
ISSN: 2155-9546 - American University of Beirutstaff.aub.edu.lb/~is08/Merrifield trout...
7
http://www.omicsonline.org/jardhome.php Gabor Mocz, University of Hawaii, USA Konrad Dabrowski, The Ohio State, University Ohio USA Covadonga R. Arias, Auburn University Alabama, USA Brian S. Shepherd, University of Wisconsin Milwaukee, USA Attila Karsi, Mississippi State University Mississippi, USA Kevin K. Schrader, Oxford University MississippiM, USA Paul V. Zimba, C Texas A&M University-Corpus Christi Texas, USA Kenneth D. Cain, University of Idaho Moscow, USA Klaus Becker, University of Hohenheim Stuttgart, Germany Toshihiro Nakai, Hiroshima University, Japan Keisuke G. Takahashi, Tohoku University Miyagi, Japan Marisa Manzano, University of Udine Udine, Italy Jorge Fernandes, Bodø University College, Norway Shiro Itoi, Nihon University, Fujisawa, JAPAN Daniel L. Merrifield, The University of Plymouth, UK Francesco buonocore, University of Tuscia Viterbo, Italy S.A.M. Martin, University of Aberdeen Aberdeen, UK Molly Twinomwe Kibenge, University of Prince Edward Island, Canada Monique Mancuso, Institute for Coastal Marine Environment of Messina, Italy Mauro Lenzi, Lagoon Ecology and Aquaculture Laboratory Orbetello, Italy Bruno Gomez-Gil, Mazatlán Unit for Aquaculture & Environmental Management, Mexico Tiantian Zhang, University of Bedfordshire, UK Patrick T.K. Woo, University of Guelph, Canada Antonino Nizza, University of Naples Naples, Italy John L. Zhou, London South Bank University, UK A quaculture plays a vital role in the economical betterment of a country. Aquaculture also termed as aqua farming is the cultivation of plants or animals such as fish, shellfish and seaweed, in natural or controlled marine or freshwater environments. Journal of Aquaculture Research & Development is an International, open access journal which publishes hypothesis-driven papers on applied or scientific research relevant to freshwater aquaculture, brackish water aquaculture and marine aquaculture. Journal of Aquaculture Research & Development - Open Access using online manuscript submission, review and tracking systems of Editorial Manager® for quality and quick review processing. Submit your manuscript at http://www.editorialmanager.com/environsci/ ISSN: 2155-9546 OMICS Publishing Group 5716 Corsa Ave., Suite 110, Westlake, Los Angeles, CA 91362-7354, USA, Phone: +1- 650-268-9744, Fax: +1-650-618-1414, Toll free: +1-800-216-6499
Transcript of ISSN: 2155-9546 - American University of Beirutstaff.aub.edu.lb/~is08/Merrifield trout...

http://www.omicsonline.org/jardhome.php
Gabor Mocz,University of Hawaii,
USA
Konrad Dabrowski,The Ohio State, University Ohio
USA
Covadonga R. Arias,Auburn University
Alabama, USA
Brian S. Shepherd,University of Wisconsin
Milwaukee, USA
Attila Karsi,Mississippi State University
Mississippi, USA
Kevin K. Schrader,Oxford UniversityMississippiM, USA
Paul V. Zimba,C Texas A&M
University-Corpus Christi Texas, USA
Kenneth D. Cain,University of Idaho
Moscow, USA
Klaus Becker,University of Hohenheim
Stuttgart, Germany
Toshihiro Nakai,Hiroshima University,
Japan
Keisuke G. Takahashi,Tohoku University
Miyagi, Japan
Marisa Manzano,University of Udine
Udine, Italy
Jorge Fernandes,Bodø University College, Norway
Shiro Itoi,Nihon University, Fujisawa, JAPAN
Daniel L. Merrifield,The University of
Plymouth, UK
Francesco buonocore,
University of TusciaViterbo, Italy
S.A.M. Martin,University of Aberdeen
Aberdeen, UK
Molly Twinomwe Kibenge,
University of Prince Edward Island, Canada
Monique Mancuso,Institute for Coastal
Marine Environment of Messina, Italy
Mauro Lenzi,Lagoon Ecology and
Aquaculture LaboratoryOrbetello, Italy
Bruno Gomez-Gil,Mazatlán Unit for
Aquaculture & Environmental
Management, Mexico
Tiantian Zhang,University of
Bedfordshire, UK
Patrick T.K. Woo,University of Guelph,
Canada
Antonino Nizza,University of Naples
Naples, Italy
John L. Zhou,London South Bank
University, UK
Aquaculture plays a vital role in the economical betterment of a
country. Aquaculture also termed as aqua farming is the cultivation of plants or animals such as fish, shellfish and seaweed, in natural or controlled marine or freshwater environments.
Journal of Aquaculture Research & Development is an International, open access journal which publishes hypothesis-driven papers on applied or scientific research relevant to freshwater aquaculture, brackish water aquaculture and marine aquaculture.
