
KERAGAMAN GEN MAJOR HISTOCOMPATIBILITY COMPLEX … · ketahanan penyakit, respon imun dan sifat...
46
KERAGAMAN GEN MAJOR HISTOCOMPATIBILITY COMPLEX (MHC) DRB3.2 LOCUS PSTI PADA POPULASI KAMBING KACANG SKRIPSI Oleh EVY HARJUNA SAAD I 111 11 256 FAKULTAS PETERNAKAN UNIVERSITAS HASANUDDIN MAKASSAR 2015
Transcript of KERAGAMAN GEN MAJOR HISTOCOMPATIBILITY COMPLEX … · ketahanan penyakit, respon imun dan sifat...

KERAGAMAN GEN MAJOR HISTOCOMPATIBILITY
COMPLEX (MHC) DRB3.2 LOCUS PSTI PADA
POPULASI KAMBING KACANG
SKRIPSI
Oleh
EVY HARJUNA SAAD
I 111 11 256
FAKULTAS PETERNAKAN
UNIVERSITAS HASANUDDIN
MAKASSAR
2015

KERAGAMAN GEN MAJOR HISTOCOMPATIBILITY
COMPLEX (MHC) DRB3.2 LOCUS PSTI PADA
POPULASI KAMBING KACANG
SKRIPSI
Oleh
EVY HARJUNA SAAD
I 111 11 256
Skripsi sebagai Salah Satu Syarat untuk Memperoleh
Gelar Sarjana Peternakan pada Fakultas Peternakan
Universitas Hasanuddin
FAKULTAS PETERNAKAN
UNIVERSITAS HASANUDDIN
MAKASSAR
2015

PERNYATAAN KEASLIAN
1. Yang bertandatangan dibawah ini:
Nama : Evy Harjuna Saad
NIM : I 111 11 256
Menyatakan dengan sebenarnya bahwa:
a. Karya skripsi yang saya tulis adalah asli
b. Apabila sebagian atau seluruhnya dari karya skripsi, terutama dalam Bab
Hasil dan Pembahasan, tidak asli atau plagiasi maka bersedia dibatalkan
dan dikenakan sanksi akademik yang berlaku.
2. Demikian pernyataan keaslian ini dibuat untuk dapat digunakan seperlunya.
Makassar, Desember 2015
Evy Harjuna Saad

i
HALAMAN PENGESAHAN
Judul Penelitian : Keragaman Gen Major Histocompatibility
(MHC) DRB3.2 Locus PstI pada PopulKacang
Nama : Evy Harjuna Saad
Nomor Induk Mahasiswa : I 111 11 256
Fakultas : Peternakan
Skripsi ini telah diperiksa dan disetujui Oleh:
Prof. Rr. Sri Rachma AB., M. Sc, Ph. D Dr.Muh. Ihsan A. Dagong,
Pembimbing Utama Pembimbing Anggot

ii
KATA PENGANTAR
Puji dan syukur penulis panjatkan ke hadirat Allah SWT, karena atas berkat
rahmat, cinta, dan taufik-Nya sehingga dapat menyelesaikan skripsi dengan judul
“Keragaman Gen Major Histocompatibility Complex (MHC) DRB3.2 Locus PstI
pada Populasi Kambing Kacang”.
Melalui kesempatan ini penulis dengan rendah hati mengucapkan terima
kasih kepada semua pihak yang telah membantu dan membimbing penulis dalam
menyelesaikan skripsi ini kepada:
1. Ibu Prof. Rr. Sri Rachma AB., M. Sc, Ph. D selaku Pembimbing Utama dan
bapak Dr. Muh. Ihsan A. Dagong, S.Pt., M.Si. selaku Pembimbing Anggota,
atas segala bantuan dan keikhlasannya untuk memberikan bimbingan, nasehat
dan saran-saran sejak awal penelitian sampai selesainya penulisan skripsi ini.
2. Kedua orang tua tercinta ayahanda Saade, S. Pd., ibunda St.Norma, S. Pd.
yang memberikan cinta kasih, dukungan mental dan memberikan doa
restunya dan saudara-saudaraku Helmy Saad, S.H., Sardiman Saad, Ewin
Juspiadi Saad, dan Asmul Shahman serta kakak ipar penulis Andi Batari Ayu,
S.Kom yang telah memberikan motivasi untuk selalu lebih semangat dan
kemenakan tercinta Aneirah Satirah Helmy
3. Bapak Prof. Dr. Ir. H. Sudirman Baco, M.Sc selaku Dekan Fakultas
Peternakan Universitas Hasanuddin.
4. Ibu Prof. Dr. drh. Hj. Ratmawati Malaka, M. Sc selaku Wakil Dekan I, Ibu Ir.
Hastang, M.Si selaku Wakil Dekan II, Bapak Prof. Dr. Ir. Jasmal A Syamsu,
M.Si selaku Wakil Dekan III Fakultas Peternakan Universitas Hasanuddin.

iii
5. Bapak Prof. Dr. Ir. Lellah Rahim, M.Sc., Bapak Prof. Dr. Sjamsuddin
Garantjang, M.Agr., dan Bapak Muhammad Yusuf, S.Pt., Ph.D., selaku
dosen pembahas yang telah banyak memberikan saran-saran dan masukan
untuk perbaikan skripsi ini.
6. Ibu Almarhumah Dr. Harfiah, S. Pt., MP., selaku Penasehat Akademik yang
selama hidupnya memberikan arahan dan bimbingannya.
7. Sahabat-sahabat masa kecil Ria Pratiwi Amd. Kep., Sri Julyarti Halid, S.E.,
Nurawaliyah, S.Pd., Laode Manarfah, Abidin Ma’ruf, Muh. Aras Prabowo,
S.E., dan A. Anni Ma’rifah, S.Pd., terima kasih atas segala dukungan dan
semangatnya.
8. Sahabat-sahabat St. Nur Ramadhani, S. Pt., May Rismi Anisa, Asrinti, Suarti,
Andi Nurfaini, Mustabsyirah Usman, Yuliana Padli, Yusri, Rajma Fastawa,
S.Pt., Trianta Tahir, dan Samsul Mardi terima kasih atas segala cinta,
pengorbanan, bantuan, pengertian, candatawa, serta kebersamaannya selama
ini.
9. Teman-teman Posko KKN Gelombang 87 Hildah Khurniyah, Nurmala Sari,
Rezky Amelia Madina, S.Sos., Avif Munandar, Ainul Anugrah, S.E., Muh.
Arman Kadir dan teman-teman sekecamatan Patimpeng.
10. Keluargan besar “SOLANDEVEN” khususnya kelas “NU3C”. Keluarga
Besar Himpunan Mahasiswa Produksi Ternak (HIMAPROTEK) terima kasih
atas kenangan yang telah terukir selama penulis bersama kalian.

iv
11. Kepada teman-teman Penelitian Genetika Molekuler, Kak Abduh, Mardha,
Fira, Umma, Inci, Nia, dan Awal terima kasih atas bantuan dan perhatiannya
selama penelitian dan penyelesaian skripsi.
12. Teman-teman asisten Ternak Potong yaitu St. Nur Ramadhani, S.Pt., Armi
Auliah Utami, S.Pt., Abdi Eriansyah, kakanda Ahmad David, Darussalam,
Andi Nurul Ainun, S.Pt., Erwin Jufri, Ayu Anggaraini, Suprapto, Appeyani,
Arman.
13. Laboratorium Terpadu Peternakan UNHAS, Kak Nurul Purnomo, Kak Tri
terima kasih atas bantuan dan perhatiannya selama penelitian dan
penyelesaian skripsi.
14. Semua pihak yang tidak dapat penulis sebut satu persatu, terima kasih banyak
atas segala bantuannya.
Penulis menyadari bahwa penyusunan skripsi ini masih terdapat kekurangan
dan kesalahan. Penulis mengharapkan kritikan dan saran yang sifatnya
membangun demi kesempurnaan skripsi ini.
Makassar, Desember 2015
Evy Harjuna Saad

