Ionotropic and metabotropic receptors - Università di Roma LUMSA · Recettori ionotropi della...
55
Ionotropic and metabotropic receptors LESSON NR. 10 - PSYCHOBIOLOGY
Transcript of Ionotropic and metabotropic receptors - Università di Roma LUMSA · Recettori ionotropi della...

Ionotropic and metabotropic
receptorsLESSON NR. 10 - PSYCHOBIOLOGY

• Channels regulated by ligand – IONOTROPIC RECEPTORS
They are membrane protein complexes, characterized by the presence on their surface not only of
the aqueous pore, but also of a specific region, said receptor site (binding site), corresponding to a
sort of pocket that receives in a stereo-specific manner one molecule, generally called ligand.
Are opposed to the metabotropic receptors that instead indicate a category of receptors, aqueous
pore-free and acting through a cascade of events, on ionotropic receptors.

IONOTROPIC RECEPTORS
Some of these receptors have the binding site on the extracellular side and then the ligand must
arrive from the outside, in other, the latter is placed inside the cell, and the ligand is produced in the
cytoplasm.
The random match between channel and ligand leads to the formation of the receptor-ligand
complex, and is therefore, entirely analogous to the formation of the enzyme substrate complex.
In both cases, in fact, the binding takes place thanks to the
weak forces, and leads to a conformational change of the
entire protein and to the opening of the channel.
There are also other important similarities:
• The formation of the binding site-ligand complex takes
place in a totally random way and therefore dependent
on the concentration of the ligand in the extra- / intra-
cellular space. This phenomenon is of great importance
in chemical synapses.

• In many cases, the binding site can
accommodate several molecules from the
ligand. These substances may have
endogenous origin (adjustment molecules) or
exogenous (drugs). They can be further
classified into:
• agonists
• competitive antagonist
• Non-competitive
antagonists
• Once formed, the receptor-ligand complex is
reversible, with the ligand that can be re-
released into the environment or degraded
IONOTROPIC RECEPTORS

• Some receptors are provided with
additional binding sites that recognize
different molecules from the ligand. In
analogy with the phenomenon of
enzymatic allosteric regulation, also in
this case the site is defined allosteric and
can be occupied by allosteric modulators,
classified as:
• allosteric activator
• allosteric inhibitor
• Even in these cases, the origin of
allosteric modulators, can be endogenous
or exogenous.
IONOTROPIC RECEPTORS

Also for the ionotropic receptors, there is the
general rule of ion channels about the
refractoriness of time between an opening
and the next.
IONOTROPIC RECEPTORS
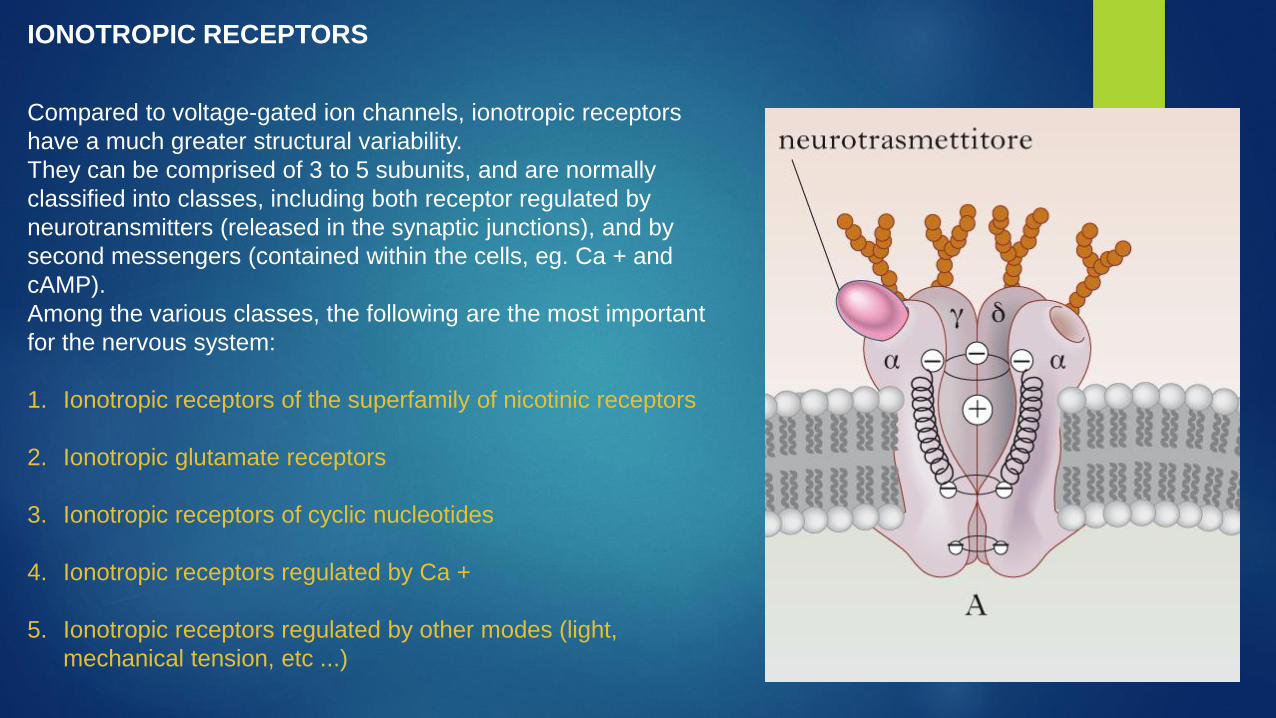
Compared to voltage-gated ion channels, ionotropic receptors
have a much greater structural variability.
They can be comprised of 3 to 5 subunits, and are normally
classified into classes, including both receptor regulated by
neurotransmitters (released in the synaptic junctions), and by
second messengers (contained within the cells, eg. Ca + and
cAMP).
Among the various classes, the following are the most important
for the nervous system:
1. Ionotropic receptors of the superfamily of nicotinic receptors
2. Ionotropic glutamate receptors
3. Ionotropic receptors of cyclic nucleotides
4. Ionotropic receptors regulated by Ca +
5. Ionotropic receptors regulated by other modes (light,
mechanical tension, etc ...)
IONOTROPIC RECEPTORS

