
JOURNAL OF Vol. No. of pp. for in U.S.A. Proton Conduction ... · Proton Conduction and Bafilomycin...
6
THE JOURNAL OF BIOLWICAL CHEMISTRY 0 1994 by The American Society for Biochemistry and Molecular Biology, Inc. Vol. 269, No. 38, Issue of September 23, pp. 2351S23523, 1994 Printed in U.S.A. Proton Conduction and Bafilomycin Binding by the Vo Domain of the Coated Vesicle V-ATPase* (Received for publication, March 16, 1994, and in revised form, July 13, 1994) Jimin Zhang, Yu Feng8, and Michael ForgacO From the Department of Cellular and Molecular Phvsiology, rifts University School of Medicine, Boston, Massachusetts 02111 We have previously demonstrated that the V, domain of the coated vesicle V-ATPase, a 250-kDa integral com- plex, does not form a functional proton channel (Zhang, J., Myers, M., and Forgac, M. (1992) J. Biol. Chem. 267, 9773-9778). In the present study we describe dissocia- tion of the V, complex and separation of the V, subunits by gel filtration. Dicyclohexylcarbodiimide-inhibitable passive proton conductance of reconstituted vesicles containing reassembled V, subunits was measured in response to a K+/valinomycin-generated membrane po- tential. We observed that reconstitutedvesicles contain- ing the 17/19-kDa subunits carried out passive proton transport, with the addition of the 38- and 100-kDa sub- units increasing proton conductance. Reconstituted vesicles containing the 38- andor 100-kDa subunits showed no proton transport. Partial separation of the 17- and 19-kDa subunits revealed that the 17-kDa sub- unit alone carried out proton transport, with increased conductance on the addition of the 19-kDa subunit. These results indicate that the V, domain possesses the information necessary to form a dicyclohexylcarbodiim- ide-inhibitable passive proton channel. Bafilomycin binding by native and reassembled V, complexes was also measured by their ability to protect V-ATPase ac- tivity against bafilomycin inhibition.The native V, do- main, the isolated 100-kDa subunit and the 100/38-kDa subunits were able to protect against inhibition by bafilomycin, suggesting that the binding site for bafilo- mycin resides on the 100-kDa subunit. The vacuolar class of (H+)-ATPases’ (or V-ATPases) is respon- sible for acidification of intracellular compartments in eukary- otic cells, which is in turn crucial for a variety of cellular pro- cesses including receptor-mediated endocytosis, intracellular membrane traffic, macromolecular processing anddegrada- tion, and coupled transport in vacuolar compartments (for re- view see Refs. 1 and 2). V-ATPases have been purified from a number of sources, including clathrin-coated vesicles (31, chro- maffin granules (4), kidney microsomes (51, lysosomes (61, and the central vacuoles of Neurospora (71, yeast (8), and plants (9, lo), and appear to share a common structural motif. * This work was supported by National Institutes of Health Grant GM 34478. Fluorescence facilities were provided through National In- stitutes of Health Grant DK 34928. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. $ Supported through a postdoctoral fellowship from the Massachu- setts affiliate of the American Heart Association. 5 To whom correspondence should be addressed. The abbreviations used are: (H+)-ATPase, proton-translocating adenosine triphosphatase; C,,E,, polyoxyethylene-9-lauryl ether; ylcarbodiimide; DTT, dithiothreitol. ACMA, 9-amino-6-chloro-2-methoxyacridine; DCCD, N,N”dicyclohex- The coated vesicle V-ATPase is composed of two domains. The peripheral VI domain, which is located on the cytoplasmic side of the membrane, contains subunits of molecular mass 73(A)-, 58(B)-, 40-, 34-, and 33-kDa and forms a 500-kDa com- plex with the structure A3B,40134,33, (11-13). The V, domain, although not separately active as an ATPase, contains both catalytic and noncatalytic nucleotide binding sites located on the A and B subunits, respectively (3, 14, 15). The integral V, domain contains subunits of molecular mass 100, 38, 19, and 17(c)-kDa and forms a 250-kDa complex with the structure 100,38,19,c, (16, 13). The V, domain remains assembled as a 250-kDa complex even after dissociation of the V, subunits and solubilization in C,,E, (16). The 17-kDa c subunit is responsible for the sensitivity of proton translocation to DCCD (17). We have previously demonstrated that clathrin-coated vesicles contain a significant population of V, domains not com- plexed with V, (16). Moreover, reconstitution studies indicate that neither these free V, domains nor the V, domains gener- ated upon dissociation of V, form functional passive DCCD- inhibitable proton channels (16). To test whether the V, domain is capable of forming such DCCD-inhibitable passive proton channels, we have established conditions for dissociation of the V, complex and separation of the loo-, 38-, and 19/17-kDa subunits by gel filtration and have investigated the proton conduction properties of reconstituted vesicles containing dif- ferent combinations of V, subunits. The most specific inhibitors of V-ATPase thus far identified are bafilomycin (18) and the structurally related macrolide con- canamycin (19). Previous work has suggested that bafilomycin is able to bind to the V, domain of the V-ATPase (20). Using reconstituted vesicles containing the isolated V, domain and the separated V, subunits to protect V-ATPase activity from inhibition by bafilomycin, we have obtained further evidence for the presence of the bafilomycin binding site on the V, do- main and have gained additional information concerning sub- units responsible for binding of bafilomycin. EXPERIMENTAL PROCEDURES Materials-Calf brains were obtained fresh from a local slaughter- house. C,,E,, cholesterol, cholic acid, DCCD, valinomycin, and ATP (grade 11) were purchased from Sigma. Phosphatidylcholine and phos- phatidylserinewere obtained as chloroform solutions from Avanti Polar Lipids, Inc. and stored at -20 “C. 9-Amino-6-chloro-2-methoxyacridine (ACMA) was purchased from Molecular Probes. Bafilomycin was the kind gift of Dr. Karlheinz Altendorf, Universityof Osnabruck. Preparation of Stripped Clathrin-coated Vesicles and Isolation of the V, Domain-Clathrin-coated vesicles were prepared from calf brain as described previously (3). Vesicles were stripped of their clathrin coat by dilution (40-fold) into 5 m~ Tris (pH 8.5),150 mM sucrose, and 0.5 mM EDTA followed by incubation for 1 h at 23 “C and sedimentation for 1 h buffer (50 mM NaCl, 30mM KCl, 20 mM HEPES (pH 7.0), 0.2 mM EGTA, at 100,000 x g. The stripped vesicles were resuspended in solubilization 10% glycerol, 2 mM 2-mercaptoethanol) and solubilized at 1.0 mg of proteidml insolubilizationbuffercontaining 1.0% C,,Eg, 0.4 mg of phosphatidylcholine/ml, and 0.2 mg of phosphatidylserine/ml for 5 min 23518
Transcript of JOURNAL OF Vol. No. of pp. for in U.S.A. Proton Conduction ... · Proton Conduction and Bafilomycin...