v
ABSTRAK
EVY HARJUNA SAAD (I 111 11 256). Keragaman Gen Major
Histocompatibility Complex (MHC) DRB3.2 Locus PstI pada Populasi Kambing
Kacang. Dibimbing oleh SRI RACHMA APRILITA BUGIWATI dan
MUHAMMAD IHSAN DAGONG.
Major Histocompatibility Complex (MHC) merupakan sekumpulan gen penting yang
mengendalikan respon imun dan memegang peranan penting dalam sistem kekebalan
tubuh.Keragaman gen MHC-DRB3.2 dianggap bertanggung jawab dalam perbedaan
antara individu dalam respon kekebalan terhadap agen infeksius. Penelitian ini bertujuan
untuk mengetahui keragaman gen Major Histocompatibility Complex (MHC) DRB3.2
Locus PstI pada kambing Kacang. Materi utama dalam penelitian ini menggunakan
sampel darah kambing Kacang yang dikoleksi dari populasi Laboratorium Ternak Potong
Fakultas Peternakan Universitas Hasanuddin, Makassar (32 ekor), Kabupaten Maros (16
ekor), dan Kabupaten Jeneponto (30 ekor). Amplifikasi fragmen DNA menggunakan
mesin PCR. Keragaman gen MHC DRB3.2 dideteksi dengan memotong amplimer
dengan menggunakan enzim restriksi PstI. Hasil penelitian menunjukkan keragaman
genetik dengan ditemukannya dua alel yaitu alel P dan p. ditemukan tiga genotype yaitu
PP, Pp, pp. Nilai heterosigositas menunjukkan populasi kambing kacang di Laboratorim
Ternak Potong (0,5034) dan di kabupaten Maros (0,5141) memiliki keragaman genetik
yang tinggi (Ho>0,50). Nilai chi-square pada ketiga populasi menunjukkan populasi di
kabupaten Jeneponto berada pada keseimbangan Hardy-Weinberg dan populasi kambing
kacang yang ada di LaboratoriumTernak Potong dan di Kabupaten Maros tidak berada
pada keseimbangan Hardy-Weinberg. Hasil penelitian ini menyimpulkan bahwa gen
MHC yang diidentifikasi dari penelitian ini dapat dijadikan sebagai informasi genetik
(kandidat gen) dalam penelitian selanjutnya untuk mencari hubungan antara variasi alel
gen MHC dengan tingkat respon imun kambing kacang terhadap penyakit.
Kata kunci: MHC DRB3.2, sistem imun, kambing Kacang, polimorfisme, PstI.

vi
ABSTRACT
EVY HARJUNA SAAD (I 111 11 256). Polymorphism of Major
Histocompatibility Complex (MHC) Genes Locus DRB3.2 PstI in Kacang Goats
Population. Supervised by SRI RACHMA APRILITA BUGIWATI as Main
Supervisor and MUHAMMAD IHSAN DAGONG as Co-Supervisor.
Major Histocompatibility Complex (MHC) is a set of essential genes that control the
immune response and plays an important role in the immune system. The diversity of
MHC-DRB3.2 genes considered responsible for the differences among individuals in the
immune response against infectious agents. The aim of this study was to determine the
diversity of Major Histocompatibility Complex (MHC) genes locus DRB3.2 PstI on
Kacang goats population. The main material in this study using blood samples from
Kacang goats, which were collected from Animal Production Laboratory, Faculty of
Animal Husbandry Hasanuddin University, Makassar (32 head), Maros (16 head), and
Jeneponto regency (30 head). Target DNA fragments amplified using PCR machine.
Diversity of MHCDRB3.2 genes detected by cutting amplimer using PstI restriction
enzymes. The results showed the genetic diversity with two alleles were identified,
namely P and p. With three genotypes of the PP, Pp, pp. Heterozygosity values show the
goat population at the Animal Production Laboratory (0.5034) and in the Maros regency
(0.5141) has a high genetic diversity (Ho> 0.50). Chi-square value of the three
populations showed that the population in the Jeneponto were in Hardy-Weinberg
equilibrium and goat populations that exist in the Animal Production Laboratoryand
Maros were not in Hardy-Weinberg equilibrium. Results of this study concluded that the
MHC genes identified from this study can be used as genetic information (candidate
genes) in future studies to find the relationship between variations in the MHC gene
alleles with the level of immune response in Kacang goat against the disease.
Key words: MHC DRB3.2, the immune system, Kacang goat , polymorphism, PstI.

vii
DAFTAR ISI
DAFTAR ISI ..................................................................................................... vii
DAFTAR TABEL ........................................................................................... ix
DAFTAR GAMBAR ........................................................................................... x
PENDAHULUAN ........................................................................................... 1
TINJAUAN PUSTAKA
Gambaran Umum Kambing Kacang ....................................................... 4
Sistem Imun ........................................................................................... 4
Keragaman Genetik ................................................................................... 5
Marker Assisted Selection (MAS) ............................................................. 6
Major Histocompatibility Complex (MHC) .............................................. 8
DRB3.2 locus PstI ..................................................................................... 10
METODE PENELITIAN
Waktu dan Tempat ................................................................................... 12
Materi Penelitian ...................................................................................... 12
Tahapan Penelitian…………………………………………………….... 13
HASIL DAN PEMBAHASAN
Amplifikasi Gen MHC DRB3.2 Locust PstI............................................. 17
Frekuensi Genotip dan Alel ...................................................................... 19
Nilai Heterozigositas ................................................................................. 22
Keseimbangan Hardy-Weinberg ............................................................... 24
KESIMPULAN DAN SARAN
Kesimpulan ........................................................................................... 26

viii
Saran ........................................................................................... 26
DAFTAR PUSTAKA .......................................................................................... 27
LAMPIRAN ........................................................................................... 31
RIWAYAT HIDUP ........................................................................................... 33

ix
DAFTAR TABEL
No. Teks Halaman
1. Sequen primer beserta enzim restriksi endonuklease untuk PCR-
RFLP................................................................................................. 9
2. Frekuensi Genotipe Gen MHC DRB3.2 Locust PstI ………………. 20
3. Frekuensi Alel Gen MHC DRB3.2 Locust PstI…………………….. 21
4. Nilai Heterizigositas Harapan dan Heterozigositas Pengamatan…… 22
5. Uji Keseimbangan Hardy-Weinberg………………………………... 24

x
DAFTAR GAMBAR
No. Teks Halaman
1. Hasil Amplifikasi Gen MHC DRB3.2 Locus PstI………………….. 17
2. pengamatan keragaman gen MHC DRB3.2 Locus PstI ………….… 18
3. Letak sequens primer forward dan reverse MHC DRB3.2.……….... 19

1
PENDAHULUAN
Kambing Kacang merupakan salah satu kambing lokal Indonesia yang
tersebar luas di seluruh kawasan Indonesia dan banyak diternakkan masyarakat
dalam skala kecil. Kambing ini cocok sebagai penghasil daging dan kulit, bersifat
prolifik, tahan terhadap berbagai kondisi, mampu beradaptasi dengan baik di
berbagai lingkungan yang berbeda termasuk dalam kondisi pemeliharaan yang
sangat sederhana (Batubara, dkk, 2009). Kambing Kacang mampu bertahan
dalam kondisi apapun namun memiliki kerentanan terhadap beberapa penyakit
akibat cacing, parasit internal, dan caplak. Adanya informasi ketahanan terhadap
penyakit pada kambing Kacang dapat digunakan sebagai salah satu acuan untuk
seleksi menggunakan penanda molekuler. Seleksi merupakan cara untuk
memperbaiki mutu genetik kambing Kacang agar dapat dihasilkan bibit unggul
yang mempunyai ketahanan yang tinggi terhadap penyakit.
Imunogenetik merupakan konsep pendekatan genetik yang mengendalikan
perbedaan reaktivitas respon imun dan kerentanan tubuh terhadap suatu penyakit
(Judajana, 1999). Kendali genetik tersebut akan menentukan perbedaan
reaktivitas imun pada setiap individu dalam suatu populasi sehingga berpengaruh
terhadap ketahanan dan kerentanan individu terhadap penyakit (Angyalosi,
Neveb, et al, 2001). Salah satu lingkup imunogenetik tersebut adalah sistem
Major Histocompatibility Complex (MHC).
Major Histocompatibility Complex (MHC) merupakan antigen limfosit
yang terdapat pada sel berinti terutama pada sel limfosit. MHC pada kambing

