
Large-scale reconstitution of a retina-to-brain pathway in adult rats ...
15
Large-scale reconstitution of a retina-to-brain pathway in adult rats using gene therapy and bridging grafts: An anatomical and behavioral analysis ☆ Si-Wei You d,1 , Mats Hellström e,1 , Margaret A. Pollett a , Chrisna LeVaillant a , Colette Moses a , Paul J. Rigby c , Marissa Penrose b , Jennifer Rodger b,f , Alan R. Harvey a,f, ⁎ a School of Anatomy, Physiology and Human Biology, The University of Western Australia, Australia b School of Animal Biology, The University of Western Australia, Australia c Centre for Microscopy, Characterisation and Analysis, The University of Western Australia, Australia d Department of Ophthalmology, Xijing Hospital, The Fourth Military Medical University, Xi’an, China e Laboratory for Transplantation and Regenerative Medicine, Dept. of Obstetrics and Gynecology, Sahlgrenska Academy, University of Gothenburg, Sweden f Western Australian Neuroscience Research Institute, Australia abstract article info Article history: Received 5 February 2016 Received in revised form 1 March 2016 Accepted 2 March 2016 Available online 10 March 2016 Peripheral nerve (PN) grafts can be used to bridge tissue defects in the CNS. Using a PN-to-optic nerve (ON) graft model, we combined gene therapy with pharmacotherapy to promote the long-distance regeneration of injured adult retinal ganglion cells (RGCs). Autologous sciatic nerve was sutured onto the transected ON and the distal end immediately inserted into contralateral superior colliculus (SC). Control rats received intraocular injections of saline or adeno-associated virus (AAV) encoding GFP. In experimental groups, three bi-cistronic AAV vectors encoding ciliary neurotrophic factor (CNTF) were injected into different regions of the grafted eye. Each vector encoded a different fluorescent reporter to assess retinotopic order in the regenerate projection. To encourage sprouting/synaptogenesis, after 6 weeks some AAV–CNTF injected rats received an intravitreal injection of re- combinant brain-derived neurotrophic factor (rBDNF) or AAV–BDNF. Four months after surgery, cholera toxin B was used to visualize regenerate RGC axons. RGC viability and axonal regrowth into SC were significantly great- er in AAV–CNTF groups. In some cases, near the insertion site, regenerate axonal density resembled retinal termi- nal densities seen in normal SC. Complex arbors were seen in superficial but not deep SC layers and many terminals were immunopositive for presynaptic proteins vGlut2 and SV2. There was improvement in visual func- tion via the grafted eye with significantly greater pupillary constriction in both AAV–CNTF+BDNF groups. In both control and AAV–CNTF+rBDNF groups the extent of light avoidance correlated with the maximal distance of axonal penetration into superficial SC. Despite the robust regrowth of RGC axons back into the SC, axons originating from different parts of the retina were intermixed at the PN graft/host SC interface, indicating that there remained a lack of order in this extensive regenerate projection. © 2016 Elsevier Inc. All rights reserved. Keywords: Gene therapy Peripheral nerve transplantation Adult rat visual system Axonal regeneration Superior colliculus Neurotrophic factors Retinal ganglion cells 1. Introduction Injury to the adult mammalian central nervous system (CNS) re- sults in long-lasting functional deficits, a consequence of intrinsic and extrinsic factors that together impact on plasticity and regener- ative ability. In the rodent primary visual system, often used as an ex- perimental model of CNS repair, regrowth of retinal ganglion cell (RGC) axons after ON crush can be enhanced using a variety of ther- apeutic manipulations (Harvey et al., 2006; Berry et al., 2008; Watanabe, 2010; Fischer and Leibinger, 2012). In mice this is most effective after conditional retinal deletion of phosphatase and tensin homologue (PTEN) and in some cases additional deletion of suppres- sor of cytokine signalling 3 (SOCS3) (Sun et al., 2011; de Lima et al., 2012; Luo et al., 2013). At present this latter type of approach, while important and instructive, is species-specific and dependent on the generation of floxed mouse lines and the use of adeno- associated viral (AAV) vectors to express cre to delete relevant gene(s). It is difficult to translate to different species and to other CNS injury models. Experimental Neurology 279 (2016) 197–211 Abbreviations: AAV, adeno-associated virus; AChE, acetyl cholinesterase; BDNF, brain- derived neurotrophic factor; CNTF, ciliary neurotrophic factor; CTB, cholera toxin B sub- unit; GFP, green fluorescent protein; ON, optic nerve; PN, peripheral nerve; RGC, retinal ganglion cell; SC, superior colliculus; SGS, stratum griseum superficiale. ☆ Funded by the WA Neurotrauma Research Programme, an NHMRC Senior Research Fellowship APP1002258 to JR, and the National Natural Science Foundation of China (no. 81470631) awarded to S-W Y. ⁎ Corresponding author at: School of Anatomy, Physiology and Human Biology, The University of Western Australia, M309, 35 Stirling Highway, Perth, WA 6009, Australia. E-mail address: [email protected] (A.R. Harvey). 1 Co-first authors. http://dx.doi.org/10.1016/j.expneurol.2016.03.006 0014-4886/© 2016 Elsevier Inc. All rights reserved. Contents lists available at ScienceDirect Experimental Neurology journal homepage: www.elsevier.com/locate/yexnr
Transcript of Large-scale reconstitution of a retina-to-brain pathway in adult rats ...

Experimental Neurology 279 (2016) 197–211
Contents lists available at ScienceDirect
Experimental Neurology
j ourna l homepage: www.e lsev ie r .com/ locate /yexnr
Large-scale reconstitution of a retina-to-brain pathway in adult rats usinggene therapy and bridging grafts: An anatomical andbehavioral analysis☆
Si-Wei You d,1, Mats Hellström e,1, Margaret A. Pollett a, Chrisna LeVaillant a, Colette Moses a, Paul J. Rigby c,Marissa Penrose b, Jennifer Rodger b,f, Alan R. Harvey a,f,⁎a School of Anatomy, Physiology and Human Biology, The University of Western Australia, Australiab School of Animal Biology, The University of Western Australia, Australiac Centre for Microscopy, Characterisation and Analysis, The University of Western Australia, Australiad Department of Ophthalmology, Xijing Hospital, The Fourth Military Medical University, Xi’an, Chinae Laboratory for Transplantation and Regenerative Medicine, Dept. of Obstetrics and Gynecology, Sahlgrenska Academy, University of Gothenburg, Swedenf Western Australian Neuroscience Research Institute, Australia
Abbreviations:AAV, adeno-associated virus; AChE, acederived neurotrophic factor; CNTF, ciliary neurotrophic funit; GFP, green fluorescent protein; ON, optic nerve; PNganglion cell; SC, superior colliculus; SGS, stratum griseum☆ Funded by the WA Neurotrauma Research Programm
Fellowship APP1002258 to JR, and the National Natural S81470631) awarded to S-W Y.⁎ Corresponding author at: School of Anatomy, Physi
University of Western Australia, M309, 35 Stirling HighwaE-mail address: [email protected] (A.R. Harvey
1 Co-first authors.
http://dx.doi.org/10.1016/j.expneurol.2016.03.0060014-4886/© 2016 Elsevier Inc. All rights reserved.
a b s t r a c t
a r t i c l e i n f oArticle history:Received 5 February 2016Received in revised form 1 March 2016Accepted 2 March 2016Available online 10 March 2016
Peripheral nerve (PN) grafts can be used to bridge tissue defects in the CNS. Using a PN-to-optic nerve (ON) graftmodel, we combined gene therapy with pharmacotherapy to promote the long-distance regeneration of injuredadult retinal ganglion cells (RGCs). Autologous sciatic nerve was sutured onto the transected ON and the distalend immediately inserted into contralateral superior colliculus (SC). Control rats received intraocular injectionsof saline or adeno-associated virus (AAV) encoding GFP. In experimental groups, three bi-cistronic AAV vectorsencoding ciliary neurotrophic factor (CNTF) were injected into different regions of the grafted eye. Each vectorencoded a different fluorescent reporter to assess retinotopic order in the regenerate projection. To encouragesprouting/synaptogenesis, after 6 weeks some AAV–CNTF injected rats received an intravitreal injection of re-combinant brain-derived neurotrophic factor (rBDNF) or AAV–BDNF. Four months after surgery, cholera toxinBwas used to visualize regenerate RGC axons. RGC viability and axonal regrowth into SCwere significantly great-er in AAV–CNTF groups. In some cases, near the insertion site, regenerate axonal density resembled retinal termi-nal densities seen in normal SC. Complex arbors were seen in superficial but not deep SC layers and manyterminalswere immunopositive for presynaptic proteins vGlut2 and SV2. Therewas improvement in visual func-tion via the grafted eye with significantly greater pupillary constriction in both AAV–CNTF+BDNF groups. Inboth control and AAV–CNTF+rBDNF groups the extent of light avoidance correlated with the maximal distanceof axonal penetration into superficial SC. Despite the robust regrowth of RGC axons back into the SC, axonsoriginating from different parts of the retina were intermixed at the PN graft/host SC interface, indicating thatthere remained a lack of order in this extensive regenerate projection.
© 2016 Elsevier Inc. All rights reserved.
Keywords:Gene therapyPeripheral nerve transplantationAdult rat visual systemAxonal regenerationSuperior colliculusNeurotrophic factorsRetinal ganglion cells
1. Introduction
Injury to the adult mammalian central nervous system (CNS) re-sults in long-lasting functional deficits, a consequence of intrinsic
tyl cholinesterase; BDNF, brain-actor; CTB, cholera toxin B sub-, peripheral nerve; RGC, retinalsuperficiale.e, an NHMRC Senior Research
cience Foundation of China (no.
ology and Human Biology, They, Perth, WA 6009, Australia.).
and extrinsic factors that together impact on plasticity and regener-ative ability. In the rodent primary visual system, often used as an ex-perimental model of CNS repair, regrowth of retinal ganglion cell(RGC) axons after ON crush can be enhanced using a variety of ther-apeutic manipulations (Harvey et al., 2006; Berry et al., 2008;Watanabe, 2010; Fischer and Leibinger, 2012). In mice this is mosteffective after conditional retinal deletion of phosphatase and tensinhomologue (PTEN) and in some cases additional deletion of suppres-sor of cytokine signalling 3 (SOCS3) (Sun et al., 2011; de Lima et al.,2012; Luo et al., 2013). At present this latter type of approach,while important and instructive, is species-specific and dependenton the generation of floxed mouse lines and the use of adeno-associated viral (AAV) vectors to express cre to delete relevantgene(s). It is difficult to translate to different species and to other CNSinjury models.

198 S.-W. You et al. / Experimental Neurology 279 (2016) 197–211
Another way of enhancing mammalian RGC regeneration is to graftsegments of peripheral nerve (PN) on to the cut ON. Regeneration ofadult RGC axons through these grafts has been achieved in wild-typerats (Vidal-Sanz et al., 1987; Villegas-Perez et al., 1988; Thanos, 1992;Avilés-Trigueros et al., 2000), hamsters (Carter et al., 1989, 1994;Sasaki et al., 1999), mice (Cui and Harvey, 2000), ferrets (Quan et al.,1999) and cats (Watanabe and Fukuda, 2002). When the distal end ofPN grafts is inserted into visual centers in the rat or hamster brain,RGC axons re-establish synaptic contacts with central target neuronsand some non-patterned visual behaviors have been restored(Vidal-Sanz et al., 1987; Keirstead et al., 1989; Carter et al., 1989,1994; Sauvé et al., 1995; Sasaki et al., 1999; Heiduschka and Thanos,2000; Vidal-Sanz et al., 2002), however there is little evidence of resto-ration of higher level patterned visual function or restitution of visualtopography (Sauvé et al., 2001).
