
Investigation of immunomodulatory properties of ...
150
Investigation of immunomodulatory properties of neurovirulent viruses – in vitro and in vivo effects of canine distemper virus Visar Qeska University of Veterinary Medicine, Hannover Department of Pathology Center for Systems Neuroscience
Transcript of Investigation of immunomodulatory properties of ...

Investigation of immunomodulatory
properties of neurovirulent viruses – in vitro
and in vivo effects of canine distemper virus
Visar Qeska
University of Veterinary Medicine, Hannover
Department of Pathology
Center for Systems Neuroscience




University of Veterinary Medicine Hannover
Department of Pathology
and
Center for Systems Neuroscience
Investigation of immunomodulatory properties of
neurovirulent viruses – in vitro and in vivo effects of canine
distemper virus
Thesis
Submitted in partial fulfillment of the requirements for the degree
Doctor of Philosophy (PhD)
awarded by the University of Veterinary Medicine Hannover
by
Visar Qeska
(Republic of Kosovo)
Hannover 2013

Supervisor: Prof. Dr. Wolfgang Baumgärtner
Supervision group: Prof. Dr. Wolfgang Baumgärtner
Prof. Dr. Andrea Tipold
Prof. Dr. Martin Stangel
1st Evaluation: Prof. Dr. Wolfgang Baumgärtner
Department of Pathology, University of Veterinary Medicine, Hannover
Prof. Dr. Andrea Tipold Small Animal Clinic, University of Veterinary Medicine, Hannover
Prof. Dr. Martin Stangel Department of Neurology, Hannover Medical School, Hannover 2
nd Evaluation: Prof. Dr. Susanne Alldinger
Department of Pathology, Faculty of Veterinary Medicine, Justus-Liebig University, Giessen
Date of final exam: 12.04.2013
Visar Qeska was supported by Young cell scheme VI, European Council and Kosovo
government, Department of Pathology, Veterinary University Hannover, Germany, and
Center for Systems Neuroscience (ZSN), Hannover . This study was in part supported
by the German Research Foundation (FOR 1103, BA 815/10-2 and BE 4200/1-2).

To Lejla

.
“Truth is not a democracy”
(Neil deGrasse Tyson)

List of publications
Parts of the thesis have been published / submitted in peer-reviewed journals previously:
Qeska V, Baumgärtner W, Beineke A, 2013. Species-specific properties and translational aspects of
canine dendritic cells. Vet. Immunol. Immunopathol. 151, 181-192
Qeska V, Barthel Y, Iseringhausen M, Stein VM, Tipold A, Baumgärtner W, Beineke A. Depletion of
Foxp3+ regulatory T cells as a putative prerequisite for lesion initiation in canine distemper virus induced
demyelinating leukoencephalitis. Vet. Res. Submitted.


i
Table of content
Chapter 1: Aims of the study ..................................................................................................................... 1
Chapter 2: General introduction ................................................................................................................ 3
2.1 Canine distemper virus (CDV) ....................................................................................................... 3
2.1.1 Viral properties of canine distemper virus ................................................................................. 4
2.1.2 Canine distemper virus infection, receptors and cell tropism ................................................... 6
2.1.3 Pathogenesis and clinical manifestation of canine distemper .................................................. 7
2.1.4 Pathology of lymphoid organs and mechanism of immunosupression ..................................... 9
2.1.5 Neuropathology of canine distemper ...................................................................................... 11
2.1.6 Immune responses in canine distemper demyelinating leukoencephalitis ............................. 14
2.1.7 Cytokine expression in canine distemper ............................................................................... 15
2.2 Dendritic cells ................................................................................................................................ 16
2.2.1 Dendritic cells in dogs ............................................................................................................. 18
2.2.2 Dendritic cells in viral diseases ............................................................................................... 20
2.3 Regulatory T cells ......................................................................................................................... 23
2.3.1 Regulatory T cells in viral diseases ......................................................................................... 26
Chapter 3: Regulatory T cells in canine distemper infection . ............................................................. 29
Chapter 4: Canine dendritic cells ............................................................................................................ 67
Chapter 5: Dendritic cells in canine distemper infection ...................................................................... 69
Chapter 6: General discussion ................................................................................................................ 93
6.1 The role of innate immunity in demyelinating disorders .......................................................... 93
6.2 Role of regulatory T cells in neurological diseases .................................................................. 96
6.3 Role of dendritic cells in demyelinating disorders .................................................................... 98
6.4 Interaction of dendritic cells and regulatory T cells in demyelinating and chronic viral diseases ............................................................................ 100
6.5 Conclusion ................................................................................................................................... 103
Chapter 7: Summary ............................................................................................................................... 105
Chapter 8: Zusammenfassung .............................................................................................................. 109
Chapter 9: References ............................................................................................................................ 113
Chapter 10: Acknowledgements ........................................................................................................... 135

ii
List of abbreviations
APC antigen presenting cell
bmDCs bone marrow derived dendritic cells
CCL22 C-C motif chemokine 22
CCR4 C-C chemokine receptor type 4
CDV canine distemper virus
CNS central nervous system
DCs dendritic cells
DL demyelinating leukoencephalitis
EAE experimental autoimmune encephalitis
Flt3L Fms-related tyrosine kinase 3 ligand
Foxp3 forkhead box P3
FV Friend retrovirus
GM-CSF granulocyte macrophage-colony stimulating factor
HIV human immunodeficiency virus
IHC Immunohistochemistry
IFN interferon
IL interleukin
LC Langerhans cells
MHC II major histocompatibility complex class II
moDC monocyte-derived dendritic cells
MS multiple sclerosis
MV measles virus
PBMC peripheral blood mononuclear cells
PMS periodical microstructure
rc recombinant canine
rh recombinant human
SLAM signalling lymphocyte activation molecule
TME Theiler’s murine encephalomyelitis
TMEV Theiler’s murine encephalomyelitis virus

iii
List of figures
Figure 1 Schematic diagram of the morbillivirus genome
and cell receptors for canine distemper virus (CDV)………………5
Figure 2 Effects of canine distemper virus (CDV) upon immune cells…....10
Figure 3 Possible mechanisms of demyelination in canine distemper……13
Figure 4 Dendritic cell (DC) lineages of dogs………………………………..19
Figure 5 Proposed mechanisms of interaction between
measles virus (MV) and dendritic cells (DC)…………………..…..23
Figure 6 Schematic diagram of mechanisms involved
in regulatory T cell (Treg)-mediated immunosuppression……….25
Figure 7 Dualism of regulatory T cells (Treg) in virus infection……………26
Figure 8 Proposed homeostatic feedback loops between
dendritic cells (DCs) and regulatory T cells (Treg)……………….101


1
Chapter 1: Aims of the study
Chapter 1: Aims and hypothesis of the present study
Canine distemper virus (CDV) infection causes a long lasting immunosuppression in
dogs. Due to its profound lymphotropism and preferential infection of CD150-expressing
immune cells, generalized lymphoid depletion and severe lymphopenia can be found
during the acute distemper phase, which is similar to human measles virus (MV)
infection (von Messling et al. 2006; Beineke et al. 2009; Sellin et al. 2009). Moreover,
due to demyelination that occurs with disease progression, canine distemper represents
a spontaneous model for human myelin disorders such as multiple sclerosis (MS)
(Vandevelde and Zurbriggen, 2005; Sips et al. 2007). Several publications
demonstrated the important role of regulatory T cells (Treg) and dendritic cells (DC) for
the development of chronic central nervous system (CNS) disorders (Wu and Laufer,
2007; Zozulya and Wiendl, 2008). Interestingly, although a therapeutic effect of these
immunomodulatory cells can be observed in autoimmune disorders, both beneficial and
detrimental effects have been described in infectious disorders (Belkaid and Rouse,
2005; Schneider-Schaulies and Dittmer, 2006; Zozulya and Wiendl, 2008). Krakowka et
al. (1982) discussed the existence of a mononuclear suppressor cell population that
causes inhibition of immune responses in CDV-infected dogs. However, this cell
population, which potentially includes Treg and/or DCs, has not been identified yet.
During the chronic phase of CDV-infection, viral antigen can be found in cells with a DC-
like morphology in splenic germinal centers (Wünschmann et al. 2000). Thus, similar to
the proposed effects of MV upon human DCs, it is hypothesized that CDV also inhibits
the differentiation of canine DCs, which might subsequerntly lead to persistent
depressive effects upon the immune system (Beineke et al. 2009; Céspedes et al. 2010;
Reuter et al. 2010). Moreover, Treg have been demonstrated to cause long lasting
immunosuppression in human measles (Sellin et al. 2009; Griffin 2010). Since so far
none of the above mentioned cell populations have been investigated in canine
distemper infection, the aims of the project were to determine disease phase-dependant
phenotypical changes and the associated cytokine expression in lymphoid organs of
CDV-infected dogs and to testify the hypothesis that a peripheral depletion of Treg

2
Chapter 1: Aims of the study
causes a lack of CNS-infiltrating immunomodulatory cells in the predemyelinating phase
of CDV-infection. The latter might represent a potential prerequisite for immune
mediated demyelination. Moreover, the effect of CDV upon canine DCs was
investigated in vitro to depict further potential parallels between canine distemper and
human measles.

3
Chapter 2: General introduction
2 Chapter 2: General introduction
2.1 Canine distemper virus (CDV)
Morbilliviruses belong to the Paramyxoviradae family and include a number of highly
pathogenic viruses, such as measles virus (MV), rinderpest virus, canine distemper
virus (CDV), and peste-des-petits-ruminants virus, which cause devastating diseases in
humans and animals (Beineke et al. 2009; Langedijk et al. 2011). In the last decades,
morbilliviruses additionally emerged as causative agents of several mass-mortalities in
bottlenose dolphins, Siberian seals, harbor seals, and striped dolphins (Osterhaus et al.
1990; Lipscomb et al. 1994; Saliki et al. 2002; Beineke et al. 2010; Stimmer et al. 2010).
Canine distemper is a fatal disease of carnivores with a worldwide distribution which
affects mainly dogs. CDV-infection is also found in other animals including felidae,
mustelidae, procyonidae, phocidae, tayassuidae, and non-human primates (Macaca
fuscata; Pringle 1999; Sips et al. 2007; Beineke et al. 2009). Efforts to prevent the CDV
infection by vaccination are largely successful (Patel et al. 2012). However, even with a
broad vaccination regiment, distemper outbreaks have been reported in France,
Germany, USA, Japan and Finland (Mori et al. 1994; Johnson et al. 1995; Beineke et al.
2009). Moreover, canine distemper has been observed in vaccinated animals following
infection with genetically different CDV strains (Simon-Martínez et al. 2008). CDV-
infection represents a systemic disease which affects the respiratory and
gastrointestinal tract, skin, lymphoid tissues, and CNS (Krakowka et al. 1980;
Baumgärtner et al. 1989). Moreover, among other morbilliviruses, CDV-infection shows
a high incidence of CNS complications (Rudd et al. 2006). Pathological changes that
occur during CDV-induced demyelinating leukoencephalitis (DL) show remarkable
similarities with human multiple sclerosis (MS), making DL a naturally occurring
translational model for human demyelinating disorders (Baumgärtner and Alldinger,
2005). Additionally, the disease course and pathogenesis in canine distemper resemble
those of human MV infection including, fever, rash, respiratory signs, lymphopenia, and

4
Chapter 2: General introduction
profound immunosuppression with generalized depletion of lymphoid organs during the
acute disease phase (von Messling et al. 2006; Beineke et al. 2009; Sellin et al. 2009).
Thus, CDV-infection of dogs is further appreaciated as a model to investigate
morbillivirus induced alterations of the immune system.
2.1.1 Viral properties of canine distemper virus
CDV is an enveloped, negative-sense, single-stranded RNA virus. Similar to other
paramyxoviruses CDV contains six structural proteins: the nucleocapsid (N), phospho
(P), large (L), matrix (M), hemagglutinin (H) and fusion (F) protein, and two accessory
non-structural proteins (C and V) found as extratranscriptional units within the P gene
(Örvell, 1980; Dhiman et al. 2004; Röthlisberger et al. 2010) (Fig. 1). The lipid envelope
surrounding the virion contains two surface proteins (F and H), which mediate virus
entry into the cell. During morbillivirus infection, the initial interaction with the host cell is
mediated by the envelope-anchored attachment protein, the H protein, an essential viral
component, which, assisted by the viral fusion protein, initiates virus cell entry (Stern et
al. 1995; von Messling et al. 2001; Langedijk et al. 2011). The N, P and L proteins are
responsible for virus replication, while the M protein connects the surface glycoproteins
and N protein during viral maturation (von Messling et al. 2001; Röthlisberger et al.
2010). Co-expression of both H and F glycoprotein are sufficient and necessary to
induce cell fusion. Moreover, the H protein represents the major factor determining CDV
cell tropism (Stern et al. 1995; Plattet et al. 2005). The cell fusion in paramyxovirus
infection seems to be a complex process. The current model proposes that the H
protein undergoes conformational changes after binding with the host cell, which also
affects the structure of the F protein resulting in insertion of hydrophobic fusion peptide
into the cell membrane with the result of final binding to the host cell (Lamb 1993;
Plattet et al. 2005). On the other side, von Messling et al. (2001) reported that the
fusogenicity is solely determined by properties of the H protein. Moreover, the F protein

5
Chapter 2: General introduction
might have a key role in viral persistence (Plattet et al. 2005; Plattet et al. 2007).
Additionally, M and N proteins which are important for viral budding might represent co-
factors for the ability of CDV to induce persistent infection (Stettler et al. 1997; Plattet et
al. 2007). Additionally, the role of V proteins accounts for rapid viral multiplication in
lymphocytes and the inhibition of interferon signaling pathways (von Messling et al.
2006; Röthlisberger et al. 2010).
Figure 1. Schematic diagram of the morbillivirus genome and cell receptors for canine distemper
virus (CDV). A) The morbillivirus virion contains the RNA genome and six structural proteins: the
nucleocapsid (N), pospho (P), matrix (M), fusion (F), hemagglutinin (H) and large (L) protein. The H and F
proteins are associated with the envelope. The V and C proteins are non-structural proteins. Additionally,
CDV uses the signalling lymphocyte activation molecule (SLAM) as a receptor for cell entry. Additional
receptors which are supposed to support infection are CD9 and CD46 B) The non-segmented RNA
genome contains six genes. The P gene encodes for the P, C and V proteins. The P and C proteins are
translated from overlapping reading frames on a functionally bicistronic mRNA and the V protein is
translated from V mRNA, which is formed after insertion of a single nucleotide by RNA editing. Modified
from Yanagi et al. (2006).
After virus attachment, the virus replicates in the cytoplasm of the host cell, and
cytoplasmic as well as intranuclear inclusions can be found in many cell types
(Baumgärtner et al. 1989). Until now, only one serotype and several co-circulating CDV

6
Chapter 2: General introduction
genotypes with differences in virulence and cell tropism have been described (Haas et
al. 1999).
2.1.2 Canine distemper virus infection, receptors and cell tropism
For MV several proteins that serve as receptors for virus entry have been described.
The signaling lymphocyte activation molecule (SLAM), also known as CD150, has been
identified as an ultimate and universal morbillivirus receptor (Fig. 1) (Hahm et al. 2004;
von Messling et al. 2006; Sato et al. 2012). SLAM is a glycosylated transmembrane
protein that is constitutively expressed on immature thymocytes, DCs, CD45ROhigh
memory T cells and a proportion of B cells, and is rapidly induced on T and B cells after
activation (Cocks et al. 1995; Tatsuo et al. 2000; Beineke et al. 2009). The expression
of SLAM on leukocytes in MV and CDV-infection accounts for the preferential infection
of lymphoid organs (lymphotropism) with subsequent lymphoid depletion and
immunosuppression (Wünschmann et al. 2000; Minagawa et al. 2001; Wenzlow et al.
2007; Langedijk et al. 2011). For instance, during early CDV-infection, SLAM is up-
regulated on lymphoid cells, which is supposed to enhance virus amplification in the
host (Wenzlow et al. 2007). Beside the SLAM receptor, certain strains of MV are able to
infect cells by interaction with the CD46 receptor (Naniche et al. 1993; Erlenhöfer et al.
2002). CD46 serves as a receptor predominantly for attenuated viruses and only for few
wild type strains in vitro (Erlenhöfer et al., 2002). Whether wild type MV in vivo interacts
with CD46 or not, and whether all MV-strains have the ability to use SLAM as a
receptor, is still not known (Dörig et al. 1993; Erlenhöfer et al. 2002; Yanagi et al. 2006).
The role of CD46 in CDV-infection has not yet been confirmed, although the lack of
SLAM in certain CDV-target cells supports the assumption of SLAM independent
infection pathways (Wenzlow et al. 2007, Beineke et al. 2009). So far, CD46 molecules
have been identified only in neoplastic lymphoid cells of dogs (Suter et al., 2005).
Additional receptors, such as CD9, are discussed as possible factors for CDV-infection

7
Chapter 2: General introduction
of Vero cells. CD9, a tetraspan transmembrane protein, was shown to induce cell-to-cell
fusion, but not virus-to-cell fusion (Löffler et al. 1997; Schmid et al. 2000). Since no
direct binding of the virus with CD9 can be demonstrated, this receptor is supposed to
represent a co-factor for viral infection as part of the receptor complex or by effecting
the expression of other receptor molecule (Löffler et al. 1997). Recently, nectin-4 was
identified as a new receptor for MV (Mühlebach et al. 2011). Its role in canine distemper
remains to be determined.
CDV is a pantropic virus that shows a broad cell tropism. Accordingly, CDV can be
found in cells of the respiratory, gastrointestinal and urinary tract, as well as in lymphoid
tissues, endocrine organs and the central nervous system (CNS; Baumgärtner et al.
1989; Gröne et al. 2004; Beineke et al. 2009). Moreover, infection of various cell types
can be found in canine distemper (Vandevelde and Zurbriggen, 2005; Seehusen et al.
2007). In the CNS, astrocytes, microglia, and oligodendrocytes, can get infected
regardless of the CDV strain, while the infection of neurons is strain dependent (Pearce-
Kelling et al. 1991; Orlando et al. 2008). During the acute disease stage, astrocytes
represent the main cell population infected by CDV (Alldinger et al. 2000; Seehusen et
al. 2007). Interestingly, even though CNS lesions are characterized by white matter
vacuolization and demyelination, only a limited number of oligodendrocytes can be
found to be infected (Zurbriggen et al. 1998; Vandevelde and Zurbriggen, 2005).
2.1.3 Pathogenesis and clinical manifestation of canine distemper
The disease course including the duration and severity of clinical signs depends mainly
on the virulence of the strain as well as on the age and immune status of the animal.
Transmission of the virus is facilitated by sneezing, coughing and close contact.
Accordingly, animals are infected primarily by inhalation of viruses and infective
droplets, respectively (Krakowka et al. 1980). Initially, virus replicates in lymphoid tissue
of the upper respiratory tract. Here, monocytes and macrophages are the first cells that

8
Chapter 2: General introduction
get infected and propagate the virus (Appel et al. 1970). The incubation period varies
from one to four weeks (Krakowka et al. 1980; Beineke et al. 2009). Animals display a
broad spectrum of clinical signs including lethargy, anorexia, dehydration, weight loss,
pneumonia, and neurological signs. Furthermore, development of a biphasic fever
represents a characteristic clinical finding (Wright et al. 1974). During the first viremic
phase (three to six days post-infection), generalized infection of all lymphoid tissues
with lymphopenia, profound immunosuppression and transient fever is observed. The
second viremia takes place several days later, and is associated with high fever and
infection of parenchymal tissues such as the respiratory tract, gastrointestinal tract, skin,
and CNS (Appel et al. 1969, Krakowka et al. 1980, Beineke et al. 2009). During this
disease stage, various signs such as conjunctivitis, nasal discharge, anorexia,
neurological disturbances, gastrointestinal signs and respiratory signs can be observed
(Krakowka et al. 1980; Beineke et al. 2009). Respiratory signs are a consequence of
virus-induced rhinitis and interstitial pneumonia, which can exceed to suppurative
bronchopneumonia due to secondary bacterial infection. Vomiting, diarrhea and
dehydration follow infection of the gastrointestinal tract (Greene and Apple, 1998;
Decaro et al. 2004;). Neurologic signs depend on viral distribution in the CNS and
include hyperesthesia, cervical rigidity, seizures, cerebellar and vestibular signs, as well
as paraparesis or tetraparesis with sensory ataxia (Deem et al. 2000; von Rüden et al.
2012). Neurological manifestations include encephalopathy, acute encephalitis,
subacute to chronic demyelinating encephalitis, and polioencephalitis (Nesseler et al.
1999; Rudd et al. 2010; Wyss-Fluehmann et al. 2010). Recovery depends on the
immune state of the animal. Particularly, a strong and effective cellular immune
response can eliminate the virus prior to the infection of parenchymal tissues, while
weak and delayed cellular and humoral immune responses lead to virus spread and
persistence, respectively

9
Chapter 2: General introduction
2.1.4 Pathology of lymphoid organs and mechanism of immunosupression
Systemic CDV-infection causes depletion of multiple lymphoid tissues such as spleen,
lymph nodes, thymus and mucosa associated lymphatic tissues (MALT) of dogs.
Microscopic changes in lymphoid organs include loss of B and T cell areas, formation of
giant cells, intracytoplasmic inclusion bodies in lymphoid cells, follicular necrosis, and
thymic atrophy (Fig. 2) (Iwatsuki et al. 1995; Wünschmann et al. 2000; Beineke et al.
2009). Moreover, syncytia formation and cell death of immune cells lead to complete
loss of secondary follicles (Iwatsuki et al. 1995). During the acute disease stage, CDV
antigen is found mostly in lymphocytes and macrophages, located predominantly in T
cell areas (Apple et al., 1969; Iwatsuki et al., 1995, Wünschmann et al., 2000;
Schobesberger et al., 2005). At this, CD4+ T cells are the primarily affected cell type and
the first population that gets depleted (Iwatsuki et al., 1995; Wünschmann et al., 2000).
Lymphopenia is associated with a rapid loss of CD4+ T helper cells, CD8+ cytotoxic T
cells, CD21+ B cells, and macrophages (Wünschmann et al. 2000). Changes in
lymphoid organs correlate with the amount of CDV antigen (Wünschmann et al. 2000).
During the chronic disease phase, the virus is cleared from lymphoid organs and
reconstitution of lymphoid cells takes place (Krakowka et al. 1980). However, despite
regeneration and lymphoid repopulation, long lasting phenotypical alterations are
observable in lymphoid organs in chronically infected dogs (Wünschmann et al. 2000).
During chronic infection, viral antigen can be found in germinal centers within
leukocytes with DC-like morphology (Wünschmann et al. 2000). Thus, similar to the
proposed effects of MV upon human DCs, it is hypothesized that CDV also modulates
the function of canine DCs, which leads to persistent immune depressive effects
(Wünschmann et al. 2000; Beineke et al., 2009). For instance, CDV-infection of thymic
DCs may result in impaired maturation of T cells with release of CD5-negative T cells.
The CD5-negative T cell population potentially contains autoreactive lymphocytes,
which might be responsible for the induction of myelin specific immunity in canine
distemper (Wünschmann et al. 2000). However, so far, functional properties of DCs in

10
Chapter 2: General introduction
canine distemper have not been investigated. Mechanisms of CDV-induced
immunosuppression remain largely undetermined (Beineke et al. 2009). In human
measles, apoptosis of immune cells is considered as one cause for severe leukopenia
(Okada et al. 2000). Similarly, apoptosis of infected and non-infected immune cells in
lymphoid organs contribute to lymphoid depletion and impaired immune responses in
CDV-infected dogs (Okada et al. 2000; Schobesberger et al. 2005). This indicates that
directly virus-mediated as well as virus-independent mechanisms might be involved in
leukocyte apoptosis. Proposed mechanisms for apoptotic cell death include an over-
activation of immune responses and activation of Fas-pathways, respectively
(Schobesberger et al. 2005; Beineke et al. 2009).
Figure 2. Effects of canine distemper virus (CDV) upon immune cells. CDV-infection of leukocytes is
supposed to cause an inhibition of plasma cell differentiation, reduced proliferation of lymphocytes,
increased lymphocyte apoptosis induction and diminished function of antigen presenting cells (APCs),
such as dendritic cells.
After viral elimination from the peripheral blood, decreased antigen presentation and
lymphocyte maturation is supposed to contribute to persistent immunosuppression

11
Chapter 2: General introduction
despite repopulation of lymphoid organs (Beineke et al. 2009; Carvalho et al. 2012).
CDV causes modulation of antigen presenting abilities of monocytes by the inhibition of
IL-1 (Krakowka et al., 1987). Furthermore, CDV-N protein potentially modulates antigen
presentation by inhibiting the production of IL-12 in DCs, as described for MV-infected
DCs (Schneider-Schaulies and Dittmer, 2006), while morbillivirus V proteins act as
interferon antagonists and cytokine inhibitors (von Messling, 2006). Krakowka et al.,
(1982) discussed the existence of a mononuclear suppressor cell population that
causes long lasting immunosupression. Interestingly, regulatory T cells (Treg) have
been demonstrated to cause long lasting immunosuppression in infectious diseases,
such as human measles (Piccirillo and Shevach 2001; Sellin et al. 2009; Reuter et al.
2012). However, until now the function of Treg in the pathogenesis of canine distemper
has not been investigated yet. Similarly, the role of DCs in CDV-induced
immunopathology remains enigmatic.
Further studies have to focus on CDV-dependent and independent mechanisms of
immunosuppression as well as upon the role of DCs and Treg for immune alteration and
neuroinflammation in canine distemper
2.1.5 Neuropathology of canine distemper
Infection of the CNS represents the a serious complication of canine distemper, often
with poor prognosis (Carvalho et al. 2012). Dependent upon the host immune response
and virus strain, polioencephalitis and demyelinating leukoencephalitis (DL) can be
discriminated (Pearce-Kelling et al. 1990; Orlando et al. 2008). Polioencephalitis is a
rare manifestation of CDV-infection and can be subclassified as old dog encephalitis,
inclusion body encephalitis and post vaccinal encephalitis (Bestetti et al. 1978;
Vandevelde et al. 1980; Nesseler et al. 1999). Grey matter lesions are predominantly
located in cortical areas and brain stem nuclei with neurons and astrocytes representing
the most affected cell populations (Nesseler et al. 1999). Histologically neuronal
degeneration and necrosis with gliosis, inclusion bodies and infiltration of macrophages

12
Chapter 2: General introduction
and lymphocytes can be found (Nesseler et al. 1997; Beineke et al. 2009).
Leukoencephalitis is the more common manifestation and shows a progressive disease
course. White matter lesions are subclassified as acute lesions, subacute lesions
without inflammation, subacute lesions with inflammation and chronic lesions with
inflammation (Alldinger et al. 1993; Wünschmann et al. 1999; Wünschmann et al. 2000;
Beineke et al. 2009). Demyelinating foci are located predominately in proximity to the
ventricles, cerebellar velum, cerebellar peduncles and optic tract (Summers and Appel,
1994). Acute DL is characterized by focal to multifocal or diffuse vacuolization of the
white matter which develops during the period of severe immunosuppression
(Wünschmann et al. 1999; Seehusen et al. 2007). Lesions consist of mild gliosis with
reactive astrocytes and few gemistocytes (Baumgärtner et al. 1989; Alldinger et al.
1993). Here, astrocytes represent the main target cells for CDV-infection (Alldinger et al.
2006; Seehusen et al. 2007; Carvalho et al. 2012). Spread of the virus among
astrocytes does not require infectious particles (Wyss-Fluehmann et al. 2010; Carvalho
et al. 2012). The usage of gap junctions of the astrocytic synapse-like network
represents a possible mechanism for CDV spread as described for herpes simplex virus
(Wyss-Fluehmann et al. 2010). Acute and subacute lesions without inflammation are
characterized by a lack of perivascular cuffing (Tipold et al. 1999; Wünschmann et al.
1999). The initial vacuolization might be caused by restricted infection of
oligodendrocytes. Experiments in vitro and in vivo revealed a down-regulation of myelin-
specific genes (Zurbriggen et al. 1998; Vandevelde and Zurbriggen, 2005). During the
subacute disease course astrocytic hypertrophy and hyperplasia (astrogliosis and
astrocytosis) with formation of gemistocytes and multinucleated astrocytes as well as
gitter cells can be observed, although mononuclear perivascular infiltrates are initially
still absent (Tipold et al. 1999; Seehusen et al. 2007; Seehusen and Baumgärtner,
2010). Subsequent inflammatory stages of DL coincide with recovery of the immune
system. CNS lesions are characterized by the presence of perivascular infiltrations of
lymphocytes, plasma cells and macrophages (Vandevelde et al. 1982). Chronic CNS
lesions are associated with prominent perivascular infiltrations (more than three layers

13
Chapter 2: General introduction
of monocytic inflammatory cells) and profound myelin loss (Vendevelde et al. 1982;
Beineke et al. 2009; Vandevelde and Zurbriggen, 2005). A correlation between
microglial activation and loss of myelin has been described (Stein et al. 2004). Virus-
activated microglia release myelinotoxic substances which leads to bystander
demyelination (Fig. 3; Alldinger et al. 1996; Vandevelde and Zurbriggen, 2005; Beineke
et al. 2009). Recent studies revealed the existence of oligodendrocytes in demyelinating
lesions, indicating that primary demyelination precedes the loss of myelin-forming cells
in DL (Schobesberger et al. 2002). Moreover, an increased apoptotic rate particularly in
the granular layer of the cerebellar grey matter in DL indicates the possibility of
demyelination as a secondary process following Wallerian degeneration or loss of
astrocytic support, respectively (Moro et al. 2003; Beineke et al. 2009; Del Puerto et al.
2010; Seehusen and Baumgärtner, 2010).
Figure 3. Possible mechanisms of demyelination in canine distemper. A) During early infection
demyelination is a consequence of direct and indirect effects (e.g. loss of trophic support) upon
oligodendrocyte which causes myelin alteration and loss. B) During advanced stages immune mediated
processes including the release of myelinotoxic substances by activated microglia (bystander
demyelination) contribute to progressive myelin damage. Modified from Carvalho et al. (2012).

14
Chapter 2: General introduction
2.1.6 Immune responses in canine distemper demyelinating leukoencephalitis
The host defense during CDV-infection initially relies on innate immune responses,
although for complete virus elimination the activation of humoral and cellular immune
responses are required (Carvalho et al. 2012). Protective humoral immunity in canine
distemper is achieved by the production of antibodies against viral nucleoproteins,
followed by the development of specific immunoglobulins against viral envelope proteins
(Miele and Krakowka, 1983; Rima et al. 1991).
Lesion development of DL represents a biphasic event with initial tissue damage directly
induced by the virus and subsequent immune mediated inflammation as a consequence
of viral persistence and delayed type hypersensitivity (Baumgärtner et al. 1989;
Alldinger et al. 1996). The infiltration of CD8+ T cells correlates with virus replication and
the appearance of early immune responses against the N-protein (Tipold et al. 1999).
These T cells contribute to viral clearance but also to initial tissue damage by antibody
independent cytotoxicity (Wünschmann et al. 1999). In subacute lesions prominent
numbers of CD4+ T cells and B cells can be found. While CD8+ T cells might function as
cytotoxic effectors, CD4+ T cells are supposed to contribute to delayed type
hypersensitivity reactions in advanced lesions (Wünschmann et al. 1999). Additionally,
major histocompatibility complex class II (MHC II) is upregulated within areas with low
or absent viral antigen in chronic foci, which indicates virus-independent, immune
mediated mechanisms of demyelination (Alldinger et al. 1996). Thus, while during the
acute phase myelin damage is ascribed as directly virus mediated, demyelination in
chronic lesions is a result of collateral damage, e.g. due to an over-activation of
microglia/macrophages (bystander demyelination; Beineke et al. 2009) (Fig 3).
Repopulation of peripheral lymphoid organs is supposed to be a prerequisite for CNS-
infiltration of immune cells during inflammatory stages (Wünschmann et al. 2000). For
instance, with recovery of the immune system an infiltration of CD4+ T cells, B cells and
IgG-producing plasma cells in the brain as well as CDV-specific humoral immune
responses in the cerebrospinal fluid can be observed (Vandevelde et al. 1982; Beineke

15
Chapter 2: General introduction
et al. 2009). The increased antibody production by plasma cells might enhance
demyelination by an antibody dependent T cell mediated cytotoxicity (Alldinger et al.
1996; Wünschmann et al. 1999).
2.1.7 Cytokine expression in canine distemper
Cytokines are important signalling molecules involved in cell communication and
orchestration of immune responses in infectious and immune mediated disorders
(Rothwell 1997). In DL, infiltration of immune cells and accompanied demyelination is
followed by a tremendous up-regulation of several cytokines (Spitzbarth et al. 2012).
Cytokine expression can be directly induced by the virus or by autocrine and paracrine
regulatory loops during CDV-infection (Gröne et al. 2002; Markus et al. 2002; Beineke
et al. 2009).
During early DL lesion development pro-inflammatory cytokines, such as IL-6, IL-12 and
TNF-α, are up-regulated, while the anti-inflammatory cytokines, IL-10 and TGF-β,
remain unchanged (Markus et al. 2002; Beineke et al. 2008). The pro-inflammatory
cytokine environment in the brain during acute CDV-infection is indicative of insufficient
counter regulatory mechanisms, potentially causing early immune over-activation and
initial tissue damage in the brain. Similarly, expression of neuroprotective and Treg
inhibitory cytokines such as IL-10 and TGF-β is insufficient in canine spinal cord injury,
potentially leading to an activation of CNS resident immune cells (Spitzbarth et al.
2011). Initial infiltration of CD8+ T cells in the brain is associated with the expression of
chemoattractant cytokines such as IL-8 (Gröne et al. 1998; Tipold et al. 1999). In
advanced stages of DL, production of IL-12 within the CDV-infected CNS might trigger
Th1-biased immune responses (Gröne et al. 2000; Wünschmann et al. 2000; Beineke et
al. 2009; Spitzbarth et al. 2012). In addition, IL-12 is known to play a role in
demyelinating diseases such as MS and experimental autoimmune encephalomyelitis
(EAE). IL-12 also accounts for the maturation of monocyte-derived dendritic cells
(moDCs) and probably the activation of microglia (Fox and Rostami, 2000; Sugiura et

16
Chapter 2: General introduction
al. 2010; Spitzbarth et al. 2012). In the cerebrospinal fluid of CDV-infected dogs,
independent of the disease course, pro-inflammatory cytokines (TNF and IL-6) and anti-
inflammatory cytokines (IL-10 and TGF-β) are often observed simultaneously (Frisk et
al. 1999; Beineke et al. 2009).
Cytokine analysis of whole blood samples of dogs with DL revealed an expression of IL-
2, IL-6, TNF and TGF-β. IL-6 can be detected in the blood of dogs with early CNS
lesions, while TGF-β is found predominately during late stages of the disease, indicative
of a delayed onset of peripheral immunomodulatory processes in advanced disease
stages (Gröne et al. 1998). IL-1, IL-6 and TNF affect the permeability of the blood brain
barrier and represent a prerequisite for infiltration of leukocytes and enhancement of
neuroinflammation (Gröne et al. 1998; Beineke et al. 2008; Beineke et al. 2009). The
lack of IFN-γ expression in peripheral blood leukocytes of CDV-infected dogs might
account for an inadequate antiviral immunity in affected dogs (Gröne et al. 1998).
Similarly, in experimental CDV-infection of ferrets, early infection (3 days post infection)
is characterized by a lack of significant cytokine responses, probably as a consequence
of virus-mediated immunosuppression (Svitek and von Messling, 2007). Interestingly, in
the same animal model, as reported in MV-infected children, with disease progression a
switch from Th1 to Th2 cytokine responses was observed (Svitek and von Messling,
2007). Referring to this, prolonged IL-10 expression of peripheral leukocytes is
supposed to cause long lasting immune alterations in measles patients (Svitek and von
Messling, 2007). However, so far, cytokine responses in lymphoid organs of CDV-
infected dogs have not been investigated.
2.2 Dendritic cells
The term DCs referred to a heterogeneous group of multifunctional leukocytes (Bodey
et al. 1997). They represent the most potent antigen-presenting cell (APC) population.
So far, no other functions of DCs then antigen presentation and regulation of immune
responses are known (Steinman, 2007). They serve as sentinels of the immune system

17
Chapter 2: General introduction
and initiate immune responses (Banchereau et al. 2000). DCs exist in at least two
states, the immature and mature stage (Banchereau et al. 2000). They are found in
various tissues, such as skin, lymphoid organs, airways and intestinal tract. The cells
are able to capture and process antigens, migrate to T cell areas of lymphoid organs
and present antigens on their cell surface via MHC molecules. The antigen uptake by
DCs is conducted by receptor-mediated endocytosis and phagocytosis using different
receptors such as Fc-receptors, lectin receptors, macrophage mannose receptors,
ICAM-3, and toll-like receptors (Bhardwaj, 2003). Following antigen uptake, DCs
undergo a process of maturation and migrate to the draining lymph node. After
recognition of the antigen by T cells, Th1 or Th2 immune responses are initiated. The
polarization of T cells depends on co-stimulatory molecules and cytokine expression of
the DC (Macatonia et al. 1995, Bhardwaj, 2003). DCs have a unique ability to present
antigens from non-replicating viruses to CD8+ T cells (Smed-Sörensen et al. 2012). In
comparison of other APCs (e.g. macrophages or B cells), DCs have a 1000 fold higher
efficiency to activate resting T cells, which demonstrates the pivotal role of DCs for the
initiation of adaptive immune responses against infectious agents (Klagge and
Schneider-Schaulies, 1999; Bhardwaj, 2003). In addition to antigen recognition by T cell
receptors, interaction between CD28 on the T cell surface with co-stimulatory molecules
on DCs is required for optimal T cell activation (Björck et al. 1997; Weis and Wardrop,
2010). Although all DCs share a common ability to process and induce immune
responses indirectly by activating T cells, they differ by the expression of surface
markers, localization and cytokine production (Wu and Liu, 2007). A deregulation of DC
function is involved in immune-mediated tissue damage and immunosuppression in
human and veterinary medicine such as histocytic tumours, leischmaniasis, atopic
dermatitis, inflammatory bowel disease (Vanloubbeeck et al. 2003; Moore, 2008;
Cerquetella et al. 2010; Ricklin et al. 2010; Silva et al. 2012).

18
Chapter 2: General introduction
2.2.1 Dendritic cells in dogs
Similar to other species, canine DCs generated in vitro are non-adherent cells with
characteristic cytoplasmic projections (dendrites). They form clusters in culture and
have the ability to induce mixed leukocyte reactions (Goodel et al., 1985). So far, two
subsets of DCs, myeloid and lymphoid lineage DCs (Fig.4), have been recognized in
dogs. Myeloid DCs express MHC II, CD34 and CD14 and derive from monocyte and
bone marrow cells, while lymphoid DCs are MHC II+, CD34+ and CD14- (Tizard, 2009).
Canine DCs can be generated in vitro from CD14+ peripheral blood mononuclear cells
(PBMC) and the bone marrow by separation of CD34+ progenitor cells. As in humans
and mice, differentiation of precursor cells into DCs is induced by stimulation with
different cytokines, such as recombinant GM-CSF and IL-4 (Hägglund et al. 2000;
Ibisch et al. 2005; Bonnefont-Rebeix et al. 2006; Wijewardana et al. 2006; Wang et al.
2007a,b; Bund et al. 2010; Sugiura et al. 2010; Mielcarek et al. 2011; Fitting et al.
2011).
Canine DCs show an abundant formation of the Golgi apparatus and endoplasmic
reticulum but lack large lysosomal organelles (Ibisch et al. 2005). A unique
ultrastructural feature of canine DCs is the presence of periodical microstructures in the
cytoplasm (Ibisch et al. 2005; Isotani et al. 2006). Additionally, in contrast to human and
mouse Langerhans cells canine Langerhans cells lack classical Birbeck granules
(Moore et al. 1996). Canine moDCs express CD14 which contrast with human and
murine moDCs (moDCs; Ibisch et al. 2005; Wijewardana et al. 2006; Ricklin Gutzwiller
et al. 2010). CD14 expression of bone marrow-derived DCs (bmDCs) is under debate
(Hägglund et al. 2000; Weber et al. 2003; Ricklin Gutzwiller et al. 2010).
Compared to monocytes and macrophages canine moDCs and bmDCs show high
expression levels of MHC II, CD1a, and CD40, as well as of the co-stimulatory
molecules CD80 and CD86 (Ibisch et al. 2005; Bonnefont-Rebeix et al. 2006; Wang et
al. 2007a; Ricklin Gutzwiller et al. 2010; Sugiura et al. 2010). Phenotypical analyses
also enable the discrimination between canine bmDCs and moDCs (Ricklin Gutzwiller et
al. 2010).

