Lab on a Chip - Institute of Atomic and molecular Sciences
17
Cite this: Lab Chip, 2013, 13, 676 Improved silicon nanowire field-effect transistors for fast protein–protein interaction screening3 Received 9th July 2012, Accepted 31st October 2012 DOI: 10.1039/c2lc40772h www.rsc.org/loc Ti-Yu Lin, a Bor-Ran Li, bc Sheng-Ta Tsai, d Chien-Wei Chen, bc Chung-Hsuan Chen, d Yit- Tsong Chen* bc and Chien-Yuan Pan* ae Understanding how proteins interact with each other is the basis for studying the biological mechanisms behind various physiological activities. Silicon nanowire field-effect transistors (SiNW-FETs) are sensitive sensors used to detect biomolecular interactions in real-time. However, the majority of the applications that use SiNW-FETs are for known interactions between different molecules. To explore the capability of SiNW-FETs as fast screening devices to identify unknown interacting molecules, we applied mass spectrometry (MS) to analyze molecules reversibly bound to the SiNW-FETs. Calmodulin (CaM) is a Ca 2+ - sensing protein that is ubiquitously expressed in cells and its interaction with target molecules is Ca 2+ - dependent. By modifying the SiNW-FET surface with glutathione, glutathione S-transferase (GST)-tagged CaM binds reversibly to the SiNW-FET. We first verified the Ca 2+ -dependent interaction between GST–CaM and purified troponin I, which is involved in muscle contraction, through the conductance changes of the SiNW-FET. Furthermore, the cell lysate containing overexpressed Ca 2+ /CaM-dependent protein kinase IIa induced a conductance change in the GST–CaM-modified SiNW-FET. The bound proteins were eluted and subsequently identified by MS as CaM and kinase. In another example, candidate proteins from neuronal cell lysates interacting with calneuron I (CalnI), a CaM-like protein, were captured with a GST–CalnI- modified SiNW-FET. The proteins that interacted with CalnI were eluted and verified by MS. The Ca 2+ - dependent interaction between GST–CalnI and one of the candidates, heat shock protein 70, was re- confirmed via the SiNW-FET measurement. Our results demonstrate the effectiveness of combining MS with SiNW-FETs to quickly screen interacting molecules from cell lysates. Introduction In this post-genomic era, constructing a protein–protein interaction map is crucial in our attempt to understand the mechanisms underlying various physiological activities. Biologists have adopted several methods, such as coimmuno- precipitation, the yeast two-hybrid system, 1 and fluorescence resonance energy transfer, 2 to verify protein–protein interac- tions. Recently, highly sensitive biosensors, such as surface plasmon resonance (SPR) and field-effect transistors (FET), have been used to investigate molecular interactions. SPR identifies the molecular interactions by measuring the SPR angle shift that results from surface plasmon changes. 3 The SiNW-FET changes its electrical conductance when target molecules bind with the receptor-modified sensing device and vary the surface potential of the biosensor. 4 Unlike SPR, which requires an expensive optical device for detecting minute angle shifts, SiNW-FETs utilize established fabrication methods that are already in use in the silicon industry 5–7 and measure the conductance with a simple electronic device. By modifying the SiNW-FET surface with specific functional molecules, SiNW-FET devices can selec- tively detect proteins, 6 metal ions, 8 RNA, 9 cellular bioelectri- city, 10 and neuronal electrical signals. 11 SiNW-FET biosensors can easily detect samples at sub-nanomolar levels and are able to identify interacting proteins from sera or cell lysates in real- time. 7,12 However, most of the reports in the literature are focused on establishing the validity of the method and confirming known interactions. Adopting the technique of reversible surface modifica- tions, 6,7 a single SiNW-FET device can be repetitively used for quantitative analysis, and the bound molecules can be eluted for further analysis. Our previous studies have revealed the usefulness of this device in detecting protein–protein interac- tions. 6,7 Because of the high sensitivity of SiNW-FET devices, a minute amount of sample is sufficient for a SiNW-FET assay; however, traditional staining methods, such as Western blots a Institute of Zoology, National Taiwan University, Taipei, Taiwan b Department of Chemistry, National Taiwan University, Taipei, Taiwan c Institute of Atomic and Molecular Sciences, Academia Sinica, P.O. Box 23-166, Taipei 106, Taiwan E-mail: [email protected]; Fax: (+886) 2-2363-6359 d Genomics Research Center, Academia Sinica, Taipei, Taiwan e Department of Life Science, National Taiwan University, 1 Sec. 4 Roosevelt Road, Taipei 106, Taiwan E-mail: [email protected]; Fax: (+886) 2-2363-6837 3 Electronic supplementary information (ESI) available. See DOI: 10.1039/ c2lc40772h Lab on a Chip PAPER 676 | Lab Chip, 2013, 13, 676–684 This journal is ß The Royal Society of Chemistry 2013 Downloaded by National Taiwan University on 25 January 2013 Published on 31 October 2012 on http://pubs.rsc.org | doi:10.1039/C2LC40772H View Article Online View Journal | View Issue
Transcript of Lab on a Chip - Institute of Atomic and molecular Sciences

