Is repetitive DNA involved in male gametogenesis ofNarcissus papyraceus?
6
Protoplasma (1993) 174:128-133 Springer-Ver[ag i993 Printed in Austria Is repetitive DNA involved in male gametogenesis of Narcissus papyraceus? Paola Bassi* Dipartimento di Biologia Vegetale, Universitfi degli Studi "La Sapienza", Roma Received December 28, 1992 Accepted February 26, 1993 Dedicated to Prof. E. Battaglia on the occasion of his 75th birthday Summary. The male gametophyte of the Spermatophyta is of par- ticular interest when studying quantitative modifications of repetitive DNA which accompany plant differentiation. In fact, this well doc- umented system offers the opportunity to investigate the phenom- enon at the nuclear level. In particular, it permits comparison be- tween single haploid nuclei which, though derived from the same mitotic division, have different, well-defined morphological and functional processes. The aim of the present work was to investigate the behaviour of repetitive DNA during male gametogenesis in Nar- cissus papyraceus. By the use of A + T and G + C specific fiuoro- chromes (DAPI and Chromomycin Aa), and by in situ DNA diges- tion with appropriate restriction endonucleases (EcoRI-k and Hae III), it is shown that during the course of gametogenesis, gen- erative and vegetative nuclei behave differently with respect to par- ticular sections of the genome. Characteristic agglomerations of spe- cific A + T rich DNA sequences can be observed, localized by the two distal ends of the spindle-shaped generative nucleus. The surface area occupied by the agglomerations differs significantly at the ends in all observed cases. The above agglomerations are present in most of the generative nuclei and are probably composed of repetitive DNA sequences. Some embryological questions are discussed in the light of these results. Keywords: Cell differentiation; Fluorochromes; In situ endonuclease digestion; Male gametogenesis; Narcissus papyraceus; Repetitive DNA. Abbreviations: DAPI 4',6-diamidino-2-phenylindole; Eco RI'k Eco RI with "star" activity (N/AATTN). Introduction Recent research has shown that the repetitive/non-cod- ing fraction of the genome is not static within the cell, * Correspondence and reprints: Dipartimento di Biologia Vegetale, Universit~ degli Studi "La Sapienza", Piazzale A, Moro 5, 1-00185 Roma, Italy. but is characterized by continuous quantitative mod- ifications, such as amplification, under-replication and loss of specific DNA sequences. The above modifications are particularly evident in plants where they frequently accompany and charac- terize biological phenomena such as tissue differentia- tion and de-differentiation, changes of phase, ageing or tumour-induction. For instance, several A + T rich DNA sequences are under-represented during root-tip development in Viciafaba (Bassi et al. 1984) while, in the same system, selective amplification of G + C rich DNA sequences precedes cell de-differentiation after cutting the root (Natali etal. 1986). Flowering also seems to be accompanied by selective amplification of several highly repetitive and moderately repetitive DNA sequences, while cellular ageing appears to be characterized by loss of specific fractions of hetero- chromatin. These and many other examples have been reviewed recently (Bassi 1990). The observation that quantitative modification of re- petitive DNA in plants is often influenced, or even induced, by particular environmental stimuli is partic- ularly interesting. It has also been hypothesized that one possible function of the repetitive/non-coding frac- tion of the genome is to constitute a "link between the environment and gene expression: something able to intercept the different environmental signals, modify- ing itself accordingly, to later influence, through a top- ological action, gene expression itself'' (Bassi 1990, 1991). Given the above, it is evident how important it is to
-
Upload
paola-bassi -
Category
Documents
-
view
214 -
download
1
Transcript of Is repetitive DNA involved in male gametogenesis ofNarcissus papyraceus?

Protoplasma (1993) 174:128-133
�9 Springer-Ver[ag i993 Printed in Austria
Is repetitive DNA involved in male gametogenesis of Narcissus papyraceus?