19
Chapter 2: General introduction
Figure 4. Dendritic cell (DC) lineages of dogs. Two DC lineages have been identified in dogs: the
myeloid and lymphoid lineage. The myeloid lineage consists of three different subsets which differ in
localization, function and phenotype (Langerhans cells, bone marrow derived DCs and monocyte derived
DCs). Plasmacytoid DCs originate from the lymphoid lineage.
Canine Langerhans cells express CD1c, CD11c, CD80, MHC II and E-cadherin in situ,
which parallels the surface molecule expression pattern of human cells. However, in
contrast to humans, canine Langerhans cells lack S100, ATPase and ICAM-1 (Moore et
al. 1996; Zaba et al. 2009; Baines et al. 2008; Ricklin-Gutzwiller et al. 2010). Different
subsets of canine DCs exhibit differences in the ability to induce mixed leukocyte
reactions and cytokine expression (Andrea et al. 1995; Syme et al. 2005; Wang et al.
2007b; Ricklin Gutzwiller et al. 2010).
Understanding species-specific properties of canine DCs is pivotal for future studies
upon the role of this cell type in infectious disorders.

20
Chapter 2: General introduction
2.2.2 Dendritic cells in viral diseases
The immune response to viruses is a complex interplay between the pathogen and
innate and adaptive immune responses which aims to eradicate the infectious agent
with minimal damage to the host (Lambotin et al. 2010). The interaction between
different viruses and DCs includes the alteration of DC functions, such as endocytosis,
vesicle trafficking, immunological synapse formation, apoptosis induction and cytokine
production (Harman et al. 2006; Cunningham et al. 2010). Virus infection of DCs can
lead to a productive infection and subsequent release of infectious particles, as
observed for human immunodeficiency virus (HIV), MV, Epstein-Barr virus, and human
cytomegalovirus (Li et al. 2002; Beck et al. 2003; Donaghy et al. 2003; Schneider-
Schaulies et al. 2003; Rinaldo and Piazza, 2004). Alternatively, virus particles can be
transferred from DCs directly to other cell types, as observed for HIV infection (Rinaldo
and Piazza, 2004). As demonstrated in murine models for Sendai virus-, Moloney
leukemia virus-, herpes simplex virus-, and influenza virus-infection, DC-mediated
priming of the immune response leads to viral elimination (Kast et al. 1998; Hengel et al.
1987; Nonacs et al. 1992; Klagge and Schneider-Schaulies, 1999). For instance, in
influenza virus infection, virus antigen can be found on all APCs, but only DCs are able
to induce effective immune responses (Hamilton-Easton and Eichelberger, 1995;
Klagge and Schneider-Schaulies, 1999). In contrast to macrophages, influenza virus-
infected DCs do not undergo rapid cell death. Interestingly, influenza virus-infected
monocytes are unable to differentiate into DCs, which leads to the assumption that
virus-mediated DC inhibition might account for an impairment of virus-specific immunity
(Boliar and Chambers, 2010). In addition, several other viruses that cause persistent
infection, such as human cytomegalovirus, murine cytomegalovirus and Epstein-Barr
virus are able to manipulate DCs, which leads to inadequate protective immune
responses (Rinaldo and Piazza, 2004).
Herpes simplex virus infection of DCs leads to productive infection with down-regulation
of the co-stimulatory molecules CD80, CD86 and CD40, but not of MHC I and MHC II,

21
Chapter 2: General introduction
which suggests that herpes simplex virus proteins target signal transduction pathways
that control the expression of co-stimulatory molecules (Mikloska et al. 2001). HIV has
been demonstrated to interact with DCs and modulate their function, which represents a
prototypical model of DC-virus interaction (Rinaldo and Piazza, 2004). DCs are
supposed to be the early targets of HIV and by the ability to cluster T cells they can
spread the virus within the host (Klagge and Schneider-Schaulies, 1999). In addition, in
advanced stages of HIV infection, DCs become the virus reservoir and therefore
contribute to virus persistence. Once the DCs incorporate the HIV, they transport the
virus to the draining lymph node to induce an immune response (Klagge and Schneider-
Schaulies, 1999). Subsequently, DCs undergo a killing process due to feedback
mechanisms that remove APCs after stimulation of T cell responses. Interestingly, only
immature DCs can get infected, while mature DCs do not support the replication of HIV
(Knight et al. 1997; Klagge and Schneider-Schaulies, 1999).
Previously it was shown that during MV infection epithelial cells of the upper respiratory
tract are the first cells to be infected (Esolen et al. 1993). Since epithelial cells express
only CD46, which is a receptor only for attenuated MV-strains, and lymphocytes, which
are one of the most affected cells in measles, are not present in large numbers in
respiratory epithelium, it was concluded that other cells might account for early MV entry
(Tatsuo et al. 2000; de Swart et al. 2007; de Witte et al. 2008). Moreover,
undifferentiated monocytes, which express CD46, are relatively resistant to MV
replication (Fugier-Vivier et al. 1997). CD150, which is necessary for virus entry, is
expressed in T cells, B cells, macrophages, and DCs (de Swart et al. 2007). In vivo
infection of DCs has been described in animal experiments using cotton rats, transgenic
mice and macaques. (de Swart et al. 2007) Thus, infection of DCs of the respiratory
tract and subsequent migration of these cells to draining lymph nodes is supposed to
contribute to virus spread in the organism. Furthermore, dysfunction of DCs following
MV infection is supposed to represent one of the factors for long lasting and profound
immune suppression in measles patients.(Servet-Delprat et al. 2000) However, DC

22
Chapter 2: General introduction
infection in human patients has not been confirmed until now (Hahm et al. 2005; de
Swart et al. 2007; Griffin, 2010).
Different studies have demonstrated MV infection of different subtypes of myeloid DCs
in vitro (Fugier-Vivier et al. 1997; Murabayashi et al. 2002; Ohgimoto et al. 2007). Here,
the increased susceptibility of mature DCs for MV infection is in part a consequence of
higher CD150 expression levels (Klagge et al. 2004). The H protein of MV determines
the tropism for moDCs. The induction of syncytia formation of infected DCs is a
characteristic feature of MV wild type strains (Fugier-Vivier et al. 1997; Murabayashi et
al. 2002; Griffin 2010), while vaccine strains can indeed infect and replicate in DCs,
although only small amounts of infectious virus are produced due to an instability of the
M protein (Ohgimoto et al. 2007; Griffin 2010). Interference of MV with APCs represents
an important cause for immunosuppression. During MV infection of cultured immature
DCs, infected and non-infected cell undergo a maturation process (Zilliox et al. 2006).
This maturation is associated with an up-regulation of CD40, CD80, CD86 and MHC II,
while CD1a and CD34 are down-regulated (Fig. 5) (Schnorr et al. 1997; Servet-Delprat
et al., 2000; Zilliox et al. 2006).
The infection also results in rapid production of type I interferon (IFN) which also
contributes to DC maturation, but without the ability to prevent viral spread (Schneider-
Schaulies et al. 2002;Schneider-Schaulies and Meulen, 2002; Zilliox et al. 2006).
Moreover, in murine models, MV infection impairs the differentiation of DCs in vivo,
characterized by a down-regulation of co-stimulatory molecules, MHC class I and MHC
II (Oldstone et al. 1999; Hahm et al. 2005; Trifilo et al. 2006). In addition to the
modulation of antigen presenting function of DCs, MV infection suppresses the
production of IL-12 (Fugier-Vivier et al. 1997). Impairment of IL-12 expression in MV-
infected DCs coincides with a high percentage of apoptotic DCs and inhibition of CD40
signalling (Fugier-Vivier et al. 1997; Servet-Delprat et al. 2000). Inhibition of IL-12
secretion can be observed with disease progression predominantly in the late stage of
the disease, which might lead to insufficient Th1 immune responses in measles patients
(Schneider-Schaulies et al. 2002)

23
Chapter 2: General introduction
Figure 5. Proposed mechanisms of interaction between measles virus (MV) and dendritic cells
(DC). MV induces maturation of DCs, characterized by an up-regulation of co-stimulatory molecules
(CD80/86), CD40 and MHC II, while impairing the production of IL-12 and T cell stimulatory ability.
Modified from Kerdiles et al. (2006).
2.3 Regulatory T cells
Immune homeostasis is mainly regulated by Treg (Vignali et al. 2009). Treg are
essential for maintaining immune tolerance and therefore prevent autoimmune diseases
and limit chronic inflammatory processes (Vignali et al. 2009). Expression of the
forkhead-winged helix transcription factor Foxp3 regulates the transcription of genes
involved in immune modulation and represents a Treg-specific marker molecule
(Brunkow et al. 2001; Hori et al. 2003; Biller et al. 2007; Feuerer et al. 2009).
Additionally Treg are characterized by the expression of CD25, CTLA-4, and GITR
(Zheng and Rudensky, 2007). Foxp3+ Treg use the αβ T cell antigen receptor (TCR) for
antigen recognition and have a broad TCR repertoire (Feuerer et al. 2009; Relland et al.

24
Chapter 2: General introduction
2012). So far, two main sources of Tregs have been described: thymic Foxp3+ Treg
generated in the thymus (natural Treg) and Treg which are induced in the periphery by
different T cell derived factors (adaptive Treg; Mills 2004; Feuerer et al. 2009; Miyara
and Sakaguchi, 2007). For example, adaptive Tregs can originate from CD4+ effector T
cells due to IL-2 and TGF-β stimulation. These T cells, although unstable, show an
expression of Foxp3 and immunosuppressive properties (Chen et al. 2003; Fantini et al.
2004; Floess et al. 2007; Feuerer et al. 2010). Immature and mature DCs have the
ability to induce proliferation of Treg in vitro and in vivo (Yamazaki et al. 2007). In the
CNS, activated microglia and DCs have the ability to attract thymic Treg via the
production of CCL22, which interacts with CCR4 on Treg (Vulcano et al. 2001; Kipnis et
al. 2004).
Treg can also be induced under inflammatory conditions by astrocytes and neurons
(Lowther and Hafler, 2012). A novel population of natural Treg has recently been
identified in the peripheral blood of human beings. These cells express CD4 or CD8 but
lack Foxp3-expression (Feger et al. 2007; Zozulya and Wiendl, 2008).
Several mechanisms are involved in Treg-mediated immunosuppression, which can be
grouped as suppression by cytokines, suppression by cytolysis, suppression by
metabolic disruption, and suppression by the modulation of DC function (Fig. 6) (Vignali
et al. 2009; Miyara and Sakaguchi, 2007). Treg can inhibit the proliferation and function
of Natual Killer T cells, CD4+ T cells, CD8+ T cells, and B cells, as well as the maturation
and antigen presenting capacity of DCs (Piccirillo and Shevach, 2001; Azuma et al.
2003; Misra et al. 2004; Lim et al. 2005). A major function of Treg is to respond to
signals associated with tissue destruction and to minimize collateral tissue damage
(Belkaid and Rouse, 2005). Treg are involved in gastrointestinal immune homeostasis,
as demonstrated in mouse colitis models (Belkaid and Rouse, 2005). Similar beneficial
effects have been observed in mouse models of Leishmania major infection (Aseffa et
al. 2002; Liu et al. 2003; Belkaid and Rouse, 2005; Rai et al. 2012). Here, the disease
severity is enhanced in the absence of Treg, while application of CD4+CD25+ Treg
reverses pathological lesions (Liu et al. 2003; Rai et al. 2012)

25
Chapter 2: General introduction
Figure 6. Schematic diagram of mechanisms involved in regulatory T cell (Treg)-mediated
immunosuppression. Treg-mediated mechanisms of immunosuppression can be grouped in the
following categories: (i) suppression of immune responses by inhibitory cytokines (IL-10, IL-35 and TGF-
β); (ii) cytolysis by the release of granzymes (Grz); (iii) metabolic disruption by cytokine deprivation via IL-
2 receptor α (CD25) with subsequent lymphocyte apoptosis, cyclic AMP-mediated inhibition, or
CD39/CD73 and adenosine receptor (A2A)-mediated immunosuppression; (iv) modulation of dendritic
cells (DC) by down-regulation of MHC II and CD80/86 which leads to a reduced antigen presenting
capacity as well as via cytotoxic T lymphocyte antigen-4 (CTLA4)–CD80/CD86-mediated induction of
indoleamine 2,3-dioxygenase (IDO) which is a potent immunosuppressive molecule. Modified from
Vignali et al. (2008) and Miyara and Sakaguchi, (2007).
26
Chapter 2: General introduction
2.3.1 Regulatory T cells in viral diseases
Immunologic self tolerance is critical for the prevention of autoimmunity and
maintenance of immune homeostasis in the CNS (Valencia et al. 2006). However, the
role of Treg during different infectious diseases remains dubious, since both beneficial
(reduction of immune mediated tissue damage) and detrimental effects (reduction of
protective immune responses) of Treg have been described in infectious disorders (Fig.
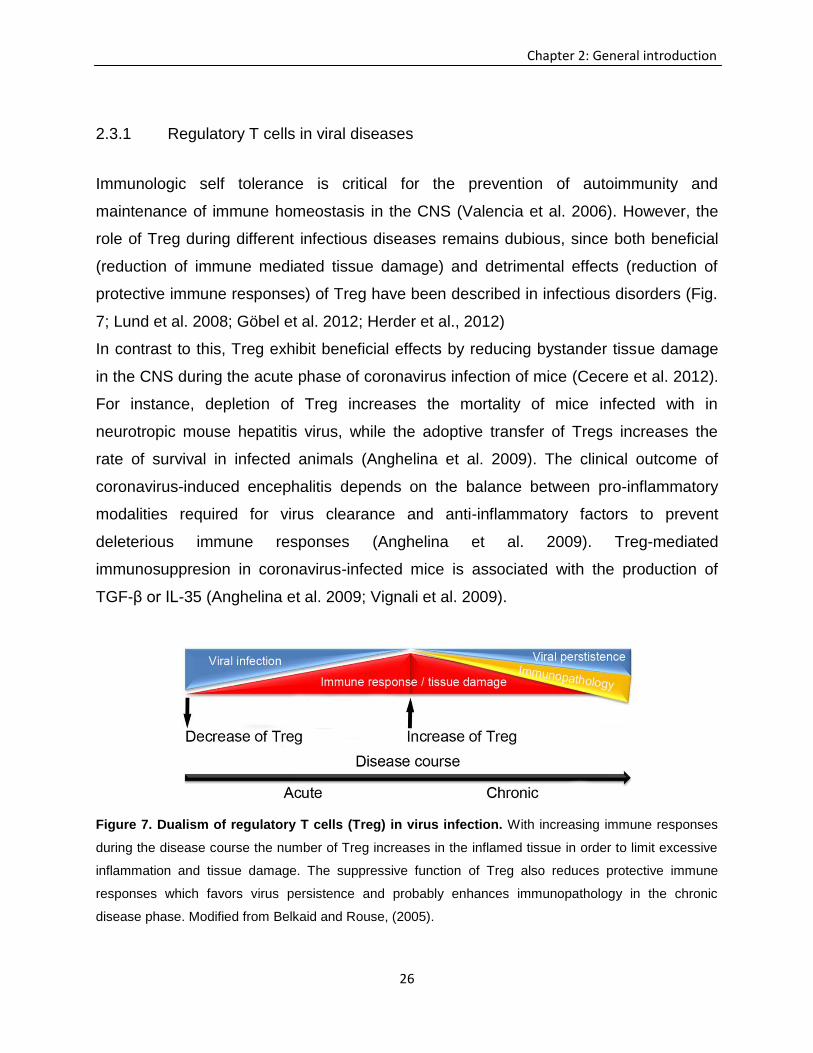
7; Lund et al. 2008; Göbel et al. 2012; Herder et al., 2012)
In contrast to this, Treg exhibit beneficial effects by reducing bystander tissue damage
in the CNS during the acute phase of coronavirus infection of mice (Cecere et al. 2012).
For instance, depletion of Treg increases the mortality of mice infected with in
neurotropic mouse hepatitis virus, while the adoptive transfer of Tregs increases the
rate of survival in infected animals (Anghelina et al. 2009). The clinical outcome of
coronavirus-induced encephalitis depends on the balance between pro-inflammatory
modalities required for virus clearance and anti-inflammatory factors to prevent
deleterious immune responses (Anghelina et al. 2009). Treg-mediated
immunosuppresion in coronavirus-infected mice is associated with the production of
TGF-β or IL-35 (Anghelina et al. 2009; Vignali et al. 2009).
Figure 7. Dualism of regulatory T cells (Treg) in virus infection. With increasing immune responses
during the disease course the number of Treg increases in the inflamed tissue in order to limit excessive
inflammation and tissue damage. The suppressive function of Treg also reduces protective immune
responses which favors virus persistence and probably enhances immunopathology in the chronic
disease phase. Modified from Belkaid and Rouse, (2005).

27
Chapter 2: General introduction
In persistent viral infection (e.g. HIV), where an equilibrium between viral proliferation
and the immune response is established, viral removal becomes difficult, which leads to
life threatening diseases (Dittmer et al. 2004). Induction and/or expansion of Treg cells
by viruses is a highly efficient strategy to prevent effector T cell activation (Mills, 2004;
Schneider-Schaulies and Dittmer, 2006). The role of Treg in MV infection is under
debate, since differing findings have observed in animal experiments and human
patients (Yu et al. 2008; Li et al. 2008; Sellin et al. 2009). Probably the interplay
between the immunoregulatory and effector response during MV infection could be
critical for pathogenesis, and the adequate balance between these two arms of
immunity may play an essential role for the disease outcome (Sellin et al. 2009).

28

29
Chapter 3: Regulatory T cells in CDV infection
3 Chapter 3: Regulatory T cells in canine distemper
virus infection
Depletion of Foxp3+ regulatory T cells as a putative prerequisite for
lesion initiation in canine distemper virus induced demyelinating
leukoencephalitis
Visar Qeska1,2*, Yvonne Barthel1*, Maximilian Iseringhausen1*, Andrea Tipold2,3,
Veronika M. Stein3, Wolfgang Baumgärtner1,2, Andreas Beineke1,
1Department of Pathology, University of Veterinary Medicine Hannover, Bünteweg 17,
D-30559 Hannover, Germany
2Center for Systems Neuroscience, Hannover, Germany
3Department of Small Animal Medicine and Surgery, University of Veterinary Medicine
Hannover, Bünteweg 9, D-30559 Hannover, Germany.
*Authors have contributed equally
Corresponding author:
Prof. Dr. Andreas Beineke
Department of Pathology, University of Veterinary Medicine Hannover
Vet. Res. 2013, submitted

30
Chapter 3: Regulatory T cells in CDV infection
Abstract
Canine distemper virus (CDV) infection causes demyelinating leukoencephalitis in dogs,
sharing similarities with human myelin disorders and is therefore appreciated as a
translational animal model for multiple sclerosis (MS). In viral neurological diseases, an
ambiguous function of regulatory T cells (Treg), with both beneficial effects by reducing
immunopathology and detrimental effects by inhibiting antiviral immunity, has been
described. However, the role of Treg in the pathogenesis of canine distemper has not
been investigated yet. The aim of the present study was to testify the hypothesis that
peripheral lymphoid depletion influences immunomodulatory mechanisms in the brain of
CDV-infected dogs. Immunohistochemistry revealed a lack of Foxp3+ Treg in
predemyelinating and early demyelinating lesions which was associated with the
accumulation of CD3+ T cells, L1+ macrophages/microglia and GFAP+ astrocytes.
Together with CD79α+ B cells, a delayed infiltration of Foxp3+ Treg was observed in
chronic demyelinating lesions. Splenic depletion of Foxp3+ Treg was associated with an
increased mRNA-expression of tumor necrosis factor in the acute disease phase,
indicative of a pro-inflammatory microenvironment and lack of immunological counter
regulation in peripheral lymphoid organs. In conclusion, disturbed immune regulatory
mechanisms represent a potential cause for excessive neuroinflammation and early
lesion development in canine distemper leukoencephalitis, as discussed for immune
mediated myelin disorders such as MS.
Introduction
Distemper in dogs is caused by the canine distemper virus (CDV), a morbillivirus which
is closely related to the human measles virus [1–3]. The disease course and
pathogenesis of canine distemper are similar to human measles, including fever, rash,
respiratory signs, lymphopenia, and profound immunosuppression with generalized

31
Chapter 3: Regulatory T cells in CDV infection
depletion of lymphoid organs during the acute disease phase [4–6]. Central nervous
system (CNS) infection and neurological complications can be observed more
frequently in infected dogs compared to measles patients, usually affecting children [7–
9]. Depending on CDV strain, host immune status, and age, naturally infected dogs
develop demyelinating leukoencephalomyelitis, which shares similarities with human
myelin disorders, such as multiple sclerosis (MS) as well as measles virus associated
post-infectious encephalomyelitis and subacute sclerosing panencephalitis [2,10,11].
Regulatory T cells (Treg), characterized by expression of the transcription factor
forkhead box P3 (Foxp3), play a key role in the maintenance of immunological tolerance
and therefore prevent autoimmune CNS disease [12–17]. However, in infectious CNS
diseases Treg exhibit both beneficial effects by reducing immune mediated tissue
damage and detrimental effects due to their immunosuppressive properties, causing
disease exacerbation or persistence, respectively [18,19]. For instance, Treg reduce
antiviral immunity in experimental Theiler’s murine encephalomyelitis [20,21], a rodent
model for demyelinating disorders as well as in Friend retrovirus mouse model [22] and
experimental herpesvirus infection of mice [23,24]. However, the impact of Treg upon
morbillivirus-induced immunological alterations during early infection and CNS
manifestation remains enigmatic [25], since reports that Treg are increased in measles
patients [26,27] have been contradicted by others [28]. Moreover, different rodent
models for measles virus infection came to ambiguous conclusions regarding Treg-
related effects upon immune responses, probably attributed to disease course-
dependant effects or mouse strain-specific responses to virus infection [5,29–31]. Thus,
in addition to rodent models, there is an increasing interest in spontaneous and
experimental canine diseases as translational large animal models for human CNS
disorders [32–34].
Demyelination in canine distemper represents a biphasic process with directly virus
induced neurodegeneration, microglial activation and CD8-mediated cytotoxicity during
the early phase [35–37]. In comparison, during the chronic phase, reconstitution of

32
Chapter 3: Regulatory T cells in CDV infection
peripheral lymphoid organs facilitates immune mediated mechanisms with delayed type
hypersensitivity and progressive myelin loss in the CNS of CDV-infected dogs [4,38,39].
A proinflammatory cytokine environment in the brain during acute CDV-infection is
indicative of insufficient counter regulatory mechanisms, potentially causing early
immune over-activation and initial tissue damage in the brain [40–43]. Similarly,
expression of neuroprotective and Treg-specific cytokines such as IL-10 and TGF-β is
insufficient in canine spinal cord injury, leading to an activation of CNS resident immune
cells [44]. Similar to the mechanisms in demyelinating leukoencephalomyelitis in CDV-
infection, an early stimulation of microglia and lack of immunoregulation is discussed as
a requirement for myelin damage in MS patients [45–48]. However, so far, the role of
immunomodulatory cells, especially Treg, in the pathogenesis of canine distemper has
not been investigated.
Since the initiation of inflammation in myelin disorders is influenced by an
immunological imbalance of the peripheral immune system [49–51], the aim of the
present study was to determine disease phase-dependant phenotypical changes and
cytokine expression in lymphoid organs in canine distemper. Special emphasis was
given to testify the hypothesis that peripheral depletion of Treg causes a lack of CNS-
infiltrating immunomodulatory cells in the predemyelinating phase of CDV infection,
which has the ability to enhance early neuroinflammation and represents a potential
prerequisite for immune mediated demyelination.
Materials and Methods
Animals and tissue selection
A total of 23 dogs of different breeds and age with spontaneous CDV-infection and five
control animals were selected for this study (Table 1). Animals were clinically examined
at the Small Animal Clinic of the University of Veterinary Medicine Hannover (Germany)
and sacrificed by by an overdose of pentobarbital (100 mg/kg intravenously) on the

33
Chapter 3: Regulatory T cells in CDV infection
owner’s request due to worsening of clinical signs related to CDV-infection, such as
neurological (seizures), respiratory (coughing, sneezing) and intestinal signs (diarrhea),
respectively, and poor prognosis. Infected dogs which died spontaneously as a
consequence of systemic distemper, showed seizures prior to death and were directly
submitted to necropsy by pet owners (animals 16, 17, 21, 27; Table 1). Main
pathological findings in the brain and extra-neuronal tissues of affected animals are
listed in table 1. All dogs were examined for research purposes at the Department of
Pathology of the University of Veterinary Medicine Hannover (Germany) with the
permission by the owners. Five non-infected healthy animals (beagles) without
neurological signs were obtained from an animal experiment performed at the Institute
for Parasitology of the University of Veterinary Medicine Hannover, which was approved
and authorized by the local authorities (Niedersächsisches Landesamt für
Verbraucherschutz- und Lebensmittelsicherheit (LAVES), Oldenburg, Germany,
permission number 08A580) and used as controls (Table1).
Infected animals were grouped according to the most advanced and dominating brain
lesion (SI-SIII, see below). For morphological and phenotypical characterization,
paraffin embedded spleen tissue was available from all CDV-infected and control
animals. Out of these, immunohistochemical analyses of paraffin embedded brain tissue
were performed in 15 CDV-infected and 5 control dogs. For cytokine expression
analyses by reverse transcriptase-quantitative polymerase chain reaction (RT-qPCR),
frozen spleen tissue was available from 11 CDV-infected and five control dogs (Table
1).
Histology and phenotyping
Paraffin embedded tissue slices (4μm thickness) from the spleen and brain were
stained with hematoxylin and eosin (HE) for morphological examination. The brain
tissue was additionally stained with luxol fast blue (LFB) for detecting myelin loss and
myelinophagia (active demyelination). Antigen detection was performed by the avidin-

34
Chapter 3: Regulatory T cells in CDV infection
biotin-peroxidase complex method as previously described [35,52]. In brief, paraffin
embedded tissues were deparaffinized in Roticlear (Carl Roth GmbH, Karlsruhe,
Germany) and rehydrated through graded alcohols. Endogenous peroxidase activity
was suppressed with 0.5% H2O2 in methanol, followed by incubation with primary
antibody overnight at 4°C. Specificity controls included substitution of the respective
monoclonal antibody with ascitic fluid from nonimmunized BALB/cJ mice or rabbit
normal serum and in the case of the anti-Foxp3 antibody, a rat immunoglobulin isotype
control, was used. Spleen tissue of a healthy dog was used as positive control for the
detection of lymphoid cells. Except for the lectin BS-1, incubation with primary
antibodies was followed by incubation with biotinylated secondary antibodies for 30
minutes at room temperature. Subsequently, the avidin-biotin-peroxidase complex
(VECTASTAIN Elite ABC Kit; Vector Laboratories, PK 6100, Burlingame, CA) was
added and incubated for 30 minutes at room temperature. Antigen-antibody reactions
were visualized by incubation with 3,3’-diaminobenzidine-tetrahydrochloride-H2O2 in 0.1
mol/L imidazole, pH 7.1 for 5 minutes, followed by counterstaining with hematoxylin.
Histological evaluation and phenotypical characterization of white matter lesions
in the cerebellum
HE- and LFB-staining of white matter lesions of dogs suffering from CDV-infection and
of control dogs were evaluated morphologically by light microscopy. The cerebellar
white matter of 15 naturally infected dogs and five control dogs were examined and
lesions were classified as described by Wünschmann et al. (1999) with slight
modifications [37]. Briefly, groups were classified as acute non-inflammatory
encephalitis (SI), subacute non-inflammatory encephalitis (SII) and subacute to chronic
inflammatory encephalitis (SIII). Acute white matter lesions (SI) were characterized by
hypercellularity and vacuolization whereas SII and SIII lesions showed active
demyelination as demonstrated by decreased intralesional LFB-staining and the
presence of LFB+-myelinophages (gitter cells).
35
Chapter 3: Regulatory T cells in CDV infection
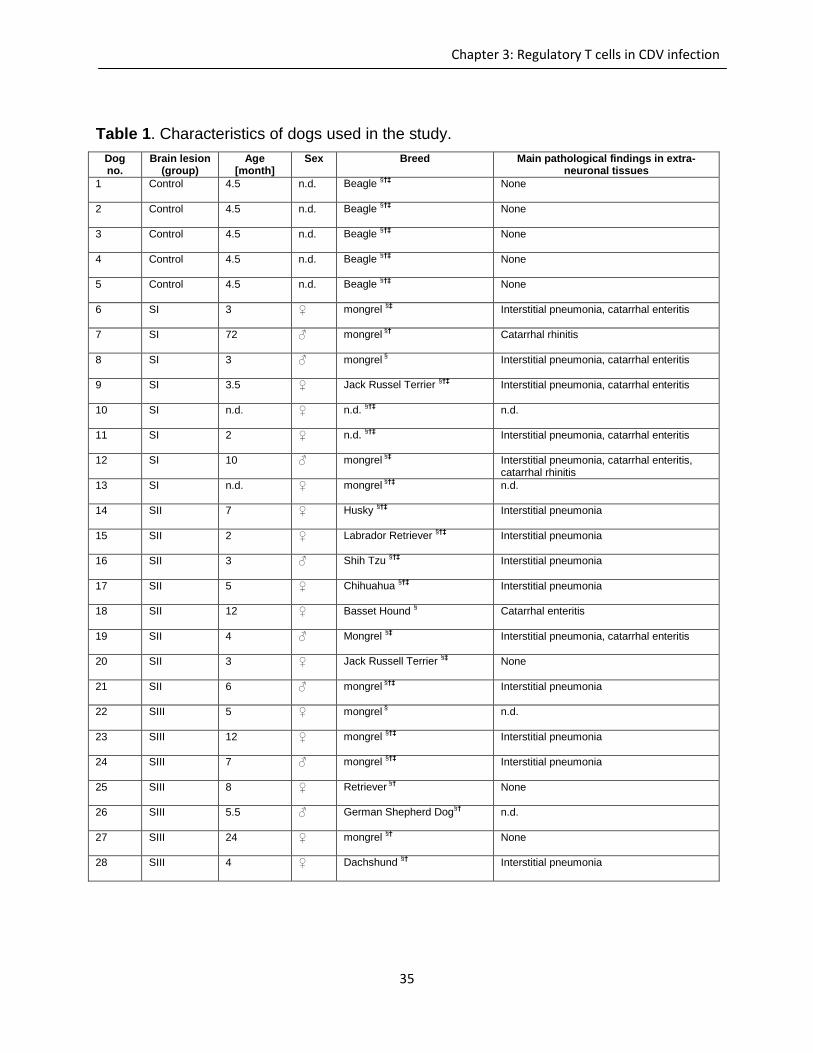
Table 1. Characteristics of dogs used in the study.
Dog no.
Brain lesion (group)
Age [month]
Sex Breed Main pathological findings in extra-neuronal tissues
1 Control
4.5 n.d. Beagle §†‡
None
2 Control 4.5
n.d. Beagle §†‡
None
3 Control 4.5
n.d. Beagle §†‡
None
4 Control 4.5
n.d. Beagle §†‡
None
5 Control 4.5
n.d. Beagle §†‡
None
6 SI
3 ♀ mongrel §‡
Interstitial pneumonia, catarrhal enteritis
7 SI 72
♂ mongrel §†
Catarrhal rhinitis
8 SI 3
♂ mongrel §
Interstitial pneumonia, catarrhal enteritis
9 SI 3.5
♀ Jack Russel Terrier §†‡
Interstitial pneumonia, catarrhal enteritis
10 SI n.d.
♀ n.d. §†‡
n.d.
11 SI 2
♀ n.d. §†‡
Interstitial pneumonia, catarrhal enteritis
12 SI
10 ♂ mongrel §‡
Interstitial pneumonia, catarrhal enteritis, catarrhal rhinitis
13 SI n.d.
♀ mongrel §†‡
n.d.
14 SII 7
♀ Husky §†‡
Interstitial pneumonia
15 SII 2
♀ Labrador Retriever §†‡
Interstitial pneumonia
16 SII 3
♂ Shih Tzu §†‡
Interstitial pneumonia
17 SII 5
♀ Chihuahua §†‡
Interstitial pneumonia
18 SII 12
♀ Basset Hound §
Catarrhal enteritis
19 SII 4
♂ Mongrel §‡
Interstitial pneumonia, catarrhal enteritis
20 SII 3
♀ Jack Russell Terrier §‡
None
21 SII 6
♂ mongrel §†‡
Interstitial pneumonia
22 SIII 5
♀ mongrel §
n.d.
23 SIII 12
♀ mongrel §†‡
Interstitial pneumonia
24 SIII 7
♂ mongrel §†‡
Interstitial pneumonia
25 SIII 8
♀ Retriever §†
None
26 SIII 5.5
♂ German Shepherd Dog§†
n.d.
27 SIII 24
♀ mongrel §†
None
28 SIII 4
♀ Dachshund §†
Interstitial pneumonia

36
Chapter 3: Regulatory T cells in CDV infection
n.d. = not determined; SI = acute non-inflammatory encephalitis; SII = subacute non-inflammatory encephalitis; SIII =
subacute to chronic inflammatory encephalitis; ♀ = female; ♂ = male; §paraffin embedded spleen tissue available;
†used for immunohistochemical analyses of brain tissue;
‡frozen spleen tissue available
While SII lesions were dominated by glial responses (microgliosis and astrogliosis)
without perivascular cuffing, SIII lesions displayed marked lymphohistiocytic infiltration
within the neuroparenchyma and perivascular spaces, indicative of an advanced
disease process. The control group (C) showed no histopathological alterations.
Immunohistochemistry and lectin histochemistry for quantifying inflammatory cell
infiltrations and glial responses in the brain were evaluated by using a morphometric
grid at a high power field resolution (number of cells/0.0625 mm²). The absolute number
of labeled cells was counted in white matter lesions (SI-SIII) and controls (C). Small
lesions were counted in total, otherwise a maximum of ten randomly distributed high
power fields within large lesions were counted [53].
Histological scoring of lymphoid depletion and characterization of
phenotypical changes in the splenic white pulp
The degree of lymphoid depletion in spleen sections of CDV-infected and control
animals, expressed as the cellularity score, was evaluated semi-quantitatively by light
microscopy as described by Wünschmann et al. (2000) [38]: 4 = normal architecture, 3
= mild, 2 = moderate, 1 = severe depletion. In a normal spleen (cellularity score 4; Fig.
S1A), the white pulp areas showed a regular architecture, with numerous primary and
secondary follicles. Mildly depleted spleens (cellularity score 3; Fig. S1B) were
characterized by a reduced size of primary follicles and loss of secondary follicles,
whereas marked reduction of the white pulp size and loss of secondary follicles were
indicative of moderate depletion (cellularity score 2; Fig. S1C). In severely depleted
spleens, the white pulp was no longer recognizable (cellularity score 1; Fig. S1D).
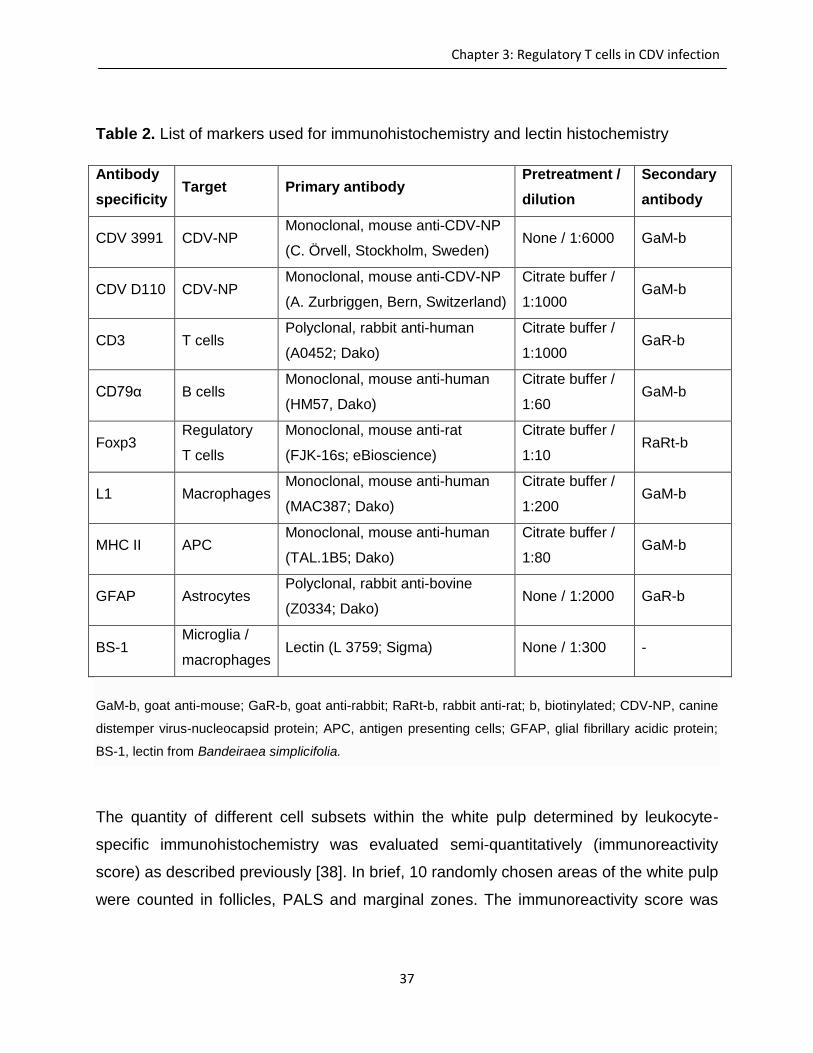
37
Chapter 3: Regulatory T cells in CDV infection
Table 2. List of markers used for immunohistochemistry and lectin histochemistry
Antibody
specificity Target Primary antibody
Pretreatment /
dilution
Secondary
antibody
CDV 3991 CDV-NP Monoclonal, mouse anti-CDV-NP
(C. Örvell, Stockholm, Sweden) None / 1:6000 GaM-b
CDV D110 CDV-NP Monoclonal, mouse anti-CDV-NP
(A. Zurbriggen, Bern, Switzerland)
Citrate buffer /
1:1000 GaM-b
CD3 T cells Polyclonal, rabbit anti-human
(A0452; Dako)
Citrate buffer /
1:1000 GaR-b
CD79α B cells Monoclonal, mouse anti-human
(HM57, Dako)
Citrate buffer /
1:60 GaM-b
Foxp3 Regulatory
T cells
Monoclonal, mouse anti-rat
(FJK-16s; eBioscience)
Citrate buffer /
1:10 RaRt-b
L1 Macrophages Monoclonal, mouse anti-human
(MAC387; Dako)
Citrate buffer /
1:200 GaM-b
MHC II APC Monoclonal, mouse anti-human
(TAL.1B5; Dako)
Citrate buffer /
1:80 GaM-b
GFAP Astrocytes Polyclonal, rabbit anti-bovine
(Z0334; Dako) None / 1:2000 GaR-b
BS-1 Microglia /
macrophages Lectin (L 3759; Sigma) None / 1:300 -
GaM-b, goat anti-mouse; GaR-b, goat anti-rabbit; RaRt-b, rabbit anti-rat; b, biotinylated; CDV-NP, canine
distemper virus-nucleocapsid protein; APC, antigen presenting cells; GFAP, glial fibrillary acidic protein;
BS-1, lectin from Bandeiraea simplicifolia.
The quantity of different cell subsets within the white pulp determined by leukocyte-
specific immunohistochemistry was evaluated semi-quantitatively (immunoreactivity
score) as described previously [38]. In brief, 10 randomly chosen areas of the white pulp
were counted in follicles, PALS and marginal zones. The immunoreactivity score was

38
Chapter 3: Regulatory T cells in CDV infection
determined as follows: 1 = 1-25% positive cells, 2 = 26-50% positive cells, 3 = 51-75%
positive cells, and 4 = 76-100% positive cells. The immunoreactivity index was
calculated by multiplying the cellularity score by the immunoreactivity score [38].
To determine virus load and distribution in splenic follicles, PALS and marginal zones by
CDV-specific immunohistochemistry, the following scale was used: 0 = no virus
detected, 1 = few infected cells, 2 = some infected cells, 3 = numerous infected cells; 4
= almost all cells infected [38].
Cytokine expression analysis by reverse transcriptase-quantitative
polymerase chain reaction
Primer design
Primers for the generation of standards and primers for measuring the quantity of
specific cytokines are listed in tables 2 and 3. PCR primer sequences for detecting
glyceraldehyde-3-phosphate dehydrogenase (GAPDH), elongation factor-1α (EF-1α),
hypoxanthine-guanine phosphoribosyltransferase (HPRT), tumor necrosis factor (TNF),
transforming growth factor-β (TGF-β), interleukin (IL)-2, IL-6, IL-8, and IL-10 as well as
for CDV were taken from the literature [42,44,54–57]. Primers were designed using
Beacon Designer software version 2.1 (Premier Biosoft International, Palo Alto, CA) or
the primer-blasting tool of basic local alignment search tool (BLAST). All primers were
purchased from Eurofins MWG Operon (Ebersberg, Germany).
Nucleic acids isolation
Total RNA was isolated from spleens using the RNeasy Mini Kit (Qiagen) as previously
described [44,54]. For the generation of serial standards dilution, total RNA was
extracted from the canine macrophage cell line DH82 (for GAPDH, EF-1α, TGF-β, TNF,
IL-6, IL-8, and IL-10), persistently CDV-infected DH82 cells (for CDV and HPRT) and a
canine lymph node (for IL-2) using TRIZOL (Invitrogen). To isolate total RNA from
spleen tissue of CDV-infected dogs and control animals, 10 sections (thickness 25µm)

