
JBC Papers in Press. Published on April 18, 2002 as ... · PDF filegs/sap/PBS at 37°C, and...
42
Megalin Functions as an Endocytic Sonic Hedgehog Receptor Robert A. McCarthy 1 , Jeremy L. Barth 1 , Mastan R. Chintalapudi, Christian Knaak and W. Scott Argraves 2 Running title: Megalin as a N-Shh receptor Key words: gp330, LRP2, patched, zebrafish Medical University of South Carolina Department of Cell Biology 171 Ashley Avenue Charleston, SC 29425-2204 USA 1 These authors contributed equally to this work. 2 To whom all correspondence should be addressed. E-mail: [email protected] Copyright 2002 by The American Society for Biochemistry and Molecular Biology, Inc. JBC Papers in Press. Published on April 18, 2002 as Manuscript M201933200 by guest on April 16, 2018 http://www.jbc.org/ Downloaded from
-
Upload
hoangxuyen -
Category
Documents
-
view
214 -
download
0
Transcript of JBC Papers in Press. Published on April 18, 2002 as ... · PDF filegs/sap/PBS at 37°C, and...

Megalin Functions as an Endocytic Sonic Hedgehog Receptor
Robert A. McCarthy1, Jeremy L. Barth1, Mastan R. Chintalapudi, Christian Knaak
and W. Scott Argraves2
Running title: Megalin as a N-Shh receptor
Key words: gp330, LRP2, patched, zebrafish
Medical University of South Carolina
Department of Cell Biology
171 Ashley Avenue
Charleston, SC 29425-2204 USA
1These authors contributed equally to this work.
2To whom all correspondence should be addressed. E-mail: [email protected]
Copyright 2002 by The American Society for Biochemistry and Molecular Biology, Inc.
JBC Papers in Press. Published on April 18, 2002 as Manuscript M201933200 by guest on A
pril 16, 2018http://w
ww
.jbc.org/D
ownloaded from

2
Summary
Embryos deficient in the morphogen sonic hedgehog (Shh) or the endocytic receptor
megalin exhibit common neurodevelopmental abnormalities. Therefore, we have investigated
the possibility that a functional relationship exists between the two proteins. During embryonic
development megalin was found to be expressed along the apical surfaces of neuroepithelial
cells and was co-expressed with Shh in the ventral floor plate of the neural tube. Using ELISA,
homologous ligand displacement and surface plasmon resonance techniques it was found that
the amino-terminal fragment of Shh (N-Shh) bound to megalin with high affinity. Megalin
expressing cells internalized N-Shh through a mechanism that was inhibited by antagonists of
megalin, namely, RAP and megalin antibodies. Heparin also inhibited N-Shh endocytosis,
implicating proteoglycans in the internalization process as has been described for other megalin
ligands. Use of chloroquine to inhibit lysosomal proteinase activity showed that N-Shh
endocytosed via megalin was not efficiently targeted to the lysosomes for degradation. The
ability of megalin-internalized N-Shh to bypass lysosomes may relate to the finding that the
interaction between N-Shh and megalin was resistant to dissociation with low pH. Together
these findings show that megalin is an efficient endocytic receptor for N-Shh. Furthermore, they
implicate megalin as a new regulatory component of the Shh signaling pathway.
by guest on April 16, 2018
http://ww
w.jbc.org/
Dow
nloaded from

3
Introduction
Sonic hedgehog (Shh) is a secreted signaling molecule that is expressed in spatially
restricted patterns during embryonic development. Shh signaling has been shown to regulate a
wide range of developmental patterning events in Drosophila and vertebrates involved in lung
(1), nervous system (2), eye (3), midbrain (4), forebrain and facial morphogenesis (5, 6). During
early vertebrate development, Shh signaling at the midline leads to patterning of the ventral
neural tube and adjacent somites. Mice lacking Shh activity have anomalies of midline
structures such as the notochord and floorplate of the early brain (7). Later these mice display
an absence of ventral neuronal cells and cranial motor neurons (8). The result of errant Shh
signaling in humans has been directly linked to basal cell carcinoma (9, 10) and
holoprosencephaly (HPE) (11).
Post-translational modification of the 45 kDa Shh polypeptide produces an ~19-kDa
amino-terminal fragment, designated N-Shh which has palmitic acid and cholesterol moieties
covalently coupled to its amino and carboxy termini, respectively (12-14). N-Shh is secreted
and represents the biologically active form of the protein, capable of initiating signaling. The
current model for Shh signaling involves a pair of multiple-pass plasma membrane proteins,
patched (Ptc or Ptc-1) and smoothened (Smo) (reviewed in (15)): Ptc functions as the Shh-
binding subunit/receptor and Smo as the signal transducing subunit. When bound to Smo, Ptc
acts as a repressor of Smo signaling activity. Following N-Shh interaction with Ptc, bound Ptc
releases from Smo and de-represses the signaling activity of Smo. The expression of Ptc-1,
Gli-2, HNF3beta, Nkx2.2 and netrin-1 have been shown to be activated by Shh, and genes
including Pax-3, Gli-3 and ephrin A5 have been shown to be suppressed by Shh (16, 17).
by guest on April 16, 2018
http://ww
w.jbc.org/
Dow
nloaded from

4
Megalin (also known as gp330 and LRP-2) is an endocytic receptor belonging to the low
density lipoprotein (LDL) receptor family (18). The receptor is expressed on apical surfaces of
numerous epithelia where it functions to mediate endocytosis of ligands, targeting them for
lysosomal degradation or transcytosis (18). Mice deficient in the expression of megalin
demonstrate the critical neurodevelopmental role for this protein (19). These mice display
numerous craniofacial abnormalities including absence of olfactory bulbs, absence of the corpus
callosum and fusion of forebrain hemispheres, collectively an HPE phenotype (19). During
development, megalin-deficient embryos (9.5 dpc) have pronounced cell death in several
structures including cranial nerves, neural crest and the optic vesicle (19). The spectrum of
defects that constitute the megalin-deficient phenotype suggests that megalin expression is
required for normal viability of the neural epithelium at an early embryonic stage.
The phenotype of megalin-deficient mice suggests a role for megalin in regulating cell
fate specification in the patterning of the neural tube and is consistent with phenotypes
observed in mice deficient in Shh and the Shh signal transducer, Smo (8, 20). For example,
Shh-deficient embryos lack cranial motor neurons (8). Inhibition of Shh signaling in the neural
tube has been shown to result in extensive apoptosis of neural epithelial cells (21). Shh has
also been shown to regulate proliferation and inhibit differentiation of central nervous system
(CNS) precursor cells (22). Smo mutants also display neural tube-related defects including
increased apoptosis of cells within the neural tube, absence of secondary motor neurons,
synopthalmia and ventral forebrain defects (20, 23). The shared aspects of the megalin-, Shh-
and Smo-deficient phenotypes suggest that Shh and megalin impact common mechanisms that
underlie CNS development. Here we report findings from experiments directed at determining
whether a functional relationship exists between megalin and Shh.
by guest on April 16, 2018
http://ww
w.jbc.org/
Dow
nloaded from

5
Experimental Procedures
Cells. MSV-transformed Brown Norway rat yolk sac cells (BN cells) were provided by Dr. Pierre
Verroust (Hospital Tenon, Paris, France). Mouse embryonal teratocarcinoma F9 cells (ATCC
CRL1720) were differentiated by treatment with retinoic acid and dibutyryl cyclic AMP
(Bt2/cAMP) for 6 days as previously described (24). C3H10T1/2 (ATCC CCL-226) were obtain
from the American Type Culture Collection (Manassas, VA).
Antibodies. Rabbit polyclonal and mouse monoclonal antibodies to megalin, rb6286 and 1H2,
have been described previously (25). Rabbit anti-megalin IgGs were purified by protein-G-
Sepharose and megalin-Sepharose chromatography (26). Mouse monoclonal RAP antibody
7F1 has been described previously (27). Mouse monoclonal N-Shh antibody 5E1 IgG was
isolated from the conditioned culture medium of a hybridoma cell line obtained from the
Developmental Studies Hybridoma Bank (The Johns Hopkins University School of Medicine and
the University of Iowa). Goat anti-glutathione-S-transferase (GST) was obtained from
Amersham Pharmacia Biotech (Piscataway NJ). Fluorescein isothiocyanate (FITC) and
indocarbocyanine (Cy3) labeled secondary IgGs were purchased from Jackson
ImmunoResearch Labs, Inc. (West Grove, PA).
Proteins. Megalin was purified from porcine kidney as described previously (28). Human RAP
was expressed in bacteria and purified as described by Kounnas et al. (29). Recombinant
murine N-Shh (residues 25-198) was obtained from R&D Systems (Minneapolis, MN). A
plasmid construct was created to express GST-N-Shh fusion protein in bacteria. Briefly, this
involved using RT-PCR to generate a cDNA encoding amino acids 20-198 of Shh from cDNA
template prepared from 9.5 dpc mouse embryo RNA. The Shh cDNA fragment was inserted
by guest on April 16, 2018
http://ww
w.jbc.org/
Dow
nloaded from

6
into the bacterial expression vector pGEX-2TK (Amersham Pharmacia Biotech) such that the
resulting plasmid encoded a fusion protein composed of GST followed by a thrombin cleavage
site (LVPRGS), a five amino acid phosphorylation target site (RRASV) and the N-Shh
polypeptide. The construct was transformed into BL21 bacteria and the fusion protein was
isolated using glutathione-Sepharose affinity chromatography. Recombinant GST was
produced from cells transformed with the empty pGEX-2TK vector. Both recombinant protein
preparations were adsorbed onto a Detoxigel Endotoxin Removing Gel (Pierce, Rockford, IL).
The biological activity of GST-N-Shh was assayed in C3H10T1/2 cells using the method of
Williams et al. (30).
Radiolabeling of GST-N-Shh and GST-RAP. GST-N-Shh and GST-RAP were labeled with [γ -
32P]-ATP using heart muscle kinase (HMK) based on manufacturer recommendations (Sigma)
and the protocol of Stefansson et al. (31). Briefly, 50 µg of protein was incubated with 125 units
of HMK in 1X HMK buffer (20 mM PIPES, pH 6.5, 1 mM DTT, 20 mM NaCl, 12 mM MgCl2) plus
0.1% denatured BSA and 50 µCi [γ -32P]-ATP (Amersham Pharmacia Biotech) for 1h. Labeled
fusion protein was purified by size exclusion chromatography using PD-10 columns. Typical
specific activities were 0.5-2 x 108 cpm/nmol.
Wholemount embryo immunolabeling. Zebrafish were maintained and embryos collected by
standard methods (32). Embryos were fixed for 15 min in 4% paraformaldehyde in phosphate-
buffered saline (PBS). Embryos were washed two times in PBS, permeabilized by washing
three times in PBS containing 0.1% saponin (sap/PBS) at 37°C, and blocked for 30 min (37°C)
in PBS containing 5% goat serum and 0.1% saponin (gs/sap/PBS). Embryos were incubated
with primary antibody (5 µg/ml) in gs/sap/PBS first for 1 h at 37°C, then overnight at 4°C and
by guest on April 16, 2018
http://ww
w.jbc.org/
Dow
nloaded from

