
Isolation of the Nuclear Gene Encoding a Subunit of the Yeast
4
THE JOURNAL OF BtOLOClCAL CHEMISTRY 0 1986 by The American Society of Biological Chemists, Inc Val. 261, No. 22, Issue of August 5, pp. 10348-10351, 1986 Printed in U.S.A. Isolation of the Nuclear Gene Encoding a Subunit of the Yeast Mitochondrial RNA Polymerase* (Received for publication, December 18, 1985) Jeffrey L. Kelly$, Arno L. Greenleafj, and I. R. Lehman From the Departmentof Biochemistry, Stanford University School of Medicine, Stanford, California 94305 Antisera directed against the purified 145,000-dal- ton subunit of the Saccharomyces cerevisiae mitochon- drial RNA polymerase have been used to immuno- screen a library of yeast genomic inserts constructed in the fusion protein expression vector, Xgtll. A 4- kilobase pair yeast DNA fragment inserted into one of the recombinant bacteriophages appears to contain most or all of the gene that encodes the 145,000-dalton subunit. In most instances, the role of a eukaryotic enzyme in vivo must be inferred from its activity in uitro. However, in the lower eukaryote, Saccharomyces cereuisiae, it is experimen- tally straightforward to obtain directgenetic support for the function of an enzyme. Thus, antisera directed against the purified enzyme can be used to clone the genes encoding the polypeptides recognized by the antisera, thereby allowing construction of haploid mutants that have been disrupted in the gene(s) of interest (1-4). The phenotype of such mutants can then be examined directly. We have been investigating a yeast mitochondrial RNA polymerase activity (5). The near homogeneous enzyme con- sists of a single 145-kDa polypeptide. We have prepared antisera directed against the enzyme that recognize the 145- kDa polypeptide and inhibit the RNA polymerase activity. This paper describes our use of these antisera to clone the gene encoding the 145-kDa polypeptide from a yeast genomic library constructed in the expression vector Xgtll (1, 6). EXPERIMENTAL PROCEDURES‘ RESULTS AND DISCUSSION Immunoscreening of the Xgtll Library-A rabbit antiserum directed against the 145-kDa RNA polymerase activity (5) was used to probe a Xgtll library consisting of 2 X lo6 * This work was supported by grants from the National Institutes of Health (GM06196to I. R. L.) and the American Heart Association (Established Investigatorship 82-195 to A. L. G.). The costs of pub- lication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “aduertise- rnent” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. $ Present address: Dept. of Plant Pathology, University of Califor- nia at Davis, Davis, CA 95616. 5 Present address: Biochemistry Dept., Duke University Medical Center, Durham, NC 27710. The “Experimental Procedures” are presented in miniprint at the end of this paper. Miniprint is easily read with the aid of a standard magnifying glass. Full size photocopies are available from the Journal of Biological Chemistry, 9650 Rockville Pike, Bethesda, MD 20814. Request Document No. 85M-4108, cite the authors, and include a check or money order for $1.00 per set of photocopies. Full size photocopies are also included in the microfilm edition of the Journal that is available from Waverly Press. 0 3 6 kb phageL I I I I 1 1 102,601,1202 1 1 1602.1702 I 1 2202 401 1 1 f :I inserts cross- hybridize 201 1 v 1802 F inserts 1901 d_o not 1 v cross- - I hybridize 2101 with 102 FIG. 1. Relationship of yeast inserts in clones selected by immunoscreening. Phage DNA was prepared from hgtll recombi- nant clones selected by immunoscreening and analyzed by EcoRI digestion and gel blot/hybridization with isolated insert fragments as described under “Experimental Procedures.” The yeast inserts are not all present in the same orientation inrelation to the P-galactosid- ase sequence of the hgtll vector. recombinant phage containing yeast genomic DNA inserts. After screening lo6 recombinants, 14 independent clones were isolated, each of which produced strong signals. Twelve of these were selected for further analysis. DNA was prepared from each and analyzed by restriction enzyme mapping and Southern hybridization (8). Of the 12 