39
Chapter 3: Regulatory T cells in CDV infection
from frozen spleen tissue were cut on a cryostat (Microm, Heidelberg, Germany). Total
RNA was extracted using the RNeasy Mini Kit (Qiagen) according to the manufacturer’s
instructions. RNA concentrations were calculated by measuring the optical density at
260 nm (GeneQuantTM pro, Amersham Biosciences Europe GmbH, Freiburg, Germany).
Subsequently, total RNA was reversely transcribed to complementary DNA using the
Omniscript kit (Qiagen) with RNase Out (Invitrogen) and Random Primers (Promega,
Mannheim, Germany) following the manufacturers’ instructions
Polymerase chain reaction for the generation of standard dilutions
For the production of standards, PCR was performed using a Biometra TProfessional
basic thermocycler (Biometra GmbH, Göttingen Germany), as described before
[44,54,57]. Annealing temperature was adjusted to 50°C (IL-2), 56°C (HPRT), 57°C
(TGF-β), 58°C (TNF, IL-6), 59°C (GAPDH, IL-10, CDV), 60°C (EF-1α, IL-8) for two
minutes, and amplification was achieved using AmpliTaq DNA Polymerase (Applied
Biosystems, Carlsbad, CA) in 1 x GeneAmp PCR Buffer II (Applied Biosystems), with
1.25 mmol/L MgCl2, 0.2 mmol/L dNTP mix (Biosystems, Carlsbad, CA), and 300 nmol/L
of each primer. Polymerase chain reaction products of standards were subsequently
analyzed by agarose gel electrophoresis and extracted using NucleoSpin Extract II Kit
(Macherey-Nagel, Düren, Germany) for production of a standard dilution from 102 to 108
copies per microliter.
Reverse transcriptase-quantitative polymerase chain reaction
RT-qPCR and data analysis were performed using the Mx3005P QPCR System (Agilent
Technologies, Waldbronn, Germany) [44,54,57,58]. In addition to the standard dilution,
complementary DNA of spleen samples and negative controls were measured in
duplicate on the same run. Quantification was carried out in 25μl of Brilliant SYBR
Green qPCR Core Reagent Kit (Agilent Technologies). Amplification was performed
using 0.05 U/μl SureStart Taq DNA Polymerase in 1 x Core PCR buffer, with 2.5
mmol/L (CDV, GAPDH, TNF, IL-6, IL-10) or 5 mmol/L (EF-1a, HPRT, IL-2, IL-8, TGF-β)
MgCl2, 8.0% glycerol, 3% dimethyl sulfoxide (4% for TNF-α, IL-2), 150 nmol/L of each

40
Chapter 3: Regulatory T cells in CDV infection
primer (Table 4), 30 nmol/L Rox as reference dye, and 200 μmol/L dNTP mix.
Specificity of the products was assessed by melting curve analysis. Calculated copy
numbers of each gene were normalized to an amount of 100ng of transcribed RNA and
gene expression values were normalized against the three housekeeping genes,
GAPDH, EF-1α, and HPRT, using the software geNorm (Ghent University Hospital
Center for Medical Genetics; available at http://medgen.ugent.be/~jvdesomp/genorm/)
[59]. In brief, the software detects the most stable reference genes of which the
geometric means were used to calculate a normalization factor for the genes of interest.
Statistics
To determine distribution of data, a Shapiro-Wilk test and visualization assessment
were performed. If data were not normally distributed, non-parametrical tests (Mann-
Whitney U-test) were applied and data between two groups were compared. For
statistical analyses and graph development, SPSS software was used.
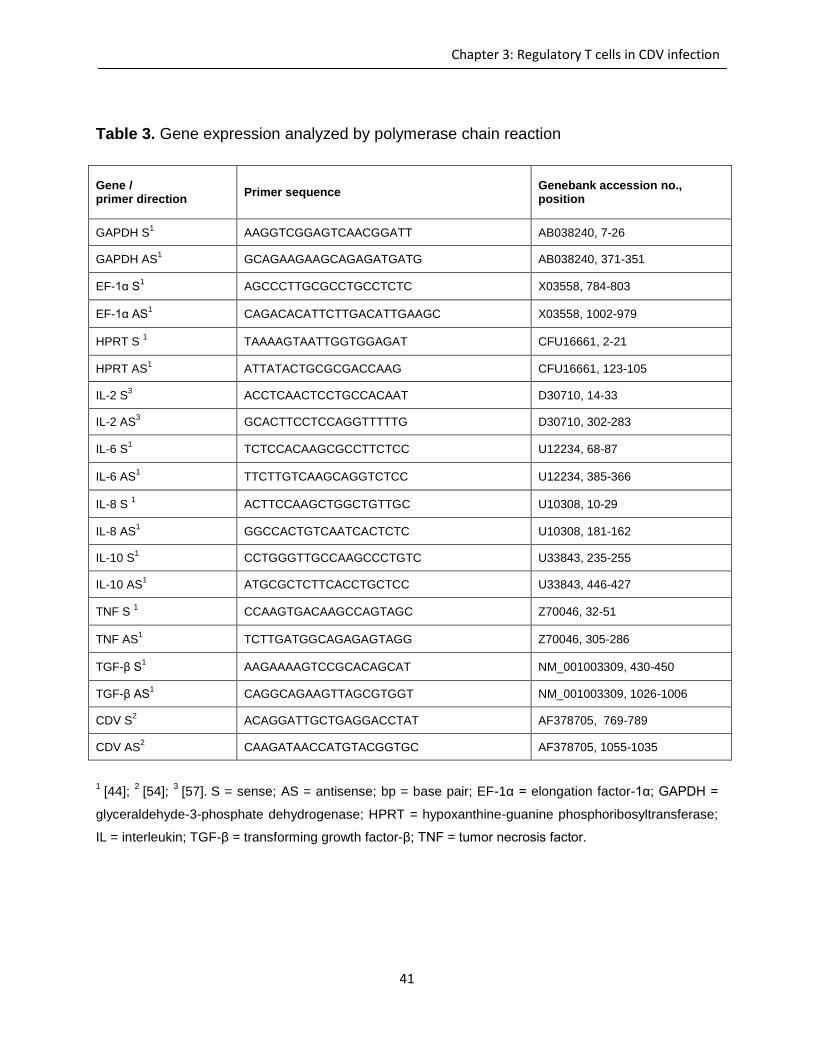
41
Chapter 3: Regulatory T cells in CDV infection
Table 3. Gene expression analyzed by polymerase chain reaction
Gene / primer direction
Primer sequence Genebank accession no., position
GAPDH S1
AAGGTCGGAGTCAACGGATT AB038240, 7-26
GAPDH AS1
GCAGAAGAAGCAGAGATGATG AB038240, 371-351
EF-1α S1
AGCCCTTGCGCCTGCCTCTC X03558, 784-803
EF-1α AS1
CAGACACATTCTTGACATTGAAGC X03558, 1002-979
HPRT S 1
TAAAAGTAATTGGTGGAGAT CFU16661, 2-21
HPRT AS1
ATTATACTGCGCGACCAAG CFU16661, 123-105
IL-2 S3
ACCTCAACTCCTGCCACAAT D30710, 14-33
IL-2 AS3
GCACTTCCTCCAGGTTTTTG D30710, 302-283
IL-6 S1
TCTCCACAAGCGCCTTCTCC U12234, 68-87
IL-6 AS1
TTCTTGTCAAGCAGGTCTCC U12234, 385-366
IL-8 S 1
ACTTCCAAGCTGGCTGTTGC U10308, 10-29
IL-8 AS1
GGCCACTGTCAATCACTCTC U10308, 181-162
IL-10 S1
CCTGGGTTGCCAAGCCCTGTC U33843, 235-255
IL-10 AS1 ATGCGCTCTTCACCTGCTCC U33843, 446-427
TNF S 1
CCAAGTGACAAGCCAGTAGC Z70046, 32-51
TNF AS1 TCTTGATGGCAGAGAGTAGG Z70046, 305-286
TGF-β S1
AAGAAAAGTCCGCACAGCAT NM_001003309, 430-450
TGF-β AS1 CAGGCAGAAGTTAGCGTGGT NM_001003309, 1026-1006
CDV S2
ACAGGATTGCTGAGGACCTAT AF378705, 769-789
CDV AS2
CAAGATAACCATGTACGGTGC AF378705, 1055-1035
1 [44];
2 [54];
3 [57].
S = sense; AS = antisense; bp = base pair; EF-1α = elongation factor-1α; GAPDH =
glyceraldehyde-3-phosphate dehydrogenase; HPRT = hypoxanthine-guanine phosphoribosyltransferase;
IL = interleukin; TGF-β = transforming growth factor-β; TNF = tumor necrosis factor.
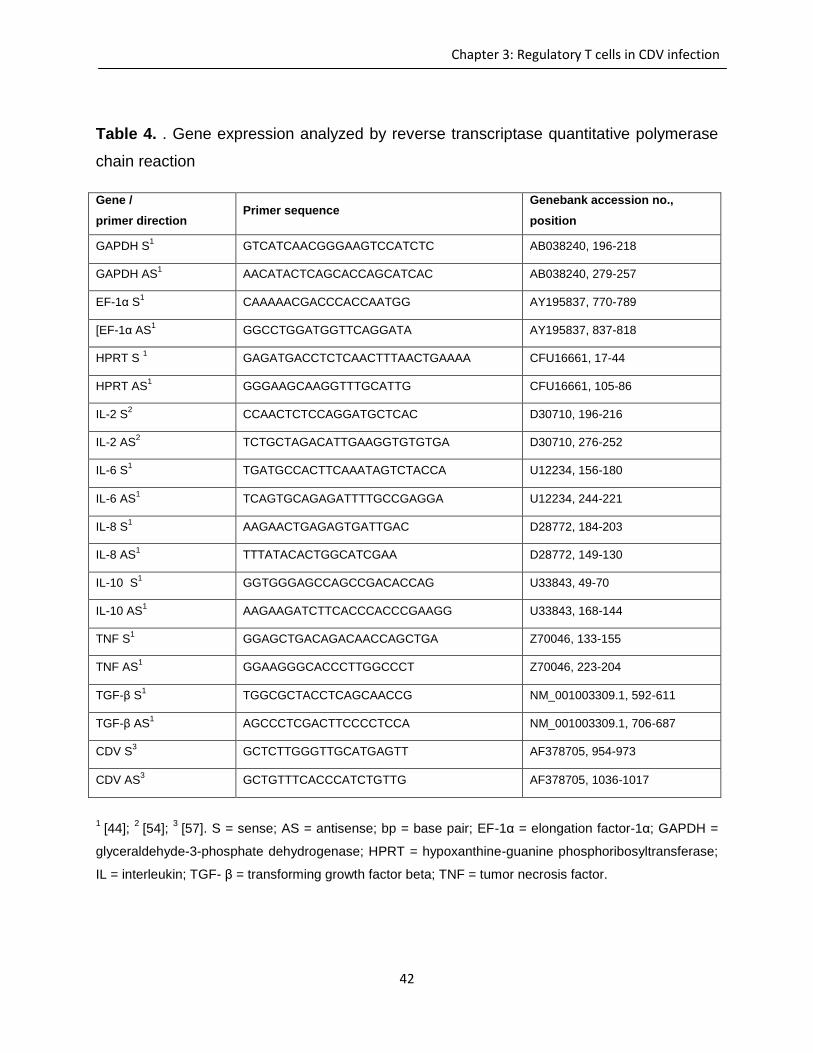
42
Chapter 3: Regulatory T cells in CDV infection
Table 4. . Gene expression analyzed by reverse transcriptase quantitative polymerase
chain reaction
Gene /
primer direction Primer sequence
Genebank accession no.,
position
GAPDH S1
GTCATCAACGGGAAGTCCATCTC AB038240, 196-218
GAPDH AS1
AACATACTCAGCACCAGCATCAC AB038240, 279-257
EF-1α S1
CAAAAACGACCCACCAATGG AY195837, 770-789
[EF-1α AS1
GGCCTGGATGGTTCAGGATA AY195837, 837-818
HPRT S 1
GAGATGACCTCTCAACTTTAACTGAAAA CFU16661, 17-44
HPRT AS1
GGGAAGCAAGGTTTGCATTG CFU16661, 105-86
IL-2 S2
CCAACTCTCCAGGATGCTCAC D30710, 196-216
IL-2 AS2
TCTGCTAGACATTGAAGGTGTGTGA D30710, 276-252
IL-6 S1
TGATGCCACTTCAAATAGTCTACCA U12234, 156-180
IL-6 AS1
TCAGTGCAGAGATTTTGCCGAGGA U12234, 244-221
IL-8 S1
AAGAACTGAGAGTGATTGAC D28772, 184-203
IL-8 AS1
TTTATACACTGGCATCGAA D28772, 149-130
IL-10 S1
GGTGGGAGCCAGCCGACACCAG U33843, 49-70
IL-10 AS1
AAGAAGATCTTCACCCACCCGAAGG U33843, 168-144
TNF S1
GGAGCTGACAGACAACCAGCTGA Z70046, 133-155
TNF AS1 GGAAGGGCACCCTTGGCCCT Z70046, 223-204
TGF-β S1
TGGCGCTACCTCAGCAACCG NM_001003309.1, 592-611
TGF-β AS1 AGCCCTCGACTTCCCCTCCA NM_001003309.1, 706-687
CDV S3
GCTCTTGGGTTGCATGAGTT AF378705, 954-973
CDV AS3
GCTGTTTCACCCATCTGTTG AF378705, 1036-1017
1 [44];
2 [54];
3 [57]. S = sense; AS = antisense; bp = base pair; EF-1α = elongation factor-1α; GAPDH =
glyceraldehyde-3-phosphate dehydrogenase; HPRT = hypoxanthine-guanine phosphoribosyltransferase;
IL = interleukin; TGF- β = transforming growth factor beta; TNF = tumor necrosis factor.

43
Chapter 3: Regulatory T cells in CDV infection
Results
Characterization of white matter lesions in the cerebellum of canine
distemper virus infected dogs
Detection of canine distemper virus in brain lesions
The CDV-NP was detected in all investigated cerebellar lesions by
immunohistochemistry. The virus amount was most prominent in SII lesions (Fig. 1). In
agreement with previous studies [37,53], with disease progression, the number of CDV-
infected cells significantly decreased in SIII lesions compared to non-inflammatory SII
lesions (p=0.019).
Results of statistical analyses (comparison between controls and infected animals) are
listed in table S1.
Phenotypical analyses of inflammatory responses in brain lesions
Progressive hypercellularity was observed in white matter lesions during the disease
course. According to the scheme described in material and methods, 28 brain lesions
were classified as acute non-inflammatory (SI; Fig. 2), 17 plaques as subacute non-
inflammatory (SII) and 25 plaques as subacute to chronic inflammatory (SIII; Fig. 2). A
significant infiltration of CD3+ T cells was found in all lesion types (SI-SIII), whereas a
significant number of infiltrating Foxp3+ cells was found only in advanced inflammatory
brain lesions (SIII; Fig. 3B), indicative of delayed infiltration of Treg. With the onset of
active demyelination (presence of LFB+ gitter cells) and perivascular lymphohistiocytic
inflammation in SIII lesions, the amount of CD3+ T cells significantly increased
compared to SI (p≤0.001) and SII lesions (p=0.006), respectively (Fig. 3A). Similar to
Foxp3+ Treg, CD79α+ B cells were found only in SIII lesions (Fig. 3C). As a
consequence of continuous leukocyte infiltration and glial activation, MHC II was
significantly up-regulated in all lesion types (SI-SIII) compared to control brains (Fig.
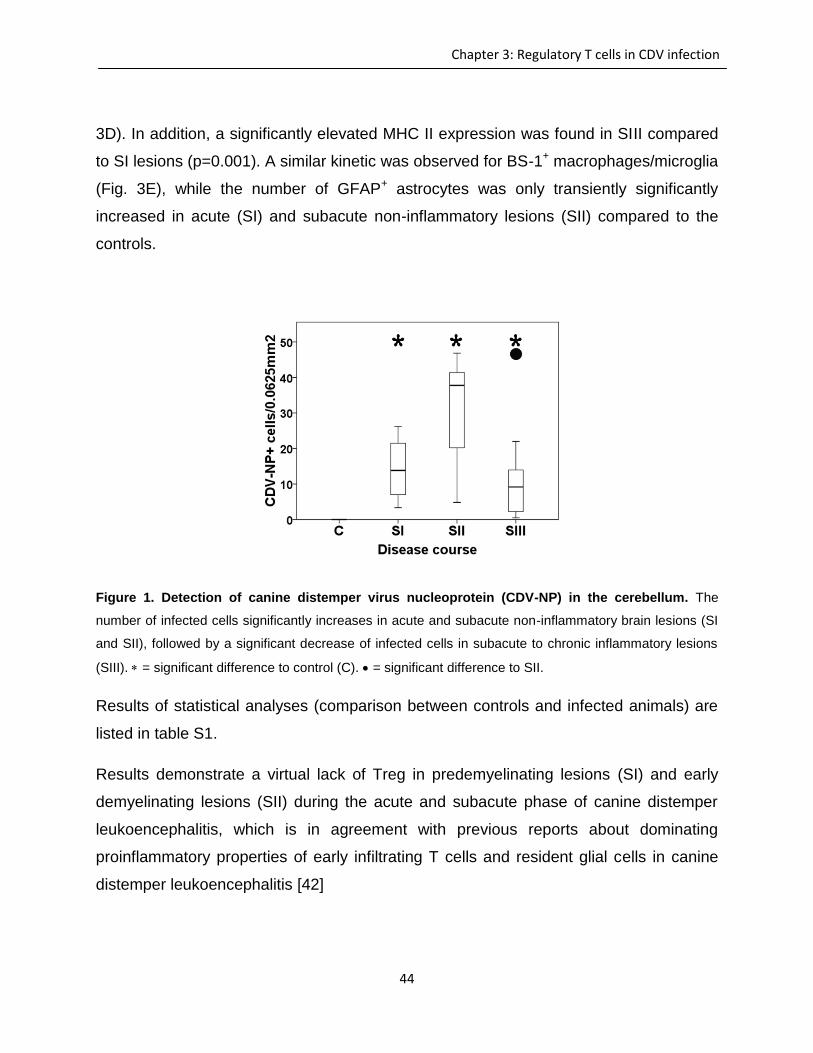
44
Chapter 3: Regulatory T cells in CDV infection
3D). In addition, a significantly elevated MHC II expression was found in SIII compared
to SI lesions (p=0.001). A similar kinetic was observed for BS-1+ macrophages/microglia
(Fig. 3E), while the number of GFAP+ astrocytes was only transiently significantly
increased in acute (SI) and subacute non-inflammatory lesions (SII) compared to the
controls.
Figure 1. Detection of canine distemper virus nucleoprotein (CDV-NP) in the cerebellum. The
number of infected cells significantly increases in acute and subacute non-inflammatory brain lesions (SI
and SII), followed by a significant decrease of infected cells in subacute to chronic inflammatory lesions
(SIII). = significant difference to control (C). = significant difference to SII.
Results of statistical analyses (comparison between controls and infected animals) are
listed in table S1.
Results demonstrate a virtual lack of Treg in predemyelinating lesions (SI) and early
demyelinating lesions (SII) during the acute and subacute phase of canine distemper
leukoencephalitis, which is in agreement with previous reports about dominating
proinflammatory properties of early infiltrating T cells and resident glial cells in canine
distemper leukoencephalitis [42]
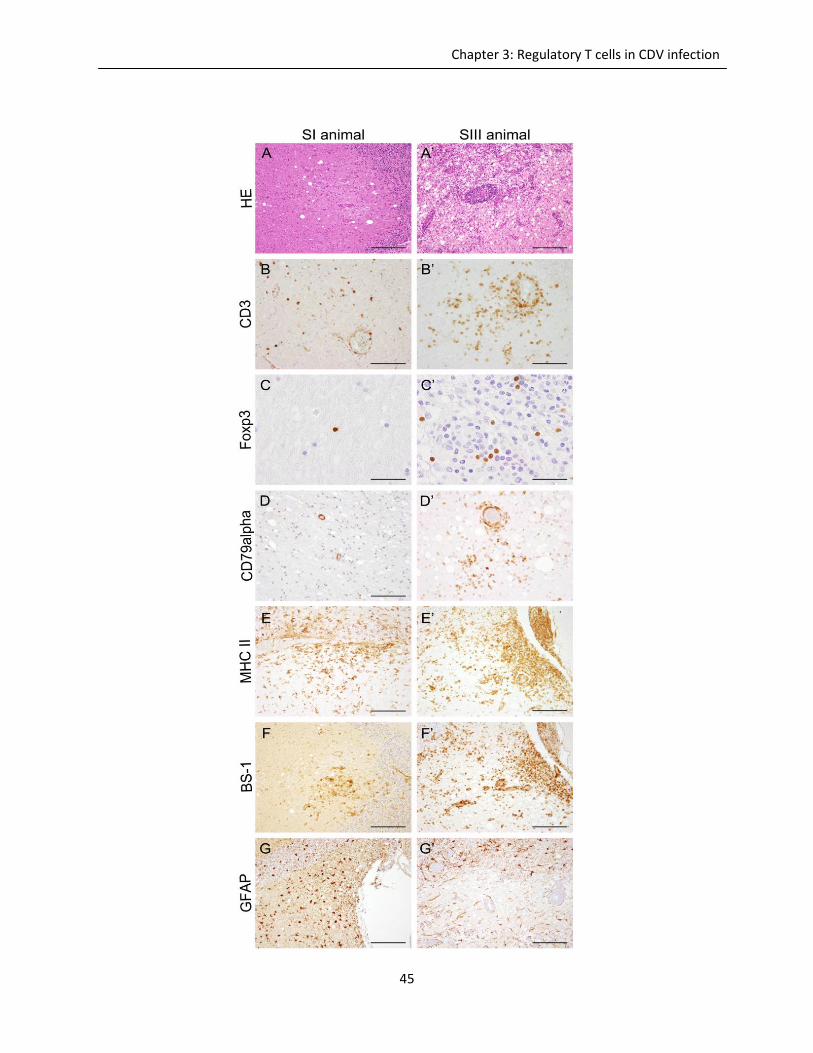
45
Chapter 3: Regulatory T cells in CDV infection

46
Chapter 3: Regulatory T cells in CDV infection
Figure 2. Characterization of white mater lesions in canine distemper virus infected dogs. A) Mild
vacuolization and cellularity in acute white matter lesions (SI). A’) Increased perivascular and
parenchymal infiltration of immune cells in subacute to chronic demyelinating brain lesions (SIII);
hematoxylin and eosin (HE) staining. B) Low numbers of CD3+ T cells in early predemyelinating lesions.
B’) Prominent CD3+ T cells infiltrates in advanced inflammatory lesions, immunohistochemistry. C) Few
individual Foxp3+ regulatory T cells in SI lesions. C’) Increased Foxp3-expression in advanced
inflammatory brain lesions, immunohistochemistry. D) Few CD79α+ B cells in SI lesions. D’) Increased
influx of CD79α+ B cells in SIII lesions, immunohistochemistry. E) Early MHC II-upregulation of glial cells
in predemyelinating lesions. E’) Additional infiltration of MHC II+ leukocytes in advanced inflammatory
lesions, immunohistochemistry. F) Low numbers of BS-1+ macrophages/microglia in SI lesions. F’)
Significant increase of BS-1+ microglia and infiltrating BS-1
+ macrophages in SIII lesions, lectin
histochemistry. G) Early prominent increase of GFAP-expression in astrocytes in predemyelinating
lesions. G’) Loss of intralesional GFAP+ astrocytes in advanced lesions, immunohistochemistry. Scale
bars: A, A’, E, E’, F, F’, G, G’ = 200μm; B, B’, C, C’, D, D’ = 20μm
.
To testify the hypothesis that the delayed onset Treg infiltration in the brain is a
consequence of peripheral Treg depletion, phenotypical changes and the associated
cytokine expression in the spleen, as a lymphoid organ consistently affected in systemic
distemper, were investigated [38].
Characterization of immunological changes in the spleen of canine
distemper virus infected dogs
Detection of canine distemper virus in the spleen
The quantity and distribution of CDV protein in the spleen were determined by
immunohistochemistry. In addition, the amount of virus RNA was measured by RT-
qPCR. Similar to the brain, virus load in the spleen was highest during early infection
phases. Using immunohistochemistry, CDV-infected cells were found in animals with
early brain lesions (SI and SII) with most prominent virus expression in SII group
animals.
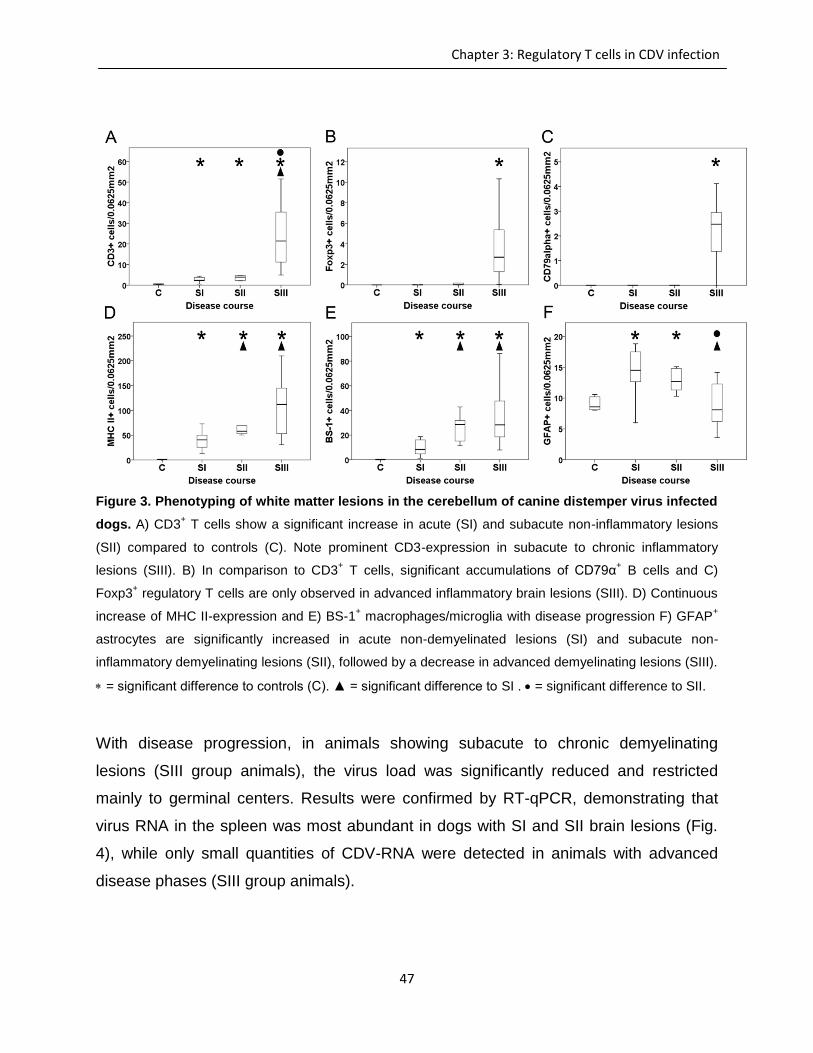
47
Chapter 3: Regulatory T cells in CDV infection
Figure 3. Phenotyping of white matter lesions in the cerebellum of canine distemper virus infected
dogs. A) CD3+ T cells show a significant increase in acute (SI) and subacute non-inflammatory lesions
(SII) compared to controls (C). Note prominent CD3-expression in subacute to chronic inflammatory
lesions (SIII). B) In comparison to CD3+ T cells, significant accumulations of CD79α
+ B cells and C)
Foxp3+ regulatory T cells are only observed in advanced inflammatory brain lesions (SIII). D) Continuous
increase of MHC II-expression and E) BS-1+ macrophages/microglia with disease progression F) GFAP
+
astrocytes are significantly increased in acute non-demyelinated lesions (SI) and subacute non-
inflammatory demyelinating lesions (SII), followed by a decrease in advanced demyelinating lesions (SIII).
= significant difference to controls (C). ▲ = significant difference to SI . = significant difference to SII.
With disease progression, in animals showing subacute to chronic demyelinating
lesions (SIII group animals), the virus load was significantly reduced and restricted
mainly to germinal centers. Results were confirmed by RT-qPCR, demonstrating that
virus RNA in the spleen was most abundant in dogs with SI and SII brain lesions (Fig.
4), while only small quantities of CDV-RNA were detected in animals with advanced
disease phases (SIII group animals).
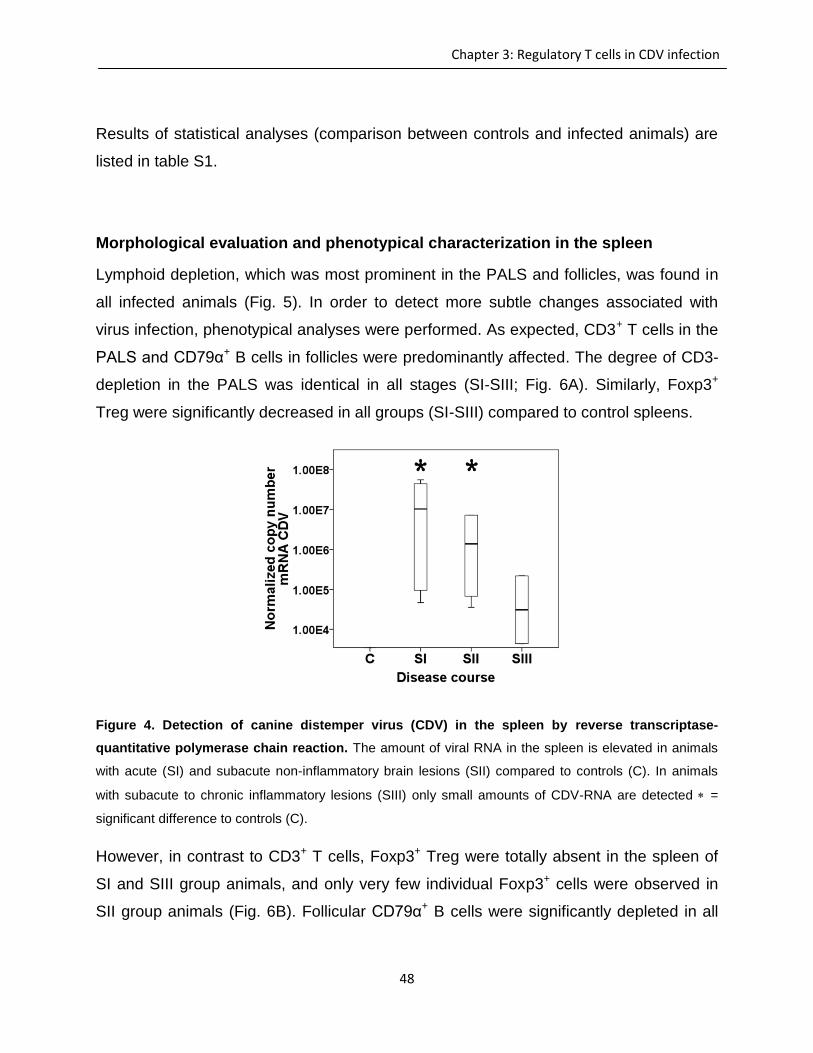
48
Chapter 3: Regulatory T cells in CDV infection
Results of statistical analyses (comparison between controls and infected animals) are
listed in table S1.
Morphological evaluation and phenotypical characterization in the spleen
Lymphoid depletion, which was most prominent in the PALS and follicles, was found in
all infected animals (Fig. 5). In order to detect more subtle changes associated with
virus infection, phenotypical analyses were performed. As expected, CD3+ T cells in the
PALS and CD79α+ B cells in follicles were predominantly affected. The degree of CD3-
depletion in the PALS was identical in all stages (SI-SIII; Fig. 6A). Similarly, Foxp3+
Treg were significantly decreased in all groups (SI-SIII) compared to control spleens.
Figure 4. Detection of canine distemper virus (CDV) in the spleen by reverse transcriptase-
quantitative polymerase chain reaction. The amount of viral RNA in the spleen is elevated in animals
with acute (SI) and subacute non-inflammatory brain lesions (SII) compared to controls (C). In animals
with subacute to chronic inflammatory lesions (SIII) only small amounts of CDV-RNA are detected =
significant difference to controls (C).
However, in contrast to CD3+ T cells, Foxp3+ Treg were totally absent in the spleen of
SI and SIII group animals, and only very few individual Foxp3+ cells were observed in
SII group animals (Fig. 6B). Follicular CD79α+ B cells were significantly depleted in all

49
Chapter 3: Regulatory T cells in CDV infection
infected animals (SI-SIII) compared to controls. Suggestive of B cell repopulation, a
statistical tendency (p=0.054) of increased CD79α+ B cell numbers in the spleen of SIII
compared to SI group animals was observed (Fig. 6C). L1+ macrophages were
significantly depleted in the splenic marginal zone of SII group animals (Fig. 6D).
Associated with lymphoid depletion a reduced expression of MHC II in all compartments
of the white pulp (follicle, PALS and marginal zone) was detected in all CDV-infected
dogs. However, a significant increase of total MHC II+ cells in SIII group compared to
SII, was found in follicles (p=0.029; Fig. 6E), PALS (p=0.040; Fig. 6F) and marginal
zones (p=0.021; data not shown), indicative of partial repopulation of lymphoid organs in
advanced stages of canine distemper as previously described [38].
Results of statistical analyses (comparison between controls and infected animals) are
listed in table S1.
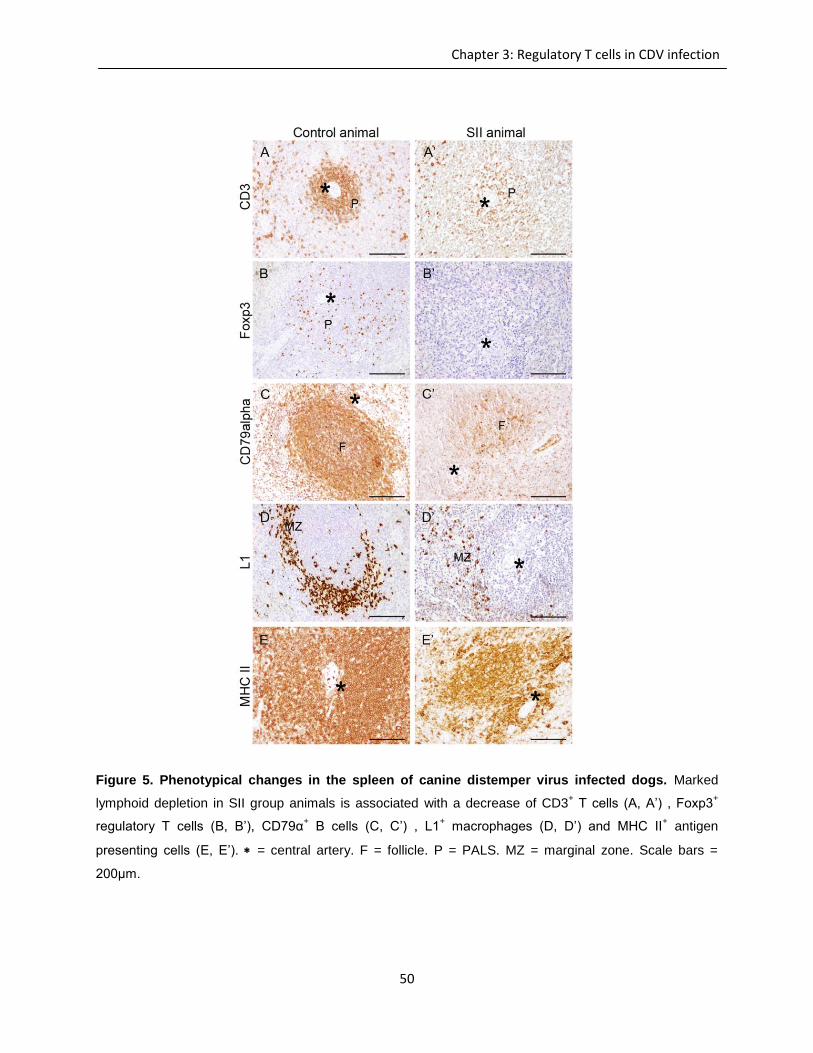
50
Chapter 3: Regulatory T cells in CDV infection
Figure 5. Phenotypical changes in the spleen of canine distemper virus infected dogs. Marked
lymphoid depletion in SII group animals is associated with a decrease of CD3+ T cells (A, A’) , Foxp3
+
regulatory T cells (B, B’), CD79α+ B cells (C, C’) , L1
+ macrophages (D, D’) and MHC II
+ antigen
presenting cells (E, E’). = central artery. F = follicle. P = PALS. MZ = marginal zone. Scale bars =
200μm.
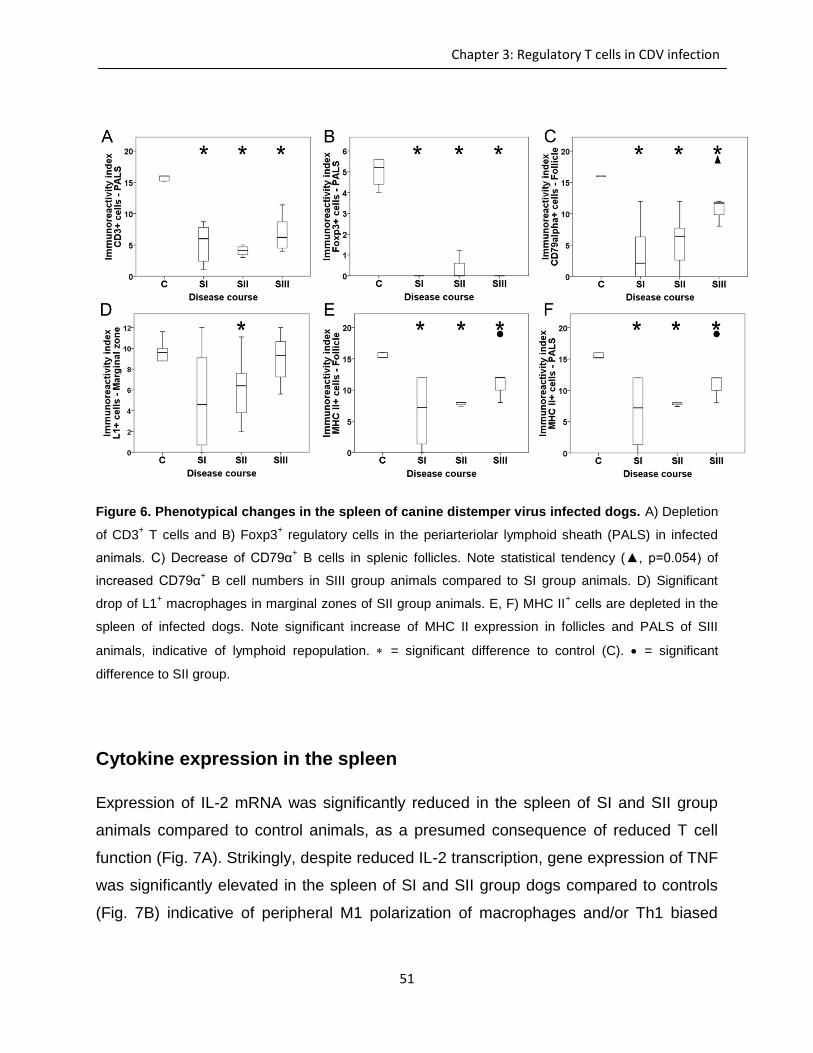
51
Chapter 3: Regulatory T cells in CDV infection
Figure 6. Phenotypical changes in the spleen of canine distemper virus infected dogs. A) Depletion
of CD3+ T cells and B) Foxp3
+ regulatory cells in the periarteriolar lymphoid sheath (PALS) in infected
animals. C) Decrease of CD79α+ B cells in splenic follicles. Note statistical tendency (▲, p=0.054) of
increased CD79α+ B cell numbers in SIII group animals compared to SI group animals. D) Significant
drop of L1+ macrophages in marginal zones of SII group animals. E, F) MHC II
+ cells are depleted in the
spleen of infected dogs. Note significant increase of MHC II expression in follicles and PALS of SIII
animals, indicative of lymphoid repopulation. = significant difference to control (C). = significant
difference to SII group.
Cytokine expression in the spleen
Expression of IL-2 mRNA was significantly reduced in the spleen of SI and SII group
animals compared to control animals, as a presumed consequence of reduced T cell
function (Fig. 7A). Strikingly, despite reduced IL-2 transcription, gene expression of TNF
was significantly elevated in the spleen of SI and SII group dogs compared to controls
(Fig. 7B) indicative of peripheral M1 polarization of macrophages and/or Th1 biased
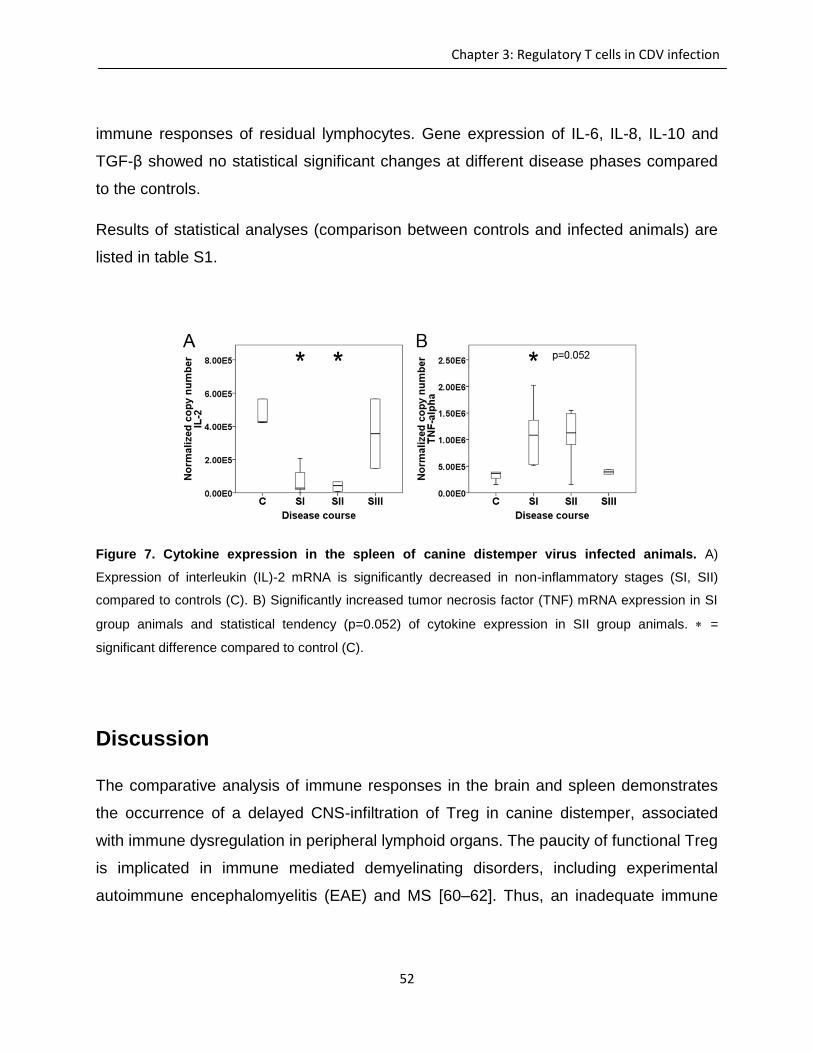
52
Chapter 3: Regulatory T cells in CDV infection
immune responses of residual lymphocytes. Gene expression of IL-6, IL-8, IL-10 and
TGF-β showed no statistical significant changes at different disease phases compared
to the controls.
Results of statistical analyses (comparison between controls and infected animals) are
listed in table S1.
Figure 7. Cytokine expression in the spleen of canine distemper virus infected animals. A)
Expression of interleukin (IL)-2 mRNA is significantly decreased in non-inflammatory stages (SI, SII)
compared to controls (C). B) Significantly increased tumor necrosis factor (TNF) mRNA expression in SI
group animals and statistical tendency (p=0.052) of cytokine expression in SII group animals. =
significant difference compared to control (C).
Discussion
The comparative analysis of immune responses in the brain and spleen demonstrates
the occurrence of a delayed CNS-infiltration of Treg in canine distemper, associated
with immune dysregulation in peripheral lymphoid organs. The paucity of functional Treg
is implicated in immune mediated demyelinating disorders, including experimental
autoimmune encephalomyelitis (EAE) and MS [60–62]. Thus, an inadequate immune