7
finally for an additional 1 h at 37°C with rocking. Embryos were washed three times with
gs/sap/PBS at 37°C, and incubated in Cy3-coupled goat anti-mouse or anti-rabbit secondary
antibody (Jackson ImmunoResearch Laboratories) 1.5 µg/ml in gs/sap/PBS for 1 h at 37°C.
Samples were washed three times in gs/sap/PBS, dehydrated in methanol, followed by clearing
in Murray’s Clear (1:2 benzylalcohol: benzylbenzoate). Laser scanning confocal microscopy
(LSCM) was performed using a BioRad MRC-1400 confocal microscope and BioRad
LaserSharp2000 software.
Immunoblotting and ligand overlay assay. Detergent extraction of cells and immunoblot
detection of megalin was performed as described previously (24). RAP ligand blot overlay
assay was performed as described by Battey et al. (27).
Solid-phase binding assays. Enzyme-linked immunosorbent assay (ELISA) was performed
essentially as described previously (33). Briefly, varying concentrations of N-Shh (R&D
Systems) or GST-N-Shh in 150 mM NaCl, 50 mM Tris pH 7.4, 3% nonfat milk, 0.05% Tween 20
were incubated for 1 h at 37oC in microtiter wells coated with megalin (3 µg/ml). Bound N-Shh
was detected using the monoclonal antibody 5E1, sheep anti-mouse IgG-horse radish
peroxidase (Amersham Pharmacia Biotech, Piscataway, NJ) and the chromogenic substrate
3,3',5,5'-tetramethyl-benzidine (TMB) (Kirkegaard & Perry, Gaithersburg, MD).
For homologous ligand competition assays, [32P]-labeled GST-N-Shh (1 nM) was
incubated in microtiter wells coated with megalin (3 µg/ml) in the presence of increasing
concentrations of unlabeled competitor (GST-N-Shh or RAP). All other conditions were similar
to those described in Williams et al. (34). The algorithm Ligand (35) within SigmaPlot 7.101 was
by guest on April 16, 2018
http://ww
w.jbc.org/
Dow
nloaded from

8
used to analyze the competition data and to determine dissociation and inhibition constants (Kd)
for receptor-ligand interactions.
Kinetic analysis of N-Shh-megalin binding. Kinetic analysis of the interaction of GST-N-Shh
with purified megalin was performed using surface plasmon resonance (SPR) measurements
made on a BIAcore 3000 instrument (BIAcore, Inc., Uppsala, Sweden). BIAcore sensor chips
(type CM5) were activated with a 1:1 mixture of 0.2 M N-ethyl-N’-(3-dimethylaminopropyl)
carbodiimide and 0.05 M N-hydroxysuccinimide in water. Megalin (50 µg/ml, 83 nM in 10 mM
sodium acetate, pH 4.8) was immobilized onto a CM5 sensor chip using the amine-coupling kit
as described by the supplier, BIAcore. Unreacted sites were blocked with 1M ethanolamine, pH
8.5. The SPR signal from immobilized megalin generated BIAcore response units ranging from
20,000-28,000. Control flow cells were activated and blocked in the absence of protein.
Binding was evaluated over a range of GST-N-Shh concentrations (25-500 nM) in 150 mM
NaCl, 0.005% polysorbate 20 and 100 mM Hepes (pH 7.4) plus and minus 1 mM CaCl2 at 25°C.
Binding of GST-N-Shh to megalin-immobilized flow cells was corrected for binding to control
flow cells. Binding data were fitted to a 1:1 (Langmuir) binding model using the BIAevaluation
3.1 software (BIAcore).
To evaluate the effect of pH on the dissociation of megalin-ligand complexes, GST-N-
Shh or RAP (each at 3 µM in 100 mM Hepes pH 7.4, 150 mM NaCl [HBSN]) were passed at 10
µl/minute for 2 minutes over sensor chips containing immobilized megalin. Subsequently,
protein-free HBSN or sodium acetate buffer pH 4.5 (sodium ion concentration adjusted to 150
mM) was applied for 5 minutes. The kinetic dissociation profiles obtained under neutral and
acid pH conditions were used to calculate off-rates (koff) using the BIAevaluation 3.1 program.
by guest on April 16, 2018
http://ww
w.jbc.org/
Dow
nloaded from

9
Between replicate experiments the chip surface was regenerated with a 10-sec pulse of 10 mM
glycine, pH 2.2 at 5 µl/min.
Confocal microscopic analysis of N-Shh uptake. BN cells were plated at 0.25 x105 cells/cm2
in eight-well chamber slides (Nunc Nalge, Naperville, IL) in Eagles Minimal Essential Medium
(MEM), containing 10% fetal bovine serum, nonessential amino acids, 100 units/ml penicillin
and 100 µg/ml streptomycin (complete medium). Cells were grown for 16 h at 37°C, 5% CO2
and the medium was replaced with serum-free medium (MEM, nonessential amino acids, 100
units/ml penicillin and 100 µg/ml streptomycin, insulin, transferrin and selenic acid). After a 1.5
h incubation the medium was replaced with serum-free medium containing 1.5% BSA and either
GST-N-Shh (20 nM) or GST (20 nM) plus or minus competitors and cultured for 2 h.
Competitors included RAP (1 µM), GST (1 µM).
For immunological detection GST-N-Shh- and GST-treated cells were rinsed in
Dulbecco’s phosphate buffered (DPBS) saline, pH 7.4 (DPBS), fixed for 20 min in 3.7%
paraformaldehyde with 0.2% Triton-X100 in DPBS, and then rinsed with DPBS. Cells were
incubated with 2% BSA in DPBS for 1 h and treated with goat anti-GST IgG (Amersham) at 1
µg/ml in 2% BSA/DPBS for 1 h, then FITC-donkey anti-goat IgG at 3 µg/ml in DPBS for 1 h, and
then rinsed in DPBS. For nuclear staining, cells were treated with RNase A (100 µg/ml) for 20
min at 37°C, rinsed in DPBS, and then treated with TOTO-3 (Molecular Probes, Eugene, OR) at
1 µg/ml in DPBS for 10 min at 37°C. Cells were rinsed in DPBS, mounted in Vector Shield
mounting solution (Vector Laboratories, Burlingame, CA) and then examined using LSCM.
by guest on April 16, 2018
http://ww
w.jbc.org/
Dow
nloaded from

10
Cellular internalization and degradation assays. BN were seeded into wells of 24-well plates
at 0.5 x105 cells/cm2 and grown for 16 h at 37°C, 5% CO2 in complete medium and then for 1.5
h in serum free medium. Medium was then replaced with serum-free medium plus 1.5% BSA
and [32P]-labeled GST-N-Shh (3 nM) with or without indicated agents (i.e., RAP, IgG or heparin),
and cells were grown for 2-6 h. For experiments measuring the effect of chloroquine treatment,
chloroquine was added at 0.1 mM concomitantly with the addition of radiolabeled ligands, and
uptake allowed to proceed for 6 h. Quantification of the amount of bound, internalized and
degraded ligands was performed as described previously (36). Radioactivity in the cell medium
that was soluble in 10% trichloroacetic acid (TCA) was taken to represent degraded ligand.
Total ligand degradation was corrected for the amount of degradation that occurred in
radioligand-containing medium in the absence of cells. To determine the amount of [32P]-ligand
that was bound and internalized, cells were washed three times with DPBS and then treated
with serum-free medium containing 0.5 mg/ml trypsin, 0.5 mg/ml proteinase K (Sigma) and 0.5
mM EDTA for 2-4 minutes at 4°C. The cell suspension was then centrifuged at 6000 x g for 4
min and the amount of radioactivity in the supernatant was taken to represent the bound fraction
while the amount in the cell pellet was taken as the internalized fraction.
Uptake experiments with differentiated and control F9 cells were performed as above
with the exception that cells were seeded at 1.0 x105 cells/cm2, and the growth medium was
DMEM, 10% FBS (or insulin, transferrin and selenic acid) containing 100 units/ml penicillin and
100 µg/ml streptomycin.
by guest on April 16, 2018
http://ww
w.jbc.org/
Dow
nloaded from

