
Journal of Experimental Psychology: Animal Behavior...
19
Journal of Experimental Psychology: Animal Behavior Processes \ VOL. 5, No. 2 APRIL 1979 Influence of Acoustic Experience on the Ontogeny of Frequency Generalization Gradients in the Chicken Lynne M. Kerr, E. Michael Ostapoff, and Edwin W Rubel Yale University The role of auditory experience in the ontogeny of perceptual coding was investigated in hatchling chickens. In Experiment 1, auditory frequency gen- eralization gradients were determined by using a habituation-generalization paradigm and an orienting response. One-day and 3-4-day chicks were habituated to 800-Hz tones and then tested at five frequencies ranging from 800 to 1,000 Hz. One-day chicks displayed reliably flatter generalization gradients -than 3-4-day chicks. In Experiment 2 an auditory deprivation method is described in which an ear impression compound was injected into the external ear. This method (a) is fully calibrated and provides approx- imately 40 dB of attenuation across a .125- to 4.0-kHz range, (b) appears to be fully reversible, and (c) is applicable prior to normal auditory experi- ence as well as at subsequent stages. In Experiment 3, this method was used to investigate the effects of auditory deprivation on the "perceptual sharpen- ing" described in Experiment 1. Chicks were deprived from Embryonic Day 184 to 3-4 days posthatch, at which time normal, deprived, and sham- operated chicks were tested as in Experiment 1. The generalization gradients of the deprived chicks were significantly flatter than, those of the 3-4 day normal and sham-operated chicks and were similar to those of 1-day chicks. This effect cannot be attributed to (a) operative procedures, (b) changes in auditory thresholds, or (c) changes in response rate with age or condi- tion. Thus the perceptual sharpening normally occurring between 1 and 3-4 days posthatch fails to occur during that time in the absence of a normal acoustic environment. According to one view, both perceptual thai, 1975), and fewer stimulus classifica- and cognitive development involve responses tions are made by younger children than to increasingly specific environmental stim- by older children (Bornstein, Kessen, & uli (Ganz, 1968; E. Gibson, 1969; Tees, Weiskopf, 1976; J. Gibson & Gibson, 1955). 1976; Werner, 1948). Such "perceptual Taken together, these and other results (see sharpening" is exemplified by a number of Rubel & Rosenthal, 1975; Tees, 1976) in- phenomena; for example, during normal dicate that sharpening of perceptual pro- ontogeny, species-typical behavioral re- cesses occurs during normal ontogeny, sponses are evoked by a decreasing number In addition, many experiments indicate of stimuli (Gottlieb, 1971; Hailman, 1967; that experience can influence perceptual Hinde, 1970; Messmer & Messmer, 1956). sharpening. Gottlieb (1976a) showed, for The generalization functions found in young example, that in simultaneous preference subjects are significantly flatter than those tests, normal ducklings select normal ma- found in older organisms (Rubel & Rosen- ternal calls over both low- and high-fre- Copyright 1979 by the American Psychological Association, Inc. 0097-7403/79/0502-0097$00.75 97
Transcript of Journal of Experimental Psychology: Animal Behavior...

Journal of Experimental Psychology:Animal Behavior Processes
\
VOL. 5, No. 2 APRIL 1979
Influence of Acoustic Experience on the Ontogenyof Frequency Generalization Gradients in the Chicken
Lynne M. Kerr, E. Michael Ostapoff, and Edwin W RubelYale University
The role of auditory experience in the ontogeny of perceptual coding wasinvestigated in hatchling chickens. In Experiment 1, auditory frequency gen-eralization gradients were determined by using a habituation-generalizationparadigm and an orienting response. One-day and 3-4-day chicks werehabituated to 800-Hz tones and then tested at five frequencies ranging from800 to 1,000 Hz. One-day chicks displayed reliably flatter generalizationgradients -than 3-4-day chicks. In Experiment 2 an auditory deprivationmethod is described in which an ear impression compound was injected intothe external ear. This method (a) is fully calibrated and provides approx-imately 40 dB of attenuation across a .125- to 4.0-kHz range, (b) appearsto be fully reversible, and (c) is applicable prior to normal auditory experi-ence as well as at subsequent stages. In Experiment 3, this method was usedto investigate the effects of auditory deprivation on the "perceptual sharpen-ing" described in Experiment 1. Chicks were deprived from Embryonic Day184 to 3-4 days posthatch, at which time normal, deprived, and sham-operated chicks were tested as in Experiment 1. The generalization gradientsof the deprived chicks were significantly flatter than, those of the 3-4 daynormal and sham-operated chicks and were similar to those of 1-day chicks.This effect cannot be attributed to (a) operative procedures, (b) changesin auditory thresholds, or (c) changes in response rate with age or condi-tion. Thus the perceptual sharpening normally occurring between 1 and3-4 days posthatch fails to occur during that time in the absence of anormal acoustic environment.
According to one view, both perceptual thai, 1975), and fewer stimulus classifica-and cognitive development involve responses tions are made by younger children thanto increasingly specific environmental stim- by older children (Bornstein, Kessen, &uli (Ganz, 1968; E. Gibson, 1969; Tees, Weiskopf, 1976; J. Gibson & Gibson, 1955).1976; Werner, 1948). Such "perceptual Taken together, these and other results (seesharpening" is exemplified by a number of Rubel & Rosenthal, 1975; Tees, 1976) in-phenomena; for example, during normal dicate that sharpening of perceptual pro-ontogeny, species-typical behavioral re- cesses occurs during normal ontogeny,sponses are evoked by a decreasing number In addition, many experiments indicateof stimuli (Gottlieb, 1971; Hailman, 1967; that experience can influence perceptualHinde, 1970; Messmer & Messmer, 1956). sharpening. Gottlieb (1976a) showed, forThe generalization functions found in young example, that in simultaneous preferencesubjects are significantly flatter than those tests, normal ducklings select normal ma-found in older organisms (Rubel & Rosen- ternal calls over both low- and high-fre-
Copyright 1979 by the American Psychological Association, Inc. 0097-7403/79/0502-0097$00.75
97

98 L. KERR, M. OSTAPOFF, AND E. RUBEL
quency attenuated calls. Ducklings that aredevocalized, acoustically deprived, and iso-lated, however, select high-frequency-attenu-ated calls as often as they do normal calls.These studies as well as other theoreticaland empirical contributions (e.g., Ganz,1968; Lashley & Wade, 1946) stronglysuggest that experience affects the degreeto which animals exhibit perceptual sharp-ening.
A question that remains unanswered inmost deprivation work to date, however, isthe role that experience plays in develop-ment per se, a criticism initially madeby Solomon and Lessac (1968). Gottlieb(1976b) formalized and extended this criti-cism by outlining the three possible roles ofexperience: maintenance, facilitation, and/or induction. By investigating the normalontogeny of a behavioral or neural process,it becomes possible to differentiate betweenthe maintenance role of experiential influ-ences and the latter two. Additional experi-ments involving prolonged deprivation andrecovery periods are necessary to ascertaininductive versus facilitative effects.
The present report describes three experi-ments. In the first experiment, inhibition ofdistress calling was used to investigate
We would like to thank Anne Kessen for help indata collection, Jim Cox for help with statisticalanalyses, and Morton Rosenthal for help with theinitial experiments. Oswald Steward, ThomasParks, Hunter Jackson, and Fred Engelhardt madehelpful comments on the manuscript. In addition,Gilbert Gottlieb provided many helpful commentsboth during the course of the experiments and onearlier manuscript drafts. This research was sup-ported by Grant BNS 76-03006 from the NationalScience Foundation and funds from the DeafnessResearch Foundation.
Lynne M. Kerr is now at the Physiology De-partment, University of Virginia.
E. Michael Ostapoff is now at Michigan StateUniversity.
Edwin W Rubel is now affiliated with the De-partments of Otolaryngology and Physiology atthe University of Virginia.
Requests for reprints should be sent to Edwin WRubel, Department of Otolaryngology, Universityof Virginia Medical Center, Box 430, Charlottes-ville, Virginia 22908.
habituation and generalization functions totonal stimuli of varying frequencies in 1-dayand 3-4-day hatchling chicks. In Experi-ment 2, we present the details and calibra-tion of our acoustic deprivation procedure.In Experiment 3, this deprivation methodwas used to assess the role of normal acous-tic experience on the developmental sharp-ening of frequency generalization gradients.
Experiment 1
When a hatchling chick is placed by itselfin a cool, well-lit chamber, it continuallyemits distress calls. Following presentationoi a novel stimulus of moderate intensity,the chick orients to the stimulus and ceasesdistress calling for some period of time.Several factors make this a useful behaviorfor investigations of avian neonates (e.g.,Hoffman, Schiff, Adams, & Searle, 1966;Rubel, 1970). First, the time period with-out distress calls, as well as the number ofcalls within any time segment, can be easilyand objectively recorded. Second, distresscalls are by far the most intense vocaliza-tions emitted by the chicks; they are stereo-typed in waveform, and they are emitted assingle notes of 100-200 msec duration withno less than 150 msec between notes. Fi-nally, neither the rate nor the topographyof the vocalizations changes dramaticallyover the first several days after hatching(Kaufman & Hinde, 1961). In this experi-ment, we sought, first, to determine whetherthe orienting response to tonal stimuli, asmeasured by inhibition of distress vocaliza-tions, was suitable for investigations of ha-bituation and generalization to acousticstimuli in hatchling chicks.
The second purpose of this experimentwas to determine if 1-day and 3-4-daychicks differed in habituation and/or gen-eralization of the orienting response to puretonal stimuli. To strictly relate changes inbehavior to an organism's perception of thesize of stimulus classes, it is necessary touse stimuli that differ only along one ormore quantifiable physical dimensions (seeRubel & Rosenthal, 1975). Thus, in theexperiments reported here, we investigated