53
Chapter 3: Regulatory T cells in CDV infection
homeostasis as a consequence of Treg depletion is assumed to allow excessive glial
and T cell responses during the predemyelinating and early demyelinating phase of
CDV-induced leukoencephalitis as previously described [36,42].
Manipulation of Treg in the periphery has significant consequences for the fate of viral
infection in the brain. In agreement with the idea of a misbalanced CNS immune
response in canine distemper, insufficient expansion of peripheral Treg leads to strong
immunopathology in acute severe dengue fever and West Nile virus infection [63,64].
The protective role of Treg against an overwhelming immune response in the CNS also
becomes obvious in animal models of stroke and EAE [65,66] and in human
immunodeficiency virus-1-associated neurodegeneration, where they reduce
astrogliosis and microglia-mediated inflammation [67]. Furthermore, functional deficits
or reduced numbers of Treg are suspected to contribute to demyelination in acute MS
lesions [68]. Silencing of microglia by Treg is mediated by IL-4 which leads to reduced
production of toxic factors such as nitric oxide [69]. Moreover, interaction with Treg
induces a M2 phenotype of microglia/macrophages which exhibit immunomodulatory
properties and potentially promote regeneration in the injured CNS [70,71]. Accordingly,
the observed absence of CNS-infiltrating Treg is in agreement with the previous findings
of low anti-inflammatory cytokine expression [42] and release of myelinotoxic molecules
by microglia in early distemper brain lesions [43].
Interestingly, the increase of Foxp3+ Treg in the CNS during advanced stages of canine
distemper coincides with the persistent infection, which corresponds to findings in
experimental measles virus infection of mice [5,30]. Different explanations about the
origin of Treg in the brain during chronic CDV-infection have to be considered. Since
prominent depletion of Treg was observed in the spleen of chronically infected dogs,
brain infiltration might result from an accumulation of residual Treg in other peripheral
organs. In addition, it has been shown that the inflammatory environment critically
influences the balance of Treg versus effector T cell differentiation which might lead to
local induction and expansion of Foxp3+ Treg within the brain of CDV-infected animals.

54
Chapter 3: Regulatory T cells in CDV infection
For instance, virus-specific CD4+ T cells can acquire a Treg phenotype including Foxp3
expression in the CNS during mouse hepatitis virus infection [72]. Moreover, neurons
and activated astrocytes are able to induce Treg as a mechanism for decreasing
excessive inflammation and demyelination in EAE [73,74]. However, as observed in the
present study, Treg seem to be unable to terminate neuroinflammation in chronic
demyelinating lesions of CDV-infected dogs, indicative of an insufficient amount or
dysfunction of these cells. Comparably, large numbers of brain-infiltrating Treg are
present in EAE, but their function is impaired by the elevated expression of
proinflammatory molecules, such as IL-6 and TNF [85, 86]. As demonstrated in previous
studies, the presence of these cytokines in the cerebrospinal fluid is associated with
disease exacerbation in CDV-infected dogs [77]. Thus, it is tempting to speculate that
the pro-inflammatory milieu in the canine brain inhibits the neuroprotective efficacy of
Treg in advanced stages. Alternatively, in spite of a potential beneficial effect upon
immune mediated brain damage in chronic distemper, immunosuppression by Treg
might contribute to viral persistence triggering progressive myelin loss as observed in
coronavirus-induced demyelination of mice [78].
Similar to human measles, a striking feature of CDV-infection of dogs is a marked loss
of immune cells and impaired immune response which favors viral persistence in
lymphoid organs and CNS [4,38,79,80]. Lymphoid depletion in canine distemper is
induced by necrosis and apoptosis directly mediated by the virus as well as by virus
independent mechanisms, probably by an over-activation of the innate immune
response [79]. Accordingly, the observed increased TNF expression might enhance
splenic depletion by induction of lymphocyte apoptosis in acutely infected dogs.
Moreover, TNF has been shown to downregulate the suppressive function of Treg by
decreasing Foxp3 expression [81]. Thus, the pro-inflammatory microenvironment might
contribute to the observed depletion of Foxp3+ cells in the spleen and, in addition,
potentially foster Th1-biased immune responses and/or M1 polarization of macrophages
[82,83]. Such primed immune cells lead to antiviral immunity, but might also account for

55
Chapter 3: Regulatory T cells in CDV infection
early lesion development by promoting excessive neuroinflammation in canine
distemper [36,37,42].
As shown in previous studies, the low number of inflammatory cells in early non-
inflammatory CNS lesions coincided with the degree of peripheral lymphoid depletion,
whereas CNS-infiltration of inflammatory cells during the chronic demyelinating phase is
associated with reconstitution of lymphoid organs [38]. Partial repopulation in chronically
infected dogs in the present survey was demonstrated by an increase of MHC II+
lymphoid cells in splenic T and B cell areas, associated with a statistical trend of
elevated numbers of CD79α+ B cells in follicles. Following measles virus infection of
children, an early Th1 response is quickly succeeded by a prolonged Th2 or mixed
Th1/Th2 response [84]. Th2 cytokine predominance after resolution of measles rash
produces an environment favoring B cell maturation which facilitates humoral immunity
important for lifelong protection against reinfection, while depressing macrophage
activation and Th1 responses which are required for combating pathogens [85]. The
importance of a robust Th1 immunity for virus elimination has also been demonstrated
in CDV-infected ferrets, a model for morbillivirus-induced immunosuppression [86].
Taken together, B cell immunity and Th1/Th2 imbalance rather than mounting Treg
responses might contribute to prolonged immune alteration in canine distemper, which
contrasts with the currently discussed role of Treg in human measles [24,25]. However,
further studies of advanced disease stages are needed to depict further similarities and
differences between canine distemper and human measles.
Interestingly, according to the currently discussed role of B cells and antibodies in the
pathogenesis of demyelinating disorders [47,75,76] an increased intrathecal antibody
production is suspected to accelerate myelin destruction in chronic canine distemper
[89]. Thus, besides its impact on virus-specific immunity, observed reconstitution of
splenic B cells and recruitment of peripheral CD79α+ B cells to the brain might also
represent an initiator for CNS-restricted plasma cell differentiation and subsequent
myelin loss in dogs during the chronic distemper phase. Referring to this, a

56
Chapter 3: Regulatory T cells in CDV infection
compartmentalization of immune responses in the brain with local antigen presentation,
plasma cell formation and antibody production behind a closed blood brain barrier is
currently discussed for progressive MS [90,91].
In conclusion, for the first time the interplay between Treg in peripheral lymphoid tissue
and brain of CDV-infected dogs has been demonstrated. Results of the present study
are indicative of inadequate immunoregulation which represents a potential prerequisite
for early lesion development. As suggested for MS [75], the development of novel
therapies targeting Treg represent a promising strategy in canine CNS disorders.
However, further studies are needed to determine disease phase-specific and probably
ambiguous functions of Treg in the pathogenesis of canine distemper, which might also
have relevance for the understanding of human demyelinating diseases.
Acknowledgements
The authors would like to thank Danuta Waschke, Bettina Buck and Petra Grünig for
their excellent technical support during the laboratory work and Dr. Karl Rohn for
statistical analyses. This study was supported by the German Research Foundation
(FOR 1103, BA 815/10-2 and BE 4200/1-2).

57
Chapter 3: Regulatory T cells in CDV infection
References
1. Pringle CR (1999) Virus taxonomy at the XIth International Congress of Virology, Sydney,
Australia, 1999. Arch Virol 144: 2065–2070.
2. Sips GJ, Chesik D, Glazenburg L, Wilschut J, Keyser JD, et al. (2007) Involvement of
morbilliviruses in the pathogenesis of demyelinating disease. Rev Med Virol 17: 223–244.
3. Wyss-Fluehmann G, Zurbriggen A, Vandevelde M, Plattet P (2010) Canine distemper virus
persistence in demyelinating encephalitis by swift intracellular cell-to-cell spread in astrocytes is
controlled by the viral attachment protein. Acta Neuropathol 119: 617–630.
4. Beineke A, Puff C, Seehusen F, Baumgärtner W, (2009) Pathogenesis and immunopathology of
systemic and nervous canine distemper. Vet Immunol Immunopathol 127: 1–18.
5. Sellin CI, Jégou JF, Renneson J, Druelle J, Wild TF, et al. (2009) Interplay between virus-specific
effector response and Foxp3 regulatory T cells in measles virus immunopathogenesis. PloS one
4: e4948.
6. von Messling V, Svitek N, Cattaneo R (2006) Receptor (SLAM [CD150]) recognition and the V
protein sustain swift lymphocyte-based invasion of mucosal tissue and lymphatic organs by a
morbillivirus. J Virol 80: 6084–6092.
7. Confer AW, Kahn DE, Koestner A, Krakowka S (1975) Comparison of canine distemper viral
strains: an electron microscopic study. Am J Vet Res 36: 741–748.
8. Young VA, Rall GF (2009) Making it to the synapse: Measles virus spread in and among neurons.
Curr Top Microbiol Immunol 330: 3–30.
9. Summers A, Greisen AH, Appel GJM (1984) Canine Distemper Encephalomyelitis: Variation With
Virus Strain. J Comp Pathol 94: 65–75.
10. Ishizaki Y, Yukaya N, Kusuhara K, Kira R, Torisu H, et al. (2010) PD1 as a common candidate
susceptibility gene of subacute sclerosing panencephalitis. Hum Genet 127: 411–419.
11. Vandevelde M, Zurbriggen A (2005) Demyelination in canine distemper virus infection: a review.
Acta Neuropathol 109: 56–68.
12. Feuerer M, Hill JA, Kretschmer K, von Boehmer H, Mathis D, et al. (2010) Genomic definition of
multiple ex vivo regulatory T cell subphenotypes. Proc Natl Acad Sci U S A 107: 5919–5924.
13. Sakaguchi S, Ono M, Setoguchi R, Yagi H, Hori S, et al. (2006) Foxp3+ CD25+ CD4+ natural
regulatory T cells in dominant self-tolerance and autoimmune disease. Immunol Rev 212: 8–27.
14. Feuerer M, Hill JA, Mathis D, Benoist C, (2009) Foxp3+ regulatory T cells: differentiation,
specification, subphenotypes. Nat Immunol 10: 689–695.
15. MacDonald AJ, Duffy M, Brady MT, McKiernan S, Hall W, et al. (2002) CD4 T helper type 1 and
regulatory T cells induced against the same epitopes on the core protein in hepatitis C virus-
infected persons. J Infect Dis 185: 720–727.

58
Chapter 3: Regulatory T cells in CDV infection
16. Sakaguchi S (2003) Regulatory T cells: mediating compromises between host and parasite. Nat
Immunol 4: 10–11.
17. Vignali AAD, Collison WL, Workman JC (2009) How regulatory T cells work. Nat Rev Immunol 8:
523–532.
18. Lund MJ, Hsing L, Pham TT, Rudensky Y (2008) Coordination of Early Protective Immunity to
Viral Infection by Regulatory T Cells. Science 320: 1220–1224.
19. Göbel K, Bittner S, Melzer N, Pankratz S, Dreykluft A, et al. (2012) CD4(+) CD25(+) FoxP3(+)
regulatory T cells suppress cytotoxicity of CD8(+) effector T cells: implications for their capacity to
limit inflammatory central nervous system damage at the parenchymal level. J Neuroinflammation
9: 41.
20. Herder V, Hansmann F, Stangel M, Schaudien D, Rohn K, et al. (2012) Cuprizone inhibits
demyelinating leukomyelitis by reducing immune responses without virus exacerbation in an
infectious model of multiple sclerosis. J Neuroimmunol 244: 84–93.
21. Richards MH, Getts MT, Podojil JR, Jin Y, Kim S, et al. (2012) Virus Expanded Regulatory T
Cells Control Disease Severity in the Theiler’s Virus Mouse Model of MS. J Autoimmun 36: 142–
154.
22. Zelinskyy G, Dietze K, Sparwasser T, Dittmer U (2009) Regulatory T Cells Suppress Antiviral
Immune Responses and Increase Viral Loads during Acute Infection with a Lymphotropic
Retrovirus. PLoS Pathog 5: e1000406.
23. Toka FN, Suvas S, Rouse BT (2004) CD4+ CD25
+ T Cells Regulate Vaccine-Generated Primary
and Memory CD8+ T-Cell Responses against Herpes Simplex Virus Type 1. J Virol 78: 13082–
13089.
24. Fernandez MA, Puttur FK, Wang YM, Howden W, Alexander SI, et al. (2008) T Regulatory Cells
Contribute to the Attenuated Primary CD8+ and CD4+ T Cell Responses to Herpes Simplex Virus
Type 2 in Neonatal Mice. J Immunol 180: 1556–1564.
25. Aydin OF, Ichiyama T, Anlar B (2010) Serum and cerebrospinal fluid cytokine concentrations in
subacute sclerosing panencephalitis. Brain Dev 32: 463–466.
26. Yu X, Cheng Y, Shi B, Qian F, Qi T, et al. (2008) Measles Virus Infection in Adults Induces
Production of IL-10 and Is Associated with Increased CD4 + CD25 + Regulatory T Cells. J
Immunol 181 : 7356–7366.
27. Moss WJ, Ryon JJ, Monze M, Griffin DE (2002) Differential regulation of interleukin (IL)-4, IL-5,
and IL-10 during measles in Zambian children. J Infect Dis 186: 879–887.
28. Li MZ, Xu FD, Huang XH, Chen XC, Deng QW, et al. (2008) [Immunosuppression induced by
measles virus in adult patients is not related to CD4+ CD25+ regulatory T cell induction].
Zhonghua Shi Yan He Lin Chuang Bing Du Xue Za Zhi 22: 211–213.

59
Chapter 3: Regulatory T cells in CDV infection
29. Wu XM, Liao YW, Wang HY, Ji KQ, Li GF, et al. (2012) Integrin alphavbeta6 is involved in
measles protein-induced airway immune suppression. Cytokine 59: 59–64.
30. Reuter D, Sparwasser T, Hünig T, Schneider-Schaulies J (2012) Foxp3+ Regulatory T Cells
Control Persistence of Viral CNS Infection. PLoS ONE 7: e33989.
31. Koga R, Ohno S, Ikegame S, Yanagi Y (2010) Measles virus-induced immunosuppression in
SLAM knock-in mice. Journal of virology 84: 5360–5367.
32. Kling J (2007) Biotech for your companion? Nat Biotechnol 25: 1343–1345.
33. Paoloni M, Khanna C (2008) Translation of new cancer treatments from pet dogs to humans. Nat
Rev Cancer 8: 147–156.
34. Spitzbarth I, Baumgärtner W, Beineke A (2012) The role of pro- and anti-inflammatory cytokines
in the pathogenesis of spontaneous canine CNS diseases. Vet Immunol Immunopathol 147: 6–
24.
35. Alldinger S, Wünschmann A, Baumgärtner W, Voss C, Kremmer E (1996) Up-regulation of major
histocompatibility complex class II antigen expression in the central nervous system of dogs with
spontaneous canine distemper virus encephalitis. Acta Neuropathol 92: 273–280.
36. Tipold A, Moore P, Zurbriggen A, Burgener I, Barben G, et al. (1999) Early T cell response in the
central nervous system in canine distemper virus infection. Acta Neuropathol 97: 45–56.
37. Wünschmann A, Alldinger S, Kremmer E, Baumgärtner W (1999) Identification of CD4+ and
CD8+ T cell subsets and B cells in the brain of dogs with spontaneous acute, subacute-, and
chronic-demyelinating distemper encephalitis. Vet Immunol Immunopathol 67: 101–116.
38. Wünschmann A, Kremmer E, Baumgärtner W (2000) Phenotypical characterization of T and B
cell areas in lymphoid tissues of dogs with spontaneous distemper. Vet Immunol Immunopathol
73: 83–98.
39. Alldinger S, Baumgärtner W, Orvell C (1993) Restricted expression of viral surface proteins in
canine distemper encephalitis. Acta Neuropathol 85: 635–645.
40. Beineke A, Markus S, Borlak J, Thum T, Baumgärtner W (2008) Increase of pro-inflammatory
cytokine expression in non-demyelinating early cerebral lesions in nervous canine distemper.
Viral Immunol 21: 401–410.
41. Imbschweiler I, Seehusen F, Peck C-T, Omar M, Baumgärtner W, et al. (2012) Increased p75
neurotrophin receptor expression in the canine distemper virus model of multiple sclerosis
identifies aldynoglial Schwann cells that emerge in response to axonal damage. Glia 60: 358–
371.
42. Markus S (2002) Increased expression of pro-inflammatory cytokines and lack of up-regulation of
anti-inflammatory cytokines in early distemper CNS lesions. J Neuroimmunol 125: 30–41.

60
Chapter 3: Regulatory T cells in CDV infection
43. Stein VM, Czub M, Schreiner N, Moore PF, Vandevelde M, et al. (2004) Microglial cell activation
in demyelinating canine distemper lesions. J Neuroimmunol 153: 122–131.
44. Spitzbarth I, Bock P, Haist V, Stein VM, Tipold A, et al. (2011) Prominent microglial activation in
the early proinflammatory immune response in naturally occurring canine spinal cord injury. J
Neuropathol Exp Neurol 70: 703–714.
45. Gay FW, Drye TJ, Dick GW, Esiri MM (1997) The application of multifactorial cluster analysis in
the staging of plaques in early multiple sclerosis. Identification and characterization of the primary
demyelinating lesion. Brain 120 : 1461–1483.
46. Barnett MH, Prineas JW (2004) Relapsing and remitting multiple sclerosis: pathology of the newly
forming lesion. Ann Neurol 55: 458–468.
47. Barnett MH, Sutton I (2006) The pathology of multiple sclerosis: a paradigm shift. Curr Opin
Neurol 19: 242–247.
48. Marik C, Felts PA, Bauer J, Lassmann H, Smith KJ (2007) Lesion genesis in a subset of patients
with multiple sclerosis: a role for innate immunity? Brain 130: 2800–2815.
49. Navarrete-Talloni MJ, Kalkuhl A, Deschl U, Ulrich R, Kummerfeld M, et al. (2010) Transient
peripheral immune response and central nervous system leaky compartmentalization in a viral
model for multiple sclerosis. Brain Pathol 20: 890–901.
50. Tsunoda I, Libbey JE, Kuang L-Q, Terry EJ, Fujinami RS (2005) Massive apoptosis in lymphoid
organs in animal models for primary and secondary progressive multiple sclerosis. Am J Pathol
167: 1631–1646.
51. De Vos AF, van Riel DA, van Meurs M, Brok HPM, Boon L, et al. (2005) Severe T-cell depletion
from the PALS leads to altered spleen composition in common marmosets with experimental
autoimmune encephalomyelitis (EAE). J neuroimmunol 161: 29–39.
52. Gröters S, Alldinger S, Baumgärtner W (2005) Up-regulation of mRNA for matrix
metalloproteinases-9 and -14 in advanced lesions of demyelinating canine distemper
leukoencephalitis. Acta Neuropathol 110: 369–382.
53. Seehusen F, Orlando EA, Wewetzer K, Baumgärtner W (2007) Vimentin-positive astrocytes in
canine distemper: a target for canine distemper virus especially in chronic demyelinating lesions?
Acta Neuropathol 114: 597–608.
54. Puff C, Krudewig C, Imbschweiler I, Baumgärtner W, Alldinger S (2009) Influence of persistent
canine distemper virus infection on expression of RECK, matrix-metalloproteinases and their
inhibitors in a canine macrophage/monocytic tumour cell line (DH82). Vet J 182: 100–107.
55. von Smolinski D, Leverkoehne I, von Samson-Himmelstjerna G, Gruber AD (2005) Impact of
formalin-fixation and paraffin-embedding on the ratio between mRNA copy numbers of differently
expressed genes. Histochem Cell Biol 124: 177–188.

61
Chapter 3: Regulatory T cells in CDV infection
56. Matsumoto Y, Mohamed A, Onodera T, Kato H, Ohashi T, et al. (1994) Molecular cloning and
expression of canine interleukin 8 cDNA. Cytokine 6: 455–461.
57. Schwartz M, Puff C, Stein VM, Baumgärtner W, Tipold A (2011) Pathogenetic factors for
excessive IgA production: Th2-dominated immune response in canine steroid-responsive
meningitis-arteritis. Vet J 187: 260–266.
58. Schwartz M, Puff C, Stein VM, Baumgärtner W, Tipold A (2010) Marked MMP-2 transcriptional
up-regulation in mononuclear leukocytes invading the subarachnoidal space in aseptic
suppurative steroid-responsive meningitis-arteritis in dogs. Vet Immunol Immunopathol 133: 198–
206.
59. Rozen S, Skaletsky H (2000) Primer3 on the WWW for general users and for biologist
programmers. Methods Mol Biol 132: 365–386.
60. Huan J, Culbertson N, Spencer L, Bartholomew R, Burrows GG, et al. (2005) Decreased FOXP3
levels in multiple sclerosis patients. J Neurosci Res 81: 45–52.
61. Viglietta V, Baecher-Allan C, Weiner HL, Hafler DA (2004) Loss of functional suppression by
CD4+CD25+ regulatory T cells in patients with multiple sclerosis. J Exp Med 199: 971–979.
62. Tischner D, Weishaupt A, van den Brandt J, Müller N, Beyersdorf N, et al. (2006) Polyclonal
expansion of regulatory T cells interferes with effector cell migration in a model of multiple
sclerosis. Brain 129: 2635–2647.
63. Lanteri MC, Brien KMO, Purtha WE, Cameron MJ, Lund JM, et al. (2009) Tregs control the
development of symptomatic West Nile virus infection in humans and mice. J Clin Invest 119:
3266-3277.
64. Lühn K, Simmons CP, Moran E, Dung NTP, Chau TNB, et al. (2007) Increased frequencies of
CD4+ CD25(high) regulatory T cells in acute dengue infection. J Exp Med 204: 979–985.
65. Liesz A, Suri-Payer E, Veltkamp C, Doerr H, Sommer C, et al. (2009) Regulatory T cells are key
cerebroprotective immunomodulators in acute experimental stroke. Nat Med 15: 192–199.
66. Beyersdorf N, Gaupp S, Balbach K, Schmidt J, Toyka KV, et al. (2005) Selective targeting of
regulatory T cells with CD28 superagonists allows effective therapy of experimental autoimmune
encephalomyelitis. J Exp Med 202: 445–455.
67. Liu J, Gong N, Huang X, Reynolds AD, Mosley RL, et al. (2009) Neuromodulatory activities of
CD4+CD25+ regulatory T cells in a murine model of HIV-1 associated neurodegeneration. J
Immunol 182: 3855–3865.
68. Fritzsching B, Haas J, König F, Kunz P, Fritzsching E, et al. (2011) Intracerebral human
regulatory T cells: analysis of CD4+ CD25+ FOXP3+ T cells in brain lesions and cerebrospinal
fluid of multiple sclerosis patients. PloS one 6: e17988.

62
Chapter 3: Regulatory T cells in CDV infection
69. Zhao W, Beers DR, Liao B, Henkel JS, Appel SH (2012) Regulatory T lymphocytes from ALS
mice suppress microglia and effector T lymphocytes through different cytokine-mediated
mechanisms. Neurobiol Dis 48: 418–428.
70. Beers DR, Henkel JS, Zhao W, Wang J, Huang A, et al. (2011) Endogenous regulatory T
lymphocytes ameliorate amyotrophic lateral sclerosis in mice and correlate with disease
progression in patients with amyotrophic lateral sclerosis. Brain 134: 1293–1314.
71. Tiemessen MM, Jagger AL, Evans HG, Herwijnen MJCV, John S, et al. (2007) CD4+ CD25+
Foxp3+ regulatory T cells induce alternative activation of human monocytes / macrophages. Proc
Natl Acad Sci U S A 104: 19446–19451.
72. Zhao J, Zhao J, Fett C, Trandem K, Fleming E, et al. (2011) IFN-γ- and IL-10-expressing virus
epitope-specific Foxp3(+) T reg cells in the central nervous system during encephalomyelitis. J
Exp Med 208: 1571–1577.
73. Liu Y, Teige I, Birnir B, Issazadeh-Navikas S (2006) Neuron-mediated generation of regulatory T
cells from encephalitogenic T cells suppresses EAE. Nat Med 12: 518–525.
74. Trajkovic V, Vuckovic O, Stosic-Grujicic S, Miljkovic D, Popadic D, et al. (2004) Astrocyte-induced
regulatory T cells mitigate CNS autoimmunity. Glia 47: 168–179.
75. Korn T, Reddy J, Gao W, Bettelli E, Awasthi A, et al. (2007) Myelin-specific regulatory T cells
accumulate in the CNS but fail to control autoimmune inflammation. Nat Med 13: 423–431.
76. Pasare C, Medzhitov R (2003) Toll pathway-dependent blockade of CD4+CD25+ T cell-mediated
suppression by dendritic cells. Science 299: 1033–1036.
77. Frisk AL, Baumgärtner W, Gröne A (1999) Dominating interleukin-10 mRNA expression induction
in cerebrospinal fluid cells of dogs with natural canine distemper virus induced demyelinating and
non-demyelinating CNS lesions. J Neuroimmunol 97: 102–109.
78. Anghelina D, Zhao J, Trandem K, Perlman S (2009) Role of regulatory T cells in coronavirus-
induced acute encephalitis. Virology 385: 358–367.
79. Schobesberger M, Summerfield A, Doherr MG, Zurbriggen A, Griot C (2005) Canine distemper
virus-induced depletion of uninfected lymphocytes is associated with apoptosis. Vet Immunol
Immunopathol 104: 33–44.
80. Gröne A, Frisk AL, Baumgärtner W (1998) Cytokine mRNA expression in whole blood samples
from dogs with natural canine distemper virus infection. Vet Immunol Immunopathol 65: 11–27.
81. Valencia X, Stephens G, Goldbach-Mansky R, Wilson M, Shevach EM, et al. (2006) TNF
downmodulates the function of human CD4+CD25hi T-regulatory cells. Blood 108: 253–261.
82. Hunt PW, Landay AL, Sinclair E, Martinson J a, Hatano H, et al. (2011) A low T regulatory cell
response may contribute to both viral control and generalized immune activation in HIV
controllers. PloS one 6: e15924.

63
Chapter 3: Regulatory T cells in CDV infection
83. Lin J, Li M, Wang Z, He S, Ma X, et al. (2010) The role of CD4+CD25+ regulatory T cells in
macrophage-derived foam-cell formation. J Lipid Res 51: 1208–1217.
84. Ryon JJ, Moss WJ, Monze M, Griffin DE (2002) Functional and Phenotypic Changes in
Circulating Lymphocytes from Hospitalized Zambian Children with Measles. Clin Diagn Lab
Immunol 9: 994–1003.
85. Carsillo M, Klapproth K, Niewiesk S (2009) Cytokine imbalance after measles virus infection has
no correlation with immune suppression. J Virol 83: 7244–7251.
86. Svitek N, von Messling V (2007) Early cytokine mRNA expression profiles predict Morbillivirus
disease outcome in ferrets. Virology 362: 404–410.
87. Cross AH, Waubant E (2011) MS and the B cell controversy. Biochim Biophys Acta 1812: 231–
238.
88. Ulrich R, Kalkuhl A, Deschl U, Baumgärtner W (2010) Machine learning approach identifies new
pathways associated with demyelination in a viral model of multiple sclerosis. J Cell Mol Med 14:
434–448.
89. Vandevelde M, Zurbriggen A, Steck A, Bichsel P (1986) Studies on the intrathecal humoral
immune response in canine distemper encephalitis. J Neuroimmunol 11: 41–51.
90. Meinl E, Krumbholz M, Derfuss T, Junker A, Hohlfeld R (2008) Compartmentalization of
inflammation in the CNS: a major mechanism driving progressive multiple sclerosis. J Neurol Sci
274: 42–44.
91. Massacesi L (2002) Compartmentalization of the immune response in the central nervous system
and natural history of multiple sclerosis. Implications for therapy. Clin Neurol Neurosurg 104:
177–181.
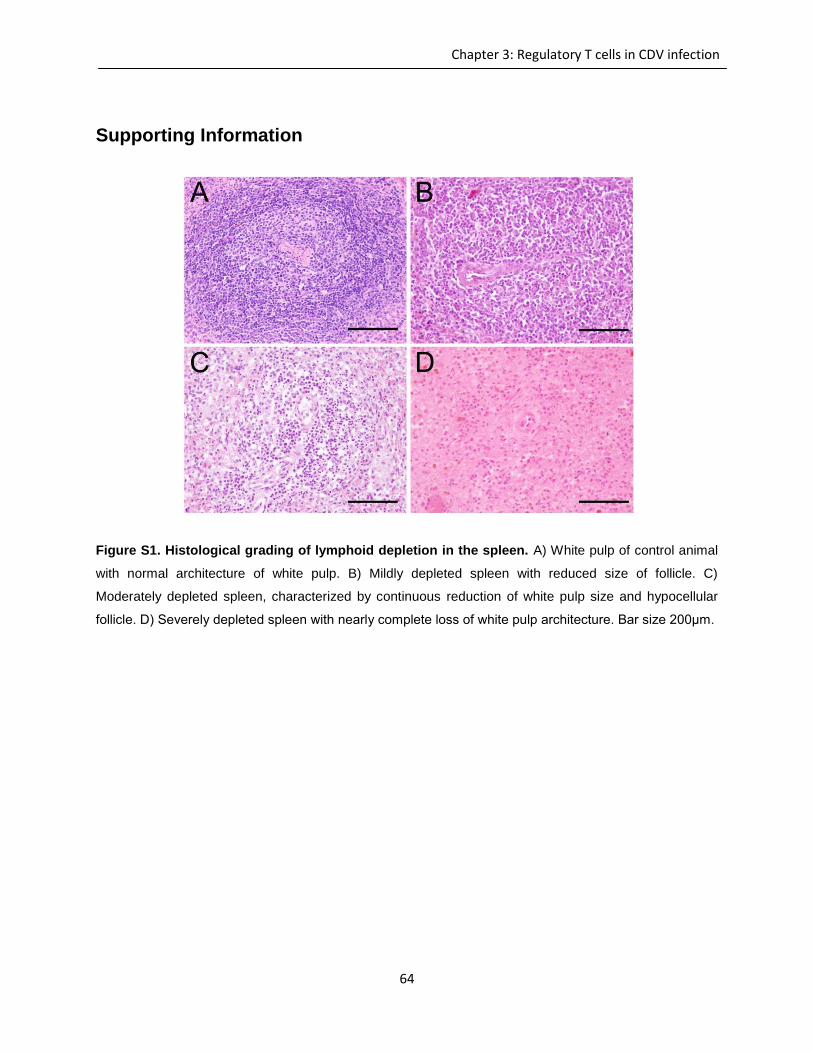
64
Chapter 3: Regulatory T cells in CDV infection
Supporting Information
Figure S1. Histological grading of lymphoid depletion in the spleen. A) White pulp of control animal
with normal architecture of white pulp. B) Mildly depleted spleen with reduced size of follicle. C)
Moderately depleted spleen, characterized by continuous reduction of white pulp size and hypocellular
follicle. D) Severely depleted spleen with nearly complete loss of white pulp architecture. Bar size 200μm.
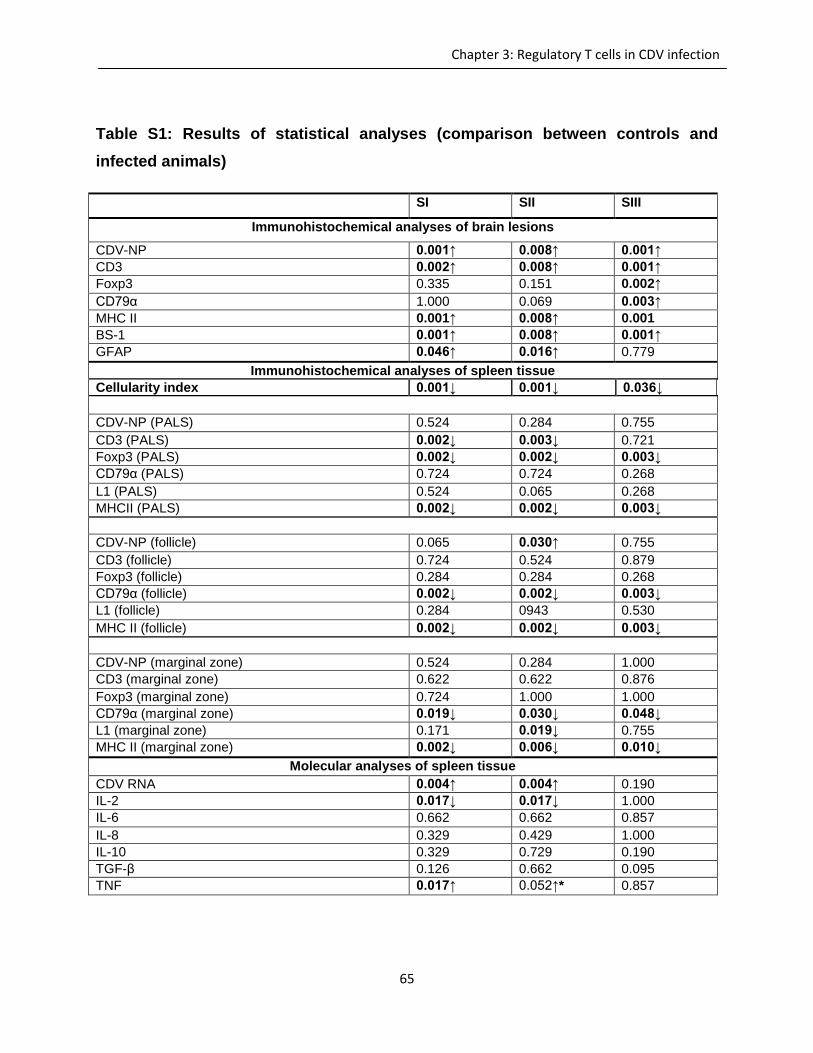
65
Chapter 3: Regulatory T cells in CDV infection
Table S1: Results of statistical analyses (comparison between controls and
infected animals)
SI SII SIII
Immunohistochemical analyses of brain lesions
CDV-NP 0.001↑ 0.008↑ 0.001↑
CD3 0.002↑ 0.008↑ 0.001↑
Foxp3 0.335 0.151 0.002↑
CD79α 1.000 0.069 0.003↑
MHC II 0.001↑ 0.008↑ 0.001
BS-1 0.001↑ 0.008↑ 0.001↑
GFAP 0.046↑ 0.016↑ 0.779
Immunohistochemical analyses of spleen tissue
Cellularity index 0.001↓ 0.001↓ 0.036↓
CDV-NP (PALS) 0.524 0.284 0.755
CD3 (PALS) 0.002↓ 0.003↓ 0.721
Foxp3 (PALS) 0.002↓ 0.002↓ 0.003↓
CD79α (PALS) 0.724 0.724 0.268
L1 (PALS) 0.524 0.065 0.268
MHCII (PALS) 0.002↓ 0.002↓ 0.003↓
CDV-NP (follicle) 0.065 0.030↑ 0.755
CD3 (follicle) 0.724 0.524 0.879
Foxp3 (follicle) 0.284 0.284 0.268
CD79α (follicle) 0.002↓ 0.002↓ 0.003↓
L1 (follicle) 0.284 0943 0.530
MHC II (follicle) 0.002↓ 0.002↓ 0.003↓
CDV-NP (marginal zone) 0.524 0.284 1.000
CD3 (marginal zone) 0.622 0.622 0.876
Foxp3 (marginal zone) 0.724 1.000 1.000
CD79α (marginal zone) 0.019↓ 0.030↓ 0.048↓
L1 (marginal zone) 0.171 0.019↓ 0.755
MHC II (marginal zone) 0.002↓ 0.006↓ 0.010↓
Molecular analyses of spleen tissue
CDV RNA 0.004↑ 0.004↑ 0.190
IL-2 0.017↓ 0.017↓ 1.000
IL-6 0.662 0.662 0.857
IL-8 0.329 0.429 1.000
IL-10 0.329 0.729 0.190
TGF-β 0.126 0.662 0.095
TNF 0.017↑ 0.052↑* 0.857

66
Chapter 3: Regulatory T cells in CDV infection
Table S1. Bold values display significant changes compared to control, ↑= significantly increased,↓=
significantly decreased compared to control; ↑* = statistical tendency compared to control; CD3 = T cells;
Foxp3 = regulatory T cells; CD79α = B cells; L1 = macrophages; MHC II = antigen presenting cells; IL =
interleukin; TNF = tumor necrosis factor; TGF-β = transforming growth factor-β

67
Chapter 4: Canine dendritic cells
4 Chapter 4: Canine dendritic cells
Review paper doi -10.1016/j.vetimm.2012.12.003
Species-specific properties and translational aspects of canine
dendritic cells
Qeska V.1,2, Baumgärtner W. 1,2, Beineke A. 1,*
1Department of Pathology, University of Veterinary Medicine Hannover, Bünteweg 17,
30559 Hannover, Germany
2Center for Systems Neuroscience Hannover
Abstract
Dogs are affected by spontaneously occurring neoplastic and inflammatory diseases
which often share many similarities with pathological conditions in humans and are thus
appreciated as important translational animal models. Dendritic cells (DCs) represent
the most potent antigen presenting cell population. Besides their physiological function
in the initiation of primary T cell responses and B cell immunity, a deregulation of DC
function is involved in immune-mediated tissue damage, immunosuppression and
transplantation complication in human and veterinary medicine. DCs represent a
promising new target for cancer immunotherapy in dogs. However, the therapeutic use
of canine DCs is restricted because of a lack of standardized isolation techniques and
limited information about dog-specific properties of this cell type. This article reviews
current protocols for the isolation and in vitro generation of canine monocyte- and bone

68
Chapter 4: Canine dendritic cells
marrow-derived DCs. DCs of dogs are characterized by unique morphological features,
such as the presence of cytoplasmic projections and periodic microstructures. Canine
DCs can be discriminated from other hematopoietic cells also based on phenotypic
properties and their high T cell stimulatory capability in mixed leukocyte reactions.
Furthermore, the classification of canine DC-derived neoplasms and the role of DCs in
the pathogeneses of selected infectious, allergic and autoimmune diseases, which
share similarities with human disorders, are discussed. Future research is needed to
expand the existing knowledge about DC function in canine diseases as a prerequisite
for the development of future therapies interfering with the immune response.
Keywords: dendritic cells, cytokines, phenotyping, functional assays, translational
research

69
Chapter 5: Dendritic cells in CDV infection
5 Chapter 5: Dendritic cells in canine distemper virus
infection
Canine distemper virus infection leads to reduced antigen
presenting function of monocyte-derived dendritic cells
V. Qeska1,2, Y. Barthel1, C. Urhausen3, V.M. Stein4, K. Rohn5, A. Tipold4,2, W.
Baumgärtner1,2, A. Beineke1
1Department of Pathology, University of Veterinary Medicine Hannover, Bünteweg 17,
D-30559 Hannover, Germany
2Center for Systems Neuroscience, Hannover, Germany
3Unit for Reproductive Medicine, Small Animal Clinic, University of Veterinary Medicine
Hannover, Bünteweg 15, D-30559 Hannover, Germany
4Department of Small Animal Medicine and Surgery, University of Veterinary Medicine
Hannover, Bünteweg 9, D-30559 Hannover, Germany.
5Department of Biometry, Epidemiology and Information Processing, University of
Veterinary Medicine Hannover, Bünteweg 2, D-30559 Hannover, German

70
Chapter 5: Dendritic cells in CDV infection
Abstract
Canine distemper virus (CDV) shows a profound lymphotropism that causes
immunosuppression and increased susceptibility of affected dogs to opportunistic
infections. Similar to human measles virus, CDV is supposed to inhibit terminal
differentiation of dendritic cells (DCs), responsible for disturbed repopulation of
lymphoid tissues and diminished antigen presenting function in dogs. In order to testify
the hypothesis that CDV-infection leads to an impairment of co-stimulatory functions of
professional antigen presenting cells, canine DCs have been generated from peripheral
blood monocytes in vitro and infected with CDV. Virus infection was confirmed and
quantified by transmission electron microscopy, CDV-specific immunofluorescence and
virus titration. Phenotypical changes of cultured cells were determined by flow
cytometry. In addition, apoptotic changes and cellular damage were quantified by the
terminal desoxynucleotidyl transferase-mediated dUTP-biotin nick end labeling-method
and lactate dehydrogenase assay, while cell proliferation was determined by 5-bromo-
2’-deoxyuridine-incorporation. Results demonstrated a significant time dependant
increase of the infectivity rate of DCs at 24, 72 and 120 hours post infection (hpi). As
observed by flow cytometry at 120 hpi, CDV-infection of canine DCs led to a down-
regulation of co-stimulatory molecules CD80 and CD86 as well as of the major
histocompatibility complex class II, indicative of disturbed antigen presenting properties.
As a potential mechanism to evade host immune responses, infected DCs showed no
evidence of apoptosis or cell lysis at 24, 72 and 120 hpi. Similarly, cell proliferation rate
of infected cells was unaffected at any investigated time points. These data suggest that
CDV-infection of DCs plays a role for pathogenesis of long lasting immune alterations
and virus persistence in canine distemper.