Cite this: Lab Chip, 2013, 13, 676
Improved silicon nanowire field-effect transistors forfast protein–protein interaction screening3
Received 9th July 2012,Accepted 31st October 2012
DOI: 10.1039/c2lc40772h
www.rsc.org/loc
Ti-Yu Lin,a Bor-Ran Li,bc Sheng-Ta Tsai,d Chien-Wei Chen,bc Chung-Hsuan Chen,d Yit-Tsong Chen*bc and Chien-Yuan Pan*ae
Understanding how proteins interact with each other is the basis for studying the biological mechanisms
behind various physiological activities. Silicon nanowire field-effect transistors (SiNW-FETs) are sensitive
sensors used to detect biomolecular interactions in real-time. However, the majority of the applications
that use SiNW-FETs are for known interactions between different molecules. To explore the capability of
SiNW-FETs as fast screening devices to identify unknown interacting molecules, we applied mass
spectrometry (MS) to analyze molecules reversibly bound to the SiNW-FETs. Calmodulin (CaM) is a Ca2+-
sensing protein that is ubiquitously expressed in cells and its interaction with target molecules is Ca2+-
dependent. By modifying the SiNW-FET surface with glutathione, glutathione S-transferase (GST)-tagged
CaM binds reversibly to the SiNW-FET. We first verified the Ca2+-dependent interaction between GST–CaM
and purified troponin I, which is involved in muscle contraction, through the conductance changes of the
SiNW-FET. Furthermore, the cell lysate containing overexpressed Ca2+/CaM-dependent protein kinase IIa
induced a conductance change in the GST–CaM-modified SiNW-FET. The bound proteins were eluted and
subsequently identified by MS as CaM and kinase. In another example, candidate proteins from neuronal
cell lysates interacting with calneuron I (CalnI), a CaM-like protein, were captured with a GST–CalnI-
modified SiNW-FET. The proteins that interacted with CalnI were eluted and verified by MS. The Ca2+-
dependent interaction between GST–CalnI and one of the candidates, heat shock protein 70, was re-
confirmed via the SiNW-FET measurement. Our results demonstrate the effectiveness of combining MS
with SiNW-FETs to quickly screen interacting molecules from cell lysates.
Introduction
In this post-genomic era, constructing a protein–proteininteraction map is crucial in our attempt to understand themechanisms underlying various physiological activities.Biologists have adopted several methods, such as coimmuno-precipitation, the yeast two-hybrid system,1 and fluorescenceresonance energy transfer,2 to verify protein–protein interac-tions. Recently, highly sensitive biosensors, such as surfaceplasmon resonance (SPR) and field-effect transistors (FET),have been used to investigate molecular interactions. SPRidentifies the molecular interactions by measuring the SPRangle shift that results from surface plasmon changes.3 TheSiNW-FET changes its electrical conductance when target
molecules bind with the receptor-modified sensing device andvary the surface potential of the biosensor.4
Unlike SPR, which requires an expensive optical device fordetecting minute angle shifts, SiNW-FETs utilize establishedfabrication methods that are already in use in the siliconindustry5–7 and measure the conductance with a simpleelectronic device. By modifying the SiNW-FET surface withspecific functional molecules, SiNW-FET devices can selec-tively detect proteins,6 metal ions,8 RNA,9 cellular bioelectri-city,10 and neuronal electrical signals.11 SiNW-FET biosensorscan easily detect samples at sub-nanomolar levels and are ableto identify interacting proteins from sera or cell lysates in real-time.7,12 However, most of the reports in the literature arefocused on establishing the validity of the method andconfirming known interactions.
Adopting the technique of reversible surface modifica-tions,6,7 a single SiNW-FET device can be repetitively used forquantitative analysis, and the bound molecules can be elutedfor further analysis. Our previous studies have revealed theusefulness of this device in detecting protein–protein interac-tions.6,7 Because of the high sensitivity of SiNW-FET devices, aminute amount of sample is sufficient for a SiNW-FET assay;however, traditional staining methods, such as Western blots
aInstitute of Zoology, National Taiwan University, Taipei, TaiwanbDepartment of Chemistry, National Taiwan University, Taipei, TaiwancInstitute of Atomic and Molecular Sciences, Academia Sinica, P.O. Box 23-166,
Taipei 106, Taiwan E-mail: [email protected]; Fax: (+886) 2-2363-6359dGenomics Research Center, Academia Sinica, Taipei, TaiwaneDepartment of Life Science, National Taiwan University, 1 Sec. 4 Roosevelt Road,
Taipei 106, Taiwan E-mail: [email protected]; Fax: (+886) 2-2363-6837
3 Electronic supplementary information (ESI) available. See DOI: 10.1039/c2lc40772h
Lab on a Chip
PAPER
676 | Lab Chip, 2013, 13, 676–684 This journal is � The Royal Society of Chemistry 2013
Dow
nloa
ded
by N
atio
nal T
aiw
an U
nive
rsity
on
25 J
anua
ry 2
013
Publ
ishe
d on
31
Oct
ober
201
2 on
http
://pu
bs.r
sc.o
rg |
doi:1
0.10
39/C
2LC
4077
2H
View Article OnlineView Journal | View Issue

and isotope labeling, are not sensitive enough to verify theeluted proteins.
In this report, we integrate mass spectrometry (MS) with theSiNW-FET assay to develop a new and fast strategy foridentifying the interacting proteins in real-time. Calmodulin(CaM) has four Ca2+-chelating sites, and its interactions withtarget proteins are Ca2+-dependent.13,14 We immobilizedglutathione S-transferase (GST)-tagged CaM on the glutathione(GSH)-modified SiNW-FET (GSH/SiNW-FET), and the Ca2+-dependent interaction between GST–CaM and Ca2+/CaM-dependent protein kinase IIa (CaMKIIa) was confirmed bymeasuring the conductance change of the SiNW-FET. Thebound proteins were then verified using MS analysis.Calneuron I (CalnI), a CaM-like protein, is involved inmodulating synaptic transmission.15 By binding GST–CalnIDHT (without the hydrophobic tail) onto the GSH/SiNW-FET (CalnIDHT/SiNW-FET), several proteins that inter-act with CalnIDHT in the neuronal cell lysate were identifiedby MS. The interaction between CalnIDHT and one of thecandidates, heat shock 70 kDa protein 1A/1B (Hsp70), wasfurther confirmed by a CalnIDHT/SiNW-FET assay usingpurified protein. These results indicate the advantages ofusing this fast, reversible, and sensitive approach for identify-ing interacting proteins by combining SiNW-FET and MS toreplace the traditional GST pull-down assay and immunopre-cipitation techniques.
Experimental
Reagents
Reagents for cell culture, Lipofectamine 2000 and mousemonoclonal anti-CaM antibody, were obtained from InvitrogenInc. (Carlsbad, CA, USA). Cardiac troponin I (TnI) and Hsp70were purchased from Abcam (Cambridge, MA, USA). Mousemonoclonal anti-green fluorescence protein (GFP) antibodywas obtained from Santa Cruz Biotech (Santa Cruz, CA, USA).All other chemicals were reagent grade and were purchasedfrom Sigma-Aldrich, unless otherwise indicated.
Fabrication of the SiNW-FET devices
The p-type SiNW-FETs used in this study were prepared usinga bottom-up technique. First, single-crystalline boron-dopedSiNWs (Si : B = 4000 : 1) were catalytically synthesized with theassistance of 20 nm gold nanoparticles in a chemical vapordeposition reaction via the vapor–liquid–solid growth mechan-ism (460 uC, 15 min in 10 standard cubic centimeter perminute (sccm) Ar, 6 sccm SiH4 (10% in He), and 7.5 sccm B2H6
(100 ppm in He) with a total chamber pressure of 25 torr).5 Thesingle-crystallinity of the synthesized SiNWs was examinedusing high-resolution transmission electron microscopy andelectron diffraction (JEM-2000FXP). The SiNW growth sub-strate, consisting of a dense ‘‘lawn’’ of SiNWs, was direction-ally slid onto another silicon substrate (acceptor substrate)that had been lithographically coated with patterned photo-resist (S1805) and Cr/Au peripheral electrodes. After the SiNWswere successfully transferred to the acceptor substrate,16 the
patterned photoresist was removed with acetone. Finally, Ni/Almicro-electrodes were defined using a photolithographictechnique. In this step, the metal contact regions were cleanedwith oxygen plasma (100 sccm and 30 W); meanwhile, thenative silica sheath of a SiNW in the contact area was removedwith a buffered oxide etching (BOE) solution for 3 s. Ni/Allayers (50/90 nm) were consecutively deposited onto thepattern-defined area by thermal evaporation. After lift-off,the SiNW-FET devices were further annealed in forming gases(10% H2 and 90% N2) at 450 uC for 3 min to ensure a decentelectrical contact between the SiNW and the metal electrodes.6
The as-fabricated FET devices are referred to as multi-SiNW-FETs and typically, each FET unit contains y5–10 SiNWs.
Surface modifications of the SiNW-FETs
The detailed modification steps can be found in a previousreport.7 In brief, a Si wafer chip containing the fabricatedSiNW-FET arrays first underwent oxygen plasma cleaning andthen was immersed in 3-aminopropyltrimethoxysilane(APTMS) containing ethanol for 30 min to form a self-assembling monolayer of APTMS on the chip. Subsequently,3-maleimidobenzoic acid N-hydroxysuccinimide ester wasused as a linker to react with APTMS at one end to form anamide bond and to immobilize the sulfhydryl group of GSH (1mM solution for 30 min) on the other end, thus forming theGSH/SiNW-FET. To immobilize CaM on the GSH/SiNW-FET, adiluted phosphate solution (PS, containing 0.76 mM Na2HPO4
and 0.24 mM NaH2PO4 at pH 7.4, Debye screening length of lD
y6.1 nm) supplemented with 200 nM GST–CaM was pumpedwith a syringe pump (KD Scientific, KD-101) at a flow rate of0.3 ml h21 for 30 min through a polydimethylsiloxane (PDMS)microfluidic channel (6.25 6 0.5 6 0.05 mm3) that wasdesigned to couple with the SiNW-FET device arrays (Fig. 1).
Because the surfaces of the SiNWs and the supportingsilicon-wafer substrates were both coated with an insulatinglayer of SiO2, the GSH was modified not only on the minuteSiNW surface but also on the widespread surrounding
Fig. 1 Schematic illustration of the experimental setup. A. The attachment ofthe PDMS module on the SiNW-FET biosensor chip to guide the flow of thesample solution. A Ag/AgCl electrode was used as a solution gate and was keptat ground potential during electrical measurements. B. The capture region forprotein binding. The microfluidic channel has dimensions of 6.25 6 0.5 6 0.05mm3. The GSH modification and the protein binding occurred on the captureregion (6.25 6 0.5 mm2) underneath the microfluidic channel. C. A set of multi-SiNW-FET units under the capture region. Each unit contains 5–10 SiNWs. Thelength and diameter of each SiNW are 2 mm and 20 nm, respectively.
This journal is � The Royal Society of Chemistry 2013 Lab Chip, 2013, 13, 676–684 | 677
Lab on a Chip Paper
Dow
nloa
ded
by N
atio
nal T
aiw
an U
nive
rsity
on
25 J
anua
ry 2
013
Publ
ishe
d on
31
Oct
ober
201
2 on
http
://pu
bs.r
sc.o
rg |
doi:1
0.10
39/C
2LC
4077
2HView Article Online