Journal of Aquaculture Research & Development - Open Access using online manuscript submission, review and tracking systems of Editorial Manager® for quality and quick review processing. Submit your manuscript athttp://www.editorialmanager.com/environsci/
ISSN: 2155-9546
OMICS Publishing Group5716 Corsa Ave., Suite 110, Westlake, Los Angeles, CA 91362-7354, USA, Phone: +1- 650-268-9744, Fax: +1-650-618-1414, Toll free: +1-800-216-6499

Open Access
Special Issue 1 • 2011J Aquac Res DevelopmentISSN: 2155-9546 JARD, an open access journal
Research Article Open Access
Harper et al. J Aquac Res Development 2011, S1http://dx.doi.org/10.4172/2155-9546.S1-004
Research Article Open Access
AquacultureResearch & Development
Keywords: Fish, Vibriosis, Disease, Health, Microbiology, Bactocell
IntroductionEvidence suggests that the gastrointestinal (GI) tract of fish is a major
portal of entry for a range of fish pathogens including Vibrio (Listonella) anguillarum, the causative agent of vibriosis [1,2]. Recent studies have sought to use probiotics to fortify the intestinal microbiome of fish and enhance the defensive barrier against such diseases [3,4]. Such studies have shown a great deal of success and several probiotic strains have been demonstrated to elevate disease resistance of trout to pathogenic bacteria [3,5-7]. The mechanisms which mediate these benefits are thought to result from host immunostimulation and competitive exclusion. Such conclusions are based on empirical evidence but information regarding probiotic-pathogen antagonism in fish studies are primarily in vitro based, with evidence that probiotics are able to outcompete or antagonise (via the production of bacteriocins, organic acids, siderophores, competition for adhesion sites etc) pathogens on agar plates, broth cultures or in adhesion assays. However, such information does not fully represent probiotic vs. pathogen interactions in situ (within the fish GI tract) where localised responses such as gut associated lymphoid tissues (GALT) and antimicrobial properties of intestinal mucus are likely to be an important factor. Therefore, we aim to further the present knowledge by observing the antagonistic relationship of probiotics and pathogens in the intestine of fish by using the ex vivo intestinal sac method [8,9]. Due to previous evidence suggesting a positive role as a probiotic for fish [10-12], Pediococcus acidilactici was used as the probiotic and because of pathogenicity and industrial relevance, V. anguillarum was used as the pathogen in the present study.
Materials and MethodsExperimental fish and diets
Female rainbow trout were acquired from a local fish farm (Hatchlands Fisheries, Greyshoot Lane, Rattery, South Brent, UK) and transported to the Aquaculture and Fish Nutrition Research facility at The Univeristy of Plymouth, UK. Fish were offered a commercial trout diet (Skretting, Skretting, 43% protein, 20% lipid) ad libitum for four weeks. After grading, batches of 20 fish (240-250g) were randomly distributed into six 150 L fibreglass tanks, each provided with 98% re-circulated aerated freshwater at a rate of 150 L hr-1. Tanks were randomly divided among two treatments with three replicate tanks per treatment. Fish in treatment-1 were offered a control diet whilst fish in treatment-2 were offered the same diet supplemented with P.
*Corresponding author: Daniel L. Merrifield, The School of Biomedical and Biological Sciences, The University of Plymouth, Plymouth, UK. Tel: +44 -0- 1752 584888; E-mail: [email protected]
Received July 04, 2011; Accepted September 01, 2011; Published September 10, 2011
Citation: Harper GM, Monfort M, Saoud IP, Emery M, Mustafa S, et al. (2011) An ex vivo approach to studying the interactions of probiotic Pediococcus acidilactici and Vibrio (Listonella) anguillarum in the anterior intestine of rainbow trout Oncorhynchus mykiss. J Aquac Res Development S1:004. doi:10.4172/2155-9546.S1-004
Copyright: © 2011 Harper GM, et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
AbstractThe aim of the present study was to observe the antagonistic relationship of the probiotic Pediococcus acidilactici
and the pathogen Vibrio (Listonella) anguillarum in the intestine of rainbow trout (Oncorhynchus mykiss) by using the ex vivo intestinal sac method. Rainbow trout (240-250g) were fed either a control diet (commercial diet: 43% protein, 20% lipid) or a probiotic diet (control diet supplemented with P. acidilactici [Bactocell] at log 7 CFU g-1) for two weeks. The anterior intestine was then isolated for an ex vivo challenge study and intestinal sacs formed by shutting one end of the intestinal portion. The sacs were filled with PBS solutions containing either no bacteria (as a control), the probiotic (P. acidilactici), the pathogen Vibrio anguillarum, or probiotic+pathogen and incubated for one hour. At the end of the exposure bacterial levels in the lumen were determined by culture based approaches and colonisation of the mucosa assessed with PCR-DGGE and electron microscopy (EM). Intestinal morphology (observation of gross morphological damage, ultrstructural differences and the quantification of goblet cells and intra epithelial lecuocyte numbers) and bacterial infection/translocation was assessed using light microscopy (LM) and EM.
Results revealed that V. anguillarum caused extensive histological damage to the gut but P. acidilactici did not. No signs of translocation of either the probiotic or pathogen were observed. Microbiological analyses indicated that P. acidilactici was able to outcompete V. anguillarum in the rainbow trout intestine and also to populate or colonise the mucosa. Additionally, elevated leucocyte levels and goblet cells in the epithelium of P. acidilactici fed fish, and intestines exposed to P. acidilactici, suggests that P. acidilactici might have potential use in controlling vibriosis. In vivo disease-challenge studies are warranted to ascertain if V. anguillarum infections can be controlled in rainbow trout using dietary applications of P. acidilactici.
