
IV. Célula procarionte y Eucarionte · único cromosoma bacteriano circular. ... citoplasma se...
50
IV. Célula procarionte y Eucarionte Asignatura: Biología Mg. Bárbara Cuevas Montuschi Universidad la República Escuela de Enfermería y Salud Pública Licenciatura en Enfermería
Transcript of IV. Célula procarionte y Eucarionte · único cromosoma bacteriano circular. ... citoplasma se...

IV. Célula procarionte y Eucarionte
Asignatura: Biología
Mg. Bárbara Cuevas Montuschi
Universidad la República Escuela de Enfermería y Salud Pública Licenciatura en Enfermería

Modelos Celulares 1. En ambos tipos celulares el ADN es el material genético. 2. Ambos tipos celulares poseen membranas plasmáticas como límite
celular. 3. Poseen ribosomas para la síntesis proteica. 4. Poseen un metabolismo básico similar 5. Ambos tipos celulares son muy diversos en formas y estructuras.

Todas las células, almacenan en forma de ADN, ácido desoxirribonucleico, a información necesaria para controlar sus actividades (reproducción, metabolismo), y para establecer su propia estructura. El ADN, es un polímero formado por una secuencia lineal, de monómeros, llamados nucleótidos. Esta secuencia de nucleótidos, especifica una secuencia de aminoácidos (estructura primaria de una proteína). La especificidad de la secuencia de aminoácidos determinada por la secuencia de bases del ADN esta regida por el código genético. La secuencia de bases del ADN, que codifica una proteína, es un GEN.

Todas las células están cubiertas por una membrana externa, llamada membrana plasmática, que las separa de otras células y del medio circundante con el cual intercambian materia y energía. Este intercambio esta altamente regulado y es selectivo. De esta forma la membrana plasmática debe actuar no sólo como limite celular sino también como barrera selectiva. Por lo tanto la célula, mantiene una composición química muy ordenada y diferente a la del entorno.
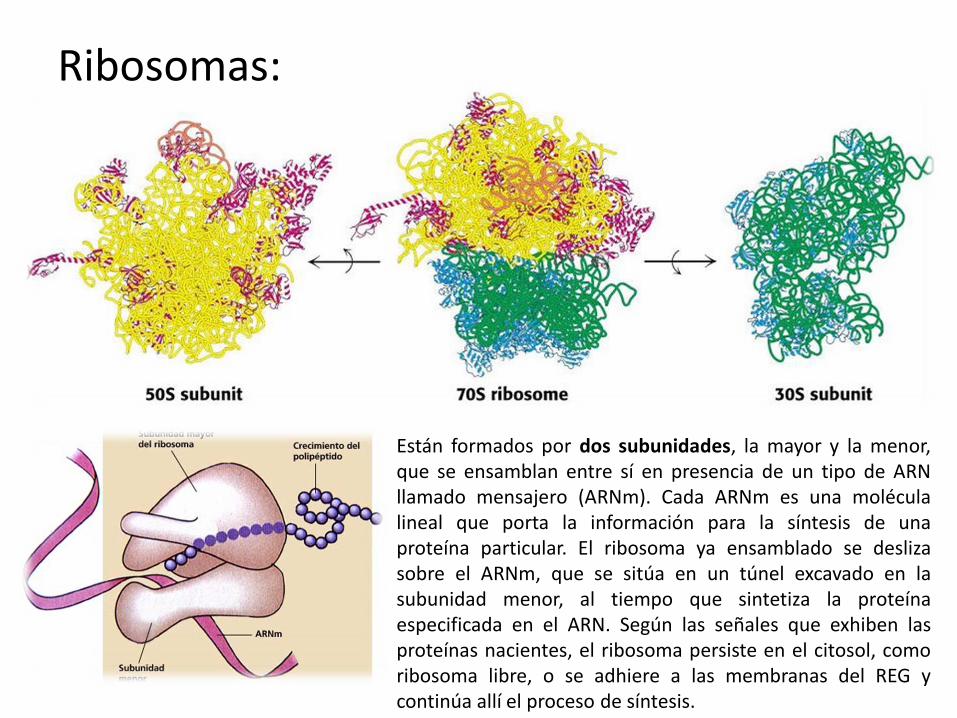
Ribosomas:
Están formados por dos subunidades, la mayor y la menor, que se ensamblan entre sí en presencia de un tipo de ARN llamado mensajero (ARNm). Cada ARNm es una molécula lineal que porta la información para la síntesis de una proteína particular. El ribosoma ya ensamblado se desliza sobre el ARNm, que se sitúa en un túnel excavado en la subunidad menor, al tiempo que sintetiza la proteína especificada en el ARN. Según las señales que exhiben las proteínas nacientes, el ribosoma persiste en el citosol, como ribosoma libre, o se adhiere a las membranas del REG y continúa allí el proceso de síntesis.

Todas las células poseen un metabolismo o conjunto de reacciones químicas, que posibilitan el mantenimiento de la vida. Este metabolismo para sustentarse necesita de una o más fuentes de energía. Las células, necesitan de distintivos tipos de moléculas energéticas: * Monedas energéticas, como el ATP. * Moléculas combustibles, como la glucosa o los ácidos grasos. * Moléculas de reserva de energía, como el glucógeno o el almidón.

