunoreactivity (119873 = 11Figure 5(b)) appeared as punctate labeling throughout theentire CA1 area The staining patterns for both transportersin the tissue slice cultures were thus similar to those reportedfrom acute tissue slice preparations of the hippocampus [3648 49]
The relatively homogeneous distribution of immunore-activity throughout the preparation found in control slicecultures was abandoned following a lesion Here distinctaccumulations of GLAST (119873 = 17 Figure 5(a)) and GLT-1immunoreactivity (119873 = 12 Figure 5(b)) were found in thescar region In both cases these preferentially colocalizedwith thick GFAP-positive bundles running in parallel to thelesion (Figure 5) Apart from the marked accumulation ofGLAST reactivity in parallel to the scar GLAST immunore-activity appeared generally increased in some lesioned slicepreparations (Figure 5(a)) but unaltered in others bothwithinthe scar as well as in its periphery For GLT-1 a decline in theintensity of immunoreactivity was observed with increasingdistance from the lesion (Figure 5(b))
In summary these results reveal distinct alterations inthe immunoreactivity of GLAST and GLT-1 following amechanical lesion In the scar region clusters of GLT-1 andGLAST immunoreactivity are present primarily colocalizedwith strongly GFAP-positive fibers Apart from these distinctclusters GLT-1 immunoreactivity appears to decline withdistance from the scar whereas GLAST immunoreactivityseems largely uniform throughout the preparation
34 Sodium Transients Induced by Activation of GlutamateUptake Glutamate uptake is accompanied by the inwardflux of sodium resulting in increase in the intracellularsodium concentration of astrocytes To monitor its functionwe assayed sodium increases induced by D-aspartate anonmetabolized substrate of sodium-dependent glutamateuptake which is also transported [1] in astrocytes in thestratum radiatum of organotypic slices To this end SR101-stained slices were additionally loaded with the sodium-sensitive fluorophore SBFI-AM (Figure 6(a) cf Figure 3(b))In situ calibrations of the SBFI fluorescence revealed a linearincrease in its fluorescence ratio at intracellular sodiumconcentrations between 10 and 40mM (119899 = 107 Figure 6(b))as reported before [33]
Repetitive pressure application of D-aspartate for 500msinduced reliable increases in the intracellular sodium con-centration in astrocytes indicating activation of sodium-dependent glutamate uptake (119899 = 488 Figure 6(c)) Theexperiments were performed in the presence of the sodium-channel blocker TTX (tetrodotoxin 05 120583M) to suppressaction potential generation The amplitude of D-aspartate-induced sodium increases was dependent on the concen-tration applied (Figure 6(c)) and saturated at about 2mM(119899 = 19 Figure 6(d)) as reported before for mouse corticalastrocytes in culture [50] At a concentration of 1mM theamplitude of D-aspartate-induced sodium transients wasnear maximum and amounted to 38 plusmn 02mM (119899 =161 Figures 6(c) and 6(d)) a value comparable to that inastrocytes in acute slices obtained in our lab [33]
To probe for changes in glutamate uptake we comparedsodium signals induced by 1mMD-aspartate between controland lesioned slices (Figure 7) In lesioned slices astrocyteslocated in the periphery of the lesion (100ndash350 120583m distance)responded with a sodium transient to application of 1mMD-aspartate (see cell 2 in Figures 7(a) and 7(b)) The amplitudeof D-aspartate-induced sodium signals in cells in the lesionperiphery was similar to that observed in control slices (38 plusmn03mM 119899 = 111 see cell 1 in Figures 7(a) 7(b) and 7(c))Application of D-aspartate also evoked sodium transientsin weakly SR101-positive SBFI-filled astrocytes located inthe glial scar area (lt100 120583m from the lesion see cells 3ndash5in Figures 7(a) and 7(b)) These cells however displayedsignificantly smaller peak amplitudes (16 plusmn 021mM 119899 =26 119875 lt 0001 Figures 7(b) and 7(c)) than astrocytes inthe periphery and in control slices
The kinetics of D-aspartate-induced sodium transientsdiffered between all three groups This was true for theslope of the sodium increase (10ndash90 change) which wassignificantly reduced in astrocytes from lesioned slices versusastrocytes from control slices (09 plusmn 005mMs in controlversus 06 plusmn 006mMs in the periphery and 02 plusmn 004mMsin the scar region 119899 = 161 111 and 26 resp 119875 lt 0001Figures 7(b) and 7(c)) In addition the decay back to baselinewas significantly slower following lesion (Figures 7(b) and7(c)) The time needed for the signal to decay by 67 wasapproximately doubled (decay time 120591 was 44 plusmn 2 s in control120591 = 81 plusmn 4 s in periphery of the lesion and 120591 = 95 plusmn 10 s inscar cells Figures 7(b) and 7(c))
These results show that mechanical injury results indistinct alterations in the kinetics andor amplitudes ofD-aspartate-induced intracellular sodium transients in reac-tive astrocytes depending on their distance to the lesion
4 Discussion
Our results show that within 6-7 days after setting a scratchwound in organotypic hippocampal slices a dense glial scarhad formed along the lesion site Up to approximately 100 120583mdistance from the lesion the scar tissue was characterized bya substantial elevation ofGFAP expression a pronounced cel-lular hypertrophy and reduced uptake of SR101 by astrocytes
10 ISRN Neurology
Con
trol
GLAST
Lesio
ned
GFAP Merge
StrR
StrP
StrO
Lesio
n sit
e
StrR
(a)
Con
trol
Lesio
ned
GLT-1 GFAP Merge
StrR
StrP
StrO
StrR
StrP
StrLM
Lesio
n sit
e
(b)
Figure 5 GLAST andGLT-1 immunoreactivity aftermechanical injury (a) Fluorescence images of immunohistochemical stainings detectingthe glutamate transporters GLAST (left) GFAP (center) as counterstaining and the corresponding merged image (GFAP green GLASTmagenta) in control (top row) and lesioned slice cultures (bottom row) (b) GLT-1 (left) GFAP (center) double labeling of a control (toprow) and a mechanically lesioned slice (bottom row) The corresponding merge (GFAP green GLT-1 magenta) is found on the right Notethat after lesion glutamate transporter immunoreactive clusters are accumulated in the direct vicinity of the lesion site (arrowheads) GFAPglial fibrillary acidic protein GLAST glutamate aspartate transporter GLT-1 glutamate-transporter-1 StrO stratum oriens StrP stratumpyramidale StrR stratum radiatum StrLM stratum lacunosum-moleculare and scale bars (a) 20 120583m (b) 40 120583m
While prominent clusters of GLAST and GLT-1 immunore-activity were observed alongGFAP-positive structures in scarcells D-aspartate-induced intracellular sodium signals werestrongly dampened indicating a significant reduction in glu-tamate uptake capacity close to the lesion In the periphery ofthe lesion (100ndash350120583m) astrocytes showed less pronounced
reactivity maintained their ability to efficiently take up SR101and displayed only minor changes in glutamate transporterimmunoreactivity and function Thus depending on theirdistance from the lesion astrocytes showed different gradesof reactivity and displayed discrete changes in glutamatetransporter immunoreactivity and uptake capacity
ISRN Neurology 11
SR101 MergeSBFI
StrRStrP
(a)
03
04
05
06
07
0 5 10 15
0
20 25 30 40 0
100
20
40
80
60
0 10 20 30 400
20
40
60
80
100
10 = 55 mM
107 cells in 6 experiments
F (340
380
) (ra
tio)
Intracellular Na+ concentration (mM)Intracellular Na+ concentration (mM)
ΔF
(340
380
) (
)
ΔF
(340
380
) (
) R2 = 097
(b)
D-aspartate
100 120583M500 120583M
750 120583M1000 120583M
1 mM
10 s
Δ
(c)
0 1 2 3 4 50
1
2
3
4
5
Δ so
dium
(mM
)
D-aspartate (mM)
lowast
(d)
Figure 6 Sodium changes induced by activation of glutamate uptake (a) Staining of astrocytes by colabeling with SR101 and SBFI Leftepifluorescence image of theCA1-area of an SR101-loaded organotypic sliceCenter SBFI fluorescence taken from the same areaRight overlaySBFI-loaded SR101-positive astrocytes appear in white (arrowheads) StrP stratum pyramidale StrR stratum radiatum scale bar 20 120583m (b)Left calibration of SBFI fluorescence in an astrocyte Depicted is the change in intracellular SBFI fluorescence ratio in the calibration solutionscontaining ionophores and in response to stepwise changes in the sodium concentration Right mean values plusmn SEM of the normalizedchanges in SBFI fluorescence at different sodium concentrations The black line represents a linear fit of the data points between 10 and40mM sodium illustrating that a 10 change in SBFI fluorescence represents a change in the sodium concentration by 55mM (c) Sodiumtransients in a single astrocyte induced by 500ms applications of D-aspartate at different concentrations (d) Mean values plusmn S E M (119899 = 19)of the peak amplitude of sodium transients at different concentrations of D-aspartate The curve represents a sigmoidal fit of the data
41 Characteristics of Lesion-Induced Reactive Gliosis inOrganotypic Cultures Organotypic tissue slice cultures ofthe hippocampus represent a well-established model systemin which the basic tissue architecture is maintained butwhich still hosts the major advantages of cell cultures suchas good accessibility and control of experimental conditionsThey have been used extensively for the analysis of neuronal
properties and developing neuronal networks [28 51 52]or the study of excitotoxic neuronal damage (eg [53 54])Recent studies also demonstrated that basic morphologicalcharacteristics of astrocytes and the typical glia-synapseorganization are well preserved in organotypic slice cultures(eg [44 45 55 56]) In addition we found a nearly completeoverlap of SR101 staining with the expression of GFP in slice
12 ISRN Neurology
SBFIControl Lesioned
SR101
1
Lesio
n sit
e
SR101 SBFI
2 45
32 4
5
1
StrR
StrP StrP
StrR
3
(a)
ControlScar
Lesioned
1
D-asp
2
D-asp
D-asp
5
4
3Periphery1mM
20 s
Δ Δ
Δ
Δ
Δ
(b)
0102030405060708090
100110
Δ so
dium
tim
e (m
Ms
)
00
02
04
06
08
10
12
0
1
2
3
4
5
6
ns
Control Periphery Scar Control Periphery Scar Control Periphery Scar
Amplitude Slope Decay time
Δ so
dium
(mM
)
120591(s
)lowastlowastlowast
lowastlowastlowastlowastlowastlowast
lowastlowastlowastlowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowast
(c)
Figure 7 D-aspartate-induced astrocyte sodium transients following lesion (a) CA1 area of a control (left) and a lesioned (right) slicepreparation Shown are epifluorescence images of SR101 and SBFI fluorescences Circled areas and numbers indicate the regions of interestanalyzed in the experiment depicted in (b) StrP stratum pyramidale StrR stratum radiatum and scale bar 40 120583m (b) Sodium transientsinduced by pressure application of 1mMD-aspartate (D-asp) for 500ms in the different cells in the control and lesioned slice as indicated in(a) (c) Histograms showing mean values plusmn S E M of the peak amplitude (left) slope (center) and decay time (right) of D-aspartate-inducedsodium transients in astrocytes in control slices (119899 = 161) as well as in cells in the scar region and its periphery (119899 = 111 and 119899 = 26 resp)lowastlowast119875 lt 001 lowastlowastlowast119875 lt 0001 StrP stratum pyramidale StrR stratum radiatum and scale bar 40120583m
cultures obtained from GFPGFAP mice demonstrating thatSR101 is well suited to identify astrocytes in this prepara-tion as described before for acute tissue slices and in vivo[29 30]
Performing a scratch through the CA1 region of organ-otypic slices resulted in the formation of a glial scar along thelesion site as judged by the prominent increase in expressionof GFAP and S100120573 In addition astrocyte cell bodies showed
a significant hypertrophy Furthermore astrocytes in theperiphery of the scar (100ndash350 120583m from the lesion) showedmild to moderate reactivity displaying swollen cell bodiesand only moderate upregulation of GFAP These changesare hallmarks of reactive gliosis observed after tissue injury[10 13 57] Generally the degree of injury and the distance ofthe astrocytes from the site of injury define the degree of theiractivation [11 13]
ISRN Neurology 13
Our study also revealed that strongly reactive astrocytesin the scar region only weakly stain with SR101 The reducedability of strongly activated astrocytes to accumulate SR101indicates that strong glial activation is accompanied by adownregulation of the organic anion transporter responsi-ble for uptake of SR101 This is reminiscent of immatureastrocytes in the early postnatal hippocampus which areSR101-negative [30] and in line with a wealth of experimentalevidence suggesting that reactive gliosis represents a processcomprising a dedifferentiation of astrocytes [17]
42 Changes in Glutamate Transporter Expression followingLesion Glutamate uptake in the hippocampus is mainlyachieved by the glial glutamate transporters GLT-1 andGLAST [1 2] A central factor in many brain pathologiesincluding traumatic brain injury is an increase in extracellu-lar glutamate and excitotoxicity it has been suggested that achange in the expression levels of glial glutamate transportersmight play a critical role in the failure of glutamate clearance[6 58ndash60] Earlier work has found an overall downregulationof protein levels of both GLAST and GLT-1 following astro-cyte activation [22ndash27] indicating that this is causal to theelevation of extracellular glutamate In contrast to this notionother studies reported increased glutamate transport capacityof reactive astrocytes and suggested a protective influence[61 62]
To visualize possible differences in the spatial expressionprofile of GLT-1 and GLAST in strongly reactive astrocytesalong the scar as compared to moderately reactive astro-cytes in its periphery we performed immunohistochemicalstainings These revealed an accumulation of GLT-1 andGLAST immunoreactivity along thick GFAP-positive fiberswhich was particularly pronounced for GLAST Besides theseclusters overall GLAST immunoreactivity seemed unalteredas compared to unlesioned slices whereas GLT-1 immunore-activity seemedweaker and declined with increasing distancefrom the scar Clustering of glutamate transporters has beendescribed by several reports [48 63 64] In developinghippocampal astrocytes cluster formation was preferentiallyfound in branches opposed to synapses and was increasedwith increased neuronal activity indicating that it is neces-sary to copewith synaptic release of glutamate [36] Along thesame lines it was observed that sustained astroglial activationby ciliary neurotrophic factor (CNTF) in the rat striatuminduced a concentration of GLAST and GLT-1 into raftmicrodomains and improved glutamate clearance indicatingthat cluster formation increased the cellular capacity for glu-tamate uptake [61] However another study found glutamate-induced clustering of GLT-1 that induced its endocytosis andintracellular trafficking without changing the total expressionlevels as detected by western blots arguing for a decrease infunctional glial glutamate uptake capacity [49] In the presentstudy antibodies against GLT-1 and GLAST were employedfor immunohistochemistry after permeabilization of theplasma membrane The clusters of GLAST and GLT-1 alongGFAP-positive fibers in the scar region (as GLAST andGLT-1immunoreactivity in general) could thus represent glutamatetransporters in the plasmamembrane or in intracellular com-partments or both Consequently the observed clustering
does not allow a prediction about possible functional changesin glutamate uptake
43 Lesion-Induced Changes in Functional Glutamate UptakeCapacity We probed for the functional activation of gluta-mate transport by application of the transportable agonistD-aspartate which results in an increase in the intracellularsodium concentration of astrocytes [8 9] The amplitudesof D-aspartate-induced sodium transients in astrocytes incontrol organotypic slices were similar to those reportedfrom astrocytes in acute slices obtained in our lab [33]indicating similar cellular glutamate transport capacity inboth preparations
Following a lesion D-aspartate-induced intracellularsodium signalswere clearly dampened in scar cellsMoreoverthe slope of the increase in sodium was significantly reducedand the decay back to baseline was slowed Because D-aspartate was applied at nearly saturating concentrationthe reduction in peak amplitudes suggests a reduction infunctional glutamate uptake capacity in cells close to thelesion which might be mediated by a decrease in the overallnumber of glutamate transporters available at the plasmamembrane In contrast the alteration in the kinetics of theD-aspartate-induced sodium signals can be explained by thecellular hypertrophy that was observed in reactive astrocytesAt an equal transport-mediated sodium influx across themembrane the slope of resulting changes in the sodiumconcentration will be decreased in cells with a larger volume
Furthermore the decay of intracellular sodium transientsback to baseline is mainly governed by the activity of theNa+K+-ATPase as well as by diffusion [8] The slower recov-ery is in line with earlier studies reporting a downregulationof the sodium pump following reactive gliosis [65 66]Moreover sodium is not buffered in the cell and travels solelyby means of diffusion in the cytoplasm Hindered diffusionresulting from increased cytoplasmic protein content orimpaired gap junction coupling which both occur uponastrocytic activation might thus also partially contribute todelayed sodium recovery [12 34 67]
In contrast to scar cells the amplitude of D-aspartate-induced intracellular sodium signals was not significantlyaltered in moderately activated astrocytes in the periphery ofthe lesion indicating that the overall number of functionalglutamate transporters was not significantly altered Becausethese cells showed hypertrophy as well the slowed kinetics ofthe sodium transients are in line with an increased cellularvolume as well as a possible downregulation of Na+K+-ATPase andor a slowed diffusion as argued above
Taken together our results indicate a significant reduc-tion in glutamate uptake capacity in strongly activated astro-cytes in the scar region Thus the prominent clustering ofGLAST and GLT-1 immunoreactivity along GFAP-positivestructures close to the lesion is likely to reflect a loss ofglutamate transporters from the plasma membrane upontransporter internalization as reported by Nakagawa et al[49] The authors of the latter study speculated that large andprolonged increases in glutamate concentrations are neces-sary to induce such cluster formation and endocytosis in thetissue as they would possibly only occur under pathological
14 ISRN Neurology
conditions Indeed ambient glutamate concentrations wereshown to be significantly increased after traumatic braininjury [68ndash70] Along these lines it can be assumed thatin our model system extracellular glutamate concentrationsrose less severely in the periphery of the lesion preventinga comparable clustering and loss of functional glutamatetransport activity in moderately activated astrocytes
5 Conclusions
Our results show complex changes in glutamate transporterexpression and function during astrocyte activation followingmechanical injury They confirm that the degree of astrocyteactivation depends on the distance from the insult and com-prises different endpoints regarding cellular morphology andphysiology Furthermore our results highlight that astrocyteswhich show different grades of reactivity also display discretechanges in glutamate transporter expression and functionWhile immunohistochemistry revealed a prominent clus-tering of GLT-1 and GLAST immunoreactivity in the scarregion our functional assay clearly showed that glutamateuptake capacity is strongly reduced in scar cells while itis largely maintained in moderately activated astrocytes inthe periphery Thus mild-to-moderate astrogliosis in theperiphery of a mechanical lesion does not necessarily seemto be accompanied by a significant change in glial glutamateuptake capacity At the glial scar itself in strongly reactiveastrocytes a clustering of glutamate transporters is observedthat apparently goes along with a severe functional reductionin astroglial glutamate uptake which may contribute toglutamate-mediated excitotoxicity in this region
Conflict of Interests
The authors of the paper declare that they do not haveany direct financial relation with the commercial identitiesmentioned in the paper that might lead to a conflict ofinterests
Acknowledgments
This study was supported by the Deutsche Forschungsge-meinschaft (DFG Ro23274-3) The authors thank SimoneDurry and Claudia Roderigo for expert technical help
References
[1] N C Danbolt ldquoGlutamate uptakerdquo Progress in Neurobiologyvol 65 no 1 pp 1ndash105 2001
[2] A V Tzingounis and J I Wadiche ldquoGlutamate transportersconfining runaway excitation by shaping synaptic transmis-sionrdquo Nature Reviews Neuroscience vol 8 no 12 pp 935ndash9472007
[3] D E Bergles J S Diamond and C E Jahr ldquoClearance ofglutamate inside the synapse and beyondrdquo Current Opinion inNeurobiology vol 9 no 3 pp 293ndash298 1999
[4] C M Anderson and R A Swanson ldquoAstrocyte glutamatetransport review of properties regulation and physiologicalfunctionsrdquo Glia vol 32 no 1 pp 1ndash14 2000
[5] P Marcaggi and D Attwell ldquoRole of glial amino acid trans-porters in synaptic transmission and brain energeticsrdquoGlia vol47 no 3 pp 217ndash225 2004
[6] N J Maragakis and J D Rothstein ldquoGlutamate transportersanimal models to neurologic diseaserdquo Neurobiology of Diseasevol 15 no 3 pp 461ndash473 2004
[7] G Gegelashvili and A Schousboe ldquoCellular distribution andkinetic properties of high-affinity glutamate transportersrdquoBrainResearch Bulletin vol 45 no 3 pp 233ndash238 1998
[8] J W Deitmer and C R Rose ldquoIon changes and signalling inperisynaptic gliardquo Brain Research Reviews vol 63 no 1-2 pp113ndash129 2010
[9] S Kirischuk V Parpura and A Verkhratsky ldquoSodium dynam-ics another key to astroglial excitabilityrdquo Trends in Neuro-sciences vol 35 no 8 pp 497ndash506 2012
[10] M V Sofroniew ldquoMolecular dissection of reactive astrogliosisand glial scar formationrdquo Trends in Neurosciences vol 32 no12 pp 638ndash647 2009
[11] J L Ridet S K Malhotra A Privat and F H Gage ldquoReactiveastrocytes cellular and molecular cues to biological functionrdquoTrends in Neurosciences vol 20 no 12 pp 570ndash577 1997
[12] J Middeldorp and E M Hol ldquoGFAP in health and diseaserdquoProgress in Neurobiology vol 93 no 3 pp 421ndash443 2011
[13] M Pekny and M Nilsson ldquoAstrocyte activation and reactivegliosisrdquo Glia vol 50 no 4 pp 427ndash434 2005
[14] A Bordey S A Lyons J J Hablitz and H SontheimerldquoElectrophysiological characteristics of reactive astrocytes inexperimental cortical dysplasiardquo Journal of Neurophysiologyvol 85 no 4 pp 1719ndash1731 2001
[15] R Jabs G Seifert and C Steinhauser ldquoAstrocytic functionand its alteration in the epileptic brainrdquo Epilepsia vol 49supplement 2 pp 3ndash12 2008
[16] A Buffo I Rite P Tripathi et al ldquoOrigin and progeny ofreactive gliosis a source of multipotent cells in the injuredbrainrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 105 no 9 pp 3581ndash3586 2008
[17] S Robel B Berninger and M Gotz ldquoThe stem cell potential ofglia lessons from reactive gliosisrdquoNature Reviews Neurosciencevol 12 no 2 pp 88ndash104 2011
[18] H Kawano J Kimura-Kuroda Y Komuta et al ldquoRole of thelesion scar in the response to damage and repair of the centralnervous systemrdquo Cell and Tissue Research vol 349 no 1 pp169ndash180 2012
[19] G Yiu and Z He ldquoGlial inhibition of CNS axon regenerationrdquoNature Reviews Neuroscience vol 7 no 8 pp 617ndash627 2006
[20] M T Fitch and J Silver ldquoCNS injury glial scars and inflamma-tion inhibitory extracellular matrices and regeneration failurerdquoExperimental Neurology vol 209 no 2 pp 294ndash301 2008
[21] L Li A Lundkvist D Andersson et al ldquoProtective role ofreactive astrocytes in brain ischemiardquo Journal of Cerebral BloodFlow amp Metabolism vol 28 no 3 pp 468ndash481 2008
[22] F K H van Landeghem T Weiss M Oehmichen and AVonDeimling ldquoDecreased expression of glutamate transportersin astrocytes after human traumatic brain injuryrdquo Journal ofNeurotrauma vol 23 no 10 pp 1518ndash1528 2006
[23] V L Raghavendra Rao M K Baskaya A Dogan J DRothstein and R J Dempsey ldquoTraumatic brain injury down-regulates glial glutamate transporter (GLT- 1 and GLAST)proteins in rat brainrdquo Journal of Neurochemistry vol 70 no 5pp 2020ndash2027 1998
ISRN Neurology 15
[24] J-C Chen H Hsu-Chou J-L Lu et al ldquoDown-regulation ofthe glial glutamate transporter GLT-1 in rat hippocampus andstriatum and its modulation by a group III metabotropic glu-tamate receptor antagonist following transient global forebrainischemiardquoNeuropharmacology vol 49 no 5 pp 703ndash714 2005
[25] S D Ginsberg L J Martin and J D Rothstein ldquoRegional deaf-ferentation down-regulates subtypes of glutamate transporterproteinsrdquo Journal of Neurochemistry vol 65 no 6 pp 2800ndash2803 1995
[26] J E Springer R D Azbill R J Mark J G Begley GWaeg andM P Mattson ldquo4-Hydroxynonenal a lipid peroxidation prod-uct rapidly accumulates following traumatic spinal cord injuryand inhibits glutamate uptakerdquo Journal of Neurochemistry vol68 no 6 pp 2469ndash2476 1997
[27] M B Moretto N S Arteni D Lavinsky et al ldquoHypoxic-ischemic insult decreases glutamate uptake by hippocampalslices from neonatal rats Prevention by guanosinerdquo Experimen-tal Neurology vol 195 no 2 pp 400ndash406 2005
[28] L Stoppini P-A Buchs and D Muller ldquoA simple method fororganotypic cultures of nervous tissuerdquo Journal of NeuroscienceMethods vol 37 no 2 pp 173ndash182 1991
[29] A Nimmerjahn F Kirchhoff J N D Kerr and F HelmchenldquoSulforhodamine 101 as a specific marker of astroglia in theneocortex in vivordquo Nat Methods vol 1 no 1 pp 31ndash37 2004
[30] K W Kafitz S D Meier J Stephan and C R Rose ldquoDevel-opmental profile and properties of sulforhodamine 101-Labeledglial cells in acute brain slices of rat hippocampusrdquo Journal ofNeuroscience Methods vol 169 no 1 pp 84ndash92 2008
[31] C Schnell Y Hagos and S Hulsmann ldquoActive sulforhodamine101 uptake into hippocampal astrocytesrdquo PLoS One vol 7 no 11Article ID e49398 2012
[32] S D Meier Y Kovalchuk and C R Rose ldquoProperties ofthe new fluorescent Na+ indicator CoroNa Green comparisonwith SBFI and confocal Na+ imagingrdquo Journal of NeuroscienceMethods vol 155 no 2 pp 251ndash259 2006
[33] J Langer and C R Rose ldquoSynaptically induced sodium signalsin hippocampal astrocytes in siturdquo Journal of Physiology vol587 part 24 pp 5859ndash5877 2009
[34] J Langer J Stephan M Theis and C R Rose ldquoGap junctionsmediate intercellular spread of sodium between hippocampalastrocytes in siturdquo Glia vol 60 no 2 pp 239ndash252 2012
[35] C R Rose andB R Ransom ldquoIntracellular sodiumhomeostasisin rat hippocampal astrocytesrdquo Journal of Physiology vol 491part 2 no 2 pp 291ndash305 1996
[36] A M Benediktsson G S Marrs J C Tu et al ldquoNeuronalactivity regulates glutamate transporter dynamics in developingastrocytesrdquo Glia vol 60 no 2 pp 175ndash188 2012
[37] M Zhou G P Schools and H K Kimelberg ldquoDevelopment ofGLAST(+) astrocytes andNG2(+) glia in rat hippocampus CA1mature astrocytes are electrophysiologically passiverdquo Journal ofNeurophysiology vol 95 no 1 pp 134ndash143 2006
[38] I Suarez G Bodega and B Fernandez ldquoModulation of glu-tamate transporters (GLAST GLT-1 and EAAC1) in the ratcerebellum following portocaval anastomosisrdquo Brain Researchvol 859 no 2 pp 293ndash302 2000
[39] X Zhu D E Bergles and A Nishiyama ldquoNG2 cells generateboth oligodendrocytes and gray matter astrocytesrdquo Develop-ment vol 135 no 1 pp 145ndash157 2008
[40] B Brunne S ZhaoADerouiche et al ldquoOriginmaturation andastroglial transformation of secondary radial glial cells in thedeveloping dentate gyrusrdquo Glia vol 58 no 13 pp 1553ndash15692010
[41] Y Wu A-Q Zhang and D T Yew ldquoAge related changes ofvarious markers of astrocytes in senescence-accelerated micehippocampusrdquo Neurochemistry International vol 46 no 7 pp565ndash574 2005
[42] S Magavi D Friedmann G Banks A Stolfi and C LoisldquoCoincident generation of pyramidal neurons and protoplasmicastrocytes in neocortical columnsrdquo Journal of Neuroscience vol32 no 14 pp 4762ndash4772 2012
[43] M Olabarria H N Noristani A Verkhratsky and J JRodrıguez ldquoConcomitant astroglial atrophy and astrogliosis ina triple transgenic animal model of Alzheimerrsquos diseaserdquo Gliavol 58 no 7 pp 831ndash838 2010
[44] A M Benediktsson S J Schachtele S H Green and M EDailey ldquoBallistic labeling and dynamic imaging of astrocytesin organotypic hippocampal slice culturesrdquo Journal of Neuro-science Methods vol 141 no 1 pp 41ndash53 2005
[45] I Lushnikova G Skibo D Muller and I Nikonenko ldquoSynapticpotentiation induces increased glial coverage of excitatorysynapses in CA1 hippocampusrdquoHippocampus vol 19 no 8 pp753ndash762 2009
[46] E Raponi F Agenes C Delphin et al ldquoS100B expressiondefines a state in which GFAP-expressing cells lose their neuralstem cell potential and acquire a more mature developmentalstagerdquo Glia vol 55 no 2 pp 165ndash177 2007
[47] C G Schipke C Boucsein C Ohlemeyer F Kirchhoff andH Kettenmann ldquoAstrocyte Ca2+ waves trigger responses inmicroglial cells in brain slicesrdquo The FASEB Journal vol 16 no2 pp 255ndash257 2002
[48] J Zhou and M L Sutherland ldquoGlutamate transporter clusterformation in astrocytic processes regulates glutamate uptakeactivityrdquo Journal of Neuroscience vol 24 no 28 pp 6301ndash63062004
[49] T Nakagawa Y Otsubo Y Yatani H Shirakawa and S KanekoldquoMechanisms of substrate transport-induced clustering of aglial glutamate transporter GLT-1 in astroglial-neuronal cul-turesrdquo European Journal of Neuroscience vol 28 no 9 pp 1719ndash1730 2008
[50] J-Y Chatton P Marquet and P J Magistretti ldquoA quantitativeanalysis of L-glutamate-regulated Na+ dynamics in mousecortical astrocytes Implications for cellular bioenergeticsrdquoEuropean Journal of Neuroscience vol 12 no 11 pp 3843ndash38532000
[51] E Forster S Zhao and M Frotscher ldquoLaminating the hip-pocampusrdquo Nature Reviews Neuroscience vol 7 no 4 pp 259ndash267 2006
[52] B H Gahwiler M Capogna D Debanne R A McKinney andS M Thompson ldquoOrganotypic slice cultures a technique hascome of agerdquo Trends in Neurosciences vol 20 no 10 pp 471ndash477 1997
[53] P E Kunkler and R P Kraig ldquoReactive astrocytosis fromexcitotoxic injury in hippocampal organ culture parallels thatseen in vivordquo Journal of Cerebral Blood Flow and Metabolismvol 17 no 1 pp 26ndash43 1997
[54] I E Holopainen ldquoOrganotypic hippocampal slice cultures amodel system to study basic cellular andmolecularmechanismsof neuronal cell death neuroprotection and synaptic plasticityrdquoNeurochemical Research vol 30 no 12 pp 1521ndash1528 2005
[55] MHaber L Zhou andK KMurai ldquoCooperative astrocyte anddendritic spine dynamics at hippocampal excitatory synapsesrdquoJournal of Neuroscience vol 26 no 35 pp 8881ndash8891 2006
16 ISRN Neurology
[56] H Nishida and S Okabe ldquoDirect astrocytic contacts regulatelocal maturation of dendritic spinesrdquo Journal of Neurosciencevol 27 no 2 pp 331ndash340 2007
[57] A Buffo C Rolando and S Ceruti ldquoAstrocytes in the dam-aged brain molecular and cellular insights into their reactiveresponse and healing potentialrdquoBiochemical Pharmacology vol79 no 2 pp 77ndash89 2010
[58] J-H Yi and A S Hazell ldquoExcitotoxic mechanisms and the roleof astrocytic glutamate transporters in traumatic brain injuryrdquoNeurochemistry International vol 48 no 5 pp 394ndash403 2006
[59] C Werner and K Engelhard ldquoPathophysiology of traumaticbrain injuryrdquo British Journal of Anaesthesia vol 99 no 1 pp4ndash9 2007
[60] G Gegelashvili M B Robinson D Trotti and T RauenldquoRegulation of glutamate transporters in health and diseaserdquoProgress in Brain Research vol 132 pp 267ndash286 2001
[61] C Escartin E Brouillet PGubellini et al ldquoCiliary neurotrophicfactor activates astrocytes redistributes their glutamate trans-porters GLAST and GLT-1 to raft microdomains and improvesglutamate handling in vivordquo Journal of Neuroscience vol 26 no22 pp 5978ndash5989 2006
[62] C Vermeiren M Najimi N Vanhoutte et al ldquoAcute up-regulation of glutamate uptake mediated by mGluR5a in reac-tive astrocytesrdquo Journal of Neurochemistry vol 94 no 2 pp405ndash416 2005
[63] C L Poitry-Yamate L Vutskits and T Rauen ldquoNeuronal-induced and glutamate-dependent activation of glial glutamatetransporter functionrdquo Journal of Neurochemistry vol 82 no 4pp 987ndash997 2002
[64] P M Beart and R D OrsquoShea ldquoTransporters for L-glutamatean update on their molecular pharmacology and pathologicalinvolvementrdquo British Journal of Pharmacology vol 150 no 1pp 5ndash17 2007
[65] F D Lima M A Souza A F Furian et al ldquoNa+K+-ATPaseactivity impairment after experimental traumatic brain injuryrelationship to spatial learning deficits and oxidative stressrdquoBehavioural Brain Research vol 193 no 2 pp 306ndash310 2008
[66] G A Gusarova H E Trejo L A Dada et al ldquoHypoxia leadsto NaK-ATPase downregulation via Ca2+ release-activatedCa2+ channels and AMPK activationrdquo Molecular and CellularBiology vol 31 no 17 pp 3546ndash3556 2011
[67] S S L Chew C S Johnson C R Green and H V Danesh-Meyer ldquoRole of connexin43 in central nervous system injuryrdquoExperimental Neurology vol 225 no 2 pp 250ndash261 2010
[68] P Nilsson L Hillered U Ponten and U Ungerstedt ldquoChangesin cortical extracellular levels of energy-related metabolites andamino acids following concussive brain injury in ratsrdquo Journal ofCerebral Blood Flow and Metabolism vol 10 no 5 pp 631ndash6371990
[69] H Katoh K Sima H Nawashiro K Wada and H ChigasakildquoThe effect of MK-801 on extracellular neuroactive amino acidsin hippocampus after closed head injury followed by hypoxia inratsrdquo Brain Research vol 758 no 1-2 pp 153ndash162 1997
[70] H Koizumi H Fujisawa H Ito T Maekawa X Di and RBullock ldquoEffects of mild hypothermia on cerebral blood flow-independent changes in cortical extracellular levels of aminoacids following contusion trauma in the ratrdquoBrain Research vol747 no 2 pp 304ndash312 1997
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

2 ISRN Neurology
human brain reported an overall downregulation of proteinlevels of both GLAST and GLT-1 andor a reduction infunctional glutamate uptake in response to brain injury andastrogliosis respectively (eg [22ndash27]) The reduction in theexpression of glial glutamate transporters has been suggestedto contribute to the elevation of extracellular glutamate con-centrations and to glutamate-mediated excitotoxicity whichis observed under many pathological conditions
Injury-induced changes in the expression of the gluta-mate transporter subtypes might however also depend onthe degree of astrogliosis This might be especially relevantand visible upon a mechanical lesion where reactivity ofastrocytes as judged for example based on their GFAPexpression and morphology ranges from severe astroglio-sis in the scar region to only moderate-mild astrogliosisdistant from the lesion [10 11] To study this questionwe analyzed expression of GLAST and GLT-1 following amechanical lesion in organotypic tissue slice cultures of themouse hippocampus Furthermore we employed ratiometricsodium imaging as functional assay for glutamate uptake inastrocytes Our results show that mechanical tissue injurygenerates subsets of reactive astrocytes depending on thedistance from the lesion site which differ in bothmorpholog-ical features and their ability to take up SR101 Furthermorethese subsets show discrete changes in glutamate transporterexpression and glutamate uptake capacity indicating that glu-tamate clearance might be largely functional in the peripheryof the lesion but strongly hampered in the scar region
2 Materials and Methods
21 Ethics Statement This study was carried out in strictaccordance with the institutional guidelines of the Hein-rich Heine University Duesseldorf Germany as well as theEuropean Community Council Directive (86609EEC) Allexperiments were communicated to and approved by theAnimal Welfare Office at the Animal Care and Use Facilityof the Heinrich Heine University Duesseldorf Germany(institutional act number O5205) In accordance with theGerman Animal Welfare Act (Tierschutzgesetz Articles 4and 7) no formal additional approval for the postmortemremoval of brain tissue was necessary For generation ofacute slices mice were quickly decapitated (following therecommendation of the European Commission published inEuthanasia of experimental animals Luxembourg Office forOfficial Publications of the European Communities 1997ISBN 92ndash827-9694-9)
22 Preparation of Tissue Slice Cultures Acute tissue slicesof the hippocampus were prepared from Balbc mice (Musmusculus) of both genders at postnatal days 7 to 8 (P7-8)using standard procedures For some experiments transgenicanimals (FVBN-Tg[GFAPGFP]14MesJ) expressing greenfluorescence protein (GFP) under the GFAP promoter wereused (obtained from Jackson Laboratory Harbor USA)After decapitation of the animals brains were quickly excisedand hemisected in ice-cold artificial cerebrospinal fluid(ACSF) composed of (in mM) 125 NaCl 25 KCl 2 CaCl
2
1 MgCl2 125 NaH
2PO4 26 NaHCO
3 and 20 glucose bub-
bled with 95 O2and 5 CO
2 and adjusted to a pH of 74
Hemisections were trimmed and transverse slices (200120583m)comprising the entorhinal cortex hippocampus fimbriaand thalamus were prepared using a vibratome (MicromHM650V Thermo Fischer Scientific Walldorf Germany)Slices were transferred to ACSF at 35∘C for 30 minutes
Organotypic hippocampal slice cultures were preparedand cultured according to the protocol introduced by Stop-pini et al [28] with minor modifications To this end acuteslices were transferred to a Millicell culture insert (PICMORG 50 hydrophilized PTFE pore size 04 120583m Merck Mil-lipore Darmstadt Germany) and maintained at the interfaceof a serum-based culture medium free of antibiotics in ahumidified incubator atmosphere of 5 CO
2at 37∘C The
culture medium was composed of 30 sterile filtered normalhorse serum (NHS GIBCOLife Technologies DarmstadtGermany) 30Dulbeccorsquos modified eagle medium (DMEMGIBCOLife Technologies Darmstadt Germany) and 40Hankrsquos balanced salt solution (HBSS GIBCOLife Tech-nologies Darmstadt Germany) supplemented with 38mMglucose (pH adjusted to 73-74) The medium was changedthree times a week and the insert surface was washed withmedium once a week
After at least 12 days in culture a mechanical lesionwas performed using a sterile scalpel blade The lesion waspositioned in the CA1 area perpendicular to the stratumpyramidale included the strata oriens pyramidale radiatumand lacunosum moleculare and spanned the entire depthof the slice (cf Figure 2) After lesioning slice cultureswere maintained for another 6-7 days (designated as 6-7days postlesion) Unlesioned control slices were cultured inparallel for a corresponding number of days
For visualization of cell death ACSF containing05 120583gmL propidium iodide (PI) was applied to the slicesurface and incubated for 3 hours at 37∘C and 5 CO
2
followed by a wash with ACSF Documentation was eitheraccomplished with an epifluorescence microscope (NikonEclipse 90i Nikon Instruments Dusseldorf Germany) orat an Olympus Fluoview300 laser scanning microscope(Olympus Hamburg Germany)
Astrocyte soma size in organotypic control and lesionedslices was calculated from images of GFP-fluorescencederived fromGFPGFAPmice which were obtained at a con-focal microscope (Olympus Fluoview300 Olympus Ham-burg Germany see also below) To this end a semiquantita-tive approach was used in which somata of single cells weremanually encircled (cf Figure 4(b)) and the resulting areawas calculated using ImageJ software (NIH Bethesda USA)
All chemicals were purchased from Sigma-AldrichChemical (Munich Germany) unless stated otherwise
23 Imaging Experiments For imaging experiments organ-otypic slices were excised from the Millicell inserts andincubated in ACSF containing 25120583M SR101 for 30minat 35∘C to stain astrocytes (see above) SR101 is a highlyspecific and widely used tool for the identification of matureastrocytes in many brain regions including the hippocampus
ISRN Neurology 3
[29 30] It has recently been shown to be taken up intomaturehippocampal astrocytes via an active transport mechanisminvolving organic anion transport polypeptides [31] Wide-field fluorescence imaging was performed using a variablescan digital imaging system (TILL Photonics MartinsriedGermany) attached to an upright microscope (BX51WiOlympus Europe Hamburg Germany) and a CCD camera(TILL Imago VGA Till Photonics Martinsried Germany)Imageswere collectedwith anAchroplan 40x objective (waterimmersion NA 08 Zeiss Gottingen Germany) SR101 wasexcited at 575 nm and emission was collected above 590 nmExcitation wavelength for detection of GFP was 488 nm andemission was collected above 510 nm
For intracellular sodium imaging slices were addition-ally incubated with the membrane permeable form of thesodium-sensitive fluorescent dye SBFI (SBFI-AM sodium-binding benzofuran isophthalate-acetoxymethyl ester Mol-ecular ProbesLife Technologies Darmstadt Germany) asdescribed earlier [32ndash34] Ratiometric sodium imaging wasperformed by alternate excitation of SBFI at 340 nm (weaklysodium-sensitive wavelength) and at 380 nm (sodium-sensitive wavelength) at 4Hz Emission (gt440 nm) wascollected in defined regions of interest (ROI) representingcell bodies Standard dynamic background correction wasperformed as described earlier [33 34] After backgroundcorrection the fluorescence ratio (F
340F380
) was calculatedfor the individual ROIs and analyzed offline using OriginPro8G Software (OriginLab Corporation Northampton MAUSA)
Changes in SBFI fluorescence ratio were expressed aschanges in sodium concentration based on in situ calibrationsas reported before [32 33 35] To equilibrate extra- andintracellular Na+-concentrations SBFI-loaded slices wereperfused with saline containing ionophores (3 120583M grami-cidin D 100 120583Mmonensin) and the Na+K+-ATPase blockerouabain (100 120583M) as well as different concentrations of Na+
The glutamate transporter agonist D-aspartate wasapplied by a pressure application device (PDES-02D NPIElectronic GmbH Tamm Germany) coupled to standardmicropipettes (Hilgenberg Waldkappel Germany) placed20ndash100120583m from cell bodies of selected cells
24 Antibodies Antibodies employed in this study are listedin Table 1 The following antisera were utilized whichrepresent well-established commercially available standardmarkers guinea-pig GLT-1 antiserum directed against theC-terminus of rat GLT-1 (Chemicon InternationalMilliporeCorp Carrigtwohill Ireland eg [36]) and guinea-pigGLAST antiserum directed against the C-terminus of ratGLAST (Chemicon InternationalMillipore Corp Carrigt-wohill Ireland eg [37]) Validation and a detailed charac-terization of both antibodies were provided before [38ndash40]For identification of astrocytes polyclonal rabbit antibodiesagainst GFAP (GFAP-pAb Dako Cytomation Denmark[40 41]) and S100120573 (Abcam United Kingdom [42]) wereemployedWhen double staining with the latter antibody wasperformed a monoclonal mouse antibody detecting GFAPwas used (GFAP-mAb Sigma Aldrich Germany [40 43])
Standard fluorochrome-conjugated antibodies (Alexa-Fluor InvitrogenLife Technologies Darmstadt Germany)were employed as secondary antibodies for immunohisto-chemistry
25 Immunohistochemistry Prior to immunohistochemicalprocessing organotypic slices were immersion fixed for30min at RT in 4 paraformaldehyde (PFA) in phosphate-buffered saline (PBS) following three washes every30min with PBS Cell membranes were permeabilizedand unspecific binding sites were blocked in PBS containing025 triton-X100 (TX) and 2 normal goat serum (NGSGIBCOLife Technologies Darmstadt Germany) for 90minat 4∘C followed by incubation with the primary antibodyGFAP-pAb (1 1000 Dako Cytomation Glostrup Denmark)diluted in the same solution over night at 4∘C In case ofS100120573GFAP double stainings slices were incubated witha mixture of the primary antibodies GFAP-mAb (1 1000Dako Cytomation Glostrup Denmark) and rabbit-S100120573(1 100 Abcam United Kingdom) After five washes in PBScontaining 025 TX and 2 NGS slices were incubatedeither with guinea pig-anti-GLAST or with guinea pig-anti-GLT-1 (both 1 1000 diluted in 025 TX2 NGSPBS) for4 hrs at RT Excess primary antibody was removed with fivewashes using 2 NGSPBS Antirabbit-AlexaFluor594 andantiguinea pig-AlexaFluor488 or antimouse-AlexaFluor488(1 100 in blocking solution) were used for visualizationof antibody binding and incubated for 2 hrs at RT Theslices were subjected to DAPI staining (410158406-diamidino-2-phenylindole 05120583m Invitrogen) washed three times andmounted on glass slides with mowiolDABCO (CalbiochemFluka distributed by Sigma-Aldrich Chemical MunichGermany)
Identical conditions were applied to all performed stain-ings regarding tissue processing and staining procedureNegative controls were run in parallel to each staining byeither omitting all or just one of the primary antibodiesControl stainings in which one of the primary antibodieswas omitted showed the identical labeling pattern for theremaining antibody as in the double stainings Omitting bothprimary antibodies never resulted in a staining
Documentation of immunofluorescence was either per-formed with an epifluorescence microscope (Nikon Eclipse90i Nikon Instruments Dusseldorf Germany) or a confocallaser scanningmicroscope (Olympus Fluoview300OlympusHamburg Germany) The epifluorescence microscope wasequipped with a standard DAPI (EX 340ndash380 DM 400BA 435ndash485) FITC (EX 465ndash495 DM 505 BA 515ndash555)and TRITC (EX 54025 DM 365 BA 60555) filter setIllumination was provided by an Intensilight fiber lamp(C-GHFI Nikon Instruments Dusseldorf Germany) andemission was detected with a monochrome digital cam-era (DS-Qi1Mc Nikon Instruments Dusseldorf Germany)Images were collected with either a 20x075 (PlanApoVCNikon Instruments Dusseldorf Germany) air objective ora 60x140 oil immersion objective (PlanApoVC NikonInstruments Dusseldorf Germany) NIS-Elements software(Nikon Instruments Dusseldorf Germany) was used forimage acquisition For confocal microscopy an Olympus
4 ISRN Neurology
Table 1 List of antibodies used
Antibody Structure of theimmunogen Company Order number Species Type Dilution
GLAST(EAAT1)
KPYQLIAQDNEPEKPVADSETKM ChemiconMillipore AB1782lowast Guinea
Pig IgGPolyclonal 1 1000
GLT-1(EAAT2)
AANGKSADCSVEEEPWKREK ChemiconMillipore AB1783 Guinea
Pig IgGPolyclonal 1 10001 2000
GFAP ns (full-length proteinfrom cow spinal cord) Dako Cytomation Z 0334 Rabbit
IgGPolyclonal 1 10001 2000
GFAP
clone G-A-5(LQSLTCDVESLRGTNESLERQMREQEERHAREAASYQEALTRLEEEGQSLKDEMARHLQEYQELLNVKLALDIEIATY)
Sigma-Aldrich G3893 MouseIgG
Monoclonal 1 10001 2000
S100szlig ns (recombinantfull-length cow protein) Abcam ab868 Rabbit
IgGPolyclonal 1 100
MouseIgG AF488 conjugated Invitrogen (Life Technologies) A11029 Goat mdash 1 100
RabbitIgG AF594 conjugated Invitrogen (Life Technologies) A11012 Goat mdash 1 100
Guineapig IgG AF488 conjugated Invitrogen (Life Technologies) A11073 Goat mdash 1 100
GLAST glutamate aspartate transporter GLT-1 glutamate transporter 1 GFAP glial fibrillary protein AF AlexaFluor appl application IHC immunohisto-chemistry WB western blot ns not specifiedlowastThemanufacturer has lately stopped the sale and distribution of the GLAST antibody used here
BX51WI microscope coupled to a confocal laser scanningsystem (FV300) equipped with a multiline argon (488 nm)and a helium-neon laser (543 nm both Melles Griot Ben-sheim Germany) was used Images were collected witheither a 20x050 (UMPlanFl Olympus Hamburg Ger-many) a 40x080 water immersion (LUMPlan OlympusHamburg Germany) or a 60x140 oil immersion objective(PlanApoVC Nikon Instruments Dusseldorf Germany) AKalman filter 4 was employed at every scan Simultaneousor sequential scanning of both fluorophores revealed nodifference in their staining patterns indicating absence ofcross-excitation or spectral bleed through The thickness ofz-plane sections was 1120583m and the number of optical sectionsvaried depending on the preparation
Figures show extended focus images as specified inthe figure legends which were calculated from z-stacksof optical sections using ImageJ software (NIH BethesdaUSA) Images were overlaid employing Adobe PhotoshopCS2 (Adobe Systems Cologne Germany)
26 Data Presentation and Statistics Unless otherwise spec-ified data are expressed as means plusmn SEM Data were statis-tically analyzed by Studentrsquos 119905-test employing the proceduresimplemented in OriginPro 8G Software (OriginLab Corpo-ration Northampton MA USA) A 119875 value of lt005 wasconsidered significant If not stated otherwise 119899representsthe number of analyzed cells and119873 the number of slices Eachset of experimentswas performedon at least three tissue slicesobtained from different animals
3 Results
31 Identification of Astrocytes in Organotypic Slice CulturesThe aim of the present study was to analyze changes inglutamate transporter expression and function in astrocytesin response to a mechanical injury To this end we preparedorganotypic tissue slice cultures of themouse hippocampus awell-establishedmodel system inwhich the layering and basiccellular organization of the neural network are maintainedover time [28] After 19ndash25 days in culture organotypic tissueslices had flattened to a thickness of 40ndash50 120583m and werecomposed of 3 to 4 cell layers At this stage propidium iodideassays detected only few dead cells dispersed throughoutthe slices (3 plusmn 06 cells per CA1 subfield 18 plusmn 34 in entirepreparation 119873 = 6 data not shown) indicating that thepreparation was viable and in a stable condition
To visualize astrocytes in cultured slices we used hip-pocampi of transgenic mice in which green fluorescentprotein (GFP) is expressed under the control of the promoterof the astrocyte-specific intermediate filament glial fibrillaryacidic protein (GFAP FVBN-Tg[GFAPGFP] ldquoGFPGFAPmicerdquo) A visualization of theGFP fluorescence demonstratedthat the general distribution and organization of astrocyteswere preserved in the strata pyramidale and radiatum ofthe CA1 region of organotypic cultures confirming earlierreports (119873 = 3 Figure 1(a) [44 45]) The vast majority ofGFP-expressing cells (sim95) also labeled with the astrocyte-specific vital marker SR101 in organotypic slices (119873 = 3Figures 1(a) and 1(b)) Furthermore immunohistochemicalstainings for GFAP in slice cultures derived from wildtype
ISRN Neurology 5
Slic
e cul
ture
(20
dic
)
GFP-GFAP MergeSR101
lowast lowast lowast lowast
lowast
lowast
lowast lowast
StrP
StrR
(a)
StrR
StrP
StrLM
StrM
SR101
StrO
(b)
GFAP S100120573
StrP
StrR
StrP
StrR
StrP
StrR
StrP
StrR
DAPI Merge
(c)
Figure 1 Astrocytes in organotypic hippocampal slice cultures (a) Organotypic slice cultures at 20 days in culture (dic) Preparations werederived from animals in which GFP was expressed under control of the GFAP promoter Shown is the expression of GFP (left) the labelingpattern of SR101 (center) and the corresponding merged image (GFAP green SR101 magenta) Asterisks point out SR101-labeled cells thatdo not express GFP arrowheads point out GFP-positive but SR101-negative cells Images represent maximum intensity projections of 15optical sections taken at 1120583m intervals (b) SR101 fluorescence of an organotypic slice at 21 dic Image represents a stack of 20 optical sectionstaken at 1 120583m intervals (c) Immunohistochemical double staining for GFAP and S100120573 as well as DAPI fluorescence and merged image ofthe fluorescence of GFAP (green) and S100120573 (magenta) of part of the CA1 region encompassing the strata pyramidale and radiatum of anorganotypic slice at 20 dic Note the radially oriented GFAP-positive fibers crossing the pyramidal cell layer Images were taken at a wide-field microscope ((a)ndash(c)) Dashed lines indicate the approximate boundaries of the stratum pyramidale StrO stratum oriens StrP stratumpyramidale StrR stratum radiatum StrLM stratum lacunosum moleculare and StrM stratum moleculare Scale bars (a) 50 120583m (b) 100120583mand (c) 50 120583m
animals showed that themajority of GFAP-positive cells werealso positive for S100120573 (119873 = 12 Figure 1(c)) a marker formature astrocytes [46]
Taken together the nearly complete overlap in the cellularstaining pattern for the vital dye SR101 with the expressionof GFP in GFPGFAP mice as well as the overlap in theimmunofluorescence for GFAP and S100120573 in organotypicslices indicates that these markers are well suited to identifyastrocytes in this preparation
32 Astrocyte Markers and Astrocyte Morphology followingMechanical Lesion To induce astrogliosis in organotypicslice cultures we used a scalpel blade and performed
a scratch under semisterile conditions through the entireCA1 region oriented perpendicular to the stratum pyramidale(Figure 2(a)) 6-7 days after the lesion a general enhance-ment of GFAP immunoreactivity was observed indicatingastrogliosis (Figure 2(b))The greatest enhancement of GFAPimmunoreactivity occurred within a distance of less than100 120583malong the lesion (119873 = 26 Figures 2(b) and 2(c)) Herea densemeshwork of thick GFAP-positive labels was presentindicative of the formation of a glial scar Furthermore theregion close to the lesion site exhibited a discernible increasein S100120573 immunoreactivity (119873 = 12 Figure 2(c)) Both theprominent expressions of GFAP and S100120573 are characteristicsof strongly reactive astrocytes which represent the major
6 ISRN Neurology
GFAP
DAPI
CA1 subregion
GFAP
DAPI
StrR
StrP StrP
StrR
StrOStrO
StrO
StrP
StrR
StrO
StrP
StrR
Control Lesioned
GFAP
Lesio
n sit
e
Hi
StrMStrG
StrP
StrR
StrLM
Scratch wound
StrO
Lesio
n sit
e
Lesioned
(a) (c)
(b)
350120583m350120583m
S100120573
Figure 2 Lesion-induced reactive gliosis in organotypic slice cultures (a) Schematic overview of the hippocampus indicating the positionof the scratch woundThe box delineates the area primarily analyzed (b) GFAP immunofluorescence and DAPI staining of the CA1 area in acontrol (left) and a lesioned (right) slice The boxes indicate the areas enlarged below The dashed line in the lower right indicates the lesion(c) Double staining for GFAP (top) and S100120573 (bottom) adjacent to the lesion site Note the gradual increase in GFAP and S100120573 expressiontowards the lesion (b) widefield microscopy (c) confocal microscopy (optical sections taken at 1120583m intervals) StrO stratum oriens StrPstratum pyramidale StrR stratum radiatum and StrLM stratum lacunosum moleculare Scale bars (b) 200 120583m (top) and 50 120583m (bottom) (c)10120583m
ISRN Neurology 7
StrR
StrP
StrLM
StrO
StrP StrRStrO StrLM
Lesio
n sit
e
SR101-staining pattern
(a)
SBFI
Lesio
n sit
e
SR101 GFAP-GFP
(b)
Figure 3 SR101 labeling in lesioned slices (a) SR101 fluorescence in the CA1 area in a slice 7 days after lesion The delineated area comprisesthe stratum pyramidale and the box indicates the area enlarged on the right side Right SR101 fluorescence at the lesion site illustrating thatSR101 labeling is reduced along the lesionThe dotted line indicates the transition between the scar area and the adjacent moderately reactivetissue The boxed area is further enlarged at the bottom (b) Image series obtained at a wide-field fluorescence microscope The slice wasprepared from a transgenic GFPGFAP mouse and the lesion site is indicated by the dotted line Left Image of SR101 fluorescence centerGFP fluorescence indicating GFAP expression and right SBFI fluorescence SR101 labeling is only weak in GFPSBFI-positive cells in the scarregion (see arrowheads) StrO stratum oriens StrP stratum pyramidale StrR stratum radiatum and StrLM stratum lacunosum-moleculareScale bars 100120583m ((a) upper left) and 20 120583m (other images)
cellular component of the glial scar forming after mechanicaltissue injury [10] More distal to the lesion (gt100ndash350 120583m) aless dramatic rearrangement of GFAP immunoreactivity wasobserved
A clear difference in astrocyte properties between thescar region and its periphery was also seen following SR101labeling In the periphery of the scar (100ndash350 120583m distancefrom the lesion) SR101 resulted in a reliable and brightcellular staining similar to what had been observed incontrol unlesioned slices (119873 = 12 Figure 3(a) cf Figure 1)
In contrast to this cellular SR101 staining was detectable butonly faint within a distance of less than 100 120583m on both sidesof the lesion (Figure 3(a) meanwidth of SR101-free area 111plusmn3 120583m 119873 = 5) In slices from GFPGFAP transgenic animals(119873 = 5 Figure 3(b)) GFAP-positive weakly SR101-positivescar cells also stained with the sodium-sensitive fluorescenceindicator SBFI-AM (acetoxymethyl ester of sodium bindingbenzofuran isophthalate see below 119873 = 3) indicatingthat they were able to take up and de-esterify this dye(Figure 3(b))
8 ISRN Neurology
lowastlowastlowast
lowastlowastlowast
80
lowast
Control ScarPeriphery
120
160
0
40
200
Som
a siz
e (120583
m2)
(a)
Lesioned
StrP
StrR
StrP
Control
StrR
StrLM
Lesio
n sit
e
(b)
Figure 4 Lesion-induced changes in somatic area (a) Quantification of somatic area based on GFAPGFP fluorescence in control andlesioned slices The soma size increased both in astrocytes in the scar region (ldquoscarrdquo) and in astrocytes in the periphery of the lesion(ldquoperipheryrdquo lowastlowastlowast119875 lt 0001 lowast119875 lt 005) (b) Confocal images of GFP fluorescence in cultured slices obtained from GFPGFAP transgenicanimals under control conditions (left) and subjected to a lesion (right) Cells surrounded by boxes are shown at higher magnification in theinsets and the dashed lines around the soma illustrate the areas which served to determine their size StrP stratum pyramidale StrR stratumradiatum StrLM stratum lacunosum moleculare and scale bars 40 120583m and 10120583m (inset)
In addition to GFAP-positive cells the scar region hostedmicroglial cells identified by their vital staining with TexasRed-coupled lectin from lycopersicon esculentum (tomatolectin119873 = 4 not shown) Tomato lectin-positive microgliahowever did not take up SBFI-AM (119873 = 4 not shown) con-firming earlier reports thatmicroglial cells do not incorporateAM-ester dyes in situ [47]
To further characterize reactive gliosis in response to thescratch wound we determined the area of astrocyte somatain confocal images of the GFP fluorescence in control andlesioned slices obtained fromGFPGFAP transgenic animalsThesemeasurements revealed that compared to astrocytes in
control slices the soma area of astrocytes in the periphery ofa lesion was increased by 18 and that of astrocytes in thescar regionwas increased by 24 (control 163plusmn2 120583m2 lesionperiphery 193plusmn3 120583m2 scar tissue 201plusmn4 120583m2119875 lt 0001 119899 =945 496 and 439 in 7 control and 9 lesioned slices Figures4(a) and 4(b)) Thus astrocytes show a second hallmark ofreactive gliosis after lesion namely a hypertrophy of their cellbodies which is observed both in the scar tissue as well as inthe periphery of the lesion
Taken together these data show that 6-7 days after themechanical lesion a glial scar has formed along the lesionsite This scar encompasses strongly activated astrocytes
ISRN Neurology 9
characterized by a robust increase in GFAP and S100120573expressionThese proximal reactive astrocytes locatedwithina distance of less than 100120583m from the lesion exhibit longGFAP-positive processes have significantly swollen somataand only weakly stain with SR101 Astrocytes in the peripheryof the scar (100ndash350 120583m from the lesion) display swollen cellbodies as well but show onlymoderate upregulation of GFAPand maintain their ability to efficiently take up SR101
33 Expression Levels and Spatial Distribution of GLT-1 andGLAST To determine the spatial distribution of the gluta-mate transporters GLAST and GLT-1 in organotypic slicesand changes therein in response to the lesion immunohisto-chemistry was employed In control slices GLAST (119873 = 13Figure 5(a)) as well as GLT-1 immunoreactivity (119873 = 11Figure 5(b)) appeared as punctate labeling throughout theentire CA1 area The staining patterns for both transportersin the tissue slice cultures were thus similar to those reportedfrom acute tissue slice preparations of the hippocampus [3648 49]
The relatively homogeneous distribution of immunore-activity throughout the preparation found in control slicecultures was abandoned following a lesion Here distinctaccumulations of GLAST (119873 = 17 Figure 5(a)) and GLT-1immunoreactivity (119873 = 12 Figure 5(b)) were found in thescar region In both cases these preferentially colocalizedwith thick GFAP-positive bundles running in parallel to thelesion (Figure 5) Apart from the marked accumulation ofGLAST reactivity in parallel to the scar GLAST immunore-activity appeared generally increased in some lesioned slicepreparations (Figure 5(a)) but unaltered in others bothwithinthe scar as well as in its periphery For GLT-1 a decline in theintensity of immunoreactivity was observed with increasingdistance from the lesion (Figure 5(b))
In summary these results reveal distinct alterations inthe immunoreactivity of GLAST and GLT-1 following amechanical lesion In the scar region clusters of GLT-1 andGLAST immunoreactivity are present primarily colocalizedwith strongly GFAP-positive fibers Apart from these distinctclusters GLT-1 immunoreactivity appears to decline withdistance from the scar whereas GLAST immunoreactivityseems largely uniform throughout the preparation
34 Sodium Transients Induced by Activation of GlutamateUptake Glutamate uptake is accompanied by the inwardflux of sodium resulting in increase in the intracellularsodium concentration of astrocytes To monitor its functionwe assayed sodium increases induced by D-aspartate anonmetabolized substrate of sodium-dependent glutamateuptake which is also transported [1] in astrocytes in thestratum radiatum of organotypic slices To this end SR101-stained slices were additionally loaded with the sodium-sensitive fluorophore SBFI-AM (Figure 6(a) cf Figure 3(b))In situ calibrations of the SBFI fluorescence revealed a linearincrease in its fluorescence ratio at intracellular sodiumconcentrations between 10 and 40mM (119899 = 107 Figure 6(b))as reported before [33]
Repetitive pressure application of D-aspartate for 500msinduced reliable increases in the intracellular sodium con-centration in astrocytes indicating activation of sodium-dependent glutamate uptake (119899 = 488 Figure 6(c)) Theexperiments were performed in the presence of the sodium-channel blocker TTX (tetrodotoxin 05 120583M) to suppressaction potential generation The amplitude of D-aspartate-induced sodium increases was dependent on the concen-tration applied (Figure 6(c)) and saturated at about 2mM(119899 = 19 Figure 6(d)) as reported before for mouse corticalastrocytes in culture [50] At a concentration of 1mM theamplitude of D-aspartate-induced sodium transients wasnear maximum and amounted to 38 plusmn 02mM (119899 =161 Figures 6(c) and 6(d)) a value comparable to that inastrocytes in acute slices obtained in our lab [33]
To probe for changes in glutamate uptake we comparedsodium signals induced by 1mMD-aspartate between controland lesioned slices (Figure 7) In lesioned slices astrocyteslocated in the periphery of the lesion (100ndash350 120583m distance)responded with a sodium transient to application of 1mMD-aspartate (see cell 2 in Figures 7(a) and 7(b)) The amplitudeof D-aspartate-induced sodium signals in cells in the lesionperiphery was similar to that observed in control slices (38 plusmn03mM 119899 = 111 see cell 1 in Figures 7(a) 7(b) and 7(c))Application of D-aspartate also evoked sodium transientsin weakly SR101-positive SBFI-filled astrocytes located inthe glial scar area (lt100 120583m from the lesion see cells 3ndash5in Figures 7(a) and 7(b)) These cells however displayedsignificantly smaller peak amplitudes (16 plusmn 021mM 119899 =26 119875 lt 0001 Figures 7(b) and 7(c)) than astrocytes inthe periphery and in control slices
The kinetics of D-aspartate-induced sodium transientsdiffered between all three groups This was true for theslope of the sodium increase (10ndash90 change) which wassignificantly reduced in astrocytes from lesioned slices versusastrocytes from control slices (09 plusmn 005mMs in controlversus 06 plusmn 006mMs in the periphery and 02 plusmn 004mMsin the scar region 119899 = 161 111 and 26 resp 119875 lt 0001Figures 7(b) and 7(c)) In addition the decay back to baselinewas significantly slower following lesion (Figures 7(b) and7(c)) The time needed for the signal to decay by 67 wasapproximately doubled (decay time 120591 was 44 plusmn 2 s in control120591 = 81 plusmn 4 s in periphery of the lesion and 120591 = 95 plusmn 10 s inscar cells Figures 7(b) and 7(c))
These results show that mechanical injury results indistinct alterations in the kinetics andor amplitudes ofD-aspartate-induced intracellular sodium transients in reac-tive astrocytes depending on their distance to the lesion
4 Discussion
Our results show that within 6-7 days after setting a scratchwound in organotypic hippocampal slices a dense glial scarhad formed along the lesion site Up to approximately 100 120583mdistance from the lesion the scar tissue was characterized bya substantial elevation ofGFAP expression a pronounced cel-lular hypertrophy and reduced uptake of SR101 by astrocytes
10 ISRN Neurology
Con
trol
GLAST
Lesio
ned
GFAP Merge
StrR
StrP
StrO
Lesio
n sit
e
StrR
(a)
Con
trol
Lesio
ned
GLT-1 GFAP Merge
StrR
StrP
StrO
StrR
StrP
StrLM
Lesio
n sit
