
JOURNAL OF BIOLOGICAL No. Q The of Printed U. …THE JOURNAL OF BIOLOGICAL CHEMISTRY Q 1985 by The...
10
THE JOURNAL OF BIOLOGICAL CHEMISTRY Q 1985 by The American Society of Biological Chemists, Inc. Vol. 260, No. 24, Issue of October 26, pp. 13208-13217,1985 Printed in U. SA. Immunochemical Studies on Blood Groups THE COMBINING SITE SPECIFICITIES OF MOUSE MONOCLONAL HYBRIDOMA ANTI-A AND ANTI-B* (Received for publication, March 13, 1985) Hua-Tang Chen and Elvin A. Kabat From the Departments of Microbiology, Human Genetics and Development, and Neurology, and the Cancer Center, Columbia University, New York, New York 10032 Mouse monoclonal hybridomas, five anti-blood group A, three anti-B, and oneanti-AB, produced by various methods of immunization, have been characterized by quantitative precipitin tests and the fine structures of their combining sites have been mapped by oligosac- charide inhibition assays. The combining sites of anti- bodies of each specificity differed among themselves. Three of the five monoclonals were specific for difu- cosy1 and two for monofucosyl A determinants. All but the anti-AB were strictly specific for blood group A or blood group B erythrocytes; all of the anti-A monoclo- nals gave essentially equivalent titers in hemagglutin- ation tests with AI and A2 erythrocytes except for a monoclonal anti-A prepared by immunization with a human gastric cancer cell line. The data provide addi- tional evidence for the heterogeneity of the antibody response to the different antigenic determinants pres- ent on blood A and B substances and emphasize the importance of difucosyl determinants which comprise most of the determinants on the water-soluble blood group substances. Since the discovery of the four human blood groups, A, B, 0, and AB (1, 2), human sera containing anti-A and anti-B have been used in blood grouping. Such anti-A and anti-B sera contain heterogeneous populations of antibody combin- ing sites. Initially, sera containing high titers of anti-A and anti-B were selected for blood grouping, but when it was found that injection of hog or human blood group A substance into individuals of groups B and 0 and of horse or human blood group B substances intoindividuals of groups A and 0 yielded high titer antisera (3, 4), anti-A from immunized B individuals and anti-B from immunized A individuals came into widespread use. Although these antisera contained het- erogeneous populations of antibody molecules, their combin- ing sites were studied by assays of the capacities of various * This work was aided by National Science Foundation Grant NSF- PCM-81-02321,National Institute of Allergyand Infectious Diseases Grant 1ROl AI-19042 (to E. A. K.), and by Cancer Support Grant CA13696 to Columbia University. This is paper LXXIII in the series. This study and the following one by Gooi et al. (49) were carried out and the papers written completely independently. The manuscripts were then exchanged to permit comments by both groups. Results with the two anti-A monoclonals which happened to have been studied in both laboratories generally, with one exception, gave the same relative results allowing for the differences in assay methods, whenever the same oligosaccharides were used. Each study supple- ments the other and the two together provide an initial measure of the diversity of monoclonal mouse anti-A blood grouping reagents. The costs of publication of this article were defrayed in part by the payment of page charges. This articlemust therefore be .hereby marked “aduertisernent” in accordance with 18 U.S.C. Section 1734 solely to indicate thls Lct. isolated blood group oligosaccharides to inhibit the precipitin reaction of anti-A by bloodgroup A substances and of anti-B by blood group B substances (5-8), the site specificities being considered most complementary to the oligosaccharidesgiving inhibition at the lowest concentrations. Thus many anti-A sera were found (7, 9, 10) to be most complementary to the following. L-FUC~ .1” 1 L ~-GalNAcal+3~-Gal~l-+3D-GlcNAc-~~1 Type 1 or L-FUCO D-GalNAcal+3~-Galpl~D-GlcNAc-O~ Type 2 and anti-B sera to L-FUCCY 4 ; ~-Galal+3~-Ga1/31+3D-GlcNAc-OR~ Type 1 and L-FUCCY J; ~-Galal+3~-Gal~l~D-GlcNAc-OR~ Type 2 R1 is 3-linked D-galactit01 formed by alkaline borohydride and R2 is 6-linked 3-hexenetetrols formed by alkaline P-elimina- tion (peeling) and reduction by borohydride of a 3,6-linked galactose. In both of these structures,the nonreducing trisac- charides were immunodominant and contributed a large part of the binding energy of the intact determinant. Since the trisaccharide portions of both the A and B structures are identical, specificity differences, except for the terminal non- reducing GalNAcl and Gal, among various anti-A and among anti-B sera involve the fourth sugar and the linkage of the trisaccharide to it. Any differences among antibody molecules with sites larger than the trisaccharide might often not be detectable in view of the heterogeneity and the predominant The abbreviations used are: GalNAc, N-acetyl-D-galactosamine; ELISA, enzyme-linked immunosorbent assays; Ap, alkaline phos- phatase; BSA, bovine serum albumin; Gal, D-galactose; GlcNAc, N-acetyl-D-glucosamine; Fuc, L-fucose; AbN, antibody nitrogen. When not given, all sugars are in D except for fucosewhich is L. 01, GalNAce(l-+3) Gd$-O(CH2)&OOCH~; 02, Gala(l-3)Galp-O- (CH&COOCHs. Fuca FUCCY 13208
Transcript of JOURNAL OF BIOLOGICAL No. Q The of Printed U. …THE JOURNAL OF BIOLOGICAL CHEMISTRY Q 1985 by The...

THE JOURNAL OF BIOLOGICAL CHEMISTRY Q 1985 by The American Society of Biological Chemists, Inc.
Vol. 260, No. 24, Issue of October 26, pp. 13208-13217,1985 Printed in U. SA.
Immunochemical Studies on Blood Groups THE COMBINING SITE SPECIFICITIES OF MOUSE MONOCLONAL HYBRIDOMA ANTI-A AND ANTI-B*
(Received for publication, March 13, 1985)
Hua-Tang Chen and Elvin A. Kabat From the Departments of Microbiology, Human Genetics and Development, and Neurology, and the Cancer Center, Columbia University, New York, New York 10032
Mouse monoclonal hybridomas, five anti-blood group A, three anti-B, and one anti-AB, produced by various methods of immunization, have been characterized by quantitative precipitin tests and the fine structures of their combining sites have been mapped by oligosac- charide inhibition assays. The combining sites of anti- bodies of each specificity differed among themselves. Three of the five monoclonals were specific for difu- cosy1 and two for monofucosyl A determinants. All but the anti-AB were strictly specific for blood group A or blood group B erythrocytes; all of the anti-A monoclo- nals gave essentially equivalent titers in hemagglutin- ation tests with AI and A2 erythrocytes except for a monoclonal anti-A prepared by immunization with a human gastric cancer cell line. The data provide addi- tional evidence for the heterogeneity of the antibody response to the different antigenic determinants pres- ent on blood A and B substances and emphasize the importance of difucosyl determinants which comprise most of the determinants on the water-soluble blood group substances.
Since the discovery of the four human blood groups, A, B, 0, and AB (1, 2), human sera containing anti-A and anti-B have been used in blood grouping. Such anti-A and anti-B sera contain heterogeneous populations of antibody combin- ing sites. Initially, sera containing high titers of anti-A and anti-B were selected for blood grouping, but when it was found that injection of hog or human blood group A substance into individuals of groups B and 0 and of horse or human blood group B substances into individuals of groups A and 0 yielded high titer antisera (3, 4), anti-A from immunized B individuals and anti-B from immunized A individuals came into widespread use. Although these antisera contained het- erogeneous populations of antibody molecules, their combin- ing sites were studied by assays of the capacities of various
* This work was aided by National Science Foundation Grant NSF- PCM-81-02321, National Institute of Allergy and Infectious Diseases Grant 1ROl AI-19042 (to E. A. K.), and by Cancer Support Grant CA13696 to Columbia University. This is paper LXXIII in the series. This study and the following one by Gooi et al. (49) were carried out and the papers written completely independently. The manuscripts were then exchanged to permit comments by both groups. Results with the two anti-A monoclonals which happened to have been studied in both laboratories generally, with one exception, gave the same relative results allowing for the differences in assay methods, whenever the same oligosaccharides were used. Each study supple- ments the other and the two together provide an initial measure of the diversity of monoclonal mouse anti-A blood grouping reagents. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be .hereby marked “aduertisernent” in accordance with 18 U.S.C. Section 1734 solely to indicate thls Lct.
isolated blood group oligosaccharides to inhibit the precipitin reaction of anti-A by blood group A substances and of anti-B by blood group B substances (5-8), the site specificities being considered most complementary to the oligosaccharides giving inhibition at the lowest concentrations. Thus many anti-A sera were found (7, 9, 10) to be most complementary to the following.