THE JOURNAL OF BIOLWICAL CHEMISTRY 0 1994 by The American Society for Biochemistry and Molecular Biology, Inc.
Vol. 269, No. 38, Issue of September 23, pp. 2351S23523, 1994 Printed in U.S.A.
Proton Conduction and Bafilomycin Binding by the Vo Domain of the Coated Vesicle V-ATPase*
(Received for publication, March 16, 1994, and in revised form, July 13, 1994)
Jimin Zhang, Yu Feng8, and Michael ForgacO From the Department of Cellular and Molecular Phvsiology, rifts University School of Medicine, Boston, Massachusetts 02111
We have previously demonstrated that the V, domain of the coated vesicle V-ATPase, a 250-kDa integral com- plex, does not form a functional proton channel (Zhang, J., Myers, M., and Forgac, M. (1992) J. Biol. Chem. 267, 9773-9778). In the present study we describe dissocia- tion of the V, complex and separation of the V, subunits by gel filtration. Dicyclohexylcarbodiimide-inhibitable passive proton conductance of reconstituted vesicles containing reassembled V, subunits was measured in response to a K+/valinomycin-generated membrane po- tential. We observed that reconstituted vesicles contain- ing the 17/19-kDa subunits carried out passive proton transport, with the addition of the 38- and 100-kDa sub- units increasing proton conductance. Reconstituted vesicles containing the 38- andor 100-kDa subunits showed no proton transport. Partial separation of the 17- and 19-kDa subunits revealed that the 17-kDa sub- unit alone carried out proton transport, with increased conductance on the addition of the 19-kDa subunit. These results indicate that the V, domain possesses the information necessary to form a dicyclohexylcarbodiim- ide-inhibitable passive proton channel. Bafilomycin binding by native and reassembled V, complexes was also measured by their ability to protect V-ATPase ac- tivity against bafilomycin inhibition. The native V, do- main, the isolated 100-kDa subunit and the 100/38-kDa subunits were able to protect against inhibition by bafilomycin, suggesting that the binding site for bafilo- mycin resides on the 100-kDa subunit.
The vacuolar class of (H+)-ATPases’ (or V-ATPases) is respon- sible for acidification of intracellular compartments in eukary- otic cells, which is in turn crucial for a variety of cellular pro- cesses including receptor-mediated endocytosis, intracellular membrane traffic, macromolecular processing and degrada- tion, and coupled transport in vacuolar compartments (for re- view see Refs. 1 and 2). V-ATPases have been purified from a number of sources, including clathrin-coated vesicles (31, chro- maffin granules (4), kidney microsomes (51, lysosomes (61, and the central vacuoles of Neurospora (71, yeast (8), and plants (9, lo), and appear to share a common structural motif.
* This work was supported by National Institutes of Health Grant GM 34478. Fluorescence facilities were provided through National In- stitutes of Health Grant DK 34928. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
$ Supported through a postdoctoral fellowship from the Massachu- setts affiliate of the American Heart Association.
5 To whom correspondence should be addressed. The abbreviations used are: (H+)-ATPase, proton-translocating
adenosine triphosphatase; C,,E,, polyoxyethylene-9-lauryl ether;
ylcarbodiimide; DTT, dithiothreitol. ACMA, 9-amino-6-chloro-2-methoxyacridine; DCCD, N,N”dicyclohex-
The coated vesicle V-ATPase is composed of two domains. The peripheral VI domain, which is located on the cytoplasmic side of the membrane, contains subunits of molecular mass 73(A)-, 58(B)-, 40-, 34-, and 33-kDa and forms a 500-kDa com- plex with the structure A3B,40134,33, (11-13). The V, domain, although not separately active as an ATPase, contains both catalytic and noncatalytic nucleotide binding sites located on the A and B subunits, respectively (3, 14, 15). The integral V, domain contains subunits of molecular mass 100, 38, 19, and 17(c)-kDa and forms a 250-kDa complex with the structure 100,38,19,c, (16, 13). The V, domain remains assembled as a 250-kDa complex even after dissociation of the V, subunits and solubilization in C,,E, (16). The 17-kDa c subunit is responsible for the sensitivity of proton translocation to DCCD (17).
We have previously demonstrated that clathrin-coated vesicles contain a significant population of V, domains not com- plexed with V, (16). Moreover, reconstitution studies indicate that neither these free V, domains nor the V, domains gener- ated upon dissociation of V, form functional passive DCCD- inhibitable proton channels (16). To test whether the V, domain is capable of forming such DCCD-inhibitable passive proton channels, we have established conditions for dissociation of the V,, complex and separation of the loo-, 38-, and 19/17-kDa subunits by gel filtration and have investigated the proton conduction properties of reconstituted vesicles containing dif- ferent combinations of V, subunits.
The most specific inhibitors of V-ATPase thus far identified are bafilomycin (18) and the structurally related macrolide con- canamycin (19). Previous work has suggested that bafilomycin is able to bind to the V, domain of the V-ATPase (20). Using reconstituted vesicles containing the isolated V, domain and the separated V, subunits to protect V-ATPase activity from inhibition by bafilomycin, we have obtained further evidence for the presence of the bafilomycin binding site on the V, do- main and have gained additional information concerning sub- units responsible for binding of bafilomycin.
