
JAUNDICE BILIRUBIN IN CONGENITAL NONHEMOLYTIC … · Journal of Clinical Investigation Vol. 42, No....
16
METABOLISM AND DISPOSITION OF C 14 - BILIRUBIN IN CONGENITAL NONHEMOLYTIC JAUNDICE Rudi Schmid, Lydia Hammaker J Clin Invest. 1963; 42(11):1720-1734. https://doi.org/10.1172/JCI104858. Research Article Find the latest version: http://jci.me/104858-pdf
Transcript of JAUNDICE BILIRUBIN IN CONGENITAL NONHEMOLYTIC … · Journal of Clinical Investigation Vol. 42, No....

METABOLISM AND DISPOSITION OF C14-BILIRUBIN IN CONGENITAL NONHEMOLYTICJAUNDICE
Rudi Schmid, Lydia Hammaker
J Clin Invest. 1963;42(11):1720-1734. https://doi.org/10.1172/JCI104858.
Research Article
Find the latest version:
http://jci.me/104858-pdf

Journal of Clinical InvestigationVol. 42, No. 11, 1963
METABOLISMAND DISPOSITION OF C14-BILIRUBIN IN CONGENITALNONHEMOLYTICJAUNDICE *
By RUDI SCHMID AND LYDIA HAMMAKER
(From the Thorndike Memorial Laboratory and the Second and Fourth [Harvard] MedicalServices, Boston City Hospital, and the Department of Medicine, Harvard
Medical School, Boston, Mass.)
(Submitted for publication June 14, 1963; accepted July 18, 1963)
In mammals, unconjugated bilirubin is rapidlycleared from the circulation and almost quanti-tatively excreted as a conjugate in the bile (2-4).The efficiency and rapidity of this process indicatethat conjugation and subsequent biliary excretionrepresent the principal pathway of bilirubin dis-position. If alternate pathways of bilirubin me-tabolism exist under physiologic conditions, theirlevel of functional efficiency must be so low as torender detection difficult. When hepatic dysfunc-tion causes interference with the efficiency ofconjugation and excretion of bilirubin, however,resulting in hyperbilirubinemia, such alternatemetabolic pathways may be expected to play amore important role in disposing of the accumu-lated pigment load (5). Moreover, when forma-tion and hence biliary excretion of conjugated pig-ment is virtually abolished owing to an enzymaticdefect in the hepatic conjugating apparatus, al-ternate pathways of bilirubin metabolism must as-sume the principal role in disposing of the bilepigment formed from the continuous breakdownof hemoglobin (6). Such metabolic anomaliesassociated with severe unconjugated hyperbili-rubinemia exist as a rare congenital syndrome inman (Crigler-Najjar syndrome) (7, 8) and as arecessively inherited enzymatic defect in a mutantstrain of Wistar rats (Gunn rats) (8, 9). Inboth instances, the level of unconjugated hyper-bilirubinemia remains remarkably constant overmonths and years (8), indicating that a "steadystate" has been established between pigment pro-duction and disposition.
In the present investigation, a patient with con-genital unconjugated hyperbilirubinemia and agroup of hyperbilirubinemic Gunn rats were in-
* Presented in part at the annual meeting of theAssociation of American Physicians, Atlantic City, N. J.,May 2, 1962 (1). Supported by U. S. Public HealthService grant A-1833.
jected with C14-bilirubin to determine the turn-over of the pigment and the metabolic dispositionof the label. Separate and quantitative collectionof bile, urine, and feces devoid of bile was achievedin the animal experiments by external biliarydrainage. Equilibration of the injected labeledpigment with the total miscible pool of bilirubinin the body permitted estimation of the extravas-cular bilirubin pool, and, in the rats, of the tissuedistribution of the pigment.
METHODS
Human studies
The patient was a 4a-year-old boy whose icterus wasfirst detected when he was 3 days old and who has sinceremained severely jaundiced (8). He experienced minorincidental illnesses, including measles, otitis, and severalupper respiratory infections, but developed normallyand failed to exhibit somatic or mental abnormalities re-ferable to his icterus. At the time of this study, heweighed 20 kg and measured 41i inches. Physical ex-amination revealed a deeply jaundiced boy without de-tectable abnormalities. Liver, spleen, and kidneys werenot palpable, secondary stigmata of hepatic disease werelacking, and repeated neurological examinations failed toelicit positive findings. The hemoglobin concentrationwas 12.1 g per 100 ml, hematocrit, 39%, reticulocytecount, 0.8% and leukocyte count, 9,000 per mm' with anormal differential. Routine urine and stool examina-tions were negative. Conventional liver-function tests,including Bromsulphalein clearance, alkaline phospha-tase activity, thymol turbidity, and cephalin flocculationwere within normal limits. Serum albumin concentrationwas 4.6 g per 100 ml (10).
The patient has been followed by the senior authorsince shortly after birth; during this period, the follow-ing special investigations were carried out. Serum bili-rubin concentrations (11) were determined on over 70individual blood samples. Values for direct-reactingbilirubin ranged from 0.3 to 1.0 mg per 100 ml and fortotal bilirubin, fluctuated between 20 and 30 mg per 100ml except during acute febrile illnesses, when bilirubinlevels tended to be slightly higher, with occasional valuesexceeding 30 mg per 100 ml. On paper chromatography(12), serum bilirubin yielded exclusively the azo deriva-
1720

METABOLISMAND DISPOSITION OF C14-BILIRUBIN
TABLE I
Urinary excretion of free N-acetyl-p-aminophenol (NAPA), NAPAglucuronide, and totalconjugated p-aminophenol after intravenous injection of NAPA
Percentage of injected dose excretedin urine in 6 hours
TotalSerum Free NAPA conjugated
Subject Age bilirubin NAPA glucuronide p-aminophenol
mg/100 mlPatient 8 mos 24.4 5.5 8.9 49.9Control boy 8 mos 0.3 4.0 28.0 78.5Control boy 8 mos 0.1 3.3 28.2 58.0Patient's father 32 yrs 0.5 2.7 33.8 53.8Patient's mother 28 yrs 0.3 2.6 46.2 69.6Normal men* 0.6 2.4 33.3 59.4
* Values are mean of 15 subjects.
tive of unconjugated bilirubin. Urine did not containdemonstrable bilirubin (13), and at the age of 6 months,fecal urobilinogen (14) was 0.23 mg per day. Bile as-pirated through a biluminal tube from the duodenumbefore and 20 minutes after intraduodenal infusion of15 ml 50%O MgSO4 contained 1.54 and 0.69 mg bilirubinper 100 ml (11), respectively; on paper chromatography(12), azo derivatives of conjugated bilirubin could notbe detected. An oral cholecystogram revealed normalvisualization of the gall bladder.
At the age of 8 months, the patient was given an ivinjection of 10 mg per kg body weight of N-acetyl-p-aminophenol (NAPA) (15). The amounts of NAPA,NAPA glucuronide, and total conjugated p-aminophenolexcreted in the urine during the subsequent 6-hour pe-riod were estimated (16), and the results expressed aspercentage of the administered dose. Similar studieswere carried out in two healthy boys of the same ageand in the patient's nonicteric parents. The results aregiven in Table I.
On the day before the present investigation,' the pa-tient's plasma volume was determined with T-1824 (17).He was then given an iv injection of 8.26 mg twice-re-crystallized C14-bilirubin (18), containing 1,134,000 dis-integrations per minute (dpm) per mg, dissolved in 14ml of 5% human albumin. During the subsequent 9days, 4-ml samples of blood were drawn at frequent in-tervals, first every 15 minutes to 1 hour, then daily, andfinally, every second day. In the serum obtained fromeach sample, bilirubin concentration and radioactivity (19)were determined and the specific activity of the pigmentwas calculated. Pooled samples of the sera collectedfrom days 2 through 9 were used for separation of theprotein fractions by continuous-flow electrophoresis (19)and for crystallization and radioassay of the extractedpigment (18, 20, 21).
All stools passed for 7 days after administration ofC14-bilirubin were collected and kept frozen until analy-
1 We are grateful to Dr. Sydney Gellis and his stafffor their permission to study this patient on the PediatricService of the Boston City Hospital and for their helpand cooperation.
sis. The stool samples were homogenized with 2 voldistilled water, and samples were used for the followingdeterminations: 1) Total radioactivity (21). 2) Crystal-lization of fecal bilirubin. Samples of the fecal homoge-nate were weakly acidified with acetic acid and repeat-edly extracted with a 2: 1 mixture of methanol and chlo-roform. Unlabeled carrier bilirubin was added, the com-bined pigment crystallized, and the specific activity ofthe crystals determined (18). 3) Fecal urobilinogen(14). 4) Fecal mesobilifuscin. Mesobilifuscin methylester was prepared by a modification of Siedel andM6ller's method (22). The purified dried pigment wasused for elemental analysis and for assay of specific ra-dioactivity (23).
All urine passed for 5 days after the administrationof the C14-bilirubin was collected, and the radioactivitydetermined (21). The remaining urine collected on days1, 2, 3, and 5 was pooled, mixed with concentrated HClto a final concentration of 1 N HCl, and then boiled for1 hour. Mesobilifuscin methyl ester was prepared asdescribed above for elemental analysis and for assay ofradioactivity. Conjugated carrier bilirubin was added tothe urine sample collected on day 4 for isolation, crystal-lization, and radioassay of the pigment.
Studies in rats
Preparation of experimental animals and collection ofsamples. Three male and five female homozygous,jaundiced Gunn rats were used. Their weights rangedfrom 200 to 400 g, the initial serum bilirubin, from 5.6to 10.9 mg per 100 ml, and their hematocrits, from 40 to49%o. Total blood volume was assumed to be 5% ofbody weight (24), and plasma volume was calculated onthe basis of the hematocrit.
In the first group of five male and female animals,external biliary drainage was established and a polyethylenesquare was sutured around the anus (21). The rats werethen placed in restraining cages, permitting separate andquantitative collection of bile, urine, and feces for unin-terrupted periods ranging from 53 to 96 hours. Pelletsof rat chow and drinking solution (0.2%o NaCl, 0.02%KCl, and 5%c glucose in water) were offered f reely.
1721