Ionotropic receptors of the superfamily of nicotinic receptors
They are all made up of 5 main sub-units, with both
terminals in the extracellular environment. For this reason
they are also called pentameric receptors. These sub-units
may be the same or different from each other and
combined in various ways depending on the receptor in
question.
Each subunit contains four regions of alpha-helical
transmembrane (M1-M4), connected to each other by
short loops that create aqueous pore on the region M3.
In addition, on the extra-cellular side, these receptors have
one or more binding sites for neurotransmitters, and / or
for allosteric modulators.
As regards the type of transported ions, their level of
selectivity is relatively low, and then carrying cations (Na
+, K + and Ca +) or anions (Cl-)
The ionotropic nicotinic receptors have a wide variety of
isoforms and therefore a very high number of receptor
subtypes.
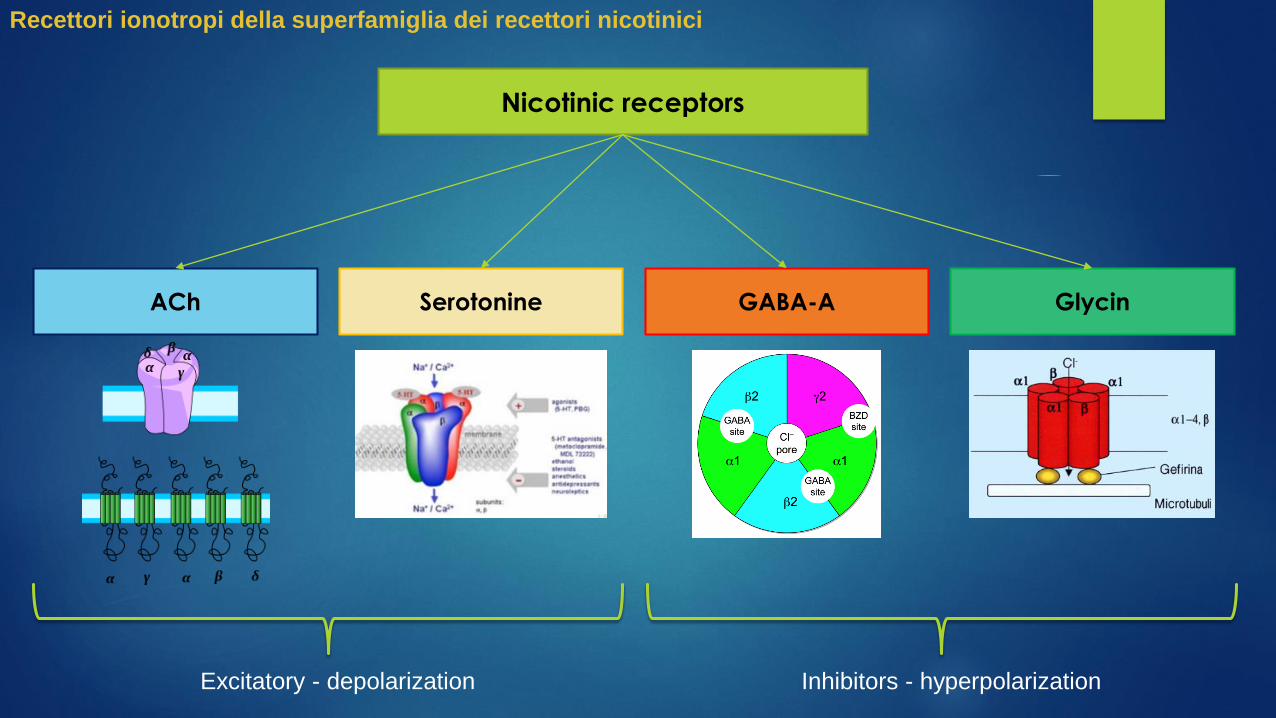
Nicotinic receptors
ACh Serotonine GABA-A
Recettori ionotropi della superfamiglia dei recettori nicotinici
Glycin
Excitatory - depolarization Inhibitors - hyperpolarization

ACh
The nicotinic receptors for the acetylcholine (ACh), get their name because of their potent agonist
nicotine.
Since they were the first ionotropic receptors to be studied and classified, they serve as a
paradigm for all pentameric ionotropic receptors.

ACh
The structure and functionality of
the receptors for ACh are different
depending on the tissue in which
they are expressed.
It is possible in fact to have:
muscle receptors (with subunits -
alpha, beta, gamma, delta and
epsilon)
neuronal receptors (with subunits -
alpha and beta)

Serotonine
The 5-HT3 receptor is a channel-receptor activated by ligand serotonin
(5-hydroxytryptamine) that allows the flow of Na + and K +; It has a
similar structure to the ones of the nicotinic cholinergic receptors, with 5
sub-units called 5-HT3a-e. The subunit 5-HT3a is the bearer of the
binding site with serotonin and is thus present in each receptor in
combination with the other subunits.
The binding of serotonin on two receptor sites determines the opening of
the channel with consequent depolarization.
These receptors are located on the parasympathetic endings in the
gastrointestinal tract and also in the splanchnic and vagal afferents.
In the central nervous system (CNS), on the other hand, there is a high
density of 5-HT3 receptors in the nucleus of the solitary tract and the
area postrema (where the vomiting center), but also in the nucleus
accumbens, amygdala, hippocampus , entorhinal cortex and frontal)
The 5-HT3 receptors in the gastrointestinal tract and in the CNS are
involved in the emetic response and form the anatomical basis for
antiemetic properties of 5-HT 3 receptor antagonists.

GABA-A
The GABA-A is the receptor for the gamma-
aminobutyric acid (GABA) which is the most
important inhibitory neurotransmitter in the
brain. GABA is the endogenous agonist of
the receptor and binds to the binding site,
mediating an allosteric modification that
does open the channel to anions, especially
Cl-.
The channel is formed by different subunits,
slightly different depending on the nervous
district in which is located, but in general
there are:
2 α subunits
2 β subunits
1 γ subunit

GABA-A
There are many agonists and antagonists of GABA A
receptor, which bind to different subunits in different
specific binding sites, dedicated to them, the most
important include:
Benzodiazepines (anxiolytics)
Barbiturates (sedatives, hypnotics)
Steroids (hormones derived from cortisol)
Ethanol (anxiolytic-like effect)
Picrotoxin (blocker)

GABA-A
Even in the case of the GABA-A, the various
sub-units can present different isoforms.
In particular we have 6 isoforms to the alpha
unit, 4 for the beta isoforms, isoforms and 4
for the gamma.
These isoforms are combined with each
other in various ways giving origin to a wide
repertoire of receptor subtypes expressed in
a specific way in the various districts of the
CNS
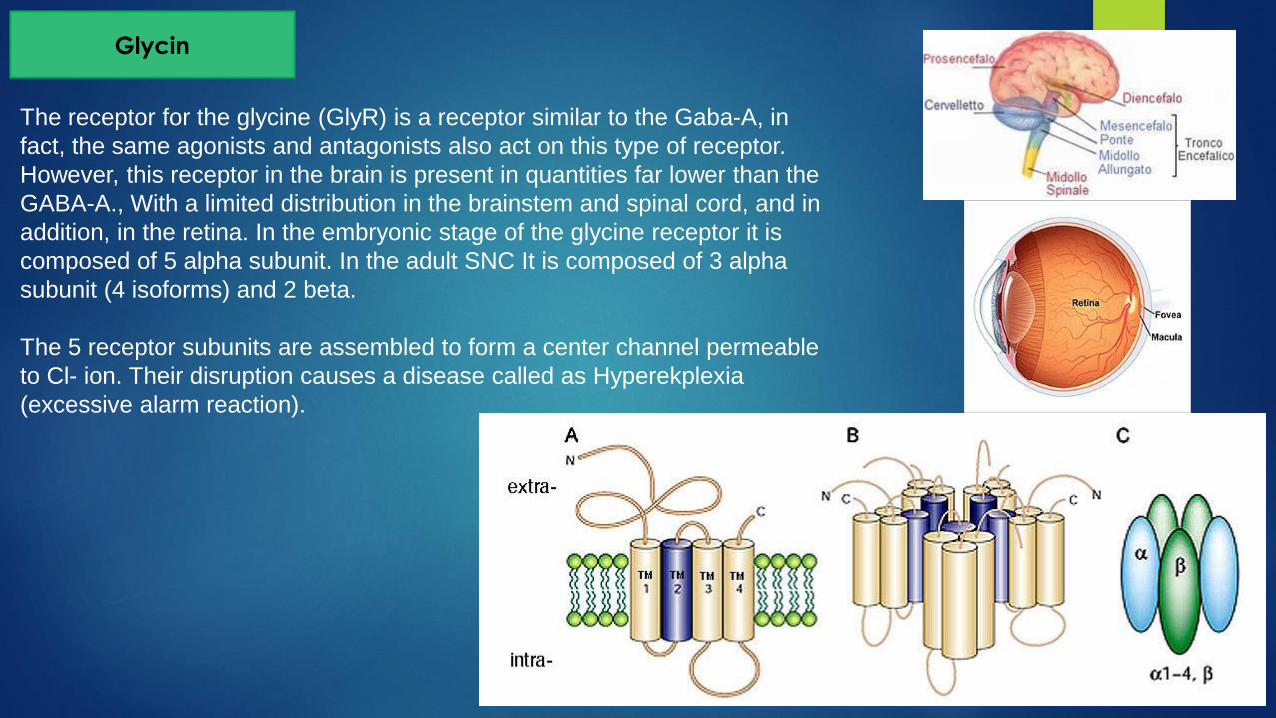
Glycin
The receptor for the glycine (GlyR) is a receptor similar to the Gaba-A, in
fact, the same agonists and antagonists also act on this type of receptor.
However, this receptor in the brain is present in quantities far lower than the
GABA-A., With a limited distribution in the brainstem and spinal cord, and in
addition, in the retina. In the embryonic stage of the glycine receptor it is
composed of 5 alpha subunit. In the adult SNC It is composed of 3 alpha
subunit (4 isoforms) and 2 beta.
The 5 receptor subunits are assembled to form a center channel permeable
to Cl- ion. Their disruption causes a disease called as Hyperekplexia
(excessive alarm reaction).