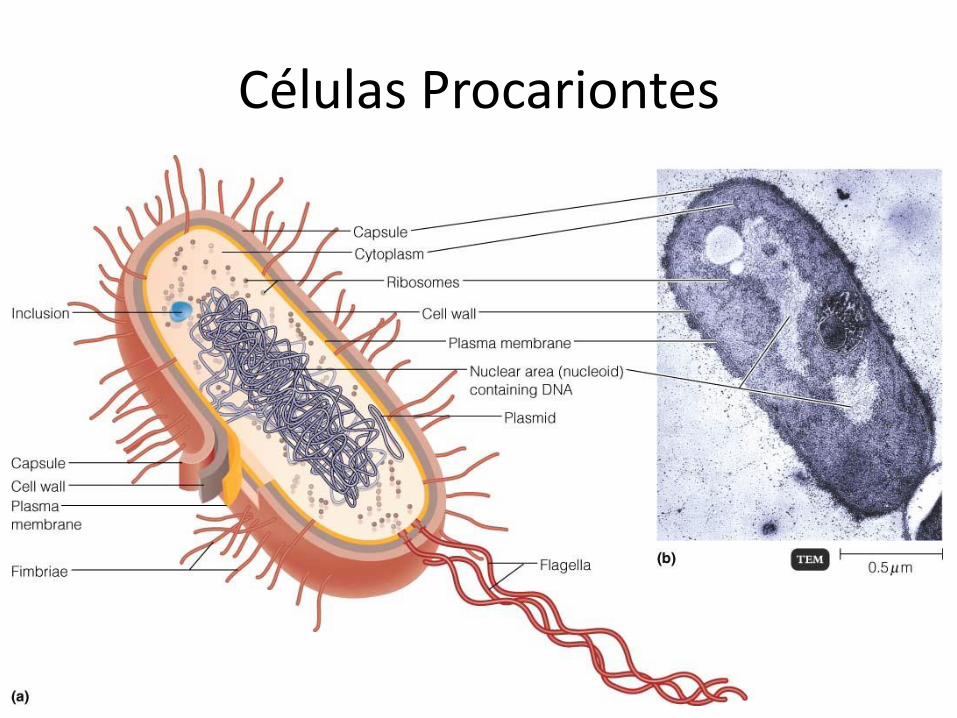
Células Procariontes


Las bacterias pueden definirse como organismos unicelulares procariontes que se reproducen por fisión binaria. Contienen toda su información genética en un único cromosoma bacteriano circular. También poseen sistemas productores de energía y biosintéticos necesarios para el crecimiento y la reproducción. Poseen como característica particular una pared rígida de peptidoglicanos. Son generalmente de vida libre y poseen ADN extracromosómico en forma de plásmidos, estos codifican genes de resistencia a antibióticos o factores "sexuales" como los pili.

Cromosoma bacteriano: nucleoide
El cromosoma de las células procariontes (cromosoma bacteriano) consiste en una sola molécula de ADN cerrada y dispuesta de modo compacto, aunque a diferencia de las células eucariontes no se encuentra rodeado de membrana nuclear. El cromosoma generalmente adopta una disposición redondeada y está unido a la membrana citoplasmática. El ADN se encuentra dividido en unidades funcionales o genes, los cuales pueden ser de dos tipos: a) aquellos cuya secuencia de bases codifica cadenas polipeptídicas o moléculas de ADN (genes estructurales), y b) los que únicamente tienen función reguladora de los anteriores (genes reguladores).
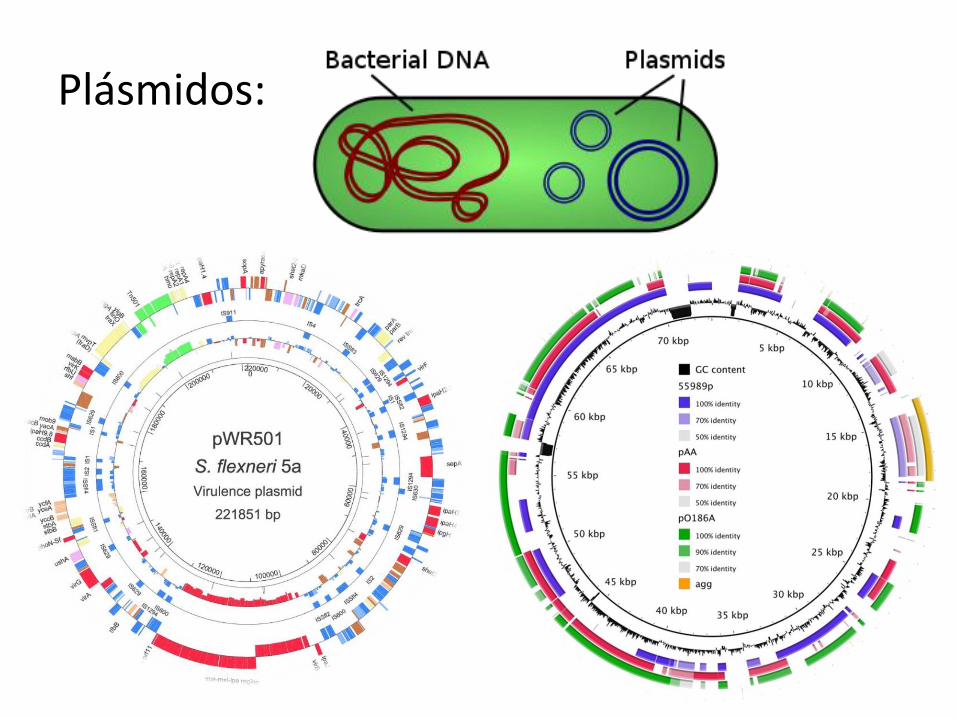
Plásmidos:

Un plásmido es una molécula de ADN extracromosómico que se replica en forma autónoma, por lo que al igual que el cromosoma es un replicón. Puede haber hasta 50 copias de un plásmido en una bacteria. Funcionalmente los plásmidos son elementos genéticos accesorios, es decir que la bacteria puede vivir sin ellos. Sin embargo, la información que contienen puede contribuir a la adaptación de la bacteria al medio y a la evolución de la misma. Los plásmidos pueden contener genes que codifican factores de resistencia a antibióticos (los plásmidos R) y factores de patogenicidad como exotoxinas. La evolución bacteriana a través de los plásmidos es factible, ya que pueden ser intercambiados entre distintas bacterias (por ejemplo, el plásmido F). Es decir que ciertos genes pueden transferirse de una bacteria otra mediante el pasaje de plásmidos, a este mecanismo se lo denomina conjugación.
Para que la conjugación pueda llevarse a cabo las dos bacterias deben ponerse en contacto físico. Esto es posible debido a que una de las bacterias, posee pili sexuales (pelos) en su envoltura.
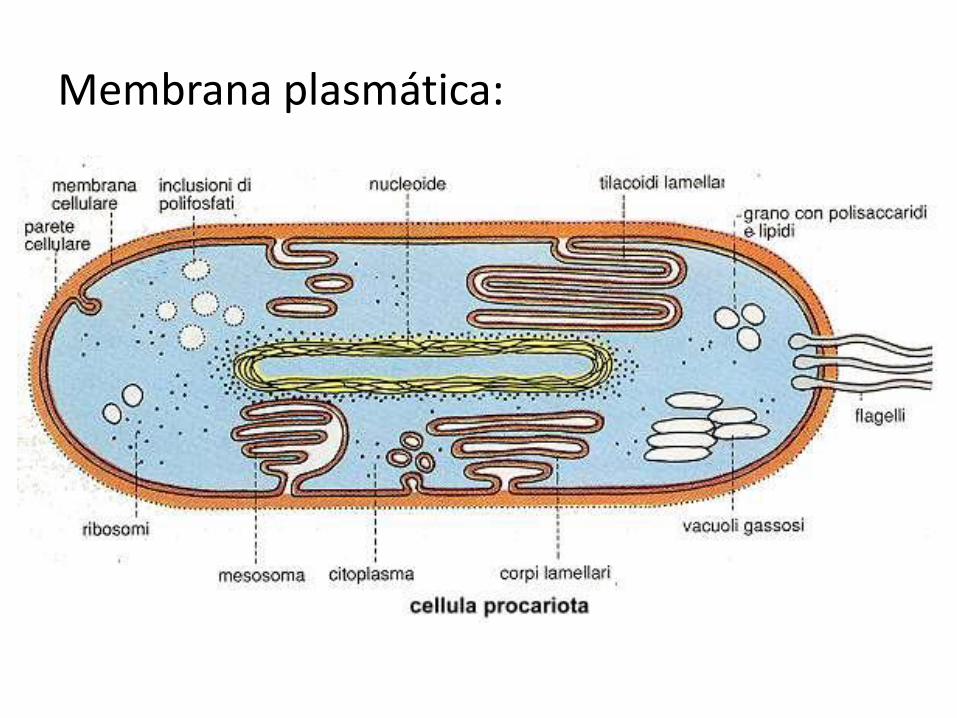
Membrana plasmática:

Pared celular de las bacterias: peptidoglicano

Por fuera de la membrana celular, se encuentra una pared celular rígida de peptidoglicano, que esta presente en todas las bacterias excepto los micoplasmas. La presencia de la pared protege a la bacteria de la diferencia de presión osmótica entre el medio interno de la bacteria y el medio exterior. De no existir la pared la bacteria estallaría. Además la pared cumple funciones de protección como por ejemplo contra sustancias tóxicas . Existen dos tipos de pared bacteriana que pueden diferenciarse por la Tinción de Gram. El primer grupo de bacterias son aquellas capaces de retener el colorante cristal violeta luego de la decoloración con alcohol-cetona. Estas bacterias son llamadas Grampositivas. El segundo grupo esta conformado por aquellas bacterias incapaces de retener el colorante luego del tratamiento decolorante, por lo tanto son llamadas Gramnegativas.