In these earlier rodent studies, the PN graft approach was rarelycombined with other growth-enhancing therapeutic strategies. Usuallyless than 20% of the RGC population remained alive, and perhaps 10–20% of these surviving neurons regenerated an axon (about 2000 RGCaxons). Thus the number of axons re-entering host visual centers wassmall and maximal extent of outgrowth was limited. Furthermore, inmany studies PN grafts were inserted after a several week delay intoipsilateral visual centers, and regenerative growth via PN grafts wasenhanced by removal of the remaining normal eye, a procedure thatobviously cannot be applied in any clinical context.
In adult rat studies, by combining pharmacotherapy and AAV genetherapy to deliver a secretable form of ciliary neurotrophic factor(CNTF) to injured RGCs, on average about 10,000 RGCs can be inducedto regrow axons at least 1–1.5 cm into blind-ended PN grafts (Leaveret al., 2006; Hellström and Harvey, 2011, 2014). In the present study,an autologous sciatic nerve was sutured onto the transected ON, thedistal end inserted acutely into contralateral superior colliculus (SC)and AAV–CNTF vectors injected into the eye at the time of surgery(Hellström et al., 2011b). To track and identify retinotopically distinctRGC axons, and thus reveal any topographic order in the regenerate ret-inal projection,we used three bi-cistronic AAV–CNTF vectors expressingdifferent fluorescent reporters, each injected into different regions ofthe grafted eye.
AAV–CNTF-injected animals were allocated to 3 groups. The firstgroup received no further intervention. In the other groups, to encour-age sprouting and synaptogenesis within the SC (Sawai et al., 1996;Cohen-Cory et al., 2010), the grafted eye was additionally injectedwith recombinant brain-derived neurotrophic factor (rBDNF) or AAV–BDNF 6 weeks after the initial surgery. For all animals, visual behaviorsmediated by the grafted eye were tested at 15 weeks post-transplantation, and the total regenerate projection was visualized byinjecting grafted eyeswith the anterograde tracer cholera toxin B (CTB).
2. Materials and methods
Forty-six PVG/c rats (female, 8–10 weeks of age, Animal ResourcesCentre, WA) were used in this study. One additional rat was used as anon-operated control. The animals were housed in standard sizedcages with filtered tops along with sterile food and water access adlibitum in a 12-h light–dark cycle (red lighting: 10 am to 10 pm, whitelighting: 10 pm to 10 am). Care and treatment of all animals were instrict accordance with NHMRC guidelines and all experimental proce-dures were approved by The University of Western Australia AnimalEthics Committee. The overall experimental plan, including details ofgroups, is shown in Fig. 1.
2.1. Viral vectors
Recombinant AAV2 vectors were produced by Vector Biolabs(Philadelphia, USA) from plasmid pTRUF12 containing BDNF and plas-mid pTRUF12.1 (gift, Prof. Joost Verhaagen) containing a secretable
form of CNTF (Sendtner et al., 1992) linked via IRES to 3 different fluo-rescent reporter proteins; green fluorescent protein (GFP), mCherryand Venus (gift, RIKEN Brain Science Institute, Japan). Both plasmidscontained the cytomegalovirus-chicken beta actin promoter, butpTRUF12.1 lacked an intron sequence to enable packaging of the largerCNTF construct. Successful expression of the three reporters was seen inHEK-293T cells using lipofectamine mediated delivery of each plasmid.A control AAV2–GFP vector was also produced using the pTRUF12.1plasmid.
The AAV–BDNF–GFP and AAV–CNTF–GFP vectors were successfullycharacterized previously (Leaver et al., 2006; Hellström and Harvey,2011).We confirmed functionality of the newAAV-vectors by transduc-ingHEK-293T cells in vitro under standard culturing conditionswith re-duced fetal calf serum (FCS; 2%). After 5 days, cells were visualizedunder fluorescence and harvested for mRNA (qPCR) and protein(ELISA) analysis.
Total RNAwas extracted from cells using standard Qiagen protocols.Validated primer pairs were used to quantify mRNA transcript expres-sions of BDNF, CNTF, mCherry and GFP, and internal reference genesPpia, TBP, and YWAHZ. qPCR runs were performed on the Rotor-Gene6000 (Qiagen, USA), using Bio-Rad iQ SYBR 2× Mastermix (Bio-Rad,Australia). Normalized data were analyzed for changes between groupsby Kruskal–Wallis ANOVA, and correlations confirmed using Biogazellesoftware. ELISAwas used to detect BDNF and CNTF release from the cellsafter AAV transduction. For BDNF detection ChemiKine BDNF SandwichELISA (Millipore Bioscience Research Reagents) was used, and for CNTFmeasurements the TSZ ELISA kit (Waltham. MA Cat No M1473) wasused, in both cases followingmanufacturer's instructions. Optical densi-ty was determined in a plate reader set at 450 nm. Sample values werecalculated from the standard curve. Optical densitywas a linear functionof concentration between 15 and 1000 pg ml−1 with a coefficient ofvariation b10%.
2.2. Surgical procedures
Rats were anesthetized with a 1:1 mixture (1.5 ml/kg) of ketamine(100 mg/ml) and xylazine (20 mg/ml). Eye ointment containing atro-pine sulphate (10 mg/g, Troy Ilium) was applied to protect the corneaduring surgery. The sciatic nerve was exposed in the left leg and a seg-ment of about 3.5 cm was isolated. The leg wound was moistenedwith sterile saline and margins temporarily re-opposed to keep the iso-lated peripheral nerve (PN) in situ until the grafting procedure. The leftON was exposed by protruding the eye and opening the conjunctivum.The ONwas completely severed about 1.5mm from the optic disc, whiletaking care not to disrupt the blood flow to the retina. The PN segmentwas removed from the leg and the proximal end was transplanted ontothe ON stump (end-to-end) using a single suture (10-0 Ethilon, Johnson& Johnson, North Ryde, NSW, Australia).
The head of the animal was then fitted in a head holder using amax-illary clamp and part of the right posterior hemispherewas exposed andaspirated to reveal the surface of the superior colliculus (SC). The PNwas then laid on the skull and the distal end was prepared so that indi-vidual nerve bundles protruded from the epineurial sheath. The bundleswere teased apart prior to gentle insertion into the rostral part of theright SC just below the pial surface using a sterile glass pipette(Fig. 2A, B). The surgical cavity was packedwith small pieces of gelfoamand the overlying scalp was closed with sterile fine sutures (6-0). Thenthe leg wound was closed with sterile wound clips. Rats also received asubcutaneous injection of buprenorphine (0.02 mg/kg, Temgesic;Reckitt & Colman, Hull, UK) followed by Carprofen (5 mg/kg, Provet,WA an intramuscular injection of Benacillin (0.3 mg/kg, Troy Ilium;NSW, Australia). Rats were placed on a heat pad andmonitored until re-covery from the anesthesia at regular intervals thereafter. The followingparameters were checked regularly post-surgery: signs of post-operative infection or bleeding, patency of sutures, self-grooming, stableor weekly weight gain, general alertness and responsiveness, normal

Fig. 1.Overall plan, detailing protocols for each of the four experimental groups. AAV–CNTF, adeno-associated viral vector encoding a secretable form of ciliary neurotrophic factor; rCNTF,recombinant CNTF; CPTcAMP, 8-(4-Chlorophenylthio)-adenosine-3′,5′-cyclic monophosphate, a cell permeable analogue of cyclic AMP; AAV–BDNF, AAV encoding brain-derivedneurotrophic factor.
199S.-W. You et al. / Experimental Neurology 279 (2016) 197–211
communal behavior and activity, signs of autotomy (self-inflicted dam-age to toes of left, operated hindlimb).
2.3. Eye injections
At the time of the PN surgery (rats still under ketamine/xylazine an-aesthesia), the grafted eye was injected with either saline (n = 6) orAAV–GFP (n = 6) (both 4 μl, group 1), or AAV–CNTF (groups 2–4)(Fig. 1). For all eye injections (see also below) care was taken to mini-mize risk of damaging the lens. For the AAV–CNTF injections, 1.5 μl ofeach the three different bi-cistronic vectors (AAV–CNTF–GFP, AAV–CNTF–mCherry and AAV–CNTF–Venus) was injected intravitreally(titer for each: 5 × 1012, 2.5 × 1013, 3.7 × 1013 gc/ml respectively).AAV–CNTF–GFP was injected into the ventro-lateral part of eye, AAV–CNTF–mCherry into the ventro-nasal region, and AAV–CNTF–Venuswas injected dorsally. This procedure resulted in transduction of differ-ent populations of regenerate RGCs with different reporter genes.
In the three AAV–CNTF injected groups, to provide interim trophicsupport prior to onset of vector-mediated gene expression, rCNTF pro-tein and a cAMP analogue (8-(4-Chlorophenylthio)-adenosine-3′,5′-cy-clic monophosphate; CPT-cAMP), a combination known to promoteRGC viability and axonal regrowth (Cui et al., 2003; Park et al., 2004;Hellström and Harvey, 2014), was injected intravitreally at 3 and10 days post-injury (rCNTF, 0.5 μg/μl, Peprotech, Rehovot, Israel; CPT-cAMP, 0.1 mM, Sigma) (Hellström et al., 2011a, 2011b). The remaininggroup (group 1, initial saline or AAV–GFP injection) received injections
of 3 μl saline at 3 and 10 days after surgery. At 6 weeks, underisofluorane anaesthesia, all AAV–CNTF injected rats received a furtherleft eye injection of either saline (n = 11) (4 μl, groups 1 and 2), AAV–BDNF (n = 11) (4 μl, 5 × 1013 gc/ml, group3) or rBDNF (Peprotech,4 μg in 4 μl, group 4) (n= 12) (Fig. 1). Animals were then left for a fur-ther 9–10 weeks before behavioral analysis (Fig. 1).
2.4. Behavioral testing
Prior to testing visual behaviors mediated via the grafted eye, visionthrough the right unoperated eye was temporarily obstructed by sutur-ing the eyelids closed using three to four 6/0 sutures (Ethicon) underisofluorane anesthesia. All tests were carried out by two observersblinded to the experimental group being tested.
2.4.1. Light–dark exploration testA light–dark tunnel was used to investigate locomotor activity and
light sensitivity. A 2 m long enclosed Perspex tunnel (approx15 cm × 15 cm square) was divided into a light (transparent) half anda dark (black) half. Each rat was placed at the furthest end of the lighthalf and the tunnel was shut at both ends with sliding Perspex doors.Over the course of 10 min the number of times the animal crossedfrom the light to the dark sidewas recorded, aswell as the total amountof time the rat spent in the light.