11
Results
Neurodevelopmental expression of megalin. Despite indications that megalin is critical to
neurodevelopment (19), little was known about the expression of the receptor during early
development. Laser scanning confocal microscopic analysis of 16-hour zebrafish embryos
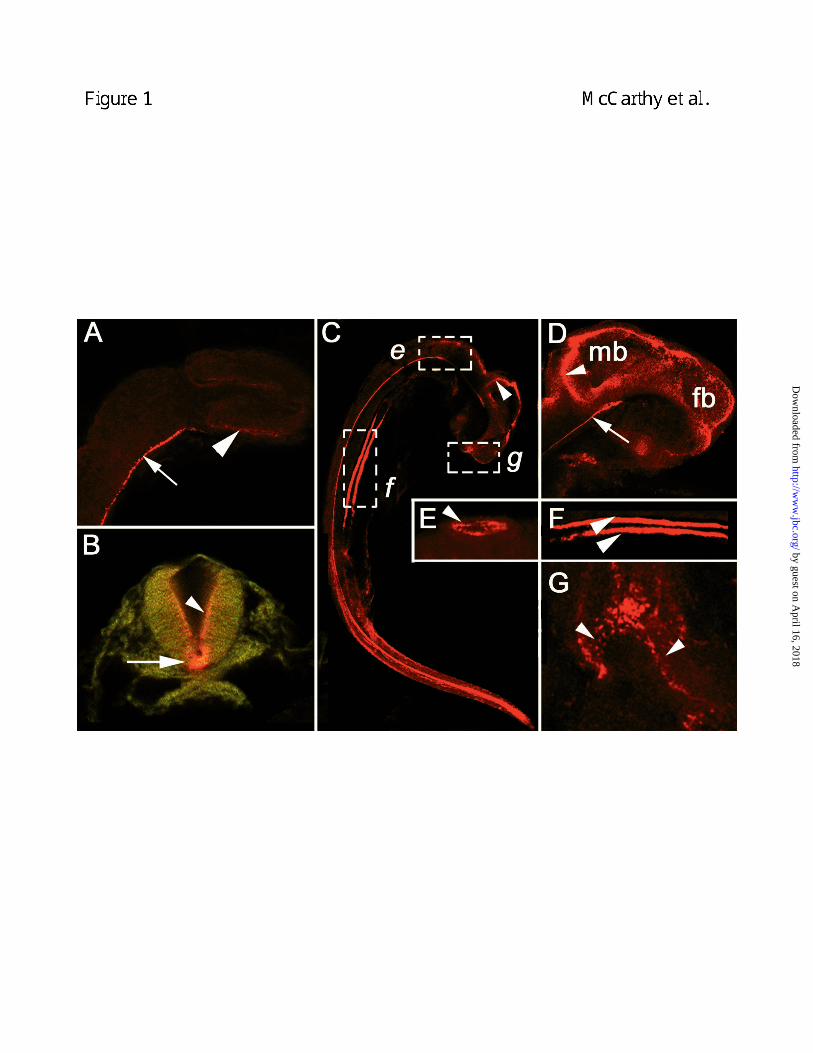
revealed that megalin was prominent in the floorplate of the neural tube (arrow, Fig. 1A) and the
apical surface of the optic cup (arrowhead, Fig. 1A). By 24-hours, megalin expression was
detected in cells of the ventral floorplate (arrow, Fig. 1B) and on the apical surface of cells lining
the lumen of the neural tube (arrowhead, Fig. 1B). At 33-hours, ventral floorplate expression
persisted and megalin was also extensively expressed on cells comprising the lumenal surfaces
of the forebrain and midbrain (arrowhead, Fig. 1D) with strong expression at the midbrain-
hindbrain border (arrowhead, Fig. 1C). At the base of the midbrain, intense staining for megalin
was seen at the most anterior extent of the floorplate (arrow, Fig. 1D). Outside the central
nervous system, megalin was detected on the apical surfaces of cells lining the lumen of the otic
vesicle of the developing ear (arrowhead, Fig. 1E). In the area of the developing mouth of 48-
hour embryos, megalin was distributed medially and laterally in the frontonasal and maxillary
processes, respectively (Fig. 1G). These findings demonstrate that early embryonic expression
of megalin occurs at specific organizing centers for morphogenesis including the ventral neural
tube, optic and otic vesicles and orofacial regions.
Many of the observed embryonic sites of megalin expression were the same as those
known to express Shh including ventral floorplate, eye, otic vesicle and frontonasal process (2-
5, 37). A notable exception was the absence of megalin expression in the notochord
(arrowheads, Fig. 2A and B). Also, megalin expression in the neural tube extended more dorsal
than Shh (inset, Fig. 2 and Fig. 1B), detected in areas of the neural tube known to express the
by guest on April 16, 2018
http://ww
w.jbc.org/
Dow
nloaded from

12
receptors for Shh, Ptc-1 and -2 (38). Taken together, megalin is expressed in tissues that
express Shh or in adjacent tissues regulated by Shh signaling. These observations support the
possibility that a functional relationship exists between megalin and Shh during early
neurodevelopment.
Megalin is a N-Shh-binding receptor. The similarity of megalin- and Shh-null phenotypes and
the early embryonic distribution of megalin in relation to sites of Shh production led us to
investigate whether megalin and N-Shh were capable of directly binding to one another.
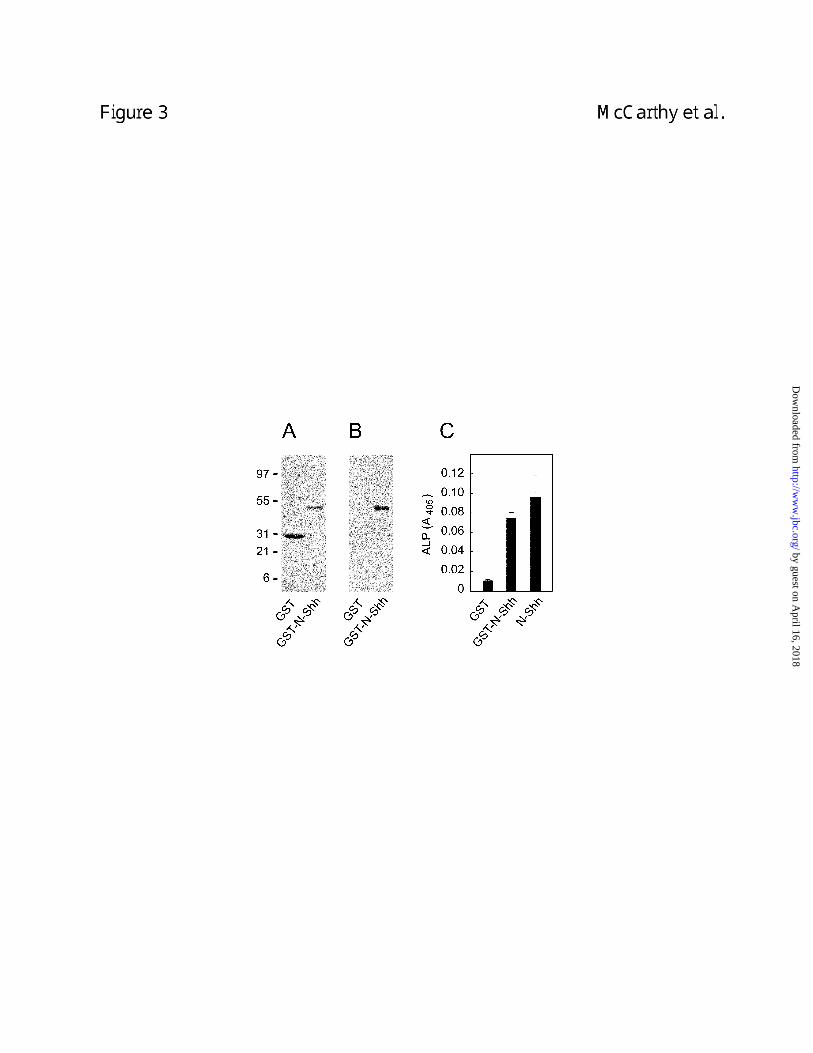
Enzyme-linked immunosorbant assay (ELISA) showed that a recombinant GST-N-Shh (Fig. 3)
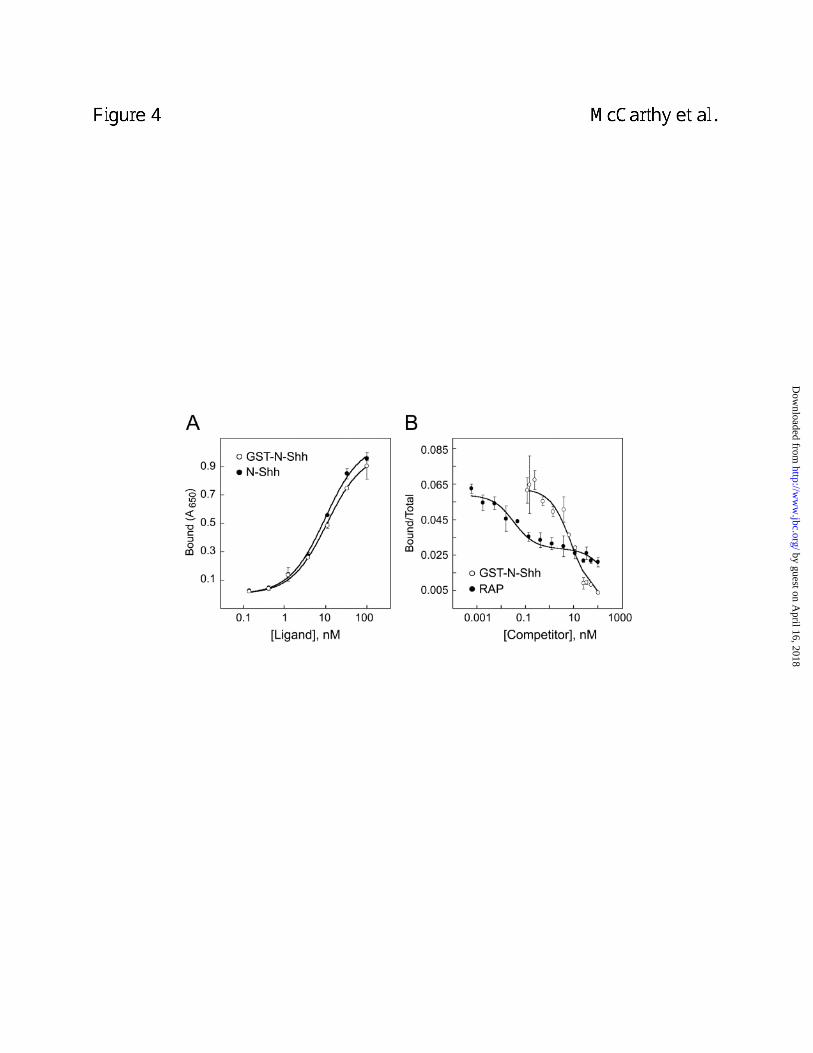
and a commercial preparation of N-Shh bound to purified megalin with similar apparent affinities
(Fig. 4A). Binding between GST-N-Shh and megalin was also tested using a homologous
ligand competition assay. [32P]-GST-N-Shh bound to megalin and the binding was inhibited in a
dose dependent manner by the addition of unlabeled GST-N-Shh (Fig. 4B). A dissociation
constant (Kd) of 81.3 nM was obtained from fitting the data to a single site model using the
Ligand algorithm. Binding of [32P]-GST-N-Shh to megalin was also inhibited by RAP, a well-
established antagonist of megalin-ligand interaction (39). Interestingly, the RAP competition
data could best be fit to a two-site model with inhibition constants (Ki) of 3.0 and 2341.9 nM.
One interpretation for these findings is that RAP binds to megalin at multiple sites and that one
of these binding interactions is a stronger inhibitor of N-Shh binding to megalin. Such an
interpretation is consistent with the fact that the megalin family member, LRP, has multiple RAP
binding sites (34).
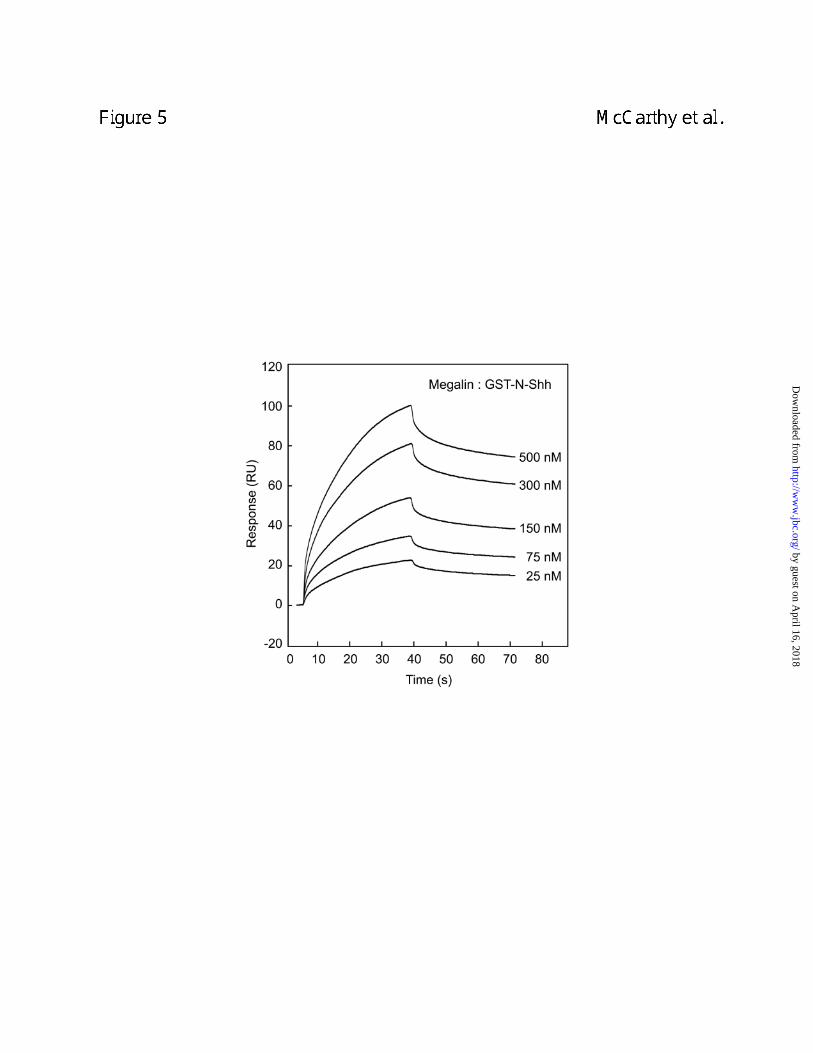
Binding of Shh to megalin was also evaluated using surface plasmon resonance (SPR).
As shown in Figure 5, GST-N-Shh bound to megalin immobilized on a sensor chip. GST alone
displayed no measurable binding to megalin (data not shown). Optimal fitting of SPR data
by guest on April 16, 2018
http://ww
w.jbc.org/
Dow
nloaded from