Células Eucariontes

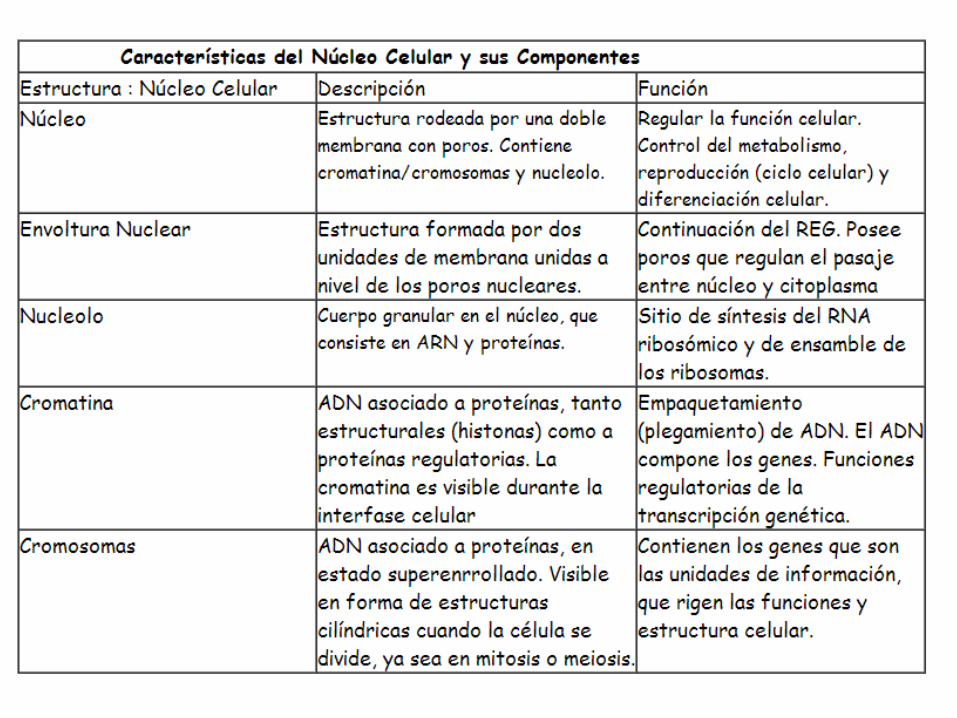
Núcleo celular

La cromatina es un material fibroso formado por la asociación entre ADN y proteínas, de las cuales las proteínas básicas llamadas histonas son las más abundantes. Las largas moléculas de ADN pueden ser empacadas dentro del núcleo gracias a que se enrollan alrededor de complejos de histonas. Las fibras de cromatina presentan diferentes grados de plegamiento, que resultan en una estructura cada vez más compacta. El grado mayor de plegamiento se alcanza durante la división celular. Entonces la cromatina da lugar a la formación de unas estructuras bien visibles al microscopio óptico, los cromosomas.

Retículo endoplasmático

El RER está formado por cisternas o bolsas aplanadas, conectadas unas a otras mediante túbulos membranosos. Sobre la cara de las membranas que da al citoplasma se apoyan ribosomas. Las proteínas sintetizadas por estos organelos atraviesan las membranas del REG. Algunas de ellas son volcadas hacia el lumen o cavidad del retículo, mientras que otras quedan ancladas a la membrana. Las proteínas que llevan la señal adecuada y consiguientemente son sintetizadas en ribosomas que se adhieren al retículo, son las siguientes: proteínas integrantes del sistema de endomembranas, proteínas lisosomales, proteínas destinadas a la membrana plasmática y proteínas de secreción. Al mismo tiempo que las cadenas peptídicas son translocadas hacia el lumen del REG sufren casi siempre un proceso de glicosilación, que consiste en la adición de una o más moléculas de oligosacárido.
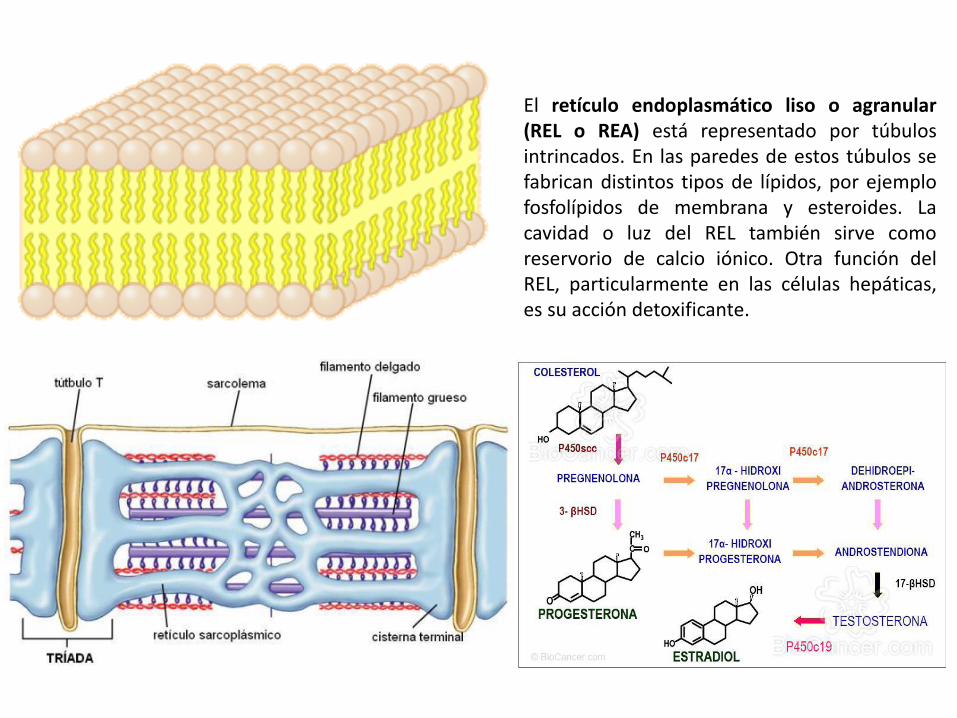
El retículo endoplasmático liso o agranular (REL o REA) está representado por túbulos intrincados. En las paredes de estos túbulos se fabrican distintos tipos de lípidos, por ejemplo fosfolípidos de membrana y esteroides. La cavidad o luz del REL también sirve como reservorio de calcio iónico. Otra función del REL, particularmente en las células hepáticas, es su acción detoxificante.