An ex vivo approach to studying the interactions of probiotic Pediococcus acidilactici and Vibrio (Listonella) anguillarum in the anterior intestine of rainbow trout Oncorhynchus mykiss Glenn M. Harper1, Imad P. Saoud2, Matthew Emery3, Sanaa Mustafa3, Mark Rawling3, Ben Eynon3, Simon J. Davies3 and Daniel L. Merrifield3*, Matthias Monfort1Electron Microscopy Centre, The University of Plymouth2American University of Beirut, Beirut, Lebanon3The School of Biomedical and Biological Sciences, The University of Plymouth, Plymouth, UK

Citation: Harper GM, Monfort M, Saoud IP, Emery M, Mustafa S, et al. (2011) An ex vivo approach to studying the interactions of probiotic Pediococcus acidilactici and Vibrio (Listonella) anguillarum in the anterior intestine of rainbow trout Oncorhynchus mykiss. J Aquac Res Development S1:004. doi:10.4172/2155-9546.S1-004
Page 2 of 6
Special Issue 1 • 2011J Aquac Res DevelopmentISSN: 2155-9546 JARD, an open access journal
acidilactici. Diets were produced by either top dressing the commercial feed with fish oil containing P. acidilactici (probiotic group) or just fish oil (no added bacteria; control group). P. acidilactici, sourced from the commercial product Bactocell (Lallemand Inc), was cultured in MRS broth at 37oC for 72 hr. Cultures were then centrifuged (2150 x g for 10 min) and washed in phosphate-buffered saline (PBS; pH 7.3, Oxoid) three times prior to re-suspension in fish oil which was sprayed onto the preheated (40oC) basal diet to give a final concentration of log 7 CFU g-1. The diets were offered to the fish for two weeks at 1.2% body weight per day divided into two feeding events (0930 and 1630). Water temperature was maintained at 15 ± 1oC, oxygen concentration at >85% saturation and pH at 7.5 ± 0.5.
Bacterial cultures
P. acidilactici cultures for the ex vivo antagonism assay were prepared as described above and resuspended in PBS at log 9 CFU g-1. V. anguillarum cultures, obtained from Laboratoire nationale de pathologic des animaux aquatique (Brest, France), were cultivated on blood agar (Oxoid) incubated at 25oC for 72 hr. A pure culture was re-streaked onto fresh blood agar plates to induce high virulence; pure cultures were grown in nutrient broth (Oxoid) containing 1.5% NaCl at pH 8.5. Cultures were then centrifuged (2150 x g for 10 min) and washed in PBS three times prior to re-suspension in PBS at log 9 CFU g-1 to produce the final assay inoculums. Finally, a 50/50 mixture of these solutions was used to produce the bacterial suspension for the antagonism assay.
Ex vivo assay
After two weeks feeding on the experimental diets fish were euthanized by an overdose (200 mg L-1 water for 10 min) of MS222 (Pharmaq, Fordingbridge, UK) and destruction of the brain. The anterior intestine of rainbow trout was chosen to use within the assays as evidence indicates it is an important gastric region for V. anguillarum colonisation, infection and damage [8,13] and research shows that P. acidilactici can provide histological benefits to this region in rainbow trout [11]. The anterior intestine of the fish was isolated as described elsewhere [8] prior to rinsing with PBS to remove intestinal contents. The posterior end was tightly tied with cotton thread before filling (ca. 800 μl) with the appropriate bacterial assay solution, tying the anterior end and suspending the sealed intestinal tube in sterile PBS. The intestinal sacs were then incubated at 20oC for 1 hr. Five replicate assays (5 intestinal samples derived from 5 separate fish) were performed for each bacterial exposure. After the 1 hr exposure, 5 mm from each end of the intestinal sac was excised, discarded and samples of the remaining intestinal tissue were retained for molecular microbiological and microscopical analysis. The bacterial solution from within the intestine was retained for plate counts.
Post ex vivo assay microbiological analysis
To determine the level of bacteria (probiotic and pathogen) within the intestinal lumen at the end of the exposure the luminal contents were serially diluted with PBS and plated onto MRS agar and nutrient agar (Oxoid) containing 1.5% NaCl at pH 8.5 to enumerate the levels of P. acidilactici and V. anguillarum, respectively. To confirm identification, a representative number of isolates were sequenced as described elsewhere (Ferguson et al. 2010). Furthermore, DNA extraction, PCR and DGGE analysis was conducted on intestinal tissues (as described elsewhere [12,14]) to investigate potential colonisation and the effect on the autochthonous bacterial community.
Microscopical analysis
Tissue samples for light microscopy (LM) were fixed in 10% formalin for 24h, dehydrated in graded ethanol concentrations and embedded in paraffin wax. For each specimen, multiple sets of sections (5 μm thick) were stained with haematoxylin and eosin (HE) or May-Grünwald/Giemsa (MGG). The number of leucocytes (histologically stained cells) infiltrated in the epithelium across a standardised distance of 100 enterocytes (only nucleated cells) was calculated by averaging the cell numbers from all specimens [12] and additionally, the number of the goblet cells (number of cells per 100 enterocytes) was also quantified.
Tissues for electron microscopy (EM) were fixed in 2.5 % glutaraldehyde in sodium cacodylate buffer (0.1M, ph7.2). Samples for scanning electron microscopy (SEM) were washed with 1% S-carboxymethyl-L-cysteine (Sigma) for 30 sec prior to fixation to remove epithelial mucus. EM samples were processed as described elsewhere [15,16] and were screened with a JSM 1200 EX transmission electron microscope at 120kV (Jeol, Tokyo, Japan) or JSM 5600 LV scanning electron microscope (Jeol). High magnification SEM micrographs were analysed in order to observe any potential bacterial interactions with the host epithelium and also to describe any qualitative effects on the epithelial structure. All digital images for morphological analysis were analyzed with Image J 1.36 (National Institutes of Health, USA).