13
obtained from measuring the binding of various concentrations of GST-N-Shh to immobilized
megalin were best achieved using a single class binding site model. As a result, an affinity
constant (KD) of 21 nM (n = 7, chi2 of fit <10) was determined for GST-N-Shh binding to megalin
in the presence of calcium. Recombinant N-Shh cleaved with thrombin to remove the amino-
terminal GST moiety and commercial N-Shh were both found to bind megalin immobilized on a
sensor chip with affinities similar to those observed for the fusion protein (data not shown).
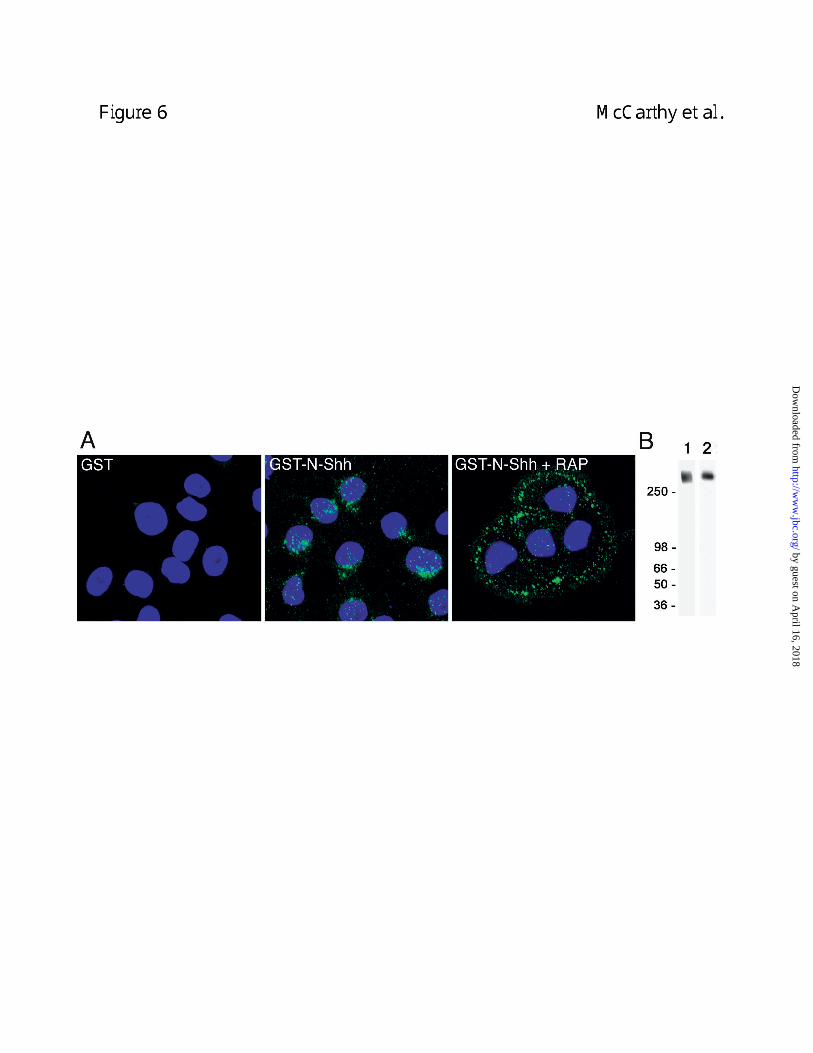
Megalin mediates endocytosis of N-Shh. The role of megalin in mediating endocytosis of N-
Shh was next evaluated. As shown in Figure 6, confocal analysis of BN cells cultured in the
presence of GST-N-Shh showed intracellular GST-N-Shh staining in a punctate pattern
consistent with vesicular localization. Cells incubated with GST showed little to no intracellular
staining (Fig. 6A). When BN cells were cultured in the presence of both GST-N-Shh and RAP,
little if any intracellular staining was observed (Fig. 6A). Instead, RAP treated cells displayed
punctate foci of staining located on the cell periphery. This staining pattern is consistent with a
plasma membrane or pericellular localization. Therefore, when megalin activity is abrogated, N-
Shh appears to bind to pericellular matrix or cell surface and uptake is blocked.
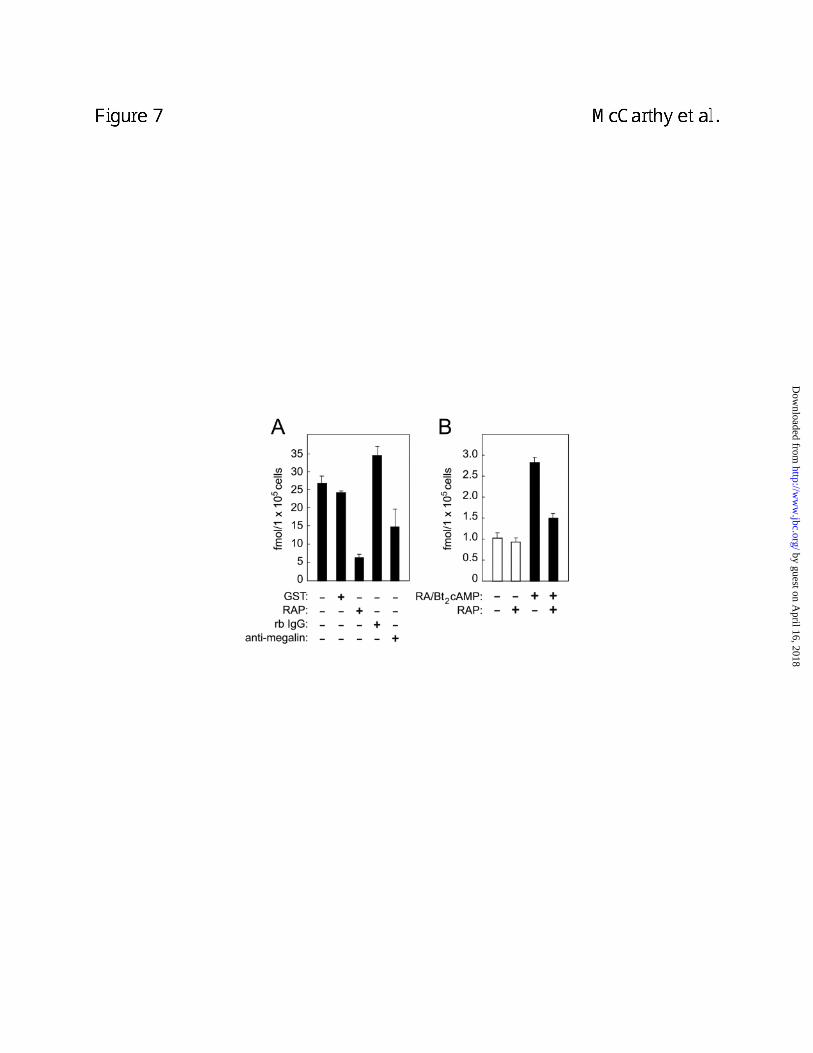
We subsequently evaluated the ability of BN cells to mediate endocytosis of radiolabeled
N-Shh. As shown in Figure 7A, BN cells internalized [32P]-GST-N-Shh. The uptake of [32P]-
GST-N-Shh could be blocked by either RAP or antibodies to megalin. The observed inhibitory
effects support the interpretation that megalin mediates N-Shh endocytosis. Furthermore,
inhibition by megalin antibodies alleviates a concern that the inhibitory effects of RAP might not
have been megalin-specific. In this regard it is also important to note that we established
megalin is the only detectable RAP-binding member of the LDLR family in BN cells (Fig. 6B).
Therefore RAP can be considered a specific inhibitor of megalin in BN cells.
by guest on April 16, 2018
http://ww
w.jbc.org/
Dow
nloaded from