clones, 5 recombinants appeared to have identical inserts, and these shared DNA sequences with two others (Fig. 1). Four clones appeared to have unique inserts, and the last produced no EcoRI frag- ments of 510 kilobase pairs. Phage 102 was chosen as a representative of theabundantclass of clones producing strong signals. Phage 102-induced Proteins Are Antigenically Related to the 145-kDa Polypeptide-As a rapid means of identifying which of the clones isolated in the Xgtll library screen carried inserts encoding antigenic determinants related to thepurified 145- kDa RNApolymerase, representative phage clones were used to affinity-purify antibodies from the polyclonal rabbit serum, and the purified antibodies were used to probe immunoblots of the purified 145-kDa protein. When phage 102 was used to affinity-purify antibodies, the antibodies recognized a poly- peptide migratingat 145 kDa in both highly purified and less purified fractions of the RNApolymerase. Analysis of the Cloned RNA Polymerase Gene-To investi- gate further the yeast DNA-encoded protein whose synthesis was induced in Escherichia coli by phage102, strains lysogenic for several of the phages were constructed. Extracts of the induced lysogenic strains were then prepared and analyzed by electrophoresisin NaD~dSO,~-polyacrylarnide slab gels fol- lowed by immunoblotting. Antiserum against the purified 145- The abbreviations used are: NaDodS04, sodium dodecyl sulfate; kb, kilobase pair; IPTG, isopropyl-0-o-thiogalactoside. 10348
Transcript of Isolation of the Nuclear Gene Encoding a Subunit of the Yeast

THE JOURNAL OF BtOLOClCAL CHEMISTRY 0 1986 by The American Society of Biological Chemists, Inc Val. 261, No. 22, Issue of August 5, pp. 10348-10351, 1986
Printed in U.S.A.
Isolation of the Nuclear Gene Encoding a Subunit of the Yeast Mitochondrial RNA Polymerase*
(Received for publication, December 18, 1985)
Jeffrey L. Kelly$, Arno L. Greenleafj, and I. R. Lehman From the Department of Biochemistry, Stanford University School of Medicine, Stanford, California 94305
Antisera directed against the purified 145,000-dal- ton subunit of the Saccharomyces cerevisiae mitochon- drial RNA polymerase have been used to immuno- screen a library of yeast genomic inserts constructed in the fusion protein expression vector, Xgtll. A 4- kilobase pair yeast DNA fragment inserted into one of the recombinant bacteriophages appears to contain most or all of the gene that encodes the 145,000-dalton subunit.
In most instances, the role of a eukaryotic enzyme in vivo must be inferred from its activity in uitro. However, in the lower eukaryote, Saccharomyces cereuisiae, it is experimen- tally straightforward to obtain direct genetic support for the function of an enzyme. Thus, antisera directed against the purified enzyme can be used to clone the genes encoding the polypeptides recognized by the antisera, thereby allowing construction of haploid mutants that have been disrupted in the gene(s) of interest (1-4). The phenotype of such mutants can then be examined directly.
We have been investigating a yeast mitochondrial RNA polymerase activity (5). The near homogeneous enzyme con- sists of a single 145-kDa polypeptide. We have prepared antisera directed against the enzyme that recognize the 145- kDa polypeptide and inhibit the RNA polymerase activity. This paper describes our use of these antisera to clone the gene encoding the 145-kDa polypeptide from a yeast genomic library constructed in the expression vector Xgtll (1, 6).
EXPERIMENTAL PROCEDURES‘
RESULTS AND DISCUSSION
Immunoscreening of the Xgtll Library-A rabbit antiserum directed against the 145-kDa RNA polymerase activity (5) was used to probe a Xgtll library consisting of 2 X lo6
* This work was supported by grants from the National Institutes of Health (GM06196 to I. R. L.) and the American Heart Association (Established Investigatorship 82-195 to A. L. G.). The costs of pub- lication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “aduertise- rnent” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
$ Present address: Dept. of Plant Pathology, University of Califor- nia at Davis, Davis, CA 95616.