Fig. 2. A, Schematic view of surgical design. The peripheral nerve (PN) graft was sutured onto the left severed optic nerve, placed over the skull and the distal end inserted into thesuperficial superior colliculus (SC) on the contralateral (right) side. LGN, lateral geniculate nucleus. B, view of PN graft successfully inserted into the right SC (arrowhead). IC, inferiorcolliculus. Rostral is to the left. C, longitudinal section of a PN graft showing that the transplant is completely filled with regenerate, cholera toxin B (CTB) labelled RGC axons(peroxidase stain). D, CTB Alexa 647 labelled RGC axons in PN bridging graft (D1), some of which are also immunostained for either mCherry (red) or GFP/Venus (green) (D2, samefield), indicating transduction of parent RGCs with either AAV–CNTF–mCherry or AAV–CNTF–GFP/Venus. Scales: B = 2 mm; C = 200 μm; D = 100 μm.
200 S.-W. You et al. / Experimental Neurology 279 (2016) 197–211
2.4.2. Pupillary responseRats were acclimatized in the dark (4 lx) for aminimumof 30min to
allow maximum pupillary dilatation and were video recorded using aninfrared light source camera. While recording, bright light was intro-duced (1000 lx) or removed instantaneously using a DC light sourceand the pupillary reaction was recorded for 15 min after the stimulus.Images from the video were digitized and stills were captured every5–10 s to acquire a baselinemeasurement.When the lightwas switchedon, stills were taken at intervals ranging from 40 ms to 1 s (thisdepended on the animal's movement and the focus of the image – notall images could be used) for the first 10 s, at which point pupil sizehad plateaued and measurements returned to every 5–10 s. Measure-ments of pupil and iris areas were taken using ImageJ software andthe iris/pupil ratio was plotted against time.
2.4.3. Optokinetic responseRats were placed on a round platform (9 cm diameter, 14 cm high)
surrounded by a cylinder (29 cmdiameter, 29.7 cmhigh), thewalls cov-ered by stripes of different thickness. The striped cylinder was rotatedclockwise then anticlockwise at a speed of 2 revs per minute for 2 minin each direction. A pause of 30 s was inserted between the 2 directions.Behavior was recorded using a video camera positioned above the rat.The number and duration of any head movements to track the movingstripes were analyzed offline.
2.5. CTB injections and perfusion
When behavioral testing was complete, the sutures over theunoperated eye were removed and under isofluorane anesthesia the
grafted eye was injected with 3 μl of 0.2% CTB conjugated to a far redfluorophore (Alexa Fluor 647; Life Technologies C34778). Six daysafter the CTB injection, rats were deeply anesthetized (Lethabarb,Virbac, NSW; i.p; 200 μl/100 g) and transcardially perfused with coldphosphate-buffered saline (PBS) (pH 7.4) containing 0.1% heparin,then a phosphate buffer containing 4% paraformaldehyde. The retinasfrom grafted eyes were removed from their eyecup and postfixed for1 h in the same fixative. The PN grafts were also dissected away toallow visualization of AAV–CNTF transduced and CTB labelled regener-ate RGC axons within the bridge. Finally, the PN graft was carefully cutto ensure that the distal 3–4 mm of each graft remained attached tothe right host SC (Fig. 2B) and could therefore be cut along with thehost SC. Nerves and brains were post-fixed in the same fixative for 1and 2 h respectively. After perfusion, one control AAV–GFP injectedrat was found to have a broken PN graft. This animal was not processedfor histological analysis.
2.6. Histological processing
2.6.1. RetinasWhole mount retinas were washed once in PBS and blocked with
antibody diluent (10% normal goat serum and 0.2% Triton-X in PBS)for 1 h at RT, followed by incubation with primary antibodies in anti-body diluent overnight at 4 °C. Retinas were washed with PBS(3 × 10 min) and incubated with secondary antibodies in antibody dil-uent for 2 h at room temperature (RT). Sectionswere thenwashedwithPBS (3 × 10 min) and coverslipped with fluorescent mounting media(Dako). Primary antibodies used for fluorescent immunohistochemistrywere rabbit anti-β-tubulin III (Covance PRB-435P, 1:2000), chickenanti-GFP (Abcam 13970; 1:500) and mouse anti-mCherry (Clontech

201S.-W. You et al. / Experimental Neurology 279 (2016) 197–211
Living Colors® 632543; 1:600). Secondary antibodies usedwere donkeyanti-rabbit AMCA (Jackson JI711156152; 1:400), goat anti-chickenAlexaFluor 488 (Life Technologies A11039; 1:400), and goat anti-mouse AlexaFluor 568 (Life Technologies A11031; 1:400).
2.6.2. Peripheral nervesPN graft segments were laid flat on a rectangle of 2% agarose which
was adhered to a piece of cardboard with OCT. The entire nerve wascovered in OCT and frozen in isopentane over dry ice, then stored at−20 °C until required. Longitudinal PN sections (16 μm) werecryosectioned, collected onto double subbed slides and stored at−20 °C. For immunofluorescence, sections air dried for 30 min at RTwere blocked with antibody diluent (10% normal goat serum and 0.2%Triton-X in PBS) for 1 h followed by incubation with primary antibodiesin antibody diluent overnight at 4 °C. Sections were washed with PBS(3 × 10 min) and incubated with secondary antibodies in the same dil-uent for 2 h at RT. Sectionswere thenwashedwith PBS (3× 10min) andcoverslipped with fluorescent mounting media (Dako). Because thelevel of intrinsic fluorescence was not consistent, sections were immu-nostained using antibodies tomCherry andGFP (the latter antibody rec-ognizes both GFP and the related Venus fluorophores). Primaryantibodies used were mouse anti-mCherry (Clontech, 1:1200), rabbitanti-GFP (Millipore AB3080; 1:500) and chicken anti-GFP (Millipore,1:200). Secondary antibodies were goat anti-mouse AlexaFluor 568(Life Technologies, 1:400), goat anti-rabbit AlexaFluor 488 (Life Tech-nologies A11008; 1:400) and goat anti-chicken AlexaFluor 488 (LifeTechnologies A11039; 1:400).
For CTB immunoperoxidase, sections were washed once with PBSand then incubated in 0.6% hydrogen peroxide in PBS for 20 min. Sec-tions were washed with PBS (3 × 10 min) and incubated overnight at4 °C with anti-CTB antibody (List Biolabs #703), 1:8000 in diluent con-taining 2% normal horse serum, 2.5% BSA and 2% Triton X-100. Sectionswere again washed 3 timeswith PBS and incubated on a shaker at roomtemperature for 2 h with biotinylated anti-goat IgG secondary antibody(Vector labs BA-9500, 1:800 in the same diluent as before). After anoth-er 3 PBS washes, sections were incubated for 1 h with Vectastain EliteABC reagent, (Vector Labs PK-6100, 1:200). After further PBS washes,sections were reacted with Pierce DAB metal enhanced substrate(Thermo Scientific, 34002) for 1 min. Sections were washed, mountedand coverslipped in DePeX.
2.6.3. BrainsBrains were sunk in sucrose overnight, then blocked out in egg yolk
whichwas set in 10% formalin vapor, again overnight. Sectionswere cutsagittally at 40 μm thickness and immunochemistry performed on freefloating sections. For CTB immunoperoxidase, sections were processedexactly as described above for PN graft segments.
For immunofluorescence, sectionswere blockedwith antibody dilu-ent (10% normal goat serum and 0.2% Triton-X in PBS; except for CTB)for 1 h at RT, followed by incubation with primary antibodies in anti-body diluent overnight at 4 °C. Sections were washed with PBS(3 × 10 min) and incubated with secondary antibodies in antibody dil-uent for 2 h at RT. Primary antibodies used were: rabbit anti-GFP(1:400), mouse mCherry (1:600), rabbit anti-glial fibrillary acidic pro-tein (GFAP) (Dako Z0334; 1:200), rabbit anti-vesicular glutamate trans-porter (vGluT)1 (SYSY Cat No 135 303; 1:500) rabbit anti-vGlut2(DNPI)(SYSY Cat No 135 403; 1:500), mouse anti-SV2 (DSHB; 1:200),mouse anti-phosphacan (DSHB clone 3F8; 1:100), rabbit anti-laminin(Sigma L9393, 1:500) and goat anti-CTB (1:2000). Secondary antibodiesused were goat anti-rabbit Alexa 488 (1:400), goat anti-mouse Cy3(Jackson; 1:300), anti-rabbit Cy3 (Jackson; 1:300) and goat anti-mouse AlexaFluor 568 (1:400).
When CTB antibody was used for immunofluorescence, sectionswere first treated with TRIS–EDTA for 3 min at 100 °C and washed be-fore blocking with antibody diluent (5% normal horse serum, 2% TritonX and 2.5% BSA). After overnight primary antibody incubation at 4 °C,
sectionswerewashed 3 timeswith PBS and incubatedwith biotinylatedanti-goat antibody (Vector BA9500; 1:800) at room temperature for 2 h.After further PBS washes they were incubated with streptavidin-FITC(Jackson Labs 016-010-084; 1:100) at room temperature for 2 h. Sec-tions were then washed with PBS (3 × 10 min) and coverslipped withfluorescent mounting media (Dako). To help delineate the superficiallayers of the SC (SGS), some sagittal sectionswere stained for acetylcho-linesterase (AChE) using an established protocol described in detailelsewhere (Harvey and MacDonald, 1985).
2.7. Confocal microscopy
Retinal wholemounts and tissue sections were imaged with a NikonA1Si confocalmicroscope using a Nikon 60×NA 1.40 PlanApoVC oil im-mersion objective. Samples were simultaneously excited with 405 nmand 488 nm lasers and fluorescence emission detected using the spec-tral detector over the range 410 nm to 600 nmwith a 10 nmgrating res-olution and either 8 or 16 line averages. Fluorescent probes werespectrally unmixed in Nikon NIS Elements software (version 4.20)using single label control samples for reference spectra. For tissue sec-tions, Z stacks were collected at 0.5 μm slice intervals through the entiresection thickness and maximum intensity projections prepared for pre-sentation. Images were collected sequentially and single stain controlswere imaged to ensure there was no spectral bleed through.
2.8. Quantitative analyses
2.8.1. RGC countsRGCs were counted in wholemounts immunostained for β-III tubu-
lin. At least 12 fields (each 0.14 mm2) were photographed, the fieldsrandomly chosen from all parts of the retina. RGC counts were madefrom these images rather than live from the wholemounts because offading of the AMCA fluorescence. In the images some regions of the ret-inawere not in focus; thesewere not included in the sampled areamea-surements and RGCs could not be counted. On average, 11.6 fields werecounted, or about 3% of the total retinal area.
2.8.2. Mapping of regenerate retinal terminals in the SCDrawings were made of every sixth CTB stained sagittal 40 μm sec-
tion through the reinnervated SC. For each section,wemarked themax-imum distance from the PN graft insertion site reached by regenerateRGC axons, in both superficial SC and in deeper non-visual SC and teg-mentum. The maximum distance grown by regenerating axons was re-corded for each animal. To estimate the proportion of superficial SC thatwas reinnervated in each rat, the drawings of axons/terminal arbors inthe superficial SC layers were reconstructed into a dorsal view of theright SC, and the location of the PN insertion noted. For each SC recon-struction, using ImageJ software a polygon selection was made of anarea bounding all axons/terminal arbors. This area was then expressedas a % of the total superior colliculus area.