Compared to voltage-gated ion channels, ionotropic receptors
have a much greater structural variability.
They can be comprised of 3 to 5 subunits, and are normally
classified into classes, including both receptor regulated by
neurotransmitters (released in the synaptic junctions), and by
second messengers (contained within the cells, eg. Ca + and
cAMP).
Among the various classes, the following are the most important
for the nervous system:
1. Ionotropic receptors of the superfamily of nicotinic receptors
2. Ionotropic glutamate receptors
3. Ionotropic receptors of cyclic nucleotides
4. Ionotropic receptors regulated by Ca +
5. Ionotropic receptors regulated by other modes (light,
mechanical tension, etc ...)
IONOTROPIC RECEPTORS
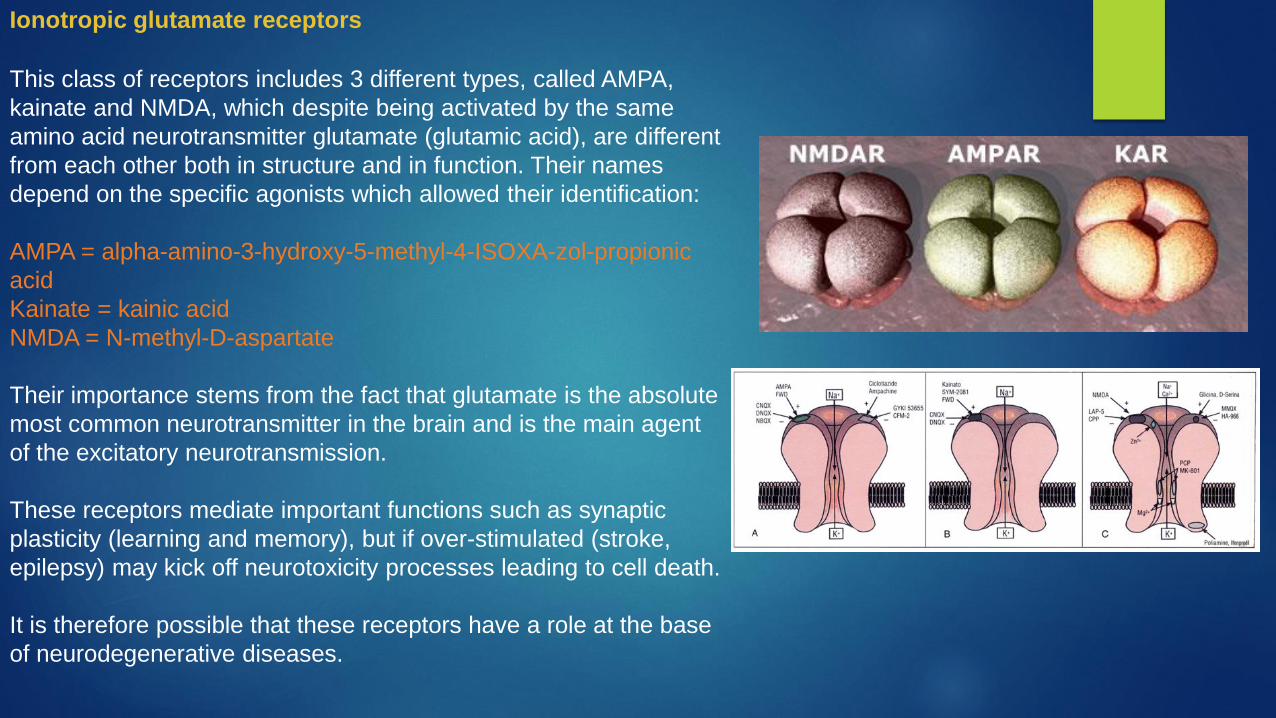
This class of receptors includes 3 different types, called AMPA,
kainate and NMDA, which despite being activated by the same
amino acid neurotransmitter glutamate (glutamic acid), are different
from each other both in structure and in function. Their names
depend on the specific agonists which allowed their identification:
AMPA = alpha-amino-3-hydroxy-5-methyl-4-ISOXA-zol-propionic
acid
Kainate = kainic acid
NMDA = N-methyl-D-aspartate
Their importance stems from the fact that glutamate is the absolute
most common neurotransmitter in the brain and is the main agent
of the excitatory neurotransmission.
These receptors mediate important functions such as synaptic
plasticity (learning and memory), but if over-stimulated (stroke,
epilepsy) may kick off neurotoxicity processes leading to cell death.
It is therefore possible that these receptors have a role at the base
of neurodegenerative diseases.
Ionotropic glutamate receptors

Regarding their specificity, the aqueous pores of AMPA and kainate
receptors have a lower specificity and allow the passage of both K +
that of Na +, and to a lesser amount of Ca2 +.
In contrast the NMDA receptor has a marked specificity for Ca2 +
and much lower for the other cations.
AMPA, KainateNMDA
Ionotropic glutamate receptors