5 Present address: Biochemistry Dept., Duke University Medical Center, Durham, NC 27710.
The “Experimental Procedures” are presented in miniprint at the end of this paper. Miniprint is easily read with the aid of a standard magnifying glass. Full size photocopies are available from the Journal of Biological Chemistry, 9650 Rockville Pike, Bethesda, MD 20814. Request Document No. 85M-4108, cite the authors, and include a check or money order for $1.00 per set of photocopies. Full size photocopies are also included in the microfilm edition of the Journal that is available from Waverly Press.
0 3 6 kb p h a g e L
I I I I 1 1
102,601,1202 1 1 1602.1702
I 1 2202
401 1 1 f :I inserts cross- hybridize
201 1 v
1802 F inserts
1901 d_o not
1 v cross- - I hybridize 2101 with 102
FIG. 1. Relationship of yeast inserts in clones selected by immunoscreening. Phage DNA was prepared from hgtll recombi- nant clones selected by immunoscreening and analyzed by EcoRI digestion and gel blot/hybridization with isolated insert fragments as described under “Experimental Procedures.” The yeast inserts are not all present in the same orientation in relation to the P-galactosid- ase sequence of the h g t l l vector.
recombinant phage containing yeast genomic DNA inserts. After screening lo6 recombinants, 14 independent clones were isolated, each of which produced strong signals. Twelve of these were selected for further analysis. DNA was prepared from each and analyzed by restriction enzyme mapping and Southern hybridization (8). Of the 12 clones, 5 recombinants appeared to have identical inserts, and these shared DNA sequences with two others (Fig. 1). Four clones appeared to have unique inserts, and the last produced no EcoRI frag- ments of 510 kilobase pairs. Phage 102 was chosen as a representative of the abundant class of clones producing strong signals.
Phage 102-induced Proteins Are Antigenically Related to the 145-kDa Polypeptide-As a rapid means of identifying which of the clones isolated in the Xgtll library screen carried inserts encoding antigenic determinants related to the purified 145- kDa RNA polymerase, representative phage clones were used to affinity-purify antibodies from the polyclonal rabbit serum, and the purified antibodies were used to probe immunoblots of the purified 145-kDa protein. When phage 102 was used to affinity-purify antibodies, the antibodies recognized a poly- peptide migrating a t 145 kDa in both highly purified and less purified fractions of the RNA polymerase.
Analysis of the Cloned RNA Polymerase Gene-To investi- gate further the yeast DNA-encoded protein whose synthesis was induced in Escherichia coli by phage 102, strains lysogenic for several of the phages were constructed. Extracts of the induced lysogenic strains were then prepared and analyzed by electrophoresis in NaD~dSO,~-polyacrylarnide slab gels fol- lowed by immunoblotting. Antiserum against the purified 145-
The abbreviations used are: NaDodS04, sodium dodecyl sulfate; kb, kilobase pair; IPTG, isopropyl-0-o-thiogalactoside.