ACOUSTIC EXPERIENCE AND STIMULUS GENERALIZATION 99
generalization gradients to auditory stimulithat differ only in frequency. Because thestimuli differ only along one quantitativedimension, the resulting generalization gra-dients are clear indicators of the relationbetween perceptual processes and that phys-ical dimension. In our earlier study (Rubel& Rosenthal, 1975), differences in generali-zation gradients were attributed to a per-ceptual sharpening process that results insmaller or more discrete functional stimulusclasses in older animals. In that study, gen-eralization of habituation was measured bychanges in the number of eye-opening re-sponses to auditory stimuli. In the presentstudy, changes in distress-calling behaviorwere used. It was felt that if similar re-sults occurred with a completely differ-ent behavioral measure, this interpretationwould be strengthened; if the generality ofthis ontogenetic event could be extendedacross response measures, the importanceof further investigations regarding the roleof early experience in development wouldalso be amplified.
In this experiment, 1-day and 3-4-daychicks were first given habituation trainingby repeated presentation of a pure-tonestimulus at 800 Hz. Immediately followinghabituation training, different groups ofchicks at each age were tested with stimuliof 800 Hz (the controls), 825, 850, 900, or1,000 Hz. It is important to note that pre-vious behavioral studies using eye-openingas the response measure (Rubel & Rosen-thai, 1975), pilot studies using distress callinhibition (Ostapoff & Rubel, Note 1),and electrophysiological studies (Saunders,Coles, & Gates, 1973; present Experiment2) all indicate that 1-day and 3-4-day-oldchicks do not differ in initial responsivenessor habituation to pure-tone stimuli in thisfrequency range.
Half of the chicks received most of thehabituation trials at a fixed interval, and theother half had the intertrial interval contin-gent upon vocalizations.1 During generaliza-tion testing, stimulus presentation was madecontingent on vocalization for all chicks.
Method
Subjects
Subjects were 100 White Leghorn chicks (Cal-lus domesticus) obtained as fertilized eggs froma commercial breeder, incubated in a forced-draftincubator at 37.5 °C (S5%-65% relative humidity)and turned four times daily. Hatchling chicks weretransferred to communal brooders (with at least12 chicks per brooder) where temperature wasmaintained at 32 ± 4 °C. Food and water wereavailable ad lib. Brooders were on a 12 :hr light/dark cycle, and experiments were run only duringthe light hours. Subjects were tested during Day 1(6-24 hr) or Days 3-4 (48-96 hr) posthatch.Each age-group consisted of SO subjects.
Materials
The experimental chamber was 22.22 X 20.32 X34.90 cm. One side was clear Plexiglas, and theother three were lined with lead-insulated acous-tical foam, painted white. Chicks stood 14.61 cmfrom the bottom of the chamber on .25-in. (.64-cm) hardware cloth. The top of the experimentalchamber, which fit tightly over the animal com-partment, held a 20.32-cm speaker covered by awire screen. The chamber, illuminated by diffuselight from a fluorescent lamp, was situated in asound-attenuated room (IAC 1200 series) whichwas maintained at an average temperature of21.1 °C. Background white noise was constantlymaintained at 55 dB (re 20 /»N/m2, B scale) mea-sured at chick ear level by a General Radio soundlevel meter (Type 1565C). A television camerasituated in front of and slightly above the chamberallowed the experimenter to continually monitorthe bird's behavior. A Sony Cardioid microphone(Type F-98) fit tightly into a hole in the experi-mental chamber 8.89 cm below the subject's stand-
1 Presentation of the acoustic stimulus duringhabituation trials was made contingent on thechick's vocalizations (contingent presentation) orindependent of the chick's behavior, at a fixedintertrial interval (noncontingent presentation).Several studies have suggested that the two modesof stimulus presentation produce different results(Carlton & Vogel, 1967; Lubow, Schnur, & Rifkin,1976; Lubow & Siebert, 1969). The latest of thesefound that rats contingently preexposed to a stim-ulus exhibited more learning than passively (non-contingently) preexposed rats. These results weretaken as indicating that when a subject is inactive control of a stimulus, that stimulus is at-tended to more than a stimulus passively receivedby the subject. If it is indeed possible to selectivelyvary the attentional component, then one mightexpect to find differences in habituation or general-ization (e.g., Fantz, 1965).

100 L. KERR, M. OSTAPOFF, AND E. RUBEL
ing level. This was connected to a Tektronix Type502 dual beam oscilloscope and audio monitorfrom which the experimenter monitored the chicks'calls.
Stimuli
Each stimulus consisted of a 1.6-sec train of fourpulses. Each of the four pulses was 250 msec induration, with a rise-fall time of 10 msec and aninterpulse interval of 200 msec. Stimuli weregenerated by a Wavetek function generator (Model134). All frequencies were presented at 90 dB(re 20 /uN/m"), or 35 dB above background level.Stimuli were calibrated in five places within theexperimental chamber by a General Radio electretcondenser microphone (Type 1560-p42) and by aGeneral Radio wave analyzer (Type 1900-A) atthe narrow bandwidth setting. No frequency usedin this experiment varied more than 3 dB overthe sound field.
Procedure
Subjects were removed individually from theircommunal brooders and placed in the experimentalchamber where each subject was given 5 min toadapt to the chamber. Vocalizations from thechick were amplified, displayed on an oscillo-scope, and passed to a voltage discriminator setto trigger only on distress calls. As discussed inthe Introduction, when chicks are placed in theexperimental chamber they emit distress calls. Itshould be noted that the distress calls tend to beemitted in bursts lasting from 5 to more than 60sec, with interburst intervals varying from a fewseconds to several minutes. Upon presentation ofthe stimulus, the chick typically stops distress call-ing and orients to the stimulus. The principal re-sponse measure was the length of time of absenceof distress calling, recorded on a Tektronix dualbeam storage oscilloscope (Type 5103N, D13) andmeasured up to 8 sec following the end of stimuluspresentation. This measure is referred to as La-tency. A second measure was the number of callsfollowing presentation of the stimulus, recordedduring the same 8-sec interval (referred to asNo. Calls).
Habituation
All subjects were given habituation training toan 800-Hz stimulus. Two habituation procedureswere followed. Half of the subjects in each agegroup (25 subjects in each one) received a con-tingent (or self-initiated) habituation paradigm,and half received a fixed-interval schedule. Thecontingent paradigm was used for generalizationtesting of all subjects.
Contingent condition. In this paradigm, a mini-mum intertrial interval (ITI) of 30 sec was im-
posed following each trial. The first distress callfollowing each ITI automatically triggered a 700-msec delay and then stimulus presentation. Habitu-ation training consisted of 15 trials of which 12included presentation of the 800-Hz stimulus and3 were "mock" (nonstimulus) trials. These mocktrials were included to give the experimenter abaseline of distress call activity and were alwayspresented at trial Positions 5, 10, and 12.
Noncontingent condition. In the noncontingentcondition, two contingent trials were initially pre-sented to ensure that the subject was orienting tothe stimulus. These were followed by 11 trials ata fixed intertrial interval of 30 sec which includedthe three mock trials, occurring in the same posi-tions (5, 10, and 12) as in the contingent condi-tion. The last two trials were again presentedunder the contingent condition in order to deter-mine whether the chicks in the two conditions hadshown similar response decrement. Thus, thesechicks differed from those in the contingent condi-tion during Trials 3-13 in that they received sevenhabituation and three mock trials on a fixed-inter-val schedule of 30 sec, not contingent upon theirown distress-calling behavior. Since distress callstend to come in long bursts and are followed byrelatively long periods of silence, inhibition of dis-tress calls during the noncontingent condition can-not be accurately measured; the stimulus might bepresented during one of the silent periods. There-fore, Latency and No. Calls were not recordedfor the trials presented on a fixed-interval schedule.
Two constraints were placed on the chicks inthe two conditions. If a chick failed to orient tothe stimulus during one of the first two trials, itwas discarded from the study. Orientation wasarbitrarily defined as a minimum 5-sec latencyprior to resumption of distress calling. Second, ifa chick failed to make any distress calls within any5-min period of testing, it was also eliminated fromthe study. A total of 17 1-day and 19 3-4-daychicks were eliminated by these criteria and werereplaced by new chicks.
Generalisation Testing
The generalization testing procedure, identicalfor all animals, was run under the contingent con-dition. Generalization testing began 30 sec after thelast habituation trial. Each chick was presentedwith five successive generalization test trials, withthe stimulus at the same frequency on each trial.As in contingent habituation, 30 sec after the pre-vious trial, the next large distress call triggeredthe 700-msec delay and then stimulus presentation.A total of 10 subjects from each age-group weretested with the stimulus at 800 (habituation fre-quency), 825, 850, 900, and 1,000 Hz. Of these 10subjects in each age by frequency group, 5 had re-ceived habituation training under the contingentcondition and 5 under the noncontingent condition.

ACOUSTIC EXPERIENCE AND STIMULUS GENERALIZATION 101
Results and Discussion
Habituation Training
Contingent condition. Results of habitu-ation training for the two age groups asmeasured by latency to resume calling andby the number of calls within 8 sec areshown in Figures 1 and 2, respectively(connected symbols). The trials on the leftare the 12 habituation trials; the mean re-sponses during the mock trials, which wereactually interspersed with the habituationtrials (see Method), are presented on theright. Inspection of Figure 1 reveals amarked decrease in Latency over trialswhich was similar for the two age groups,indicating that habituation training was suc-cessful and comparable in the two agegroups. A two-way analysis of variance(Age X Trials), performed to confirmthese observations, yielded a significantmain effect for trials, F(ll, 528) =68.46,p < .01, whereas neither the difference be-tween age groups nor the Age X Trialsinteraction approached statistical signifi-cance, F(l, 48) = 1.58 and F(ll, 528) =1.46, respectively.
Inspection of Figure 2 reveals a marked
5 5zuI- 4
3 4 da
4 6 8
T R I A L10 mock
Figure 1. Mean Latency responses to the 12 stim-ulus-presentation trials during habituation trainingfor 1-day (dashed line) and 3-4-day (solid line)chicks. (The group means of the three mock trials,which were actually interspersed with the stimulus-presentation trials are presented on the right, opencircles for 1-day and filled circles for 3-4-daychicks. The unconnected symbols above Trials 1,2, 11, and 12 represent the results from the 1-day[open squares] and 3-4-day [filled squares] chicksin the noncontingent condition.)
14-
12-U)-I 10-
o 8-
2 4 H£
z-
,~. >f? 5
•3-4 day
6 8T R I A L
10 12 mock
Figure 2. Mean No. Calls during the 12 stimulus-presentation trials of habituation training for 1-day(circles and dashed lines) and 3-4-day (squaresand solid lines) chicks. (The group means of thethree mock trials, which were actually interspersedwith the 12 stimulus presentation trials, are pre-sented on the right side of the figure, circles for1-day and squares for 3-4-day chicks. The un-connected symbols above Trials 1, 2, 11, and 12represent the results from the 1-day [open circles]and 3-4-day [open squares] chicks in the non-contingent condition.)
increase in No. Calls over habituation forthe two age groups. As with the Latencymeasure, the 12 habituation trials are onthe left, and the mean responses to mocktrials are presented on the right. An Age XTrials analysis of variance yielded a signifi-cant main effect for trials, F(ll, 528) =46.27, p < .01, whereas neither the differ-ence between age groups nor the Age XTrials interaction approached statistical re-liability, F(l, 48) = .67 and F(ll, 528) =1.48, respectively. As the No. Calls responsemeasure was highly correlated with Latency(M 1-day r = -.75; M 3-4-day r = -.81),these results are hereinafter not describedin detail.
Examination of the mock trials, the meanresponse to which is presented on the right-hand side of Figures 1 and 2, shows that1-day and 3-4-day chicks were also similarin their baseline level of distress calling, La-tency, F(l, 48) - 1.72. The lack of a sta-tistically reliable difference between agegroups on both habituation training andmock trials suggests that the two age groupswere similar in both habituation and overallresponse rate.