L - F U C ~
.1” 1 L
~-GalNAcal+3~-Gal~l-+3D-GlcNAc-~~1 Type 1
or
L-FUCO
D-GalNAcal+3~-Galp l~D-GlcNAc-O~ Type 2
and anti-B sera to
L-FUCCY
4; ~-Galal+3~-Ga1/31+3D-GlcNAc-OR~ Type 1
and
L-FUCCY
J; ~ - G a l a l + 3 ~ - G a l ~ l ~ D - G l c N A c - O R ~ Type 2
R1 is 3-linked D-galactit01 formed by alkaline borohydride and R2 is 6-linked 3-hexenetetrols formed by alkaline P-elimina- tion (peeling) and reduction by borohydride of a 3,6-linked galactose. In both of these structures, the nonreducing trisac- charides were immunodominant and contributed a large part of the binding energy of the intact determinant. Since the trisaccharide portions of both the A and B structures are identical, specificity differences, except for the terminal non- reducing GalNAcl and Gal, among various anti-A and among anti-B sera involve the fourth sugar and the linkage of the trisaccharide to it. Any differences among antibody molecules with sites larger than the trisaccharide might often not be detectable in view of the heterogeneity and the predominant
The abbreviations used are: GalNAc, N-acetyl-D-galactosamine; ELISA, enzyme-linked immunosorbent assays; Ap, alkaline phos- phatase; BSA, bovine serum albumin; Gal, D-galactose; GlcNAc, N-acetyl-D-glucosamine; Fuc, L-fucose; AbN, antibody nitrogen. When not given, all sugars are in D except for fucose which is L. 01, GalNAce(l-+3) Gd$-O(CH2)&OOCH~; 02, Gala(l-3)Galp-O- (CH&COOCHs.
Fuca FUCCY
13208

Immunochemical Studies on Blood Groups 13209
contribution of the nonreducing trisaccharide portion to the total binding energy.
However, some oligosaccharides isolated from water-soluble blood group A and B substances contain a second fucose- linked a 1 4 or a1+3 to the GlcNAc in the type 1 and 2 structures above, respectively. With five anti-A and three anti-B sera, the corresponding A and B difucosyl oligosaccha- rides with the second fucose-linked a 1 4 to the D-GlcNAc of the type 2 chain ( 6 , l l ) were considerably poorer inhibitors than the monofucosyl type 1 and type 2 oligosaccharides. However, with one anti-A serum, the type 2 difucosyl oligo- saccharide was appreciably more active than the correspond- ing type 2 monofucosyl oligosaccharide (11). It is evident that heterogeneity of the antibody response in antisera complicates evaluation of the contribution of various antibodies with different site structures. By contrast with monoclonal anti-I and anti-i human sera, individual combining sites have been precisely mapped (12-15).
The introduction of the hybridoma technic by Kohler and Milstein (16) has provided a spectrum of mouse ascitic fluids or tissue culture supernatants containing monoclonal anti-A and anti-B which will probably replace human anti-A and anti-B typing sera. Indeed, commercial production of such monoclonals has begun in Canada (17), Great Britain (18- 20), and Sweden (21). For research purposes, it is of impor- tance to define the combining site specificities of these various monoclonals. This will eventually permit a precise evaluation of the utility of various reagents and will help to define the repertoire of the antibody response to water-soluble blood group substances, to A and B erythrocytes, and to A and B determinants synthesized chemically and coupled to protein or to 0 erythrocytes, the materials generally used by various laboratories for immunization. Accordingly, we have mapped the combining sites of five monoclonal anti-A, one monoclonal anti-AB, and three monoclonal anti-B by ELISA using oli- gosaccharides to inhibit A anti-A and B anti-B binding and have studied their reactivity with various blood group A or B substances by the quantitative precipitin reaction. Individual monoclonals differed in their combining sites indicating, as inferred from studies on whole antisera, that there is a con- siderable repertoire of different anti-A and anti-B combining sites and that determinants may be type 1 or type 2 and involve one or two fucoses. All of these give satisfactory and comparable results as blood grouping reagents.
MATERIALS AND METHODS
Mouse Monoclonal Hybridoma Antibodies-Anti-A AC-1001 and anti-B BC-1004 were obtained from Dr. Don Baker, University of Alberta, Edmonton, Alberta, Canada (17). A15/3D3.92.1, MH2/ 6D4.12.3.1 (we refer to it as MH2/6D4), A15/1A4.7.3, and NB10/ 5B2.4.5 were provided by Drs. A. D. Lowe, and E. Lennox, Medical Research Council Laboratory of Molecular Biology, Cambridge, Eng- land (18-20). Anti-A A003=40/5G7 and anti-B B003=46/2D7 were provided by Drs. A. Lundblad and L. Messeter, Department of Clin- ical Chemistry and University Hospital, University of Lund, Lund, Sweden (21). AH16 was from Dr. Sen-Itiroh Hakomori, University of Washington, Seattle (22). Anti-AB A15/1A4.7.3 was shown to be monoclonal (20) since absorption with A or with B erythrocytes reduced both anti-A and anti-B levels proportionally. Table I gives the methods of immunization employed. All monoclonals were of the IgM class except for AH16 which was IgG3 (22).
Blood Group Substances-The purified blood group substances used were prepared in this laboratory from human ovarian cyst fluid or saliva and from horse, bovine, or hog gastric mucosa (23-27).
Mom- and Oligosaccharides-The following were available: D- galactose, N-acetyl-D-galactosamine, N-acetyl-D-glucosamine, and L- fucose (Sigma); methyl-a-D-galactoside (Sigma); melibiose (East- man Kodak Company); galactinol; RL1.34; A5II; Beach P1RL0.62; GalNAca(l+3)[Fuca(1+2)]Gal; Gala(l+3)[Fuca(1+2)]Gal; and
TABLE I Immunization schedules for producing monoclonal anti-A, anti-B,
and anti-AB ~ ~
Monoclonal Antigen used for immuni- zation Reference
Anti-A AC-1001 AI erythrocytes and 01
coupled to BSA or to 0 erythrocytes
substance
(HT-29) from hu- man group A colon carcinoma case
insoluble cyst 14
cell line MKN 45
A15/3D3.92.1 Al erythrocytes and A 18
MH2/6D4 Tissue culture cells 19
A003=40/5G7 A2 substance phenol- 20
AH16 Human gastric cancer 21
Anti-B BC-1004 02 coupled to BSA or 17
to 0 erythrocytes
substance NB10/5B2.4.5 B erythrocytes and B
B003=46/2D7 B substance 20
A15/1A4.7.3 A, erythrocytes and A Anti-AB
substance
MSM&RL0.56. Beach BR~0.44 used were prepared in the laboratory as described previously (28-32). 01 and 02 were from Dr. Don Baker (17). A-tetrasaccharide, A-pentasaccharide, A-hexasaccharide, A- heptasaccharide, and B-pentasaccharide were gifts from Dr. A. Lund- blad (33, 34).
ELISA Reagents-Alkaline phosphatase, p-nitrophenyl phosphate as the disodium salt (phosphatase substrate), and glutaraldehyde were from Sigma, and affinity purified goat antibody to mouse p(1gM) were from Kirkegaard and Perry Laboratories Inc., Gaithersburg, MD. Alkaline phosphatase-labeled protein A was a gift from Dr. K. 0. Lloyd of Sloan-Kettering Institute.
Hemagglutination Assays-These were. carried out using a Takatsy microtitrator with 0.025-ml loops and a 2% suspension of human AI, A2, or B erythrocytes.
Quantitative Precipitin Assay-As described previously (35, 36), a constant amount of mouse monoclonal hybridoma antibody contain- ing 5 to 8 pg of antibody nitrogen was added to varying quantities of blood group substance (0-70 pg), the total volume was adjusted to 200 pl with saline (0.15 M NaCl), and the contents of each tube were mixed and incubated at 37 "C for 1 h and kept a t 4 "C for 5 days with daily mixing. The tubes were centrifuged at 4 "C at 2000 rpm, the supernatants were decanted, and the precipitates were washed twice with 0.5 ml of chilled saline. Total nitrogen in the washed precipitates was determined by ninhydrin assay (37).
Inhibition Assays by ELZSA-Coupling of alkaline phosphatase to goat anti-mouse IgM was performed by the one-step glutaraldehyde procedure (38, 39). The required quantities of enzyme and ailtibody were mixed in the given volume and dialyzed overnight against 0.05 M phosphate-buffered saline, pH 7.2, at 4 'C. The 25% glutaraldehyde was added to a final concentration of 0.2% while the solution was gently stirred and the reaction mixture was kept a t room temperature for 2 h. After dialysis against the same buffer overnight at 4 "C, the mixture was diluted with 5% BSA, 0.05 M Tris/M?/Na azide buffer to the appropriate concentration and stored in the refrigerator a t 4 "C.