2
disebut Caprine Lymphosite Antigen (CLA). Gen MHC adalah gen multigenik
karena beberapa gen terkait dengan MHC mengkode berbagai molekul gen MHC
yang berbeda. Gen MHC pada setiap populasi juga memiliki banyak alel sehingga
disebut gen polimorfik yang dapat menimbulkan perubahan-perubahan genetik
pada induknya.
Keragaman genetik dalam suatu populasi kambing Kacang akan
mengakibatkan perbedaan aktivitas imun pada setiap individu dalam suatu
populasi sehingga berpengaruh terhadap ketahanan dan kerentanan individu
terhadap penyakit. Sistem imun kambing Kacang diatur oleh gen MHC DRB3.2
Locus PstI. Polimorfisme CLA-DRB3.2 dianggap bertanggung jawab untuk
perbedaan antara individu dalam respon kekebalan terhadap agen infeksius.
Sharif, et al (1998) menyatakan bahwa asosiasi alel sapi dari gen MHC-DRB3
ekson 2 (BoLA (Bovine Lymphosite Antigent) DRB3.2) berhubungan dengan
ketahanan penyakit, respon imun dan sifat produksi. Keragaman gen MHC
DRB3.2 Locus PstI (CLA DRB3.2 Locust PstI) dapat dijadikan kandidat gen
sebagai penanda genetik untuk melihat hubungan tingkat ketahanan terhadap
penyakit akibat cacing, parasit internal, dan caplak. Namun sampai saat ini belum
ada informasi mengenai keragaman gen MHC DRB3.2 Locus PstI pada populasi
kambing Kacang. Berdasarkan hal tersebut maka perlu dilakukan penelitian untuk
mengidentifikasi keragaman gen Major Histompatibility Complex (MHC) pada
populasi kambing Kacang.
Penelitian ini bertujuan untuk mengetahui keragaman gen Major
Histocompatibility Complex (MHC) DRB3.2 Locus PstI pada populasi kambing

3
Kacang. Kegunaan penelitian ini yaitu menambah informasi mengenai
keragaman genetik pada sistem pertahanan tubuh pada kambing Kacang sehingga
dapat dijadikan dasar dalam proses seleksi agar dapat dihasilkan bibit unggul yang
mempunyai daya tahan terhadap penyakit akibat cacing, parasit internal, dan
caplak.

4
TINJAUAN PUSTAKA
Gambaran Umum Kambing Kacang
Kambing Kacang merupakan kambing asli Indonesia yang bersifat
prolifik, tahan terhadap berbagai kondisi, dan mampu beradaptasi dengan baik di
berbagai lingkungan yang berbeda termasuk dalam kondisi pemeliharaan yang
sangat sederhana. Pemanfaatan kambing lokal dengan potensi genetik yang baik,
namun belum dieksploitasi secara optimal dapat memberikan hasil yang baik
(Batubara,dkk., 2009).
Sistem Imun
Sistem imun adalah semua mekanisme yang digunakan tubuh untuk
mempertahankan keutuhan tubuh, sebagai perlindungan terhadap bahaya yang
dapat ditimbulkan berbagai bahan dalam lingkungan hidup (Anthony, et al, 2007).
Pada ternak sistem imun dibutuhkan untuk mempertahankan keutuhannya
terhadap bahaya yang dapat ditimbulkan dari lingkungan (Baratawidjaja, 2006).
Secara umum sistem imun terbagi dalam dua yaitu : alamiah dan adaptif
(spesifik). Sistem imun alamiah seperti air liur, selaput lendir, serta asam
lambung termasuk di dalamnya. Sedangkan pada sistem imun adaptif terdapat
sistem dan struktur fungsi yang lebih kompleks dan beragam. Sistem imun
adaptif terdiri dari sub sistem seluler yaitu keluarga sel limfosit T (T penolong dan
T sitotoksik) dan keluarga sel mononuklear (berinti tunggal). Sub sistem kedua
adalah sub sistem humoral, yang terdiri dari kelompok protein globulin terlarut
yaitu: imunoglobulin. Imunoglobulin dihasilkan oleh sel limfosit B melalui suatu
proses aktivasi khusus (Pandjassarame, 2009).

5
Cara kerja sistem imun pada kambing yaitu stimulasi antigenik
menginduksi respons imun yang dilakukan sistem seluler secara bersama-sama
diperankan oleh makrofag, limfosit B, dan limfosit T. Makrofag memproses
antigen dan menyerahkannya kepada limfosit. Limfosit B, yang berperan sebagai
mediator imunitas humoral, yang mengalami transformasi menjadi sel plasma dan
memproduksi antibodi. Limfosit T mengambil peran pada imunitas seluler dan
mengalami diferensiasi fungsi yang berbeda sebagai subpopulasi (Sharma, et al,
2005).
Keragaman Genetik
Keanekaragaman genetik (genetic diversity) adalah jumlah total variasi
genetik dalam keseluruhan spesies yang mendiami sebagian atau seluruh
permukaan bumi yang berbeda dari variabilitas genetik yang menjelaskan
kecenderungan kemampuan suatu karakter/sifat untuk bervariasi yang
dikendalikan secara genetik (Tisdell, 2003).
Keragaman genetik di antara populasi dari suatu spesies bisa sangat besar.
Demikian juga perbedaan genetik di antara individu dalam populasi alami sering
juga besar. Keragaman genetik dalam suatu individu bilamana ada dua alel untuk
gen yang sama namun memiliki perbedaan konfigurasi DNA yang menduduki
lokus yang sama pada suatu kromosom (Sufro,1994).
Polimorfisme pada suatu populasi digunakan sebagai salah satu indeks
keragaman genetik. Sifat polimorfik ini ditentukan dengan mengidentifikasi
jumlah alel pada suatu populasi. Dengan adanya identifikasi jumlah alel maka

6
akan dapat ditentukan frekuensi alel dan nilai heterozigositas suatu populasi
(Barendse, et al., 2008).
Keanekaragaman genetik memainkan peran yang sangat penting dalam
sintasan dan adaptabilitas suatu spesies karena ketika lingkungan suatu spesies
berubah, variasi gen yang kecil diperlukan agar spesies dapat bertahan hidup dan
beradaptasi. Spesies yang memiliki derajat keanekaragaman genetik yang tinggi
pada populasinya akan memiliki lebih banyak variasi alel yang dapat diseleksi.
Seleksi yang memiliki sangat sedikit variasi cenderung memiliki risiko lebih besar
(Anonim, 2008).
Marker Assisted Selection (MAS)
Marka gen adalah variasi sekuen DNA yang mencirikan terjadinya variasi
sifat fenotipe, baik yang secara langsung mempengaruhi sifat tersebut maupun
secara tidak langsung karena terjadi linkage (pautan) dengan sekuen DNA yang
mempengaruhi sifat fenotip. Ide dasar yang melatarbelakangi perlunya seleksi
berdasarkan marka gen adalah adanya kemungkinan gen-gen dengan pengaruh
signifikan yang menjadi target khusus dalam seleksi. Kegunaan utama marka gen
adalah untuk seleksi/pemuliaan hewan berdasarkan variasi pada aras DNA
terpilih. Dari peta semacam inilah muncul suatu pendekatan molekuler untuk
melakukan pemuliaan hewan guna memperoleh suatu individu unggul. Teknik ini
dikenal dengan pendekatan Marker Assisted Selection (MAS), yaitu suatu
pendekatan langsung untuk memperoleh hewan-hewan yang secara genetik
superior. MAS digunakan dalam seleksi berdasarkan pada marka yang
berhubungan dengan gen yang dikehendaki (indirect marker). Pendekatan MAS

7
dapat digunakan pada hewan, tumbuhan maupun manusia, dengan berbagai
macam tujuan (Sutarno, 2006).
MAS (Marker Assisted Selection) yaitu proses seleksi tidak langsung pada
sifat yang ingin diseleksi dan juga merupakan alat untuk menduga dan membantu
seleksi penotipe sifat yang menjadi target pemuliaan dengan menggunakan
penanda yang terkait dengan sifat tersebut (Elfianis, 2015).
Menurut Soller (1983) informasi tentang variasi genetik dapat dijadikan
dasar dalam seleksi hewan melalui teknik yang dikenal dengan Marker Assisted
Selection (MAS) atau seleksi berdasarkan penanda gen. Variasi genetik juga
dapat dijadikan dasar untuk konservasi jenis. Suatu jenis tertentu mungkin
dihasilkan dari suatu proses adaptasi terhadap keadaan lingkungan yang
mengarahkan pada terbentuknya kombinasi alel yang unik. Penggunaan Marker
Assisted Selection (MAS) didasarkan pada gagasan bahwa terdapat gen yang
memegang peranan utama dan menjadi sasaran atau target secara spesifik dalam
seleksi (Van der Warf, 2000).
Efisiensi dari MAS dalam peningkatan kualitas hewan produksi tergantung
pada beberapa faktor antara lain proporsi varian sifat tambahan yang disebabkan
oleh marka, dan ketepatan teknik seleksi. Namun demikian, Edwards dan Page
(1994) serta Lande dan Thompson (1990) menyatakan bahwa peningkatan sifat
genetik sampai 50% dapat dipastikan terjadi dengan teknik MAS. Peningkatan ini
terjadi karena lebih akuratnya teknik MAS dalam seleksi, dan pengurangan waktu
seleksi antar generasi karena gen dapat diidentifikasi sejak awal kelahiran atau
bahkan semasa masih dalam embrio. Pendekatan marka gen telah banyak