EXPERIMENTAL PROCEDURES Materials-Calf brains were obtained fresh from a local slaughter-
house. C,,E,, cholesterol, cholic acid, DCCD, valinomycin, and ATP (grade 11) were purchased from Sigma. Phosphatidylcholine and phos- phatidylserine were obtained as chloroform solutions from Avanti Polar Lipids, Inc. and stored at -20 “C. 9-Amino-6-chloro-2-methoxyacridine (ACMA) was purchased from Molecular Probes. Bafilomycin was the kind gift of Dr. Karlheinz Altendorf, University of Osnabruck.
Preparation of Stripped Clathrin-coated Vesicles and Isolation of the V, Domain-Clathrin-coated vesicles were prepared from calf brain as described previously (3). Vesicles were stripped of their clathrin coat by dilution (40-fold) into 5 m~ Tris (pH 8.5), 150 mM sucrose, and 0.5 mM EDTA followed by incubation for 1 h at 23 “C and sedimentation for 1 h
buffer (50 mM NaCl, 30 mM KCl, 20 mM HEPES (pH 7.0), 0.2 mM EGTA, at 100,000 x g . The stripped vesicles were resuspended in solubilization
10% glycerol, 2 mM 2-mercaptoethanol) and solubilized at 1.0 mg of proteidml in solubilization buffer containing 1.0% C,,Eg, 0.4 mg of phosphatidylcholine/ml, and 0.2 mg of phosphatidylserine/ml for 5 min
23518

Proton Conduction and Bafilomycin Binding by V, 23519
a t 23 "C followed by 15 min a t 4 "C and sedimentation at 150,000 x g for 1 h. 0.7 ml of supernatant was applied to each 11-ml 12-25% glycerol density gradient prepared in solubilization buffer containing 0.02% C,,E,, 8 pg of phosphatidylcholine/ml and spun at 38,000 rpm in a Beckman SW-41 rotor for 16 h. The resultant gradients were fraction- ated from the bottom to give 19-20 fractions. The V, containing frac- tions from the gradient (fractions 12-14) were pooled and concentrated 20-fold using an Amicon cell with a YM30 membrane.
Dissociation and Reassembly of the V, Domain-Dissociation of the V, domain was carried out at a final protein concentration of 160 pg/ml, 3 M sodium trichloroacetate, 1% sodium deoxycholate, 1 mM EDTA, 10 m~ Tris (pH K O ) , 1 mM DTT. The mixture (6-8 ml) was stirred at room temperature for 20 h and then applied to a 2.5 x 120-cm Sephadex G-200 column at 4 "C equilibrated with 1 M sodium trichloroacetate, 2% sodium cholate, 1 mM EDTA, 1 mM D m , 10 m~ Tris (pH 8.0) and eluted with the same buffer a t a flow rate of 3-5 mvh. Fractions containing the 100-kDa subunit, the 38-kDa subunit, and the 17/19-kDa subunits were pooled separately and dialyzed against two changes of 50 volumes of dialysis buffer (30 m~ NaC1, 0.5% sodium cholate, 10% glycerol, 1 mM DTT, 10 mM Tris (pH 8.0)) for 16 h at 4 "C. The pooled fractions were concentrated by application to a 0.7-1111 DEAE-cellulose column equili- brated with the above dialysis buffer and eluted with 1 ml of 500 mM NaCl in the dialysis buffer followed by dialysis overnight against the dialysis buffer a t 4 "C to remove excess salt and store at -80 "C until use. The yield following dissociation and purification was approxi- mately 90% for the 100-kDa subunit, 70% for the 38-kDa subunit, and 30% for the 17/19-kDa subunits. Reassembly was carried out by com- bining the pooled concentrated V,, subunits in the above dialysis buffer containing KC1 added to a final concentration of 500 mM, followed by incubation for 2 h at room temperature with stirring. The subunits were combined to give a final molar ratio approximating that of the initial V,, complex (i.e. 100,38,19,17,).
Reconstitution of Native and Reassembled V,-Reconstitution of the native and reassembled V, was carried out essentially as described previously (16). To 0.5 ml of the isolated V, domain (20-30 pg of protein) or 0.5 ml of various mixtures of V, subunits was added 0.1 ml of 5% sodium cholate, 4 mg of phosphatidylcholine/ml, and 2 mg of phosphatidylserine/ml followed by 0.14 ml of 10% sodium cholate, 7 mg of phosphatidylcholine/ml, 4 mg of cholesterollml in 150 mM NaCl, 2 mM 2-mercaptoethanol. The mixture was incubated for 5 min at 23 "C and 15 min at 4 "C followed by dialysis for 3 days at 4 "C against 5 changes of 200 volumes of buffer containing 150 mM KCl, 20 mM HEPES pH 7.0, 0.2 mM EGTA, 10% glycerol, 2 m~ 2-mercaptoethanol using Spectral Por-2 dialysis tubing with a M, cutoff of 12,000-14,000. Reconstitution of V, subunits for the bafilomycin protection assay was carried out by dialysis against solubilization buffer containing 50 mM NaCl and 30 mM KC1 instead of 150 mM KC].
Measurement of Passive Proton Flux in Reconstituted Vesicles Con- taining Reassembled V, Subunits-Measurement of passive proton flux in reconstituted vesicles containing V, subunits was carried out as described previously (16). Vesicles were diluted 1:20 into 150 mM NaCl, 20 mM HEPES pH 7.0, 0.2 mM EGTA, 2 mM 2-mercaptoethanol, 10% glycerol, 0.5 mg bovine serum albumin (fatty acid-free) per ml and 2 UM
ACMA. The fatty acid-free bovine serum albumin helped to reduce the nonspecific passive proton permeability of the reconstituted vesicles by absorbing trace lipid oxidation products that increased proton perme- ability. Proton flux was initiated by the addition of 20 ILM valinomycin, which resulted in the generation of a negative interior membrane po- tential. Proton uptake was monitored by fluorescence quenching of ACMA using excitation and emission wavelengths of 410 and 490 nm, respectively.