Paola Bassi*
Dipartimento di Biologia Vegetale, Universitfi degli Studi "La Sapienza", Roma
Received December 28, 1992 Accepted February 26, 1993
Dedicated to Prof. E. Battaglia on the occasion of his 75th birthday
Summary. The male gametophyte of the Spermatophyta is of par- ticular interest when studying quantitative modifications of repetitive DNA which accompany plant differentiation. In fact, this well doc- umented system offers the opportunity to investigate the phenom- enon at the nuclear level. In particular, it permits comparison be- tween single haploid nuclei which, though derived from the same mitotic division, have different, well-defined morphological and functional processes. The aim of the present work was to investigate the behaviour of repetitive DNA during male gametogenesis in Nar- cissus papyraceus. By the use of A + T and G + C specific fiuoro- chromes (DAPI and Chromomycin Aa), and by in situ DNA diges- tion with appropriate restriction endonucleases (EcoRI-k and Hae III), it is shown that during the course of gametogenesis, gen- erative and vegetative nuclei behave differently with respect to par- ticular sections of the genome. Characteristic agglomerations of spe- cific A + T rich DNA sequences can be observed, localized by the two distal ends of the spindle-shaped generative nucleus. The surface area occupied by the agglomerations differs significantly at the ends in all observed cases. The above agglomerations are present in most of the generative nuclei and are probably composed of repetitive DNA sequences. Some embryological questions are discussed in the light of these results.
Keywords: Cell differentiation; Fluorochromes; In situ endonuclease digestion; Male gametogenesis; Narcissus papyraceus; Repetitive DNA.
Abbreviations: DAPI 4',6-diamidino-2-phenylindole; Eco RI'k Eco RI with "star" activity (N/AATTN).
Introduction
Recent research has shown that the repeti t ive/non-cod- ing fraction o f the genome is not static within the cell,
* Correspondence and reprints: Dipartimento di Biologia Vegetale, Universit~ degli Studi "La Sapienza", Piazzale A, Moro 5, 1-00185 Roma, Italy.
but is characterized by cont inuous quantitative mod-
ifications, such as amplification, under-replication and
loss o f specific D N A sequences. The above modificat ions are particularly evident in plants where they frequently accompany and charac- terize biological phenomena such as tissue differentia-
t ion and de-differentiation, changes o f phase, ageing or tumour- induct ion. For instance, several A + T rich D N A sequences are under-represented during root-t ip
development in V i c i a f a b a (Bassi et al. 1984) while, in the same system, selective amplification o f G + C rich
D N A sequences precedes cell de-differentiation after cutting the roo t (Natali etal. 1986). Flowering also
seems to be accompanied by selective amplification of several highly repetitive and moderate ly repetitive D N A sequences, while cellular ageing appears to be characterized by loss o f specific fractions o f hetero- chromatin. These and many other examples have been reviewed recently (Bassi 1990). The observat ion that quanti tat ive modif icat ion o f re-
petitive D N A in plants is often influenced, or even induced, by part icular environmental stimuli is partic- ularly interesting. I t has also been hypothesized that one possible funct ion o f the repeti t ive/non-coding frac-
t ion o f the genome is to constitute a "l ink between the environment and gene expression: something able to intercept the different environmental signals, modify- ing itself accordingly, to later influence, th rough a top- ological action, gene expression itself ' ' (Bassi 1990, 1991). Given the above, it is evident how impor tan t it is to

Paola Bassi: Is repetitive DNA involved in mate gametogenesis of Narcissus papyraceus? 129
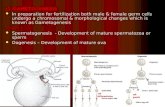
better unders tand the role played in p lant deve lopment by the repet i t ive/non-coding fract ion of the genome, and also of the mechanisms of action. In spite o f the fact that m a n y studies have tackled this problem, and tha t numerous hypotheses have been formula ted , pre- cise in format ion is still lacking. This is part ial ly a result o f the var ious models , par t icular ly for plants, involving m a n y poor ly identifiable variables. The male game tophy te o f the S p e r m a t o p h y t a is par- ticularly interesting in the s tudy o f these p rob lems be- cause it offers the oppor tun i ty o f s tudying the phe- n o m e n o n directly at the nuclear level. In part icular , it permits compar i son of repetit ive D N A fract ions in sin- gle haploid nuclei which, though derived f rom the same mitot ic division (and therefore, theoretically, geneti- cally identical), have different, well-defined m o r p h o - logical and funct ional development . The a im o f the present work was to investigate the behav iour o f re- petitive D N A during male gametogenesis in Narc issus
papyraceus .