e
(b)
Figure 5 GLAST andGLT-1 immunoreactivity aftermechanical injury (a) Fluorescence images of immunohistochemical stainings detectingthe glutamate transporters GLAST (left) GFAP (center) as counterstaining and the corresponding merged image (GFAP green GLASTmagenta) in control (top row) and lesioned slice cultures (bottom row) (b) GLT-1 (left) GFAP (center) double labeling of a control (toprow) and a mechanically lesioned slice (bottom row) The corresponding merge (GFAP green GLT-1 magenta) is found on the right Notethat after lesion glutamate transporter immunoreactive clusters are accumulated in the direct vicinity of the lesion site (arrowheads) GFAPglial fibrillary acidic protein GLAST glutamate aspartate transporter GLT-1 glutamate-transporter-1 StrO stratum oriens StrP stratumpyramidale StrR stratum radiatum StrLM stratum lacunosum-moleculare and scale bars (a) 20 120583m (b) 40 120583m
While prominent clusters of GLAST and GLT-1 immunore-activity were observed alongGFAP-positive structures in scarcells D-aspartate-induced intracellular sodium signals werestrongly dampened indicating a significant reduction in glu-tamate uptake capacity close to the lesion In the periphery ofthe lesion (100ndash350120583m) astrocytes showed less pronounced
reactivity maintained their ability to efficiently take up SR101and displayed only minor changes in glutamate transporterimmunoreactivity and function Thus depending on theirdistance from the lesion astrocytes showed different gradesof reactivity and displayed discrete changes in glutamatetransporter immunoreactivity and uptake capacity
ISRN Neurology 11
SR101 MergeSBFI
StrRStrP
(a)
03
04
05
06
07
0 5 10 15
0
20 25 30 40 0
100
20
40
80
60
0 10 20 30 400
20
40
60
80
100
10 = 55 mM
107 cells in 6 experiments
F (340
380
) (ra
tio)
Intracellular Na+ concentration (mM)Intracellular Na+ concentration (mM)
ΔF
(340
380
) (
)
ΔF
(340
380
) (
) R2 = 097
(b)
D-aspartate
100 120583M500 120583M
750 120583M1000 120583M
1 mM
10 s
Δ
(c)
0 1 2 3 4 50
1
2
3
4
5
Δ so
dium
(mM
)
D-aspartate (mM)
lowast
(d)
Figure 6 Sodium changes induced by activation of glutamate uptake (a) Staining of astrocytes by colabeling with SR101 and SBFI Leftepifluorescence image of theCA1-area of an SR101-loaded organotypic sliceCenter SBFI fluorescence taken from the same areaRight overlaySBFI-loaded SR101-positive astrocytes appear in white (arrowheads) StrP stratum pyramidale StrR stratum radiatum scale bar 20 120583m (b)Left calibration of SBFI fluorescence in an astrocyte Depicted is the change in intracellular SBFI fluorescence ratio in the calibration solutionscontaining ionophores and in response to stepwise changes in the sodium concentration Right mean values plusmn SEM of the normalizedchanges in SBFI fluorescence at different sodium concentrations The black line represents a linear fit of the data points between 10 and40mM sodium illustrating that a 10 change in SBFI fluorescence represents a change in the sodium concentration by 55mM (c) Sodiumtransients in a single astrocyte induced by 500ms applications of D-aspartate at different concentrations (d) Mean values plusmn S E M (119899 = 19)of the peak amplitude of sodium transients at different concentrations of D-aspartate The curve represents a sigmoidal fit of the data
41 Characteristics of Lesion-Induced Reactive Gliosis inOrganotypic Cultures Organotypic tissue slice cultures ofthe hippocampus represent a well-established model systemin which the basic tissue architecture is maintained butwhich still hosts the major advantages of cell cultures suchas good accessibility and control of experimental conditionsThey have been used extensively for the analysis of neuronal
properties and developing neuronal networks [28 51 52]or the study of excitotoxic neuronal damage (eg [53 54])Recent studies also demonstrated that basic morphologicalcharacteristics of astrocytes and the typical glia-synapseorganization are well preserved in organotypic slice cultures(eg [44 45 55 56]) In addition we found a nearly completeoverlap of SR101 staining with the expression of GFP in slice
12 ISRN Neurology
SBFIControl Lesioned
SR101
1
Lesio
n sit
e
SR101 SBFI
2 45
32 4
5
1
StrR
StrP StrP
StrR
3
(a)
ControlScar
Lesioned
1
D-asp
2
D-asp
D-asp
5
4
3Periphery1mM
20 s
Δ Δ
Δ
Δ
Δ
(b)
0102030405060708090
100110
Δ so
dium
tim
e (m
Ms
)
00
02
04
06
08
10
12
0
1
2
3
4
5
6
ns
Control Periphery Scar Control Periphery Scar Control Periphery Scar
Amplitude Slope Decay time
Δ so
dium
(mM
)
120591(s
)lowastlowastlowast
lowastlowastlowastlowastlowastlowast
lowastlowastlowastlowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowast
(c)
Figure 7 D-aspartate-induced astrocyte sodium transients following lesion (a) CA1 area of a control (left) and a lesioned (right) slicepreparation Shown are epifluorescence images of SR101 and SBFI fluorescences Circled areas and numbers indicate the regions of interestanalyzed in the experiment depicted in (b) StrP stratum pyramidale StrR stratum radiatum and scale bar 40 120583m (b) Sodium transientsinduced by pressure application of 1mMD-aspartate (D-asp) for 500ms in the different cells in the control and lesioned slice as indicated in(a) (c) Histograms showing mean values plusmn S E M of the peak amplitude (left) slope (center) and decay time (right) of D-aspartate-inducedsodium transients in astrocytes in control slices (119899 = 161) as well as in cells in the scar region and its periphery (119899 = 111 and 119899 = 26 resp)lowastlowast119875 lt 001 lowastlowastlowast119875 lt 0001 StrP stratum pyramidale StrR stratum radiatum and scale bar 40120583m
cultures obtained from GFPGFAP mice demonstrating thatSR101 is well suited to identify astrocytes in this prepara-tion as described before for acute tissue slices and in vivo[29 30]
Performing a scratch through the CA1 region of organ-otypic slices resulted in the formation of a glial scar along thelesion site as judged by the prominent increase in expressionof GFAP and S100120573 In addition astrocyte cell bodies showed
a significant hypertrophy Furthermore astrocytes in theperiphery of the scar (100ndash350 120583m from the lesion) showedmild to moderate reactivity displaying swollen cell bodiesand only moderate upregulation of GFAP These changesare hallmarks of reactive gliosis observed after tissue injury[10 13 57] Generally the degree of injury and the distance ofthe astrocytes from the site of injury define the degree of theiractivation [11 13]
ISRN Neurology 13
Our study also revealed that strongly reactive astrocytesin the scar region only weakly stain with SR101 The reducedability of strongly activated astrocytes to accumulate SR101indicates that strong glial activation is accompanied by adownregulation of the organic anion transporter responsi-ble for uptake of SR101 This is reminiscent of immatureastrocytes in the early postnatal hippocampus which areSR101-negative [30] and in line with a wealth of experimentalevidence suggesting that reactive gliosis represents a processcomprising a dedifferentiation of astrocytes [17]
42 Changes in Glutamate Transporter Expression followingLesion Glutamate uptake in the hippocampus is mainlyachieved by the glial glutamate transporters GLT-1 andGLAST [1 2] A central factor in many brain pathologiesincluding traumatic brain injury is an increase in extracellu-lar glutamate and excitotoxicity it has been suggested that achange in the expression levels of glial glutamate transportersmight play a critical role in the failure of glutamate clearance[6 58ndash60] Earlier work has found an overall downregulationof protein levels of both GLAST and GLT-1 following astro-cyte activation [22ndash27] indicating that this is causal to theelevation of extracellular glutamate In contrast to this notionother studies reported increased glutamate transport capacityof reactive astrocytes and suggested a protective influence[61 62]
To visualize possible differences in the spatial expressionprofile of GLT-1 and GLAST in strongly reactive astrocytesalong the scar as compared to moderately reactive astro-cytes in its periphery we performed immunohistochemicalstainings These revealed an accumulation of GLT-1 andGLAST immunoreactivity along thick GFAP-positive fiberswhich was particularly pronounced for GLAST Besides theseclusters overall GLAST immunoreactivity seemed unalteredas compared to unlesioned slices whereas GLT-1 immunore-activity seemedweaker and declined with increasing distancefrom the scar Clustering of glutamate transporters has beendescribed by several reports [48 63 64] In developinghippocampal astrocytes cluster formation was preferentiallyfound in branches opposed to synapses and was increasedwith increased neuronal activity indicating that it is neces-sary to copewith synaptic release of glutamate [36] Along thesame lines it was observed that sustained astroglial activationby ciliary neurotrophic factor (CNTF) in the rat striatuminduced a concentration of GLAST and GLT-1 into raftmicrodomains and improved glutamate clearance indicatingthat cluster formation increased the cellular capacity for glu-tamate uptake [61] However another study found glutamate-induced clustering of GLT-1 that induced its endocytosis andintracellular trafficking without changing the total expressionlevels as detected by western blots arguing for a decrease infunctional glial glutamate uptake capacity [49] In the presentstudy antibodies against GLT-1 and GLAST were employedfor immunohistochemistry after permeabilization of theplasma membrane The clusters of GLAST and GLT-1 alongGFAP-positive fibers in the scar region (as GLAST andGLT-1immunoreactivity in general) could thus represent glutamatetransporters in the plasmamembrane or in intracellular com-partments or both Consequently the observed clustering
does not allow a prediction about possible functional changesin glutamate uptake
43 Lesion-Induced Changes in Functional Glutamate UptakeCapacity We probed for the functional activation of gluta-mate transport by application of the transportable agonistD-aspartate which results in an increase in the intracellularsodium concentration of astrocytes [8 9] The amplitudesof D-aspartate-induced sodium transients in astrocytes incontrol organotypic slices were similar to those reportedfrom astrocytes in acute slices obtained in our lab [33]indicating similar cellular glutamate transport capacity inboth preparations
Following a lesion D-aspartate-induced intracellularsodium signalswere clearly dampened in scar cellsMoreoverthe slope of the increase in sodium was significantly reducedand the decay back to baseline was slowed Because D-aspartate was applied at nearly saturating concentrationthe reduction in peak amplitudes suggests a reduction infunctional glutamate uptake capacity in cells close to thelesion which might be mediated by a decrease in the overallnumber of glutamate transporters available at the plasmamembrane In contrast the alteration in the kinetics of theD-aspartate-induced sodium signals can be explained by thecellular hypertrophy that was observed in reactive astrocytesAt an equal transport-mediated sodium influx across themembrane the slope of resulting changes in the sodiumconcentration will be decreased in cells with a larger volume
Furthermore the decay of intracellular sodium transientsback to baseline is mainly governed by the activity of theNa+K+-ATPase as well as by diffusion [8] The slower recov-ery is in line with earlier studies reporting a downregulationof the sodium pump following reactive gliosis [65 66]Moreover sodium is not buffered in the cell and travels solelyby means of diffusion in the cytoplasm Hindered diffusionresulting from increased cytoplasmic protein content orimpaired gap junction coupling which both occur uponastrocytic activation might thus also partially contribute todelayed sodium recovery [12 34 67]
In contrast to scar cells the amplitude of D-aspartate-induced intracellular sodium signals was not significantlyaltered in moderately activated astrocytes in the periphery ofthe lesion indicating that the overall number of functionalglutamate transporters was not significantly altered Becausethese cells showed hypertrophy as well the slowed kinetics ofthe sodium transients are in line with an increased cellularvolume as well as a possible downregulation of Na+K+-ATPase andor a slowed diffusion as argued above
Taken together our results indicate a significant reduc-tion in glutamate uptake capacity in strongly activated astro-cytes in the scar region Thus the prominent clustering ofGLAST and GLT-1 immunoreactivity along GFAP-positivestructures close to the lesion is likely to reflect a loss ofglutamate transporters from the plasma membrane upontransporter internalization as reported by Nakagawa et al[49] The authors of the latter study speculated that large andprolonged increases in glutamate concentrations are neces-sary to induce such cluster formation and endocytosis in thetissue as they would possibly only occur under pathological
14 ISRN Neurology
conditions Indeed ambient glutamate concentrations wereshown to be significantly increased after traumatic braininjury [68ndash70] Along these lines it can be assumed thatin our model system extracellular glutamate concentrationsrose less severely in the periphery of the lesion preventinga comparable clustering and loss of functional glutamatetransport activity in moderately activated astrocytes
5 Conclusions
Our results show complex changes in glutamate transporterexpression and function during astrocyte activation followingmechanical injury They confirm that the degree of astrocyteactivation depends on the distance from the insult and com-prises different endpoints regarding cellular morphology andphysiology Furthermore our results highlight that astrocyteswhich show different grades of reactivity also display discretechanges in glutamate transporter expression and functionWhile immunohistochemistry revealed a prominent clus-tering of GLT-1 and GLAST immunoreactivity in the scarregion our functional assay clearly showed that glutamateuptake capacity is strongly reduced in scar cells while itis largely maintained in moderately activated astrocytes inthe periphery Thus mild-to-moderate astrogliosis in theperiphery of a mechanical lesion does not necessarily seemto be accompanied by a significant change in glial glutamateuptake capacity At the glial scar itself in strongly reactiveastrocytes a clustering of glutamate transporters is observedthat apparently goes along with a severe functional reductionin astroglial glutamate uptake which may contribute toglutamate-mediated excitotoxicity in this region
Conflict of Interests
The authors of the paper declare that they do not haveany direct financial relation with the commercial identitiesmentioned in the paper that might lead to a conflict ofinterests
Acknowledgments
This study was supported by the Deutsche Forschungsge-meinschaft (DFG Ro23274-3) The authors thank SimoneDurry and Claudia Roderigo for expert technical help
References
[1] N C Danbolt ldquoGlutamate uptakerdquo Progress in Neurobiologyvol 65 no 1 pp 1ndash105 2001
[2] A V Tzingounis and J I Wadiche ldquoGlutamate transportersconfining runaway excitation by shaping synaptic transmis-sionrdquo Nature Reviews Neuroscience vol 8 no 12 pp 935ndash9472007
[3] D E Bergles J S Diamond and C E Jahr ldquoClearance ofglutamate inside the synapse and beyondrdquo Current Opinion inNeurobiology vol 9 no 3 pp 293ndash298 1999
[4] C M Anderson and R A Swanson ldquoAstrocyte glutamatetransport review of properties regulation and physiologicalfunctionsrdquo Glia vol 32 no 1 pp 1ndash14 2000
[5] P Marcaggi and D Attwell ldquoRole of glial amino acid trans-porters in synaptic transmission and brain energeticsrdquoGlia vol47 no 3 pp 217ndash225 2004
[6] N J Maragakis and J D Rothstein ldquoGlutamate transportersanimal models to neurologic diseaserdquo Neurobiology of Diseasevol 15 no 3 pp 461ndash473 2004
[7] G Gegelashvili and A Schousboe ldquoCellular distribution andkinetic properties of high-affinity glutamate transportersrdquoBrainResearch Bulletin vol 45 no 3 pp 233ndash238 1998
[8] J W Deitmer and C R Rose ldquoIon changes and signalling inperisynaptic gliardquo Brain Research Reviews vol 63 no 1-2 pp113ndash129 2010
[9] S Kirischuk V Parpura and A Verkhratsky ldquoSodium dynam-ics another key to astroglial excitabilityrdquo Trends in Neuro-sciences vol 35 no 8 pp 497ndash506 2012
[10] M V Sofroniew ldquoMolecular dissection of reactive astrogliosisand glial scar formationrdquo Trends in Neurosciences vol 32 no12 pp 638ndash647 2009
[11] J L Ridet S K Malhotra A Privat and F H Gage ldquoReactiveastrocytes cellular and molecular cues to biological functionrdquoTrends in Neurosciences vol 20 no 12 pp 570ndash577 1997
[12] J Middeldorp and E M Hol ldquoGFAP in health and diseaserdquoProgress in Neurobiology vol 93 no 3 pp 421ndash443 2011
[13] M Pekny and M Nilsson ldquoAstrocyte activation and reactivegliosisrdquo Glia vol 50 no 4 pp 427ndash434 2005
[14] A Bordey S A Lyons J J Hablitz and H SontheimerldquoElectrophysiological characteristics of reactive astrocytes inexperimental cortical dysplasiardquo Journal of Neurophysiologyvol 85 no 4 pp 1719ndash1731 2001
[15] R Jabs G Seifert and C Steinhauser ldquoAstrocytic functionand its alteration in the epileptic brainrdquo Epilepsia vol 49supplement 2 pp 3ndash12 2008
[16] A Buffo I Rite P Tripathi et al ldquoOrigin and progeny ofreactive gliosis a source of multipotent cells in the injuredbrainrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 105 no 9 pp 3581ndash3586 2008
[17] S Robel B Berninger and M Gotz ldquoThe stem cell potential ofglia lessons from reactive gliosisrdquoNature Reviews Neurosciencevol 12 no 2 pp 88ndash104 2011
[18] H Kawano J Kimura-Kuroda Y Komuta et al ldquoRole of thelesion scar in the response to damage and repair of the centralnervous systemrdquo Cell and Tissue Research vol 349 no 1 pp169ndash180 2012
[19] G Yiu and Z He ldquoGlial inhibition of CNS axon regenerationrdquoNature Reviews Neuroscience vol 7 no 8 pp 617ndash627 2006
[20] M T Fitch and J Silver ldquoCNS injury glial scars and inflamma-tion inhibitory extracellular matrices and regeneration failurerdquoExperimental Neurology vol 209 no 2 pp 294ndash301 2008
[21] L Li A Lundkvist D Andersson et al ldquoProtective role ofreactive astrocytes in brain ischemiardquo Journal of Cerebral BloodFlow amp Metabolism vol 28 no 3 pp 468ndash481 2008
[22] F K H van Landeghem T Weiss M Oehmichen and AVonDeimling ldquoDecreased expression of glutamate transportersin astrocytes after human traumatic brain injuryrdquo Journal ofNeurotrauma vol 23 no 10 pp 1518ndash1528 2006
[23] V L Raghavendra Rao M K Baskaya A Dogan J DRothstein and R J Dempsey ldquoTraumatic brain injury down-regulates glial glutamate transporter (GLT- 1 and GLAST)proteins in rat brainrdquo Journal of Neurochemistry vol 70 no 5pp 2020ndash2027 1998
ISRN Neurology 15
[24] J-C Chen H Hsu-Chou J-L Lu et al ldquoDown-regulation ofthe glial glutamate transporter GLT-1 in rat hippocampus andstriatum and its modulation by a group III metabotropic glu-tamate receptor antagonist following transient global forebrainischemiardquoNeuropharmacology vol 49 no 5 pp 703ndash714 2005
[25] S D Ginsberg L J Martin and J D Rothstein ldquoRegional deaf-ferentation down-regulates subtypes of glutamate transporterproteinsrdquo Journal of Neurochemistry vol 65 no 6 pp 2800ndash2803 1995
[26] J E Springer R D Azbill R J Mark J G Begley GWaeg andM P Mattson ldquo4-Hydroxynonenal a lipid peroxidation prod-uct rapidly accumulates following traumatic spinal cord injuryand inhibits glutamate uptakerdquo Journal of Neurochemistry vol68 no 6 pp 2469ndash2476 1997
[27] M B Moretto N S Arteni D Lavinsky et al ldquoHypoxic-ischemic insult decreases glutamate uptake by hippocampalslices from neonatal rats Prevention by guanosinerdquo Experimen-tal Neurology vol 195 no 2 pp 400ndash406 2005
[28] L Stoppini P-A Buchs and D Muller ldquoA simple method fororganotypic cultures of nervous tissuerdquo Journal of NeuroscienceMethods vol 37 no 2 pp 173ndash182 1991
[29] A Nimmerjahn F Kirchhoff J N D Kerr and F HelmchenldquoSulforhodamine 101 as a specific marker of astroglia in theneocortex in vivordquo Nat Methods vol 1 no 1 pp 31ndash37 2004
[30] K W Kafitz S D Meier J Stephan and C R Rose ldquoDevel-opmental profile and properties of sulforhodamine 101-Labeledglial cells in acute brain slices of rat hippocampusrdquo Journal ofNeuroscience Methods vol 169 no 1 pp 84ndash92 2008
[31] C Schnell Y Hagos and S Hulsmann ldquoActive sulforhodamine101 uptake into hippocampal astrocytesrdquo PLoS One vol 7 no 11Article ID e49398 2012
[32] S D Meier Y Kovalchuk and C R Rose ldquoProperties ofthe new fluorescent Na+ indicator CoroNa Green comparisonwith SBFI and confocal Na+ imagingrdquo Journal of NeuroscienceMethods vol 155 no 2 pp 251ndash259 2006
[33] J Langer and C R Rose ldquoSynaptically induced sodium signalsin hippocampal astrocytes in siturdquo Journal of Physiology vol587 part 24 pp 5859ndash5877 2009
[34] J Langer J Stephan M Theis and C R Rose ldquoGap junctionsmediate intercellular spread of sodium between hippocampalastrocytes in siturdquo Glia vol 60 no 2 pp 239ndash252 2012
[35] C R Rose andB R Ransom ldquoIntracellular sodiumhomeostasisin rat hippocampal astrocytesrdquo Journal of Physiology vol 491part 2 no 2 pp 291ndash305 1996
[36] A M Benediktsson G S Marrs J C Tu et al ldquoNeuronalactivity regulates glutamate transporter dynamics in developingastrocytesrdquo Glia vol 60 no 2 pp 175ndash188 2012
[37] M Zhou G P Schools and H K Kimelberg ldquoDevelopment ofGLAST(+) astrocytes andNG2(+) glia in rat hippocampus CA1mature astrocytes are electrophysiologically passiverdquo Journal ofNeurophysiology vol 95 no 1 pp 134ndash143 2006
[38] I Suarez G Bodega and B Fernandez ldquoModulation of glu-tamate transporters (GLAST GLT-1 and EAAC1) in the ratcerebellum following portocaval anastomosisrdquo Brain Researchvol 859 no 2 pp 293ndash302 2000
[39] X Zhu D E Bergles and A Nishiyama ldquoNG2 cells generateboth oligodendrocytes and gray matter astrocytesrdquo Develop-ment vol 135 no 1 pp 145ndash157 2008
[40] B Brunne S ZhaoADerouiche et al ldquoOriginmaturation andastroglial transformation of secondary radial glial cells in thedeveloping dentate gyrusrdquo Glia vol 58 no 13 pp 1553ndash15692010
[41] Y Wu A-Q Zhang and D T Yew ldquoAge related changes ofvarious markers of astrocytes in senescence-accelerated micehippocampusrdquo Neurochemistry International vol 46 no 7 pp565ndash574 2005
[42] S Magavi D Friedmann G Banks A Stolfi and C LoisldquoCoincident generation of pyramidal neurons and protoplasmicastrocytes in neocortical columnsrdquo Journal of Neuroscience vol32 no 14 pp 4762ndash4772 2012
[43] M Olabarria H N Noristani A Verkhratsky and J JRodrıguez ldquoConcomitant astroglial atrophy and astrogliosis ina triple transgenic animal model of Alzheimerrsquos diseaserdquo Gliavol 58 no 7 pp 831ndash838 2010
[44] A M Benediktsson S J Schachtele S H Green and M EDailey ldquoBallistic labeling and dynamic imaging of astrocytesin organotypic hippocampal slice culturesrdquo Journal of Neuro-science Methods vol 141 no 1 pp 41ndash53 2005
[45] I Lushnikova G Skibo D Muller and I Nikonenko ldquoSynapticpotentiation induces increased glial coverage of excitatorysynapses in CA1 hippocampusrdquoHippocampus vol 19 no 8 pp753ndash762 2009
[46] E Raponi F Agenes C Delphin et al ldquoS100B expressiondefines a state in which GFAP-expressing cells lose their neuralstem cell potential and acquire a more mature developmentalstagerdquo Glia vol 55 no 2 pp 165ndash177 2007
[47] C G Schipke C Boucsein C Ohlemeyer F Kirchhoff andH Kettenmann ldquoAstrocyte Ca2+ waves trigger responses inmicroglial cells in brain slicesrdquo The FASEB Journal vol 16 no2 pp 255ndash257 2002
[48] J Zhou and M L Sutherland ldquoGlutamate transporter clusterformation in astrocytic processes regulates glutamate uptakeactivityrdquo Journal of Neuroscience vol 24 no 28 pp 6301ndash63062004
[49] T Nakagawa Y Otsubo Y Yatani H Shirakawa and S KanekoldquoMechanisms of substrate transport-induced clustering of aglial glutamate transporter GLT-1 in astroglial-neuronal cul-turesrdquo European Journal of Neuroscience vol 28 no 9 pp 1719ndash1730 2008
[50] J-Y Chatton P Marquet and P J Magistretti ldquoA quantitativeanalysis of L-glutamate-regulated Na+ dynamics in mousecortical astrocytes Implications for cellular bioenergeticsrdquoEuropean Journal of Neuroscience vol 12 no 11 pp 3843ndash38532000
[51] E Forster S Zhao and M Frotscher ldquoLaminating the hip-pocampusrdquo Nature Reviews Neuroscience vol 7 no 4 pp 259ndash267 2006
[52] B H Gahwiler M Capogna D Debanne R A McKinney andS M Thompson ldquoOrganotypic slice cultures a technique hascome of agerdquo Trends in Neurosciences vol 20 no 10 pp 471ndash477 1997
[53] P E Kunkler and R P Kraig ldquoReactive astrocytosis fromexcitotoxic injury in hippocampal organ culture parallels thatseen in vivordquo Journal of Cerebral Blood Flow and Metabolismvol 17 no 1 pp 26ndash43 1997
[54] I E Holopainen ldquoOrganotypic hippocampal slice cultures amodel system to study basic cellular andmolecularmechanismsof neuronal cell death neuroprotection and synaptic plasticityrdquoNeurochemical Research vol 30 no 12 pp 1521ndash1528 2005
[55] MHaber L Zhou andK KMurai ldquoCooperative astrocyte anddendritic spine dynamics at hippocampal excitatory synapsesrdquoJournal of Neuroscience vol 26 no 35 pp 8881ndash8891 2006
16 ISRN Neurology
[56] H Nishida and S Okabe ldquoDirect astrocytic contacts regulatelocal maturation of dendritic spinesrdquo Journal of Neurosciencevol 27 no 2 pp 331ndash340 2007
[57] A Buffo C Rolando and S Ceruti ldquoAstrocytes in the dam-aged brain molecular and cellular insights into their reactiveresponse and healing potentialrdquoBiochemical Pharmacology vol79 no 2 pp 77ndash89 2010
[58] J-H Yi and A S Hazell ldquoExcitotoxic mechanisms and the roleof astrocytic glutamate transporters in traumatic brain injuryrdquoNeurochemistry International vol 48 no 5 pp 394ndash403 2006
[59] C Werner and K Engelhard ldquoPathophysiology of traumaticbrain injuryrdquo British Journal of Anaesthesia vol 99 no 1 pp4ndash9 2007
[60] G Gegelashvili M B Robinson D Trotti and T RauenldquoRegulation of glutamate transporters in health and diseaserdquoProgress in Brain Research vol 132 pp 267ndash286 2001
[61] C Escartin E Brouillet PGubellini et al ldquoCiliary neurotrophicfactor activates astrocytes redistributes their glutamate trans-porters GLAST and GLT-1 to raft microdomains and improvesglutamate handling in vivordquo Journal of Neuroscience vol 26 no22 pp 5978ndash5989 2006
[62] C Vermeiren M Najimi N Vanhoutte et al ldquoAcute up-regulation of glutamate uptake mediated by mGluR5a in reac-tive astrocytesrdquo Journal of Neurochemistry vol 94 no 2 pp405ndash416 2005
[63] C L Poitry-Yamate L Vutskits and T Rauen ldquoNeuronal-induced and glutamate-dependent activation of glial glutamatetransporter functionrdquo Journal of Neurochemistry vol 82 no 4pp 987ndash997 2002
[64] P M Beart and R D OrsquoShea ldquoTransporters for L-glutamatean update on their molecular pharmacology and pathologicalinvolvementrdquo British Journal of Pharmacology vol 150 no 1pp 5ndash17 2007
[65] F D Lima M A Souza A F Furian et al ldquoNa+K+-ATPaseactivity impairment after experimental traumatic brain injuryrelationship to spatial learning deficits and oxidative stressrdquoBehavioural Brain Research vol 193 no 2 pp 306ndash310 2008
[66] G A Gusarova H E Trejo L A Dada et al ldquoHypoxia leadsto NaK-ATPase downregulation via Ca2+ release-activatedCa2+ channels and AMPK activationrdquo Molecular and CellularBiology vol 31 no 17 pp 3546ndash3556 2011
[67] S S L Chew C S Johnson C R Green and H V Danesh-Meyer ldquoRole of connexin43 in central nervous system injuryrdquoExperimental Neurology vol 225 no 2 pp 250ndash261 2010
[68] P Nilsson L Hillered U Ponten and U Ungerstedt ldquoChangesin cortical extracellular levels of energy-related metabolites andamino acids following concussive brain injury in ratsrdquo Journal ofCerebral Blood Flow and Metabolism vol 10 no 5 pp 631ndash6371990
[69] H Katoh K Sima H Nawashiro K Wada and H ChigasakildquoThe effect of MK-801 on extracellular neuroactive amino acidsin hippocampus after closed head injury followed by hypoxia inratsrdquo Brain Research vol 758 no 1-2 pp 153ndash162 1997
[70] H Koizumi H Fujisawa H Ito T Maekawa X Di and RBullock ldquoEffects of mild hypothermia on cerebral blood flow-independent changes in cortical extracellular levels of aminoacids following contusion trauma in the ratrdquoBrain Research vol747 no 2 pp 304ndash312 1997
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

ISRN Neurology 3
[29 30] It has recently been shown to be taken up intomaturehippocampal astrocytes via an active transport mechanisminvolving organic anion transport polypeptides [31] Wide-field fluorescence imaging was performed using a variablescan digital imaging system (TILL Photonics MartinsriedGermany) attached to an upright microscope (BX51WiOlympus Europe Hamburg Germany) and a CCD camera(TILL Imago VGA Till Photonics Martinsried Germany)Imageswere collectedwith anAchroplan 40x objective (waterimmersion NA 08 Zeiss Gottingen Germany) SR101 wasexcited at 575 nm and emission was collected above 590 nmExcitation wavelength for detection of GFP was 488 nm andemission was collected above 510 nm
For intracellular sodium imaging slices were addition-ally incubated with the membrane permeable form of thesodium-sensitive fluorescent dye SBFI (SBFI-AM sodium-binding benzofuran isophthalate-acetoxymethyl ester Mol-ecular ProbesLife Technologies Darmstadt Germany) asdescribed earlier [32ndash34] Ratiometric sodium imaging wasperformed by alternate excitation of SBFI at 340 nm (weaklysodium-sensitive wavelength) and at 380 nm (sodium-sensitive wavelength) at 4Hz Emission (gt440 nm) wascollected in defined regions of interest (ROI) representingcell bodies Standard dynamic background correction wasperformed as described earlier [33 34] After backgroundcorrection the fluorescence ratio (F
340F380
) was calculatedfor the individual ROIs and analyzed offline using OriginPro8G Software (OriginLab Corporation Northampton MAUSA)
Changes in SBFI fluorescence ratio were expressed aschanges in sodium concentration based on in situ calibrationsas reported before [32 33 35] To equilibrate extra- andintracellular Na+-concentrations SBFI-loaded slices wereperfused with saline containing ionophores (3 120583M grami-cidin D 100 120583Mmonensin) and the Na+K+-ATPase blockerouabain (100 120583M) as well as different concentrations of Na+
The glutamate transporter agonist D-aspartate wasapplied by a pressure application device (PDES-02D NPIElectronic GmbH Tamm Germany) coupled to standardmicropipettes (Hilgenberg Waldkappel Germany) placed20ndash100120583m from cell bodies of selected cells
24 Antibodies Antibodies employed in this study are listedin Table 1 The following antisera were utilized whichrepresent well-established commercially available standardmarkers guinea-pig GLT-1 antiserum directed against theC-terminus of rat GLT-1 (Chemicon InternationalMilliporeCorp Carrigtwohill Ireland eg [36]) and guinea-pigGLAST antiserum directed against the C-terminus of ratGLAST (Chemicon InternationalMillipore Corp Carrigt-wohill Ireland eg [37]) Validation and a detailed charac-terization of both antibodies were provided before [38ndash40]For identification of astrocytes polyclonal rabbit antibodiesagainst GFAP (GFAP-pAb Dako Cytomation Denmark[40 41]) and S100120573 (Abcam United Kingdom [42]) wereemployedWhen double staining with the latter antibody wasperformed a monoclonal mouse antibody detecting GFAPwas used (GFAP-mAb Sigma Aldrich Germany [40 43])
Standard fluorochrome-conjugated antibodies (Alexa-Fluor InvitrogenLife Technologies Darmstadt Germany)were employed as secondary antibodies for immunohisto-chemistry
25 Immunohistochemistry Prior to immunohistochemicalprocessing organotypic slices were immersion fixed for30min at RT in 4 paraformaldehyde (PFA) in phosphate-buffered saline (PBS) following three washes every30min with PBS Cell membranes were permeabilizedand unspecific binding sites were blocked in PBS containing025 triton-X100 (TX) and 2 normal goat serum (NGSGIBCOLife Technologies Darmstadt Germany) for 90minat 4∘C followed by incubation with the primary antibodyGFAP-pAb (1 1000 Dako Cytomation Glostrup Denmark)diluted in the same solution over night at 4∘C In case ofS100120573GFAP double stainings slices were incubated witha mixture of the primary antibodies GFAP-mAb (1 1000Dako Cytomation Glostrup Denmark) and rabbit-S100120573(1 100 Abcam United Kingdom) After five washes in PBScontaining 025 TX and 2 NGS slices were incubatedeither with guinea pig-anti-GLAST or with guinea pig-anti-GLT-1 (both 1 1000 diluted in 025 TX2 NGSPBS) for4 hrs at RT Excess primary antibody was removed with fivewashes using 2 NGSPBS Antirabbit-AlexaFluor594 andantiguinea pig-AlexaFluor488 or antimouse-AlexaFluor488(1 100 in blocking solution) were used for visualizationof antibody binding and incubated for 2 hrs at RT Theslices were subjected to DAPI staining (410158406-diamidino-2-phenylindole 05120583m Invitrogen) washed three times andmounted on glass slides with mowiolDABCO (CalbiochemFluka distributed by Sigma-Aldrich Chemical MunichGermany)
Identical conditions were applied to all performed stain-ings regarding tissue processing and staining procedureNegative controls were run in parallel to each staining byeither omitting all or just one of the primary antibodiesControl stainings in which one of the primary antibodieswas omitted showed the identical labeling pattern for theremaining antibody as in the double stainings Omitting bothprimary antibodies never resulted in a staining
Documentation of immunofluorescence was either per-formed with an epifluorescence microscope (Nikon Eclipse90i Nikon Instruments Dusseldorf Germany) or a confocallaser scanningmicroscope (Olympus Fluoview300OlympusHamburg Germany) The epifluorescence microscope wasequipped with a standard DAPI (EX 340ndash380 DM 400BA 435ndash485) FITC (EX 465ndash495 DM 505 BA 515ndash555)and TRITC (EX 54025 DM 365 BA 60555) filter setIllumination was provided by an Intensilight fiber lamp(C-GHFI Nikon Instruments Dusseldorf Germany) andemission was detected with a monochrome digital cam-era (DS-Qi1Mc Nikon Instruments Dusseldorf Germany)Images were collected with either a 20x075 (PlanApoVCNikon Instruments Dusseldorf Germany) air objective ora 60x140 oil immersion objective (PlanApoVC NikonInstruments Dusseldorf Germany) NIS-Elements software(Nikon Instruments Dusseldorf Germany) was used forimage acquisition For confocal microscopy an Olympus
4 ISRN Neurology
Table 1 List of antibodies used
Antibody Structure of theimmunogen Company Order number Species Type Dilution
GLAST(EAAT1)
KPYQLIAQDNEPEKPVADSETKM ChemiconMillipore AB1782lowast Guinea
Pig IgGPolyclonal 1 1000
GLT-1(EAAT2)
AANGKSADCSVEEEPWKREK ChemiconMillipore AB1783 Guinea
Pig IgGPolyclonal 1 10001 2000
GFAP ns (full-length proteinfrom cow spinal cord) Dako Cytomation Z 0334 Rabbit
IgGPolyclonal 1 10001 2000
GFAP
clone G-A-5(LQSLTCDVESLRGTNESLERQMREQEERHAREAASYQEALTRLEEEGQSLKDEMARHLQEYQELLNVKLALDIEIATY)
Sigma-Aldrich G3893 MouseIgG
Monoclonal 1 10001 2000
S100szlig ns (recombinantfull-length cow protein) Abcam ab868 Rabbit
IgGPolyclonal 1 100
MouseIgG AF488 conjugated Invitrogen (Life Technologies) A11029 Goat mdash 1 100
RabbitIgG AF594 conjugated Invitrogen (Life Technologies) A11012 Goat mdash 1 100
Guineapig IgG AF488 conjugated Invitrogen (Life Technologies) A11073 Goat mdash 1 100
GLAST glutamate aspartate transporter GLT-1 glutamate transporter 1 GFAP glial fibrillary protein AF AlexaFluor appl application IHC immunohisto-chemistry WB western blot ns not specifiedlowastThemanufacturer has lately stopped the sale and distribution of the GLAST antibody used here
BX51WI microscope coupled to a confocal laser scanningsystem (FV300) equipped with a multiline argon (488 nm)and a helium-neon laser (543 nm both Melles Griot Ben-sheim Germany) was used Images were collected witheither a 20x050 (UMPlanFl Olympus Hamburg Ger-many) a 40x080 water immersion (LUMPlan OlympusHamburg Germany) or a 60x140 oil immersion objective(PlanApoVC Nikon Instruments Dusseldorf Germany) AKalman filter 4 was employed at every scan Simultaneousor sequential scanning of both fluorophores revealed nodifference in their staining patterns indicating absence ofcross-excitation or spectral bleed through The thickness ofz-plane sections was 1120583m and the number of optical sectionsvaried depending on the preparation
Figures show extended focus images as specified inthe figure legends which were calculated from z-stacksof optical sections using ImageJ software (NIH BethesdaUSA) Images were overlaid employing Adobe PhotoshopCS2 (Adobe Systems Cologne Germany)
26 Data Presentation and Statistics Unless otherwise spec-ified data are expressed as means plusmn SEM Data were statis-tically analyzed by Studentrsquos 119905-test employing the proceduresimplemented in OriginPro 8G Software (OriginLab Corpo-ration Northampton MA USA) A 119875 value of lt005 wasconsidered significant If not stated otherwise 119899representsthe number of analyzed cells and119873 the number of slices Eachset of experimentswas performedon at least three tissue slicesobtained from different animals
3 Results
31 Identification of Astrocytes in Organotypic Slice CulturesThe aim of the present study was to analyze changes inglutamate transporter expression and function in astrocytesin response to a mechanical injury To this end we preparedorganotypic tissue slice cultures of themouse hippocampus awell-establishedmodel system inwhich the layering and basiccellular organization of the neural network are maintainedover time [28] After 19ndash25 days in culture organotypic tissueslices had flattened to a thickness of 40ndash50 120583m and werecomposed of 3 to 4 cell layers At this stage propidium iodideassays detected only few dead cells dispersed throughoutthe slices (3 plusmn 06 cells per CA1 subfield 18 plusmn 34 in entirepreparation 119873 = 6 data not shown) indicating that thepreparation was viable and in a stable condition
To visualize astrocytes in cultured slices we used hip-pocampi of transgenic mice in which green fluorescentprotein (GFP) is expressed under the control of the promoterof the astrocyte-specific intermediate filament glial fibrillaryacidic protein (GFAP FVBN-Tg[GFAPGFP] ldquoGFPGFAPmicerdquo) A visualization of theGFP fluorescence demonstratedthat the general distribution and organization of astrocyteswere preserved in the strata pyramidale and radiatum ofthe CA1 region of organotypic cultures confirming earlierreports (119873 = 3 Figure 1(a) [44 45]) The vast majority ofGFP-expressing cells (sim95) also labeled with the astrocyte-specific vital marker SR101 in organotypic slices (119873 = 3Figures 1(a) and 1(b)) Furthermore immunohistochemicalstainings for GFAP in slice cultures derived from wildtype
ISRN Neurology 5
Slic
e cul
ture
(20
dic
)
GFP-GFAP MergeSR101
lowast lowast lowast lowast
lowast
lowast
lowast lowast
StrP
StrR
(a)
StrR
StrP
StrLM
StrM
SR101
StrO
(b)
GFAP S100120573
StrP
StrR
StrP
StrR
StrP
StrR
StrP
StrR
DAPI Merge
(c)
Figure 1 Astrocytes in organotypic hippocampal slice cultures (a) Organotypic slice cultures at 20 days in culture (dic) Preparations werederived from animals in which GFP was expressed under control of the GFAP promoter Shown is the expression of GFP (left) the labelingpattern of SR101 (center) and the corresponding merged image (GFAP green SR101 magenta) Asterisks point out SR101-labeled cells thatdo not express GFP arrowheads point out GFP-positive but SR101-negative cells Images represent maximum intensity projections of 15optical sections taken at 1120583m intervals (b) SR101 fluorescence of an organotypic slice at 21 dic Image represents a stack of 20 optical sectionstaken at 1 120583m intervals (c) Immunohistochemical double staining for GFAP and S100120573 as well as DAPI fluorescence and merged image ofthe fluorescence of GFAP (green) and S100120573 (magenta) of part of the CA1 region encompassing the strata pyramidale and radiatum of anorganotypic slice at 20 dic Note the radially oriented GFAP-positive fibers crossing the pyramidal cell layer Images were taken at a wide-field microscope ((a)ndash(c)) Dashed lines indicate the approximate boundaries of the stratum pyramidale StrO stratum oriens StrP stratumpyramidale StrR stratum radiatum StrLM stratum lacunosum moleculare and StrM stratum moleculare Scale bars (a) 50 120583m (b) 100120583mand (c) 50 120583m
animals showed that themajority of GFAP-positive cells werealso positive for S100120573 (119873 = 12 Figure 1(c)) a marker formature astrocytes [46]
Taken together the nearly complete overlap in the cellularstaining pattern for the vital dye SR101 with the expressionof GFP in GFPGFAP mice as well as the overlap in theimmunofluorescence for GFAP and S100120573 in organotypicslices indicates that these markers are well suited to identifyastrocytes in this preparation
32 Astrocyte Markers and Astrocyte Morphology followingMechanical Lesion To induce astrogliosis in organotypicslice cultures we used a scalpel blade and performed
a scratch under semisterile conditions through the entireCA1 region oriented perpendicular to the stratum pyramidale(Figure 2(a)) 6-7 days after the lesion a general enhance-ment of GFAP immunoreactivity was observed indicatingastrogliosis (Figure 2(b))The greatest enhancement of GFAPimmunoreactivity occurred within a distance of less than100 120583malong the lesion (119873 = 26 Figures 2(b) and 2(c)) Herea densemeshwork of thick GFAP-positive labels was presentindicative of the formation of a glial scar Furthermore theregion close to the lesion site exhibited a discernible increasein S100120573 immunoreactivity (119873 = 12 Figure 2(c)) Both theprominent expressions of GFAP and S100120573 are characteristicsof strongly reactive astrocytes which represent the major
6 ISRN Neurology
GFAP
DAPI
CA1 subregion
GFAP
DAPI
StrR
StrP StrP
StrR
StrOStrO
StrO
StrP
StrR
StrO
StrP
StrR
Control Lesioned
GFAP
Lesio
n sit
e
Hi
StrMStrG
StrP
StrR
StrLM
Scratch wound
StrO
Lesio
n sit
e
Lesioned
(a) (c)
(b)
350120583m350120583m
S100120573
Figure 2 Lesion-induced reactive gliosis in organotypic slice cultures (a) Schematic overview of the hippocampus indicating the positionof the scratch woundThe box delineates the area primarily analyzed (b) GFAP immunofluorescence and DAPI staining of the CA1 area in acontrol (left) and a lesioned (right) slice The boxes indicate the areas enlarged below The dashed line in the lower right indicates the lesion(c) Double staining for GFAP (top) and S100120573 (bottom) adjacent to the lesion site Note the gradual increase in GFAP and S100120573 expressiontowards the lesion (b) widefield microscopy (c) confocal microscopy (optical sections taken at 1120583m intervals) StrO stratum oriens StrPstratum pyramidale StrR stratum radiatum and StrLM stratum lacunosum moleculare Scale bars (b) 200 120583m (top) and 50 120583m (bottom) (c)10120583m
ISRN Neurology 7
StrR
StrP
StrLM
StrO
StrP StrRStrO StrLM
Lesio
n sit
e
SR101-staining pattern
(a)
SBFI
Lesio
n sit
e
SR101 GFAP-GFP
(b)
Figure 3 SR101 labeling in lesioned slices (a) SR101 fluorescence in the CA1 area in a slice 7 days after lesion The delineated area comprisesthe stratum pyramidale and the box indicates the area enlarged on the right side Right SR101 fluorescence at the lesion site illustrating thatSR101 labeling is reduced along the lesionThe dotted line indicates the transition between the scar area and the adjacent moderately reactivetissue The boxed area is further enlarged at the bottom (b) Image series obtained at a wide-field fluorescence microscope The slice wasprepared from a transgenic GFPGFAP mouse and the lesion site is indicated by the dotted line Left Image of SR101 fluorescence centerGFP fluorescence indicating GFAP expression and right SBFI fluorescence SR101 labeling is only weak in GFPSBFI-positive cells in the scarregion (see arrowheads) StrO stratum oriens StrP stratum pyramidale StrR stratum radiatum and StrLM stratum lacunosum-moleculareScale bars 100120583m ((a) upper left) and 20 120583m (other images)
cellular component of the glial scar forming after mechanicaltissue injury [10] More distal to the lesion (gt100ndash350 120583m) aless dramatic rearrangement of GFAP immunoreactivity wasobserved
A clear difference in astrocyte properties between thescar region and its periphery was also seen following SR101labeling In the periphery of the scar (100ndash350 120583m distancefrom the lesion) SR101 resulted in a reliable and brightcellular staining similar to what had been observed incontrol unlesioned slices (119873 = 12 Figure 3(a) cf Figure 1)
In contrast to this cellular SR101 staining was detectable butonly faint within a distance of less than 100 120583m on both sidesof the lesion (Figure 3(a) meanwidth of SR101-free area 111plusmn3 120583m 119873 = 5) In slices from GFPGFAP transgenic animals(119873 = 5 Figure 3(b)) GFAP-positive weakly SR101-positivescar cells also stained with the sodium-sensitive fluorescenceindicator SBFI-AM (acetoxymethyl ester of sodium bindingbenzofuran isophthalate see below 119873 = 3) indicatingthat they were able to take up and de-esterify this dye(Figure 3(b))
8 ISRN Neurology
lowastlowastlowast
lowastlowastlowast
80
lowast
Control ScarPeriphery
120
160
0
40
200
Som
a siz
e (120583
m2)
(a)
Lesioned
StrP
StrR
StrP
Control
StrR
StrLM
Lesio
n sit
e
(b)
Figure 4 Lesion-induced changes in somatic area (a) Quantification of somatic area based on GFAPGFP fluorescence in control andlesioned slices The soma size increased both in astrocytes in the scar region (ldquoscarrdquo) and in astrocytes in the periphery of the lesion(ldquoperipheryrdquo lowastlowastlowast119875 lt 0001 lowast119875 lt 005) (b) Confocal images of GFP fluorescence in cultured slices obtained from GFPGFAP transgenicanimals under control conditions (left) and subjected to a lesion (right) Cells surrounded by boxes are shown at higher magnification in theinsets and the dashed lines around the soma illustrate the areas which served to determine their size StrP stratum pyramidale StrR stratumradiatum StrLM stratum lacunosum moleculare and scale bars 40 120583m and 10120583m (inset)
In addition to GFAP-positive cells the scar region hostedmicroglial cells identified by their vital staining with TexasRed-coupled lectin from lycopersicon esculentum (tomatolectin119873 = 4 not shown) Tomato lectin-positive microgliahowever did not take up SBFI-AM (119873 = 4 not shown) con-firming earlier reports thatmicroglial cells do not incorporateAM-ester dyes in situ [47]
To further characterize reactive gliosis in response to thescratch wound we determined the area of astrocyte somatain confocal images of the GFP fluorescence in control andlesioned slices obtained fromGFPGFAP transgenic animalsThesemeasurements revealed that compared to astrocytes in
control slices the soma area of astrocytes in the periphery ofa lesion was increased by 18 and that of astrocytes in thescar regionwas increased by 24 (control 163plusmn2 120583m2 lesionperiphery 193plusmn3 120583m2 scar tissue 201plusmn4 120583m2119875 lt 0001 119899 =945 496 and 439 in 7 control and 9 lesioned slices Figures4(a) and 4(b)) Thus astrocytes show a second hallmark ofreactive gliosis after lesion namely a hypertrophy of their cellbodies which is observed both in the scar tissue as well as inthe periphery of the lesion
Taken together these data show that 6-7 days after themechanical lesion a glial scar has formed along the lesionsite This scar encompasses strongly activated astrocytes
ISRN Neurology 9
characterized by a robust increase in GFAP and S100120573expressionThese proximal reactive astrocytes locatedwithina distance of less than 100120583m from the lesion exhibit longGFAP-positive processes have significantly swollen somataand only weakly stain with SR101 Astrocytes in the peripheryof the scar (100ndash350 120583m from the lesion) display swollen cellbodies as well but show onlymoderate upregulation of GFAPand maintain their ability to efficiently take up SR101
33 Expression Levels and Spatial Distribution of GLT-1 andGLAST To determine the spatial distribution of the gluta-mate transporters GLAST and GLT-1 in organotypic slicesand changes therein in response to the lesion immunohisto-chemistry was employed In control slices GLAST (119873 = 13Figure 5(a)) as well as GLT-1 immunoreactivity (119873 = 11Figure 5(b)) appeared as punctate labeling throughout theentire CA1 area The staining patterns for both transportersin the tissue slice cultures were thus similar to those reportedfrom acute tissue slice preparations of the hippocampus [3648 49]
The relatively homogeneous distribution of immunore-activity throughout the preparation found in control slicecultures was abandoned following a lesion Here distinctaccumulations of GLAST (119873 = 17 Figure 5(a)) and GLT-1immunoreactivity (119873 = 12 Figure 5(b)) were found in thescar region In both cases these preferentially colocalizedwith thick GFAP-positive bundles running in parallel to thelesion (Figure 5) Apart from the marked accumulation ofGLAST reactivity in parallel to the scar GLAST immunore-activity appeared generally increased in some lesioned slicepreparations (Figure 5(a)) but unaltered in others bothwithinthe scar as well as in its periphery For GLT-1 a decline in theintensity of immunoreactivity was observed with increasingdistance from the lesion (Figure 5(b))
In summary these results reveal distinct alterations inthe immunoreactivity of GLAST and GLT-1 following amechanical lesion In the scar region clusters of GLT-1 andGLAST immunoreactivity are present primarily colocalizedwith strongly GFAP-positive fibers Apart from these distinctclusters GLT-1 immunoreactivity appears to decline withdistance from the scar whereas GLAST immunoreactivityseems largely uniform throughout the preparation
34 Sodium Transients Induced by Activation of GlutamateUptake Glutamate uptake is accompanied by the inwardflux of sodium resulting in increase in the intracellularsodium concentration of astrocytes To monitor its functionwe assayed sodium increases induced by D-aspartate anonmetabolized substrate of sodium-dependent glutamateuptake which is also transported [1] in astrocytes in thestratum radiatum of organotypic slices To this end SR101-stained slices were additionally loaded with the sodium-sensitive fluorophore SBFI-AM (Figure 6(a) cf Figure 3(b))In situ calibrations of the SBFI fluorescence revealed a linearincrease in its fluorescence ratio at intracellular sodiumconcentrations between 10 and 40mM (119899 = 107 Figure 6(b))as reported before [33]
Repetitive pressure application of D-aspartate for 500msinduced reliable increases in the intracellular sodium con-centration in astrocytes indicating activation of sodium-dependent glutamate uptake (119899 = 488 Figure 6(c)) Theexperiments were performed in the presence of the sodium-channel blocker TTX (tetrodotoxin 05 120583M) to suppressaction potential generation The amplitude of D-aspartate-induced sodium increases was dependent on the concen-tration applied (Figure 6(c)) and saturated at about 2mM(119899 = 19 Figure 6(d)) as reported before for mouse corticalastrocytes in culture [50] At a concentration of 1mM theamplitude of D-aspartate-induced sodium transients wasnear maximum and amounted to 38 plusmn 02mM (119899 =161 Figures 6(c) and 6(d)) a value comparable to that inastrocytes in acute slices obtained in our lab [33]
To probe for changes in glutamate uptake we comparedsodium signals induced by 1mMD-aspartate between controland lesioned slices (Figure 7) In lesioned slices astrocyteslocated in the periphery of the lesion (100ndash350 120583m distance)responded with a sodium transient to application of 1mMD-aspartate (see cell 2 in Figures 7(a) and 7(b)) The amplitudeof D-aspartate-induced sodium signals in cells in the lesionperiphery was similar to that observed in control slices (38 plusmn03mM 119899 = 111 see cell 1 in Figures 7(a) 7(b) and 7(c))Application of D-aspartate also evoked sodium transientsin weakly SR101-positive SBFI-filled astrocytes located inthe glial scar area (lt100 120583m from the lesion see cells 3ndash5in Figures 7(a) and 7(b)) These cells however displayedsignificantly smaller peak amplitudes (16 plusmn 021mM 119899 =26 119875 lt 0001 Figures 7(b) and 7(c)) than astrocytes inthe periphery and in control slices
The kinetics of D-aspartate-induced sodium transientsdiffered between all three groups This was true for theslope of the sodium increase (10ndash90 change) which wassignificantly reduced in astrocytes from lesioned slices versusastrocytes from control slices (09 plusmn 005mMs in controlversus 06 plusmn 006mMs in the periphery and 02 plusmn 004mMsin the scar region 119899 = 161 111 and 26 resp 119875 lt 0001Figures 7(b) and 7(c)) In addition the decay back to baselinewas significantly slower following lesion (Figures 7(b) and7(c)) The time needed for the signal to decay by 67 wasapproximately doubled (decay time 120591 was 44 plusmn 2 s in control120591 = 81 plusmn 4 s in periphery of the lesion and 120591 = 95 plusmn 10 s inscar cells Figures 7(b) and 7(c))
These results show that mechanical injury results indistinct alterations in the kinetics andor amplitudes ofD-aspartate-induced intracellular sodium transients in reac-tive astrocytes depending on their distance to the lesion
4 Discussion
Our results show that within 6-7 days after setting a scratchwound in organotypic hippocampal slices a dense glial scarhad formed along the lesion site Up to approximately 100 120583mdistance from the lesion the scar tissue was characterized bya substantial elevation ofGFAP expression a pronounced cel-lular hypertrophy and reduced uptake of SR101 by astrocytes
10 ISRN Neurology
Con
trol
GLAST
Lesio
ned
GFAP Merge
StrR
StrP
StrO
Lesio
n sit
e
StrR
(a)
Con
trol
Lesio
ned
GLT-1 GFAP Merge
StrR
StrP
StrO
StrR
StrP
StrLM
Lesio
n sit
e
(b)
Figure 5 GLAST andGLT-1 immunoreactivity aftermechanical injury (a) Fluorescence images of immunohistochemical stainings detectingthe glutamate transporters GLAST (left) GFAP (center) as counterstaining and the corresponding merged image (GFAP green GLASTmagenta) in control (top row) and lesioned slice cultures (bottom row) (b) GLT-1 (left) GFAP (center) double labeling of a control (toprow) and a mechanically lesioned slice (bottom row) The corresponding merge (GFAP green GLT-1 magenta) is found on the right Notethat after lesion glutamate transporter immunoreactive clusters are accumulated in the direct vicinity of the lesion site (arrowheads) GFAPglial fibrillary acidic protein GLAST glutamate aspartate transporter GLT-1 glutamate-transporter-1 StrO stratum oriens StrP stratumpyramidale StrR stratum radiatum StrLM stratum lacunosum-moleculare and scale bars (a) 20 120583m (b) 40 120583m
While prominent clusters of GLAST and GLT-1 immunore-activity were observed alongGFAP-positive structures in scarcells D-aspartate-induced intracellular sodium signals werestrongly dampened indicating a significant reduction in glu-tamate uptake capacity close to the lesion In the periphery ofthe lesion (100ndash350120583m) astrocytes showed less pronounced
reactivity maintained their ability to efficiently take up SR101and displayed only minor changes in glutamate transporterimmunoreactivity and function Thus depending on theirdistance from the lesion astrocytes showed different gradesof reactivity and displayed discrete changes in glutamatetransporter immunoreactivity and uptake capacity
ISRN Neurology 11
SR101 MergeSBFI
StrRStrP
(a)
03
04
05
06
07
0 5 10 15
0
20 25 30 40 0
100
20
40
80
60
0 10 20 30 400
20
40
60
80
100
10 = 55 mM
107 cells in 6 experiments
F (340
380
) (ra
tio)
Intracellular Na+ concentration (mM)Intracellular Na+ concentration (mM)
ΔF
(340
380
) (
)
ΔF
(340
380
) (
) R2 = 097
(b)
D-aspartate
100 120583M500 120583M
750 120583M1000 120583M
1 mM
10 s
Δ
(c)
0 1 2 3 4 50
1
2
3
4
5
Δ so
dium
(mM
)
D-aspartate (mM)
lowast
(d)
Figure 6 Sodium changes induced by activation of glutamate uptake (a) Staining of astrocytes by colabeling with SR101 and SBFI Leftepifluorescence image of theCA1-area of an SR101-loaded organotypic sliceCenter SBFI fluorescence taken from the same areaRight overlaySBFI-loaded SR101-positive astrocytes appear in white (arrowheads) StrP stratum pyramidale StrR stratum radiatum scale bar 20 120583m (b)Left calibration of SBFI fluorescence in an astrocyte Depicted is the change in intracellular SBFI fluorescence ratio in the calibration solutionscontaining ionophores and in response to stepwise changes in the sodium concentration Right mean values plusmn SEM of the normalizedchanges in SBFI fluorescence at different sodium concentrations The black line represents a linear fit of the data points between 10 and40mM sodium illustrating that a 10 change in SBFI fluorescence represents a change in the sodium concentration by 55mM (c) Sodiumtransients in a single astrocyte induced by 500ms applications of D-aspartate at different concentrations (d) Mean values plusmn S E M (119899 = 19)of the peak amplitude of sodium transients at different concentrations of D-aspartate The curve represents a sigmoidal fit of the data
41 Characteristics of Lesion-Induced Reactive Gliosis inOrganotypic Cultures Organotypic tissue slice cultures ofthe hippocampus represent a well-established model systemin which the basic tissue architecture is maintained butwhich still hosts the major advantages of cell cultures suchas good accessibility and control of experimental conditionsThey have been used extensively for the analysis of neuronal
properties and developing neuronal networks [28 51 52]or the study of excitotoxic neuronal damage (eg [53 54])Recent studies also demonstrated that basic morphologicalcharacteristics of astrocytes and the typical glia-synapseorganization are well preserved in organotypic slice cultures(eg [44 45 55 56]) In addition we found a nearly completeoverlap of SR101 staining with the expression of GFP in slice
12 ISRN Neurology
SBFIControl Lesioned
SR101
1
Lesio
n sit
e
SR101 SBFI
2 45
32 4
5
1
StrR
StrP StrP
StrR
3
(a)
ControlScar
Lesioned
1
D-asp
2
D-asp
D-asp
5
4
3Periphery1mM
20 s
Δ Δ
Δ
Δ
Δ
(b)
0102030405060708090
100110
Δ so
dium
tim
e (m
Ms
)
00
02
04
06
08
10
12
0
1
2
3
4
5
6
ns
Control Periphery Scar Control Periphery Scar Control Periphery Scar
Amplitude Slope Decay time
Δ so
dium
(mM
)
120591(s
)lowastlowastlowast
lowastlowastlowastlowastlowastlowast
lowastlowastlowastlowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowast
(c)
Figure 7 D-aspartate-induced astrocyte sodium transients following lesion (a) CA1 area of a control (left) and a lesioned (right) slicepreparation Shown are epifluorescence images of SR101 and SBFI fluorescences Circled areas and numbers indicate the regions of interestanalyzed in the experiment depicted in (b) StrP stratum pyramidale StrR stratum radiatum and scale bar 40 120583m (b) Sodium transientsinduced by pressure application of 1mMD-aspartate (D-asp) for 500ms in the different cells in the control and lesioned slice as indicated in(a) (c) Histograms showing mean values plusmn S E M of the peak amplitude (left) slope (center) and decay time (right) of D-aspartate-inducedsodium transients in astrocytes in control slices (119899 = 161) as well as in cells in the scar region and its periphery (119899 = 111 and 119899 = 26 resp)lowastlowast119875 lt 001 lowastlowastlowast119875 lt 0001 StrP stratum pyramidale StrR stratum radiatum and scale bar 40120583m
cultures obtained from GFPGFAP mice demonstrating thatSR101 is well suited to identify astrocytes in this prepara-tion as described before for acute tissue slices and in vivo[29 30]
Performing a scratch through the CA1 region of organ-otypic slices resulted in the formation of a glial scar along thelesion site as judged by the prominent increase in expressionof GFAP and S100120573 In addition astrocyte cell bodies showed
a significant hypertrophy Furthermore astrocytes in theperiphery of the scar (100ndash350 120583m from the lesion) showedmild to moderate reactivity displaying swollen cell bodiesand only moderate upregulation of GFAP These changesare hallmarks of reactive gliosis observed after tissue injury[10 13 57] Generally the degree of injury and the distance ofthe astrocytes from the site of injury define the degree of theiractivation [11 13]
ISRN Neurology 13
Our study also revealed that strongly reactive astrocytesin the scar region only weakly stain with SR101 The reducedability of strongly activated astrocytes to accumulate SR101indicates that strong glial activation is accompanied by adownregulation of the organic anion transporter responsi-ble for uptake of SR101 This is reminiscent of immatureastrocytes in the early postnatal hippocampus which areSR101-negative [30] and in line with a wealth of experimentalevidence suggesting that reactive gliosis represents a processcomprising a dedifferentiation of astrocytes [17]
42 Changes in Glutamate Transporter Expression followingLesion Glutamate uptake in the hippocampus is mainlyachieved by the glial glutamate transporters GLT-1 andGLAST [1 2] A central factor in many brain pathologiesincluding traumatic brain injury is an increase in extracellu-lar glutamate and excitotoxicity it has been suggested that achange in the expression levels of glial glutamate transportersmight play a critical role in the failure of glutamate clearance[6 58ndash60] Earlier work has found an overall downregulationof protein levels of both GLAST and GLT-1 following astro-cyte activation [22ndash27] indicating that this is causal to theelevation of extracellular glutamate In contrast to this notionother studies reported increased glutamate transport capacityof reactive astrocytes and suggested a protective influence[61 62]
To visualize possible differences in the spatial expressionprofile of GLT-1 and GLAST in strongly reactive astrocytesalong the scar as compared to moderately reactive astro-cytes in its periphery we performed immunohistochemicalstainings These revealed an accumulation of GLT-1 andGLAST immunoreactivity along thick GFAP-positive fiberswhich was particularly pronounced for GLAST Besides theseclusters overall GLAST immunoreactivity seemed unalteredas compared to unlesioned slices whereas GLT-1 immunore-activity seemedweaker and declined with increasing distancefrom the scar Clustering of glutamate transporters has beendescribed by several reports [48 63 64] In developinghippocampal astrocytes cluster formation was preferentiallyfound in branches opposed to synapses and was increasedwith increased neuronal activity indicating that it is neces-sary to copewith synaptic release of glutamate [36] Along thesame lines it was observed that sustained astroglial activationby ciliary neurotrophic factor (CNTF) in the rat striatuminduced a concentration of GLAST and GLT-1 into raftmicrodomains and improved glutamate clearance indicatingthat cluster formation increased the cellular capacity for glu-tamate uptake [61] However another study found glutamate-induced clustering of GLT-1 that induced its endocytosis andintracellular trafficking without changing the total expressionlevels as detected by western blots arguing for a decrease infunctional glial glutamate uptake capacity [49] In the presentstudy antibodies against GLT-1 and GLAST were employedfor immunohistochemistry after permeabilization of theplasma membrane The clusters of GLAST and GLT-1 alongGFAP-positive fibers in the scar region (as GLAST andGLT-1immunoreactivity in general) could thus represent glutamatetransporters in the plasmamembrane or in intracellular com-partments or both Consequently the observed clustering
does not allow a prediction about possible functional changesin glutamate uptake
43 Lesion-Induced Changes in Functional Glutamate UptakeCapacity We probed for the functional activation of gluta-mate transport by application of the transportable agonistD-aspartate which results in an increase in the intracellularsodium concentration of astrocytes [8 9] The amplitudesof D-aspartate-induced sodium transients in astrocytes incontrol organotypic slices were similar to those reportedfrom astrocytes in acute slices obtained in our lab [33]indicating similar cellular glutamate transport capacity inboth preparations
Following a lesion D-aspartate-induced intracellularsodium signalswere clearly dampened in scar cellsMoreoverthe slope of the increase in sodium was significantly reducedand the decay back to baseline was slowed Because D-aspartate was applied at nearly saturating concentrationthe reduction in peak amplitudes suggests a reduction infunctional glutamate uptake capacity in cells close to thelesion which might be mediated by a decrease in the overallnumber of glutamate transporters available at the plasmamembrane In contrast the alteration in the kinetics of theD-aspartate-induced sodium signals can be explained by thecellular hypertrophy that was observed in reactive astrocytesAt an equal transport-mediated sodium influx across themembrane the slope of resulting changes in the sodiumconcentration will be decreased in cells with a larger volume
Furthermore the decay of intracellular sodium transientsback to baseline is mainly governed by the activity of theNa+K+-ATPase as well as by diffusion [8] The slower recov-ery is in line with earlier studies reporting a downregulationof the sodium pump following reactive gliosis [65 66]Moreover sodium is not buffered in the cell and travels solelyby means of diffusion in the cytoplasm Hindered diffusionresulting from increased cytoplasmic protein content orimpaired gap junction coupling which both occur uponastrocytic activation might thus also partially contribute todelayed sodium recovery [12 34 67]