Statistics
Statistical analyses were conducted using an independent samples t-test with SPSS V16.0 (Chicago IL, USA). Significance was accepted at P< 0.05 and trends towards significance were acknowledged at P <0.10.
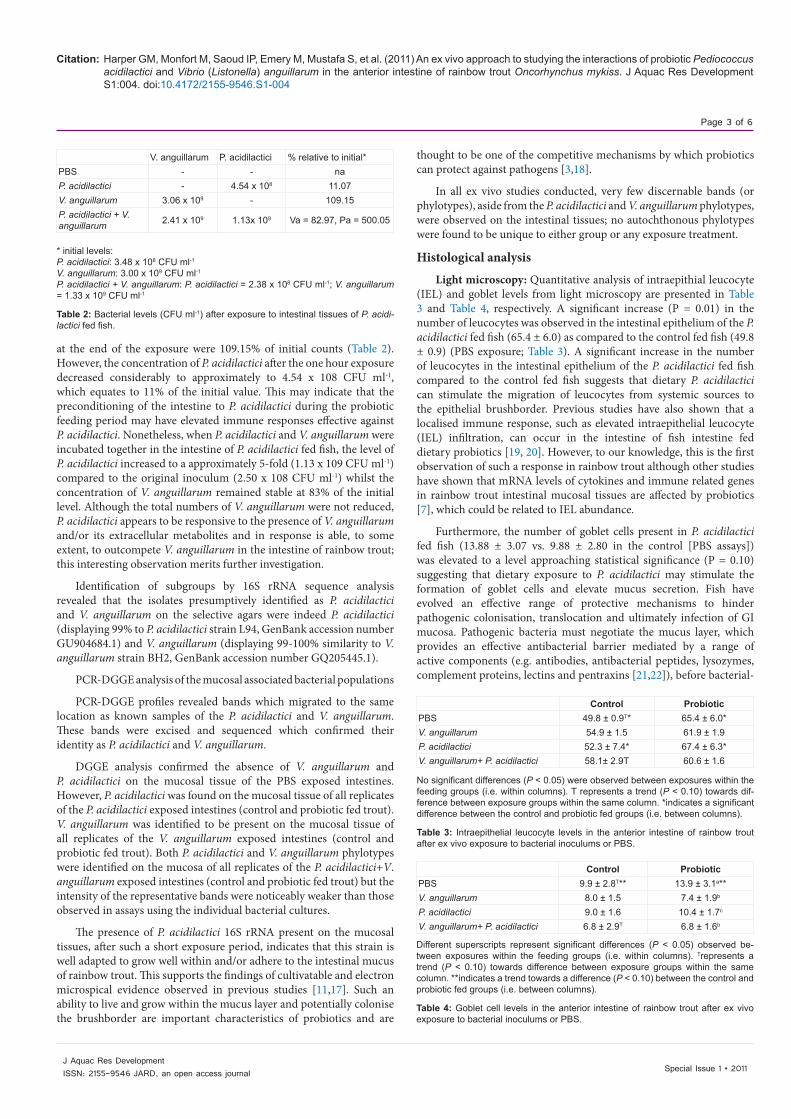
Results and DiscussionBacterial levels
After the 1 hour exposure period in the intestinal lumen of the control fed fish, the level of V. anguillarum decreased down to 1.38 x 109 colony forming units (CFU) ml-1 (from 3.00 x 109 CFU ml-1) which is approximately half of the initial value (46% of the initial level, Table 1). The concentration of P. acidilactici, after the 1 hour exposure to the control intestine, remained stable when incubated alone (remaining at 90% of the initial concentration), however, when P. acidilactici was incubated with V. anguillarum the concentration of P. acidilactici more than doubled to 5.58 x 108 CFU ml-1 (236% of the initial concentration). V. anguillarum levels remained similar to those of the initial inocula (117.92% of the initial level).
In the intestine of the probiotic fed fish, the concentration of V. anguillarum remained stable after the 1 hour exposure period; counts
V. anguillarum P. acidilactici % relative to initial*PBS - - naP. acidilactici - 3.14 x 108 90.26V. anguillarum 1.38 x 109 - 46.11P. acidilactici +V. anguillarum 1.56 x 109 5.58 x 108 Va = 117.92, Pa = 235.93
* initial levels:P. acidilactici: 3.48 x 108 CFU ml-1V. anguillarum: 3.00 x 109 CFU ml-1P. acidilactici +V. anguillarum: P. acidilactici = 2.38 x 108 CFU ml-1; V. anguillarum = 1.33 x 109 CFU ml-1
Table 1: Bacterial levels (CFU ml-1) after exposure to intestinal tissues of control fed fish.