Materials and methods
Plant material
Pollen grains and root tips of N. papyraceus Ker-Gawl (Amarylli- daceae) from an established garden population in the University of Rome were used.
Fixation and staining
Pollen grains at various stages of development were fixed in 1 : 3 acetic-alcohol for 16h and then squashed on a slide. Root tips were similarIy fixed in I:3 acetic-alcohoI for 16h and squashed (Bassi etal. 1982). Some slides were stained following the Feulgen and Giemsa techniques (Vosa 1985); others were stained with the fluo- rochromes DAPI (4',6-diamidino-2-phenylindole; Serva, Heidelberg, Federal Republic of Germany) and Chromomycin A3 (Serva), sep- arately and using a double-staining technique described by Schweizer (1976).
Restriction endonuelease digestions of cytological preparations
In situ restriction endonuclease digestion with Eco RI -i" (N/AATTN) and Hae III (GG/CC) from Boehringer (Mannheim, Federal Re- public of Germany) were performed according to Lopez-Fernandez et al. (1991). Enz)wne solutions were prepared by dissoMng t00 units of each enzyme in 100gl of the incubation buffers as indicated by
Fig. I. Pollen grain nuclei of Narcissus papyraceus stained by "Feulgen" technique, g Generative nucleus; v vegetative nucleus. Bar: 10 gm

130 Paola Bassi: Is repetitive DNA involved in male gametogenesis of Narcissus papyraceus?
Fig. 2 a~l. Pollen grain nuclei of Narcissuspapyraceus at different stages of development stained with DAPI. Arrows indicate two agglomerations of DAPI-positive DNA. g Generative nucleus; v vegetative nucleus. Bar: 10 ~m

Paola Bassi: Is repetitive DNA involved in male gametogenesis of Narcissus papyraceus? 131
the manufacturer. The "star" activity of Eco RI was obtained by the use of 25raM Tris-HC1 pH8.5, 2raM MgCI2, 12% glyceroI buffer. Treatment was carried out by placing enzyme solution on the cytological preparations and covering them with a coverslip to spread the enzyme evenly. The incubation was performed for 16h at 37 ~ in a moist chamber. Control slides were treated under the same conditions with incubation buffer only. All slides were stained using "Feulgen" techniques and fluorochromes. Observations and micrographs were made on a Zeiss Axiophot mi- croscope. Exitor/barrier filter combinations used for the fluorescence observations were those suggested by the epi-illumination manufac- turers. An HBO 50W lamp and Kodak 100 Asa and Kodak 400 Asa films for normal light and epifluorescence were used.
Image analysis
The patterns obtained were analyzed using computer image analysis software and a Cambridge Instruments Quantimet 970. For con- venience, the analysis was performed on photographs since the aim was to calculate the relative percentage of various zones within the individual nucleus stained by the fiuorochrome. Statistics were per- formed on data from a significant number of nuclei.
Results
Staining with the "Feu lgen" technique reveals no rel- evant detail in the generative or vegetative nuclei, con-
firming published data (Fig. 1). Ana logous results were obtained with Giemsa. In contrast , the use o f A + T
(DAPI) and G + C (Chromomyc in A3) specific fluo- rochromes seems to indicate that, in the course o f ga- metogenesis, the generative and vegetative nuclei be- have differently in relation to specific genome fractions. In particular, following the first gametogenetic divi- sion, there is a period during which nuclear D N A seems to be homogeneous ly stained by D A P I . After a while, however, the format ion o f agglomerat ions o f D A P I - positive D N A sequences can be observed in one o f the nuclei (specifically that destined to become the gen-
erative nucleus). Dur ing subsequent differentiation
these D N A agglomerat ions are increasingly localized
at the two poles o f the developing generative nucleus,
and, finally, clearly restricted to the distal ends o f the
spindle-shaped generative nucleus (Fig. 2 a-d) .