Inhibition assays were carried out as follows. A polystyrene plate (Corning, New York) was coated with blood group substance: 1 pg/ ml Hog 4 10% for the A and anti-A system; and 2.5 pg/ml phenol- insoluble Tij I1 or 10 pg/ml Horse 4 25% for the B and anti-B system. 100 pl were placed in each well; the wells with no antigen served as a blank. After 2 h a t 37 "C, the wells were washed twice with saline containing 0.05% Tween 20 and 0.02% Na azide, the plate was blocked with 200 p1 of 1% BSA containing 0.05% Tween 20 and 0.02% Na azide at room temperature for 1 h, and the plate was again washed twice. Varying quantities of the sugar inhibitors and the amount of monoclonal antibody giving maximum precipitation were added; the wells with no inhibitor served as a standard. The total volume was

13210 Immunochemical Studies on Blood Groups
adjusted to 100 gl with 0.02 M borate buffered saline, pH 8.3, mixed, incubated at 37 "C for 3 h, then washed 3 times. 100 pl of an appropriately diluted alkaline phosphatase-labeled anti-mouse IgM or alkaline phosphate-labeled protein A was added and the plate was incubated at 37 "C for 3 h. After washing 3 times, 100 p1 Gf p- nitrophenyl phosphate solution, 30 mg/50 ml diethanolamine buffer, pH 9.8, were added and the reaction was placed at room temperature for 1 h. Twenty-five pl of 3 N NaOH were added to stop the reaction and the A at 410 nm was read immediately.
Percentage inhibition was calculated as follows: % inhibition = ((standard A - blank A ) - ( A with inhibitor - blank A))/(standard A - blank A) .
Calculation of AAC"The standard formula for AAG" = 273Rln ( x / y ) (15, 40, 41) is used; x is the amount of inhibitor in nanomoles giving 50% inhibition at 0 "C and y the amount of A- or of B- trisaccharide giving 50% inhibition for each monoclonal.
RESULTS
Hemagglutination Assays-Table I1 gives the hemagglutin- ation titers of the various anti-A, anti-B, and anti-AB mono- clonals. The titers of four anti-A monoclonals toward AI and A2 erythrocytes only varied over a %fold range. With AH16, produced to human gastric cancer cells, the titer with Az erythrocytes was 16 times lower than A, erythrocytes. The anti-A and anti-B monoclonals were completely A- or B- specific even with the undiluted material. With the anti-AB monoclonal the titers toward AI, Az, and B erythrocytes were 2048, 1024, and 1024, respectively.
Quantitative Precipitin Studies-Fig. 1 shows the quanti- tative precipitin curves obtained with various blood group substances and the five monoclonal anti-A samples studied. All five monoclonals showed blood group A specificity and did not react with B, H, Le", and Leb substances. The blood group A substance from hog gastric mucosa and the A, substances from human ovarian cyst fluids reacted strongly but tended to show relatively small differences in reactivity per micro- gram of A substance added. With AC-1001 and A15/3D3.92.1, the substances tend to fall into three groups but the members of each group differ. The most potent (group 1) in precipitat- ing were cyst 9, MSM lo%, MSS 10% 2X with both; Hog 4 10% was in group 1 with the former and Hog 75 10% in group 1 with latter monoclonal. The differences between groups 1 and 2 were about 25%. With A003=40/5G7, the distinction between the groups was not as clear but MSM 10% and Hog 4 10% reacted slightly better than the others, and with MH2/ 6D4 and AH16 the two groups were not seen.
The most striking finding was the reaction of the monoclo- nals with human blood group A2 substances. With AC-1001, A003=40/5G7, MH2/6D4, and AH16, Az substance phenol- insoluble cyst 14 was considerably more potent than saliva Az
TABLE I1 Hemagglutination titers of monoclonal mouse anti-A, anti-B, and
anti-AB Titers with human erythrocytes"
Monoclonal Antibody
Anti A A1 A2 B
AC-1001 2,048 1,024 0 A15/3D3.92.1 8,192 4,096 MH2/6D4
0 2,048 1,024 0
A003=40/5G7 16,384 16,384 0 AH16 256 16 0
Anti-B BC-1004 0 0 16,384 NB10/5B2.4.3 0 0 1,024 B003=46/2D7 0 0 8,192
A15/1A4.7.3 2,048 1,024 1,024 Anti-AB
a Highest dilution giving definite agglutination.
blood group substance phenol-insoluble W. G. and, with the first three monoclonals, phenol-insoluble cyst 14 was as po- tent as the less strongly reactive A, substances; with AH16 it was considerably less active. However, anti-A A15/3D3.92.1 reacted considerably less well with Az than with AI substances, and both A2 substances, phenol-insoluble cyst 14 and phenol- insoluble W. G., gave indistinguishable quantitative precipitin curves and showed about 30 and 40% of the activity of the group 1 and group 2 substances, respectively.
With the anti-AB monoclonal, A15/1A4.7.3, the precipitin curves were completely different (Fig. 2). In order of decreas- ing activity, the A substances were Hog 4 lo%, MSM lo%, MSS 10% 2X, Hog 75 lo%, phenol-insoluble McDon 15%, and phenol-insoluble cyst 9; among B substances Horse 4 25% was the only B substance tested which gave any precip- itation; phenol-insoluble Tij 11, phenol-insoluble PM, and phenol-insoluble Beach did not react. A15/1A4.7.3 did not precipitate with A, substances, phenol-insoluble cyst 14 and phenol-insoluble W. G. Each of the blood group substances precipitated a different amount of AbN at the maximum ranging from 5.5 to 3.0 pg of total N precipitated.
With all monoclonals, the amount of N precipitated re- mained at the maximum once reached, throughout the entire range studied, except for the reaction of Hog 75 10% and phenol-insoluble cyst 14 with A003=40/5G7 with which an inhibition zone in antigen excess was seen. With AC-1001, Hog 75 10% precipitated only 5 pg of N at the maximum, whereas the others ranged between 5.8 and 6.3 pg of N.
Quantitative precipitin studies could only be carried out on two of the three anti-B monoclonals since NB10/5B2.4.5 did not precipitate with B substances. The two other anti-B monoclonals, BC-1004 and B003=46/2D7, were B-specific (Fig. 2) in that they did not react with A, H, Le", or Leb substances. Typical precipitin curves were obtained with hu- man ovarian cyst B substances, phenol-insoluble Tij I1 and phenol-insoluble Beach, with human saliva B, phenol-insol- uble PM and Horse 4 25%. Precipitin curves of all four substances fell into a narrow range with BC-1004 but differed considerably with B003=46/2D7, the most active being about 3-fold better than the least potent. All B substances precipi- tated comparable amounts of total N at the maximum within experimental error.
Water-soluble Cow 35 of 10% ppt did not precipitate with anti-AB A15/1A4.7.3, nor with the two anti-B ascites; it reacted as well as the Az substances with A15/3D3.92.1 and A003=40/5G7 and less strongly than Az substances with the other anti-A ascites.