Citation: Harper GM, Monfort M, Saoud IP, Emery M, Mustafa S, et al. (2011) An ex vivo approach to studying the interactions of probiotic Pediococcus acidilactici and Vibrio (Listonella) anguillarum in the anterior intestine of rainbow trout Oncorhynchus mykiss. J Aquac Res Development S1:004. doi:10.4172/2155-9546.S1-004
Page 3 of 6
Special Issue 1 • 2011J Aquac Res DevelopmentISSN: 2155-9546 JARD, an open access journal
at the end of the exposure were 109.15% of initial counts (Table 2). However, the concentration of P. acidilactici after the one hour exposure decreased considerably to approximately to 4.54 x 108 CFU ml-1, which equates to 11% of the initial value. This may indicate that the preconditioning of the intestine to P. acidilactici during the probiotic feeding period may have elevated immune responses effective against P. acidilactici. Nonetheless, when P. acidilactici and V. anguillarum were incubated together in the intestine of P. acidilactici fed fish, the level of P. acidilactici increased to a approximately 5-fold (1.13 x 109 CFU ml-1) compared to the original inoculum (2.50 x 108 CFU ml-1) whilst the concentration of V. anguillarum remained stable at 83% of the initial level. Although the total numbers of V. anguillarum were not reduced, P. acidilactici appears to be responsive to the presence of V. anguillarum and/or its extracellular metabolites and in response is able, to some extent, to outcompete V. anguillarum in the intestine of rainbow trout; this interesting observation merits further investigation.
Identification of subgroups by 16S rRNA sequence analysis revealed that the isolates presumptively identified as P. acidilactici and V. anguillarum on the selective agars were indeed P. acidilactici (displaying 99% to P. acidilactici strain L94, GenBank accession number GU904684.1) and V. anguillarum (displaying 99-100% similarity to V. anguillarum strain BH2, GenBank accession number GQ205445.1).
PCR-DGGE analysis of the mucosal associated bacterial populations
PCR-DGGE profiles revealed bands which migrated to the same location as known samples of the P. acidilactici and V. anguillarum. These bands were excised and sequenced which confirmed their identity as P. acidilactici and V. anguillarum.
DGGE analysis confirmed the absence of V. anguillarum and P. acidilactici on the mucosal tissue of the PBS exposed intestines. However, P. acidilactici was found on the mucosal tissue of all replicates of the P. acidilactici exposed intestines (control and probiotic fed trout). V. anguillarum was identified to be present on the mucosal tissue of all replicates of the V. anguillarum exposed intestines (control and probiotic fed trout). Both P. acidilactici and V. anguillarum phylotypes were identified on the mucosa of all replicates of the P. acidilactici+V. anguillarum exposed intestines (control and probiotic fed trout) but the intensity of the representative bands were noticeably weaker than those observed in assays using the individual bacterial cultures.
The presence of P. acidilactici 16S rRNA present on the mucosal tissues, after such a short exposure period, indicates that this strain is well adapted to grow well within and/or adhere to the intestinal mucus of rainbow trout. This supports the findings of cultivatable and electron microspical evidence observed in previous studies [11,17]. Such an ability to live and grow within the mucus layer and potentially colonise the brushborder are important characteristics of probiotics and are
thought to be one of the competitive mechanisms by which probiotics can protect against pathogens [3,18].
In all ex vivo studies conducted, very few discernable bands (or phylotypes), aside from the P. acidilactici and V. anguillarum phylotypes, were observed on the intestinal tissues; no autochthonous phylotypes were found to be unique to either group or any exposure treatment.
Histological analysis
Light microscopy: Quantitative analysis of intraepithial leucocyte (IEL) and goblet levels from light microscopy are presented in Table 3 and Table 4, respectively. A significant increase (P = 0.01) in the number of leucocytes was observed in the intestinal epithelium of the P. acidilactici fed fish (65.4 ± 6.0) as compared to the control fed fish (49.8 ± 0.9) (PBS exposure; Table 3). A significant increase in the number of leucocytes in the intestinal epithelium of the P. acidilactici fed fish compared to the control fed fish suggests that dietary P. acidilactici can stimulate the migration of leucocytes from systemic sources to the epithelial brushborder. Previous studies have also shown that a localised immune response, such as elevated intraepithelial leucocyte (IEL) infiltration, can occur in the intestine of fish intestine fed dietary probiotics [19, 20]. However, to our knowledge, this is the first observation of such a response in rainbow trout although other studies have shown that mRNA levels of cytokines and immune related genes in rainbow trout intestinal mucosal tissues are affected by probiotics [7], which could be related to IEL abundance.
Furthermore, the number of goblet cells present in P. acidilactici fed fish (13.88 ± 3.07 vs. 9.88 ± 2.80 in the control [PBS assays]) was elevated to a level approaching statistical significance (P = 0.10) suggesting that dietary exposure to P. acidilactici may stimulate the formation of goblet cells and elevate mucus secretion. Fish have evolved an effective range of protective mechanisms to hinder pathogenic colonisation, translocation and ultimately infection of GI mucosa. Pathogenic bacteria must negotiate the mucus layer, which provides an effective antibacterial barrier mediated by a range of active components (e.g. antibodies, antibacterial peptides, lysozymes, complement proteins, lectins and pentraxins [21,22]), before bacterial-
V. anguillarum P. acidilactici % relative to initial*PBS - - naP. acidilactici - 4.54 x 108 11.07V. anguillarum 3.06 x 109 - 109.15P. acidilactici + V. anguillarum 2.41 x 109 1.13x 109 Va = 82.97, Pa = 500.05
* initial levels:P. acidilactici: 3.48 x 108 CFU ml-1V. anguillarum: 3.00 x 109 CFU ml-1P. acidilactici + V. anguillarum: P. acidilactici = 2.38 x 108 CFU ml-1; V. anguillarum = 1.33 x 109 CFU ml-1
Table 2: Bacterial levels (CFU ml-1) after exposure to intestinal tissues of P. acidi-lactici fed fish.