The above phenomenon has been observed in the ma- jori ty o f the pollen grains examined. In addition, in some cases, D N A agglomerat ion is only seen at one extremity o f the generative nucleus. A statistical anal- ysis, carried out on more than 2,500 pollen grains, shows that 53.0% o f them contain D N A agglomera- tions at bo th extremities o f the generative nucleus,
11.8% had agglomerates at only one extremity, while
31.2% were wi thout agglomerations. The remaining
4.0% were unstained or degenerate pollen grains (Table 1).
The surface area occupied by DAPI-pos i t ive D N A ag- glomerat ions varies between pollen grains. Neverthe- less, the two surface areas at the distal ends o f a single spindle-shaped generative nucleus are significantly dif- ferent f rom one another in all observed cases (Fig. 2 d).
This difference has been assessed by automat ic image analysis. The data show that the two D N A agglom- erations occupy on average 10.0% + 2.0 and
4.3% + 1.5 o f the entire generative nuclear surface (Fig. 3).
In mos t cases, DAPI-pos i t ive D N A agglomerat ions
were not stained by Chromomyc in A3, either separately or by the double-staining technique. In some cases, however, par t o f the agglomeration, particularly the more distal zone, may also be Chromomyc in A3-po- sitive.
Parallel D A P I staining did no t show any part icular agglomerat ion o f species-specific he t e rochromadn o f the meristematic nuclei o f Narcissus.
In situ restriction endonuclease digestion carried out
with Eco RI~- produces a gap coincident with the DAPI-pos i t ive agglomerat ion region. In most cases this
Table 1. Quantitative distribution of different types of pollen grains in relation to DAPI-positive agglomerations in the generative nucleus
Type Nmnber Percent
a I390 53.0 b 309 11.8 c 819 31.2 d 105 4.0 Total 2623 100.0
a Pollen grains with DNA aggIomerations at both extremities; b pollen grains with DNA agglomerations at one extremity only; e pollen grains without DNA agglomerations; d other cases (see text)
85,7
t = 1 0 , 0 %
3%
Fig. 3. Surface area occupied by DAPPpositive DNA agglomerations as percentage of the total surface area of the spindle-shaped gen- erative nucleus. Measurements were made using an arbitrary ref- erence standard corresponding to the maximum levels observed, a Major agglomeration; b minor agglomeration

i32 Paola Bassi: Is repetitive DNA involved in male gametogenesis of Narcissus papyraceus?
Fig. 4. Pollen grain nucleus of Narcissus papyraceus digested with Eco RI ~ (N/AATTN) and stained with DAPI. The arrow indicates the zone of the generative nucleus digested by the enzyme, g Gen- erative nucleus; v vegetative nucleus. Bar: 10 gm
gap is seen, especially at one end of the spindle-shaped generative nucleus (Fig. 4). In contrast, digestion with Hae III does not seem to provoke visible digestion.
Discussion
The results of the present study seem to indicate that A + T rich DNA fractions may be involved in male gametogenesis of Narcissus papyraceus. In particular, different technical approaches, such as fluorochrome staining and in situ restriction endonuclease digestion, both confirm the formation of peculiar agglomerations of DNA sequences at the distal ends of the generative nucleus. These agglomerations, probably, comprise re- petitive DNA sequences. In particular, the fact that the gap produced by Eco RI 4t is usually manifest at one end of the spindle-shaped generative nucleus may be explained by the different amount of A + T rich DNA sequences present at the two ends of the nucleus. The ability of in situ Eco R I * digestion to visibly cut the DNA agglomerations, while HaeIII does not, would seem to provide further evidence for the A + T
rich nature of the DNA in these agglomerations. The contrasting action of the restriction endonucleases could, however, depend on other factors such as the degree of methylation of the target DNA. Not only is plant DNA known to be highly methylated, but it has also been suggested that patterns of methylation may correspond to areas of low transcriptional activity such as repetitive fractions of the genome. The "star" ac- tivity of Eco RI is much less susceptible to DNA meth- ylation than other restriction enzymes including Hae III. In addition, in situ digestion is also affected by a large number of factors depending on the physical structure of the DNA itself (supercoiling around histones, po- sition in the nucleus, etc.). Further