14
Uptake of GST-N-Shh was also evaluated in murine F9 cells. F9 cells express little or no
megalin but can be differentiated with retinoic acid and Bt2/cAMP causing induced megalin
expression and decreased expression of other LDLR family members (24). As shown in Figure
7B, differentiated cells exhibited an increased capacity to internalize [32P]-GST-N-Shh. RAP
effectively inhibited internalization of [32P]-GST-N-Shh by differentiated F9 but had little effect on
the relatively low level of internalization in undifferentiated cells. These findings further support
the interpretation that megalin mediates endocytosis of N-Shh.
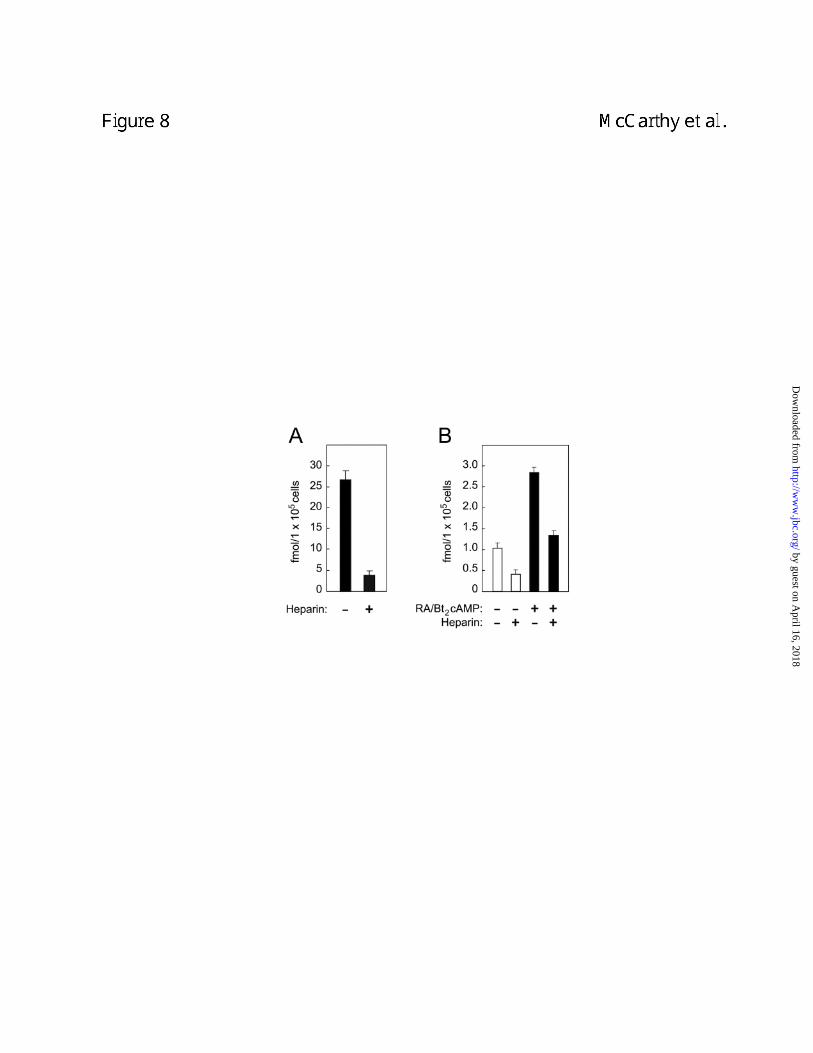
N-Shh endocytosis involves proteoglycans. In light of the fact that cell surface
proteoglycans have been implicated as partners with megalin and other LDLR family members
in the uptake of numerous ligands (18) we investigated their possible involvement in N-Shh
endocytosis. Heparin was an effective inhibitor of the uptake of [32P]-GST-N-Shh by both BN
cells and differentiated F9 cells (Figs. 8A and 8B). This suggests the involvement of cell
surface proteoglycans in the process of N-Shh endocytosis.
N-Shh is not efficiently targeted to lysosomes by megalin. One well-characterized
consequence of megalin-mediated endocytosis is targeting of ligands to the lysosome for
degradation. Inhibition of lysosomal proteinase activity using the drug chloroquine did not inhibit
[32P]-GST-N-Shh degradation in BN cells (Fig. 9). By contrast, in control experiments
chloroquine efficiently inhibited the degradation of [32P]-RAP (Fig. 9), a megalin ligand that is
targeted to the lysosomes following megalin-mediated endocytosis. Interestingly, there was a
significant level of chloroquine-insensitive N-Shh degradation suggesting that degradation of N-
Shh may occur extracellularly.
by guest on April 16, 2018
http://ww
w.jbc.org/
Dow
nloaded from

15
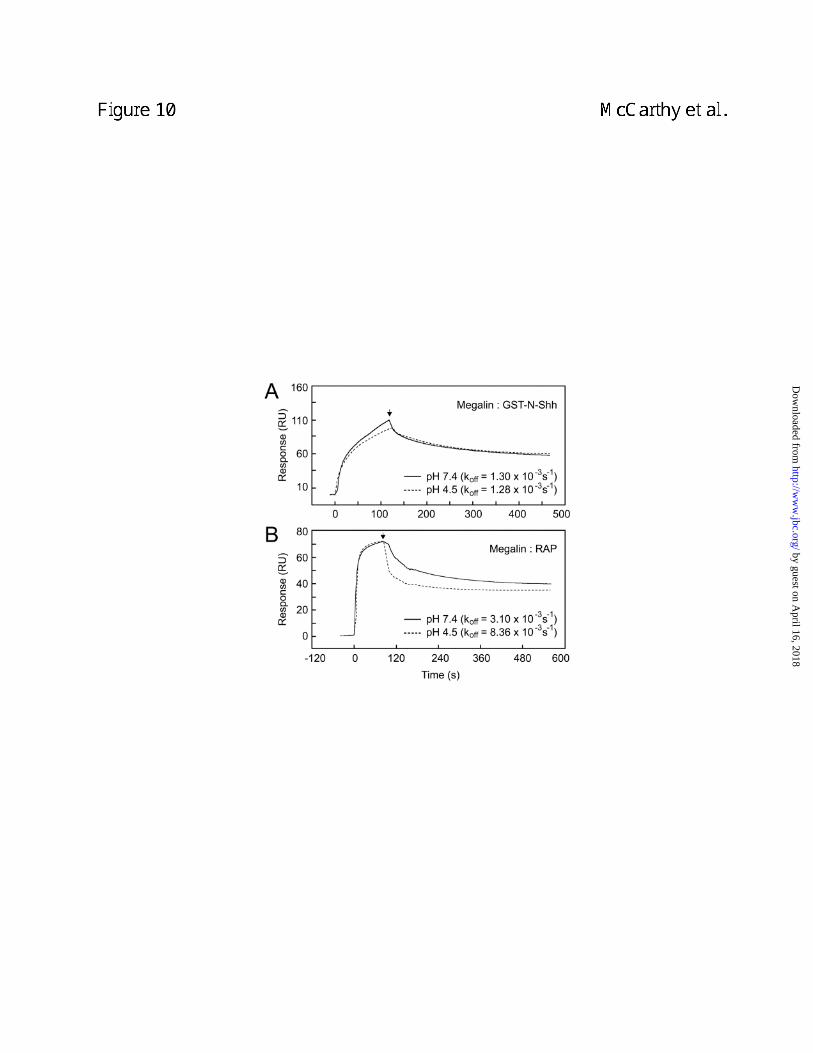
Evaluation of lowered pH on dissociation of the N-Shh-megalin complex. The effect of low
pH on the dissociation of the N-Shh-megalin complex was evaluated by SPR on a BIAcore
instrument. Little difference was evident in the dissociation rate constant (koff) for the N-Shh-
megalin interaction under acidic versus neutral pH conditions, 1.3 x 10-3 s-1 and 1.28 x 10-3 s-1,
respectively (Fig. 10A). By contrast, dissociation of the RAP-megalin complex increased ~3-
fold from 3.1 x 10-3 s-1 under neutral pH conditions to 8.36 x 10-3 s-1 under acidic pH conditions
(Fig. 10B). These findings indicate that the N-Shh-megalin interaction is resistant to
dissociation by acidic pH as low as 4.5 and suggest that N-Shh may not readily dissociate from
megalin under acidic pH within endosomes.
by guest on April 16, 2018
http://ww
w.jbc.org/
Dow
nloaded from

16
Discussion
Here we establish that a functional relationship exists between the endocytic receptor
megalin and the morphogen N-Shh. Specifically, we find that N-Shh binds to megalin with high
affinity and that the interaction is resistant to dissociation by low pH. We also show that one
consequence of the interaction is endocytosis of N-Shh. Megalin-mediated uptake of N-Shh
can be blocked by heparin, suggesting the involvement of heparan sulfate proteoglycans
(HSPGs) in the internalization process.
HSPGs have been implicated in N-Shh signaling (40, 41) and in the process of megalin-
mediated endocytosis of a number of its ligands (18). In the latter case, evidence suggests that
HSPGs serve to sequester ligands at or near the cell surface and thereby either facilitate
presentation of ligands to megalin or augment the affinity of ligands for megalin (18). Our
observation that N-Shh appeared to accumulate pericellularly on BN cells after blocking the
ligand-binding activity of megalin suggests the existence of an additional cell surface or
pericellular N-Shh-binding molecule. Considering recent evidence that Ptc is not detected at
significant levels on the cell surface (42), this other N-Shh binding component may very well be
HSPGs.
The likely significance of the interaction of N-Shh with megalin is that it impacts Shh
signaling. Three possibilities are that the interaction leads to: 1) direct signal transduction by
megalin, 2) modulation of the availability of N-Shh for its receptors or 3) transcytosis of N-Shh
important for long range N-Shh signaling. Direct signal transduction by megalin is supported by
recent evidence that other members of the LDLR family mediate signaling (43). For example,
LRP has been shown to interact with the heparin-binding growth factor, midkine, and regulate
by guest on April 16, 2018
http://ww
w.jbc.org/
Dow
nloaded from