102 L. KERR, M. OSTAPOFF, AND E. RUBEL
The latency during the mock trials ap-pears to be lower than the last three trialsof habituation training (Figure 1). This in-dicates that habituation never reached anasymptote equivalent to the baseline levelof distress-call activity for either of the agegroups. An analysis of variance on TrialType (mock trials vs. final three stimuluspresentation) X Age was performed to con-firm this observation. This yielded a signifi-cant within-subject trial type effect, F(l,96) = 8.59, p < .01, for the mock trialsversus the last three habituation trials. Thecomparison between age groups did not ap-proach statistical significance, F(l, 96) =.814, which was also true for the Trial TypeX Age interaction, F(\, 96) = .05.
Noncontingent condition. In Figures 1and 2, the unconnected symbols over Trials1, 2, 11, and 12, which were presented "con-tingently," represent the results for the non-contingent subjects in each age group. In-spection of these points reveals that thecontingent and noncontingent habituationparadigms yielded very similar results. ForLatency, a Condition X Age X Trials anal-ysis of variance on the first two and the lasttwo trials did not approach statistical relia-bility for condition, F(l, 96) = .12, agegroup, F(l, 96) = 1.88, the Age X Condi-tion interaction, F(l, 96) = .77, the Condi-tion X Trials interaction, F(3, 288) = .63,the Age X Trials interaction, F(3, 288) =.31, or the Age X Trials X Condition in-teraction, F(3, 288) = .76. The trials effectwas statistically significant, F(3, 288) =427.47, p < .01. This same pattern was ob-served in the Age X Condition X Trialsanalysis of variance for the No. Calls mea-sure. The age, F(l, 96) = 1.25, and thecondition, F(l, 96) =.09, effects as wellas the Age X Condition, F(l, 96) = .19,the Age X Trials, F(3, 288) = .09, theCondition X Trials, F(3, 288) = .80, andthe Age X Condition X Trials, F(3, 288)= 1.13, interactions were nonsignificant.The trials effect was significant, F(3, 288)= 258.41, p < .01. Thus, there was no dif-ference between chicks in the contingent andnoncontingent conditions despite the factthat the mean intertrial interval for chicks in
the contingent condition was longer than itwas for those in the noncontingent condition.2
Generalisation Testing
The generalization testing results for thetwo habituation-condition groups are notpresented separately since inspection of thedata revealed no systematic differences anda Condition X Age X Frequency X Trialsanalysis of variance yielded no reliable dif-ferences between chicks in the two condi-tions.
The mean Latencies for chicks in thetwo age-groups over the five generalizationtrials at each frequency are shown in Figure3. Inspection of this figure reveals that the1-day chicks displayed a markedly flattergeneralization gradient than the 3-4-daychicks. An Age X Frequency X Trials anal-ysis of variance was performed to confirmthese observations. This analysis yielded asignificant main effect for age, F(l, 80) =5.88, p < .05, frequency, F(4, 80) = 8.55,p < .01, and trials, F(4, 320) = 3.78, p <.01. The Age X Frequency interaction didnot yield statistical significance at the .05level, F(4, 80) = 2.25, .05 < p < .10; how-ever, the Frequency X Trials interactionwas significant, F(16, 320) = 2.27, p < .01.The Age X Trials interaction and the AgeX Frequency X Trials interaction did notapproach statistical significance, F(4, 320)= .61 and F(16, 320) = .86, respectively.
2 As discussed above, the intertrial interval forthe contingent condition was a variable (30 secminimum) intertrial interval as opposed to thefixed 30-sec interval for the noncontingent condi-tion. In another experiment performed in thesame manner (Kerr & Rubel, Note 2), the meanintertrial interval for 3- to 4-day chicks in thecontingent condition was initially 80 sec (ap-proximately) and highly variable. By approxi-mately the fifth trial of habituation, the inter-trial intervals became much less variable and ap-proached the experimentally imposed 30-sec delayof the noncontingent condition. Assuming that thechicks in the present experiment behaved similarly,it is interesting that in spite of a slightly longerand highly variable intertrial interval, we foundno differences between chicks in the two condi-tions, at either the end of habituation training orin their generalization gradients.

ACOUSTIC EXPERIENCE AND STIMULUS GENERALIZATION 103
These results indicate that the age groupswere reliably different in their responses tothe test stimuli, and chicks in each agegroup displayed reliable generalization func-tions. In addition, there was a reliable re-sponse decrement over the five generaliza-tion test trials. The failure to attain thetraditional .05 level of significance on theAge X Frequency interaction was probablydue to the fact that the 1-day and the 3-4-day chicks responded similarly to 1,000 Hz.The analysis of variance interaction termtests whether the two linear trends areparallel; the absence of a significant inter-action is understandable since the two func-tions begin and end at the same point anddepart from one another only in the middle(see below).
Since it is important to determine thefrequencies at which each age-group showsa significant generalization decrement andthe frequencies at which the age groups dif-fer in response to the test stimuli, the New-man-Keuls method for individual compari-sons (Winer, 1962), was employed. Thefirst set of analyses investigated the sourceot the reliable differences along the fre-quency dimension by comparing responsesto the control frequency (800 Hz) with theresponses to other frequencies within eachage group. The results can be summarizedas follows. All significant qs reported arereliable at the .01 level. Three-four-daychicks showed a significantly longer La-tency at 850 Hz (q = 3.97), 900 Hz (q =5.77), and 1,000 Hz (q = 5.64), whereasthe 1-day chicks did not show a reliable in-crease in Latency until 1,000 Hz (q = 3.89).These results, considered together, indicatethat the younger chicks displayed flattergeneralization gradients in the frequencyrange near the control stimulus (habitua-tion-training stimulus, 800 Hz).
The Newman-Keuls method was nextused to investigate the locus of the age dif-ferences. The 1-day chicks had a signifi-cantly lower Latency than 3-4-day chicksat 850 and 900 Hz (q = 2.75 and q = 4.51,respectively), which also confirms the ob-servation that 1-day chicks displayed flat-ter generalization gradients than 3-4-day
oz
<01
3-4 Day
I Day
800 825 850 900
F R E Q U E N C Y (Hz)
1000
Figure 3. Generalization gradients for meanLatency responses of 1-day (circles and dashedlines) and 3-4-day (squares and solid lines) chicks.
chicks. There were no significant differ-ences between age groups at 800, 825, or1,000 Hz.
The responses of the chicks on the firstgeneralization trial alone are the purest in-dicator of stimulus generalization; there-fore, an Age X Frequency analysis of vari-ance was also performed on the Trial 1 dataalone. This analysis yielded a significantmain effect for age, F(l, 80) = 6.25, p <.01, and frequency, F(4, 80) = 10.80, p <.01, and, again, the Age X Frequency inter-action was not significant, F(4, 80) = 1.51.Individual comparisons were again per-formed and produced results similar to thosedescribed above. (The pattern of results re-ported above for the Latency measure wasduplicated in the results for the No. Callsmeasure, which is not discussed.)
In summary, chicks in both the 1-dayand 3-4-day groups displayed similar ha-bituation functions, responses to mock trials,and responses to generalization test fre-quencies of 800, 825, and 1,000 Hz. Thus,the age groups do not appear to differ inauditory responsiveness in general, in theirbasal rate of distress calling, or in acquisi-tion of habituation. Taken together, thesefactors indicate that the distress-call-inhibi-tion procedure is suitable for investigatinghabituation and generalization in hatchlingchicks.
Using the distress-call-inhibition proce-dure as our response measure, we havefound that 1-day chicks display flatter gen-eralization gradients than 3-4-day chicks