In contrast to scar cells the amplitude of D-aspartate-induced intracellular sodium signals was not significantlyaltered in moderately activated astrocytes in the periphery ofthe lesion indicating that the overall number of functionalglutamate transporters was not significantly altered Becausethese cells showed hypertrophy as well the slowed kinetics ofthe sodium transients are in line with an increased cellularvolume as well as a possible downregulation of Na+K+-ATPase andor a slowed diffusion as argued above
Taken together our results indicate a significant reduc-tion in glutamate uptake capacity in strongly activated astro-cytes in the scar region Thus the prominent clustering ofGLAST and GLT-1 immunoreactivity along GFAP-positivestructures close to the lesion is likely to reflect a loss ofglutamate transporters from the plasma membrane upontransporter internalization as reported by Nakagawa et al[49] The authors of the latter study speculated that large andprolonged increases in glutamate concentrations are neces-sary to induce such cluster formation and endocytosis in thetissue as they would possibly only occur under pathological
14 ISRN Neurology
conditions Indeed ambient glutamate concentrations wereshown to be significantly increased after traumatic braininjury [68ndash70] Along these lines it can be assumed thatin our model system extracellular glutamate concentrationsrose less severely in the periphery of the lesion preventinga comparable clustering and loss of functional glutamatetransport activity in moderately activated astrocytes
5 Conclusions
Our results show complex changes in glutamate transporterexpression and function during astrocyte activation followingmechanical injury They confirm that the degree of astrocyteactivation depends on the distance from the insult and com-prises different endpoints regarding cellular morphology andphysiology Furthermore our results highlight that astrocyteswhich show different grades of reactivity also display discretechanges in glutamate transporter expression and functionWhile immunohistochemistry revealed a prominent clus-tering of GLT-1 and GLAST immunoreactivity in the scarregion our functional assay clearly showed that glutamateuptake capacity is strongly reduced in scar cells while itis largely maintained in moderately activated astrocytes inthe periphery Thus mild-to-moderate astrogliosis in theperiphery of a mechanical lesion does not necessarily seemto be accompanied by a significant change in glial glutamateuptake capacity At the glial scar itself in strongly reactiveastrocytes a clustering of glutamate transporters is observedthat apparently goes along with a severe functional reductionin astroglial glutamate uptake which may contribute toglutamate-mediated excitotoxicity in this region
Conflict of Interests
The authors of the paper declare that they do not haveany direct financial relation with the commercial identitiesmentioned in the paper that might lead to a conflict ofinterests
Acknowledgments
This study was supported by the Deutsche Forschungsge-meinschaft (DFG Ro23274-3) The authors thank SimoneDurry and Claudia Roderigo for expert technical help
References
[1] N C Danbolt ldquoGlutamate uptakerdquo Progress in Neurobiologyvol 65 no 1 pp 1ndash105 2001
[2] A V Tzingounis and J I Wadiche ldquoGlutamate transportersconfining runaway excitation by shaping synaptic transmis-sionrdquo Nature Reviews Neuroscience vol 8 no 12 pp 935ndash9472007
[3] D E Bergles J S Diamond and C E Jahr ldquoClearance ofglutamate inside the synapse and beyondrdquo Current Opinion inNeurobiology vol 9 no 3 pp 293ndash298 1999
[4] C M Anderson and R A Swanson ldquoAstrocyte glutamatetransport review of properties regulation and physiologicalfunctionsrdquo Glia vol 32 no 1 pp 1ndash14 2000
[5] P Marcaggi and D Attwell ldquoRole of glial amino acid trans-porters in synaptic transmission and brain energeticsrdquoGlia vol47 no 3 pp 217ndash225 2004
[6] N J Maragakis and J D Rothstein ldquoGlutamate transportersanimal models to neurologic diseaserdquo Neurobiology of Diseasevol 15 no 3 pp 461ndash473 2004
[7] G Gegelashvili and A Schousboe ldquoCellular distribution andkinetic properties of high-affinity glutamate transportersrdquoBrainResearch Bulletin vol 45 no 3 pp 233ndash238 1998
[8] J W Deitmer and C R Rose ldquoIon changes and signalling inperisynaptic gliardquo Brain Research Reviews vol 63 no 1-2 pp113ndash129 2010
[9] S Kirischuk V Parpura and A Verkhratsky ldquoSodium dynam-ics another key to astroglial excitabilityrdquo Trends in Neuro-sciences vol 35 no 8 pp 497ndash506 2012
[10] M V Sofroniew ldquoMolecular dissection of reactive astrogliosisand glial scar formationrdquo Trends in Neurosciences vol 32 no12 pp 638ndash647 2009
[11] J L Ridet S K Malhotra A Privat and F H Gage ldquoReactiveastrocytes cellular and molecular cues to biological functionrdquoTrends in Neurosciences vol 20 no 12 pp 570ndash577 1997
[12] J Middeldorp and E M Hol ldquoGFAP in health and diseaserdquoProgress in Neurobiology vol 93 no 3 pp 421ndash443 2011
[13] M Pekny and M Nilsson ldquoAstrocyte activation and reactivegliosisrdquo Glia vol 50 no 4 pp 427ndash434 2005
[14] A Bordey S A Lyons J J Hablitz and H SontheimerldquoElectrophysiological characteristics of reactive astrocytes inexperimental cortical dysplasiardquo Journal of Neurophysiologyvol 85 no 4 pp 1719ndash1731 2001
[15] R Jabs G Seifert and C Steinhauser ldquoAstrocytic functionand its alteration in the epileptic brainrdquo Epilepsia vol 49supplement 2 pp 3ndash12 2008
[16] A Buffo I Rite P Tripathi et al ldquoOrigin and progeny ofreactive gliosis a source of multipotent cells in the injuredbrainrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 105 no 9 pp 3581ndash3586 2008
[17] S Robel B Berninger and M Gotz ldquoThe stem cell potential ofglia lessons from reactive gliosisrdquoNature Reviews Neurosciencevol 12 no 2 pp 88ndash104 2011
[18] H Kawano J Kimura-Kuroda Y Komuta et al ldquoRole of thelesion scar in the response to damage and repair of the centralnervous systemrdquo Cell and Tissue Research vol 349 no 1 pp169ndash180 2012
[19] G Yiu and Z He ldquoGlial inhibition of CNS axon regenerationrdquoNature Reviews Neuroscience vol 7 no 8 pp 617ndash627 2006
[20] M T Fitch and J Silver ldquoCNS injury glial scars and inflamma-tion inhibitory extracellular matrices and regeneration failurerdquoExperimental Neurology vol 209 no 2 pp 294ndash301 2008
[21] L Li A Lundkvist D Andersson et al ldquoProtective role ofreactive astrocytes in brain ischemiardquo Journal of Cerebral BloodFlow amp Metabolism vol 28 no 3 pp 468ndash481 2008
[22] F K H van Landeghem T Weiss M Oehmichen and AVonDeimling ldquoDecreased expression of glutamate transportersin astrocytes after human traumatic brain injuryrdquo Journal ofNeurotrauma vol 23 no 10 pp 1518ndash1528 2006
[23] V L Raghavendra Rao M K Baskaya A Dogan J DRothstein and R J Dempsey ldquoTraumatic brain injury down-regulates glial glutamate transporter (GLT- 1 and GLAST)proteins in rat brainrdquo Journal of Neurochemistry vol 70 no 5pp 2020ndash2027 1998
ISRN Neurology 15
[24] J-C Chen H Hsu-Chou J-L Lu et al ldquoDown-regulation ofthe glial glutamate transporter GLT-1 in rat hippocampus andstriatum and its modulation by a group III metabotropic glu-tamate receptor antagonist following transient global forebrainischemiardquoNeuropharmacology vol 49 no 5 pp 703ndash714 2005
[25] S D Ginsberg L J Martin and J D Rothstein ldquoRegional deaf-ferentation down-regulates subtypes of glutamate transporterproteinsrdquo Journal of Neurochemistry vol 65 no 6 pp 2800ndash2803 1995
[26] J E Springer R D Azbill R J Mark J G Begley GWaeg andM P Mattson ldquo4-Hydroxynonenal a lipid peroxidation prod-uct rapidly accumulates following traumatic spinal cord injuryand inhibits glutamate uptakerdquo Journal of Neurochemistry vol68 no 6 pp 2469ndash2476 1997
[27] M B Moretto N S Arteni D Lavinsky et al ldquoHypoxic-ischemic insult decreases glutamate uptake by hippocampalslices from neonatal rats Prevention by guanosinerdquo Experimen-tal Neurology vol 195 no 2 pp 400ndash406 2005
[28] L Stoppini P-A Buchs and D Muller ldquoA simple method fororganotypic cultures of nervous tissuerdquo Journal of NeuroscienceMethods vol 37 no 2 pp 173ndash182 1991
[29] A Nimmerjahn F Kirchhoff J N D Kerr and F HelmchenldquoSulforhodamine 101 as a specific marker of astroglia in theneocortex in vivordquo Nat Methods vol 1 no 1 pp 31ndash37 2004
[30] K W Kafitz S D Meier J Stephan and C R Rose ldquoDevel-opmental profile and properties of sulforhodamine 101-Labeledglial cells in acute brain slices of rat hippocampusrdquo Journal ofNeuroscience Methods vol 169 no 1 pp 84ndash92 2008
[31] C Schnell Y Hagos and S Hulsmann ldquoActive sulforhodamine101 uptake into hippocampal astrocytesrdquo PLoS One vol 7 no 11Article ID e49398 2012
[32] S D Meier Y Kovalchuk and C R Rose ldquoProperties ofthe new fluorescent Na+ indicator CoroNa Green comparisonwith SBFI and confocal Na+ imagingrdquo Journal of NeuroscienceMethods vol 155 no 2 pp 251ndash259 2006
[33] J Langer and C R Rose ldquoSynaptically induced sodium signalsin hippocampal astrocytes in siturdquo Journal of Physiology vol587 part 24 pp 5859ndash5877 2009
[34] J Langer J Stephan M Theis and C R Rose ldquoGap junctionsmediate intercellular spread of sodium between hippocampalastrocytes in siturdquo Glia vol 60 no 2 pp 239ndash252 2012
[35] C R Rose andB R Ransom ldquoIntracellular sodiumhomeostasisin rat hippocampal astrocytesrdquo Journal of Physiology vol 491part 2 no 2 pp 291ndash305 1996
[36] A M Benediktsson G S Marrs J C Tu et al ldquoNeuronalactivity regulates glutamate transporter dynamics in developingastrocytesrdquo Glia vol 60 no 2 pp 175ndash188 2012
[37] M Zhou G P Schools and H K Kimelberg ldquoDevelopment ofGLAST(+) astrocytes andNG2(+) glia in rat hippocampus CA1mature astrocytes are electrophysiologically passiverdquo Journal ofNeurophysiology vol 95 no 1 pp 134ndash143 2006
[38] I Suarez G Bodega and B Fernandez ldquoModulation of glu-tamate transporters (GLAST GLT-1 and EAAC1) in the ratcerebellum following portocaval anastomosisrdquo Brain Researchvol 859 no 2 pp 293ndash302 2000
[39] X Zhu D E Bergles and A Nishiyama ldquoNG2 cells generateboth oligodendrocytes and gray matter astrocytesrdquo Develop-ment vol 135 no 1 pp 145ndash157 2008
[40] B Brunne S ZhaoADerouiche et al ldquoOriginmaturation andastroglial transformation of secondary radial glial cells in thedeveloping dentate gyrusrdquo Glia vol 58 no 13 pp 1553ndash15692010
[41] Y Wu A-Q Zhang and D T Yew ldquoAge related changes ofvarious markers of astrocytes in senescence-accelerated micehippocampusrdquo Neurochemistry International vol 46 no 7 pp565ndash574 2005
[42] S Magavi D Friedmann G Banks A Stolfi and C LoisldquoCoincident generation of pyramidal neurons and protoplasmicastrocytes in neocortical columnsrdquo Journal of Neuroscience vol32 no 14 pp 4762ndash4772 2012
[43] M Olabarria H N Noristani A Verkhratsky and J JRodrıguez ldquoConcomitant astroglial atrophy and astrogliosis ina triple transgenic animal model of Alzheimerrsquos diseaserdquo Gliavol 58 no 7 pp 831ndash838 2010
[44] A M Benediktsson S J Schachtele S H Green and M EDailey ldquoBallistic labeling and dynamic imaging of astrocytesin organotypic hippocampal slice culturesrdquo Journal of Neuro-science Methods vol 141 no 1 pp 41ndash53 2005
[45] I Lushnikova G Skibo D Muller and I Nikonenko ldquoSynapticpotentiation induces increased glial coverage of excitatorysynapses in CA1 hippocampusrdquoHippocampus vol 19 no 8 pp753ndash762 2009
[46] E Raponi F Agenes C Delphin et al ldquoS100B expressiondefines a state in which GFAP-expressing cells lose their neuralstem cell potential and acquire a more mature developmentalstagerdquo Glia vol 55 no 2 pp 165ndash177 2007
[47] C G Schipke C Boucsein C Ohlemeyer F Kirchhoff andH Kettenmann ldquoAstrocyte Ca2+ waves trigger responses inmicroglial cells in brain slicesrdquo The FASEB Journal vol 16 no2 pp 255ndash257 2002
[48] J Zhou and M L Sutherland ldquoGlutamate transporter clusterformation in astrocytic processes regulates glutamate uptakeactivityrdquo Journal of Neuroscience vol 24 no 28 pp 6301ndash63062004
[49] T Nakagawa Y Otsubo Y Yatani H Shirakawa and S KanekoldquoMechanisms of substrate transport-induced clustering of aglial glutamate transporter GLT-1 in astroglial-neuronal cul-turesrdquo European Journal of Neuroscience vol 28 no 9 pp 1719ndash1730 2008
[50] J-Y Chatton P Marquet and P J Magistretti ldquoA quantitativeanalysis of L-glutamate-regulated Na+ dynamics in mousecortical astrocytes Implications for cellular bioenergeticsrdquoEuropean Journal of Neuroscience vol 12 no 11 pp 3843ndash38532000
[51] E Forster S Zhao and M Frotscher ldquoLaminating the hip-pocampusrdquo Nature Reviews Neuroscience vol 7 no 4 pp 259ndash267 2006
[52] B H Gahwiler M Capogna D Debanne R A McKinney andS M Thompson ldquoOrganotypic slice cultures a technique hascome of agerdquo Trends in Neurosciences vol 20 no 10 pp 471ndash477 1997
[53] P E Kunkler and R P Kraig ldquoReactive astrocytosis fromexcitotoxic injury in hippocampal organ culture parallels thatseen in vivordquo Journal of Cerebral Blood Flow and Metabolismvol 17 no 1 pp 26ndash43 1997
[54] I E Holopainen ldquoOrganotypic hippocampal slice cultures amodel system to study basic cellular andmolecularmechanismsof neuronal cell death neuroprotection and synaptic plasticityrdquoNeurochemical Research vol 30 no 12 pp 1521ndash1528 2005
[55] MHaber L Zhou andK KMurai ldquoCooperative astrocyte anddendritic spine dynamics at hippocampal excitatory synapsesrdquoJournal of Neuroscience vol 26 no 35 pp 8881ndash8891 2006
16 ISRN Neurology
[56] H Nishida and S Okabe ldquoDirect astrocytic contacts regulatelocal maturation of dendritic spinesrdquo Journal of Neurosciencevol 27 no 2 pp 331ndash340 2007
[57] A Buffo C Rolando and S Ceruti ldquoAstrocytes in the dam-aged brain molecular and cellular insights into their reactiveresponse and healing potentialrdquoBiochemical Pharmacology vol79 no 2 pp 77ndash89 2010
[58] J-H Yi and A S Hazell ldquoExcitotoxic mechanisms and the roleof astrocytic glutamate transporters in traumatic brain injuryrdquoNeurochemistry International vol 48 no 5 pp 394ndash403 2006
[59] C Werner and K Engelhard ldquoPathophysiology of traumaticbrain injuryrdquo British Journal of Anaesthesia vol 99 no 1 pp4ndash9 2007
[60] G Gegelashvili M B Robinson D Trotti and T RauenldquoRegulation of glutamate transporters in health and diseaserdquoProgress in Brain Research vol 132 pp 267ndash286 2001
[61] C Escartin E Brouillet PGubellini et al ldquoCiliary neurotrophicfactor activates astrocytes redistributes their glutamate trans-porters GLAST and GLT-1 to raft microdomains and improvesglutamate handling in vivordquo Journal of Neuroscience vol 26 no22 pp 5978ndash5989 2006
[62] C Vermeiren M Najimi N Vanhoutte et al ldquoAcute up-regulation of glutamate uptake mediated by mGluR5a in reac-tive astrocytesrdquo Journal of Neurochemistry vol 94 no 2 pp405ndash416 2005
[63] C L Poitry-Yamate L Vutskits and T Rauen ldquoNeuronal-induced and glutamate-dependent activation of glial glutamatetransporter functionrdquo Journal of Neurochemistry vol 82 no 4pp 987ndash997 2002
[64] P M Beart and R D OrsquoShea ldquoTransporters for L-glutamatean update on their molecular pharmacology and pathologicalinvolvementrdquo British Journal of Pharmacology vol 150 no 1pp 5ndash17 2007
[65] F D Lima M A Souza A F Furian et al ldquoNa+K+-ATPaseactivity impairment after experimental traumatic brain injuryrelationship to spatial learning deficits and oxidative stressrdquoBehavioural Brain Research vol 193 no 2 pp 306ndash310 2008
[66] G A Gusarova H E Trejo L A Dada et al ldquoHypoxia leadsto NaK-ATPase downregulation via Ca2+ release-activatedCa2+ channels and AMPK activationrdquo Molecular and CellularBiology vol 31 no 17 pp 3546ndash3556 2011
[67] S S L Chew C S Johnson C R Green and H V Danesh-Meyer ldquoRole of connexin43 in central nervous system injuryrdquoExperimental Neurology vol 225 no 2 pp 250ndash261 2010
[68] P Nilsson L Hillered U Ponten and U Ungerstedt ldquoChangesin cortical extracellular levels of energy-related metabolites andamino acids following concussive brain injury in ratsrdquo Journal ofCerebral Blood Flow and Metabolism vol 10 no 5 pp 631ndash6371990
[69] H Katoh K Sima H Nawashiro K Wada and H ChigasakildquoThe effect of MK-801 on extracellular neuroactive amino acidsin hippocampus after closed head injury followed by hypoxia inratsrdquo Brain Research vol 758 no 1-2 pp 153ndash162 1997
[70] H Koizumi H Fujisawa H Ito T Maekawa X Di and RBullock ldquoEffects of mild hypothermia on cerebral blood flow-independent changes in cortical extracellular levels of aminoacids following contusion trauma in the ratrdquoBrain Research vol747 no 2 pp 304ndash312 1997
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
4 ISRN Neurology
Table 1 List of antibodies used
Antibody Structure of theimmunogen Company Order number Species Type Dilution
GLAST(EAAT1)
KPYQLIAQDNEPEKPVADSETKM ChemiconMillipore AB1782lowast Guinea
Pig IgGPolyclonal 1 1000
GLT-1(EAAT2)
AANGKSADCSVEEEPWKREK ChemiconMillipore AB1783 Guinea
Pig IgGPolyclonal 1 10001 2000
GFAP ns (full-length proteinfrom cow spinal cord) Dako Cytomation Z 0334 Rabbit
IgGPolyclonal 1 10001 2000
GFAP
clone G-A-5(LQSLTCDVESLRGTNESLERQMREQEERHAREAASYQEALTRLEEEGQSLKDEMARHLQEYQELLNVKLALDIEIATY)
Sigma-Aldrich G3893 MouseIgG
Monoclonal 1 10001 2000
S100szlig ns (recombinantfull-length cow protein) Abcam ab868 Rabbit
IgGPolyclonal 1 100
MouseIgG AF488 conjugated Invitrogen (Life Technologies) A11029 Goat mdash 1 100
RabbitIgG AF594 conjugated Invitrogen (Life Technologies) A11012 Goat mdash 1 100
Guineapig IgG AF488 conjugated Invitrogen (Life Technologies) A11073 Goat mdash 1 100
GLAST glutamate aspartate transporter GLT-1 glutamate transporter 1 GFAP glial fibrillary protein AF AlexaFluor appl application IHC immunohisto-chemistry WB western blot ns not specifiedlowastThemanufacturer has lately stopped the sale and distribution of the GLAST antibody used here
BX51WI microscope coupled to a confocal laser scanningsystem (FV300) equipped with a multiline argon (488 nm)and a helium-neon laser (543 nm both Melles Griot Ben-sheim Germany) was used Images were collected witheither a 20x050 (UMPlanFl Olympus Hamburg Ger-many) a 40x080 water immersion (LUMPlan OlympusHamburg Germany) or a 60x140 oil immersion objective(PlanApoVC Nikon Instruments Dusseldorf Germany) AKalman filter 4 was employed at every scan Simultaneousor sequential scanning of both fluorophores revealed nodifference in their staining patterns indicating absence ofcross-excitation or spectral bleed through The thickness ofz-plane sections was 1120583m and the number of optical sectionsvaried depending on the preparation
Figures show extended focus images as specified inthe figure legends which were calculated from z-stacksof optical sections using ImageJ software (NIH BethesdaUSA) Images were overlaid employing Adobe PhotoshopCS2 (Adobe Systems Cologne Germany)
26 Data Presentation and Statistics Unless otherwise spec-ified data are expressed as means plusmn SEM Data were statis-tically analyzed by Studentrsquos 119905-test employing the proceduresimplemented in OriginPro 8G Software (OriginLab Corpo-ration Northampton MA USA) A 119875 value of lt005 wasconsidered significant If not stated otherwise 119899representsthe number of analyzed cells and119873 the number of slices Eachset of experimentswas performedon at least three tissue slicesobtained from different animals
3 Results
31 Identification of Astrocytes in Organotypic Slice CulturesThe aim of the present study was to analyze changes inglutamate transporter expression and function in astrocytesin response to a mechanical injury To this end we preparedorganotypic tissue slice cultures of themouse hippocampus awell-establishedmodel system inwhich the layering and basiccellular organization of the neural network are maintainedover time [28] After 19ndash25 days in culture organotypic tissueslices had flattened to a thickness of 40ndash50 120583m and werecomposed of 3 to 4 cell layers At this stage propidium iodideassays detected only few dead cells dispersed throughoutthe slices (3 plusmn 06 cells per CA1 subfield 18 plusmn 34 in entirepreparation 119873 = 6 data not shown) indicating that thepreparation was viable and in a stable condition
To visualize astrocytes in cultured slices we used hip-pocampi of transgenic mice in which green fluorescentprotein (GFP) is expressed under the control of the promoterof the astrocyte-specific intermediate filament glial fibrillaryacidic protein (GFAP FVBN-Tg[GFAPGFP] ldquoGFPGFAPmicerdquo) A visualization of theGFP fluorescence demonstratedthat the general distribution and organization of astrocyteswere preserved in the strata pyramidale and radiatum ofthe CA1 region of organotypic cultures confirming earlierreports (119873 = 3 Figure 1(a) [44 45]) The vast majority ofGFP-expressing cells (sim95) also labeled with the astrocyte-specific vital marker SR101 in organotypic slices (119873 = 3Figures 1(a) and 1(b)) Furthermore immunohistochemicalstainings for GFAP in slice cultures derived from wildtype
ISRN Neurology 5
Slic
e cul
ture
(20
dic
)
GFP-GFAP MergeSR101
lowast lowast lowast lowast
lowast
lowast
lowast lowast
StrP
StrR
(a)
StrR
StrP
StrLM
StrM
SR101
StrO
(b)
GFAP S100120573
StrP
StrR
StrP
StrR
StrP
StrR
StrP
StrR
DAPI Merge
(c)
Figure 1 Astrocytes in organotypic hippocampal slice cultures (a) Organotypic slice cultures at 20 days in culture (dic) Preparations werederived from animals in which GFP was expressed under control of the GFAP promoter Shown is the expression of GFP (left) the labelingpattern of SR101 (center) and the corresponding merged image (GFAP green SR101 magenta) Asterisks point out SR101-labeled cells thatdo not express GFP arrowheads point out GFP-positive but SR101-negative cells Images represent maximum intensity projections of 15optical sections taken at 1120583m intervals (b) SR101 fluorescence of an organotypic slice at 21 dic Image represents a stack of 20 optical sectionstaken at 1 120583m intervals (c) Immunohistochemical double staining for GFAP and S100120573 as well as DAPI fluorescence and merged image ofthe fluorescence of GFAP (green) and S100120573 (magenta) of part of the CA1 region encompassing the strata pyramidale and radiatum of anorganotypic slice at 20 dic Note the radially oriented GFAP-positive fibers crossing the pyramidal cell layer Images were taken at a wide-field microscope ((a)ndash(c)) Dashed lines indicate the approximate boundaries of the stratum pyramidale StrO stratum oriens StrP stratumpyramidale StrR stratum radiatum StrLM stratum lacunosum moleculare and StrM stratum moleculare Scale bars (a) 50 120583m (b) 100120583mand (c) 50 120583m
animals showed that themajority of GFAP-positive cells werealso positive for S100120573 (119873 = 12 Figure 1(c)) a marker formature astrocytes [46]
Taken together the nearly complete overlap in the cellularstaining pattern for the vital dye SR101 with the expressionof GFP in GFPGFAP mice as well as the overlap in theimmunofluorescence for GFAP and S100120573 in organotypicslices indicates that these markers are well suited to identifyastrocytes in this preparation
32 Astrocyte Markers and Astrocyte Morphology followingMechanical Lesion To induce astrogliosis in organotypicslice cultures we used a scalpel blade and performed
a scratch under semisterile conditions through the entireCA1 region oriented perpendicular to the stratum pyramidale(Figure 2(a)) 6-7 days after the lesion a general enhance-ment of GFAP immunoreactivity was observed indicatingastrogliosis (Figure 2(b))The greatest enhancement of GFAPimmunoreactivity occurred within a distance of less than100 120583malong the lesion (119873 = 26 Figures 2(b) and 2(c)) Herea densemeshwork of thick GFAP-positive labels was presentindicative of the formation of a glial scar Furthermore theregion close to the lesion site exhibited a discernible increasein S100120573 immunoreactivity (119873 = 12 Figure 2(c)) Both theprominent expressions of GFAP and S100120573 are characteristicsof strongly reactive astrocytes which represent the major
6 ISRN Neurology
GFAP
DAPI
CA1 subregion
GFAP
DAPI
StrR
StrP StrP
StrR
StrOStrO
StrO
StrP
StrR
StrO
StrP
StrR
Control Lesioned
GFAP
Lesio
n sit
e
Hi
StrMStrG
StrP
StrR
StrLM
Scratch wound
StrO
Lesio
n sit
e
Lesioned
(a) (c)
(b)
350120583m350120583m
S100120573
Figure 2 Lesion-induced reactive gliosis in organotypic slice cultures (a) Schematic overview of the hippocampus indicating the positionof the scratch woundThe box delineates the area primarily analyzed (b) GFAP immunofluorescence and DAPI staining of the CA1 area in acontrol (left) and a lesioned (right) slice The boxes indicate the areas enlarged below The dashed line in the lower right indicates the lesion(c) Double staining for GFAP (top) and S100120573 (bottom) adjacent to the lesion site Note the gradual increase in GFAP and S100120573 expressiontowards the lesion (b) widefield microscopy (c) confocal microscopy (optical sections taken at 1120583m intervals) StrO stratum oriens StrPstratum pyramidale StrR stratum radiatum and StrLM stratum lacunosum moleculare Scale bars (b) 200 120583m (top) and 50 120583m (bottom) (c)10120583m
ISRN Neurology 7
StrR
StrP
StrLM
StrO
StrP StrRStrO StrLM
Lesio
n sit
e
SR101-staining pattern
(a)
SBFI
Lesio
n sit
e
SR101 GFAP-GFP
(b)
Figure 3 SR101 labeling in lesioned slices (a) SR101 fluorescence in the CA1 area in a slice 7 days after lesion The delineated area comprisesthe stratum pyramidale and the box indicates the area enlarged on the right side Right SR101 fluorescence at the lesion site illustrating thatSR101 labeling is reduced along the lesionThe dotted line indicates the transition between the scar area and the adjacent moderately reactivetissue The boxed area is further enlarged at the bottom (b) Image series obtained at a wide-field fluorescence microscope The slice wasprepared from a transgenic GFPGFAP mouse and the lesion site is indicated by the dotted line Left Image of SR101 fluorescence centerGFP fluorescence indicating GFAP expression and right SBFI fluorescence SR101 labeling is only weak in GFPSBFI-positive cells in the scarregion (see arrowheads) StrO stratum oriens StrP stratum pyramidale StrR stratum radiatum and StrLM stratum lacunosum-moleculareScale bars 100120583m ((a) upper left) and 20 120583m (other images)
cellular component of the glial scar forming after mechanicaltissue injury [10] More distal to the lesion (gt100ndash350 120583m) aless dramatic rearrangement of GFAP immunoreactivity wasobserved
A clear difference in astrocyte properties between thescar region and its periphery was also seen following SR101labeling In the periphery of the scar (100ndash350 120583m distancefrom the lesion) SR101 resulted in a reliable and brightcellular staining similar to what had been observed incontrol unlesioned slices (119873 = 12 Figure 3(a) cf Figure 1)
In contrast to this cellular SR101 staining was detectable butonly faint within a distance of less than 100 120583m on both sidesof the lesion (Figure 3(a) meanwidth of SR101-free area 111plusmn3 120583m 119873 = 5) In slices from GFPGFAP transgenic animals(119873 = 5 Figure 3(b)) GFAP-positive weakly SR101-positivescar cells also stained with the sodium-sensitive fluorescenceindicator SBFI-AM (acetoxymethyl ester of sodium bindingbenzofuran isophthalate see below 119873 = 3) indicatingthat they were able to take up and de-esterify this dye(Figure 3(b))
8 ISRN Neurology
lowastlowastlowast
lowastlowastlowast
80
lowast
Control ScarPeriphery
120
160
0
40
200
Som
a siz
e (120583
m2)
(a)
Lesioned
StrP
StrR
StrP
Control
StrR
StrLM
Lesio
n sit
e
(b)
Figure 4 Lesion-induced changes in somatic area (a) Quantification of somatic area based on GFAPGFP fluorescence in control andlesioned slices The soma size increased both in astrocytes in the scar region (ldquoscarrdquo) and in astrocytes in the periphery of the lesion(ldquoperipheryrdquo lowastlowastlowast119875 lt 0001 lowast119875 lt 005) (b) Confocal images of GFP fluorescence in cultured slices obtained from GFPGFAP transgenicanimals under control conditions (left) and subjected to a lesion (right) Cells surrounded by boxes are shown at higher magnification in theinsets and the dashed lines around the soma illustrate the areas which served to determine their size StrP stratum pyramidale StrR stratumradiatum StrLM stratum lacunosum moleculare and scale bars 40 120583m and 10120583m (inset)
In addition to GFAP-positive cells the scar region hostedmicroglial cells identified by their vital staining with TexasRed-coupled lectin from lycopersicon esculentum (tomatolectin119873 = 4 not shown) Tomato lectin-positive microgliahowever did not take up SBFI-AM (119873 = 4 not shown) con-firming earlier reports thatmicroglial cells do not incorporateAM-ester dyes in situ [47]
To further characterize reactive gliosis in response to thescratch wound we determined the area of astrocyte somatain confocal images of the GFP fluorescence in control andlesioned slices obtained fromGFPGFAP transgenic animalsThesemeasurements revealed that compared to astrocytes in
control slices the soma area of astrocytes in the periphery ofa lesion was increased by 18 and that of astrocytes in thescar regionwas increased by 24 (control 163plusmn2 120583m2 lesionperiphery 193plusmn3 120583m2 scar tissue 201plusmn4 120583m2119875 lt 0001 119899 =945 496 and 439 in 7 control and 9 lesioned slices Figures4(a) and 4(b)) Thus astrocytes show a second hallmark ofreactive gliosis after lesion namely a hypertrophy of their cellbodies which is observed both in the scar tissue as well as inthe periphery of the lesion
Taken together these data show that 6-7 days after themechanical lesion a glial scar has formed along the lesionsite This scar encompasses strongly activated astrocytes
ISRN Neurology 9
characterized by a robust increase in GFAP and S100120573expressionThese proximal reactive astrocytes locatedwithina distance of less than 100120583m from the lesion exhibit longGFAP-positive processes have significantly swollen somataand only weakly stain with SR101 Astrocytes in the peripheryof the scar (100ndash350 120583m from the lesion) display swollen cellbodies as well but show onlymoderate upregulation of GFAPand maintain their ability to efficiently take up SR101
33 Expression Levels and Spatial Distribution of GLT-1 andGLAST To determine the spatial distribution of the gluta-mate transporters GLAST and GLT-1 in organotypic slicesand changes therein in response to the lesion immunohisto-chemistry was employed In control slices GLAST (119873 = 13Figure 5(a)) as well as GLT-1 immunoreactivity (119873 = 11Figure 5(b)) appeared as punctate labeling throughout theentire CA1 area The staining patterns for both transportersin the tissue slice cultures were thus similar to those reportedfrom acute tissue slice preparations of the hippocampus [3648 49]
The relatively homogeneous distribution of immunore-activity throughout the preparation found in control slicecultures was abandoned following a lesion Here distinctaccumulations of GLAST (119873 = 17 Figure 5(a)) and GLT-1immunoreactivity (119873 = 12 Figure 5(b)) were found in thescar region In both cases these preferentially colocalizedwith thick GFAP-positive bundles running in parallel to thelesion (Figure 5) Apart from the marked accumulation ofGLAST reactivity in parallel to the scar GLAST immunore-activity appeared generally increased in some lesioned slicepreparations (Figure 5(a)) but unaltered in others bothwithinthe scar as well as in its periphery For GLT-1 a decline in theintensity of immunoreactivity was observed with increasingdistance from the lesion (Figure 5(b))
In summary these results reveal distinct alterations inthe immunoreactivity of GLAST and GLT-1 following amechanical lesion In the scar region clusters of GLT-1 andGLAST immunoreactivity are present primarily colocalizedwith strongly GFAP-positive fibers Apart from these distinctclusters GLT-1 immunoreactivity appears to decline withdistance from the scar whereas GLAST immunoreactivityseems largely uniform throughout the preparation
34 Sodium Transients Induced by Activation of GlutamateUptake Glutamate uptake is accompanied by the inwardflux of sodium resulting in increase in the intracellularsodium concentration of astrocytes To monitor its functionwe assayed sodium increases induced by D-aspartate anonmetabolized substrate of sodium-dependent glutamateuptake which is also transported [1] in astrocytes in thestratum radiatum of organotypic slices To this end SR101-stained slices were additionally loaded with the sodium-sensitive fluorophore SBFI-AM (Figure 6(a) cf Figure 3(b))In situ calibrations of the SBFI fluorescence revealed a linearincrease in its fluorescence ratio at intracellular sodiumconcentrations between 10 and 40mM (119899 = 107 Figure 6(b))as reported before [33]
Repetitive pressure application of D-aspartate for 500msinduced reliable increases in the intracellular sodium con-centration in astrocytes indicating activation of sodium-dependent glutamate uptake (119899 = 488 Figure 6(c)) Theexperiments were performed in the presence of the sodium-channel blocker TTX (tetrodotoxin 05 120583M) to suppressaction potential generation The amplitude of D-aspartate-induced sodium increases was dependent on the concen-tration applied (Figure 6(c)) and saturated at about 2mM(119899 = 19 Figure 6(d)) as reported before for mouse corticalastrocytes in culture [50] At a concentration of 1mM theamplitude of D-aspartate-induced sodium transients wasnear maximum and amounted to 38 plusmn 02mM (119899 =161 Figures 6(c) and 6(d)) a value comparable to that inastrocytes in acute slices obtained in our lab [33]
To probe for changes in glutamate uptake we comparedsodium signals induced by 1mMD-aspartate between controland lesioned slices (Figure 7) In lesioned slices astrocyteslocated in the periphery of the lesion (100ndash350 120583m distance)responded with a sodium transient to application of 1mMD-aspartate (see cell 2 in Figures 7(a) and 7(b)) The amplitudeof D-aspartate-induced sodium signals in cells in the lesionperiphery was similar to that observed in control slices (38 plusmn03mM 119899 = 111 see cell 1 in Figures 7(a) 7(b) and 7(c))Application of D-aspartate also evoked sodium transientsin weakly SR101-positive SBFI-filled astrocytes located inthe glial scar area (lt100 120583m from the lesion see cells 3ndash5in Figures 7(a) and 7(b)) These cells however displayedsignificantly smaller peak amplitudes (16 plusmn 021mM 119899 =26 119875 lt 0001 Figures 7(b) and 7(c)) than astrocytes inthe periphery and in control slices
The kinetics of D-aspartate-induced sodium transientsdiffered between all three groups This was true for theslope of the sodium increase (10ndash90 change) which wassignificantly reduced in astrocytes from lesioned slices versusastrocytes from control slices (09 plusmn 005mMs in controlversus 06 plusmn 006mMs in the periphery and 02 plusmn 004mMsin the scar region 119899 = 161 111 and 26 resp 119875 lt 0001Figures 7(b) and 7(c)) In addition the decay back to baselinewas significantly slower following lesion (Figures 7(b) and7(c)) The time needed for the signal to decay by 67 wasapproximately doubled (decay time 120591 was 44 plusmn 2 s in control120591 = 81 plusmn 4 s in periphery of the lesion and 120591 = 95 plusmn 10 s inscar cells Figures 7(b) and 7(c))
These results show that mechanical injury results indistinct alterations in the kinetics andor amplitudes ofD-aspartate-induced intracellular sodium transients in reac-tive astrocytes depending on their distance to the lesion
4 Discussion
Our results show that within 6-7 days after setting a scratchwound in organotypic hippocampal slices a dense glial scarhad formed along the lesion site Up to approximately 100 120583mdistance from the lesion the scar tissue was characterized bya substantial elevation ofGFAP expression a pronounced cel-lular hypertrophy and reduced uptake of SR101 by astrocytes
10 ISRN Neurology
Con
trol
GLAST
Lesio
ned
GFAP Merge
StrR
StrP
StrO
Lesio
n sit
e
StrR
(a)
Con
trol
Lesio
ned
GLT-1 GFAP Merge
StrR
StrP
StrO
StrR
StrP
StrLM
Lesio
n sit
e
(b)
Figure 5 GLAST andGLT-1 immunoreactivity aftermechanical injury (a) Fluorescence images of immunohistochemical stainings detectingthe glutamate transporters GLAST (left) GFAP (center) as counterstaining and the corresponding merged image (GFAP green GLASTmagenta) in control (top row) and lesioned slice cultures (bottom row) (b) GLT-1 (left) GFAP (center) double labeling of a control (toprow) and a mechanically lesioned slice (bottom row) The corresponding merge (GFAP green GLT-1 magenta) is found on the right Notethat after lesion glutamate transporter immunoreactive clusters are accumulated in the direct vicinity of the lesion site (arrowheads) GFAPglial fibrillary acidic protein GLAST glutamate aspartate transporter GLT-1 glutamate-transporter-1 StrO stratum oriens StrP stratumpyramidale StrR stratum radiatum StrLM stratum lacunosum-moleculare and scale bars (a) 20 120583m (b) 40 120583m
While prominent clusters of GLAST and GLT-1 immunore-activity were observed alongGFAP-positive structures in scarcells D-aspartate-induced intracellular sodium signals werestrongly dampened indicating a significant reduction in glu-tamate uptake capacity close to the lesion In the periphery ofthe lesion (100ndash350120583m) astrocytes showed less pronounced
reactivity maintained their ability to efficiently take up SR101and displayed only minor changes in glutamate transporterimmunoreactivity and function Thus depending on theirdistance from the lesion astrocytes showed different gradesof reactivity and displayed discrete changes in glutamatetransporter immunoreactivity and uptake capacity
ISRN Neurology 11
SR101 MergeSBFI
StrRStrP
(a)
03
04
05
06
07
0 5 10 15
0
20 25 30 40 0
100
20
40
80
60
0 10 20 30 400
20
40
60
80
100
10 = 55 mM
107 cells in 6 experiments
F (340
380
) (ra
tio)
Intracellular Na+ concentration (mM)Intracellular Na+ concentration (mM)
ΔF
(340
380
) (
)
ΔF
(340
380
) (
) R2 = 097
(b)
D-aspartate
100 120583M500 120583M
750 120583M1000 120583M
1 mM
10 s
Δ
(c)
0 1 2 3 4 50
1
2
3
4
5
Δ so
dium
(mM
)
D-aspartate (mM)
lowast
(d)
Figure 6 Sodium changes induced by activation of glutamate uptake (a) Staining of astrocytes by colabeling with SR101 and SBFI Leftepifluorescence image of theCA1-area of an SR101-loaded organotypic sliceCenter SBFI fluorescence taken from the same areaRight overlaySBFI-loaded SR101-positive astrocytes appear in white (arrowheads) StrP stratum pyramidale StrR stratum radiatum scale bar 20 120583m (b)Left calibration of SBFI fluorescence in an astrocyte Depicted is the change in intracellular SBFI fluorescence ratio in the calibration solutionscontaining ionophores and in response to stepwise changes in the sodium concentration Right mean values plusmn SEM of the normalizedchanges in SBFI fluorescence at different sodium concentrations The black line represents a linear fit of the data points between 10 and40mM sodium illustrating that a 10 change in SBFI fluorescence represents a change in the sodium concentration by 55mM (c) Sodiumtransients in a single astrocyte induced by 500ms applications of D-aspartate at different concentrations (d) Mean values plusmn S E M (119899 = 19)of the peak amplitude of sodium transients at different concentrations of D-aspartate The curve represents a sigmoidal fit of the data
41 Characteristics of Lesion-Induced Reactive Gliosis inOrganotypic Cultures Organotypic tissue slice cultures ofthe hippocampus represent a well-established model systemin which the basic tissue architecture is maintained butwhich still hosts the major advantages of cell cultures suchas good accessibility and control of experimental conditionsThey have been used extensively for the analysis of neuronal
properties and developing neuronal networks [28 51 52]or the study of excitotoxic neuronal damage (eg [53 54])Recent studies also demonstrated that basic morphologicalcharacteristics of astrocytes and the typical glia-synapseorganization are well preserved in organotypic slice cultures(eg [44 45 55 56]) In addition we found a nearly completeoverlap of SR101 staining with the expression of GFP in slice
12 ISRN Neurology
SBFIControl Lesioned
SR101
1
Lesio
n sit
e
SR101 SBFI
2 45
32 4
5
1
StrR
StrP StrP
StrR
3
(a)
ControlScar
Lesioned
1
D-asp
2
D-asp
D-asp
5
4
3Periphery1mM
20 s
Δ Δ
Δ
Δ
Δ
(b)
0102030405060708090
100110
Δ so
dium
tim
e (m
Ms
)
00
02
04
06
08
10
12
0
1
2
3
4
5
6
ns
Control Periphery Scar Control Periphery Scar Control Periphery Scar
Amplitude Slope Decay time
Δ so
dium
(mM
)
120591(s
)lowastlowastlowast
lowastlowastlowastlowastlowastlowast
lowastlowastlowastlowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowast
(c)
Figure 7 D-aspartate-induced astrocyte sodium transients following lesion (a) CA1 area of a control (left) and a lesioned (right) slicepreparation Shown are epifluorescence images of SR101 and SBFI fluorescences Circled areas and numbers indicate the regions of interestanalyzed in the experiment depicted in (b) StrP stratum pyramidale StrR stratum radiatum and scale bar 40 120583m (b) Sodium transientsinduced by pressure application of 1mMD-aspartate (D-asp) for 500ms in the different cells in the control and lesioned slice as indicated in(a) (c) Histograms showing mean values plusmn S E M of the peak amplitude (left) slope (center) and decay time (right) of D-aspartate-inducedsodium transients in astrocytes in control slices (119899 = 161) as well as in cells in the scar region and its periphery (119899 = 111 and 119899 = 26 resp)lowastlowast119875 lt 001 lowastlowastlowast119875 lt 0001 StrP stratum pyramidale StrR stratum radiatum and scale bar 40120583m
cultures obtained from GFPGFAP mice demonstrating thatSR101 is well suited to identify astrocytes in this prepara-tion as described before for acute tissue slices and in vivo[29 30]
Performing a scratch through the CA1 region of organ-otypic slices resulted in the formation of a glial scar along thelesion site as judged by the prominent increase in expressionof GFAP and S100120573 In addition astrocyte cell bodies showed
a significant hypertrophy Furthermore astrocytes in theperiphery of the scar (100ndash350 120583m from the lesion) showedmild to moderate reactivity displaying swollen cell bodiesand only moderate upregulation of GFAP These changesare hallmarks of reactive gliosis observed after tissue injury[10 13 57] Generally the degree of injury and the distance ofthe astrocytes from the site of injury define the degree of theiractivation [11 13]
ISRN Neurology 13
Our study also revealed that strongly reactive astrocytesin the scar region only weakly stain with SR101 The reducedability of strongly activated astrocytes to accumulate SR101indicates that strong glial activation is accompanied by adownregulation of the organic anion transporter responsi-ble for uptake of SR101 This is reminiscent of immatureastrocytes in the early postnatal hippocampus which areSR101-negative [30] and in line with a wealth of experimentalevidence suggesting that reactive gliosis represents a processcomprising a dedifferentiation of astrocytes [17]
42 Changes in Glutamate Transporter Expression followingLesion Glutamate uptake in the hippocampus is mainlyachieved by the glial glutamate transporters GLT-1 andGLAST [1 2] A central factor in many brain pathologiesincluding traumatic brain injury is an increase in extracellu-lar glutamate and excitotoxicity it has been suggested that achange in the expression levels of glial glutamate transportersmight play a critical role in the failure of glutamate clearance[6 58ndash60] Earlier work has found an overall downregulationof protein levels of both GLAST and GLT-1 following astro-cyte activation [22ndash27] indicating that this is causal to theelevation of extracellular glutamate In contrast to this notionother studies reported increased glutamate transport capacityof reactive astrocytes and suggested a protective influence[61 62]
To visualize possible differences in the spatial expressionprofile of GLT-1 and GLAST in strongly reactive astrocytesalong the scar as compared to moderately reactive astro-cytes in its periphery we performed immunohistochemicalstainings These revealed an accumulation of GLT-1 andGLAST immunoreactivity along thick GFAP-positive fiberswhich was particularly pronounced for GLAST Besides theseclusters overall GLAST immunoreactivity seemed unalteredas compared to unlesioned slices whereas GLT-1 immunore-activity seemedweaker and declined with increasing distancefrom the scar Clustering of glutamate transporters has beendescribed by several reports [48 63 64] In developinghippocampal astrocytes cluster formation was preferentiallyfound in branches opposed to synapses and was increasedwith increased neuronal activity indicating that it is neces-sary to copewith synaptic release of glutamate [36] Along thesame lines it was observed that sustained astroglial activationby ciliary neurotrophic factor (CNTF) in the rat striatuminduced a concentration of GLAST and GLT-1 into raftmicrodomains and improved glutamate clearance indicatingthat cluster formation increased the cellular capacity for glu-tamate uptake [61] However another study found glutamate-induced clustering of GLT-1 that induced its endocytosis andintracellular trafficking without changing the total expressionlevels as detected by western blots arguing for a decrease infunctional glial glutamate uptake capacity [49] In the presentstudy antibodies against GLT-1 and GLAST were employedfor immunohistochemistry after permeabilization of theplasma membrane The clusters of GLAST and GLT-1 alongGFAP-positive fibers in the scar region (as GLAST andGLT-1immunoreactivity in general) could thus represent glutamatetransporters in the plasmamembrane or in intracellular com-partments or both Consequently the observed clustering
does not allow a prediction about possible functional changesin glutamate uptake
43 Lesion-Induced Changes in Functional Glutamate UptakeCapacity We probed for the functional activation of gluta-mate transport by application of the transportable agonistD-aspartate which results in an increase in the intracellularsodium concentration of astrocytes [8 9] The amplitudesof D-aspartate-induced sodium transients in astrocytes incontrol organotypic slices were similar to those reportedfrom astrocytes in acute slices obtained in our lab [33]indicating similar cellular glutamate transport capacity inboth preparations
Following a lesion D-aspartate-induced intracellularsodium signalswere clearly dampened in scar cellsMoreoverthe slope of the increase in sodium was significantly reducedand the decay back to baseline was slowed Because D-aspartate was applied at nearly saturating concentrationthe reduction in peak amplitudes suggests a reduction infunctional glutamate uptake capacity in cells close to thelesion which might be mediated by a decrease in the overallnumber of glutamate transporters available at the plasmamembrane In contrast the alteration in the kinetics of theD-aspartate-induced sodium signals can be explained by thecellular hypertrophy that was observed in reactive astrocytesAt an equal transport-mediated sodium influx across themembrane the slope of resulting changes in the sodiumconcentration will be decreased in cells with a larger volume
Furthermore the decay of intracellular sodium transientsback to baseline is mainly governed by the activity of theNa+K+-ATPase as well as by diffusion [8] The slower recov-ery is in line with earlier studies reporting a downregulationof the sodium pump following reactive gliosis [65 66]Moreover sodium is not buffered in the cell and travels solelyby means of diffusion in the cytoplasm Hindered diffusionresulting from increased cytoplasmic protein content orimpaired gap junction coupling which both occur uponastrocytic activation might thus also partially contribute todelayed sodium recovery [12 34 67]
In contrast to scar cells the amplitude of D-aspartate-induced intracellular sodium signals was not significantlyaltered in moderately activated astrocytes in the periphery ofthe lesion indicating that the overall number of functionalglutamate transporters was not significantly altered Becausethese cells showed hypertrophy as well the slowed kinetics ofthe sodium transients are in line with an increased cellularvolume as well as a possible downregulation of Na+K+-ATPase andor a slowed diffusion as argued above
Taken together our results indicate a significant reduc-tion in glutamate uptake capacity in strongly activated astro-cytes in the scar region Thus the prominent clustering ofGLAST and GLT-1 immunoreactivity along GFAP-positivestructures close to the lesion is likely to reflect a loss ofglutamate transporters from the plasma membrane upontransporter internalization as reported by Nakagawa et al[49] The authors of the latter study speculated that large andprolonged increases in glutamate concentrations are neces-sary to induce such cluster formation and endocytosis in thetissue as they would possibly only occur under pathological
14 ISRN Neurology
conditions Indeed ambient glutamate concentrations wereshown to be significantly increased after traumatic braininjury [68ndash70] Along these lines it can be assumed thatin our model system extracellular glutamate concentrationsrose less severely in the periphery of the lesion preventinga comparable clustering and loss of functional glutamatetransport activity in moderately activated astrocytes
5 Conclusions
Our results show complex changes in glutamate transporterexpression and function during astrocyte activation followingmechanical injury They confirm that the degree of astrocyteactivation depends on the distance from the insult and com-prises different endpoints regarding cellular morphology andphysiology Furthermore our results highlight that astrocyteswhich show different grades of reactivity also display discretechanges in glutamate transporter expression and functionWhile immunohistochemistry revealed a prominent clus-tering of GLT-1 and GLAST immunoreactivity in the scarregion our functional assay clearly showed that glutamateuptake capacity is strongly reduced in scar cells while itis largely maintained in moderately activated astrocytes inthe periphery Thus mild-to-moderate astrogliosis in theperiphery of a mechanical lesion does not necessarily seemto be accompanied by a significant change in glial glutamateuptake capacity At the glial scar itself in strongly reactiveastrocytes a clustering of glutamate transporters is observedthat apparently goes along with a severe functional reductionin astroglial glutamate uptake which may contribute toglutamate-mediated excitotoxicity in this region
Conflict of Interests
The authors of the paper declare that they do not haveany direct financial relation with the commercial identitiesmentioned in the paper that might lead to a conflict ofinterests
Acknowledgments
This study was supported by the Deutsche Forschungsge-meinschaft (DFG Ro23274-3) The authors thank SimoneDurry and Claudia Roderigo for expert technical help
References
[1] N C Danbolt ldquoGlutamate uptakerdquo Progress in Neurobiologyvol 65 no 1 pp 1ndash105 2001
[2] A V Tzingounis and J I Wadiche ldquoGlutamate transportersconfining runaway excitation by shaping synaptic transmis-sionrdquo Nature Reviews Neuroscience vol 8 no 12 pp 935ndash9472007
[3] D E Bergles J S Diamond and C E Jahr ldquoClearance ofglutamate inside the synapse and beyondrdquo Current Opinion inNeurobiology vol 9 no 3 pp 293ndash298 1999
[4] C M Anderson and R A Swanson ldquoAstrocyte glutamatetransport review of properties regulation and physiologicalfunctionsrdquo Glia vol 32 no 1 pp 1ndash14 2000
[5] P Marcaggi and D Attwell ldquoRole of glial amino acid trans-porters in synaptic transmission and brain energeticsrdquoGlia vol47 no 3 pp 217ndash225 2004
[6] N J Maragakis and J D Rothstein ldquoGlutamate transportersanimal models to neurologic diseaserdquo Neurobiology of Diseasevol 15 no 3 pp 461ndash473 2004
[7] G Gegelashvili and A Schousboe ldquoCellular distribution andkinetic properties of high-affinity glutamate transportersrdquoBrainResearch Bulletin vol 45 no 3 pp 233ndash238 1998
[8] J W Deitmer and C R Rose ldquoIon changes and signalling inperisynaptic gliardquo Brain Research Reviews vol 63 no 1-2 pp113ndash129 2010
[9] S Kirischuk V Parpura and A Verkhratsky ldquoSodium dynam-ics another key to astroglial excitabilityrdquo Trends in Neuro-sciences vol 35 no 8 pp 497ndash506 2012
[10] M V Sofroniew ldquoMolecular dissection of reactive astrogliosisand glial scar formationrdquo Trends in Neurosciences vol 32 no12 pp 638ndash647 2009
[11] J L Ridet S K Malhotra A Privat and F H Gage ldquoReactiveastrocytes cellular and molecular cues to biological functionrdquoTrends in Neurosciences vol 20 no 12 pp 570ndash577 1997
[12] J Middeldorp and E M Hol ldquoGFAP in health and diseaserdquoProgress in Neurobiology vol 93 no 3 pp 421ndash443 2011
[13] M Pekny and M Nilsson ldquoAstrocyte activation and reactivegliosisrdquo Glia vol 50 no 4 pp 427ndash434 2005
[14] A Bordey S A Lyons J J Hablitz and H SontheimerldquoElectrophysiological characteristics of reactive astrocytes inexperimental cortical dysplasiardquo Journal of Neurophysiologyvol 85 no 4 pp 1719ndash1731 2001
[15] R Jabs G Seifert and C Steinhauser ldquoAstrocytic functionand its alteration in the epileptic brainrdquo Epilepsia vol 49supplement 2 pp 3ndash12 2008
[16] A Buffo I Rite P Tripathi et al ldquoOrigin and progeny ofreactive gliosis a source of multipotent cells in the injuredbrainrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 105 no 9 pp 3581ndash3586 2008
[17] S Robel B Berninger and M Gotz ldquoThe stem cell potential ofglia lessons from reactive gliosisrdquoNature Reviews Neurosciencevol 12 no 2 pp 88ndash104 2011
[18] H Kawano J Kimura-Kuroda Y Komuta et al ldquoRole of thelesion scar in the response to damage and repair of the centralnervous systemrdquo Cell and Tissue Research vol 349 no 1 pp169ndash180 2012
[19] G Yiu and Z He ldquoGlial inhibition of CNS axon regenerationrdquoNature Reviews Neuroscience vol 7 no 8 pp 617ndash627 2006
[20] M T Fitch and J Silver ldquoCNS injury glial scars and inflamma-tion inhibitory extracellular matrices and regeneration failurerdquoExperimental Neurology vol 209 no 2 pp 294ndash301 2008
[21] L Li A Lundkvist D Andersson et al ldquoProtective role ofreactive astrocytes in brain ischemiardquo Journal of Cerebral BloodFlow amp Metabolism vol 28 no 3 pp 468ndash481 2008
[22] F K H van Landeghem T Weiss M Oehmichen and AVonDeimling ldquoDecreased expression of glutamate transportersin astrocytes after human traumatic brain injuryrdquo Journal ofNeurotrauma vol 23 no 10 pp 1518ndash1528 2006
[23] V L Raghavendra Rao M K Baskaya A Dogan J DRothstein and R J Dempsey ldquoTraumatic brain injury down-regulates glial glutamate transporter (GLT- 1 and GLAST)proteins in rat brainrdquo Journal of Neurochemistry vol 70 no 5pp 2020ndash2027 1998
ISRN Neurology 15
[24] J-C Chen H Hsu-Chou J-L Lu et al ldquoDown-regulation ofthe glial glutamate transporter GLT-1 in rat hippocampus andstriatum and its modulation by a group III metabotropic glu-tamate receptor antagonist following transient global forebrainischemiardquoNeuropharmacology vol 49 no 5 pp 703ndash714 2005
[25] S D Ginsberg L J Martin and J D Rothstein ldquoRegional deaf-ferentation down-regulates subtypes of glutamate transporterproteinsrdquo Journal of Neurochemistry vol 65 no 6 pp 2800ndash2803 1995
[26] J E Springer R D Azbill R J Mark J G Begley GWaeg andM P Mattson ldquo4-Hydroxynonenal a lipid peroxidation prod-uct rapidly accumulates following traumatic spinal cord injuryand inhibits glutamate uptakerdquo Journal of Neurochemistry vol68 no 6 pp 2469ndash2476 1997
[27] M B Moretto N S Arteni D Lavinsky et al ldquoHypoxic-ischemic insult decreases glutamate uptake by hippocampalslices from neonatal rats Prevention by guanosinerdquo Experimen-tal Neurology vol 195 no 2 pp 400ndash406 2005
[28] L Stoppini P-A Buchs and D Muller ldquoA simple method fororganotypic cultures of nervous tissuerdquo Journal of NeuroscienceMethods vol 37 no 2 pp 173ndash182 1991
[29] A Nimmerjahn F Kirchhoff J N D Kerr and F HelmchenldquoSulforhodamine 101 as a specific marker of astroglia in theneocortex in vivordquo Nat Methods vol 1 no 1 pp 31ndash37 2004
[30] K W Kafitz S D Meier J Stephan and C R Rose ldquoDevel-opmental profile and properties of sulforhodamine 101-Labeledglial cells in acute brain slices of rat hippocampusrdquo Journal ofNeuroscience Methods vol 169 no 1 pp 84ndash92 2008
[31] C Schnell Y Hagos and S Hulsmann ldquoActive sulforhodamine101 uptake into hippocampal astrocytesrdquo PLoS One vol 7 no 11Article ID e49398 2012
[32] S D Meier Y Kovalchuk and C R Rose ldquoProperties ofthe new fluorescent Na+ indicator CoroNa Green comparisonwith SBFI and confocal Na+ imagingrdquo Journal of NeuroscienceMethods vol 155 no 2 pp 251ndash259 2006
[33] J Langer and C R Rose ldquoSynaptically induced sodium signalsin hippocampal astrocytes in siturdquo Journal of Physiology vol587 part 24 pp 5859ndash5877 2009
[34] J Langer J Stephan M Theis and C R Rose ldquoGap junctionsmediate intercellular spread of sodium between hippocampalastrocytes in siturdquo Glia vol 60 no 2 pp 239ndash252 2012
[35] C R Rose andB R Ransom ldquoIntracellular sodiumhomeostasisin rat hippocampal astrocytesrdquo Journal of Physiology vol 491part 2 no 2 pp 291ndash305 1996
[36] A M Benediktsson G S Marrs J C Tu et al ldquoNeuronalactivity regulates glutamate transporter dynamics in developingastrocytesrdquo Glia vol 60 no 2 pp 175ndash188 2012
[37] M Zhou G P Schools and H K Kimelberg ldquoDevelopment ofGLAST(+) astrocytes andNG2(+) glia in rat hippocampus CA1mature astrocytes are electrophysiologically passiverdquo Journal ofNeurophysiology vol 95 no 1 pp 134ndash143 2006
[38] I Suarez G Bodega and B Fernandez ldquoModulation of glu-tamate transporters (GLAST GLT-1 and EAAC1) in the ratcerebellum following portocaval anastomosisrdquo Brain Researchvol 859 no 2 pp 293ndash302 2000
[39] X Zhu D E Bergles and A Nishiyama ldquoNG2 cells generateboth oligodendrocytes and gray matter astrocytesrdquo Develop-ment vol 135 no 1 pp 145ndash157 2008
[40] B Brunne S ZhaoADerouiche et al ldquoOriginmaturation andastroglial transformation of secondary radial glial cells in thedeveloping dentate gyrusrdquo Glia vol 58 no 13 pp 1553ndash15692010
[41] Y Wu A-Q Zhang and D T Yew ldquoAge related changes ofvarious markers of astrocytes in senescence-accelerated micehippocampusrdquo Neurochemistry International vol 46 no 7 pp565ndash574 2005
[42] S Magavi D Friedmann G Banks A Stolfi and C LoisldquoCoincident generation of pyramidal neurons and protoplasmicastrocytes in neocortical columnsrdquo Journal of Neuroscience vol32 no 14 pp 4762ndash4772 2012
[43] M Olabarria H N Noristani A Verkhratsky and J JRodrıguez ldquoConcomitant astroglial atrophy and astrogliosis ina triple transgenic animal model of Alzheimerrsquos diseaserdquo Gliavol 58 no 7 pp 831ndash838 2010
[44] A M Benediktsson S J Schachtele S H Green and M EDailey ldquoBallistic labeling and dynamic imaging of astrocytesin organotypic hippocampal slice culturesrdquo Journal of Neuro-science Methods vol 141 no 1 pp 41ndash53 2005
[45] I Lushnikova G Skibo D Muller and I Nikonenko ldquoSynapticpotentiation induces increased glial coverage of excitatorysynapses in CA1 hippocampusrdquoHippocampus vol 19 no 8 pp753ndash762 2009
[46] E Raponi F Agenes C Delphin et al ldquoS100B expressiondefines a state in which GFAP-expressing cells lose their neuralstem cell potential and acquire a more mature developmentalstagerdquo Glia vol 55 no 2 pp 165ndash177 2007
[47] C G Schipke C Boucsein C Ohlemeyer F Kirchhoff andH Kettenmann ldquoAstrocyte Ca2+ waves trigger responses inmicroglial cells in brain slicesrdquo The FASEB Journal vol 16 no2 pp 255ndash257 2002
[48] J Zhou and M L Sutherland ldquoGlutamate transporter clusterformation in astrocytic processes regulates glutamate uptakeactivityrdquo Journal of Neuroscience vol 24 no 28 pp 6301ndash63062004
[49] T Nakagawa Y Otsubo Y Yatani H Shirakawa and S KanekoldquoMechanisms of substrate transport-induced clustering of aglial glutamate transporter GLT-1 in astroglial-neuronal cul-turesrdquo European Journal of Neuroscience vol 28 no 9 pp 1719ndash1730 2008
[50] J-Y Chatton P Marquet and P J Magistretti ldquoA quantitativeanalysis of L-glutamate-regulated Na+ dynamics in mousecortical astrocytes Implications for cellular bioenergeticsrdquoEuropean Journal of Neuroscience vol 12 no 11 pp 3843ndash38532000
[51] E Forster S Zhao and M Frotscher ldquoLaminating the hip-pocampusrdquo Nature Reviews Neuroscience vol 7 no 4 pp 259ndash267 2006
[52] B H Gahwiler M Capogna D Debanne R A McKinney andS M Thompson ldquoOrganotypic slice cultures a technique hascome of agerdquo Trends in Neurosciences vol 20 no 10 pp 471ndash477 1997
[53] P E Kunkler and R P Kraig ldquoReactive astrocytosis fromexcitotoxic injury in hippocampal organ culture parallels thatseen in vivordquo Journal of Cerebral Blood Flow and Metabolismvol 17 no 1 pp 26ndash43 1997
[54] I E Holopainen ldquoOrganotypic hippocampal slice cultures amodel system to study basic cellular andmolecularmechanismsof neuronal cell death neuroprotection and synaptic plasticityrdquoNeurochemical Research vol 30 no 12 pp 1521ndash1528 2005
[55] MHaber L Zhou andK KMurai ldquoCooperative astrocyte anddendritic spine dynamics at hippocampal excitatory synapsesrdquoJournal of Neuroscience vol 26 no 35 pp 8881ndash8891 2006
16 ISRN Neurology
[56] H Nishida and S Okabe ldquoDirect astrocytic contacts regulatelocal maturation of dendritic spinesrdquo Journal of Neurosciencevol 27 no 2 pp 331ndash340 2007
[57] A Buffo C Rolando and S Ceruti ldquoAstrocytes in the dam-aged brain molecular and cellular insights into their reactiveresponse and healing potentialrdquoBiochemical Pharmacology vol79 no 2 pp 77ndash89 2010
[58] J-H Yi and A S Hazell ldquoExcitotoxic mechanisms and the roleof astrocytic glutamate transporters in traumatic brain injuryrdquoNeurochemistry International vol 48 no 5 pp 394ndash403 2006
[59] C Werner and K Engelhard ldquoPathophysiology of traumaticbrain injuryrdquo British Journal of Anaesthesia vol 99 no 1 pp4ndash9 2007
[60] G Gegelashvili M B Robinson D Trotti and T RauenldquoRegulation of glutamate transporters in health and diseaserdquoProgress in Brain Research vol 132 pp 267ndash286 2001
[61] C Escartin E Brouillet PGubellini et al ldquoCiliary neurotrophicfactor activates astrocytes redistributes their glutamate trans-porters GLAST and GLT-1 to raft microdomains and improvesglutamate handling in vivordquo Journal of Neuroscience vol 26 no22 pp 5978ndash5989 2006
[62] C Vermeiren M Najimi N Vanhoutte et al ldquoAcute up-regulation of glutamate uptake mediated by mGluR5a in reac-tive astrocytesrdquo Journal of Neurochemistry vol 94 no 2 pp405ndash416 2005
[63] C L Poitry-Yamate L Vutskits and T Rauen ldquoNeuronal-induced and glutamate-dependent activation of glial glutamatetransporter functionrdquo Journal of Neurochemistry vol 82 no 4pp 987ndash997 2002
[64] P M Beart and R D OrsquoShea ldquoTransporters for L-glutamatean update on their molecular pharmacology and pathologicalinvolvementrdquo British Journal of Pharmacology vol 150 no 1pp 5ndash17 2007
[65] F D Lima M A Souza A F Furian et al ldquoNa+K+-ATPaseactivity impairment after experimental traumatic brain injuryrelationship to spatial learning deficits and oxidative stressrdquoBehavioural Brain Research vol 193 no 2 pp 306ndash310 2008
[66] G A Gusarova H E Trejo L A Dada et al ldquoHypoxia leadsto NaK-ATPase downregulation via Ca2+ release-activatedCa2+ channels and AMPK activationrdquo Molecular and CellularBiology vol 31 no 17 pp 3546ndash3556 2011
[67] S S L Chew C S Johnson C R Green and H V Danesh-Meyer ldquoRole of connexin43 in central nervous system injuryrdquoExperimental Neurology vol 225 no 2 pp 250ndash261 2010
[68] P Nilsson L Hillered U Ponten and U Ungerstedt ldquoChangesin cortical extracellular levels of energy-related metabolites andamino acids following concussive brain injury in ratsrdquo Journal ofCerebral Blood Flow and Metabolism vol 10 no 5 pp 631ndash6371990
[69] H Katoh K Sima H Nawashiro K Wada and H ChigasakildquoThe effect of MK-801 on extracellular neuroactive amino acidsin hippocampus after closed head injury followed by hypoxia inratsrdquo Brain Research vol 758 no 1-2 pp 153ndash162 1997
[70] H Koizumi H Fujisawa H Ito T Maekawa X Di and RBullock ldquoEffects of mild hypothermia on cerebral blood flow-independent changes in cortical extracellular levels of aminoacids following contusion trauma in the ratrdquoBrain Research vol747 no 2 pp 304ndash312 1997
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
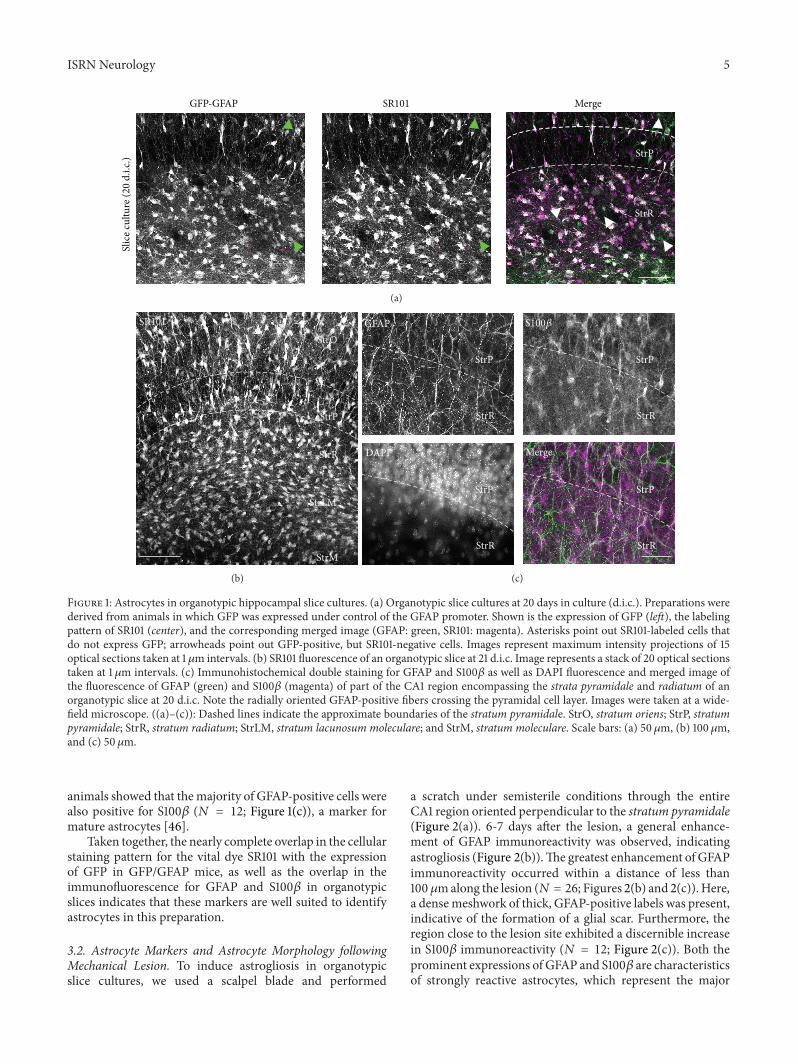
ISRN Neurology 5
Slic
e cul
ture
(20
dic
)
GFP-GFAP MergeSR101
lowast lowast lowast lowast
lowast
lowast
lowast lowast
StrP
StrR
(a)
StrR
StrP
StrLM
StrM
SR101
StrO
(b)
GFAP S100120573
StrP
StrR
StrP
StrR
StrP
StrR
StrP
StrR
DAPI Merge
(c)