10348
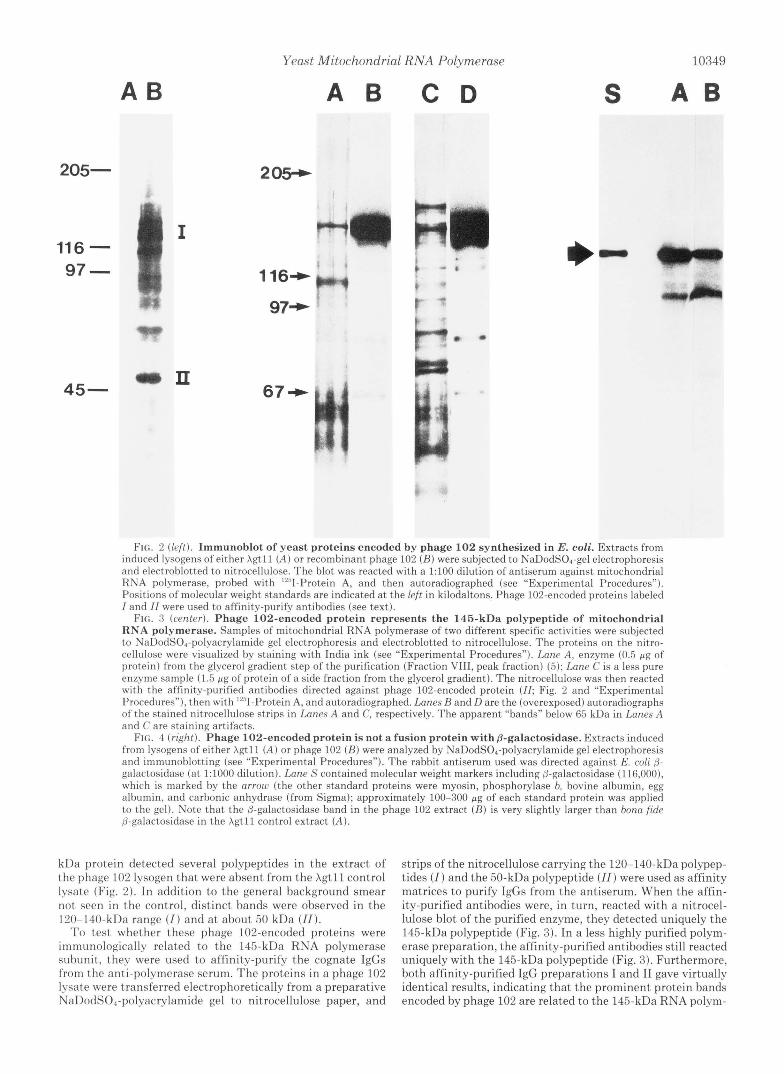
Yeast Mitochondrial RNA Polymerase 10349
A B
205-
I 116 - 97 -
A B C D S A B
205-
97+
-11 45- 67 - i
FIG. 2 (left). Immunoblot of yeast proteins encoded by phage 102 synthesized in E. coli. Extracts from induced lysogens of either Xgt11 ( A ) or recombinant phage 102 ( R ) were subjected to NaDodSOI-gel electrophoresis and electroblotted to nitrocellulose. The blot was reacted with a 1:100 dilution of antiserum against mitochondrial RNA polymerase, probed with ‘“I-Protein A, and then autoradiographed (see “Experimental Procedures”). Positions of molecular weight standards are indicated at the left in kilodaltons. Phage 102-encoded proteins labeled I and 11 were used to affinity-purify antibodies (see text).
FIG. 3 (center). Phage 102-encoded protein represents the 145-kDa polypeptide of mitochondrial RNA polymerase. Samples of mitochondrial RNA polymerase of two different specific activities were subjected to NaDodS04-polyacrylamide gel electrophoresis and electroblotted to nitrocellulose. The proteins on the nitro- cellulose were visualized by staining with India ink (see “Experimental Procedures”). Lane A, enzyme (0.5 pg of protein) from the glycerol gradient step of the purification (Fraction VIII, peak fraction) (5); Lane C is a less pure enzyme sample (1.5 pg of protein of a side fraction from the glycerol gradient). The nitrocellulose was then reacted with the affinity-purified antibodies directed against phage 102-encoded protein ( I I ; Fig. 2 and “Experimental Procedures”). then with ’2sII-Protein A, and autoradiographed. Lanes R and D are the (overexposed) autoradiographs of the stained nitrocellulose strips in Lanes A and C, respectively. The apparent “bands” below 65 kDa in Lanes A and C are staining artifacts.