substrate area. In the following step of immobilizing GST-conjugated proteins on the GSH/SiNW-FET for subsequentsensing measurements, the GST-conjugated proteins wereanchored in all areas (6.25 6 0.5 mm2) underneath the PDMSmicrofluidic channel on the biosensor chip containing SiNW-FET arrays. The zone that binds the target proteins wasdesignated as the ‘‘capture region’’, as illustrated schemati-cally in Fig. 1.
SiNW-FET electrical measurements
All sensing measurements were performed using a detectionsystem that combines a current preamplifier (1211, DLInstrument) and a lock-in amplifier (SR830, StanfordResearch System) operating at a source–drain voltage (Vds) of30 mV, a modulation frequency of 79 Hz, and a time constantof 100 ms. A Ag/AgCl electrode (MF2052, BAS) connected to thePDMS microfluidic channel was used as a solution gate. Tominimize electrical noise, this microfluidic channel systemwas sustained at a ground potential throughout the entire real-time electrical measurements (Fig. 1). Every reaction wasrepeated using three different SiNW-FET devices. The con-ductance response curves shown in each subfigure wereobtained using the same SiNW-FET device.
Elution of bound GST-conjugated proteins
After the detection of the interacting proteins using a SiNW-FET, the complexes of the interacting protein and the GST-conjugated protein were eluted with GSH (1 mM). The elutedcomplexes were from the entire capture region (6.25 6 0.5mm2) on the biosensor chip (Fig. 1), including those bound onthe SiNW surface and on the surrounding substrate region.
DNA constructs
To generate the DNA constructs for the overexpression of GST–CaM, GST–CaM1234, and GST–CalnIDHT (CalnI without theC-terminal hydrophobic tail) in Escherichia coli (E. coli), CaM,CaM1234, and CalnIDHT were amplified from CaM–pcDNA3.1,CaM1234–pcDNA3.1, and CalnIDHT–pcDNA3.1,17 respectively,using the polymerase chain reaction and then subcloned intopGEX4T-1 (GE Healthcare). The GFP–CaMKIIa plasmid was agenerous gift from Professor Tobias Meyer (StanfordUniversity, CA, USA).
To generate the DNA constructs for the overexpression ofGST–CaM in 293T cells, GST–CaM was amplified from CaM–pGEX4T-1 by PCR and then subcloned into pcDNA3.1.
GST fusion protein purification
Overnight cultures of BL21 cells transformed with CaM–pGEX4T-1, CaM1234–pGEX4T-1, or CalnIDHT–pGEX4T-1 werediluted 100-fold with fresh loading broth supplemented with100 mg ml21 ampicillin. The cultures were incubated at 30 uCuntil the OD600 reached 0.6, at which point isopropyl b-D-1-thiogalactopyranoside (IPTG) was added to a final concentra-tion of 1 mM. The cultures were then incubated at 30 uCovernight. Cells were harvested, resuspended in phosphatebuffered saline (PBS, containing 137 mM NaCl, 2.7 mM KCl, 10mM Na2HPO4, and 2 mM KH2PO4, pH 7.4) and lysed bysonication. To dissolve the overproduced GST-fusion proteins,Triton X-100 (1%) was added into the homogenates at 4 uC for
30 min. The homogenates were centrifuged at 15 400g, 4 uC for10 min, and the supernatants were subjected to affinitypurification using GSH sepharose 4B (GE Healthcare). TheGST-fusion proteins were eluted with reduced GSH (10 mM)and analyzed using dodecylsulfate polyacrylamide gel electro-phoresis (SDS-PAGE). The concentrations of the purifiedproteins were determined using a Bradford protein assay(Bio-Rad, Hercules, CA).
Cell lysate preparation
293T cells (1 6 107) were transiently transfected withpcDNA3.1, GST–CaM, GFP, or GFP–CaMKIIa usingLipofectamineTM 2000 as described in the manufacturer’sinstructions. One day after transfection, the cells wereharvested and lysed for 30 min on ice in RIPA lysis buffer(150 mM NaCl, 1% NP-40, 0.5% sodium deoxycholate, 1% SDS,50 mM Tris, pH 7.5). For the neuronal cell preparation, apregnant Sprague-Dawley rat (E14.5) was sacrificed with aprocedure that complied with the animal welfare regulationsof the National Taiwan University. The embryo corticalneurons were isolated as described previously18 and lysed for30 min on ice in RIPA buffer. The cell lysates were centrifugedat 15 400g, 4 uC for 10 min and the supernatants were dilutedto a concentration of 0.1 mg ml21 with PS for SiNW-FETmeasurements. The protein concentrations were determinedby the Bradford protein assay.
Western blot analysis
Cell lysates or purified proteins were subjected to SDS-PAGEanalysis and then transferred to polyvinylidene fluoridemembranes. The membranes were incubated in PBST block-ing buffer (PBS buffer containing 5% nonfat dry milk and0.1% Tween 20) at room temperature for 1 h. The membraneswere probed with appropriate dilutions of the primaryantibody in PBST at room temperature for 1 h. The unboundantibody was removed by extensive washes with PBST. Thewashed membranes were incubated with horseradish perox-idase-conjugated secondary antibody in PBST at room tem-perature for 1 h. Following extensive washes with PBST, theantibody-reacted proteins were visualized by chemilumines-cence using ECL Western blotting detection reagents.
Mass spectrometry
Samples were dissolved in 0.1% anionic acid labile surfactant(Protea, WV, USA) and vortexed for 10 s. Subsequently, theproteins were reduced in 5 mM dithiothreitol (DTT) at 56 uCfor 30 min and then alkylated in 55 mM iodoacetamide atroom temperature in the dark for 30 min. The proteins weredigested overnight with trypsin at 37 uC. After acidification bytrifluoroacetic acid, the peptide mixtures were collected anddesalted on Oasis HLB cartridges (Waters, Milford, MA). Theeluted peptides were dried using a SpeedVac for LC-MS/MSanalysis. All dried fractions were analyzed on a LTQ-FT Ultra(Thermo Fisher Scientific) equipped with a nanoelectrosprayion source (New Objective, Woburn, MA, USA).Chromatographic separation was performed on a self-packedreversed phase C18 nano-column (75 mm I.D. 6 300 mm, 5mm, 100 Å) at 300 ml min21. Full-scan MS spectra (m/z 300–1800) were acquired with a mass resolution of 100 000 at m/z
678 | Lab Chip, 2013, 13, 676–684 This journal is � The Royal Society of Chemistry 2013
Paper Lab on a Chip
Dow
nloa
ded
by N
atio
nal T
aiw
an U
nive
rsity
on
25 J
anua
ry 2
013
Publ
ishe
d on
31
Oct
ober
201
2 on
http
://pu
bs.r
sc.o
rg |
doi:1
0.10
39/C
2LC
4077
2HView Article Online