8
digunakan dengan baik untuk sifat-sifat seperti resistensi terhadap penyakit,
fertilitas dan reproduksi, dan produksi susu (Sutarno, 2006).
Major Histocompatibility Complex (MHC)
Major Histocompatibility Complex (MHC) adalah sekumpulan gen yang
ditemukan pada semua jenis vertebrata. Protein MHC yang disandikan berperan
dalam mengikat dan mempresentasikan antigen peptida ke sel T. Molekul
permukaan sel yang bertanggung jawab terhadap rejeksi transplan dinamakan
molekul histokompatibilitas, dan gen yang mengkodenya disebut gen
histokompatibilitas. Nama ini kemudian disebut dengan histokompatibilitas mayor
karena ternyata MHC bukan satu-satunya penentu rejeksi karena terdapat pula
molekul lain yaitu molekul histokompitibilitas minor yang walaupun lebih lemah
juga ikut menentukan rejeksi. Pada saat ini telah diketahui bahwa molekul MHC
merupakan titik sentral inisiasi respons imun (Anonim, 2012).
Major Histocompatibility Complex (MHC) merupakan kelompok lokus
yang terdiri atas kumpulan gen penting (major) yang mengendalikan respon imun.
Respon imun terutama disebabkan oleh adanya aksi limfosit yang dihasilkan
dalam sel sumsum tulang. Aksi limfosit dalam sistem kekebalan dipacu oleh
adanya antigen. Peranan MHC dalam sistem kekebalan seluler diawali dari
masuknya antigen ke dalam tubuh melalui proses up take oleh makrofag yang
kemudian memicu limfosit T untuk mematikan sel yang terinfeksi. Sementara itu,
dalam sistem kekebalan humoral, MHC berperan dalam membantu pembentukan
antibodi oleh limfosit B (Tizard, 2000).

9
MHC atau antigen histokompatibilitas utama adalah antigen yang terdapat
pada sel limfosit yang bersifat lebih imunogenik dibandingkan antigen lainnya.
Antigen ini ditemukan pertama pada leukosit darah, nomenklatur MHC pada
setiap hewan berbeda. Diawali dengan singkatan yang merujuk pada jenis hewan
dan dilanjutkan dengan huruf L (Limfosit) dan A (Antigen) (Tizard, 2000). MHC
yang terdapat pada kambing yaitu Caprine Limfosit Antigent (CLA) yang telah
terbukti sama dengan domba dan sapi. (Takada, et al., 1998).
Menurut Kuncorojakti (2011), molekul gen MHC dibedakan menjadi tiga
kelas, yaitu MHC kelas I, II, dan III. MHC kelas I berisi beberapa gen yang
berperan dalam respon imun selular.MHC kelas II merupakan gabungan gen yang
berperan penting dalam respon imun selular dan humoral. Kelas II gen MHC
secara ekstensif dicirikan dengan domba dan sapi (Takada, et al., 1998). MHC
kelas II menciri dengan jumlah alel yang besar pada setiap lokus dan terdapat
perbedaan jumlah asam amino yang besar pada setiap alelnya. MHC kelas III
mengandung beberapa gen yang memiliki fungsi luas, berperan dalam
pembentukan komponen protein dan sistem komplemen, hanya beberapa
diantaranya yang terlibat dalam respon imun (Guillemot, et al, 1988). Daerah
MHC sangat luas, sekitar 3500 kb di lengan pendek kromosom 6, meliputi regio
yang mengkode MHC kelas I, II, III, dan protein lain, serta gen lain yang belum
dikenal, yang mempunyai peran penting pada fungsi sistem imun (Anonim, 2012).
Gen MHC berhubungan dengan gen imunoglobulin dan gen reseptor sel T
(TCR = T-cell receptors) yang tergabung dalam keluarga supergen
imunoglobulin, tetapi pada perkembangannya tidak mengalami penataan kembali

10
gen seperti halnya gen imunoglobulin dan TCR (Anonim, 2012). Menurut Caron,
Abplanalpand, and Taylor (1997), MHC berhubungan dengan kepekaan terhadap
infeksi parasit. Keragaman ini berhubungan dengan keragaman reseptor pada
limfosit T, yang berkontribusi pada perbedaan respon kekebalan pada individu
(Sommer, 2005).
DRB3.2 locusPst I
Major histocompatibility Complex (MHC) adalah gen yang wilayahnya
paling luas atau gen genom keluarga ditemukan di sebagian besar vertebrata yang
mengkodekan molekul MHC. Molekul MHC memegang peranan penting dalam
sistem kekebalan tubuh dan autoimunitas. Ada dua kelas umum molekul MHC:
Kelas Idan Kelas II. Molekul kelas I MHC ditemukan pada hampir semua sel dan
protein ini untuk mematikan sel T. kelas IIMolekul MHC ditemukan pada sel-sel
kekebalan tertentu (Traherne,et al., 2006).
MHC-DRB berperan dalam sistem imunitas yang penting dalam
pengenalan pathogen serta membawa pathogen tersebut (Petlane, et al., 2012).
Sistem Caprine Limfosit Antigen (CLA) adalah kompleks histocompatibility
utama kambing, telah terbukti mirip dengan sapi yang dinyatakan memiliki dua
kelas II antigen, DQ dan DR (Takada, et al., 1998). Molekul MHC DR subtipe
telah diidentifikasi sebagai salah satu kelas prinsip II protein yang ditemukan pada
sel kambing (Schwaiger, et al., 1993).
Sejauh ini, setidaknya ada dua lokus DRB yang telah ditandai (Schwaiger
et al., 1993; Amills et al., 1995). CLA-DRB3 ekson 2 (CLA-DRB3.2)
mengkodekan β1 domain dari DR molekul dan menampilkan tingkat yang sangat

11
tinggi dari polimorfisme dengan lebih dari 25 urutan yang berbeda. Polimorfisme
CLA-DRB3.2 dianggap bertanggung jawab untuk perbedaan antara individu
dalam respon kekebalan terhadap agen infeksius. Penelitian sebelumnya
menyatakan, asosiasi alel dari sapi MHC-DRB3 ekson 2 (BoLA DRB3.2)
berhubungan dengan ketahanan penyakit, respon imun, dan produksi sifat (Sharif,
et al.,1998). Namun, sedikit yang diketahui tentang asosiasi antara CLA-DRB3.2
alel dan pertahanan penyakit (misalnya Cowdriosis dan infeksi nematoda) dan
ciri-ciri produksi (misalnya daging dan susu) pada kambing.

12
METODOLOGI PENELITIAN
Waktu dan Tempat
Penelitian ini dilaksanakan pada bulan Juni-Juli 2015 bertempat di
Kandang Kambing Laboratorium Ternak Potong dan Laboratorium Bioteknologi
Terpadu, Fakultas Peternakan Universitas Hasanuddin, Makassar.
Materi Penelitian
Bahan utama dari penelitian ini adalah 32 sampel darah dari 32 ekor induk
kambing Kacang umur 2 tahun di Unit Kandang Kambing Laboratorium Ternak
Potong Fakultas Peternakan Universitas Hasanuddin dan 46 sampel DNA induk
kambing Kacang umur 2 tahun dari koleksi DNA Laboratorium Bioteknologi
Terpadu Fakultas Peternakan Universitas Hasanuddin. Bahan pendukung antara
lain: enzim restriksi PstI, bahan ekstraksi DNA (lysis buffer, proteinaseK, wash
buffer I,wash buffer II, elution buffer, ethanol absolute 96%), bahan PCR (dNTP
mix, MgCl2,enzim Taq DNA polymerase), bahan elektroforesis (agarose,Na2
EDTA, ethidium bromide, marker DNA, DNA loading dye), tissue, dan primer
gen MHC DRB3.2Locust Pst I.
Tabel 1. Sequen primer beserta enzim restriksi endonuklease untuk PCR-
RFLP
Primer Sequen DNA Enzim
Restriksi
Panjang
PCR Sumber
MHC F : 5’-TATCCCGTCTCTGCAGCACATTTC-3’
PstI 285 bp Ahmed,
2006 R : 5’-TCGCCGCTGCACACTGAAACTCTC-3’
Alat yang digunakan yaitu : Kit DNA ekstraksi (Thermo Scientific),
venoject, tabung vacutainer, mesin PCR, centrifuge, alat pendingin, tabung