Figure 1 Astrocytes in organotypic hippocampal slice cultures (a) Organotypic slice cultures at 20 days in culture (dic) Preparations werederived from animals in which GFP was expressed under control of the GFAP promoter Shown is the expression of GFP (left) the labelingpattern of SR101 (center) and the corresponding merged image (GFAP green SR101 magenta) Asterisks point out SR101-labeled cells thatdo not express GFP arrowheads point out GFP-positive but SR101-negative cells Images represent maximum intensity projections of 15optical sections taken at 1120583m intervals (b) SR101 fluorescence of an organotypic slice at 21 dic Image represents a stack of 20 optical sectionstaken at 1 120583m intervals (c) Immunohistochemical double staining for GFAP and S100120573 as well as DAPI fluorescence and merged image ofthe fluorescence of GFAP (green) and S100120573 (magenta) of part of the CA1 region encompassing the strata pyramidale and radiatum of anorganotypic slice at 20 dic Note the radially oriented GFAP-positive fibers crossing the pyramidal cell layer Images were taken at a wide-field microscope ((a)ndash(c)) Dashed lines indicate the approximate boundaries of the stratum pyramidale StrO stratum oriens StrP stratumpyramidale StrR stratum radiatum StrLM stratum lacunosum moleculare and StrM stratum moleculare Scale bars (a) 50 120583m (b) 100120583mand (c) 50 120583m
animals showed that themajority of GFAP-positive cells werealso positive for S100120573 (119873 = 12 Figure 1(c)) a marker formature astrocytes [46]
Taken together the nearly complete overlap in the cellularstaining pattern for the vital dye SR101 with the expressionof GFP in GFPGFAP mice as well as the overlap in theimmunofluorescence for GFAP and S100120573 in organotypicslices indicates that these markers are well suited to identifyastrocytes in this preparation
32 Astrocyte Markers and Astrocyte Morphology followingMechanical Lesion To induce astrogliosis in organotypicslice cultures we used a scalpel blade and performed
a scratch under semisterile conditions through the entireCA1 region oriented perpendicular to the stratum pyramidale(Figure 2(a)) 6-7 days after the lesion a general enhance-ment of GFAP immunoreactivity was observed indicatingastrogliosis (Figure 2(b))The greatest enhancement of GFAPimmunoreactivity occurred within a distance of less than100 120583malong the lesion (119873 = 26 Figures 2(b) and 2(c)) Herea densemeshwork of thick GFAP-positive labels was presentindicative of the formation of a glial scar Furthermore theregion close to the lesion site exhibited a discernible increasein S100120573 immunoreactivity (119873 = 12 Figure 2(c)) Both theprominent expressions of GFAP and S100120573 are characteristicsof strongly reactive astrocytes which represent the major
6 ISRN Neurology
GFAP
DAPI
CA1 subregion
GFAP
DAPI
StrR
StrP StrP
StrR
StrOStrO
StrO
StrP
StrR
StrO
StrP
StrR
Control Lesioned
GFAP
Lesio
n sit
e
Hi
StrMStrG
StrP
StrR
StrLM
Scratch wound
StrO
Lesio
n sit
e
Lesioned
(a) (c)
(b)
350120583m350120583m
S100120573
Figure 2 Lesion-induced reactive gliosis in organotypic slice cultures (a) Schematic overview of the hippocampus indicating the positionof the scratch woundThe box delineates the area primarily analyzed (b) GFAP immunofluorescence and DAPI staining of the CA1 area in acontrol (left) and a lesioned (right) slice The boxes indicate the areas enlarged below The dashed line in the lower right indicates the lesion(c) Double staining for GFAP (top) and S100120573 (bottom) adjacent to the lesion site Note the gradual increase in GFAP and S100120573 expressiontowards the lesion (b) widefield microscopy (c) confocal microscopy (optical sections taken at 1120583m intervals) StrO stratum oriens StrPstratum pyramidale StrR stratum radiatum and StrLM stratum lacunosum moleculare Scale bars (b) 200 120583m (top) and 50 120583m (bottom) (c)10120583m
ISRN Neurology 7
StrR
StrP
StrLM
StrO
StrP StrRStrO StrLM
Lesio
n sit
e
SR101-staining pattern
(a)
SBFI
Lesio
n sit
e
SR101 GFAP-GFP
(b)
Figure 3 SR101 labeling in lesioned slices (a) SR101 fluorescence in the CA1 area in a slice 7 days after lesion The delineated area comprisesthe stratum pyramidale and the box indicates the area enlarged on the right side Right SR101 fluorescence at the lesion site illustrating thatSR101 labeling is reduced along the lesionThe dotted line indicates the transition between the scar area and the adjacent moderately reactivetissue The boxed area is further enlarged at the bottom (b) Image series obtained at a wide-field fluorescence microscope The slice wasprepared from a transgenic GFPGFAP mouse and the lesion site is indicated by the dotted line Left Image of SR101 fluorescence centerGFP fluorescence indicating GFAP expression and right SBFI fluorescence SR101 labeling is only weak in GFPSBFI-positive cells in the scarregion (see arrowheads) StrO stratum oriens StrP stratum pyramidale StrR stratum radiatum and StrLM stratum lacunosum-moleculareScale bars 100120583m ((a) upper left) and 20 120583m (other images)
cellular component of the glial scar forming after mechanicaltissue injury [10] More distal to the lesion (gt100ndash350 120583m) aless dramatic rearrangement of GFAP immunoreactivity wasobserved
A clear difference in astrocyte properties between thescar region and its periphery was also seen following SR101labeling In the periphery of the scar (100ndash350 120583m distancefrom the lesion) SR101 resulted in a reliable and brightcellular staining similar to what had been observed incontrol unlesioned slices (119873 = 12 Figure 3(a) cf Figure 1)
In contrast to this cellular SR101 staining was detectable butonly faint within a distance of less than 100 120583m on both sidesof the lesion (Figure 3(a) meanwidth of SR101-free area 111plusmn3 120583m 119873 = 5) In slices from GFPGFAP transgenic animals(119873 = 5 Figure 3(b)) GFAP-positive weakly SR101-positivescar cells also stained with the sodium-sensitive fluorescenceindicator SBFI-AM (acetoxymethyl ester of sodium bindingbenzofuran isophthalate see below 119873 = 3) indicatingthat they were able to take up and de-esterify this dye(Figure 3(b))
8 ISRN Neurology
lowastlowastlowast
lowastlowastlowast
80
lowast
Control ScarPeriphery
120
160
0
40
200
Som
a siz
e (120583
m2)
(a)
Lesioned
StrP
StrR
StrP
Control
StrR
StrLM
Lesio
n sit
e
(b)
Figure 4 Lesion-induced changes in somatic area (a) Quantification of somatic area based on GFAPGFP fluorescence in control andlesioned slices The soma size increased both in astrocytes in the scar region (ldquoscarrdquo) and in astrocytes in the periphery of the lesion(ldquoperipheryrdquo lowastlowastlowast119875 lt 0001 lowast119875 lt 005) (b) Confocal images of GFP fluorescence in cultured slices obtained from GFPGFAP transgenicanimals under control conditions (left) and subjected to a lesion (right) Cells surrounded by boxes are shown at higher magnification in theinsets and the dashed lines around the soma illustrate the areas which served to determine their size StrP stratum pyramidale StrR stratumradiatum StrLM stratum lacunosum moleculare and scale bars 40 120583m and 10120583m (inset)
In addition to GFAP-positive cells the scar region hostedmicroglial cells identified by their vital staining with TexasRed-coupled lectin from lycopersicon esculentum (tomatolectin119873 = 4 not shown) Tomato lectin-positive microgliahowever did not take up SBFI-AM (119873 = 4 not shown) con-firming earlier reports thatmicroglial cells do not incorporateAM-ester dyes in situ [47]
To further characterize reactive gliosis in response to thescratch wound we determined the area of astrocyte somatain confocal images of the GFP fluorescence in control andlesioned slices obtained fromGFPGFAP transgenic animalsThesemeasurements revealed that compared to astrocytes in
control slices the soma area of astrocytes in the periphery ofa lesion was increased by 18 and that of astrocytes in thescar regionwas increased by 24 (control 163plusmn2 120583m2 lesionperiphery 193plusmn3 120583m2 scar tissue 201plusmn4 120583m2119875 lt 0001 119899 =945 496 and 439 in 7 control and 9 lesioned slices Figures4(a) and 4(b)) Thus astrocytes show a second hallmark ofreactive gliosis after lesion namely a hypertrophy of their cellbodies which is observed both in the scar tissue as well as inthe periphery of the lesion
Taken together these data show that 6-7 days after themechanical lesion a glial scar has formed along the lesionsite This scar encompasses strongly activated astrocytes
ISRN Neurology 9
characterized by a robust increase in GFAP and S100120573expressionThese proximal reactive astrocytes locatedwithina distance of less than 100120583m from the lesion exhibit longGFAP-positive processes have significantly swollen somataand only weakly stain with SR101 Astrocytes in the peripheryof the scar (100ndash350 120583m from the lesion) display swollen cellbodies as well but show onlymoderate upregulation of GFAPand maintain their ability to efficiently take up SR101
33 Expression Levels and Spatial Distribution of GLT-1 andGLAST To determine the spatial distribution of the gluta-mate transporters GLAST and GLT-1 in organotypic slicesand changes therein in response to the lesion immunohisto-chemistry was employed In control slices GLAST (119873 = 13Figure 5(a)) as well as GLT-1 immunoreactivity (119873 = 11Figure 5(b)) appeared as punctate labeling throughout theentire CA1 area The staining patterns for both transportersin the tissue slice cultures were thus similar to those reportedfrom acute tissue slice preparations of the hippocampus [3648 49]
The relatively homogeneous distribution of immunore-activity throughout the preparation found in control slicecultures was abandoned following a lesion Here distinctaccumulations of GLAST (119873 = 17 Figure 5(a)) and GLT-1immunoreactivity (119873 = 12 Figure 5(b)) were found in thescar region In both cases these preferentially colocalizedwith thick GFAP-positive bundles running in parallel to thelesion (Figure 5) Apart from the marked accumulation ofGLAST reactivity in parallel to the scar GLAST immunore-activity appeared generally increased in some lesioned slicepreparations (Figure 5(a)) but unaltered in others bothwithinthe scar as well as in its periphery For GLT-1 a decline in theintensity of immunoreactivity was observed with increasingdistance from the lesion (Figure 5(b))
In summary these results reveal distinct alterations inthe immunoreactivity of GLAST and GLT-1 following amechanical lesion In the scar region clusters of GLT-1 andGLAST immunoreactivity are present primarily colocalizedwith strongly GFAP-positive fibers Apart from these distinctclusters GLT-1 immunoreactivity appears to decline withdistance from the scar whereas GLAST immunoreactivityseems largely uniform throughout the preparation
34 Sodium Transients Induced by Activation of GlutamateUptake Glutamate uptake is accompanied by the inwardflux of sodium resulting in increase in the intracellularsodium concentration of astrocytes To monitor its functionwe assayed sodium increases induced by D-aspartate anonmetabolized substrate of sodium-dependent glutamateuptake which is also transported [1] in astrocytes in thestratum radiatum of organotypic slices To this end SR101-stained slices were additionally loaded with the sodium-sensitive fluorophore SBFI-AM (Figure 6(a) cf Figure 3(b))In situ calibrations of the SBFI fluorescence revealed a linearincrease in its fluorescence ratio at intracellular sodiumconcentrations between 10 and 40mM (119899 = 107 Figure 6(b))as reported before [33]
Repetitive pressure application of D-aspartate for 500msinduced reliable increases in the intracellular sodium con-centration in astrocytes indicating activation of sodium-dependent glutamate uptake (119899 = 488 Figure 6(c)) Theexperiments were performed in the presence of the sodium-channel blocker TTX (tetrodotoxin 05 120583M) to suppressaction potential generation The amplitude of D-aspartate-induced sodium increases was dependent on the concen-tration applied (Figure 6(c)) and saturated at about 2mM(119899 = 19 Figure 6(d)) as reported before for mouse corticalastrocytes in culture [50] At a concentration of 1mM theamplitude of D-aspartate-induced sodium transients wasnear maximum and amounted to 38 plusmn 02mM (119899 =161 Figures 6(c) and 6(d)) a value comparable to that inastrocytes in acute slices obtained in our lab [33]
To probe for changes in glutamate uptake we comparedsodium signals induced by 1mMD-aspartate between controland lesioned slices (Figure 7) In lesioned slices astrocyteslocated in the periphery of the lesion (100ndash350 120583m distance)responded with a sodium transient to application of 1mMD-aspartate (see cell 2 in Figures 7(a) and 7(b)) The amplitudeof D-aspartate-induced sodium signals in cells in the lesionperiphery was similar to that observed in control slices (38 plusmn03mM 119899 = 111 see cell 1 in Figures 7(a) 7(b) and 7(c))Application of D-aspartate also evoked sodium transientsin weakly SR101-positive SBFI-filled astrocytes located inthe glial scar area (lt100 120583m from the lesion see cells 3ndash5in Figures 7(a) and 7(b)) These cells however displayedsignificantly smaller peak amplitudes (16 plusmn 021mM 119899 =26 119875 lt 0001 Figures 7(b) and 7(c)) than astrocytes inthe periphery and in control slices
The kinetics of D-aspartate-induced sodium transientsdiffered between all three groups This was true for theslope of the sodium increase (10ndash90 change) which wassignificantly reduced in astrocytes from lesioned slices versusastrocytes from control slices (09 plusmn 005mMs in controlversus 06 plusmn 006mMs in the periphery and 02 plusmn 004mMsin the scar region 119899 = 161 111 and 26 resp 119875 lt 0001Figures 7(b) and 7(c)) In addition the decay back to baselinewas significantly slower following lesion (Figures 7(b) and7(c)) The time needed for the signal to decay by 67 wasapproximately doubled (decay time 120591 was 44 plusmn 2 s in control120591 = 81 plusmn 4 s in periphery of the lesion and 120591 = 95 plusmn 10 s inscar cells Figures 7(b) and 7(c))
These results show that mechanical injury results indistinct alterations in the kinetics andor amplitudes ofD-aspartate-induced intracellular sodium transients in reac-tive astrocytes depending on their distance to the lesion
4 Discussion
Our results show that within 6-7 days after setting a scratchwound in organotypic hippocampal slices a dense glial scarhad formed along the lesion site Up to approximately 100 120583mdistance from the lesion the scar tissue was characterized bya substantial elevation ofGFAP expression a pronounced cel-lular hypertrophy and reduced uptake of SR101 by astrocytes
10 ISRN Neurology
Con
trol
GLAST
Lesio
ned
GFAP Merge
StrR
StrP
StrO
Lesio
n sit
e
StrR
(a)
Con
trol
Lesio
ned
GLT-1 GFAP Merge
StrR
StrP
StrO
StrR
StrP
StrLM
Lesio
n sit
e
(b)
Figure 5 GLAST andGLT-1 immunoreactivity aftermechanical injury (a) Fluorescence images of immunohistochemical stainings detectingthe glutamate transporters GLAST (left) GFAP (center) as counterstaining and the corresponding merged image (GFAP green GLASTmagenta) in control (top row) and lesioned slice cultures (bottom row) (b) GLT-1 (left) GFAP (center) double labeling of a control (toprow) and a mechanically lesioned slice (bottom row) The corresponding merge (GFAP green GLT-1 magenta) is found on the right Notethat after lesion glutamate transporter immunoreactive clusters are accumulated in the direct vicinity of the lesion site (arrowheads) GFAPglial fibrillary acidic protein GLAST glutamate aspartate transporter GLT-1 glutamate-transporter-1 StrO stratum oriens StrP stratumpyramidale StrR stratum radiatum StrLM stratum lacunosum-moleculare and scale bars (a) 20 120583m (b) 40 120583m
While prominent clusters of GLAST and GLT-1 immunore-activity were observed alongGFAP-positive structures in scarcells D-aspartate-induced intracellular sodium signals werestrongly dampened indicating a significant reduction in glu-tamate uptake capacity close to the lesion In the periphery ofthe lesion (100ndash350120583m) astrocytes showed less pronounced
reactivity maintained their ability to efficiently take up SR101and displayed only minor changes in glutamate transporterimmunoreactivity and function Thus depending on theirdistance from the lesion astrocytes showed different gradesof reactivity and displayed discrete changes in glutamatetransporter immunoreactivity and uptake capacity
ISRN Neurology 11
SR101 MergeSBFI
StrRStrP
(a)
03
04
05
06
07
0 5 10 15
0
20 25 30 40 0
100
20
40
80
60
0 10 20 30 400
20
40
60
80
100
10 = 55 mM
107 cells in 6 experiments
F (340
380
) (ra
tio)
Intracellular Na+ concentration (mM)Intracellular Na+ concentration (mM)
ΔF
(340
380
) (
)
ΔF
(340
380
) (
) R2 = 097
(b)
D-aspartate
100 120583M500 120583M
750 120583M1000 120583M
1 mM
10 s
Δ
(c)
0 1 2 3 4 50
1
2
3
4
5
Δ so
dium
(mM
)
D-aspartate (mM)
lowast
(d)
Figure 6 Sodium changes induced by activation of glutamate uptake (a) Staining of astrocytes by colabeling with SR101 and SBFI Leftepifluorescence image of theCA1-area of an SR101-loaded organotypic sliceCenter SBFI fluorescence taken from the same areaRight overlaySBFI-loaded SR101-positive astrocytes appear in white (arrowheads) StrP stratum pyramidale StrR stratum radiatum scale bar 20 120583m (b)Left calibration of SBFI fluorescence in an astrocyte Depicted is the change in intracellular SBFI fluorescence ratio in the calibration solutionscontaining ionophores and in response to stepwise changes in the sodium concentration Right mean values plusmn SEM of the normalizedchanges in SBFI fluorescence at different sodium concentrations The black line represents a linear fit of the data points between 10 and40mM sodium illustrating that a 10 change in SBFI fluorescence represents a change in the sodium concentration by 55mM (c) Sodiumtransients in a single astrocyte induced by 500ms applications of D-aspartate at different concentrations (d) Mean values plusmn S E M (119899 = 19)of the peak amplitude of sodium transients at different concentrations of D-aspartate The curve represents a sigmoidal fit of the data
41 Characteristics of Lesion-Induced Reactive Gliosis inOrganotypic Cultures Organotypic tissue slice cultures ofthe hippocampus represent a well-established model systemin which the basic tissue architecture is maintained butwhich still hosts the major advantages of cell cultures suchas good accessibility and control of experimental conditionsThey have been used extensively for the analysis of neuronal
properties and developing neuronal networks [28 51 52]or the study of excitotoxic neuronal damage (eg [53 54])Recent studies also demonstrated that basic morphologicalcharacteristics of astrocytes and the typical glia-synapseorganization are well preserved in organotypic slice cultures(eg [44 45 55 56]) In addition we found a nearly completeoverlap of SR101 staining with the expression of GFP in slice
12 ISRN Neurology
SBFIControl Lesioned
SR101
1
Lesio
n sit
e
SR101 SBFI
2 45
32 4
5
1
StrR
StrP StrP
StrR
3
(a)
ControlScar
Lesioned
1
D-asp
2
D-asp
D-asp
5
4
3Periphery1mM
20 s
Δ Δ
Δ
Δ
Δ
(b)
0102030405060708090
100110
Δ so
dium
tim
e (m
Ms
)
00
02
04
06
08
10
12
0
1
2
3
4
5
6
ns
Control Periphery Scar Control Periphery Scar Control Periphery Scar
Amplitude Slope Decay time
Δ so
dium
(mM
)
120591(s
)lowastlowastlowast
lowastlowastlowastlowastlowastlowast
lowastlowastlowastlowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowast
(c)
Figure 7 D-aspartate-induced astrocyte sodium transients following lesion (a) CA1 area of a control (left) and a lesioned (right) slicepreparation Shown are epifluorescence images of SR101 and SBFI fluorescences Circled areas and numbers indicate the regions of interestanalyzed in the experiment depicted in (b) StrP stratum pyramidale StrR stratum radiatum and scale bar 40 120583m (b) Sodium transientsinduced by pressure application of 1mMD-aspartate (D-asp) for 500ms in the different cells in the control and lesioned slice as indicated in(a) (c) Histograms showing mean values plusmn S E M of the peak amplitude (left) slope (center) and decay time (right) of D-aspartate-inducedsodium transients in astrocytes in control slices (119899 = 161) as well as in cells in the scar region and its periphery (119899 = 111 and 119899 = 26 resp)lowastlowast119875 lt 001 lowastlowastlowast119875 lt 0001 StrP stratum pyramidale StrR stratum radiatum and scale bar 40120583m
cultures obtained from GFPGFAP mice demonstrating thatSR101 is well suited to identify astrocytes in this prepara-tion as described before for acute tissue slices and in vivo[29 30]
Performing a scratch through the CA1 region of organ-otypic slices resulted in the formation of a glial scar along thelesion site as judged by the prominent increase in expressionof GFAP and S100120573 In addition astrocyte cell bodies showed
a significant hypertrophy Furthermore astrocytes in theperiphery of the scar (100ndash350 120583m from the lesion) showedmild to moderate reactivity displaying swollen cell bodiesand only moderate upregulation of GFAP These changesare hallmarks of reactive gliosis observed after tissue injury[10 13 57] Generally the degree of injury and the distance ofthe astrocytes from the site of injury define the degree of theiractivation [11 13]
ISRN Neurology 13
Our study also revealed that strongly reactive astrocytesin the scar region only weakly stain with SR101 The reducedability of strongly activated astrocytes to accumulate SR101indicates that strong glial activation is accompanied by adownregulation of the organic anion transporter responsi-ble for uptake of SR101 This is reminiscent of immatureastrocytes in the early postnatal hippocampus which areSR101-negative [30] and in line with a wealth of experimentalevidence suggesting that reactive gliosis represents a processcomprising a dedifferentiation of astrocytes [17]
42 Changes in Glutamate Transporter Expression followingLesion Glutamate uptake in the hippocampus is mainlyachieved by the glial glutamate transporters GLT-1 andGLAST [1 2] A central factor in many brain pathologiesincluding traumatic brain injury is an increase in extracellu-lar glutamate and excitotoxicity it has been suggested that achange in the expression levels of glial glutamate transportersmight play a critical role in the failure of glutamate clearance[6 58ndash60] Earlier work has found an overall downregulationof protein levels of both GLAST and GLT-1 following astro-cyte activation [22ndash27] indicating that this is causal to theelevation of extracellular glutamate In contrast to this notionother studies reported increased glutamate transport capacityof reactive astrocytes and suggested a protective influence[61 62]
To visualize possible differences in the spatial expressionprofile of GLT-1 and GLAST in strongly reactive astrocytesalong the scar as compared to moderately reactive astro-cytes in its periphery we performed immunohistochemicalstainings These revealed an accumulation of GLT-1 andGLAST immunoreactivity along thick GFAP-positive fiberswhich was particularly pronounced for GLAST Besides theseclusters overall GLAST immunoreactivity seemed unalteredas compared to unlesioned slices whereas GLT-1 immunore-activity seemedweaker and declined with increasing distancefrom the scar Clustering of glutamate transporters has beendescribed by several reports [48 63 64] In developinghippocampal astrocytes cluster formation was preferentiallyfound in branches opposed to synapses and was increasedwith increased neuronal activity indicating that it is neces-sary to copewith synaptic release of glutamate [36] Along thesame lines it was observed that sustained astroglial activationby ciliary neurotrophic factor (CNTF) in the rat striatuminduced a concentration of GLAST and GLT-1 into raftmicrodomains and improved glutamate clearance indicatingthat cluster formation increased the cellular capacity for glu-tamate uptake [61] However another study found glutamate-induced clustering of GLT-1 that induced its endocytosis andintracellular trafficking without changing the total expressionlevels as detected by western blots arguing for a decrease infunctional glial glutamate uptake capacity [49] In the presentstudy antibodies against GLT-1 and GLAST were employedfor immunohistochemistry after permeabilization of theplasma membrane The clusters of GLAST and GLT-1 alongGFAP-positive fibers in the scar region (as GLAST andGLT-1immunoreactivity in general) could thus represent glutamatetransporters in the plasmamembrane or in intracellular com-partments or both Consequently the observed clustering
does not allow a prediction about possible functional changesin glutamate uptake
43 Lesion-Induced Changes in Functional Glutamate UptakeCapacity We probed for the functional activation of gluta-mate transport by application of the transportable agonistD-aspartate which results in an increase in the intracellularsodium concentration of astrocytes [8 9] The amplitudesof D-aspartate-induced sodium transients in astrocytes incontrol organotypic slices were similar to those reportedfrom astrocytes in acute slices obtained in our lab [33]indicating similar cellular glutamate transport capacity inboth preparations
Following a lesion D-aspartate-induced intracellularsodium signalswere clearly dampened in scar cellsMoreoverthe slope of the increase in sodium was significantly reducedand the decay back to baseline was slowed Because D-aspartate was applied at nearly saturating concentrationthe reduction in peak amplitudes suggests a reduction infunctional glutamate uptake capacity in cells close to thelesion which might be mediated by a decrease in the overallnumber of glutamate transporters available at the plasmamembrane In contrast the alteration in the kinetics of theD-aspartate-induced sodium signals can be explained by thecellular hypertrophy that was observed in reactive astrocytesAt an equal transport-mediated sodium influx across themembrane the slope of resulting changes in the sodiumconcentration will be decreased in cells with a larger volume
Furthermore the decay of intracellular sodium transientsback to baseline is mainly governed by the activity of theNa+K+-ATPase as well as by diffusion [8] The slower recov-ery is in line with earlier studies reporting a downregulationof the sodium pump following reactive gliosis [65 66]Moreover sodium is not buffered in the cell and travels solelyby means of diffusion in the cytoplasm Hindered diffusionresulting from increased cytoplasmic protein content orimpaired gap junction coupling which both occur uponastrocytic activation might thus also partially contribute todelayed sodium recovery [12 34 67]
In contrast to scar cells the amplitude of D-aspartate-induced intracellular sodium signals was not significantlyaltered in moderately activated astrocytes in the periphery ofthe lesion indicating that the overall number of functionalglutamate transporters was not significantly altered Becausethese cells showed hypertrophy as well the slowed kinetics ofthe sodium transients are in line with an increased cellularvolume as well as a possible downregulation of Na+K+-ATPase andor a slowed diffusion as argued above
Taken together our results indicate a significant reduc-tion in glutamate uptake capacity in strongly activated astro-cytes in the scar region Thus the prominent clustering ofGLAST and GLT-1 immunoreactivity along GFAP-positivestructures close to the lesion is likely to reflect a loss ofglutamate transporters from the plasma membrane upontransporter internalization as reported by Nakagawa et al[49] The authors of the latter study speculated that large andprolonged increases in glutamate concentrations are neces-sary to induce such cluster formation and endocytosis in thetissue as they would possibly only occur under pathological
14 ISRN Neurology
conditions Indeed ambient glutamate concentrations wereshown to be significantly increased after traumatic braininjury [68ndash70] Along these lines it can be assumed thatin our model system extracellular glutamate concentrationsrose less severely in the periphery of the lesion preventinga comparable clustering and loss of functional glutamatetransport activity in moderately activated astrocytes
5 Conclusions
Our results show complex changes in glutamate transporterexpression and function during astrocyte activation followingmechanical injury They confirm that the degree of astrocyteactivation depends on the distance from the insult and com-prises different endpoints regarding cellular morphology andphysiology Furthermore our results highlight that astrocyteswhich show different grades of reactivity also display discretechanges in glutamate transporter expression and functionWhile immunohistochemistry revealed a prominent clus-tering of GLT-1 and GLAST immunoreactivity in the scarregion our functional assay clearly showed that glutamateuptake capacity is strongly reduced in scar cells while itis largely maintained in moderately activated astrocytes inthe periphery Thus mild-to-moderate astrogliosis in theperiphery of a mechanical lesion does not necessarily seemto be accompanied by a significant change in glial glutamateuptake capacity At the glial scar itself in strongly reactiveastrocytes a clustering of glutamate transporters is observedthat apparently goes along with a severe functional reductionin astroglial glutamate uptake which may contribute toglutamate-mediated excitotoxicity in this region
Conflict of Interests
The authors of the paper declare that they do not haveany direct financial relation with the commercial identitiesmentioned in the paper that might lead to a conflict ofinterests
Acknowledgments
This study was supported by the Deutsche Forschungsge-meinschaft (DFG Ro23274-3) The authors thank SimoneDurry and Claudia Roderigo for expert technical help
References
[1] N C Danbolt ldquoGlutamate uptakerdquo Progress in Neurobiologyvol 65 no 1 pp 1ndash105 2001
[2] A V Tzingounis and J I Wadiche ldquoGlutamate transportersconfining runaway excitation by shaping synaptic transmis-sionrdquo Nature Reviews Neuroscience vol 8 no 12 pp 935ndash9472007
[3] D E Bergles J S Diamond and C E Jahr ldquoClearance ofglutamate inside the synapse and beyondrdquo Current Opinion inNeurobiology vol 9 no 3 pp 293ndash298 1999
[4] C M Anderson and R A Swanson ldquoAstrocyte glutamatetransport review of properties regulation and physiologicalfunctionsrdquo Glia vol 32 no 1 pp 1ndash14 2000
[5] P Marcaggi and D Attwell ldquoRole of glial amino acid trans-porters in synaptic transmission and brain energeticsrdquoGlia vol47 no 3 pp 217ndash225 2004
[6] N J Maragakis and J D Rothstein ldquoGlutamate transportersanimal models to neurologic diseaserdquo Neurobiology of Diseasevol 15 no 3 pp 461ndash473 2004
[7] G Gegelashvili and A Schousboe ldquoCellular distribution andkinetic properties of high-affinity glutamate transportersrdquoBrainResearch Bulletin vol 45 no 3 pp 233ndash238 1998
[8] J W Deitmer and C R Rose ldquoIon changes and signalling inperisynaptic gliardquo Brain Research Reviews vol 63 no 1-2 pp113ndash129 2010
[9] S Kirischuk V Parpura and A Verkhratsky ldquoSodium dynam-ics another key to astroglial excitabilityrdquo Trends in Neuro-sciences vol 35 no 8 pp 497ndash506 2012
[10] M V Sofroniew ldquoMolecular dissection of reactive astrogliosisand glial scar formationrdquo Trends in Neurosciences vol 32 no12 pp 638ndash647 2009
[11] J L Ridet S K Malhotra A Privat and F H Gage ldquoReactiveastrocytes cellular and molecular cues to biological functionrdquoTrends in Neurosciences vol 20 no 12 pp 570ndash577 1997
[12] J Middeldorp and E M Hol ldquoGFAP in health and diseaserdquoProgress in Neurobiology vol 93 no 3 pp 421ndash443 2011
[13] M Pekny and M Nilsson ldquoAstrocyte activation and reactivegliosisrdquo Glia vol 50 no 4 pp 427ndash434 2005
[14] A Bordey S A Lyons J J Hablitz and H SontheimerldquoElectrophysiological characteristics of reactive astrocytes inexperimental cortical dysplasiardquo Journal of Neurophysiologyvol 85 no 4 pp 1719ndash1731 2001
[15] R Jabs G Seifert and C Steinhauser ldquoAstrocytic functionand its alteration in the epileptic brainrdquo Epilepsia vol 49supplement 2 pp 3ndash12 2008
[16] A Buffo I Rite P Tripathi et al ldquoOrigin and progeny ofreactive gliosis a source of multipotent cells in the injuredbrainrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 105 no 9 pp 3581ndash3586 2008
[17] S Robel B Berninger and M Gotz ldquoThe stem cell potential ofglia lessons from reactive gliosisrdquoNature Reviews Neurosciencevol 12 no 2 pp 88ndash104 2011
[18] H Kawano J Kimura-Kuroda Y Komuta et al ldquoRole of thelesion scar in the response to damage and repair of the centralnervous systemrdquo Cell and Tissue Research vol 349 no 1 pp169ndash180 2012
[19] G Yiu and Z He ldquoGlial inhibition of CNS axon regenerationrdquoNature Reviews Neuroscience vol 7 no 8 pp 617ndash627 2006
[20] M T Fitch and J Silver ldquoCNS injury glial scars and inflamma-tion inhibitory extracellular matrices and regeneration failurerdquoExperimental Neurology vol 209 no 2 pp 294ndash301 2008
[21] L Li A Lundkvist D Andersson et al ldquoProtective role ofreactive astrocytes in brain ischemiardquo Journal of Cerebral BloodFlow amp Metabolism vol 28 no 3 pp 468ndash481 2008
[22] F K H van Landeghem T Weiss M Oehmichen and AVonDeimling ldquoDecreased expression of glutamate transportersin astrocytes after human traumatic brain injuryrdquo Journal ofNeurotrauma vol 23 no 10 pp 1518ndash1528 2006
[23] V L Raghavendra Rao M K Baskaya A Dogan J DRothstein and R J Dempsey ldquoTraumatic brain injury down-regulates glial glutamate transporter (GLT- 1 and GLAST)proteins in rat brainrdquo Journal of Neurochemistry vol 70 no 5pp 2020ndash2027 1998
ISRN Neurology 15
[24] J-C Chen H Hsu-Chou J-L Lu et al ldquoDown-regulation ofthe glial glutamate transporter GLT-1 in rat hippocampus andstriatum and its modulation by a group III metabotropic glu-tamate receptor antagonist following transient global forebrainischemiardquoNeuropharmacology vol 49 no 5 pp 703ndash714 2005
[25] S D Ginsberg L J Martin and J D Rothstein ldquoRegional deaf-ferentation down-regulates subtypes of glutamate transporterproteinsrdquo Journal of Neurochemistry vol 65 no 6 pp 2800ndash2803 1995
[26] J E Springer R D Azbill R J Mark J G Begley GWaeg andM P Mattson ldquo4-Hydroxynonenal a lipid peroxidation prod-uct rapidly accumulates following traumatic spinal cord injuryand inhibits glutamate uptakerdquo Journal of Neurochemistry vol68 no 6 pp 2469ndash2476 1997
[27] M B Moretto N S Arteni D Lavinsky et al ldquoHypoxic-ischemic insult decreases glutamate uptake by hippocampalslices from neonatal rats Prevention by guanosinerdquo Experimen-tal Neurology vol 195 no 2 pp 400ndash406 2005
[28] L Stoppini P-A Buchs and D Muller ldquoA simple method fororganotypic cultures of nervous tissuerdquo Journal of NeuroscienceMethods vol 37 no 2 pp 173ndash182 1991
[29] A Nimmerjahn F Kirchhoff J N D Kerr and F HelmchenldquoSulforhodamine 101 as a specific marker of astroglia in theneocortex in vivordquo Nat Methods vol 1 no 1 pp 31ndash37 2004
[30] K W Kafitz S D Meier J Stephan and C R Rose ldquoDevel-opmental profile and properties of sulforhodamine 101-Labeledglial cells in acute brain slices of rat hippocampusrdquo Journal ofNeuroscience Methods vol 169 no 1 pp 84ndash92 2008
[31] C Schnell Y Hagos and S Hulsmann ldquoActive sulforhodamine101 uptake into hippocampal astrocytesrdquo PLoS One vol 7 no 11Article ID e49398 2012
[32] S D Meier Y Kovalchuk and C R Rose ldquoProperties ofthe new fluorescent Na+ indicator CoroNa Green comparisonwith SBFI and confocal Na+ imagingrdquo Journal of NeuroscienceMethods vol 155 no 2 pp 251ndash259 2006
[33] J Langer and C R Rose ldquoSynaptically induced sodium signalsin hippocampal astrocytes in siturdquo Journal of Physiology vol587 part 24 pp 5859ndash5877 2009
[34] J Langer J Stephan M Theis and C R Rose ldquoGap junctionsmediate intercellular spread of sodium between hippocampalastrocytes in siturdquo Glia vol 60 no 2 pp 239ndash252 2012
[35] C R Rose andB R Ransom ldquoIntracellular sodiumhomeostasisin rat hippocampal astrocytesrdquo Journal of Physiology vol 491part 2 no 2 pp 291ndash305 1996
[36] A M Benediktsson G S Marrs J C Tu et al ldquoNeuronalactivity regulates glutamate transporter dynamics in developingastrocytesrdquo Glia vol 60 no 2 pp 175ndash188 2012
[37] M Zhou G P Schools and H K Kimelberg ldquoDevelopment ofGLAST(+) astrocytes andNG2(+) glia in rat hippocampus CA1mature astrocytes are electrophysiologically passiverdquo Journal ofNeurophysiology vol 95 no 1 pp 134ndash143 2006
[38] I Suarez G Bodega and B Fernandez ldquoModulation of glu-tamate transporters (GLAST GLT-1 and EAAC1) in the ratcerebellum following portocaval anastomosisrdquo Brain Researchvol 859 no 2 pp 293ndash302 2000
[39] X Zhu D E Bergles and A Nishiyama ldquoNG2 cells generateboth oligodendrocytes and gray matter astrocytesrdquo Develop-ment vol 135 no 1 pp 145ndash157 2008
[40] B Brunne S ZhaoADerouiche et al ldquoOriginmaturation andastroglial transformation of secondary radial glial cells in thedeveloping dentate gyrusrdquo Glia vol 58 no 13 pp 1553ndash15692010
[41] Y Wu A-Q Zhang and D T Yew ldquoAge related changes ofvarious markers of astrocytes in senescence-accelerated micehippocampusrdquo Neurochemistry International vol 46 no 7 pp565ndash574 2005
[42] S Magavi D Friedmann G Banks A Stolfi and C LoisldquoCoincident generation of pyramidal neurons and protoplasmicastrocytes in neocortical columnsrdquo Journal of Neuroscience vol32 no 14 pp 4762ndash4772 2012
[43] M Olabarria H N Noristani A Verkhratsky and J JRodrıguez ldquoConcomitant astroglial atrophy and astrogliosis ina triple transgenic animal model of Alzheimerrsquos diseaserdquo Gliavol 58 no 7 pp 831ndash838 2010
[44] A M Benediktsson S J Schachtele S H Green and M EDailey ldquoBallistic labeling and dynamic imaging of astrocytesin organotypic hippocampal slice culturesrdquo Journal of Neuro-science Methods vol 141 no 1 pp 41ndash53 2005
[45] I Lushnikova G Skibo D Muller and I Nikonenko ldquoSynapticpotentiation induces increased glial coverage of excitatorysynapses in CA1 hippocampusrdquoHippocampus vol 19 no 8 pp753ndash762 2009
[46] E Raponi F Agenes C Delphin et al ldquoS100B expressiondefines a state in which GFAP-expressing cells lose their neuralstem cell potential and acquire a more mature developmentalstagerdquo Glia vol 55 no 2 pp 165ndash177 2007
[47] C G Schipke C Boucsein C Ohlemeyer F Kirchhoff andH Kettenmann ldquoAstrocyte Ca2+ waves trigger responses inmicroglial cells in brain slicesrdquo The FASEB Journal vol 16 no2 pp 255ndash257 2002
[48] J Zhou and M L Sutherland ldquoGlutamate transporter clusterformation in astrocytic processes regulates glutamate uptakeactivityrdquo Journal of Neuroscience vol 24 no 28 pp 6301ndash63062004
[49] T Nakagawa Y Otsubo Y Yatani H Shirakawa and S KanekoldquoMechanisms of substrate transport-induced clustering of aglial glutamate transporter GLT-1 in astroglial-neuronal cul-turesrdquo European Journal of Neuroscience vol 28 no 9 pp 1719ndash1730 2008
[50] J-Y Chatton P Marquet and P J Magistretti ldquoA quantitativeanalysis of L-glutamate-regulated Na+ dynamics in mousecortical astrocytes Implications for cellular bioenergeticsrdquoEuropean Journal of Neuroscience vol 12 no 11 pp 3843ndash38532000
[51] E Forster S Zhao and M Frotscher ldquoLaminating the hip-pocampusrdquo Nature Reviews Neuroscience vol 7 no 4 pp 259ndash267 2006
[52] B H Gahwiler M Capogna D Debanne R A McKinney andS M Thompson ldquoOrganotypic slice cultures a technique hascome of agerdquo Trends in Neurosciences vol 20 no 10 pp 471ndash477 1997
[53] P E Kunkler and R P Kraig ldquoReactive astrocytosis fromexcitotoxic injury in hippocampal organ culture parallels thatseen in vivordquo Journal of Cerebral Blood Flow and Metabolismvol 17 no 1 pp 26ndash43 1997
[54] I E Holopainen ldquoOrganotypic hippocampal slice cultures amodel system to study basic cellular andmolecularmechanismsof neuronal cell death neuroprotection and synaptic plasticityrdquoNeurochemical Research vol 30 no 12 pp 1521ndash1528 2005
[55] MHaber L Zhou andK KMurai ldquoCooperative astrocyte anddendritic spine dynamics at hippocampal excitatory synapsesrdquoJournal of Neuroscience vol 26 no 35 pp 8881ndash8891 2006
16 ISRN Neurology
[56] H Nishida and S Okabe ldquoDirect astrocytic contacts regulatelocal maturation of dendritic spinesrdquo Journal of Neurosciencevol 27 no 2 pp 331ndash340 2007
[57] A Buffo C Rolando and S Ceruti ldquoAstrocytes in the dam-aged brain molecular and cellular insights into their reactiveresponse and healing potentialrdquoBiochemical Pharmacology vol79 no 2 pp 77ndash89 2010
[58] J-H Yi and A S Hazell ldquoExcitotoxic mechanisms and the roleof astrocytic glutamate transporters in traumatic brain injuryrdquoNeurochemistry International vol 48 no 5 pp 394ndash403 2006
[59] C Werner and K Engelhard ldquoPathophysiology of traumaticbrain injuryrdquo British Journal of Anaesthesia vol 99 no 1 pp4ndash9 2007
[60] G Gegelashvili M B Robinson D Trotti and T RauenldquoRegulation of glutamate transporters in health and diseaserdquoProgress in Brain Research vol 132 pp 267ndash286 2001
[61] C Escartin E Brouillet PGubellini et al ldquoCiliary neurotrophicfactor activates astrocytes redistributes their glutamate trans-porters GLAST and GLT-1 to raft microdomains and improvesglutamate handling in vivordquo Journal of Neuroscience vol 26 no22 pp 5978ndash5989 2006
[62] C Vermeiren M Najimi N Vanhoutte et al ldquoAcute up-regulation of glutamate uptake mediated by mGluR5a in reac-tive astrocytesrdquo Journal of Neurochemistry vol 94 no 2 pp405ndash416 2005
[63] C L Poitry-Yamate L Vutskits and T Rauen ldquoNeuronal-induced and glutamate-dependent activation of glial glutamatetransporter functionrdquo Journal of Neurochemistry vol 82 no 4pp 987ndash997 2002
[64] P M Beart and R D OrsquoShea ldquoTransporters for L-glutamatean update on their molecular pharmacology and pathologicalinvolvementrdquo British Journal of Pharmacology vol 150 no 1pp 5ndash17 2007
[65] F D Lima M A Souza A F Furian et al ldquoNa+K+-ATPaseactivity impairment after experimental traumatic brain injuryrelationship to spatial learning deficits and oxidative stressrdquoBehavioural Brain Research vol 193 no 2 pp 306ndash310 2008
[66] G A Gusarova H E Trejo L A Dada et al ldquoHypoxia leadsto NaK-ATPase downregulation via Ca2+ release-activatedCa2+ channels and AMPK activationrdquo Molecular and CellularBiology vol 31 no 17 pp 3546ndash3556 2011
[67] S S L Chew C S Johnson C R Green and H V Danesh-Meyer ldquoRole of connexin43 in central nervous system injuryrdquoExperimental Neurology vol 225 no 2 pp 250ndash261 2010
[68] P Nilsson L Hillered U Ponten and U Ungerstedt ldquoChangesin cortical extracellular levels of energy-related metabolites andamino acids following concussive brain injury in ratsrdquo Journal ofCerebral Blood Flow and Metabolism vol 10 no 5 pp 631ndash6371990
[69] H Katoh K Sima H Nawashiro K Wada and H ChigasakildquoThe effect of MK-801 on extracellular neuroactive amino acidsin hippocampus after closed head injury followed by hypoxia inratsrdquo Brain Research vol 758 no 1-2 pp 153ndash162 1997
[70] H Koizumi H Fujisawa H Ito T Maekawa X Di and RBullock ldquoEffects of mild hypothermia on cerebral blood flow-independent changes in cortical extracellular levels of aminoacids following contusion trauma in the ratrdquoBrain Research vol747 no 2 pp 304ndash312 1997
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
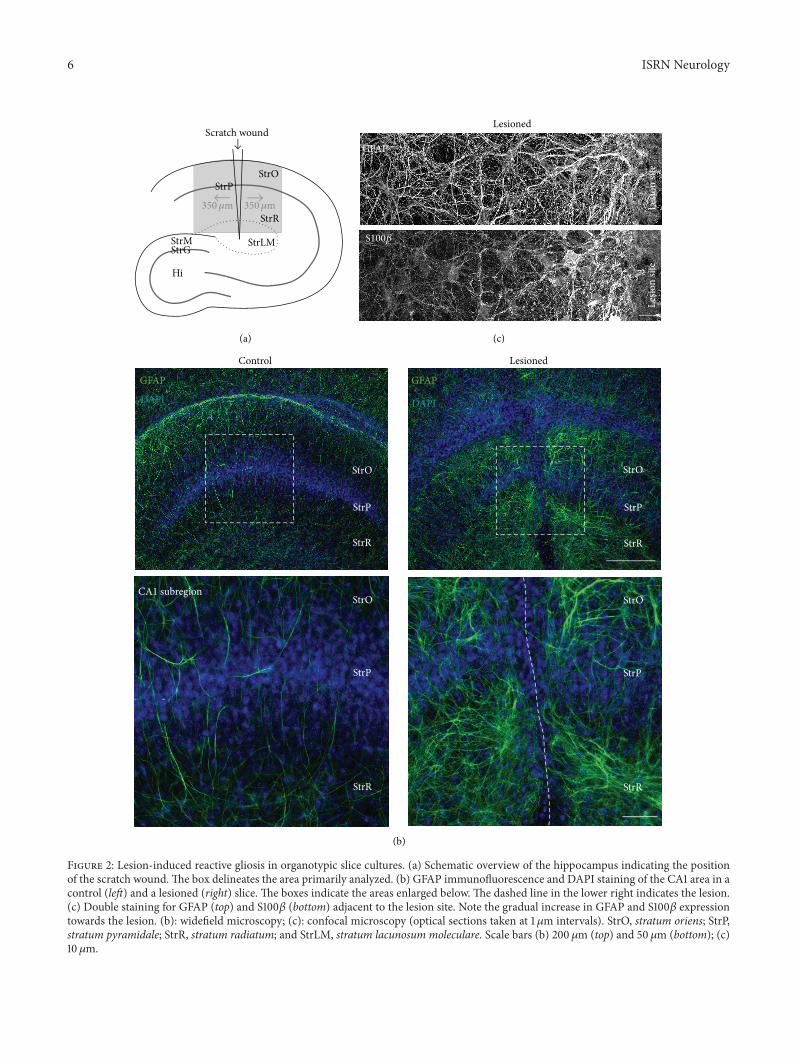
6 ISRN Neurology
GFAP
DAPI
CA1 subregion
GFAP
DAPI
StrR
StrP StrP
StrR
StrOStrO
StrO
StrP
StrR
StrO
StrP
StrR
Control Lesioned
GFAP
Lesio
n sit
e
Hi
StrMStrG
StrP
StrR
StrLM
Scratch wound
StrO
Lesio
n sit
e
Lesioned
(a) (c)
(b)
350120583m350120583m
S100120573
Figure 2 Lesion-induced reactive gliosis in organotypic slice cultures (a) Schematic overview of the hippocampus indicating the positionof the scratch woundThe box delineates the area primarily analyzed (b) GFAP immunofluorescence and DAPI staining of the CA1 area in acontrol (left) and a lesioned (right) slice The boxes indicate the areas enlarged below The dashed line in the lower right indicates the lesion(c) Double staining for GFAP (top) and S100120573 (bottom) adjacent to the lesion site Note the gradual increase in GFAP and S100120573 expressiontowards the lesion (b) widefield microscopy (c) confocal microscopy (optical sections taken at 1120583m intervals) StrO stratum oriens StrPstratum pyramidale StrR stratum radiatum and StrLM stratum lacunosum moleculare Scale bars (b) 200 120583m (top) and 50 120583m (bottom) (c)10120583m
ISRN Neurology 7
StrR
StrP
StrLM
StrO
StrP StrRStrO StrLM
Lesio
n sit
e
SR101-staining pattern
(a)
SBFI
Lesio
n sit
e
SR101 GFAP-GFP
(b)
Figure 3 SR101 labeling in lesioned slices (a) SR101 fluorescence in the CA1 area in a slice 7 days after lesion The delineated area comprisesthe stratum pyramidale and the box indicates the area enlarged on the right side Right SR101 fluorescence at the lesion site illustrating thatSR101 labeling is reduced along the lesionThe dotted line indicates the transition between the scar area and the adjacent moderately reactivetissue The boxed area is further enlarged at the bottom (b) Image series obtained at a wide-field fluorescence microscope The slice wasprepared from a transgenic GFPGFAP mouse and the lesion site is indicated by the dotted line Left Image of SR101 fluorescence centerGFP fluorescence indicating GFAP expression and right SBFI fluorescence SR101 labeling is only weak in GFPSBFI-positive cells in the scarregion (see arrowheads) StrO stratum oriens StrP stratum pyramidale StrR stratum radiatum and StrLM stratum lacunosum-moleculareScale bars 100120583m ((a) upper left) and 20 120583m (other images)
cellular component of the glial scar forming after mechanicaltissue injury [10] More distal to the lesion (gt100ndash350 120583m) aless dramatic rearrangement of GFAP immunoreactivity wasobserved
A clear difference in astrocyte properties between thescar region and its periphery was also seen following SR101labeling In the periphery of the scar (100ndash350 120583m distancefrom the lesion) SR101 resulted in a reliable and brightcellular staining similar to what had been observed incontrol unlesioned slices (119873 = 12 Figure 3(a) cf Figure 1)
In contrast to this cellular SR101 staining was detectable butonly faint within a distance of less than 100 120583m on both sidesof the lesion (Figure 3(a) meanwidth of SR101-free area 111plusmn3 120583m 119873 = 5) In slices from GFPGFAP transgenic animals(119873 = 5 Figure 3(b)) GFAP-positive weakly SR101-positivescar cells also stained with the sodium-sensitive fluorescenceindicator SBFI-AM (acetoxymethyl ester of sodium bindingbenzofuran isophthalate see below 119873 = 3) indicatingthat they were able to take up and de-esterify this dye(Figure 3(b))
8 ISRN Neurology
lowastlowastlowast
lowastlowastlowast
80
lowast
Control ScarPeriphery
120
160
0
40
200
Som
a siz
e (120583
m2)
(a)
Lesioned
StrP
StrR
StrP
Control
StrR
StrLM
Lesio
n sit
e
(b)
Figure 4 Lesion-induced changes in somatic area (a) Quantification of somatic area based on GFAPGFP fluorescence in control andlesioned slices The soma size increased both in astrocytes in the scar region (ldquoscarrdquo) and in astrocytes in the periphery of the lesion(ldquoperipheryrdquo lowastlowastlowast119875 lt 0001 lowast119875 lt 005) (b) Confocal images of GFP fluorescence in cultured slices obtained from GFPGFAP transgenicanimals under control conditions (left) and subjected to a lesion (right) Cells surrounded by boxes are shown at higher magnification in theinsets and the dashed lines around the soma illustrate the areas which served to determine their size StrP stratum pyramidale StrR stratumradiatum StrLM stratum lacunosum moleculare and scale bars 40 120583m and 10120583m (inset)
In addition to GFAP-positive cells the scar region hostedmicroglial cells identified by their vital staining with TexasRed-coupled lectin from lycopersicon esculentum (tomatolectin119873 = 4 not shown) Tomato lectin-positive microgliahowever did not take up SBFI-AM (119873 = 4 not shown) con-firming earlier reports thatmicroglial cells do not incorporateAM-ester dyes in situ [47]
To further characterize reactive gliosis in response to thescratch wound we determined the area of astrocyte somatain confocal images of the GFP fluorescence in control andlesioned slices obtained fromGFPGFAP transgenic animalsThesemeasurements revealed that compared to astrocytes in
control slices the soma area of astrocytes in the periphery ofa lesion was increased by 18 and that of astrocytes in thescar regionwas increased by 24 (control 163plusmn2 120583m2 lesionperiphery 193plusmn3 120583m2 scar tissue 201plusmn4 120583m2119875 lt 0001 119899 =945 496 and 439 in 7 control and 9 lesioned slices Figures4(a) and 4(b)) Thus astrocytes show a second hallmark ofreactive gliosis after lesion namely a hypertrophy of their cellbodies which is observed both in the scar tissue as well as inthe periphery of the lesion
Taken together these data show that 6-7 days after themechanical lesion a glial scar has formed along the lesionsite This scar encompasses strongly activated astrocytes
ISRN Neurology 9
characterized by a robust increase in GFAP and S100120573expressionThese proximal reactive astrocytes locatedwithina distance of less than 100120583m from the lesion exhibit longGFAP-positive processes have significantly swollen somataand only weakly stain with SR101 Astrocytes in the peripheryof the scar (100ndash350 120583m from the lesion) display swollen cellbodies as well but show onlymoderate upregulation of GFAPand maintain their ability to efficiently take up SR101
33 Expression Levels and Spatial Distribution of GLT-1 andGLAST To determine the spatial distribution of the gluta-mate transporters GLAST and GLT-1 in organotypic slicesand changes therein in response to the lesion immunohisto-chemistry was employed In control slices GLAST (119873 = 13Figure 5(a)) as well as GLT-1 immunoreactivity (119873 = 11Figure 5(b)) appeared as punctate labeling throughout theentire CA1 area The staining patterns for both transportersin the tissue slice cultures were thus similar to those reportedfrom acute tissue slice preparations of the hippocampus [3648 49]
The relatively homogeneous distribution of immunore-activity throughout the preparation found in control slicecultures was abandoned following a lesion Here distinctaccumulations of GLAST (119873 = 17 Figure 5(a)) and GLT-1immunoreactivity (119873 = 12 Figure 5(b)) were found in thescar region In both cases these preferentially colocalizedwith thick GFAP-positive bundles running in parallel to thelesion (Figure 5) Apart from the marked accumulation ofGLAST reactivity in parallel to the scar GLAST immunore-activity appeared generally increased in some lesioned slicepreparations (Figure 5(a)) but unaltered in others bothwithinthe scar as well as in its periphery For GLT-1 a decline in theintensity of immunoreactivity was observed with increasingdistance from the lesion (Figure 5(b))
In summary these results reveal distinct alterations inthe immunoreactivity of GLAST and GLT-1 following amechanical lesion In the scar region clusters of GLT-1 andGLAST immunoreactivity are present primarily colocalizedwith strongly GFAP-positive fibers Apart from these distinctclusters GLT-1 immunoreactivity appears to decline withdistance from the scar whereas GLAST immunoreactivityseems largely uniform throughout the preparation
34 Sodium Transients Induced by Activation of GlutamateUptake Glutamate uptake is accompanied by the inwardflux of sodium resulting in increase in the intracellularsodium concentration of astrocytes To monitor its functionwe assayed sodium increases induced by D-aspartate anonmetabolized substrate of sodium-dependent glutamateuptake which is also transported [1] in astrocytes in thestratum radiatum of organotypic slices To this end SR101-stained slices were additionally loaded with the sodium-sensitive fluorophore SBFI-AM (Figure 6(a) cf Figure 3(b))In situ calibrations of the SBFI fluorescence revealed a linearincrease in its fluorescence ratio at intracellular sodiumconcentrations between 10 and 40mM (119899 = 107 Figure 6(b))as reported before [33]
Repetitive pressure application of D-aspartate for 500msinduced reliable increases in the intracellular sodium con-centration in astrocytes indicating activation of sodium-dependent glutamate uptake (119899 = 488 Figure 6(c)) Theexperiments were performed in the presence of the sodium-channel blocker TTX (tetrodotoxin 05 120583M) to suppressaction potential generation The amplitude of D-aspartate-induced sodium increases was dependent on the concen-tration applied (Figure 6(c)) and saturated at about 2mM(119899 = 19 Figure 6(d)) as reported before for mouse corticalastrocytes in culture [50] At a concentration of 1mM theamplitude of D-aspartate-induced sodium transients wasnear maximum and amounted to 38 plusmn 02mM (119899 =161 Figures 6(c) and 6(d)) a value comparable to that inastrocytes in acute slices obtained in our lab [33]
To probe for changes in glutamate uptake we comparedsodium signals induced by 1mMD-aspartate between controland lesioned slices (Figure 7) In lesioned slices astrocyteslocated in the periphery of the lesion (100ndash350 120583m distance)responded with a sodium transient to application of 1mMD-aspartate (see cell 2 in Figures 7(a) and 7(b)) The amplitudeof D-aspartate-induced sodium signals in cells in the lesionperiphery was similar to that observed in control slices (38 plusmn03mM 119899 = 111 see cell 1 in Figures 7(a) 7(b) and 7(c))Application of D-aspartate also evoked sodium transientsin weakly SR101-positive SBFI-filled astrocytes located inthe glial scar area (lt100 120583m from the lesion see cells 3ndash5in Figures 7(a) and 7(b)) These cells however displayedsignificantly smaller peak amplitudes (16 plusmn 021mM 119899 =26 119875 lt 0001 Figures 7(b) and 7(c)) than astrocytes inthe periphery and in control slices
The kinetics of D-aspartate-induced sodium transientsdiffered between all three groups This was true for theslope of the sodium increase (10ndash90 change) which wassignificantly reduced in astrocytes from lesioned slices versusastrocytes from control slices (09 plusmn 005mMs in controlversus 06 plusmn 006mMs in the periphery and 02 plusmn 004mMsin the scar region 119899 = 161 111 and 26 resp 119875 lt 0001Figures 7(b) and 7(c)) In addition the decay back to baselinewas significantly slower following lesion (Figures 7(b) and7(c)) The time needed for the signal to decay by 67 wasapproximately doubled (decay time 120591 was 44 plusmn 2 s in control120591 = 81 plusmn 4 s in periphery of the lesion and 120591 = 95 plusmn 10 s inscar cells Figures 7(b) and 7(c))
These results show that mechanical injury results indistinct alterations in the kinetics andor amplitudes ofD-aspartate-induced intracellular sodium transients in reac-tive astrocytes depending on their distance to the lesion
4 Discussion
Our results show that within 6-7 days after setting a scratchwound in organotypic hippocampal slices a dense glial scarhad formed along the lesion site Up to approximately 100 120583mdistance from the lesion the scar tissue was characterized bya substantial elevation ofGFAP expression a pronounced cel-lular hypertrophy and reduced uptake of SR101 by astrocytes
10 ISRN Neurology
Con
trol
GLAST
Lesio
ned
GFAP Merge
StrR
StrP
StrO
Lesio
n sit
e
StrR
(a)
Con
trol
Lesio
ned
GLT-1 GFAP Merge
StrR
StrP
StrO
StrR
StrP
StrLM
Lesio
n sit
e
(b)
Figure 5 GLAST andGLT-1 immunoreactivity aftermechanical injury (a) Fluorescence images of immunohistochemical stainings detectingthe glutamate transporters GLAST (left) GFAP (center) as counterstaining and the corresponding merged image (GFAP green GLASTmagenta) in control (top row) and lesioned slice cultures (bottom row) (b) GLT-1 (left) GFAP (center) double labeling of a control (toprow) and a mechanically lesioned slice (bottom row) The corresponding merge (GFAP green GLT-1 magenta) is found on the right Notethat after lesion glutamate transporter immunoreactive clusters are accumulated in the direct vicinity of the lesion site (arrowheads) GFAPglial fibrillary acidic protein GLAST glutamate aspartate transporter GLT-1 glutamate-transporter-1 StrO stratum oriens StrP stratumpyramidale StrR stratum radiatum StrLM stratum lacunosum-moleculare and scale bars (a) 20 120583m (b) 40 120583m
While prominent clusters of GLAST and GLT-1 immunore-activity were observed alongGFAP-positive structures in scarcells D-aspartate-induced intracellular sodium signals werestrongly dampened indicating a significant reduction in glu-tamate uptake capacity close to the lesion In the periphery ofthe lesion (100ndash350120583m) astrocytes showed less pronounced
reactivity maintained their ability to efficiently take up SR101and displayed only minor changes in glutamate transporterimmunoreactivity and function Thus depending on theirdistance from the lesion astrocytes showed different gradesof reactivity and displayed discrete changes in glutamatetransporter immunoreactivity and uptake capacity
ISRN Neurology 11
SR101 MergeSBFI
StrRStrP
(a)
03
04
05
06
07
0 5 10 15
0
20 25 30 40 0
100
20
40
80
60
0 10 20 30 400
20
40
60
80
100
10 = 55 mM
107 cells in 6 experiments
F (340
380
) (ra
tio)
Intracellular Na+ concentration (mM)Intracellular Na+ concentration (mM)
ΔF
(340
380
) (
)
ΔF
(340
380
) (
) R2 = 097
(b)
D-aspartate
100 120583M500 120583M
750 120583M1000 120583M
1 mM
10 s
Δ
(c)
0 1 2 3 4 50
1
2
3
4
5
Δ so
dium
(mM
)
D-aspartate (mM)
lowast
(d)
Figure 6 Sodium changes induced by activation of glutamate uptake (a) Staining of astrocytes by colabeling with SR101 and SBFI Leftepifluorescence image of theCA1-area of an SR101-loaded organotypic sliceCenter SBFI fluorescence taken from the same areaRight overlaySBFI-loaded SR101-positive astrocytes appear in white (arrowheads) StrP stratum pyramidale StrR stratum radiatum scale bar 20 120583m (b)Left calibration of SBFI fluorescence in an astrocyte Depicted is the change in intracellular SBFI fluorescence ratio in the calibration solutionscontaining ionophores and in response to stepwise changes in the sodium concentration Right mean values plusmn SEM of the normalizedchanges in SBFI fluorescence at different sodium concentrations The black line represents a linear fit of the data points between 10 and40mM sodium illustrating that a 10 change in SBFI fluorescence represents a change in the sodium concentration by 55mM (c) Sodiumtransients in a single astrocyte induced by 500ms applications of D-aspartate at different concentrations (d) Mean values plusmn S E M (119899 = 19)of the peak amplitude of sodium transients at different concentrations of D-aspartate The curve represents a sigmoidal fit of the data
41 Characteristics of Lesion-Induced Reactive Gliosis inOrganotypic Cultures Organotypic tissue slice cultures ofthe hippocampus represent a well-established model systemin which the basic tissue architecture is maintained butwhich still hosts the major advantages of cell cultures suchas good accessibility and control of experimental conditionsThey have been used extensively for the analysis of neuronal
properties and developing neuronal networks [28 51 52]or the study of excitotoxic neuronal damage (eg [53 54])Recent studies also demonstrated that basic morphologicalcharacteristics of astrocytes and the typical glia-synapseorganization are well preserved in organotypic slice cultures(eg [44 45 55 56]) In addition we found a nearly completeoverlap of SR101 staining with the expression of GFP in slice
12 ISRN Neurology
SBFIControl Lesioned
SR101
1
Lesio
n sit
e
SR101 SBFI
2 45
32 4
5
1
StrR
StrP StrP
StrR
3
(a)
ControlScar
Lesioned
1
D-asp
2
D-asp
D-asp
5
4
3Periphery1mM
20 s
Δ Δ
Δ
Δ
Δ
(b)
0102030405060708090
100110
Δ so
dium
tim
e (m
Ms
)
00
02
04
06
08
10
12
0
1
2
3
4
5
6
ns
Control Periphery Scar Control Periphery Scar Control Periphery Scar
Amplitude Slope Decay time
Δ so
dium
(mM
)
120591(s
)lowastlowastlowast
lowastlowastlowastlowastlowastlowast
lowastlowastlowastlowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowast
(c)
Figure 7 D-aspartate-induced astrocyte sodium transients following lesion (a) CA1 area of a control (left) and a lesioned (right) slicepreparation Shown are epifluorescence images of SR101 and SBFI fluorescences Circled areas and numbers indicate the regions of interestanalyzed in the experiment depicted in (b) StrP stratum pyramidale StrR stratum radiatum and scale bar 40 120583m (b) Sodium transientsinduced by pressure application of 1mMD-aspartate (D-asp) for 500ms in the different cells in the control and lesioned slice as indicated in(a) (c) Histograms showing mean values plusmn S E M of the peak amplitude (left) slope (center) and decay time (right) of D-aspartate-inducedsodium transients in astrocytes in control slices (119899 = 161) as well as in cells in the scar region and its periphery (119899 = 111 and 119899 = 26 resp)lowastlowast119875 lt 001 lowastlowastlowast119875 lt 0001 StrP stratum pyramidale StrR stratum radiatum and scale bar 40120583m
cultures obtained from GFPGFAP mice demonstrating thatSR101 is well suited to identify astrocytes in this prepara-tion as described before for acute tissue slices and in vivo[29 30]
Performing a scratch through the CA1 region of organ-otypic slices resulted in the formation of a glial scar along thelesion site as judged by the prominent increase in expressionof GFAP and S100120573 In addition astrocyte cell bodies showed
a significant hypertrophy Furthermore astrocytes in theperiphery of the scar (100ndash350 120583m from the lesion) showedmild to moderate reactivity displaying swollen cell bodiesand only moderate upregulation of GFAP These changesare hallmarks of reactive gliosis observed after tissue injury[10 13 57] Generally the degree of injury and the distance ofthe astrocytes from the site of injury define the degree of theiractivation [11 13]
ISRN Neurology 13
Our study also revealed that strongly reactive astrocytesin the scar region only weakly stain with SR101 The reducedability of strongly activated astrocytes to accumulate SR101indicates that strong glial activation is accompanied by adownregulation of the organic anion transporter responsi-ble for uptake of SR101 This is reminiscent of immatureastrocytes in the early postnatal hippocampus which areSR101-negative [30] and in line with a wealth of experimentalevidence suggesting that reactive gliosis represents a processcomprising a dedifferentiation of astrocytes [17]
42 Changes in Glutamate Transporter Expression followingLesion Glutamate uptake in the hippocampus is mainlyachieved by the glial glutamate transporters GLT-1 andGLAST [1 2] A central factor in many brain pathologiesincluding traumatic brain injury is an increase in extracellu-lar glutamate and excitotoxicity it has been suggested that achange in the expression levels of glial glutamate transportersmight play a critical role in the failure of glutamate clearance[6 58ndash60] Earlier work has found an overall downregulationof protein levels of both GLAST and GLT-1 following astro-cyte activation [22ndash27] indicating that this is causal to theelevation of extracellular glutamate In contrast to this notionother studies reported increased glutamate transport capacityof reactive astrocytes and suggested a protective influence[61 62]
To visualize possible differences in the spatial expressionprofile of GLT-1 and GLAST in strongly reactive astrocytesalong the scar as compared to moderately reactive astro-cytes in its periphery we performed immunohistochemicalstainings These revealed an accumulation of GLT-1 andGLAST immunoreactivity along thick GFAP-positive fiberswhich was particularly pronounced for GLAST Besides theseclusters overall GLAST immunoreactivity seemed unalteredas compared to unlesioned slices whereas GLT-1 immunore-activity seemedweaker and declined with increasing distancefrom the scar Clustering of glutamate transporters has beendescribed by several reports [48 63 64] In developinghippocampal astrocytes cluster formation was preferentiallyfound in branches opposed to synapses and was increasedwith increased neuronal activity indicating that it is neces-sary to copewith synaptic release of glutamate [36] Along thesame lines it was observed that sustained astroglial activationby ciliary neurotrophic factor (CNTF) in the rat striatuminduced a concentration of GLAST and GLT-1 into raftmicrodomains and improved glutamate clearance indicatingthat cluster formation increased the cellular capacity for glu-tamate uptake [61] However another study found glutamate-induced clustering of GLT-1 that induced its endocytosis andintracellular trafficking without changing the total expressionlevels as detected by western blots arguing for a decrease infunctional glial glutamate uptake capacity [49] In the presentstudy antibodies against GLT-1 and GLAST were employedfor immunohistochemistry after permeabilization of theplasma membrane The clusters of GLAST and GLT-1 alongGFAP-positive fibers in the scar region (as GLAST andGLT-1immunoreactivity in general) could thus represent glutamatetransporters in the plasmamembrane or in intracellular com-partments or both Consequently the observed clustering
does not allow a prediction about possible functional changesin glutamate uptake
43 Lesion-Induced Changes in Functional Glutamate UptakeCapacity We probed for the functional activation of gluta-mate transport by application of the transportable agonistD-aspartate which results in an increase in the intracellularsodium concentration of astrocytes [8 9] The amplitudesof D-aspartate-induced sodium transients in astrocytes incontrol organotypic slices were similar to those reportedfrom astrocytes in acute slices obtained in our lab [33]indicating similar cellular glutamate transport capacity inboth preparations