Quantitative Inhibition Assays-Quantitative inhibition as- says by ELISA showed differences in fine structure among anti-A combining sites (Fig. 3). Inhibition assays with AH16 were carried out with alkaline phosphatase-labeled protein A (22). The five anti-A sera showed different patterns; AC-1001 and MH2/6D4 were identical in order of reactivity with the A-heptasaccharide being best followed by the A-penta-, the A-hexa-, the A-tetrasaccharide, and MSMA3RL0.56 which were equal, and by 01 and the A-trisaccharide. The relative inhibiting powers of the other oligosaccharides to the A- heptasaccharide differed; with AC-1001 the hepta-, penta-, and hexasaccharides were very close, the three differing over less than a 2-fold range, whereas with MH2/6D4 the A- heptasaccharide was three times more potent per nanomole than the A-pentasaccharide and six times better than the A- hexasaccharide. MsMAzR~0.56 and the A-tetrasaccharide were 0.3 as active as the A-heptasaccharide with AC-1001 but only 1/10 as potent in MH2/6D4. The ratios of nanomoles required for 50% inhibition of MsMAsR~0.56 and the A-
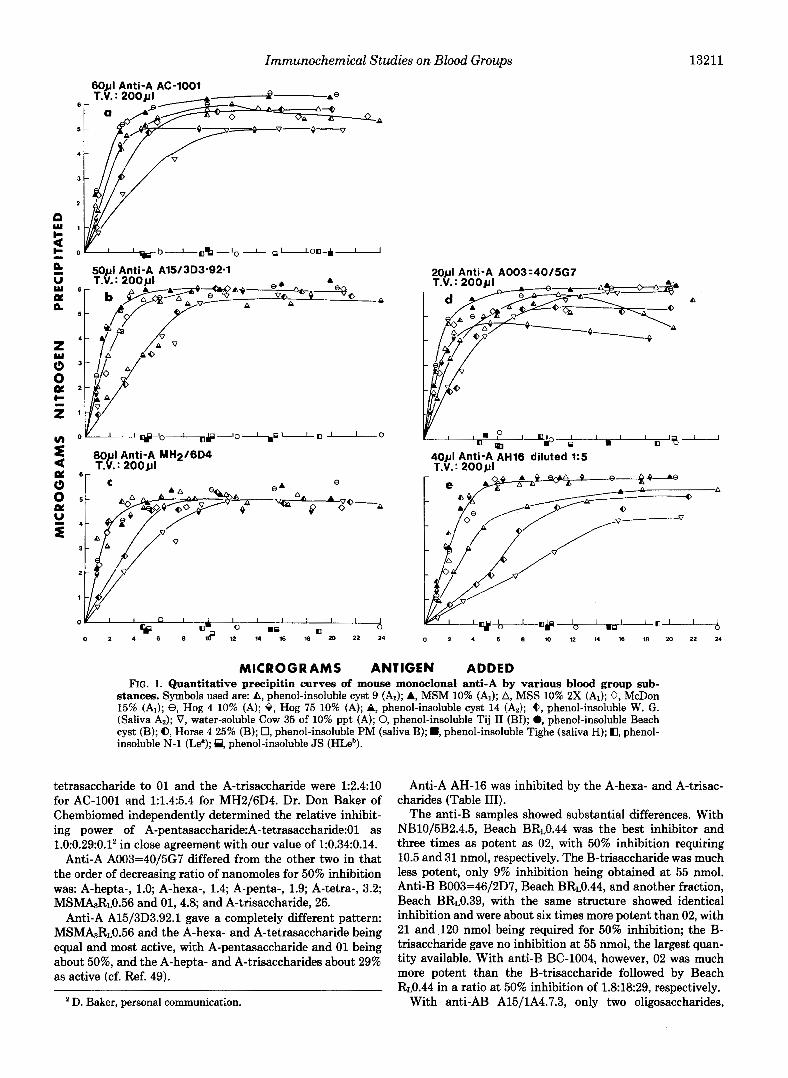
Immunochemical Studies on Blood Groups 13211
E
-A
I 1 q p " 0 ~ 0 ~ " - ' 0 + 0 " D -0
80pl Anti-A MH2/6D4 T.V.: 2 0 0 ~ 1
r
401.11 Anti-A AH16 diluted 1:5 T.V. : 2 0 0 ~ 1
I l o = ? I a p I , I I I~"..A
0 2 4 6 B l0 12 14 16 18 20 22 24
tetrasaccharide to 01 and the A-trisaccharide were 1:2.4:10 for AC-1001 and 1:1.4:5.4 for MH2/6D4. Dr. Don Baker of Chembiomed independently determined the relative inhibit- ing power of A-pentasaccharide:A-tetrasaccharide:Ol as 1.0:0.29:0.12 in close agreement with our value of 1:0.34:0.14.
Anti-A A003=40/5G7 differed from the other two in that the order of decreasing ratio of nanomoles for 50% inhibition was: A-hepta-, 1.0; A-hexa-, 1.4; A-penta-, 1.9; A-tetra-, 3.2; MSMA3R~0.56 and 01, 4.8; and A-trisaccharide, 26.
Anti-A A15/3D3.92.1 gave a completely different pattern: MSMA3RL0.56 and the A-hexa- and A-tetrasaccharide being equal and most active, with A-pentasaccharide and 01 being about 50%, and the A-hepta- and A-trisaccharides about 29% as active (cf. Ref. 49).
A
MICROGRAMS ANTIGEN ADDED FIG. 1. Quantitative precipitin curves of mouse monoclonal anti-A by various blood group sub-
stances. Symbols used are: A, phenol-insoluble cyst 9 (AI); A, MSM 10% (AI); A, MSS 10% 2X (A1); 0, McDon 15% (A1); 8, Hog 4 10% (A); 8, Hog 75 10% (A); A, phenol-insoluble cyst 14 (Az); 6, phenol-insoluble W. G. (Saliva A2); V, water-soluble Cow 35 of 10% ppt (A); 0, phenol-insoluble Tij I1 (BI); 0, phenol-insoluble Beach cyst (B); 0, Horse 4 25% (B); 0, phenol-insoluble PM (saliva B); B, phenol-insoluble Tighe (saliva H); CI, phenol- insoluble N-1 (Le"); @, phenol-insoluble JS (HLeb).
D. Baker, personal communication.
Anti-A AH-16 was inhibited by the A-hexa- and A-trisac- charides (Table 111).
The anti-B samples showed substantial differences. With NB10/5B2.4.5, Beach BR~0.44 was the best inhibitor and three times as potent as 02, with 50% inhibition requiring 10.5 and 31 nmol, respectively. The B-trisaccharide was much less potent, only 9% inhibition being obtained at 55 nmol. Anti-B B003=46/2D7, Beach BRL0.44, and another fraction, Beach BR~0.39, with the same structure showed identical inhibition and were about six times more potent than 02, with 21 and 120 nmol being required for 50% inhibition; the B- trisaccharide gave no inhibition at 55 nmol, the largest quan- tity available. With anti-B BC-1004, however, 02 was much more potent than the B-trisaccharide followed by Beach R~0.44 in a ratio at 50% inhibition of 1.81829, respectively.
With anti-AB A15/1A4.7.3, only two oligosaccharides,
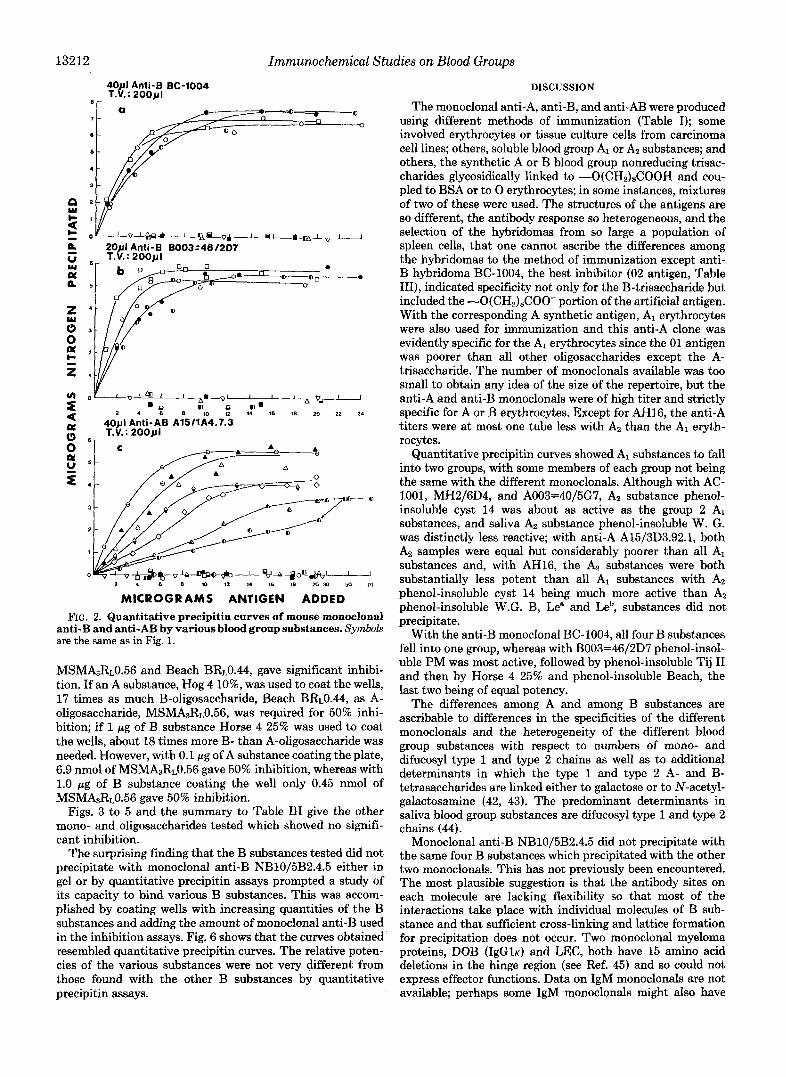
13212 Immunochemical Studies on Blood Groups
40411 Anti-B BC-1004 T.V. : 20041 I
= a
4 0 ~ 1 Anti-AB A15tlA4.7.3 T.V.: 2 0 0 ~ 1
*-VI I ' ' B-"' i a 0 . 2 4 6 8 10 I2 I4 16 18 20 22 24
MICROGRAMS ANTIGEN ADDED FIG. 2. Quantitative precipitin curves of mouse monoclonal
anti-B and anti-AB by various blood group substances. Symbols are the same as in Fig. 1.