experiments are therefore necessary to improve understanding of the action of restriction enzymes on the DAPI-positive DNA agglomerates observed in this work. The above agglomerations could be the result of se- lective amplification of several specific DNA sequences; the experimental data so far, however, do not allow us to exclude the hypothesis that they are caused by re- arrangement of some DNA sequences present in the genome: cytophotometric and cytofluorimetric anal- yses are in progress to resolve this question. In any case, the data obtained could indicate that, during male gametogenesis of N. papyraceus, repetitive DNA behaves differently in the generative and the veg- etative nuclei. This contrasting behaviour could pre- sumably be related to the different morphological and functional destinies of the two nuclei. In particular, repetitive fractions of the genome might have a specific function in events which characterize male gametogenesis. For instance, the fact that the DNA agglomerations present at the two ends of an individual spindle-shaped generative nucleus always differ in size, or, that generative nuclei have only one DNA agglomeration, may indicate involvement in the entry strategy of the generative nucleus into the pollen tube. Repetitive DNA could also be involved in the type and orientation of the mitotic division that takes place within the pollen tube, or in the diverging destinies of the two spermatic nuclei. Some published data (Roman 1947) seem to confirm this latter hypothesis (see also Bruceknox and Ducker 1991). Other published data such as, fbr instance, those ob- tained from doubled haploid plants of Nicotiana syl- vestris(De Paepe et al. 1982), might find an explanation in the different behaviour of repetitive DNA in the nuclei of the male gametophyte.

Paola Bassi: Is repetitive DNA involved in male gametogenesis of Narcissus papyraceus? 133
Finally, the percentage of generative nuclei showing
this p h e n o m e n o n might be l inked to the percentage of
fertile pollen grains in a popula t ion .
Pre l iminary experiments conducted following the same
methodology on pollen grains of other Narcissus spp.
do no t show analogous results. On the other hand,
some unrela ted vegetable species show similar char-
acteristics to the system here.
Fur ther experiments using molecular biological tech-
niques, quant i ta t ive D N A analysis, and in vitro cul-
tures are now in progress with the aim of better un-
ders tanding the na ture of the p h e n o m e n o n and inves-
t igating its implications.
A c k n o w l e d g e m e n t s
The author is grateful to Prof. W. Nagl for critically reading the manuscript, to the Department of Biology of the University "Tor Vergata" of Rome for the use of the image analysis equipment and to Dr. R. Butler for help with English style and grammar.
R e f e r e n c e s
Bassi P (1990) Quantitative variations of nuclear DNA during plant development: a critical analysis. Biol Rev 65:185-225 (t991) Repetitive/non-coding DNA: a possible link between en-
vironment and gene expression in plants? Biol Zentralbl 10: 1- 13
- Cremonini R, Cionini PG (1982) Cytological localization of in- verted repeated DNA sequences in Viciafaba. Chromosoma 85: 453459
- Cionini PG, Cremonini R, Seghizzi P (1984) Underrepresentation of nuclear DNA sequences in differentiating root cells of Vicia faba. Protoplasma 123:70-77
Bruceknox R, Ducker SC (1991) The evolution of gametes from motility to double fertilization. In: Blackmore S, Barnes SH (eds) Pollen and spores, vo144. Clarendon Press, Oxford, pp 345-361
De Paepe R, Prat D, Huguet T (1982) Heritable nuclear DNA changes in doubled haploid plants obtained by pollen culture of Nicotiana sylvestris. Plant Sci Lett 28:11-28
Lopez-Fernandez C, Gosfilvez J, Ferrucci L, Mezzanotte R (1991) Restriction endonucleases in the study of eukaryotic chromo- somes. Genetica 83:257-274
Natali L, Cavallini A, Cremonini R, Bassi P, Cionini PG (1986) Amplification of nuclear DNA sequences during induced plant cell dedifferentiation. Cell Diff 18:157-161
Roman H (1947) Mitotic non-disjunction in the case of interchanges involving the B-type chromosome in maize. Genetics 32:391 409
Schweizer D (1976) Reverse fluorescent chromosome banding with chromomycin and DAN. Chromosoma 58:307-324
Vosa CG (1985) Chromosome banding in plants. In: Sharma AK, Sharma A (eds) Advances in chromosome and cell genetics. IBH Publishers, Bombay, pp 79-104