Aparato de Golgi
El aparato o complejo de Golgi consta de bolsas circulares apiladas unas sobre otras. Allí se sintetizan algunos glúcidos complejos y además se reciben vesículas de transporte con las moléculas elaboradas por el REG y el REL. Las vesículas o vacuolas son pequeñas bolsas membranosas que se desprenden como brotes de los dos sectores del retículo endoplasmático y sirven como “empaque” y vehículo de transporte para los productos allí fabricados. Las vesículas de transporte son desplazadas por el citosol hasta llegar al aparato de Golgi. Este complejo presenta una cara de recepción (cara cis), a la cual se fusionan las vesículas, entregando su cargamento. Ya dentro del aparato de Golgi, los distintos cargamentos son modificados, concentrados y clasificados. Así, según su destino, son empacados en nuevas vesículas que brotan de la cara opuesta del aparato de Golgi, la cara de emisión (cara trans). Algunas de las vesículas emitidas por el aparato de Golgi son lisosomas; otras son vesículas de secreción.

Lisosomas
Los lisosomas contienen las proteínas lisosomales originadas en el REG. Las mismas son enzimas hidrolíticas y participan en procesos de digestión
intracelular. Las vesículas de secreción se fusionan con la membrana plasmática. Esto permite volcar su contenido en el medio extracelular, donde cumplirá una función, ya sea hormonal, enzimática, estructural, etc. La fusión de vesículas con la membrana plasmática también es el medio para que ésta incorpore nuevos componentes, lípidos o proteínas, sintetizados en el sistema de endomembranas y transportados como parte integrante de la membrana vesicular.
http://highered.mcgraw-hill.com/olcweb/cgi/pluginpop.cgi?it=swf::535::535::/sites/dl/free/0072437316/120067/bio01.swf::Lysosomes
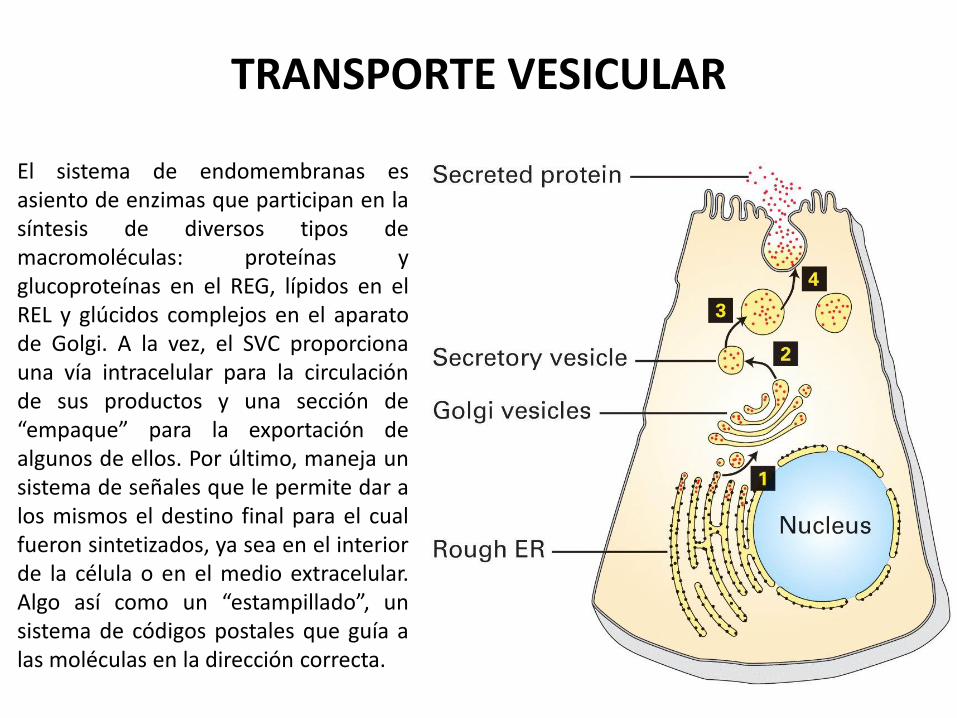
TRANSPORTE VESICULAR
El sistema de endomembranas es asiento de enzimas que participan en la síntesis de diversos tipos de macromoléculas: proteínas y glucoproteínas en el REG, lípidos en el REL y glúcidos complejos en el aparato de Golgi. A la vez, el SVC proporciona una vía intracelular para la circulación de sus productos y una sección de “empaque” para la exportación de algunos de ellos. Por último, maneja un sistema de señales que le permite dar a los mismos el destino final para el cual fueron sintetizados, ya sea en el interior de la célula o en el medio extracelular. Algo así como un “estampillado”, un sistema de códigos postales que guía a las moléculas en la dirección correcta.