Bafilomycin Protection Assay-The ability of reconstituted vesicles containing either the isolated V, domain or various combinations of V, subunits to protect against inhibition of V-ATPase by bafilomycin was assayed as follows. Reconstituted vesicles (50 pl) were preincubated with 5 nM bafilomycin (a concentration causing approximately 75% inhibition of V-ATPase activity) in solubilization buffer containing 50 mM NaCl, 30 mM KC], 20 mM HEPES pH 7.0, 10% glycerol, 0.2 mM EGTA, and 2 mM 2-mercaptoethanol at 23 "C for 3 min. Stripped vesicles (15 pg of protein) containing the V-ATPase are then added to the mixture and incubated for 3 min at 23 "C followed by addition of 2 UM ACMA. ATP-dependent proton transport was initiated by addition of 0.5 mM ATP and 1.0 mM MgSO,, and the fluorescence change was moni- tored as described above. Control samples received reconstituted vesicles containing only lipid but no protein in order to correct for any nonspecific adsorption of bafilomycin by the added lipid.
RESULTS
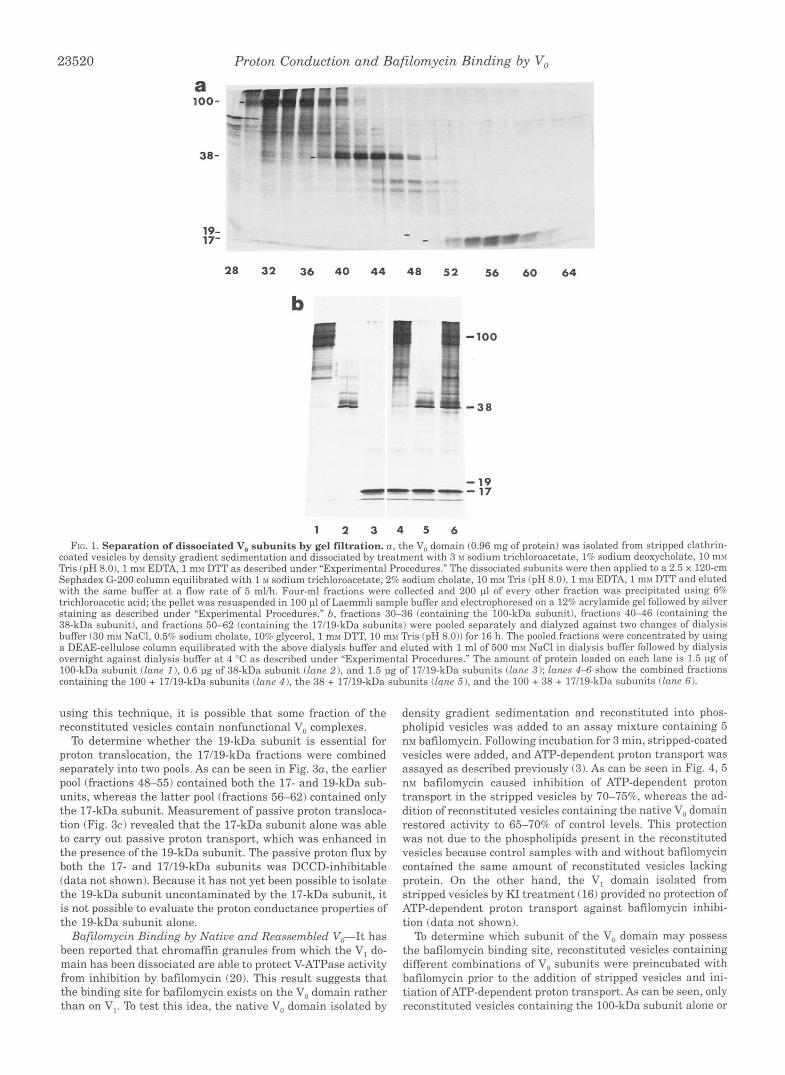
Dissociation of Vu and Zsolation of the Vu Subunits-We have previously demonstrated that clathrin-coated vesicles contain a significant population of free V, domains that, like the V, gen- erated by dissociation of V,, does not form a functional proton channel (16). The V,, domain solubilized in C,,E, was isolated from clathrin-coated vesicles by glycerol density gradient sedi- mentation as described previously (16). Using a modification of the protocol employed in isolation of the F, subunits of the E. coli F-ATPase (21), the V, domain was dissociated by treatment with 3.0 M sodium trichloroacetate, 10 mM Tris (pH 8.01,l.O mM EDTA, 1% deoxycholate, and 1 mM DTT. Following incubation overnight, the mixture was applied to a 2.5 x 120-cm Sephadex G-200 column and eluted with 1.0 M sodium trichloroacetate, 10 mM Tris (pH 8.01, 2% sodium cholate, 1 m~ EDTA, and 1 mM DTT. The fractions were analyzed by SDS-polyacrylamide gel electrophoresis on a 12% acrylamide gel.
As can be seen in Fig. la, the 100- and 38-kDa subunits were effectively separated from each other and from the 17-kDa subunit. The 19-kDa subunit does not visualize well by either silver staining or Coomassie blue staining, but it could be seen to partially resolve from the 17-kDa subunit in more darkly stained gels (see Fig. 3a). As we have noted previously (161, because of the migration position of the 250-kDa V, complex on the glycerol density gradients, additional polypeptides are pres- ent in the fractions derived from the gel filtration column.
Reassembly, Reconstitution, and Proton Danslocation by the Vu Domain-The fractions containing the loo-, 38-, and 17119- kDa subunits were pooled separately, dialyzed a t 4 "C against dialysis buffer (30 mM NaCl, 0.5% sodium cholate, 10% glycerol, 1 mM DTT, 10 mM Tris (pH 8.011, and concentrated by applica- tion to a 0.7-ml DEAE-cellulose column eluted with the above dialysis buffer containing 0.5 M NaC1. The fractions were then dialyzed against dialysis buffer at 4 "C overnight. Fig. l b shows pooled fractions containing the 100-kDa subunit (fractions 30- 36; lane 11, the 38-kDa subunit (fractions 40-46; lane 21, the 17/19-kDa subunits (fractions 50-62; lane 3) ; and the combined fractions containing the 100 + 17/19-kDa subunits (lane 41, the 38 + 17/19-kDa subunits (lane 51, and the 100 + 38 + 17/19-kDa subunits (lane 6). Following combination of the V, subunits, assembly was initiated by the addition of KC1 to a final con- centration of 0.5 M. Samples were stirred a t 23 "C for 2 h. The V, subunits were then reconstituted into phospholipid vesicles by cholate dialysis as described previously (16). The reconsti- tuted vesicles were prepared in the presence of 150 mM KC1 such that dilution of the vesicles into buffer containing NaCl in place of KC1 and the addition of valinomycin resulted in the generation of a negative interior membrane potential (16). Pas- sive proton flux was measured using the fluorescence dye ACMA, which on accumulation in acidic compartments under- goes fluorescence quenching.