2.9. Statistics
RGC survival and the extent of regeneration (maximumdistance andarea reinnervated) were analyzed by ANOVA followed by post hoc(Tukeys) pairwise comparisons. In the light dark tunnel, the total num-ber of crosses and % time spent in light was analyzed by one wayANOVA, followed by post hoc (Tukey's) pairwise comparison. Pupillaryresponse was analysed by repeated measures ANOVA with time as therepeatedmeasure. The optokinetic response was not obvious in any an-imals and data were not analyzed further. Correlations were analyzedusing the restricted maximum likelihood method in JMP software. Forall analyses, significance was set at p b 0.05.

202 S.-W. You et al. / Experimental Neurology 279 (2016) 197–211
3. Results
Strong post-IRES expression of Venus, mCherry and GFPwas obtain-ed in cultured HEK293T cells transfected with the respective plasmidscarrying the transgene (Fig. 3A). AAV-transgene induced fluorescencewas confirmed 3 and 5 days after transduction of HEK293T cellsin vitro (not shown). qPCR data revealed a 15-fold increase in BDNFmRNA expression in the AAV–BDNF transduced cells compared tonon-transduced hek293T cells. CNTF mRNA expression was increased
Fig. 3.AAV–CNTF bicistronic vectors used for intravitreal injectionwere linked via IRES to threecells expressing high levels of each fluorescent protein after transduction using plasmids and liRGCs in an AAV–CNTF injected retina almost 4months after optic nerve transection and PN tranAAV–CNTF–Venus (c1–c3), AAV–CNTF–mCherry (c4) or AAV–CNTF–GFP (c5–c6). Plates c2, c3,tibodies to either mCherry or GFP (see text). The c2 plate is a confocal image showing intrinsiceach retina is shown by the arrows; dorsal, lower nasal (N) and lower temporal (T) respectivelyAAV–GFP group (control). In these box plots the bottom and top lines represent the first and thshown by the continuous line.
19- (AAV–CNTF–GFP), 21- (AAV–CNTF–mCherry) and 23-fold (AAV–CNTF–Venus) compared to non-transduced HEK293T cells. ELISA con-firmed release of both BDNF and CNTF from AAV transduced HEK-293T cells (not shown). All bi-cistronic CNTF vectors showed expressionand secretion – ranging from 30 to 40 pg/ml in conditioned media. Invivo, these vectors were intravitreally injected into dorsal, ventro-nasal and ventro-temporal regions of PN grafted eyes respectively,resulting in transduction of essentially distinct populations of RGCs indifferent parts of the retina (Fig. 3B).
different reporter genes (Venus,mCherry or green fluorescent protein –GFP). A, HEK-293Tpofectamine. B, retinal wholemount showing a high density of viable β-III tubulin positivesplantation. C, retinal wholemount image (rat #29) and examples of RGCs transducedwithc6 show intrinsic fluorescence, c1, c4, c5 are fromwholemounts immunostained with an-yellow Venus fluorescence at 528 nm. The approximate location of each reporter label in. D, the number of surviving RGCs in the three AAV–CNTF groups and combined saline andird quartiles and the line inside the box is the median. The overall mean for all animals is

203S.-W. You et al. / Experimental Neurology 279 (2016) 197–211
3.1. Retinal ganglion cell viability
Retinal wholemounts were immunostained for β-III tubulin(Fig. 3C), permitting an estimate of RGC viability in each animal fromeach group (Fig. 3D). On average there were 198.8 (±27.2 SEM)RGCs/mm2 and 202.5 (±29.8 SEM) RGCs/mm2 in the saline and AAV–GFP control groups respectively. These values were not significantly dif-ferent from each other. Pooling both control groups gave a mean valueof 201 RGCs/mm2, or about 11,600 viable RGCs per retina. Average num-bers of surviving RGCs 16 weeks after PN transplantation in the AAV–CNTF+ saline, AAV–CNTF+AAV–BDNF and AAV–CNTF–rBDNF groupswere 493.5 (±20.1 SEM) RGCs/mm2, 474.3 (±20.1 SEM) RGCs/mm2,and 422.6 (±19.2 SEM) RGCs/mm2 respectively (Fig. 3D). All valueswere significantly higher than both control groups (p b 0.05), butthere was no significant difference between the three AAV–CNTFgroups. Thus the level of RGC viability is established in the first fewdays after the PN–ON graft procedure as a consequence of the initialpharmacotherapy and AAV–CNTF injections, and later application ofBDNF does not further influence long-term RGC survival. The highest
Fig. 4. Examples of peripheral nerve (PN) bridge insertion into the contralateral superficial supcholera toxin B (CTB) into the grafted eye. CTB axons were identified using peroxidase immunreceived an AAV–BDNF injection 6 weeks later; D, E, AAV–CNTF injected rat (#13) that receivalways restricted to the superficial layers (stratum griseum superficiale, SGS) of the SC imminnervated by retinal axons. The SGS is easily recognizable in adjacent sections stained for aceshown in A. F–H shows a PN graft (rat #38) which likely failed because a scar formed aroundintense laminin immunoreactivity (arrow in G) and increased inhibitory proteoglycan deposA2 = 300 μm; A1 and A2 = 200 μm.
RGC density in any animal (AAV–CNTF + saline group) was617 RGCs/mm2, or about 35,800 viable RGCs (one third of the initialganglion cell population) (Hellström and Harvey, 2011).
3.2. Regeneration of RGC axons
3.2.1. PN graftsThe impact of various treatments on RGC regeneration was first
assessed in longitudinal sections of the PN bridging grafts. CTB labelledaxons were seen in control saline and AAV–GFP injected animals butnumbers were considerably less than that seen in all AAV–CNTF groups(Fig. 2C, D1). Inmost of these animals the PN graftswere densely packedwith regenerate axons. RGC axonal density remained high at the distalend of the grafts inserted into the SC (Fig. 4). In four rats (1 saline, 1AAV–CNTF + AAV–BDNF, 2 AAV–CNTF + rBDNF) CTB label was weakwith few labelled axons. In the 3 AAV–CNTF animals RGC viability wasnot obviously different from other animals in their respective groups,thus the sparse CTB label is regarded as indicative of poor intraocular in-jections and reduced RGC uptake of the anterograde marker. Finally,
erior colliculus (SC); regenerated RGC axons were anterogradely labelled after injection ofohistochemistry. A–B, AAV–CNTF injected rat (#30); C, AAV–CNTF injected rat (#37) thated a recombinant BDNF injection 6 weeks later. Note the extremely dense reinnervationediately adjacent to the PN insertion (arrows: A, C, D). These are the layers normally
tylcholinesterase (B, E). A1 and A2 are higher power images of the regenerate innervationthe insertion site. Many RGC axons can be seen at the distal end of the graft (F), there isition (phosphacan, arrow in H). SO, stratum opticum. Scales for all figures except A1 and

204 S.-W. You et al. / Experimental Neurology 279 (2016) 197–211
immunostaining of grafts for mCherry or GFP/Venus revealedintermixing of axons from disparate regions of the retina (Figs. 2D2, 3).
3.2.2. Entry of regenerating RGC axons into contralateral midbrainFig. 4 shows examples of regenerating CTB-labelled RGC axons at the
site of re-entry into the contralateral SC, from three different AAV–CNTFinjected animals. In these examples the distal end of each PN graft waslocated primarily in superficial SC. Note the extremely dense reinnerva-tion immediately adjacent to the PN insertion in each case (Fig. 4A, C, D).These localized regions of dense reinnervationwere only seen in the su-perficial layers (stratum griseum superficiale, SGS) of the SC, and mostcommonly in the AAV–CNTF + saline and AAV–CNTF + rBDNF groups(Table 1). Dense or moderate/dense patches were seen in 7/11 (64%)rats in the AAV–CNTF + saline group and in 6/8 (75%) rats in theAAV–CNTF + rBDNF group. Interestingly such patches were less fre-quent in rats with secondary intravitreal injection of AAV–BDNF (30%of cases, Table 1). Identification of SGS was facilitated by staining adja-cent sections for AChE (Fig. 4B, E), an established marker for this layer(Harvey and MacDonald, 1985). In normal animals, the superficiallayers of the SC are the primary target for retinal axons, indicating selec-tivity in the reinnervation of the deafferented SC.
Immunostaining for laminin revealed intense staining in the PNgrafttissue but in most cases no evidence of a confining boundary betweengraft and host SC. On occasion however, the PN graft ended in a bulb,with thousands of regenerating RGC axons apparently trapped withinthe PN tissue (Fig. 4F). In this example (from the AAV–CNTF + rBDNFgroup) there is a clear laminin scar surrounding the PN–SC interface(arrow, Fig. 4G) which is also highly immunoreactive for the inhibitory
Table 1Summary showing the consistency and extent of regeneration of retinal ganglion cell axons intoof axons adjacent to the peripheral nerve graft insertion and scattered elsewhere in the SC is indsections). The furthest extent of outgrowth in superficial and deep SC is also shown. SGS, strat
Dense near insertion SGS - # sections
Saline-no AAV No 1No 0Low 2No 3
AAV–GFP only No 4Moderate 3Low 3No 2No 1
AAV–CNTF+saline No 0Dense 7Low 6Moderate/dense 11Low 7Dense 7Moderate/dense 6No 6Dense 7Moderate 8Moderate 7
AAV–CNTF+AAV–BDNF Dense 6Low 6No 3No 3Moderate 4Moderate 8No 5dense 6No 7Moderate/dense 6
AAV–CNTF+rBDNF Very dense 4Moderate/dense 5Moderate 4Small–dense 4Very dense 8Low 7Moderate/dense 7Moderate/dense 8
proteoglycan phosphacan (arrow, Fig. 4H). This type of interface wasseen in five PN-SC animals, but only in two caseswas there complete in-hibition of RGC outgrowth. In the others, because the distal PNwas sep-arated into various bundles prior to insertion, reinnervation of the SCwas seen although the density of reinnervation was likely reduced be-cause of obstruction of at least some regrown RGC axons.
3.2.3. Distribution of axons and terminal arborsFig. 5 shows examples of regenerative RGC growth in the contralat-
eral midbrain at sites distant from the PN–SC interface. Within the SC,complex terminal arbors laden with boutons were only seen in SGSand the stratum zonale (SZ). Heavy to moderate CTB label in the super-ficial SC is shown in Fig. 5A–D. Qualitatively, the density of scatteredRGC axons and terminals was similar in the AAV–CNTF + saline andAAV–CNTF + rBDNF groups (Table 1), Complex arbors were seen inall AAV–CNTF groups (Fig. 5D–G), although the most complex arborswere generally seen in animals that also received intraretinal supplyof BDNF (Fig. 5B, C, H). When PN grafts were inserted more deeplyinto the SC (Fig. 5H1) many axons regrew into intermediate and deepSC, but terminals were far less abundant here compared to the SGS(Fig. 5J). On occasion, individual arbors could be seen arising from singlenon-branched axons in the stratum opticum (SO) that abruptly turneddorsally to branch at different depths within the SGS (arrows, Fig. 5E–G). Such turnswere never observed to be oriented in a ventral direction.Fig. 5H1–H3 shows three different magnifications of a SC section from aPN grafted rat injected with AAV–CNTF and rBDNF. In this example RGCaxons grew out of the graft both superficially and deep, axons runningfor several mm from the PN–SC interface. Again, RGC fibers coursed
the contralateral superior colliculus (SC) in adult rats in each treatment group. The densityicated, as are the number of sagittal sections found to contain axons (1:6 series, 40 μmthickum griseum superficiale.