Control ProbioticPBS 49.8 ± 0.9T* 65.4 ± 6.0*V. anguillarum 54.9 ± 1.5 61.9 ± 1.9P. acidilactici 52.3 ± 7.4* 67.4 ± 6.3*V. anguillarum+ P. acidilactici 58.1± 2.9T 60.6 ± 1.6
No significant differences (P < 0.05) were observed between exposures within the feeding groups (i.e. within columns). T represents a trend (P < 0.10) towards dif-ference between exposure groups within the same column. *indicates a significant difference between the control and probiotic fed groups (i.e. between columns).
Table 3: Intraepithelial leucocyte levels in the anterior intestine of rainbow trout after ex vivo exposure to bacterial inoculums or PBS.
Control ProbioticPBS 9.9 ± 2.8T** 13.9 ± 3.1a**V. anguillarum 8.0 ± 1.5 7.4 ± 1.9b
P. acidilactici 9.0 ± 1.6 10.4 ± 1.7c
V. anguillarum+ P. acidilactici 6.8 ± 2.9T 6.8 ± 1.6b
Different superscripts represent significant differences (P < 0.05) observed be-tween exposures within the feeding groups (i.e. within columns). Trepresents a trend (P < 0.10) towards difference between exposure groups within the same column. **indicates a trend towards a difference (P < 0.10) between the control and probiotic fed groups (i.e. between columns).
Table 4: Goblet cell levels in the anterior intestine of rainbow trout after ex vivo exposure to bacterial inoculums or PBS.

Citation: Harper GM, Monfort M, Saoud IP, Emery M, Mustafa S, et al. (2011) An ex vivo approach to studying the interactions of probiotic Pediococcus acidilactici and Vibrio (Listonella) anguillarum in the anterior intestine of rainbow trout Oncorhynchus mykiss. J Aquac Res Development S1:004. doi:10.4172/2155-9546.S1-004
Page 4 of 6
Special Issue 1 • 2011J Aquac Res DevelopmentISSN: 2155-9546 JARD, an open access journal
host cell interactions can occur. Thus, elevated mucus secretion, particularily in combination with elevated IELs, is likely to elevate host defences to enteric infections. Indeed, the lower number of goblet cells observed in the intestines of probiotic fed fish intestines exposed to V. anguilliarum and V. anguillarum+P. acidilactici and control fed fish intestines exposed to V. anguillarum+P. acidilactici is suggestive of a rapid secretion of mucus (thus reducing the numbers of goblet cells in the brushborder) in response to the presence of the mucosal associated pathogen V. anguillarum.
Transmission electron microscopy: TEM revealed the presence of presumed V. anguillarum (morphologically corresponding to known V. anguillarum cells in terms of shape, size and electron density), in high numbers present in close association with the brushborder in V. anguillarum exposed tissues (Figure 1a), which indicates a high affinity of the pathogen to the intestine of rainbow trout. Although P. acidilactici-like cells were also observed in close association with the brushborder, they were noticeably fewer than V. anguillarum in number (Figure 1b). No signs of histological damage or translocation was observed in the intestines of fish exposed to P. acidilactici; this finding is of high importance as lack of translocation and cell damage have been previously proposed as key selection criteria when evaluating the use of probiotics in fish [2]. In contrast, TEM analysis showed some typical signs of histological damage caused by V. anguillarum; disorganised microvilli, intracellular gaps and necrotic enterocytes were observed in close proximity to large V. anguillarum populations (Figure 2). Similar results have been reported in Atlantic salmon intestines exposed to V. anguillarum [8]. Previous studies have reported observations of pathogen translocation through the enterocytes, or paracellular translocation between enterocytes [2,23] yet, no signs of translocation of V. anguillarum were observed in the present study. This indicates that the process of uptake, or invasion, may require more time than the one hour tested in the present study.
Scanning electron microscopy: SEM revealed complex mucosal foldings at low magnification. At greater magnification it was possible to observe enterocytes present on the surface of the epithelium and also on the apical brush border microvilli. In the intestines of the control fed fish exposed to PBS, very few signs of damage were observed and the enterocyte and microvilli formations were generally intact appearing healthy (Figure 3A). However, exposure to V. anguillarum resulted in
observable tissue damage. The presence of necrotic enterocytes, irregular shaped enterocytes and missing/damaged microvilli formations were observed (Figure 3B). No signs of pathogenic damage were observed in the tissue samples exposed to P. acidilacticiwhich appeared similar to the PBS controls (Figure 3C). Intestinal tissue exposed to both V. anguillarum+P. acidilactici showed a range of damage, similar to that of the V. anguillarum group, where irregular and nectrotic cells were observed.
In the trout group previously fed the probiotic diet, the PBS exposure again did not result in any clear signs of damage. Some signs of damage were observed in the intestinal tissue of the probiotic fed fish when exposed to V. anguillarum. However, from a qualitative point of view, this appeared less than that observed in the control fed trout exposed to V. anguillarum. No tissue damage was observed following exposure to P. acidilactici. Furthermore, relatively little damage was observed in the intestinal tissue of probiotic fed fish exposed to both V. anguillarum+P. acidilactici (Figure 3D). This suggests that a good method to reduce V. anguillarum damage in the GI tract of rainbow trout is likely to be feeding on P. acidilactici supplemented diets prior to, and during, V. anguillarum infection. Further in vivo studies are required to test this hypothesis.