RUDI SCHMID AND LYDIA HAMMAKER
One rat was pretreated with oral neomycin (0.1 g perkg body wt per day) for 5 days immediately before theexperiment. A second group of three Gunn rats wasplaced in restraining cages without cannulation of thebile duct, but with the necessary provisions for separateand quantitative collection of feces and urine for unin-terrupted periods ranging from 96 to 148 hours.
At time zero, the rats were injected intravenously with170 to 1,100 ,ug of twice-recrystallized C14-bilirubin(SA, 800 to 2,420 dpm per pug) dissolved in 1.0 to 2.8 mlof normal rat serum. Volumes of 0.2 to 0.3 ml of bloodwere collected from the tip of the tail, first at intervalsof less than an hour and later, of 1 to 6 hours; duringthe latter part of the experiments, 2 to 3 blood sampleswere obtained per day. All urine and feces were col-lected continuously throughout the experiments. Atautopsy, stool contained in the small and large intestineswas recovered and analyzed separately. Bile wasdrained into tubes kept at 40 C in the dark and changedat frequent intervals.
Analytical procedures. Serum from each blood col-lection was used for determination of bilirubin concen-tration and radioactivity, and the specific activity of se-rum bilirubin was calculated. In pooled serum, obtainedby exsanguinating the animals at the end of the experi-ment, serum bilirubin concentration and radioactivitywere measured, and bilirubin was extracted and crystal-lized for determination of specific activity. Total radio-activity of urine was determined by methods previouslydescribed (21). In four instances pooled urine sampleswere mixed with unconjugated or conjugated carrierbilirubin for extraction, crystallization, and radioassay ofthe pigment. All samples of feces were homogenizedwith 4 vol of water, and portions were used for assayof total radioactivity (21). After addition of unlabeledcarrier bilirubin to either the fecal homogenates or totheir methanol-chloroform extracts, the pigment fromseven stool collections was crystallized and its specificactivity determined. All organic solvents used con-tained small amounts of hydroquinone to protect thebilirubin against oxidative breakdown (25).
In all bile specimens, total radioactivity was determined,and apparent bilirubin concentration was estimated by thediazo reaction. Five samples of Gunn rat bile weremixed with conjugated or unconjugated carrier biliru-bin, and the pigment was extracted and crystallized forradioassay. In three instances, 4.5 to 10 ml of Gunn ratbile containing known amounts of radioactivity was in-jected intravenously into large male Sprague-Dawleyrats with an external biliary fistula. In the recipients,all bile was collected for periods ranging from 6 to 18hours after iv injection. Bilirubin concentration andtotal radioactivity in these bile specimens were deter-mined, and bilirubin was crystallized for radioassay.Conjugated C14-bilirubin excreted in the bile of thenormal recipients was expressed as percentage of thetotal radioactivity in the injected Gunn rat bile. Forcomparison, normal rat bile containing conjugated C4-bilirubin was injected into two normal recipients with
external fistula, and the bile collected over the subse-quent 5 hours was similarly analyzed.
Attempts at chromatographic identification of the ra-dioactive catabolites in Gunn rat bile were unsuccessful,but adequate separation from the labeled pigments innormal rat bile was achieved by stepwise, ascending,thin-layer chromatography (26). Equal volumes of la-beled Gunn rat bile and unlabeled normal rat bile, or ofunlabeled Gunn rat bile and labeled normal rat bile weremixed and lyophilized, and the residue extracted with a2: 1 mixture of methanol and chloroform. After centri-fugation, 0.01-ml samples of the supernatant fluid wereapplied to a dry, 1-mm-thick layer of MN-cellulose 300g/CM 2 pasted on a 20- X 20-cm glass plate. The threesuccessively employed developing systems consisted of1) chloroform, 2) a 2: 1 mixture of butanol and hep-tane, and 3), a 10: 15: 10 mixture of butanol, methanol,and water. The dried chromatogram was divided intoconsecutive 2-cm squares; from these, the cellulose layerwas scraped and suspended in 1 ml of 1 M HyaminelOX for radioassay. Radioactivity in each square wasexpressed as percentage of the total isotope on the chro-matogram.
Samples of bile from a normal and a Gunn rat, previ-ously inj ected with C4-bilirubin, were extracted re-peatedly with butanol or chloroform at pH 5.0. Therespective extracts were pooled for determination ofbilirubin content and total radioactivity. Bile from eachof these two animals, buffered to pH 6.8, was incubatedwith 50 mg of bacterial 6-glucuronidase 3 at 370 C for4 hours. The pH was then adjusted to 5.0, and the in-cubated substance extracted repeatedly with chloroformfor determination of bilirubin content and radioactivity.Control incubations were carried out with heat-inacti-vated enzyme.
Gmelin (27) and pentdyopent (28) reactions werecarried out on native normal and Gunn rat bile and ontheir chloroform and methanol-chloroform extracts.For spectroscopic analysis, Gunn rat bile or normal ratbile was diluted with water to a final volume of 4.0 ml.Two ml of the diluted bile was mixed with 0.5 ml diazoreagent and 2.5 ml methanol. The color reaction wasallowed to develop in the dark for 30 minutes, and thenoptical density in the visible range was determined in aBeckman DU spectrophotometer. Spectral analysis wasalso carried out on butanol extracts of native -normaland Gunn rat bile at pH 5.0 and on chloroform extractsof comparable bile specimens previously hydrolyzed inthe dark for 20 minutes with 1 N NaOH.
Tissue analysis for radioactivity was done as follows.All rats were exsanguinated between 53 and 148 hoursafter the iv injection of C14-bilirubin, and the followingorgans and tissues were removed for radioactive assay:liver, spleen, kidneys, brain, abdominal skin, femoralmuscle, retroperitoneal or epidydimal adipose tissue,small and large bowel. The intestine was opened, andafter all fecal material collected was either combined
2 Brinkmann Instruments, Inc., Great Neck, N. Y.3 Sigma Chemical Corp., St. Louis, Mo.
1 722