71
Chapter 5: Dendritic cells in CDV infection
1. Introduction
Canine distemper virus (CDV), a morbillivirus, causes persistent infection of peripheral
lymphoid organs and the central nervous system of carnivores (Krakowka 1982;
Beineke et al. 2009). Similar to measles virus (MV) infection of human beings clinical
findings in CDV-infected dogs include fever, rash, respiratory signs, and lymphopenia.
Affected dogs show an increased susceptibility to opportunistic infections as a
consequence of generalized lymphoid depletion and profound immunosuppression,
respectively (von Messling et al. 2006; Beineke et al. 2009; Sellin et al. 2009). Several
pathogens, including human herpesvirus type 1 as well as human and feline
immunodeficiency viruses, target DCs and have evolved strategies to modulate their
cytokine expression and antigen presenting capacity, thereby promoting virus immune
evasion and persistence (Steinman and Banchereau, 2007; Tompkins and Tompkins
2008). Additionally, the interaction between different viruses and DCs causes an
alteration of endocytosis, vesicle trafficking and immunological synapse formation, and
apoptosis induction of DCs (Klagge and Schneider-Schaulies 1999; Rinaldo and Piazza
2004; Harman et al. 2006; Schneider-Schaulies and Dittmer 2006; Cunningham et al.
2010). Immunosuppressive effects of MV is supposed to be a consequence of disturbed
function of antigen presenting cells, including DCs (Oldstone et al. 1999; Hahm et al.
2005; Trifilo et al. 2006; Zilliox et al. 2006). Moreover, following infection of the
respiratory tract, DCs might mediate viral transport to secondary lymphoid organs in
measles patients (Steinman 2007; Abt et al. 2009). MV-infection of DCs has been
demonstrated in vitro and in animal experiments using cotton rats, transgenic mice and
macaques. However, until now, DC infection by MV has not been confirmed in human
beings in vivo (Hahm et al. 2005; de Swart et al. 2007; Griffin 2010).
During the chronic disease stage of canine distemper, DCs seem to serve as the
primary host cells for the virus, which might promote viral persistence in lymphoid
organs (Wünschmann et al. 2000). This process is assumed to inhibit terminal
differentiation of DCs, responsible for disturbed repopulation of lymphoid tissues and

72
Chapter 5: Dendritic cells in CDV infection
diminished antigen presenting function in dogs, as suggested for MV-infection (Kerdiles
et al. 2006; Schneider-Schaulies and Dittmer 2006). In addition, CDV-infection of thymic
epithelial DCs may result in compromised maturation of T cells, promoting the release
of immature, potentially autoreactive lymphocytes, demonstrating a potential central role
of DCs in CDV-induced immunosuppression and immunopathology (Wünschmann et al.
2000). However, until now, the effect of CDV upon the function of canine DCs has not
been investigated. Thus, the aim of the present study was to investigate the
susceptibility of canine DCs for CDV and to testify the hypothesis that CDV-infection
leads to an impairment of co-stimulatory functions of canine DCs in vitro.
2. Materials and Methods
2.1 Generation of monocyte derived dendritic cells
Monocyte dendritic cells (moDCs) were generated as previously described, with minor
modifications (Yoshida et al. 2003; Ibisch et al. 2005; Bonnefont-Rebeix et al. 2006).
Briefly, 20ml of fresh heparinized blood was taken from clinically healthy dogs (n=16).
Peripheral blood mononuclear cells (PBMC) were obtained by density gradient
centrifugation (500xg) using Histopaque®-1077 (Sigma-Aldrich, Germany) at room
temperature for 30 minutes. PBMC were carefully collected from the interface, washed
twice with phosphate buffered saline (PBS) + 0.02% EDTA, counted and adjusted to a
concentration of 2x106 cells/ml, seeded in RPMI 1640 medium (PAA, Austria)
supplemented with 100UI/ml penicillin, 100µg/ml streptomycin and 10% fetal calf serum
(FCS) and incubated for 24h at 37°C and 5%CO2. Subsequently, non-adherent cells
were removed by gentle washing with PBS and adherent cells were incubated under
standard conditions for 6 more days, supplemented with 10.6µg/ml recombinant human
(rh) GM-CSF (RND Systems, MN, USA) and 20µg/ml recombinant canine (rc) IL-4
(RND Systems, MN, USA). Fresh medium was added every third day. During medium

73
Chapter 5: Dendritic cells in CDV infection
change, non-adherent cells were collected, centrifuged, and supernatant (SNT) was
discarded. Diluted cell pellet was transferred back to the culture flask.
2.2 Immuophenotyping of dendritic cells
Antibodies used for phenotypic analyses were either cell surface markers specific for
dog or cross-reacting with canine antigens. Monoclonal antibodies CD1a, CD11c,
CD14, CD80, CD86, and major histocompatibility complex (MHC) II were used for
labeling monocytes and DCs, respectively, while CD3- and CD21-specific antibodies
were used for labeling T cells and B cells, respectively. IgG1 and IgG2a were used as
isotype controls. All antibodies used for phenotypic analysis are listed in table 1.
Monocytes and DCs were stained as already described (Stein et al. 2004). Briefly, after
the cultivation period cells were collected, centrifuged (250xg, 4°C, 10 minutes) and
washed. Non-specific antibody binding was blocked by pretreatment of cells with 10
mg/ml human normal IgG (Globuman Berna, Switzerland). Primary antibodies and
isotype controls were incubated for 30 minutes at 4°C and afterwards centrifuged
(250xg, 20°C, 10 minutes) and washed twice with cell wash solution (BD Dickinson,
Germany). Cells labeled with conjugated markers (CD14, CD21 and IgG2a) were
resuspended in FACS flow solution (BD Dickinson, Germany) and stored at 4°C until
use. Non-conjugated primary antibodies were incubated with the secondary antibody
(goat anti-mouse pycoerythrin; Jackson ImmunoResearch, Dianova, Germany) for 30
minutes at 4°C, followed by centrifugation (250xg, 20°C, 10 minutes) and two washing
steps. Subsequently, cells were resuspended in FACS flow solution and analyzed
immediately. Cells were gated using forward scatter height (FSC-H) and side scatter
height (SSC-H) not exceeding 2% positive staining with serotypes. Phenotyping of cells
was performed using the FACSCalibur flow cytometer (Becton Dickinson, CA, USA) and
data were analyzed with FlowJo software (Tree star, OR, USA). Monocytes were
analyzed at day one in culture after the cell adherence and DCs were analyzed at day
seven in culture
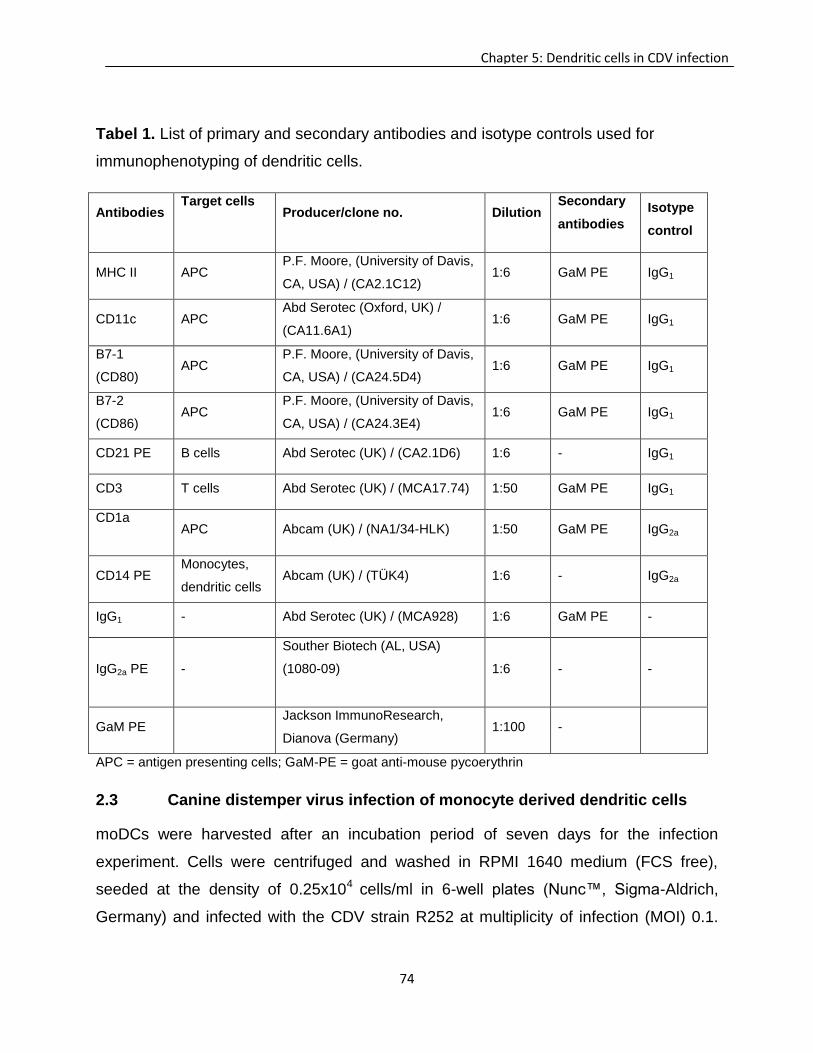
74
Chapter 5: Dendritic cells in CDV infection
Tabel 1. List of primary and secondary antibodies and isotype controls used for
immunophenotyping of dendritic cells.
Antibodies Target cells
Producer/clone no. Dilution Secondary
antibodies
Isotype
control
MHC II APC P.F. Moore, (University of Davis,
CA, USA) / (CA2.1C12) 1:6 GaM PE IgG1
CD11c APC Abd Serotec (Oxford, UK) /
(CA11.6A1) 1:6 GaM PE IgG1
B7-1
(CD80) APC
P.F. Moore, (University of Davis,
CA, USA) / (CA24.5D4) 1:6 GaM PE IgG1
B7-2
(CD86) APC
P.F. Moore, (University of Davis,
CA, USA) / (CA24.3E4) 1:6 GaM PE IgG1
CD21 PE B cells Abd Serotec (UK) / (CA2.1D6) 1:6 - IgG1
CD3 T cells Abd Serotec (UK) / (MCA17.74) 1:50 GaM PE IgG1
CD1a
APC Abcam (UK) / (NA1/34-HLK) 1:50 GaM PE IgG2a
CD14 PE Monocytes,
dendritic cells Abcam (UK) / (TÜK4) 1:6 - IgG2a
IgG1 - Abd Serotec (UK) / (MCA928) 1:6 GaM PE -
IgG2a PE -
Souther Biotech (AL, USA)
(1080-09)
1:6 - -
GaM PE Jackson ImmunoResearch,
Dianova (Germany) 1:100 -
APC = antigen presenting cells; GaM-PE = goat anti-mouse pycoerythrin
2.3 Canine distemper virus infection of monocyte derived dendritic cells
moDCs were harvested after an incubation period of seven days for the infection
experiment. Cells were centrifuged and washed in RPMI 1640 medium (FCS free),
seeded at the density of 0.25x104 cells/ml in 6-well plates (Nunc™, Sigma-Aldrich,
Germany) and infected with the CDV strain R252 at multiplicity of infection (MOI) 0.1.

75
Chapter 5: Dendritic cells in CDV infection
After an incubation period of 2 hours under standard conditions (37°C, 5%CO2), cells
were centrifuged and washed twice with PBS to remove unbound virus. Subsequently
fresh conditioned medium containing cytokines (rhGM-CSF, rcIL-4) and 10% FCS was
added to the culture. Medium was changed every third day as described above.
Infectivity rate of the cells was investigated at 24, 72 and 120 hours post infection (hpi).
2.4 Immunofluorescence
For the detection of infected cells and quantification of the infectivity rate, respectively,
cells were labeled using a monoclonal mouse anti-CDV-specific antibody (D110; dilution
1:200; A. Zurbriggen, Switzerland). Briefly, cells were transferred to a glass slide by
cytospin centrifugation (250xg, 5 minutes). After fixation with paraformaldehyde 4% for
30 minutes at room temperature, cells were washed with phosphate buffered saline
Triton X (PBST). Non-specific blocking was performed with goat and horse serum (5%
each) for 20 minutes. Subsequently, cells were incubated with the primary antibody
(dilution 1:100) for 4 hours at room temperature, followed by incubation with the
secondary antibody (goat anti-mouse Cy3; 1:100; Jackson, ImmunoResearch, Dianova,
Germany) for 1 hour at room temperature in a dark chamber. For counterstaining cells
were incubated with bisbenzimidine (1:100; Sigma-Aldrich, Germany) for 15 minutes at
room temperature. The percentages of CDV-infected cells were determined at 24, 72
and 120 hpi in duplicates (n=4 for each time point) by immunofluorescence microscopy
(Olympus IX-70, Olympus Life Science Europe GmbH, Germany).
2.5 Virus titration
At 24, 72 and 120 hpi, the SNT was harvested to calculate the 50% log10 tissue culture
infectious dose/ml (TCID50/ml) of cell free SNT of CDV-infected cells. Briefly, SNT was
centrifuged at 300xg at 4°C, aliquoted and stored at -80°C until used. SNT was diluted
logarithmically from 100 to 10-8 in RPMI 1640 medium containing 10% FCS and titrated

76
Chapter 5: Dendritic cells in CDV infection
in 96-well microtiter plates (Nunc™, Sigma-Aldrich, Germany) containing
Vero.dogSLAM cells (1.5x104 cells/well). After an incubation period of five days under
standard conditions cells were examined and evaluated for presence of cytopathogenic
effects. The TCID50 was calculated as described (Frisk et al. 1999; Techangamsuwan et
al. 2009). All samples were evaluated in triplicates (n=5; for each time point).
2.6 Proliferation assay
The proliferation rate was determined in control samples (non-infected DCs) and CDV-
infected DCs at 24, 72 and 120hpi in duplicates by the 5-bromo-2’-deoxyuridine (BrdU)-
assay (In Situ Proliferation Kit, FLUOS, Roche, Switzerland) following the
manufacturer’s instruction. Briefly, infected and non-infected cells were incubated
with10μM BrdU for 12 hours (37°C, 5%CO2). After centrifugation (200xg, 5 minutes)
cells were washed and fixated for 30 minutes (4°C) with ethanol fixative (3 volumes of
glycine solution [50mM, pH 2.0] with 7 volumes of ethanol). Subsequently, cells were
denaturated with 4M HCl for 20 minutes at room temperature. Denaturation was
stopped by adding 2ml PBS and centrifugation (300xg, 10 minutes). Non-specific
antibody binding was blocked with goat and horse serum (5% each) for 20 minutes and
incubation with 50μl anti-BrdU-FLUOS-antibody for 45 minutes at room temperature.
Cells were counterstained by bisbenzimidine (1:100, Sigma-Aldrich, Germany) for 15
minutes at room temperature. For the detection and quantification of proliferating cells,
ten randomly chosen areas (40x magnification; n=4 for each time point) were evaluated
by immunofluorescence microscopy (Olympus IX-70; Olympus Life Science Europe
GmbH, Germany).
2.7 Detection and quantification of apoptosis
Apoptotic cells were identified by the terminal desoxynucleotidyl transferase-mediated
dUTP-biotin nick end labeling (TUNEL)-method (ApopTag® Plus Peroxidase In Situ

77
Chapter 5: Dendritic cells in CDV infection
Apoptosis Detection Kit, Chemicon, MA, USA) according to the manufacturer’s
instructions (Urhausen et al. 2011). Briefly, infected and non-infected DCs were
transferred to glass slides by cytospin centrifugation (250xg, 5 minutes). Cells were
fixated with 1% paraformaldehyde for 10 minutes at room temperature and post-fixated
with ice cold (-20°C) ethanol : acetic acid (2:1) for 5 minutes. Subsequently, endogenous
peroxidase was quenched with 3% H2O2 for 5 minutes followed by 2 washing steps in
PBS. After short incubation (10 seconds) in washing equilibration buffer, the terminal
desoxynucleotidyl transferase (TdT) enzyme (working strength TdT enzyme) was added
and samples were incubated in a humidified chamber for 1 hour. Reaction was stopped
by transferring the slides into working strength stop/wash buffer and three subsequent
washing steps. Afterwards, anti-dioxigenin conjugate was applied and slides were
incubated at room temperature for 30 minutes. Following cytospin slides were washed
four times in PBS and incubated at room temperature in diaminobenzidin-
tetrahydrochloride peroxidase substrate for 5 minutes. Mayer’s hematoxylin was used
for nuclear counterstaining following dehydration in a series of graded alcohols. Slides
were mounted under coverslip and ten randomly chosen areas (40x magnification; n=4
for each time point) were investigated by light microscopy. Mouse spleen tissue served
as a positive control, while DC cytospin slides that lacked TdT enzyme was used as
negative controls.
2.8 Lactate dehydrogenase assay
For detecting cell lysis, a lactate dehydrogenase assay (LDH; CytoTox 96® Non-
radioactive Cytotoxicity Assay assay Promega, WI, USA) was performed according to
manufacturer`s instructions. Briefly, after centrifugation of cells, SNT from non-infected
and CDV-infected cells at 24, 72, and 120 hpi were carefully collected and stored at -
80°C until further use. 50μl of SNT and 50μl of substrate mix were added to a 96-well
microtiter plate, and incubated for 30 minutes. Reaction was stopped with 1M acetic
acid (stop solution) and absorbance was measured at 490nm using an ELISA reader

78
Chapter 5: Dendritic cells in CDV infection
(Fluorostat Optima, BMG LABTECH, Germany). Data (n=4 for each time point) were
analyzed using the Optima data analysis software (BMG LABTECH, Germany).
2.9 Transmission electron microscopy
Isolated monocytes at day one and generated DCs at day seven in culture were
centrifuged (250xg, 4°C, 10 minutes) and collected in 1.5ml tubes, while non-infected
and CDV-infected DCs at 24, 72, and 120hpi in 6-well plates (Nunc™, Sigma-Aldrich,
Germany) were centrifuged (250xg, 4°C, 10 minutes). Subsequently cells were fixated
with 2.5% glutaraldehyde and incubated overnight at 4°C. Post-fixation was performed
in 1% aqueous osmium tetroxide and after 5 washes in cacodylate buffer (5 minutes
each) samples were dehydrated through series of graded alcohols and embedded in
Epon 812 medium. Semi-thin sections were cut at microtome (Ultracut Reichert-Jung,
Leica Microsystems, Germany) and stained with uranyl citrate for 15 minutes. After
eight washing steps samples were incubated with lead citrate for 7 minutes. Ultra-thin
sections were cut with a diamond knife (Diatome, PA, USA) and transferred to copper
grids. Samples were examined by a transmission electron microscope (EM 10C, Zeiss,
Germany).
2.10 Statistical analyses
To determine the distribution of data a Shapiro-Wilk test and visualization assessment
were performed. For not normally distributed values (phenotypic analyses, LDH assay)
a non-parametric test (Mann-Whitney U-test) was used. For normally distributed data
(evaluation of semi-thin sections, infectivity rate, BrdU-assay, TUNEL-assay) a student’s
T-test was performed. For statistical analyses and visualization of data the SPSS
software (IBM, NY, USA) was used.

79
Chapter 5: Dendritic cells in CDV infection
1 Results
3.1 Generation and characterization of monocyte derived dendritic cells in
culture
3.1.1. Morphological characterization
After seven days in culture in the presence of rcIL-4 and rhGM-CSF, isolated PBMC
showed a typical DC-like morphology with long cytoplasmic processes as demonstrated
by phase contrast microscopy (Fig.1A). In agreement with this, counting of different cell
types on semi-thin sections at day one in culture revealed that the majority of cells
represented monocytes (p=0.001) and only few lymphocytes and neutrophils were
detected (Fig.2A). In contrast, at day seven in culture the majority of generated cells
showed a DC-like morphology (Fig.1B) with long cytoplasmic processes, while the
amount of monocyte-like cells was comparatively low (p=0.001; Fig.2B). Comparing day
one and day seven a significant decrease of monocytes (p=0.001) and lymphocytes
(p=0.031) was observed over time (data not shown). At day one in culture individual
cells were identified as neutrophils, while no neutrophils were observed at day seven.
DC differentiation in vitro was confirmed by transmission electron microscopy which
revealed a typical DC-like morphology, including long cytoplasmic processes, abundant
golgi apparatus formation, only few lysosomes and the presence of periodical
microstructures as a distinct ultrastructural feature of canine monocyte derived DCs
(Fig.3).
3.1.2. Flow cytometry
Phenotypical analyses of PBMC (day one) and moDCs (day seven) were performed by
flow cytometry (Fig.4). The percentage of gated cells was determined to characterize
culture purity and the geometrical mean fluorescent intensity (GMFI) for the
quantification of surface marker expression of monocytes and DCs, respectively. The
majority of cultured cells at day one and day seven expressed CD14 and CD11c,
indicative of monocytic origin. Moreover, CD3+ T cells showed a significant decrease at
day seven compared to day one in culture (p=0.01). An increased percentage of cells
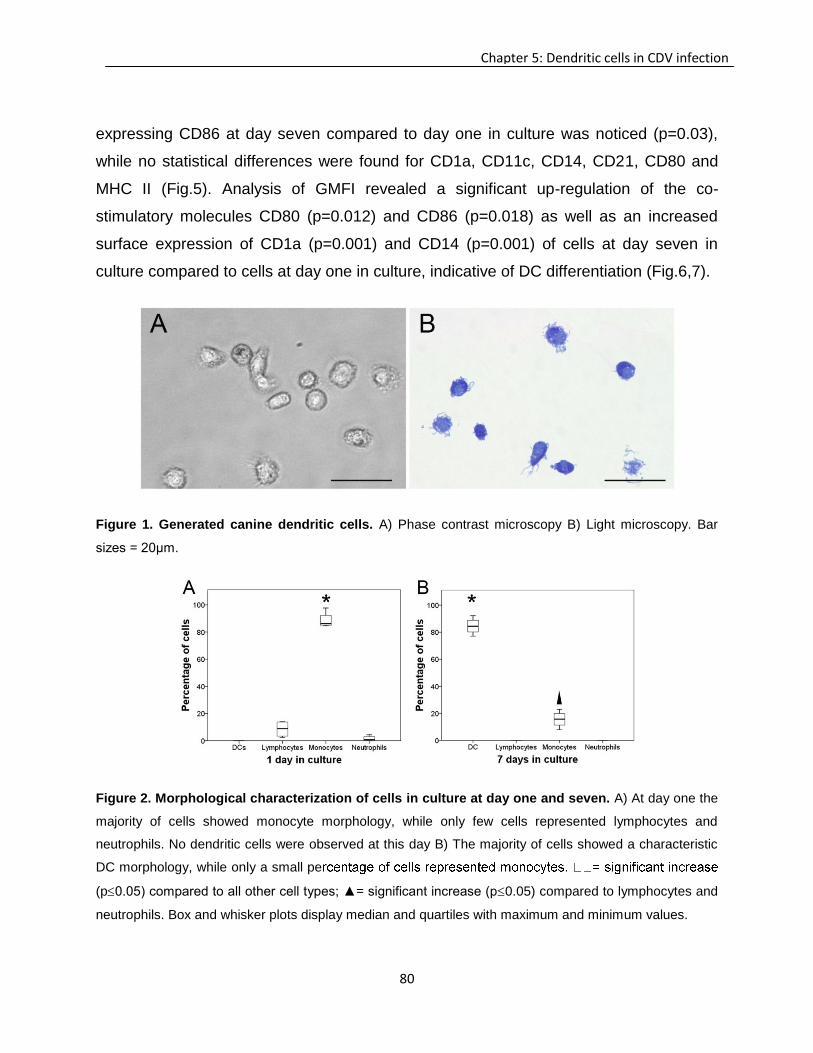
80
Chapter 5: Dendritic cells in CDV infection
expressing CD86 at day seven compared to day one in culture was noticed (p=0.03),
while no statistical differences were found for CD1a, CD11c, CD14, CD21, CD80 and
MHC II (Fig.5). Analysis of GMFI revealed a significant up-regulation of the co-
stimulatory molecules CD80 (p=0.012) and CD86 (p=0.018) as well as an increased
surface expression of CD1a (p=0.001) and CD14 (p=0.001) of cells at day seven in
culture compared to cells at day one in culture, indicative of DC differentiation (Fig.6,7).
Figure 1. Generated canine dendritic cells. A) Phase contrast microscopy B) Light microscopy. Bar
sizes = 20μm.
Figure 2. Morphological characterization of cells in culture at day one and seven. A) At day one the
majority of cells showed monocyte morphology, while only few cells represented lymphocytes and
neutrophils. No dendritic cells were observed at this day B) The majority of cells showed a characteristic
DC morphology, while only a small pe
(p0.05) compared to all other cell types; ▲= significant increase (p0.05) compared to lymphocytes and
neutrophils. Box and whisker plots display median and quartiles with maximum and minimum values.
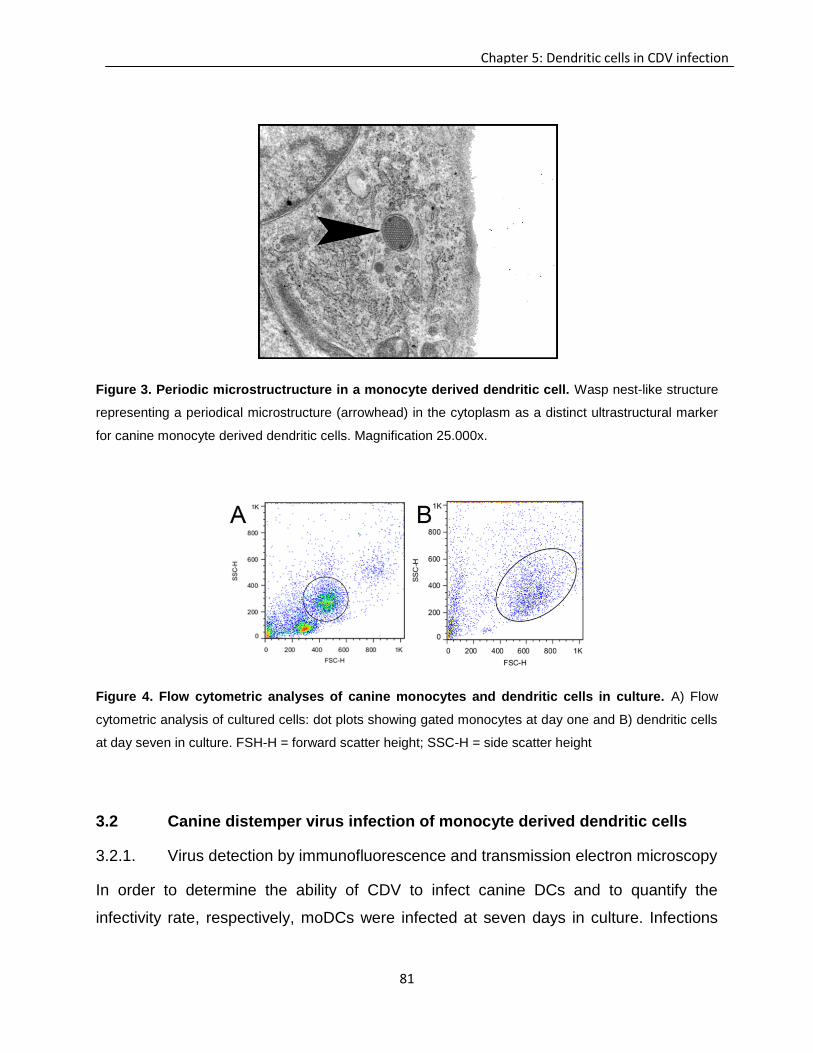
81
Chapter 5: Dendritic cells in CDV infection
Figure 3. Periodic microstructructure in a monocyte derived dendritic cell. Wasp nest-like structure
representing a periodical microstructure (arrowhead) in the cytoplasm as a distinct ultrastructural marker
for canine monocyte derived dendritic cells. Magnification 25.000x.
Figure 4. Flow cytometric analyses of canine monocytes and dendritic cells in culture. A) Flow
cytometric analysis of cultured cells: dot plots showing gated monocytes at day one and B) dendritic cells
at day seven in culture. FSH-H = forward scatter height; SSC-H = side scatter height
3.2 Canine distemper virus infection of monocyte derived dendritic cells
3.2.1. Virus detection by immunofluorescence and transmission electron microscopy
In order to determine the ability of CDV to infect canine DCs and to quantify the
infectivity rate, respectively, moDCs were infected at seven days in culture. Infections
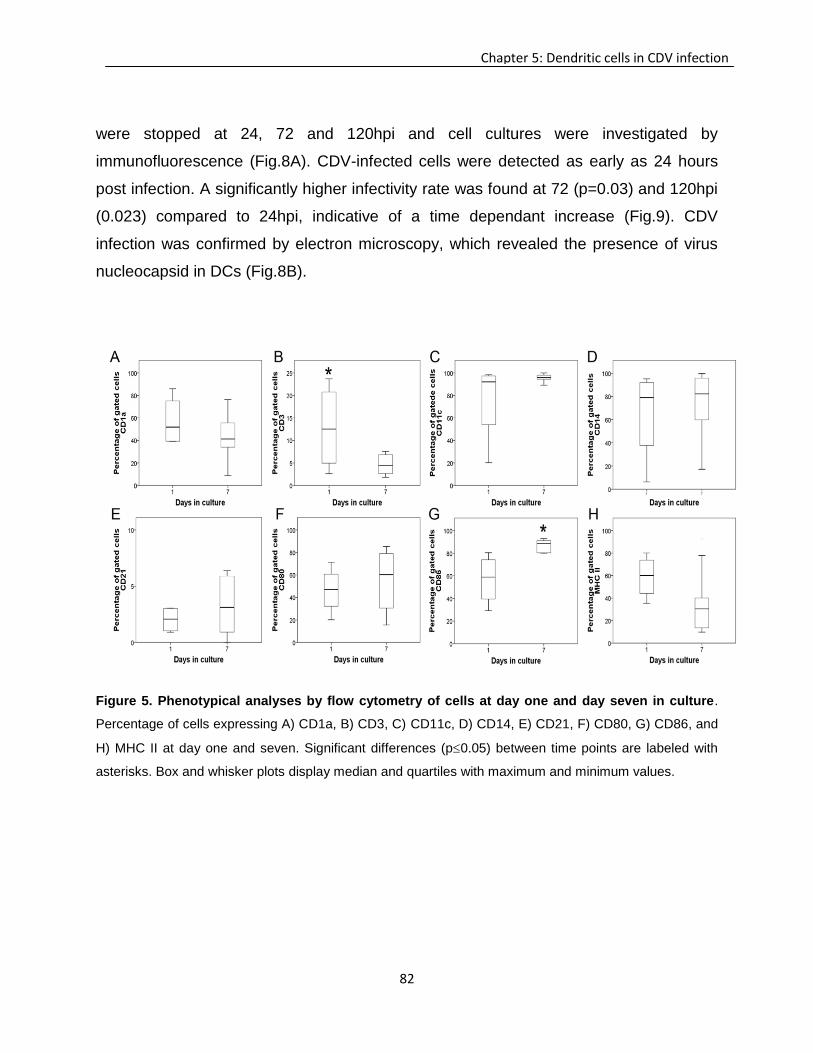
82
Chapter 5: Dendritic cells in CDV infection
were stopped at 24, 72 and 120hpi and cell cultures were investigated by
immunofluorescence (Fig.8A). CDV-infected cells were detected as early as 24 hours
post infection. A significantly higher infectivity rate was found at 72 (p=0.03) and 120hpi
(0.023) compared to 24hpi, indicative of a time dependant increase (Fig.9). CDV
infection was confirmed by electron microscopy, which revealed the presence of virus
nucleocapsid in DCs (Fig.8B).
Figure 5. Phenotypical analyses by flow cytometry of cells at day one and day seven in culture.
Percentage of cells expressing A) CD1a, B) CD3, C) CD11c, D) CD14, E) CD21, F) CD80, G) CD86, and
H) MHC II at day one and seven. Significant differences (p0.05) between time points are labeled with
asterisks. Box and whisker plots display median and quartiles with maximum and minimum values.
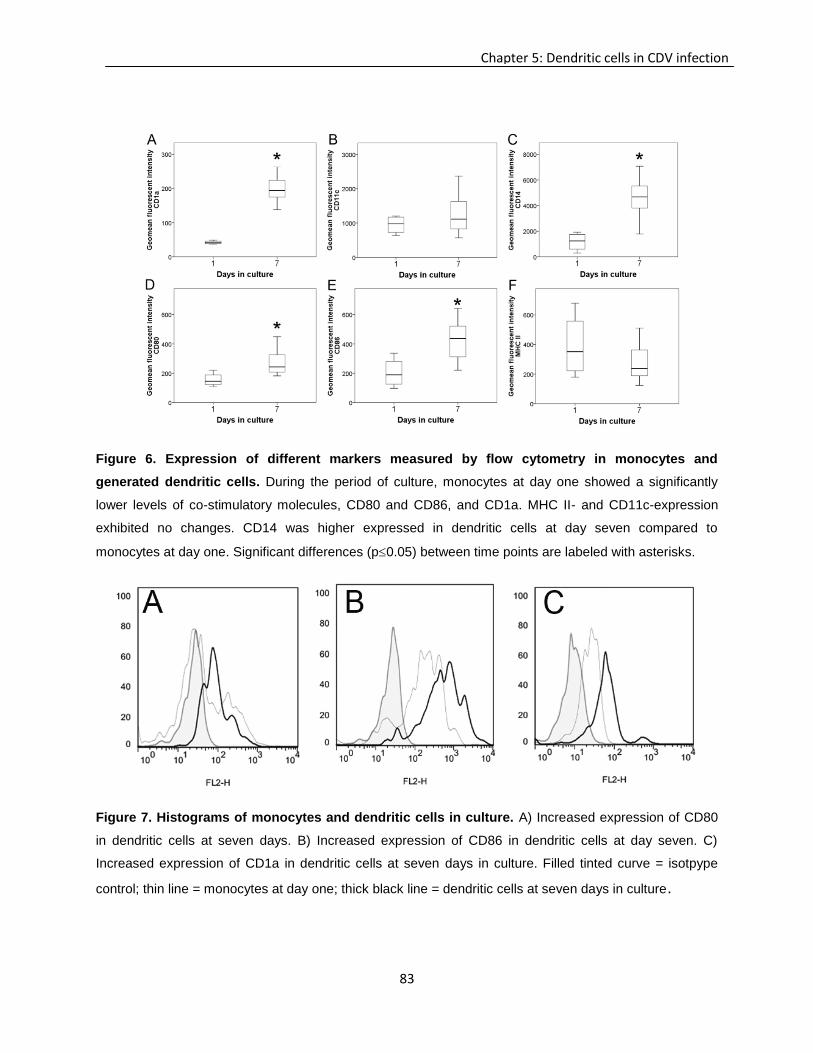
83
Chapter 5: Dendritic cells in CDV infection
Figure 6. Expression of different markers measured by flow cytometry in monocytes and
generated dendritic cells. During the period of culture, monocytes at day one showed a significantly
lower levels of co-stimulatory molecules, CD80 and CD86, and CD1a. MHC II- and CD11c-expression
exhibited no changes. CD14 was higher expressed in dendritic cells at day seven compared to
monocytes at day one. Significant differences (p0.05) between time points are labeled with asterisks.
Figure 7. Histograms of monocytes and dendritic cells in culture. A) Increased expression of CD80
in dendritic cells at seven days. B) Increased expression of CD86 in dendritic cells at day seven. C)
Increased expression of CD1a in dendritic cells at seven days in culture. Filled tinted curve = isotpype
control; thin line = monocytes at day one; thick black line = dendritic cells at seven days in culture.
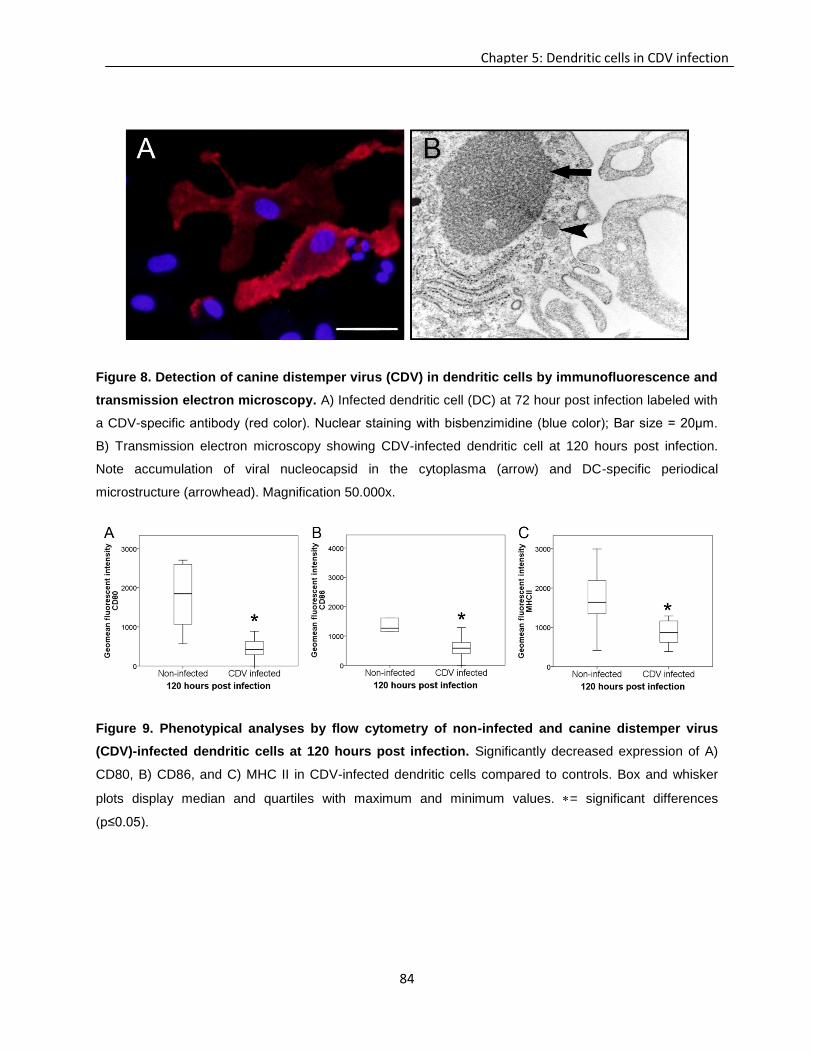
84
Chapter 5: Dendritic cells in CDV infection
Figure 8. Detection of canine distemper virus (CDV) in dendritic cells by immunofluorescence and
transmission electron microscopy. A) Infected dendritic cell (DC) at 72 hour post infection labeled with
a CDV-specific antibody (red color). Nuclear staining with bisbenzimidine (blue color); Bar size = 20μm.
B) Transmission electron microscopy showing CDV-infected dendritic cell at 120 hours post infection.
Note accumulation of viral nucleocapsid in the cytoplasma (arrow) and DC-specific periodical
microstructure (arrowhead). Magnification 50.000x.
Figure 9. Phenotypical analyses by flow cytometry of non-infected and canine distemper virus
(CDV)-infected dendritic cells at 120 hours post infection. Significantly decreased expression of A)
CD80, B) CD86, and C) MHC II in CDV-infected dendritic cells compared to controls. Box and whisker
plots display median and quartiles with maximum and minimum values. = significant differences
(p≤0.05).
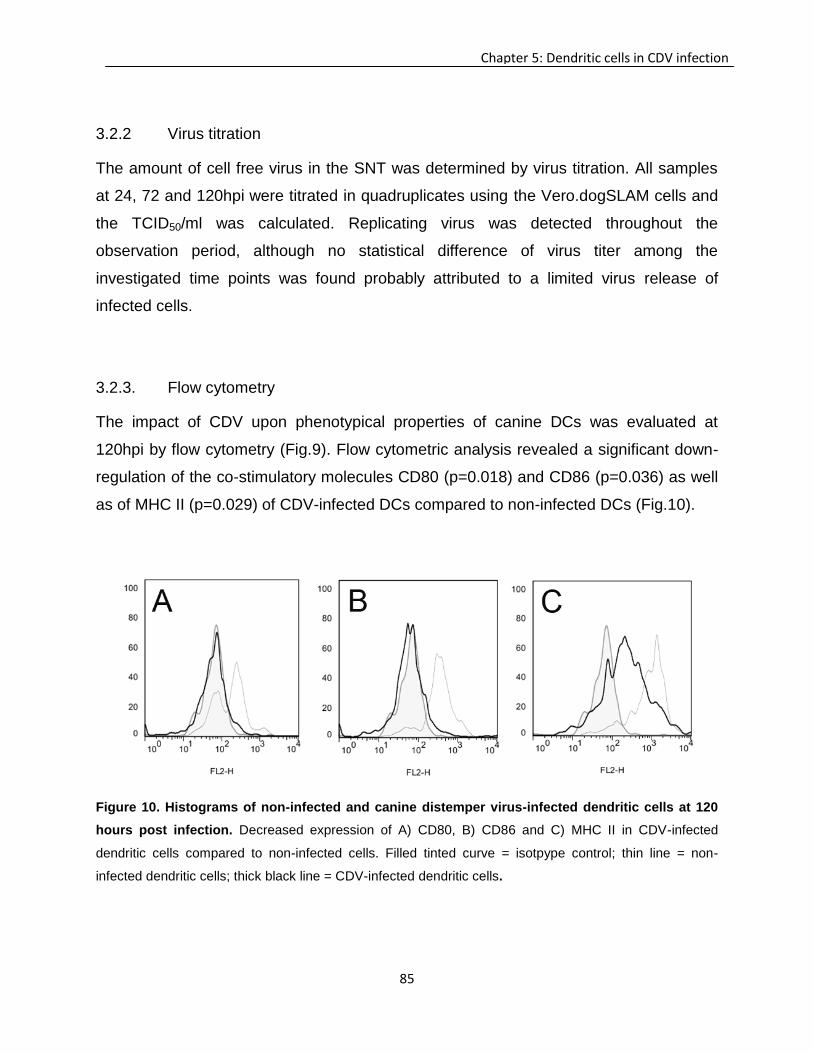
85
Chapter 5: Dendritic cells in CDV infection
3.2.2 Virus titration
The amount of cell free virus in the SNT was determined by virus titration. All samples
at 24, 72 and 120hpi were titrated in quadruplicates using the Vero.dogSLAM cells and
the TCID50/ml was calculated. Replicating virus was detected throughout the
observation period, although no statistical difference of virus titer among the
investigated time points was found probably attributed to a limited virus release of
infected cells.
3.2.3. Flow cytometry
The impact of CDV upon phenotypical properties of canine DCs was evaluated at
120hpi by flow cytometry (Fig.9). Flow cytometric analysis revealed a significant down-
regulation of the co-stimulatory molecules CD80 (p=0.018) and CD86 (p=0.036) as well
as of MHC II (p=0.029) of CDV-infected DCs compared to non-infected DCs (Fig.10).
Figure 10. Histograms of non-infected and canine distemper virus-infected dendritic cells at 120
hours post infection. Decreased expression of A) CD80, B) CD86 and C) MHC II in CDV-infected
dendritic cells compared to non-infected cells. Filled tinted curve = isotpype control; thin line = non-
infected dendritic cells; thick black line = CDV-infected dendritic cells.