Conclusion
Ex vivo studies, as demonstrated by the present study, reveal important probiotic-pathogen-host interactions which yield important information in respect to the understanding the complex multifactoral mechanisms which underpin and mediate probiotic benefits against disease resistance. In vitro and ex vivo studies are essential in reducing the number of animals used in scientific studies and thus a preliminary ex vivo approach to investigate the efficacy and potential of probiotics in respect to disease resistance should be utilised prior to large scale in vivo disease challenge studies. Little information is available regarding the effect of P. acidilactici against rainbow trout bacterial infections and disease resistance. The present study shows that P. acidilactici can outcompete V. anguillarum in the intestine of rainbow trout. Furthermore, the recovery of P. acidilactici from the mucosa, elevated levels of leucocyte and goblet cells in the epithelium of P. acidilactici fed fish and qualitative observations indicating a possible reduction of epithelial tissue damage, suggests potential for V. anguillarum infections
Figure 1: TEM demonstrating localised bacterial populations in close proximity and association with the epithelial brush border in the anterior intestine of rainbow trout. Scale bars 2 µm (except B inset: 5µm). TEM showing V. anguillarum-like cells (A; probiotic fed fish exposed to V. anguillarum) and P. acidilactici-like cells (B; probiotic fed fish exposed to P. acidilactici) in the lumen and in close association with microvilli. Inset figures show respective cell morphologies of known samples of the bacterial strains used within the assays.

Citation: Harper GM, Monfort M, Saoud IP, Emery M, Mustafa S, et al. (2011) An ex vivo approach to studying the interactions of probiotic Pediococcus acidilactici and Vibrio (Listonella) anguillarum in the anterior intestine of rainbow trout Oncorhynchus mykiss. J Aquac Res Development S1:004. doi:10.4172/2155-9546.S1-004
Page 5 of 6
Special Issue 1 • 2011J Aquac Res DevelopmentISSN: 2155-9546 JARD, an open access journal
to be controlled in rainbow trout with the dietary application of P. acidilactici. In vivo disease challenge studies are warranted to validate this hypothesis.
Acknowledgements
This study was funded by a grant awarded to Dr Merrifield under the University of Plymouth’s International Research Networking and Collaboration Scheme, 2010. The authors wish to thank the technical staff at the microbiology research laboratory at the University of Plymouth for their support and expertise.
References
1. Groff J, LaPatra S (2000) Infectious diseases impacting the commercial culture of salmonids. J Appl Aquacult 10: 17-90.
2. Ringø E, Myklebust R, Mayhew TM, Olsen RE (2007). Bacterial translocation and pathogenesis in the digestive tract of larvae and fry. Aquacult 268: 251-264.
3. Merrifield DL, Dimitroglou A, Foey A, Davies S J, Baker RTM, et al. (2010) The current status and future focus of probiotic and prebiotic applications for salmonids. Aquacult 302: 1-18.
4. Dimitroglou A, Merrifield DL, Carnevali O, Picchietti S, Avella M, et al. (2011) Microbial manipulations to improve fish health and production - a Mediterranean perspective. Fish Shellfish Immunol 30: 1-16.
5. Balcázar JL, de Blas I, Ruiz-Zazuela I, Vandrell D, Gironés O, et al. (2007) Enhancement of the immune response and protection induced by probiotic lactic acid bacteria against furunculosis in rainbow trout (Oncorhynchus mykiss) FEMS. Immunol Med Microbiol 51: 185-193.
6. Brunt J, Newaj-Fyzul A, Austin B (2007) The development of probiotics for the control of multiple bacterial diseases of rainbow trout, Oncorhynchus mykiss (Walbaum). J Fish Dis 30: 573-579.
7. Pérez-Sánchez T, Balcázar JL, Merrifield DL, Carnevali O, Gioacchini G, et al. (2011) Expression of immune-related genes in rainbow trout (Oncorhynchus mykiss) induced by probiotic bacteria during Lactococcus garvieae infection. Fish Shellfish Immunol 31: 196-201.
8. Ringø E, Salinas I, Olsen RE, Nyhaug A, Myklebust R, et al. (2007a) Histological changes in Atlantic salmon (Salmo salar L.) intestine following in vitro exposure to pathogenic and probiotic bacterial strains. Cell Tis Res 328: 109-116.
9. Salinas I, Myklebust R, Esteban MA, Olsen RE, Meseguer J, et al. (2008) In vitro studies of Lactobacillus delbrueckii subsp. lactis in Atlantic salmon (Salmo salar L.) foregut: tissue responses and evidence of protection against Aeromonas salmonicida subsp. salmonicida epithelial damage. Vet Microbiol 128: 167-177.
10. Aubin J, Gatesoupe F-J, Labbe L, Lebrun L (2005) Trial of probiotics to prevent the vertebral column compression syndrome in rainbow trout (Oncorhynchus mykiss Walbaum). Aquacult Res 36: 758-767.
11. Merrifield DL, Harper GM, Dimitroglou A, Ringø E, Davies SJ (2010) Possible influence of probiotic adhesion to intestinal mucosa on the activity and morphology of rainbow trout (Oncorhynchus mykiss) enterocytes. Aquacult Res 41: 1268-1272.