17
midkine-dependent survival of embryonic neurons (44). LRP has also been shown to interact
with PDGF-BB and function as a co-receptor in the process of PDGF signaling (45, 46).
Additionally, VLDLR and apoER2 interact with the neuronal protein reelin and mediate signaling
through the cyotplasmic adaptor protein Dab1 (47). With respect to the second possibility,
megalin-mediated endocytosis of N-Shh may modulate the extracellular levels of N-Shh and
thereby regulate availability to Ptc. For example, megalin might compete with Ptc for limiting
levels of N-Shh and thereby reduce Ptc dissociation from Smo leading to decreased Smo
signaling. Alternatively, megalin may deliver N-Shh to vesicular pools of Ptc and thus regulate
the potential of this Ptc to complex with Smo. The third possibility is consistent with the
emerging role of megalin as a mediator of transepithelial transport of various macromolecules.
For example, thyroglobulin (TG), transcobalamin-vitamin B12 complex and retinol-binding protein
(RBP) in complex with retinol/vitamin A are internalized by megalin, but avoid lysosomal
degradation and are delivered to the basolateral membrane from which they get released (48-
50). The mechanism by which megalin ligands bypass lysosomal degradation is not known.
One possibility is that the interaction between megalin and these ligands might not be readily
dissociated by acidic pH such as occurs in endocytic vesicles. As a consequence, the ligands
traffic together with the receptor and are either transported to apical or basolateral aspects of
the cell. Our finding that the megalin-N-Shh interaction is insensitive to low pH suggests that N-
Shh may also traffic in complex with megalin and thus be recycled and/or transcytosed. This
possibility is further supported by our findings from chloroquine experiments indicating that
endocytosed-N-Shh bypasses lysosomes.
Megalin-mediated transcytosis of N-Shh may facilitate long range signaling by N-Shh
during early development. For example, N-Shh expressed in the floor plate may bind to megalin
expressed on the apical surface of the neural tube epithelium and mediate transepithelial
by guest on April 16, 2018
http://ww
w.jbc.org/
Dow
nloaded from

18
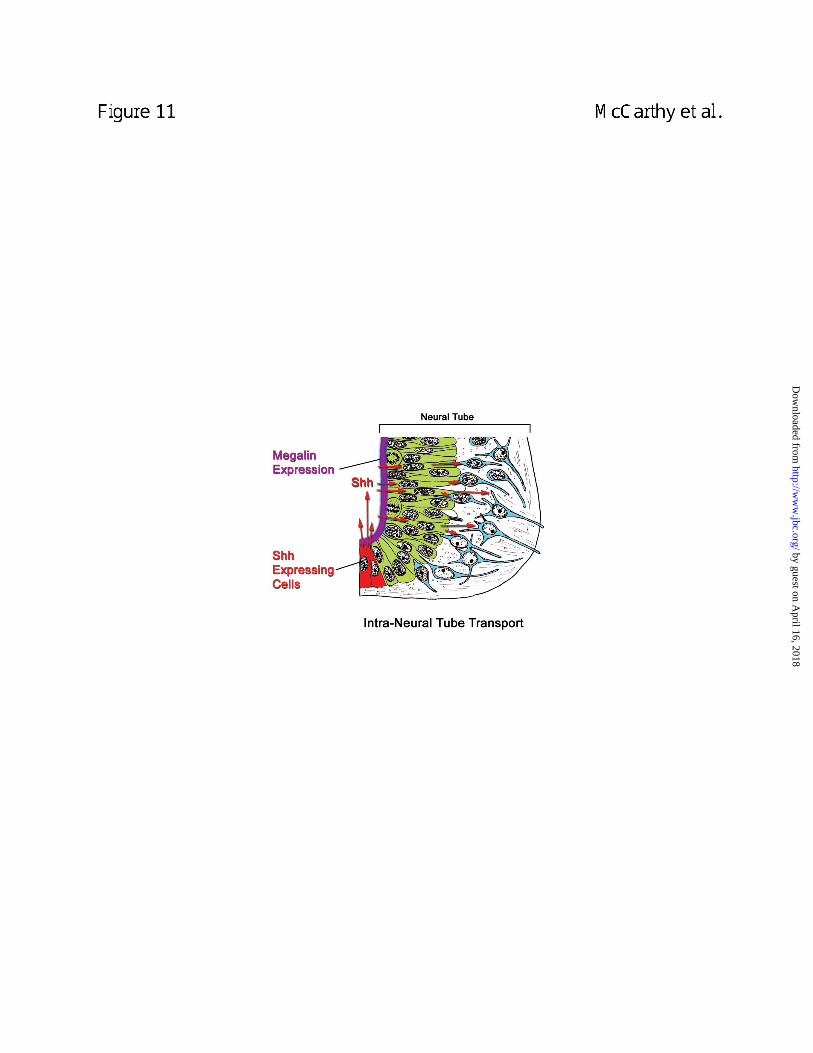
transport of N-Shh (Fig. 11). This process could account for delivery of N-Shh to cells in the
ventral region of the neural tube that undergo differentiation to form ventral nerves, a process
dependent on both N-Shh signal transduction and megalin expression (17, 19). A similar
process has been described in Drosophila involving transport of the morphogen Wingless
protein over large distances through imaginal disc epithelia (51). In this case, membrane
vesicles called argosomes, derived from the basolateral membranes, are transported
throughout imaginal disc epithelia. The argosomes are thought to originate from either
multivesicular endosomes and/or endosome transcytosis. Importantly, Wingless signaling has
been shown to involve a megalin family member, LRP6/arrow, although its exact role in the
process remains to be determined (52, 53).
In addition to the potential role of megalin in mediating long distance signaling via
transcytosis of N-Shh across the neural epithelium, its ability to endocytose N-Shh may also
impact N-Shh signaling in the early neural epithelium directly. Whether the mechanism for this
involves effects on the bioavailability of N-Shh or on the regulation of Ptc as described above,
the end result may be to influence N-Shh-dependent survival and differentiation of neural
epithelial cells (7, 8, 21). This hypothesis is supported by the megalin-deficient mouse
phenotype which demonstrates that megalin is required for normal viability and development of
the neuroepithelium (19).
by guest on April 16, 2018
http://ww
w.jbc.org/
Dow
nloaded from

19
References
1. Miller, L. A., Wert, S. E., and Whitsett, J. A. (2001) Immunolocalization of sonic
hedgehog (shh) in developing mouse lung, J Histochem Cytochem 49(12), 1593-604
2. Charrier, J. B., Lapointe, F., Douarin, N. M., and Teillet, M. A. (2001) Anti-apoptotic role
of Sonic hedgehog protein at the early stages of nervous system organogenesis, Development
128(20), 4011-20
3. Zhang, X. M., and Yang, X. J. (2001) Temporal and spatial effects of Sonic hedgehog
signaling in chick eye morphogenesis, Dev Biol 233(2), 271-90
4. Agarwala, S., Sanders, T. A., and Ragsdale, C. W. (2001) Sonic hedgehog control of
size and shape in midbrain pattern formation, Science 291(5511), 2147-50
5. Schneider, R. A., Hu, D., Rubenstein, J. L., Maden, M., and Helms, J. A. (2001) Local
retinoid signaling coordinates forebrain and facial morphogenesis by maintaining FGF8 and
SHH, Development 128(14), 2755-67
6. Hu, D., and Helms, J. A. (1999) The role of sonic hedgehog in normal and abnormal
craniofacial morphogenesis, Development 126(21), 4873-84
7. Chiang, C., Litingtung, Y., Lee, E., Young, K. E., Corden, J. L., Westphal, H., and
Beachy, P. A. (1996) Cyclopia and defective axial patterning in mice lacking Sonic hedgehog
gene function, Nature 383, 407-413
8. Litingtung, Y., and Chiang, C. (2000) Control of Shh activity and signaling in the neural
tube, Dev Dyn 219(2), 143-54
9. Fan, H., Oro, A. E., Scott, M. P., and Khavari, P. A. (1997) Induction of basal cell
carcinoma features in transgenic human skin expressing Sonic Hedgehog, Nat Med 3(7), 788-
92
by guest on April 16, 2018
http://ww
w.jbc.org/
Dow
nloaded from

20
10. Xie, J., Murone, M., Luoh, S. M., Ryan, A., Gu, Q., Zhang, C., Bonifas, J. M., Lam, C.
W., Hynes, M., Goddard, A., Rosenthal, A., Epstein, E. H., Jr., and de Sauvage, F. J. (1998)
Activating Smoothened mutations in sporadic basal-cell carcinoma, Nature 391(6662), 90-2
11. Nanni, L., Ming, J. E., Bocian, M., Steinhaus, K., Bianchi, D. W., Die-Smulders, C.,
Giannotti, A., Imaizumi, K., Jones, K. L., Campo, M. D., Martin, R. A., Meinecke, P., Pierpont, M.
E., Robin, N. H., Young, I. D., Roessler, E., and Muenke, M. (1999) The mutational spectrum of
the sonic hedgehog gene in holoprosencephaly: SHH mutations cause a significant proportion
of autosomal dominant holoprosencephaly, Hum Mol Genet 8(13), 2479-88
12. McMahon, A. P. (2000) More surprises in the hedgehog signaling pathway, Cell 100,
185-188
13. Porter, J. A., Young, K. E., and Beachy, P. A. (1996) Cholesterol modification of
hedgehog signaling proteins in animal development, Science 274(5285), 255-9
14. Chamoun, Z., Mann, R. K., Nellen, D., von Kessler, D. P., Bellotto, M., Beachy, P. A.,
and Basler, K. (2001) Skinny hedgehog, an acyltransferase required for palmitoylation and
activity of the hedgehog signal, Science 293(5537), 2080-4
15. Ingham, P. W., and McMahon, A. P. (2001) Hedgehog signaling in animal development:
paradigms and principles, Genes Dev 15(23), 3059-87
16. Hynes, M., Ye, W., Wang, K., Stone, D., Murone, M., Sauvage, F., and Rosenthal, A.
(2000) The seven-transmembrane receptor smoothened cell-autonomously induces multiple
ventral cell types, Nat Neurosci 3(1), 41-6
17. Litingtung, Y., and Chiang, C. (2000) Specification of ventral neuron types is mediated
by an antagonistic interaction between Shh and Gli3, Nat Neurosci 3(10), 979-85
18. Argraves, W. S. (2001) Members of the low density lipoprotein receptor family control
diverse physiological processes, Frontiers in Bioscience 6(March 1), D406-416
by guest on April 16, 2018
http://ww
w.jbc.org/
Dow
nloaded from