As can be seen in Fig. 2a, reconstituted vesicles containing the 17/19-kDa subunits possessed significantly higher passive proton permeability than phospholipid vesicles containing no protein. Reconstituted vesicles containing, in addition to the 17/19-kDa subunits, the 100- and 38-kDa subunits, either alone or in combination, showed increased passive proton transport. By contrast, the 100- or 38-kDa subunits, either alone or in combination, did not show significant passive proton conduc- tion (data not shown). To determine whether the observed pas- sive proton translocation was specific, reconstituted vesicles were treated with 50 UM DCCD for 30 min at 4 "C prior to measurement of proton flux. As can be seen in Fig. 2b, DCCD effectively inhibited passive proton translocation in reconsti- tuted vesicles containing the V, subunits. Because it is not possible to determine the number of functional proton channels
23520
a 100-
38-
19- 1 7-
Proton Conduction and Bafilomycin Binding by V,
1 - 100
“”- 17 - 19
__-- ” -
1 2 3 4 5 6 Frc. 1. Separation of dissociated V,, subunits by gel filtration. a , the V, domain (0.96 mg of protein) was isolated from stripped clathrin-
coated vesicles by density gradient sedimentation and dissociated by treatment with 3 XI sodium trichloroacetate, 1% sodium deoxycholate, 10 mx! Tris (pH 8.01, 1 mM EDTA, 1 my DTT as described under “Experimental Procedures.” The dissociated subunits were then applied to a 2.5 x 120-cm Sephadex G-200 column equilibrated with 1 M sodium trichloroacetate, 2% sodium cholate, 10 m\l Tris (pH 8.01, 1 mxf EDTA, 1 mhr DTT and eluted with the same buffer a t a flow rate of 5 mlh. Four-ml fractions were collected and 200 pl of every other fraction was precipitated using 6% trichloroacetic acid; the pellet was resuspended in 100 pl of Laemmli sample buffer and electrophoresed on a 12% acrylamide gel followed by silver staining as described under “Experimental Procedures.” b, fractions 30-36 (containing the 100-kDa subunit), fractions 4 0 4 6 (containing the 38-kDa subunit), and fractions 50-62 (containing the 17/19-kDa subunits) were pooled separately and dialyzed against two changes of dialysis buffer (30 mv NaCI, 0.5% sodium cholate, 10% glycerol, 1 mlr DTT, 10 mx1 Tris (pH 8.0)) for 16 h. The pooled fractions were concentrated by using a DEAE-cellulose column equilibrated with the above dialysis buffer and eluted with 1 ml of 500 mv NaCl in dialysis buffer followed by dialysis overnight against dialysis buffer a t 4 “C as described under “Experimental Procedures.” The amount of protein loaded on each lane is 1.5 pg of 100-kDa subunit (lane I ), 0.6 pg of 38-kDa subunit (lane 2) , and 1.5 pg of 17/19-kDa subunits (lane 3); lanes 4-6 show the combined fractions containing the 100 + 17/19-kDa subunits (lane 4 ) . the 38 + 17/19-kDa subunits (lane 5). and the 100 + 38 + 17/19-kDa subunits (lane 61.
using this technique, it is possible that some fraction of the reconstituted vesicles contain nonfunctional V, complexes.
To determine whether the 19-kDa subunit is essential for proton translocation, the 17/19-kDa fractions were combined separately into two pools. As can be seen in Fig. 3a, the earlier pool (fractions 48-55) contained both the 17- and 19-kDa sub- units, whereas the latter pool (fractions 56-62) contained only the 17-kDa subunit. Measurement of passive proton transloca- tion (Fig. 3c) revealed that the 17-kDa subunit alone was able to carry out passive proton transport, which was enhanced in the presence of the 19-kDa subunit. The passive proton flux by both the 17- and 17/19-kDa subunits was DCCD-inhibitable (data not shown). Because it has not yet been possible to isolate the 19-kDa subunit uncontaminated by the 17-kDa subunit, it is not possible to evaluate the proton conductance properties of the 19-kDa subunit alone.
Bafilomycin Binding by Native and Reassembled V,-It has been reported that chromaffin granules from which the VI do- main has been dissociated are able to protect V-ATPase activity from inhibition by bafilomycin (20). This result suggests that the binding site for bafilomycin exists on the V, domain rather than on V,. To test this idea, the native V, domain isolated by
density gradient sedimentation and reconstituted into phos- pholipid vesicles was added to an assay mixture containing 5 nM bafilomycin. Following incubation for 3 min, stripped-coated vesicles were added, and ATP-dependent proton transport was assayed as described previously (3). As can be seen in Fig. 4, 5 nM bafilomycin caused inhibition of ATP-dependent proton transport in the stripped vesicles by 70-75%, whereas the ad- dition of reconstituted vesicles containing the native V, domain restored activity to 65-70% of control levels. This protection was not due to the phospholipids present in the reconstituted vesicles because control samples with and without bafilomycin contained the same amount of reconstituted vesicles lacking protein. On the other hand, the VI domain isolated from stripped vesicles by KI treatment (16) provided no protection of ATP-dependent proton transport against bafilomycin inhibi- tion (data not shown).