Following a lesion D-aspartate-induced intracellularsodium signalswere clearly dampened in scar cellsMoreoverthe slope of the increase in sodium was significantly reducedand the decay back to baseline was slowed Because D-aspartate was applied at nearly saturating concentrationthe reduction in peak amplitudes suggests a reduction infunctional glutamate uptake capacity in cells close to thelesion which might be mediated by a decrease in the overallnumber of glutamate transporters available at the plasmamembrane In contrast the alteration in the kinetics of theD-aspartate-induced sodium signals can be explained by thecellular hypertrophy that was observed in reactive astrocytesAt an equal transport-mediated sodium influx across themembrane the slope of resulting changes in the sodiumconcentration will be decreased in cells with a larger volume
Furthermore the decay of intracellular sodium transientsback to baseline is mainly governed by the activity of theNa+K+-ATPase as well as by diffusion [8] The slower recov-ery is in line with earlier studies reporting a downregulationof the sodium pump following reactive gliosis [65 66]Moreover sodium is not buffered in the cell and travels solelyby means of diffusion in the cytoplasm Hindered diffusionresulting from increased cytoplasmic protein content orimpaired gap junction coupling which both occur uponastrocytic activation might thus also partially contribute todelayed sodium recovery [12 34 67]
In contrast to scar cells the amplitude of D-aspartate-induced intracellular sodium signals was not significantlyaltered in moderately activated astrocytes in the periphery ofthe lesion indicating that the overall number of functionalglutamate transporters was not significantly altered Becausethese cells showed hypertrophy as well the slowed kinetics ofthe sodium transients are in line with an increased cellularvolume as well as a possible downregulation of Na+K+-ATPase andor a slowed diffusion as argued above
Taken together our results indicate a significant reduc-tion in glutamate uptake capacity in strongly activated astro-cytes in the scar region Thus the prominent clustering ofGLAST and GLT-1 immunoreactivity along GFAP-positivestructures close to the lesion is likely to reflect a loss ofglutamate transporters from the plasma membrane upontransporter internalization as reported by Nakagawa et al[49] The authors of the latter study speculated that large andprolonged increases in glutamate concentrations are neces-sary to induce such cluster formation and endocytosis in thetissue as they would possibly only occur under pathological
14 ISRN Neurology
conditions Indeed ambient glutamate concentrations wereshown to be significantly increased after traumatic braininjury [68ndash70] Along these lines it can be assumed thatin our model system extracellular glutamate concentrationsrose less severely in the periphery of the lesion preventinga comparable clustering and loss of functional glutamatetransport activity in moderately activated astrocytes
5 Conclusions
Our results show complex changes in glutamate transporterexpression and function during astrocyte activation followingmechanical injury They confirm that the degree of astrocyteactivation depends on the distance from the insult and com-prises different endpoints regarding cellular morphology andphysiology Furthermore our results highlight that astrocyteswhich show different grades of reactivity also display discretechanges in glutamate transporter expression and functionWhile immunohistochemistry revealed a prominent clus-tering of GLT-1 and GLAST immunoreactivity in the scarregion our functional assay clearly showed that glutamateuptake capacity is strongly reduced in scar cells while itis largely maintained in moderately activated astrocytes inthe periphery Thus mild-to-moderate astrogliosis in theperiphery of a mechanical lesion does not necessarily seemto be accompanied by a significant change in glial glutamateuptake capacity At the glial scar itself in strongly reactiveastrocytes a clustering of glutamate transporters is observedthat apparently goes along with a severe functional reductionin astroglial glutamate uptake which may contribute toglutamate-mediated excitotoxicity in this region
Conflict of Interests
The authors of the paper declare that they do not haveany direct financial relation with the commercial identitiesmentioned in the paper that might lead to a conflict ofinterests
Acknowledgments
This study was supported by the Deutsche Forschungsge-meinschaft (DFG Ro23274-3) The authors thank SimoneDurry and Claudia Roderigo for expert technical help
References
[1] N C Danbolt ldquoGlutamate uptakerdquo Progress in Neurobiologyvol 65 no 1 pp 1ndash105 2001
[2] A V Tzingounis and J I Wadiche ldquoGlutamate transportersconfining runaway excitation by shaping synaptic transmis-sionrdquo Nature Reviews Neuroscience vol 8 no 12 pp 935ndash9472007
[3] D E Bergles J S Diamond and C E Jahr ldquoClearance ofglutamate inside the synapse and beyondrdquo Current Opinion inNeurobiology vol 9 no 3 pp 293ndash298 1999
[4] C M Anderson and R A Swanson ldquoAstrocyte glutamatetransport review of properties regulation and physiologicalfunctionsrdquo Glia vol 32 no 1 pp 1ndash14 2000
[5] P Marcaggi and D Attwell ldquoRole of glial amino acid trans-porters in synaptic transmission and brain energeticsrdquoGlia vol47 no 3 pp 217ndash225 2004
[6] N J Maragakis and J D Rothstein ldquoGlutamate transportersanimal models to neurologic diseaserdquo Neurobiology of Diseasevol 15 no 3 pp 461ndash473 2004
[7] G Gegelashvili and A Schousboe ldquoCellular distribution andkinetic properties of high-affinity glutamate transportersrdquoBrainResearch Bulletin vol 45 no 3 pp 233ndash238 1998
[8] J W Deitmer and C R Rose ldquoIon changes and signalling inperisynaptic gliardquo Brain Research Reviews vol 63 no 1-2 pp113ndash129 2010
[9] S Kirischuk V Parpura and A Verkhratsky ldquoSodium dynam-ics another key to astroglial excitabilityrdquo Trends in Neuro-sciences vol 35 no 8 pp 497ndash506 2012
[10] M V Sofroniew ldquoMolecular dissection of reactive astrogliosisand glial scar formationrdquo Trends in Neurosciences vol 32 no12 pp 638ndash647 2009
[11] J L Ridet S K Malhotra A Privat and F H Gage ldquoReactiveastrocytes cellular and molecular cues to biological functionrdquoTrends in Neurosciences vol 20 no 12 pp 570ndash577 1997
[12] J Middeldorp and E M Hol ldquoGFAP in health and diseaserdquoProgress in Neurobiology vol 93 no 3 pp 421ndash443 2011
[13] M Pekny and M Nilsson ldquoAstrocyte activation and reactivegliosisrdquo Glia vol 50 no 4 pp 427ndash434 2005
[14] A Bordey S A Lyons J J Hablitz and H SontheimerldquoElectrophysiological characteristics of reactive astrocytes inexperimental cortical dysplasiardquo Journal of Neurophysiologyvol 85 no 4 pp 1719ndash1731 2001
[15] R Jabs G Seifert and C Steinhauser ldquoAstrocytic functionand its alteration in the epileptic brainrdquo Epilepsia vol 49supplement 2 pp 3ndash12 2008
[16] A Buffo I Rite P Tripathi et al ldquoOrigin and progeny ofreactive gliosis a source of multipotent cells in the injuredbrainrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 105 no 9 pp 3581ndash3586 2008
[17] S Robel B Berninger and M Gotz ldquoThe stem cell potential ofglia lessons from reactive gliosisrdquoNature Reviews Neurosciencevol 12 no 2 pp 88ndash104 2011
[18] H Kawano J Kimura-Kuroda Y Komuta et al ldquoRole of thelesion scar in the response to damage and repair of the centralnervous systemrdquo Cell and Tissue Research vol 349 no 1 pp169ndash180 2012
[19] G Yiu and Z He ldquoGlial inhibition of CNS axon regenerationrdquoNature Reviews Neuroscience vol 7 no 8 pp 617ndash627 2006
[20] M T Fitch and J Silver ldquoCNS injury glial scars and inflamma-tion inhibitory extracellular matrices and regeneration failurerdquoExperimental Neurology vol 209 no 2 pp 294ndash301 2008
[21] L Li A Lundkvist D Andersson et al ldquoProtective role ofreactive astrocytes in brain ischemiardquo Journal of Cerebral BloodFlow amp Metabolism vol 28 no 3 pp 468ndash481 2008
[22] F K H van Landeghem T Weiss M Oehmichen and AVonDeimling ldquoDecreased expression of glutamate transportersin astrocytes after human traumatic brain injuryrdquo Journal ofNeurotrauma vol 23 no 10 pp 1518ndash1528 2006
[23] V L Raghavendra Rao M K Baskaya A Dogan J DRothstein and R J Dempsey ldquoTraumatic brain injury down-regulates glial glutamate transporter (GLT- 1 and GLAST)proteins in rat brainrdquo Journal of Neurochemistry vol 70 no 5pp 2020ndash2027 1998
ISRN Neurology 15
[24] J-C Chen H Hsu-Chou J-L Lu et al ldquoDown-regulation ofthe glial glutamate transporter GLT-1 in rat hippocampus andstriatum and its modulation by a group III metabotropic glu-tamate receptor antagonist following transient global forebrainischemiardquoNeuropharmacology vol 49 no 5 pp 703ndash714 2005
[25] S D Ginsberg L J Martin and J D Rothstein ldquoRegional deaf-ferentation down-regulates subtypes of glutamate transporterproteinsrdquo Journal of Neurochemistry vol 65 no 6 pp 2800ndash2803 1995
[26] J E Springer R D Azbill R J Mark J G Begley GWaeg andM P Mattson ldquo4-Hydroxynonenal a lipid peroxidation prod-uct rapidly accumulates following traumatic spinal cord injuryand inhibits glutamate uptakerdquo Journal of Neurochemistry vol68 no 6 pp 2469ndash2476 1997
[27] M B Moretto N S Arteni D Lavinsky et al ldquoHypoxic-ischemic insult decreases glutamate uptake by hippocampalslices from neonatal rats Prevention by guanosinerdquo Experimen-tal Neurology vol 195 no 2 pp 400ndash406 2005
[28] L Stoppini P-A Buchs and D Muller ldquoA simple method fororganotypic cultures of nervous tissuerdquo Journal of NeuroscienceMethods vol 37 no 2 pp 173ndash182 1991
[29] A Nimmerjahn F Kirchhoff J N D Kerr and F HelmchenldquoSulforhodamine 101 as a specific marker of astroglia in theneocortex in vivordquo Nat Methods vol 1 no 1 pp 31ndash37 2004
[30] K W Kafitz S D Meier J Stephan and C R Rose ldquoDevel-opmental profile and properties of sulforhodamine 101-Labeledglial cells in acute brain slices of rat hippocampusrdquo Journal ofNeuroscience Methods vol 169 no 1 pp 84ndash92 2008
[31] C Schnell Y Hagos and S Hulsmann ldquoActive sulforhodamine101 uptake into hippocampal astrocytesrdquo PLoS One vol 7 no 11Article ID e49398 2012
[32] S D Meier Y Kovalchuk and C R Rose ldquoProperties ofthe new fluorescent Na+ indicator CoroNa Green comparisonwith SBFI and confocal Na+ imagingrdquo Journal of NeuroscienceMethods vol 155 no 2 pp 251ndash259 2006
[33] J Langer and C R Rose ldquoSynaptically induced sodium signalsin hippocampal astrocytes in siturdquo Journal of Physiology vol587 part 24 pp 5859ndash5877 2009
[34] J Langer J Stephan M Theis and C R Rose ldquoGap junctionsmediate intercellular spread of sodium between hippocampalastrocytes in siturdquo Glia vol 60 no 2 pp 239ndash252 2012
[35] C R Rose andB R Ransom ldquoIntracellular sodiumhomeostasisin rat hippocampal astrocytesrdquo Journal of Physiology vol 491part 2 no 2 pp 291ndash305 1996
[36] A M Benediktsson G S Marrs J C Tu et al ldquoNeuronalactivity regulates glutamate transporter dynamics in developingastrocytesrdquo Glia vol 60 no 2 pp 175ndash188 2012
[37] M Zhou G P Schools and H K Kimelberg ldquoDevelopment ofGLAST(+) astrocytes andNG2(+) glia in rat hippocampus CA1mature astrocytes are electrophysiologically passiverdquo Journal ofNeurophysiology vol 95 no 1 pp 134ndash143 2006
[38] I Suarez G Bodega and B Fernandez ldquoModulation of glu-tamate transporters (GLAST GLT-1 and EAAC1) in the ratcerebellum following portocaval anastomosisrdquo Brain Researchvol 859 no 2 pp 293ndash302 2000
[39] X Zhu D E Bergles and A Nishiyama ldquoNG2 cells generateboth oligodendrocytes and gray matter astrocytesrdquo Develop-ment vol 135 no 1 pp 145ndash157 2008
[40] B Brunne S ZhaoADerouiche et al ldquoOriginmaturation andastroglial transformation of secondary radial glial cells in thedeveloping dentate gyrusrdquo Glia vol 58 no 13 pp 1553ndash15692010
[41] Y Wu A-Q Zhang and D T Yew ldquoAge related changes ofvarious markers of astrocytes in senescence-accelerated micehippocampusrdquo Neurochemistry International vol 46 no 7 pp565ndash574 2005
[42] S Magavi D Friedmann G Banks A Stolfi and C LoisldquoCoincident generation of pyramidal neurons and protoplasmicastrocytes in neocortical columnsrdquo Journal of Neuroscience vol32 no 14 pp 4762ndash4772 2012
[43] M Olabarria H N Noristani A Verkhratsky and J JRodrıguez ldquoConcomitant astroglial atrophy and astrogliosis ina triple transgenic animal model of Alzheimerrsquos diseaserdquo Gliavol 58 no 7 pp 831ndash838 2010
[44] A M Benediktsson S J Schachtele S H Green and M EDailey ldquoBallistic labeling and dynamic imaging of astrocytesin organotypic hippocampal slice culturesrdquo Journal of Neuro-science Methods vol 141 no 1 pp 41ndash53 2005
[45] I Lushnikova G Skibo D Muller and I Nikonenko ldquoSynapticpotentiation induces increased glial coverage of excitatorysynapses in CA1 hippocampusrdquoHippocampus vol 19 no 8 pp753ndash762 2009
[46] E Raponi F Agenes C Delphin et al ldquoS100B expressiondefines a state in which GFAP-expressing cells lose their neuralstem cell potential and acquire a more mature developmentalstagerdquo Glia vol 55 no 2 pp 165ndash177 2007
[47] C G Schipke C Boucsein C Ohlemeyer F Kirchhoff andH Kettenmann ldquoAstrocyte Ca2+ waves trigger responses inmicroglial cells in brain slicesrdquo The FASEB Journal vol 16 no2 pp 255ndash257 2002
[48] J Zhou and M L Sutherland ldquoGlutamate transporter clusterformation in astrocytic processes regulates glutamate uptakeactivityrdquo Journal of Neuroscience vol 24 no 28 pp 6301ndash63062004
[49] T Nakagawa Y Otsubo Y Yatani H Shirakawa and S KanekoldquoMechanisms of substrate transport-induced clustering of aglial glutamate transporter GLT-1 in astroglial-neuronal cul-turesrdquo European Journal of Neuroscience vol 28 no 9 pp 1719ndash1730 2008
[50] J-Y Chatton P Marquet and P J Magistretti ldquoA quantitativeanalysis of L-glutamate-regulated Na+ dynamics in mousecortical astrocytes Implications for cellular bioenergeticsrdquoEuropean Journal of Neuroscience vol 12 no 11 pp 3843ndash38532000
[51] E Forster S Zhao and M Frotscher ldquoLaminating the hip-pocampusrdquo Nature Reviews Neuroscience vol 7 no 4 pp 259ndash267 2006
[52] B H Gahwiler M Capogna D Debanne R A McKinney andS M Thompson ldquoOrganotypic slice cultures a technique hascome of agerdquo Trends in Neurosciences vol 20 no 10 pp 471ndash477 1997
[53] P E Kunkler and R P Kraig ldquoReactive astrocytosis fromexcitotoxic injury in hippocampal organ culture parallels thatseen in vivordquo Journal of Cerebral Blood Flow and Metabolismvol 17 no 1 pp 26ndash43 1997
[54] I E Holopainen ldquoOrganotypic hippocampal slice cultures amodel system to study basic cellular andmolecularmechanismsof neuronal cell death neuroprotection and synaptic plasticityrdquoNeurochemical Research vol 30 no 12 pp 1521ndash1528 2005
[55] MHaber L Zhou andK KMurai ldquoCooperative astrocyte anddendritic spine dynamics at hippocampal excitatory synapsesrdquoJournal of Neuroscience vol 26 no 35 pp 8881ndash8891 2006
16 ISRN Neurology
[56] H Nishida and S Okabe ldquoDirect astrocytic contacts regulatelocal maturation of dendritic spinesrdquo Journal of Neurosciencevol 27 no 2 pp 331ndash340 2007
[57] A Buffo C Rolando and S Ceruti ldquoAstrocytes in the dam-aged brain molecular and cellular insights into their reactiveresponse and healing potentialrdquoBiochemical Pharmacology vol79 no 2 pp 77ndash89 2010
[58] J-H Yi and A S Hazell ldquoExcitotoxic mechanisms and the roleof astrocytic glutamate transporters in traumatic brain injuryrdquoNeurochemistry International vol 48 no 5 pp 394ndash403 2006
[59] C Werner and K Engelhard ldquoPathophysiology of traumaticbrain injuryrdquo British Journal of Anaesthesia vol 99 no 1 pp4ndash9 2007
[60] G Gegelashvili M B Robinson D Trotti and T RauenldquoRegulation of glutamate transporters in health and diseaserdquoProgress in Brain Research vol 132 pp 267ndash286 2001
[61] C Escartin E Brouillet PGubellini et al ldquoCiliary neurotrophicfactor activates astrocytes redistributes their glutamate trans-porters GLAST and GLT-1 to raft microdomains and improvesglutamate handling in vivordquo Journal of Neuroscience vol 26 no22 pp 5978ndash5989 2006
[62] C Vermeiren M Najimi N Vanhoutte et al ldquoAcute up-regulation of glutamate uptake mediated by mGluR5a in reac-tive astrocytesrdquo Journal of Neurochemistry vol 94 no 2 pp405ndash416 2005
[63] C L Poitry-Yamate L Vutskits and T Rauen ldquoNeuronal-induced and glutamate-dependent activation of glial glutamatetransporter functionrdquo Journal of Neurochemistry vol 82 no 4pp 987ndash997 2002
[64] P M Beart and R D OrsquoShea ldquoTransporters for L-glutamatean update on their molecular pharmacology and pathologicalinvolvementrdquo British Journal of Pharmacology vol 150 no 1pp 5ndash17 2007
[65] F D Lima M A Souza A F Furian et al ldquoNa+K+-ATPaseactivity impairment after experimental traumatic brain injuryrelationship to spatial learning deficits and oxidative stressrdquoBehavioural Brain Research vol 193 no 2 pp 306ndash310 2008
[66] G A Gusarova H E Trejo L A Dada et al ldquoHypoxia leadsto NaK-ATPase downregulation via Ca2+ release-activatedCa2+ channels and AMPK activationrdquo Molecular and CellularBiology vol 31 no 17 pp 3546ndash3556 2011
[67] S S L Chew C S Johnson C R Green and H V Danesh-Meyer ldquoRole of connexin43 in central nervous system injuryrdquoExperimental Neurology vol 225 no 2 pp 250ndash261 2010
[68] P Nilsson L Hillered U Ponten and U Ungerstedt ldquoChangesin cortical extracellular levels of energy-related metabolites andamino acids following concussive brain injury in ratsrdquo Journal ofCerebral Blood Flow and Metabolism vol 10 no 5 pp 631ndash6371990
[69] H Katoh K Sima H Nawashiro K Wada and H ChigasakildquoThe effect of MK-801 on extracellular neuroactive amino acidsin hippocampus after closed head injury followed by hypoxia inratsrdquo Brain Research vol 758 no 1-2 pp 153ndash162 1997
[70] H Koizumi H Fujisawa H Ito T Maekawa X Di and RBullock ldquoEffects of mild hypothermia on cerebral blood flow-independent changes in cortical extracellular levels of aminoacids following contusion trauma in the ratrdquoBrain Research vol747 no 2 pp 304ndash312 1997
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
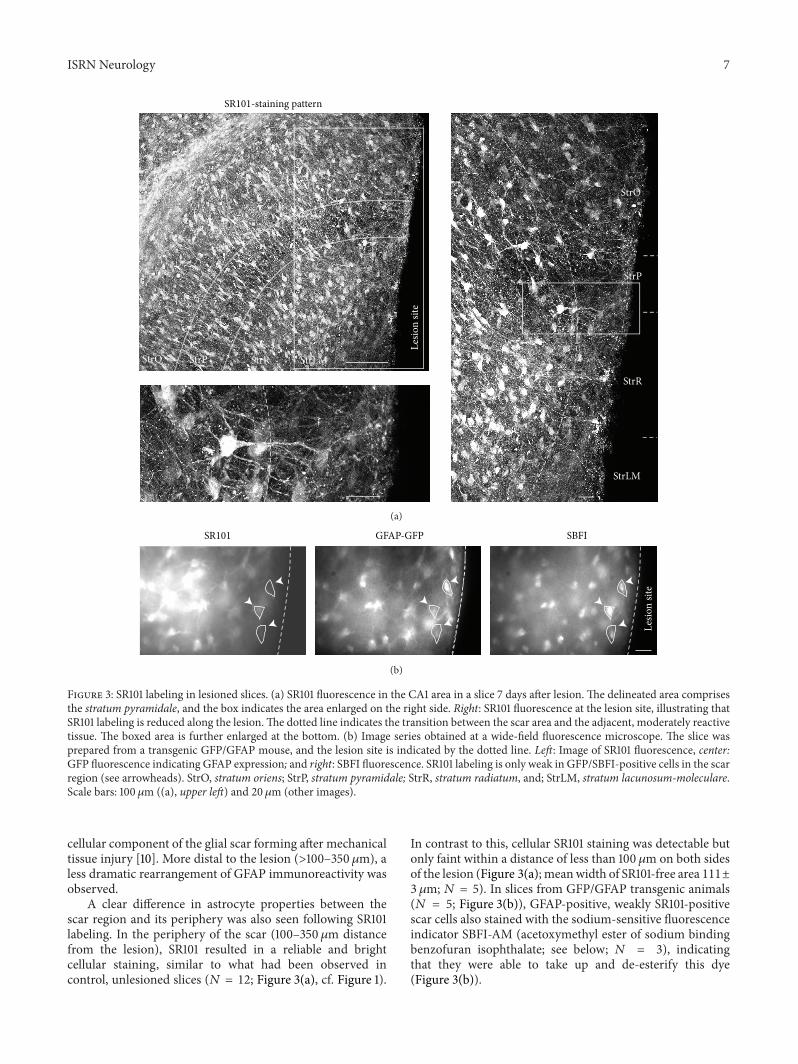
ISRN Neurology 7
StrR
StrP
StrLM
StrO
StrP StrRStrO StrLM
Lesio
n sit
e
SR101-staining pattern
(a)
SBFI
Lesio
n sit
e
SR101 GFAP-GFP
(b)
Figure 3 SR101 labeling in lesioned slices (a) SR101 fluorescence in the CA1 area in a slice 7 days after lesion The delineated area comprisesthe stratum pyramidale and the box indicates the area enlarged on the right side Right SR101 fluorescence at the lesion site illustrating thatSR101 labeling is reduced along the lesionThe dotted line indicates the transition between the scar area and the adjacent moderately reactivetissue The boxed area is further enlarged at the bottom (b) Image series obtained at a wide-field fluorescence microscope The slice wasprepared from a transgenic GFPGFAP mouse and the lesion site is indicated by the dotted line Left Image of SR101 fluorescence centerGFP fluorescence indicating GFAP expression and right SBFI fluorescence SR101 labeling is only weak in GFPSBFI-positive cells in the scarregion (see arrowheads) StrO stratum oriens StrP stratum pyramidale StrR stratum radiatum and StrLM stratum lacunosum-moleculareScale bars 100120583m ((a) upper left) and 20 120583m (other images)
cellular component of the glial scar forming after mechanicaltissue injury [10] More distal to the lesion (gt100ndash350 120583m) aless dramatic rearrangement of GFAP immunoreactivity wasobserved
A clear difference in astrocyte properties between thescar region and its periphery was also seen following SR101labeling In the periphery of the scar (100ndash350 120583m distancefrom the lesion) SR101 resulted in a reliable and brightcellular staining similar to what had been observed incontrol unlesioned slices (119873 = 12 Figure 3(a) cf Figure 1)
In contrast to this cellular SR101 staining was detectable butonly faint within a distance of less than 100 120583m on both sidesof the lesion (Figure 3(a) meanwidth of SR101-free area 111plusmn3 120583m 119873 = 5) In slices from GFPGFAP transgenic animals(119873 = 5 Figure 3(b)) GFAP-positive weakly SR101-positivescar cells also stained with the sodium-sensitive fluorescenceindicator SBFI-AM (acetoxymethyl ester of sodium bindingbenzofuran isophthalate see below 119873 = 3) indicatingthat they were able to take up and de-esterify this dye(Figure 3(b))
8 ISRN Neurology
lowastlowastlowast
lowastlowastlowast
80
lowast
Control ScarPeriphery
120
160
0
40
200
Som
a siz
e (120583
m2)
(a)
Lesioned
StrP
StrR
StrP
Control
StrR
StrLM
Lesio
n sit
e
(b)
Figure 4 Lesion-induced changes in somatic area (a) Quantification of somatic area based on GFAPGFP fluorescence in control andlesioned slices The soma size increased both in astrocytes in the scar region (ldquoscarrdquo) and in astrocytes in the periphery of the lesion(ldquoperipheryrdquo lowastlowastlowast119875 lt 0001 lowast119875 lt 005) (b) Confocal images of GFP fluorescence in cultured slices obtained from GFPGFAP transgenicanimals under control conditions (left) and subjected to a lesion (right) Cells surrounded by boxes are shown at higher magnification in theinsets and the dashed lines around the soma illustrate the areas which served to determine their size StrP stratum pyramidale StrR stratumradiatum StrLM stratum lacunosum moleculare and scale bars 40 120583m and 10120583m (inset)
In addition to GFAP-positive cells the scar region hostedmicroglial cells identified by their vital staining with TexasRed-coupled lectin from lycopersicon esculentum (tomatolectin119873 = 4 not shown) Tomato lectin-positive microgliahowever did not take up SBFI-AM (119873 = 4 not shown) con-firming earlier reports thatmicroglial cells do not incorporateAM-ester dyes in situ [47]
To further characterize reactive gliosis in response to thescratch wound we determined the area of astrocyte somatain confocal images of the GFP fluorescence in control andlesioned slices obtained fromGFPGFAP transgenic animalsThesemeasurements revealed that compared to astrocytes in
control slices the soma area of astrocytes in the periphery ofa lesion was increased by 18 and that of astrocytes in thescar regionwas increased by 24 (control 163plusmn2 120583m2 lesionperiphery 193plusmn3 120583m2 scar tissue 201plusmn4 120583m2119875 lt 0001 119899 =945 496 and 439 in 7 control and 9 lesioned slices Figures4(a) and 4(b)) Thus astrocytes show a second hallmark ofreactive gliosis after lesion namely a hypertrophy of their cellbodies which is observed both in the scar tissue as well as inthe periphery of the lesion
Taken together these data show that 6-7 days after themechanical lesion a glial scar has formed along the lesionsite This scar encompasses strongly activated astrocytes
ISRN Neurology 9
characterized by a robust increase in GFAP and S100120573expressionThese proximal reactive astrocytes locatedwithina distance of less than 100120583m from the lesion exhibit longGFAP-positive processes have significantly swollen somataand only weakly stain with SR101 Astrocytes in the peripheryof the scar (100ndash350 120583m from the lesion) display swollen cellbodies as well but show onlymoderate upregulation of GFAPand maintain their ability to efficiently take up SR101
33 Expression Levels and Spatial Distribution of GLT-1 andGLAST To determine the spatial distribution of the gluta-mate transporters GLAST and GLT-1 in organotypic slicesand changes therein in response to the lesion immunohisto-chemistry was employed In control slices GLAST (119873 = 13Figure 5(a)) as well as GLT-1 immunoreactivity (119873 = 11Figure 5(b)) appeared as punctate labeling throughout theentire CA1 area The staining patterns for both transportersin the tissue slice cultures were thus similar to those reportedfrom acute tissue slice preparations of the hippocampus [3648 49]
The relatively homogeneous distribution of immunore-activity throughout the preparation found in control slicecultures was abandoned following a lesion Here distinctaccumulations of GLAST (119873 = 17 Figure 5(a)) and GLT-1immunoreactivity (119873 = 12 Figure 5(b)) were found in thescar region In both cases these preferentially colocalizedwith thick GFAP-positive bundles running in parallel to thelesion (Figure 5) Apart from the marked accumulation ofGLAST reactivity in parallel to the scar GLAST immunore-activity appeared generally increased in some lesioned slicepreparations (Figure 5(a)) but unaltered in others bothwithinthe scar as well as in its periphery For GLT-1 a decline in theintensity of immunoreactivity was observed with increasingdistance from the lesion (Figure 5(b))
In summary these results reveal distinct alterations inthe immunoreactivity of GLAST and GLT-1 following amechanical lesion In the scar region clusters of GLT-1 andGLAST immunoreactivity are present primarily colocalizedwith strongly GFAP-positive fibers Apart from these distinctclusters GLT-1 immunoreactivity appears to decline withdistance from the scar whereas GLAST immunoreactivityseems largely uniform throughout the preparation
34 Sodium Transients Induced by Activation of GlutamateUptake Glutamate uptake is accompanied by the inwardflux of sodium resulting in increase in the intracellularsodium concentration of astrocytes To monitor its functionwe assayed sodium increases induced by D-aspartate anonmetabolized substrate of sodium-dependent glutamateuptake which is also transported [1] in astrocytes in thestratum radiatum of organotypic slices To this end SR101-stained slices were additionally loaded with the sodium-sensitive fluorophore SBFI-AM (Figure 6(a) cf Figure 3(b))In situ calibrations of the SBFI fluorescence revealed a linearincrease in its fluorescence ratio at intracellular sodiumconcentrations between 10 and 40mM (119899 = 107 Figure 6(b))as reported before [33]
Repetitive pressure application of D-aspartate for 500msinduced reliable increases in the intracellular sodium con-centration in astrocytes indicating activation of sodium-dependent glutamate uptake (119899 = 488 Figure 6(c)) Theexperiments were performed in the presence of the sodium-channel blocker TTX (tetrodotoxin 05 120583M) to suppressaction potential generation The amplitude of D-aspartate-induced sodium increases was dependent on the concen-tration applied (Figure 6(c)) and saturated at about 2mM(119899 = 19 Figure 6(d)) as reported before for mouse corticalastrocytes in culture [50] At a concentration of 1mM theamplitude of D-aspartate-induced sodium transients wasnear maximum and amounted to 38 plusmn 02mM (119899 =161 Figures 6(c) and 6(d)) a value comparable to that inastrocytes in acute slices obtained in our lab [33]
To probe for changes in glutamate uptake we comparedsodium signals induced by 1mMD-aspartate between controland lesioned slices (Figure 7) In lesioned slices astrocyteslocated in the periphery of the lesion (100ndash350 120583m distance)responded with a sodium transient to application of 1mMD-aspartate (see cell 2 in Figures 7(a) and 7(b)) The amplitudeof D-aspartate-induced sodium signals in cells in the lesionperiphery was similar to that observed in control slices (38 plusmn03mM 119899 = 111 see cell 1 in Figures 7(a) 7(b) and 7(c))Application of D-aspartate also evoked sodium transientsin weakly SR101-positive SBFI-filled astrocytes located inthe glial scar area (lt100 120583m from the lesion see cells 3ndash5in Figures 7(a) and 7(b)) These cells however displayedsignificantly smaller peak amplitudes (16 plusmn 021mM 119899 =26 119875 lt 0001 Figures 7(b) and 7(c)) than astrocytes inthe periphery and in control slices
The kinetics of D-aspartate-induced sodium transientsdiffered between all three groups This was true for theslope of the sodium increase (10ndash90 change) which wassignificantly reduced in astrocytes from lesioned slices versusastrocytes from control slices (09 plusmn 005mMs in controlversus 06 plusmn 006mMs in the periphery and 02 plusmn 004mMsin the scar region 119899 = 161 111 and 26 resp 119875 lt 0001Figures 7(b) and 7(c)) In addition the decay back to baselinewas significantly slower following lesion (Figures 7(b) and7(c)) The time needed for the signal to decay by 67 wasapproximately doubled (decay time 120591 was 44 plusmn 2 s in control120591 = 81 plusmn 4 s in periphery of the lesion and 120591 = 95 plusmn 10 s inscar cells Figures 7(b) and 7(c))
These results show that mechanical injury results indistinct alterations in the kinetics andor amplitudes ofD-aspartate-induced intracellular sodium transients in reac-tive astrocytes depending on their distance to the lesion
4 Discussion
Our results show that within 6-7 days after setting a scratchwound in organotypic hippocampal slices a dense glial scarhad formed along the lesion site Up to approximately 100 120583mdistance from the lesion the scar tissue was characterized bya substantial elevation ofGFAP expression a pronounced cel-lular hypertrophy and reduced uptake of SR101 by astrocytes
10 ISRN Neurology
Con
trol
GLAST
Lesio
ned
GFAP Merge
StrR
StrP
StrO
Lesio
n sit
e
StrR
(a)
Con
trol
Lesio
ned
GLT-1 GFAP Merge
StrR
StrP
StrO
StrR
StrP
StrLM
Lesio
n sit
e
(b)
Figure 5 GLAST andGLT-1 immunoreactivity aftermechanical injury (a) Fluorescence images of immunohistochemical stainings detectingthe glutamate transporters GLAST (left) GFAP (center) as counterstaining and the corresponding merged image (GFAP green GLASTmagenta) in control (top row) and lesioned slice cultures (bottom row) (b) GLT-1 (left) GFAP (center) double labeling of a control (toprow) and a mechanically lesioned slice (bottom row) The corresponding merge (GFAP green GLT-1 magenta) is found on the right Notethat after lesion glutamate transporter immunoreactive clusters are accumulated in the direct vicinity of the lesion site (arrowheads) GFAPglial fibrillary acidic protein GLAST glutamate aspartate transporter GLT-1 glutamate-transporter-1 StrO stratum oriens StrP stratumpyramidale StrR stratum radiatum StrLM stratum lacunosum-moleculare and scale bars (a) 20 120583m (b) 40 120583m
While prominent clusters of GLAST and GLT-1 immunore-activity were observed alongGFAP-positive structures in scarcells D-aspartate-induced intracellular sodium signals werestrongly dampened indicating a significant reduction in glu-tamate uptake capacity close to the lesion In the periphery ofthe lesion (100ndash350120583m) astrocytes showed less pronounced
reactivity maintained their ability to efficiently take up SR101and displayed only minor changes in glutamate transporterimmunoreactivity and function Thus depending on theirdistance from the lesion astrocytes showed different gradesof reactivity and displayed discrete changes in glutamatetransporter immunoreactivity and uptake capacity
ISRN Neurology 11
SR101 MergeSBFI
StrRStrP
(a)
03
04
05
06
07
0 5 10 15
0
20 25 30 40 0
100
20
40
80
60
0 10 20 30 400
20
40
60
80
100
10 = 55 mM
107 cells in 6 experiments
F (340
380
) (ra
tio)
Intracellular Na+ concentration (mM)Intracellular Na+ concentration (mM)
ΔF
(340
380
) (
)
ΔF
(340
380
) (
) R2 = 097
(b)
D-aspartate
100 120583M500 120583M
750 120583M1000 120583M
1 mM
10 s
Δ
(c)
0 1 2 3 4 50
1
2
3
4
5
Δ so
dium
(mM
)
D-aspartate (mM)
lowast
(d)
Figure 6 Sodium changes induced by activation of glutamate uptake (a) Staining of astrocytes by colabeling with SR101 and SBFI Leftepifluorescence image of theCA1-area of an SR101-loaded organotypic sliceCenter SBFI fluorescence taken from the same areaRight overlaySBFI-loaded SR101-positive astrocytes appear in white (arrowheads) StrP stratum pyramidale StrR stratum radiatum scale bar 20 120583m (b)Left calibration of SBFI fluorescence in an astrocyte Depicted is the change in intracellular SBFI fluorescence ratio in the calibration solutionscontaining ionophores and in response to stepwise changes in the sodium concentration Right mean values plusmn SEM of the normalizedchanges in SBFI fluorescence at different sodium concentrations The black line represents a linear fit of the data points between 10 and40mM sodium illustrating that a 10 change in SBFI fluorescence represents a change in the sodium concentration by 55mM (c) Sodiumtransients in a single astrocyte induced by 500ms applications of D-aspartate at different concentrations (d) Mean values plusmn S E M (119899 = 19)of the peak amplitude of sodium transients at different concentrations of D-aspartate The curve represents a sigmoidal fit of the data
41 Characteristics of Lesion-Induced Reactive Gliosis inOrganotypic Cultures Organotypic tissue slice cultures ofthe hippocampus represent a well-established model systemin which the basic tissue architecture is maintained butwhich still hosts the major advantages of cell cultures suchas good accessibility and control of experimental conditionsThey have been used extensively for the analysis of neuronal
properties and developing neuronal networks [28 51 52]or the study of excitotoxic neuronal damage (eg [53 54])Recent studies also demonstrated that basic morphologicalcharacteristics of astrocytes and the typical glia-synapseorganization are well preserved in organotypic slice cultures(eg [44 45 55 56]) In addition we found a nearly completeoverlap of SR101 staining with the expression of GFP in slice
12 ISRN Neurology
SBFIControl Lesioned
SR101
1
Lesio
n sit
e
SR101 SBFI
2 45
32 4
5
1
StrR
StrP StrP
StrR
3
(a)
ControlScar
Lesioned
1
D-asp
2
D-asp
D-asp
5
4
3Periphery1mM
20 s
Δ Δ
Δ
Δ
Δ
(b)
0102030405060708090
100110
Δ so
dium
tim
e (m
Ms
)
00
02
04
06
08
10
12
0
1
2
3
4
5
6
ns
Control Periphery Scar Control Periphery Scar Control Periphery Scar
Amplitude Slope Decay time
Δ so
dium
(mM
)
120591(s
)lowastlowastlowast
lowastlowastlowastlowastlowastlowast
lowastlowastlowastlowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowast
(c)
Figure 7 D-aspartate-induced astrocyte sodium transients following lesion (a) CA1 area of a control (left) and a lesioned (right) slicepreparation Shown are epifluorescence images of SR101 and SBFI fluorescences Circled areas and numbers indicate the regions of interestanalyzed in the experiment depicted in (b) StrP stratum pyramidale StrR stratum radiatum and scale bar 40 120583m (b) Sodium transientsinduced by pressure application of 1mMD-aspartate (D-asp) for 500ms in the different cells in the control and lesioned slice as indicated in(a) (c) Histograms showing mean values plusmn S E M of the peak amplitude (left) slope (center) and decay time (right) of D-aspartate-inducedsodium transients in astrocytes in control slices (119899 = 161) as well as in cells in the scar region and its periphery (119899 = 111 and 119899 = 26 resp)lowastlowast119875 lt 001 lowastlowastlowast119875 lt 0001 StrP stratum pyramidale StrR stratum radiatum and scale bar 40120583m
cultures obtained from GFPGFAP mice demonstrating thatSR101 is well suited to identify astrocytes in this prepara-tion as described before for acute tissue slices and in vivo[29 30]
Performing a scratch through the CA1 region of organ-otypic slices resulted in the formation of a glial scar along thelesion site as judged by the prominent increase in expressionof GFAP and S100120573 In addition astrocyte cell bodies showed
a significant hypertrophy Furthermore astrocytes in theperiphery of the scar (100ndash350 120583m from the lesion) showedmild to moderate reactivity displaying swollen cell bodiesand only moderate upregulation of GFAP These changesare hallmarks of reactive gliosis observed after tissue injury[10 13 57] Generally the degree of injury and the distance ofthe astrocytes from the site of injury define the degree of theiractivation [11 13]
ISRN Neurology 13
Our study also revealed that strongly reactive astrocytesin the scar region only weakly stain with SR101 The reducedability of strongly activated astrocytes to accumulate SR101indicates that strong glial activation is accompanied by adownregulation of the organic anion transporter responsi-ble for uptake of SR101 This is reminiscent of immatureastrocytes in the early postnatal hippocampus which areSR101-negative [30] and in line with a wealth of experimentalevidence suggesting that reactive gliosis represents a processcomprising a dedifferentiation of astrocytes [17]
42 Changes in Glutamate Transporter Expression followingLesion Glutamate uptake in the hippocampus is mainlyachieved by the glial glutamate transporters GLT-1 andGLAST [1 2] A central factor in many brain pathologiesincluding traumatic brain injury is an increase in extracellu-lar glutamate and excitotoxicity it has been suggested that achange in the expression levels of glial glutamate transportersmight play a critical role in the failure of glutamate clearance[6 58ndash60] Earlier work has found an overall downregulationof protein levels of both GLAST and GLT-1 following astro-cyte activation [22ndash27] indicating that this is causal to theelevation of extracellular glutamate In contrast to this notionother studies reported increased glutamate transport capacityof reactive astrocytes and suggested a protective influence[61 62]
To visualize possible differences in the spatial expressionprofile of GLT-1 and GLAST in strongly reactive astrocytesalong the scar as compared to moderately reactive astro-cytes in its periphery we performed immunohistochemicalstainings These revealed an accumulation of GLT-1 andGLAST immunoreactivity along thick GFAP-positive fiberswhich was particularly pronounced for GLAST Besides theseclusters overall GLAST immunoreactivity seemed unalteredas compared to unlesioned slices whereas GLT-1 immunore-activity seemedweaker and declined with increasing distancefrom the scar Clustering of glutamate transporters has beendescribed by several reports [48 63 64] In developinghippocampal astrocytes cluster formation was preferentiallyfound in branches opposed to synapses and was increasedwith increased neuronal activity indicating that it is neces-sary to copewith synaptic release of glutamate [36] Along thesame lines it was observed that sustained astroglial activationby ciliary neurotrophic factor (CNTF) in the rat striatuminduced a concentration of GLAST and GLT-1 into raftmicrodomains and improved glutamate clearance indicatingthat cluster formation increased the cellular capacity for glu-tamate uptake [61] However another study found glutamate-induced clustering of GLT-1 that induced its endocytosis andintracellular trafficking without changing the total expressionlevels as detected by western blots arguing for a decrease infunctional glial glutamate uptake capacity [49] In the presentstudy antibodies against GLT-1 and GLAST were employedfor immunohistochemistry after permeabilization of theplasma membrane The clusters of GLAST and GLT-1 alongGFAP-positive fibers in the scar region (as GLAST andGLT-1immunoreactivity in general) could thus represent glutamatetransporters in the plasmamembrane or in intracellular com-partments or both Consequently the observed clustering
does not allow a prediction about possible functional changesin glutamate uptake
43 Lesion-Induced Changes in Functional Glutamate UptakeCapacity We probed for the functional activation of gluta-mate transport by application of the transportable agonistD-aspartate which results in an increase in the intracellularsodium concentration of astrocytes [8 9] The amplitudesof D-aspartate-induced sodium transients in astrocytes incontrol organotypic slices were similar to those reportedfrom astrocytes in acute slices obtained in our lab [33]indicating similar cellular glutamate transport capacity inboth preparations
Following a lesion D-aspartate-induced intracellularsodium signalswere clearly dampened in scar cellsMoreoverthe slope of the increase in sodium was significantly reducedand the decay back to baseline was slowed Because D-aspartate was applied at nearly saturating concentrationthe reduction in peak amplitudes suggests a reduction infunctional glutamate uptake capacity in cells close to thelesion which might be mediated by a decrease in the overallnumber of glutamate transporters available at the plasmamembrane In contrast the alteration in the kinetics of theD-aspartate-induced sodium signals can be explained by thecellular hypertrophy that was observed in reactive astrocytesAt an equal transport-mediated sodium influx across themembrane the slope of resulting changes in the sodiumconcentration will be decreased in cells with a larger volume
Furthermore the decay of intracellular sodium transientsback to baseline is mainly governed by the activity of theNa+K+-ATPase as well as by diffusion [8] The slower recov-ery is in line with earlier studies reporting a downregulationof the sodium pump following reactive gliosis [65 66]Moreover sodium is not buffered in the cell and travels solelyby means of diffusion in the cytoplasm Hindered diffusionresulting from increased cytoplasmic protein content orimpaired gap junction coupling which both occur uponastrocytic activation might thus also partially contribute todelayed sodium recovery [12 34 67]
In contrast to scar cells the amplitude of D-aspartate-induced intracellular sodium signals was not significantlyaltered in moderately activated astrocytes in the periphery ofthe lesion indicating that the overall number of functionalglutamate transporters was not significantly altered Becausethese cells showed hypertrophy as well the slowed kinetics ofthe sodium transients are in line with an increased cellularvolume as well as a possible downregulation of Na+K+-ATPase andor a slowed diffusion as argued above
Taken together our results indicate a significant reduc-tion in glutamate uptake capacity in strongly activated astro-cytes in the scar region Thus the prominent clustering ofGLAST and GLT-1 immunoreactivity along GFAP-positivestructures close to the lesion is likely to reflect a loss ofglutamate transporters from the plasma membrane upontransporter internalization as reported by Nakagawa et al[49] The authors of the latter study speculated that large andprolonged increases in glutamate concentrations are neces-sary to induce such cluster formation and endocytosis in thetissue as they would possibly only occur under pathological
14 ISRN Neurology
conditions Indeed ambient glutamate concentrations wereshown to be significantly increased after traumatic braininjury [68ndash70] Along these lines it can be assumed thatin our model system extracellular glutamate concentrationsrose less severely in the periphery of the lesion preventinga comparable clustering and loss of functional glutamatetransport activity in moderately activated astrocytes
5 Conclusions
Our results show complex changes in glutamate transporterexpression and function during astrocyte activation followingmechanical injury They confirm that the degree of astrocyteactivation depends on the distance from the insult and com-prises different endpoints regarding cellular morphology andphysiology Furthermore our results highlight that astrocyteswhich show different grades of reactivity also display discretechanges in glutamate transporter expression and functionWhile immunohistochemistry revealed a prominent clus-tering of GLT-1 and GLAST immunoreactivity in the scarregion our functional assay clearly showed that glutamateuptake capacity is strongly reduced in scar cells while itis largely maintained in moderately activated astrocytes inthe periphery Thus mild-to-moderate astrogliosis in theperiphery of a mechanical lesion does not necessarily seemto be accompanied by a significant change in glial glutamateuptake capacity At the glial scar itself in strongly reactiveastrocytes a clustering of glutamate transporters is observedthat apparently goes along with a severe functional reductionin astroglial glutamate uptake which may contribute toglutamate-mediated excitotoxicity in this region
Conflict of Interests
The authors of the paper declare that they do not haveany direct financial relation with the commercial identitiesmentioned in the paper that might lead to a conflict ofinterests
Acknowledgments
This study was supported by the Deutsche Forschungsge-meinschaft (DFG Ro23274-3) The authors thank SimoneDurry and Claudia Roderigo for expert technical help
References
[1] N C Danbolt ldquoGlutamate uptakerdquo Progress in Neurobiologyvol 65 no 1 pp 1ndash105 2001
[2] A V Tzingounis and J I Wadiche ldquoGlutamate transportersconfining runaway excitation by shaping synaptic transmis-sionrdquo Nature Reviews Neuroscience vol 8 no 12 pp 935ndash9472007
[3] D E Bergles J S Diamond and C E Jahr ldquoClearance ofglutamate inside the synapse and beyondrdquo Current Opinion inNeurobiology vol 9 no 3 pp 293ndash298 1999
[4] C M Anderson and R A Swanson ldquoAstrocyte glutamatetransport review of properties regulation and physiologicalfunctionsrdquo Glia vol 32 no 1 pp 1ndash14 2000
[5] P Marcaggi and D Attwell ldquoRole of glial amino acid trans-porters in synaptic transmission and brain energeticsrdquoGlia vol47 no 3 pp 217ndash225 2004
[6] N J Maragakis and J D Rothstein ldquoGlutamate transportersanimal models to neurologic diseaserdquo Neurobiology of Diseasevol 15 no 3 pp 461ndash473 2004
[7] G Gegelashvili and A Schousboe ldquoCellular distribution andkinetic properties of high-affinity glutamate transportersrdquoBrainResearch Bulletin vol 45 no 3 pp 233ndash238 1998
[8] J W Deitmer and C R Rose ldquoIon changes and signalling inperisynaptic gliardquo Brain Research Reviews vol 63 no 1-2 pp113ndash129 2010
[9] S Kirischuk V Parpura and A Verkhratsky ldquoSodium dynam-ics another key to astroglial excitabilityrdquo Trends in Neuro-sciences vol 35 no 8 pp 497ndash506 2012
[10] M V Sofroniew ldquoMolecular dissection of reactive astrogliosisand glial scar formationrdquo Trends in Neurosciences vol 32 no12 pp 638ndash647 2009
[11] J L Ridet S K Malhotra A Privat and F H Gage ldquoReactiveastrocytes cellular and molecular cues to biological functionrdquoTrends in Neurosciences vol 20 no 12 pp 570ndash577 1997
[12] J Middeldorp and E M Hol ldquoGFAP in health and diseaserdquoProgress in Neurobiology vol 93 no 3 pp 421ndash443 2011
[13] M Pekny and M Nilsson ldquoAstrocyte activation and reactivegliosisrdquo Glia vol 50 no 4 pp 427ndash434 2005
[14] A Bordey S A Lyons J J Hablitz and H SontheimerldquoElectrophysiological characteristics of reactive astrocytes inexperimental cortical dysplasiardquo Journal of Neurophysiologyvol 85 no 4 pp 1719ndash1731 2001
[15] R Jabs G Seifert and C Steinhauser ldquoAstrocytic functionand its alteration in the epileptic brainrdquo Epilepsia vol 49supplement 2 pp 3ndash12 2008
[16] A Buffo I Rite P Tripathi et al ldquoOrigin and progeny ofreactive gliosis a source of multipotent cells in the injuredbrainrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 105 no 9 pp 3581ndash3586 2008
[17] S Robel B Berninger and M Gotz ldquoThe stem cell potential ofglia lessons from reactive gliosisrdquoNature Reviews Neurosciencevol 12 no 2 pp 88ndash104 2011
[18] H Kawano J Kimura-Kuroda Y Komuta et al ldquoRole of thelesion scar in the response to damage and repair of the centralnervous systemrdquo Cell and Tissue Research vol 349 no 1 pp169ndash180 2012
[19] G Yiu and Z He ldquoGlial inhibition of CNS axon regenerationrdquoNature Reviews Neuroscience vol 7 no 8 pp 617ndash627 2006
[20] M T Fitch and J Silver ldquoCNS injury glial scars and inflamma-tion inhibitory extracellular matrices and regeneration failurerdquoExperimental Neurology vol 209 no 2 pp 294ndash301 2008
[21] L Li A Lundkvist D Andersson et al ldquoProtective role ofreactive astrocytes in brain ischemiardquo Journal of Cerebral BloodFlow amp Metabolism vol 28 no 3 pp 468ndash481 2008
[22] F K H van Landeghem T Weiss M Oehmichen and AVonDeimling ldquoDecreased expression of glutamate transportersin astrocytes after human traumatic brain injuryrdquo Journal ofNeurotrauma vol 23 no 10 pp 1518ndash1528 2006
[23] V L Raghavendra Rao M K Baskaya A Dogan J DRothstein and R J Dempsey ldquoTraumatic brain injury down-regulates glial glutamate transporter (GLT- 1 and GLAST)proteins in rat brainrdquo Journal of Neurochemistry vol 70 no 5pp 2020ndash2027 1998
ISRN Neurology 15
[24] J-C Chen H Hsu-Chou J-L Lu et al ldquoDown-regulation ofthe glial glutamate transporter GLT-1 in rat hippocampus andstriatum and its modulation by a group III metabotropic glu-tamate receptor antagonist following transient global forebrainischemiardquoNeuropharmacology vol 49 no 5 pp 703ndash714 2005
[25] S D Ginsberg L J Martin and J D Rothstein ldquoRegional deaf-ferentation down-regulates subtypes of glutamate transporterproteinsrdquo Journal of Neurochemistry vol 65 no 6 pp 2800ndash2803 1995
[26] J E Springer R D Azbill R J Mark J G Begley GWaeg andM P Mattson ldquo4-Hydroxynonenal a lipid peroxidation prod-uct rapidly accumulates following traumatic spinal cord injuryand inhibits glutamate uptakerdquo Journal of Neurochemistry vol68 no 6 pp 2469ndash2476 1997
[27] M B Moretto N S Arteni D Lavinsky et al ldquoHypoxic-ischemic insult decreases glutamate uptake by hippocampalslices from neonatal rats Prevention by guanosinerdquo Experimen-tal Neurology vol 195 no 2 pp 400ndash406 2005
[28] L Stoppini P-A Buchs and D Muller ldquoA simple method fororganotypic cultures of nervous tissuerdquo Journal of NeuroscienceMethods vol 37 no 2 pp 173ndash182 1991
[29] A Nimmerjahn F Kirchhoff J N D Kerr and F HelmchenldquoSulforhodamine 101 as a specific marker of astroglia in theneocortex in vivordquo Nat Methods vol 1 no 1 pp 31ndash37 2004
[30] K W Kafitz S D Meier J Stephan and C R Rose ldquoDevel-opmental profile and properties of sulforhodamine 101-Labeledglial cells in acute brain slices of rat hippocampusrdquo Journal ofNeuroscience Methods vol 169 no 1 pp 84ndash92 2008
[31] C Schnell Y Hagos and S Hulsmann ldquoActive sulforhodamine101 uptake into hippocampal astrocytesrdquo PLoS One vol 7 no 11Article ID e49398 2012
[32] S D Meier Y Kovalchuk and C R Rose ldquoProperties ofthe new fluorescent Na+ indicator CoroNa Green comparisonwith SBFI and confocal Na+ imagingrdquo Journal of NeuroscienceMethods vol 155 no 2 pp 251ndash259 2006
[33] J Langer and C R Rose ldquoSynaptically induced sodium signalsin hippocampal astrocytes in siturdquo Journal of Physiology vol587 part 24 pp 5859ndash5877 2009
[34] J Langer J Stephan M Theis and C R Rose ldquoGap junctionsmediate intercellular spread of sodium between hippocampalastrocytes in siturdquo Glia vol 60 no 2 pp 239ndash252 2012
[35] C R Rose andB R Ransom ldquoIntracellular sodiumhomeostasisin rat hippocampal astrocytesrdquo Journal of Physiology vol 491part 2 no 2 pp 291ndash305 1996
[36] A M Benediktsson G S Marrs J C Tu et al ldquoNeuronalactivity regulates glutamate transporter dynamics in developingastrocytesrdquo Glia vol 60 no 2 pp 175ndash188 2012
[37] M Zhou G P Schools and H K Kimelberg ldquoDevelopment ofGLAST(+) astrocytes andNG2(+) glia in rat hippocampus CA1mature astrocytes are electrophysiologically passiverdquo Journal ofNeurophysiology vol 95 no 1 pp 134ndash143 2006
[38] I Suarez G Bodega and B Fernandez ldquoModulation of glu-tamate transporters (GLAST GLT-1 and EAAC1) in the ratcerebellum following portocaval anastomosisrdquo Brain Researchvol 859 no 2 pp 293ndash302 2000
[39] X Zhu D E Bergles and A Nishiyama ldquoNG2 cells generateboth oligodendrocytes and gray matter astrocytesrdquo Develop-ment vol 135 no 1 pp 145ndash157 2008
[40] B Brunne S ZhaoADerouiche et al ldquoOriginmaturation andastroglial transformation of secondary radial glial cells in thedeveloping dentate gyrusrdquo Glia vol 58 no 13 pp 1553ndash15692010
[41] Y Wu A-Q Zhang and D T Yew ldquoAge related changes ofvarious markers of astrocytes in senescence-accelerated micehippocampusrdquo Neurochemistry International vol 46 no 7 pp565ndash574 2005
[42] S Magavi D Friedmann G Banks A Stolfi and C LoisldquoCoincident generation of pyramidal neurons and protoplasmicastrocytes in neocortical columnsrdquo Journal of Neuroscience vol32 no 14 pp 4762ndash4772 2012
[43] M Olabarria H N Noristani A Verkhratsky and J JRodrıguez ldquoConcomitant astroglial atrophy and astrogliosis ina triple transgenic animal model of Alzheimerrsquos diseaserdquo Gliavol 58 no 7 pp 831ndash838 2010
[44] A M Benediktsson S J Schachtele S H Green and M EDailey ldquoBallistic labeling and dynamic imaging of astrocytesin organotypic hippocampal slice culturesrdquo Journal of Neuro-science Methods vol 141 no 1 pp 41ndash53 2005
[45] I Lushnikova G Skibo D Muller and I Nikonenko ldquoSynapticpotentiation induces increased glial coverage of excitatorysynapses in CA1 hippocampusrdquoHippocampus vol 19 no 8 pp753ndash762 2009
[46] E Raponi F Agenes C Delphin et al ldquoS100B expressiondefines a state in which GFAP-expressing cells lose their neuralstem cell potential and acquire a more mature developmentalstagerdquo Glia vol 55 no 2 pp 165ndash177 2007
[47] C G Schipke C Boucsein C Ohlemeyer F Kirchhoff andH Kettenmann ldquoAstrocyte Ca2+ waves trigger responses inmicroglial cells in brain slicesrdquo The FASEB Journal vol 16 no2 pp 255ndash257 2002
[48] J Zhou and M L Sutherland ldquoGlutamate transporter clusterformation in astrocytic processes regulates glutamate uptakeactivityrdquo Journal of Neuroscience vol 24 no 28 pp 6301ndash63062004
[49] T Nakagawa Y Otsubo Y Yatani H Shirakawa and S KanekoldquoMechanisms of substrate transport-induced clustering of aglial glutamate transporter GLT-1 in astroglial-neuronal cul-turesrdquo European Journal of Neuroscience vol 28 no 9 pp 1719ndash1730 2008
[50] J-Y Chatton P Marquet and P J Magistretti ldquoA quantitativeanalysis of L-glutamate-regulated Na+ dynamics in mousecortical astrocytes Implications for cellular bioenergeticsrdquoEuropean Journal of Neuroscience vol 12 no 11 pp 3843ndash38532000
[51] E Forster S Zhao and M Frotscher ldquoLaminating the hip-pocampusrdquo Nature Reviews Neuroscience vol 7 no 4 pp 259ndash267 2006
[52] B H Gahwiler M Capogna D Debanne R A McKinney andS M Thompson ldquoOrganotypic slice cultures a technique hascome of agerdquo Trends in Neurosciences vol 20 no 10 pp 471ndash477 1997
[53] P E Kunkler and R P Kraig ldquoReactive astrocytosis fromexcitotoxic injury in hippocampal organ culture parallels thatseen in vivordquo Journal of Cerebral Blood Flow and Metabolismvol 17 no 1 pp 26ndash43 1997
[54] I E Holopainen ldquoOrganotypic hippocampal slice cultures amodel system to study basic cellular andmolecularmechanismsof neuronal cell death neuroprotection and synaptic plasticityrdquoNeurochemical Research vol 30 no 12 pp 1521ndash1528 2005
[55] MHaber L Zhou andK KMurai ldquoCooperative astrocyte anddendritic spine dynamics at hippocampal excitatory synapsesrdquoJournal of Neuroscience vol 26 no 35 pp 8881ndash8891 2006
16 ISRN Neurology
[56] H Nishida and S Okabe ldquoDirect astrocytic contacts regulatelocal maturation of dendritic spinesrdquo Journal of Neurosciencevol 27 no 2 pp 331ndash340 2007
[57] A Buffo C Rolando and S Ceruti ldquoAstrocytes in the dam-aged brain molecular and cellular insights into their reactiveresponse and healing potentialrdquoBiochemical Pharmacology vol79 no 2 pp 77ndash89 2010
[58] J-H Yi and A S Hazell ldquoExcitotoxic mechanisms and the roleof astrocytic glutamate transporters in traumatic brain injuryrdquoNeurochemistry International vol 48 no 5 pp 394ndash403 2006
[59] C Werner and K Engelhard ldquoPathophysiology of traumaticbrain injuryrdquo British Journal of Anaesthesia vol 99 no 1 pp4ndash9 2007
[60] G Gegelashvili M B Robinson D Trotti and T RauenldquoRegulation of glutamate transporters in health and diseaserdquoProgress in Brain Research vol 132 pp 267ndash286 2001
[61] C Escartin E Brouillet PGubellini et al ldquoCiliary neurotrophicfactor activates astrocytes redistributes their glutamate trans-porters GLAST and GLT-1 to raft microdomains and improvesglutamate handling in vivordquo Journal of Neuroscience vol 26 no22 pp 5978ndash5989 2006
[62] C Vermeiren M Najimi N Vanhoutte et al ldquoAcute up-regulation of glutamate uptake mediated by mGluR5a in reac-tive astrocytesrdquo Journal of Neurochemistry vol 94 no 2 pp405ndash416 2005
[63] C L Poitry-Yamate L Vutskits and T Rauen ldquoNeuronal-induced and glutamate-dependent activation of glial glutamatetransporter functionrdquo Journal of Neurochemistry vol 82 no 4pp 987ndash997 2002
[64] P M Beart and R D OrsquoShea ldquoTransporters for L-glutamatean update on their molecular pharmacology and pathologicalinvolvementrdquo British Journal of Pharmacology vol 150 no 1pp 5ndash17 2007
[65] F D Lima M A Souza A F Furian et al ldquoNa+K+-ATPaseactivity impairment after experimental traumatic brain injuryrelationship to spatial learning deficits and oxidative stressrdquoBehavioural Brain Research vol 193 no 2 pp 306ndash310 2008
[66] G A Gusarova H E Trejo L A Dada et al ldquoHypoxia leadsto NaK-ATPase downregulation via Ca2+ release-activatedCa2+ channels and AMPK activationrdquo Molecular and CellularBiology vol 31 no 17 pp 3546ndash3556 2011
[67] S S L Chew C S Johnson C R Green and H V Danesh-Meyer ldquoRole of connexin43 in central nervous system injuryrdquoExperimental Neurology vol 225 no 2 pp 250ndash261 2010
[68] P Nilsson L Hillered U Ponten and U Ungerstedt ldquoChangesin cortical extracellular levels of energy-related metabolites andamino acids following concussive brain injury in ratsrdquo Journal ofCerebral Blood Flow and Metabolism vol 10 no 5 pp 631ndash6371990
[69] H Katoh K Sima H Nawashiro K Wada and H ChigasakildquoThe effect of MK-801 on extracellular neuroactive amino acidsin hippocampus after closed head injury followed by hypoxia inratsrdquo Brain Research vol 758 no 1-2 pp 153ndash162 1997
[70] H Koizumi H Fujisawa H Ito T Maekawa X Di and RBullock ldquoEffects of mild hypothermia on cerebral blood flow-independent changes in cortical extracellular levels of aminoacids following contusion trauma in the ratrdquoBrain Research vol747 no 2 pp 304ndash312 1997
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
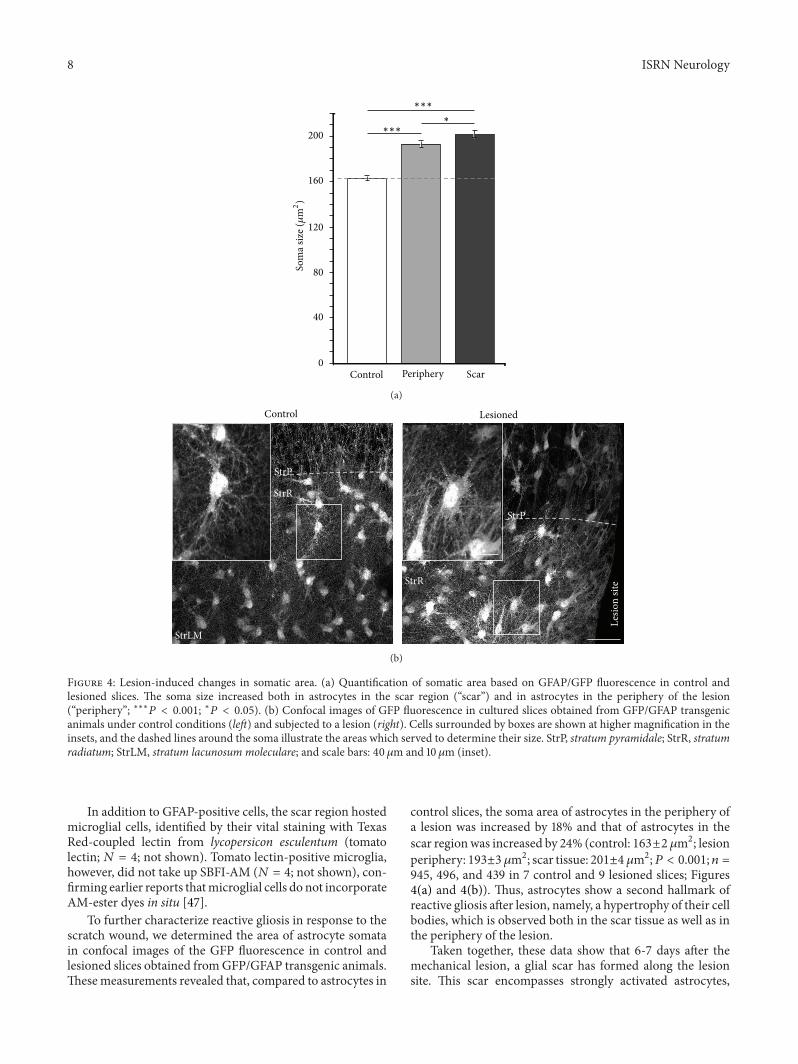
8 ISRN Neurology
lowastlowastlowast
lowastlowastlowast
80
lowast
Control ScarPeriphery
120
160
0
40
200
Som
a siz
e (120583
m2)
(a)
Lesioned
StrP
StrR
StrP
Control
StrR
StrLM
Lesio
n sit
e
(b)
Figure 4 Lesion-induced changes in somatic area (a) Quantification of somatic area based on GFAPGFP fluorescence in control andlesioned slices The soma size increased both in astrocytes in the scar region (ldquoscarrdquo) and in astrocytes in the periphery of the lesion(ldquoperipheryrdquo lowastlowastlowast119875 lt 0001 lowast119875 lt 005) (b) Confocal images of GFP fluorescence in cultured slices obtained from GFPGFAP transgenicanimals under control conditions (left) and subjected to a lesion (right) Cells surrounded by boxes are shown at higher magnification in theinsets and the dashed lines around the soma illustrate the areas which served to determine their size StrP stratum pyramidale StrR stratumradiatum StrLM stratum lacunosum moleculare and scale bars 40 120583m and 10120583m (inset)
In addition to GFAP-positive cells the scar region hostedmicroglial cells identified by their vital staining with TexasRed-coupled lectin from lycopersicon esculentum (tomatolectin119873 = 4 not shown) Tomato lectin-positive microgliahowever did not take up SBFI-AM (119873 = 4 not shown) con-firming earlier reports thatmicroglial cells do not incorporateAM-ester dyes in situ [47]
To further characterize reactive gliosis in response to thescratch wound we determined the area of astrocyte somatain confocal images of the GFP fluorescence in control andlesioned slices obtained fromGFPGFAP transgenic animalsThesemeasurements revealed that compared to astrocytes in
control slices the soma area of astrocytes in the periphery ofa lesion was increased by 18 and that of astrocytes in thescar regionwas increased by 24 (control 163plusmn2 120583m2 lesionperiphery 193plusmn3 120583m2 scar tissue 201plusmn4 120583m2119875 lt 0001 119899 =945 496 and 439 in 7 control and 9 lesioned slices Figures4(a) and 4(b)) Thus astrocytes show a second hallmark ofreactive gliosis after lesion namely a hypertrophy of their cellbodies which is observed both in the scar tissue as well as inthe periphery of the lesion
Taken together these data show that 6-7 days after themechanical lesion a glial scar has formed along the lesionsite This scar encompasses strongly activated astrocytes
ISRN Neurology 9
characterized by a robust increase in GFAP and S100120573expressionThese proximal reactive astrocytes locatedwithina distance of less than 100120583m from the lesion exhibit longGFAP-positive processes have significantly swollen somataand only weakly stain with SR101 Astrocytes in the peripheryof the scar (100ndash350 120583m from the lesion) display swollen cellbodies as well but show onlymoderate upregulation of GFAPand maintain their ability to efficiently take up SR101
33 Expression Levels and Spatial Distribution of GLT-1 andGLAST To determine the spatial distribution of the gluta-mate transporters GLAST and GLT-1 in organotypic slicesand changes therein in response to the lesion immunohisto-chemistry was employed In control slices GLAST (119873 = 13Figure 5(a)) as well as GLT-1 immunoreactivity (119873 = 11Figure 5(b)) appeared as punctate labeling throughout theentire CA1 area The staining patterns for both transportersin the tissue slice cultures were thus similar to those reportedfrom acute tissue slice preparations of the hippocampus [3648 49]
The relatively homogeneous distribution of immunore-activity throughout the preparation found in control slicecultures was abandoned following a lesion Here distinctaccumulations of GLAST (119873 = 17 Figure 5(a)) and GLT-1immunoreactivity (119873 = 12 Figure 5(b)) were found in thescar region In both cases these preferentially colocalizedwith thick GFAP-positive bundles running in parallel to thelesion (Figure 5) Apart from the marked accumulation ofGLAST reactivity in parallel to the scar GLAST immunore-activity appeared generally increased in some lesioned slicepreparations (Figure 5(a)) but unaltered in others bothwithinthe scar as well as in its periphery For GLT-1 a decline in theintensity of immunoreactivity was observed with increasingdistance from the lesion (Figure 5(b))
In summary these results reveal distinct alterations inthe immunoreactivity of GLAST and GLT-1 following amechanical lesion In the scar region clusters of GLT-1 andGLAST immunoreactivity are present primarily colocalizedwith strongly GFAP-positive fibers Apart from these distinctclusters GLT-1 immunoreactivity appears to decline withdistance from the scar whereas GLAST immunoreactivityseems largely uniform throughout the preparation
34 Sodium Transients Induced by Activation of GlutamateUptake Glutamate uptake is accompanied by the inwardflux of sodium resulting in increase in the intracellularsodium concentration of astrocytes To monitor its functionwe assayed sodium increases induced by D-aspartate anonmetabolized substrate of sodium-dependent glutamateuptake which is also transported [1] in astrocytes in thestratum radiatum of organotypic slices To this end SR101-stained slices were additionally loaded with the sodium-sensitive fluorophore SBFI-AM (Figure 6(a) cf Figure 3(b))In situ calibrations of the SBFI fluorescence revealed a linearincrease in its fluorescence ratio at intracellular sodiumconcentrations between 10 and 40mM (119899 = 107 Figure 6(b))as reported before [33]
Repetitive pressure application of D-aspartate for 500msinduced reliable increases in the intracellular sodium con-centration in astrocytes indicating activation of sodium-dependent glutamate uptake (119899 = 488 Figure 6(c)) Theexperiments were performed in the presence of the sodium-channel blocker TTX (tetrodotoxin 05 120583M) to suppressaction potential generation The amplitude of D-aspartate-induced sodium increases was dependent on the concen-tration applied (Figure 6(c)) and saturated at about 2mM(119899 = 19 Figure 6(d)) as reported before for mouse corticalastrocytes in culture [50] At a concentration of 1mM theamplitude of D-aspartate-induced sodium transients wasnear maximum and amounted to 38 plusmn 02mM (119899 =161 Figures 6(c) and 6(d)) a value comparable to that inastrocytes in acute slices obtained in our lab [33]
To probe for changes in glutamate uptake we comparedsodium signals induced by 1mMD-aspartate between controland lesioned slices (Figure 7) In lesioned slices astrocyteslocated in the periphery of the lesion (100ndash350 120583m distance)responded with a sodium transient to application of 1mMD-aspartate (see cell 2 in Figures 7(a) and 7(b)) The amplitudeof D-aspartate-induced sodium signals in cells in the lesionperiphery was similar to that observed in control slices (38 plusmn03mM 119899 = 111 see cell 1 in Figures 7(a) 7(b) and 7(c))Application of D-aspartate also evoked sodium transientsin weakly SR101-positive SBFI-filled astrocytes located inthe glial scar area (lt100 120583m from the lesion see cells 3ndash5in Figures 7(a) and 7(b)) These cells however displayedsignificantly smaller peak amplitudes (16 plusmn 021mM 119899 =26 119875 lt 0001 Figures 7(b) and 7(c)) than astrocytes inthe periphery and in control slices