From the structural point of view, these receptors are composed of
4 or 5 sub-main units, characterized by a common basic
organization of the polypeptide chain, with the N-terminal end in
the extracellular environment and the C-terminal end in the intra-
cellular environment.
Each subunit contains always 3 trans-membrane regions (M1, M3
and M4) and a loop (M2) located in the intracellular side the
purpose of which is to control the permeability of the aqueous pore.
Ionotropic glutamate receptors
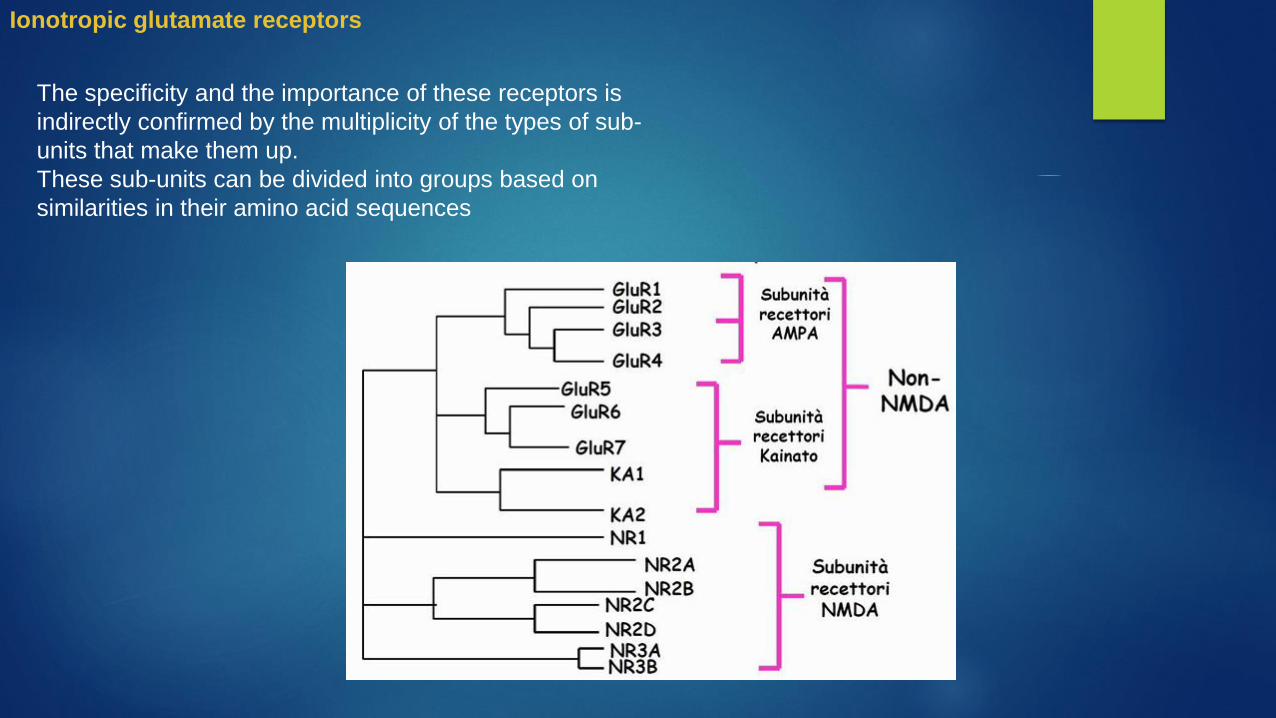
The specificity and the importance of these receptors is
indirectly confirmed by the multiplicity of the types of sub-
units that make them up.
These sub-units can be divided into groups based on
similarities in their amino acid sequences
Ionotropic glutamate receptors
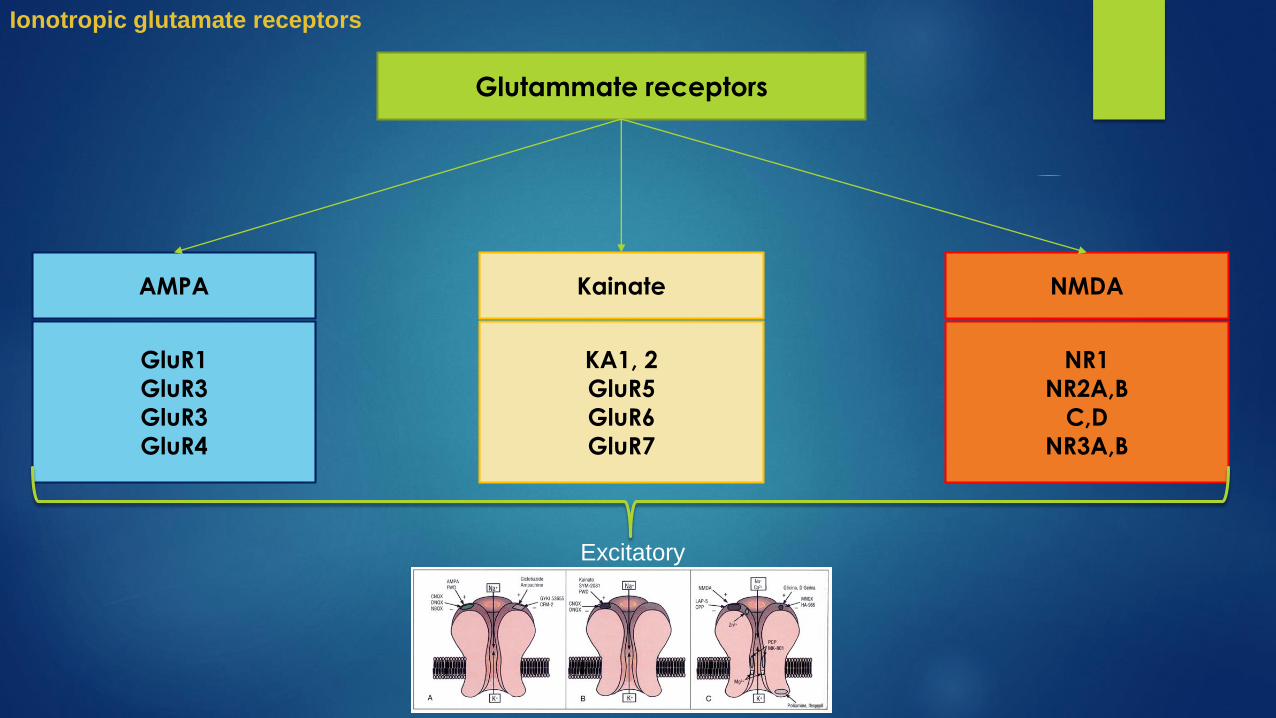
Glutammate receptors
AMPA Kainate NMDA
GluR1
GluR3
GluR3
GluR4
KA1, 2
GluR5
GluR6
GluR7
NR1
NR2A,B
C,D
NR3A,B
Excitatory
Ionotropic glutamate receptors

AMPA
The AMPA receptors are the ionotropic
more present in our brain and the main
mediators of fast excitatory transmission.
They consist of four sub-units (GluR1-4), of
which 2 are always GluR1.
Each type of sub-units is presented with
several variations and subtypes that
originate from post-transcriptional
modifications of RNA, such as:
alternative splicing
RNA editing
Or by post-translational modifications of the
polypeptide chain, such as
phosphorylation.
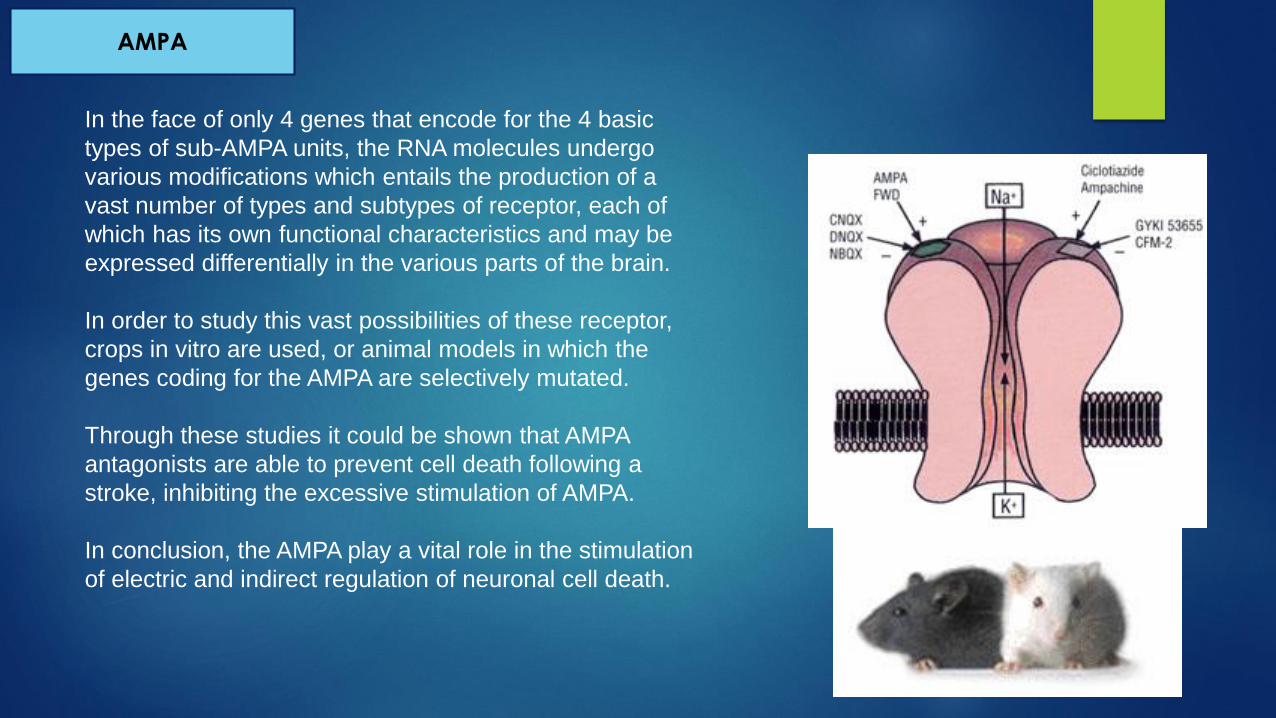
In the face of only 4 genes that encode for the 4 basic
types of sub-AMPA units, the RNA molecules undergo
various modifications which entails the production of a
vast number of types and subtypes of receptor, each of
which has its own functional characteristics and may be
expressed differentially in the various parts of the brain.
In order to study this vast possibilities of these receptor,
crops in vitro are used, or animal models in which the
genes coding for the AMPA are selectively mutated.
Through these studies it could be shown that AMPA
antagonists are able to prevent cell death following a
stroke, inhibiting the excessive stimulation of AMPA.
In conclusion, the AMPA play a vital role in the stimulation
of electric and indirect regulation of neuronal cell death.
AMPA