21
19. Willnow, T. E., Hilpert, J., Armstrong, S. A., Rohlmann, A., Hammer, R. E., Burns, D. K.,
and Herz, J. (1996) Defective forebrain development in mice lacking gp330/megalin, Proc Natl
Acad Sci U S A 93(16), 8460-4
20. Chen, W., Burgess, S., and Hopkins, N. (2001) Analysis of the zebrafish smoothened
mutant reveals conserved and divergent functions of hedgehog activity, Development 128(12),
2385-96
21. Ahlgren, S. C., and Bronner-Fraser, M. (1999) Inhibition of sonic hedgehog signaling in
vivo results in craniofacial neural crest cell death, Curr Biol 9(22), 1304-14
22. Rowitch, D. H., B, S. J., Lee, S. M., Flax, J. D., Snyder, E. Y., and McMahon, A. P.
(1999) Sonic hedgehog regulates proliferation and inhibits differentiation of CNS precursor cells,
J Neurosci 19(20), 8954-65
23. Varga, Z. M., Amores, A., Lewis, K. E., Yan, Y. L., Postlethwait, J. H., Eisen, J. S., and
Westerfield, M. (2001) Zebrafish smoothened functions in ventral neural tube specification and
axon tract formation, Development 128(18), 3497-509
24. Stefansson, S., Chappell, D. A., Argraves, K. M., Strickland, D. K., and Argraves, W. S.
(1995) Glycoprotein 330/low density lipoprotein receptor-related protein-2 mediates endocytosis
of low density lipoproteins via interaction with apolipoprotein B100, J Biol Chem 270(33), 19417-
21
25. Kounnas, M. Z., Chappell, D. A., Strickland, D. K., and Argraves, W. S. (1993)
Glycoprotein 330, a member of the low density lipoprotein receptor family, binds lipoprotein
lipase in vitro, J Biol Chem 268(19), 14176-81
26. Kounnas, M. Z., Haudenschild, C. C., Strickland, D. K., and Argraves, W. S. (1994)
Immunological localization of glycoprotein 330, low density lipoprotein receptor related protein
and 39 kDa receptor associated protein in embryonic mouse tissues, In Vivo 8(3), 343-51
by guest on April 16, 2018
http://ww
w.jbc.org/
Dow
nloaded from

22
27. Battey, F. D., Gafvels, M. E., Fitz Gerald, D. J., Argraves, W. S., Chappell, D. A.,
Strauss, J. F. r., and Strickland, D. K. (1994) The 39-kDa receptor-associated protein regulates
ligand binding by the very low density lipoprotein receptor, J Biol Chem 269(37), 23268-73
28. Kounnas, M. Z., Stefansson, S., Loukinova, E., Argraves, K. M., Strickland, D. K., and
Argraves, W. S. (1994) An overview of the structure and function of glycoprotein 330, a receptor
related to the alpha 2-macroglobulin receptor, Ann N Y Acad Sci 737, 114-23
29. Kounnas, M. Z., Argraves, W. S., and Strickland, D. K. (1992) The 39-kDa receptor-
associated protein interacts with two members of the low density lipoprotein receptor family,
alpha 2-macroglobulin receptor and glycoprotein 330, J Biol Chem 267(29), 21162-6
30. Williams, K. P., Rayhorn, P., Chi-Rosso, G., Garber, E. A., Strauch, K. L., Horan, G. S.,
Reilly, J. O., Baker, D. P., Taylor, F. R., Koteliansky, V., and Pepinsky, R. B. (1999) Functional
antagonists of sonic hedgehog reveal the importance of the N terminus for activity, J Cell Sci
112(Pt 23), 4405-14
31. Stefansson, S., Muhammad, S., Cheng, X. F., Battey, F. D., Strickland, D. K., and
Lawrence, D. A. (1998) Plasminogen activator inhibitor-1 contains a cryptic high affinity binding
site for the low density lipoprotein receptor-related protein, J Biol Chem 273(11), 6358-66
32. Westerfield, M. (1995) The Zebrafish Book: A guide for the laboratory use of zebrafish
(Danio rerio)., 3rd ed. Ed., University of Oregon Press, Eugene, Oregon
33. Hammad, S. M., Ranganathan, S., Loukinova, E., Twal, W. O., and Argraves, W. S.
(1997) Interaction of apolipoprotein J-amyloid beta-peptide complex with low density lipoprotein
receptor-related protein-2/megalin. A mechanism to prevent pathological accumulation of
amyloid beta-peptide, J Biol Chem 272(30), 18644-9
34. Williams, S. E., Ashcom, J. D., Argraves, W. S., and Strickland, D. K. (1992) A novel
mechanism for controlling the activity of alpha 2-macroglobulin receptor/low density lipoprotein
by guest on April 16, 2018
http://ww
w.jbc.org/
Dow
nloaded from

23
receptor-related protein. Multiple regulatory sites for 39-kDa receptor-associated protein, J Biol
Chem 267(13), 9035-40
35. Munson, P. J., and Rodbard, D. (1980) Ligand: a versatile computerized approach for
characterization of ligand-binding systems, Anal Biochem 107(1), 220-39
36. Stefansson, S., Kounnas, M. Z., Henkin, J., Mallampalli, R. K., Chappell, D. A.,
Strickland, D. K., and Argraves, W. S. (1995) gp330 on type II pneumocytes mediates
endocytosis leading to degradation of pro-urokinase, plasminogen activator inhibitor-1 and
urokinase-plasminogen activator inhibitor-1 complex, J Cell Sci 108(Pt 6), 2361-8
37. Helms, J. A., Kim, C. H., Hu, D., Minkoff, R., Thaller, C., and Eichele, G. (1997) Sonic
hedgehog participates in craniofacial morphogenesis and is down-regulated by teratogenic
doses of retinoic acid, Dev Biol 187(1), 25-35
38. Lewis, K. E., Concordet, J. P., and Ingham, P. W. (1999) Characterisation of a second
patched gene in the zebrafish Danio rerio and the differential response of patched genes to
Hedgehog signalling, Dev Biol 208(1), 14-29
39. Willnow, T. E. (1998) Receptor-associated protein (RAP): a specialized chaperone for
endocytic receptors, Biol Chem 379(8-9), 1025-31
40. Gritli-Linde, A., Lewis, P., McMahon, A. P., and Linde, A. (2001) The whereabouts of a
morphogen: direct evidence for short- and graded long-range activity of hedgehog signaling
peptides, Dev Biol 236(2), 364-86
41. The, I., Bellaiche, Y., and Perrimon, N. (1999) Hedgehog movement is regulated through
tout velu-dependent synthesis of a heparan sulfate proteoglycan, Mol Cell 4(4), 633-9
42. Incardona, J. P., Lee, J. H., Robertson, C. P., Enga, K., Kapur, R. P., and Roelink, H.
(2000) Receptor-mediated endocytosis of soluble and membrane-tethered sonic hedgehog by
patched-1, Proc Natl Acad Sci U S A 97(22), 12044-9
by guest on April 16, 2018
http://ww
w.jbc.org/
Dow
nloaded from

24
43. Strickland, D. K., Gonias, S., and Argraves, W. S. (2002) Diverse roles for the LDL
receptor family, Trends in Endocrinology & Metabolism 13(2), 66-74
44. Muramatsu, H., Zou, K., Sakaguchi, N., Ikematsu, S., Sakuma, S., and Muramatsu, T.
(2000) LDL Receptor-Related Protein as a Component of the Midkine Receptor, Biochem
Biophys Res Commun 270(3), 936-941
45. Boucher, P., Liu, P., Gotthardt, M., Hiesberger, T., Anderson, R. G., and Herz, J. (2002)
PDGF mediates tyrosine phosphorylation of the cytoplasmic domain of the LDL receptor-related
protein (LRP) in caveolae, J Biol Chem 19, 19
46. Loukinova, E., Ranganathan, S., Kuznetsov, S., Gorlatov, N., Migliorini, M. M., Loukinov,
D., Ulery, P. G., Mikhailenko, I., Lawrence, D. A., and Strickland, D. K. (2002) PDGF-induced
tyrosine phosphorylation of the LDL receptor-related protein (LRP): Evidence for integrated co-
receptor function betweenLRP and the PDGF receptor, J Biol Chem 19, 19
47. Trommsdorff, M., Gotthardt, M., Hiesberger, T., Shelton, J., Stockinger, W., Nimpf, J.,
Hammer, R. E., Richardson, J. A., and Herz, J. (1999) Reeler/Disabled-like disruption of
neuronal migration in knockout mice lacking the VLDL receptor and ApoE receptor 2, Cell 97(6),
689-701
48. Marino, M., and McCluskey, R. T. (2000) Megalin-mediated transcytosis of thyroglobulin
by thyroid cells is a calmodulin-dependent process, Thyroid 10(6), 461-9
49. Nielsen, R., Sorensen, B. S., Birn, H., Christensen, E. I., and Nexo, E. (2001)
Transcellular transport of vitamin b(12) in llc-pk1 renal proximal tubule cells, J Am Soc Nephrol
12(6), 1099-106
50. Marino, M., Andrews, D., Brown, D., and McCluskey, R. T. (2001) Transcytosis of
Retinol-Binding Protein across Renal Proximal Tubule Cells after Megalin (gp 330)-Mediated
Endocytosis, J Am Soc Nephrol 12(4), 637-48
by guest on April 16, 2018
http://ww
w.jbc.org/
Dow
nloaded from