The kinetics of D-aspartate-induced sodium transientsdiffered between all three groups This was true for theslope of the sodium increase (10ndash90 change) which wassignificantly reduced in astrocytes from lesioned slices versusastrocytes from control slices (09 plusmn 005mMs in controlversus 06 plusmn 006mMs in the periphery and 02 plusmn 004mMsin the scar region 119899 = 161 111 and 26 resp 119875 lt 0001Figures 7(b) and 7(c)) In addition the decay back to baselinewas significantly slower following lesion (Figures 7(b) and7(c)) The time needed for the signal to decay by 67 wasapproximately doubled (decay time 120591 was 44 plusmn 2 s in control120591 = 81 plusmn 4 s in periphery of the lesion and 120591 = 95 plusmn 10 s inscar cells Figures 7(b) and 7(c))
These results show that mechanical injury results indistinct alterations in the kinetics andor amplitudes ofD-aspartate-induced intracellular sodium transients in reac-tive astrocytes depending on their distance to the lesion
4 Discussion
Our results show that within 6-7 days after setting a scratchwound in organotypic hippocampal slices a dense glial scarhad formed along the lesion site Up to approximately 100 120583mdistance from the lesion the scar tissue was characterized bya substantial elevation ofGFAP expression a pronounced cel-lular hypertrophy and reduced uptake of SR101 by astrocytes
10 ISRN Neurology
Con
trol
GLAST
Lesio
ned
GFAP Merge
StrR
StrP
StrO
Lesio
n sit
e
StrR
(a)
Con
trol
Lesio
ned
GLT-1 GFAP Merge
StrR
StrP
StrO
StrR
StrP
StrLM
Lesio
n sit
e
(b)
Figure 5 GLAST andGLT-1 immunoreactivity aftermechanical injury (a) Fluorescence images of immunohistochemical stainings detectingthe glutamate transporters GLAST (left) GFAP (center) as counterstaining and the corresponding merged image (GFAP green GLASTmagenta) in control (top row) and lesioned slice cultures (bottom row) (b) GLT-1 (left) GFAP (center) double labeling of a control (toprow) and a mechanically lesioned slice (bottom row) The corresponding merge (GFAP green GLT-1 magenta) is found on the right Notethat after lesion glutamate transporter immunoreactive clusters are accumulated in the direct vicinity of the lesion site (arrowheads) GFAPglial fibrillary acidic protein GLAST glutamate aspartate transporter GLT-1 glutamate-transporter-1 StrO stratum oriens StrP stratumpyramidale StrR stratum radiatum StrLM stratum lacunosum-moleculare and scale bars (a) 20 120583m (b) 40 120583m
While prominent clusters of GLAST and GLT-1 immunore-activity were observed alongGFAP-positive structures in scarcells D-aspartate-induced intracellular sodium signals werestrongly dampened indicating a significant reduction in glu-tamate uptake capacity close to the lesion In the periphery ofthe lesion (100ndash350120583m) astrocytes showed less pronounced
reactivity maintained their ability to efficiently take up SR101and displayed only minor changes in glutamate transporterimmunoreactivity and function Thus depending on theirdistance from the lesion astrocytes showed different gradesof reactivity and displayed discrete changes in glutamatetransporter immunoreactivity and uptake capacity
ISRN Neurology 11
SR101 MergeSBFI
StrRStrP
(a)
03
04
05
06
07
0 5 10 15
0
20 25 30 40 0
100
20
40
80
60
0 10 20 30 400
20
40
60
80
100
10 = 55 mM
107 cells in 6 experiments
F (340
380
) (ra
tio)
Intracellular Na+ concentration (mM)Intracellular Na+ concentration (mM)
ΔF
(340
380
) (
)
ΔF
(340
380
) (
) R2 = 097
(b)
D-aspartate
100 120583M500 120583M
750 120583M1000 120583M
1 mM
10 s
Δ
(c)
0 1 2 3 4 50
1
2
3
4
5
Δ so
dium
(mM
)
D-aspartate (mM)
lowast
(d)
Figure 6 Sodium changes induced by activation of glutamate uptake (a) Staining of astrocytes by colabeling with SR101 and SBFI Leftepifluorescence image of theCA1-area of an SR101-loaded organotypic sliceCenter SBFI fluorescence taken from the same areaRight overlaySBFI-loaded SR101-positive astrocytes appear in white (arrowheads) StrP stratum pyramidale StrR stratum radiatum scale bar 20 120583m (b)Left calibration of SBFI fluorescence in an astrocyte Depicted is the change in intracellular SBFI fluorescence ratio in the calibration solutionscontaining ionophores and in response to stepwise changes in the sodium concentration Right mean values plusmn SEM of the normalizedchanges in SBFI fluorescence at different sodium concentrations The black line represents a linear fit of the data points between 10 and40mM sodium illustrating that a 10 change in SBFI fluorescence represents a change in the sodium concentration by 55mM (c) Sodiumtransients in a single astrocyte induced by 500ms applications of D-aspartate at different concentrations (d) Mean values plusmn S E M (119899 = 19)of the peak amplitude of sodium transients at different concentrations of D-aspartate The curve represents a sigmoidal fit of the data
41 Characteristics of Lesion-Induced Reactive Gliosis inOrganotypic Cultures Organotypic tissue slice cultures ofthe hippocampus represent a well-established model systemin which the basic tissue architecture is maintained butwhich still hosts the major advantages of cell cultures suchas good accessibility and control of experimental conditionsThey have been used extensively for the analysis of neuronal
properties and developing neuronal networks [28 51 52]or the study of excitotoxic neuronal damage (eg [53 54])Recent studies also demonstrated that basic morphologicalcharacteristics of astrocytes and the typical glia-synapseorganization are well preserved in organotypic slice cultures(eg [44 45 55 56]) In addition we found a nearly completeoverlap of SR101 staining with the expression of GFP in slice
12 ISRN Neurology
SBFIControl Lesioned
SR101
1
Lesio
n sit
e
SR101 SBFI
2 45
32 4
5
1
StrR
StrP StrP
StrR
3
(a)
ControlScar
Lesioned
1
D-asp
2
D-asp
D-asp
5
4
3Periphery1mM
20 s
Δ Δ
Δ
Δ
Δ
(b)
0102030405060708090
100110
Δ so
dium
tim
e (m
Ms
)
00
02
04
06
08
10
12
0
1
2
3
4
5
6
ns
Control Periphery Scar Control Periphery Scar Control Periphery Scar
Amplitude Slope Decay time
Δ so
dium
(mM
)
120591(s
)lowastlowastlowast
lowastlowastlowastlowastlowastlowast
lowastlowastlowastlowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowast
(c)
Figure 7 D-aspartate-induced astrocyte sodium transients following lesion (a) CA1 area of a control (left) and a lesioned (right) slicepreparation Shown are epifluorescence images of SR101 and SBFI fluorescences Circled areas and numbers indicate the regions of interestanalyzed in the experiment depicted in (b) StrP stratum pyramidale StrR stratum radiatum and scale bar 40 120583m (b) Sodium transientsinduced by pressure application of 1mMD-aspartate (D-asp) for 500ms in the different cells in the control and lesioned slice as indicated in(a) (c) Histograms showing mean values plusmn S E M of the peak amplitude (left) slope (center) and decay time (right) of D-aspartate-inducedsodium transients in astrocytes in control slices (119899 = 161) as well as in cells in the scar region and its periphery (119899 = 111 and 119899 = 26 resp)lowastlowast119875 lt 001 lowastlowastlowast119875 lt 0001 StrP stratum pyramidale StrR stratum radiatum and scale bar 40120583m
cultures obtained from GFPGFAP mice demonstrating thatSR101 is well suited to identify astrocytes in this prepara-tion as described before for acute tissue slices and in vivo[29 30]
Performing a scratch through the CA1 region of organ-otypic slices resulted in the formation of a glial scar along thelesion site as judged by the prominent increase in expressionof GFAP and S100120573 In addition astrocyte cell bodies showed
a significant hypertrophy Furthermore astrocytes in theperiphery of the scar (100ndash350 120583m from the lesion) showedmild to moderate reactivity displaying swollen cell bodiesand only moderate upregulation of GFAP These changesare hallmarks of reactive gliosis observed after tissue injury[10 13 57] Generally the degree of injury and the distance ofthe astrocytes from the site of injury define the degree of theiractivation [11 13]
ISRN Neurology 13
Our study also revealed that strongly reactive astrocytesin the scar region only weakly stain with SR101 The reducedability of strongly activated astrocytes to accumulate SR101indicates that strong glial activation is accompanied by adownregulation of the organic anion transporter responsi-ble for uptake of SR101 This is reminiscent of immatureastrocytes in the early postnatal hippocampus which areSR101-negative [30] and in line with a wealth of experimentalevidence suggesting that reactive gliosis represents a processcomprising a dedifferentiation of astrocytes [17]
42 Changes in Glutamate Transporter Expression followingLesion Glutamate uptake in the hippocampus is mainlyachieved by the glial glutamate transporters GLT-1 andGLAST [1 2] A central factor in many brain pathologiesincluding traumatic brain injury is an increase in extracellu-lar glutamate and excitotoxicity it has been suggested that achange in the expression levels of glial glutamate transportersmight play a critical role in the failure of glutamate clearance[6 58ndash60] Earlier work has found an overall downregulationof protein levels of both GLAST and GLT-1 following astro-cyte activation [22ndash27] indicating that this is causal to theelevation of extracellular glutamate In contrast to this notionother studies reported increased glutamate transport capacityof reactive astrocytes and suggested a protective influence[61 62]
To visualize possible differences in the spatial expressionprofile of GLT-1 and GLAST in strongly reactive astrocytesalong the scar as compared to moderately reactive astro-cytes in its periphery we performed immunohistochemicalstainings These revealed an accumulation of GLT-1 andGLAST immunoreactivity along thick GFAP-positive fiberswhich was particularly pronounced for GLAST Besides theseclusters overall GLAST immunoreactivity seemed unalteredas compared to unlesioned slices whereas GLT-1 immunore-activity seemedweaker and declined with increasing distancefrom the scar Clustering of glutamate transporters has beendescribed by several reports [48 63 64] In developinghippocampal astrocytes cluster formation was preferentiallyfound in branches opposed to synapses and was increasedwith increased neuronal activity indicating that it is neces-sary to copewith synaptic release of glutamate [36] Along thesame lines it was observed that sustained astroglial activationby ciliary neurotrophic factor (CNTF) in the rat striatuminduced a concentration of GLAST and GLT-1 into raftmicrodomains and improved glutamate clearance indicatingthat cluster formation increased the cellular capacity for glu-tamate uptake [61] However another study found glutamate-induced clustering of GLT-1 that induced its endocytosis andintracellular trafficking without changing the total expressionlevels as detected by western blots arguing for a decrease infunctional glial glutamate uptake capacity [49] In the presentstudy antibodies against GLT-1 and GLAST were employedfor immunohistochemistry after permeabilization of theplasma membrane The clusters of GLAST and GLT-1 alongGFAP-positive fibers in the scar region (as GLAST andGLT-1immunoreactivity in general) could thus represent glutamatetransporters in the plasmamembrane or in intracellular com-partments or both Consequently the observed clustering
does not allow a prediction about possible functional changesin glutamate uptake
43 Lesion-Induced Changes in Functional Glutamate UptakeCapacity We probed for the functional activation of gluta-mate transport by application of the transportable agonistD-aspartate which results in an increase in the intracellularsodium concentration of astrocytes [8 9] The amplitudesof D-aspartate-induced sodium transients in astrocytes incontrol organotypic slices were similar to those reportedfrom astrocytes in acute slices obtained in our lab [33]indicating similar cellular glutamate transport capacity inboth preparations
Following a lesion D-aspartate-induced intracellularsodium signalswere clearly dampened in scar cellsMoreoverthe slope of the increase in sodium was significantly reducedand the decay back to baseline was slowed Because D-aspartate was applied at nearly saturating concentrationthe reduction in peak amplitudes suggests a reduction infunctional glutamate uptake capacity in cells close to thelesion which might be mediated by a decrease in the overallnumber of glutamate transporters available at the plasmamembrane In contrast the alteration in the kinetics of theD-aspartate-induced sodium signals can be explained by thecellular hypertrophy that was observed in reactive astrocytesAt an equal transport-mediated sodium influx across themembrane the slope of resulting changes in the sodiumconcentration will be decreased in cells with a larger volume
Furthermore the decay of intracellular sodium transientsback to baseline is mainly governed by the activity of theNa+K+-ATPase as well as by diffusion [8] The slower recov-ery is in line with earlier studies reporting a downregulationof the sodium pump following reactive gliosis [65 66]Moreover sodium is not buffered in the cell and travels solelyby means of diffusion in the cytoplasm Hindered diffusionresulting from increased cytoplasmic protein content orimpaired gap junction coupling which both occur uponastrocytic activation might thus also partially contribute todelayed sodium recovery [12 34 67]
In contrast to scar cells the amplitude of D-aspartate-induced intracellular sodium signals was not significantlyaltered in moderately activated astrocytes in the periphery ofthe lesion indicating that the overall number of functionalglutamate transporters was not significantly altered Becausethese cells showed hypertrophy as well the slowed kinetics ofthe sodium transients are in line with an increased cellularvolume as well as a possible downregulation of Na+K+-ATPase andor a slowed diffusion as argued above
Taken together our results indicate a significant reduc-tion in glutamate uptake capacity in strongly activated astro-cytes in the scar region Thus the prominent clustering ofGLAST and GLT-1 immunoreactivity along GFAP-positivestructures close to the lesion is likely to reflect a loss ofglutamate transporters from the plasma membrane upontransporter internalization as reported by Nakagawa et al[49] The authors of the latter study speculated that large andprolonged increases in glutamate concentrations are neces-sary to induce such cluster formation and endocytosis in thetissue as they would possibly only occur under pathological
14 ISRN Neurology
conditions Indeed ambient glutamate concentrations wereshown to be significantly increased after traumatic braininjury [68ndash70] Along these lines it can be assumed thatin our model system extracellular glutamate concentrationsrose less severely in the periphery of the lesion preventinga comparable clustering and loss of functional glutamatetransport activity in moderately activated astrocytes
5 Conclusions
Our results show complex changes in glutamate transporterexpression and function during astrocyte activation followingmechanical injury They confirm that the degree of astrocyteactivation depends on the distance from the insult and com-prises different endpoints regarding cellular morphology andphysiology Furthermore our results highlight that astrocyteswhich show different grades of reactivity also display discretechanges in glutamate transporter expression and functionWhile immunohistochemistry revealed a prominent clus-tering of GLT-1 and GLAST immunoreactivity in the scarregion our functional assay clearly showed that glutamateuptake capacity is strongly reduced in scar cells while itis largely maintained in moderately activated astrocytes inthe periphery Thus mild-to-moderate astrogliosis in theperiphery of a mechanical lesion does not necessarily seemto be accompanied by a significant change in glial glutamateuptake capacity At the glial scar itself in strongly reactiveastrocytes a clustering of glutamate transporters is observedthat apparently goes along with a severe functional reductionin astroglial glutamate uptake which may contribute toglutamate-mediated excitotoxicity in this region
Conflict of Interests
The authors of the paper declare that they do not haveany direct financial relation with the commercial identitiesmentioned in the paper that might lead to a conflict ofinterests
Acknowledgments
This study was supported by the Deutsche Forschungsge-meinschaft (DFG Ro23274-3) The authors thank SimoneDurry and Claudia Roderigo for expert technical help
References
[1] N C Danbolt ldquoGlutamate uptakerdquo Progress in Neurobiologyvol 65 no 1 pp 1ndash105 2001
[2] A V Tzingounis and J I Wadiche ldquoGlutamate transportersconfining runaway excitation by shaping synaptic transmis-sionrdquo Nature Reviews Neuroscience vol 8 no 12 pp 935ndash9472007
[3] D E Bergles J S Diamond and C E Jahr ldquoClearance ofglutamate inside the synapse and beyondrdquo Current Opinion inNeurobiology vol 9 no 3 pp 293ndash298 1999
[4] C M Anderson and R A Swanson ldquoAstrocyte glutamatetransport review of properties regulation and physiologicalfunctionsrdquo Glia vol 32 no 1 pp 1ndash14 2000
[5] P Marcaggi and D Attwell ldquoRole of glial amino acid trans-porters in synaptic transmission and brain energeticsrdquoGlia vol47 no 3 pp 217ndash225 2004
[6] N J Maragakis and J D Rothstein ldquoGlutamate transportersanimal models to neurologic diseaserdquo Neurobiology of Diseasevol 15 no 3 pp 461ndash473 2004
[7] G Gegelashvili and A Schousboe ldquoCellular distribution andkinetic properties of high-affinity glutamate transportersrdquoBrainResearch Bulletin vol 45 no 3 pp 233ndash238 1998
[8] J W Deitmer and C R Rose ldquoIon changes and signalling inperisynaptic gliardquo Brain Research Reviews vol 63 no 1-2 pp113ndash129 2010
[9] S Kirischuk V Parpura and A Verkhratsky ldquoSodium dynam-ics another key to astroglial excitabilityrdquo Trends in Neuro-sciences vol 35 no 8 pp 497ndash506 2012
[10] M V Sofroniew ldquoMolecular dissection of reactive astrogliosisand glial scar formationrdquo Trends in Neurosciences vol 32 no12 pp 638ndash647 2009
[11] J L Ridet S K Malhotra A Privat and F H Gage ldquoReactiveastrocytes cellular and molecular cues to biological functionrdquoTrends in Neurosciences vol 20 no 12 pp 570ndash577 1997
[12] J Middeldorp and E M Hol ldquoGFAP in health and diseaserdquoProgress in Neurobiology vol 93 no 3 pp 421ndash443 2011
[13] M Pekny and M Nilsson ldquoAstrocyte activation and reactivegliosisrdquo Glia vol 50 no 4 pp 427ndash434 2005
[14] A Bordey S A Lyons J J Hablitz and H SontheimerldquoElectrophysiological characteristics of reactive astrocytes inexperimental cortical dysplasiardquo Journal of Neurophysiologyvol 85 no 4 pp 1719ndash1731 2001
[15] R Jabs G Seifert and C Steinhauser ldquoAstrocytic functionand its alteration in the epileptic brainrdquo Epilepsia vol 49supplement 2 pp 3ndash12 2008
[16] A Buffo I Rite P Tripathi et al ldquoOrigin and progeny ofreactive gliosis a source of multipotent cells in the injuredbrainrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 105 no 9 pp 3581ndash3586 2008
[17] S Robel B Berninger and M Gotz ldquoThe stem cell potential ofglia lessons from reactive gliosisrdquoNature Reviews Neurosciencevol 12 no 2 pp 88ndash104 2011
[18] H Kawano J Kimura-Kuroda Y Komuta et al ldquoRole of thelesion scar in the response to damage and repair of the centralnervous systemrdquo Cell and Tissue Research vol 349 no 1 pp169ndash180 2012
[19] G Yiu and Z He ldquoGlial inhibition of CNS axon regenerationrdquoNature Reviews Neuroscience vol 7 no 8 pp 617ndash627 2006
[20] M T Fitch and J Silver ldquoCNS injury glial scars and inflamma-tion inhibitory extracellular matrices and regeneration failurerdquoExperimental Neurology vol 209 no 2 pp 294ndash301 2008
[21] L Li A Lundkvist D Andersson et al ldquoProtective role ofreactive astrocytes in brain ischemiardquo Journal of Cerebral BloodFlow amp Metabolism vol 28 no 3 pp 468ndash481 2008
[22] F K H van Landeghem T Weiss M Oehmichen and AVonDeimling ldquoDecreased expression of glutamate transportersin astrocytes after human traumatic brain injuryrdquo Journal ofNeurotrauma vol 23 no 10 pp 1518ndash1528 2006
[23] V L Raghavendra Rao M K Baskaya A Dogan J DRothstein and R J Dempsey ldquoTraumatic brain injury down-regulates glial glutamate transporter (GLT- 1 and GLAST)proteins in rat brainrdquo Journal of Neurochemistry vol 70 no 5pp 2020ndash2027 1998
ISRN Neurology 15
[24] J-C Chen H Hsu-Chou J-L Lu et al ldquoDown-regulation ofthe glial glutamate transporter GLT-1 in rat hippocampus andstriatum and its modulation by a group III metabotropic glu-tamate receptor antagonist following transient global forebrainischemiardquoNeuropharmacology vol 49 no 5 pp 703ndash714 2005
[25] S D Ginsberg L J Martin and J D Rothstein ldquoRegional deaf-ferentation down-regulates subtypes of glutamate transporterproteinsrdquo Journal of Neurochemistry vol 65 no 6 pp 2800ndash2803 1995
[26] J E Springer R D Azbill R J Mark J G Begley GWaeg andM P Mattson ldquo4-Hydroxynonenal a lipid peroxidation prod-uct rapidly accumulates following traumatic spinal cord injuryand inhibits glutamate uptakerdquo Journal of Neurochemistry vol68 no 6 pp 2469ndash2476 1997
[27] M B Moretto N S Arteni D Lavinsky et al ldquoHypoxic-ischemic insult decreases glutamate uptake by hippocampalslices from neonatal rats Prevention by guanosinerdquo Experimen-tal Neurology vol 195 no 2 pp 400ndash406 2005
[28] L Stoppini P-A Buchs and D Muller ldquoA simple method fororganotypic cultures of nervous tissuerdquo Journal of NeuroscienceMethods vol 37 no 2 pp 173ndash182 1991
[29] A Nimmerjahn F Kirchhoff J N D Kerr and F HelmchenldquoSulforhodamine 101 as a specific marker of astroglia in theneocortex in vivordquo Nat Methods vol 1 no 1 pp 31ndash37 2004
[30] K W Kafitz S D Meier J Stephan and C R Rose ldquoDevel-opmental profile and properties of sulforhodamine 101-Labeledglial cells in acute brain slices of rat hippocampusrdquo Journal ofNeuroscience Methods vol 169 no 1 pp 84ndash92 2008
[31] C Schnell Y Hagos and S Hulsmann ldquoActive sulforhodamine101 uptake into hippocampal astrocytesrdquo PLoS One vol 7 no 11Article ID e49398 2012
[32] S D Meier Y Kovalchuk and C R Rose ldquoProperties ofthe new fluorescent Na+ indicator CoroNa Green comparisonwith SBFI and confocal Na+ imagingrdquo Journal of NeuroscienceMethods vol 155 no 2 pp 251ndash259 2006
[33] J Langer and C R Rose ldquoSynaptically induced sodium signalsin hippocampal astrocytes in siturdquo Journal of Physiology vol587 part 24 pp 5859ndash5877 2009
[34] J Langer J Stephan M Theis and C R Rose ldquoGap junctionsmediate intercellular spread of sodium between hippocampalastrocytes in siturdquo Glia vol 60 no 2 pp 239ndash252 2012
[35] C R Rose andB R Ransom ldquoIntracellular sodiumhomeostasisin rat hippocampal astrocytesrdquo Journal of Physiology vol 491part 2 no 2 pp 291ndash305 1996
[36] A M Benediktsson G S Marrs J C Tu et al ldquoNeuronalactivity regulates glutamate transporter dynamics in developingastrocytesrdquo Glia vol 60 no 2 pp 175ndash188 2012
[37] M Zhou G P Schools and H K Kimelberg ldquoDevelopment ofGLAST(+) astrocytes andNG2(+) glia in rat hippocampus CA1mature astrocytes are electrophysiologically passiverdquo Journal ofNeurophysiology vol 95 no 1 pp 134ndash143 2006
[38] I Suarez G Bodega and B Fernandez ldquoModulation of glu-tamate transporters (GLAST GLT-1 and EAAC1) in the ratcerebellum following portocaval anastomosisrdquo Brain Researchvol 859 no 2 pp 293ndash302 2000
[39] X Zhu D E Bergles and A Nishiyama ldquoNG2 cells generateboth oligodendrocytes and gray matter astrocytesrdquo Develop-ment vol 135 no 1 pp 145ndash157 2008
[40] B Brunne S ZhaoADerouiche et al ldquoOriginmaturation andastroglial transformation of secondary radial glial cells in thedeveloping dentate gyrusrdquo Glia vol 58 no 13 pp 1553ndash15692010
[41] Y Wu A-Q Zhang and D T Yew ldquoAge related changes ofvarious markers of astrocytes in senescence-accelerated micehippocampusrdquo Neurochemistry International vol 46 no 7 pp565ndash574 2005
[42] S Magavi D Friedmann G Banks A Stolfi and C LoisldquoCoincident generation of pyramidal neurons and protoplasmicastrocytes in neocortical columnsrdquo Journal of Neuroscience vol32 no 14 pp 4762ndash4772 2012
[43] M Olabarria H N Noristani A Verkhratsky and J JRodrıguez ldquoConcomitant astroglial atrophy and astrogliosis ina triple transgenic animal model of Alzheimerrsquos diseaserdquo Gliavol 58 no 7 pp 831ndash838 2010
[44] A M Benediktsson S J Schachtele S H Green and M EDailey ldquoBallistic labeling and dynamic imaging of astrocytesin organotypic hippocampal slice culturesrdquo Journal of Neuro-science Methods vol 141 no 1 pp 41ndash53 2005
[45] I Lushnikova G Skibo D Muller and I Nikonenko ldquoSynapticpotentiation induces increased glial coverage of excitatorysynapses in CA1 hippocampusrdquoHippocampus vol 19 no 8 pp753ndash762 2009
[46] E Raponi F Agenes C Delphin et al ldquoS100B expressiondefines a state in which GFAP-expressing cells lose their neuralstem cell potential and acquire a more mature developmentalstagerdquo Glia vol 55 no 2 pp 165ndash177 2007
[47] C G Schipke C Boucsein C Ohlemeyer F Kirchhoff andH Kettenmann ldquoAstrocyte Ca2+ waves trigger responses inmicroglial cells in brain slicesrdquo The FASEB Journal vol 16 no2 pp 255ndash257 2002
[48] J Zhou and M L Sutherland ldquoGlutamate transporter clusterformation in astrocytic processes regulates glutamate uptakeactivityrdquo Journal of Neuroscience vol 24 no 28 pp 6301ndash63062004
[49] T Nakagawa Y Otsubo Y Yatani H Shirakawa and S KanekoldquoMechanisms of substrate transport-induced clustering of aglial glutamate transporter GLT-1 in astroglial-neuronal cul-turesrdquo European Journal of Neuroscience vol 28 no 9 pp 1719ndash1730 2008
[50] J-Y Chatton P Marquet and P J Magistretti ldquoA quantitativeanalysis of L-glutamate-regulated Na+ dynamics in mousecortical astrocytes Implications for cellular bioenergeticsrdquoEuropean Journal of Neuroscience vol 12 no 11 pp 3843ndash38532000
[51] E Forster S Zhao and M Frotscher ldquoLaminating the hip-pocampusrdquo Nature Reviews Neuroscience vol 7 no 4 pp 259ndash267 2006
[52] B H Gahwiler M Capogna D Debanne R A McKinney andS M Thompson ldquoOrganotypic slice cultures a technique hascome of agerdquo Trends in Neurosciences vol 20 no 10 pp 471ndash477 1997
[53] P E Kunkler and R P Kraig ldquoReactive astrocytosis fromexcitotoxic injury in hippocampal organ culture parallels thatseen in vivordquo Journal of Cerebral Blood Flow and Metabolismvol 17 no 1 pp 26ndash43 1997
[54] I E Holopainen ldquoOrganotypic hippocampal slice cultures amodel system to study basic cellular andmolecularmechanismsof neuronal cell death neuroprotection and synaptic plasticityrdquoNeurochemical Research vol 30 no 12 pp 1521ndash1528 2005
[55] MHaber L Zhou andK KMurai ldquoCooperative astrocyte anddendritic spine dynamics at hippocampal excitatory synapsesrdquoJournal of Neuroscience vol 26 no 35 pp 8881ndash8891 2006
16 ISRN Neurology
[56] H Nishida and S Okabe ldquoDirect astrocytic contacts regulatelocal maturation of dendritic spinesrdquo Journal of Neurosciencevol 27 no 2 pp 331ndash340 2007
[57] A Buffo C Rolando and S Ceruti ldquoAstrocytes in the dam-aged brain molecular and cellular insights into their reactiveresponse and healing potentialrdquoBiochemical Pharmacology vol79 no 2 pp 77ndash89 2010
[58] J-H Yi and A S Hazell ldquoExcitotoxic mechanisms and the roleof astrocytic glutamate transporters in traumatic brain injuryrdquoNeurochemistry International vol 48 no 5 pp 394ndash403 2006
[59] C Werner and K Engelhard ldquoPathophysiology of traumaticbrain injuryrdquo British Journal of Anaesthesia vol 99 no 1 pp4ndash9 2007
[60] G Gegelashvili M B Robinson D Trotti and T RauenldquoRegulation of glutamate transporters in health and diseaserdquoProgress in Brain Research vol 132 pp 267ndash286 2001
[61] C Escartin E Brouillet PGubellini et al ldquoCiliary neurotrophicfactor activates astrocytes redistributes their glutamate trans-porters GLAST and GLT-1 to raft microdomains and improvesglutamate handling in vivordquo Journal of Neuroscience vol 26 no22 pp 5978ndash5989 2006
[62] C Vermeiren M Najimi N Vanhoutte et al ldquoAcute up-regulation of glutamate uptake mediated by mGluR5a in reac-tive astrocytesrdquo Journal of Neurochemistry vol 94 no 2 pp405ndash416 2005
[63] C L Poitry-Yamate L Vutskits and T Rauen ldquoNeuronal-induced and glutamate-dependent activation of glial glutamatetransporter functionrdquo Journal of Neurochemistry vol 82 no 4pp 987ndash997 2002
[64] P M Beart and R D OrsquoShea ldquoTransporters for L-glutamatean update on their molecular pharmacology and pathologicalinvolvementrdquo British Journal of Pharmacology vol 150 no 1pp 5ndash17 2007
[65] F D Lima M A Souza A F Furian et al ldquoNa+K+-ATPaseactivity impairment after experimental traumatic brain injuryrelationship to spatial learning deficits and oxidative stressrdquoBehavioural Brain Research vol 193 no 2 pp 306ndash310 2008
[66] G A Gusarova H E Trejo L A Dada et al ldquoHypoxia leadsto NaK-ATPase downregulation via Ca2+ release-activatedCa2+ channels and AMPK activationrdquo Molecular and CellularBiology vol 31 no 17 pp 3546ndash3556 2011
[67] S S L Chew C S Johnson C R Green and H V Danesh-Meyer ldquoRole of connexin43 in central nervous system injuryrdquoExperimental Neurology vol 225 no 2 pp 250ndash261 2010
[68] P Nilsson L Hillered U Ponten and U Ungerstedt ldquoChangesin cortical extracellular levels of energy-related metabolites andamino acids following concussive brain injury in ratsrdquo Journal ofCerebral Blood Flow and Metabolism vol 10 no 5 pp 631ndash6371990
[69] H Katoh K Sima H Nawashiro K Wada and H ChigasakildquoThe effect of MK-801 on extracellular neuroactive amino acidsin hippocampus after closed head injury followed by hypoxia inratsrdquo Brain Research vol 758 no 1-2 pp 153ndash162 1997
[70] H Koizumi H Fujisawa H Ito T Maekawa X Di and RBullock ldquoEffects of mild hypothermia on cerebral blood flow-independent changes in cortical extracellular levels of aminoacids following contusion trauma in the ratrdquoBrain Research vol747 no 2 pp 304ndash312 1997
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

ISRN Neurology 9
characterized by a robust increase in GFAP and S100120573expressionThese proximal reactive astrocytes locatedwithina distance of less than 100120583m from the lesion exhibit longGFAP-positive processes have significantly swollen somataand only weakly stain with SR101 Astrocytes in the peripheryof the scar (100ndash350 120583m from the lesion) display swollen cellbodies as well but show onlymoderate upregulation of GFAPand maintain their ability to efficiently take up SR101
33 Expression Levels and Spatial Distribution of GLT-1 andGLAST To determine the spatial distribution of the gluta-mate transporters GLAST and GLT-1 in organotypic slicesand changes therein in response to the lesion immunohisto-chemistry was employed In control slices GLAST (119873 = 13Figure 5(a)) as well as GLT-1 immunoreactivity (119873 = 11Figure 5(b)) appeared as punctate labeling throughout theentire CA1 area The staining patterns for both transportersin the tissue slice cultures were thus similar to those reportedfrom acute tissue slice preparations of the hippocampus [3648 49]
The relatively homogeneous distribution of immunore-activity throughout the preparation found in control slicecultures was abandoned following a lesion Here distinctaccumulations of GLAST (119873 = 17 Figure 5(a)) and GLT-1immunoreactivity (119873 = 12 Figure 5(b)) were found in thescar region In both cases these preferentially colocalizedwith thick GFAP-positive bundles running in parallel to thelesion (Figure 5) Apart from the marked accumulation ofGLAST reactivity in parallel to the scar GLAST immunore-activity appeared generally increased in some lesioned slicepreparations (Figure 5(a)) but unaltered in others bothwithinthe scar as well as in its periphery For GLT-1 a decline in theintensity of immunoreactivity was observed with increasingdistance from the lesion (Figure 5(b))
In summary these results reveal distinct alterations inthe immunoreactivity of GLAST and GLT-1 following amechanical lesion In the scar region clusters of GLT-1 andGLAST immunoreactivity are present primarily colocalizedwith strongly GFAP-positive fibers Apart from these distinctclusters GLT-1 immunoreactivity appears to decline withdistance from the scar whereas GLAST immunoreactivityseems largely uniform throughout the preparation
34 Sodium Transients Induced by Activation of GlutamateUptake Glutamate uptake is accompanied by the inwardflux of sodium resulting in increase in the intracellularsodium concentration of astrocytes To monitor its functionwe assayed sodium increases induced by D-aspartate anonmetabolized substrate of sodium-dependent glutamateuptake which is also transported [1] in astrocytes in thestratum radiatum of organotypic slices To this end SR101-stained slices were additionally loaded with the sodium-sensitive fluorophore SBFI-AM (Figure 6(a) cf Figure 3(b))In situ calibrations of the SBFI fluorescence revealed a linearincrease in its fluorescence ratio at intracellular sodiumconcentrations between 10 and 40mM (119899 = 107 Figure 6(b))as reported before [33]
Repetitive pressure application of D-aspartate for 500msinduced reliable increases in the intracellular sodium con-centration in astrocytes indicating activation of sodium-dependent glutamate uptake (119899 = 488 Figure 6(c)) Theexperiments were performed in the presence of the sodium-channel blocker TTX (tetrodotoxin 05 120583M) to suppressaction potential generation The amplitude of D-aspartate-induced sodium increases was dependent on the concen-tration applied (Figure 6(c)) and saturated at about 2mM(119899 = 19 Figure 6(d)) as reported before for mouse corticalastrocytes in culture [50] At a concentration of 1mM theamplitude of D-aspartate-induced sodium transients wasnear maximum and amounted to 38 plusmn 02mM (119899 =161 Figures 6(c) and 6(d)) a value comparable to that inastrocytes in acute slices obtained in our lab [33]
To probe for changes in glutamate uptake we comparedsodium signals induced by 1mMD-aspartate between controland lesioned slices (Figure 7) In lesioned slices astrocyteslocated in the periphery of the lesion (100ndash350 120583m distance)responded with a sodium transient to application of 1mMD-aspartate (see cell 2 in Figures 7(a) and 7(b)) The amplitudeof D-aspartate-induced sodium signals in cells in the lesionperiphery was similar to that observed in control slices (38 plusmn03mM 119899 = 111 see cell 1 in Figures 7(a) 7(b) and 7(c))Application of D-aspartate also evoked sodium transientsin weakly SR101-positive SBFI-filled astrocytes located inthe glial scar area (lt100 120583m from the lesion see cells 3ndash5in Figures 7(a) and 7(b)) These cells however displayedsignificantly smaller peak amplitudes (16 plusmn 021mM 119899 =26 119875 lt 0001 Figures 7(b) and 7(c)) than astrocytes inthe periphery and in control slices
The kinetics of D-aspartate-induced sodium transientsdiffered between all three groups This was true for theslope of the sodium increase (10ndash90 change) which wassignificantly reduced in astrocytes from lesioned slices versusastrocytes from control slices (09 plusmn 005mMs in controlversus 06 plusmn 006mMs in the periphery and 02 plusmn 004mMsin the scar region 119899 = 161 111 and 26 resp 119875 lt 0001Figures 7(b) and 7(c)) In addition the decay back to baselinewas significantly slower following lesion (Figures 7(b) and7(c)) The time needed for the signal to decay by 67 wasapproximately doubled (decay time 120591 was 44 plusmn 2 s in control120591 = 81 plusmn 4 s in periphery of the lesion and 120591 = 95 plusmn 10 s inscar cells Figures 7(b) and 7(c))
These results show that mechanical injury results indistinct alterations in the kinetics andor amplitudes ofD-aspartate-induced intracellular sodium transients in reac-tive astrocytes depending on their distance to the lesion
4 Discussion
Our results show that within 6-7 days after setting a scratchwound in organotypic hippocampal slices a dense glial scarhad formed along the lesion site Up to approximately 100 120583mdistance from the lesion the scar tissue was characterized bya substantial elevation ofGFAP expression a pronounced cel-lular hypertrophy and reduced uptake of SR101 by astrocytes
10 ISRN Neurology
Con
trol
GLAST
Lesio
ned
GFAP Merge
StrR
StrP
StrO
Lesio
n sit
e
StrR
(a)
Con
trol
Lesio
ned
GLT-1 GFAP Merge
StrR
StrP
StrO
StrR
StrP
StrLM
Lesio
n sit
e
(b)
Figure 5 GLAST andGLT-1 immunoreactivity aftermechanical injury (a) Fluorescence images of immunohistochemical stainings detectingthe glutamate transporters GLAST (left) GFAP (center) as counterstaining and the corresponding merged image (GFAP green GLASTmagenta) in control (top row) and lesioned slice cultures (bottom row) (b) GLT-1 (left) GFAP (center) double labeling of a control (toprow) and a mechanically lesioned slice (bottom row) The corresponding merge (GFAP green GLT-1 magenta) is found on the right Notethat after lesion glutamate transporter immunoreactive clusters are accumulated in the direct vicinity of the lesion site (arrowheads) GFAPglial fibrillary acidic protein GLAST glutamate aspartate transporter GLT-1 glutamate-transporter-1 StrO stratum oriens StrP stratumpyramidale StrR stratum radiatum StrLM stratum lacunosum-moleculare and scale bars (a) 20 120583m (b) 40 120583m
While prominent clusters of GLAST and GLT-1 immunore-activity were observed alongGFAP-positive structures in scarcells D-aspartate-induced intracellular sodium signals werestrongly dampened indicating a significant reduction in glu-tamate uptake capacity close to the lesion In the periphery ofthe lesion (100ndash350120583m) astrocytes showed less pronounced
reactivity maintained their ability to efficiently take up SR101and displayed only minor changes in glutamate transporterimmunoreactivity and function Thus depending on theirdistance from the lesion astrocytes showed different gradesof reactivity and displayed discrete changes in glutamatetransporter immunoreactivity and uptake capacity
ISRN Neurology 11
SR101 MergeSBFI
StrRStrP
(a)
03
04
05
06
07
0 5 10 15
0
20 25 30 40 0
100
20
40
80
60
0 10 20 30 400
20
40
60
80
100
10 = 55 mM
107 cells in 6 experiments
F (340
380
) (ra
tio)
Intracellular Na+ concentration (mM)Intracellular Na+ concentration (mM)
ΔF
(340
380
) (
)
ΔF
(340
380
) (
) R2 = 097
(b)
D-aspartate
100 120583M500 120583M
750 120583M1000 120583M
1 mM
10 s
Δ
(c)
0 1 2 3 4 50
1
2
3
4
5
Δ so
dium
(mM
)
D-aspartate (mM)
lowast
(d)
Figure 6 Sodium changes induced by activation of glutamate uptake (a) Staining of astrocytes by colabeling with SR101 and SBFI Leftepifluorescence image of theCA1-area of an SR101-loaded organotypic sliceCenter SBFI fluorescence taken from the same areaRight overlaySBFI-loaded SR101-positive astrocytes appear in white (arrowheads) StrP stratum pyramidale StrR stratum radiatum scale bar 20 120583m (b)Left calibration of SBFI fluorescence in an astrocyte Depicted is the change in intracellular SBFI fluorescence ratio in the calibration solutionscontaining ionophores and in response to stepwise changes in the sodium concentration Right mean values plusmn SEM of the normalizedchanges in SBFI fluorescence at different sodium concentrations The black line represents a linear fit of the data points between 10 and40mM sodium illustrating that a 10 change in SBFI fluorescence represents a change in the sodium concentration by 55mM (c) Sodiumtransients in a single astrocyte induced by 500ms applications of D-aspartate at different concentrations (d) Mean values plusmn S E M (119899 = 19)of the peak amplitude of sodium transients at different concentrations of D-aspartate The curve represents a sigmoidal fit of the data
41 Characteristics of Lesion-Induced Reactive Gliosis inOrganotypic Cultures Organotypic tissue slice cultures ofthe hippocampus represent a well-established model systemin which the basic tissue architecture is maintained butwhich still hosts the major advantages of cell cultures suchas good accessibility and control of experimental conditionsThey have been used extensively for the analysis of neuronal
properties and developing neuronal networks [28 51 52]or the study of excitotoxic neuronal damage (eg [53 54])Recent studies also demonstrated that basic morphologicalcharacteristics of astrocytes and the typical glia-synapseorganization are well preserved in organotypic slice cultures(eg [44 45 55 56]) In addition we found a nearly completeoverlap of SR101 staining with the expression of GFP in slice
12 ISRN Neurology
SBFIControl Lesioned
SR101
1
Lesio
n sit
e
SR101 SBFI
2 45
32 4
5
1
StrR
StrP StrP
StrR
3
(a)
ControlScar
Lesioned
1
D-asp
2
D-asp
D-asp
5
4
3Periphery1mM
20 s
Δ Δ
Δ
Δ
Δ
(b)
0102030405060708090
100110
Δ so
dium
tim
e (m
Ms
)
00
02
04
06
08
10
12
0
1
2
3
4
5
6
ns
Control Periphery Scar Control Periphery Scar Control Periphery Scar
Amplitude Slope Decay time
Δ so
dium
(mM
)
120591(s
)lowastlowastlowast
lowastlowastlowastlowastlowastlowast
lowastlowastlowastlowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowast
(c)
Figure 7 D-aspartate-induced astrocyte sodium transients following lesion (a) CA1 area of a control (left) and a lesioned (right) slicepreparation Shown are epifluorescence images of SR101 and SBFI fluorescences Circled areas and numbers indicate the regions of interestanalyzed in the experiment depicted in (b) StrP stratum pyramidale StrR stratum radiatum and scale bar 40 120583m (b) Sodium transientsinduced by pressure application of 1mMD-aspartate (D-asp) for 500ms in the different cells in the control and lesioned slice as indicated in(a) (c) Histograms showing mean values plusmn S E M of the peak amplitude (left) slope (center) and decay time (right) of D-aspartate-inducedsodium transients in astrocytes in control slices (119899 = 161) as well as in cells in the scar region and its periphery (119899 = 111 and 119899 = 26 resp)lowastlowast119875 lt 001 lowastlowastlowast119875 lt 0001 StrP stratum pyramidale StrR stratum radiatum and scale bar 40120583m
cultures obtained from GFPGFAP mice demonstrating thatSR101 is well suited to identify astrocytes in this prepara-tion as described before for acute tissue slices and in vivo[29 30]
Performing a scratch through the CA1 region of organ-otypic slices resulted in the formation of a glial scar along thelesion site as judged by the prominent increase in expressionof GFAP and S100120573 In addition astrocyte cell bodies showed
a significant hypertrophy Furthermore astrocytes in theperiphery of the scar (100ndash350 120583m from the lesion) showedmild to moderate reactivity displaying swollen cell bodiesand only moderate upregulation of GFAP These changesare hallmarks of reactive gliosis observed after tissue injury[10 13 57] Generally the degree of injury and the distance ofthe astrocytes from the site of injury define the degree of theiractivation [11 13]
ISRN Neurology 13
Our study also revealed that strongly reactive astrocytesin the scar region only weakly stain with SR101 The reducedability of strongly activated astrocytes to accumulate SR101indicates that strong glial activation is accompanied by adownregulation of the organic anion transporter responsi-ble for uptake of SR101 This is reminiscent of immatureastrocytes in the early postnatal hippocampus which areSR101-negative [30] and in line with a wealth of experimentalevidence suggesting that reactive gliosis represents a processcomprising a dedifferentiation of astrocytes [17]
42 Changes in Glutamate Transporter Expression followingLesion Glutamate uptake in the hippocampus is mainlyachieved by the glial glutamate transporters GLT-1 andGLAST [1 2] A central factor in many brain pathologiesincluding traumatic brain injury is an increase in extracellu-lar glutamate and excitotoxicity it has been suggested that achange in the expression levels of glial glutamate transportersmight play a critical role in the failure of glutamate clearance[6 58ndash60] Earlier work has found an overall downregulationof protein levels of both GLAST and GLT-1 following astro-cyte activation [22ndash27] indicating that this is causal to theelevation of extracellular glutamate In contrast to this notionother studies reported increased glutamate transport capacityof reactive astrocytes and suggested a protective influence[61 62]
To visualize possible differences in the spatial expressionprofile of GLT-1 and GLAST in strongly reactive astrocytesalong the scar as compared to moderately reactive astro-cytes in its periphery we performed immunohistochemicalstainings These revealed an accumulation of GLT-1 andGLAST immunoreactivity along thick GFAP-positive fiberswhich was particularly pronounced for GLAST Besides theseclusters overall GLAST immunoreactivity seemed unalteredas compared to unlesioned slices whereas GLT-1 immunore-activity seemedweaker and declined with increasing distancefrom the scar Clustering of glutamate transporters has beendescribed by several reports [48 63 64] In developinghippocampal astrocytes cluster formation was preferentiallyfound in branches opposed to synapses and was increasedwith increased neuronal activity indicating that it is neces-sary to copewith synaptic release of glutamate [36] Along thesame lines it was observed that sustained astroglial activationby ciliary neurotrophic factor (CNTF) in the rat striatuminduced a concentration of GLAST and GLT-1 into raftmicrodomains and improved glutamate clearance indicatingthat cluster formation increased the cellular capacity for glu-tamate uptake [61] However another study found glutamate-induced clustering of GLT-1 that induced its endocytosis andintracellular trafficking without changing the total expressionlevels as detected by western blots arguing for a decrease infunctional glial glutamate uptake capacity [49] In the presentstudy antibodies against GLT-1 and GLAST were employedfor immunohistochemistry after permeabilization of theplasma membrane The clusters of GLAST and GLT-1 alongGFAP-positive fibers in the scar region (as GLAST andGLT-1immunoreactivity in general) could thus represent glutamatetransporters in the plasmamembrane or in intracellular com-partments or both Consequently the observed clustering
does not allow a prediction about possible functional changesin glutamate uptake
43 Lesion-Induced Changes in Functional Glutamate UptakeCapacity We probed for the functional activation of gluta-mate transport by application of the transportable agonistD-aspartate which results in an increase in the intracellularsodium concentration of astrocytes [8 9] The amplitudesof D-aspartate-induced sodium transients in astrocytes incontrol organotypic slices were similar to those reportedfrom astrocytes in acute slices obtained in our lab [33]indicating similar cellular glutamate transport capacity inboth preparations
Following a lesion D-aspartate-induced intracellularsodium signalswere clearly dampened in scar cellsMoreoverthe slope of the increase in sodium was significantly reducedand the decay back to baseline was slowed Because D-aspartate was applied at nearly saturating concentrationthe reduction in peak amplitudes suggests a reduction infunctional glutamate uptake capacity in cells close to thelesion which might be mediated by a decrease in the overallnumber of glutamate transporters available at the plasmamembrane In contrast the alteration in the kinetics of theD-aspartate-induced sodium signals can be explained by thecellular hypertrophy that was observed in reactive astrocytesAt an equal transport-mediated sodium influx across themembrane the slope of resulting changes in the sodiumconcentration will be decreased in cells with a larger volume
Furthermore the decay of intracellular sodium transientsback to baseline is mainly governed by the activity of theNa+K+-ATPase as well as by diffusion [8] The slower recov-ery is in line with earlier studies reporting a downregulationof the sodium pump following reactive gliosis [65 66]Moreover sodium is not buffered in the cell and travels solelyby means of diffusion in the cytoplasm Hindered diffusionresulting from increased cytoplasmic protein content orimpaired gap junction coupling which both occur uponastrocytic activation might thus also partially contribute todelayed sodium recovery [12 34 67]
In contrast to scar cells the amplitude of D-aspartate-induced intracellular sodium signals was not significantlyaltered in moderately activated astrocytes in the periphery ofthe lesion indicating that the overall number of functionalglutamate transporters was not significantly altered Becausethese cells showed hypertrophy as well the slowed kinetics ofthe sodium transients are in line with an increased cellularvolume as well as a possible downregulation of Na+K+-ATPase andor a slowed diffusion as argued above
Taken together our results indicate a significant reduc-tion in glutamate uptake capacity in strongly activated astro-cytes in the scar region Thus the prominent clustering ofGLAST and GLT-1 immunoreactivity along GFAP-positivestructures close to the lesion is likely to reflect a loss ofglutamate transporters from the plasma membrane upontransporter internalization as reported by Nakagawa et al[49] The authors of the latter study speculated that large andprolonged increases in glutamate concentrations are neces-sary to induce such cluster formation and endocytosis in thetissue as they would possibly only occur under pathological
14 ISRN Neurology
conditions Indeed ambient glutamate concentrations wereshown to be significantly increased after traumatic braininjury [68ndash70] Along these lines it can be assumed thatin our model system extracellular glutamate concentrationsrose less severely in the periphery of the lesion preventinga comparable clustering and loss of functional glutamatetransport activity in moderately activated astrocytes
5 Conclusions
Our results show complex changes in glutamate transporterexpression and function during astrocyte activation followingmechanical injury They confirm that the degree of astrocyteactivation depends on the distance from the insult and com-prises different endpoints regarding cellular morphology andphysiology Furthermore our results highlight that astrocyteswhich show different grades of reactivity also display discretechanges in glutamate transporter expression and functionWhile immunohistochemistry revealed a prominent clus-tering of GLT-1 and GLAST immunoreactivity in the scarregion our functional assay clearly showed that glutamateuptake capacity is strongly reduced in scar cells while itis largely maintained in moderately activated astrocytes inthe periphery Thus mild-to-moderate astrogliosis in theperiphery of a mechanical lesion does not necessarily seemto be accompanied by a significant change in glial glutamateuptake capacity At the glial scar itself in strongly reactiveastrocytes a clustering of glutamate transporters is observedthat apparently goes along with a severe functional reductionin astroglial glutamate uptake which may contribute toglutamate-mediated excitotoxicity in this region
Conflict of Interests
The authors of the paper declare that they do not haveany direct financial relation with the commercial identitiesmentioned in the paper that might lead to a conflict ofinterests
Acknowledgments
This study was supported by the Deutsche Forschungsge-meinschaft (DFG Ro23274-3) The authors thank SimoneDurry and Claudia Roderigo for expert technical help
References
[1] N C Danbolt ldquoGlutamate uptakerdquo Progress in Neurobiologyvol 65 no 1 pp 1ndash105 2001
[2] A V Tzingounis and J I Wadiche ldquoGlutamate transportersconfining runaway excitation by shaping synaptic transmis-sionrdquo Nature Reviews Neuroscience vol 8 no 12 pp 935ndash9472007
[3] D E Bergles J S Diamond and C E Jahr ldquoClearance ofglutamate inside the synapse and beyondrdquo Current Opinion inNeurobiology vol 9 no 3 pp 293ndash298 1999
[4] C M Anderson and R A Swanson ldquoAstrocyte glutamatetransport review of properties regulation and physiologicalfunctionsrdquo Glia vol 32 no 1 pp 1ndash14 2000
[5] P Marcaggi and D Attwell ldquoRole of glial amino acid trans-porters in synaptic transmission and brain energeticsrdquoGlia vol47 no 3 pp 217ndash225 2004
[6] N J Maragakis and J D Rothstein ldquoGlutamate transportersanimal models to neurologic diseaserdquo Neurobiology of Diseasevol 15 no 3 pp 461ndash473 2004
[7] G Gegelashvili and A Schousboe ldquoCellular distribution andkinetic properties of high-affinity glutamate transportersrdquoBrainResearch Bulletin vol 45 no 3 pp 233ndash238 1998
[8] J W Deitmer and C R Rose ldquoIon changes and signalling inperisynaptic gliardquo Brain Research Reviews vol 63 no 1-2 pp113ndash129 2010
[9] S Kirischuk V Parpura and A Verkhratsky ldquoSodium dynam-ics another key to astroglial excitabilityrdquo Trends in Neuro-sciences vol 35 no 8 pp 497ndash506 2012
[10] M V Sofroniew ldquoMolecular dissection of reactive astrogliosisand glial scar formationrdquo Trends in Neurosciences vol 32 no12 pp 638ndash647 2009
[11] J L Ridet S K Malhotra A Privat and F H Gage ldquoReactiveastrocytes cellular and molecular cues to biological functionrdquoTrends in Neurosciences vol 20 no 12 pp 570ndash577 1997
[12] J Middeldorp and E M Hol ldquoGFAP in health and diseaserdquoProgress in Neurobiology vol 93 no 3 pp 421ndash443 2011
[13] M Pekny and M Nilsson ldquoAstrocyte activation and reactivegliosisrdquo Glia vol 50 no 4 pp 427ndash434 2005
[14] A Bordey S A Lyons J J Hablitz and H SontheimerldquoElectrophysiological characteristics of reactive astrocytes inexperimental cortical dysplasiardquo Journal of Neurophysiologyvol 85 no 4 pp 1719ndash1731 2001
[15] R Jabs G Seifert and C Steinhauser ldquoAstrocytic functionand its alteration in the epileptic brainrdquo Epilepsia vol 49supplement 2 pp 3ndash12 2008
[16] A Buffo I Rite P Tripathi et al ldquoOrigin and progeny ofreactive gliosis a source of multipotent cells in the injuredbrainrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 105 no 9 pp 3581ndash3586 2008
[17] S Robel B Berninger and M Gotz ldquoThe stem cell potential ofglia lessons from reactive gliosisrdquoNature Reviews Neurosciencevol 12 no 2 pp 88ndash104 2011
[18] H Kawano J Kimura-Kuroda Y Komuta et al ldquoRole of thelesion scar in the response to damage and repair of the centralnervous systemrdquo Cell and Tissue Research vol 349 no 1 pp169ndash180 2012
[19] G Yiu and Z He ldquoGlial inhibition of CNS axon regenerationrdquoNature Reviews Neuroscience vol 7 no 8 pp 617ndash627 2006
[20] M T Fitch and J Silver ldquoCNS injury glial scars and inflamma-tion inhibitory extracellular matrices and regeneration failurerdquoExperimental Neurology vol 209 no 2 pp 294ndash301 2008
[21] L Li A Lundkvist D Andersson et al ldquoProtective role ofreactive astrocytes in brain ischemiardquo Journal of Cerebral BloodFlow amp Metabolism vol 28 no 3 pp 468ndash481 2008
[22] F K H van Landeghem T Weiss M Oehmichen and AVonDeimling ldquoDecreased expression of glutamate transportersin astrocytes after human traumatic brain injuryrdquo Journal ofNeurotrauma vol 23 no 10 pp 1518ndash1528 2006
[23] V L Raghavendra Rao M K Baskaya A Dogan J DRothstein and R J Dempsey ldquoTraumatic brain injury down-regulates glial glutamate transporter (GLT- 1 and GLAST)proteins in rat brainrdquo Journal of Neurochemistry vol 70 no 5pp 2020ndash2027 1998
ISRN Neurology 15
[24] J-C Chen H Hsu-Chou J-L Lu et al ldquoDown-regulation ofthe glial glutamate transporter GLT-1 in rat hippocampus andstriatum and its modulation by a group III metabotropic glu-tamate receptor antagonist following transient global forebrainischemiardquoNeuropharmacology vol 49 no 5 pp 703ndash714 2005
[25] S D Ginsberg L J Martin and J D Rothstein ldquoRegional deaf-ferentation down-regulates subtypes of glutamate transporterproteinsrdquo Journal of Neurochemistry vol 65 no 6 pp 2800ndash2803 1995
[26] J E Springer R D Azbill R J Mark J G Begley GWaeg andM P Mattson ldquo4-Hydroxynonenal a lipid peroxidation prod-uct rapidly accumulates following traumatic spinal cord injuryand inhibits glutamate uptakerdquo Journal of Neurochemistry vol68 no 6 pp 2469ndash2476 1997
[27] M B Moretto N S Arteni D Lavinsky et al ldquoHypoxic-ischemic insult decreases glutamate uptake by hippocampalslices from neonatal rats Prevention by guanosinerdquo Experimen-tal Neurology vol 195 no 2 pp 400ndash406 2005
[28] L Stoppini P-A Buchs and D Muller ldquoA simple method fororganotypic cultures of nervous tissuerdquo Journal of NeuroscienceMethods vol 37 no 2 pp 173ndash182 1991
[29] A Nimmerjahn F Kirchhoff J N D Kerr and F HelmchenldquoSulforhodamine 101 as a specific marker of astroglia in theneocortex in vivordquo Nat Methods vol 1 no 1 pp 31ndash37 2004
[30] K W Kafitz S D Meier J Stephan and C R Rose ldquoDevel-opmental profile and properties of sulforhodamine 101-Labeledglial cells in acute brain slices of rat hippocampusrdquo Journal ofNeuroscience Methods vol 169 no 1 pp 84ndash92 2008
[31] C Schnell Y Hagos and S Hulsmann ldquoActive sulforhodamine101 uptake into hippocampal astrocytesrdquo PLoS One vol 7 no 11Article ID e49398 2012
[32] S D Meier Y Kovalchuk and C R Rose ldquoProperties ofthe new fluorescent Na+ indicator CoroNa Green comparisonwith SBFI and confocal Na+ imagingrdquo Journal of NeuroscienceMethods vol 155 no 2 pp 251ndash259 2006
[33] J Langer and C R Rose ldquoSynaptically induced sodium signalsin hippocampal astrocytes in siturdquo Journal of Physiology vol587 part 24 pp 5859ndash5877 2009
[34] J Langer J Stephan M Theis and C R Rose ldquoGap junctionsmediate intercellular spread of sodium between hippocampalastrocytes in siturdquo Glia vol 60 no 2 pp 239ndash252 2012
[35] C R Rose andB R Ransom ldquoIntracellular sodiumhomeostasisin rat hippocampal astrocytesrdquo Journal of Physiology vol 491part 2 no 2 pp 291ndash305 1996
[36] A M Benediktsson G S Marrs J C Tu et al ldquoNeuronalactivity regulates glutamate transporter dynamics in developingastrocytesrdquo Glia vol 60 no 2 pp 175ndash188 2012
[37] M Zhou G P Schools and H K Kimelberg ldquoDevelopment ofGLAST(+) astrocytes andNG2(+) glia in rat hippocampus CA1mature astrocytes are electrophysiologically passiverdquo Journal ofNeurophysiology vol 95 no 1 pp 134ndash143 2006
[38] I Suarez G Bodega and B Fernandez ldquoModulation of glu-tamate transporters (GLAST GLT-1 and EAAC1) in the ratcerebellum following portocaval anastomosisrdquo Brain Researchvol 859 no 2 pp 293ndash302 2000
[39] X Zhu D E Bergles and A Nishiyama ldquoNG2 cells generateboth oligodendrocytes and gray matter astrocytesrdquo Develop-ment vol 135 no 1 pp 145ndash157 2008
[40] B Brunne S ZhaoADerouiche et al ldquoOriginmaturation andastroglial transformation of secondary radial glial cells in thedeveloping dentate gyrusrdquo Glia vol 58 no 13 pp 1553ndash15692010
[41] Y Wu A-Q Zhang and D T Yew ldquoAge related changes ofvarious markers of astrocytes in senescence-accelerated micehippocampusrdquo Neurochemistry International vol 46 no 7 pp565ndash574 2005
[42] S Magavi D Friedmann G Banks A Stolfi and C LoisldquoCoincident generation of pyramidal neurons and protoplasmicastrocytes in neocortical columnsrdquo Journal of Neuroscience vol32 no 14 pp 4762ndash4772 2012
[43] M Olabarria H N Noristani A Verkhratsky and J JRodrıguez ldquoConcomitant astroglial atrophy and astrogliosis ina triple transgenic animal model of Alzheimerrsquos diseaserdquo Gliavol 58 no 7 pp 831ndash838 2010
[44] A M Benediktsson S J Schachtele S H Green and M EDailey ldquoBallistic labeling and dynamic imaging of astrocytesin organotypic hippocampal slice culturesrdquo Journal of Neuro-science Methods vol 141 no 1 pp 41ndash53 2005
[45] I Lushnikova G Skibo D Muller and I Nikonenko ldquoSynapticpotentiation induces increased glial coverage of excitatorysynapses in CA1 hippocampusrdquoHippocampus vol 19 no 8 pp753ndash762 2009
[46] E Raponi F Agenes C Delphin et al ldquoS100B expressiondefines a state in which GFAP-expressing cells lose their neuralstem cell potential and acquire a more mature developmentalstagerdquo Glia vol 55 no 2 pp 165ndash177 2007
[47] C G Schipke C Boucsein C Ohlemeyer F Kirchhoff andH Kettenmann ldquoAstrocyte Ca2+ waves trigger responses inmicroglial cells in brain slicesrdquo The FASEB Journal vol 16 no2 pp 255ndash257 2002
[48] J Zhou and M L Sutherland ldquoGlutamate transporter clusterformation in astrocytic processes regulates glutamate uptakeactivityrdquo Journal of Neuroscience vol 24 no 28 pp 6301ndash63062004
[49] T Nakagawa Y Otsubo Y Yatani H Shirakawa and S KanekoldquoMechanisms of substrate transport-induced clustering of aglial glutamate transporter GLT-1 in astroglial-neuronal cul-turesrdquo European Journal of Neuroscience vol 28 no 9 pp 1719ndash1730 2008
[50] J-Y Chatton P Marquet and P J Magistretti ldquoA quantitativeanalysis of L-glutamate-regulated Na+ dynamics in mousecortical astrocytes Implications for cellular bioenergeticsrdquoEuropean Journal of Neuroscience vol 12 no 11 pp 3843ndash38532000
[51] E Forster S Zhao and M Frotscher ldquoLaminating the hip-pocampusrdquo Nature Reviews Neuroscience vol 7 no 4 pp 259ndash267 2006
[52] B H Gahwiler M Capogna D Debanne R A McKinney andS M Thompson ldquoOrganotypic slice cultures a technique hascome of agerdquo Trends in Neurosciences vol 20 no 10 pp 471ndash477 1997
[53] P E Kunkler and R P Kraig ldquoReactive astrocytosis fromexcitotoxic injury in hippocampal organ culture parallels thatseen in vivordquo Journal of Cerebral Blood Flow and Metabolismvol 17 no 1 pp 26ndash43 1997
[54] I E Holopainen ldquoOrganotypic hippocampal slice cultures amodel system to study basic cellular andmolecularmechanismsof neuronal cell death neuroprotection and synaptic plasticityrdquoNeurochemical Research vol 30 no 12 pp 1521ndash1528 2005
[55] MHaber L Zhou andK KMurai ldquoCooperative astrocyte anddendritic spine dynamics at hippocampal excitatory synapsesrdquoJournal of Neuroscience vol 26 no 35 pp 8881ndash8891 2006
16 ISRN Neurology
[56] H Nishida and S Okabe ldquoDirect astrocytic contacts regulatelocal maturation of dendritic spinesrdquo Journal of Neurosciencevol 27 no 2 pp 331ndash340 2007
[57] A Buffo C Rolando and S Ceruti ldquoAstrocytes in the dam-aged brain molecular and cellular insights into their reactiveresponse and healing potentialrdquoBiochemical Pharmacology vol79 no 2 pp 77ndash89 2010
[58] J-H Yi and A S Hazell ldquoExcitotoxic mechanisms and the roleof astrocytic glutamate transporters in traumatic brain injuryrdquoNeurochemistry International vol 48 no 5 pp 394ndash403 2006
[59] C Werner and K Engelhard ldquoPathophysiology of traumaticbrain injuryrdquo British Journal of Anaesthesia vol 99 no 1 pp4ndash9 2007
[60] G Gegelashvili M B Robinson D Trotti and T RauenldquoRegulation of glutamate transporters in health and diseaserdquoProgress in Brain Research vol 132 pp 267ndash286 2001
[61] C Escartin E Brouillet PGubellini et al ldquoCiliary neurotrophicfactor activates astrocytes redistributes their glutamate trans-porters GLAST and GLT-1 to raft microdomains and improvesglutamate handling in vivordquo Journal of Neuroscience vol 26 no22 pp 5978ndash5989 2006
[62] C Vermeiren M Najimi N Vanhoutte et al ldquoAcute up-regulation of glutamate uptake mediated by mGluR5a in reac-tive astrocytesrdquo Journal of Neurochemistry vol 94 no 2 pp405ndash416 2005
[63] C L Poitry-Yamate L Vutskits and T Rauen ldquoNeuronal-induced and glutamate-dependent activation of glial glutamatetransporter functionrdquo Journal of Neurochemistry vol 82 no 4pp 987ndash997 2002
[64] P M Beart and R D OrsquoShea ldquoTransporters for L-glutamatean update on their molecular pharmacology and pathologicalinvolvementrdquo British Journal of Pharmacology vol 150 no 1pp 5ndash17 2007
[65] F D Lima M A Souza A F Furian et al ldquoNa+K+-ATPaseactivity impairment after experimental traumatic brain injuryrelationship to spatial learning deficits and oxidative stressrdquoBehavioural Brain Research vol 193 no 2 pp 306ndash310 2008
[66] G A Gusarova H E Trejo L A Dada et al ldquoHypoxia leadsto NaK-ATPase downregulation via Ca2+ release-activatedCa2+ channels and AMPK activationrdquo Molecular and CellularBiology vol 31 no 17 pp 3546ndash3556 2011
[67] S S L Chew C S Johnson C R Green and H V Danesh-Meyer ldquoRole of connexin43 in central nervous system injuryrdquoExperimental Neurology vol 225 no 2 pp 250ndash261 2010
[68] P Nilsson L Hillered U Ponten and U Ungerstedt ldquoChangesin cortical extracellular levels of energy-related metabolites andamino acids following concussive brain injury in ratsrdquo Journal ofCerebral Blood Flow and Metabolism vol 10 no 5 pp 631ndash6371990
[69] H Katoh K Sima H Nawashiro K Wada and H ChigasakildquoThe effect of MK-801 on extracellular neuroactive amino acidsin hippocampus after closed head injury followed by hypoxia inratsrdquo Brain Research vol 758 no 1-2 pp 153ndash162 1997
[70] H Koizumi H Fujisawa H Ito T Maekawa X Di and RBullock ldquoEffects of mild hypothermia on cerebral blood flow-independent changes in cortical extracellular levels of aminoacids following contusion trauma in the ratrdquoBrain Research vol747 no 2 pp 304ndash312 1997
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
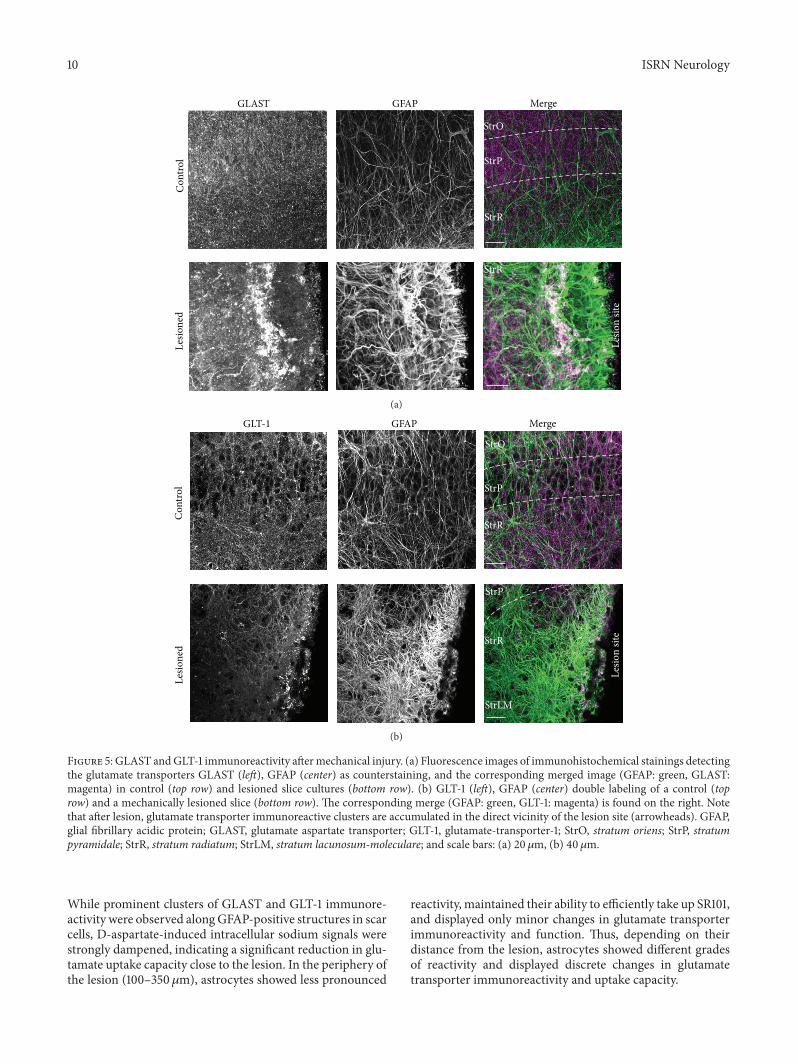
10 ISRN Neurology
Con
trol
GLAST
Lesio
ned
GFAP Merge
StrR
StrP
StrO
Lesio
n sit
e
StrR
(a)
Con
trol
Lesio
ned
GLT-1 GFAP Merge
StrR
StrP
StrO
StrR
StrP
StrLM
Lesio
n sit
e
(b)
Figure 5 GLAST andGLT-1 immunoreactivity aftermechanical injury (a) Fluorescence images of immunohistochemical stainings detectingthe glutamate transporters GLAST (left) GFAP (center) as counterstaining and the corresponding merged image (GFAP green GLASTmagenta) in control (top row) and lesioned slice cultures (bottom row) (b) GLT-1 (left) GFAP (center) double labeling of a control (toprow) and a mechanically lesioned slice (bottom row) The corresponding merge (GFAP green GLT-1 magenta) is found on the right Notethat after lesion glutamate transporter immunoreactive clusters are accumulated in the direct vicinity of the lesion site (arrowheads) GFAPglial fibrillary acidic protein GLAST glutamate aspartate transporter GLT-1 glutamate-transporter-1 StrO stratum oriens StrP stratumpyramidale StrR stratum radiatum StrLM stratum lacunosum-moleculare and scale bars (a) 20 120583m (b) 40 120583m