FIG. 4 (right). Phage 102-encoded protein is not a fusion protein with &galactosidase. Extracts induced from lysogens of either Xgtll ( A ) or phage 102 ( B ) were analyzed by NaDodS04-polyacrylamide gel electrophoresis and immunoblotting (see “Experimental Procedures”). The rabbit antiserum used was directed against E. coli @- galactosidase (at 1:1000 dilution). Lane S cont.ained molecular weight markers including @-galactosidase (116,000), which is marked by the arrow (the other standard proteins were myosin, phosphorylase b. bovine albumin, egg albumin, and carbonic anhydrase (from Sigma); approximately 100-300 pcg of each standard protein was applied to the gel). Note that the @-galactosidase band in the phage 102 extract ( R ) is very slightly larger t.han bona fide B-galactosidase in the Xgtll control extract ( A ) .
kDa protein detected several pol-ypeptides in the extract of the phage 102 lysogen that were absent from the Xgtll control lysate (Fig. 2). In addition to t,he general background smear not seen in the control, distinct bands were observed in the 120-140-kDa range (Z) and at about 50 kDa (ZI).
To test whether these phage 102-encoded proteins were immunologically related to the 145-kDa RNA polymerase subunit, they were used to affinity-purifv the cognate IgGs from the anti-polvmerase serum. The proteins in a phage 102 lysate were transferred electrophoretically from a preparative NaDodS0,-polyacrylamide gel to nitrocellulose paper, and
strips of the nitrocellulose carrying the 120-140-kDa polypep- tides ( I ) and the 50-kDa polypeptide (ZZ) were used as affinity matrices to purify IgGs from the antiserum. When the affin- ity-purified antibodies were, in turn, reacted with a nitrocel- lulose blot of the purified enzyme, they detected uniquely the 145-kDa polypeptide (Fig. 3). In a less highly purified polym- erase preparation, the affinity-purified antibodies still reacted uniquely with the 145-kDa polypeptide (Fig. 3). Furthermore, both affinity-purified IgG preparations I and I1 gave virtually identical results, indicating that the prominent protein bands encoded by phage 102 are relat,ed to the 145-kDa RNA polym-

10350 Yeast Mitochondrial RNA Polymerase
I I I I I I
28 NO SERA
24
20
16
12
E
4
0 0 5 10 15 20 25 30
TIME ( m i d
FIG. 5. Inhibition of RNA polymerase by affinity-purified antibodies. Fraction VI11 (0.5 ng) (5) was incubated with either 2.5 fig of preimmune or 1.25 pg of affinity-purified IgG for 60 min at 15 "C and assayed for RNA polymerase activity (5).
R R R R R R R R B S B B B
1 I Genome I I I x x 1 1 X ' ' A 1 ' (303)
H H H H H H H H
1 kb H
FIG. 6. Restriction map of the genomic region containing the phage 102 yeast DNA insert. The open box represents the approximately 4-kb yeast DNA fragment inserted in phage 102 at the Xgtll EcoRI site. Restriction endonuclease sites are indicated R, EcoRI; H, HindIII; B, BamH1; S, SalI; X , XhoI.
erase species, and that the smaller polypeptides are presum- ably metastable degradation products of the largest (approx- imately 140 kDa) polypeptide synthesized in E. coli. These results demonstrate that the yeast DNA insert in phage 102 encodes a protein product strongly related antigenically to the purified yeast RNA polymerase subunit.
If the phage 102-encoded proteins consist of a yeast DNA- encoded sequence fused with the large N-terminal fragment of @-galactosidase, as expected from the structure of Xgtll recombinants (13), they should also react with antibodies directed against @-galactosidase. However, in contrast to this expectation, the anti-6-galactosidase serum did not react with the yeast DNA-encoded proteins (Fig. 4). The serum did detect a protein the size of @-galactosidase and another band of apparently 85 kDa, both of which were also present in the Xgtll control extract. (The anti-0-galactosidase serum did detect a 160-kDa fusion protein encoded by phage 201, one of the phages with an insert unrelated to that of phage 102 (data not shown).) We conclude that the prominent bands ranging up to 140 kDa in the phage 102 lysate carry little if any 0- galactosidase amino acid sequence; either they are encoded by a transcript initiated independently of the 6-galactosidase promoter, or they represent products of translation re-start
or protein processing. Whatever the mechanism of their ori- gin, the size of the largest proteins (approximately 140 kDa) suggests that most of the 4-kb yeast DNA insert carried by phage 102 represents a protein-coding sequence.