25
51. Greco, V., Hannus, M., and Eaton, S. (2001) Argosomes. A Potential Vehicle for the
Spread of Morphogens through Epithelia, Cell 106(5), 633-45
52. Tamai, K., Semenov, M., Kato, Y., Spokony, R., Liu, C., Katsuyama, Y., Hess, F., Saint-
Jeannet, J. P., and He, X. (2000) LDL-receptor-related proteins in Wnt signal transduction,
Nature 407(6803), 530-5
53. Wehrli, M., Dougan, S. T., Caldwell, K., O'Keefe, L., Schwartz, S., Vaizel-Ohayon, D.,
Schejter, E., Tomlinson, A., and DiNardo, S. (2000) arrow encodes an LDL-receptor-related
protein essential for Wingless signalling, Nature 407(6803), 527-30
54. Balinsky, B. I. (1975) An Introduction to Embryology, 4 Ed., W.B. Saunders,
Philiadelphia
by guest on April 16, 2018
http://ww
w.jbc.org/
Dow
nloaded from

26
Acknowledgements
This work was supported by grants from the National Institutes of Health (HL61873) and the
American Heart Association, National (9950344N) to W.S.A. The authors thank Sandra Klatt for
technical assistance on this project, Drs. Paul Rayhorn and Kevin P. Williams (Biogen, Inc.,
Cambridge, MA) for technical advice concerning the differentiation of the C3H10T1/2 cells with
N-Shh and Dr. Waleed O. Twal for technical advice on BIAcore experimentation.
by guest on April 16, 2018
http://ww
w.jbc.org/
Dow
nloaded from

27
Figure Legends
Figure 1. Megalin expression during early embryogenesis. Shown are confocal
micrographs of a 16-hour (A), a 24-hour (B) and a 33-hour (C-G) zebrafish embryo. In A,
megalin is expressed in the central region of the developing eyecup (arrowhead). At all stages,
megalin is expressed in the floorplate (arrows in A, B and D). At 24 h megalin is prominently
expressed in the floorplate and more dorsally in cells that line the lumen of the neural tube (B,
arrowhead). In a 33-hour embryo, megalin is extensively expressed on the lumenal surfaces of
the ventricles of the midbrain (mb) and forebrain (fb), with intense staining beginning at the
midbrain-hindbrain border (arrowhead, C and D). Megalin expression is intense on the interior
epithelial surface of the otic vesicle of the developing ear (E). Megalin is also expressed in the
paired pronephric ducts (arrowheads, F) of the forming kidney where it is also associated with
the lumenal side of the epithelium. A frontal view of the developing oral region (G) shows that
megalin is expressed in the ridge of the frontonasal process and maxillary processes
(arrowheads).
Figure 2. Comparative analysis of Shh and megalin expression during early
embryogenesis. Shown are confocal micrographs of lateral views of 19-hour (20 somite)
zebrafish embryos stained with antibodies to N-Shh (A) and megalin (B). Both proteins are co-
expressed in the floor plate of the brain (arrows). N-Shh is expressed in the notochord
(arrowhead, A) whereas megalin expression is not detectable in the notochord (arrowhead, B).
Cross sectional views of regions caudal to the hindbrain (insets) show that megalin is
expressed more dorsally in the neural tube whereas N-Shh is confined to the floor plate.
by guest on April 16, 2018
http://ww
w.jbc.org/
Dow
nloaded from

28
Figure 3. Analysis of the integrity of recombinant GST-N-Shh. Panel A shows a Coomassie
staining of 5 µg of GST-N-Shh and GST. Panel B shows anti-N-Shh immunoblotting of 50 ng of
each protein. Panel C shows that GST-N-Shh is capable of stimulating10T1/2 cells to express
alkaline phosphatase (ALP), a marker of osteoblast differentiation. C3H10T1/2 cells were
treated for 5 days with 111 nM of commercial N-Shh, GST-N-Shh or GST and alkaline
phosphatase levels measured as described in Williams et al. (30).
Figure 4. ELISA and competitive radioligand binding assay demonstrate that N-Shh
binds to megalin and RAP inhibits the binding. In A, ELISA shows that both GST-N-Shh
(open circles) and commercially available N-Shh (filled circles) bind to megalin. In B,
homologous ligand displacement assay (open circles) was used to demonstrate the interaction
between [32P]-GST-N-Shh and megalin. In B, heterologous ligand displacement assay (filled
circles) was used to show that RAP inhibits binding of [32P]-GST-N-Shh to megalin. Curves
shown in B were based on fits of the data calculated using the computer program Ligand.
Figure 5. SPR analysis of Shh binding to megalin. Shown are SPR sensorgrams of GST-N-
Shh at varying concentrations (25-500 nM) binding to megalin immobilized on a sensor chip.
Data depicted is normalized to 100 response units and is representative of 6 separate
experiments. To obtain affinity constants (KD), SPR profiles in a given series were
simultaneously fit to a 1:1 binding site model using BIAevaluation 3.1 software.
Figure 6. N-Shh is endocytosed by BN cells and the uptake is inhibited by the megalin
antagonist RAP. In A, BN cells were incubated with GST-N-Shh or GST (20 nM) in the
presence of absence of RAP (1 µM) for 2 h and immunostained with anti-GST and FITC-
by guest on April 16, 2018
http://ww
w.jbc.org/
Dow
nloaded from

29
labeled-anti-IgG (green). Nuclei were stained using TOTO-3 (blue). RAP treatment did not
effect binding of GST-N-Shh to the cell but inhibited its internalization. Shown are confocal
micrographs from two independent experiments. Panel B shows that megalin is the principal
RAP-binding protein present in detergent extracts of BN cells. Aliquots of BN cell extract were
immunoblotted with anti-megalin IgG (lane 1) or incubated with RAP (1 µM) and the bound RAP
then detected with mouse monoclonal anti-RAP IgG (lane 2). No other RAP-binding proteins
were evident even after prolonged exposure of the RAP overlay blot.
Figure 7. Megalin antagonists inhibit uptake of [32P]-GST-N-Shh by BN cells and
differentiated F9 cells. In A, BN cells were incubated for 2 h with [32P]-GST-N-Shh alone or in
the presence of RAP (1 µM), anti-megalin IgG (150 µg/ml), normal rabbit IgG (150 µg/ml).
In B, undifferentiated F9 cells (white bars) or F9 cells differentiated with retinoic acid and (black
bars) were incubated for 2 h with [32P]-GST-N-Shh alone or in the presence of RAP (1 µM).
Figure 8. Heparin inhibits uptake of [32P]-GST-N-Shh by BN cells and differentiated F9
cells. In A, BN cells were incubated for 2 h with [32P]-GST-N-Shh alone or in the presence of
heparin (1 µM). In B, undifferentiated F9 cells (white bars) or F9 cells differentiated with retinoic
acid and Bt2/cAMP (black bars) were incubated for 2 h with [32P]-GST-N-Shh alone or in the
presence of heparin (1 µM).
Figure 9. Chloroquine treament does not inhibit degradation of internalized N-Shh. BN
cells were incubated with 3 nM [32P]-GST-N-Shh (top panels) or 3 nM [32P]-GST-RAP (bottom
panels) alone or in the presence of RAP (1 µM), GST (1 µM) or chloroquine (0.1 mM).
Measurements of bound, internalized and degraded radiolabeled ligand were made after 3-h
by guest on April 16, 2018
http://ww
w.jbc.org/
Dow
nloaded from

30
incubation. Note that RAP inhibits binding and internalization of [32P]-GST-N-Shh and [32P]-
RAP. By contrast, RAP and chloroquine both block degradation of labeled-RAP, but not of
labeled-N-Shh.
Figure 10. The N-Shh-megalin complex is resistant to dissociation by acidic pH. N-Shh
(A) or RAP (B) were allowed to associate for 2 min at neutral pH with megalin immobilized on a
sensor chip. The kinetics of N-Shh-megalin and RAP-megalin complex dissociation were
measured after the addition of protein-free buffer of neutral pH buffer or protein-free buffers of
acidic pH that ranged from 4.5 - 6.44. Shown are representative sensorgrams from experiments
that evaluated the effect of neutral pH and pH 4.5 buffers on the dissociation of each complex.
Arrows indicate the point of addition of protein-free flow buffer of indicated pH. Off rates (koff,
inset) were calculated using BIAevaluation 3.1 software.
Figure 11. Model of megalin-mediated transcytosis of N-Shh across neural tube
epithelial cells. Shown is a diagram of a neural tube in which cells of the ependymal layer
(green) transport N-Shh to Shh-responsive cells in the mantle layer (blue). N-Shh expressing
floorplate cells are red, lumenal epithelial cells are green, apical megalin expression is indicated
by purple and Shh-responsive cells are depicted in blue. Red arrows indicate the path of Shh
trafficking. The neural tube portion of the diagram was adapted from Balinsky (54).
by guest on April 16, 2018
http://ww
w.jbc.org/
Dow
nloaded from
by guest on April 16, 2018
http://ww
w.jbc.org/
Dow
nloaded from
argraves
Figure 1 McCarthy et al.

by guest on April 16, 2018
http://ww
w.jbc.org/
Dow
nloaded from
argraves
Figure 2 McCarthy et al.
by guest on April 16, 2018
http://ww
w.jbc.org/
Dow
nloaded from
argraves
Figure 3 McCarthy et al.
by guest on April 16, 2018
http://ww
w.jbc.org/
Dow
nloaded from
argraves
Figure 4 McCarthy et al.
by guest on April 16, 2018
http://ww
w.jbc.org/
Dow
nloaded from
argraves
Figure 5 McCarthy et al.
by guest on April 16, 2018
http://ww
w.jbc.org/
Dow
nloaded from
argraves
Figure 6 McCarthy et al.
by guest on April 16, 2018
http://ww
w.jbc.org/
Dow
nloaded from
argraves
Figure 7 McCarthy et al.
by guest on April 16, 2018
http://ww
w.jbc.org/
Dow
nloaded from
argraves
Figure 8 McCarthy et al.

by guest on April 16, 2018
http://ww
w.jbc.org/
Dow
nloaded from
argraves
Figure 9 McCarthy et al.
by guest on April 16, 2018
http://ww
w.jbc.org/
Dow
nloaded from
argraves
Figure 10 McCarthy et al.
argraves
by guest on April 16, 2018
http://ww
w.jbc.org/
Dow
nloaded from
argraves
Figure 11 McCarthy et al.

Scott ArgravesRobert A. McCarthy, Jeremy L. Barth, Mastan R. Chintalapudi, Christian Knaak and W.
Megalin functions as an endocytic sonic hedgehog receptor
published online April 18, 2002J. Biol. Chem.
10.1074/jbc.M201933200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
by guest on April 16, 2018
http://ww
w.jbc.org/
Dow
nloaded from


