400. The obtained spectra were analyzed using Mascotsoftware version 2.2 (Matrix Science) against the proteinsequences of GST–CaM, GFP–CaMKIIa, GST–CalnIDHT, orSwiss-Prot database (Rattus norvegicus, total of 524 420sequences).
Results and discussion
Ca2+ chelation determines the interaction capabilities of CaMand other Ca2+-binding proteins to their targets. CaM has 4Ca2+ binding sites; the key amino acids, 2 for each site, weremutated in CaM1234 to block the Ca2+ binding ability.14 Toverify if the SiNW-FET is sufficiently sensitive to discriminatethe Ca2+ binding status of CaM, both GST–CaM and GST–CaM1234 were expressed in E. coli and purified with amolecular weight of y43 kDa, as indicated by a Western blot(Fig. 2A). The binding of either GST–CaM or GST–CaM1234 ontothe GSH/SiNW-FET induced a similar conductance increase,y7%, in a Ca2+-free buffer (Fig. 2B). Increasing the concentra-tion of Ca2+ to 100 mM only resulted in a small conductancedrop (y1%) in the GST–CaM1234-modified SiNW-FET(CaM1234/SiNW-FET), but a pronounced drop (y7%) in the
CaM/SiNW-FET (Fig. 2C). The CaM1234 might have a residualCa2+ binding ability and induces a small conductance change.Removing the Ca2+ from the buffer returned the conductanceto the original values (data not shown).
The interaction between CaM and TnI, a protein involved inmuscle contraction, is Ca2+-dependent.13 To verify the effec-tiveness of the SiNW-FET for detecting the Ca2+-dependentinteraction between CaM and TnI, TnI was applied onto theCaM/SiNW-FET or CaM1234/SiNW-FET in the presence orabsence of Ca2+. The conductance of the CaM/SiNW-FETdecreased by y1.5% in the presence of Ca2+ when TnI enteredthe device but was not changed for the CaM1234/SiNW-FET(Fig. 2D). These results indicate that the SiNW-FET is sensitivefor not only discriminating the Ca2+-binding status of aprotein but also recognizing Ca2+-dependent protein–proteininteractions.
Cell lysates containing proteins of interest are usuallyadopted for biomolecular interaction studies by techniques,like GST pull-down, which use GST-tagged proteins to verifythe interaction, or immunoprecipitation, which appliesspecific antibodies to identify the bound molecules. To verifythe ability of SiNW-FETs to identify molecular interactions incell lysates, GST–CaM and GFP-tagged CaMKIIa were sepa-
Fig. 2 Ca2+-dependent interaction between CaM and TnI. A. Purified GST-tagged proteins. The GST-tagged CaM (GST–CaM) and CaM1234 (GST–CaM1234) expressed inE. coli were analyzed by SDS-PAGE. The left lane is the protein marker with the marked band at 43 kDa. B. Real-time electrical measurement. GST–CaM (blue) or GST–CaM1234 (red), 400 nM in PS supplemented with EDTA, was applied to a GSH/SiNW-FET, as indicated by the arrow, and the conductance response was recorded. C.Effects of Ca2+ binding. Buffer containing 100 mM Ca2+ was applied to the CaM/SiNW-FET or CaM1234/SiNW-FET, as indicated by the arrow. D. Interaction betweenCaM and TnI. Cardiac TnI (150 nM) was applied to the CaM/SiNW-FET or CaM1234/SiNW-FET in the presence or absence of 100 mM Ca2+, as indicated by the arrow.
This journal is � The Royal Society of Chemistry 2013 Lab Chip, 2013, 13, 676–684 | 679
Lab on a Chip Paper
Dow
nloa
ded
by N
atio
nal T
aiw
an U
nive
rsity
on
25 J
anua
ry 2
013
Publ
ishe
d on
31
Oct
ober
201
2 on
http
://pu
bs.r
sc.o
rg |
doi:1
0.10
39/C
2LC
4077
2HView Article Online

Fig. 3 Detections of protein–protein interactions in cell lysates. 293T cells expressing GFP, GST–CaM, or GFP–CaMKIIa were lysed and centrifuged to collect the celllysate. A. Western blot of cell lysates with an antibody against CaM from cells expressing empty vector (mock) or GST–CaM. B. GSH/SiNW-FET response to cell lysatecontaining GST–CaM. Boiled cell lysate (0.1 mg ml21) was applied to the GSH/SiNW-FET with the conductance being recorded as the baseline. Native cell lysate (0.1mg ml21) from mock or GST–CaM expressing cells was applied, as indicated by the arrow. C. Expression of GFP–CaMKIIa. Cell lysates from cells expressing GFP or GFP–CaMKIIa were verified by Western blot with antibody against GFP. The numbers on the left indicate the molecular weight marker. D. Interaction between CaM andCaMKIIa. Lysate (0.1 mg ml21) containing GFP or GFP–CaMKIIa in the presence or absence of Ca2+ was applied onto the CaM/SiNW-FET, as indicated by the arrow,and the conductance was measured. E. MS analysis of the bound molecules. The bound molecules eluted from the capture region of the SiNW-FET biosensor chipwere analyzed by MS. The identified peptide fragments and the coverage of GST–CaM and GFP–CaMKIIa are listed. Lengths for the fused proteins: GST–CaM: GST 1–227 and CaM 228–375; GFP–CaMKIIa: GFP 1–239 and CaMKIIa 267–745.
680 | Lab Chip, 2013, 13, 676–684 This journal is � The Royal Society of Chemistry 2013
Paper Lab on a Chip
Dow
nloa
ded
by N
atio
nal T
aiw
an U
nive
rsity
on
25 J
anua
ry 2
013
Publ
ishe
d on
31
Oct
ober
201
2 on
http
://pu
bs.r
sc.o
rg |
doi:1
0.10
39/C
2LC
4077
2HView Article Online