Scattered density SGS furthest Deep furthest
Very sparse 250 NoneNoVery sparse 150 800Very sparse 900 NoneSparse 1620 2000Very sparse 1030 1200Very sparse 800 950Very sparse 850 1100Very sparse 200 1080No (bulb)Moderate/high 1905 2170Low 1640 2535Moderate/high 1940 3010Moderate 2300 2580Low/moderate 1350 1640Low/moderate 1190 NoneModerate/high 2160 3580Moderate/high 1670 2800High 2030 2710Low 1430 NoneModerate 1270 2300Low/moderate 1340 1350Moderate/high 950 300Sparse 675 1100Low 1570 1350Moderate 1670 2180Low 1080 2400Low 1560 100Sparse 720 2740Low 1500 2080Moderate/high 1880 2700Moderate/high 2430 3280Low/moderate 2350 NoneLow 940 2080High 1680 2530Sparse (bulb) 700 1650Moderate 1900 3450Moderate/high 1920 2800

Fig. 5.A–H, Examples of re-establishment of complex axon arbors in the superficial superior colliculus. A, AAV–CNTF injected rat (#37) that received anAAV–BDNF injection 6weeks later;B–C, AAV–CNTF injected rats (#20, #13) that received a recombinant (r) BDNF injection 6 weeks later. D, moderate label in rat (#34) injected with AAV–CNTF followed by saline at6 weeks, E–G, individual arbors commonly arose from non-branched axons in the stratum opticum (SO) that suddenly turned dorsally to branch at different depths within the SGS(arrows). H1–H3 show superficial and deep regenerate RGC axons in a rat (#24) injected with AAV–CNTF and then rBDNF. Note the substantial outgrowth for several mm from the PNgraft, but complex arbor formation was limited to the SGS. I, Retinal innervation of the pretectum in an AAV–CNTF+rBDNF animal (#20). J, regenerate axons in the deep intermediatelayers of the SC (#45, AAV–CNTF+AAV–BDNF group). Note the lack of complex arbors. Scale bars for all except H1 = 100 μm; scale bar for H1 = 500 μm.
205S.-W. You et al. / Experimental Neurology 279 (2016) 197–211
through SO but highly complex arbors were only evident in the superfi-cial SC (Fig. 5H3).
In about 20% of all AAV–CNTF injected rats, occasional, scattered re-generate RGC axons were seen rostral to the midbrain SC. In four cases(two AAV–CNTF + saline, one AAV–BDNF and one AAV–CNTF + rBDNF animal), part of the PN graft entered at the rostral SC border andthe density of RGC axons was relatively high rostro-lateral to the SC,
in the brachium/nucleus of the optic tract region. An example isshown in Fig. 5I.
3.2.4. QuantificationOutgrowth of RGC axons back into the midbrain was quantified in a
number of ways. Only those animals in which there was good CTB labelwithin PN grafts (Fig. 2C) and at the distal end of each graft (Fig. 4) were
206 S.-W. You et al. / Experimental Neurology 279 (2016) 197–211
considered for analysis. In addition, in three animals (one saline, twoAAV–CNTF + rBDNF) the SC morphology was poor, perhaps due toin vivo degenerative changes or post-mortem artifact associated withpreparation for histology; these rats could not be used for quantifica-tion. Finally, there were two rats in which there was almost no RGC ax-onal outgrowth from the PN graft due to the presence of bulb endingswith obstructive matrix – see Fig. 4I–K). As a consequence, a total of 8saline or AAV–GFP (control data were pooled from these groups), 10AAV–CNTF + saline, 10 AAV–CNTF + AAV-BDNF, and 8 AAV–CNTF +rBDNF animals were quantified.
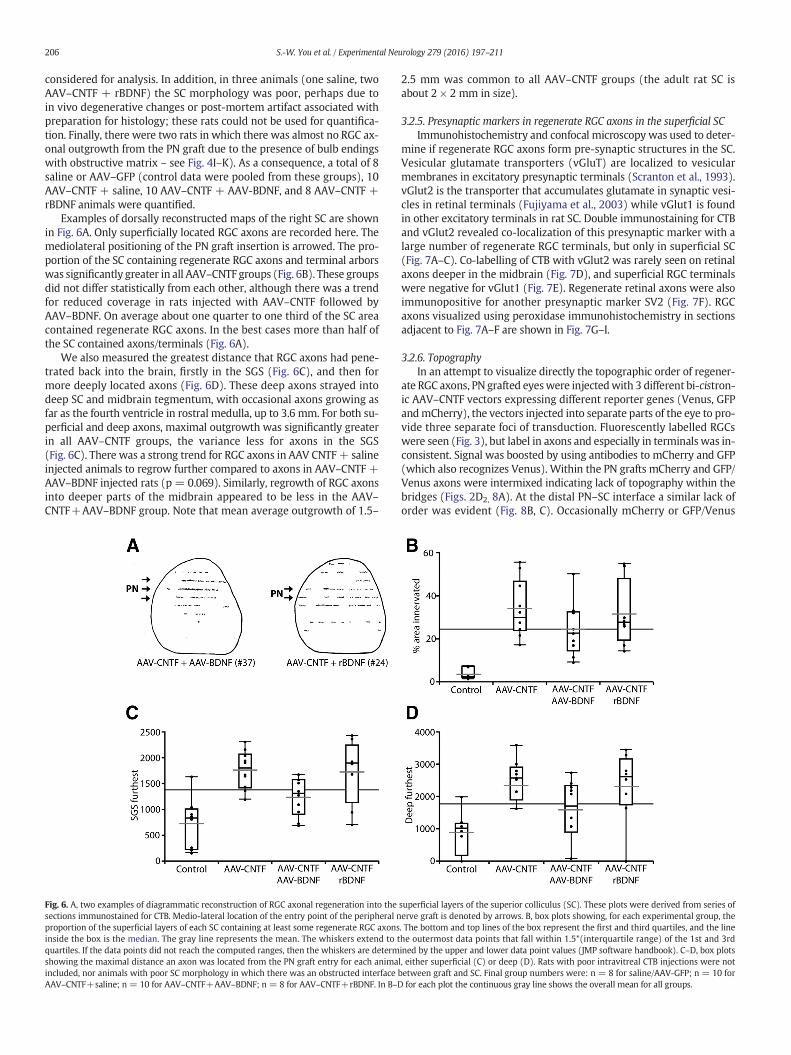
Examples of dorsally reconstructed maps of the right SC are shownin Fig. 6A. Only superficially located RGC axons are recorded here. Themediolateral positioning of the PN graft insertion is arrowed. The pro-portion of the SC containing regenerate RGC axons and terminal arborswas significantly greater in all AAV–CNTF groups (Fig. 6B). These groupsdid not differ statistically from each other, although there was a trendfor reduced coverage in rats injected with AAV–CNTF followed byAAV–BDNF. On average about one quarter to one third of the SC areacontained regenerate RGC axons. In the best cases more than half ofthe SC contained axons/terminals (Fig. 6A).
We also measured the greatest distance that RGC axons had pene-trated back into the brain, firstly in the SGS (Fig. 6C), and then formore deeply located axons (Fig. 6D). These deep axons strayed intodeep SC and midbrain tegmentum, with occasional axons growing asfar as the fourth ventricle in rostral medulla, up to 3.6 mm. For both su-perficial and deep axons, maximal outgrowth was significantly greaterin all AAV–CNTF groups, the variance less for axons in the SGS(Fig. 6C). There was a strong trend for RGC axons in AAV CNTF+ salineinjected animals to regrow further compared to axons in AAV–CNTF +AAV–BDNF injected rats (p = 0.069). Similarly, regrowth of RGC axonsinto deeper parts of the midbrain appeared to be less in the AAV–CNTF+AAV–BDNF group. Note that mean average outgrowth of 1.5–
Fig. 6. A, two examples of diagrammatic reconstruction of RGC axonal regeneration into thesections immunostained for CTB. Medio-lateral location of the entry point of the peripheral nproportion of the superficial layers of each SC containing at least some regenerate RGC axonsinside the box is the median. The gray line represents the mean. The whiskers extend to tquartiles. If the data points did not reach the computed ranges, then the whiskers are determshowing the maximal distance an axon was located from the PN graft entry for each animaincluded, nor animals with poor SC morphology in which there was an obstructed interfaceAAV–CNTF+saline; n = 10 for AAV–CNTF+AAV–BDNF; n = 8 for AAV–CNTF+rBDNF. In B–D
2.5 mm was common to all AAV–CNTF groups (the adult rat SC isabout 2 × 2 mm in size).
3.2.5. Presynaptic markers in regenerate RGC axons in the superficial SCImmunohistochemistry and confocal microscopy was used to deter-
mine if regenerate RGC axons form pre-synaptic structures in the SC.Vesicular glutamate transporters (vGluT) are localized to vesicularmembranes in excitatory presynaptic terminals (Scranton et al., 1993).vGlut2 is the transporter that accumulates glutamate in synaptic vesi-cles in retinal terminals (Fujiyama et al., 2003) while vGlut1 is foundin other excitatory terminals in rat SC. Double immunostaining for CTBand vGlut2 revealed co-localization of this presynaptic marker with alarge number of regenerate RGC terminals, but only in superficial SC(Fig. 7A–C). Co-labelling of CTB with vGlut2 was rarely seen on retinalaxons deeper in the midbrain (Fig. 7D), and superficial RGC terminalswere negative for vGlut1 (Fig. 7E). Regenerate retinal axons were alsoimmunopositive for another presynaptic marker SV2 (Fig. 7F). RGCaxons visualized using peroxidase immunohistochemistry in sectionsadjacent to Fig. 7A–F are shown in Fig. 7G–I.
3.2.6. TopographyIn an attempt to visualize directly the topographic order of regener-
ate RGC axons, PN grafted eyeswere injectedwith 3 different bi-cistron-ic AAV–CNTF vectors expressing different reporter genes (Venus, GFPandmCherry), the vectors injected into separate parts of the eye to pro-vide three separate foci of transduction. Fluorescently labelled RGCswere seen (Fig. 3), but label in axons and especially in terminals was in-consistent. Signal was boosted by using antibodies to mCherry and GFP(which also recognizes Venus). Within the PN grafts mCherry and GFP/Venus axons were intermixed indicating lack of topography within thebridges (Figs. 2D2, 8A). At the distal PN–SC interface a similar lack oforder was evident (Fig. 8B, C). Occasionally mCherry or GFP/Venus
superficial layers of the superior colliculus (SC). These plots were derived from series oferve graft is denoted by arrows. B, box plots showing, for each experimental group, the. The bottom and top lines of the box represent the first and third quartiles, and the linehe outermost data points that fall within 1.5*(interquartile range) of the 1st and 3rdined by the upper and lower data point values (JMP software handbook). C–D, box plotsl, either superficial (C) or deep (D). Rats with poor intravitreal CTB injections were notbetween graft and SC. Final group numbers were: n = 8 for saline/AAV-GFP; n = 10 forfor each plot the continuous gray line shows the overall mean for all groups.