METABOLISMAND DISPOSITION OF C'4-BILIRUBIN
301
0 24 48 72 96 120 144 168 192 216H OU R S
FIG. 1. SA OF SERUM BILIRUBIN IN THE 41-YEAR-OLD PATIENT WITHCONGENITAL UNCONJUGATEDHYPERBILIRUBINEMIA. At time zero, 4.3 tic C1-bilirubin was injected intravenously.
or separated for small and large bowel, the tissue was
washed in 0.9% saline. Tissue radioactivity was deter-mined as described for feces, and isotope concentrationwas expressed in disintegrations per minute per gramwet weight. For calculation of bilirubin concentration,tissue radioactivity was divided by the specific activityof the serum bilirubin at death, and a correction factorwas applied for labeled pigment bound to tissue albu-min (29), on the assumption that the binding ratio was
the same as that in the circulating plasma. In two rats,the pigment crystals of the renal papilla (30) were dis-solved in chloroform for determination of total bilirubincontent and radioactivity. In three instances, samples of
liver were homogenized with 4 vol of 1: 2 chloroform andmethanol for crystallization and radioassay of thepigment.
Radioassay of all pigment, serum, excreta, and tissueextracts was performed in a Packard Tri-Carb liquidscintillation spectrometer. Radioactivity was expressedin disintegrations per minute with toluene-C"4 as an in-ternal standard (18). Counting time ranged from 2 X10 to 2 X 30 minutes, depending on the activity present.Samples were considered to contain significant radioac-tivity when the number of counts in the counting vialsexceeded by at least 3 SD that of appropriately pre-
pared background vials (31).
TABLE II
Turnover of C'4-bilirubin and recovery of excreted C'4-activity in a 42-year-old boywith congenital unconjugated hyperbilirubinemia
Feces UrineC'4-activity
Days after recovered inadministration of Turnover of Wet Uro- Total crystalline C'4-activity
C14-bilirubin Serum bilirubin C14-bilirubin wt bilinogen C14-activity bilirubin Volume recovered
mg/100 ml dpm/24 hrs g/24 hrs mg/24 hrs dpm/24 hrs dpmn/24 hrs ml/24 hrs dpm/24 hrs1 25.1-27.1 880,000 No stool passed 610 65,580
[101*2 25.1-26.7 811,000 No stool passed 285 45,540
[21*3 24.6 741,000 35.3 8.3 0 0 425 46,1604 26.0 684,000 49.3 15.0 462,000 7,380 350 52,8505 26.0 625,000 51.6 27.0 1,190,000 13,780 530 60,5306 26.7 568,000 31.1 12.6 628,000 9,780 Not collected7 26.1-27.5 512,000 57.7 21.0 816,000 25,150 Not collected
[2]*Total (mean) 25.9 4,821,000 225.0 83.9 3,096,000 56,090 2,200 270,660
* Number of individual determinations.
100
70'
50
dpm/pg
1723
BILIRUBIN T/2 156 HRS.TOTAL MISCIBLE POOL 568 mg.DAILY TURNOVER 60.1 mg.
02016.4
10
7
5-

RUDI SCHMIDAND LYDIA HAMMAKER
12 24 36 48 60 72 84 96
HOU R S
;ERUM BILIRUBIN IN A 400-G MALE GUNNRAT WITH
RAINAGE. At time zero, 0.64 tic CG4-bilirubin was in-
RESULTS
Human studies
During this investigation, the patient's serum
bilirubin concentration remained stable (TableII), indicating a "steady state" between bilirubinformation and disposition. After injection of C14-bilirubin, mixing of the label with the rapidly ex-
changing bilirubin space resulted in a sharp ini-tial fall of specific activity (Figure 1), while a
second, slower phase of equilibration occurredduring the subsequent 24 hours. Specific activitydeclined logarithmically from approximately 30hours on (Figure 1), indicating that first-orderkinetics had been reached that could be character-ized by a single exponential rate of disappearance.In the pooled sera, collected from days 2 to 9,bilirubin SA was 11.8 dpm per jug when estimatedon the basis of total serum radioactivity and diazoreaction, and 11.4 dpm per ug after extraction andcrystallization of the pigment. All radioactivitymigrated with the albumin fraction on electropho-retic separation of the serum proteins (19).
From these data and a measured plasma volumeof 1,080 ml, the following values were calculated(32): circulating plasma pool of bilirubin, 280mg; total miscible pool, 568 mg; bilirubin halflife, 156 hours; and bilirubin turnover, 60.1 mg
per 24 hours.
No stools were passed during the first 2 daysof the study, but between days 3 and 7, 225 g offeces were collected, representing the total amountpassed during this period (Table II). For thepurpose of comparing fecal isotope excretion withthe turnover of C14-bilirubin in the body pool, an
average delay of 2 days was allowed for intestinaltransit time. Since the specimen passed on day3 failed to exhibit significant radioactivity, theactual transit time during the early phase of thisstudy may have been longer. For the presentcomparison, however, this appeared to be irrele-vant because the biological half-time of the labelas calculated from the fecal isotope excretion(Table II) was similar to that determined on thebasis of pigment turnover in the serum (Figure1). During the 5 days of stool collection, a totalof 3,096,000 dpm was eliminated by the fecalroute, corresponding to 82% of the calculated iso-tope turnover in the body pool during the com-
parable period. Only 56,090 dpm were recoveredas crystallized bilirubin from these stool specimens(Table II). On the basis of the specific activityof serum bilirubin on the days corresponding tothe fecal collections (Figure 1), the total amountof crystalline bilirubin recovered from the feceswas calculated at less than 5 mg.
Cumulative fecal excretion of urobilinogen forthe 5 days was 83.9 mg, representing 28% of
dpm/pg200
100
50
FIG. 2. SA OF SEXTERNAL BILIARY D
jected intravenously.
1724
METABOLISMAND DISPOSITION OF C14-BILIRUBIN1
dpm/pg
50
BILIRUBIN T/2 41 HRSTOTAL MISCIBLE POOL 400mgDAILY TURNOVER 62 mg
20
0 10 20 30 40 50 60 70 80 90 100
HOUR S
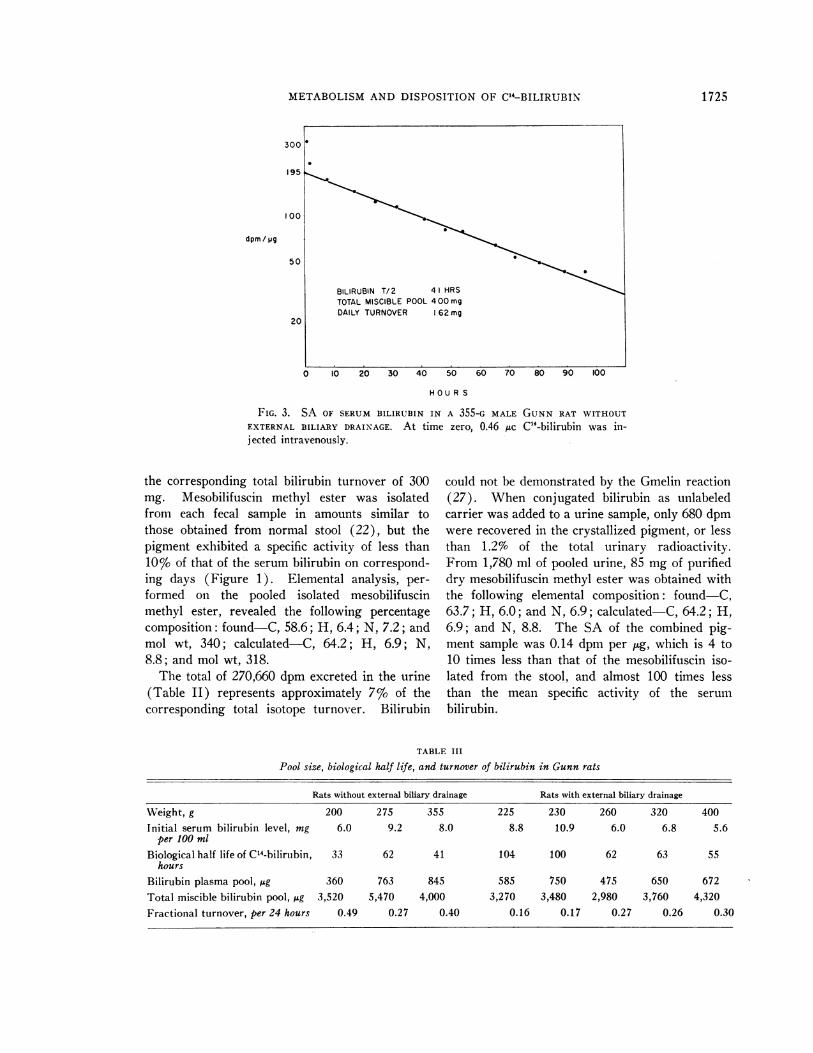
FIG. 3. SA OF SERUMBILIRUBIN IN A 355-G MALE GUNNRAT WITHOUT
EXTERNAL BILIARY DRAINAGE. At time zero, 0.46 MAc C14-bilirubin was in-jected intravenously.
the corresponding total bilirubin turnover of 300mg. Mesobilifuscin methyl ester was isolatedfrom each fecal sample in amounts similar tothose obtained from normal stool (22), but thepigment exhibited a specific activity of less than10% of that of the serum bilirubin on correspond-ing days (Figure 1). Elemental analysis, per-
formed on the pooled isolated mesobilifuscinmethyl ester, revealed the following percentagecomposition: found-C, 58.6; H, 6.4; N, 7.2; andmol wt, 340; calculated-C, 64.2; H, 6.9; N,8.8; and mol wt, 318.
The total of 270,660 dpm excreted in the urine(Table II) represents approximately 7% of thecorresponding total isotope turnover. Bilirubin
could not be demonstrated by the Gmelin reaction(27). When conjugated bilirubin as unlabeledcarrier was added to a urine sample, only 680 dpmwere recovered in the crystallized pigment, or lessthan 1.29 of the total urinary radioactivity.From 1,780 ml of pooled urine, 85 mg of purifieddry mesobilifuscin methyl ester was obtained withthe following elemental composition: found-C,63.7; H, 6.0; and N, 6.9; calculated-C, 64.2; H,6.9; and N, 8.8. The SA of the combined pig-ment sample was 0.14 dpm per uig, which is 4 to10 times less than that of the mesobilifuscin iso-lated from the stool, and almost 100 times lessthan the mean specific activity of the serum
bilirubin.
TABLE III
Pool size, biological half life, and turnover of bilirubin in Gunn rats
Rats without external biliary drainage Rats with external biliary drainage
Weight, g 200 275 355 225 230 260 320 400Initial serum bilirubin level, mg 6.0 9.2 8.0 8.8 10.9 6.0 6.8 5.6
per 100 mlBiological half life of C14-bilirubin, 33 62 41 104 100 62 63 55
hoursBilirubin plasma pool, jig 360 763 845 585 750 475 650 672Total miscible bilirubin pool, jig 3,520 5,470 4,000 3,270 3,480 2,980 3,760 4,320Fractional turnover, per 24 hours 0.49 0.27 0.40 0.16 0.17 0.27 0.26 0.30
1725