MsMbR~0.56 and Beach BR~0.44, gave significant inhibi- tion. If an A substance, Hog 4 lo%, was used to coat the wells, 17 times as much B-oligosaccharide, Beach BRt0.44, as A- oligosaccharide, MSMA3R~0.56, was required for 50% inhi- bition; if 1 pg of B substance Horse 4 25% was used to coat the wells, about 18 times more B- than A-oligosaccharide was needed. However, with 0.1 pg of A substance coating the plate, 6.9 nmol of MSMA3R~0.56 gave 50% inhibition, whereas with 1.0 pg of B substance coating the well only 0.45 nmol of MSMA3RL0.56 gave 50% inhibition.
Figs. 3 to 5 and the summary to Table I11 give the other mono- and oligosaccharides tested which showed no signifi- cant inhibition.
The surprising finding that the B substances tested did not precipitate with monoclonal anti-B NBlOj5B2.4.5 either in gel or by quantitative precipitin assays prompted a study of its capacity to bind various B substances. This was accom- plished by coating wells with increasing quantities of the B substances and adding the amount of monoclonal anti-B used in the inhibition assays. Fig. 6 shows that the curves obtained resembled quantitative precipitin curves. The relative poten- cies of the various substances were not very different from those found with the other B substances by quantitative precipitin assays.
DISCUSSION
The monoclonal anti-A, anti-B, and anti-AB were produced using different methods of immunization (Table I); some involved erythrocytes or tissue culture cells from carcinoma cell lines; others, soluble blood group A, or A, substances; and others, the synthetic A or B blood group nonreducing trisac- charides glycosidically linked to -O(CH2)sCOOH and cou- pled to BSA or to 0 erythrocytes; in some instances, mixtures of two of these were used. The structures of the antigens are so different, the antibody response so heterogeneous, and the selection of the hybridomas from so large a population of spleen cells, that one cannot ascribe the differences among the hybridomas to the method of immunization except anti- B hybridoma BC-1004, the best inhibitor (02 antigen, Table III), indicated specificity not only for the B-trisaccharide but included the -O(CH,),COO- portion of the artificial antigen. With the corresponding A synthetic antigen, A, erythrocytes were also used for immunization and this anti-A clone was evidently specific for the AI erythrocytes since the 01 antigen was poorer than all other oligosaccharides except the A- trisaccharide. The number of monoclonals available was too small to obtain any idea of the size of the repertoire, but the anti-A and anti-B monoclonals were of high titer and strictly specific for A or B erythrocytes. Except for AH16, the anti-A titers were at most one tube less with A, than the AI eryth- rocytes.
Quantitative precipitin curves showed AI substances to fall into two groups, with some members of each group not being the same with the different monoclonals. Although with AC- 1001, MH2/6D4, and A003=40/5G7, Az substance phenol- insoluble cyst 14 was about as active as the group 2 A, substances, and saliva A, substance phenol-insoluble W. G. was distinctly less reactive; with anti-A A15/3D3.92.1, both A, samples were equal but considerably poorer than all A, substances and, with AH16, the A2 substances were both substantially less potent than all AI substances with A$ phenol-insoluble cyst 14 being much more active than Az phenol-insoluble W.G. B, Le" and Leb, substances did not precipitate.
With the anti-B monoclonal BC-1004, all four B substances fell into one group, whereas with B003=46/2D7 phenol-insol- uble PM was most active, followed by phenol-insoluble Tij I1 and then by Horse 4 25% and phenol-insoluble Beach, the last two being of equal potency.
The differences among A and among B substances are ascribable to differences in the specificities of the different monoclonals and the heterogeneity of the different blood group substances with respect to numbers of mono- and difucosyl type 1 and type 2 chains as well as to additional determinants in which the type 1 and type 2 A- and B- tetrasaccharides are linked either to galactose or to N-acetyl- galactosamine (42, 43). The predominant determinants in saliva blood group substances are difucosyl type 1 and type 2 chains (44).
Monoclonal anti-B NB10/5B2.4.5 did not precipitate with the same four B substances which precipitated with the other two monoclonals. This has not previously been encountered. The most plausible suggestion is that the antibody sites on each molecule are lacking flexibility so that most of the interactions take place with individual molecules of B sub- stance and that sufficient cross-linking and lattice formation for precipitation does not occur. Two monoclonal myeloma proteins, DOB (IgGlK) and LEC, both have 15 amino acid deletions in the hinge region (see Ref. 45) and so could not express effector functions. Data on IgM monoclonals are not available; perhaps some IgM monoclonals might also have
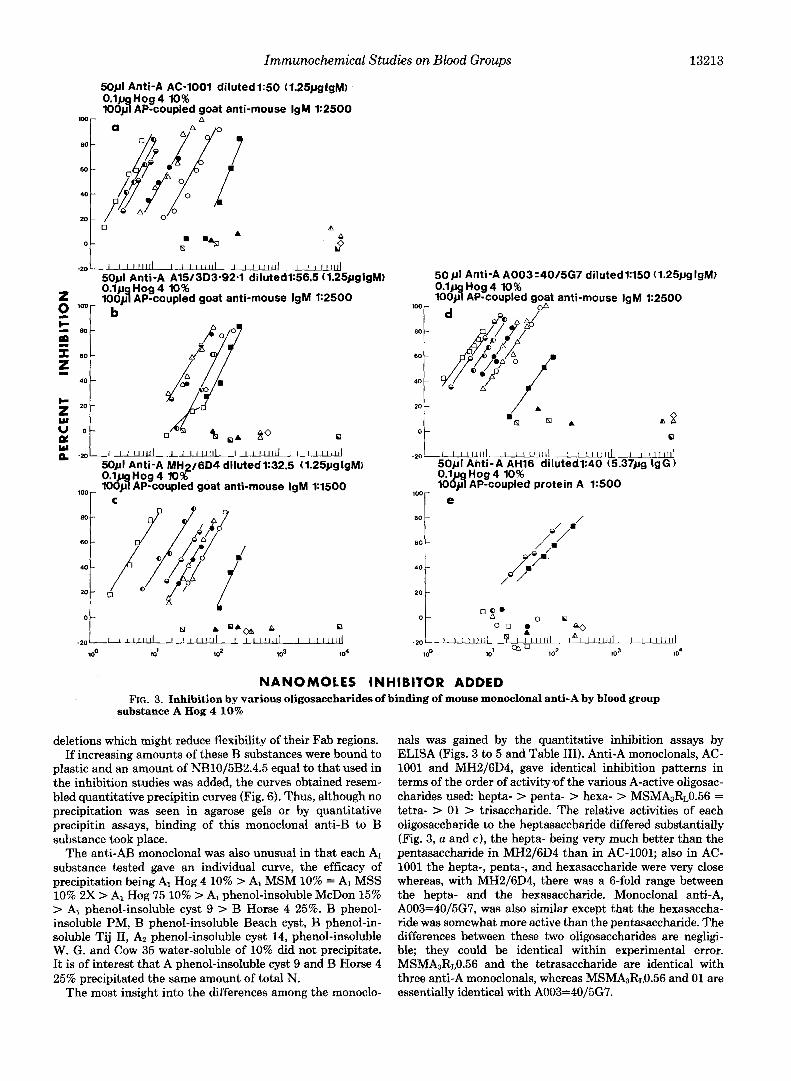
Immunochemical Studies on Blood Groups 13213
mor
80 -
W -
40 -
20 -
0 -
5 0 ~ 1 Anti-A AC-1001 dilutedl:50 (1.25pglgM)
1OOd AP-coupled goat anti-mouse IgM 1:2500 0 . 1 ~ Hog 4 10%
0 A A I9
-201 I L l l l l l f l I I 1 1 1 1 1 1 1 I I 1 1 1 1 1 1 1 I I l l l l u l
z 100yq AP-coupled goat anti-mouse IgM 1:2500
5 0 ~ 1 Anti-A A15/3D3-92.1 dilutedl:56.5 (1.25pglgM) 0 . 1 ~ Hog4 10%
2 loor b
2 Z
6 0 -
I
40 -
5 2 0 -
s W u 0 -
0 -20-
100 r
80 -
60 -
40 -
20 -
I I 1 1 1 1 1 1 1 I I 1 1 1 1 1 1 1 I I I 1 1 1 1 1 1 I l 1 1 1 1 1 1 ~
5 0 ~ 1 Anti-A MH216D4 dilutedt32.5 (1.25~gIgM) loa AP-coupled goat anti-mouse IgM 1:1500 0.1 Hog4 10%
C
5opl Anti-A A003=40/5G7 diluted1:ltiO (1.25~gIgM) 0 . 1 ~ Hog 4 10% 1OOd AP-coupled goat anti-mouse IgM 1:2500
1 W r . na
A B
O t 0
- 2 0 0 5 0 ~ 1 Anti-A AH16 dilutedl:40 (5.37119 IgG) 0.lua Hoa 4 10%
100- lOdy1 APrcoupled protein A 1:500 e
80 -
60
40
- &' - 4.y
20 -
/
/
0 - 0 2 . 0 0
0 0 A 0
-20 I I l I l l l l / B I 1 1 1 1 1 1 1 I*l 1 1 1 1 1 1 1 I I1111111
1 00 IO1 102 lo3 lo4
NANOMOLES INHIBITOR ADDED FIG. 3. Inhibition by various oligosaccharides of binding of mouse monoclonal anti-A by blood group
substance A Hog 4 10%
deletions which might reduce flexibility of their Fab regions. If increasing amounts of these B substances were bound to
plastic and an amount of NB10/5B2.4.5 equal to that used in the inhibition studies was added, the curves obtained resem- bled quantitative precipitin curves (Fig. 6). Thus, although no precipitation was seen in agarose gels or by quantitative precipitin assays, binding of this monoclonal anti-B to B substance took place.