Kainato
The kainate (kainic acid) is a convulsant agent, that
besides being an agonist of AMPA receptors, is able to
activate the channels that have this molecule as selective
agonist, and are precisely denominated channels of
kainate.
These channels are generally formed by a complex of
tetrameric 5 KA1-2 and iGluR5-7 possible sub-units that fit
together.
These subunits may variously be combined between
them, but the presence of GluR5 or GluR6 is stable in
order to obtain a receptor that, when activated selectively
by the agonist, is able to generate a current intense
cationic modulation.
Both of these two subunits can exist in two different
variants: in the sequence of amino acids in the pore-
channel wall, are located both the variable with arginine
which makes this little permeable to calcium receptor, both
the glutamine residues that make it highly permeable to
sodium and calcium.

Less abundant in the CNS of AMPA receptors, these receptors are mainly
expressed in the striatum, in the reticular nucleus of the thalamus,
hypothalamus, in the deep layers of the cerebral cortex, the layer of granular
cells of the cerebellum, in the dentate gyrus and in the glossy layer of ' CA3
hippocampal area.
The kainate receptors appear to play an important role in the development and
plasticity of the CNS since at birth they vary in number and in the expression of
the different sub-units and are also involved in long-term potentiation (LTP).
The KA receptors coexist with other receptor subtypes in postsynaptic level.
Activation of these receptors also seem to modulate the release of GABA in
hypothalamus and hippocampus.
Kainate

NMDA
NMDA receptors have a much slower kinetics (in the order of
hundreds of milliseconds) of the AMPA and KA receptors and are
highly permeable to calcium.
As a rule, they act together with AMPA receptors and Ka, but their
specific characteristics are the basis of the involvement of these
in all higher cognitive processes but also diseases such as
psychosis or schizophrenia.
They are typically composed of four sub-units, each of which
presents variation caused exclusively by alternative splicing. The
different subunits include: NR1, NR2A-D (common) and NR3A-B,
NR4 (inhibitory).
The NR1 subunit is always present in all of the NMDA receptors.
The peculiarity of this subunit is given by the sequence of amino
acids which delimits the wall of the pore-channel where there are
asparagine sites that make this receptor highly permeable to
calcium and give this receptor other properties such as the one to
bind magnesium ions, which in the non-opening of the receptor,
are binded within the aqueous pore blocking completely the
functionality. The NR1 subunit is ubiquitously in all brain regions
while NR2A-D subunits are present preferentially in the cortex, in
hippocampus and cerebellum.

NMDA
The NMDA receptors have two different binding
sites of the main endogenous ligands that
respectively bind the L-glutamate, L-aspartate, L-
omocistinate and chinolinate, and as coactivators
ligands: glycine, D-serine and D-alanine.
As said, within the channel, there is a site for the
binding of magnesium ions.
The activation of the NMDA receptor can take place
only if at the same time both the glutamate and the
glycine interact in their binding sites, in addition to
these conditions, however, are necessary additional
contingencies.

NMDA
In total there are three, then, the situations necessary for the
channel activation:
1. Receptor binding ligand or glutamate agonists.
2. Glycine present on the second binding site
3. Removal of Mg ++ ions. This is possible because on the
postsynaptic membrane near the receptor there are also
present rapid kinetic AMPA receptors which, when
activated, lead to a rapid entry of calcium (less) and
sodium that induce a rapid depolarization of the
membrane which promotes the removal of magnesium
ions, making possible the functioning of the receptor.
The freeze due to magnesium can also be removed by
endogenous polyamines: spermidine and spermine; at low
and high concentrations, respectively, enhance and inhibit
receptor activity. Along with these substances, also ketamine
and phencyclidine act as non-competitive antagonists,
causing phenomena similar to the positive symptoms of
schizophrenia.
In conclusion, the NMDA receptors are subjected to both the
ligand control and the membrane potential control.

Compared to voltage-gated ion channels, ionotropic receptors
have a much greater structural variability.
They can be comprised of 3 to 5 subunits, and are normally
classified into classes, including both receptor regulated by
neurotransmitters (released in the synaptic junctions), and by
second messengers (contained within the cells, eg. Ca + and
cAMP).
Among the various classes, the following are the most important
for the nervous system:
1. Ionotropic receptors of the superfamily of nicotinic receptors
2. Ionotropic glutamate receptors
3. Ionotropic receptors of cyclic nucleotides
4. Ionotropic receptors regulated by Ca +
5. Ionotropic receptors regulated by other modes (light,
mechanical tension, etc ...)
IONOTROPIC RECEPTORS

Receptors of cyclic nucleotides
CNG HCN
Cyclic Nucleotide-
Gated
Hyperpolarization and
Cyclic Nucleotide-
Gated
Excitatory
Ionotropic receptors of cyclic nucleotides

They are receptors of cyclic nucleotides (cAMP and cGMP), in tetrameric
structure in which two subunits are homologues. Both the ends amino and
carboxyl, are intracellular. These receptors are composed of 6
transmembrane helices.
Moreover, there is a sequence of amino acids that enters from the
extracytoplasmic side, it crosses the membrane partly folds in on itself, and
flows out from the same side of the membrane (P LOOP).
This amino acid sequence with the S6 helix participates in the formation of
pore-channel wall.
The binding of the ligands occurs on the intracellular side. On the other
hand, cAMP and cGMP are formed within the cell and their binding site has
to be intracellular.
Receptors in the HCN S4 region contains a voltage sensor completely
analogous to those of the voltage-regulated receptor.
The receptors of the cyclic nucleotides perform important tasks in the
photoreceptors of the retina, in the olfactory sensory epithelium cells, over
that, in various districts of the CNS and P. HCN receptors are also
particularly important for the regulation of the heartbeat.
CNG HCN

Compared to voltage-gated ion channels, ionotropic receptors
have a much greater structural variability.
They can be comprised of 3 to 5 subunits, and are normally
classified into classes, including both receptor regulated by
neurotransmitters (released in the synaptic junctions), and by
second messengers (contained within the cells, eg. Ca + and
cAMP).
Among the various classes, the following are the most important
for the nervous system:
1. Ionotropic receptors of the superfamily of nicotinic receptors
2. Ionotropic glutamate receptors
3. Ionotropic receptors of cyclic nucleotides
4. Ionotropic receptors regulated by Ca +
5. Ionotropic receptors regulated by other modes (light,
mechanical tension, etc ...)
IONOTROPIC RECEPTORS