Peroxisomas Contienen enzimas que participan en procesos oxidativos; algunas de ellas tienden a formar un cuerpo cristalino. Son, junto con las mitocondrias, los organelos donde se consume el oxígeno que llega a las células, aunque, a diferencia de lo que ocurre en las mitocondrias, las reacciones que transcurren en los peroxisomas no generan ATP. Especialmente en células hepáticas, las reacciones oxidativas de los peroxisomas posibilitan la metabolización de sustancias tóxicas, como el etanol o alcohol etílico. El peróxido de hidrógeno (agua oxigenada) se forma como producto de estas reacciones de detoxificación. Una enzima característica del peroxisoma es la catalasa, que cataliza la descomposición del exceso de peróxido de hidrógeno allí generado.

Detoxificación celular y beta oxidación de los ácidos grasos:

Mitocondrias y cloroplastos:
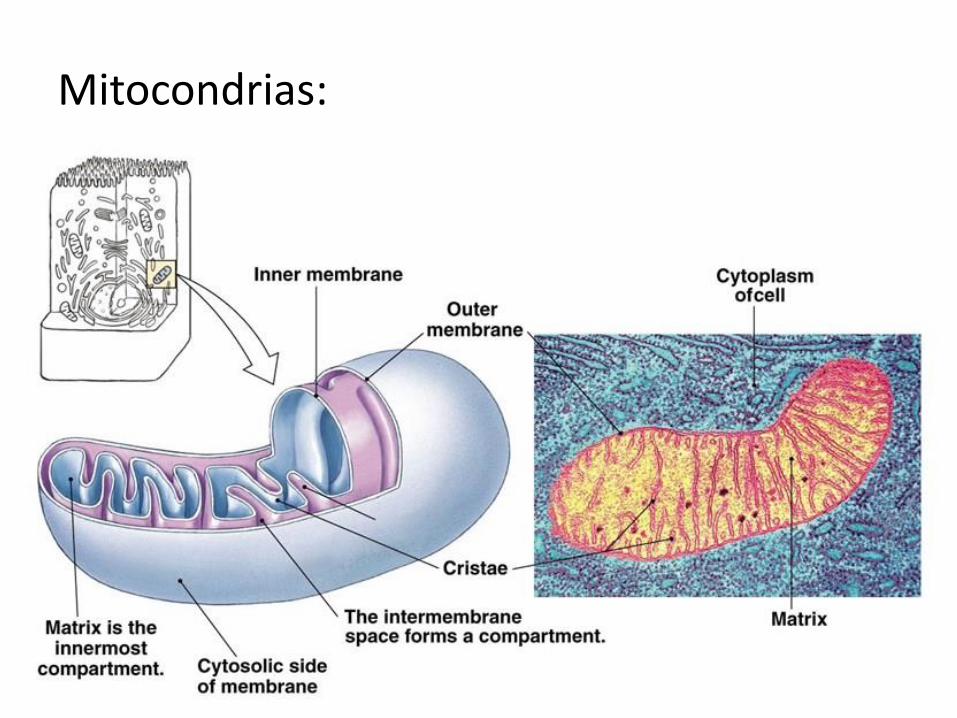
Mitocondrias:


Cloroplastos:



Vacuola:


Citoplasma
El citoplasma es la zona que se ubica entre la membrana plasmática y el núcleo. La parte líquida del citoplasma se denomina citosol o matriz citoplasmática. La matriz citoplasmática es un medio acuoso que contiene iones y moléculas pequeñas disueltas y también muchos tipos de macromoléculas en suspensión; entre ellas, enzimas, que participan en importantes vías metabólicas. En algunos casos, el citosol tiene inclusiones, que son depósitos de distintas sustancias. Por ejemplo, en las células del tejido adiposo grandes gotas de triglicéridos se reservan en el citosol. Sin embargo, el citoplasma está lejos de ser una masa homogénea e informe. Por el contrario, el citosol está interrumpido y recorrido por una serie de estructuras complejas y especializadas: el citoesqueleto, el sistema de endomembranas y los organelos citoplasmáticos.