86
Chapter 5: Dendritic cells in CDV infection
3.2.4 Quantification of cell proliferation, apoptosis and lysis
To determine the possible effect of CDV upon cell division the proliferation rate of
infected and non-infected DCs was measured by BrdU-incorporation. Although
proliferating cells (BrdU+ cells) were present at 24, 72 and 120hpi, no statistical
differences of proliferation rates were found between non-infected and CDV-infected
DCs at any time point.
TUNEL-assay was performed to detect apoptotic changes at 24, 72 and 120hpi.
Strikingly, similar apoptotic rates of infected DCs and non-infected cells were observed
(data not shown). The absence of an overt apoptosis induction and cell damage
following infection was confirmed by LDH-assay, which revealed no statistical
differences between infected and non-infected cells at any time point (data not shown).
4 Discussion
Results of the present study demonstrate the ability of CDV to infect canine DCs and to
modulate their antigen presenting properties, which has the potential to influence host
innate and adaptive immune responses. Many viruses have developed strategies to
alter antigen presentation or allostimulatory properties of DCs in order to evade immune
responses, as described for influenza virus, HIV, and herpes simplex virus (Mikloska et
al. 2001; Rinaldo and Piazza, 2004; Granucci, 2005). Furthermore, infection of DCs is
supposed to account for long lasting and profound immunosuppression in measles
patients (Fugier-Vivier et al. 1997; Murabayashi et al. 2002; Ohgimoto et al. 2007).
Similarly, CDV-mediated modulation of DC function might represent a mechanism to
suppress protective immunity, which favors persistent infection in infected dogs. In
agreement with this, Wünschmann et al. (2000) reported the occurrence of CDV-
infected DC-like cells within lymphoid organs in advanced stages of canine distemper.
Based on these observations in vivo and the present results in vitro it is hypothesized
that infection of DCs represents a mechanism of viral persistence in peripheral organs

87
Chapter 5: Dendritic cells in CDV infection
similar to observations in neurons and glial cells in canine distemper (Nesseler et al.
1997; Zurbriggen et al. 1998; Nesseler et al. 1999; Wünschmann et al. 2000; Beineke
et al. 2009; Céspedes et al. 2010).
CDV-infection of canine moDCs leads to a down-regulation of co-stimulatory molecules
(CD80 and CD86) as well as of MHC II, which might impair T cell activation in affected
dogs (Björck et al. 1997). According to this, in addition to lymphopenia, remaining
peripheral blood lymphocytes of dogs suffering from CDV infection show an extensively
reduced mitogen-induced lymphocyte proliferation (Krakowka et al. 1975; Krakowka and
Wallace, 1979). Besides reducing T cell responses, CDV inhibits IL-1 production and
increases prostaglandin E2 release, which impairs antigen presentation and subsequent
B cell differentiation, plasma cell formation and immunoglobulin production in CDV-
infected dogs (Krakowka et al. 1987). In addition, diminished T helper cell function as a
consequence of impaired antigen presentation in persistently infected dogs might
contribute also to disturbed germinal center formation and a reduced class switch from
IgM to IgG (Winters et al. 1983).
Dysregulation of antigen presenting properties is supposed to account for
immunosuppression in human measles (Servet-Delprat et al. 2000). Mechanisms
include the down-regulation of IL-12, which leads to a failure to activate T cells by DCs
(Fugier-Vivier et al. 1997; Servet-Delprat et al. 2000; Schneider-Schaulies et al. 2002).
Possibly, CDV nucleoprotein acts indirectly on T cell function by modulating antigen
presentation of DCs via interaction with the Fc receptor (CD32) and subsequent
diminished IL-12 production (Schneider-Schaulies and Dittmer, 2006). Similar to the
findings of the present study, MV-infection of mice causes a down-regulation of co-
stimulatory molecules as well as of MHC class I and II molecules in splenic DCs
(Oldstone et al. 1999; Hahm et al. 2005; Trifilo et al. 2006). However, in contrast to the
present results, MV-infection of human DCs in vitro leads to an up-regulation of co-
stimulatory molecules. These differences demonstrate that despite numerous
similarities between the pathogeneses of human measles and canine distemper virus-

88
Chapter 5: Dendritic cells in CDV infection
specific but also species-specific properties have to be considered for critical
interpretation of data in translational research.
Human cytomegalovirus, murine cytomegalovirus and Epstein-Barr virus infect and
manipulate DCs to circumvent cell death, which causes persistent infection (Rinaldo
and Piazza, 2004). According to this, the observed lack of apoptosis and lysis of CDV-
infected DCs represents a potential prerequisite for virus persistence in lymphoid
organs in chronically infected dogs. Furthermore, DCs have a limited lifespan and
defects in DC apoptosis might trigger autoimmune disease as demonstrated in
transgenic FasL- and Fas-deficient mice (Cohen and Eisenberg, 1992; Kushwah and
Hu, 2010). Referring to this, disturbed DC function in canine distemper is supposed to
compromise maturation and selection of T cells, promoting the release of immature CD5
negative T cells, including potentially autoreactive cells (Wünschmann et al. 2000).
In conclusion, for the first time CDV infection of canine DCs has been demonstrated in
vitro. Modulation of co-stimulatory molecules and reduced antigen presenting function of
CDV-infected DCs might account for inhibitory effects upon the immune system of dogs.
Getting insights into the interaction between viruses and DCs is fundamental to
understand the pathogeneses of infectious disorders, which has implication for
prevention (e.g. vaccination) and treatment strategies (Hou et al. 2007). Thus, further
studies are needed to investigate the role of DCs in the pathogenesis of
immunopathology and virus persistence in canine distemper.
Acknowledgment
The authors would like to thank Kerstin Schöne, Regina Carlson, Kerstin Rohn, Danuta
Waschke, Bettina Buck, and Petra Grünig for their excellent technical support during the
laboratory work. This study was supported by the German Research Foundation (FOR
1103, BA 815/10-2 and BE 4200/1-2).

89
Chapter 5: Dendritic cells in CDV infection
References:
Abt M, Gassert E, Schneider-Schaulies S, 2009. Measles virus modulates chemokine release and
chemotactic responses of dendritic cells. J Gen Virol 90, 909–914.
Beineke A, Puff C, Seehusen F, Baumgärtner W, 2009. Pathogenesis and immunopathology of systemic
and nervous canine distemper. Vet Immuno Immunopathol 127, 1–18.
Björck P, Banchereau J, Flores-Romo L, 1997. CD40 ligation counteracts Fas-induced apoptosis of
human dendritic cells. Int Immunol 9, 365–372.
Bonnefont-Rebeix C, Miranda de Carvalho C, Bernaud J, Chabanne L, Marchal T, Rigal D, 2006. CD86
molecule is a specific marker for canine monocyte-derived dendritic cells. Vet Immuno
Immunopathol 109, 167–176.
Cohen PL, Eisenberg RA, 1992. The Lpr and Gld genes in systemic autoimmunity: life and death in the
Fas lane. Immunol Today 13, 427–428.
Cunningham AL, Donaghy H, Harman AN, Kim M, Turville SG, 2010. Manipulation of dendritic cell
function by viruses. Curr Opin Microbiol 13, 524–529.
Céspedes PF, Cruz P, Navarro CO, 2010. Modulación de la respuesta inmune durante la infección por
virus distemper canino : implicancias terapéuticas y en el desarrollo de vacunas - Modulation of
immune response during canine distemper virus infection: therapeutic and vaccine development
implication. Arch Med Vet 28, 15–28.
De Swart RL, Ludlow M, de Witte L, Yanagi Y, van Amerongen G, McQuaid S, Yüksel S, Geijtenbeek
TBH, Paul Duprex W, Osterhaus ADME, 2007. Predominant infection of CD150+ lymphocytes
and dendritic cells during measles virus infection of macaques. PLoS Pathog 3, e178.
Frisk AL, Baumgärtner W, Gröne A, 1999. Dominating interleukin-10 mRNA expression induction in
cerebrospinal fluid cells of dogs with natural canine distemper virus induced demyelinating and
non-demyelinating CNS lesions. J Neuroimmunol 97, 102–109.
Fugier-Vivier I, Servet-Delprat C, Rivailler P, Rissoan MC, Liu YJ, Rabourdin-Combe C, 1997. Measles
virus suppresses cell-mediated immunity by interfering with the survival and functions of dendritic
and T cells. J Exp Med 186, 813–23.
Granucci F, Foti M, Ricciardi-Castagnoli P, 2005. Dendritic cell biology. Adv Immunol 88, 193–233.
Griffin DE, 2010. Measles virus-induced suppression of immune responses. Immunol Rev 236, 176–189.
Hahm B, Trifilo MJ, Zuniga EI, Oldstone MBA, 2005. Viruses evade the immune system through type I
Interferon-mediated STAT2-dependent, but STAT1-independent, signaling. Immunity 22, 247–
257.

90
Chapter 5: Dendritic cells in CDV infection
Harman AN, Wilkinson J, Bye CR, Bosnjak L, Stern JL, Nicholle M, Lai J, Cunningham AL, 2006. HIV
induces maturation of monocyte-derived dendritic cells and langerhans cells. J Immunol 177,
7103–7113.
Hou W, So E, Kim BS, 2007. Role of dendritic cells in differential susceptibility to viral demyelinating
disease. PLoS Pathog 3, e124.
Ibisch C, Pradal G, Bach JM, Lieubeau B, 2005. Functional canine dendritic cells can be generated in
vitro from peripheral blood mononuclear cells and contain a cytoplasmic ultrastructural marker. J
Immunol Methods 298, 175–182.
Kerdiles YM, Sellin CI, Druelle J, Horvat B, 2006. Immunosuppression caused by measles virus: role of
viral proteins. Rev Med Virol 16, 49–63.
Klagge IM, Schneider-Schaulies S, 1999. Virus interactions with dendritic cells. J Gen Virol 80, 823–833.
Krakowka S, Cockerell G, Koestner A, 1975. Effects of canine distemper virus infection on lymphoid
function in vitro and in vivo. Infec Immun 11, 1069–1078.
Krakowka S, Wallace AL, 1979. Lymphocyte-associated immune responses to canine distemper and
measles viruses in distemper-infected gnotobiotic dogs. Am J Vet Res 40, 669–672.
Krakowka S, 1982. Mechanisms of in vitro immunosuppression in canine distemper virus infection. J Clin
Lab Immunol 8, 187–196.
Krakowka S, Ringler SS, Lewis M, Olsen RG, Axthelm MK 1987. Immunosuppression by canine
distemper virus: modulation of in vitro immunoglobulin synthesis, interleukin release and
prostaglandin E2 production. Vet Immuno Immunopathol 15, 181–201.
Kushwah R, Hu J, 2010. Dendritic cell apoptosis: regulation of tolerance versus immunity. J Immunol,
185, 795–802.
von Messling V, Svitek N, Cattaneo R, 2006. Receptor (SLAM [CD150]) recognition and the V protein
sustain swift lymphocyte-based invasion of mucosal tissue and lymphatic organs by a
morbillivirus. J Virol 80, 6084–6092.
Mikloska Z, Bosnjak L, Cunningham AL, 2001. Immature monocyte-derived dendritic cells are
productively infected with herpes simplex virus Type 1. J Virol 75, 5958–5964.
Murabayashi N, Kurita-Taniguchi M, Ayata M, Matsumoto M, Ogura H, Seya T, 2002. Susceptibility of
human dendritic cells (DCs) to measles virus (MV) depends on their activation stages in
conjunction with the level of CDw150: role of toll stimulators in DC maturation and MV
amplification. Microb Infect 4, 785–794.
Nesseler A, Baumgärtner W, Gaedke K, Zurbriggen A, 1997. Abundant expression of viral nucleoprotein
mRNA and restricted translation of the corresponding viral protein in inclusion body
polioencephalitis of canine distemper. J Comp Pathol 116, 291–301.

91
Chapter 5: Dendritic cells in CDV infection
Nesseler A, Baumgärtner W, Zurbriggen A, Orvell C. 1999. Restricted virus protein translation in canine
distemper virus inclusion body polioencephalitis. Vet Microbiol 69, 23–28.
Ohgimoto K, Ohgimoto S, Ihara T, Mizuta H, Ishido S, Ayata M, Ogura H, Hotta H, 2007. Difference in
production of infectious wild-type measles and vaccine viruses in monocyte-derived dendritic
cells. Virus Res 123, 1–8.
Oldstone MBA, Lewicki H, Thomas D, Tishon A, Dales S, Patterson J, Manchester M, Homann D,
Naniche D, Holz A, 1999. Measles virus infection in a transgenic model: and central nervous
system disease. Cell 98, 629–640.
Rinaldo CR, Piazza P, 2004. Virus infection of dendritic cells: portal for host invasion and host defense.
Trends Microbiol 12, 337–345.
Schneider-Schaulies S, Bieback K, Avota E, Klagge I, ter Meulen V, 2002. Regulation of gene expression
in lymphocytes and antigen-presenting cells by measles virus: consequences for
immunomodulation. J Mol Med 80, 73–85.
Schneider-Schaulies S, Dittmer U, 2006. Silencing T cells or T-cell silencing: concepts in virus-induced
Iimunosuppression. J Gen Virol 87, 1423–1438.
Sellin CI, Jégou J-F, Renneson J, Druelle J, Wild TF, Marie JC, Horvat B, 2009. Interplay between virus-
specific effector response and Foxp3 regulatory T cells in measles virus immunopathogenesis.
PloS One 4, e4948.
Servet-Delprat C, Vidalain PO, Bausinger H, Manié S, Le Deist F, Azocar O, Hanau D, Fischer A,
Rabourdin-Combe C, 2000. Measles virus induces abnormal differentiation of CD40 ligand-
activated human dendritic cells. J Immunol 164, 1753–1760.
Stein VM, Czub M, Schreiner N, Moore PF, Vandevelde M, Zurbriggen A, Tipold A, 2004. Microglial cell
activation in demyelinating canine distemper lesions. J Neuroimmunol 153, 122–131.
Steinman RM, 2007. Dendritic cells: versatile controllers of the immune system. Nat Med 13, 1155–1159.
Steinman RM, Banchereau J, 2007. Taking dendritic cells into medicine. Nat 449, 419–426.
Techangamsuwan S, Haas L, Rohn K, Baumgärtner W, Wewetzer K, 2009. Distinct cell tropism of canine
distemper virus strains to adult olfactory ensheathing cells and schwann cells in vitro. Virus Res
144, 195–201.
Tompkins MB, Tompkins WA, 2008. Lentivirus-induced immune dysregulation. Vet Immuno
Immunopathol 123, 45–55.
Trifilo MJ, Hahm B, Zuniga EI, Edelmann KH, Oldstone MBA, 2006. Dendritic cell inhibition: memoirs from
immunosuppressive viruses. J Infect Dis 194 Suppl 1, S3–10.
Urhausen C, Beineke A, Piechotta X, Karre I, Beyerbach M, Günzel-Apel AR, 2011. Apoptosis in the
uterotubal junction and oviductal isthmus during the estrous cycle of the bitch. Anat Rec 294,
342–348.

92
Chapter 5: Dendritic cells in CDV infection
Winters KA, Mathes LE, Krakowka S, Olsen RG, 1983. Immunoglobulin class response to canine
distemper virus in gnotobiotic dogs. Vet Immuno Immunopathol 5, 209–215.
Wünschmann A, Kremmer E, Baumgärtner W, 2000. Phenotypical characterization of T and B cell areas
in lymphoid tissues of dogs with spontaneous distemper. Vet Immuno Immunopathol 73, 83–98.
Yoshida H, Momoi Y, Taga N, Ide K, Yamazoe K, Iwasaki T, Kudo T, 2003. Generation of canine
dendritic cells from peripheral blood mononuclear cells. J Vet Med Sci 65, 663–669.
Zilliox MJ, Parmigiani G, Griffin DE, 2006. Gene expression patterns in dendritic cells infected with
measles virus compared with other pathogens. Proc Natl Acad Sci USA 103, 3363–3368.
Zurbriggen A, Schmid I, Graber HU, Vandevelde M, 1998. Oligodendroglial pathology in canine
distemper. Acta Neuropathol 95, 71–77.

93
Chapter 6: General discussion
6 Chapter 6: General Discussion
Based upon the observation that immune alterations in canine distemper are a
consequence of direct and indirect viral effects and thus a disturbed immunomodulation
accounts for lesion development, the aims of the present study were (i) to investigate
phase-dependant changes of Treg and their association with peripheral immune
responses in canine distemper to testify the hypothesis that insufficient Treg numbers
contribute to uncontrolled neuroinflammation and early lesion initiation in the CNS of
infected dogs. Since DCs have been demonstrated to influence immune homeostasis in
several infectious diseases and immunopathologies (ii) current concepts of canine DCs
and their impact upon canine diseases have been reviewed. In addition, (iii) the impact
of CDV upon antigen presenting function of canine DCs and the proposed role of DCs in
virus persistence have been investigated in vitro.
6.1 The role of innate immunity in demyelinating disorders
Results of the present study demonstrate a lack of Treg infiltration in predemyelinating
and early demyelinating lesions in CDV-infected dogs, which represents a potential
cause for excessive neuroinflammation, including glial and innate immune responses,
respectively. Microglia and astrocytes contribute to innate immune responses within the
CNS. Together with infiltrating Natual Killer cells, granulocytes, macrophages and DCs,
resident glial cells initiate responses to various stimuli and thus represent an important
prerequisite for antigen-specific adaptive immune responses in neurological disorders
(Gandhi et al. 2010). Similar to observations in the present survey, microglial activation
can be observed in early demyelinating lesions in MS patients. However, it remains
undetermined whether microglia induces primary tissue damage or have to be activated
first by encephalitogenic T cells (Marik et al. 2007; Gandhi et al. 2010).
In canine distemper, microglial activation leads to bystander demyelination during the
progressive disease phase (Griot et al. 1990; Botteron et al. 1992). Early signs of

94
Chapter 6: General discussion
activation of resident CNS cells represent the prominent up-regulation of MHC II prior to
T and B cell infiltration in canine and phocine distemper (Beineke et al. 2008; Stimmer
et al. 2010). Referring to this, a dominance of pro-inflammatory cytokine environment in
the brain during the acute distemper phase is indicative of M1-polarization of microglia
(Beineke et al. 2008; Markus et al. 2002). This hypothesis is supported by the
observation of an increased expression of adhesion molecules, phagocytic activity and
release of reactive oxygen species by microglia in experimentally CDV-infected dogs
(Stein et al. 2004; Stein et al. 2008). In EAE and Theiler’s murine encephalomyelitis
(TME), M1-type microglial cells contribute to lesion development, while M2-type cells
exhibit immunomodulatory properties and attenuation of clinical signs (Mikita et al.
2011; Gerhauser et al. 2012). Thus activation of potentially neurotoxic microglial
populations might represent an initiating event for subsequent immune mediated tissue
damage of the cerebellar white matter in canine distemper. Similarly, a dominating role
of innate immunity in acute and subacute traumatic spinal cord injury has been
observed in dogs (Spitzbarth et al. 2011).
Similar to findings in canine distemper, the cerebral cortex is not primarily affected by
myelin loss in TME and phocine distemper (Stimmer et al. 2010; Kummerfeld et al.
2012). The reason for region-specific differences in demyelinating disorders remains
undetermined. Possible explanations for this phenomenon include topographical
differences of the CNS microenvironment or functionality of glial cells, such as a
reduced myelin degrading proteolytic activity of microglia as observed in C57BL/6 mice
(Liuzzi et al. 1995). Functional differences of microglial cells have also been observed in
different CNS regions of the dog (Stein et al. 2004; Stein et al. 2006; Stein et al. 2007;
Ensinger et al. 2010). Moreover, regional differences of the vulnerability of axons might
account for topographical differences of glial responses in virus-induced demyelinating
disorders, such as canine distemper and TME (Tsunoda and Fujinami 2002; Seehusen
and Baumgärtner 2010; Kummerfeld et al. 2012). Regional differences of myelin loss
can also be observed in toxic demyelination models, such as the cuprizone model
(Herder et al. 2011). While microglia and macrophages contribute to demyelination in

95
Chapter 6: General discussion
the corpus callosum (Biancotti et al. 2008), a delayed infiltration of CD107b+ microglial
cells and CD11b-mRNA-expression can be found in the spinal cord of cuprizone treated
mice (Herder et al. 2011). In contrast to the potential neurotoxic function of glial cells in
canine distemper, this process might represent a reaction to toxin-induced
oligodendroglial damage in the spinal cord, which potentially promotes tissue repair and
remyelination (M2-type microglia; Kotter et al. 2001; Mantovani et al. 2005; Kigerl et al.
2009). In analogy to the concept of region-specific lesion development in canine
distemper and human MS, recent studies have demonstrated topographical differences
of de- and remyelination within the brain of cuprizone fed mice, which might be
attributed to unequal densities or functions of microglia, astrocytes and oligodendocyte
progenitor cells in the respective CNS areas (Skripuletz et al. 2008; Gudi et al.
2009;Skripuletz et al. 2010).
The role of microglia in neurological disorders is discussed controversially, since both
detrimental and beneficial effects have been described. For instance, phagocytosis of
myelin debris by microglia and macrophages represent an important prerequisite for
regeneration following traumatic CNS injury (Yang and Schnaar, 2008), while excessive
cytokine expression and release of reactive oxygen species by these cells enhances
tissue damage (Banati and Kreutzberg, 1993; Stein et al. 2006; Ensinger et al. 2010;
Stein et al. 2011). As demonstrated in canine organotypic spinal cord slice cultures and
early MS lesions, an activation of microglia by phagocytosis of myelin debris leads to a
pro-inflammatory and potentially neurotoxic M1-phenotype of microglia (Pinteaux-Jones
et al. 2008; Spitzbarth et al. 2011). This process is supposed to be initiated by the
interaction between TLR on glial cells and the released cellular compounds (damage
associated molecular patterns) as a consequence of tissue damage (Kigerl et al. 2007;
Kigerl and Popovich, 2009).
Besides their function in innate immunity, microglia, macrophages, and DCs play a
pivotal role in the induction of specific immune responses in peripheral lymphoid organs.
In TME an early migration of CD68+-antigen presenting cells from the CNS to the

96
Chapter 6: General discussion
cervical lymph node can be observed (Navarrete-Talloni et al. 2010). Subsequently,
activated peripheral lymphocytes infiltrate the CNS and get reactivated by APCs in the
CNS (McMahon et al. 2005). In agreement with this, prominent MHC II expression in
early virus-induced lesions might account for antigen presenting capacity of glial cells in
canine and phocine distemper (Beineke et al. 2008; Stimmer et al. 2010). However, in
contrast to non-lymphotropic viruses (e.g. Theiler’s murine encephalomyelitis virus
(TMEV)), pathogen-specific immune responses in peripheral lymphoid organs are not
induced in MS and TME, probably as a consequence of massive lymphoid depletion as
observed in the present study.
6.2 Role of regulatory T cells in neurological diseases
Treg play a key role in controlling immune responses under physiological conditions and
in various systemic and CNS inflammatory diseases (Sakaguchi et al. 2006; Liesz et al.
2009). This has been demonstrated by depletion of natural Treg which leads to an
activation of self-reactive T cell clones, inducing severe and widespread autoimmune
inflammatory diseases (Sakaguchi et al. 2006). Moreover, functional impairment or an
insufficient amount of Treg is suspected to contribute to demyelination in acute MS
lesions (Fritzsching et al. 2011). Impaired Treg function could lead to disturbed
tolerance against autoantigens, resulting in autoimmunity (Viglietta et al. 2004; Baecher-
Allan and Hafler, 2006; Zozulya and Wiendl, 2008). These studies demonstrate that
manipulation of Treg in the periphery influences the outcome of CNS infection.
Well orchestrated immune responses against invading pathogens is essential within the
CNS since also minor damage can have significant consequences for neurological
functions (Lowther and Hafler, 2012). For instance, Treg reduce antiviral immunity in
experimental Theiler’s murine encephalomyelitis (TME; Herder et al. 2012; Richards et
al. 2012). Treg protect the CNS against an overshooting immune response, as
demonstrated in EAE as well as in animal models of stroke and HIV-associated
neurodegeneration. Besides dampening leukocyte responses, Treg are also able to

97
Chapter 6: General discussion
reduce astrogliosis and microglia-mediated inflammation (Beyersdorf et al. 2005; Liesz
et al. 2009; Liu et al. 2009). For example, the absence of Treg in mouse models of
stroke leads to an increased severity of tissue damage due to an excessive activation of
microglia and expression of TNF and IFN-γ (Liesz et al. 2009). In mouse models of
coronavirus-induced acute encephalitis, Treg help to limit immune mediated tissue
damage during the acute stage, demonstrating a protective role of Treg (Anghelina et
al. 2009). Moreover, during acute herpes simplex virus (HSV) infection of mice, Treg
facilitate early immune responses to local viral infection by allowing a timely entry of
immune cells into infected tissues, including the brain (Lund et al. 2008). In contrast, a
lack of Treg in HSV-infected mice is associated with delayed infiltration of DCs and T
cells, that accelerates fatal infection and increases the viral load in the CNS (Lund et al.
2008). However, although the generation of Treg in CNS inflammatory diseases is
regarded as physiological process to prevent immunopathology (MacDonald et al. 2002;
Sakaguchi 2003; Dittmer et al. 2004), Treg can also have detrimental effects for the
host by their suppressive effects upon protective immune responses which leads to an
incomplete elimination of pathogens and persistent infections, respectively (Dittmer et
al. 2004; Belkaid and Rouse, 2005). For instance, in experimental MV-infection of mice,
an elevated number of Treg increases viral replication in the brain, whereas transient
depletion of Treg significantly reduces the number of infected neurons (Reuter et al.
2012).
In the present study, permanent depletion of Treg in the spleen of all disease stages of
canine distemper has been observed, which causes a delayed infiltration of Treg
infiltration in the brain in advanced stages of canine distemper. The origin of CNS-
infiltrating Treg in the chronic stage of canine distemper remains unclear. It remains to
determine whether Treg accumulation in the brain of investigated dogs represents a
cumulative effect due to migration of residual peripheral Treg or a de novo generation of
Treg within the inflamed CNS. In HIV encephalitis mouse models, Treg migrate across
the blood brain barrier and are retained within neuroinflammatory sites (Gong et al.
2011). The lack of Treg observed in early stages of CDV-induced leukoencephalitis

98
Chapter 6: General discussion
potentially contributes to uncontrolled inflammatory responses by resident cells.
Similarly, an insufficient expansion of peripheral Treg might lead to strong
immunopathology in acute severe dengue fever and West Nile virus infection (Lühn et
al. 2007; Lanteri et al. 2009). Interestingly, the increase of Foxp3+ Treg in the CNS
during advanced inflammatory stages coincides with persistent infection, similar to
findings in experimental MV infection of mice (Sellin et al. 2009; Reuter et al. 2012).
Probably a compartmentalization of immune responses in the CNS as described in MS
(Massacesi 2002; Meinl et al. 2008) might induce the proliferation and expansion of
Foxp3+ cells locally within the brain of CDV-infected animals. In EAE, large numbers of
Treg are present in the inflamed brain, but their function is impaired by the elevated
expression of pro-inflammatory molecules, such as IL-6 and TNF, which counters Treg
immunosuppressive effects (Pasare and Medzhitov, 2003; Korn et al. 2007). Results of
the present study are indicative of inadequate immunoregulation which represents a
potential prerequisite for early lesion development. However, further studies are needed
to determine disease phase-specific and probably ambiguous functions of Treg in the
pathogenesis of canine distemper, which might also have relevance for the
understanding of human demyelinating diseases.
6.3 Role of dendritic cells in demyelinating disorders
DCs play a central role for the initiation and maintenance of both innate and adaptive
immune responses to infectious agents, including bacteria and viruses (Granucci et al.
2005; Hou et al. 2007). They are distributed in the skin, mucosa, and in lymphoid
organs, where they efficiently take up and process diverse microbial antigens and
present them as MHC–peptide complexes to T cells (Hou et al. 2007). Despite their
importance for immune homeostasis in peripheral organs, DCs have been assumed not
to be important for neuroinflammation because of their absence in the CNS parenchyma
(Hart and Fabre, 1981; Deshpande et al. 2007). However this view hase been
challenged by other studies which have shown the presence of DCs in meninges and

99
Chapter 6: General discussion
choroid plexus of healthy rodents, and their ability to activate naïve T cells that enter the
inflamed CNS (McMenamin 1999; McMahon et al. 2005). DCs are the primary APCs
directing T cell functions and therefore are extremely important in directing the immune
pathology characteristic of MS (Gandhi et al. 2010). Several studies have described an
accumulation of DCs in CNS during inflammation in MS and EAE pathogenesis (Bailey
et al. 2007; Cudrici et al. 2007; Wu and Laufer, 2007). In EAE, DCs primed with myelin
antigens in the periphery activate autoaggressive T cells and can lead to an overt
autoimmune disease (Dittel et al. 1999). DCs have been shown to be functionally
abnormal in MS (Wu and Laufer, 2007). For instance, DCs of MS patients exhibit an
activated phenotype with increased expression of activation markers, such as CD40
and CD80, and promote a pro-inflammatory environment (Huang et al. 1999; Karni et al.
2006). Moreover, DCs seem to affect the disease course of MS, since secondary
progressive MS patients show higher DC expression levels of CD80, IL-12 and TNF
compared to patients with relapsing-remitting MS (Karni et al. 2006). In MS, expression
of CD40 by DCs was found to be related to the increased production of IL-12 and IL-18
(Balashov et al. 1997; Karni et al. 2002). Furthermore, co-stimulatory molecules, CD80
and CD86, do not promote identical signals where expression of CD80 in DCs promotes
Th1 response and is involved in the exacerbation of EAE, and is reported to correlate
with disease duration, while CD86 elicits Th2 humural response (Freeman et al. 1995;
Kuchroo et al. 1995; Kouwenhoven et al. 2001). With progression of EAE, DCs tend to
show a down-regulation of antigen presentation properties, characterized by reduced
expression of MHC II and co-stimulatory molecules (Deshpande et al. 2007). Although,
DCs in MS patients and healthy controls show no differences in expression of IL-12, an
important inducer of IFN-γ and Th1 responses, respectively (Huang et al. 2001), several
studies reported a down-regulation of co-stimulatory molecules of plasmacytoid DCs
and moDCs of MS patients (Huang et al. 2001; Stasiolek et al. 2006).
In TME, infection rate of DCs varies between resistant and susceptible mice strains.
Altered DC function is supposed to influence antiviral immunity which causes persistent
infection and chronic demyelinating disease (Hou et al. 2007). In susceptible mice,

100
Chapter 6: General discussion
TMEV preferentially infects immature DCs, blocking their differentiation and activation,
while failing to induce apoptosis. TMEV-infected DCs may also contribute to disease
pathogenesis by altering the cytokine profile by T cells (Hou et al. 2007).
In the present study for the first time CDV-infection of DCs has been demonstrated.
Phenotyical analyses by flow cytometry revealed an impairment of antigen presentation
properties with down-regulation of co-stimulatory molecules of infected moDCs. These
changes might influence peripheral immune responses which potentially reduce antiviral
immunity in CDV-induced leukoencephalitis. Furthermore, the absence of apoptosis
induction of CDV-infected DCs might represent a mechanism of immune evasion and
viral persistence, respectively.
6.4 Interaction of dendritic cells and regulatory T cells in
demyelinating and chronic viral diseases
Function of DCs strongly depends on the signal, which can induce pro-inflammatory or
tolerogenic properties (Finkelman et al. 1996). The ability of DCs to induce immune
tolerance depends on their origin, activation state, maturation signals and the cytokine
environment at the time they encounter T lymphocytes (Janikashvili et al. 2011). The
tolerogenic function of DCs is mediated by T cell deletion, induction of anergy, or the
expansion of antigen-specific Treg (Steinman and Nussenzweig, 2002; Luo et al. 2007;
Yamazaki et al. 2007; Darrasse-Jèze et al. 2009; Janikashvili et al. 2011). The ability of
Treg to survive in the periphery requires the presence of co-stimulation through the
CD28 and B7 pathway (Perrin et al. 1995; Chang et al. 1999; Tada et al. 1999; Salomon
et al. 2000). In EAE, DCs support Treg-mediated immunosuppression which ameliorate
autoimmune responses (Deshpande et al. 2007). Furthermore, the number of Treg in
vivo is directly proportional to the number of DCs (Darrasse-Jèze et al. 2009). On the
other side, a decrease of Treg has been demonstrated to induce DC proliferation
(Darrasse-Jèze et al. 2009). DCs are the only APC population that can mediate
peripheral activation and expansion of Treg in vivo (Walker et al. 2003; Tarbell et al.