It is a diverse group of ionotropic receptors whose aqueous pore is
selective for K + and opens when the intracellular binding site,
binds a Ca2 + ion.
Their molecular structure is similar to that of the channels
regulated by voltage, they are then formed by tetrameric
complexes with 6 transmembrane domains.
The α1 subunit is the responsible for all electrophysiological and
pharmacological properties of these channels since the pore and
the voltage sensor are located on it.
The 4th segment represents the potential sensor and between the
5th and 6th α-helix there is the binding site that characterizes the
ion selectivity.
There are 9 isoforms of the α1 subunit.
Ionotropic receptors regulated by Ca +

The receptors regulated by Ca2 +, are normally classified
according to their permeability for K + ions, specifically we can
have:
High permeability (BK receptors)
Intermediate permeability (IK receptors)
Low permeability (SK receptors)
Ionotropic receptors regulated by Ca +

Compared to voltage-gated ion channels, ionotropic receptors
have a much greater structural variability.
They can be comprised of 3 to 5 subunits, and are normally
classified into classes, including both receptor regulated by
neurotransmitters (released in the synaptic junctions), and by
second messengers (contained within the cells, eg. Ca + and
cAMP).
Among the various classes, the following are the most important
for the nervous system:
1. Ionotropic receptors of the superfamily of nicotinic receptors
2. Ionotropic glutamate receptors
3. Ionotropic receptors of cyclic nucleotides
4. Ionotropic receptors regulated by Ca +
5. Ionotropic receptors regulated by other modes (light,
mechanical tension, etc ...)
IONOTROPIC RECEPTORS
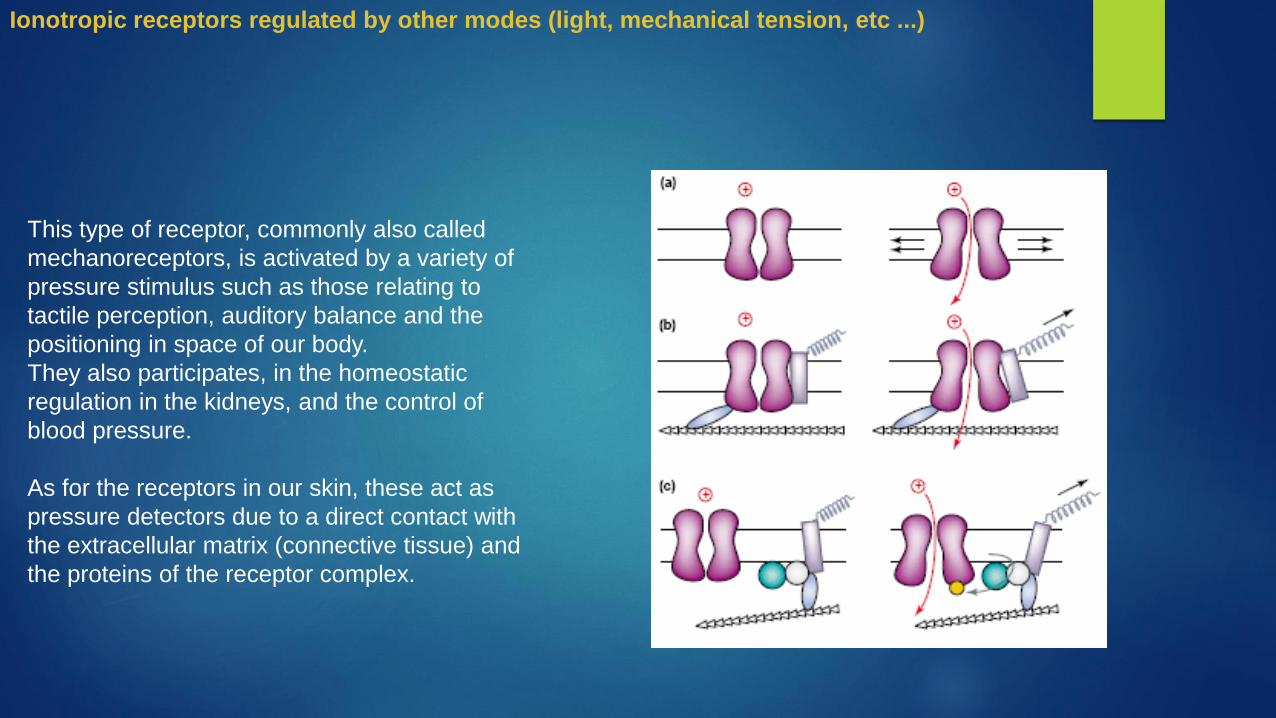
This type of receptor, commonly also called
mechanoreceptors, is activated by a variety of
pressure stimulus such as those relating to
tactile perception, auditory balance and the
positioning in space of our body.
They also participates, in the homeostatic
regulation in the kidneys, and the control of
blood pressure.
As for the receptors in our skin, these act as
pressure detectors due to a direct contact with
the extracellular matrix (connective tissue) and
the proteins of the receptor complex.
Ionotropic receptors regulated by other modes (light, mechanical tension, etc ...)

A second type of mechanoreceptors regard
those places in the organ of Corti, in 'inner ear.
These receptors constitute the final step of a
series of vibrations that depart from the
eardrum, and following transmission through
the ossicular chain, up to the oval window,
finally come to move the liquid contained in the
organ of Corti.
These shifts move a membrane called the
tectorial membrane, which is in direct contact
with the hair cells (stereocilia) which
depending of their movement, open and close
their channels, sensitive to Na + or Ca2 +.
Ionotropic receptors regulated by other modes (light, mechanical tension, etc ...)

Metabotropic receptors
They are receptors that respond to the arrival of extracellular ligands and give way to a cascade of
metabolic processes within the cytoplasm.
They are very heterogeneous from a structural point of view, and can be comprised of a single
polypeptide chain, or of 2 different sub-units which in the presence of the ligand, unite with each
other.
From the functional point of view we can classify them into two large families:
Receptors linked to enzyme activity
Receptors bound to G protein