RUDI SCHMID AND LYDIA HAMMAKER
TABLE IV
Bilirubin concentration in organs of eight Gunn ratsinjected with ClS-bilirubin*
Bilirubin
Tissue Range Mean Corrected meant
lg or mlSerum 54-104 84 84Liver 59-175 92 84Kidney 27-100 44 37Spleen 4-50 29Brain 1-14 6 <6Intestine 20-61 43 37Adipose 22-400 102 99Skin 10-48 28 19Muscle 3-29 12 10
* Pigment concentration per gram wet weight, calculatedfrom tissue radioactivity divided by specific activity ofserum bilirubin at death (see text).
t Correction applied for pigment bound to tissue albumin(29), assuming that the binding ratio was the same as inthe plasma.
Studies in rats
Isotope turnover in vivo. In the eight Gunnrats injected with C14-bilirubin, equilibration ofthe labeled pigment with the total miscible bili-rubin pool required approximately 7 to 10 hours.Thereafter, disappearance of the isotope con-formed to a first-order reaction (Figures 2 and 3),permitting determination of the biological half lifeof C14-bilirubin (32) (Table III). In the pooledsera, obtained by exsanguinating the animals, bili-rubin SA was 28 dpm per jug when estimated onthe basis of total serum radioactivity and diazoreaction, and 32 dpm per ,ug when the pigment wasextracted and crystallized. On the basis of plasmavolume (24), serum concentration, and biologi-cal half life, the total miscible pool and the frac-tional turnover of bilirubin were calculated (32)for each rat. As shown in Table III, the totalmiscible pool was on the average 6 times largerthan the amount of pigment in the circulation, andthe fractional turnover per 24 hours ranged from0.16 to 0.49.
Bilirubin content of tissues. Bilirubin concen-tration in the major organs varied considerably(Table IV). After correction for pigment pre-sumed to be bound to the albumin present in thesetissues (29), it was evident that significant por-tions of the extravascular bilirubin pool werecontained in the liver, kidneys, intestine, and par-ticularly in adipose tissue, whereas in brain, radio-activity was barely measurable. In the three in-
stances in which C14-bilirubin was crystallizedfrom the liver, SA of the crystals was 26, 243,and 269 dpm per ,tg; SA of bilirubin in the corn-parable serum samples was 24, 256, and 201 dpmper ,ug. The bilirubin crystals recovered intactfrom the renal papilla exhibited less than half ofthe specific activity of the serum bilirubin obtainedat death, indicating that this pigment fractionturned over more slowly than the rest of themiscible pool.
Recovery of isotope in excreta. Total turnoverof C14-isotope for the whole experimental periodwas calculated in each rat, and the values werecompared with the combined recovery of radio-activity in the excreta. In the three rats withoutexternal bile drainage, combined recovery of ra-dioactivity in urine and feces ranged from 70 to100% (average, 83%o) of the respective valuescalculated for isotope turnover. In all instances,feces passed during the first day after administra-tion of the labeled pigment contained significantradioactivity. Values calculated for the biologicalhalf life of the label on the basis of fecal isotopeexcretion approximated those obtained from therate of disappearance of radioactivity from theserum.
In the five animals with external biliary drain-age, 32.5 to 114%o (average, 68%o) of the isotopecalculated as lost from the body pool was recoveredin the combined excreta. The lowest value wasobtained in a rat which, for unknown reasons,exhibited a gradual decrease in bile flow and infecal and urinary output. At sacrifice, this rat'shematocrit was below 30%, suggesting that theextracellular fluid compartment had expanded.In these rats, exclusion of the bile from the in-testinal tract resulted in bulky stools, and fecalisotope excretion tended to reach a plateau ratherthan to fall commensurately with the progressivedecrease in total isotope turnover (Figure 4).This was particularly evident in the rat pretreatedwith neomycin, which passed unusually volumi-nous stools containing a total of 76% of the iso-tope calculated to have been lost from the bodypool. In the remaining four rats of this group,radioactivity recovered in the feces averaged 17%cand in the urine, 3%o of the calculated isotopeturnover (Figure 4).
Characterization of the excreted isotope. Inall instances, the urine exhibited a negative
1726