Citoesqueleto
El citoesqueleto es propio de las células eucarióticas. Es una estructura tridimensional dinámica que se extiende a través del citoplasma. Por lo tanto la idea de que el citoplasma de la célula es una masa amorfa y gelatinosa es equivocada. Esta matriz fibrosa de proteínas se extiende por el citoplasma entre el núcleo y la cara interna de la membrana plasmática, ayudando a definir la forma de la célula e interviniendo en la locomoción y división celular. Funciones del citoesqueleto: - Estabilidad celular y forma celular - Locomoción celular - División celular - Movimiento de los orgánulos internos - Regulación metabólica


Pared Celular

Uniones Celulares

Las uniones celulares mantienen unidas a las células para formar tejidos

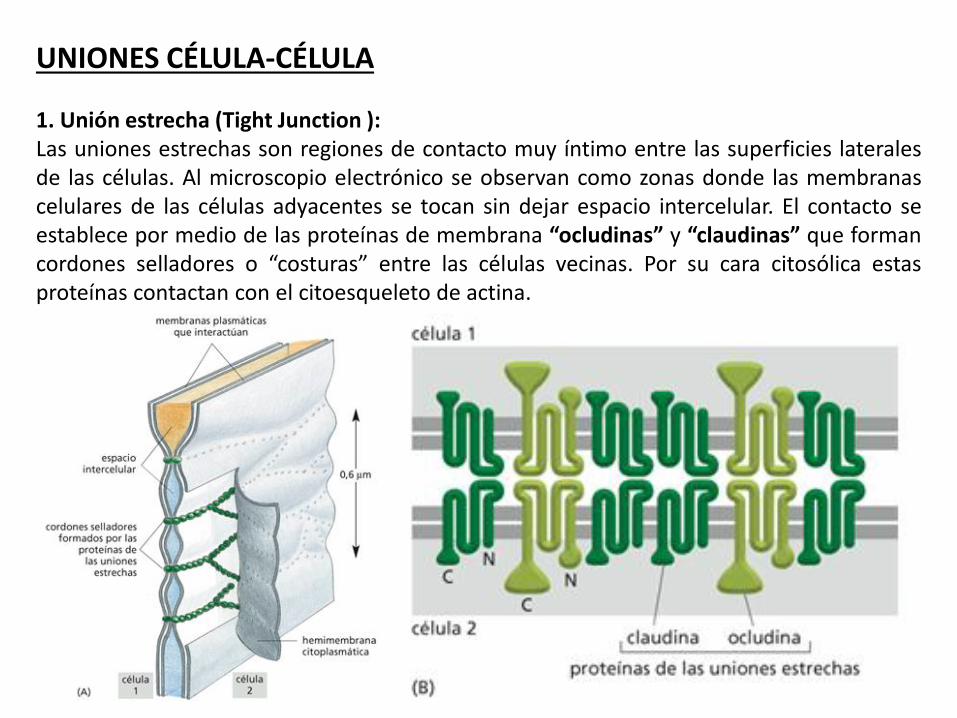
UNIONES CÉLULA-CÉLULA 1. Unión estrecha (Tight Junction ): Las uniones estrechas son regiones de contacto muy íntimo entre las superficies laterales de las células. Al microscopio electrónico se observan como zonas donde las membranas celulares de las células adyacentes se tocan sin dejar espacio intercelular. El contacto se establece por medio de las proteínas de membrana “ocludinas” y “claudinas” que forman cordones selladores o “costuras” entre las células vecinas. Por su cara citosólica estas proteínas contactan con el citoesqueleto de actina.

2. Unión adherente : Las uniones adherentes contribuyen a dar cohesión al tejido. Se encuentran en la superficie lateral formando un cinturón que rodea a la célula y la mantiene adherida a las células vecinas. La adhesión depende de unas proteínas de membrana llamadas “cadherinas”. Éstas presentan una zona o dominio que se proyecta hacia el exterior, haciendo contacto con las cadherinas de la célula adyacente, como si fueran los dientes de un cierre. El dominio de las cadherinas que se proyecta hacia el citosol se conecta con microfilamentos, los cuales cumplen con funciones de fijación y contracción.