rately expressed in 293T cells, from which the cell lysates werecollected. Tagged GFP may provide a green fluorescence signalto visualize the binding of CaMKIIa to the device. Theexpression of GST–CaM was verified using a Western blot asa single band at y43 kDa (Fig. 3A). The conductance of theGSH/SiNW-FET in the presence of denatured cell lysates wasused as a baseline. No elevation was observed when controllysates (cells not expressing GST–CaM) were added; afteradding a cell lysate containing GST–CaM, the conductance ofthe same SiNW-FET device was elevated by y3% (Fig. 3B).These results suggest that the GSH/SiNW-FET specificallyinteracted with the overexpressed GST–CaM in the cell lysate.
CaMKII, a serine/threonine protein kinase, is activated bybinding to Ca2+-bound CaM (Ca2+/CaM).19 Cell lysate contain-ing overexpressed GFP or GFP–CaMKIIa in the presence orabsence of Ca2+ (100 mM) was added to a CaM/SiNW-FET, andthe conductance was measured. The appearance of GFP (y24kDa) and GFP–CaMKIIa (y82 KDa) in the cell lysates wasidentified using a Western blot with an antibody against GFP(Fig. 3C). The conductance of the CaM/SiNW-FET using adenatured cell lysate was used as a baseline. Lysates withoutCa2+ or containing only GFP did not induce any significantconductance change. The addition of a lysate containingoverexpressed GFP–CaMKIIa in the presence of Ca2+ elevatedthe conductance of CaM/SiNW-FET by y4% (Fig. 3D). Theseresults indicate that protein binding to the surface-modifiedSiNW-FET is both specific and sensitive and that the SiNW-FET has the ability to verify suspected interacting proteinsfrom cell lysates.
The charge effect on the SiNW determines the increase ordecrease in the conductance of the SiNW-FET. The SiNW usedin this report is boron-doped and is a p-type semiconductor.Therefore, the binding of negatively charged GST–CaM orGST–CaM1234 (both have isoelectric points (pI) of y5) to theGSH/SiNW-FET at pH 7.4 caused an increase in conductance(Fig. 2B). Conversely, the binding of Ca2+ to the CaM/SiNW-FET, but not to CaM1234/SiNW-FET, resulted in a decrease inthe conductance (Fig. 2C). Likewise, the bindings of TnI (pIy9) and CaMKIIa (pI y6) to the Ca2+/CaM/SiNW-FETdecreased (Fig. 2D) and increased (Fig. 3D) the conductance,respectively, further supporting that the SiNW-FET is a p-typetransistor.
Conventional GST pull-down assays require 25 ml of GSH–agarose beads (surface area y750 mm2), y10 mg of a GST-tagged protein, a lysate prepared from 107 cells, and a lengthyWestern blot procedure. All of these components are requiredto detect the protein–protein interaction. In comparison, theone-dimensional, ultrasensitive SiNW-FET nanowire biosensorrequires only a very small amount of sample. In the providedexamples, less than 1 mg of purified GST–CaM (Fig. 2) and alysate prepared from 104 cells expressing GST–CaM or GFP–CaMKIIa (Fig. 3) were sufficient for the SiNW-FET-basedprotein binding assay. The amount of bound interactingproteins eluted from the capture region of the SiNW-FETbiosensor (6.25 6 0.5 mm2, Fig. 1) was below the thresholdamount for subsequent identification by Western blot analysis
(data not shown). Therefore, we utilized MS to analyze thebound proteins on the SiNW-FET device.
The immobilized GST-conjugated proteins on the GSH/SiNW-FET can easily be eluted with high concentrations ofGSH (¢1 mM). By examining the charged fragmentarypeptides, MS analysis provides a fast and very sensitiveapproach for identifying the interacting proteins.20 Followingthe binding detected in Fig. 2D, the MS fragmentation spectra(Fig. S13) of the eluted solution revealed that the majorproteins were GST–CaM and CaMKIIa (Fig. 3E). These resultsconfirm that the conductance change was induced by thespecific interaction between GST–CaM and CaMKIIa but notfrom other molecules in the lysate.
The binding of the GST–CaM and CaMKIIa proteins fromthe cell lysates to the GSH/SiNW-FET and the Ca2+/CaM/SiNW-FET, respectively, extend the applications of the SiNW-FET topractical biological assays. Unlike traditional immunoprecipi-tation or GST pull-down assays, which require tediousprocedures and specific antibodies to detect protein–proteininteractions, the functionalized SiNW-FET directly detects theoccurrence of molecular interactions by monitoring itsconductivity change.
CalnI, a CaM-like protein, is expressed in the brain at a latedevelopmental stage21 and is involved in modulating synaptictransmission.15 CalnI has two functional EF-hand motifs at
Fig. 4 Schematic illustration of the integration of MS with a SiNW-FET forscreening interacting proteins. To apply the SiNW-FET as a screening system toidentify the proteins interacting with CalnIDHT, GST–CalnIDHT is bound onto aGSH/SiNW-FET and neuronal cell lysate is applied. After washing out the non-specifically adhesive molecules on the SiNW-FET surface, the bound moleculesare eluted and analyzed by MS.
This journal is � The Royal Society of Chemistry 2013 Lab Chip, 2013, 13, 676–684 | 681
Lab on a Chip Paper
Dow
nloa
ded
by N
atio
nal T
aiw
an U
nive
rsity
on
25 J
anua
ry 2
013
Publ
ishe
d on
31
Oct
ober
201
2 on
http
://pu
bs.r
sc.o
rg |
doi:1
0.10
39/C
2LC
4077
2HView Article Online