101
Chapter 6: General discussion
2004). The interaction of DCs and Treg is regulated by a feedback loop which depends
on MHC expression by DCs (Fig.8) (Darrasse-Jèze et al. 2009). DC stimulation in the
presence of TGF-β induces the generation of Treg from naïve CD4+ T cells (Shevach,
2002). In general, DC-T cell contact stimulates the secretion of small amounts of IL-2 by
T cells, which in conjunction with MHC II independently from DC co-stimulatory signals,
is sufficient for the induction and proliferation of Treg (Walker et al. 2003; Lange et al.
2007; Zou et al. 2010). Interestingly, matured DCs, although representing potent
inducer of cellular immune responses, have an higher ability to trigger expansion and
activation of Treg than immature DCs, as demonstrated in in vitro studies (Walker et al.
2003; Lange et al. 2007). These tolerogenic DCs are found in vivo in Peyer’s patches,
lungs, and the anterior chamber of the eye. They exhibit a mature DC phenotype but
secrete IL-10 and thus trigger Treg function (Lutz and Schuler, 2002).
Figure 8. Proposed homeostatic feedback loops between dendritic cells (DCs) and regulatory T
cells (Treg). A) Increased numbers of DCs increase the number of Treg via MHC II expression; B) Treg
are able to decrease the number of DCs due to expression of interleukin (IL)-10 and transforming growth
factor (TGF)-β; C) Reduction of DCs declines the amount of Treg due to decreased MHC II expression;
D) Loss of Treg induces DCs by an increased availability of Fms-related tyrosine kinase 3 ligand (Flt3L).
Modified from Darrasse-Jèze et al. (2009).
Co-stimulatory molecules of DCs, such as CD40, CD80 and CD86, play an important
role for the interaction with Treg. Here, CTLA-4 expressed on Treg impairs antigen

102
Chapter 6: General discussion
presentation properties of DCs (Salomon et al. 2000; Godfrey et al. 2004; Fallarino et al.
2006; Janikashvili et al. 2011). For instance, Treg can suppress the up-regulation co-
stimulatory molecules CD40, CD80 and CD86 and inhibit the production of pro-
inflammatory cytokines of murine DCs, which leads to a reduced ability to induce T cell
activation (Chaux et al. 1997). In humans it has been reported that a Treg-mediated
inhibition of DC maturation is associated with a down-regulation of CD80 and CD86
without affecting CD40 expression (Serra et al. 2003). Treg-mediated suppression of
DC maturation and down-regulation of co-stimulatory molecules depends on several
mechanisms, such as CTLA-4 ligation as well as the induction of indoleamine 2,3-
dioxygenase (IDO), lymphocyte activation gene 3 (LAG3) and neuropilin-1 (Fallarino et
al. 2006; Huang et al. 2004; Paust et al. 2004; Miyara and Sakaguchi, 2007; Sarris et al.
2008; Janikashvili et al. 2011). The precise role of LAG3, which interacts with MHC II,
remains unknown, since LAG3-deficient mice fail to develop enhanced autoimmune
responses unlike CTLA-4-deficient mice (Huang et al. 2004; Miyara and Sakaguchi,
2007). Furthermore, Treg can modulate DC function by the production of inhibitory
cytokines such as IL-10, IL-35 and TGF-β (Miyara and Sakaguchi, 2007; Vignali et al.
2009).
As demonstrated in vitro studies, CDV modulates the expression of co-stimulatory
molecules and MHC II in canine moDCs. Based on previous observations
(Wünschmann et al. 2000), cells with DC-like morphology in splenic germinal centers
were the only cell population to be infected with CDV during chronic stage. Moreover,
after lymphoid repopulation of the spleen during chronic disease stage, Foxp3+ cells
remained depleted. These results led to the hypothesis that impaired antigen presenting
properties of DCs might contribute to an insufficient function or expansion of Treg in
canine distemper.

103
Chapter 6: General discussion
6.5 Conclusion
In conclusion, results of the present study support the hypothesis that an inadequate
immunoregulation as a consequence of Treg depletion promotes early lesion CNS
development in canine distemper. These findings are in agreement with previous
reports that the initiation of inflammation in myelin disorders is influenced by an
immunological imbalance of the peripheral immune system (Tsunoda et al., 2005;
Navarrete-Talloni et al., 2010). As suggested for human MS, the development of novel
therapies targeting Treg represents a promising strategy in canine neurological
disorders. However, the ambiguous functions of Treg, exhibiting both beneficial and
detrimental effects upon the host immune response, have to be considered in canine
CNS diseases with a confirmed (e.g. canine distemper) or suspected infectious etiology
(e.g. granulomatous meningoencephalitis). Since dogs are appreciated as translational
animal models for several human diseases (Kling, 2007; Spitzbarth et al., 2012),
observations in canine distemper might have relevance also for human demyelinating
disorders.
Getting insight into the interaction between viruses and DCs is fundamental to
understand the pathogenesis of infectious disorders, which has implication for
prevention (e.g. vaccination) and treatment strategies (Hou, So, and Kim 2007). For the
first time, CDV infection of cultured canine DCs has been demonstrated in the present
survey. Modulation of co-stimulatory molecules and reduced antigen presenting function
of CDV-infected DCs might account for profound and long lasting inhibitory effects upon
the immune system in canine distemper. However, further studies are needed to
expand the existing knowledge about the impact of infected DCs upon leukocyte
activation and polarization of immune responses in canine distemper and other canine
neurological disorders, as a prerequisite for the development of future therapies.

104

105
Chapter 7: Summary
Visar Qeska - “Investigation of immunomodulatory properties of neurovirulent viruses –
in vitro and in vivo effects of canine distemper virus”
7 Chapter 7: Summary
Distemper in dogs is caused by the canine distemper virus (CDV). The disease course
and pathogenesis of canine distemper are similar to human measles, including fever,
rash, respiratory signs, lymphopenia, and profound immunosuppression with
generalized depletion of lymphoid organs during the acute disease phase. Central
nervous system (CNS) infection and neurological complications can be observed
frequently in infected dogs. Depending on CDV strain, host immune status and age,
naturally infected dogs develop demyelinating leukoencephalomyelitis, which shares
similarities with human myelin disorders. Therefore canine distemper is appreciated as
a translational animal model for multiple sclerosis (MS). In virus-induced neurological
diseases, an ambiguous function of regulatory T cells (Treg), with both beneficial effects
by reducing immunopathology and detrimental effects by inhibiting antiviral immunity,
has been described. However, the role of Treg in the pathogenesis of canine distemper
has not been investigated yet. Moreover, despite the demonstration of infected dendritic
cell (DC)-like cells in canine distemper, the effect of CDV upon canine DCs remains
undetermined.
The aim of the first part of the present study was to testify the hypothesis that peripheral
lymphoid depletion influences immunomodulatory mechanisms in the brain of CDV-
infected dogs. Immunohistochemistry revealed a lack of Foxp3+ Treg in
predemyelinating and early demyelinating lesions which was associated with the
accumulation of CD3+ T cells, L1+ macrophages/microglia and GFAP+ astrocytes.
Together with CD79α+ B cells, a delayed infiltration of Foxp3+ Treg was observed in
chronic demyelinating lesions. Splenic depletion of Foxp3+ Treg was associated with an
increased mRNA-expression of tumor necrosis factor in the acute disease phase,
indicative of a pro-inflammatory microenvironment and lack of immunological counter

106
Chapter 7: Summary
regulation in peripheral lymphoid organs. In conclusion, disturbed immune regulatory
mechanisms represent a potential cause for excessive neuroinflammation and early
lesion development in canine distemper leukoencephalitis, as discussed for immune
mediated myelin disorders such as MS.
In the second part of the thesis species-specific properties and translational aspects of
canine DCs have been reviewed, including the current knowledge about in vitro
generation and characterization of canine DCs. In addition, the role of DCs in the
pathogenesis of selected canine neoplastic, infectious (including canine distemper),
allergic and autoimmune diseases, which share similarities with human disorders and
thus have significance for translational medicine, have been discussed.
The impact of CDV upon professional antigen presenting cells was investigated in the
third part of the thesis. Similar to human measles virus, CDV is supposed to inhibit
terminal differentiation of DCs, responsible for disturbed repopulation of lymphoid
tissues and diminished antigen presenting function in dogs. In order to testify the
hypothesis that CDV-infection leads to an impairment of co-stimulatory functions of
professional antigen presenting cells, canine DCs have been generated from peripheral
blood monocytes in vitro and infected with CDV. Virus infection was confirmed and
quantified by transmission electron microscopy, CDV-specific immunofluorescence and
virus titration. Phenotypical changes of cultured cells were determined by flow
cytometry. In addition, apoptotic changes and cellular damage were quantified by the
terminal desoxynucleotidyl transferase-mediated dUTP-biotin nick end labeling-method
and lactate dehydrogenase-assay, while cell proliferation was determined by 5-bromo-
2’-deoxyuridine-incorporation. Results demonstrated a significant time dependant
increase of the infectivity rate of DCs at 24, 72 and 120 hours post infection (hpi). As
observed by flow cytometry at 120 hpi, CDV-infection of canine DCs led to a down-
regulation of co-stimulatory molecules CD80 and CD86 as well as of MHC II, indicating
disturbed antigen presenting properties. As a potential mechanism to evade host
immune responses, infected DCs showed no evidence of apoptosis or cell lysis at 24,
72 and 120 hpi. Similarly, cell proliferation rate of infected cells was unaffected at any

107
Chapter 7: Summary
investigated time points. These data suggest that CDV-infection of DCs plays a role for
pathogenesis of long lasting immune alterations and virus persistence in canine
distemper.
In conclusion, results of the present study support the hypothesis that an inadequate
immunoregulation as a consequence of Treg depletion promotes the development of
early CNS lesion in canine distemper. These findings are in agreement with previous
reports, that initiation of inflammation in myelin disorders is influenced by an
immunological imbalance of the peripheral immune system. As suggested for human
MS, the development of novel therapies targeting Treg represents a promising strategy
in canine neurological disorders. Moreover, getting insights into the interaction between
viruses and DCs is fundamental to understand the pathogenesis of infectious disorders,
which has implication for prevention (e.g. vaccination) and treatment strategies. Since
dogs are appreciated as translational animal models for several human diseases
observations in canine distemper might have relevance also for human CNS disorders

108

109
Chapter 8: Zusammenfassung
Visar Qeska – “Untersuchung der immunomodulatorischen Eigenschaften von
neurovirulent viren – in vitro und in vivo Effekte des Staupevirus”
8 Chapter 8: Zusammenfassung
Die Infektion mit dem Hundestaupevirus (canine distemper virus, CDV) führt bei
Hunden zu einer fieberhaften Erkrankung mit respiratorischen, enteralen und
zentralnervösen Symptomen. Vergleichbar mit der Masernvirusinfektion des Menschen
entwickelt sich in der akuten Phase der Hundestaupe außerdem eine Depletion
sämtlicher lymphatischer Organe mit daraus resultierender Leukopenie und
Immunsuppression. In Abhängigkeit vom Immunstatus und Alter des Hundes sowie vom
Virusstamm entwickelt sich eine demyelinisierende Leukoenzephalomyelitis, welche
Ähnlichkeiten mit den Läsionen bei der Multiplen Sklerose (MS) aufweist. Die
Hundestaupe stellt daher ein translationales Tiermodell für humane
Entmarkungskrankheiten dar. Regulatorische T-Zellen (regulatory T cells, Treg) führen
einerseits zu einer Reduktion immunpathologischer Prozesse, andererseits konnte
insbesondere bei viralen Infektionen des Zentralen Nervensystems (ZNS) eine
Hemmung der protektiven Immunität und damit eine Exazerbation der Krankheit
beobachtet werden. Die Bedeutung von Treg im Verlauf der CDV-Infektion ist bislang
unklar. Außerdem ist die Rolle der Dendritischen Zellen (DZ) für die Pathogenese der
Hundestaupe bisher nur unzureichend untersucht worden.
Das Ziel des ersten Teils der Arbeit war die Überprüfung der Hypothese, dass eine
Treg-Depletion in peripheren lymphatischen Organen zu einer gestörten
Immunmodulation im Gehirn von CDV-infizierten Hunden führt. Mittels Immunhistologie
konnte ein Verlust von Foxp3+ Treg in Verbindung mit einer Ansammlung von CD3+ T-
Zellen, L1+ Makrophagen/Mikroglia und GFAP+ Astrozyten in prädemyelinisierenden
und frühen demyelinisierenden Herden im ZNS von infizierten Hunden nachgewiesen
werden. Gemeinsam mit CD79α+ B-Zellen konnte eine verzögerte Infiltration von
Foxp3+ Treg in der chronischen Entmarkungsphase festgestellt werden. Die Depletion

110
Chapter 8: Zusammenfassung
von Foxp3+ Treg in der Milz war mit einer vermehrten mRNS-Expression des Tumor-
Nekrose-Faktors vergesellschaftet. Die Ergebnisse sprechen für das Vorliegen einer
gestörten Immunregulation in peripheren lymphatischen Organen und dem ZNS, welche
zu einer exzessiven Neuroinflammation und möglicherweise Initiation der
Gehirnläsionen führt. Vergleichbare Vorgänge werden gegenwärtig für die Pathogenese
der MS diskutiert.
Im zweiten Teil der Arbeit wurden in einem Übersichtsartikel tierartspezifische und
translationale Aspekte von kaninen DZ zusammengefasst und die Rolle der Zellen in
neoplastischen, infektiösen (inklusive Hundestaupe), allergischen und autoimmunen
Krankheiten des Hundes diskutiert. Ein Schwerpunkt war hierbei, Unterschiede und
Gemeinsamkeiten zwischen kaninen und humanen Krankheiten aufzuzeigen.
Im dritten Teil der Arbeit wurde der Einfluss des CDV auf professionell
antigenpräsentierende Zellen näher untersucht. Hintergrund hierbei war die
Beobachtung, dass, vergleichbar mit den Prozessen bei der Masernvirusinfektion, CDV
eine Differenzierungsstörung von DZ hervorruft und dadurch eine verminderte
Antigenpräsentation und Lymphozytenaktivierung bei Hunden ausgelöst wird. Zur
Überprüfung wurden Monozyten aus dem Blut von klinisch gesunden Hunden isoliert, in
Kultur zu DZ differenziert und anschließend mit CDV infiziert. Die Infektion der DZ
wurde mittels Transmissionselektronenmikroskopie verifiziert und der Virusgehalt
mittels Immunfluoreszenz und Virustitration quantifiziert. Außerdem wurden
durchflusszytometrische Analysen durchgeführt. Zelluläre Schäden wurden mittels der
terminal desoxynucleotidyl transferase-mediated dUTP-biotin nick end labeling
(TUNEL)-Methode und dem Laktatdehydrogenase (LDH)-Test ermittelt. Der
Bromdesoxyuridin (BrdU)-Test wurde zur Quantifizierung der Zellproliferation in vitro
eingesetzt. Die Ergebnisse zeigten eine signifikante Zunahme der Infektionsrate 24, 72
und 120 Stunden nach der Infektion (hours post infection, hpi). Mittels
Durchflusszytometrie konnte eine verminderte Expression der ko-stimulatorischen
Moleküle CD80 und CD86 sowie des MHC II in infizierten DZ nachgewiesen werden.

111
Chapter 8: Zusammenfassung
Als möglicher Persistenzmechanismus zeigten die Zellen 24, 72 und 120 hpi keine
gesteigerte Apoptose (TUNEL-Methode) bzw. Lyse (LDH-Test). Außerdem blieb die
Proliferation infizierter Zellen im Vergleich zu nicht infizierten Zellen mittels BrdU-Test
an allen Untersuchungszeitpunkten unverändert. Die Daten sprechen für das Vorliegen
einer virusinduzierten Störung der Ko-stimulation bzw. Antigenpräsentation von DZ.
Außerdem besitzt das CDV die Eigenschaft potentiell in kaninen DZ zu persistieren.
Die Ergebnisse der Studien unterstützen die Hypothese einer gestörten
Immunregulation im Zusammenhang mit dem Verlust von Treg, wodurch eine
unkontrollierte Entzündungsreaktion im ZNS von CDV-infizierten Tieren ausgelöst wird.
Außerdem ist möglicherweise die Infektion von DZ für die lang andauernden
Alterationen des Immunsystems bei der Hundestaupe mitverantwortlich. In diesem
Zusammenhang haben neuere Untersuchungen gezeigt, dass die Neuroinflammation
bei Entmarkungskrankheiten maßgeblich durch eine gestörte Immunhomöostase in
peripheren lymphatischen Organen beeinflusst wird. Techniken zur Modulation von
Treg und DZ stellen daher innovative Therapieansätze für neurologische Krankheiten in
der Human- und Tiermedizin dar.

112

113
Chapter 9: References
9 Chapter 9: References
Alldinger S, Baumgärtner W, Orvell C, 1993. Restricted expression of viral surface proteins in canine
distemper encephalitis. Acta Neuropathol 85, 635–645.
Alldinger S, Wünschmann A, Baumgärtner W, Voss C, Kremmer E, 1996. Up-regulation of major
histocompatibility complex class II antigen expression in the central nervous system of dogs with
spontaneous canine distemper virus encephalitis. Acta Neuropathol 92, 273–280.
Alldinger S, Fonfara S, Kremmer E, Baumgärtner W, 2000. Up-regulation of the hyaluronate receptor
CD44 in canine distemper demyelinated plaques. Acta Neuropathol 99, 138-46.
Alldinger S, Gröters S, Miao Q, Fonfara S, Kremmer E, Baumgärtner W, 2006. Roles of an extracellular
matrix (ECM) receptor and ECM processing enzymes in demyelinating canine distemper
encephalitis. Dtsch Tierarztl Wochenschr 113, 151-156.
Andrea BAD, Ma X, Aste-Amezaga M, Paganin C, Trinchieri G, 1995. Stimulatory and inhibitory effects of
interleukin ( IL ) -4 and ID13 on the production of cytokines by human peripheral blood
mononuclear cells : priming for ID12 and tumor necrosis factor alpha production. J Exp Med 181,
537–546.
Anghelina D, Zhao J, Trandem K, Perlman S, 2009. Role of regulatory T cells in coronavirus-induced
acute encephalitis. Virology 385, 358–367.
Appel MJ, 1969. Pathogenesis of canine distemper. Am J Vet Res30(7):1167-82.
Appel MJ, 1970. Distemper pathogenesis in dogs. J Am Vet Med Assoc 156, 1681-4.
Aseffa A, Gumy A, Launois P, MacDonald RH, Louis JA, Tacchini-Cottier F, 2002. The early IL-4
response to Leishmania major and the resulting Th2 cell maturation steering progressive disease
in BALB/c mice are subject to the control of regulatory CD4+CD25+ T cells. J Immunol 169,
3232–3241.
Azuma T, Takahashi T, Kunisato A, 2003. Human CD4 + CD25 + Regulatory T cells suppress NKT cell
functions. Cancer Res 63, 4516–4520.
Baecher-Allan C, Hafler DA, 2006. Human regulatory T cells and their role in autoimmune disease.
Immunol Rev 212, 203–216.
Bailey SL, Bettina S, Eileen JM, Miller SD, 2007. CNS myeloid DCs presenting endogenous myelin
peptides ‘preferentially’ polarize CD4+ T(H)-17 cells in relapsing EAE. Nat Immunol 8, 172-180.
Baines SJ, Mcinnes EF, Mcconnell I, 2008. E-cadherin expression in canine cutaneous histiocytomas.
Vet Rec 162, 509-513
Balashov KE, Smith DR, Khoury SJ, Hafler DA, Weiner HL, 1997. Increased interleukin 12 production in
progressive multiple sclerosis: induction by activated CD4+ T cells via CD40 ligand. Proc Natl
Acad Sci USA 94, 599–603.

114
Chapter 9: References
Banati RB, Kreutzberg GW, 1993. Flow Cytometry: Measurement of proteolytic and cytotoxic activity of
microglia. Clin Neuropathol 12, 285–288.
Banchereau J, Briere F, Caux C, Davoust J, Lebecque S, Liu YJ, Pulendran B, Palucka K, 2000.
Imunobiology of dendritic cells. Annu Rev Immunol 18, 767–811.
Baumgärtner W, Orvell C, Reinacher M, 1989. Naturally occurring canine distemper virus encephalitis:
distribution and expression of viral polypeptides in nervous tissues. Acta Neuropathol 78, 504-
512.
Baumgärtner W, Alldinger S, 2005. The pathogenesis of canine distemper virus induced demyelination —
a biphasic process. In: Lavi, E, Constantinescu, CS (Eds), Experimental Models of Multiple
Sclerosis Springer, New York, pp 871–887
Beck K, Meyer-König U, Weidmann M, Nern C, Hufert FT, 2003. Human cytomegalovirus impairs
dendritic cell function: a novel mechanism of human cytomegalovirus immune escape. Eur J
Immunol 33, 1528–1538.
Beineke A, Markus S, Borlak J, Thum T, Baumgärtner W, 2008. Increase of pro-inflammatory cytokine
expression in non-demyelinating early cerebral lesions in nervous canine distemper. Viral
Immunol 21, 401–410.
Beineke A, Puff C, Seehusen F, Baumgärtner W, 2009. Pathogenesis and immunopathology of systemic
and nervous canine distemper. Vet Immunol Immunopathol 127, 1–18.
Beineke A, Siebert U, Wohlsein P, Baumgärtner W, 2010. Immunology of whales and dolphins. Vet
Immunol Immunopathol 133, 81–94.
Belkaid Y, Rouse BT, 2005. Natural regulatory T cells in infectious disease. Nat Immunol 6, 353–60.
Bestetti G, Fatzer R, Frankhauser R, 1978. Encephalitis following vaccination against distemper and
infectious hepatitis in the dog An optical and ultrastructural study. Acta Neuropathol 43, 69-75.
Beyersdorf N, Gaupp S, Balbach K, Schmidt J, Toyka KV, Lin CH, Hanke T, Hünig T, Kerkau T, Gold R,
2005. Selective targeting of regulatory T cells with CD28 superagonists allows effective therapy of
experimental autoimmune encephalomyelitis. J Exp Med 202, 445–455.
Bhardwaj N, 2003. The modulation of immunity by dendritic cells. Clin Appl Immunol 3, 173-182.
Biancotti JC, Kumar S, de Vellis J, 2008. Activation of inflammatory response by a combination of growth
factors in cuprizone-induced demyelinated brain leads to myelin repair. Neurochem Res 33,
2615–2628.
Biller BJ, Elmslie RE, Burnett RC, Avery AC, Dow SW, 2007. Use of FoxP3 expression to identify
regulatory T cells in healthy dogs and dogs with cancer. Vet Immunol Immunopathol 116, 69–78.
Bodey B, Bodey BJr, Kaiser HE, 1997. Dendritic type, accessory cells within the mammalian thymic
microenvironment Antigen presentation in the dendritic neuro-endocrine-immune cellular network.
In Vivo 11, 351-370.

115
Chapter 9: References
Björck P, Banchereau J, Flores-Romo L, 1997. CD40 ligation counteracts Fas-induced apoptosis of
human dendritic cells. Int Immunol 9, 365–372
Boliar S, Chambers TM, 2010. A new strategy of immune evasion by influenza A virus: inhibition of
monocyte differentiation into dendritic cells. Vet Immunol Immunopathol 136, 201–210.
Bonnefont-Rebeix C, de Carvalho CM, Bernaud J, Chabanne L, Marchal T,Rigal D, 2006. CD86 molecule
is a specific marker for canine monocyte-derived dendritic cells. Vet Immunol Immunopathol 109,
167–176.
Botteron C, Zurbriggen A, Griot C, Vandevelde M, 1992. Canine distemper virus-immune complexes
induce bystander degeneration of oligodendrocytes. Acta Neuropathol 83, 402–407.
Brunkow ME, Jeffery EW, Hjerrild KA, Paeper B, Clark LB, Yasayko SA, Wilkinson JE, Galas D, Ziegler
SF, Ramsdell F, 2001. Disruption of a new forkhead/winged-helix protein, scurfin, results in the
fatal lymphoproliferative disorder of the scurfy mouse. Nat Genet 27, 68–73.
Bund D, Buhmann R, Gökmen F, Kremser A, Dreyssig J, Kolb HJ, Schmetzer HM, 2010. Canine-DCs
using different serum-free methods as an approach to provide an animal-model for
immunotherapeutic strategies. Cell Immunol 263, 88–98.
Carvalho OV, Botelho CV, Ferreira CG, Scherer PO, Soares-Martins JA, Almeida MR, Silva Júnior A,
2012. Immunopathogenic and neurological mechanisms of canine distemper virus. Adv Virol
2012, 1–10.
Cecere TE, Todd SM, Leroith T, 2012. Regulatory T cells in arterivirus and coronavirus infections: do they
protect against disease or enhance it? Viruses 4, 833–846.
Cerquetella, M, Spaterna, A, Laus, F, Tesei, B, Rossi, G, Antonelli, E, Villanacci V, Bassotti G, 2010.
Inflammatory bowel disease in the dog: differences and similarities with humans. World J
Gastroenterol 16, 1050-1056.
Chang TT, Jabs C, Sobel RA, Kuchroo VK, Sharpe AH, 1999. Studies in B7-deficient mice reveal a
critical role for B7 costimulation in both induction and effector phases of experimental
autoimmune encephalomyelitis. J Exp Med 190, 733–740.
Chaux P, Favre N, Martin M, Martin F, 1997. Tumor-infiltrating dendritic cells are defective in their
antigen-presenting function and inducible B7 expression in rats. Int J Cancer 72, 619–624
Chen W, Jin W, Hardegen N, Lei KJ, Li L, Marinos N, McGrady G, Wahl SM, 2003. Conversion of
peripheral CD4+CD25- naive T cells to CD4+CD25+ regulatory T cells by TGF-beta induction of
transcription factor Foxp3. J Exp Med 198, 1875–1886.
Cocks BG, Chang CCJ, Carballido JM, Yssel H, de Vries JE, Aversa G, 1995. A novel receptor involved
in T-cell activation. Nature 376, 260–263.
http://www.ncbi.nlm.nih.gov/pubmed?term=Wilkinson%20JE%5BAuthor%5D&cauthor=true&cauthor_uid=11138001

116
Chapter 9: References
Cudrici C, Ito T, Zafranskaia E, Niculescu F, Mullen KM, Vlaicu S, Judge SIV, Calabresi PA, Rus H, 2007.
Dendritic cells are abundant in non-lesional gray matter in multiple sclerosis. Exp Mol Pathol 83,
198–206.
Cunningham AL, Donaghy H, Harman AN, Kim M, Turville SG, 2010. Manipulation of dendritic cell
function by viruses. Curr Opin Microbiol 13, 524–529.
Céspedes PF, Cruz P, Navarro CO, 2010. Modulación de la respuesta inmune durante la infección por
virus distemper canino : implicancias terapéuticas y en el desarrollo de vacunas - Modulation of
immune response during canine distemper virus infection: therapeutic and vaccine development
implication. Arch Med Vet 28, 15–28.
Darrasse-Jèze G, Deroubaix S, Mouquet H, Victora GD, Eisenreich T, Yao KH, Masilamani RF, Dustin
ML, Rudensky A, Liu K, Nussenzweig MC, 2009. Feedback control of regulatory T cell
homeostasis by dendritic cells in vivo. J Exp Med 206, 1853–1862.
Decaro N, Camero M, Greco G, Zizzo N, Tinelli A, Campolo M, Pratelli A, Buonavoglia C, 2004. Canine
distemper and related diseases: report of a severe outbreak in a kennel. New Microbiol 2, 177-
181.
Del Puerto HL, Martins AS, Moro L, Milsted A, Alves F, Braz GF, Vasconcelos AC, 2010. Caspase-3/-8/-
9, Bax and Bcl-2 expression inthe cerebellum, lymph nodes and leukocytes of dogs naturally
infected with canine distemper virus. Genet Mol Res 9, 151–161.
Deem SL, Spelman LH, Yates RA, Montali RJ, 2000. Canine distemper in terrestrial carnivores: a review
J Zoo Wildl Med 31, 441–451.
Deshpande P, King IL, Segal BM, 2007. CNS CD11c+ Cells from mice with encephalomyelitis polarize
Th17 cells and support CD25+CD4+ T cell-mediated immunosuppression, suggesting dual roles
in the disease process. J Immunol 178, 6695–6699.
de Swart RL, Ludlow M, de Witte L, Yanagi Y, van Amerongen G, McQuaid S, Yüksel S, Geijtenbeek TB,
Duprex WP, Osterhaus AD, 2007. Predominant infection of CD150+ lymphocytes and dendritic
cells during measles virus infection of macaques. PLoS Pathog 3, e178.
de Witte L, de Vries RD, van der Vlist M, Yüksel S, Litjens M, de Swart RL, Geijtenbeek TB, 2008. DC-
SIGN and CD150 have distinct roles in transmission of measles virus from dendritic cells to T-
lymphocytes. PLoS Pathog 4, e1000049.
Dhiman N, Jacobson RM, Poland GA, 2004. Measles virus receptors: SLAM and CD46. Rev Med Virol
14, 217–229.
Dittel B, Visintin I, Merchantm RM, Janeway CA Jr, 1999. Presentation of the self antigen myelin basic
protein by dendritic cells leads to experimental autoimmune encephalomyelitis. J Immunol 163,
32–39.
http://www.ncbi.nlm.nih.gov/pubmed?term=de%20Witte%20L%5BAuthor%5D&cauthor=true&cauthor_uid=18020706

117
Chapter 9: References
Dittmer U, He H, Messer RJ, Schimmer S, Olbrich AR, Ohlen C, Greenberg PD, Stromnes, IM, Iwashiro
M, Sakaguchi S, Evans LH, Peterson KE, Yang G, Hasenkrug KJ, 2004. Functional impairment of
CD8(+) T cells by regulatory T cells during persistent retroviral infection. Immunity 20, 293–303.
Donaghy H, Gazzard B, Gotch F, Patterson S, 2003. Dysfunction and infection of freshly isolated blood
myeloid and plasmacytoid dendritic cells in patients infected with HIV-1. Blood 101, 4505–4511.
Dörig RE, Marcil A, Chopra A, Richardson CD, 1993. The human CD46 molecule is a receptor for
measles virus (Edmonston strain). Cell 75, 295–305.
Ensinger EM, Boekhoff TMA, Carlson R, Beineke A, Rohn K, Tipold A, Stein VM, 2010. Regional
topographical differences of canine microglial immunophenotype and function in the healthy
spinal cord. J Neuroimmunol 227, 144–152.
Erlenhöfer C, Duprex WP, Rima BK, ter Meulen V, Schneider-Schaulies J, 2002. Analysis of receptor
(CD46, CD150) usage by measles virus. J Gen Virol 83, 1431–1436.
Esolen LM, Ward BJ, Moench TR, Griffin DE, 1993. Infection of monocytes during measles. J Infect Dis
168, 47–52.
Fallarino F, Grohmann U, You S, Mcgrath BC, Cavener DR, Vacca C, Orabona, C, Bianchi R, Belladona
ML, Volpi C, Santamaria P, Fioretti MC, Puccetti O, 2006. The combined effects of tryptophan
starvation and tryptophan catabolites down-regulate T cell receptor zeta-chain and induce a
regulatory phenotype in naive T cells. J Immunol 176, 6752–6761.
Fantini MC, Becker C, Monteleone G, Pallone F, Galle PR, Neurath MF, 2004. Cutting edge: TGF-beta
induces a regulatory phenotype in CD4+CD25- T cells through Foxp3 induction and down-
regulation of Smad7. J Immunol 172, 5149–5153.
Feger U, Tolosa E, Huang YH, Waschbisch A, Biedermann T, Melms A, Wiendl H, 2007. HLA-G
expression defines a novel regulatory T-cell subset present in human peripheral blood and sites
of inflammation. Blood, 110, 568–577.
Feuerer M, Hill JA, Mathis D, Benoist C, 2009. Foxp3+ regulatory T cells: differentiation, specification,
subphenotypes. Nat Immunol 10, 689–695.
Feuerer M, Hill JA, Kretschmer K, von Boehmer H, Mathis D, Benoist C, 2010. Genomic definition of
multiple ex vivo regulatory T cell subphenotypes. Proc Nat Acad Sci U S A 107, 5919–5924.
Finkelman F, Lees A, Birnbaum R, Gause WC, Morris SC, 1996. Dendritic cells can present antigen in
vivo in a tolerogenic or immunogenic fashion. J Immunol 157, 1406–1414.
Fitting J, Killian D, Junghanss C, Willenbrock S, Murua Escobar H, Lange S, Nolte I, Barth S, Tur MK,
2011. Generation of recombinant antibody fragments that target canine dendritic cells by phage
display technology. Vet Comp Oncol 9, 183–195.
http://www.ncbi.nlm.nih.gov/pubmed?term=Greenberg%20PD%5BAuthor%5D&cauthor=true&cauthor_uid=15030773
http://www.ncbi.nlm.nih.gov/pubmed?term=Hasenkrug%20KJ%5BAuthor%5D&cauthor=true&cauthor_uid=15030773
http://www.ncbi.nlm.nih.gov/pubmed?term=Richardson%20CD%5BAuthor%5D&cauthor=true&cauthor_uid=8402913
http://www.ncbi.nlm.nih.gov/pubmed?term=Monteleone%20G%5BAuthor%5D&cauthor=true&cauthor_uid=15100250
http://www.ncbi.nlm.nih.gov/pubmed?term=Waschbisch%20A%5BAuthor%5D&cauthor=true&cauthor_uid=17371944
http://www.ncbi.nlm.nih.gov/pubmed?term=Biedermann%20T%5BAuthor%5D&cauthor=true&cauthor_uid=17371944

118
Chapter 9: References
Floess S, Freyer J, Siewert C, Baron U, Olek S, Polansky J, Schlawe K, Chang HD, Bopp T, Schmitt E,
Klein-Hessling S, Serfling E, Hamann A, Huehn J, 2007. Epigenetic control of the foxp3 locus in
regulatory T cells. PLoS Biol 5, pe38.
Fox RJ, Rostami AM, 2000. Anti-Interleukin-12 antibody: potential role in preventing relapses of multiple
sclerosis. Biodrugs 13, 233-241.
Freeman GJ, Boussiotis VA, Anumanthan A, Bernstein GM, Ke XY, Rennert PD, Gray GS, Gribben JG,
Nadler LM, 1995. B7-1 and B7-2 do not deliver identical costimulatory signals, since B7-2 but not
B7-1 preferentially costimulates the initial production of IL-4. Immunity 2, 523–532.
Frenzke M, Sawatsky B, Wong XX, Delpeut S, Mateo M, Cattaneo R, von Messling V, 2012. Nectin-4
dependent measles virus spread to the cynomolgus monkey tracheal epithelium: role of infected
immune cells infiltrating the lamina propria. J Virol JVI, 03037-12.
Frisk AL, Baumgärtner W, Gröne A, 1999. Dominating interleukin-10 mRNA expression induction in
cerebrospinal fluid cells of dogs with natural canine distemper virus induced demyelinating and
non-demyelinating CNS lesions. J Neuroimmunol 97, 102–109.
Fritzsching B, Haas J, König F, Kunz P, Fritzsching E, Pöschl J, Krammer PH, Brück W, Suri-Payer E,
Wildemann B, 2011. Intracerebral human regulatory T cells: analysis of CD4+ CD25+ FOXP3+ T
cells in brain lesions and cerebrospinal fluid of multiple sclerosis patients. PloS One, 6, pe17988.
Fugier-Vivier I, Servet-Delprat C, Rivailler P, Rissoan MC, Liu YJ, Rabourdin-Combe C, 1997. Measles
virus suppresses cell-mediated immunity by interfering with the survival and functions of dendritic
and T cells. J Exp Med 186, 813–823.
Gandhi R, Laroni A, Weiner HL, 2010. Role of the innate immune system in the pathogenesis of multiple
sclerosis. J Neuroimmunol 221, 7–14.
Gerhauser I, Hansmann F, Puff C, Kumnok J, Schaudien D, Wewetzer K, Baumgärtner W, 2012. Theiler's
murine encephalomyelitis virus induced phenotype switch of microglia in vitro. J Neuroimmunol
15, 49-55.
Godfrey WR, Ge YG, Spoden DJ, Levine BL, June CH, Blazar BR, Porter SB, 2004. In vitro-expanded
human CD4(+)CD25(+) T-regulatory cells can markedly inhibit allogeneic dendritic cell-stimulated
MLR cultures. Blood 104, 453–461.
Goodell EM, Blumenstock DA, Bowers WE, 1985. Canine dendritic cells from peripheral blood and lymph
nodes. Vet Immunol Immunopathol 8, 301-310.
Gong N, Liu J, Reynolds AD, Gorantla S, Mosley RL, Gendelman HE, 2011. Brain ingress of regulatory T
Cells in a murine model of HIV-1 encephalitis. J Neuroimmunol 230, 33–41.
Granucci F, Foti M, Ricciardi-Castagnoli P, 2005. Dendritic cell biology. Adv Immunol 88, 193–233.
Greene CE, Appel MJ, 1998. Canine distemper In: Greeene CE (Ed), Infectious diseases of dog and cat,
WB Saunders Company, Philadephia, London, Toronto, Montreal, Sydney, Tokyo, pp 9-22
http://www.ncbi.nlm.nih.gov/pubmed?term=K%C3%B6nig%20F%5BAuthor%5D&cauthor=true&cauthor_uid=21437244
http://www.ncbi.nlm.nih.gov/pubmed?term=Br%C3%BCck%20W%5BAuthor%5D&cauthor=true&cauthor_uid=21437244

119
Chapter 9: References
Griffin, DE, 2010. Measles virus-induced suppression of immune responses. Immunol Rev 236, 176–189.
Griot C, Vandevelde M, Richard A, Peterhans E, Stocker R, 1990. Selective degeneration of
oligodendrocytes mediated by reactive oxygen species. Free Radic Res Commun 11, 181–193.
Grosjean I, Caux C, Bella C, Berger I, Wild F, Banchereau J, Kaiserlian D, 1997. Measles virus infects
human dendritic cells and blocks their allostimulatory properties for CD4+ T cells. J Exp Med 186,
801–812.
Gröne A, Frisk AL, Baumgärtner W, 1998. Cytokine mRNA expression in whole blood samples from dogs
with natural canine distemper virus infection. Vet Immunol Immunopathol 65, 11–27.
Gröne A, Alldinger S, Baumgärtner W, 2000. Interleukin-1b, -6, -12 and tumor necrosis factor-a
expression in brains of dogs with canine distemper virus infection. J Neuroimmunol 110, 20–30.
Gröne A, Fonfara S, Baumgärtner W, 2002. Cell type-dependent cytokine expression after canine
distemper virus infection. Viral Immunol 15, 493–505.
Gröne A, Doherr MG, Zurbriggen A, 2004. Canine distemper virus infection of canine footpad epidermis.
Vet Dermatol 15, 159-167.
Gudi V, Moharregh-Khiabani D, Skripuletz T, Koutsoudaki PN, Kotsiari A, Skuljec J, Trebst C, Stangel M,
2009. Regional differences between grey and white matter in cuprizone induced demyelination.
Brain Res 1283, 127–138.
Göbel K, Bittner S, Melzer N, Pankratz S, Dreykluft A, Schuhmann MK, Meuth SG, Wiendl H, 2012.
CD4(+) CD25(+) FoxP3(+) regulatory T cells suppress cytotoxicity of CD8(+) effector T cells:
implications for their capacity to limit inflammatory central nervous system damage at the
parenchymal level. J Neuroinflammation 9, 41.
Haas L, Liermann H, Harder TC, Barrett T, Löchelt M, von Messling V, Baumgärtner W, Greiser-Wilke I,
1999. Analysis of the H gene, the central untranslated region and the proximal coding part of the
F gene of wild-type and vaccine canine distemper viruses. Vet Microbiol 69, 15-18.
Hart DN, Fabre JW, 1981. Demonstration and characterization of Ia-positive dendritic cells in the
interstitial connective tissues of rat heart and other tissues, but not brain. J Exp Med 154, 347–
361.
Hahm B, Arbour N, Oldstone MBA, 2004. Measles virus interacts with human SLAM receptor on dendritic
cells to cause immunosuppression. Virology 323, 292–302.
Hahm B, Trifilo MJ, Zuniga EI, Oldstone MB, 2005. Viruses evade the immune system through type I
interferon-mediated STAT2-dependent, but STAT1-independent, signaling. Immunity 22, 247–
257
Hamilton-Easton A, Eichelberger M, 1995. Virus-specic antigen presentation by different subsets of cells
from lung and mediastinal lymph node tissues of inluenza-infected mice. J Virol 69, 6359-6366.