104 L. KERR, M. OSTAPOFF, AND E. RUBEL
when tested with acoustic stimuli differingalong the frequency dimension. This findingcannot be ascribed to differences betweenthe age groups in overall response rate, asthe two groups were similar, as has beennoted, in their initial response rate, final ha-bituation level, response rate during mocktrials, and their responses to 800, 825, and1,000 Hz during generalization testing. An-other possible explanation for our resultsis that auditory sensitivity to higher fre-quencies improves with age and that the re-sponse of the 3- to 4-day group at 1000 Hzwas artificially truncated by a ceiling effecton the Latency measure. This explanationis made unlikely by the fact that both groupsshow much longer Latencies at the begin-ning of habituation than during testing with1,000 Hz. Therefore, the finding that thetwo age groups respond similarly to 800,825, and 1,000 Hz but differ in their re-sponses to 850 and 900 Hz can best be at-tributed to a tendency for the youngerchicks to generalize over a broader fre-quency range.
These results parallel those of Rubel andRosenthal (1975) despite the methodologi-cal changes and the use of an inhibitoryrather than an excitatory response. In bothstudies, 1-day chicks displayed significantlyflatter generalization gradients than 3- to 4-day chicks. Thus, over the first 3-4 daysposthatch, chicks become differentially re-sponsive to smaller stimulus changes.
Experiment 2
An increasing number of investigatorshave begun to assess the importance of earlyacoustic experience on the ontogeny of be-havioral and physiological function (e.g., seeClements & Kelly, 1978; Clopton & Silver-man, 1977; Gottlieb, 1976a; Silverman &Clopton, 1977; Tees, 1976). Several meth-ods have been used to deprive animals ofacoustic stimulation. With the exception ofGottlieb's devocalization and isolation pro-cedure, previous methods do not provideeither sufficient attenuation (Clements &Kelly, 1978) or sufficient documentation toallow complete specification of an organism'sauditory environment. However, Gottlieb
and Vandenbergh's (1968) devocalizationprocedure for ducklings has not proven fea-sible in our laboratory or others (Gottlieb,Note 3) because the location and arrange-ment of the syringeal membranes is differentin chicks and ducklings.
The purpose of this experiment, then, wasto develop and calibrate a method of audi-tory deprivation that would (a) provideadequate attenuation of acoustic stimulithroughout the audible range of the do-mestic chick, (b) be applicable prior tonormal auditory experience as well as atlater stages of development, (c) be reversi-ble, thereby making later behavioral andphysiological testing possible, and (d) beamenable to calibration across the audiblefrequency range in order to specify andcontrol the degree of conductive hearing im-pairment at any frequency.
Method
Subjects and Treatments
Subjects were 19 1- to 10-day White Leghornchicks. Fertilized eggs were obtained from a com-mercial breeder, incubated in a forced-draft incu-bator at 37.5 °C, and turned four times daily until18 days of incubation. Chicks were divided intothree age groups and two treatment conditions ateach age. The age groups, 1 day, 3-4 day, and9-10 day, refer to the posthatch age at whichevoked potentials were recorded from the brainstem in order to derive Frequency X Thresholdfunctions. From these functions, the effectivenessof the ear plugs was determined. The two treat-ment conditions within each age group, Acute andChronic, refer to the time at which ear plugs wereapplied. There were three chicks in each age bytreatment group except the chronic 3- to 4-daygroup, which contained four chicks.
Chicks in the chronic condition had ear plugsinserted into one ear prior to hatching. At ap-proximately 18.5 days of incubation (436-456 hrof incubation), the egg was removed from theincubator, and the blunt end of the shell coveringthe air space was removed. This time is at, orjust prior to, the "tenting stage" (Oppenheim,1973), that is, before entry of the bill into theair space. Embryos whose bill had already en-tered the air space were not used in this study.The inner shell membrane was swabbed with warmsaline to visualize major blood vessels and thencut, avoiding major vascular damage. The em-bryo's head was gently pulled out of the shell, andthe area around one ear was dried with cottonswabs. A commercial, silicon-base ear impres-

ACOUSTIC EXPERIENCE AND STIMULUS GENERALIZATION 105
sion compound (Tru-mold, Scientific Plastics,New York, New York) was then injected intothe external ear with a blunted 20-ga. hypodermicneedle. The compound was allowed to set for 3-5min, after which the embryo in the remainingshell was placed on a wet paper towel in thehatching incubator and allowed to hatch withother deprived and normal chicks. Opening theshell and membranes expedites hatching by 6-18hr. However, all ages reported here and in Ex-periment 3 are calculated from the age at whichnormal chicks hatch. Hatchling chicks, with theear plug in place, remained in the incubator atlowered humidity for 18-24 hr and then weretransferred to a communal brooder until used forelectrophysiological recording.
Chicks in the acute condition were allowed tohatch normally and then were transferred to abrooder until used for electrophysiological re-cording. Ear plugs were inserted during the re-cording session (see Threshold Determinationbelow) by simply injecting the Tru-mold com-pound into the external ear of the hatchling chick.
Surgical and Recording Procedures
At 1 day (18-24 hr), 3-4 days, or 9-10 days ofage, the chicks were anesthetized and ThresholdX Frequency functions were determined on thebasis of brain stem evoked potential averages,with and without the ear plugs in place. Theevoked potentials used for threshold determina-tion were recorded with a bipolar electrode posi-tioned at the surface of the medulla in the regionwhere the cochlear nerve fibers course around thecerebellar peduncle (Rubel & Parks, 1975), Thisposition was attained by remotely lowering theelectrode through the cerebellum while presentingclick stimuli of moderate intensity to the ipsilateralear. As the electrode approached and finally con-tacted the medullary surface, the evoked potentialgrew in amplitude and "unit-cluster" responsescould be seen on the oscilloscope. At the point ofmaximum peak-to-peak evoked potential amplitude,the electrode was stopped, and its depth, and thelatency, amplitude, and polarity of the evokedpotential, were recorded. In addition, the "bestfrequency" of the "unit-cluster" activity was de-termined with superthreshold tone bursts in orderto ensure that it was within the 800-3,000 Hzrange, which signifies that the electrode positionwas well within the boundaries of the responsivearea (Rubel & Parks, 1975). This procedure ne-cessitated two, or at most three, electrode penetra-tions in order to find the adequate position, andhistological procedures confirmed that it resultedin a relatively standard placement around the mid-dle of the ascending branch of the cochlear nerve,prior to its entrance into n. magnocellularis. Theevoked potentials were probably of primarily pre-synaptic, eighth-nerve origin, although some con-
tribution of n. magnocellularis, n. angularis, or n.laminaris postsynaptic activity cannot be ruled out.
The following is a more detailed description ofthe general procedure outlined above. Chicks wereanesthetized with intraperitoneal injections of .0015ml/gm body weight of Equi-Thesin (Jensen-Salsbery) potentiated with intramuscular injec-tions of .008 ml/gm body weight of Vetalar (Parke-Davis). Also, .005 mg of atropine was injectedto reduce trachea! secretions. Nociceptive reflexeswere monitored during the surgical procedure andbetween stimulus runs; subsequent doses (twothirds or three fourths of the original dose) ofVetalar were administered as needed. Featherswere clipped from the head area, including theear-flaps. Following a midline incision, skin over-lying the posterior cranium was retracted. Dentalacrylic, attached to the beak, was used to securethe bird's head to a specially designed headholder.Neck muscles were retracted, and the bone wasremoved to expose the cerebellar cortex. In onechick, the middorsal sinus was ligated in twoplaces between the anterior cerebral vein and themiddle cerebral vein. The sinus was then cut, andthe entire cerebellum was aspirated to expose thefloor of the fourth ventricle. Cochlear nerve fiberscoursing rostromedially over the medulla and alarge blood vessel which lies above the rostro-medial portion of n. magnocellularis served asvisual landmarks for electrode placement. In allsubsequent experiments, the electrode was intro-duced through the intact cerebellum at an ap-propriate position. This procedure resulted in lesspulsation and, therefore, superior recording prop-erties.
The animal was positioned under the micro-drive in the sound-attenuated room, and with theaid of a Zeiss operating microscope, the electrodewas positioned over the cerebellum at an anglesuch that the electrode tip could be driven downinto the area where the eighth nerve courses overthe dorsal surface of the medulla. The electrodeswere of the concentric bipolar stainless steel type,with approximately 200 /um of exposed tip ending .2mm below the concentric shaft which was of 26-ga. hypodermic tubing. Approximately 200 /*m ofthe tubing was uninsulated, and the remainder wascoated three times with Epoxylite and tested. Po-tentials were recorded differentially, with a stain-less steel wire inserted in the exposed neck musclesserving as ground. Potentials were amplified, fil-tered, and displayed on one oscilloscope after fil-tering to pass 1-3,000 Hz. Signals were also passedto a Fabritek (1052 LS) signal averager. A secondoscilloscope was used to display multiunit activityby filtering potentials to pass .1-15 kHz. Stimuluspresentation was monitored on a second channelof each oscilloscope. The oscilloscope traces andFabritek accumulation were triggered 10 msecprior to stimulus onset. Output of the averager waspermanently recorded on a Beckman 10-in. (25.4-cm) strip chart recorder.

106 L. KERR, M. OSTAPOFF, AND E. RUBEL
Stimulus Calibration
The stimuli used for determining brain stem po-tential thresholds with and without the ear plugin place were IS msec (or 25 msec in the case offive chicks) pure-tone bursts with S-msec rise andfall times and a 2/sec rate of presentation (.5-secISI). Sine waves from a Wavetek (Model 134)function generator were led through a Wisconsin-type (Ludwig) electronic switch, attenuated(Hewlett-Packard Model 350), impedance matchedto the earphones, and delivered through earphoneassemblies sealed to each external auditory meatusto form a "closed system." Sound frequencies weredetermined on-line with a General Radio digitalperiod-frequency meter. The earphone assembliesconsisted of Briiel and Kjaer (B & K) probe kitearphones (B & K Model UA0040) sealed to 4-cm brass adapters which could be accurately posi-tioned against the meatus and sealed with either Au-dalin hearing-aid sealer or Vaseline. Each earphoneassembly was calibrated in a "closed system"over the range from .1 to 8 kHz with a B & K.25-in. condenser microphone and a B & K fre-quency analyzer (Model 2107). All harmonicswere at least 50 dB attenuated.
Threshold Determination
Evoked potential thresholds were determinedfor 10 frequencies which span the auditory rangeof the domestic chicken and were presented in arandom order. The frequencies were .125, 250, .500,.750, 1.0, 1.5, 2.0, 3.0, 4.0, and 5.0 kHz. To deter-mine the threshold for a given frequency, we firstpresented 128 repetitions of the tone burst at sub-threshold level and then at 5- or 10-dB incrementsin intensity until a threshold response was ob-tained (to within 5 dB). The attenuator reading atthreshold was then converted to sound pressurelevel (re 20 /iN/m2). Threshold was defined as apeak-to-peak amplitude of the averaged evokedpotential at least twice the amplitude of theaveraged variations observed in the 10 msec priorto stimulus onset. Threshold determination forall 10 frequencies constituted a single "stimulusrun" and took 40-60 min to complete. As notedbelow, at least two stimulus runs were completedfor each chick, one for determining the thresholdswith the ear plug in place and one for evaluatingthresholds without the ear plug.
Chronic condition. Chicks in the Chronic condi-tion had an ear plug inserted in one ear prior tohatching. The first stimulus run for these chickswas completed with the original ear plug in place(Plugged). The electrode was then withdrawnfrom the brain, and the ear plug was removedwith a forceps. The electrode was then repositionedin the same place, and a second stimulus run wasconducted (Open). Successive stimulus runs werebegun immediately after changing the ear plugcondition and replacing the electrode (5-15 min).The 9-10-day chronic chicks also had an additional
Open stimulus run to the ear that had beenplugged. This run was delayed 30-45 min after theend of the first Open testing.
Acute condition. Chicks in this condition hadnormal auditory experience (no ear plug) through-out development. Thus, these chicks were firsttested for thresholds with the ear in the normal(or Open) condition, and then another stimulusrun was conducted with the electrode in the sameposition but after the ear plug was in place(Plugged). The electrode was, as above, with-drawn between stimulus runs and then was care-fully replaced in the same region in an attempt torecord from a similar position in the nerve onall stimulus runs for each ear. It should be notedthat for chicks in the Acute condition, it waspossible to determine for both ears the conductivehearing loss at each frequency caused by the earplug. Thus, in most of the subjects, both earswere tested, and the results were treated as in-dependent samples. In the 1-day group, one animalhad both ears tested, which resulted in an w = 4,whereas in the 3-4- and 9-10-day groups both earswere tested in each chick (ns = 6).
Results and Discussion
Figure 4 shows the mean thresholds (dB,re 20 /«N/m2) across frequencies for theOpen and Plugged stimulus runs of eachage by condition group. In addition to show-ing that the ear plug resulted in a markedelevation of averaged evoked potentialthresholds at all frequencies tested, thesedata indicate that the attenuation was es-sentially constant regardless of whether theplug was inserted just prior to testing(Acute condition) or up to 12 days prior totesting (Chronic condition).3 Furthermore,inspection of the figure reveals that therewas little elevation of the thresholds as afunction of the length of time the plug wasin place. That is, comparing the Open con-ditions in the acute and chronic groupsyielded no major differences at any age,which suggests that the ear plug did notcause damage to the middle ear, cochlea,or ganglionic structures at least up to 10days posthatching. The slight elevation of
3 Because of the small sample sizes and unequalns, statistical comparisons across the treatmentconditions or age-groups were not consideredmeaningful in this experiment. Thus, conclusionsare drawn only regarding the major variables, andthese conclusions can be verified by examination ofthe figures.