These data are consistent with the conclusion that the yeast DNA insert in phage 102 encodes a large polypeptide that represents most of the 145-kDa yeast RNA polymerase. Therefore, it was to be expected that the IgGs that were affinity purified using the proteins synthesized in E. coli (above) should inhibit the highly purified yeast RNA polym- erase. As shown in Fig. 5 , the affinity-purified antibodies directed against the 120-140-kDa bands ( I ) inhibited the rate of the RNA polymerase reaction. The affinity-purified anti- bodies directed against the 50-kDa bands (11) were less inhib- itory.
Isolation of the Genomic Region around the Phage 102 Insert-To obtain clones that contained larger regions of the yeast genome and included the insert in phage 102, a library constructed of approximately 15-kb yeast genomic sequences inserted into the vector EMBLBA (9) was screened using the yeast insert present in phage 102 as a probe. In a screen of 15,000 recombinants, 15 independent clones were isolated, of which 12 were selected for analysis. The restriction enzyme map of the genomic insert contained in a representative phage is shown in Fig. 6. It should be noted that the EcoRI sites at both ends of the insert in phage 102 correspond to genomic EcoRI sites.
In summary, our results indicate that we have cloned the gene for the 145-kDa subunit of the yeast mitochondrial RNA polymerase. The availability of this clone has already permit- ted genetic tests of its role in vivo (14).
Acknowledgment-We are particularly grateful to Dr. Michael Snyder for his help with many aspects of this work.
REFERENCES 1. Young, R. A., and Davis, R. W. (1983) Proc. Natl. Acad. Sci. U.
2. Young, R. A., and Davis, R. W. (1984) Science 222, 778-782 3. Johnson, L. M., Snyder, M., Chang, L. M. S., Davis, R. W., and
4. Goto, T., and Wang, J. C. (1984) Cell 36, 1073-1080 5. Kelly, J. L., and Lehman, I. R. (1986) J. Biol. Chem. 261,10340-
10347 6. Snyder, M., and Davis, R. W. (1985) in Hybridoma Technology in
the Biosciences and Medicine (Springer, T. A., ed) pp. 397-406, Plenum Publishing Corp., New York
7. Davis, R. W., Botstein, D., and Roth, J. R. (1982) Advanced Bacterial Genetics, Cold Spring Harbor Laboratory, Cold Spring Harbor, NY
S. A. 80, 1194-1198
Campbell, J. (1985) Cell 43, 369-377
8. Southern, E. M. (1975) J. Mol. Biol. 98, 503-517 9. Snyder, M., Elledge, S., and Davis, R. W. (1986) Proc. Natl. Acad.
Sci. U. S. A. 83, 730-734 10. Rigby, P. W. J., Dieckmann, M., Rhodes, C., and Berg, P. (1977)
J. Mol. Biol. 113, 237-251 11. Weeks, J. R., Coulter, D. E., and Greenleaf, A. L. (1982) J. Biol.
Chem. 257,5884-5891 12. Hancock, K., and Tsang, V. C. W. (1983) A d . Biochem. 133,
157-162 13. Young, R. A,, and Davis, R. W. (1985) in Genetic.Engineering
(Setlow, J., and Hollaender, A., eds) Vol. 7, pp. 29-41, Plenum Publishing Corp., New York
14. Greenleaf, A. L., Kelly, J. L., and Lehman, I. R. (1986) Proc. Natl. Acad. Sci. U. S. A. 83, 3391-3394

Yeast Mitochondrial RNA Polymerase 10351
M. Snyder, personal communication.


