Fig. 7.A–F, confocal images of regenerate RGC terminals. A, CTB labelled RGC axons in the superficial superior colliculus immunostainedwith streptavidin-FITC. B, same field as A showingimmunofluorescence for vesicular glutamate transporter 2 (vGlut2). C, merged A and B images showing colocalization (e.g. arrow) of CTB and vGlut2 immunoreactivity. Numerous otherco-labelled (yellow) profiles can be seen. D, Deeper CTB labelled RGC axons (green) not immunoreactive for vGlut2. E, superficial regenerate RGC terminals not co-labelled with vGlut1. F,RGC terminals adjacent to peripheral nerve (PN) graft entry site immunopositive for both CTB (green) and the synaptic vesicle protein SV2 (red). G–I show adjacent sections to D–Frespectively showing RGC axons visualized using peroxidase immunohistochemistry. Scale bars: A–C, F = 10 μm; D, E = 20 μm; G = 200 μm; H, I = 100 μm.
207S.-W. You et al. / Experimental Neurology 279 (2016) 197–211
positive axons and terminal arborswere seenwithin the SC (Fig. 8D, E2).Fig. 8E shows a low power view of CTB 647 fluorescent axons in distalPN with axons growing into adjacent SC (white arrow). At higherpower, mCherry and GFP/Venus labelled RGC profiles can be seenadjacent to each other in rostral SC (Fig. E2), suggesting a lack of orderin the pattern of reinnervation. Note that mCherry positive axonsoriginating from lower nasal retina (Fig. 3C) should project to caudalnot rostral, SC.
3.3. Behavior
Weused three different behavioral tests to detect possible functionaloutcomes associated with the regeneration of RGC axons back into thecontralateral midbrain. These were a light–dark exploration test, mea-surement of the pupillary light response and an optokinetic test. Duringtesting, vision through the right unoperated eye was temporarilyobstructed by eyelid sutures. Eight rats had small cataracts (2 eachfrom the AAV–CNTF + saline and AAV–CNTF + rBDNF groups, and 4in the AAV–CNTF + AAV–BDNF group) which made it difficult to accu-ratelymonitor changes in pupillary size. These animalswere not includ-ed in the pupillary response analysis, however we were able to assess alarger number of animals in the light–dark assessment (Fig. 8A). ANOVAand post hoc tests did not show any differences between GFP and salinefor any of the behavioral tests and these control groups were therefore
combined. We were unable to detect any unequivocal responses inthe optokinetic test, in any group.
In the light exploration test, overall activity levels were similaracross groups; due to large inter-animal variability in performancethere were no significant differences between groups, althoughmedianaverages were lower in the two BDNF treated cohorts (Fig. 9A). Impor-tantly however, the maximal distance of axonal regeneration into thecontralateral, superficial SC was strongly correlated with percent timein the light for rats in the control and AAV–CNTF + rBDNF groups (rfor both = 0.95, p b 0.05; Fig. 9B). The correlation for time in light andSGS reinnervation failed to reach significance (p = 0.09) for AAV–CNTF + AAV–BDNF animals. There was no correlation between behav-ioral outcomes and extent of RGC axonal outgrowth into deeper parts ofthe midbrain in any of the experimental groups.
In the pupillary light testwemeasured bothminimumdiameter andthe iris/pupil ratio over time to plot speed of pupillarymovement. Therewas an overall difference between groups (for pupil size, with AAV–CNTF + AAV BDNF and AAV–CNTF + rBDNF groups displaying morepupillary constriction compared to controls (saline and AAV–GFPgrouped) (Fig. 9C, D). Although there was no correlation betweenmax-imum axonal outgrowth and pupillary light responses in animals fromeither of the BDNF groups, there was a significant correlation in thecombined control group (r=0.97, p b 0.05) between the area of the su-perficial SC containing regenerate RGC axons and minimum pupildiameter.

Fig. 8. Identification of mCherry (red) and GFP/Venus (green) containing RGC axons and terminals in peripheral nerve grafts and superior colliculus. Because the level of intrinsicfluorescence was not consistent, sections were immunostained using antibodies to mCherry and GFP (the latter antibody recognizes both GFP and the related Venus fluorophores). A,intermixed mCherry and GFP/Venus axons in peripheral nerve (PN) graft (rat #38). B–C, intermixed regenerate RGC axons at the distal end of a PN graft (rat #24), immediatelyadjacent to the superior colliculus (SC; asterisks delineate interface between graft and brain). D, mCherry positive RGC axons in intermediate SC. E, CTB labelled RGC axons (far redfluorescence) in PN graft with some axons in rostral SGS (white arrow, rat #24); E2, high power view of E, showing adjacent RGC terminals in the SGS immunopositive for mCherry(red arrow) or GFP/Venus (green arrow). SGS, stratum griseum superficiale. Scale bars: A, C = 200 μm; B, D, E2 = 100 μm; E = 250 μm.
208 S.-W. You et al. / Experimental Neurology 279 (2016) 197–211
Two further comparisons were made. In 4 AAV–CNTF injected ratswith measurable pupillary responses, regenerate axons were seen ros-tral to the SC in the pretectal area (e.g. Fig. 5I). While average minimalpupil diameter was slightly lower in these animals compared to otherAAV–CNTF injected animals, the difference was not significant (p =0.64, t-test). We also compared responses in animals with predomi-nantly rostrally located axons in the SC, but again this did not impacton pupillary responses compared to the remaining animals. Eight ani-mals were not included in the pupil analysis due to cataracts thatpreventedmeasurement. Given that lens injury can enhance the regen-erative response of RGCs (Leon et al., 2000) we comparedmaximal out-growth and area of SC innervation in these ratswith the animals that didnot have cataracts. No significant differences were seen, although in theAAV–CNTF + AAV–BDNF group there was a trend suggesting less out-growth in the lens damaged animals.
4. Discussion
After complete optic nerve transection, the novel combination of au-tologous PN bridging graft, gene therapy and pharmacotherapy consis-tently resulted in extensive adult RGC survival and unequivocalregeneration of axons back into the contralateral SC. In many animals,localized patches of high density axons/terminals were seen in SGS ad-jacent to the PN insertion site. Complex terminal arbors were formedonly in the superficial SC layers, the terminals expressing appropriatepresynaptic retinal markers. Despite the unprecedented number ofRGC axons re-entering the host SC we obtained no clear indication ofvisuotopic order in the regenerate projection. Nonetheless, additionalBDNF application was associated with improved visual function viathe grafted eye and at the individual animal level the extent of retinalaxon regrowth back into the SC correlated with light avoidancebehavior.
4.1. PN grafts and retinal ganglion cell survival
Combining autologous PN grafts with other therapies, such as bymodulating macrophage/microglial activity (Thanos and Mey, 1995;Avilés-Trigueros et al., 2000; Yin et al., 2003) or application of recombi-nant neurotrophic factors (Watanabe, 2010; Harvey et al., 2012), fur-ther improves adult RGC viability and regenerative potential. Thepresent study extends our previous work using blind-ended PN graftsand gene therapy (Leaver et al., 2006; Hellström and Harvey, 2014) byinserting the grafts into the contralateral SC. Connection with centraltargets potentially provides an additional source of trophic support forregenerating axons. Consistent with this, in control PN–SC grafted ani-mals RGC survival was about 50% higher than in equivalent blind-ended PN grafts. On the other hand, when AAV–CNTFwas administeredto ratswith PN–SC grafts, RGC viabilitywas not greater than that report-ed previously using blind-ended PN (Hellström and Harvey, 2011;Hellström et al., 2011b). Sequential intra-retinal administration ofBDNF did not further enhance RGC viability indicating that, in the pres-ence of appropriate local neurotrophic support, connection to targetsdoes not further enhance the survival of injured CNS neurons (Harveyet al., 2015).
4.2. Regeneration of RGC axons into central visual targets
In earlier studies, RGC outgrowth from PN grafted into visual centerswas often sparse and in most cases limited to a maximum of 500 μmfrom the insertion (Vidal-Sanz et al., 1987; Carter et al., 1989).With sur-vival times of 4 months or greater, regenerate RGC axons formed moreelaborate terminal arbors and grew further distances within the brain,the type and shape of the arbors influenced by the nature of the centraltarget (Carter et al., 1994; Carter and Jhaveri, 1997; Avilés-Trigueroset al., 2000). Even so, where documented, in both rat and hamster

Fig. 9. Analysis of visual responses following treatments. Rats were tested in the light/dark tunnel, and pupil mobility wasmeasured under infra-red illumination. For all animals only thegrafted (left) eye was tested; the right eye was sutured closed during testing. A: Box plot showing proportion of time rats spent in the dark versus illuminated part of apparatus. B:correlation between percent time in the light and maximal outgrowth of RGC axons in the stratum griseum superficiale (SGS). A significant relationship was seen in the control (noAAV–CNTF) group and rats injected with AAV–CNTF followed by recombinant BDNF. C: Box plot showing mean minimum pupil area measured 2 min after turning on the light,expressed as % of maximum pupil area measured after 10 min of dark adaptation. Note that several rats could not be tested due to ocular issues (e.g. cataracts) that interfered withmeasurement. D: Time course of mean pupillary constriction over time for all groups. Constriction time and extent in a normal ungrafted eye is shown for comparison. A,B: Box plotsshow median and 1.5 interquartile ranges. Short gray horizontal lines on each box show means of individual groups. Horizontal line across the entire plot shows the grand mean. D:Error bars are standard error of the mean.
209S.-W. You et al. / Experimental Neurology 279 (2016) 197–211
RGC axonal outgrowth failed in more than 50–60% of PN graftedanimals.
The outgrowth seen in the present study usingAAV–CNTF to initiallydrive regeneration was far greater, more consistent, and denser thananything reported previously. In AAV–CNTF treated rats, 4 monthspost-transplantation re-growth into the visual midbrain was seen in93% of cases. RGC axonswere found up to 3.6mm from the PN insertion,and average outgrowth ranged from 1.5–2.5 mm in the different AAV–CNTF injected groups. Near the insertion, reinnervation was so densethat individual arbors could not be distinguished. These patches weremost frequently seen in the AAV–CNTF + rBDNF group. On average25–35% of the SC contained axons/terminals, and in 4 cases more thanhalf the SC contained regenerate profiles. These are remarkable results,especially given thatwe inserted the distal end of the PN grafts into con-tralateral SC immediately after the proximal end was sutured onto thetransected ON. In many previous studies, to increase synaptic site va-cancy and hence to enhance outgrowth, PN grafts were inserted into ip-silateral visual centers and the input from the remaining, contralateraleye was removed at the time of the PN graft (Vidal-Sanz et al., 1987;Sasaki et al., 1999; Avilés-Trigueros et al., 2000) or several weeks later(Carter et al., 1989, 1994; Keirstead et al., 1989; Carter and Jhaveri,1997). Our new combinatorial therapy data show that large-scale rein-nervation of CNS targets can be achievedwithout recourse to removal ofany remaining normal inputs, clearly important in a clinical context.