• Receptors linked to enzyme activity
The receptors related to enzymatic activities are very
diverse from a structural point of view, although the most
famous and important are normally made up of two
polypeptide chains, which as a result of ligand binding,
unite their zones in the extracellular domain in a dimer.
The formation of extra-cellular receptor dimer, allows the
intracellular protein portions to initiate an enzymatic
activity, which in most cases is a protein kinase.
The kinases consists in the phosphorylation of other
proteins or in the self-phosphorylation of the receptor.
Depending on which amino acid is phosphorylated the
name of the receptors could change in:
Tyrosine kinase
Histidine kinase
Serine / threonine kinase
The phosphorylation operated by these receptors is
typically the first step in a series of important metabolic
processes that follow the arrival of ligands such as
growth factors (skin, neurotrophic, fibroblasts and
platelets) or hormones (insulin).
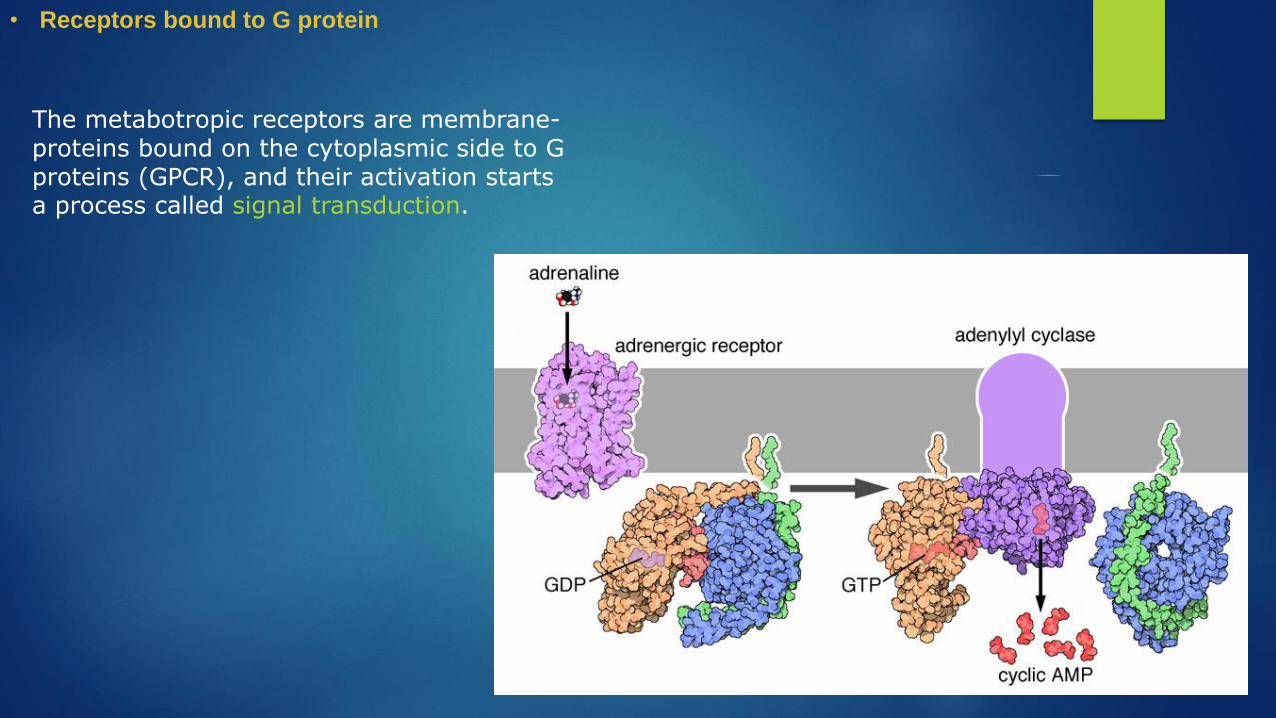
• Receptors bound to G protein
The metabotropic receptors are membrane-proteins bound on the cytoplasmic side to G proteins (GPCR), and their activation starts a process called signal transduction.
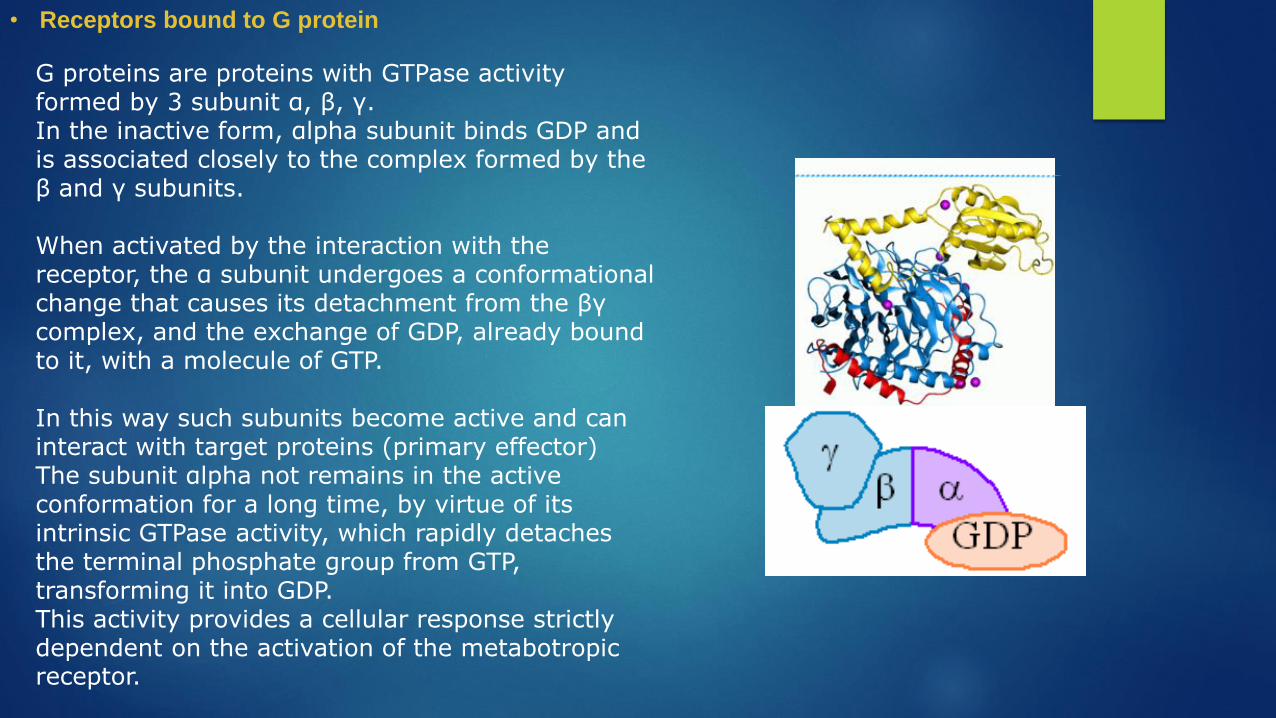
G proteins are proteins with GTPase activity formed by 3 subunit α, β, γ.In the inactive form, αlpha subunit binds GDP and is associated closely to the complex formed by the β and γ subunits.
When activated by the interaction with the receptor, the α subunit undergoes a conformational change that causes its detachment from the βγ complex, and the exchange of GDP, already bound to it, with a molecule of GTP.
In this way such subunits become active and can interact with target proteins (primary effector)The subunit αlpha not remains in the active conformation for a long time, by virtue of its intrinsic GTPase activity, which rapidly detaches the terminal phosphate group from GTP, transforming it into GDP. This activity provides a cellular response strictly dependent on the activation of the metabotropic receptor.
• Receptors bound to G protein