ACOUSTIC EXPERIENCE AND STIMULUS GENERALIZATION 107
thresholds seen in the 9- to 10-day chronicchicks at frequencies above 1.0 kHz is sug-gestive of an effect of deprivation on thedevelopment of the high-frequency respon-siveness. However, two other equally viablealternatives exist. First, slight differences inelectrode position could account for a shiftin threshold functions toward the higherfrequencies. Second, the ear plug couldcause a shift in high-frequency thresholdsthrough temporary influences on peripheraltransmission. If, immediately after removalof the ear plug, there is a slight mass load-ing of the middle ear or tympanic membranethrough liquid residues or a lag in equilibra-tion, such a shift might be expected. In thiscontext it is worth noting that the second(delayed) stimulus run with the ear plugremoved (Open) on the 9- to 10-day chronicchicks (see Threshold Determination above)failed to reveal even a slight difference be-tween the acute and chronic Open thresh-olds.
Figure 5 shows the mean attenuation (±1 SD) due to the ear plug across all sub-jects. The mean hearing loss produced bythe ear plug across all frequencies was 40.3dB (re 20 /tN/m2). This level was quiteconstant from .125 through 3.0 kHz, with aslight drop at 4.0 kHz and a sharp declineat 5.0 kHz. The decline in attenuation at4.0 and 5.0 kHz is probably a result of thelimitations of the sound-presentation system.The maximum sound levels available at4.0 and 5.0 kHz were 116 and 112 dB (re 20juN/m2), respectively, which were insuffi-cient to elicit a threshold response in thePlugged condition in any of the experiments.In these cases, to be conservative, thesemaximum intensities were used as thethresholds for the Plugged threshold func-tion. Therefore, the values given as attenua-tion at 4.0 and 5.0 kHz represent only alower limit and not necessarily a drop in theattenuation of the plug at these frequencies.
In summary, we have described a rela-tively simple method of inducing a knownamount of auditory deprivation which canbe applied before normal auditory experi-ence begins. The method is relatively effec-tive in elevating auditory thresholds and
120
100
80
60
40-
20-
-s Q'
CL 120CO
CQ I0°•O-* 80
3 60:oI 40CO
0-120
100-
80-
60
40-
20
I Day
3-4 Day
9-10 Day
.125 .25 .5 751 1.52 3 4 5
FREQUENCY (kHz)
Figure 4. Audiograms of brain stem evoked po-tential thresholds (dB, re 20 juN/m") for 1-, 3-4-,and 9-10-day hatchling chicks. (Points are meanthresholds at each frequency for all ears tested.Triangles refer to acute treatment groups andcircles refer to chronic treatment groups. Openpoints refer to stimulus runs without ear plugs[Open runs], and closed points refer to Pluggedstimulus runs for both treatment groups. Dottedlines connect frequencies at which one or morechicks failed to achieve a threshold response at theloudest sound level presented in the Plugged con-dition.)
appears to be completely reversible to 9-10days posthatch. Furthermore, by "biolog-ically calibrating" the method on the basisof brain stem averaged evoked potentialthresholds, it becomes possible to selec-

108 L. KERR, M. OSTAPOFF, AND E. RUBEL
ID 20Zuh- 10
-....•{-H
.25 .5 .75 i 1.5 2 3 4 5
FREQUENCY (kHz)
Figure 5. Mean attenuation (±1 OT) of brainstem evoked potential thresholds as a result of theear plugs, collapsed across all ages and treatments.(At each frequency, the Open threshold level wassubtracted from the corresponding Plugged thresh-old for that subject. Means of these differencescores were computed for each frequency acrossall ages and treatments to arrive at an overallmeasure of attenuation due to the ear plug. Notethat the attenuation of 4.0 and 5.0 kHz representsonly minimum values for attenuation because, ina number of experiments, no threshold responsewas recorded at those frequencies during thePlugged stimulus runs.)
tively expose animals to known amountsof exogenous acoustic stimulation in anydesired frequency range. Such procedureswill become important for evaluating thespecificity with which early acoustic experi-ences influence the behavioral, physiological,and morphological ontogeny of the auditorysystem (see Rubel, 1978).
Experiment 3
As discussed above, auditory and visualdeprivation studies have demonstrated thatdeprived animals do not perform as well asnormally reared animals on many per-ceptual tasks. Lashley and Wade (1946),and later Ganz (1968), asserted that sub-jects deprived along a single stimulus di-mension often display shallow generaliza-tion gradients along that dimension. Thepurpose of Experiment 3 was to investigatethe effects of auditory deprivation on thedevelopmental change in frequency general-ization gradients seen in Experiment 1.Chicks were bilaterally deprived by themethod described in Experiment 2. Sham-operated chicks, subjected to the entireoperation procedure except for the actualinjection of ear plugs, served as a controlgroup, along with normal chicks that were
allowed to hatch undisturbed. After hatch-ing, chicks were housed communally for3-4 days, at which time ear plugs were re-moved and the chicks were tested withthe habituation-generalization paradigm de-scribed in Experiment 1.
Method
Treatments
Subjects were ISO White Leghorn chicks.Fertilized eggs were obtained from a commercialbreeder, incubated in a forced-draft incubatormaintained at 37.S °C (relative humidity 5S%-65%),and turned four times daily until 18 days of in-cubation. Chicks were divided into three treat-ment conditions of SO birds each (normal, sham,and deprived). All chicks were tested at 3-4 days(48-96 hr) posthatch.
Chicks in the deprived condition had ear plugsinserted bilaterally on approximately EmbryonicDay 18i (441-447 hr after the start of incubation),during the tenting stage. Ear plugging of embryoswas accomplished by the procedures described inExperiment 2. (If upon opening the shell it wasdiscovered that the chick's beak had alreadybroken through the chorio-allantoic membrane,the chick was discarded from the study.) Tru-moldimpression compound was injected into both outerears. The chicks were then placed in groups of10-20 in a hatching incubator maintained at 37.7±.S °C (relative humidity 65%-75%). The hatch-ing incubator was situated in a quiet isolatedroom, and chicks were allowed to hatch withoutbeing disturbed in any way. Hatchling chicks werethen transferred to a communal brooder in thesame isolated room, and since they were housedwith the sham chicks, there were always at least12 birds in the brooder. Brooder temperature wasmaintained at 32 ±4 °C. Chicks in the sham con-dition underwent the same manipulations as thosein the deprived condition except for the actual in-jection of Tru-mold ear plugs. Chicks in the nor-mal condition were transferred to a hatching in-cubator at Day 18 of incubation, and upon hatch-ing, they were transferred to communal brooders(with at least 12 chicks in each brooder), alsomaintained at 32 ± 4 °C.
Weights and Survival Rate
Normal chicks were, on the average, heavierthan chicks in either the sham or the deprivedcondition. The mean weights and standard errorswere as follows: normal, M~42.11 g, SE = .81;sham, M = 35.18 g, SB =. 54; and deprived, M= 34.02 g, SE = A5. Chicks in the deprived andsham conditions were not significantly different,*(94) = 1.66. Normal chicks differed reliably fromsham-operated, <(88) =7.96, p < .01, and deprived,