the N-terminus for Ca2+ binding and a hydrophobic segmentat the C-terminus for membrane localization.22 It has beenreported that CalnI inhibits Ca2+ currents17 and regulatesGolgi–plasma membrane trafficking.23 To investigate themechanisms underlying these phenomena, we used theSiNW-FET device together with MS to identify the CalnI-interacting proteins in the neuronal lysate.
Fig. 4 illustrates the experimental design of integrating MSwith the SiNW-FET for fast screening of the interactingproteins. GST–CalnIDHT was immobilized on a GSH/SiNW-FET. Neuronal cell lysate was added, and the conductancechange of the SiNW-FET was monitored to observe theoccurrence of protein–protein interactions. After washing outthe non-specifically bound molecules, the bound targetmolecules were eluted and analyzed by MS. A similar approachto identify interacting proteins has been performed using SPRas the sensing device.24 However, compared to SPR, the SiNWdevice has advantages such as higher sensitivity, lowerquantity of protein required, and the potential for very largeintegrated arrays.25,26 Therefore, the conjugation of NW-FETand MS may be a promising tool for screening biomolecularinteractions.
Fig. 5 shows the conductance recording of the entireprocedure. GST–CalnIDHT was expressed in E. coli andpurified. The completion of this step was confirmed by SDS-PAGE because the major band was observed at y46 kDa
(Fig. 5A). The modification of GST–CalnIDHT on the GSH/SiNW-FET to form CalnIDHT/SiNW-FET caused an increase inthe conductance by y2% (Fig. 5B). The addition of Ca2+ (100mM) decreased the conductance of the CalnIDHT/SiNW-FET byy1% (Fig. 5C). To identify the molecules interacting withCalnI, neuronal cell lysate was added to CalnIDHT/SiNW-FET,and the conductance gradually increased by y1% (Fig. 5D).Note that for the SiNW-FET modified with only GST in thepresence of Ca2+ (Ca2+/GST/SiNW-FET), neuronal cell lysate didnot induce any significant conductance change on the samedevice (Fig. S23). The delay in the conductance change after theintroduction of cell lysates might be due to the competitionbetween the target proteins and nonspecific proteins. AfterGSH elution of the bound proteins in Fig. 5D, the majority ofthe elute was identified to be the derivatives of GST–CalnIDHT,as analyzed by MS (Fig. S13).
The identified candidates with sequence coverage of .2%and a mean error of ,2 ppm are summarized in Table S13.According to their reported functions, these proteins weredivided into seven categories: cytoskeletal regulation, vesicletransportation, protein folding, signal transduction, glycolysis,protein biosynthesis, and others. These results hint thepossible functions of CalnI involved in various physiologicalprocesses. However, further biological experiments need to beperformed to verify the roles of CalnI in modulating thesebiological functions.
Fig. 5 Identification of CalnI-interacting proteins from neuronal lysate. GST–CalnIDHT was expressed in E. coli and purified for the SiNW-FET binding assay. A. PurifiedGST–CalnIDHT characterized by SDS-PAGE and Coomassie blue staining. The left lane represents the protein marker with molecular weights shown on the left. B.Conductance change induced by GST–CalnIDHT. GST–CalnIDHT (400 nM) in Ca2+-free buffer was applied, as indicated by the arrow, onto the GSH/SiNW-FET and theconductance was measured. C. Effects of Ca2+ binding. Buffer containing 100 mM Ca2+ was applied, as indicated by the arrow, onto the CalnIDHT/SiNW-FET. D.Interaction between CalnIDHT and the rat neuronal cell lysate. Cell lysate (0.1 mg ml21) isolated from embryonic rat cortical neurons in the presence of Ca2+ wasapplied onto the Ca2+/CalnIDHT/SiNW-FET, as indicated by the arrow, and the conductance was measured. E. Interaction between CalnIDHT and Hsp70. PurifiedHsp70 (200 nM) was applied onto the CalnIDHT/SiNW-FET in the presence (Ca2+/CalnIDHT/SiNW-FET) or absence (CalnIDHT/SiNW-FET) of 100 mM Ca2+; or Ca2+/GST/SiNW-FET, as indicated by the arrow.
682 | Lab Chip, 2013, 13, 676–684 This journal is � The Royal Society of Chemistry 2013
Paper Lab on a Chip
Dow
nloa
ded
by N
atio
nal T
aiw
an U
nive
rsity
on
25 J
anua
ry 2
013
Publ
ishe
d on
31
Oct
ober
201
2 on
http
://pu
bs.r
sc.o
rg |
doi:1
0.10
39/C
2LC
4077
2HView Article Online

Using MS to identify the interacting molecules captured bythe receptor-modified SiNW-FET provides a novel strategy forfast screening. We used CalnIDHT as ‘‘bait’’ to capturemolecules from the neuronal cell lysate that were lateridentified by MS. To maximize the binding capacity, purifiedGST–CalnIDHT and cell lysates isolated from the E14.5 ratembryo cortex were used. The small observed conductancechange (y1%, Fig. 5D) induced by the neuronal lysate mightbe due to the bindings of various proteins with different pIvalues. Therefore, the elute contained many proteins, each in asmall amount, which were below the threshold for detectionby MS analysis. To collect a sufficient sample for MS analysis,the binding and washing procedures were repeated ten timesand pooled. The total neuron lysate used was approximately0.1 mg (y105 cells).
The identified interacting molecules could then be furtherconfirmed using the same protocol via the SiNW-FETdetection as discussed above. Hsp70 is involved in proteinfolding and has been reported to modulate Ca2+ channelactivity.27 To confirm that CalnI interacts directly with thecytosolic Hsp70, purified Hsp70 was applied to the CalnIDHT/SiNW-FET. The conductance increased by y2% in thepresence, but not in the absence of Ca2+. In addition,Hsp70 did not induce any conductance change in the Ca2+/GST/SiNW-FET (Fig. 5E). As further proof, denatured Hsp70has no effect on the conductance of the Ca2+/CalnIDHT/SiNW-FET (Fig. S33). Furthermore, we examined the binding ofneuronal cell lysate to the Ca2+/GST/SiNW-FET using the sameprotocol. Only a few proteins were identified (Table S23), andthey were different from those listed in Table S13. Our SiNW-FET biosensor provided both a convenient screening platformand a fast checking protocol to confirm previously identifiedproteins.
Conclusions
We have demonstrated that in tandem with MS, SiNW-FETscan serve as a promising fast biosensing platform to screeninteraction candidates from crude cell lysates. Compared tothe traditional techniques, this approach provides fast,sensitive, and accurate detections. By monitoring the con-ductance change of the SiNW-FET, we can confirm theinteraction at each screening step before going to the nextone. Furthermore, the identified candidate protein can oncemore be analyzed by the SiNW-FET to verify if the interactionis direct or indirect. In addition, by using these methods, wehave made the first step toward elucidating the neuronalinteraction network of CalnI with its binding partners.
Acknowledgements
This study was supported by the National Science Council ofTaiwan under Grant Nos. NSC 100-2627-M-002-012, NSC 100-2627-M-002-013, and NSC 99-2628-B-002-054-MY3. Technical
support from NanoCore, the Core Facilities for Nanoscienceand Nanotechnology at Academia Sinica, is acknowledged.
References
1 R. D. Gietz, B. Triggs-Raine, A. Robbins, K. C. Graham andR. A. Woods, Mol. Cell. Biochem., 1997, 172, 67–79.
2 J. A. Brzostowski, T. Meckel, J. Hong, A. Chen and T. Jin,Curr. Protoc. Protein Sci., 2009, ch. 19, unit 19 15.
3 V. Kodoyianni, BioTechniques, 2011, 50, 32–40.4 K.-I. Chen, B.-R. Li and Y.-T. Chen, Nano Today, 2011, 6,
131–154.5 F. Patolsky, G. Zheng and C. M. Lieber, Nat. Protoc., 2006, 1,
1711–1724.6 T. W. Lin, P. J. Hsieh, C. L. Lin, Y. Y. Fang, J. X. Yang, C.
C. Tsai, P. L. Chiang, C. Y. Pan and Y. T. Chen, Proc. Natl.Acad. Sci. U. S. A., 2010, 107, 1047–1052.
7 S. P. Lin, C. Y. Pan, K. C. Tseng, M. C. Lin, C. D. Chen, C.C. Tsai, S. H. Yu, Y. C. Sun, T. W. Lin and Y. T. Chen, NanoToday, 2009, 4, 235–243.
8 X. Bi, W. L. Wong, W. Ji, A. Agarwal, N. Balasubramanianand K. L. Yang, Biosens. Bioelectron., 2008, 23, 1442–1448.
9 G. J. Zhang, J. H. Chua, R. E. Chee, A. Agarwal and S.M. Wong, Biosens. Bioelectron., 2009, 24, 2504–2508.
10 T. S. Pui, A. Agarwal, F. Ye, N. Balasubramanian andP. Chen, Small, 2009, 5, 208–212.
11 F. Patolsky, B. P. Timko, G. H. Yu, Y. Fang, A. B. Greytak, G.F. Zheng and C. M. Lieber, Science, 2006, 313, 1100–1104.
12 G. F. Zheng, F. Patolsky, Y. Cui, W. U. Wang and C.M. Lieber, Nat. Biotechnol., 2005, 23, 1294–1301.
13 C. H. Keller, B. B. Olwin, D. C. LaPorte and D. R. Storm,Biochemistry, 1982, 21, 156–162.
14 B. Z. Peterson, C. D. DeMaria, J. P. Adelman and D. T. Yue,Neuron, 1999, 22, 549–558.
15 L. P. Haynes, H. V. McCue and R. D. Burgoyne, Front. Mol.Neurosci., 2012, 5, 9.
16 Z. Y. Fan, J. C. Ho, Z. A. Jacobson, R. Yerushalmi, R.L. Alley, H. Razavi and A. Javey, Nano Lett., 2008, 8, 20–25.
17 P. Y. Shih, C. L. Lin, P. W. Cheng, J. H. Liao and C. Y. Pan,Biochem. Biophys. Res. Commun., 2009, 388, 549–553.
18 C. C. Tsai, C. Y. Pan, C. C. Yang, P. Y. Shih, C. S. Wu, C.D. Chen and Y. T. Chen, J. Phys. Chem. B, 2008, 112,9165–9173.
19 T. R. Soderling, B. Chang and D. Brickey, J. Biol. Chem.,2001, 276, 3719–3722.
20 W. M. Winnik, R. M. Dekroon, J. S. Jeong, M. Mocanu, J.B. Robinette, C. Osorio, N. N. Dicheva, E. Hamlett andO. Alzate, Methods Mol. Biol., 2012, 854, 47–66.
21 Y. Q. Wu, X. Lin, C. M. Liu, M. Jamrich and L. G. Shaffer,Mol. Genet. Metab., 2001, 72, 343–350.
22 J. Hradsky, V. Raghuram, P. P. Reddy, G. Navarro, M. Hupe,V. Casado, P. J. McCormick, Y. Sharma, M. R. Kreutz andM. Mikhaylova, J. Biol. Chem., 2011, 286, 36762–36776.
23 M. Mikhaylova, P. P. Reddy, T. Munsch, P. Landgraf, S.K. Suman, K. H. Smalla, E. D. Gundelfinger, Y. Sharma andM. R. Kreutz, Proc. Natl. Acad. Sci. U. S. A., 2009, 106,9093–9098.
24 N. J. de Mol, Methods Mol. Biol., 2012, 800, 33–53.25 Z. Fan, J. C. Ho, Z. A. Jacobson, R. Yerushalmi, R. L. Alley,
H. Razavi and A. Javey, Nano Lett., 2008, 8, 20–25.
This journal is � The Royal Society of Chemistry 2013 Lab Chip, 2013, 13, 676–684 | 683
Lab on a Chip Paper
Dow
nloa
ded
by N
atio
nal T
aiw
an U
nive
rsity
on
25 J
anua
ry 2
013
Publ
ishe
d on
31
Oct
ober
201
2 on
http
://pu
bs.r
sc.o
rg |
doi:1
0.10
39/C
2LC
4077
2HView Article Online