12. Ferguson RMW, Merrifield DL, Harper GM, Rawling MD, Mustafa S, et al. (2010) The effect of Pediococcus acidilactici on the gut microbiota and immune status of on-growing red tilapia (Oreochromis niloticus). J Appl Microbiol 109: 851-862.
13. Grisez L, Chair M, Sorgeloos P, Ollevier F (1996) Mode of infection and spread of Vibrio anguillarum in turbot Scopthalamus maximus larvae after oral challenge through live feed. Dis Aquatic Org 26: 181-187.
14. Merrifield DL, Burnard D, Bradley G, Davies SJ, Baker RTM (2009) Microbial community diversity associated with the intestinal mucosa of farmed rainbow trout (Oncorhynchus mykiss Walbaum). Aquacult Res 40: 1064-1072.
15. Merrifield DL, Bradley G, Baker RTM, Dimitroglou A, Davies SJ (2009) Soybean meal alters autochthonous microbial populations, microvilli morphology and compromises intestinal enterocyte integrity of rainbow trout (Oncorhynchus mykiss Walbaum). J Fish Dis 32: 755-766.
16. Dimitroglou A, Merrifield DL, Moate R, Davies SJ, Spring P, et al. (2009) Dietary mannan oligosaccharide supplementation modulates intestinal microbial ecology and improves gut morphology of rainbow trout, Oncorhynchus mykiss (Walbaum). J Animal Sci 87: 3226-3234.
17. Merrifield DL, Bradley G, Harper G, Baker RTM, Munn CB, et al. (2011) Assessment of the effects of vegetative and lyophilised Pediococcus acidilactici on growth, feed utilisation, intestinal colonisation and health parameters of rainbow trout (Oncorhynchus mykiss Walbaum). Aqua Nutr 17: 73-79
18. Balcázar JL, de Blas I, Ruiz-Zazuela I, Cunningham D, Vandrell D, et al. (2006) The role of probiotics in aquaculture. Vet Microbiol 114: 173-186.
19. Picchietti S, Mazzini M, Taddei AR, Renna R, Fausto AM, et al. (2007) Effects of administration of probiotic strains on GALT of larval gilthead seabream: immunohistochemical and ultrastructural studies. Fish Shellfish Immunol 22: 57-67.
20. Picchietti S, Fausto AM, Randelli E, Carnevali O, Taddei AR, et al. (2009) Early treatment with Lactobacillus delbrueckii strain induces rise in intestinal T cells and granulocytes and modulates immune related genes of larval Dicentrarchus labrax (L.). Fish Shellfish Immunol 26: 368-376.
21. Dalmo DA, Ingebrigtsen K, Bøgwald J (1997) Non-specific defence mechanisms in fish, with particular reference to the reticuloendothelial system (RES) J Fish Dis 20: 241-273.
Figure 2: Transmission electron micrograph of the anterior intestine (mid gut) of rainbow trout exposed to Vibrio anguillarum (VA). Clear signs of tis-sue damage are characterised by necrotic enterocytes (NE), disorganised microvilli and irregular tight junctional complexes (TJ) resulting in intracel-lular spaces (*). E = enterocyte. Scale bar = 5 µm.
Figure 3: SEM images demonstrating the morphological changes in the an-terior intestine of rainbow trout exposed to the different bacterial inoculums in the ex vivo challenge. A: control fed fish intestine exposed to PBS, B: Control fed fish intestine exposed to V. anguillarum; C: Control fed fish intestine ex-posed P. acidilactici; D: P. acidilactici fed fish exposed to V. anguillarum+P. acidilactici. Key: * irregular microvilli, IR irregular enterocyte formations, ar-rows = spaces/gaps forming between enterocytes (arrows) exposing intra-cellular tight junctions. Scale bars 10µm.

Citation: Harper GM, Monfort M, Saoud IP, Emery M, Mustafa S, et al. (2011) An ex vivo approach to studying the interactions of probiotic Pediococcus acidilactici and Vibrio (Listonella) anguillarum in the anterior intestine of rainbow trout Oncorhynchus mykiss. J Aquac Res Development S1:004. doi:10.4172/2155-9546.S1-004
Page 6 of 6
Special Issue 1 • 2011J Aquac Res DevelopmentISSN: 2155-9546 JARD, an open access journal
22. Saurabh S, Sahoo PK (2008) Lysozyme: an important defence molecule of fish innate immune system. Aquacult Res 39: 223-239.
23. Ringø E, Olsen RE, Mayhew TM, Myklebust R (2003) Electron microscopy of the intestinal microflora of fish. Aquaculture 227: 395-415.
Submit your next manuscript and get advantages of OMICS Group submissionsUnique features:
• Userfriendly/feasiblewebsite-translationofyourpaperto50world’sleadinglanguages• AudioVersionofpublishedpaper• Digitalarticlestoshareandexplore
Special features:
• 100OpenAccessJournals• 10,000editorialteam• 21daysrapidreviewprocess• Qualityandquickeditorial,reviewandpublicationprocessing• IndexingatPubMed(partial),Scopus,DOAJ,EBSCO,IndexCopernicusandGoogleScholaretc• SharingOption:SocialNetworkingEnabled• Authors,ReviewersandEditorsrewardedwithonlineScientificCredits• Betterdiscountforyoursubsequentarticles
Submityourmanuscriptat:http://www.omicsonline.org/submission


