ACOUSTIC EXPERIENCE AND STIMULUS GENERALIZATION 109
t(88)-9.96, /><.01, chicks. Chicks in the shamand deprived conditions had approximately thesame survival rate: 78.8% and 79.5%, respectively.
MaterialsThe experimental chamber was a clear Plexiglas
cylinder 20.32 cm in diameter and 27.94 cm high.Approximately three quarters of the chamber cir-cumference was lined with translucent plastic.Chicks stood 13.97 cm from the bottom on .25-in.hardware cloth. The top of the chamber, holding a20.32-cm speaker covered by a wire screen, fittightly over the above compartment. This chamberhad acoustical properties superior to those of thesquare box used in Experiment 1, and the visualfield of the chick was much more uniform. Thechamber was illuminated by diffuse light from a40-W bulb on the ceiling of the sound-attenuatedroom in which the chamber was situated. Thesound-attenuated room was maintained at an aver-age temperature of 21.1 °C. Background whitenoise was constantly maintained at 55 dB (re 20/iN/m2, B scale) measured at chick ear level. Atelevision camera situated in front of and slightlyabove the chamber allowed the experimenter tocontinually monitor the chick's behavior. A SonyCardioid microphone (Type F-98) fit tightly intoa hole in the experimental chamber 5.08 cm belowthe chick's standing level. The microphone outputwas amplified and displayed as in Experiment 1.
StimuliStimuli were the same as in Experiment 1: a
1.6-sec train of four pure-tone bursts. All fre-quencies were presented at 90 dB (re 20 ^N/ma),35 dB above background level. No stimulus usedin this experiment varied more than 3 dB overthe sound field.
ProcedureAt 3-4 days after hatching (calculated from
the mean time at which the normal birds hatched),the chicks were removed individually from theircommunal brooders and placed in an isolationbrooder maintained at 32 ± 4 °C for 25 min priorto testing. This isolation brooder was not soundattenuated and was situated in the laboratory. Inthe case of the deprived chicks, the ear plugs wereremoved immediately prior to placement in theisolation brooder to allow adaptation to the newsound levels and air pressure equilibration. Thus,the deprived chicks experienced 25 min of a nor-mal acoustic environment prior to testing. Im-pressions of the tympanic membrane and the foot'of the columella were present on most ear plugs,which suggested good plugs and no gross damageto the middle ear.
HabituationAfter the 25-min adaptation period in the iso-
lation brooder, chicks were placed in the ex-
perimental chamber and allowed 5 min to adapt tothe chamber. Vocalizations from the chicks wereamplified and recorded as in Experiment 1. Theresponse measures, which were identical to thoseused in Experiment 1, were latency to resumecalling (Latency) and the number of calls within8 sec (No. Calls). The habituation stimulus was800 Hz for all subjects. Habituation training wasidentical to the contingent condition in Experiment1 except that the three mock (nonstimulus) trialswere presented at Positions 5, 10, and 15. Again,if the chick did not orient to the tones within thefirst two trials or if the chick did not make anydistress calls within any 5-min period of testing,it was eliminated from the study. Totals of 18deprived, 15 sham, and 16 normal chicks wereeliminated from the study by these criteria andreplaced by new subjects.
Generalisation Testing
Generalization testing began 30 sec after the lasthabituation trial, and it was the same as in Ex-periment 1. There were 50 chicks in each of thethree experimental conditions (normal, sham, anddeprived). Each chick was presented with fivesuccessive generalization test trials, with thestimulus at the same frequency on each trial.Ten subjects from each condition were tested withstimuli at each of the five frequencies: 800 (con-trol condition), 825, 850, 900, and 1,000 Hz.
Results and Discussion
Habituation Training
Results of habituation training for chicksin all three conditions are presented inFigure 6 (Latency). The 12 habituationtrials are presented on the left, and themean of the three mock trials, which wereinterspersed with the habituation trials, arepresented on the right. There was a markeddecrease in Latency over trials, indicatingthat habituation training was successful.Figure 6 also shows that the Latency of thedeprived chicks is at times lower than theLatency of the normal and sham chicks. ACondition X Trials analysis of variance wasperformed to further investigate these ob-servations. The trials effect was significant,F(ll, 1617) = 285.18, p < .01, as was thecomparison between conditions, F(2, 147)= 3.91, p < .05. The Condition X Trialsinteraction was not significant, F(22, 1617)= 1.20. The results for the No. Calls mea-sure were similar except that the difference
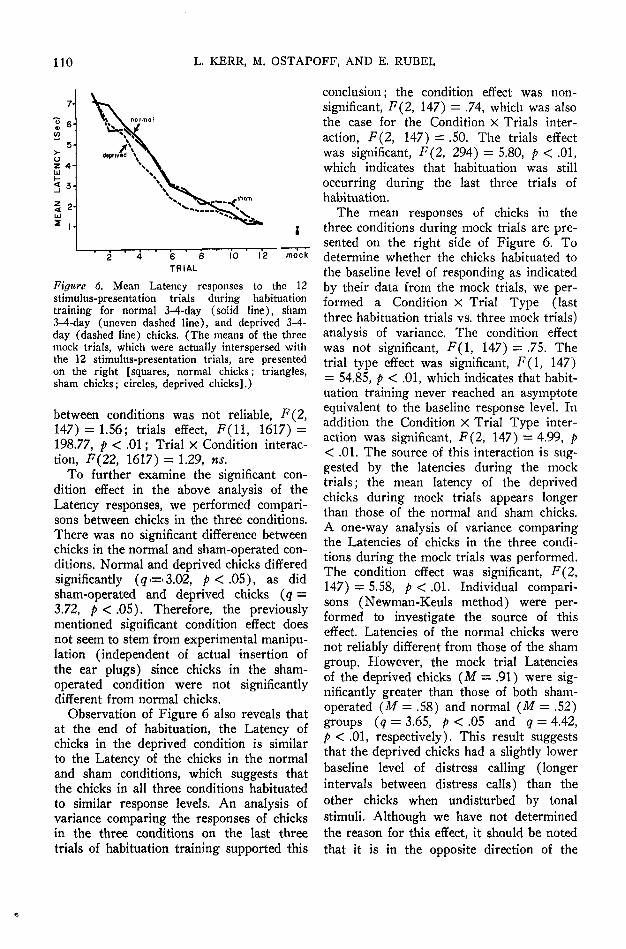
110 L. KERR, M. OSTAPOFF, AND E. RUBEL
7-
w 6-
" 5oz 4uj
< 3
z gUJ2 ,
depnv«d V
6 8TRIAL
10 12 mock
Figure 6. Mean Latency responses to the 12stimulus-presentation trials during habituationtraining for normal 3-4-day (solid line), sham3-4-day (uneven dashed line), and deprived 3-4-day (dashed line) chicks. (The means of the threemock trials, which were actually interspersed withthe 12 stimulus-presentation trials, are presentedon the right [squares, normal chicks; triangles,sham chicks; circles, deprived chicks].)
between conditions was not reliable, F{2,147) = 1.56; trials effect, F(ll, 1617) =198.77, p < .01; Trial X Condition interac-tion, F(22, 1617) = 1.29, ns.
To further examine the significant con-dition effect in the above analysis of theLatency responses, we performed compari-sons between chicks in the three conditions.There was no significant difference betweenchicks in the normal and sham-operated con-ditions. Normal and deprived chicks differedsignificantly (g=>3.02, p < .05), as didsham-operated and deprived chicks (q =3.72, p < .05). Therefore, the previouslymentioned significant condition effect doesnot seem to stem from experimental manipu-lation (independent of actual insertion ofthe ear plugs) since chicks in the sham-operated condition were not significantlydifferent from normal chicks.
Observation of Figure 6 also reveals thatat the end of habituation, the Latency ofchicks in the deprived condition is similarto the Latency of the chicks in the normaland sham conditions, which suggests thatthe chicks in all three conditions habituatedto similar response levels. An analysis ofvariance comparing the responses of chicksin the three conditions on the last threetrials of habituation training supported this
conclusion; the condition effect was non-significant, F(2, 147) = .74, which was alsothe case for the Condition X Trials inter-action, F(2, 147) = .50. The trials effectwas significant, F(2, 294) = 5.80, p < .01,which indicates that habituation was stilloccurring during the last three trials ofhabituation.
The mean responses of chicks in thethree conditions during mock trials are pre-sented on the right side of Figure 6. Todetermine whether the chicks habituated tothe baseline level of responding as indicatedby their data from the mock trials, we per-formed a Condition X Trial Type (lastthree habituation trials vs. three mock trials)analysis of variance. The condition effectwas not significant, F(l, 147) = .75. Thetrial type effect was significant, F(l, 147)= 54.85, p < .01, which indicates that habit-uation training never reached an asymptoteequivalent to the baseline response level. Inaddition the Condition X Trial Type inter-action was significant, F(2, 147) = 4.99, p< .01. The source of this interaction is sug-gested by the latencies during the mocktrials; the mean latency of the deprivedchicks during mock trials appears longerthan those of the normal and sham chicks.A one-way analysis of variance comparingthe Latencies of chicks in the three condi-tions during the mock trials was performed.The condition effect was significant, F(2,147) = 5.58, p < .01. Individual compari-sons (Newman-Keuls method) were per-formed to investigate the source of thiseffect. Latencies of the normal chicks werenot reliably different from those of the shamgroup. However, the mock trial Latenciesof the deprived chicks (M — .91) were sig-nificantly greater than those of both sham-operated (M = .58) and normal (M = .52)groups (q = 3.65, p < .05 and q = 4.42,p < .01, respectively). This result suggeststhat the deprived chicks had a slightly lowerbaseline level of distress calling (longerintervals between distress calls) than theother chicks when undisturbed by tonalstimuli. Although we have not determinedthe reason for this effect, it should be notedthat it is in the opposite direction of the
ACOUSTIC EXPERIENCE AND STIMULUS GENERALIZATION 111
differences found during generalizationtesting (see below) and therefore has littleconsequence regarding the differences be-tween groups in generalization functions.
Generalization Testing
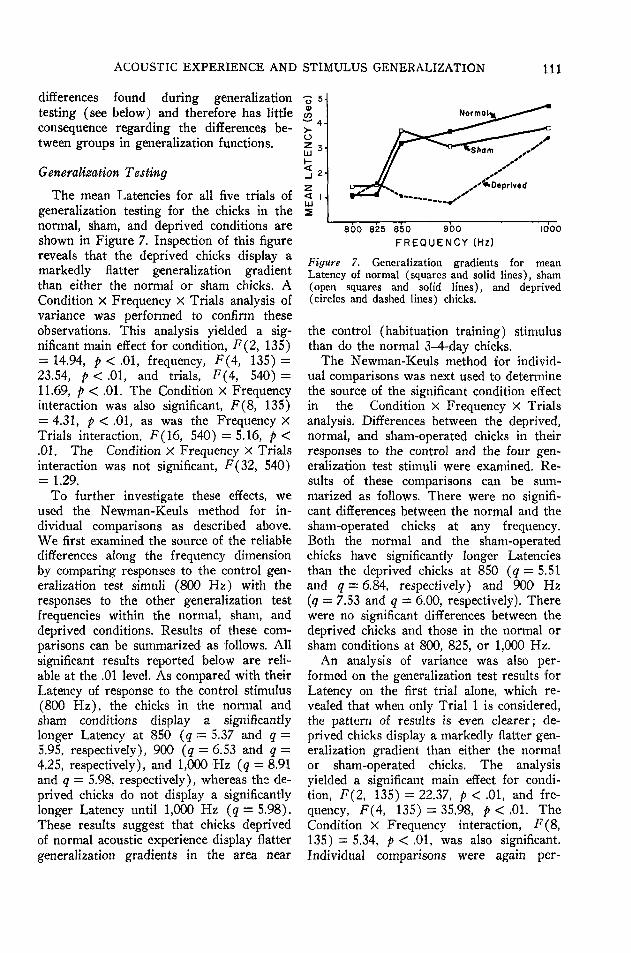
The mean Latencies for all five trials ofgeneralization testing for the chicks in thenormal, sham, and deprived conditions areshown in Figure 7. Inspection of this figurereveals that the deprived chicks display amarkedly flatter generalization gradientthan either the normal or sham chicks. ACondition X Frequency X Trials analysis ofvariance was performed to confirm theseobservations. This analysis yielded a sig-nificant main effect for condition, F(2, 135)= 14.94, p < .01, frequency, F(4, 135) =23.54, p < .01, and trials, F(4, 540) =11.69, p < .01. The Condition X Frequencyinteraction was also significant, F(8, 135)= 4.31, p < .01, as was the Frequency XTrials interaction. F(16, 540) = 5.16, p <.01. The Condition X Frequency X Trialsinteraction was not significant, F(32, 540)= 1.29.
To further investigate these effects, weused the Newman-Keuls method for in-dividual comparisons as described above.We first examined the source of the reliabledifferences along the frequency dimensionby comparing responses to the control gen-eralization test simuli (800 Hz) with theresponses to the other generalization testfrequencies within the normal, sham, anddeprived conditions. Results of these com-parisons can be summarized as follows. Allsignificant results reported below are reli-able at the .01 level. As compared with theirLatency of response to the control stimulus(800 Hz), the chicks in the normal andsham conditions display a significantlylonger Latency at 850 (q = 5.37 and q =5.95, respectively), 900 (q = 6.53 and q =4.25, respectively), and 1,000 Hz (q = 8.91and q = 5.98, respectively), whereas the de-prived chicks do not display a significantlylonger Latency until 1,000 Hz (q = 5.98).These results suggest that chicks deprivedof normal acoustic experience display flattergeneralization gradients in the area near
4-
< ILU5
Normals
800 825 850 900
F R E Q U E N C Y (Hz)1000
Figure 7. Generalization gradients for meanLatency of normal (squares and solid lines), sham(open squares and solid lines), and deprived(circles and dashed lines) chicks.
the control (habituation training) stimulusthan do the normal 3-4-day chicks.
The Newman-Keuls method for individ-ual comparisons was next used to determinethe source of the significant condition effectin the Condition X Frequency X Trialsanalysis. Differences between the deprived,normal, and sham-operated chicks in theirresponses to the control and the four gen-eralization test stimuli were examined. Re-sults of these comparisons can be sum-marized as follows. There were no signifi-cant differences between the normal and thesham-operated chicks at any frequency.Both the normal and the sham-operatedchicks have significantly longer Latenciesthan the deprived chicks at 850 (q = 5.51and q = 6.84, respectively) and 900 Hz(q = 7.53 and q = 6.00, respectively). Therewere no significant differences between thedeprived chicks and those in the normal orsham conditions at 800, 825, or 1,000 Hz.
An analysis of variance was also per-formed on the generalization test results forLatency on the first trial alone, which re-vealed that when only Trial 1 is considered,the pattern of results is even clearer; de-prived chicks display a markedly flatter gen-eralization gradient than either the normalor sham-operated chicks. The analysisyielded a significant main effect for condi-tion, F(2, 135) = 22.37, p < .01, and fre-quency, F(4, 135) = 35.98, p < .01. TheCondition X Frequency interaction, F(8,135) = 5.34, p < .01, was also significant.Individual comparisons were again per-