The anti-AB monoclonal was also unusual in that each A, substance tested gave an individual curve, the efficacy of precipitation being AI Hog 4 10% > A1 MSM 10% = A, MSS 10% 2X > AI Hog 75 10% > A1 phenol-insoluble McDon 15% > A1 phenol-insoluble cyst 9 > B Horse 4 25%. B phenol- insoluble PM, B phenol-insoluble Beach cyst, B phenol-in- soluble Tij 11, A2 phenol-insoluble cyst 14, phenol-insoluble W. G. and Cow 35 water-soluble of 10% did not precipitate. It is of interest that A phenol-insoluble cyst 9 and B Horse 4 25% precipitated the same amount of total N.
The most insight into the differences among the monoclo-
nals was gained by the quantitative inhibition assays by ELISA (Figs. 3 to 5 and Table 111). Anti-A monoclonals, AC- 1001 and MH2/6D4, gave identical inhibition patterns in terms of the order of activity.of the various A-active oligosac- charides used: hepta- > penta- > hexa- > MSMhRL0.56 = tetra- > 01 > trisaccharide. The relative activities of each oligosaccharide to the heptasaccharide differed substantially (Fig. 3, a and e), the hepta- being very much better than the pentasaccharide in MH2/6D4 than in AC-1001; also in AC- 1001 the hepta-, penta-, and hexasaccharide were very close whereas, with MH2/6D4, there was a 6-fold range between the hepta- and the hexasaccharide. Monoclonal anti-A, A003=40/5G7, was also similar except that the hexasaccha- ride was somewhat more active than the pentasaccharide. The differences between these two oligosaccharides are negligi- ble; they could be identical within experimental error. MSMA3R~0.56 and the tetrasaccharide are identical with three anti-A monoclonals, whereas MsMAsR~0.56 and 01 are essentially identical with A003=40/5G7.

13214 Immunochemical Studies on Blood Groups TABLE I11
Oligosaccharides active in inhibition assays by ELISA (see Figs. 3-5) The following mono- and oligosaccharides gave no significant inhibition with the anti-A monoclonals: 0, D-
GalNAc; A, D-GlcNAc; A, L-FUC; O, D-Gal; A, RL1.34 [GalNAcd4Gal];N, AJI (GalNAcoll43Gal~l~3GlcNAc). The anti-B monoclonals were not inhibited by: 0, D-Gak 0, D-G~NAc; A, D-GlcNAc; A, L-FUC; V, galactinol; V, melibiose; V, methyl-a-galactoside; V, Beach P1Rt0.62 ( G a l a l ~ 3 G a l ~ l 4 G l c N A c - R ) . The anti-AB monoclonal was not inhibited by: 0, D-GalNAc; 0, D - G ~ , A, D-GlcNAc; A, L-FUC; A, RJ.34; El, A& ., A-trisaccharide; 0, 01; e, A-tetrasaccharide; @, A-pentasaccharide; 0, A-hexasaccharide; 0, A-heptasaccharide; V, galactinol; V, Beach P1R~0.62; V, B-trisaccharide; A, 02; 0, B-pentasaccharide.
50 Percent Inhibition (nanomoles) Anti-A
AAG- [Kcallmole) SYm- bo1 Oligosaccharide Structure
A15/3D3. A15/303. I “””“” AC-1001 92.1 M 2 / 6 W A003=40/567 AH16 AC-1001 92.1 Mi2/6M A003=40/5G7 AH16
A-Tri
0 01
A-Tetra
A MSM3R~fl.56
% (> A-Penta
9 A-Hexa
a A-Hepta
GalNAcal+3Gal +: Fuca
GalNAcol+3Gale-O(CH2)8-COOCH3 t 2 1 Fuca
GalNAcd+3Ga1~1+46lc t 2 Fuca
1
GalNACal+3G~lBl+4GlcNAcg-R
1 Fuco t 2
GalNAcol+3Galel+4Glc t: t ;
F U C ~ FUCO
GdlNPc~l+3GalEl+3GlCNAC61+3Gal61~GlC + 2 1 Fuc.
GalNAc~l+3Galal+3GlcNAcsl,3Gal81+4Glc t; t;
FUCh Fuco
132 125 245 80 en o D 0 0 0
33 82.5 63 14.8 -29% -0.75 -0.22 -0.73 -0.91 inactive (29 nmoles)
13.5 36 45.5 9.8 - 7.8 -1.23 -0.67 -0.91 -1.13 inactive (42 nnwles)
13.5 36 45.5 14.8 -31% -1.23 -0.67 -0.91 -0.91 inactive (26 nmoles)
4.6 82.5 15.8 6 - 0 . 4 -1.81 -0.22 -1.48 -1.4 inactive (62 nmolea)
5.9 36 30 4.2 46 -1.68 -0.67 -1.14 -1.59 -0.3
3.7 2(pd 4.8 3.1 - 6.0 -1.93 >+0.10 -2.12 -1.76 inactive (60 nmoles) (21 nmoles)
Anti-B 50 Percent Inhibition (nanomoles) &AG’ (Kcalfmolej
NElOl E003. NBlOl tlOO3= BC-1OW 582.4.5 46/207
-” EC-1004 582.4.5 46/2[37
”-
0 91.24 inactive
1.G 31 120 -1.23 0 0
29 10.5 ?1 +0.26 - 0.59 -0.94
6% -1% -10.2% (79.5 (79.5 (79.5 nmoles) nmoles) nmoles]
>*1.95 inactive inactive
50 Percent Inhibition (nanomoles) Anti-AB
b b t (Kcallmole) -ATS/lA4. A1511A4.
(A anti-A) (8 anti-E) 7.3 7.3 7.3 7.3
Al511A4. A15I114.