3. Desmosoma: Los desmosomas han sido comparados con “remaches” que unen fuertemente las membranas de células adyacentes. El contacto entre las células está mediado por cadherinas. En la cara citosólica de la membrana, las cadherinas se unen a una placa densa formada por desmina (una proteína ligadora). La desmina establece contacto con filamentos intermedios de queratina. Los desmosomas proporcionan resistencia a las tensiones a que pueden verse sometidos los epitelios.
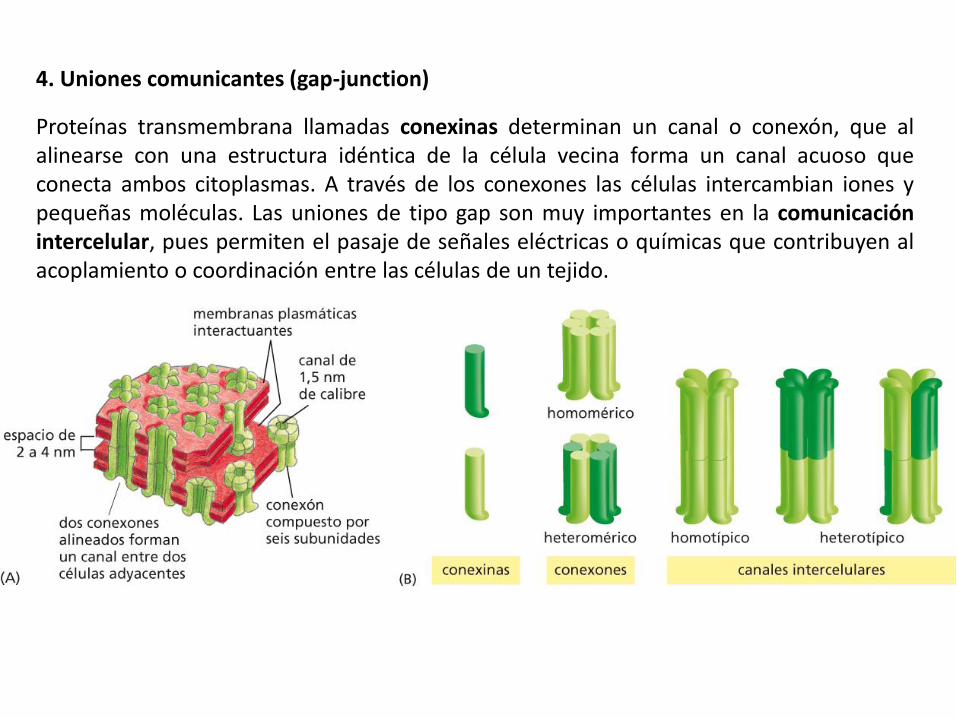
4. Uniones comunicantes (gap-junction)
Proteínas transmembrana llamadas conexinas determinan un canal o conexón, que al alinearse con una estructura idéntica de la célula vecina forma un canal acuoso que conecta ambos citoplasmas. A través de los conexones las células intercambian iones y pequeñas moléculas. Las uniones de tipo gap son muy importantes en la comunicación intercelular, pues permiten el pasaje de señales eléctricas o químicas que contribuyen al acoplamiento o coordinación entre las células de un tejido.

UNIONES CÉLULA-MATRIZ Hemidesmosomas: son uniones de anclaje entre las células y la matriz extracelular (MEC). Se encuentran en la superficie basal de los epitelios, donde éstos establecen contacto con una zona especializada de la MEC, llamada lámina basal. Su nombre deriva de su semejanza con los desmosomas, aunque solo corresponden a la mitad de estos últimos. En la unión de la célula con la matriz intervienen proteínas transmembranares de la familia de la “integrinas”. Las integrinas se unen con los microfilamentos del citoesqueleto, por un lado, y con componentes de la MEC por el otro; entre ellos, laminina y colágeno.
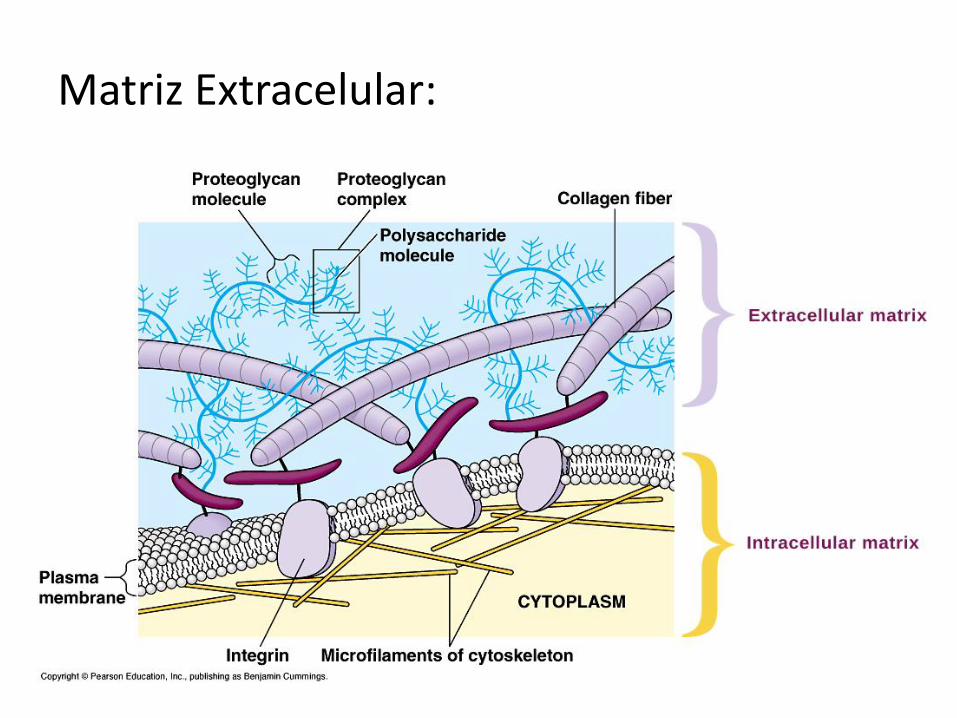
Matriz Extracelular:


