26 Y. L. Bunimovich, Y. S. Shin, W. S. Yeo, M. Amori,G. Kwong and J. R. Heath, J. Am. Chem. Soc., 2006, 128,16323–16331.
27 T. Schneider, K. Radhakrishnan, A. Krieger, G. P. Dietz,F. Nagel, M. Bahr and J. Hescheler, Journal of Receptor,Ligand and Channel Research, 2011, 29.
684 | Lab Chip, 2013, 13, 676–684 This journal is � The Royal Society of Chemistry 2013
Paper Lab on a Chip
Dow
nloa
ded
by N
atio
nal T
aiw
an U
nive
rsity
on
25 J
anua
ry 2
013
Publ
ishe
d on
31
Oct
ober
201
2 on
http
://pu
bs.r
sc.o
rg |
doi:1
0.10
39/C
2LC
4077
2HView Article Online

1
Improved Silicon Nanowire Field-effect Transistors for Fast Protein-protein
Interaction Screening
(Supplementary Information)
Ti-Yu Lina, Bor-Ran Lib,c, Sheng-Ta Tsaid, Chien-Wei Chenb,c, Chung-Hsuan
Chend, Yit-Tsong Chenb,c,*, and Chien-Yuan Pana,e,*
aInstitute of Zoology, National Taiwan University, Taipei, Taiwan bDepartment of Chemistry, National Taiwan University, Taipei, Taiwan cInstitute of Atomic and Molecular Sciences, Academia Sinica, Taipei, Taiwan dGenomics Research Center, Academia Sinica, Taipei, Taiwan
eDepartment of Life Science, National Taiwan University, Taipei, Taiwan
* CORRESPONDING AUTHORS Dr. YT Chen, Department of Chemistry, National Taiwan University and Institute of Atomic and Molecular Sciences, Academia Sinica, P.O. Box 23-166, Taipei 106, Taiwan; E-mail: [email protected], Fax: (+886) 2-2363-6359 Dr. CY Pan, Institute of Zoology and Department of Life Science, National Taiwan University, 1 Sec. 4, Roosevelt Road, Taipei 106, Taiwan; E-mail: [email protected], Fax: (+886) 2-2363-6837
Electronic Supplementary Material (ESI) for Lab on a ChipThis journal is © The Royal Society of Chemistry 2012
2
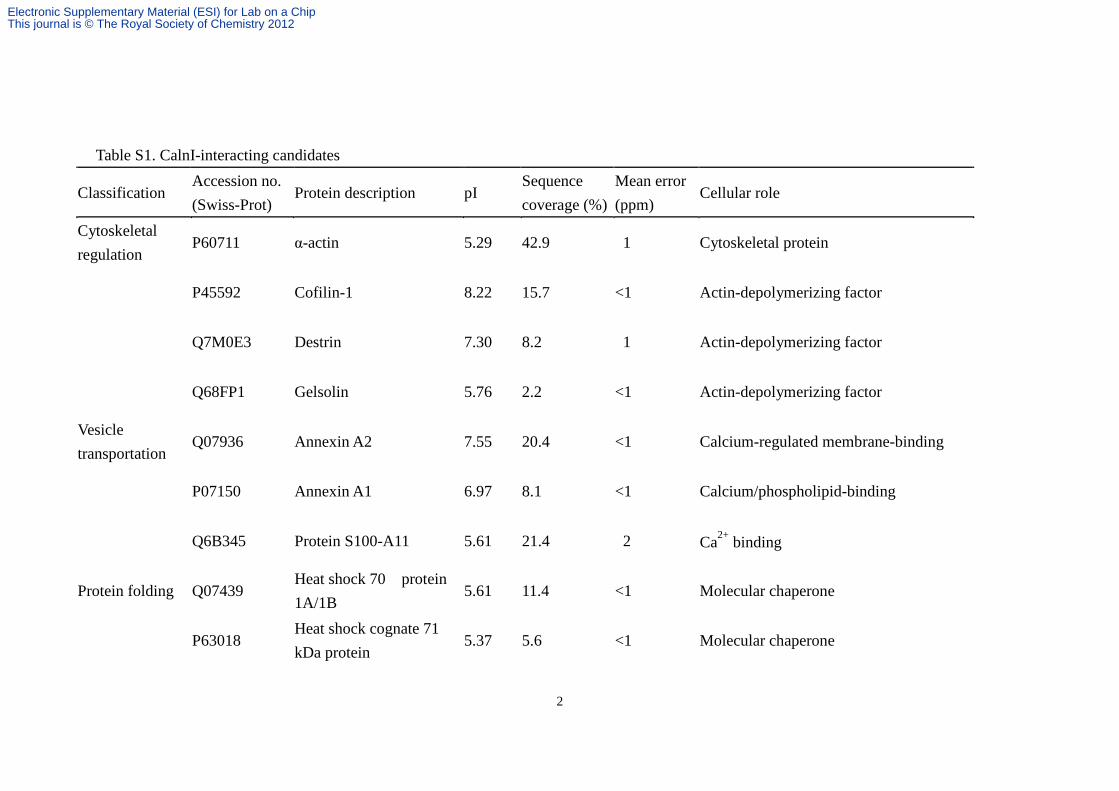
Table S1. CalnI-interacting candidates
Classification Accession no. (Swiss-Prot)
Protein description pI Sequence coverage (%)
Mean error (ppm)
Cellular role
Cytoskeletal regulation
P60711 α-actin 5.29 42.9 1 Cytoskeletal protein
P45592 Cofilin-1 8.22 15.7 <1 Actin-depolymerizing factor
Q7M0E3 Destrin 7.30 8.2 1 Actin-depolymerizing factor
Q68FP1 Gelsolin 5.76 2.2 <1 Actin-depolymerizing factor
Vesicle transportation
Q07936 Annexin A2 7.55 20.4 <1 Calcium-regulated membrane-binding
P07150 Annexin A1 6.97 8.1 <1 Calcium/phospholipid-binding
Q6B345 Protein S100-A11 5.61 21.4 2 Ca2+ binding
Protein folding Q07439 Heat shock 70 protein 1A/1B
5.61 11.4 <1 Molecular chaperone
P63018 Heat shock cognate 71 kDa protein
5.37 5.6 <1 Molecular chaperone
Electronic Supplementary Material (ESI) for Lab on a ChipThis journal is © The Royal Society of Chemistry 2012