METABOLISMAND DISPOSITION OF C14-BILIRUBIN
FECES
URINE
100,000
50,000-
100,000.
50,000-
-,:1-n-111A I mm
"I'll..rr , , . ,..
IST DAY 2N" DAY 3RODAY 4TH DAY AUTOPSY
FIG. 4. TURNOVEROF C14-BILIRUBIN AND EXCRETION OF RADIOACTIVE LABEL
IN A GUNNRAT INJECTED WITH C1-BILrRUBIN. Data for SA of serum
bilirubin in this rat are given in Figure 2.
Gmelin reaction. When urine samples were
mixed with unconjugated or conjugated carrierbilirubin for crystallization of the pigment, no sig-nificant label was detected in the crystals, and most
of the radioactivity remained in the aqueous
phase. In seven instances, fecal samples fromrats with an external biliary fistula were mixedwith unlabeled carrier bilirubin for extraction andcrystallization of the pigment. With three sam-
ples, to which albumin-bound bilirtubin had beenadded, the crystals contained 51, 49, and 26% ofthe total radioactivity present in the feces, whereasin the four instances in which the unlabeled bili-rubin was added to the combined chloroform ex-
tracts, 32, 25, 11, and 8% of the isotope was
present in the crystallized pigment. The last twovalues were obtained with feces collected at au-
topsy from the rat with the gradual decrease infecal, biliary, and urinary excretion.
In the five Gunn rats with external bile drain-age, the combined bile samples from each indi-vidual animal exhibited from 23 to 417% (aver-age, 35%o) of the radioactivity calculated to havebeen lost from the body pool (Figure 4). Whensamples of labeled Gunn rat bile were mixed withunlabeled normal rat bile for isolation and crystal-lization of the pigment, the crystals contained only3 to 13%o of the total radioactivity present, whilethe rest of the isotope remained in the aqueous
supernatant fluid. On addition of carrier biliru-
bin in solution of albumin or chloroform, 3 to 5%cof the total isotope was present in the crystallizedpigment. After injection of radioactive Gunn ratbile into normal rats with external bile drainage,any labeled bilirubin present in the administeredspecimens would have been mixed and excretedwith the recipients' endogenous bilirubin; thispermitted direct crystallization of the pigment andeliminated the need for addition of carrier bili-rubin ini vitro. As shown in Table V, only 8.1%cof the total radioactivity in the injected Gunn ratbile appeared in the recipients' bile in the form oflabeled bilirubin. Calculated on the basis of ex-
creted radioactivity, 15.1% of the isotope in therecipients' bile was recovered as C14-bilirubin.By contrast, after injection of normal rat bilecontaining conjugated C"4-bilirubin, 66% of theinjected or 80%o of the excreted radioactivity was
present in the bilirubin of the recipients' bile(Table V).
On thin-layer chromatography of extracts oflyophilized labeled Gunn rat bile, 88%c of theisotope migrated in about equal proportions withthe two butanol-containing systems, whereas only12% advanced with the chloroform fraction (Fig-ure 5). The latter contained most of the yellowpigment and exhibited a faintly positive Gmelinreaction, whereas the butanol-heptane and thebutanol-methanol-water fractions were virtuallycolorless. In contrast, comparable extracts of
d.p.m.
300,000
TURNOVEROFC14- BILIRUBIN
BILE 100,000-50,000'
RECOVERYOF LABEL IN
1727

RUDI SCHMID AND LYDIA HAMMAKER
TABLE V
Recovery of isotope in bile of normal rats injected with labeled Gunn rat bile
Injected bile specimen Collected bile specimen
Isotope in crystallized bilirubin
Duration of Total radioactivity %of injected %of radioactivityRat source Isotope content collection in bile radioactivity excreted in bile
dpm/specimen hours dpmGunn 19,944 18 7,920 6.3 15.9Gunn 20,928 18 12,875 8.7 14.1Gunn 21,520 6* 13,150 9.3 15.2Normal 68,840 5 59,232 69 80Normal 34,480 5 28,002 64 80
* Bilirubin crystallized from the bile specimen collected from 6 to 18 hours did not exhibit significant radioactivity.
labeled normal rat bile revealed 79% of the ra-
dioactivity migrating in the chloroform fraction,whereas the butanol systems contained only minorfractions of the isotope.
After incubation of Gunn rat bile with /3-glu-curonidase, chloroform-extractable radioactivitydid not differ significantly from that in comparablecontrol experiments with inactivated enzyme (Ta-ble VI). In contrast, incubation of normal ratbile containing conjugated C14-bilirubin with theenzyme resulted in a fourfold increment in chloro-form-extractable isotope (Table VI).
Native Gunn rat bile and samples extractedwith chloroform or with a 2: 1 mixture of meth-anol and chloroform gave negative to faintly posi-tive Gmelin (27) and pentdyopent (28) reactions,whereas with normal rat bile, these reactionswere strongly positive. With the modified van
den Bergh reaction, much less color developedwith Gunn rat bile than with normal bile (Fig-ure 6). Moreover, with the former, the small
amount of azo derivatives obtained exhibitedmaximal absorption at 520 mim, whereas with nor-
mal bile, the absorption peak lies in the 550 m/uband (33) (Figure 6). Since this indicated thatGunn rat bile contained constituents other thanbilirubin that reacted with the diazo reagent, ac-
tual bilirubin concentrations in these specimenscould not be estimated by this method. Additionalevidence that the diazo-positive material was notidentical with the excreted label was obtained on
extraction with butanol. Whereas from normalbile, butanol extracted a significant amount of la-beled yellow pigment exhibiting the characteristicspectral absorption of conjugated bilirubin (34)(Figure 7) and a positive diazo reaction (TableVI), comparable extracts of Gunn rat bile alsocontained radioactivity, but revealed a maximalabsorption around 400 mpk (Figure 7) and onlytraces of diazo-positive material (Table VI).Similar spectroscopic differences were noted inchloroform extracts of normal and Gunn rat bile
TABLE VI
Radioactivity extracted from bile specimens after iv injection of C'4-bilirubin
Radioactivity extracted
Sample Gunn rat bile Normal rat bile
SA of bilirubin,dpm % dpm % dpm/ug
Initial bile specimen, per ml 3,974 100 34,420 100 222Butanol extract, pH 5.0 1,270* 32* 15,150 44 168Chloroform extract, pH 5.0 239* 6* 894 2.6 162Incubated with inactivated 654* 16* 4,116 12 171
,3-glucuronidase, chloroform ex-tract, pH 5.0
Incubated with active ,1-glucu- 701* 18* 15,935 46 163ronidase, chloroform extract,pH 5.0
* Extract contained only trace of diazo-reacting material.
1728