13
eppendorf besar dan kecil, gel dokumentasi (syngene G:BOX), mikropipet, tip, rak
tabung, elektroforesis, autoclave, timbangan, dan sarung tangan.
Tahapan Penelitian
Koleksi Sampel Darah
Sampel darah diperoleh dari induk kambing Kacang berumur 2 tahun di
Unit Kandang Kambing Laboratorium Ternak Potong, Fakultas Peternakan
Universitas Hasanuddin. Pengambilan darah melalui vena jugularis sebanyak 2
ml kemudian ditampung pada tabung vacutainer yang telah berisi antikoagulan
EDTA untuk mencegah penggumpalan darah. Sampel darah yang dikumpulkan
sebanyak 32 tabung.
Ekstraksi DNA
DNA diisolasi dan dimurnikan dengan menggunakan Kit DNA ekstraksi
(Genjet Genomic DNA Extraction Thermo Scientific) dengan mengikuti protocol
ekstraksi yang disediakan. Sebanyak 200 μl sampel darah dilisis dengan
menambah400 μl larutan buffer (lysis buffer), 20 μl proitenaseK (10 mg/ml),
kemudiandicampur dan diinkubasi pada suhu 56ºC selama 60 menit pada
waterbath shaker.Setelah inkubasi larutan, ditambahkan 200 μl ethanol absolute
96% dandisentrifugasi 6.000 x g selama 1 menit.
Pemurnian DNA dilakukan menggunakan spin column dengan
penambahan500 μl larutan pencuci wash buffer I yang kemudian dilanjutkan
dengan sentrifugasi pada 8.000 x g selama 1 menit. Setelah supernatanya
dibuang, DNA kemudian dicuci lagi dengan 500 μl wash buffer II dan
disentrifugasi pada 12.000 x g selama 3 menit. Setelah supernatanya dibuang,

14
DNA kemudian dilarutkan dalam 200 μl elution buffer dan disentrifugasi pada
8.000 x g untuk selanjutnya DNA hasil ekstraksi ditampung dan disimpan pada
suhu -20 ºC.
Teknik PCR-RFLP
Komposisi reaksi PCR dikondisikan pada volume reaksi 25 μl yang terdiri
atas 100 mg DNA, 0.25 mM primer MHC DRB3.2 PstI, 150 μM dNTP, 2.5 mM
Mg2+,0.5μl Taq DNA polymerase dan 1x buffer. Kondisi mesin PCR dimulai
dengan denaturasi awal pada suhu 94ºC x 2 menit, diikuti dengan 35 siklus
berikutnya masing-masing denaturasi 94ºC x 45 detik dengan suhu annealing
yaitu : 64ºC x 60 detik yang dilanjutkan dengan ekstensi : 72ºC x 60 detik yang
kemudian diakhiri dengan satu siklus ekstensi akhir pada suhu 72ºC selama 5
menit dengan menggunakan mesin PCR (SensoQuest, Germany). Produk PCR
kemudian dielektroforesis pada gel agarose 1.5 % dengan buffer 1x TBE (89 mM
Tris, 89 mM asam borat, 2 mM Na2EDTA) yang mengandung 100 ng/ml ethidium
bromide. Kemudian divisualisasi pada UV transiluminator (gel documentation
system ;syngeneG:BOX)).
Produk PCR yang diperoleh dari masing-masing gen target kemudian
dianalisis menggunakan RFLP melalui pemotongan menggunakan enzim restriksi
PstI (CTGGA G) yang memiliki situs pemotongan pada gen MHC. Sebanyak
5l DNA produk PCR ditambahkan 0,3 l enzim restriksi (5U) ; 0,7 l buffer
enzim dan 1lmilique water sampai volume 7l, selanjutnya dilakukan inkubasi
selama 18 jampada suhu 37ºC. Analisis produk RFLP dilakukan dengan

15
elektroforesis pada gel polyacrylamide dan pewarnaan dengan perak mengikuti
metode Tegelstrom (1992).
Analisis Data
Keragaman genotipe tiap-tiap individu dapat ditentukan dari pita-pita
DNA gen yang ditemukan. Masing-masing sampel dibandingkan berdasarkan
ukuran (marker) yang sama dan dihitung frekuensi alelnya. Frekuensi alel
dihitung menggunakan rumus Nei dan Kumar (2000) :
xi =
2nii + ∑ nij
j≠i
2n
Keterangan :
Xi = frekuensi alel ke -i
nii = jumlah sampel yang bergenotip ii ( homozigot)
nij = jumlah sampel yang bergenotip ij ( heterozigot)
n = jumlah sampel
Nilai heterozigositas pengamatan (Ho) dan heterozigositas harapan
(He)berdasarkan rumus heterozigositas Nei dan Kumar (2000) dihitung dengan
menggunakan software PopGene 32 versi 1.31 (Yeh, Yang, and Boyle, 1999).
Keterangan:
Ho = heterozigositas pengamatan di antara populasi,
He = heterozigositas harapan di antara populasi,

16
𝑤𝑘 =ukuran relatif populasi,
Xkij (i≠j) = frekuensi AiAj pada populasi ke-k.
Test keseimbangan Hardy-Weinberg (HWE) dengan uji chi-square (Hartl,
1988) sebagai berikut :
Keterangan :
χ² = chi-square ,
Obs = jumlah genotipe ke-ii atau ke-ij hasil pengamatan,
Exp = jumlah genotipe ke-ii atau ke-ij yang diharapkan.

17
300 bp
100 bp
285 bp 300 bp
HASIL DAN PEMBAHASAN
Amplifikasi Gen MHC DRB3.2 Locust PstI
Gen MHC DRB3.2 Locust PstI berhasil diamplifikasi dengan
menggunakan mesin thermocycler SensoQuest Germany, dengan Suhu annealing
64ºC terhadap 72 sampel DNA kambing Kacang. Hasil amplifikasi ruas gen
dapat divisualisasikan pada gel agarose 1,5% yang dapat dilihat pada Gambar 1.
Gambar 1.Hasil Amplifikasi Gen MHC DRB3.2 Locus PstI yang divisualisasi
pada gel Agarose 1,5 %
M : Marker (100 bp); 1-8 : Amplikon gen MHC DRB3.2 Locus PstI.
Gambar 1. menunjukkan panjang fragmen yang merupakan hasil
amplifikasi dari PCR terhadap gen MHC DRB3.2 Locus PstI yaitu sepanjang 285
bp. Hal ini sesuai dengan hasil penelitian yang dilakukan oleh Ahmed (2006)
bahwa panjang produk PCR yang dihasilkan adalah 285 bp. Petlanea et al (2012)
juga mendapatkan hasil PCR pada CLA dengan panjang produk 285 bp yang
dilakukan pada kambing Saneen, PE (Peranakan Etawah), dan PESA (PE dan
Saneen) dengan menggunakan PstI sebagai enzim pemotong.
Penentuan genotipe gen MHC DRB3.2 pada kambing Kacang
menggunakan metode PCR-RFLP yang dideteksi berdasarkan banyaknya pita
200 bp
M 1 2 3 4 5 6 7 8
18
226 bp
M 1 2 3 4 5 6 7 8
yang muncul dengan PstI sebagai enzim restriksi. Hasil pendeteksian keragaman
gen MHC DRB 3.2 Locus PstI dapat dilihat pada Gambar 2.
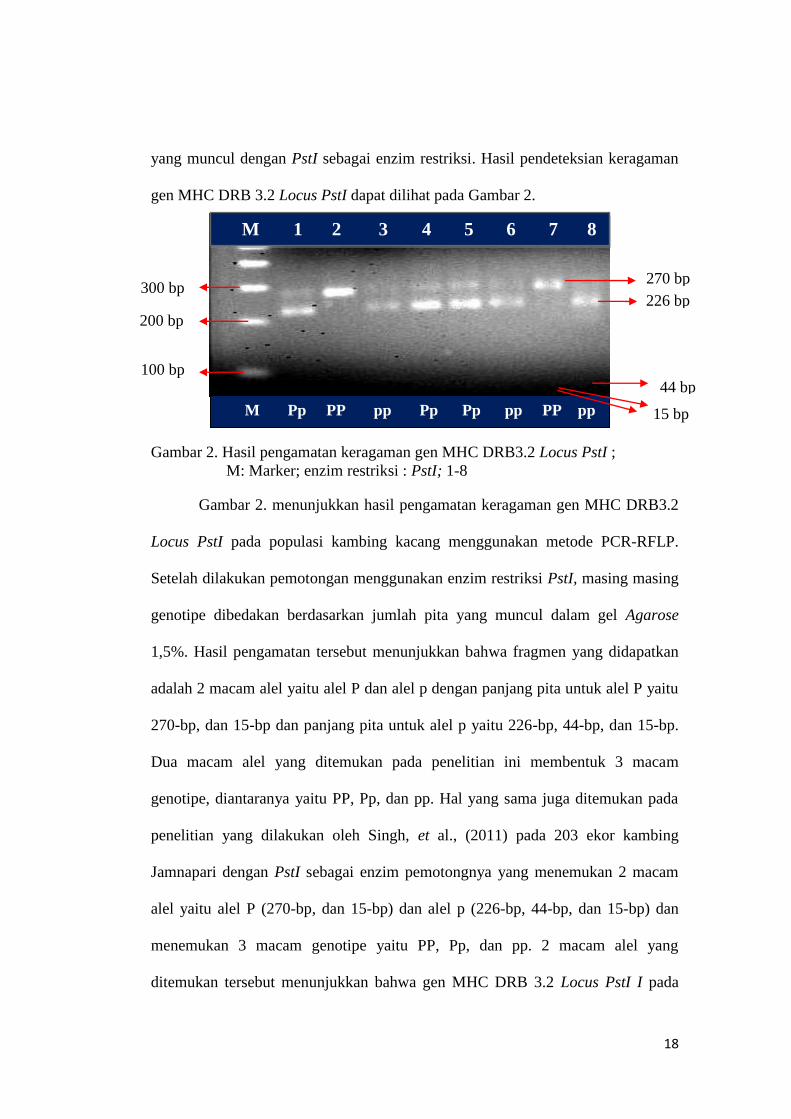
Gambar 2. Hasil pengamatan keragaman gen MHC DRB3.2 Locus PstI ;
M: Marker; enzim restriksi : PstI; 1-8
Gambar 2. menunjukkan hasil pengamatan keragaman gen MHC DRB3.2
Locus PstI pada populasi kambing kacang menggunakan metode PCR-RFLP.
Setelah dilakukan pemotongan menggunakan enzim restriksi PstI, masing masing
genotipe dibedakan berdasarkan jumlah pita yang muncul dalam gel Agarose
1,5%. Hasil pengamatan tersebut menunjukkan bahwa fragmen yang didapatkan
adalah 2 macam alel yaitu alel P dan alel p dengan panjang pita untuk alel P yaitu
270-bp, dan 15-bp dan panjang pita untuk alel p yaitu 226-bp, 44-bp, dan 15-bp.
Dua macam alel yang ditemukan pada penelitian ini membentuk 3 macam
genotipe, diantaranya yaitu PP, Pp, dan pp. Hal yang sama juga ditemukan pada
penelitian yang dilakukan oleh Singh, et al., (2011) pada 203 ekor kambing
Jamnapari dengan PstI sebagai enzim pemotongnya yang menemukan 2 macam
alel yaitu alel P (270-bp, dan 15-bp) dan alel p (226-bp, 44-bp, dan 15-bp) dan
menemukan 3 macam genotipe yaitu PP, Pp, dan pp. 2 macam alel yang
ditemukan tersebut menunjukkan bahwa gen MHC DRB 3.2 Locus PstI I pada
M Pp PP pp Pp Pp pp PP pp
Pp
270 bp
200 bp
100 bp
300 bp
44 bp
15 bp