To determine which subunit of the V, domain may possess the bafilomycin binding site, reconstituted vesicles containing different combinations of V, subunits were preincubated with bafilomycin prior to the addition of stripped vesicles and ini- tiation ofATP-dependent proton transport. As can be seen, only reconstituted vesicles containing the 100-kDa subunit alone or

Proton Conduction and Bafilomycin Binding by V, 23521
aao
T h e (min)
b a o
75
- 3 70
aI C - - E 65 C
0
60 0 > U. - f 55 - m -
50
45
t
r 40 -L
0 1 2 3 4 5 6
Time (min)
FIG. 2. Passive proton translocation by reconstituted vesicles containing V, subunits. Fractions containing the loo-, 38-, and 17/ 19-kDa subunits obtained by gel filtration as described in the legend to Fig. 1 were reassembled either separately or in combination in 500 mxr KC1 and 2 mM DTT a t 23 "C for 2 h and then reconstituted into artificial phospholipid vesicles containing 150 mhl KCl, 20 mM HEPES pH 7.0,0.2 mht EGTA, 10% glycerol, and 2 mM 2-mercaptoethanol by cholate dial- ysis as described under "Experimental Procedures." 25 pl of reconsti- tuted vesicles (containing 73 pg of lipid) were diluted 1:20 into the same buffer containing 150 my NaCl in place of KC1 plus 0.5 mg/ml bovine serum albumin (fatty acid-free) per ml and 2 p~ ACMA. At t = 0, proton flux was initiated by the addition of 20 nM valinomycin and monitored by measurement of the fluorescence intensity a t 490 nm on excitation a t 410 nm. At t = 5 min, vesicles were permeabilized to protons by the addition of 1 p~ carbonyl cyanide p-chlorophenylhydrazone. Shown in a are the results obtained with untreated reconstituted vesicles, while h shows the results obtained following treatment of reconstituted vesicles with 50 UY DCCD for 30 min a t 4 "C prior to measurement of proton flux. W, control vesicles containing only lipid and no protein; 0, vesicles containing 0.75 pg of 17/19-kDa subunits; +, vesicles containing 0.75 pg of 17/19 and 0.3 pg of 38-kDa subunits; 0 , vesicles containing 0.75
a 19- . . . . . ~
17- Ir.r(l311LI
48 50 52 54 56 58 60 62 1-17/19 -~-17-~
100 73 58 4 0
$334
19 17
17 17/19 VIVo
1 I
0 1 2 3 4 5 6
Time (min)
FIG. 3. Partial resolution of the 17- and 19-kDa subunits by gel filtration and comparison of passive proton translocation by reconstituted vesicles containing either the 17/19-kDa subunits or the 17-kDa subunit alone. a, the V,, subunits were dissociated and separated by gel filtration and 200-pl aliquots of fractions 48-62 were precipitated with trichloroacetic acid and separated by SDS-polyacryl- amide gel electrophoresis as described in the legend to Fig. 1, except that the gel was developed longer during silver staining. h, fractions 48-55 (containing both the 17- and 19-kDa subunits) and fractions 56-62 (containing only the 17-kDa subunit) were pooled, concentrated, and reconstituted separately as described under "Experimental Proce- dures" and the legend to Fig. 1. 40 p1 of reconstituted vesicles were applied to a 12% polyacrylamide gel followed by silver staining. The lane marked V,V, contains 50 pl of purified V-ATPase (1.25 pg of pro- tein). c, reconstituted vesicles (25 pl) containing either lipid alone (W), 0.4 pg of 17-kDa subunit ( 0 , or 0.5 pg of 17/19-kDa subunits ( + ) were prepared and assayed for passive proton translocation as described in the legend to Fig. 2.
in combination with the 38-kDa subunit showed significant protection of inhibition by bafilomycin. None of the other V, subunits, either alone or in combination, showed significant protection of activity. The 100-kDa subunit also showed a sim- ilar level of protection of ATPase activity from bafilomycin in- hibition using a reconstituted preparation of purified V-ATPase (data not shown). It was surprising that reconstituted vesicles containing the 17/19-kDa subunits in addition to the 100-kDa
pg of 17/19 and 0.75 pg of 100-kDa subunits; A, vesicles containing 0.75 pg of 17/19, 0.3 pg of 38, and 0.75 pg of 100-kDa subunits.

23522 Proton Conduction and Bafilomycin Binding by V,
100
" 90
-
- g 80
::
--
5 70
-- 2 30
-- E 4 0
-- c 50
-- C 6 0
--
; I
- d 20 "
10
o r -
_-
- 0 -
jt-- +Bafilornycin -4
FIG. 4. Protection of V-ATPase activity from bafilomycin inhi- bition by reconstituted vesicles containing the native V, domain and separated V, subunits. 50 pl of reconstituted vesicles (146 pg of lipid) containing the indicated V, subunits (1.5 pg of the 17/19- or 100-kDa subunits, 0.6 pg of the 38-kDa subunit) or the intact V,, domain (2-3 pg of protein) were prepared as described under "Experimental Procedures" and incubated with 5 n~ bafilomycin in 50 mM NaCl, 30 m~ KCl, 20 mM HEPES pH 7.0, 0.2 mM EGTA, 2 mM 2-mercaptoethanol,
then added, followed by incubation for 3 min at 23 "C and the addition 10% glycerol for 3 min at 23 "C. Stripped vesicles (15 pg of protein) were
of 2 UM ACMA. ATP-dependent proton transport by stripped vesicles was initiated by the addition of 0.5 mM ATP and 1.0 mM MgSO, and measured as the carbonyl cyanide p-chlorophenylhydrazone-reversible portion of the ATP-dependent decrease in fluorescence intensity at 490 nm (excitation at 410 nm). Minus bafilomycin and control samples received the same amount of reconstituted vesicles containing only lipid and no protein.
subunit were unable to prevent bafilomycin inhibition. These results suggest that, although the 100-kDa subunit appears to be capable of binding bafilomycin, this binding site is masked in reassembled complexes containing the 100- and 17/19-kDa subunits. No consistent effect of bafilomycin on passive proton transport by reassembled V, complexes was observed (data not shown). To test the reversibility of bafilomycin binding to the V-
ATPase, the order of addition of bafilomycin, reconstituted V,, and stripped vesicles was altered. Thus, instead of preincubat- ing bafilomycin and reconstituted V,, prior to the addition of stripped vesicles, bafilomycin was added directly to stripped vesicles followed by incubation for 3 min, addition of reconsti- tuted V,, incubation for an additional 3 min, and assay of ATP- dependent proton transport activity. As can be seen in Fig. 5 , the level of protection against bafilomycin inhibition provided by the reconstituted V, was independent of the order of addi- tion, indicating that inhibition of V-ATPase activity by bafilo- mycin was reversible within 3 min. The passive proton trans- location and bafilomycin binding properties of the native and reassembled V, domains are discussed below.