METABOLISMAND DISPOSITION OF Cl"-BILIRUBIN
previously subjected to alkaline hydrolysis (Fig-ure 8).
DISCUSSION
Because of the large capacity of normal mam-malian liver to conjugate and excrete bilirubin(24, 35), the pigment formed from breakdown
% OFLABEL
80
40z
0 0
BO- o
40-
0
BUT (21 ) iHEPT (I)
CHCI33 2
GUNN RATmmmf1II~~~lilill ~~ BILE-C 14
X 1 [ x s X XORMA RATY; 111111111i _ _ _ _ _ _ C11|E1XlllE o _ _ _,BILE-
..........h+1 §§ §§
|BUT(IO): H20 (10)MOOH(15)
FIG. 5. STEPWISE ASCENDING THIN-LAYER CHROMA-TOGRAPHYOF EXTRACTS PREPAREDFROM LABELED BILE OFNORMALAND GUNNRATS. For details, see text.
of hemoglobin is rapidly cleared from the circu-lation and eliminated in the bile. Since this proc-ess results in a very short biological half life ofbilirubin, an exogenous label injected in a singledose would not mix adequately with the en-dogenously produced pigment. On the other
DIAZOTIZATION OF NATIVE BILE FROMNORMALRAT (1:6)
0. D. GUNN RAT *
O.D,
0.8
BUTANOL EXTRACT OF NATIVE BILE FROMNORMALRAT (1: 3) o--GUNN RAT - *
WA V E L E N GT H, mp.
FIG. 7. SPECTRAL ABSORPTIONOF A BUTANOLEXTRACT OF
NATIVE BILE FROMNORMALAND GUNNRATS.
hand, when C14-bilirubin was administered intra-venously to a patient and to animals with uncon-jugated hyperbilirubinemia, the disappearancerate of the label from the serum revealed kinetics(Figures 1-3) suggesting that virtually completeequilibration had been achieved. This was sup-ported by the finding that in the patient and inthe three rats without external bile drainage, thebiological half life of the label as calculated fromits rate of disappearance from the serum was
OD CHLOROFORMEXTRACT OF HYDROLYZEDBILE FROM°51 NORMALRAT 1:5)1
I ~~~~~~~GUNNRAT
380 420 460 500 540
WAVE LENGTH, mp.
FIG. 8. SPECTRAL ABSORPTION OF A CHLOROFORMEX-TRACT OF ALKALI-HYDROLYZED BILE FROM NORMALANDGUNNRATS.
similar to that calculated from fecal excretion ofthe isotope. In the five rats with an external biliaryfistula, variable weight loss associated with theoperative trauma and the exclusion of bile fromthe intestinal tract created a more complex situ-ation, making it difficult to ascertain that a true"steady state" was maintained throughout theexperiment. This difficulty may explain why inthis group of rats, bilirubin appeared to exhibita slightly longer biological half life and the mis-
WAVE LENGTH, mij.
FIG. 6. SPECTRAL ABSORPTION OF DIAZO-TREATED BILEFROM NORMALAND GUNNRATS.
1729
OUR" mml

RUDI SCHMID AND LYDIA HAMMAKER
cible pool a smaller fractional turnover than inrats without surgery (Table III).
In the patient, daily bilirubin turnover amountedto 60.1 mg, closely approximating the anticipatedvalue of 58.1 mg, calculated from the total red-cellvolume, a mean erythrocyte life span of 120 days(36), and an estimated 12% increment for bilepigment formation from sources other than cir-culating hemoglobin (37). Similarly, in the threeGunn rats without external bile drainage, the mag-
nitude of bilirubin turnover agreed in general withthe values calculated from a mean erythrocyte lifespan of 60 days (38), allowing for additional pig-ment formation from random destruction of a
minor red-cell fraction (39) and from nonhemo-globin sources (37). These findings indicatethat in man and rats with this type of hyperbili-rubinemia, bile pigment formation from endoge-nous sources is quantitatively comparable to thatoccurring under physiologic conditions.
On the other hand, the two species differed sig-nificantly in their bodily distribution of bilirubin.In the patient, the total pigment pool was dividedabout equally between the circulating plasma andthe extravascular compartments. Since bilirubinis tightly bound to human albumin (19), it was
not surprising that the pigment exhibited a totalspace of distribution similar to that reported forthis protein (40, 41). The biological half life ofbilirubin, however, was two to three times shorter
(Figure 1) than that of albumin (40, 41), sug-
gesting that in the tissues, part of the pigmentbecame detached from its carrier protein and was
destroyed at an accelerated rate (19, 42). In therats, however, where the interaction between thepigment and albumin is weaker (19), five-sixthsof the miscible bilirubin pool was present in theextravascular compartments (Table III). Sincethis space of distribution is far larger than thatof extravascular albumin (29, 43), a significantfraction of the exchangeable bilirubin must havebeen bound to tissue proteins or lipids. Asshown in Table IV, this binding occurred pre-
dominantly in the liver, kidney, intestine, and adi-pose tissue, but smaller amounts of pigment were
present also in the spleen, skin, and muscle. Al-though all rats used in these experiments exhibitedneurological damage resembling kernicterus (44).their brains contained only negligible radioac-tivity. This finding supports present concepts
that after the neonatal period the blood-brain bar-rier becomes virtually impermeable to bilirubin(45), but it does not rule out the possibility of in-creased pigment concentrations occurring in se-lected local centers of the nervous system.
One of the major aims of this investigation was
the detection and functional evaluation of the al-ternate pathways for bilirubin disposition that inthe absence of effective conjugation, must be re-sponsible for the maintenance of a "steady state"between pigment formation and elimination (6).A partial answer to this problem was obtainedfrom the study of the patient and the three intactrats. In the patient, 300 mg of bilirubin wereturned over during 5 days, but less than 30%o ofit could be accounted for as fecal urobilinogenand bilirubin (Table II). The exact source ofthis urobilinogen fraction could not be definitelyestablished. Since with NAPA, glucuronide for-mation was found to be markedly reduced but notentirely absent (Table I), it is conceivable thatsmall amounts of conjugated bilirubin could havebeen formed in the liver and excreted in the bilewithout being detected on duodenal aspiration.Although this possibility could not be definitelyexcluded, urobilinogen could also have been de-rived from unconjugated bilirubin reaching theintestinal lumen either with the bile (46), oracross bowel wall, and subsequently being reducedby the fecal flora (47).
In the stool, two-thirds of the excreted isotopewas present in unidentified catabolites of biliru-bin that probably reached the intestinal tract withthe bile. That most of this labeled material couldnot have been mesobilifuscin was demonstrated bythe latter's low specific activity. This findingsupports earlier observations (22) that most ofthe fecal mesobilifuscin is derived from metabolicsources other than circulating bilirubin or itsintestinal breakdown products.
Urinary excretion played a minor role in theover-all pigment disposition, and the small amountof isotope appearing in urine was all present aswater-soluble bilirubin catabolites. Similar ob-servations were made in the three Gunn rats with-out external bile drainage, where 69 to 96% ofthe radioactivity calculated as lost from the bodypool was recovered in the feces, whereas lessthan 57% of the label appeared in the urine.
Although in these studies most of the isotope
1730