19
reverse cut size PstI
Forward
populasi kambing Kacang yang diamati sangat beragam. Hal ini sesuai dengan
pendapat Sufro (1994) yang menyatakan bahwa keragaman genetik dalam suatu
individu dapat dilihat bilamana ada dua alel untuk gen yang sama namun memiliki
perbedaan konfigurasi DNA yang menduduki lokus yang sama pada suatu
kromosom.
Pendeteksian keragaman dan penempelan primer gen MHC DRB3.2
dengan metode PCR-RFLP dan PstI sebagai enzim pemotong (CTGGA|G) dapat
dilihat pada Gambar 3.
Gambar 3. Letak sequens primer forward dan reverse MHC DRB3.2.
Gambar 3. memperlihatkan letak/runutan sequens DNA gen MHC
DRB3.2 Locus PstI dari primer yang dipakai menggunakan enzim
pemotong/restriksi PstI dengan situs pemotongan (CTGGA|G) dimana enzim PstI
memotong dan menempel pada rantai ke 210.
Frekuensi Genotipe dan Alel
Hasil analisis frekuensi genotipe pada fragmen gen MHC DRB3.2 Locus
PstI kambing Kacang dapat dilihat pada Tabel 2.

20
Tabel 2. Frekuensi Genotipe Gen MHC DRB3.2 Locus PstI
Tabel 2 menunjukkan frekuensi genotipe fragmen gen MHC pada populasi
kambing Kacang yang ada di Kandang Ternak Potong dan dari Kabupaten
Jeneponto memiliki 3 macam genotipe yaitu PP, Pp, dan pp. Sedangkan pada
populasi kambing Kacang dari Kabupaten Maros hanya memiliki 2 macam
genotipe yaitu Pp, dan pp. Frekuensi genotipe PP di Kandang Ternak Potong
(43,37) lebih tinggi dari Kabupaten Maros (0,00) dan Jeneponto (16,66).
Sedangkan frekuensi genotipe Pp di Kabupaten Maros (93,75) dan Jeneponto
(43,33) lebih tinggi dari Kandang Ternak Potong (25,00). Hal ini terjadi karena
proses perkawinan pada Kandang Ternak Potong hanya menggunakan 1 ekor
pejantan untuk beberapa betina dan diduga pejantan yang digunakan hanya
membawa genotipe PP. Sedangkan pada Kabupaten Maros dan Jeneponto
menggunakan beberapa pejantan untuk dikawinkan secara acak.
Sampel DNA kambing Kacang dari kabupaten Maros tidak ditemukan
genotipe PP. Hasil tersebut dapat terjadi karena jumlah sampel yang diteliti
jumlahnya sedikit. Hal yang sama juga ditemukan terhadap kambing perah PESA
(Peranakan Etawah-Saneen) pada penelitian Petlanea, et al., (2012) yang tidak
menemukan genotipe PP (0,00) dengan jumlah sampel yang teliti yaitu hanya 9
sampel DNA.
Populasi Jumlah
(ekor)
Frekuensi Genotipe
PP (ekor) pp (ekor) Pp (ekor)
Kambing Kacang Di Kandang Ternak
Potong 32 43,37 (14) 31,25 (10) 25,00 (8)
Kambing Kacang Di Kab. Maros 16 0,00 (0) 6,25 (1) 93,75 (15)
Kambing Kacang Di Kab. Jeneponto 30 16,66 (5) 40,00 (12) 43,33 (13)

21
Frekuensi alel adalah proporsi ataupun perbandingan keseluruhan kopi gen
yang terdiri dari suatu varian gen tertentu (alel). Frekuensi alel dihitung
menggunakan rumus Nei dan Kumar (2000) dapat dilihat pada Tabel 3.
Tabel 3. Frekuensi Alel Gen MHC DRB3.2 Locus PstI
Berdasarkan Tabel 3. Frekuensi alel p di Kabupaten Maros (0,531) dan
Jeneponto (0,616) lebih tinggi dari Kandang Ternak Potong (0,451). Viabilitas
alel p lebih tinggi karena adanya sistem perkawinan acak pada populasi kambing
Kacang di Kabupaten Maros dan Jeneponto dibandingkan populasi kambing
Kacang di Kandang Ternak Potong. Hal yang sama juga ditemukan pada
penelitian Sing, et al., (2012) yang menemukan nilai total frekuansi alel p (0,835)
lebih tinggi dibandingkan dengan nilai total frekuensi alel P (0,165). Begitu juga
pada penelitian Petlanea, dkk., (2012) menemukan frekuensi alel p lebih tinggi
dibandingkan alel P.
Nilai frekuensi alel pada Tabel 3. menunjukkan bahwa populasi yang
diamati bersifat polimorfik atau beragam karena salah satu alelnya kurang dari
0,99. Menurut Nei dan Kumar, (2000) keragaman dapat ditunjukkan dengan
adanya dua alel atau lebih dalam suatu populasi dan frekuensi alel sama dengan
atau berada di bawah 0,99. Ditambahkan pula oleh Barandse, et al (2008)
polimorfisme pada suatu populasi digunakan sebagai salah satu indeks keragaman
Lokasi Jumlah
(ekor)
Frekuensi Alel
P (%) p (%)
Kambing Kacang Di Kandang Ternak Potong 32 0,549 0,451
Kambing Kacang Di Kab. Maros 16 0,469 0,531 Kambing Kacang Di Kab. Jeneponto 30 0,384 0,616