While prominent clusters of GLAST and GLT-1 immunore-activity were observed alongGFAP-positive structures in scarcells D-aspartate-induced intracellular sodium signals werestrongly dampened indicating a significant reduction in glu-tamate uptake capacity close to the lesion In the periphery ofthe lesion (100ndash350120583m) astrocytes showed less pronounced
reactivity maintained their ability to efficiently take up SR101and displayed only minor changes in glutamate transporterimmunoreactivity and function Thus depending on theirdistance from the lesion astrocytes showed different gradesof reactivity and displayed discrete changes in glutamatetransporter immunoreactivity and uptake capacity
ISRN Neurology 11
SR101 MergeSBFI
StrRStrP
(a)
03
04
05
06
07
0 5 10 15
0
20 25 30 40 0
100
20
40
80
60
0 10 20 30 400
20
40
60
80
100
10 = 55 mM
107 cells in 6 experiments
F (340
380
) (ra
tio)
Intracellular Na+ concentration (mM)Intracellular Na+ concentration (mM)
ΔF
(340
380
) (
)
ΔF
(340
380
) (
) R2 = 097
(b)
D-aspartate
100 120583M500 120583M
750 120583M1000 120583M
1 mM
10 s
Δ
(c)
0 1 2 3 4 50
1
2
3
4
5
Δ so
dium
(mM
)
D-aspartate (mM)
lowast
(d)
Figure 6 Sodium changes induced by activation of glutamate uptake (a) Staining of astrocytes by colabeling with SR101 and SBFI Leftepifluorescence image of theCA1-area of an SR101-loaded organotypic sliceCenter SBFI fluorescence taken from the same areaRight overlaySBFI-loaded SR101-positive astrocytes appear in white (arrowheads) StrP stratum pyramidale StrR stratum radiatum scale bar 20 120583m (b)Left calibration of SBFI fluorescence in an astrocyte Depicted is the change in intracellular SBFI fluorescence ratio in the calibration solutionscontaining ionophores and in response to stepwise changes in the sodium concentration Right mean values plusmn SEM of the normalizedchanges in SBFI fluorescence at different sodium concentrations The black line represents a linear fit of the data points between 10 and40mM sodium illustrating that a 10 change in SBFI fluorescence represents a change in the sodium concentration by 55mM (c) Sodiumtransients in a single astrocyte induced by 500ms applications of D-aspartate at different concentrations (d) Mean values plusmn S E M (119899 = 19)of the peak amplitude of sodium transients at different concentrations of D-aspartate The curve represents a sigmoidal fit of the data
41 Characteristics of Lesion-Induced Reactive Gliosis inOrganotypic Cultures Organotypic tissue slice cultures ofthe hippocampus represent a well-established model systemin which the basic tissue architecture is maintained butwhich still hosts the major advantages of cell cultures suchas good accessibility and control of experimental conditionsThey have been used extensively for the analysis of neuronal
properties and developing neuronal networks [28 51 52]or the study of excitotoxic neuronal damage (eg [53 54])Recent studies also demonstrated that basic morphologicalcharacteristics of astrocytes and the typical glia-synapseorganization are well preserved in organotypic slice cultures(eg [44 45 55 56]) In addition we found a nearly completeoverlap of SR101 staining with the expression of GFP in slice
12 ISRN Neurology
SBFIControl Lesioned
SR101
1
Lesio
n sit
e
SR101 SBFI
2 45
32 4
5
1
StrR
StrP StrP
StrR
3
(a)
ControlScar
Lesioned
1
D-asp
2
D-asp
D-asp
5
4
3Periphery1mM
20 s
Δ Δ
Δ
Δ
Δ
(b)
0102030405060708090
100110
Δ so
dium
tim
e (m
Ms
)
00
02
04
06
08
10
12
0
1
2
3
4
5
6
ns
Control Periphery Scar Control Periphery Scar Control Periphery Scar
Amplitude Slope Decay time
Δ so
dium
(mM
)
120591(s
)lowastlowastlowast
lowastlowastlowastlowastlowastlowast
lowastlowastlowastlowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowast
(c)
Figure 7 D-aspartate-induced astrocyte sodium transients following lesion (a) CA1 area of a control (left) and a lesioned (right) slicepreparation Shown are epifluorescence images of SR101 and SBFI fluorescences Circled areas and numbers indicate the regions of interestanalyzed in the experiment depicted in (b) StrP stratum pyramidale StrR stratum radiatum and scale bar 40 120583m (b) Sodium transientsinduced by pressure application of 1mMD-aspartate (D-asp) for 500ms in the different cells in the control and lesioned slice as indicated in(a) (c) Histograms showing mean values plusmn S E M of the peak amplitude (left) slope (center) and decay time (right) of D-aspartate-inducedsodium transients in astrocytes in control slices (119899 = 161) as well as in cells in the scar region and its periphery (119899 = 111 and 119899 = 26 resp)lowastlowast119875 lt 001 lowastlowastlowast119875 lt 0001 StrP stratum pyramidale StrR stratum radiatum and scale bar 40120583m
cultures obtained from GFPGFAP mice demonstrating thatSR101 is well suited to identify astrocytes in this prepara-tion as described before for acute tissue slices and in vivo[29 30]
Performing a scratch through the CA1 region of organ-otypic slices resulted in the formation of a glial scar along thelesion site as judged by the prominent increase in expressionof GFAP and S100120573 In addition astrocyte cell bodies showed
a significant hypertrophy Furthermore astrocytes in theperiphery of the scar (100ndash350 120583m from the lesion) showedmild to moderate reactivity displaying swollen cell bodiesand only moderate upregulation of GFAP These changesare hallmarks of reactive gliosis observed after tissue injury[10 13 57] Generally the degree of injury and the distance ofthe astrocytes from the site of injury define the degree of theiractivation [11 13]
ISRN Neurology 13
Our study also revealed that strongly reactive astrocytesin the scar region only weakly stain with SR101 The reducedability of strongly activated astrocytes to accumulate SR101indicates that strong glial activation is accompanied by adownregulation of the organic anion transporter responsi-ble for uptake of SR101 This is reminiscent of immatureastrocytes in the early postnatal hippocampus which areSR101-negative [30] and in line with a wealth of experimentalevidence suggesting that reactive gliosis represents a processcomprising a dedifferentiation of astrocytes [17]
42 Changes in Glutamate Transporter Expression followingLesion Glutamate uptake in the hippocampus is mainlyachieved by the glial glutamate transporters GLT-1 andGLAST [1 2] A central factor in many brain pathologiesincluding traumatic brain injury is an increase in extracellu-lar glutamate and excitotoxicity it has been suggested that achange in the expression levels of glial glutamate transportersmight play a critical role in the failure of glutamate clearance[6 58ndash60] Earlier work has found an overall downregulationof protein levels of both GLAST and GLT-1 following astro-cyte activation [22ndash27] indicating that this is causal to theelevation of extracellular glutamate In contrast to this notionother studies reported increased glutamate transport capacityof reactive astrocytes and suggested a protective influence[61 62]
To visualize possible differences in the spatial expressionprofile of GLT-1 and GLAST in strongly reactive astrocytesalong the scar as compared to moderately reactive astro-cytes in its periphery we performed immunohistochemicalstainings These revealed an accumulation of GLT-1 andGLAST immunoreactivity along thick GFAP-positive fiberswhich was particularly pronounced for GLAST Besides theseclusters overall GLAST immunoreactivity seemed unalteredas compared to unlesioned slices whereas GLT-1 immunore-activity seemedweaker and declined with increasing distancefrom the scar Clustering of glutamate transporters has beendescribed by several reports [48 63 64] In developinghippocampal astrocytes cluster formation was preferentiallyfound in branches opposed to synapses and was increasedwith increased neuronal activity indicating that it is neces-sary to copewith synaptic release of glutamate [36] Along thesame lines it was observed that sustained astroglial activationby ciliary neurotrophic factor (CNTF) in the rat striatuminduced a concentration of GLAST and GLT-1 into raftmicrodomains and improved glutamate clearance indicatingthat cluster formation increased the cellular capacity for glu-tamate uptake [61] However another study found glutamate-induced clustering of GLT-1 that induced its endocytosis andintracellular trafficking without changing the total expressionlevels as detected by western blots arguing for a decrease infunctional glial glutamate uptake capacity [49] In the presentstudy antibodies against GLT-1 and GLAST were employedfor immunohistochemistry after permeabilization of theplasma membrane The clusters of GLAST and GLT-1 alongGFAP-positive fibers in the scar region (as GLAST andGLT-1immunoreactivity in general) could thus represent glutamatetransporters in the plasmamembrane or in intracellular com-partments or both Consequently the observed clustering
does not allow a prediction about possible functional changesin glutamate uptake
43 Lesion-Induced Changes in Functional Glutamate UptakeCapacity We probed for the functional activation of gluta-mate transport by application of the transportable agonistD-aspartate which results in an increase in the intracellularsodium concentration of astrocytes [8 9] The amplitudesof D-aspartate-induced sodium transients in astrocytes incontrol organotypic slices were similar to those reportedfrom astrocytes in acute slices obtained in our lab [33]indicating similar cellular glutamate transport capacity inboth preparations
Following a lesion D-aspartate-induced intracellularsodium signalswere clearly dampened in scar cellsMoreoverthe slope of the increase in sodium was significantly reducedand the decay back to baseline was slowed Because D-aspartate was applied at nearly saturating concentrationthe reduction in peak amplitudes suggests a reduction infunctional glutamate uptake capacity in cells close to thelesion which might be mediated by a decrease in the overallnumber of glutamate transporters available at the plasmamembrane In contrast the alteration in the kinetics of theD-aspartate-induced sodium signals can be explained by thecellular hypertrophy that was observed in reactive astrocytesAt an equal transport-mediated sodium influx across themembrane the slope of resulting changes in the sodiumconcentration will be decreased in cells with a larger volume
Furthermore the decay of intracellular sodium transientsback to baseline is mainly governed by the activity of theNa+K+-ATPase as well as by diffusion [8] The slower recov-ery is in line with earlier studies reporting a downregulationof the sodium pump following reactive gliosis [65 66]Moreover sodium is not buffered in the cell and travels solelyby means of diffusion in the cytoplasm Hindered diffusionresulting from increased cytoplasmic protein content orimpaired gap junction coupling which both occur uponastrocytic activation might thus also partially contribute todelayed sodium recovery [12 34 67]
In contrast to scar cells the amplitude of D-aspartate-induced intracellular sodium signals was not significantlyaltered in moderately activated astrocytes in the periphery ofthe lesion indicating that the overall number of functionalglutamate transporters was not significantly altered Becausethese cells showed hypertrophy as well the slowed kinetics ofthe sodium transients are in line with an increased cellularvolume as well as a possible downregulation of Na+K+-ATPase andor a slowed diffusion as argued above
Taken together our results indicate a significant reduc-tion in glutamate uptake capacity in strongly activated astro-cytes in the scar region Thus the prominent clustering ofGLAST and GLT-1 immunoreactivity along GFAP-positivestructures close to the lesion is likely to reflect a loss ofglutamate transporters from the plasma membrane upontransporter internalization as reported by Nakagawa et al[49] The authors of the latter study speculated that large andprolonged increases in glutamate concentrations are neces-sary to induce such cluster formation and endocytosis in thetissue as they would possibly only occur under pathological
14 ISRN Neurology
conditions Indeed ambient glutamate concentrations wereshown to be significantly increased after traumatic braininjury [68ndash70] Along these lines it can be assumed thatin our model system extracellular glutamate concentrationsrose less severely in the periphery of the lesion preventinga comparable clustering and loss of functional glutamatetransport activity in moderately activated astrocytes
5 Conclusions
Our results show complex changes in glutamate transporterexpression and function during astrocyte activation followingmechanical injury They confirm that the degree of astrocyteactivation depends on the distance from the insult and com-prises different endpoints regarding cellular morphology andphysiology Furthermore our results highlight that astrocyteswhich show different grades of reactivity also display discretechanges in glutamate transporter expression and functionWhile immunohistochemistry revealed a prominent clus-tering of GLT-1 and GLAST immunoreactivity in the scarregion our functional assay clearly showed that glutamateuptake capacity is strongly reduced in scar cells while itis largely maintained in moderately activated astrocytes inthe periphery Thus mild-to-moderate astrogliosis in theperiphery of a mechanical lesion does not necessarily seemto be accompanied by a significant change in glial glutamateuptake capacity At the glial scar itself in strongly reactiveastrocytes a clustering of glutamate transporters is observedthat apparently goes along with a severe functional reductionin astroglial glutamate uptake which may contribute toglutamate-mediated excitotoxicity in this region
Conflict of Interests
The authors of the paper declare that they do not haveany direct financial relation with the commercial identitiesmentioned in the paper that might lead to a conflict ofinterests
Acknowledgments
This study was supported by the Deutsche Forschungsge-meinschaft (DFG Ro23274-3) The authors thank SimoneDurry and Claudia Roderigo for expert technical help
References
[1] N C Danbolt ldquoGlutamate uptakerdquo Progress in Neurobiologyvol 65 no 1 pp 1ndash105 2001
[2] A V Tzingounis and J I Wadiche ldquoGlutamate transportersconfining runaway excitation by shaping synaptic transmis-sionrdquo Nature Reviews Neuroscience vol 8 no 12 pp 935ndash9472007
[3] D E Bergles J S Diamond and C E Jahr ldquoClearance ofglutamate inside the synapse and beyondrdquo Current Opinion inNeurobiology vol 9 no 3 pp 293ndash298 1999
[4] C M Anderson and R A Swanson ldquoAstrocyte glutamatetransport review of properties regulation and physiologicalfunctionsrdquo Glia vol 32 no 1 pp 1ndash14 2000
[5] P Marcaggi and D Attwell ldquoRole of glial amino acid trans-porters in synaptic transmission and brain energeticsrdquoGlia vol47 no 3 pp 217ndash225 2004
[6] N J Maragakis and J D Rothstein ldquoGlutamate transportersanimal models to neurologic diseaserdquo Neurobiology of Diseasevol 15 no 3 pp 461ndash473 2004
[7] G Gegelashvili and A Schousboe ldquoCellular distribution andkinetic properties of high-affinity glutamate transportersrdquoBrainResearch Bulletin vol 45 no 3 pp 233ndash238 1998
[8] J W Deitmer and C R Rose ldquoIon changes and signalling inperisynaptic gliardquo Brain Research Reviews vol 63 no 1-2 pp113ndash129 2010
[9] S Kirischuk V Parpura and A Verkhratsky ldquoSodium dynam-ics another key to astroglial excitabilityrdquo Trends in Neuro-sciences vol 35 no 8 pp 497ndash506 2012
[10] M V Sofroniew ldquoMolecular dissection of reactive astrogliosisand glial scar formationrdquo Trends in Neurosciences vol 32 no12 pp 638ndash647 2009
[11] J L Ridet S K Malhotra A Privat and F H Gage ldquoReactiveastrocytes cellular and molecular cues to biological functionrdquoTrends in Neurosciences vol 20 no 12 pp 570ndash577 1997
[12] J Middeldorp and E M Hol ldquoGFAP in health and diseaserdquoProgress in Neurobiology vol 93 no 3 pp 421ndash443 2011
[13] M Pekny and M Nilsson ldquoAstrocyte activation and reactivegliosisrdquo Glia vol 50 no 4 pp 427ndash434 2005
[14] A Bordey S A Lyons J J Hablitz and H SontheimerldquoElectrophysiological characteristics of reactive astrocytes inexperimental cortical dysplasiardquo Journal of Neurophysiologyvol 85 no 4 pp 1719ndash1731 2001
[15] R Jabs G Seifert and C Steinhauser ldquoAstrocytic functionand its alteration in the epileptic brainrdquo Epilepsia vol 49supplement 2 pp 3ndash12 2008
[16] A Buffo I Rite P Tripathi et al ldquoOrigin and progeny ofreactive gliosis a source of multipotent cells in the injuredbrainrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 105 no 9 pp 3581ndash3586 2008
[17] S Robel B Berninger and M Gotz ldquoThe stem cell potential ofglia lessons from reactive gliosisrdquoNature Reviews Neurosciencevol 12 no 2 pp 88ndash104 2011
[18] H Kawano J Kimura-Kuroda Y Komuta et al ldquoRole of thelesion scar in the response to damage and repair of the centralnervous systemrdquo Cell and Tissue Research vol 349 no 1 pp169ndash180 2012
[19] G Yiu and Z He ldquoGlial inhibition of CNS axon regenerationrdquoNature Reviews Neuroscience vol 7 no 8 pp 617ndash627 2006
[20] M T Fitch and J Silver ldquoCNS injury glial scars and inflamma-tion inhibitory extracellular matrices and regeneration failurerdquoExperimental Neurology vol 209 no 2 pp 294ndash301 2008
[21] L Li A Lundkvist D Andersson et al ldquoProtective role ofreactive astrocytes in brain ischemiardquo Journal of Cerebral BloodFlow amp Metabolism vol 28 no 3 pp 468ndash481 2008
[22] F K H van Landeghem T Weiss M Oehmichen and AVonDeimling ldquoDecreased expression of glutamate transportersin astrocytes after human traumatic brain injuryrdquo Journal ofNeurotrauma vol 23 no 10 pp 1518ndash1528 2006
[23] V L Raghavendra Rao M K Baskaya A Dogan J DRothstein and R J Dempsey ldquoTraumatic brain injury down-regulates glial glutamate transporter (GLT- 1 and GLAST)proteins in rat brainrdquo Journal of Neurochemistry vol 70 no 5pp 2020ndash2027 1998
ISRN Neurology 15
[24] J-C Chen H Hsu-Chou J-L Lu et al ldquoDown-regulation ofthe glial glutamate transporter GLT-1 in rat hippocampus andstriatum and its modulation by a group III metabotropic glu-tamate receptor antagonist following transient global forebrainischemiardquoNeuropharmacology vol 49 no 5 pp 703ndash714 2005
[25] S D Ginsberg L J Martin and J D Rothstein ldquoRegional deaf-ferentation down-regulates subtypes of glutamate transporterproteinsrdquo Journal of Neurochemistry vol 65 no 6 pp 2800ndash2803 1995
[26] J E Springer R D Azbill R J Mark J G Begley GWaeg andM P Mattson ldquo4-Hydroxynonenal a lipid peroxidation prod-uct rapidly accumulates following traumatic spinal cord injuryand inhibits glutamate uptakerdquo Journal of Neurochemistry vol68 no 6 pp 2469ndash2476 1997
[27] M B Moretto N S Arteni D Lavinsky et al ldquoHypoxic-ischemic insult decreases glutamate uptake by hippocampalslices from neonatal rats Prevention by guanosinerdquo Experimen-tal Neurology vol 195 no 2 pp 400ndash406 2005
[28] L Stoppini P-A Buchs and D Muller ldquoA simple method fororganotypic cultures of nervous tissuerdquo Journal of NeuroscienceMethods vol 37 no 2 pp 173ndash182 1991
[29] A Nimmerjahn F Kirchhoff J N D Kerr and F HelmchenldquoSulforhodamine 101 as a specific marker of astroglia in theneocortex in vivordquo Nat Methods vol 1 no 1 pp 31ndash37 2004
[30] K W Kafitz S D Meier J Stephan and C R Rose ldquoDevel-opmental profile and properties of sulforhodamine 101-Labeledglial cells in acute brain slices of rat hippocampusrdquo Journal ofNeuroscience Methods vol 169 no 1 pp 84ndash92 2008
[31] C Schnell Y Hagos and S Hulsmann ldquoActive sulforhodamine101 uptake into hippocampal astrocytesrdquo PLoS One vol 7 no 11Article ID e49398 2012
[32] S D Meier Y Kovalchuk and C R Rose ldquoProperties ofthe new fluorescent Na+ indicator CoroNa Green comparisonwith SBFI and confocal Na+ imagingrdquo Journal of NeuroscienceMethods vol 155 no 2 pp 251ndash259 2006
[33] J Langer and C R Rose ldquoSynaptically induced sodium signalsin hippocampal astrocytes in siturdquo Journal of Physiology vol587 part 24 pp 5859ndash5877 2009
[34] J Langer J Stephan M Theis and C R Rose ldquoGap junctionsmediate intercellular spread of sodium between hippocampalastrocytes in siturdquo Glia vol 60 no 2 pp 239ndash252 2012
[35] C R Rose andB R Ransom ldquoIntracellular sodiumhomeostasisin rat hippocampal astrocytesrdquo Journal of Physiology vol 491part 2 no 2 pp 291ndash305 1996
[36] A M Benediktsson G S Marrs J C Tu et al ldquoNeuronalactivity regulates glutamate transporter dynamics in developingastrocytesrdquo Glia vol 60 no 2 pp 175ndash188 2012
[37] M Zhou G P Schools and H K Kimelberg ldquoDevelopment ofGLAST(+) astrocytes andNG2(+) glia in rat hippocampus CA1mature astrocytes are electrophysiologically passiverdquo Journal ofNeurophysiology vol 95 no 1 pp 134ndash143 2006
[38] I Suarez G Bodega and B Fernandez ldquoModulation of glu-tamate transporters (GLAST GLT-1 and EAAC1) in the ratcerebellum following portocaval anastomosisrdquo Brain Researchvol 859 no 2 pp 293ndash302 2000
[39] X Zhu D E Bergles and A Nishiyama ldquoNG2 cells generateboth oligodendrocytes and gray matter astrocytesrdquo Develop-ment vol 135 no 1 pp 145ndash157 2008
[40] B Brunne S ZhaoADerouiche et al ldquoOriginmaturation andastroglial transformation of secondary radial glial cells in thedeveloping dentate gyrusrdquo Glia vol 58 no 13 pp 1553ndash15692010
[41] Y Wu A-Q Zhang and D T Yew ldquoAge related changes ofvarious markers of astrocytes in senescence-accelerated micehippocampusrdquo Neurochemistry International vol 46 no 7 pp565ndash574 2005
[42] S Magavi D Friedmann G Banks A Stolfi and C LoisldquoCoincident generation of pyramidal neurons and protoplasmicastrocytes in neocortical columnsrdquo Journal of Neuroscience vol32 no 14 pp 4762ndash4772 2012
[43] M Olabarria H N Noristani A Verkhratsky and J JRodrıguez ldquoConcomitant astroglial atrophy and astrogliosis ina triple transgenic animal model of Alzheimerrsquos diseaserdquo Gliavol 58 no 7 pp 831ndash838 2010
[44] A M Benediktsson S J Schachtele S H Green and M EDailey ldquoBallistic labeling and dynamic imaging of astrocytesin organotypic hippocampal slice culturesrdquo Journal of Neuro-science Methods vol 141 no 1 pp 41ndash53 2005
[45] I Lushnikova G Skibo D Muller and I Nikonenko ldquoSynapticpotentiation induces increased glial coverage of excitatorysynapses in CA1 hippocampusrdquoHippocampus vol 19 no 8 pp753ndash762 2009
[46] E Raponi F Agenes C Delphin et al ldquoS100B expressiondefines a state in which GFAP-expressing cells lose their neuralstem cell potential and acquire a more mature developmentalstagerdquo Glia vol 55 no 2 pp 165ndash177 2007
[47] C G Schipke C Boucsein C Ohlemeyer F Kirchhoff andH Kettenmann ldquoAstrocyte Ca2+ waves trigger responses inmicroglial cells in brain slicesrdquo The FASEB Journal vol 16 no2 pp 255ndash257 2002
[48] J Zhou and M L Sutherland ldquoGlutamate transporter clusterformation in astrocytic processes regulates glutamate uptakeactivityrdquo Journal of Neuroscience vol 24 no 28 pp 6301ndash63062004
[49] T Nakagawa Y Otsubo Y Yatani H Shirakawa and S KanekoldquoMechanisms of substrate transport-induced clustering of aglial glutamate transporter GLT-1 in astroglial-neuronal cul-turesrdquo European Journal of Neuroscience vol 28 no 9 pp 1719ndash1730 2008
[50] J-Y Chatton P Marquet and P J Magistretti ldquoA quantitativeanalysis of L-glutamate-regulated Na+ dynamics in mousecortical astrocytes Implications for cellular bioenergeticsrdquoEuropean Journal of Neuroscience vol 12 no 11 pp 3843ndash38532000
[51] E Forster S Zhao and M Frotscher ldquoLaminating the hip-pocampusrdquo Nature Reviews Neuroscience vol 7 no 4 pp 259ndash267 2006
[52] B H Gahwiler M Capogna D Debanne R A McKinney andS M Thompson ldquoOrganotypic slice cultures a technique hascome of agerdquo Trends in Neurosciences vol 20 no 10 pp 471ndash477 1997
[53] P E Kunkler and R P Kraig ldquoReactive astrocytosis fromexcitotoxic injury in hippocampal organ culture parallels thatseen in vivordquo Journal of Cerebral Blood Flow and Metabolismvol 17 no 1 pp 26ndash43 1997
[54] I E Holopainen ldquoOrganotypic hippocampal slice cultures amodel system to study basic cellular andmolecularmechanismsof neuronal cell death neuroprotection and synaptic plasticityrdquoNeurochemical Research vol 30 no 12 pp 1521ndash1528 2005
[55] MHaber L Zhou andK KMurai ldquoCooperative astrocyte anddendritic spine dynamics at hippocampal excitatory synapsesrdquoJournal of Neuroscience vol 26 no 35 pp 8881ndash8891 2006
16 ISRN Neurology
[56] H Nishida and S Okabe ldquoDirect astrocytic contacts regulatelocal maturation of dendritic spinesrdquo Journal of Neurosciencevol 27 no 2 pp 331ndash340 2007
[57] A Buffo C Rolando and S Ceruti ldquoAstrocytes in the dam-aged brain molecular and cellular insights into their reactiveresponse and healing potentialrdquoBiochemical Pharmacology vol79 no 2 pp 77ndash89 2010
[58] J-H Yi and A S Hazell ldquoExcitotoxic mechanisms and the roleof astrocytic glutamate transporters in traumatic brain injuryrdquoNeurochemistry International vol 48 no 5 pp 394ndash403 2006
[59] C Werner and K Engelhard ldquoPathophysiology of traumaticbrain injuryrdquo British Journal of Anaesthesia vol 99 no 1 pp4ndash9 2007
[60] G Gegelashvili M B Robinson D Trotti and T RauenldquoRegulation of glutamate transporters in health and diseaserdquoProgress in Brain Research vol 132 pp 267ndash286 2001
[61] C Escartin E Brouillet PGubellini et al ldquoCiliary neurotrophicfactor activates astrocytes redistributes their glutamate trans-porters GLAST and GLT-1 to raft microdomains and improvesglutamate handling in vivordquo Journal of Neuroscience vol 26 no22 pp 5978ndash5989 2006
[62] C Vermeiren M Najimi N Vanhoutte et al ldquoAcute up-regulation of glutamate uptake mediated by mGluR5a in reac-tive astrocytesrdquo Journal of Neurochemistry vol 94 no 2 pp405ndash416 2005
[63] C L Poitry-Yamate L Vutskits and T Rauen ldquoNeuronal-induced and glutamate-dependent activation of glial glutamatetransporter functionrdquo Journal of Neurochemistry vol 82 no 4pp 987ndash997 2002
[64] P M Beart and R D OrsquoShea ldquoTransporters for L-glutamatean update on their molecular pharmacology and pathologicalinvolvementrdquo British Journal of Pharmacology vol 150 no 1pp 5ndash17 2007
[65] F D Lima M A Souza A F Furian et al ldquoNa+K+-ATPaseactivity impairment after experimental traumatic brain injuryrelationship to spatial learning deficits and oxidative stressrdquoBehavioural Brain Research vol 193 no 2 pp 306ndash310 2008
[66] G A Gusarova H E Trejo L A Dada et al ldquoHypoxia leadsto NaK-ATPase downregulation via Ca2+ release-activatedCa2+ channels and AMPK activationrdquo Molecular and CellularBiology vol 31 no 17 pp 3546ndash3556 2011
[67] S S L Chew C S Johnson C R Green and H V Danesh-Meyer ldquoRole of connexin43 in central nervous system injuryrdquoExperimental Neurology vol 225 no 2 pp 250ndash261 2010
[68] P Nilsson L Hillered U Ponten and U Ungerstedt ldquoChangesin cortical extracellular levels of energy-related metabolites andamino acids following concussive brain injury in ratsrdquo Journal ofCerebral Blood Flow and Metabolism vol 10 no 5 pp 631ndash6371990
[69] H Katoh K Sima H Nawashiro K Wada and H ChigasakildquoThe effect of MK-801 on extracellular neuroactive amino acidsin hippocampus after closed head injury followed by hypoxia inratsrdquo Brain Research vol 758 no 1-2 pp 153ndash162 1997
[70] H Koizumi H Fujisawa H Ito T Maekawa X Di and RBullock ldquoEffects of mild hypothermia on cerebral blood flow-independent changes in cortical extracellular levels of aminoacids following contusion trauma in the ratrdquoBrain Research vol747 no 2 pp 304ndash312 1997
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
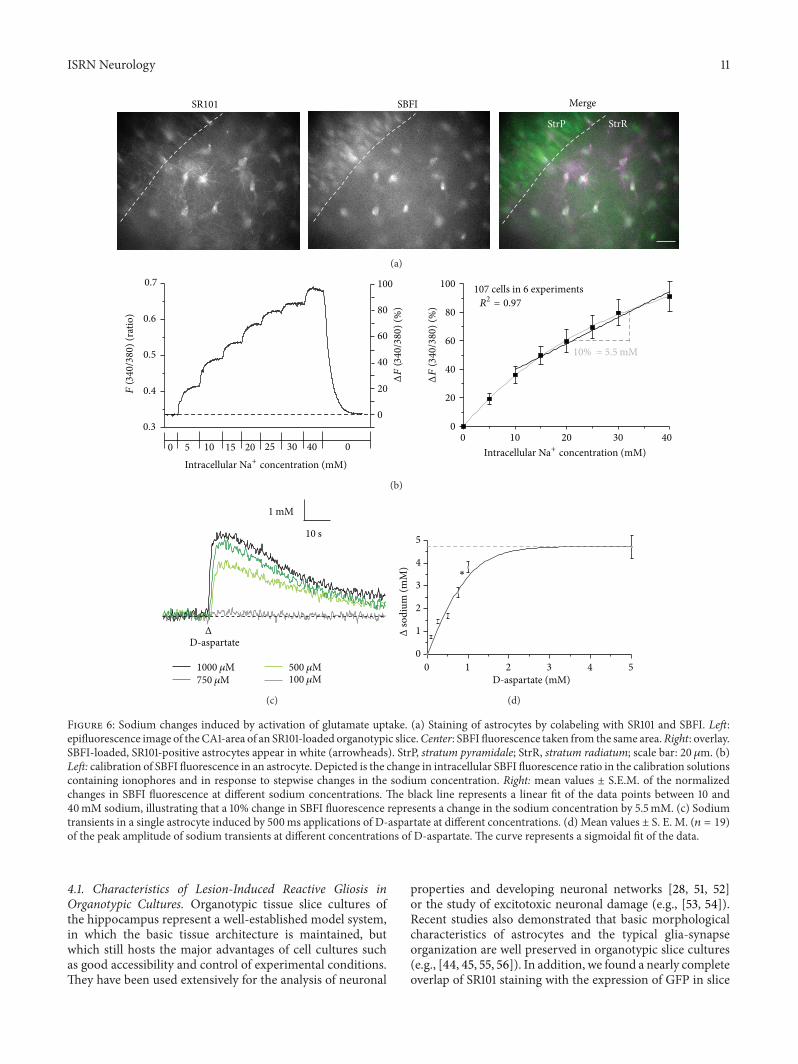
ISRN Neurology 11
SR101 MergeSBFI
StrRStrP
(a)
03
04
05
06
07
0 5 10 15
0
20 25 30 40 0
100
20
40
80
60
0 10 20 30 400
20
40
60
80
100
10 = 55 mM
107 cells in 6 experiments
F (340
380
) (ra
tio)
Intracellular Na+ concentration (mM)Intracellular Na+ concentration (mM)
ΔF
(340
380
) (
)
ΔF
(340
380
) (
) R2 = 097
(b)
D-aspartate
100 120583M500 120583M
750 120583M1000 120583M
1 mM
10 s
Δ
(c)
0 1 2 3 4 50
1
2
3
4
5
Δ so
dium
(mM
)
D-aspartate (mM)
lowast
(d)
Figure 6 Sodium changes induced by activation of glutamate uptake (a) Staining of astrocytes by colabeling with SR101 and SBFI Leftepifluorescence image of theCA1-area of an SR101-loaded organotypic sliceCenter SBFI fluorescence taken from the same areaRight overlaySBFI-loaded SR101-positive astrocytes appear in white (arrowheads) StrP stratum pyramidale StrR stratum radiatum scale bar 20 120583m (b)Left calibration of SBFI fluorescence in an astrocyte Depicted is the change in intracellular SBFI fluorescence ratio in the calibration solutionscontaining ionophores and in response to stepwise changes in the sodium concentration Right mean values plusmn SEM of the normalizedchanges in SBFI fluorescence at different sodium concentrations The black line represents a linear fit of the data points between 10 and40mM sodium illustrating that a 10 change in SBFI fluorescence represents a change in the sodium concentration by 55mM (c) Sodiumtransients in a single astrocyte induced by 500ms applications of D-aspartate at different concentrations (d) Mean values plusmn S E M (119899 = 19)of the peak amplitude of sodium transients at different concentrations of D-aspartate The curve represents a sigmoidal fit of the data
41 Characteristics of Lesion-Induced Reactive Gliosis inOrganotypic Cultures Organotypic tissue slice cultures ofthe hippocampus represent a well-established model systemin which the basic tissue architecture is maintained butwhich still hosts the major advantages of cell cultures suchas good accessibility and control of experimental conditionsThey have been used extensively for the analysis of neuronal
properties and developing neuronal networks [28 51 52]or the study of excitotoxic neuronal damage (eg [53 54])Recent studies also demonstrated that basic morphologicalcharacteristics of astrocytes and the typical glia-synapseorganization are well preserved in organotypic slice cultures(eg [44 45 55 56]) In addition we found a nearly completeoverlap of SR101 staining with the expression of GFP in slice
12 ISRN Neurology
SBFIControl Lesioned
SR101
1
Lesio
n sit
e
SR101 SBFI
2 45
32 4
5
1
StrR
StrP StrP
StrR
3
(a)
ControlScar
Lesioned
1
D-asp
2
D-asp
D-asp
5
4
3Periphery1mM
20 s
Δ Δ
Δ
Δ
Δ
(b)
0102030405060708090
100110
Δ so
dium
tim
e (m
Ms
)
00
02
04
06
08
10
12
0
1
2
3
4
5
6
ns
Control Periphery Scar Control Periphery Scar Control Periphery Scar
Amplitude Slope Decay time
Δ so
dium
(mM
)
120591(s
)lowastlowastlowast
lowastlowastlowastlowastlowastlowast
lowastlowastlowastlowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowast
(c)
Figure 7 D-aspartate-induced astrocyte sodium transients following lesion (a) CA1 area of a control (left) and a lesioned (right) slicepreparation Shown are epifluorescence images of SR101 and SBFI fluorescences Circled areas and numbers indicate the regions of interestanalyzed in the experiment depicted in (b) StrP stratum pyramidale StrR stratum radiatum and scale bar 40 120583m (b) Sodium transientsinduced by pressure application of 1mMD-aspartate (D-asp) for 500ms in the different cells in the control and lesioned slice as indicated in(a) (c) Histograms showing mean values plusmn S E M of the peak amplitude (left) slope (center) and decay time (right) of D-aspartate-inducedsodium transients in astrocytes in control slices (119899 = 161) as well as in cells in the scar region and its periphery (119899 = 111 and 119899 = 26 resp)lowastlowast119875 lt 001 lowastlowastlowast119875 lt 0001 StrP stratum pyramidale StrR stratum radiatum and scale bar 40120583m
cultures obtained from GFPGFAP mice demonstrating thatSR101 is well suited to identify astrocytes in this prepara-tion as described before for acute tissue slices and in vivo[29 30]
Performing a scratch through the CA1 region of organ-otypic slices resulted in the formation of a glial scar along thelesion site as judged by the prominent increase in expressionof GFAP and S100120573 In addition astrocyte cell bodies showed
a significant hypertrophy Furthermore astrocytes in theperiphery of the scar (100ndash350 120583m from the lesion) showedmild to moderate reactivity displaying swollen cell bodiesand only moderate upregulation of GFAP These changesare hallmarks of reactive gliosis observed after tissue injury[10 13 57] Generally the degree of injury and the distance ofthe astrocytes from the site of injury define the degree of theiractivation [11 13]
ISRN Neurology 13
Our study also revealed that strongly reactive astrocytesin the scar region only weakly stain with SR101 The reducedability of strongly activated astrocytes to accumulate SR101indicates that strong glial activation is accompanied by adownregulation of the organic anion transporter responsi-ble for uptake of SR101 This is reminiscent of immatureastrocytes in the early postnatal hippocampus which areSR101-negative [30] and in line with a wealth of experimentalevidence suggesting that reactive gliosis represents a processcomprising a dedifferentiation of astrocytes [17]
42 Changes in Glutamate Transporter Expression followingLesion Glutamate uptake in the hippocampus is mainlyachieved by the glial glutamate transporters GLT-1 andGLAST [1 2] A central factor in many brain pathologiesincluding traumatic brain injury is an increase in extracellu-lar glutamate and excitotoxicity it has been suggested that achange in the expression levels of glial glutamate transportersmight play a critical role in the failure of glutamate clearance[6 58ndash60] Earlier work has found an overall downregulationof protein levels of both GLAST and GLT-1 following astro-cyte activation [22ndash27] indicating that this is causal to theelevation of extracellular glutamate In contrast to this notionother studies reported increased glutamate transport capacityof reactive astrocytes and suggested a protective influence[61 62]
To visualize possible differences in the spatial expressionprofile of GLT-1 and GLAST in strongly reactive astrocytesalong the scar as compared to moderately reactive astro-cytes in its periphery we performed immunohistochemicalstainings These revealed an accumulation of GLT-1 andGLAST immunoreactivity along thick GFAP-positive fiberswhich was particularly pronounced for GLAST Besides theseclusters overall GLAST immunoreactivity seemed unalteredas compared to unlesioned slices whereas GLT-1 immunore-activity seemedweaker and declined with increasing distancefrom the scar Clustering of glutamate transporters has beendescribed by several reports [48 63 64] In developinghippocampal astrocytes cluster formation was preferentiallyfound in branches opposed to synapses and was increasedwith increased neuronal activity indicating that it is neces-sary to copewith synaptic release of glutamate [36] Along thesame lines it was observed that sustained astroglial activationby ciliary neurotrophic factor (CNTF) in the rat striatuminduced a concentration of GLAST and GLT-1 into raftmicrodomains and improved glutamate clearance indicatingthat cluster formation increased the cellular capacity for glu-tamate uptake [61] However another study found glutamate-induced clustering of GLT-1 that induced its endocytosis andintracellular trafficking without changing the total expressionlevels as detected by western blots arguing for a decrease infunctional glial glutamate uptake capacity [49] In the presentstudy antibodies against GLT-1 and GLAST were employedfor immunohistochemistry after permeabilization of theplasma membrane The clusters of GLAST and GLT-1 alongGFAP-positive fibers in the scar region (as GLAST andGLT-1immunoreactivity in general) could thus represent glutamatetransporters in the plasmamembrane or in intracellular com-partments or both Consequently the observed clustering
does not allow a prediction about possible functional changesin glutamate uptake
43 Lesion-Induced Changes in Functional Glutamate UptakeCapacity We probed for the functional activation of gluta-mate transport by application of the transportable agonistD-aspartate which results in an increase in the intracellularsodium concentration of astrocytes [8 9] The amplitudesof D-aspartate-induced sodium transients in astrocytes incontrol organotypic slices were similar to those reportedfrom astrocytes in acute slices obtained in our lab [33]indicating similar cellular glutamate transport capacity inboth preparations
Following a lesion D-aspartate-induced intracellularsodium signalswere clearly dampened in scar cellsMoreoverthe slope of the increase in sodium was significantly reducedand the decay back to baseline was slowed Because D-aspartate was applied at nearly saturating concentrationthe reduction in peak amplitudes suggests a reduction infunctional glutamate uptake capacity in cells close to thelesion which might be mediated by a decrease in the overallnumber of glutamate transporters available at the plasmamembrane In contrast the alteration in the kinetics of theD-aspartate-induced sodium signals can be explained by thecellular hypertrophy that was observed in reactive astrocytesAt an equal transport-mediated sodium influx across themembrane the slope of resulting changes in the sodiumconcentration will be decreased in cells with a larger volume
Furthermore the decay of intracellular sodium transientsback to baseline is mainly governed by the activity of theNa+K+-ATPase as well as by diffusion [8] The slower recov-ery is in line with earlier studies reporting a downregulationof the sodium pump following reactive gliosis [65 66]Moreover sodium is not buffered in the cell and travels solelyby means of diffusion in the cytoplasm Hindered diffusionresulting from increased cytoplasmic protein content orimpaired gap junction coupling which both occur uponastrocytic activation might thus also partially contribute todelayed sodium recovery [12 34 67]
In contrast to scar cells the amplitude of D-aspartate-induced intracellular sodium signals was not significantlyaltered in moderately activated astrocytes in the periphery ofthe lesion indicating that the overall number of functionalglutamate transporters was not significantly altered Becausethese cells showed hypertrophy as well the slowed kinetics ofthe sodium transients are in line with an increased cellularvolume as well as a possible downregulation of Na+K+-ATPase andor a slowed diffusion as argued above
Taken together our results indicate a significant reduc-tion in glutamate uptake capacity in strongly activated astro-cytes in the scar region Thus the prominent clustering ofGLAST and GLT-1 immunoreactivity along GFAP-positivestructures close to the lesion is likely to reflect a loss ofglutamate transporters from the plasma membrane upontransporter internalization as reported by Nakagawa et al[49] The authors of the latter study speculated that large andprolonged increases in glutamate concentrations are neces-sary to induce such cluster formation and endocytosis in thetissue as they would possibly only occur under pathological
14 ISRN Neurology
conditions Indeed ambient glutamate concentrations wereshown to be significantly increased after traumatic braininjury [68ndash70] Along these lines it can be assumed thatin our model system extracellular glutamate concentrationsrose less severely in the periphery of the lesion preventinga comparable clustering and loss of functional glutamatetransport activity in moderately activated astrocytes
5 Conclusions
Our results show complex changes in glutamate transporterexpression and function during astrocyte activation followingmechanical injury They confirm that the degree of astrocyteactivation depends on the distance from the insult and com-prises different endpoints regarding cellular morphology andphysiology Furthermore our results highlight that astrocyteswhich show different grades of reactivity also display discretechanges in glutamate transporter expression and functionWhile immunohistochemistry revealed a prominent clus-tering of GLT-1 and GLAST immunoreactivity in the scarregion our functional assay clearly showed that glutamateuptake capacity is strongly reduced in scar cells while itis largely maintained in moderately activated astrocytes inthe periphery Thus mild-to-moderate astrogliosis in theperiphery of a mechanical lesion does not necessarily seemto be accompanied by a significant change in glial glutamateuptake capacity At the glial scar itself in strongly reactiveastrocytes a clustering of glutamate transporters is observedthat apparently goes along with a severe functional reductionin astroglial glutamate uptake which may contribute toglutamate-mediated excitotoxicity in this region
Conflict of Interests
The authors of the paper declare that they do not haveany direct financial relation with the commercial identitiesmentioned in the paper that might lead to a conflict ofinterests
Acknowledgments
This study was supported by the Deutsche Forschungsge-meinschaft (DFG Ro23274-3) The authors thank SimoneDurry and Claudia Roderigo for expert technical help
References
[1] N C Danbolt ldquoGlutamate uptakerdquo Progress in Neurobiologyvol 65 no 1 pp 1ndash105 2001
[2] A V Tzingounis and J I Wadiche ldquoGlutamate transportersconfining runaway excitation by shaping synaptic transmis-sionrdquo Nature Reviews Neuroscience vol 8 no 12 pp 935ndash9472007
[3] D E Bergles J S Diamond and C E Jahr ldquoClearance ofglutamate inside the synapse and beyondrdquo Current Opinion inNeurobiology vol 9 no 3 pp 293ndash298 1999
[4] C M Anderson and R A Swanson ldquoAstrocyte glutamatetransport review of properties regulation and physiologicalfunctionsrdquo Glia vol 32 no 1 pp 1ndash14 2000
[5] P Marcaggi and D Attwell ldquoRole of glial amino acid trans-porters in synaptic transmission and brain energeticsrdquoGlia vol47 no 3 pp 217ndash225 2004
[6] N J Maragakis and J D Rothstein ldquoGlutamate transportersanimal models to neurologic diseaserdquo Neurobiology of Diseasevol 15 no 3 pp 461ndash473 2004
[7] G Gegelashvili and A Schousboe ldquoCellular distribution andkinetic properties of high-affinity glutamate transportersrdquoBrainResearch Bulletin vol 45 no 3 pp 233ndash238 1998
[8] J W Deitmer and C R Rose ldquoIon changes and signalling inperisynaptic gliardquo Brain Research Reviews vol 63 no 1-2 pp113ndash129 2010
[9] S Kirischuk V Parpura and A Verkhratsky ldquoSodium dynam-ics another key to astroglial excitabilityrdquo Trends in Neuro-sciences vol 35 no 8 pp 497ndash506 2012
[10] M V Sofroniew ldquoMolecular dissection of reactive astrogliosisand glial scar formationrdquo Trends in Neurosciences vol 32 no12 pp 638ndash647 2009
[11] J L Ridet S K Malhotra A Privat and F H Gage ldquoReactiveastrocytes cellular and molecular cues to biological functionrdquoTrends in Neurosciences vol 20 no 12 pp 570ndash577 1997
[12] J Middeldorp and E M Hol ldquoGFAP in health and diseaserdquoProgress in Neurobiology vol 93 no 3 pp 421ndash443 2011
[13] M Pekny and M Nilsson ldquoAstrocyte activation and reactivegliosisrdquo Glia vol 50 no 4 pp 427ndash434 2005
[14] A Bordey S A Lyons J J Hablitz and H SontheimerldquoElectrophysiological characteristics of reactive astrocytes inexperimental cortical dysplasiardquo Journal of Neurophysiologyvol 85 no 4 pp 1719ndash1731 2001
[15] R Jabs G Seifert and C Steinhauser ldquoAstrocytic functionand its alteration in the epileptic brainrdquo Epilepsia vol 49supplement 2 pp 3ndash12 2008
[16] A Buffo I Rite P Tripathi et al ldquoOrigin and progeny ofreactive gliosis a source of multipotent cells in the injuredbrainrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 105 no 9 pp 3581ndash3586 2008
[17] S Robel B Berninger and M Gotz ldquoThe stem cell potential ofglia lessons from reactive gliosisrdquoNature Reviews Neurosciencevol 12 no 2 pp 88ndash104 2011
[18] H Kawano J Kimura-Kuroda Y Komuta et al ldquoRole of thelesion scar in the response to damage and repair of the centralnervous systemrdquo Cell and Tissue Research vol 349 no 1 pp169ndash180 2012
[19] G Yiu and Z He ldquoGlial inhibition of CNS axon regenerationrdquoNature Reviews Neuroscience vol 7 no 8 pp 617ndash627 2006
[20] M T Fitch and J Silver ldquoCNS injury glial scars and inflamma-tion inhibitory extracellular matrices and regeneration failurerdquoExperimental Neurology vol 209 no 2 pp 294ndash301 2008
[21] L Li A Lundkvist D Andersson et al ldquoProtective role ofreactive astrocytes in brain ischemiardquo Journal of Cerebral BloodFlow amp Metabolism vol 28 no 3 pp 468ndash481 2008
[22] F K H van Landeghem T Weiss M Oehmichen and AVonDeimling ldquoDecreased expression of glutamate transportersin astrocytes after human traumatic brain injuryrdquo Journal ofNeurotrauma vol 23 no 10 pp 1518ndash1528 2006
[23] V L Raghavendra Rao M K Baskaya A Dogan J DRothstein and R J Dempsey ldquoTraumatic brain injury down-regulates glial glutamate transporter (GLT- 1 and GLAST)proteins in rat brainrdquo Journal of Neurochemistry vol 70 no 5pp 2020ndash2027 1998
ISRN Neurology 15
[24] J-C Chen H Hsu-Chou J-L Lu et al ldquoDown-regulation ofthe glial glutamate transporter GLT-1 in rat hippocampus andstriatum and its modulation by a group III metabotropic glu-tamate receptor antagonist following transient global forebrainischemiardquoNeuropharmacology vol 49 no 5 pp 703ndash714 2005
[25] S D Ginsberg L J Martin and J D Rothstein ldquoRegional deaf-ferentation down-regulates subtypes of glutamate transporterproteinsrdquo Journal of Neurochemistry vol 65 no 6 pp 2800ndash2803 1995
[26] J E Springer R D Azbill R J Mark J G Begley GWaeg andM P Mattson ldquo4-Hydroxynonenal a lipid peroxidation prod-uct rapidly accumulates following traumatic spinal cord injuryand inhibits glutamate uptakerdquo Journal of Neurochemistry vol68 no 6 pp 2469ndash2476 1997
[27] M B Moretto N S Arteni D Lavinsky et al ldquoHypoxic-ischemic insult decreases glutamate uptake by hippocampalslices from neonatal rats Prevention by guanosinerdquo Experimen-tal Neurology vol 195 no 2 pp 400ndash406 2005
[28] L Stoppini P-A Buchs and D Muller ldquoA simple method fororganotypic cultures of nervous tissuerdquo Journal of NeuroscienceMethods vol 37 no 2 pp 173ndash182 1991
[29] A Nimmerjahn F Kirchhoff J N D Kerr and F HelmchenldquoSulforhodamine 101 as a specific marker of astroglia in theneocortex in vivordquo Nat Methods vol 1 no 1 pp 31ndash37 2004
[30] K W Kafitz S D Meier J Stephan and C R Rose ldquoDevel-opmental profile and properties of sulforhodamine 101-Labeledglial cells in acute brain slices of rat hippocampusrdquo Journal ofNeuroscience Methods vol 169 no 1 pp 84ndash92 2008
[31] C Schnell Y Hagos and S Hulsmann ldquoActive sulforhodamine101 uptake into hippocampal astrocytesrdquo PLoS One vol 7 no 11Article ID e49398 2012
[32] S D Meier Y Kovalchuk and C R Rose ldquoProperties ofthe new fluorescent Na+ indicator CoroNa Green comparisonwith SBFI and confocal Na+ imagingrdquo Journal of NeuroscienceMethods vol 155 no 2 pp 251ndash259 2006
[33] J Langer and C R Rose ldquoSynaptically induced sodium signalsin hippocampal astrocytes in siturdquo Journal of Physiology vol587 part 24 pp 5859ndash5877 2009
[34] J Langer J Stephan M Theis and C R Rose ldquoGap junctionsmediate intercellular spread of sodium between hippocampalastrocytes in siturdquo Glia vol 60 no 2 pp 239ndash252 2012
[35] C R Rose andB R Ransom ldquoIntracellular sodiumhomeostasisin rat hippocampal astrocytesrdquo Journal of Physiology vol 491part 2 no 2 pp 291ndash305 1996
[36] A M Benediktsson G S Marrs J C Tu et al ldquoNeuronalactivity regulates glutamate transporter dynamics in developingastrocytesrdquo Glia vol 60 no 2 pp 175ndash188 2012
[37] M Zhou G P Schools and H K Kimelberg ldquoDevelopment ofGLAST(+) astrocytes andNG2(+) glia in rat hippocampus CA1mature astrocytes are electrophysiologically passiverdquo Journal ofNeurophysiology vol 95 no 1 pp 134ndash143 2006
[38] I Suarez G Bodega and B Fernandez ldquoModulation of glu-tamate transporters (GLAST GLT-1 and EAAC1) in the ratcerebellum following portocaval anastomosisrdquo Brain Researchvol 859 no 2 pp 293ndash302 2000
[39] X Zhu D E Bergles and A Nishiyama ldquoNG2 cells generateboth oligodendrocytes and gray matter astrocytesrdquo Develop-ment vol 135 no 1 pp 145ndash157 2008
[40] B Brunne S ZhaoADerouiche et al ldquoOriginmaturation andastroglial transformation of secondary radial glial cells in thedeveloping dentate gyrusrdquo Glia vol 58 no 13 pp 1553ndash15692010
[41] Y Wu A-Q Zhang and D T Yew ldquoAge related changes ofvarious markers of astrocytes in senescence-accelerated micehippocampusrdquo Neurochemistry International vol 46 no 7 pp565ndash574 2005
[42] S Magavi D Friedmann G Banks A Stolfi and C LoisldquoCoincident generation of pyramidal neurons and protoplasmicastrocytes in neocortical columnsrdquo Journal of Neuroscience vol32 no 14 pp 4762ndash4772 2012
[43] M Olabarria H N Noristani A Verkhratsky and J JRodrıguez ldquoConcomitant astroglial atrophy and astrogliosis ina triple transgenic animal model of Alzheimerrsquos diseaserdquo Gliavol 58 no 7 pp 831ndash838 2010
[44] A M Benediktsson S J Schachtele S H Green and M EDailey ldquoBallistic labeling and dynamic imaging of astrocytesin organotypic hippocampal slice culturesrdquo Journal of Neuro-science Methods vol 141 no 1 pp 41ndash53 2005
[45] I Lushnikova G Skibo D Muller and I Nikonenko ldquoSynapticpotentiation induces increased glial coverage of excitatorysynapses in CA1 hippocampusrdquoHippocampus vol 19 no 8 pp753ndash762 2009
[46] E Raponi F Agenes C Delphin et al ldquoS100B expressiondefines a state in which GFAP-expressing cells lose their neuralstem cell potential and acquire a more mature developmentalstagerdquo Glia vol 55 no 2 pp 165ndash177 2007
[47] C G Schipke C Boucsein C Ohlemeyer F Kirchhoff andH Kettenmann ldquoAstrocyte Ca2+ waves trigger responses inmicroglial cells in brain slicesrdquo The FASEB Journal vol 16 no2 pp 255ndash257 2002
[48] J Zhou and M L Sutherland ldquoGlutamate transporter clusterformation in astrocytic processes regulates glutamate uptakeactivityrdquo Journal of Neuroscience vol 24 no 28 pp 6301ndash63062004
[49] T Nakagawa Y Otsubo Y Yatani H Shirakawa and S KanekoldquoMechanisms of substrate transport-induced clustering of aglial glutamate transporter GLT-1 in astroglial-neuronal cul-turesrdquo European Journal of Neuroscience vol 28 no 9 pp 1719ndash1730 2008
[50] J-Y Chatton P Marquet and P J Magistretti ldquoA quantitativeanalysis of L-glutamate-regulated Na+ dynamics in mousecortical astrocytes Implications for cellular bioenergeticsrdquoEuropean Journal of Neuroscience vol 12 no 11 pp 3843ndash38532000
[51] E Forster S Zhao and M Frotscher ldquoLaminating the hip-pocampusrdquo Nature Reviews Neuroscience vol 7 no 4 pp 259ndash267 2006
[52] B H Gahwiler M Capogna D Debanne R A McKinney andS M Thompson ldquoOrganotypic slice cultures a technique hascome of agerdquo Trends in Neurosciences vol 20 no 10 pp 471ndash477 1997
[53] P E Kunkler and R P Kraig ldquoReactive astrocytosis fromexcitotoxic injury in hippocampal organ culture parallels thatseen in vivordquo Journal of Cerebral Blood Flow and Metabolismvol 17 no 1 pp 26ndash43 1997
[54] I E Holopainen ldquoOrganotypic hippocampal slice cultures amodel system to study basic cellular andmolecularmechanismsof neuronal cell death neuroprotection and synaptic plasticityrdquoNeurochemical Research vol 30 no 12 pp 1521ndash1528 2005
[55] MHaber L Zhou andK KMurai ldquoCooperative astrocyte anddendritic spine dynamics at hippocampal excitatory synapsesrdquoJournal of Neuroscience vol 26 no 35 pp 8881ndash8891 2006
16 ISRN Neurology
[56] H Nishida and S Okabe ldquoDirect astrocytic contacts regulatelocal maturation of dendritic spinesrdquo Journal of Neurosciencevol 27 no 2 pp 331ndash340 2007
[57] A Buffo C Rolando and S Ceruti ldquoAstrocytes in the dam-aged brain molecular and cellular insights into their reactiveresponse and healing potentialrdquoBiochemical Pharmacology vol79 no 2 pp 77ndash89 2010
[58] J-H Yi and A S Hazell ldquoExcitotoxic mechanisms and the roleof astrocytic glutamate transporters in traumatic brain injuryrdquoNeurochemistry International vol 48 no 5 pp 394ndash403 2006
[59] C Werner and K Engelhard ldquoPathophysiology of traumaticbrain injuryrdquo British Journal of Anaesthesia vol 99 no 1 pp4ndash9 2007
[60] G Gegelashvili M B Robinson D Trotti and T RauenldquoRegulation of glutamate transporters in health and diseaserdquoProgress in Brain Research vol 132 pp 267ndash286 2001
[61] C Escartin E Brouillet PGubellini et al ldquoCiliary neurotrophicfactor activates astrocytes redistributes their glutamate trans-porters GLAST and GLT-1 to raft microdomains and improvesglutamate handling in vivordquo Journal of Neuroscience vol 26 no22 pp 5978ndash5989 2006
[62] C Vermeiren M Najimi N Vanhoutte et al ldquoAcute up-regulation of glutamate uptake mediated by mGluR5a in reac-tive astrocytesrdquo Journal of Neurochemistry vol 94 no 2 pp405ndash416 2005
[63] C L Poitry-Yamate L Vutskits and T Rauen ldquoNeuronal-induced and glutamate-dependent activation of glial glutamatetransporter functionrdquo Journal of Neurochemistry vol 82 no 4pp 987ndash997 2002
[64] P M Beart and R D OrsquoShea ldquoTransporters for L-glutamatean update on their molecular pharmacology and pathologicalinvolvementrdquo British Journal of Pharmacology vol 150 no 1pp 5ndash17 2007
[65] F D Lima M A Souza A F Furian et al ldquoNa+K+-ATPaseactivity impairment after experimental traumatic brain injuryrelationship to spatial learning deficits and oxidative stressrdquoBehavioural Brain Research vol 193 no 2 pp 306ndash310 2008
[66] G A Gusarova H E Trejo L A Dada et al ldquoHypoxia leadsto NaK-ATPase downregulation via Ca2+ release-activatedCa2+ channels and AMPK activationrdquo Molecular and CellularBiology vol 31 no 17 pp 3546ndash3556 2011
[67] S S L Chew C S Johnson C R Green and H V Danesh-Meyer ldquoRole of connexin43 in central nervous system injuryrdquoExperimental Neurology vol 225 no 2 pp 250ndash261 2010
[68] P Nilsson L Hillered U Ponten and U Ungerstedt ldquoChangesin cortical extracellular levels of energy-related metabolites andamino acids following concussive brain injury in ratsrdquo Journal ofCerebral Blood Flow and Metabolism vol 10 no 5 pp 631ndash6371990
[69] H Katoh K Sima H Nawashiro K Wada and H ChigasakildquoThe effect of MK-801 on extracellular neuroactive amino acidsin hippocampus after closed head injury followed by hypoxia inratsrdquo Brain Research vol 758 no 1-2 pp 153ndash162 1997
[70] H Koizumi H Fujisawa H Ito T Maekawa X Di and RBullock ldquoEffects of mild hypothermia on cerebral blood flow-independent changes in cortical extracellular levels of aminoacids following contusion trauma in the ratrdquoBrain Research vol747 no 2 pp 304ndash312 1997
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

12 ISRN Neurology
SBFIControl Lesioned
SR101
1
Lesio
n sit
e
SR101 SBFI
2 45
32 4
5
1
StrR
StrP StrP
StrR
3
(a)
ControlScar
Lesioned
1
D-asp
2
D-asp
D-asp
5
4
3Periphery1mM
20 s
Δ Δ
Δ
Δ
Δ
(b)
0102030405060708090
100110
Δ so
dium
tim
e (m
Ms
)
00
02
04
06
08
10
12
0
1
2
3
4
5
6
ns
Control Periphery Scar Control Periphery Scar Control Periphery Scar
Amplitude Slope Decay time
Δ so
dium
(mM
)
120591(s
)lowastlowastlowast
lowastlowastlowastlowastlowastlowast
lowastlowastlowastlowastlowastlowast
lowastlowastlowast
lowastlowastlowast
lowastlowast
(c)
Figure 7 D-aspartate-induced astrocyte sodium transients following lesion (a) CA1 area of a control (left) and a lesioned (right) slicepreparation Shown are epifluorescence images of SR101 and SBFI fluorescences Circled areas and numbers indicate the regions of interestanalyzed in the experiment depicted in (b) StrP stratum pyramidale StrR stratum radiatum and scale bar 40 120583m (b) Sodium transientsinduced by pressure application of 1mMD-aspartate (D-asp) for 500ms in the different cells in the control and lesioned slice as indicated in(a) (c) Histograms showing mean values plusmn S E M of the peak amplitude (left) slope (center) and decay time (right) of D-aspartate-inducedsodium transients in astrocytes in control slices (119899 = 161) as well as in cells in the scar region and its periphery (119899 = 111 and 119899 = 26 resp)lowastlowast119875 lt 001 lowastlowastlowast119875 lt 0001 StrP stratum pyramidale StrR stratum radiatum and scale bar 40120583m
cultures obtained from GFPGFAP mice demonstrating thatSR101 is well suited to identify astrocytes in this prepara-tion as described before for acute tissue slices and in vivo[29 30]
Performing a scratch through the CA1 region of organ-otypic slices resulted in the formation of a glial scar along thelesion site as judged by the prominent increase in expressionof GFAP and S100120573 In addition astrocyte cell bodies showed
a significant hypertrophy Furthermore astrocytes in theperiphery of the scar (100ndash350 120583m from the lesion) showedmild to moderate reactivity displaying swollen cell bodiesand only moderate upregulation of GFAP These changesare hallmarks of reactive gliosis observed after tissue injury[10 13 57] Generally the degree of injury and the distance ofthe astrocytes from the site of injury define the degree of theiractivation [11 13]
ISRN Neurology 13
Our study also revealed that strongly reactive astrocytesin the scar region only weakly stain with SR101 The reducedability of strongly activated astrocytes to accumulate SR101indicates that strong glial activation is accompanied by adownregulation of the organic anion transporter responsi-ble for uptake of SR101 This is reminiscent of immatureastrocytes in the early postnatal hippocampus which areSR101-negative [30] and in line with a wealth of experimentalevidence suggesting that reactive gliosis represents a processcomprising a dedifferentiation of astrocytes [17]
42 Changes in Glutamate Transporter Expression followingLesion Glutamate uptake in the hippocampus is mainlyachieved by the glial glutamate transporters GLT-1 andGLAST [1 2] A central factor in many brain pathologiesincluding traumatic brain injury is an increase in extracellu-lar glutamate and excitotoxicity it has been suggested that achange in the expression levels of glial glutamate transportersmight play a critical role in the failure of glutamate clearance[6 58ndash60] Earlier work has found an overall downregulationof protein levels of both GLAST and GLT-1 following astro-cyte activation [22ndash27] indicating that this is causal to theelevation of extracellular glutamate In contrast to this notionother studies reported increased glutamate transport capacityof reactive astrocytes and suggested a protective influence[61 62]
To visualize possible differences in the spatial expressionprofile of GLT-1 and GLAST in strongly reactive astrocytesalong the scar as compared to moderately reactive astro-cytes in its periphery we performed immunohistochemicalstainings These revealed an accumulation of GLT-1 andGLAST immunoreactivity along thick GFAP-positive fiberswhich was particularly pronounced for GLAST Besides theseclusters overall GLAST immunoreactivity seemed unalteredas compared to unlesioned slices whereas GLT-1 immunore-activity seemedweaker and declined with increasing distancefrom the scar Clustering of glutamate transporters has beendescribed by several reports [48 63 64] In developinghippocampal astrocytes cluster formation was preferentiallyfound in branches opposed to synapses and was increasedwith increased neuronal activity indicating that it is neces-sary to copewith synaptic release of glutamate [36] Along thesame lines it was observed that sustained astroglial activationby ciliary neurotrophic factor (CNTF) in the rat striatuminduced a concentration of GLAST and GLT-1 into raftmicrodomains and improved glutamate clearance indicatingthat cluster formation increased the cellular capacity for glu-tamate uptake [61] However another study found glutamate-induced clustering of GLT-1 that induced its endocytosis andintracellular trafficking without changing the total expressionlevels as detected by western blots arguing for a decrease infunctional glial glutamate uptake capacity [49] In the presentstudy antibodies against GLT-1 and GLAST were employedfor immunohistochemistry after permeabilization of theplasma membrane The clusters of GLAST and GLT-1 alongGFAP-positive fibers in the scar region (as GLAST andGLT-1immunoreactivity in general) could thus represent glutamatetransporters in the plasmamembrane or in intracellular com-partments or both Consequently the observed clustering
does not allow a prediction about possible functional changesin glutamate uptake
43 Lesion-Induced Changes in Functional Glutamate UptakeCapacity We probed for the functional activation of gluta-mate transport by application of the transportable agonistD-aspartate which results in an increase in the intracellularsodium concentration of astrocytes [8 9] The amplitudesof D-aspartate-induced sodium transients in astrocytes incontrol organotypic slices were similar to those reportedfrom astrocytes in acute slices obtained in our lab [33]indicating similar cellular glutamate transport capacity inboth preparations
Following a lesion D-aspartate-induced intracellularsodium signalswere clearly dampened in scar cellsMoreoverthe slope of the increase in sodium was significantly reducedand the decay back to baseline was slowed Because D-aspartate was applied at nearly saturating concentrationthe reduction in peak amplitudes suggests a reduction infunctional glutamate uptake capacity in cells close to thelesion which might be mediated by a decrease in the overallnumber of glutamate transporters available at the plasmamembrane In contrast the alteration in the kinetics of theD-aspartate-induced sodium signals can be explained by thecellular hypertrophy that was observed in reactive astrocytesAt an equal transport-mediated sodium influx across themembrane the slope of resulting changes in the sodiumconcentration will be decreased in cells with a larger volume
Furthermore the decay of intracellular sodium transientsback to baseline is mainly governed by the activity of theNa+K+-ATPase as well as by diffusion [8] The slower recov-ery is in line with earlier studies reporting a downregulationof the sodium pump following reactive gliosis [65 66]Moreover sodium is not buffered in the cell and travels solelyby means of diffusion in the cytoplasm Hindered diffusionresulting from increased cytoplasmic protein content orimpaired gap junction coupling which both occur uponastrocytic activation might thus also partially contribute todelayed sodium recovery [12 34 67]
In contrast to scar cells the amplitude of D-aspartate-induced intracellular sodium signals was not significantlyaltered in moderately activated astrocytes in the periphery ofthe lesion indicating that the overall number of functionalglutamate transporters was not significantly altered Becausethese cells showed hypertrophy as well the slowed kinetics ofthe sodium transients are in line with an increased cellularvolume as well as a possible downregulation of Na+K+-ATPase andor a slowed diffusion as argued above
Taken together our results indicate a significant reduc-tion in glutamate uptake capacity in strongly activated astro-cytes in the scar region Thus the prominent clustering ofGLAST and GLT-1 immunoreactivity along GFAP-positivestructures close to the lesion is likely to reflect a loss ofglutamate transporters from the plasma membrane upontransporter internalization as reported by Nakagawa et al[49] The authors of the latter study speculated that large andprolonged increases in glutamate concentrations are neces-sary to induce such cluster formation and endocytosis in thetissue as they would possibly only occur under pathological
14 ISRN Neurology
conditions Indeed ambient glutamate concentrations wereshown to be significantly increased after traumatic braininjury [68ndash70] Along these lines it can be assumed thatin our model system extracellular glutamate concentrationsrose less severely in the periphery of the lesion preventinga comparable clustering and loss of functional glutamatetransport activity in moderately activated astrocytes
5 Conclusions
Our results show complex changes in glutamate transporterexpression and function during astrocyte activation followingmechanical injury They confirm that the degree of astrocyteactivation depends on the distance from the insult and com-prises different endpoints regarding cellular morphology andphysiology Furthermore our results highlight that astrocyteswhich show different grades of reactivity also display discretechanges in glutamate transporter expression and functionWhile immunohistochemistry revealed a prominent clus-tering of GLT-1 and GLAST immunoreactivity in the scarregion our functional assay clearly showed that glutamateuptake capacity is strongly reduced in scar cells while itis largely maintained in moderately activated astrocytes inthe periphery Thus mild-to-moderate astrogliosis in theperiphery of a mechanical lesion does not necessarily seemto be accompanied by a significant change in glial glutamateuptake capacity At the glial scar itself in strongly reactiveastrocytes a clustering of glutamate transporters is observedthat apparently goes along with a severe functional reductionin astroglial glutamate uptake which may contribute toglutamate-mediated excitotoxicity in this region