METABOLISMAND DISPOSITION OF C14-BILIRUBIN
turned oxver appeared in the feces, little directinformation was obtained about the form inwhich the labeled compounds reached the intesti-nal tract and by which routes. The followingtheoretical possibilities appeared to be worthy ofconsideration: 1) that alternate conjugatingmechanisms, such as sulfate formation (48), sub-stitute for the defective glucuronidation (30); 2)that unconjugated bilirubin is excreted in thebile (46) and perhaps across the intestinal mu-
cosa (49); and 3) that bilirubin is converted tomore polar catabolites that can be excreted with-out the need for conjugation.
In the five rats with external biliary drainage,although no specific search was made for bili-rubin sulfate (48), spectroscopy (Figures 6, 7),chromatography (Figure 5), and butanol ex-
traction (Table VI) of the bile failed to revealdetectable amounts of conjugated bilirubin. In-deed, if these animals could excrete appreciableamounts of bilirubin by sulfate formation or byother conjugating mechanisms, significant reten-tion of unconj ugated pigment in the organism
would not be expected. In all instances, the bilewas found to contain small amounts of uncon-
jugated bilirubin, but in terms of the over-all pig-ment disposition, the contribution of this excre-
tory pathway appeared to be of minor significance.More important was the observation that in spiteof complete diversion of the bile to the outside, a
portion of the injected radioactivity was presentin the feces. Moreover, a significant fraction ofthis label was contained in unconjugated bilirubin,suggesting that the latter had reached the bowellumen across the intestinal mucosa. Since uncon-
jugated bilirubin is absorbed from the gut (21)also, the fecal material appeared to contain a bili-rubin pool exchanging with the miscible body pool.By oral administration of cholestyramine,4 whichtightly binds bilirubin, a portion of this fecal pig-ment was trapped in the intestine, causing a
marked fall of the serum bilirubin level (49).These observations indicated that in Gunn rats,and perhaps in patients with unconjugated hyper-bilirubinemia, direct transfer of pigment across
the intestinal wall represents an ancillary path-way for bilirubin excretion. In the rats with ex-
ternal biliary drainage, however, the bulky con-
4 MK-135, Merck Sharp & Dohme Research Labora-tories, West Point, Pa.
sistency of the feces may have resulted in a largerthan normal excretion of labeled pigment into thegut; this was particularly evident in the rat pre-treated with neomycin.
Approximately one-third to two-fifths of thetotal isotope turnover appeared in the form ofwater-soluble catabolites of bilirubin excreted inthe bile, and to a lesser degree, in the urine.In part, these pigment derivatives were yellowand soluble in butanol (Table VI), but they lackedthe spectral properties of bilirubin (Figures 6-8)and failed to give a positive diazo reaction (TableVI). Moreover, after injection into normal rats,they were rapidly excreted in the bile without be-ing converted to conjugated bilirubin (Table V).The exact structure of these pigment derivativesis unknown, but their solubility and spectroscopicproperties appeared to be comparable to those ofthe breakdown products obtained on exposure ofbilirubin to mild alkali (18) and to light (50)in vitro. The present findings suggest that inunconjugated hyperbilirubinemia, a similar break-down of bilirubin may occur in vivo. To a limitedextent, such pigment breakdown may occur alsoin animals with an intact conjugating mechanism,as we inferred from the finding that in the bile ofnormal rats injected with C'4-bilirubin, only 80%of the excreted isotope was accounted for in theisolated labeled pigment (Table V). Further-more, in such bile specimens, the apparent specificactivity of bilirubin was distinctly higher than thatof the pigment fractions extracted with butanol orchloroform (Table VI). On the other hand, inGunn rat bile, the diazo reaction gave spectropho-tomietric readings suggesting significantly higherbilirubin concentrations than were actually present.This result implies that in these specimens, sub-stances other than bilirubin were responsible forthe occurrence of an "atypical" diazo reaction(Figure 6). The nature of these compounds hasnot been determined, nor is it known whethersimilar diazo-positive excretory products are pres-ent in normal bile. Human T-tube bile, however,may contain small amounts of diazo-reacting ma-terial that is neither hydrolyzed by alkali, nor ac-counted for as sulfate conjugates of bilirubin (48).
These observations indicate that in unconju-gated hyperbilirubinemia, several alternate path-ways of pigment disposition can substitute for thedeficient glucuronide formation (Figure 9).
1731

RUDI SCHMID AND LYDIA HAMMAKER
BREAKDOWNOFHEMEPROTEINS
POLAR, DIAZO-NEGATIVEDERIVATIVES
UNCONJUGC% NEBILIRUBIN DEPOLARDERIVATIV. URINE
GUT
BILE
FIG. 9. SCHEMATIC REPRESENTATION OF ALTERNATE
PATHS OF BILIRUBIN DISPOSITION IN HYPERBILIRUBINEMIC
GUNNRATS WITH DEFECrIVE GLUCURONIDEFORMATION.
When their combined functional rate equals therate of pigment formation, a "steady state" isachieved, resulting in a constant level of serum
bilirubin. In patients with the Crigler-Najjarsyndrome, such a balanced state is reached only atvery high bilirubin concentrations (7, 8), whichprobably reflects the unusually strong affinity andprotective action of human albumin for the pigment(19, 42). In Gunn rats, on the other hand, wherebilirubin is less tightly bound to albumin (19)and hence escapes more easily into the extravas-cular compartments, the combined rates of pig-ment breakdown and of excretion through the in-testinal wall equal that of pigment production atsignificantly lower bilirubin levels (30, 44).Since in human and rat sera, bilirubin cataboliteswere not detectable, it appears likely that they are
very rapidly cleared from the circulation, or thatpigment breakdown occurs at anatomical sites per-
mitting irreversible egress of the resulting metabo-lites. It is possible that destruction of bilirubinin these tissue sites may be enhanced by intenseillumination (51, 52), or by administration ofcertain drugs or chemicals (53, 54).
SUMMARY
1) In a patient with congenital unconjugatedhyperbilirubinemia (Crigler-Najjar syndrome)and in eight rats with hereditary icterus (Gunnrats), tracer techniques were used to estimate thebiological half life, total miscible pool, and frac-tional turnover of bilirubin. The metabolic dis-position of the C14-bilirubin was determined in
the feces and urine, and in five of the rats, in thehile.
2) In the patient, the total miscible bilirubin10ool was distributed about equally between thecirculating plasma and the extravascular space,which was attributed to the very strong interac-tion of the pigment with human albumin. In therats, however, where bilirubin is less tightly boundto albumin, five-sixths of the exchangeable pig-ment pool was present in the extravascular com-partments.
3) In the absence of a functioning conjugatingapparatus, elimination of bilirubin is accomplishedthrough alternate metabolic pathways. These in-clude breakdown of the pigment to more polar,diazo-negative derivatives, transfer of pigmentacross the intestinal mucosa into the gut, and ex-cretion of unconjugated bilirubin in the bile.When the combined functional rate of these path-ways equals that of pigment formation, a "steadystate" is reached, leading to constant levels ofserum bilirubin.
4) In hyperbilirubinemia of this type, the inter-action of the pigment with serum albumin appearsto be the major determinant regulating the serumconcentration and the mode of disposition ofbilirubin.
ACKNOWLEDGMENT
We are grateful to Drs. Norbert Freinkel and RogerLester for their advice in this study and in the prepara-tion of the manuscript.
REFERENCES
1. Schmid, R., and L. Hammaker. Metabolism anddisposition of C14-bilirubin in congenital non-he-molytic jaundice. Trans. Ass. Amer. Phycns 1962,75, 220.
2. Weinbren, K., and B. H. Billing. Hepatic clear-ance of bilirubin as an index of cellular function inthe regenerating rat liver. Brit. J. exp. Path. 1956,37, 199.
3. Arias, I. M., L. Johnson, and S. Wolfson. Biliaryexcretion of injected conjugated and unconjugatedbilirubin by normal and Gunn rats. Amer. J.Physiol. 1961, 200, 1091.
4. Ostrow, J. D., J. H. Jandl, and R. Schmid. Theformation of bilirubin from hemoglobin in vivo.J. clin. Invest. 1962, 41, 1628.
5. Williams, R., and B. H. Billing. Action of steroidtherapy in jaundice. Lancet, 1961, 2, 392.
6. Schmid, R. Some aspects of bile pigment metabolism.Clin. Chem. 1957, suppl. 3, 394.
1732