22
genetik. Sifat polimorfik ini ditentukan dengan mengidentifikasi jumlah alel
(Gambar 2.) pada suatu populasi.
Penentuan frekuensi alel pada populasi kambing Kacang menunjukkan
bahwa gen MHC DRB3.2 bersifat polimorfik (beragam). Polimorfisme CLA-
DRB3.2 dianggap bertanggung jawab untuk perbedaan antara individu dalam
respon kekebalan terhadap agen infeksius. Beberapa penelitian telah dilakukan
untuk mengetahui hubungan alel-alel CLA DRB3.2 Locus PstI dengan sifat
produksi dan imunitas. Alel-alel CLA DRB3 exon 2 berhubungan erat dengan
karakter imunitas (Ammils, et al., 1995) dan mempunyai hubungan resistensi
terhadap penyakit (Ahmed and Othman, 2006).
Nilai Heterozigositas
Nilai heterozigositas pengamatan (Hₒ) dan heterozigositas harapan (He)
berdasarkan rumus heterozigositas Nei dan Kumar (2000) dihitung menggunakan
software Pop Gene 32 versi 1.31 (Yeah, et al., 1999) dapat dilihat pada Tabel 4.
Tabel 4. Nilai Heterozigositas Harapan dan Heterozigositas Pengamatan
Berdasarkan Tabel 4 diperoleh nilai heterozigositas pengamatan (Ho)
populasi kambing Kacang di Kandang Ternak Potong lebih tinggi dibanding
heterozigositas harapan (He). Hal ini menunjukkan bahwa populasi kambing
Kacang di Kandang Ternak Potong memiliki nilai Ho yang cukup tinggi karena
nilainya berada diatas 0,50. Nilai Ho yang tinggi menunjukkan keragaman yang
Lokasi Jumlah
(ekor)
Nilai Heterozigositas
Ho He
Kambing Kacang Di Kandang Ternak Potong 32 0,5034 0,2581
Kambing Kacang Di Kab. Maros 16 0,5141 0,9375 Kambing Kacang Di Kab. Jeneponto 30 0,4808 0,4333

23
tinggi pada suatu populasi. Hal ini sesuai dengan pendapat Javanmard, et al.,
(2005) yang menyatakan bahwa nilai heterozigositas harapan yang tinggi
(Ho>0,50) menunjukkan tingginya keragaman gen pada suatu polulasi. Populasi
kambing Kacang di Kabupaten Maros nilai He lebih tinggi dari nilai Ho karena
memiliki jumlah populasi yang rendah namun memiliki keragaman yang tinggi .
Ukuran populasi kecil menyebabkan tingginya penyimpangan keragaman genetik
yang berakibat pada menurunnya jumlah individu dalam suatu populasi, serta
perkawinan kerabat (inbreeding) (Angeloni, et al., 2014). Sedangkan populasi
kambing Kacang di Kabupaten Jeneponto memiliki nilai Ho dan He yang hampir
sama. Hal ini menunjukkan bahwa populasi kambing Kacang di Kabupaten
Jeneponto memiliki keragaman genetik yang rendah karena nilai heterozigositas
hampir sama. Nilai heterozigositas merupakan cara yang paling akurat untuk
mengukur keragaman suatu populasi (Nei, 1987). Nilai heterozigositas dapat
dipengaruhi oleh jumlah sampel, jumlah alel dan frekuensi alel.
Populasi kambing Kacang di Kandang Ternak Potong dan populasi
kambing di Kabupaten Maros memiliki keragaman yang tinggi. Sedangkan
populasi kambing di Kabupaten Jeneponto memiliki keragaman yang rendah.
Tingginya keragaman gen MHC DRB3.2 Locus PstI di Kandang Ternak Potong
dan di kabupaten Maros dapat dijadikan kandidat gen sebagai penanda genetik
untuk melihat hubungan tingkat ketahanan penyakit pada populasi kambing
Kacang. Sharif et al., (1998) menyatakan bahwa keragaman tinggi dari gen MHC
DRB3.2 berhubungan dengan ketahanan penyakit, respon imun, dan sifat
produksi.

24
Keseimbangan Hardy-Weinberg
Pengujian keseimbangan Hardy-Weinberg pada populasi kambing Kacang
dilakukan dengan menggunakan uji chi-square untuk mengetahui apakah data
yang diperoleh menyimpang atau tidak dari yang diharapkan. Hasil pengujian
Keseimbangan Hardy-Weinberg pada populasi kambing Kacang dapat dilihat
pada Tabel 5.
Tabel 5. Uji Keseimbangan Hardy-Weinberg
Berdasarkan Tabel 5. nilai chi-square pada populasi kambing Kacang di
Kabupaten Jeneponto (0,30) berada dalam keseimbangan Hardy-Weinberg
(P<0,50). Hal ini terjadi karena nilai Chi-Square lebih rendah dari nilai F tabel
(0,45). Sedangkan populasi kambing Kacang yang terdapat di Kabupaten Maros
dan di Kandang Ternak Potong diperoleh nilai Chi-Square yang tidak sesuai
dengan hukum Hardy-Weinberg. Hal ini mengindikasikan bahwa telah terjadi
proses seleksi yaitu pemilihan calon pejantan dan calon induk pada kambing
Kacang di Kandang Ternak Potong dan Kabupaten Maros. Menurut
Hardjosubroto (1998), Suatu populasi dikatakan dalam keseimbangan Hardy-
Weinberg jika frekuensi genotipe dan frekuensi alel selalu konstan dari generasi
ke generasi. Pada penelitian petlanea et al., (2012) juga ditemukan hasil yang
tidak sesuai dengan hukum Hardy-Weinberg dengan perbanding jumlah populasi
kambing yang hampir sama.
Populasi N
(ekor) X2 F tabel
Derajat
Bebas A
Kambing Kacang Di Kandang Ternak
Potong 32 7,61 3,84 1 0,05 Kambing Kacang Di Kab. Maros 16 11,58 10, 827 1 0,001 Kambing Kacang Di Kab. Jeneponto 30 0,30 0,45 1 0,50

25
Hardy-Weinberg menyatakan bahwa frekuensi alel dan frekuensi genotipe
dalam suatu populasi akan tetap konstan, yakni berada dalam keseimbangan dari
satu generasi ke generasi lainnya kecuali apabila terdapat pengaruh-pengaruh
tertentu yang mengganggu keseimbangan tersebut. Pengaruh-pengaruh tersebut
meliputi perkawinan tak acak, mutasi, seleksi, dan ukuran populasi terbatas.
Dalam suatu populasi satu atau lebih pengaruh ini akan selalu ada (Burns, 1980).

26
KESIMPULAN DAN SARAN
Kesimpulan
Gen MHC DRB3.2 Locus PstI pada populasi kambing Kacang bersifat
polimorfik (beragam). Populasi kambing Kacang di Kandang Ternak Potong dan
di kabupaten Maros memiliki keragaman tinggi yang menandakan populasi
kambing Kacang di Kandang Ternak Potong dan di Kabupaten Maros dapat
dijadikan kandidat gen sebagai penanda genetik untuk melihat hubungan tingkat
ketahanan penyakit pada kambing Kacang.
Saran
Perlu dilakukan penelitian lebih lanjut untuk mengetahui bagaimana
hubungan sifat polimorfik gen MHC DRB3.2 Locus PstI dengan kemampuan
respontabilitas kambing Kacang terhadap penyakit akibat cacing, parasit internal,
dan caplak.