Regenerate retinal arbors were found at various depths within theSGS. In normal rat the terminals of different classes of RGC are tosome extent segregated, slower-conducting axons projecting to upperSGS and faster-conducting axons terminatingmore deeply in the super-ficial gray and optic layers (Sefton et al., 2015). In some species, such ascat, mouse and ferret, large Type 1 RGCs (physiologically identified as
fast-conducting Y cells in cats) predominate in the regenerate projec-tion mediated by PN grafts (Quan et al., 1999; Cui and Harvey, 2000;Watanabe and Fukuda, 2002; Watanabe, 2010). A similar observationwas recently made in mice after optic nerve crush (Duan et al., 2015).However in rats, while Type 1 RGCs may contribute a disproportionateamount in the regenerate population after CNTF or AAV–CNTF treat-ments (Rodger et al., 2012; Drummond et al., 2014), all classes of RGCregrow axons into PN grafts (Vidal-Sanz et al., 1987; Thanos and Mey,1995), and consistent with this we saw regenerate arbors that wereclearly restricted to upper SGS. More definitive association of RGC sub-type to particular regenerate terminal arbors may be possible in miceinwhich RGC classes can be characterized usingmarkers that do not de-pend onmorphological criteria such as some size, or dendritic or termi-nal architecture (Duan et al., 2015).
While many regenerating RGC axons were found in the superficialSC, axons were also seen coursing deep within the midbrain for severalmillimetres. Secondary application of AAV–BDNF or rBDNF 6 weeksafter surgery was tested to see if this would enhance terminal arbor for-mation of regenerate RGC axons within the SC. Animals were perfused9–10 weeks later, and CNTF driven RGC outgrowth presumably contin-uedduring this period.Where individual arbors could be visualized theydid appear to be more complex in BDNF treated rats, but there was alsoan intriguing trend for the extent of SC reinnervation to be less in ratsthat received AAV-BDNF injections, perhaps suggesting cessation of ax-onal elongation.
4.3. Selectivity of reinnervation
In all AAV–CNTF treated animals complex terminal arbors and ex-pression of presynaptic proteins (vGlut2 and SV2) were seen only in

210 S.-W. You et al. / Experimental Neurology 279 (2016) 197–211
RGC axons that were located in the superficial SC layers, layers that nor-mally receive retinal input. What is the nature of the axon-target selec-tivity seen here – is it a recapitulation of specific developmentalmechanisms or is it a more general response to deafferentation signalsresulting from the removal of retinal (and in most cases visual cortical)axons (Zwimpfer et al., 1992)? Spontaneously regrowing adult rat RGCaxons can selectively recognize their appropriate, localized target zoneswithin developing tectal tissue (Harvey and Tan, 1992), but in the SCafter adult ON transection there is loss of terminals and synapses, localreoccupation of vacant synaptic sites (Turner et al., 2005), and long-term changes in astrocytic and microglial reactivity (Rao and Lund,1993). We believe the PN–SC graft model will help dissect out thedrivers of regenerative specificity in the adult CNS, of general impor-tance in any study of CNS repair, especially if regrowing axons takemany weeks or months to reach their deafferented targets.
4.4. Behavior, function and visual topography
We saw no consistent optokinetic response, but similar to others(Thanos, 1992, Whiteley et al., 1998) pupillary responses to light inthe grafted eye were significantly improved in the AAV–CNTF + AAVBDNF and AAV–CNTF + rBDNF groups. The olivary pretectal nucleus isregarded as central to the pupillary light response (Sefton et al., 2015)and we occasionally saw axons in the vicinity of this nucleus, althoughresponses in these rats were not significantly different from others.Light can affect the iris sphincter directly (Xue et al., 2011) but anysuch indirect effect cannot easily explainwhywe saw significant chang-es in pupillary responsiveness in animals exposed to both CNTF andBDNF. Note that neurons in the superficial SC project to pretectal nuclei(May, 2006) and to deeper tectal neurons (Sefton et al., 2015), perhapsproviding an opportunity for indirect mediation of the reflex by regen-erate RGC axons.
In the light exploration test, due to substantial variance we saw nosignificant differences at the group level, although median time spentin the light was lower in the BDNF treated cohorts. At the group level,light avoidance behaviour using a conditioning response protocol hasbeen reported (Sasaki et al., 1993, 1999). Most importantly however,we found a significant correlation between behavior and the extent ofreinnervation of the SGS in individual in the AAV–CNTF + rBDNFgroup but intriguingly also in the control group; the further the out-growth the better the behavioral response. This positive correlation be-tween function and regenerative axonal growth has rarely beendocumented after injury to the adult mammalian CNS, including spinalcord. It is intriguing to note in this context that, for reasons that remainunclear, most adult rat RGCs regenerating axons into a PN graft appearto beONcentre cells based on careful analysis of dendriticfield architec-ture (Rodger et al., 2012), similar to that seen physiologically and mor-phologically in PN grafted cats (Watanabe and Fukuda, 2002; Yata et al.,2007; Watanabe, 2010).
Amajor aim of mammalian visual system repair is the restoration ofvisuotopicity and patterned vision. In developing zebrafish, lack of com-petition does not obviously influence retinotopic order (Gosse et al.,2008), however in mammals a gross lack of order in the regenerate ret-inotectal projection has been attributed to the sparseness of reinnerva-tion: “With few innervating axons, the opportunity for eithercompetition or cooperativity among axons in establishing and stabiliz-ing synaptic connections would be minimal” (Sauvé et al., 2001). Ourstudies were designed to substantially increase the number of RGCaxons re-entering the SC, and by injecting different AAV–CNTF vectorswith different reporters into different regions of the eye we hoped tomap the relative location in the SC of regenerate axons and terminalsoriginating from different RGC populations. There was a lack of orderin regenerating axons within PN grafts and at the PN–SC interface, andalthough terminal label was inconsistent, mCherry and GFP/Venus la-belled RGC profiles were occasionally seen adjacent to each other,
suggesting a lack of topography in the restored projection, regardlessof the high density of innervation.
The lack of order is surprising given evidence for temporary re-expression of at least some relevant positional markers in deafferentedrodent SC (Bähr andWizenmann, 1996; Knöll et al., 2001; Koeberle andBähr, 2004), with upregulation and graded rostro-caudal expression ofephrin-A2 for several months (Rodger et al., 2001, 2005b). It may bethat sustained growth induction by CNTF was detrimental to the re-establishment of topography, perhaps similar to the impact of increas-ing cAMP during retinotectal regeneration in goldfish, which increasedthe rate of axonal regrowth but contributed to disorder in the reformedprojection (Rodger et al., 2005a). Sustained high levels of CNTFmay alsobe detrimental to RGC function (see also Rodger et al. (2012)), suggest-ing that it will be necessary to regulate transgene expression of growthfactors. In future PN graft studies itmay be possible tominimize relianceon trophic factors by enhancing RGC axonal outgrowth by limiting scar-ring (Symonds et al., 2007) and proteoglycan deposition (Garcia-Alíasand Fawcett, 2012). Clearly multifactorial approaches will be needed,perhaps including intensive training (Beazley et al., 2003), to “force a re-capitulation of the orderly developmental events that got us there in thefirst place” (Koeberle and Bähr, 2004).
References
Avilés-Trigueros, M., Sauvé, Y., Lund, R.D., Vidal-Sanz, M., 2000. Selective innervation ofretinorecipient brainstem nuclei by retinal ganglion cell axons regenerating throughperipheral nerve grafts in adult rats. J. Neurosci. 20, 361–374.
Bähr, M., Wizenmann, A., 1996. Retinal ganglion cell axons recognize specific guidancecues present in the deafferented adult rat superior colliculus. J. Neurosci. 16,5106–5116.
Beazley, L.D., Rodger, J., Chen, P., Tee, L.B., Stirling, R.V., Taylor, A.L., Dunlop, S.A., 2003.Training on a visual task improves the outcome of optic nerve regeneration.J. Neurotrauma 20, 1263–1270.
Berry, M., Ahmed, Z., Lorber, B., Douglas, M., Logan, A., 2008. Regeneration of axons in thevisual system. Restor. Neurol. Neurosci. 26, 147–174.
Carter, D.A., Jhaveri, S., 1997. Retino-geniculate axons regenerating in adult hamsters areable to form morphologically distinct terminals. Exp. Neurol. 146, 315–322.
Carter, D.A., Bray, G.M., Aguayo, A.J., 1989. Regenerated retinal ganglion cell axons canform well-differentiated synapses in the superior colliculus of adult hamsters.J. Neurosci. 9, 4042–4050.
Carter, D.A., Bray, G.M., Aguayo, A.J., 1994. Long-term growth and remodeling of regener-ated retino-collicular connections in adult hamsters. J. Neurosci. 14, 590–598.
Cohen-Cory, S., Kidane, A.H., Shirkey, N.J., Marshak, S., 2010. Brain-derived neurotrophicfactor and the development of structural neuronal connectivity. Dev, Neurobiol 70,271–288.
Cui, Q., Harvey, A.R., 2000. CNTF promotes the regrowth of retinal ganglion cell axons intomurine peripheral nerve grafts. Neuroreport 11, 3999–4002.
Cui, Q., Yip, H.K., Zhao, R.C.H., So, K.-F., Harvey, A.R., 2003. Intraocular elevation of cyclicAMP potentiates CNTF-induced regeneration of adult retinal ganglion cell axons.Mol. Cell. Neurosci. 22, 49–61.
de Lima, S., Koriyama, Y., Kurimoto, T., Oliveira, J.T., Yin, Y., Li, Y., Gilbert, H.Y., Fagiolini, M.,Martinez, A.M., Benowitz, L.I., 2012. Full-length axon regeneration in the adult mouseoptic nerve and partial recovery of simple visual behaviors. Proc. Natl. Acad. Sci. U. S.A. 109, 9149–9154.
Drummond, E.S., Rodger, J., Penrose, M., Robertson, D., Hu, Y., Harvey, A.R., 2014. Effects ofintravitreal injection of either a Rho-GTPase inhibitor (BA-210), or CNTF combinedwith an analogue of cAMP, on the dendritic morphology of regenerating adult rat ret-inal ganglion cells. Restor. Neurol. Neurosci. 32, 391–402.
Duan, X., Qiao, M., Bei, F., Kim, I.J., He, Z., Sanes, J.R., 2015. Subtype-specific regeneration ofretinal ganglion cells following axotomy: effects of osteopontin and mTOR signalling.Neuron 85, 1244–1256.
Fischer, D., Leibinger, M., 2012. Promoting optic nerve regeneration. Prog. Retin. Eye Res.31, 688–701.
Fujiyama, F., Hioki, H., Tomioka, R., Taki, K., Tamamaki, N., Nomura, S., Okamoto, K.,Kaneko, T., 2003. Changes of immunocytochemical localization of vesicular glutamatetransporters in the rat visual system after the retinofugal denervation. J. Comp.Neurol. 465, 234–249.
Garcia-Alías, G., Fawcett, J.W., 2012. Training and anti-CSPG combination therapy for spi-nal cord injury. Exp. Neurol. 235, 26–32.
Gosse, N.J., Nevin, L.M., Baier, H., 2008. Retinotopic order in the absence of axon competi-tion. Nature 452, 892–895.
Harvey, A.R., MacDonald, A.M., 1985. The development of acetylcholinesterase activity innormal and transplanted superior colliculus in rats. J. Comp. Neurol. 240, 117–127.
Harvey, A.R., Tan, M.M.L., 1992. Spontaneous regeneration of retinal ganglion cell axons inadult rats. NeuroReport 3, 239–242.
Harvey, A.R., Hu, Y., Leaver, S.G., Mellough, C.B., Park, K., Verhaagen, J., Plant, G.W., Cui, Q.,2006. Gene therapy and transplantation in CNS repair: the visual system. Prog. Retin.Eye Res. 25, 449–489.