DISCUSSION
The integral V, domain of the V-ATPase complex is respon- sible for proton translocation across the vacuolar membrane. This domain is composed of four different polypeptide chains of molecular mass loo-, 38-, 19-, and 17(c)-kDa assembled into a 250-kDa complex containing six copies of the c subunit and single copies of the remaining polypeptides (13, 16). The 17- kDa c subunit is a highly hydrophobic proteolipid responsible
8 0 85 1 1 \\
4 0 4 5 5 0 1 2 3 4 5 6
Time (mln)
FIG. 5. Reversibility of bafilomycin inhibition of V-ATPase by V,. Reconstituted vesicles (50 pl containing 146 pg of lipid) containing either no protein (0) or the intact V, domain (1.2 pg of protein) (1, + ) were incubated for 3 min in 50 mM NaCl, 30 mM KCl, 20 mM HEPES pH 7.0, 0.2 mM EGTA, 2 mM 2-mercaptoethanol, 10% glycerol either in the presence (0, +) or absence (1) of 5 nM bafilomycin followed by the addition of stripped vesicles (15 pg of protein), incubation for an addi- tional 3 min, and assay ofATP-dependent proton transport by addition ofACMA and MgATP as described in the legend to Fig. 4. Alternatively,
bafilomycin followed by the addition of reconstituted vesicles (50 pl) stripped vesicles (15 pg protein) were first incubated for 3 min with 5 n~
ATP-dependent proton transport as described ( 0 ). The observation that containing V,, (2-3 pg of protein), incubation for 3 min, and assay of
the same level of protection was obtained whether the stripped vesicles were preincubated with bafilomycin or whether bafilomycin was first incubated with reconstituted V,, prior to addition of stripped vesicles indicates that reversal of bafilomycin inhibition occurs within 3 min of the addition of the reconstituted V, domain.
for the sensitivity of proton translocation to inhibition by DCCD (17). The c subunit has four putative transmembrane helices with a single buried carboxyl residue in the carboxyl- terminal most CY helix, the likely site of reaction with DCCD (22). The V-ATPase c subunit appears to have been derived by gene duplication of the homologous 8-kDa F-ATPase c subunit (23). While the 19-kDa subunit is also highly hydrophobic (131, the 38-kDa subunit lacks putative transmembrane helics (24), and thus appears to remain tightly associated with the V,, do- main through protein-protein interactions. The 100-kDa sub- unit is a transmembrane glycoprotein possessing a significant luminal domain (12, 13). Cloning studies have indicated that it is organized into an amino-terminal hydrophilic domain and a carboxyl-terminal hydrophobic domain possessing six putative transmembrane helices (25, 26).
We have previously demonstrated that clathrin-coated vesicles contain a significant population of V, domains not com- plexed with V, subunits (16). Moreover, neither this endoge- nous V, population nor the V, domains generated upon disso- ciation of the V, sector are functional as DCCD-inhibitable passive proton channels upon reconstitution into phospholipid vesicles (16). To determine whether the V, sector contains the information necessary to form a passive proton channel, the V, subunits were dissociated from each other and separated by gel filtration using conditions similar to those described for isola- tion of the F, subunits (21). Reconstitution of the V, subunits separately revealed that vesicles containing 17119-kDa sub-

Proton Conduction and Bafilomycin Binding by V,, 23523
units showed DCCD-inhibitable passive proton transport whereas the loo-, 38-, or 100/38-kDa subunits were unable to form passive proton channels. Nevertheless, the 100- and 38- kDa subunits, either alone or in combination, did enhance the passive proton conductance of vesicles containing the 17/19- kDa subunits. Partial separation of the 17- and 19-kDa sub- units reveals that the 17-kDa subunit alone is able to carry out proton conduction, which is enhanced in the presence of the 19-kDa subunit. These results suggest that the 17-kDa sub- units form the minimal unit necessary for passive proton trans- port but that the loo-, 38-, and 19-kDa subunits are able to enhance the proton conductance through reassembled V,. This is in contrast to reconstitution of the F, domain, where addition of the complete complement of a, b, and c subunits is necessary to form a DCCD-inhibitable proton channel (21).
These results are somewhat surprising in light of our previ- ous observation that the native V, domain is not a functional proton channel (16). There are several possible explanations for the difference in passive proton conduction between native and reassembled V,. First, it is possible that as a result of treatment under dissociating conditions (3.0 M trichloroacetate, 1% deoxy- cholate), one or more of the V, subunits has been altered in conformation such that the structure of the reassembled com- plex is different that of the native V, domain. Alternatively, it is possible that during isolation of the V, subunits by gel fil- tration, some inhibitor of passive proton transport has been lost such that the reassembled V, lacking this inhibitor is func- tional. If V, is treated under dissociating conditions and then immediately dialyzed and reconstituted without separation of the V, subunits by gel filtration, no DCCD-inhibitable passive proton transport is observed (data not shown). This result sug- gests the possibility that even after treatment under dissociat- ing conditions the V, subunits are able to reassemble into a complex that resembles the native V, in its lack of proton chan- nel activity. Alternatively, the V, treated under these conditions may have simply failed to assemble into a functional proton channel. We have thus far been unable to isolate an inhibitor of passive proton conductance of the reassembled V, domain by pooling and concentrating the remaining fractions from the gel filtration column, suggesting that if such an inhibitor exists, it is either unstable in the absence of other V, subunits or is irreversibly lost during gel filtration, possibly through nonspe- cific interactions with the gel matrix.
It has previously been reported that the c subunit of the coated vesicle V-ATPase, isolated by toluene extraction, is able to form a DCCD-inhibitable proton channel by itself on core- constitution with bacteriorhodopsin (27). In our hands, this toluene extraction protocol results in recovery of less than 5% of the total c subunit present in coated vesicles as judged by extraction with either chlorofordmethanol or butanol. More- over, we have been unable to observe the formation of a DCCD- inhibitable proton channel employing our assay system with material extracted by any of these procedures. Nevertheless, our results indicate that proton conductance is observed upon reconstitution of the purified 17-kDa subunit isolated by deter- gent solubilization and gel filtration as described. Thus the 17-kDa subunit does appear to contain the information neces- sary to form a DCCD-inhibitable proton channel.