(A anti-A) ( E anti-B) ” _______
GalNAc~l+3Ga1R1+4GlcNAcs-R 6.9 0.45 +: -1.52 -1.56
Fuca
Gdlo1+3Ga1B1+4G1cNAcB”
t: Fuca
115 8 0 0
The fourth anti-A monoclonal, A15/3D3.92.1, was quite clonal anti-A, TL5, studied by Gooi et al. (46), which reacts different from three others, MSMA3RL0.56, A-hexa-, and A- with the receptor for epidermal growth factor of human epi- tetrasaccharide, which were most potent and equal as inhibi- dermoid carcinoma cell line A431. AH16 differed from the tors followed by the A-pentasaccharide and 01, which were others in that it was inhibited only by the two oligosaccharides also identical, with the A-trisaccharide being somewhat less satisfying the monofucosyl type 1 structure (22) of the A active; the A-heptasaccharide was about as active as the A- glycolipid from the cancer cells used for immunization. Thus, trisaccharide. These data are not too different from the mono- one may conclude that all of the six monoclonal hybridoma
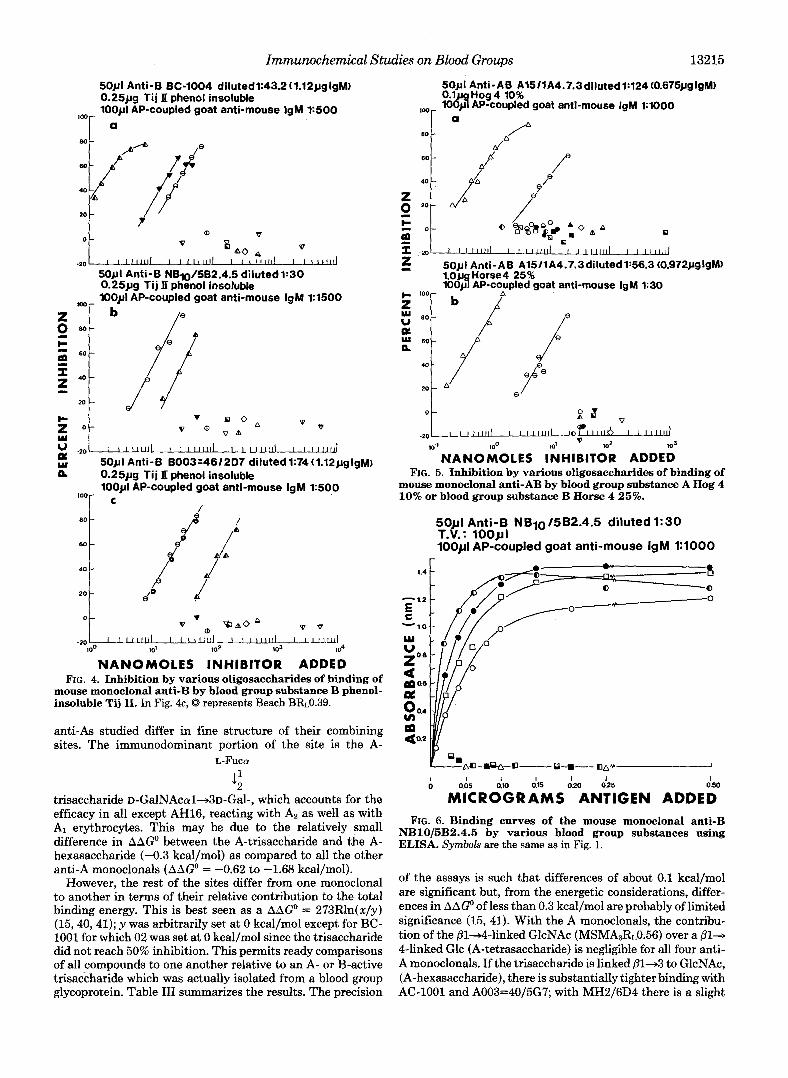
Immunochemical Studies on Blood Groups 13215
50pl Anti-B BC-1004 dilutedl:43.2(1.12~~gIgM) 0.25~19 Tij II phenol insoluble l0Opl AP-coupled goat anti-mouse IgM 1x500
ID0 - a
80 - PA / e
20 - 7
0 'A0 0 v
SOpI Anti-B NB10/5B2.4.5 dilutedl:30 0.25~19 Ti j It phenol insoluble 100~11 AP-coupled goat anti-mouse IgM 1:1500
I I l l l l l l l I I111111l I I 1 1 1 1 1 1 I I I I I I ld
b
z' ot I l 1 1 1 1 1 ( I 1 1 1 1 1 1 1 I I 1 1 1 1 1 1 1 I I 1 1 1 1 1 ~ YI
v -20
50pl Anti-B B003:46/207 dilutedl:74(1.12pgIgM) 0.25~19 Tij II phenol insoluble lWpl AP-coupled goat anti-mouse IgM 1:500 loor c
/
0 - 0 'I % A O A o v
-20 I I 1 1 1 1 ( 1 1 1 1 1 1 1 1 1 1 1 1 1 1 I I I l l d 100 10' 102 lo3 lo4
NANOMOLES INHIBITOR ADDED FIG. 4. Inhibition by various oligosaccharides of binding of
mouse monoclonal anti-B by blood group substance B phenol- insoluble Tij 11. In Fig. 4c, 0 represents Beach BR~0.39.
anti-As studied differ in fine structure of their combining sites. The immunodominant portion of the site is the A-
L-FUCOI
4 trisaccharide ~-GalNAcal+3~-Gal-, which accounts for the efficacy in all except AH16, reacting with A2 as well as with AI erythrocytes. This may be due to the relatively small difference in AAGO between the A-trisaccharide and the A- hexasaccharide (-0.3 kcal/mol) as compared to all the other anti-A monoclonals (AAGo = -0.62 to -1.68 kcal/mol).
However, the rest of the sites differ from one monoclonal to another in terms of their relative contribution to the total binding energy. This is best seen as a AAGO = 273Rln(x/y) (15,40,41); y was arbitrarily set at 0 kcal/mol except for BC- 1001 for which 02 was set at 0 kcal/mol since the trisaccharide did not reach 50% inhibition. This permits ready comparisons of all compounds to one another relative to an A- or B-active trisaccharide which was actually isolated from a blood group glycoprotein. Table I11 summarizes the results. The precision
50pI Anti-AB A1511A4.7.3dilutedl:124(0.675pgIgM) 01 Hog4 10% lo&! AP-coupled goat anti-mouse IgM 1:lOOO
I D 0 r-
A /"
5 50pl Anti-AB A15/1A4.7.3dilutedl:56.3.(0.972pglgM) l o p Horse4 25% iooJ AP -coupled goat anti-mouse IgM 1:30
P
cy
r. "* / 20
-20 lllllllliliilllllll;& 10-1 100 101 102 lo3
NANOMOLES INHIBITOR ADDED FIG. 5. Inhibition by various oligosaccharides of binding of
mouse monoclonal anti-AB by blood group substance A Hog 4 10% or blood group substance B Horse 4 25%.
50~11 Anti-B NB10/582.4.5 diluted 1:30 T.V. : 100~11 100~11 AP-coupled goat anti-mouse IgM 1:lOOO
, I 1 I I I I 010 0.15 0.20 0.25 0.50
O " ~ R O G R A M S ANTIGEN ADDED FIG. 6. Binding curves of the mouse monoclonal anti-B
NB1015B2.4.5 by various blood group substances using ELISA. Synbok are the same as in Fig. 1.
of the assays is such that differences of about 0.1 kcal/mol are significant but, from the energetic considerations, differ- ences in AAG' of less than 0.3 kcal/mol are probably of limited significance (15, 41). With the A monoclonals, the contribu- tion of the PlA-linked GlcNAc (MSMA3R~0.56) over a ,8l+ 4-linked Glc (A-tetrasaccharide) is negligible for all four anti- A monoclonals. If the trisaccharide is linked Pl+3 to GlcNAc, (A-hexasaccharide), there is substantially tighter binding with AC-1001 and A003=40/5G7; with MH2/6D4 there is a slight

13216 Immunochemical Studies on Blood Groups
increase in binding, and with A15/3D3.92.1 there is no differ- ence.
The contribution of the second fucose, if linked a143 t o the type 2 chain, was substantial by about 0.5 kcal/mol with AC-1001, MH2/6D4, and A003=40/5G7, but in A15/3D3.92.1 it actually reduced binding affinity by about 0.45 kcal/mol. If the second fucose was linked a 1 4 to the type 1 chain, with MH2/6D4 the increase in binding was about 1 kcal/mol, with AC-1001 and it was only about 0.2 kcaljmol and, with A15/ 3D3.92.1, binding affinity was essentially minimal, 50% bind- ing not being reached. The AAGO values of the A-hepta-, A- penta-, and A-hexasaccharide for AC-1001 are very close.
The inhibition data on A15/3D3.92.1 clearly indicate that the site is most specific for at least a monofucosyl tetrasac- charide GalNAcal+3Gal@l4GlcNAc@- structure with the
t 2 1
FUCCX- difucosyl compound blocking entry into the site, whereas the other three anti-A monoclonals are most specific for a difu- cosy1 oligosaccharide. MH2/6D4 and A003=40/5G7 clearly involve the type 1 difucosyl pentasaccharide; with AC-1001 it is not clear whether the specificity is for the difucosyl type 1 (A-pentasaccharide) or type 2 (A-heptasaccharide) oligosac- charides. Thus, although there has been a tendency to accept the monofucosyl type 1 and type 2 oligosaccharides as the A determinants (9, 10) and essentially to ignore the difucosyl determinants, three of the four monoclonals turn out to have a combining site most complementary to difucosyl moieties.
Two of the three anti-B monoclonals, NB10/5B2.4.5 and B003=46/2D7, clearly do not have difucosyl type 2 specificity since these compounds were inactive compared to the mono- fucosyl type 2 oligosaccharides; in both instances maximum amounts of the B-trisaccharide used did not give 50% inhi- bition; this level was attained by oligosaccharide 02 and is perhaps ascribable to the additional @-linkage to the -O(CH2)sCOOCH3 aglycone with or without the aglycone making an additional hydrophobic contribution to binding. The monofucosyl type 2 oligosaccharide, Beach BR~0.44, was considerably more active. BC-1004 was unusual in that the 02 was -1.2 kcal/mol more negative than the B-trisaccharide, establishing that a substantial portion of the binding energy was directed toward the aglycone and that its antibody com- bining site was larger than the trisaccharide as would be expected since 02, with its carboxyl coupled to 0 erythrocytes or to bovine albumin, had been used for immunization. That AAGO of the mono- and difucosyl type 2 oligosaccharides were much less potent suggests that the combining site of BC-1004 was most specific for the B-trisaccharide plus the @-linkage and that the aglycone-specific part of its site could not accom- modate B determinants larger than the trisaccharide. Despite this, it is a highly satisfactory blood group reagent. Lemieux et aL3 have studied two IgM affinity purified hybridoma anti- B monoclonals from clones 3E-4 and A1-2, produced by immunization with the same B-trisaccharide antigen as BC-1004. As with BC-1004, attaching the aglycone -(CH2)sCOOCH3 made AAGO substantially more negative, but if the same aglycone was linked B to the B-tetrasaccharide in which the B-trisaccharide was linked p l 4 to a D-GlcNAc, the AAGO became more than 1 kcal/mol more positive, indi- cating that the combining site could not accommodate a D- GlcNAc in the position occupied by the aglycone in the trisaccharide aglycone, a finding in complete agreement with
3R. U. Lemieux, A. P. Venot, U. Spohr, P. Bird, G. Mandal, N. Morishima, 0. Hindsgaul, and D. R. Bundle, manuscript in prepara- tion.
our data on BC-1004. Thus, these two monoclonals, like BC- 1004, have a site complementary to the trisaccharide glyco- sidically linked to -O(CH2)sCOO-.