Signal transduction
Ligand
Metabotropic receptor
G protein
Enzyme - inactive
GDP

Bond of the Ligand with the receptor – first messanger
conformational change in the receptor that leads him
to bind with the receptor
Signal transduction
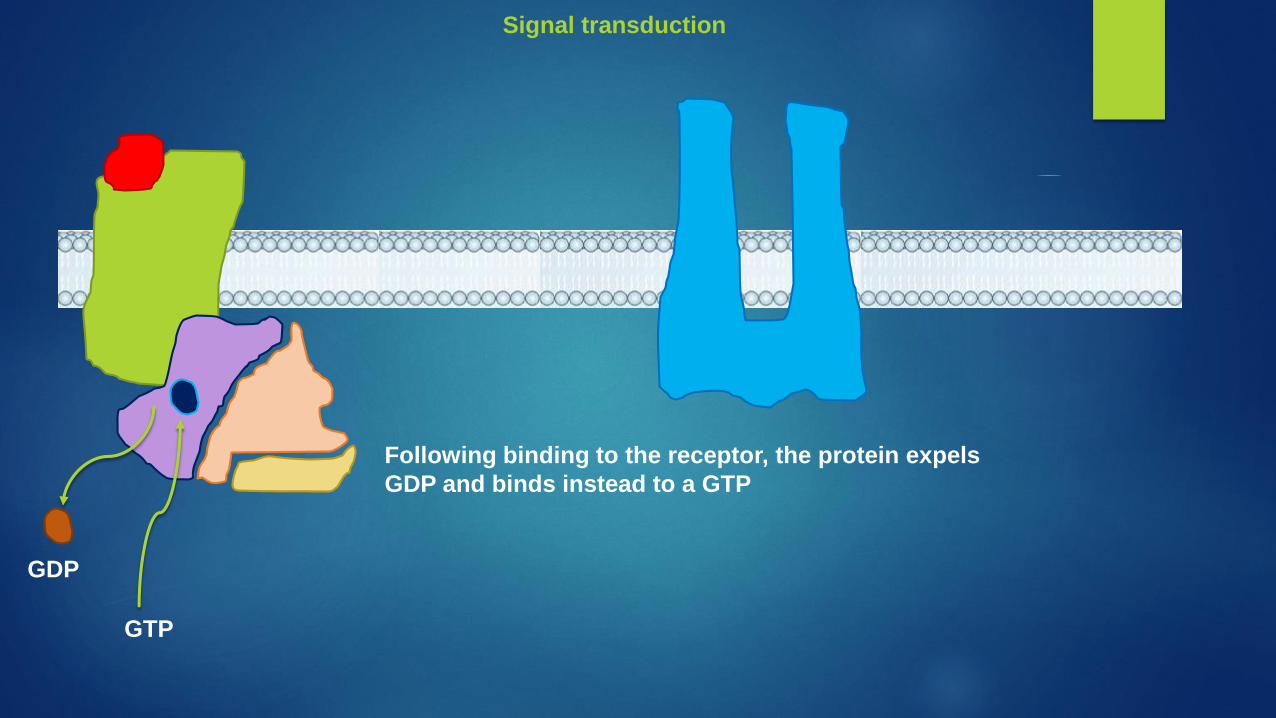
Following binding to the receptor, the protein expels
GDP and binds instead to a GTP
GDP
GTP
Signal transduction

The presence of GTP instead of GDP cause the
detachment of the protein from the receptor, and the
division into two subunits G-alpha-GTP and the
heterodimer Beta-gamma.
By the way, they remains anchored to the plasma
membrane and are free to move nearby the receptor
area.
Signal transduction

The complex G-alpga-GTP meet an enzyme that will
act as the primary effector in the transduction
process. The enzyme will start using the energy
derived from the hydrolysis of GTP to GDP to produce
new molecules that will act as second messengers to
go to activate secondary effector
Likewise, also the beta-
gamma complex will
bind to other primary
effectors to start the
production of other
metabolic processes
Signal transduction

After the hydrolysis of GTP to GDP, the
alpha complex detaches from the
primary effector ending the production
of second messengers.
Once back free in the cytoplasm the
alpha complex can meet a beta-gamma
complex free and reform the original G
protein.
Signal transduction

Signal transduction


GPCRs possess seven transmembrane domains to α-helix, a binding site for the neurotransmitter place deep in the center of the portion that faces the extracellular and intracellular domain that makes contact with G protein
And through the interaction with these proteins that metabotropic receptors exert their effects.
• Receptors
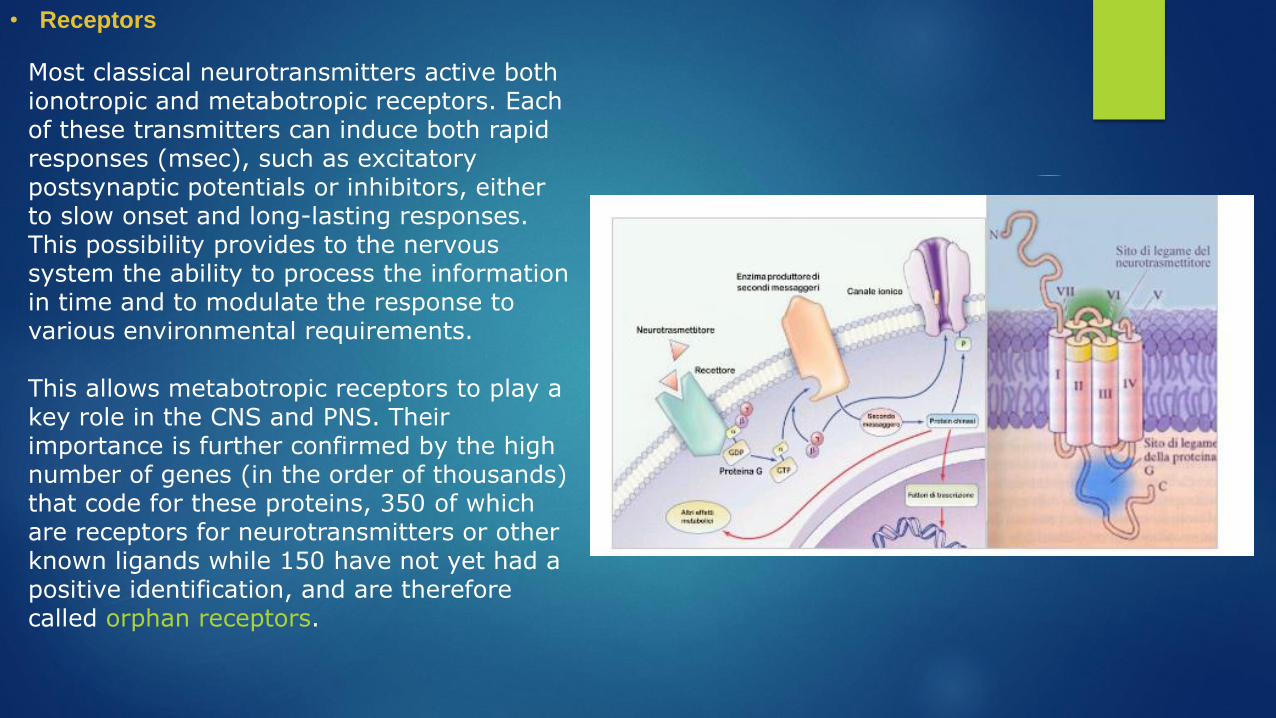
• Receptors
Most classical neurotransmitters active both ionotropic and metabotropic receptors. Each of these transmitters can induce both rapid responses (msec), such as excitatory postsynaptic potentials or inhibitors, either to slow onset and long-lasting responses. This possibility provides to the nervous system the ability to process the information in time and to modulate the response to various environmental requirements.
This allows metabotropic receptors to play a key role in the CNS and PNS. Their importance is further confirmed by the high number of genes (in the order of thousands) that code for these proteins, 350 of which are receptors for neurotransmitters or other known ligands while 150 have not yet had a positive identification, and are therefore called orphan receptors.

In general, GPCRs can be grouped into three families:
The family A, is by far the most numerous and includes most of the receptors for the monoamines and neuropeptides.
Family B consists of the secretin receptor, glucagon and calcitonin.
The C family consists mainly of metabotropic glutamate receptors and receptors sensitive to Ca2 +
An activated receptor ligand can activate many copies of G proteins with a cascading effect that amplifies the extracellular signal.
• Receptors

There are several classes of G proteins with specificity both in the target and in the activity. Gs, Gi, and Gq are the most important.
• G protein and first effector

The second messengers are responsible for the induction of new cellular activities:
cAMP activates protein kinase A (PKA), calcium ions activate, together with diacylglycerol, protein kinase C (PKC) and, through interaction with calmodulin, another regulatory protein, the calcium-calmodulin dependent protein kinase (CAMK ).
All of these kinases, cascading, phosphorylate many protein targets within the cell, changing the activity.
Another important function of PKA is the activation of CREB transcription factor that activates the transcription of genes that encode proteins that once synthesized, may bind to ion channels, enzymes and / or structural proteins, modifying the activity.
• Second messanger