Bafilomycin (18) and the structurally related concanamycin (19) are highly specific inhibitors of the V-ATPases that inhibit activity in the nanomolar to 0.1 nanomolar range, respectively. It has previously been observed that chromaffin granules from which the V, domain has been dissociated are able to protect intact chromaffin granule V-ATPase from inhibition by bafilo-
mycin (20). This result was interpreted to indicate that the bafilomycin binding site is located on the V, domain. To test this idea, reconstituted vesicles containing the isolated V, do- main were tested for their ability to protect V-ATPase activity from inhibition by bafilomycin. Our results indicate that the isolated V, domain is able to protect against bafilomycin inhi- bition. The failure of reconstituted vesicles lacking protein to prevent inhibition indicates that protection is not due to non- specific adsorption of bafilomycin to the reconstituted vesicles. In addition, the results presented in Fig. 5 demonstrate that bafilomycin binding to the V-ATPase is reversible. Comparison of the protection afforded by reconstituted vesicles containing individual V, subunits indicates that only the 100-kDa subunit (in the presence or absence of the 38-kDa subunit) is able to significantly protect activity, suggesting that the 100-kDa sub- unit may possesses a binding site for bafilomycin. It should be noted that these results provide only indirect evidence for bafilomycin binding to the 100-kDa subunit and that further experiments will be required to confirm this interpretation. Surprisingly, reconstituted vesicles containing the 17/19-kDa subunits in addition to the 100-kDa subunit show no protec- tion, suggesting that the bafilomycin binding site on the 100- kDa subunit is masked in complexes containing the 17/19-kDa subunits. Thus reassembled and native V, complexes differ in both their proton translocation activity and their ability to bind bafilomycin. Nevertheless, the results presented suggest that the V, complex does contain the information necessary to form a DCCD-inhibitable proton channel and that the 100-kDa sub- unit may possess the binding site for bafilomycin.
1. 2. 3.
4. 5. 6.
7.
8.
10. 9.
11.
12.
13.
14. 15.
16. 17. 18.
19.
20.
21. 22.
23. 24.
25.
26.
REFERENCES Forgac, M. (1989) Physiol. Reu. 69, 765-796 Forgac, M. (1992) J. Exp. Biol. 172, 155-169 Arai, H., Berne, M., Terres, G., Terres, H., Puopolo, K., and Forgac, M. (1987)
Moriyama, Y., and Nelson, N. (1989) J. Biol. Chem. 264,35773582 Gluck, S., and Caldwell, J. (1987) J. Biol. Chem. 262, 15780-15789 Arai, K., Shimaya, A., Hiratani, N., and Ohkuma, S. (1993) J. Biol. Chem. 268,
Bowman, B. J., Dschida, W. J., Harris, T., and Bowman, E. J. (1989) J. Biol.
Kane, P. M., Yamashiro, C. T., and Stevens, T. H. (1989) J. B i d . Chem. 264,
Lai, S., Randall, S. K., and Sze, H. (1988) J. Biol. Chem. 263, 16731-16737 Parry, R. V., Turner, J. C., and Rea, P. A. (1989) J. Biol. Chem. 264, 20025-
Adachi, I., Arai, H., Pimental, R., and Forgac, M. (1990) J. B i d . Chem. 265,
Adachi, I., Puopolo, K., Marquez-Sterling, N., Arai, H., and Forgac, M. (1990)
Arai, H., Terres, G., Pink, S., and Forgac, M. (1988) J. Biol. Chem. 263,
Feng, Y., and Forgac, M. (1992) J. B i d . Chem. 267,5817-5822 Manolson, M. F., Rea, P. A., and Poole, R. J. (1985) J. B i d . Chem. 260, 12273-
Zhang, J., Myers, M., and Forgac, M. (1992) J. Biol. Chem. 267, 9773-9778 Arai, H., Berne, M., and Forgac, M. (1987) J. B i d . Chem. 262, 11006-11011 Bowman, E. J., Siebers, A,, and Altendorf, K. (1988) Proc. Nutl. Acud. Sci.
Drose, S., Bindseil, K. U., Bowman, E. J., Siebers, A., Zeeck. A,, Altendorf, K.
Hanada. H., Moriyama, Y., Maeda, M., and Futai, M. (1990) Biochem. Biophy.
Schneider, E., and Altendorf, K. (1985) EMBO. J. 4, 515-518 Noumi, T., Beltran, C., Nelson, H., and Nelson, N. (1991) Proc. Natl. Acad. Sci.
Fillingame, R. H. (1992) J. Bioenerg. Biomembr. 24, 485-191 Wang, S.-Y., Moriyama, Y., Mandel, M., Hulmes, J. D., Pan, Y.-C., Danho, W.,
Perin, M. S., Fried, V. A., Stone, D. K., Xie, X . 6 , and Sudhof, T. C . (1991) J. Nelson, H., and Nelson, N. (1988) J. B i d . Chem. 263, 17638-17642
Manolson, M. F., Proteau, D., Preston, R. A., Stenbit, A,, Roberts, B. T., Hoyt, Biol. Chem. 266,3877-3881
M. A., Preuss, D., Mulholland, J., Botstein, D., and Jones, E. W. (1992) J. B i d . Chem. 267.14294-14303
Biochemistry 26,6632-6638
5649-5660
Chem. 264, 1560g15612
19236-19244
20032
960-966
J. B i d . Chem. 265,967-973
87968802
12279
U. S. A. 85, 7972-7976
(1993) Biochemistry 32,3902-3906
Res. Commun. 170,873-878
U. S. A. 88, 1938-1942
27. Sun, S.-Z., Xie, X.-S., and Stone, D. K. (1987) J. B i d . Chem. 262.14790-14793

















![Electro-catalytic synthesis of ammonia by surface proton ... · for catalyst support in terms of the ability of proton conduction [48-50]. Also, the property of not only but also](https://static.fdocuments.net/doc/165x107/5e8c0b7f5e76293fb049ef0c/electro-catalytic-synthesis-of-ammonia-by-surface-proton-for-catalyst-support.jpg)