METABOLISMAND DISPOSITION OF C"4-BILIRUBIN
7. Crigler, J. F., Jr., and V. A. Najjar. Congenitalfamilial nonhemolytic jaundice with kernicterus.Pediatrics 1952, 10, 169.
8. Schmid, R. Hyperbilirubinemia in The MetabolicBasis of Inherited Disease, J. B. Stanbury, J. B.\Vyngaarden, and D. S. Fredrickson, Eds. NewYork, Blakiston, 1960, p. 226.
9. Gunn, C. K. Hereditary acholuric jaundice. J.Hered. 1938, 29, 137.
10. Rutstein, D. D., E. F. Ingenito, and W. E. Reynolds.The determination of albumin in human bloodplasma and serum. A method based on the inter-action of albumin with an anionic dye-2- (4'-hy-droxybenzeneazo) benzoic acid. J. clin. Invest.1954, 33, 211.
11. Ducci, H., and C. J. Watson. The quantitative de-termination of the serum bilirubin with specialreference to the prompt-reacting and the chloro-form soluble types. J. Lab. clin. Med. 1945, 30,293.
12. Schmid, R. The identification of "direct-reacting"bilirubin as bilirubin glucuronide. J. biol. Chem.1957, 229, 881.
13. Hawkinson, V., C. J. Watson, and R. H. Turner. Amodification of Harrison's test for bilirubin in theurine especially suited for mass and serial use.
J. Amer. med. Ass. 1945, 129, 514.14. Schwartz, S., V. Sborov, and C. J. Watson. Stud-
ies of urobilinogen. IV. The quantitative deter-mination of urobilinogen by means of the Evelynphotoelectric colorimeter. Amer. J. clin. Path.1944, 14, 598.
15. Schmid, R., and L. Hammaker. Glucuronide forma-tion in patients with constitutional hepatic dys-function (Gilbert's disease). New Engl. J. Med.1959, 260, 1310.
16. Brodie, B. B., and J. Axelrod. The estimation ofacetanilide and its metabolic products, anilineN-acetyl-p-aminophenol and p-aminophenol (freeand total conjugated) in biological fluids and tis-sues. J. pharmacol. exp. Ther. 1948, 94, 22.
17. Mollison, P. L. Blood Transfusion in Clinical Medi-cine, 2nd ed. Oxford, Blackwell, 1956, p. 39.
18. Ostrow, J. D., L. Hammaker, and R. Schmid. Thepreparation of crystalline bilirubin-C14. J. clin.Invest. 1961, 40, 1442.
19. Ostrow, J. D., and R. Schmid. The protein-bindingof C14-bilirubin in human and murine serum. J.clin. Invest. 1963, 42, 1286.
20. Najjar, V. A., and B. Childs. The crystallization andproperties of serum bilirubin. J. biol. Chem.1953, 204, 359.
21. Lester, R., and R. Schmid. Intestinal absorptionof bile pigments. I. The enterohepatic circula-tion of bilirubin in the rat. J. clin. Invest. 1963,42, 736.
22. Gilbertsen, A. S., P. T. Lowry, V. Hawkinson, andC. J. Watson. Studies of the dipyrrylmethene("fuscin") pigments. I. The anabolic significance
of the fecal mesobilifuscin. J. clin. Invest. 1959,38, 1166.
23. Robinson, S., T. Vanier, J. F. Desforges, and R.Schmid. Jaundice in thalassemia minor: a con-sequence of "ineffective erythropoiesis." NewEngl. J. Med. 1962, 267, 523.
24. Belcher, E. H., and E. B. Harriss. Studies ofplasma volume, red cell volume and total bloodvolume in young growing rats. J. Physiol. (Lond.)1957, 139, 64.
25. Lowry, P. T., I. Bossenmaier, and C. J. Watson.A method for the isolation of bilirubin from feces.J. biol. Chem. 1953, 202, 305.
26. Stahl, E. Duennschicht-Chromatographie. IV. Mit-teilung: Einsatzschema, Randeffekt, "saure undbasische" Schichten, Stufentechnik. Arch. Pharm.(Weinheim) 1959, 292, 411.
27. Gray, C. H. The Bile Pigments. London, Methuen,1953, p. 16.
28. Gray, C. H. The Bile Pigments. London, Methuen,1953, p. 24.
29. Dewey, W. C. Vascular-extravascular exchange of1P3' plasma proteins in the rat. Amer. J. Physiol.1959, 197, 423.
30. Schmid, R., J. Axelrod, L. Hammaker, and R. L.Swarm. Congenital jaundice in rats, due to adefect in glucuronide formation. J. clin. Invest.1958, 37, 1123.
31. Snedecor, G. W. Statistical Methods Applied toExperiments in Agriculture and Biology. Ames,Iowa, Iowa State College Press, 1946.
32. Solomon, A. K. Equations for tracer experiments.J. clin. Invest. 1949, 28, 1297.
33. With, T. K. Biologie der Gallenfarbstoffe. Stutt-gart, Thieme, 1960, p. 33.
34. With, T. K. Biology of Bile Pigments. Copen-hagen, Frost-Hansen, 1954, p. 383.
35. Weech, A. A., D. Vann, and R. A. Grillo. Theclearance of bilirubin from the plasma. A meas-ure of the excreting power of the liver. J. clin.Invest. 1941, 20, 323.
36. Harris, J. W. The Red Cell. Cambridge, Mass.,Harvard University Press, 1963, p. 235.
37. London, I. M., R. West, D. Shemin, and D. Ritten-berg. On the origin of bile pigment in normalman. J. biol. Chem. 1950, 184, 351.
38. Belcher, E. H., and E. B. Harriss. Studies of redcell life span in the rat. J. Physiol. (Lond.)1959, 146, 217.
39. Hughes Jones, N. C., and B. Cheney. The use of5"Cr and 'Fe as red cell labels to determine thefate of normal erythrocytes in the rat. Clin. Sci.1961, 20, 323.
40. Beeken, W. L., W. Volwiler, P. D. Goldsworthy,L. E. Garby, W. E. Reynolds, R. Stogsdill, andR. S. Stemler. Studies of I"'l-albumin catabolismand distribution in normal young male adults. J.clin. Invest. 1962, 41, 1312.
1733

RUDI SCHMID AND LYDIA HAMMAKER
41. Takeda, Y., and E. B. Reeve. Studies of the metab-olism and distribution of albumin with autologousI13'-albumin in healthy men. J. Lab. clin. Med.1963, 61, 183.
42. Barac, G., and J. M. Gernay. Recherches sur lacenapse albumino-bilirubinique. Bull. Soc. Chim.biol. (Paris) 1949, 31, 128.
43. Campbell, R. M., D. P. Cuthbertson, C. M. E. Mat-thews, and A. S. McFarlane. Behaviour of 14C-and 131I-labelled plasma proteins in the rat. Int.J. appl. Radiat. 1956, 1, 66.
44. Blanc, W. A., and L. Johnson. Studies on kernic-terus: relationship with sulfonamide intoxication,report on kernicterus in rats with glucuronyltransferase deficiency and review of pathogenesis.J. Neuropath. exp. Neurol. 1959, 18, 165.
45. Arias, I. M. The chemical basis of kernicterus inAdvances in Clinical Chemistry, H. Sobotka andC. P. Stewart, Eds. New York, Academic Press,1960, vol. 3, p. 45.
46. Weber, A. P., L. Schalm, and J. Witmans. Bili-rubin monoglucuronide (pigment I): a complex.Acta med. scand. 1963, 173, 19.
47. Watson, C. J., M. Campbell, and P. T. Lowry.Preferential reduction of conjugated bilirubin to
urobilinogen by normal fecal flora. Proc. Soc.exp. Biol. (N. Y.) 1958, 98, 707.
43. Isselbacher, K. J., and E. A. McCarthy. Studieson bilirubin sulfate and other nonglucuronide con-jugates of bilirubin. J. clin. Invest. 1959, 38, 645.
49. Lester, R., L. Hammaker, and R. Schmid. A newtherapeutic approach to unconjugated hyperbili-rubinaemia. Lancet 1962, 2, 1257.
50. Blondheim, S. H., D. Lathrop, and J. Zabriskie.Effect of light on the absorption spectrum ofjaundiced serum. J. Lab. clin. Med. 1962, 60,31.
51. Cremer, R. J., P. W. Perryman, and D. H. Richards.Influence of light on hyperbilirubinaemia of in-fants. Lancet 1958, 1, 1094.
52. Mellone, 0. Treatment of hyperbilirubinemia of thenewborn with an intense light focus. Rev. Soc.paul. Med. vet. 1960, 57, 47.
53. Silverman, W. A., D. H. Andersen, W. A. Blanc,and D. N. Crozier. A difference in mortalityrate and incidence of kernicterus among prematureinfants allotted to two prophylactic antibacterialregimens. Pediatrics 1956, 18, 614.
54. Odell, G. B. Studies in kernicterus. I. The proteinbinding of bilirubin. J. clin. Invest. 1959, 38, 823.
1734


