211S.-W. You et al. / Experimental Neurology 279 (2016) 197–211
Harvey, A.R., Ooi, J.W.W., Rodger, J., 2012. Neurotrophic factors and the regeneration ofadult retinal ganglion cell axons. In: Goldberg, J.L., Trakhtenberg, E.F. (Eds.), AxonGrowth and Regeneration. Int Rev Neurobiol Vol. 106, pp. 1–34.
Harvey, A.R., Lovett, S.J., Majda, B.T., Yoon, J.H., Wheeler, L.P.G., Hodgetts, S.I., 2015. Neu-rotrophic factors for spinal cord repair: which, where, how and when to apply, andfor what period of time? Brain Res. 1619, 36–71.
Heiduschka, P., Thanos, S., 2000. Restoration of the retinofugal pathway. Prog. Retin. EyeRes. 19, 577–606.
Hellström, M., Harvey, A.R., 2011. Retinal ganglion cell gene therapy and visual system re-pair. Curr. Gene Ther. 11, 116–131.
Hellström, M., Harvey, A.R., 2014. Cyclic AMP and the regeneration of retinal ganglion cellaxons. Int. J. Biochem. Cell Biol. 56, 66–73.
Hellström, M., Muhling, J., Ehrlert, E., Verhaagen, J., Pollett, M.A., Hu, Y., Harvey, A.R.,2011a. Negative impact of rAAV2 mediated expression of SOCS3 on the regenerationof adult retinal ganglion cell axons. Mol. Cell. Neurosci. 46, 507–515.
Hellström, M., Pollett, M.A., Harvey, A.R., 2011b. Post-injury delivery of rAAV2-CNTF com-bined with short-term pharmacotherapy is neuroprotective and promotes extensiveaxonal regeneration after optic nerve trauma. J. Neurotrauma 28, 2475–2483.
Keirstead, S.A., Rasminsky, M., Fukuda, Y., Carter, D.A., Aguayo, A.J., Vidal-Sanz, M., 1989.Electrophysiologic responses in hamster superior colliculus evoked by regeneratingretinal axons. Science 246, 255–257.
Knöll, B., Isenmann, S., Kilic, E., Walkenhorst, J., Engel, S., Wehinger, J., Bähr, M., Drescher,U., 2001. Graded expression patterns of ephrin-As in the superior colliculus after le-sion of the adult mouse optic nerve. Mech. Dev. 106, 119–127.
Koeberle, P.D., Bähr, M., 2004. Growth and guidance cues for regenerating axons: wherehave they gone? J. Neurobiol. 59, 162–180.
Leaver, S.G., Cui, Q., Plant, G.W., Arulpragasam, A., Hisheh, S., Verhaagen, J., Harvey, A.R.,2006. AAV-mediated expression of CNTF promotes long-term survival and regenera-tion of adult rat retinal ganglion cells. Gene Ther. 13, 1328–1341.
Leon, S., Yin, Y., Nguyen, J., Irwin, N., Benowitz, L.I., 2000. Lens injury stimulates axon re-generation in the mature rat optic nerve. J. Neurosci. 20 (4615-4612).
Luo, X., Salgueiro, Y., Beckerman, S.R., Lemmon, V.P., Tsoulfas, P., Park, K.K., 2013. Three-dimensional evaluation of retinal ganglion cell axon regeneration and pathfindingin whole mouse tissue after injury. Exp. Neurol. 247, 653–662.
May, P.J., 2006. The mammalian superior colliculus: laminar structure and connections.Prog. Brain Res. 151, 321–378.
Park, K., Luo, J.-M., Hisheh, S., Harvey, A.R., Cui, Q., 2004. Cellular mechanisms associatedwith spontaneous and ciliary neurotrophic factor/cAMP-induced survival and axonalregeneration of adult retinal ganglion cells. J. Neurosci. 24, 10806–10815.
Quan, M.-Z., Kosaka, J., Watanabe, M., Wakabayashi, T., Fukuda, Y., 1999. Survival ofaxotomized retinal ganglion cells in peripheral nerve-grafted ferrets. Invest.Ophthalmol. Vis. Sci. 40, 2360–2366.
Rao, K., Lund, R.D., 1993. Optic nerve degeneration induces the expression of MHC anti-gens in the rat visual system. J. Comp. Neurol. 336, 613–627.
Rodger, J., Lindsey, K.A., Leaver, S.G., King, C.E., Dunlop, S.A., Beazley, L.D., 2001. Expressionof ephrin-A2 in the superior colliculus and EphA5 in the retina following optic nervesection in adult rat. Eur. J. Neurosci. 14, 1929–1936.
Rodger, J., Goto, H., Cui, Q., Chen, P.B., Harvey, A.R., 2005a. cAMP regulates axon outgrowthand guidance during optic nerve regeneration in goldfish. Mol. Cell. Neurosci. 30,452–464.
Rodger, J., Symonds, A.C.E., Springbett, J., Shen, W.-Y., Bartlett, C.A., Rakoczy, P.E., Beazley,L.D., Dunlop, S.A., 2005b. Eph/ephrin expression in the adult rat visual system follow-ing localized retinal lesions: localized and transneuronal up-regulation in the retinaand superior colliculus. Eur. J. Neurosci. 22, 1840–1852.
Rodger, J., Drummond, E.S., Hellström, M., Robertson, D., Harvey, A.R., 2012. Long-termgene therapy causes transgene-specific changes in the morphology of regeneratingretinal ganglion cells. PLoS One 7 (2), e31061.
Sasaki, H., Inoue, T., Iso, H., Fukuda, Y., Hayashi, Y., 1993. Light–dark discrimination aftersciatic nerve transplantation to the sectioned optic nerve in adult hamsters. Vis.Res. 7, 877–880.
Sasaki, H., Inoue, T., Iso, H., Fukuda, Y., 1999. Recovery of visual behaviors in adult ham-sters with peripheral nerve graft to the sectioned optic nerve. Exp. Neurol. 159,377–390.
Sauvé, Y., Sawai, H., Rasminsky,M., 1995. Functional synaptic connectionsmade by regen-erated retinal ganglion cell axons in the superior colliculus of adult hamsters.J. Neurosci. 15, 665–675.
Sauvé, Y., Sawai, H., Rasminsky, M., 2001. Topological specificity in reinnervation of thesuperior colliculus by regenerated retinal ganglion cell axons in adult hamsters.J. Neurosci. 21, 951–960.
Sawai, H., Clarke, D.B., Kittlerova, P., Bray, G.M., Aguayo, A.J., 1996. Brain-derived neuro-trophic factor and neurotrophin-4/5 stimulate growth of axonal branches fromregenerating retinal ganglion cells. J. Neurosci. 16, 3887–3894.
Scranton, T.W., Iwata, M., Carlson, S.S., 1993. The SV2 protein of synaptic vesicles is akeratan sulfate proteoglycan. J. Neurochem. 61, 29–44.
Sefton, A.J., Dreher, B., Harvey, A.R., Martin, P.R., 2015. Visual System. In: Paxinos, G. (Ed.),The Rat Nervous System, fourth ed., pp. 947–983.
Sendtner, M., Schmalbruch, H., Stockli, K.A., Carroll, P., Kreutzberg, G.W., Thoenen, H.,1992. Ciliary neurotrophic factor prevents degeneration of motor neurons in mousemutant progressive motor neuronopathy. Nature 358, 502–504.
Sun, F., Park, K.K., Belin, S., Wang, D., Lu, T., Chen, G., Zhang, K., Yeung, C., Feng, G.,Yankner, B.A., He, Z., 2011. Sustained axon regeneration induced by co-deletion ofPTEN and SOCS3. Nature 480, 372–376.
Symonds, A.C., King, C.E., Bartlett, C.A., Sauvé, Y., Lund, R.D., Beazley, L.D., Dunlop, S.A.,Rodger, J., 2007. EphA5 and ephrin-A2 expression during optic nerve regeneration:a ‘two-edged sword’. Eur. J. Neurosci. 25, 744–752.
Thanos, S., 1992. Adult retinofugal axons regenerating through peripheral nerve graftscan restore the light-induced pupilloconstriction reflex. Eur. J. Neurosci. 4, 691–699.
Thanos, S., Mey, J., 1995. Type-specific stabilization and target-dependent survival ofregenerating ganglion cells in the retina of adult rats. J. Neurosci. 15, 1057–1079.
Turner, J.P., Sauvé, Y., Varela-Rodriguez, C., Lund, R.D., Salt, T.E., 2005. Recruitment of localexcitatory circuits in the superior colliculus following deafferentation and the regen-eration of retinocollicular inputs. Eur. J. Neurosci. 22, 1643–1654.
Vidal-Sanz, M., Bray, G.M., Villegas-Perez, M.P., Thanos, S., Aguayo, A.J., 1987. Axonal re-generation and synapse formation in the superior colliculus by retinal ganglioncells in the adult rat. J. Neurosci. 7, 2894–2909.
Vidal-Sanz, M., Aviles-Trigueros, M., Whiteley, S.J., Sauvé, Y., Lund, R.D., 2002. Reinnerva-tion of the pretectum in adult rats by regenerated retinal ganglion cell axons: ana-tomical and functional studies. Prog. Brain Res. 137, 443–452.
Villegas-Perez, M.P., Vidal-Sanz, M., Bray, G.M., Aguayo, A.J., 1988. Influences of peripheralnerve grafts on the survival and regrowth of axotomized retinal ganglion cells inadult rats. J. Neurosci. 8, 265–280.
Watanabe, M., 2010. Regeneration of optic nerve fibers of adult mammals. Develop.Growth Differ. 52, 567–576.
Watanabe, M., Fukuda, Y., 2002. Survival and regeneration of retinal ganglion cells inadult cats. Prog. Retin. Eye Res. 21, 529–553.
Whiteley, S.J., Sauvé, Y., Avilés-Trigueros, M., Vidal-Sanz, M., Lund, R.D., 1998. Extent andduration of recovered pupillary light reflex following retinal ganglion cell axon re-generation through peripheral nerve grafts directed to the pretectum in adult rats.Exp. Neurol. 154, 560–572.
Xue, T., Do, M.T., Riccio, A., Jiang, Z., Hsieh, J., Wang, H.C., Merbs, S.L., Welsbie, D.S.,Yoshioka, T., Weissgerber, P., Stolz, S., Flockerzi, V., Freichel, M., Simon, M.I.,Clapham, D.E., Yau, K.W., 2011. Melanopsin signalling in mammalian iris and retina.Nature 479, 67–73.
Yata, T., Nakamura, M., Sagawa, H., Tokita, Y., Terasaki, H., Watanabe, M., 2007. Survivaland axonal regeneration of OFF-center retinal ganglion cells of adult cats are promot-ed with an anti-glaucoma drug, Nipradilol, but not BDNF or CNTF. Neuroscience 148,53–64.
Yin, Y., Cui, Q., Li, Y., Irwin, N., Fischer, D., Harvey, A.R., Benowitz, L.I., 2003. Macrophage-derived factors stimulate optic nerve regeneration. J. Neurosci. 23, 2284–2293.
Zwimpfer, T.J., Aguayo, A.J., Bray, G.M., 1992. Synapse formation and preferential distribu-tion in the granule cell layer by regenerating retinal ganglion cell axons guided to thecerebellum of adult hamsters. J. Neurosci. 12, 1144–1159.


