The AAGo of the anti-AB monoclonal, A15/1A4.7.3, is quite interesting. In inhibition assays of A anti-A and B anti-B precipitation, if the B monofucosyl type 2 oligosaccharide is assigned a AAGO of 0, the A monofucosyl type 2 oligosaccha- ride has a AAGO of -1.52 and -1.56 kcal/mol, respectively. Since it was not inhibited by the A-tetra-, A-penta-, A-hexa-, A-hepta-, and B-pentasaccharides (see summary to Table 111), it is clearly specific for the monofucosyl type 2 structure. Unfortunately, no monofucosyl type 1 oligosaccharide was available for comparison.
The present study clearly indicates that anti-A and anti-B sera are heterogeneous populations of antibody molecules and that monofucosyl as well as difucosyl type 1 and type 2 chains can function effectively as A and probably as B determinants. Moreover, the A and B type 2 monofucosyl tetrasaccharide determinants in which the A- and B-trisaccharides are linked @l-& to Gal or to GalNAc and the contribution of antibodies specific for these moieties to the total anti-A and anti-B repertoire require further examination when oligosaccharides become available. Study of additional anti-A monoclonals should resolve the question of the nature of the AI and Az determinants (47). Monoclonal antibodies recognizing the difucosyl and monofucosyl type 2 H, and the difucosyl type 1 Leb structures have been found (see Ref. 48).
Acknowledgment-We would like to thank Paula Borden for advice about the ELISA technics.
REFERENCES
1. Landsteiner, K. (1900) Zent. Bakteriol. Parasitenk. 2 7 , 357-362;
2. DeCastello, A. von, and Sturli, A. (1902) Munch. Med. Wochsehr.
3. Witebsky, E., Klendshoj, N. C., and McNeil, C. (1944) Proc. SOC.
4. Kabat, E. A., and Bezer, A. E. (1945) J. Exp. Med. 82,207-215 5. Bendich, A., Kabat, E. A., and Bezer, A. E. (1946) J. Exp. Med.
6. Lloyd, K. O., Kabat, E. A., and Rosenfield, R. E. (1966) Biochem-
7. Lloyd, K. O., and Kabat, E. A. (1968) Proc. Natl. Acad. Sei.
(1901) Wien Klin. Wochschr. 14,1132-1134
49,1090-1095
Expt. Biol. Med. 5 5 , 167-170
83,485-497
istry 5,1502-1507
U , S . A. 6 i , 1470-1477 8. Kabat. E. A. (19821 Am. J. Clin. Pathol. 78.281-292 9. Morgan, W. T. J. (1970) in British Biochemitry Past and Present.
(Goodwin, T. W., ed) Academic Press, New York 10. Watkins, W. M. (1980) Adu. Hum. Genet. 10, 1-136 11. Lundblad, A., and Kabat, E. A. (1971) J. Immunol. 1 0 6 , 1572-
12. Kabat, E. A., Liao, J., and Lemieux, R. U. (1978) Immunochem-
13. Kabat, E. A., Liao, J., Burzynska, M. H., Wong, T. C., Thglgersen,
14. Feizi, T., Childs, R. A., Watanabe, K., and Hakomori, S. (1979)
15. Lemieux, R. U., Wong, T. C., Liao, J., and Kabat, E. A. (1984)
16. Kohler, G., and Milstein, C. (1975) Nature 256 ,495497 17. Bundle, D. R., Gidney, M. A. J., Kassam, N., and Rahman, A. F.
18. Lowe, A. D., Lennox, E. S., and Voak, D. (1984) Vox Sang. 4 6 ,
19. Voak, D., Sacks, S., Alderson, T., Takei, F., Lennox, E., Jarvis, J., Milstein, C., and Darnborough, J. (1980) Vox Sang. 3 9 ,
20. Voak, D., Lowe, A. D., and Lennox, E. (1983) Biotest Bull. 4,
21. Messeter, L., Brodin, T., Chester, M. A., Low, B., and Lundblad,
1577
istry 15,727-731
H., and Lemieux, R. U. (1381) Mol. Immunol. 18,873-881
J. Exp. Med. 149,975-980
Mol. Immurwl. 21,751-759
R. (1982) J. Immurwl. 129,678-682
29-35
134-140.
291-299

Immunochemical Studies on Blood Groups 13217
A. (1984) Vox Sang. 45, 185-194 22. Abe, K., Levery, S. B., and Hakomori, S.-I. (1984) J. Zmmunol.
23. Beiser, S. M., and Kabat, E. A. (1952) J. Zmmunol. 68, 19-40 24. Allen, P. Z., and Kabat, E. A. (1959) J. Zmmunol. 82, 340-357 25. Vicari, G., and Kabat, E. A. (1969) J. Zmmunol. 102,821-825 26. Lloyd, K. O., Kabat, E. A., and Licerio, E. (1968) Biochemistry
27. Maisonrouge-McAuliffe, F., and Kabat, E. A. (1976) Arch. Biochem. Biophys. 175,71-80
28. Lloyd, K. O., Kabat, E. A., Layug, E. J., and Gruezo, F. (1966) Biochemistry 5,1489-1501
29. Schiffman, G., Kabat, E. A., and Leskowitz, S. (1962) J. Am. Chem. Soc. 84,73-77
30. Schiffman, G., Kabat, E. A., and Thompson, W. (1964a) Biochem- istry 3,113-120
31. Schiffman, G., Kabat, E. A., and Thompson, W. (1964b) Biochem- istry 3,587-593
32. Newman, W., and Kabat, E. A. (1976) Arch. Biochem. Biophys. 172,535-550
33. Sabharwal, H., Chester, M. A., Sjoblad, S., and Lundblad, A. (1983) Glycoconjugates, Proceedings of The Symposium on Gly- coconjugates Ronneby, Sweden, July 17-23, pp. 221-222, Rahms, Lund, Sweden
132,1951-1954
7,2976-2990
34. Messeter, L. (1983) Ph.D. thesis, University of Lund, Sweden 35. Kabat, E. A. (1961) Kabat and Mayer’s Experimental Immuno-
chemistry, 2nd Ed., Charles C Thomas, Springfield, IL 36. Kabat, E. A. (1980) Methods Enzymol. 70,3-39
37. Schiffman, G., Kabat, E. A., and Thompson, W. (1964) Biochem-
38. Avrameas, S., Ternynck, T., and Guesdon, J.-L. (1978) Scand. J. Immunol. 8, Suppl. 7,7-23
39. Hammerling, G. J., Hammerling, U., and Kearney, J.-F. (eds) (1981) Monoclonal Antibodies and T-cell Hybridomas, pp. 514- 577, Elsevier, New York
i s t ~ 3,113-120
40. Kabat, E. A. (1956) J. Zmmunol. 77, 377-385 41. Baker, D. A., Sugii, S., Kabat, E. A., Ratcliffe, R. M., Hermentin,
42. Pereira, M. E. A., and Kabat, E. A. (1979) Crit. Rev. Zmmunol. 1,
43. Wu, A. M., Kabat, E. A., Nilsson, B., Zopf, D. A., Gruezo, F. G.,
44. Sakamoto, J., Yin, B. W. T., and Lloyd, K. 0. (1984) Mol.
45. Davies, D. R., and Metzger, H. (1983) Annu. Rev. Immunol. 1,
46. Gooi, H. C., Schlessinger, J., Lax, I., Yarden, Y., Libermann, T. A., and Feizi, T. (1983) Biosci. Rep. 3, 1045-1052
47. Kisailus, E. C., and Kabat, E. A. (1978) J. Exp. Med. 147, 830- 843
48. Lloyd, K. O., Larson, G., Stromberg, H., Thurin, J., and Karlsson, K.-A. (1983) Immunogenetics 17, 537-541
49. Gooi, H. C., Hounsell, E. F., Picard, J. K., Lowe, A., Voak, D., Lennox, E., and Feizi, T. (1985) J. Biol. Chem. 260, 13218- 13224
P., and Lemieux, R. U. (1983) Biochemistry 22,2741-2750
33-78
and Liao, J., (1984) J. Biol. Chem. 259, 7178-7186
Immunol. 21,1093-1098
87-117


