112 L. KERR, M, OSTAPOFF, AND E. RUBEL
formed and revealed a pattern similar tothat described above. (Analysis performedon the No. Calls measure during generaliza-tion testing yielded a pattern identical to theresults for the Latency measure as describedabove.)
Thus, it appears that the lack of normalauditory experience over the period fromjust prior to hatching to approximately 30min before testing at 3-4 days preventedthe perceptual sharpening that normallyoccurs over this developmental period. Theflatter generalization gradients cannot beattributed to the effects of the operationprocedure per se since the responses of thesham chicks were not significantly differentfrom those of the normal chicks.
Whereas we think that the differencesbetween generalization functions of the de-prived chicks and the chicks in the normaland sham conditions can best be ascribed toa failure in the normal occurrence of per-ceptual sharpening, alternative explanationsshould be examined.
One possible interpretation of the differ-ences in generalization gradients is thatthey were due to underlying differences inoverall response levels between deprivedchicks and normal or sham-operated chicks.There are two reasons why this might beentertained. First, the deprived chicks hadslightly longer latencies on mock trials thandid the normal and sham-operated groups.These differences would, if anything, biasthe results in the opposite direction to thoseobserved, and therefore could not be ofmajor consequence. Second, the deprivedchicks displayed marginal but statisticallysignificant acceleration of habituation, whichmay suggest that the deprived chicks wereresponding at uniformly shorter latenciesthan the other groups during stimulus trials.Had this been the case, it would be ex-pected that the latencies would have beenconsistently less and the number of callsconsistently more for deprived chicks thanfor normal or sham chicks on the test trialsat 1,000 Hz as well as on the final habitua-tion trials and test trials at 800 and 825 Hz.This was not the case. However, at 850and 900 Hz these differences are qualita-
tively large, consistent, and highly signifi-cant. Also, the generalizaiton function of thedeprived chicks is qualitatively and statis-tically flat out to 900 Hz. Given the markedchange in response level of both controlgroups between 825 and 850 Hz, somechange in the response function of the de-prived chicks would be expected if under-lying parallel functions were the case.
A second interpretation is that because ofthe slightly accelerated habituation the de-prived group was overtrained, comparedwith the control groups, and that this ledto a shallower generalization gradient in thedeprived group. Thorough investigations ofthe relation between the amount of habitua-tion training and the generalization gradientslope are not available; however, in anearlier study (Rubel & Rosenthal, 1975) itwas found that overtraining did notmarkedly influence the slope of frequencygeneralization gradients in 1-day or 3-4-daychicks. If anything, the generalization func-tions appeared steeper with increased habit-uation training.
A final alternative interpretation, that thedeprived chicks are less sensitive to thehigher frequencies (above 825 Hz) than arethe normal or sham-operated chicks, alsoseems very unlikely. Differences betweenthe groups were not even marginally sig-nificant at 1,000 Hz for either Latency orNo, Calls, whereas at both 850 and 900 Hzthe differences between deprived chicks andchicks in the control groups were highlyreliable (p < .01) on both measures. Theresults of Experiment 2 also speak to thispoint; with similar deprivation conditions,evoked potential thresholds returned to nor-mal throughout the frequency range as soonas the ear plugs were removed (3-4-daychronic group).
In summary, it is unlikely that the flattergeneralization gradient found in the acousti-cally deprived subjects is attributable tooverall differences in responsiveness, to dif-ferences in habituation, or to differences insensitivity at the higher frequencies betweendeprived and normal or sham-operatedchicks. Therefore, it can be stated that theabsence of normal acoustic experience re-

ACOUSTIC EXPERIENCE AND STIMULUS GENERALIZATION 113
suits in broader stimulus classifications thanare evidenced by normal subjects. In otherwords, the perceptual sharpening that occursover the first 3-4 days after hatching isdependent on normal auditory experience.
General Discussion
We have observed that both 1-day normalchicks and acoustically deprived 3- to 4-daychicks display flatter generalization gradi-ents than normal 3- to 4-day chicks. Theseresults implicate experience in the per-ceptual sharpening that is observed duringnormal ontogeny. Other deprivation anddifferential rearing studies, in both the audi-tory and visual systems, also suggest anexperiential role in perceptual development.In the visual system, for exmaple, Peterson,(1962) observed flat generalization gradi-ents for ducklings raised in monochromaticlight while those for ducklings raised inwhite light were & function of the wave-length difference between the training andthe test stimuli (also see Oppenheim, 1968;Tracy, 1970). In another study, Ganz andFitch (1968) found that monocularly rearedcats experienced severe initial deficienciesin depth estimation and visual form dis-crimination with their deprived eyes.
Similar results have been obtained instudies of the auditory system. Tees (1967a,1967b) showed that rats, for example,acoustically restricted at an early age per-formed more poorly than normal animalson tasks involving tonal pattern and formdiscrimination but that they were similarto normal animals on intensity and frequencydiscrimination tasks. Batkin and Ansberry(1964) and Batkin, Groth, Watson, andAnsberry (1970) found that guinea pigsacoustically deprived from birth showed amarked insensitivity to sound. However, thelatter study suggests that this effect re-covers within approximately 3 wk. Clementsand Kelly (1978) found that auditory de-privation had detrimental effects on soundlocalization in the guinea pig. All of theabove mentioned studies, however, sufferunder the criticism offered by Solomon andLessac (1968) ; the experiential role in per-
ceptual development is impossible to pre-cisely determine without information con-cerning the initial and final capacities ofthe organism being studied.
Gottlieb (1976b) formalized and extendedthis position by suggesting three possibleroles of experience in normal ontogeny.First, sensory experience may serve to main-tain an ability that developed prior to, orindependent of, the sensory events in ques-tion. Gottlieb gave several examples, one ofwhich is especially germane here; youngchicks, which normally respond to speciesmaternal calls at birth, become relatively un-responsive if they are not allowed to hearmaternal calls for several days (Graves,1973). Another example was provided byGottlieb (1976a). Devocalized, isolate duck-lings are, at 65 hr posthatch, less able todiscriminate between normal and high fre-quency maternal calls than they were at48-hr posthatch. Thus, auditory experienceis necessary to maintain the highly selectiveability that the deprived ducklings exhibitedearlier in their development. The same phe-nomenon was observed by Tees (1974).Dark-reared (deprived) rats, although in-itally inferior to light-reared rats in theirdiscrimination of depth on a visual cliffapparatus, begin to improve despite a con-tinuing absence of visual experience. Even-tually, however, their performance deterio-rates. In this case, visual experience isnecessary to maintain the improved depthdiscrimination.
According to Gottlieb's classificationscheme, a second role of sensory experiencemay be jaciHtative, where it works quantita-tively to accelerate the development of abehavior that would eventually occur, al-though more slowly, in the absence of ap-propriate stimulation. Tees (1974) demon-strated that dark-reared rats develop theirdepth perception more slowly than normalrats, and Gottlieb (1971) demonstrated thatdevocalized, isolate Peking ducklings de-veloped the ability to discriminate theirspecies-typical maternal call from the woodduck maternal call more slowly than normal,communally reared ducklings. In both ofthese studies, experience acted quantita-