27
DAFTAR PUSTAKA
Ahmed, S. and E. Othman. 2006. A PCR-RFLP method for the analysis of
Egyptian goat MHC class II DRB gene. Department of Cell Biology,
National Research Center. Biotechnology. 5(1): 58-6.
Amills, M., O. Francino and A. Sanchez. 1995. Nested PCRallows the
characterization of TaqI and PstI RFLPs in thesecond exon of the caprine
MHC class II DRB gene. Vet.Immunol. Immunopathol. 48:313-321.
Angeloni, F., P. Vergeer, C. A. M. Wagemaker, and N. J. Ouborg. 2014. Within
and between population variation in inbreeding depression in the locally
threatened perennial Scabiosa columbaria. Conservion Genetic. 15: 331-
342.
Angyalosi G, R. Neveb, I. Wolowczuk, A. Delanoye, J. Herno, andC. Auriault.
2001. HLA class IIpolymorphism influences onset and severityof
pathology in Schistosoma mansoniinfectedtransgenic mice. Infect Immun
69:58-74.
Anonim. 2008. Genetic Diversity.National Biological Information
Infrastructure.NBII.www.nbii.gov. Diakses pada 23 Maret 2015.
Anonim. 2012. Imunologi Dasar : Kompleks Histokompatibilitas
Mayor.http://allergycliniconline.com/.Diakses pada 26 Maret 2015.
Anthony, L., DeFranco, M. Richard, Locksley, and R. Miranda. 2007. Immunity:
The Immune Response in Infectious and Inflammatory Disease. Oxford
University Press.
Baratawidjaja, K.G. 2006. Imunologi Dasar. 7thed. Balai Penerbit FKUI. Jakarta.
Barendse, W., B.E. Harrison, R, J. Bunchand, and M.B. Thomas. 2008. Variation
at the calpain 3 gene is associated with meat tenderness in Zebu and
composite breeds of cattle. BMC Genetic. 9:41-49.
Batubara, A, B. Tiesnamurti, F.A. Pamungkas, M. Doloksaribu dan E.
Sihite.2009.Petunjuk Teknis Potensi Beberapa Plasma Nutfah Kambing
Lokal Indonesia.Pusat Penelitian dan Pengembangan Peternakan.Bogor.
Burns, G.W. 1980.The Science of Genetics: Chapter 14. Macmillan Publishing
Co. New York.
Caron, L.A., H. Abplanalpand R.L. Taylor.1997. Resistance, susceptibility, and
immunity to Eimeria Tenella in Major Histocompatibility Complex
congenic lines.Poultry Science.76(1): 677−682.

28
Edwards, M. D.,and N. J Page. 1994. Evaluation of marker assisted selection
through computer simulation. Theoretical and Applied Genetics 88:376-
382.
Elfianis, R. 2015. Mark Assisted Selected (MAS). http://ritaelfianis.com/mas-
marker-assisted-selection/ diakses pada 26 Maret 2015.
Guillemot, F., N. Fréchin, A. Billault, A.M. Chaussé, R. Zoorob, C. Auffray, and
J.Embo. 1988. Isolation of chicken Major Histocompatibility Complex
class II (B-L) beta chain sequences: comparison with mammalian beta
chains and expression in lymphoid organs.7(4):103-109.
Hardjosubroto W, 1998. Pengantar Genetika Hewan. Fakultas Peternakan
Universitas Gadjah Mada, Yogyakarta.
Hartl, D.L. 1988. Principle of Population Genetic.Sinauer Associates, Inc.
Publisher.Sunderland.
Javanmard A, Asadzadeh N, Banabazi MH, and Tavakolian J. 2005. The Allele
and genotype frequencies of bovine pituitary specific transcription factor
and leptin genes in iranian cattle and buffalo populations using PCR-
RFLP. Iranian J Biotechnol. 3:104-108
Judajana, F. M. 1999. Imunogenetika.Dalam Imunologi Mukosal Kedokteran
Airlangga University Press.Hal.1-25.
Kuncorojakti, S. 2011. Major Histocompatibility Complex (MHC).
web.unair.ac.id. Diakses pada 26 Maret 2015.
Lande, R., and R. Thompson. 1990. Efficiency of marker-assisted selection in the
improvement of quantitative traits. Genetics. 124(1):743-756.
Nei M. 1987. Molecular evalutionery genetics. New York (NY): Columbia
University Press.
Nei, and Kumar. 2000. Molecular Evolutian and Phylogenetics.Oxford University
Press.New York.
Pandjassarame, K. 2009. Bioinformation discovery: data to knowledge in
Biology. Springer.
Petlanea, R. R. Noor, and R. R. A. Maheswari. 2012. The Genetic Diversity of
TLR4 MHC-DRB Genes in Dairy Goats Using PCR-RFLP Technique.
Media Peternakan. 66:91-95.
Sambrook,K.J.,E.F.Fritsch and T.Maniatis.1989.Molecular Cloning Laboratory
manual 3rd Ed.Cold Spring Harbour Lab. Press New York.

29
Schwaiger F. W., J. Buitkamp,E. Weyers, and J. T. Epplen.1993. TypingofMHC-
DRBgenes with the help of intronic simple repeatedDNA sequences. Mol.
Ecol. 2:55–59.
Sharif S., B. A. Mallard, B. N. Wilkie, J. M. Sargeant, H. M. Scott,J. C. M.
Dekkers,K. E. Leslie. 1998.Associations of the bovinemajor
histocompatibility complex DRB3 (BoLA-DRB3) with productiontraits in
Canadian dairy cattle.Animal Genetic. 30:157–160.
Sharma, A. K., B. Bhushan, P. Kumar, D. Sharma, V. K. Saxena, A.Sharma and
S. Kumar. 2005. DNA polymorphism of DRB 3.2locus and its association
with serum lysozyme in Rathi cattle(Bos indicus). J. Appl. Anim. Res.
28:61-64.
Singh, P.K., S.V. Singh, M.K. Singh, V.K. Saxena, A.V Singh, and J.S. Sohal.
2011. Genetic analysis of MHC Class II DRB gene in an endangered
Jamunapari breed of goats.Indian Journal of Biotechnology. 11 : 220-223.
Soller, M. and J.S. Backman.1983.Genetic polymorphism varietal identification
and genetic improvement. Theory Application Genetic. 67(1):25-33.
Sommer, S. 2005. The importance of immune gene variability (MHC) in
evolutionary ecology and conservation.Frontiers in Zoology. 2:1
Sufro, A.S.M. 1994. Keanekaragaman Genetik.Yogjakarta : Andi offset
Sutarno, 2006.Penggunaan Teknik Molekuler untuk Memperbaiki Sifat Produksi
Hewan Ternak. Universitas Sebelas Maret. Surakarta
Takada, T., Y. Kikkawa, H. Yonekawa, and T. Amano. 1998.Analysis of goat
MHC class II DRA and DRB genes:identification of the expressed gene
and new DRB alleles.Immunogenetics 48(1):408-412.
Tambasco D. D., C. C. P. Paz, M. Tambasco-Studart, A. P. Pereira, M. M.
Alencar, A. R. Freitas, L. L. Countinho, I.U. Packer and L. C. A. Regitano.
2003. Candidate genes for growth traits in beef cattle Bos Taurus x Bos
Indicus. J. Anim. Bred. Genet. 120: 51-60.
Tisdell, C. 2003. Socioeconomic causes of loss of animal genetic diversity:
analysis and assessment. Ecological Economics 45(3):365-376.
Tizard, I.R. 2000. Veterinary Immunology An Introduction. Sixth Edition.WB
Saunders Company. Harcourt Health Sciences Company. Philadelphia,
Pennsylvania.
Traherne, J. A., R. Horton, A. N. Roberts, M. Miretti, M. E. Hurles,C. A. Stewart,
J. L. Ashurst, A. M. Atrazhev, P. Coggill, S.Palmer, J. Almeida, S. Sims,
L. G. Wilming, J. Rogers, P. J.de Jong, M. Carrington, J. F. Elliott, S.

30
Sawcer, J. A. Todd, J.Trowsdale and S. Beck. 2006. Genetic analysis of
completely sequenced disease-associated MHC haplotypes identifies
shuffling of segments in recent human history. PLoS Genet. 2:81-92.
Van der Warf, J. 2000. An overview of animal breeding programs.Di dalam
Kinghorn B, Van der Werf J, editor. QTL course :Identifiying and
Incorporating Genetic Markers and Major Genes in Animal Breeding
Programs. Armidale, Australia : University of New England.
Yeh, F.C., R.C. Yang and T. Boyle.1999.POPGENE version 1.31 : Microsoft
Window-based Freeware for Population Genetic Analysis. Edmonton, AB.
Canada : University of Alberta Canada.

31
LAMPIRAN

32

33
RIWAYAT HIDUP
Evy Harjuna Saad dilahirkan pada tanggal 15 Juli 1992
di Kota Watampone, Kabupaten Bone, Provinsi Sulawesi
Selatan. Penulis adalah anak kedua dari lima bersaudara
dari pasangan Saade, S. Pd. dan St. Norma, S. Pd. Pada
tahun 1999 penulis memulai pendidikan di Sekolah Dasar
Negeri 206 Apala dan tamat pada tahun 2005. Pada tahun yang sama, penulis
melanjutkan ke Sekolah Menengah Pertama Negeri 1 Barebbo, tamat pada tahun
2008. Kemudian penulis melanjutkan ke SMAN 2 WATAMPONE pada tahun
2008 dan tamat pada tahun 2011. Pada tahun yang sama pula, penulis
melanjutkan pendidikan ke Perguruan Tinggi Negeri dan lulus melalui Jalur
SNMPTN di Program Studi Peternakan, Fakultas Peternakan, Universitas
Hasanuddin, Makassar.


