3
P04785 Protein disulfide-isomerase
4.82 3.1 <1 Catalyzes the rearrangement of disulfide bonds
P42930 Heat shock protein beta-1 6.12 4.9 <1 Molecular chaperone
P10111 Peptidyl-prolyl cis-trans isomerase A
8.34 8.5 <1 Catalyzes the cis-trans isomerization of proline imidic peptide bonds
Signal transduction
P63102 14-3-3 protein zeta/delta 4.73 16.3 <1 Adapter protein
P68511 14-3-3 protein eta 4.81 6.5 2 Adapter protein
P38983 40S ribosomal protein SA 4.80 14.9 2 Receptor for laminin
P09527 Ras-related protein Rab-7a
6.80 6.4 <1 Small GTPase signaling
Glycolysis P04764 α-enolase 6.16 10.6 <1 Glycolytic enzyme
P48500 Triosephosphate isomerase
6.89 11.2 1 Glycolytic enzyme
P04642 L-lactate dehydrogenase A chain
8.45 8.1 1 Glycolytic enzyme
P25113 Phosphoglycerate mutase 1
6.67 8.3 1 Glycolytic enzyme
Electronic Supplementary Material (ESI) for Lab on a ChipThis journal is © The Royal Society of Chemistry 2012

4
Protein biosynthesis
P05197 Elongation factor 2 6.41 4.3 2 Catalyzes ribosomal translocation during translation elongation
P62630 Elongation factor 1-alpha 1
9.10 7.6 1 Promotes the binding of aminoacyl-tRNA to the A-site of ribosomes
Others Q6P0K8 Junction plakoglobin 5.75 26.3 1 Desmosome protein
P62982 Ubiquitin-40S ribosomal protein S27a
9.68 10.3 <1 Proteolysis
P55053 Fatty acid-binding protein, epidermal
6.73 6.7 1 Lipid binding
Neuronal cell lysate was passed through the CalnIΔHT modified SiNW-FET in the presence of Ca2+ and the bound proteins were eluted for MS analysis. Proteins with sequence coverage of > 2% and a mean error of < 2% are listed and classified into 7 categories. The pIs and brief descriptions of functions are provided.
Electronic Supplementary Material (ESI) for Lab on a ChipThis journal is © The Royal Society of Chemistry 2012

5
Table S2. Nonspecific binding of the neuronal lysate to the GST/SiNW-FET device.
Accession no.
(Swiss-Prot) Protein description pI
Sequence
coverage (%) Mean error
(ppm) P68370 Tubulin α1A chain 4.94 10.6 0 P06302 Prothymosin α 3.75 25.9 <1 P69897 Tubulin β5 chain 4.78 6.8 <1 P02091 Hemoglobin subunit β1 7.88 15.6 <1 P15865 Histone H1.2 11.1 7.3 1 P63312 Thymosin β10 5.31 31.8 1 P62161 CaM 4.09 8.7 1 P62329 Thymosin β 5.02 31.8 1
The molecules in the neuronal lysate that was bound onto the GST/SiNW-FET were
eluted and used for MS analysis. Proteins with sequence coverage of > 2% and mean
errors of < 2% are listed. pI: isoelectric point
Electronic Supplementary Material (ESI) for Lab on a ChipThis journal is © The Royal Society of Chemistry 2012

6
Figure S1. The MS fragmentation spectra of the eluted GST-CaM, GFP-CaMKIIα,
and GST-CalnIΔHT.
GST-CaM
GFP-CaMKIIα
GST-CalnI∆HT
Electronic Supplementary Material (ESI) for Lab on a ChipThis journal is © The Royal Society of Chemistry 2012

7
Figure S2. A GST/SiNW-FET did not respond to the neuronal cell lysate in the
presence of Ca2+.
0 5 10 15 20 25-0.4
-0.2
0.0
0.2
0.4
∆G (%
)
Time (Min)
Neuronal Cell Lysate
Ca2+/∆ST/SiNW-FET
Electronic Supplementary Material (ESI) for Lab on a ChipThis journal is © The Royal Society of Chemistry 2012

8
Figure S3. CalnI did not interact with the denatured Hsp70. Denatured Hsp70, which
was boiled in water for 10 min, had no effect on the electrical conductance of the
Ca2+/CalnIΔHT/SiNW-FET. After replacing the denatured Hsp70 with native Hsp70,
an increase in the conductance was observed.
0 3 6 9
0
1
2
∆G (%
)
Time (Min)
Denatured Hsp70 Hsp70
Ca2+/CalnI∆HT/SiNW-FET
Electronic Supplementary Material (ESI) for Lab on a ChipThis journal is © The Royal Society of Chemistry 2012


