Conflict of Interests
The authors of the paper declare that they do not haveany direct financial relation with the commercial identitiesmentioned in the paper that might lead to a conflict ofinterests
Acknowledgments
This study was supported by the Deutsche Forschungsge-meinschaft (DFG Ro23274-3) The authors thank SimoneDurry and Claudia Roderigo for expert technical help
References
[1] N C Danbolt ldquoGlutamate uptakerdquo Progress in Neurobiologyvol 65 no 1 pp 1ndash105 2001
[2] A V Tzingounis and J I Wadiche ldquoGlutamate transportersconfining runaway excitation by shaping synaptic transmis-sionrdquo Nature Reviews Neuroscience vol 8 no 12 pp 935ndash9472007
[3] D E Bergles J S Diamond and C E Jahr ldquoClearance ofglutamate inside the synapse and beyondrdquo Current Opinion inNeurobiology vol 9 no 3 pp 293ndash298 1999
[4] C M Anderson and R A Swanson ldquoAstrocyte glutamatetransport review of properties regulation and physiologicalfunctionsrdquo Glia vol 32 no 1 pp 1ndash14 2000
[5] P Marcaggi and D Attwell ldquoRole of glial amino acid trans-porters in synaptic transmission and brain energeticsrdquoGlia vol47 no 3 pp 217ndash225 2004
[6] N J Maragakis and J D Rothstein ldquoGlutamate transportersanimal models to neurologic diseaserdquo Neurobiology of Diseasevol 15 no 3 pp 461ndash473 2004
[7] G Gegelashvili and A Schousboe ldquoCellular distribution andkinetic properties of high-affinity glutamate transportersrdquoBrainResearch Bulletin vol 45 no 3 pp 233ndash238 1998
[8] J W Deitmer and C R Rose ldquoIon changes and signalling inperisynaptic gliardquo Brain Research Reviews vol 63 no 1-2 pp113ndash129 2010
[9] S Kirischuk V Parpura and A Verkhratsky ldquoSodium dynam-ics another key to astroglial excitabilityrdquo Trends in Neuro-sciences vol 35 no 8 pp 497ndash506 2012
[10] M V Sofroniew ldquoMolecular dissection of reactive astrogliosisand glial scar formationrdquo Trends in Neurosciences vol 32 no12 pp 638ndash647 2009
[11] J L Ridet S K Malhotra A Privat and F H Gage ldquoReactiveastrocytes cellular and molecular cues to biological functionrdquoTrends in Neurosciences vol 20 no 12 pp 570ndash577 1997
[12] J Middeldorp and E M Hol ldquoGFAP in health and diseaserdquoProgress in Neurobiology vol 93 no 3 pp 421ndash443 2011
[13] M Pekny and M Nilsson ldquoAstrocyte activation and reactivegliosisrdquo Glia vol 50 no 4 pp 427ndash434 2005
[14] A Bordey S A Lyons J J Hablitz and H SontheimerldquoElectrophysiological characteristics of reactive astrocytes inexperimental cortical dysplasiardquo Journal of Neurophysiologyvol 85 no 4 pp 1719ndash1731 2001
[15] R Jabs G Seifert and C Steinhauser ldquoAstrocytic functionand its alteration in the epileptic brainrdquo Epilepsia vol 49supplement 2 pp 3ndash12 2008
[16] A Buffo I Rite P Tripathi et al ldquoOrigin and progeny ofreactive gliosis a source of multipotent cells in the injuredbrainrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 105 no 9 pp 3581ndash3586 2008
[17] S Robel B Berninger and M Gotz ldquoThe stem cell potential ofglia lessons from reactive gliosisrdquoNature Reviews Neurosciencevol 12 no 2 pp 88ndash104 2011
[18] H Kawano J Kimura-Kuroda Y Komuta et al ldquoRole of thelesion scar in the response to damage and repair of the centralnervous systemrdquo Cell and Tissue Research vol 349 no 1 pp169ndash180 2012
[19] G Yiu and Z He ldquoGlial inhibition of CNS axon regenerationrdquoNature Reviews Neuroscience vol 7 no 8 pp 617ndash627 2006
[20] M T Fitch and J Silver ldquoCNS injury glial scars and inflamma-tion inhibitory extracellular matrices and regeneration failurerdquoExperimental Neurology vol 209 no 2 pp 294ndash301 2008
[21] L Li A Lundkvist D Andersson et al ldquoProtective role ofreactive astrocytes in brain ischemiardquo Journal of Cerebral BloodFlow amp Metabolism vol 28 no 3 pp 468ndash481 2008
[22] F K H van Landeghem T Weiss M Oehmichen and AVonDeimling ldquoDecreased expression of glutamate transportersin astrocytes after human traumatic brain injuryrdquo Journal ofNeurotrauma vol 23 no 10 pp 1518ndash1528 2006
[23] V L Raghavendra Rao M K Baskaya A Dogan J DRothstein and R J Dempsey ldquoTraumatic brain injury down-regulates glial glutamate transporter (GLT- 1 and GLAST)proteins in rat brainrdquo Journal of Neurochemistry vol 70 no 5pp 2020ndash2027 1998
ISRN Neurology 15
[24] J-C Chen H Hsu-Chou J-L Lu et al ldquoDown-regulation ofthe glial glutamate transporter GLT-1 in rat hippocampus andstriatum and its modulation by a group III metabotropic glu-tamate receptor antagonist following transient global forebrainischemiardquoNeuropharmacology vol 49 no 5 pp 703ndash714 2005
[25] S D Ginsberg L J Martin and J D Rothstein ldquoRegional deaf-ferentation down-regulates subtypes of glutamate transporterproteinsrdquo Journal of Neurochemistry vol 65 no 6 pp 2800ndash2803 1995
[26] J E Springer R D Azbill R J Mark J G Begley GWaeg andM P Mattson ldquo4-Hydroxynonenal a lipid peroxidation prod-uct rapidly accumulates following traumatic spinal cord injuryand inhibits glutamate uptakerdquo Journal of Neurochemistry vol68 no 6 pp 2469ndash2476 1997
[27] M B Moretto N S Arteni D Lavinsky et al ldquoHypoxic-ischemic insult decreases glutamate uptake by hippocampalslices from neonatal rats Prevention by guanosinerdquo Experimen-tal Neurology vol 195 no 2 pp 400ndash406 2005
[28] L Stoppini P-A Buchs and D Muller ldquoA simple method fororganotypic cultures of nervous tissuerdquo Journal of NeuroscienceMethods vol 37 no 2 pp 173ndash182 1991
[29] A Nimmerjahn F Kirchhoff J N D Kerr and F HelmchenldquoSulforhodamine 101 as a specific marker of astroglia in theneocortex in vivordquo Nat Methods vol 1 no 1 pp 31ndash37 2004
[30] K W Kafitz S D Meier J Stephan and C R Rose ldquoDevel-opmental profile and properties of sulforhodamine 101-Labeledglial cells in acute brain slices of rat hippocampusrdquo Journal ofNeuroscience Methods vol 169 no 1 pp 84ndash92 2008
[31] C Schnell Y Hagos and S Hulsmann ldquoActive sulforhodamine101 uptake into hippocampal astrocytesrdquo PLoS One vol 7 no 11Article ID e49398 2012
[32] S D Meier Y Kovalchuk and C R Rose ldquoProperties ofthe new fluorescent Na+ indicator CoroNa Green comparisonwith SBFI and confocal Na+ imagingrdquo Journal of NeuroscienceMethods vol 155 no 2 pp 251ndash259 2006
[33] J Langer and C R Rose ldquoSynaptically induced sodium signalsin hippocampal astrocytes in siturdquo Journal of Physiology vol587 part 24 pp 5859ndash5877 2009
[34] J Langer J Stephan M Theis and C R Rose ldquoGap junctionsmediate intercellular spread of sodium between hippocampalastrocytes in siturdquo Glia vol 60 no 2 pp 239ndash252 2012
[35] C R Rose andB R Ransom ldquoIntracellular sodiumhomeostasisin rat hippocampal astrocytesrdquo Journal of Physiology vol 491part 2 no 2 pp 291ndash305 1996
[36] A M Benediktsson G S Marrs J C Tu et al ldquoNeuronalactivity regulates glutamate transporter dynamics in developingastrocytesrdquo Glia vol 60 no 2 pp 175ndash188 2012
[37] M Zhou G P Schools and H K Kimelberg ldquoDevelopment ofGLAST(+) astrocytes andNG2(+) glia in rat hippocampus CA1mature astrocytes are electrophysiologically passiverdquo Journal ofNeurophysiology vol 95 no 1 pp 134ndash143 2006
[38] I Suarez G Bodega and B Fernandez ldquoModulation of glu-tamate transporters (GLAST GLT-1 and EAAC1) in the ratcerebellum following portocaval anastomosisrdquo Brain Researchvol 859 no 2 pp 293ndash302 2000
[39] X Zhu D E Bergles and A Nishiyama ldquoNG2 cells generateboth oligodendrocytes and gray matter astrocytesrdquo Develop-ment vol 135 no 1 pp 145ndash157 2008
[40] B Brunne S ZhaoADerouiche et al ldquoOriginmaturation andastroglial transformation of secondary radial glial cells in thedeveloping dentate gyrusrdquo Glia vol 58 no 13 pp 1553ndash15692010
[41] Y Wu A-Q Zhang and D T Yew ldquoAge related changes ofvarious markers of astrocytes in senescence-accelerated micehippocampusrdquo Neurochemistry International vol 46 no 7 pp565ndash574 2005
[42] S Magavi D Friedmann G Banks A Stolfi and C LoisldquoCoincident generation of pyramidal neurons and protoplasmicastrocytes in neocortical columnsrdquo Journal of Neuroscience vol32 no 14 pp 4762ndash4772 2012
[43] M Olabarria H N Noristani A Verkhratsky and J JRodrıguez ldquoConcomitant astroglial atrophy and astrogliosis ina triple transgenic animal model of Alzheimerrsquos diseaserdquo Gliavol 58 no 7 pp 831ndash838 2010
[44] A M Benediktsson S J Schachtele S H Green and M EDailey ldquoBallistic labeling and dynamic imaging of astrocytesin organotypic hippocampal slice culturesrdquo Journal of Neuro-science Methods vol 141 no 1 pp 41ndash53 2005
[45] I Lushnikova G Skibo D Muller and I Nikonenko ldquoSynapticpotentiation induces increased glial coverage of excitatorysynapses in CA1 hippocampusrdquoHippocampus vol 19 no 8 pp753ndash762 2009
[46] E Raponi F Agenes C Delphin et al ldquoS100B expressiondefines a state in which GFAP-expressing cells lose their neuralstem cell potential and acquire a more mature developmentalstagerdquo Glia vol 55 no 2 pp 165ndash177 2007
[47] C G Schipke C Boucsein C Ohlemeyer F Kirchhoff andH Kettenmann ldquoAstrocyte Ca2+ waves trigger responses inmicroglial cells in brain slicesrdquo The FASEB Journal vol 16 no2 pp 255ndash257 2002
[48] J Zhou and M L Sutherland ldquoGlutamate transporter clusterformation in astrocytic processes regulates glutamate uptakeactivityrdquo Journal of Neuroscience vol 24 no 28 pp 6301ndash63062004
[49] T Nakagawa Y Otsubo Y Yatani H Shirakawa and S KanekoldquoMechanisms of substrate transport-induced clustering of aglial glutamate transporter GLT-1 in astroglial-neuronal cul-turesrdquo European Journal of Neuroscience vol 28 no 9 pp 1719ndash1730 2008
[50] J-Y Chatton P Marquet and P J Magistretti ldquoA quantitativeanalysis of L-glutamate-regulated Na+ dynamics in mousecortical astrocytes Implications for cellular bioenergeticsrdquoEuropean Journal of Neuroscience vol 12 no 11 pp 3843ndash38532000
[51] E Forster S Zhao and M Frotscher ldquoLaminating the hip-pocampusrdquo Nature Reviews Neuroscience vol 7 no 4 pp 259ndash267 2006
[52] B H Gahwiler M Capogna D Debanne R A McKinney andS M Thompson ldquoOrganotypic slice cultures a technique hascome of agerdquo Trends in Neurosciences vol 20 no 10 pp 471ndash477 1997
[53] P E Kunkler and R P Kraig ldquoReactive astrocytosis fromexcitotoxic injury in hippocampal organ culture parallels thatseen in vivordquo Journal of Cerebral Blood Flow and Metabolismvol 17 no 1 pp 26ndash43 1997
[54] I E Holopainen ldquoOrganotypic hippocampal slice cultures amodel system to study basic cellular andmolecularmechanismsof neuronal cell death neuroprotection and synaptic plasticityrdquoNeurochemical Research vol 30 no 12 pp 1521ndash1528 2005
[55] MHaber L Zhou andK KMurai ldquoCooperative astrocyte anddendritic spine dynamics at hippocampal excitatory synapsesrdquoJournal of Neuroscience vol 26 no 35 pp 8881ndash8891 2006
16 ISRN Neurology
[56] H Nishida and S Okabe ldquoDirect astrocytic contacts regulatelocal maturation of dendritic spinesrdquo Journal of Neurosciencevol 27 no 2 pp 331ndash340 2007
[57] A Buffo C Rolando and S Ceruti ldquoAstrocytes in the dam-aged brain molecular and cellular insights into their reactiveresponse and healing potentialrdquoBiochemical Pharmacology vol79 no 2 pp 77ndash89 2010
[58] J-H Yi and A S Hazell ldquoExcitotoxic mechanisms and the roleof astrocytic glutamate transporters in traumatic brain injuryrdquoNeurochemistry International vol 48 no 5 pp 394ndash403 2006
[59] C Werner and K Engelhard ldquoPathophysiology of traumaticbrain injuryrdquo British Journal of Anaesthesia vol 99 no 1 pp4ndash9 2007
[60] G Gegelashvili M B Robinson D Trotti and T RauenldquoRegulation of glutamate transporters in health and diseaserdquoProgress in Brain Research vol 132 pp 267ndash286 2001
[61] C Escartin E Brouillet PGubellini et al ldquoCiliary neurotrophicfactor activates astrocytes redistributes their glutamate trans-porters GLAST and GLT-1 to raft microdomains and improvesglutamate handling in vivordquo Journal of Neuroscience vol 26 no22 pp 5978ndash5989 2006
[62] C Vermeiren M Najimi N Vanhoutte et al ldquoAcute up-regulation of glutamate uptake mediated by mGluR5a in reac-tive astrocytesrdquo Journal of Neurochemistry vol 94 no 2 pp405ndash416 2005
[63] C L Poitry-Yamate L Vutskits and T Rauen ldquoNeuronal-induced and glutamate-dependent activation of glial glutamatetransporter functionrdquo Journal of Neurochemistry vol 82 no 4pp 987ndash997 2002
[64] P M Beart and R D OrsquoShea ldquoTransporters for L-glutamatean update on their molecular pharmacology and pathologicalinvolvementrdquo British Journal of Pharmacology vol 150 no 1pp 5ndash17 2007
[65] F D Lima M A Souza A F Furian et al ldquoNa+K+-ATPaseactivity impairment after experimental traumatic brain injuryrelationship to spatial learning deficits and oxidative stressrdquoBehavioural Brain Research vol 193 no 2 pp 306ndash310 2008
[66] G A Gusarova H E Trejo L A Dada et al ldquoHypoxia leadsto NaK-ATPase downregulation via Ca2+ release-activatedCa2+ channels and AMPK activationrdquo Molecular and CellularBiology vol 31 no 17 pp 3546ndash3556 2011
[67] S S L Chew C S Johnson C R Green and H V Danesh-Meyer ldquoRole of connexin43 in central nervous system injuryrdquoExperimental Neurology vol 225 no 2 pp 250ndash261 2010
[68] P Nilsson L Hillered U Ponten and U Ungerstedt ldquoChangesin cortical extracellular levels of energy-related metabolites andamino acids following concussive brain injury in ratsrdquo Journal ofCerebral Blood Flow and Metabolism vol 10 no 5 pp 631ndash6371990
[69] H Katoh K Sima H Nawashiro K Wada and H ChigasakildquoThe effect of MK-801 on extracellular neuroactive amino acidsin hippocampus after closed head injury followed by hypoxia inratsrdquo Brain Research vol 758 no 1-2 pp 153ndash162 1997
[70] H Koizumi H Fujisawa H Ito T Maekawa X Di and RBullock ldquoEffects of mild hypothermia on cerebral blood flow-independent changes in cortical extracellular levels of aminoacids following contusion trauma in the ratrdquoBrain Research vol747 no 2 pp 304ndash312 1997
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

ISRN Neurology 13
Our study also revealed that strongly reactive astrocytesin the scar region only weakly stain with SR101 The reducedability of strongly activated astrocytes to accumulate SR101indicates that strong glial activation is accompanied by adownregulation of the organic anion transporter responsi-ble for uptake of SR101 This is reminiscent of immatureastrocytes in the early postnatal hippocampus which areSR101-negative [30] and in line with a wealth of experimentalevidence suggesting that reactive gliosis represents a processcomprising a dedifferentiation of astrocytes [17]
42 Changes in Glutamate Transporter Expression followingLesion Glutamate uptake in the hippocampus is mainlyachieved by the glial glutamate transporters GLT-1 andGLAST [1 2] A central factor in many brain pathologiesincluding traumatic brain injury is an increase in extracellu-lar glutamate and excitotoxicity it has been suggested that achange in the expression levels of glial glutamate transportersmight play a critical role in the failure of glutamate clearance[6 58ndash60] Earlier work has found an overall downregulationof protein levels of both GLAST and GLT-1 following astro-cyte activation [22ndash27] indicating that this is causal to theelevation of extracellular glutamate In contrast to this notionother studies reported increased glutamate transport capacityof reactive astrocytes and suggested a protective influence[61 62]
To visualize possible differences in the spatial expressionprofile of GLT-1 and GLAST in strongly reactive astrocytesalong the scar as compared to moderately reactive astro-cytes in its periphery we performed immunohistochemicalstainings These revealed an accumulation of GLT-1 andGLAST immunoreactivity along thick GFAP-positive fiberswhich was particularly pronounced for GLAST Besides theseclusters overall GLAST immunoreactivity seemed unalteredas compared to unlesioned slices whereas GLT-1 immunore-activity seemedweaker and declined with increasing distancefrom the scar Clustering of glutamate transporters has beendescribed by several reports [48 63 64] In developinghippocampal astrocytes cluster formation was preferentiallyfound in branches opposed to synapses and was increasedwith increased neuronal activity indicating that it is neces-sary to copewith synaptic release of glutamate [36] Along thesame lines it was observed that sustained astroglial activationby ciliary neurotrophic factor (CNTF) in the rat striatuminduced a concentration of GLAST and GLT-1 into raftmicrodomains and improved glutamate clearance indicatingthat cluster formation increased the cellular capacity for glu-tamate uptake [61] However another study found glutamate-induced clustering of GLT-1 that induced its endocytosis andintracellular trafficking without changing the total expressionlevels as detected by western blots arguing for a decrease infunctional glial glutamate uptake capacity [49] In the presentstudy antibodies against GLT-1 and GLAST were employedfor immunohistochemistry after permeabilization of theplasma membrane The clusters of GLAST and GLT-1 alongGFAP-positive fibers in the scar region (as GLAST andGLT-1immunoreactivity in general) could thus represent glutamatetransporters in the plasmamembrane or in intracellular com-partments or both Consequently the observed clustering
does not allow a prediction about possible functional changesin glutamate uptake
43 Lesion-Induced Changes in Functional Glutamate UptakeCapacity We probed for the functional activation of gluta-mate transport by application of the transportable agonistD-aspartate which results in an increase in the intracellularsodium concentration of astrocytes [8 9] The amplitudesof D-aspartate-induced sodium transients in astrocytes incontrol organotypic slices were similar to those reportedfrom astrocytes in acute slices obtained in our lab [33]indicating similar cellular glutamate transport capacity inboth preparations
Following a lesion D-aspartate-induced intracellularsodium signalswere clearly dampened in scar cellsMoreoverthe slope of the increase in sodium was significantly reducedand the decay back to baseline was slowed Because D-aspartate was applied at nearly saturating concentrationthe reduction in peak amplitudes suggests a reduction infunctional glutamate uptake capacity in cells close to thelesion which might be mediated by a decrease in the overallnumber of glutamate transporters available at the plasmamembrane In contrast the alteration in the kinetics of theD-aspartate-induced sodium signals can be explained by thecellular hypertrophy that was observed in reactive astrocytesAt an equal transport-mediated sodium influx across themembrane the slope of resulting changes in the sodiumconcentration will be decreased in cells with a larger volume
Furthermore the decay of intracellular sodium transientsback to baseline is mainly governed by the activity of theNa+K+-ATPase as well as by diffusion [8] The slower recov-ery is in line with earlier studies reporting a downregulationof the sodium pump following reactive gliosis [65 66]Moreover sodium is not buffered in the cell and travels solelyby means of diffusion in the cytoplasm Hindered diffusionresulting from increased cytoplasmic protein content orimpaired gap junction coupling which both occur uponastrocytic activation might thus also partially contribute todelayed sodium recovery [12 34 67]
In contrast to scar cells the amplitude of D-aspartate-induced intracellular sodium signals was not significantlyaltered in moderately activated astrocytes in the periphery ofthe lesion indicating that the overall number of functionalglutamate transporters was not significantly altered Becausethese cells showed hypertrophy as well the slowed kinetics ofthe sodium transients are in line with an increased cellularvolume as well as a possible downregulation of Na+K+-ATPase andor a slowed diffusion as argued above
Taken together our results indicate a significant reduc-tion in glutamate uptake capacity in strongly activated astro-cytes in the scar region Thus the prominent clustering ofGLAST and GLT-1 immunoreactivity along GFAP-positivestructures close to the lesion is likely to reflect a loss ofglutamate transporters from the plasma membrane upontransporter internalization as reported by Nakagawa et al[49] The authors of the latter study speculated that large andprolonged increases in glutamate concentrations are neces-sary to induce such cluster formation and endocytosis in thetissue as they would possibly only occur under pathological
14 ISRN Neurology
conditions Indeed ambient glutamate concentrations wereshown to be significantly increased after traumatic braininjury [68ndash70] Along these lines it can be assumed thatin our model system extracellular glutamate concentrationsrose less severely in the periphery of the lesion preventinga comparable clustering and loss of functional glutamatetransport activity in moderately activated astrocytes
5 Conclusions
Our results show complex changes in glutamate transporterexpression and function during astrocyte activation followingmechanical injury They confirm that the degree of astrocyteactivation depends on the distance from the insult and com-prises different endpoints regarding cellular morphology andphysiology Furthermore our results highlight that astrocyteswhich show different grades of reactivity also display discretechanges in glutamate transporter expression and functionWhile immunohistochemistry revealed a prominent clus-tering of GLT-1 and GLAST immunoreactivity in the scarregion our functional assay clearly showed that glutamateuptake capacity is strongly reduced in scar cells while itis largely maintained in moderately activated astrocytes inthe periphery Thus mild-to-moderate astrogliosis in theperiphery of a mechanical lesion does not necessarily seemto be accompanied by a significant change in glial glutamateuptake capacity At the glial scar itself in strongly reactiveastrocytes a clustering of glutamate transporters is observedthat apparently goes along with a severe functional reductionin astroglial glutamate uptake which may contribute toglutamate-mediated excitotoxicity in this region
Conflict of Interests
The authors of the paper declare that they do not haveany direct financial relation with the commercial identitiesmentioned in the paper that might lead to a conflict ofinterests
Acknowledgments
This study was supported by the Deutsche Forschungsge-meinschaft (DFG Ro23274-3) The authors thank SimoneDurry and Claudia Roderigo for expert technical help
References
[1] N C Danbolt ldquoGlutamate uptakerdquo Progress in Neurobiologyvol 65 no 1 pp 1ndash105 2001
[2] A V Tzingounis and J I Wadiche ldquoGlutamate transportersconfining runaway excitation by shaping synaptic transmis-sionrdquo Nature Reviews Neuroscience vol 8 no 12 pp 935ndash9472007
[3] D E Bergles J S Diamond and C E Jahr ldquoClearance ofglutamate inside the synapse and beyondrdquo Current Opinion inNeurobiology vol 9 no 3 pp 293ndash298 1999
[4] C M Anderson and R A Swanson ldquoAstrocyte glutamatetransport review of properties regulation and physiologicalfunctionsrdquo Glia vol 32 no 1 pp 1ndash14 2000
[5] P Marcaggi and D Attwell ldquoRole of glial amino acid trans-porters in synaptic transmission and brain energeticsrdquoGlia vol47 no 3 pp 217ndash225 2004
[6] N J Maragakis and J D Rothstein ldquoGlutamate transportersanimal models to neurologic diseaserdquo Neurobiology of Diseasevol 15 no 3 pp 461ndash473 2004
[7] G Gegelashvili and A Schousboe ldquoCellular distribution andkinetic properties of high-affinity glutamate transportersrdquoBrainResearch Bulletin vol 45 no 3 pp 233ndash238 1998
[8] J W Deitmer and C R Rose ldquoIon changes and signalling inperisynaptic gliardquo Brain Research Reviews vol 63 no 1-2 pp113ndash129 2010
[9] S Kirischuk V Parpura and A Verkhratsky ldquoSodium dynam-ics another key to astroglial excitabilityrdquo Trends in Neuro-sciences vol 35 no 8 pp 497ndash506 2012
[10] M V Sofroniew ldquoMolecular dissection of reactive astrogliosisand glial scar formationrdquo Trends in Neurosciences vol 32 no12 pp 638ndash647 2009
[11] J L Ridet S K Malhotra A Privat and F H Gage ldquoReactiveastrocytes cellular and molecular cues to biological functionrdquoTrends in Neurosciences vol 20 no 12 pp 570ndash577 1997
[12] J Middeldorp and E M Hol ldquoGFAP in health and diseaserdquoProgress in Neurobiology vol 93 no 3 pp 421ndash443 2011
[13] M Pekny and M Nilsson ldquoAstrocyte activation and reactivegliosisrdquo Glia vol 50 no 4 pp 427ndash434 2005
[14] A Bordey S A Lyons J J Hablitz and H SontheimerldquoElectrophysiological characteristics of reactive astrocytes inexperimental cortical dysplasiardquo Journal of Neurophysiologyvol 85 no 4 pp 1719ndash1731 2001
[15] R Jabs G Seifert and C Steinhauser ldquoAstrocytic functionand its alteration in the epileptic brainrdquo Epilepsia vol 49supplement 2 pp 3ndash12 2008
[16] A Buffo I Rite P Tripathi et al ldquoOrigin and progeny ofreactive gliosis a source of multipotent cells in the injuredbrainrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 105 no 9 pp 3581ndash3586 2008
[17] S Robel B Berninger and M Gotz ldquoThe stem cell potential ofglia lessons from reactive gliosisrdquoNature Reviews Neurosciencevol 12 no 2 pp 88ndash104 2011
[18] H Kawano J Kimura-Kuroda Y Komuta et al ldquoRole of thelesion scar in the response to damage and repair of the centralnervous systemrdquo Cell and Tissue Research vol 349 no 1 pp169ndash180 2012
[19] G Yiu and Z He ldquoGlial inhibition of CNS axon regenerationrdquoNature Reviews Neuroscience vol 7 no 8 pp 617ndash627 2006
[20] M T Fitch and J Silver ldquoCNS injury glial scars and inflamma-tion inhibitory extracellular matrices and regeneration failurerdquoExperimental Neurology vol 209 no 2 pp 294ndash301 2008
[21] L Li A Lundkvist D Andersson et al ldquoProtective role ofreactive astrocytes in brain ischemiardquo Journal of Cerebral BloodFlow amp Metabolism vol 28 no 3 pp 468ndash481 2008
[22] F K H van Landeghem T Weiss M Oehmichen and AVonDeimling ldquoDecreased expression of glutamate transportersin astrocytes after human traumatic brain injuryrdquo Journal ofNeurotrauma vol 23 no 10 pp 1518ndash1528 2006
[23] V L Raghavendra Rao M K Baskaya A Dogan J DRothstein and R J Dempsey ldquoTraumatic brain injury down-regulates glial glutamate transporter (GLT- 1 and GLAST)proteins in rat brainrdquo Journal of Neurochemistry vol 70 no 5pp 2020ndash2027 1998
ISRN Neurology 15
[24] J-C Chen H Hsu-Chou J-L Lu et al ldquoDown-regulation ofthe glial glutamate transporter GLT-1 in rat hippocampus andstriatum and its modulation by a group III metabotropic glu-tamate receptor antagonist following transient global forebrainischemiardquoNeuropharmacology vol 49 no 5 pp 703ndash714 2005
[25] S D Ginsberg L J Martin and J D Rothstein ldquoRegional deaf-ferentation down-regulates subtypes of glutamate transporterproteinsrdquo Journal of Neurochemistry vol 65 no 6 pp 2800ndash2803 1995
[26] J E Springer R D Azbill R J Mark J G Begley GWaeg andM P Mattson ldquo4-Hydroxynonenal a lipid peroxidation prod-uct rapidly accumulates following traumatic spinal cord injuryand inhibits glutamate uptakerdquo Journal of Neurochemistry vol68 no 6 pp 2469ndash2476 1997
[27] M B Moretto N S Arteni D Lavinsky et al ldquoHypoxic-ischemic insult decreases glutamate uptake by hippocampalslices from neonatal rats Prevention by guanosinerdquo Experimen-tal Neurology vol 195 no 2 pp 400ndash406 2005
[28] L Stoppini P-A Buchs and D Muller ldquoA simple method fororganotypic cultures of nervous tissuerdquo Journal of NeuroscienceMethods vol 37 no 2 pp 173ndash182 1991
[29] A Nimmerjahn F Kirchhoff J N D Kerr and F HelmchenldquoSulforhodamine 101 as a specific marker of astroglia in theneocortex in vivordquo Nat Methods vol 1 no 1 pp 31ndash37 2004
[30] K W Kafitz S D Meier J Stephan and C R Rose ldquoDevel-opmental profile and properties of sulforhodamine 101-Labeledglial cells in acute brain slices of rat hippocampusrdquo Journal ofNeuroscience Methods vol 169 no 1 pp 84ndash92 2008
[31] C Schnell Y Hagos and S Hulsmann ldquoActive sulforhodamine101 uptake into hippocampal astrocytesrdquo PLoS One vol 7 no 11Article ID e49398 2012
[32] S D Meier Y Kovalchuk and C R Rose ldquoProperties ofthe new fluorescent Na+ indicator CoroNa Green comparisonwith SBFI and confocal Na+ imagingrdquo Journal of NeuroscienceMethods vol 155 no 2 pp 251ndash259 2006
[33] J Langer and C R Rose ldquoSynaptically induced sodium signalsin hippocampal astrocytes in siturdquo Journal of Physiology vol587 part 24 pp 5859ndash5877 2009
[34] J Langer J Stephan M Theis and C R Rose ldquoGap junctionsmediate intercellular spread of sodium between hippocampalastrocytes in siturdquo Glia vol 60 no 2 pp 239ndash252 2012
[35] C R Rose andB R Ransom ldquoIntracellular sodiumhomeostasisin rat hippocampal astrocytesrdquo Journal of Physiology vol 491part 2 no 2 pp 291ndash305 1996
[36] A M Benediktsson G S Marrs J C Tu et al ldquoNeuronalactivity regulates glutamate transporter dynamics in developingastrocytesrdquo Glia vol 60 no 2 pp 175ndash188 2012
[37] M Zhou G P Schools and H K Kimelberg ldquoDevelopment ofGLAST(+) astrocytes andNG2(+) glia in rat hippocampus CA1mature astrocytes are electrophysiologically passiverdquo Journal ofNeurophysiology vol 95 no 1 pp 134ndash143 2006
[38] I Suarez G Bodega and B Fernandez ldquoModulation of glu-tamate transporters (GLAST GLT-1 and EAAC1) in the ratcerebellum following portocaval anastomosisrdquo Brain Researchvol 859 no 2 pp 293ndash302 2000
[39] X Zhu D E Bergles and A Nishiyama ldquoNG2 cells generateboth oligodendrocytes and gray matter astrocytesrdquo Develop-ment vol 135 no 1 pp 145ndash157 2008
[40] B Brunne S ZhaoADerouiche et al ldquoOriginmaturation andastroglial transformation of secondary radial glial cells in thedeveloping dentate gyrusrdquo Glia vol 58 no 13 pp 1553ndash15692010
[41] Y Wu A-Q Zhang and D T Yew ldquoAge related changes ofvarious markers of astrocytes in senescence-accelerated micehippocampusrdquo Neurochemistry International vol 46 no 7 pp565ndash574 2005
[42] S Magavi D Friedmann G Banks A Stolfi and C LoisldquoCoincident generation of pyramidal neurons and protoplasmicastrocytes in neocortical columnsrdquo Journal of Neuroscience vol32 no 14 pp 4762ndash4772 2012
[43] M Olabarria H N Noristani A Verkhratsky and J JRodrıguez ldquoConcomitant astroglial atrophy and astrogliosis ina triple transgenic animal model of Alzheimerrsquos diseaserdquo Gliavol 58 no 7 pp 831ndash838 2010
[44] A M Benediktsson S J Schachtele S H Green and M EDailey ldquoBallistic labeling and dynamic imaging of astrocytesin organotypic hippocampal slice culturesrdquo Journal of Neuro-science Methods vol 141 no 1 pp 41ndash53 2005
[45] I Lushnikova G Skibo D Muller and I Nikonenko ldquoSynapticpotentiation induces increased glial coverage of excitatorysynapses in CA1 hippocampusrdquoHippocampus vol 19 no 8 pp753ndash762 2009
[46] E Raponi F Agenes C Delphin et al ldquoS100B expressiondefines a state in which GFAP-expressing cells lose their neuralstem cell potential and acquire a more mature developmentalstagerdquo Glia vol 55 no 2 pp 165ndash177 2007
[47] C G Schipke C Boucsein C Ohlemeyer F Kirchhoff andH Kettenmann ldquoAstrocyte Ca2+ waves trigger responses inmicroglial cells in brain slicesrdquo The FASEB Journal vol 16 no2 pp 255ndash257 2002
[48] J Zhou and M L Sutherland ldquoGlutamate transporter clusterformation in astrocytic processes regulates glutamate uptakeactivityrdquo Journal of Neuroscience vol 24 no 28 pp 6301ndash63062004
[49] T Nakagawa Y Otsubo Y Yatani H Shirakawa and S KanekoldquoMechanisms of substrate transport-induced clustering of aglial glutamate transporter GLT-1 in astroglial-neuronal cul-turesrdquo European Journal of Neuroscience vol 28 no 9 pp 1719ndash1730 2008
[50] J-Y Chatton P Marquet and P J Magistretti ldquoA quantitativeanalysis of L-glutamate-regulated Na+ dynamics in mousecortical astrocytes Implications for cellular bioenergeticsrdquoEuropean Journal of Neuroscience vol 12 no 11 pp 3843ndash38532000
[51] E Forster S Zhao and M Frotscher ldquoLaminating the hip-pocampusrdquo Nature Reviews Neuroscience vol 7 no 4 pp 259ndash267 2006
[52] B H Gahwiler M Capogna D Debanne R A McKinney andS M Thompson ldquoOrganotypic slice cultures a technique hascome of agerdquo Trends in Neurosciences vol 20 no 10 pp 471ndash477 1997
[53] P E Kunkler and R P Kraig ldquoReactive astrocytosis fromexcitotoxic injury in hippocampal organ culture parallels thatseen in vivordquo Journal of Cerebral Blood Flow and Metabolismvol 17 no 1 pp 26ndash43 1997
[54] I E Holopainen ldquoOrganotypic hippocampal slice cultures amodel system to study basic cellular andmolecularmechanismsof neuronal cell death neuroprotection and synaptic plasticityrdquoNeurochemical Research vol 30 no 12 pp 1521ndash1528 2005
[55] MHaber L Zhou andK KMurai ldquoCooperative astrocyte anddendritic spine dynamics at hippocampal excitatory synapsesrdquoJournal of Neuroscience vol 26 no 35 pp 8881ndash8891 2006
16 ISRN Neurology
[56] H Nishida and S Okabe ldquoDirect astrocytic contacts regulatelocal maturation of dendritic spinesrdquo Journal of Neurosciencevol 27 no 2 pp 331ndash340 2007
[57] A Buffo C Rolando and S Ceruti ldquoAstrocytes in the dam-aged brain molecular and cellular insights into their reactiveresponse and healing potentialrdquoBiochemical Pharmacology vol79 no 2 pp 77ndash89 2010
[58] J-H Yi and A S Hazell ldquoExcitotoxic mechanisms and the roleof astrocytic glutamate transporters in traumatic brain injuryrdquoNeurochemistry International vol 48 no 5 pp 394ndash403 2006
[59] C Werner and K Engelhard ldquoPathophysiology of traumaticbrain injuryrdquo British Journal of Anaesthesia vol 99 no 1 pp4ndash9 2007
[60] G Gegelashvili M B Robinson D Trotti and T RauenldquoRegulation of glutamate transporters in health and diseaserdquoProgress in Brain Research vol 132 pp 267ndash286 2001
[61] C Escartin E Brouillet PGubellini et al ldquoCiliary neurotrophicfactor activates astrocytes redistributes their glutamate trans-porters GLAST and GLT-1 to raft microdomains and improvesglutamate handling in vivordquo Journal of Neuroscience vol 26 no22 pp 5978ndash5989 2006
[62] C Vermeiren M Najimi N Vanhoutte et al ldquoAcute up-regulation of glutamate uptake mediated by mGluR5a in reac-tive astrocytesrdquo Journal of Neurochemistry vol 94 no 2 pp405ndash416 2005
[63] C L Poitry-Yamate L Vutskits and T Rauen ldquoNeuronal-induced and glutamate-dependent activation of glial glutamatetransporter functionrdquo Journal of Neurochemistry vol 82 no 4pp 987ndash997 2002
[64] P M Beart and R D OrsquoShea ldquoTransporters for L-glutamatean update on their molecular pharmacology and pathologicalinvolvementrdquo British Journal of Pharmacology vol 150 no 1pp 5ndash17 2007
[65] F D Lima M A Souza A F Furian et al ldquoNa+K+-ATPaseactivity impairment after experimental traumatic brain injuryrelationship to spatial learning deficits and oxidative stressrdquoBehavioural Brain Research vol 193 no 2 pp 306ndash310 2008
[66] G A Gusarova H E Trejo L A Dada et al ldquoHypoxia leadsto NaK-ATPase downregulation via Ca2+ release-activatedCa2+ channels and AMPK activationrdquo Molecular and CellularBiology vol 31 no 17 pp 3546ndash3556 2011
[67] S S L Chew C S Johnson C R Green and H V Danesh-Meyer ldquoRole of connexin43 in central nervous system injuryrdquoExperimental Neurology vol 225 no 2 pp 250ndash261 2010
[68] P Nilsson L Hillered U Ponten and U Ungerstedt ldquoChangesin cortical extracellular levels of energy-related metabolites andamino acids following concussive brain injury in ratsrdquo Journal ofCerebral Blood Flow and Metabolism vol 10 no 5 pp 631ndash6371990
[69] H Katoh K Sima H Nawashiro K Wada and H ChigasakildquoThe effect of MK-801 on extracellular neuroactive amino acidsin hippocampus after closed head injury followed by hypoxia inratsrdquo Brain Research vol 758 no 1-2 pp 153ndash162 1997
[70] H Koizumi H Fujisawa H Ito T Maekawa X Di and RBullock ldquoEffects of mild hypothermia on cerebral blood flow-independent changes in cortical extracellular levels of aminoacids following contusion trauma in the ratrdquoBrain Research vol747 no 2 pp 304ndash312 1997
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

14 ISRN Neurology
conditions Indeed ambient glutamate concentrations wereshown to be significantly increased after traumatic braininjury [68ndash70] Along these lines it can be assumed thatin our model system extracellular glutamate concentrationsrose less severely in the periphery of the lesion preventinga comparable clustering and loss of functional glutamatetransport activity in moderately activated astrocytes
5 Conclusions
Our results show complex changes in glutamate transporterexpression and function during astrocyte activation followingmechanical injury They confirm that the degree of astrocyteactivation depends on the distance from the insult and com-prises different endpoints regarding cellular morphology andphysiology Furthermore our results highlight that astrocyteswhich show different grades of reactivity also display discretechanges in glutamate transporter expression and functionWhile immunohistochemistry revealed a prominent clus-tering of GLT-1 and GLAST immunoreactivity in the scarregion our functional assay clearly showed that glutamateuptake capacity is strongly reduced in scar cells while itis largely maintained in moderately activated astrocytes inthe periphery Thus mild-to-moderate astrogliosis in theperiphery of a mechanical lesion does not necessarily seemto be accompanied by a significant change in glial glutamateuptake capacity At the glial scar itself in strongly reactiveastrocytes a clustering of glutamate transporters is observedthat apparently goes along with a severe functional reductionin astroglial glutamate uptake which may contribute toglutamate-mediated excitotoxicity in this region
Conflict of Interests
The authors of the paper declare that they do not haveany direct financial relation with the commercial identitiesmentioned in the paper that might lead to a conflict ofinterests
Acknowledgments
This study was supported by the Deutsche Forschungsge-meinschaft (DFG Ro23274-3) The authors thank SimoneDurry and Claudia Roderigo for expert technical help
References
[1] N C Danbolt ldquoGlutamate uptakerdquo Progress in Neurobiologyvol 65 no 1 pp 1ndash105 2001
[2] A V Tzingounis and J I Wadiche ldquoGlutamate transportersconfining runaway excitation by shaping synaptic transmis-sionrdquo Nature Reviews Neuroscience vol 8 no 12 pp 935ndash9472007
[3] D E Bergles J S Diamond and C E Jahr ldquoClearance ofglutamate inside the synapse and beyondrdquo Current Opinion inNeurobiology vol 9 no 3 pp 293ndash298 1999
[4] C M Anderson and R A Swanson ldquoAstrocyte glutamatetransport review of properties regulation and physiologicalfunctionsrdquo Glia vol 32 no 1 pp 1ndash14 2000
[5] P Marcaggi and D Attwell ldquoRole of glial amino acid trans-porters in synaptic transmission and brain energeticsrdquoGlia vol47 no 3 pp 217ndash225 2004
[6] N J Maragakis and J D Rothstein ldquoGlutamate transportersanimal models to neurologic diseaserdquo Neurobiology of Diseasevol 15 no 3 pp 461ndash473 2004
[7] G Gegelashvili and A Schousboe ldquoCellular distribution andkinetic properties of high-affinity glutamate transportersrdquoBrainResearch Bulletin vol 45 no 3 pp 233ndash238 1998
[8] J W Deitmer and C R Rose ldquoIon changes and signalling inperisynaptic gliardquo Brain Research Reviews vol 63 no 1-2 pp113ndash129 2010
[9] S Kirischuk V Parpura and A Verkhratsky ldquoSodium dynam-ics another key to astroglial excitabilityrdquo Trends in Neuro-sciences vol 35 no 8 pp 497ndash506 2012
[10] M V Sofroniew ldquoMolecular dissection of reactive astrogliosisand glial scar formationrdquo Trends in Neurosciences vol 32 no12 pp 638ndash647 2009
[11] J L Ridet S K Malhotra A Privat and F H Gage ldquoReactiveastrocytes cellular and molecular cues to biological functionrdquoTrends in Neurosciences vol 20 no 12 pp 570ndash577 1997
[12] J Middeldorp and E M Hol ldquoGFAP in health and diseaserdquoProgress in Neurobiology vol 93 no 3 pp 421ndash443 2011
[13] M Pekny and M Nilsson ldquoAstrocyte activation and reactivegliosisrdquo Glia vol 50 no 4 pp 427ndash434 2005
[14] A Bordey S A Lyons J J Hablitz and H SontheimerldquoElectrophysiological characteristics of reactive astrocytes inexperimental cortical dysplasiardquo Journal of Neurophysiologyvol 85 no 4 pp 1719ndash1731 2001
[15] R Jabs G Seifert and C Steinhauser ldquoAstrocytic functionand its alteration in the epileptic brainrdquo Epilepsia vol 49supplement 2 pp 3ndash12 2008
[16] A Buffo I Rite P Tripathi et al ldquoOrigin and progeny ofreactive gliosis a source of multipotent cells in the injuredbrainrdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 105 no 9 pp 3581ndash3586 2008
[17] S Robel B Berninger and M Gotz ldquoThe stem cell potential ofglia lessons from reactive gliosisrdquoNature Reviews Neurosciencevol 12 no 2 pp 88ndash104 2011
[18] H Kawano J Kimura-Kuroda Y Komuta et al ldquoRole of thelesion scar in the response to damage and repair of the centralnervous systemrdquo Cell and Tissue Research vol 349 no 1 pp169ndash180 2012
[19] G Yiu and Z He ldquoGlial inhibition of CNS axon regenerationrdquoNature Reviews Neuroscience vol 7 no 8 pp 617ndash627 2006
[20] M T Fitch and J Silver ldquoCNS injury glial scars and inflamma-tion inhibitory extracellular matrices and regeneration failurerdquoExperimental Neurology vol 209 no 2 pp 294ndash301 2008
[21] L Li A Lundkvist D Andersson et al ldquoProtective role ofreactive astrocytes in brain ischemiardquo Journal of Cerebral BloodFlow amp Metabolism vol 28 no 3 pp 468ndash481 2008
[22] F K H van Landeghem T Weiss M Oehmichen and AVonDeimling ldquoDecreased expression of glutamate transportersin astrocytes after human traumatic brain injuryrdquo Journal ofNeurotrauma vol 23 no 10 pp 1518ndash1528 2006
[23] V L Raghavendra Rao M K Baskaya A Dogan J DRothstein and R J Dempsey ldquoTraumatic brain injury down-regulates glial glutamate transporter (GLT- 1 and GLAST)proteins in rat brainrdquo Journal of Neurochemistry vol 70 no 5pp 2020ndash2027 1998
ISRN Neurology 15
[24] J-C Chen H Hsu-Chou J-L Lu et al ldquoDown-regulation ofthe glial glutamate transporter GLT-1 in rat hippocampus andstriatum and its modulation by a group III metabotropic glu-tamate receptor antagonist following transient global forebrainischemiardquoNeuropharmacology vol 49 no 5 pp 703ndash714 2005
[25] S D Ginsberg L J Martin and J D Rothstein ldquoRegional deaf-ferentation down-regulates subtypes of glutamate transporterproteinsrdquo Journal of Neurochemistry vol 65 no 6 pp 2800ndash2803 1995
[26] J E Springer R D Azbill R J Mark J G Begley GWaeg andM P Mattson ldquo4-Hydroxynonenal a lipid peroxidation prod-uct rapidly accumulates following traumatic spinal cord injuryand inhibits glutamate uptakerdquo Journal of Neurochemistry vol68 no 6 pp 2469ndash2476 1997
[27] M B Moretto N S Arteni D Lavinsky et al ldquoHypoxic-ischemic insult decreases glutamate uptake by hippocampalslices from neonatal rats Prevention by guanosinerdquo Experimen-tal Neurology vol 195 no 2 pp 400ndash406 2005
[28] L Stoppini P-A Buchs and D Muller ldquoA simple method fororganotypic cultures of nervous tissuerdquo Journal of NeuroscienceMethods vol 37 no 2 pp 173ndash182 1991
[29] A Nimmerjahn F Kirchhoff J N D Kerr and F HelmchenldquoSulforhodamine 101 as a specific marker of astroglia in theneocortex in vivordquo Nat Methods vol 1 no 1 pp 31ndash37 2004
[30] K W Kafitz S D Meier J Stephan and C R Rose ldquoDevel-opmental profile and properties of sulforhodamine 101-Labeledglial cells in acute brain slices of rat hippocampusrdquo Journal ofNeuroscience Methods vol 169 no 1 pp 84ndash92 2008
[31] C Schnell Y Hagos and S Hulsmann ldquoActive sulforhodamine101 uptake into hippocampal astrocytesrdquo PLoS One vol 7 no 11Article ID e49398 2012
[32] S D Meier Y Kovalchuk and C R Rose ldquoProperties ofthe new fluorescent Na+ indicator CoroNa Green comparisonwith SBFI and confocal Na+ imagingrdquo Journal of NeuroscienceMethods vol 155 no 2 pp 251ndash259 2006
[33] J Langer and C R Rose ldquoSynaptically induced sodium signalsin hippocampal astrocytes in siturdquo Journal of Physiology vol587 part 24 pp 5859ndash5877 2009
[34] J Langer J Stephan M Theis and C R Rose ldquoGap junctionsmediate intercellular spread of sodium between hippocampalastrocytes in siturdquo Glia vol 60 no 2 pp 239ndash252 2012
[35] C R Rose andB R Ransom ldquoIntracellular sodiumhomeostasisin rat hippocampal astrocytesrdquo Journal of Physiology vol 491part 2 no 2 pp 291ndash305 1996
[36] A M Benediktsson G S Marrs J C Tu et al ldquoNeuronalactivity regulates glutamate transporter dynamics in developingastrocytesrdquo Glia vol 60 no 2 pp 175ndash188 2012
[37] M Zhou G P Schools and H K Kimelberg ldquoDevelopment ofGLAST(+) astrocytes andNG2(+) glia in rat hippocampus CA1mature astrocytes are electrophysiologically passiverdquo Journal ofNeurophysiology vol 95 no 1 pp 134ndash143 2006
[38] I Suarez G Bodega and B Fernandez ldquoModulation of glu-tamate transporters (GLAST GLT-1 and EAAC1) in the ratcerebellum following portocaval anastomosisrdquo Brain Researchvol 859 no 2 pp 293ndash302 2000
[39] X Zhu D E Bergles and A Nishiyama ldquoNG2 cells generateboth oligodendrocytes and gray matter astrocytesrdquo Develop-ment vol 135 no 1 pp 145ndash157 2008
[40] B Brunne S ZhaoADerouiche et al ldquoOriginmaturation andastroglial transformation of secondary radial glial cells in thedeveloping dentate gyrusrdquo Glia vol 58 no 13 pp 1553ndash15692010
[41] Y Wu A-Q Zhang and D T Yew ldquoAge related changes ofvarious markers of astrocytes in senescence-accelerated micehippocampusrdquo Neurochemistry International vol 46 no 7 pp565ndash574 2005
[42] S Magavi D Friedmann G Banks A Stolfi and C LoisldquoCoincident generation of pyramidal neurons and protoplasmicastrocytes in neocortical columnsrdquo Journal of Neuroscience vol32 no 14 pp 4762ndash4772 2012
[43] M Olabarria H N Noristani A Verkhratsky and J JRodrıguez ldquoConcomitant astroglial atrophy and astrogliosis ina triple transgenic animal model of Alzheimerrsquos diseaserdquo Gliavol 58 no 7 pp 831ndash838 2010
[44] A M Benediktsson S J Schachtele S H Green and M EDailey ldquoBallistic labeling and dynamic imaging of astrocytesin organotypic hippocampal slice culturesrdquo Journal of Neuro-science Methods vol 141 no 1 pp 41ndash53 2005
[45] I Lushnikova G Skibo D Muller and I Nikonenko ldquoSynapticpotentiation induces increased glial coverage of excitatorysynapses in CA1 hippocampusrdquoHippocampus vol 19 no 8 pp753ndash762 2009
[46] E Raponi F Agenes C Delphin et al ldquoS100B expressiondefines a state in which GFAP-expressing cells lose their neuralstem cell potential and acquire a more mature developmentalstagerdquo Glia vol 55 no 2 pp 165ndash177 2007
[47] C G Schipke C Boucsein C Ohlemeyer F Kirchhoff andH Kettenmann ldquoAstrocyte Ca2+ waves trigger responses inmicroglial cells in brain slicesrdquo The FASEB Journal vol 16 no2 pp 255ndash257 2002
[48] J Zhou and M L Sutherland ldquoGlutamate transporter clusterformation in astrocytic processes regulates glutamate uptakeactivityrdquo Journal of Neuroscience vol 24 no 28 pp 6301ndash63062004
[49] T Nakagawa Y Otsubo Y Yatani H Shirakawa and S KanekoldquoMechanisms of substrate transport-induced clustering of aglial glutamate transporter GLT-1 in astroglial-neuronal cul-turesrdquo European Journal of Neuroscience vol 28 no 9 pp 1719ndash1730 2008
[50] J-Y Chatton P Marquet and P J Magistretti ldquoA quantitativeanalysis of L-glutamate-regulated Na+ dynamics in mousecortical astrocytes Implications for cellular bioenergeticsrdquoEuropean Journal of Neuroscience vol 12 no 11 pp 3843ndash38532000
[51] E Forster S Zhao and M Frotscher ldquoLaminating the hip-pocampusrdquo Nature Reviews Neuroscience vol 7 no 4 pp 259ndash267 2006
[52] B H Gahwiler M Capogna D Debanne R A McKinney andS M Thompson ldquoOrganotypic slice cultures a technique hascome of agerdquo Trends in Neurosciences vol 20 no 10 pp 471ndash477 1997
[53] P E Kunkler and R P Kraig ldquoReactive astrocytosis fromexcitotoxic injury in hippocampal organ culture parallels thatseen in vivordquo Journal of Cerebral Blood Flow and Metabolismvol 17 no 1 pp 26ndash43 1997
[54] I E Holopainen ldquoOrganotypic hippocampal slice cultures amodel system to study basic cellular andmolecularmechanismsof neuronal cell death neuroprotection and synaptic plasticityrdquoNeurochemical Research vol 30 no 12 pp 1521ndash1528 2005
[55] MHaber L Zhou andK KMurai ldquoCooperative astrocyte anddendritic spine dynamics at hippocampal excitatory synapsesrdquoJournal of Neuroscience vol 26 no 35 pp 8881ndash8891 2006
16 ISRN Neurology
[56] H Nishida and S Okabe ldquoDirect astrocytic contacts regulatelocal maturation of dendritic spinesrdquo Journal of Neurosciencevol 27 no 2 pp 331ndash340 2007
[57] A Buffo C Rolando and S Ceruti ldquoAstrocytes in the dam-aged brain molecular and cellular insights into their reactiveresponse and healing potentialrdquoBiochemical Pharmacology vol79 no 2 pp 77ndash89 2010
[58] J-H Yi and A S Hazell ldquoExcitotoxic mechanisms and the roleof astrocytic glutamate transporters in traumatic brain injuryrdquoNeurochemistry International vol 48 no 5 pp 394ndash403 2006
[59] C Werner and K Engelhard ldquoPathophysiology of traumaticbrain injuryrdquo British Journal of Anaesthesia vol 99 no 1 pp4ndash9 2007
[60] G Gegelashvili M B Robinson D Trotti and T RauenldquoRegulation of glutamate transporters in health and diseaserdquoProgress in Brain Research vol 132 pp 267ndash286 2001
[61] C Escartin E Brouillet PGubellini et al ldquoCiliary neurotrophicfactor activates astrocytes redistributes their glutamate trans-porters GLAST and GLT-1 to raft microdomains and improvesglutamate handling in vivordquo Journal of Neuroscience vol 26 no22 pp 5978ndash5989 2006
[62] C Vermeiren M Najimi N Vanhoutte et al ldquoAcute up-regulation of glutamate uptake mediated by mGluR5a in reac-tive astrocytesrdquo Journal of Neurochemistry vol 94 no 2 pp405ndash416 2005
[63] C L Poitry-Yamate L Vutskits and T Rauen ldquoNeuronal-induced and glutamate-dependent activation of glial glutamatetransporter functionrdquo Journal of Neurochemistry vol 82 no 4pp 987ndash997 2002
[64] P M Beart and R D OrsquoShea ldquoTransporters for L-glutamatean update on their molecular pharmacology and pathologicalinvolvementrdquo British Journal of Pharmacology vol 150 no 1pp 5ndash17 2007
[65] F D Lima M A Souza A F Furian et al ldquoNa+K+-ATPaseactivity impairment after experimental traumatic brain injuryrelationship to spatial learning deficits and oxidative stressrdquoBehavioural Brain Research vol 193 no 2 pp 306ndash310 2008
[66] G A Gusarova H E Trejo L A Dada et al ldquoHypoxia leadsto NaK-ATPase downregulation via Ca2+ release-activatedCa2+ channels and AMPK activationrdquo Molecular and CellularBiology vol 31 no 17 pp 3546ndash3556 2011
[67] S S L Chew C S Johnson C R Green and H V Danesh-Meyer ldquoRole of connexin43 in central nervous system injuryrdquoExperimental Neurology vol 225 no 2 pp 250ndash261 2010
[68] P Nilsson L Hillered U Ponten and U Ungerstedt ldquoChangesin cortical extracellular levels of energy-related metabolites andamino acids following concussive brain injury in ratsrdquo Journal ofCerebral Blood Flow and Metabolism vol 10 no 5 pp 631ndash6371990
[69] H Katoh K Sima H Nawashiro K Wada and H ChigasakildquoThe effect of MK-801 on extracellular neuroactive amino acidsin hippocampus after closed head injury followed by hypoxia inratsrdquo Brain Research vol 758 no 1-2 pp 153ndash162 1997
[70] H Koizumi H Fujisawa H Ito T Maekawa X Di and RBullock ldquoEffects of mild hypothermia on cerebral blood flow-independent changes in cortical extracellular levels of aminoacids following contusion trauma in the ratrdquoBrain Research vol747 no 2 pp 304ndash312 1997
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

ISRN Neurology 15
[24] J-C Chen H Hsu-Chou J-L Lu et al ldquoDown-regulation ofthe glial glutamate transporter GLT-1 in rat hippocampus andstriatum and its modulation by a group III metabotropic glu-tamate receptor antagonist following transient global forebrainischemiardquoNeuropharmacology vol 49 no 5 pp 703ndash714 2005
[25] S D Ginsberg L J Martin and J D Rothstein ldquoRegional deaf-ferentation down-regulates subtypes of glutamate transporterproteinsrdquo Journal of Neurochemistry vol 65 no 6 pp 2800ndash2803 1995
[26] J E Springer R D Azbill R J Mark J G Begley GWaeg andM P Mattson ldquo4-Hydroxynonenal a lipid peroxidation prod-uct rapidly accumulates following traumatic spinal cord injuryand inhibits glutamate uptakerdquo Journal of Neurochemistry vol68 no 6 pp 2469ndash2476 1997
[27] M B Moretto N S Arteni D Lavinsky et al ldquoHypoxic-ischemic insult decreases glutamate uptake by hippocampalslices from neonatal rats Prevention by guanosinerdquo Experimen-tal Neurology vol 195 no 2 pp 400ndash406 2005
[28] L Stoppini P-A Buchs and D Muller ldquoA simple method fororganotypic cultures of nervous tissuerdquo Journal of NeuroscienceMethods vol 37 no 2 pp 173ndash182 1991
[29] A Nimmerjahn F Kirchhoff J N D Kerr and F HelmchenldquoSulforhodamine 101 as a specific marker of astroglia in theneocortex in vivordquo Nat Methods vol 1 no 1 pp 31ndash37 2004
[30] K W Kafitz S D Meier J Stephan and C R Rose ldquoDevel-opmental profile and properties of sulforhodamine 101-Labeledglial cells in acute brain slices of rat hippocampusrdquo Journal ofNeuroscience Methods vol 169 no 1 pp 84ndash92 2008
[31] C Schnell Y Hagos and S Hulsmann ldquoActive sulforhodamine101 uptake into hippocampal astrocytesrdquo PLoS One vol 7 no 11Article ID e49398 2012
[32] S D Meier Y Kovalchuk and C R Rose ldquoProperties ofthe new fluorescent Na+ indicator CoroNa Green comparisonwith SBFI and confocal Na+ imagingrdquo Journal of NeuroscienceMethods vol 155 no 2 pp 251ndash259 2006
[33] J Langer and C R Rose ldquoSynaptically induced sodium signalsin hippocampal astrocytes in siturdquo Journal of Physiology vol587 part 24 pp 5859ndash5877 2009
[34] J Langer J Stephan M Theis and C R Rose ldquoGap junctionsmediate intercellular spread of sodium between hippocampalastrocytes in siturdquo Glia vol 60 no 2 pp 239ndash252 2012
[35] C R Rose andB R Ransom ldquoIntracellular sodiumhomeostasisin rat hippocampal astrocytesrdquo Journal of Physiology vol 491part 2 no 2 pp 291ndash305 1996
[36] A M Benediktsson G S Marrs J C Tu et al ldquoNeuronalactivity regulates glutamate transporter dynamics in developingastrocytesrdquo Glia vol 60 no 2 pp 175ndash188 2012
[37] M Zhou G P Schools and H K Kimelberg ldquoDevelopment ofGLAST(+) astrocytes andNG2(+) glia in rat hippocampus CA1mature astrocytes are electrophysiologically passiverdquo Journal ofNeurophysiology vol 95 no 1 pp 134ndash143 2006
[38] I Suarez G Bodega and B Fernandez ldquoModulation of glu-tamate transporters (GLAST GLT-1 and EAAC1) in the ratcerebellum following portocaval anastomosisrdquo Brain Researchvol 859 no 2 pp 293ndash302 2000
[39] X Zhu D E Bergles and A Nishiyama ldquoNG2 cells generateboth oligodendrocytes and gray matter astrocytesrdquo Develop-ment vol 135 no 1 pp 145ndash157 2008
[40] B Brunne S ZhaoADerouiche et al ldquoOriginmaturation andastroglial transformation of secondary radial glial cells in thedeveloping dentate gyrusrdquo Glia vol 58 no 13 pp 1553ndash15692010
[41] Y Wu A-Q Zhang and D T Yew ldquoAge related changes ofvarious markers of astrocytes in senescence-accelerated micehippocampusrdquo Neurochemistry International vol 46 no 7 pp565ndash574 2005
[42] S Magavi D Friedmann G Banks A Stolfi and C LoisldquoCoincident generation of pyramidal neurons and protoplasmicastrocytes in neocortical columnsrdquo Journal of Neuroscience vol32 no 14 pp 4762ndash4772 2012
[43] M Olabarria H N Noristani A Verkhratsky and J JRodrıguez ldquoConcomitant astroglial atrophy and astrogliosis ina triple transgenic animal model of Alzheimerrsquos diseaserdquo Gliavol 58 no 7 pp 831ndash838 2010
[44] A M Benediktsson S J Schachtele S H Green and M EDailey ldquoBallistic labeling and dynamic imaging of astrocytesin organotypic hippocampal slice culturesrdquo Journal of Neuro-science Methods vol 141 no 1 pp 41ndash53 2005
[45] I Lushnikova G Skibo D Muller and I Nikonenko ldquoSynapticpotentiation induces increased glial coverage of excitatorysynapses in CA1 hippocampusrdquoHippocampus vol 19 no 8 pp753ndash762 2009
[46] E Raponi F Agenes C Delphin et al ldquoS100B expressiondefines a state in which GFAP-expressing cells lose their neuralstem cell potential and acquire a more mature developmentalstagerdquo Glia vol 55 no 2 pp 165ndash177 2007
[47] C G Schipke C Boucsein C Ohlemeyer F Kirchhoff andH Kettenmann ldquoAstrocyte Ca2+ waves trigger responses inmicroglial cells in brain slicesrdquo The FASEB Journal vol 16 no2 pp 255ndash257 2002
[48] J Zhou and M L Sutherland ldquoGlutamate transporter clusterformation in astrocytic processes regulates glutamate uptakeactivityrdquo Journal of Neuroscience vol 24 no 28 pp 6301ndash63062004
[49] T Nakagawa Y Otsubo Y Yatani H Shirakawa and S KanekoldquoMechanisms of substrate transport-induced clustering of aglial glutamate transporter GLT-1 in astroglial-neuronal cul-turesrdquo European Journal of Neuroscience vol 28 no 9 pp 1719ndash1730 2008
[50] J-Y Chatton P Marquet and P J Magistretti ldquoA quantitativeanalysis of L-glutamate-regulated Na+ dynamics in mousecortical astrocytes Implications for cellular bioenergeticsrdquoEuropean Journal of Neuroscience vol 12 no 11 pp 3843ndash38532000
[51] E Forster S Zhao and M Frotscher ldquoLaminating the hip-pocampusrdquo Nature Reviews Neuroscience vol 7 no 4 pp 259ndash267 2006
[52] B H Gahwiler M Capogna D Debanne R A McKinney andS M Thompson ldquoOrganotypic slice cultures a technique hascome of agerdquo Trends in Neurosciences vol 20 no 10 pp 471ndash477 1997
[53] P E Kunkler and R P Kraig ldquoReactive astrocytosis fromexcitotoxic injury in hippocampal organ culture parallels thatseen in vivordquo Journal of Cerebral Blood Flow and Metabolismvol 17 no 1 pp 26ndash43 1997
[54] I E Holopainen ldquoOrganotypic hippocampal slice cultures amodel system to study basic cellular andmolecularmechanismsof neuronal cell death neuroprotection and synaptic plasticityrdquoNeurochemical Research vol 30 no 12 pp 1521ndash1528 2005
[55] MHaber L Zhou andK KMurai ldquoCooperative astrocyte anddendritic spine dynamics at hippocampal excitatory synapsesrdquoJournal of Neuroscience vol 26 no 35 pp 8881ndash8891 2006
16 ISRN Neurology
[56] H Nishida and S Okabe ldquoDirect astrocytic contacts regulatelocal maturation of dendritic spinesrdquo Journal of Neurosciencevol 27 no 2 pp 331ndash340 2007
[57] A Buffo C Rolando and S Ceruti ldquoAstrocytes in the dam-aged brain molecular and cellular insights into their reactiveresponse and healing potentialrdquoBiochemical Pharmacology vol79 no 2 pp 77ndash89 2010
[58] J-H Yi and A S Hazell ldquoExcitotoxic mechanisms and the roleof astrocytic glutamate transporters in traumatic brain injuryrdquoNeurochemistry International vol 48 no 5 pp 394ndash403 2006
[59] C Werner and K Engelhard ldquoPathophysiology of traumaticbrain injuryrdquo British Journal of Anaesthesia vol 99 no 1 pp4ndash9 2007
[60] G Gegelashvili M B Robinson D Trotti and T RauenldquoRegulation of glutamate transporters in health and diseaserdquoProgress in Brain Research vol 132 pp 267ndash286 2001
[61] C Escartin E Brouillet PGubellini et al ldquoCiliary neurotrophicfactor activates astrocytes redistributes their glutamate trans-porters GLAST and GLT-1 to raft microdomains and improvesglutamate handling in vivordquo Journal of Neuroscience vol 26 no22 pp 5978ndash5989 2006
[62] C Vermeiren M Najimi N Vanhoutte et al ldquoAcute up-regulation of glutamate uptake mediated by mGluR5a in reac-tive astrocytesrdquo Journal of Neurochemistry vol 94 no 2 pp405ndash416 2005
[63] C L Poitry-Yamate L Vutskits and T Rauen ldquoNeuronal-induced and glutamate-dependent activation of glial glutamatetransporter functionrdquo Journal of Neurochemistry vol 82 no 4pp 987ndash997 2002
[64] P M Beart and R D OrsquoShea ldquoTransporters for L-glutamatean update on their molecular pharmacology and pathologicalinvolvementrdquo British Journal of Pharmacology vol 150 no 1pp 5ndash17 2007
[65] F D Lima M A Souza A F Furian et al ldquoNa+K+-ATPaseactivity impairment after experimental traumatic brain injuryrelationship to spatial learning deficits and oxidative stressrdquoBehavioural Brain Research vol 193 no 2 pp 306ndash310 2008
[66] G A Gusarova H E Trejo L A Dada et al ldquoHypoxia leadsto NaK-ATPase downregulation via Ca2+ release-activatedCa2+ channels and AMPK activationrdquo Molecular and CellularBiology vol 31 no 17 pp 3546ndash3556 2011
[67] S S L Chew C S Johnson C R Green and H V Danesh-Meyer ldquoRole of connexin43 in central nervous system injuryrdquoExperimental Neurology vol 225 no 2 pp 250ndash261 2010
[68] P Nilsson L Hillered U Ponten and U Ungerstedt ldquoChangesin cortical extracellular levels of energy-related metabolites andamino acids following concussive brain injury in ratsrdquo Journal ofCerebral Blood Flow and Metabolism vol 10 no 5 pp 631ndash6371990
[69] H Katoh K Sima H Nawashiro K Wada and H ChigasakildquoThe effect of MK-801 on extracellular neuroactive amino acidsin hippocampus after closed head injury followed by hypoxia inratsrdquo Brain Research vol 758 no 1-2 pp 153ndash162 1997
[70] H Koizumi H Fujisawa H Ito T Maekawa X Di and RBullock ldquoEffects of mild hypothermia on cerebral blood flow-independent changes in cortical extracellular levels of aminoacids following contusion trauma in the ratrdquoBrain Research vol747 no 2 pp 304ndash312 1997
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

16 ISRN Neurology
[56] H Nishida and S Okabe ldquoDirect astrocytic contacts regulatelocal maturation of dendritic spinesrdquo Journal of Neurosciencevol 27 no 2 pp 331ndash340 2007
[57] A Buffo C Rolando and S Ceruti ldquoAstrocytes in the dam-aged brain molecular and cellular insights into their reactiveresponse and healing potentialrdquoBiochemical Pharmacology vol79 no 2 pp 77ndash89 2010
[58] J-H Yi and A S Hazell ldquoExcitotoxic mechanisms and the roleof astrocytic glutamate transporters in traumatic brain injuryrdquoNeurochemistry International vol 48 no 5 pp 394ndash403 2006
[59] C Werner and K Engelhard ldquoPathophysiology of traumaticbrain injuryrdquo British Journal of Anaesthesia vol 99 no 1 pp4ndash9 2007
[60] G Gegelashvili M B Robinson D Trotti and T RauenldquoRegulation of glutamate transporters in health and diseaserdquoProgress in Brain Research vol 132 pp 267ndash286 2001
[61] C Escartin E Brouillet PGubellini et al ldquoCiliary neurotrophicfactor activates astrocytes redistributes their glutamate trans-porters GLAST and GLT-1 to raft microdomains and improvesglutamate handling in vivordquo Journal of Neuroscience vol 26 no22 pp 5978ndash5989 2006
[62] C Vermeiren M Najimi N Vanhoutte et al ldquoAcute up-regulation of glutamate uptake mediated by mGluR5a in reac-tive astrocytesrdquo Journal of Neurochemistry vol 94 no 2 pp405ndash416 2005
[63] C L Poitry-Yamate L Vutskits and T Rauen ldquoNeuronal-induced and glutamate-dependent activation of glial glutamatetransporter functionrdquo Journal of Neurochemistry vol 82 no 4pp 987ndash997 2002
[64] P M Beart and R D OrsquoShea ldquoTransporters for L-glutamatean update on their molecular pharmacology and pathologicalinvolvementrdquo British Journal of Pharmacology vol 150 no 1pp 5ndash17 2007
[65] F D Lima M A Souza A F Furian et al ldquoNa+K+-ATPaseactivity impairment after experimental traumatic brain injuryrelationship to spatial learning deficits and oxidative stressrdquoBehavioural Brain Research vol 193 no 2 pp 306ndash310 2008
[66] G A Gusarova H E Trejo L A Dada et al ldquoHypoxia leadsto NaK-ATPase downregulation via Ca2+ release-activatedCa2+ channels and AMPK activationrdquo Molecular and CellularBiology vol 31 no 17 pp 3546ndash3556 2011
[67] S S L Chew C S Johnson C R Green and H V Danesh-Meyer ldquoRole of connexin43 in central nervous system injuryrdquoExperimental Neurology vol 225 no 2 pp 250ndash261 2010
[68] P Nilsson L Hillered U Ponten and U Ungerstedt ldquoChangesin cortical extracellular levels of energy-related metabolites andamino acids following concussive brain injury in ratsrdquo Journal ofCerebral Blood Flow and Metabolism vol 10 no 5 pp 631ndash6371990
[69] H Katoh K Sima H Nawashiro K Wada and H ChigasakildquoThe effect of MK-801 on extracellular neuroactive amino acidsin hippocampus after closed head injury followed by hypoxia inratsrdquo Brain Research vol 758 no 1-2 pp 153ndash162 1997
[70] H Koizumi H Fujisawa H Ito T Maekawa X Di and RBullock ldquoEffects of mild hypothermia on cerebral blood flow-independent changes in cortical extracellular levels of aminoacids following contusion trauma in the ratrdquoBrain Research vol747 no 2 pp 304ndash312 1997
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom


















![Spinal cord-specific deletion of the glutamate transporter ... · Consistent with previous report [19], the strong tdTomato ... Hoxb8-Cre/GLT1 flox/flox mice had slightly lower body](https://static.fdocuments.net/doc/165x107/5b0a0b357f8b9ae61b8b7d58/spinal-cord-specific-deletion-of-the-glutamate-transporter-with-previous-report.jpg)