114 L. KERR, M. OSTAPOFF, AND E. RUBEL
tively by accelerating, not causing, the oc-currence of a particular behavioral process.
The third possible role of experience isinduction, in which case a particular be-havior will not develop in the absence of aparticular experience (or set of experiences).Gottlieb offered imprinting as an examplewhere early experience is necessary for thedevelopment of attachment behavior to aspecific object.
In Gottlieb's classification scheme, ex-periences that induce a behavior are distin-guished from those that work on the be-havior by either accelerating (facilitating)or maintaining the behavior. In relation tothe experiments presented here, however,we view perceptual development as an on-going process of which any part or stagecan be induced, facilitated, or maintained.The fact that 1-day chicks do not normallydisplay frequency generalization gradientsthat are as sharp as those of 3- to 4-daychicks indicates that a major influence ofacoustic experience on this ontogeneticprocess over this time period is formativein nature. This result, coupled with the factthat 3- to 4-day deprived chicks displayflatter generalization gradients than 3- to 4-day normal chicks, rules out maintenanceor simple maturation and implicates an ac-tive role for experience. Therefore, over thefirst 3-4 days posthatching, experience maybe acting to either (a) induce the perceptualsharpening in which case no further sharp-ening of generalization gradients will occurwithout the benefit of acoustic experience,or (b) facilitate the process, in which casethe development of the perceptual sharpen-ing will eventually occur in the continuedabsence of normal acoustic experience. Itwill be important to apply the paradigmused in this study to older chicks in orderto differentiate between the possible induc-tive or facilitative roles of experience.
The types of experience necessary for theperceptual change we have examined hereis also in need of further clarification. Forinstance, is acoustic experience in generalsufficient for perceptual sharpening to occur,or does acoustic experience have to bespecific to the frequency range to which the
animals must attend? This information,which can be obtained by using this sameparadigm and precisely varying the amountand frequency range of auditory experience,may shed some light on the specificity ofexperience necessary for perceptual develop-ment.
Reference Notes
1. Ostapoff, E. M., & Rubel, E. W. Unpublishedobservations, 1975.
2. Kerr, L. M., & Rubel, E. W. Unpublished ob-servations, 1976.
3. Gottlieb, G. Personal communication, April1975.
References
Batkin, S., & Ansberry, M. Effect of auditory dep-rivation. Journal of the Acoustical Society ofAmerica, 1964, 36, 598. (Letter)
Batkin, S., Groth, H., Watson, J. R., & Ansberry,M. Effects of auditory deprivation on the de-velopment of auditory sensitivity in albino rats.Electroencephalography and Clinical Neurophvs-iology, 1970, 28, 351-359.
Bornstein, M. H., Kessen, W., & Weiskopf, S. A.Categories of hue in infancy. Science, 1976, 191,201-202.
Carlton, P. H., & Vogel, J. R. Habituation andconditioning. Journal of Comparative and Physi-ological Psychology, 1967, 63, 348-351.
Clements, M., & Kelly, J. B. Auditory spatial re-sponses of young guinea pigs (Cavia porcellus)during and after ear blocking. Journal of Com-parative and Physiological Psychology, 1978, 92,34-44.
Clopton, B. M., & Silverman, M. S. Plasticity ofbinaural interaction: II. Critical period andchanges in midline response. Journal of Neuro-physiology, 1977, 40, 1275-1280.
Fantz, R. L. Ontogeny of perception. In A. M.Schrier, H. F. Harlow, & F. Stollnitz (Eds.),Behavior of non-human primates (Vol. 2). NewYork: Academic Press, 1965.
Ganz, L. An analysis of generalization behavior inthe stimulus deprived organism. In G. Newton &S. Levine (Eds.), Early experience and be-havior: The psychobiology of development.Springfield, 111.: Charles C Thomas, 1968.
Ganz, L., & Fitch, M. The effect of visual depriva-tion on perceptual behavior. Experimental Neu-rology, 1968, 22, 638-660.
Gibson, E. J. Principles of perceptual learning anddevelopment. New York: Appleton-Century-Crofts, 1969.
Gibson, J. J., & Gibson, E. J. Perceptual learning:Differentiation or enrichment? PsychologicalReview, 1955, 62, 32-41.

ACOUSTIC EXPERIENCE AND STIMULUS GENERALIZATION 115
Gottlieb, G. Development of species identificationin birds: An inquiry into the prenatal determi-nants of perception. Chicago: University of Chi-cago Press, 1971.
Gottlieb, G. Early development of species-specificauditory perception in birds. In G. Gottlieb(Ed.), Studies on the development of behaviorand the nervous system (Vol. 3). Neural andbehavioral specificity. New York: AcademicPress, 1976. (a)
Gottlieb, G. The roles of experience in the devel-opment of behavior and the nervous system. InG. Gottlieb (Ed.), Studies on the development ofbehavior and the nervous system: (Vol. 3).Neural and behavioral specificity. New York:Academic Press, 1976. (b)
Gottlieb, G., & Vandenbergh, J. G. Ontogeny ofvocalization in duck and chick embryos. Journalof Experimental Zoology, 1968, 168, 307-326.
Graves, H. B. Early social responses in Callus:A functional analysis. Science, 1973, 182, 937-938.
Hailman, J. P. The ontogeny of an instinct. Be-haviour, 1967, Suppl. 15, 1-159.
Hinde, R. A. Animal behavior (2nd ed.). NewYork: McGraw-Hill, 1970.
Hoffman, H. S., Schiff, D., Adams, J., & Searle,J. L. Enhanced distress vocalization throughselective reinforcement. Science, 1966, 151, 352-354.
Kaufman, I. C., & Hinde, R. A. Factors influencingdistress calling in chicks with special referenceto temperature changes and social isolation. Ani-mal Behaviour, 1961, 9, 197-204.
Lashley, K, S., & Wade, M. The Pavlovian theoryof generalization. Psychological Review, 1946,S3, 72-87.
Lubow, R. E., Schnur, P., & Rifkin, B. Latent in-hibition and conditioned attention theory. Jour-nal of Experimental Psychology: Animal Be-havior Processes, 1975, 2, 163-174.
Lubow, R. E., & Siebert, L. Latent inhibitionwithin the CER paradigm. Journal of Compara-tive and Physiological Psychology, 1969, 68,136-138.
Messmer, F., & Messmer, I. Die Entwicklung derlautausserungen und einiger Verhaltensweisender Amsel. (Turdus merula merula L.) unternaturlichen Bedingungen und nach Einzelauf-zucht in schalldichten Raumen, Zeitschrift fiirTierpsychologie, 1956, 13, 341-441.
Oppenheim, R. W. Color preferences in the peck-ing response of newly hatched ducks (Anasplatyrhynchos). Journal of Comparative andPhysiological Psychology, 1968, 66(3, Pt. 2).
Oppenheim, R. W. Prehatching and hatching be-havior : Comparative and physiological consid-erations. In G. Gottlieb (Ed.), Studies on thedevelopment of behavior and the nervous sys-tem: (Vol. 1). Behavioral embryology. NewYork: Academic Press, 1973.
Peterson, N. Effect of monochromatic rearing onthe control of responding by wavelength. Sci-ence, 1962, 136, 774-775.
Rubel, E. W. Effects of early experience on fearbehaviour of Coturnix coturnix. Animal Be-haviour, 1970, 18, 427-433.
Rubel, E. W. Ontogeny of structure and functionin the vertebrate auditory system. In M. Jacob-son (Ed.), Handbook of sensory physiology:(Vol. 9). Development of sensory systems. NewYork: Springer-Verlag, 1978.
Rubel, E. W., & Parks, T. N. Organization anddevelopment of brain stem auditory nuclei ofthe chicken: Tonotopic organization of n. mag-nocellularis and n. laminaris. Journal of Com-parative Neurology, 1975, 164, 411-434.
Rubel, E. W., & Rosenthal, M. H. The ontogenyof auditory frequency generalization in thechicken. Journal of Experimental Psychology:Animal Behavior Processes, 1975, 1, 287-297.
Saunders, J. S., Coles, R. B., & Gates, G. R. Thedevelopment of auditory evoked responses in thecochlea and cochlear nuclei of the chick. BrainResearch, 1973, 63, 59-74.
Silverman, M. S., & Clopton, B. M. Plasticity ofbinaural interaction: I. Effect of early auditorydeprivation. Journal of Neurophysiology, 1977,40, 1266-1274.
Solomon, R. L., & Lessac, M. S. A control groupdesign for experimental studies of developmentalprocesses. Psychological Bulletin, 1968, 70, 145-150.
Tees, R. C. Effects of early auditory restrictionin rats on adult pattern discrimination. Journalof Comparative and Physiological Psychology,1967, 63, 389-393. (a)
Tees, R. C. The effects of early auditory restric-tion in the rat on adult duration discrimination.Journal of Auditory Research, 1967, 7, 195-207.(b)
Tees, R. C. Effect of visual deprivation on devel-opment of depth perception in the rat. Journal ofComparative and Physiological Psychology, 1974,86, 300-308.
Tees, R. C. Perceptual development in mammals.In G. Gottlieb (Ed.), Studies on the develop-ment of behavior and the nervous system: (Vol.3). Neural and behavioral specificity. New York:Academic Press, 1976.
Tracy, W. K. Wavelength generalization andpreference in monochromatically reared duck-lings. Journal of the Experimental Analysis ofBehavior, 1970, 13, 163-178.
Werner, H. Comparative psychology of mental de-velopment. Chicago: Follett, 1948.
Winer, B. J. Statistical principles in experimentaldesign. New York: McGraw-Hill, 1962.
Received June 1, 1978Revision received November 24, 1978 •


