120
Chapter 9: References
Harman AN, Wilkinson J, Bye CR, Bosnjak L, Stern JL, Nicholle M, Lai J, Cunningham AL, 2006. HIV
induces maturation of monocyte-derived dendritic cells and Langerhans cells. J Immunol 177,
7103–7113.
Hengel H, Lindner M, Wagner H, Heeg K, 1987. Frequency of herpes simplex virus specific murine
cytotoxic T lymphocyte precursors in mitogen- and antigen-driven primary T cell responses. J
Immunol 139, 4196-4202.
Herder V, Hansmann F, Stangel M, Skripuletz T, Baumgärtner W, Beineke A, 2011. Lack of cuprizone-
induced demyelination in the murine spinal cord despite oligodendroglial alterations substantiates
the concept of site-specific susceptibilities of the central nervous system. Neuropathol App
Neurobiol 37, 676–684.
Herder V, Hansmann F, Stangel M, Schaudien D, Rohn K, Baumgärtner W, Beineke A, 2012. Cuprizone
inhibits demyelinating leukomyelitis by reducing immune responses without virus exacerbation in
an infectious model of multiple sclerosis. J Neuroimmunol 244, 84–93.
Hori S, Nomura T, Sakaguchi S, 2003. Control of regulatory T cell development by the transcription factor
Foxp3. Science 299, 1057–1061.
Hou W, So EY, Kim BS, 2007. Role of dendritic cells in differential susceptibility to viral demyelinating
disease. PLoS Pathog 3, pe124.
Huang YM, Xiao BG, Ozenci V, Kouwenhoven M, Teleshova N, Fredrikson S, Link H, 1999. Multiple
sclerosis is associated with high levels of circulating dendritic cells secreting pro-inflammatory
cytokines. J Neuroimmunol 99, 82–90.
Huang YM, Stoyanova N, Jin YP, Teleshova N, Hussien Y, Xiao BG, Fredrikson S, Link H, 2001. Altered
phenotype and function of blood dendritic cells in multiple sclerosis are modulated by IFN-beta
and IL-10. Clin Exp Immunol 124, 106-114.
Huang CT, Workman CJ, Flies D, Pan X, Marson AL, Zhou G, Hipkiss EL, Ravi S, Kowalski J, Levitsky
HI, Powell JD, Pardoll DM, Drake CG, Vignali DA, 2004. Role of LAG-3 in regulatory T cells.
Immunity 21, 503–513.
Hägglund HG, McSweeney PA, Mathioudakis G, Bruno B, Georges GE, Gass MJ, Moore P, Sale GE,
Storb R, Nash RA, 2000. Ex vivo expansion of canine dendritic cells from CD34+ bone marrow
progenitor cells. Transplantation 70, 1437–1442.
Ibisch C, Pradal G, Bach JM, Lieubeau B, 2005. Functional canine dendritic cells can be generated in
vitro from peripheral blood mononuclear cells and contain a cytoplasmic ultrastructural marker. J
Immunol Methods 298, 175–182.
Isotani M, Katsuma K, Tamura K, Yamada M, Yagihara H, Azakami D, Ono K, Washizu T, Bonkobara M,
2006. Efficient generation of canine bone marrow-derived dendritic cells. J Vet Med Sci 68, 809–
814.
http://www.ncbi.nlm.nih.gov/pubmed?term=Fredrikson%20S%5BAuthor%5D&cauthor=true&cauthor_uid=10496180
http://www.ncbi.nlm.nih.gov/pubmed?term=Fredrikson%20S%5BAuthor%5D&cauthor=true&cauthor_uid=11422209

121
Chapter 9: References
Iwatsuki K, Okita M, Ochikubo F, Gemma T, Shin YS, Miyashita N, Mikami T, Kai C, 1995.
Immunohistochemical analysis of the lymphoid organs of dogs naturally infected with canine
distemper virus. J Comp Pathol 113, 185-190.
Janikashvili N, Bonnotte B, Katsanis E, Larmonier N, 2011. The dendritic cell-regulatory T lymphocyte
crosstalk contributes to tumor-induced tolerance. Clin Dev Immunol 2011, 430394.
Johnson R, Glickman LT, Emerick TJ, Patronek GJ, 1995. Canine distemper infection in pet dogs: I
Surveillance in Indiana during a suspected outbreak. J Am Anim Hosp Assoc 31, 223–229.
Karni A, Abraham M, Monsonego A, Cai G, Freeman GJ, Hafler D, Khoury SJ, Weiner HL, 2006. Innate
immunity in multiple sclerosis: Myeloid dendritic cells drive a proinflammatory immune response
1. J Immunol 177, 4196–4202.
Karni A, Koldzic DN, Bharanidharan P, Khoury SJ, Weiner HL, 2002. IL-18 Is linked to raised IFN-gamma
in multiple sclerosis and is induced by activated CD4(+) T Cells via CD40-CD40 ligand
interactions. J Neuroimmunol 125, 134–140.
Kast WM, Boog CJP, Roep BO, Voordouw AC, Melief CJM, 1988. Failure or success in the restoration of
virus-specific cytotoxic T lymphocyte response defects by dendritic cells. J Immunol 140, 3186-
3193.
Kerdiles YM, Sellin CI, Druelle J, Horvat B, 2006. Immunosuppression caused by measles virus: role of
viral proteins. Rev Med Virol 16, 49–63.
Kigerl KA, Lai W, Rivest S, Hart RP, Satoskar AR, Popovich PG, 2007. Toll-like receptor (TLR)-2 and
TLR-4 regulate inflammation, gliosis, and myelin sparing after spinal cord injury. J Neurochem
102, 37–50.
Kigerl KA, Popovich PG, 2009. Toll-like receptors in spinal cord injury. Curr Top Microbiol Immunol 336,
121–136.
Kigerl KA, Gensel JC, Ankeny DP, Alexander JK, Donnelly DJ, Popovich PG, 2009. Identification of two
distinct macrophage subsets with divergent effects causing either neurotoxicity or regeneration in
the injured mouse spinal cord. J Neurosci 29, 13435–13444.
Kipnis J, Avidan H, Caspi RR, Schwartz M, 2004. Dual effect of CD4+CD25+ regulatory T cells in
neurodegeneration: a dialogue with microglia. Proc Natl Acad Sci U S A 101, 14663–14669.
Klagge IM, Schneider-Schaulies S, 1999. Virus interactions with dendritic cells. J Gen Virol 80, 823–833.
Klagge IM, Abt M, Fries B, Schneider-Schaulies S, 2004. Impact of measles virus dendritic-cell infection
on Th-cell polarization in vitro. J Gen Virol 85, 3239–3247.
Knight SC, Askonas BA, Macatonia SE, 1997. Dendritic cells as targets for cytotoxic T lymphocytes. Adv
Exp Med Biol 417, 389-394.

122
Chapter 9: References
Korn T, Reddy J, Gao W, Bettelli E, Awasthi A, Petersen TR, Bäckström BT, Sobel RA, Wucherpfennig
KW, Strom TB, Oukka M, Kuchroo VK, 2007. Myelin-specific regulatory T cells accumulate in the
CNS but fail to control autoimmune inflammation. Nat Med 13, 423–431.
Kotter MR, Setzu A, Sim FJ, Van Rooijen N, Franklin RJ, 2001. Macrophage depletion impairs
oligodendrocyte remyelination following lysolecithin-induced demyelination. Glia 35, 204–212.
Kouwenhoven M, Teleshova N, Ozenci V, Press R, Link H, 2001. Monocytes in multiple sclerosis:
phenotype and cytokine profile. J Neuroimmunol 112 197–205.
Kuchroo VK, Das MP, Brown JA, Ranger AM, Zamvil SS, Sobel RA, Weiner HL, Nabavi N, Glimcher LH,
1995. B7-1 and B7-2 costimulatory molecules activate differentially the Th1/Th2 developmental
pathways: application to autoimmune disease therapy. Cell 80, 707–718.
Kummerfeld M, Seehusen F, Klein S, Ulrich R, Kreutzer R, Gerhauser I, Herder V, Baumgärtner W,
Beineke A, 2012. Periventricular demyelination and axonal pathology is associated with
subependymal virus spread in a murine model for multiple sclerosis. Intervirology 55, 401–416.
Krakowka S, Higgins RJ, Koestner A, 1980. Canine distemper virus: review of structural and functional
modulations in lymphoid tissues. Am J Vet Res 41, 284-292.
Krakowka S, 1982. Mechanisms of in vitro immunosuppression in canine distemper virus infection. J Clin
Lab Immunol 8, 187-196.
Krakowka S, Ringler SS, Lewis M, Olsen RG, Axthelm MK, 1987. Immunosuppression by canine
distemper virus: modulation of in vitro immunoglobulin synthesis, interleukin release and
prostaglandin E2 production. Vet Immunol Immunopathol 15, 181-201.
Lamb RA, 1993. Paramyxovirus fusion: a hypothesis for changes. Virology, 197, 1–11.
Lambotin M, Raghuraman S, Stoll-Keller F, Baumert TF, Barth H, 2010. A look behind closed doors:
interaction of persistent viruses with dendritic cells. Nat Rev Microbiol 8, 350-360.
Lange C, Dürr M, Doster H, Melms A, Bischof F, 2007. Dendritic Cell-regulatory T-cell Interactions Control
Self-directed Immunity. Immunol Cell Biol 85, 575–581.
Langedijk JPM, Janda J, Origgi FC, Örvell C, Vandevelde M, Zurbriggen A, Plattet P, 2011. Canine
distemper virus infects canine keratinocytes and immune cells by using overlapping and distinct
regions located on one side of the attachment protein. J Virol 85, 11242–11254.
Lanteri M, O'Brien KM, Purtha WE, Cameron MJ, Lund JM, Owen RE, Heitman JW, Custer B, Hirschkorn
DF, Tobler LH, Kiely N, Prince HE, Ndhlovu LC, Nixon DF, Kamel HT, Kelvin DJ, Busch MP,
Rudensky AY, Diamond MS, Norris PJ, 2009. Tregs control the development of symptomatic
West Nile virus infection in humans and mice. J Clin Invest 119, 3266-3277.
Li L, Liu D, Hutt-Fletcher L, Morgan A, Masucci MG, Levitsky V, 2002. Epstein-Barr virus inhibits the
development of dendritic cells by promoting apoptosis of their monocyte precursors in the
http://www.ncbi.nlm.nih.gov/pubmed?term=Raghuraman%20S%5BAuthor%5D&cauthor=true&cauthor_uid=20372157
http://www.ncbi.nlm.nih.gov/pubmed?term=Vandevelde%20M%5BAuthor%5D&cauthor=true&cauthor_uid=21849439

123
Chapter 9: References
presence of granulocyte macrophage-colony-stimulating factor and interleukin-4. Blood 99, 3725–
3734.
Li MZ, Xu FD, Huang XH, Chen XC, Deng QW, Liu ST, Liu Y, Xu LM, Wang HS, Cui JJ, 2008.
[Immunosuppression induced by measles virus in adult patients is not related to CD4+ CD25+
regulatory T cell induction]. Zhonghua Shi Yan He Lin Chuang Bing Du Xue Za Zhi 22, 211–213.
Liesz A, Suri-Payer E, Veltkamp C, Doerr H, Sommer, C Rivest S, Giese T, Veltkamp R, 2009.
Regulatory T cells are key cerebroprotective immunomodulators in acute experimental stroke.
Nat Med 15, 192–199.
Lim HW, Hillsamer P, Banham AH, Kim CH, 2005. Cutting edge: Direct suppression of B Cells by
CD4+CD25+ regullatory T cells. J Immunol 175, 4180-4183.
Lipscomb TP, Schulman FY, Moffett D, Kennedy S, 1994. Morbilliviral disease in Atlantic bottlenose
dolphins (Tursiops truncatus) from the 1987-1988 epizootic. J Wildl Dis 30, 567–571
Liu H, Hu B, Xu D, Liew FY, 2003. CD4+CD25+ regulatory T cells cure murine colitis: the role of IL-10,
TGF-beta, and CTLA4. J Immunol 171, 5012–5017.
Liu J, Gong N, Huang X, Reynolds AD, Mosley RL, Gendelman HE, 2009. Neuromodulatory activities of
CD4+CD25+ regulatory T cells in a murine model of HIV-1 associated neurodegeneration. J
Immunol 182, 3855–3865.
Liuzzi GM, Riccio P, Dal Canto MC, 1995. Release of myelin basic protein-degrading proteolytic activity
from microglia and macrophages after infection with Theiler’s murine encephalomyelitis virus:
comparison between susceptible and resistant mice. J Neuroimmunol 62, 91–102.
Lowther DE, Hafler DA, 2012. Regulatory T cells in the central nervous system Immunol Rev 248, 156–
169.
Ludewig B, Odermatt B, Landmann S, Hengartner H, Zinkernagel RM, 1998. Dendritic cells induce
autoimmune diabetes and maintain disease via de novo formation of local lymphoid tissue. J Exp
Med 188, 1493–1501.
Lund MJ, Hsing L, Pham TT, Rudensky Y, 2008. Coordination of early protective immunity to viral
infection by regulatory T cells. Science, 320, 1220–1224.
Luo X, Tarbell KV, Yang H, Pothoven K, Bailey SL, Ding R, Steinman RM, Suthanthiran M, 2007.
Dendritic cells with TGF-beta1 differentiate naive CD4+CD25- T cells into islet-protective Foxp3+
regulatory T cells. Proc Natl Acad Sci USA 104, 2821–2826.
Lutz MB, Schuler G, 2002. Immature, semi-mature and fully mature dendritic cells: which signals induce
tolerance or immunity? Trends in Immunol 23, 445–449.
Löffler S, Lottspeich F, Lanza F, Azorsa DO, ter Meulen V, Schneider-Schaulies J, 1997. CD9, a
tetraspan transmembrane protein, renders cells susceptible to canine distemper virus. J Virol 71,
42–49.
http://www.ncbi.nlm.nih.gov/pubmed?term=Suri-Payer%20E%5BAuthor%5D&cauthor=true&cauthor_uid=19169263

124
Chapter 9: References
Lühn K, Simmons CP, Moran E, Dung NT, Chau TN, Quyen NT, Thao le TT, Van Ngoc T, Dung NM, Wills
B, Farrar J, McMichael AJ, Dong T, Rowland-Jones S, 2007. Increased frequencies of CD4+
CD25(high) regulatory T cells in acute dengue infection. J Exp Med 204, 979–985.
Macatonia SE, Hosken NA, Litton M, Vieira P, Hsieh CS, Culpepper JA, Wysocka M, Trinchieri G, Murphy
KM, O’Garra A, 1995. Dendritic cells produce IL-12 and direct the development of Th1 cells from
naïve CD4 T cells. J Immunol 154, 5071–5079.
MacDonald AJ, Duffy M, Brady MT, McKiernan S, Hall W, Hegarty J, Curry M, Mills KH, 2002. CD4 T
helper Type 1 and regulatory T cells induced against the same epitopes on the core protein in
hepatitis C virus-infected persons. J Infect Dis 185, 720–727.
Mantovani A, Sica A, Locati M, 2005. Macrophage polarization comes of age. Immunity. 23, 344–346.
Marik C, Felts PA, Bauer J, Lassmann H, Smith KJ, 2007. Lesion genesis in a subset of patients with
multiple sclerosis : a role for innate immunity? Brain 130, 2800–2815.
Markus S, Failing K, Baumgärtner W, 2002. Increased expression of pro-inflammatory cytokines and lack
of up-regulation of anti-inflammatory cytokines in early distemper CNS lesions. J Neuroimmunol
125, 30–41.
Massacesi L, 2002. Compartmentalization of the immune response in the central nervous system and
natural history of multiple sclerosis Implications for therapy. Clin Neurol Neurosurg 104, 177–181.
McMahon EJ, Bailey SL, Castenada CV, Waldner H, Miller SD, 2005. Epitope spreading initiates in the
CNS in two mouse models of multiple sclerosis. Nat Med 11, 335–339.
McMenamin PG, 1999. Distribution and phenotype of dendritic cells and resident tissue macrophages in
the dura mater, leptomeninges, and choroid plexus of the rat brain as demonstrated in
wholemount preparations. J Comp Neurol 405, 553–562.
Meinl E, Krumbholz M, Derfuss T, Junker A, Hohlfeld R, 2008. Compartmentalization of inflammation in
the CNS: a major mechanism driving progressive multiple sclerosis. J Neurol Sci 274, 42–44.
Mielcarek M, Torok-Storb B, Storb R, 2011. Pharmacological immunosuppression reduces but does not
eliminate the need for total-body irradiation in nonmyeloablative conditioning regimens for
hematopoietic cell transplantation. Biol Blood Marrow Transplant 17, 1255–1260.
Miele JA, Krakowka S, 1983. Antibody responses to virion polypeptides in gnotobiotic dogs infected with
canine distemper virus. Infect Immunity 41, 869–871.
Mikita J, Dubourdieu-Cassagno N, Deloire MS, Vekris A, Biran M, Raffard G, Brochet B, Canron MH,
Franconi JM, Boiziau C, Petry KG, 2011. Altered M1/M2 activation patterns of monocytes in
severe relapsing experimental rat model of multiple sclerosis Amelioration of clinical status by M2
activated monocyte administration. Multi Scler 17, 2–15
Mikloska Z, Bosnjak L, Cunningham AL, 2001. Immature monocyte-derived dendritic cells are
productively infected with herpes simplex virus type 1. J Virol 75, 5958–5964.
http://www.ncbi.nlm.nih.gov/pubmed?term=Thao%20le%20TT%5BAuthor%5D&cauthor=true&cauthor_uid=17452519
http://www.ncbi.nlm.nih.gov/pubmed?term=Van%20Ngoc%20T%5BAuthor%5D&cauthor=true&cauthor_uid=17452519
http://www.ncbi.nlm.nih.gov/pubmed?term=McMichael%20AJ%5BAuthor%5D&cauthor=true&cauthor_uid=17452519
http://www.ncbi.nlm.nih.gov/pubmed?term=McMichael%20AJ%5BAuthor%5D&cauthor=true&cauthor_uid=17452519
http://www.ncbi.nlm.nih.gov/pubmed?term=MacDonald%20AJ%5BAuthor%5D&cauthor=true&cauthor_uid=11920289

125
Chapter 9: References
Mills KHG, 2004. Regulatory T cells: friend or foe in immunity to infection? Nat Rev Immunol 4, 841–855.
Minagawa H, Tanaka K, Ono N, Tatsuo H, Yanagi Y, 2001. Induction of the measles virus receptor SLAM
(CD150) on monocytes. J Gen Virol 82, 2913–2917.
Misra N, Bayry J, Lacroix-Desmazes S, Kazatchkine MD, Kaveri SV, 2004. Cutting edge: human
CD4+CD25+ T cells restrain the maturation and antigen-presenting function of dendritic cells. J
Immunol 172, 4676–4680.
Miyara M, Sakaguchi S, 2007. Natural regulatory T cells: mechanisms of suppression. Trends Mol Med
13, 108–116.
Moore PF, Schrenzel MD, Affolter VK, Olivry T, Naydan D, 1996. Canine cutaneous histiocytoma is an
epidermotropic Langerhans cell histiocytosis that expresses CD1 and specific beta 2-integrin
molecules. Am J Pathol 148, 1699–1708.
Moore PF, 2008. Immunological concepts applied to pathologic diagnosis of proliferative diseases of the
immune system. Cancer Ther 6, 263–270.
Mori T, Shin YS, Okita M, Hirayama N, Miyashita N, Gemma T, Kai C, Mikami T, 1994. The biological
characterization of field isolates of canine distemper virus from Japan. J Gen Virol 75, 2403–
2408.
Moro L, de Sousa Martins A, de Moraes Alves C, de Araújo Santos FG, dos Santos Nunes JE, Carneiro
RA, Carvalho R, Vasconcelos AC, 2003. Apoptosis in canine distemper. Arch Virol 148, 153–164.
Murabayashi N, Kurita-Taniguchi M, Ayata M, Matsumoto M, Ogura H, Seya T, 2002. Susceptibility of
human dendritic cells (DCs) to measles virus (MV) depends on their activation stages in
conjunction with the level of CDw150: role of Toll stimulators in DC maturation and MV
amplification. Microbes Infect 4, 785–794.
Mühlebach MD, Mateo M, Sinn PL, Prüfer S, Uhlig KM, Leonard VH, Navaratnarajah CK, Frenzke M,
Wong XX, Sawatsky B, Ramachandran S, McCray PB Jr, Cichutek K, von Messling V, Lopez M,
Cattaneo R, 2012. Adherens junction protein nectin-4 is the epithelial receptor for measles virus.
Nature 480, 530-533.
Naniche D, Varior-Krishnan G, Cervoni F, Wild TF, Rossi B, Rabourdin-Combe C, Gerlier D, 1993.
Human membrane cofactor protein ( CD46 ) acts as a cellular receptor for measles virus.
Microbiology 67, 6025–6032.
Navarrete-Talloni MJ, Kalkuhl A, Deschl U, Ulrich R, Kummerfeld M, Rohn K, Baumgärtner W, Beineke A,
2010. Transient peripheral immune response and central nervous system leaky
compartmentalization in a viral model for multiple sclerosis. Brain Pathol 20, 890–901.
Nesseler A, Baumgärtner W, Gaedke K, Zurbriggen A, 1997. Abundant expression of viral nucleoprotein
mRNA and restricted translation of the corresponding viral protein in inclusion body
polioencephalitis of canine distemper. J Comp Pathol 116, 291–301.

126
Chapter 9: References
Nesseler A, Baumgärtner W, Zurbriggen A, Orvell C, 1999. Restricted virus protein translation in canine
distemper virus inclusion body polioencephalitis. Vet Microbiol 69, 23–28.
Nonacs R, Humborg C, Tam JP, Steinman RM, 1992. Mechanisms of mouse spleen dendritic cell
function in the generation of influenza-specific cytolytic T lymphocytes. J Exp Med 176, 519-529.
Noyce RS, Richardson CD, 2012. Nectin 4 is the epithelial cell receptor for measles virus. Trends
Microbiol 20, 429-439.
Ohgimoto K, Ohgimoto S, Ihara T, Mizuta H, Ishido S, Ayata M, Ogura H, Hotta H, 2007. Difference in
production of infectious wild-type measles and vaccine viruses in monocyte-derived dendritic
cells. Virus Res 123, 1–8.
Okada H, Kobune F, Sato TA, Kohama T, Takeuchi Y, Abe T, Takayama N, Tsuchiya T, Tashiro M, 2000.
Extensive lymphopenia due to apoptosis of uninfected lymphocytes in acute measles patients.
Arch Virol 145, 905–920.
Oldstone MBA, Lewicki H, Thomas D, Tishon A, Dales S, Patterson J, Manchester M, Homann D,
Naniche D, Holz A, 1999. Measles virus infection in a transgenic model: and central nervous
system disease. Cell 98, 629–640.
Orlando EA, Imbschweiler I, Gerhauser I, Baumgärtner W, Wewetzer K, 2008. In vitro characterization
and preferential infection by canine distemper virus of glial precursors with Schwann cell
characteristics from adult canine brain. Neuropathol App Neurobiol 34, 621–637.
Osterhaus A, Groen J, Spijkers HE, Broeders HW, UytdeHaag FG, de Vries P, Teppema JS, Visser IK,
van de Bildt MW, Vedder EJ, 1990. Mass mortality in seals caused by a newly discovered virus-
like morbillivirus. Vet Microbiol 23, 343–350.
Pasare C, Medzhitov R, 2003. Toll pathway-dependent blockade of CD4+CD25+ T cell-mediated
suppression by dendritic cells. Science 299, 1033–1036.
Paust S, Lu L, McCarty N, Cantor H, 2004. Engagement of B7 on effector T cells by regulatory T cells
prevents autoimmune disease. Proc Natl Acad Sci USA 101, 10398–10403.
Patel JR, Heldens JG, Bakonyi T, Rusvai M, 2012. Important mammalian veterinary viral
immunodiseases and their control. Vaccine 30, 1767–1781.
Pearce-Kelling S, Mitchell WJ, Summers BA, Appel MJ, 1990. Growth of canine distemper virus in
cultured astrocytes: relationship to in vivo persistence and disease. Microb Pathogen 8, 71–82.
Perrin PJ, Scott D, Quigley L, Albert PS, Feder O, Gray GS, Abe R, June CH, Racke MK, 1995. Role of
B7:CD28/CTLA-4 in the induction of chronic relapsing experimental allergic encephalomyelitis. J
Immunol 154, 1481–1490.
Piccirillo CA, Shevach EM, 2001. Cutting edge: control of CD8+ T cell activation by CD4+CD25+
immunoregulatory cells. J Immunol 167, 1137–1140.
http://www.ncbi.nlm.nih.gov/pubmed?term=Zurbriggen%20A%5BAuthor%5D&cauthor=true&cauthor_uid=10515265

127
Chapter 9: References
Pinteaux-Jones F, Sevastou IG, Fry VA, Heales S, Baker D, Pocock JM, 2008. Myelin-induced microglial
neurotoxicity can be controlled by microglial metabotropic glutamate receptors. J Neurochem
106, 442–454.
Plattet P, Rivals JP, Zuber B, Brunner JM, Zurbriggen A, Wittek R, 2005. The fusion protein of wild-type
canine distemper virus is a major determinant of persistent infection. Virology 337, 312–326.
Plattet P, Cherpillod P, Wiener D, Zipperle L, Vandevelde M, Wittek R, Zurbriggen A, 2007. Signal
peptide and helical bundle domains of virulent canine distemper virus fusion protein restrict
fusogenicity. J Virol 81, 11413–11425.
Pringle CR, 1999. Virus taxonomy at the XIth international congress of virology, Sydney, Australia, Arch
Virol 144, 2065–2070.
Syme R, Bajwa R, Robertson L Stewart D, Glück S, 2005. Comparison of CD34 and monocyte-derived
dendritic cells from mobilized peripheral blood from cancer patients. Stem Cells 23, 74–81.
Rai AK, Thakur CP, Singh A, Seth T, Srivastava SK, Singh P, Mitra DK, 2012. Regulatory T cells
suppress T cell activation at the pathologic site of human visceral leishmaniasis. PloS One 7,
pe31551.
Relland LM, Williams JB, Relland GN, Haribhai D, Ziegelbauer J, Yassai M, Gorski J, Williams CB, 2012.
The TCR repertoires of regulatory and conventional T cells specific for the same foreign antigen
are distinct. J Immunol 189, 3566–3574.
Reuter D, Sparwasser T,Hünig T, Schneider-Schaulies J, 2012. Foxp3+ regulatory T cells control
persistence of viral CNS infection. PLoS One 7, pe33989.
Richards MH, Getts MT, Podojil JR, Jin YH, Kim BS, Miller SD, 2012. Virus expanded regulatory T cells
control disease severity in the Theiler’s virus mouse model of MS. J Autoimmun 36, 142–154.
Ricklin Gutzwiller ME, Moulin HR, Zurbriggen A, Roosje P, Summerfield A, 2010. Comparative analysis of
canine monocyte- and bone-marrow-derived dendritic cells. Vet Res 41, 40.
Ricklin ME, Roosje P, Summerfield A, 2010. Characterization of canine dendritic cells in healthy, atopic,
and non-allergic inflamed skin. J Clin Immunol 30, 845–854.
Rima KB, Duffy N, Mitchell JW, Summers AB, Appel GJM, 1991. Correlation between humoral immune
responses and presence of virus in the CNS in dogs experimentally infected with canine
distemper virus. Arch Virol 121, 1–8.
Rinaldo CR, Piazza P, 2004. Virus infection of dendritic cells: portal for host invasion and host defense.
Trends Microbiol 12, 337–345.
Rothwell NJ, 1997. Neuroimmune interactions : the role of cytokines. Brit J Pharmacol 121, 841–847.
Rudd PA, Bastien-Hamel LE, Von Messling V, 2010. Acute canine distemper encephalitis is associated
with rapid neuronal loss and local immune activation. J Gen Virol 91, 980–989.
http://www.ncbi.nlm.nih.gov/pubmed?term=Zurbriggen%20A%5BAuthor%5D&cauthor=true&cauthor_uid=15893783
http://www.ncbi.nlm.nih.gov/pubmed?term=Cherpillod%20P%5BAuthor%5D&cauthor=true&cauthor_uid=17686846
http://www.ncbi.nlm.nih.gov/pubmed?term=Vandevelde%20M%5BAuthor%5D&cauthor=true&cauthor_uid=17686846
http://www.ncbi.nlm.nih.gov/pubmed?term=Zurbriggen%20A%5BAuthor%5D&cauthor=true&cauthor_uid=17686846
http://www.ncbi.nlm.nih.gov/pubmed?term=Sparwasser%20T%5BAuthor%5D&cauthor=true&cauthor_uid=22448284
http://www.ncbi.nlm.nih.gov/pubmed?term=H%C3%BCnig%20T%5BAuthor%5D&cauthor=true&cauthor_uid=22448284

128
Chapter 9: References
Rudd PA, Cattaneo R, Von Messling V, 2006. Canine distemper virus uses both the anterograde and the
hematogenous pathway for neuroinvasion. J Virol 80, 9361–9370.
Röthlisberger A, Wiener D, Schweizer M, Peterhans E, Zurbriggen A, Plattet P, 2010. Two domains of the
V protein of virulent canine distemper virus selectively inhibit STAT1 and STAT2 nuclear import. J
Virol 84, 6328–6343.
Saliki TJ, Cooper JE, Gustavson PJ, 2002. Emerging morbillivirus infections of marine mammals. Ann NY
Acad Sci 5, 51–59.
Sakaguchi S, 2003. Regulatory T cells: mediating compromises between host and parasite. Nat Immunol
4, 10–11.
Sakaguchi S, Ono M, Setoguchi R, Yagi H, Hori S, Fehervari Z, Shimizu J, Takahashi T, Nomura T, 2006.
Foxp3+ CD25+ CD4+ natural regulatory T cells in dominant self-tolerance and autoimmune
disease. Immunol Rev 212, 8–27.
Salomon B, Lenschow DJ, Rhee L, Ashourian N, Singh B, Sharpe A, Bluestone JA, 2000. B7/CD28
costimulation is essential for the homeostasis of the CD4+CD25+ immunoregulatory T cells that
control autoimmune diabetes. Immunity 12, 431–440.
Sarris M, Andersen KG, Randow F, Mayr L, Betz AG, 2008. Neuropilin-1 expression on regulatory T cells
enhances their interactions with dendritic cells during antigen recognition Immunity 28, 402–413.
Sato H, Yoneda M, Honda T, Kai C, 2012. Morbillivirus receptors and tropism: multiple pathways for
infection. Front Microbiol 3, 75.
Schmid E, Zurbriggen A, Gassen U, Rima B, ter Meulen V, Schneider-Schaulies J, 2000. Antibodies to
CD9 , a tetraspan transmembrane protein, inhibit canine distemper virus-induced cell-cell fusion
but not virus-cell fusion. J Virol 74, 7554–7561.
Schneider-Schaulies S, Klagge IM, ter Meulen V, 2003. Dendritic cells and measles virus infection. Curr
Topics Microbiol Immunol 276, 77–101.
Schneider-schaulies S, ter Meulen V, 2002. Measles virus and immunomodulation : molecular bases and
perspectives Measles virus and immunomodulation. Exp Rev Mol Med 4, 1–18.
Schneider-Schaulies S, Bieback K, Avota E, Klagge I, ter Meulen V, 2002. Regulation of gene expression
in lymphocytes and antigen-presenting cells by measles virus: consequences for
immunomodulation. J Mol Med 80, 73–85.
Schneider-Schaulies S, Dittmer U, 2006. Silencing T cells or T-cell silencing: concepts in virus-induced
immunosuppression. J Gen Virol 87, 1423–38.
Schnorr JJ, Xanthakos S, Keikavoussi P, Kämpgen E, ter Meulen V, Schneider-Schaulies S, 1997.
Induction of maturation of human blood dendritic cell precursors by measles virus is associated
with immunosuppression. Proc Natl Acad Sci USA 94, 5326–5331.
http://www.ncbi.nlm.nih.gov/pubmed?term=Zurbriggen%20A%5BAuthor%5D&cauthor=true&cauthor_uid=20427537
http://www.ncbi.nlm.nih.gov/pubmed?term=Bluestone%20JA%5BAuthor%5D&cauthor=true&cauthor_uid=10795741
http://www.ncbi.nlm.nih.gov/pubmed?term=Zurbriggen%20A%5BAuthor%5D&cauthor=true&cauthor_uid=10906209

129
Chapter 9: References
Schobesberger M, Zurbriggen A, Doherr MG, Weissenböck H, Vandevelde M, Lassmann H, Griot C,
2002. Demyelination precedes oligodendrocyte loss in canine distemper virus-induced
encephalitis. Acta Neuropathol 103, 11-19.
Schobesberger M, Summerfield A, Doherr MG, Zurbriggen A, Griot C, 2005. Canine distemper virus-
induced depletion of uninfected lymphocytes is associated with apoptosis. Vet Immunol
Immunopathol 104, 33–44.
Seehusen, F, Orlando, EA, Wewetzer, K, Baumgärtner, W, 2007. Vimentin-positive astrocytes in canine
distemper: a target for canine distemper virus especially in chronic demyelinating lesions? Acta
Neuropathol 114, 597–608.
Seehusen F, Baumgärtner W, 2010. Axonal pathology and loss precede demyelination and accompany
chronic lesions in a spontaneously occurring animal model of multiple sclerosis. Brain Pathol 20,
551–559.
Sellin CI, Jégou JF, Renneson J, Druelle J, Wild TF, Marie JC, Horvat B, 2009. Interplay between virus-
specific effector response and Foxp3 regulatory T cells in measles virus immunopathogenesis.
PloS One 4, e4948.
Serra P, Amrani A, Yamanouchi J, Han B, Thiessen S, Utsugi T, Verdaguer J, Santamaria P, 2003. CD40
ligation releases immature dendritic cells from the control of regulatory CD4+CD25+ T cells.
Immunity 19, 877–889.
Servet-Delprat C, Vidalain PO, Bausinger H, Manié S, Le Deist F, Azocar O, Hanau D, Fischer A,
Rabourdin-Combe C, 2000. Measles virus induces abnormal differentiation of CD40 ligand-
activated human dendritic cells. J Immunol 164, 1753–1760.
Shevach EM, 2002. CD4+ CD25+ suppressor T cells: more questions than answers. Nat Rev Immunol 2,
389–400.
Shishkova Y, Harms H, Krohne G, Avota E, Schneider-Schaulies S, 2007. Immune synapses formed with
measles virus-infected dendritic cells are unstable and fail to sustain T cell activation. Cell
Microbiol 9, 1974–1986.
Silva SJ, Andrade CA, Santana CC, Santos QL, de Oliveira IC, Vassallo J, dos-Santos CLW, 2012. Low
CXCL13 expression, splenic lymphoid tissue atrophy and germinal center disruption in severe
canine visceral leishmaniasis. PLoS ONE 7, e29103.
Simon-Martínez J, Ulloa-Arvizu R, Soriano VE, Fajardo R, 2008. Identification of a genetic variant of
canine distemper virus from clinical cases in two vaccinated dogs in Mexico. Vet J 175, 423–426.
Sips G, Chesik D, Glazenburg L, Wilschut J, De Keyser J, Wilczak N, 2007. Involvement of morbilliviruses
in the pathogenesis of demyelinating disease. Rev Med Virol 17, 223–244.
http://www.ncbi.nlm.nih.gov/pubmed?term=Zurbriggen%20A%5BAuthor%5D&cauthor=true&cauthor_uid=11837742
http://www.ncbi.nlm.nih.gov/pubmed?term=Vandevelde%20M%5BAuthor%5D&cauthor=true&cauthor_uid=11837742
http://www.ncbi.nlm.nih.gov/pubmed?term=Zurbriggen%20A%5BAuthor%5D&cauthor=true&cauthor_uid=15661329
http://www.ncbi.nlm.nih.gov/pubmed?term=Yamanouchi%20J%5BAuthor%5D&cauthor=true&cauthor_uid=14670304
http://www.ncbi.nlm.nih.gov/pubmed?term=Santamaria%20P%5BAuthor%5D&cauthor=true&cauthor_uid=14670304
http://www.ncbi.nlm.nih.gov/pubmed?term=Mani%C3%A9%20S%5BAuthor%5D&cauthor=true&cauthor_uid=10657621

130
Chapter 9: References
Skripuletz T, Lindner M, Kotsiari A, Garde N, Fokuhl J, Linsmeier F, Trebst C, Stangel M, 2008. Cortical
demyelination is prominent in the murine cuprizone model and is strain-dependent. Am J Pathol
172, 1053–1061.
Skripuletz T, Bussmann JH, Gudi V, Koutsoudaki PN, Pul R, Moharregh-Khiabani D, Lindner M, Stangel
M, 2010. Cerebellar cortical demyelination in the murine cuprizone model. Brain Pathol 20, 301–
312.
Smed-Sörensen A, Chalouni C, Chatterjee B, Cohn L, Blattmann P, Nakamura N, Delamarre L,
Mellman I, 2012. Influenza A virus infection of human primary dendritic cells impairs their ability to
cross-present antigen to CD8 T cells. PLoS Pathog 8, e1002572.
Spitzbarth I, Bock P, Haist V, Stein VM, Tipold A, Wewetzer K, Baumgärtner W, Beineke A, 2011.
Prominent microglial activation in the early proinflammatory immune response in naturally
occurring canine spinal cord injury. J Neuropathol Exp Neurol 70, 703–714.
Spitzbarth I, Baumgärtner W, Beineke A, 2012. The role of pro- and anti-inflammatory cytokines in the
pathogenesis of spontaneous canine CNS diseases. Vet Immunol Immunopathol 147, 6–24.
Stasiolek M, Bayas A, Kruse N, Wieczarkowiecz A, Toyka KV, Gold R, Selmaj K, 2006. Impaired
maturation and altered regulatory function of plasmacytoid dendritic cells in multiple sclerosis.
Brain 129, 1293–1305.
Stein VM, Czub M, Schreiner N, Moore PF, Vandevelde M, Zurbriggen A, Tipold A, 2004. Microglial cell
activation in demyelinating canine distemper lesions. J Neuroimmunol 153, 122–131.
Stein VM, Baumgärtner W, Kreienbrock L, Zurbriggen A, Vandevelde M, Tipold A, 2006. Canine
microglial cells: stereotypy in immunophenotype and specificity in function? Vet Immunol
Immunopathol 113, 277–287.
Stein VM, Baumgärtner W, Schröder S, Zurbriggen A, Vandevelde M, Tipold A, 2007. Differential
expression of CD45 on canine microglial cells. J Vet Med A Physiol Pathol Clin Med 54, 314–320.
Stein VM, Schreiner NM, Moore PF, Vandevelde M, Zurbriggen A, Tipold A, 2008. Immunophenotypical
characterization of monocytes in canine distemper virus infection. Vet Microbiol 131, 237–246.
Stein VM, Puff C, Genini S, Contioso VB, Baumgärtner W, Tipold A, 2011. Variations on brain microglial
gene expression of MMPs, RECK, and TIMPs in inflammatory and non-inflammatory diseases in
dogs. Vet Immunol Immunopathol 144, 17–26.
Steinman RM, Nussenzweig MC, 2002. Avoiding horror autotoxicus: the importance of dendritic cells in
peripheral T cell tolerance. Proc Natl Acad Sci USA 99, 351–358.
Steinman RM, 2007. Dendritic cells: versatile controllers of the immune system. Nat Med 13, 1155-1159.
Stern LB, Greenberg M, Gershoni JM, Rozenblatt S, 1995. The hemagglutinin envelope protein of canine
distemper virus (CDV) confers cell tropism as illustrated by CDV and measles virus
complementation analysis. J Virol 69, 1661–1668.
http://www.ncbi.nlm.nih.gov/pubmed?term=Skripuletz%20T%5BAuthor%5D&cauthor=true&cauthor_uid=18349131
http://www.ncbi.nlm.nih.gov/pubmed?term=Skripuletz%20T%5BAuthor%5D&cauthor=true&cauthor_uid=19371354
http://www.ncbi.nlm.nih.gov/pubmed?term=Zurbriggen%20A%5BAuthor%5D&cauthor=true&cauthor_uid=16834995
http://www.ncbi.nlm.nih.gov/pubmed?term=Vandevelde%20M%5BAuthor%5D&cauthor=true&cauthor_uid=16834995
http://www.ncbi.nlm.nih.gov/pubmed?term=Zurbriggen%20A%5BAuthor%5D&cauthor=true&cauthor_uid=17650152
http://www.ncbi.nlm.nih.gov/pubmed?term=Vandevelde%20M%5BAuthor%5D&cauthor=true&cauthor_uid=17650152
http://www.ncbi.nlm.nih.gov/pubmed?term=Schreiner%20NM%5BAuthor%5D&cauthor=true&cauthor_uid=18472370
http://www.ncbi.nlm.nih.gov/pubmed?term=Vandevelde%20M%5BAuthor%5D&cauthor=true&cauthor_uid=18472370

131
Chapter 9: References
Stettler M, Beck K, Wagner A, Vandevelde M, Zurbriggen A, 1997. Determinants of persistence in canine
distemper viruses. Vet Microbiol 57, 83–93.
Stimmer L, Siebert U, Wohlsein P, Fontaine JJ, Baumgärtner W, Beineke A, 2010. Viral protein
expression and phenotyping of inflammatory responses in the central nervous system of phocine
distemper virus-infected harbor seals (Phoca vitulina). Vet Microbiol 145, 23–33.
Sugiura K, Wijewardana V, Fujimoto M, Akazawa T, Yahata M, Mito K, Hatoya S, Inoue N, Inaba T, 2010.
Effect of IL-12 on canine dendritic cell maturation following differentiation induced by granulocyte-
macrophage CSF and IL-4. Vet Immunol Immunopathol 137, 322–326.
Suter SE, Chein MB, von Messling V, Yip B, Cattaneo R, Vernau W, Madewell BR, London CA, 2005. In
vitro canine distemper virus infection of canine lymphoid cells: a prelude to oncolytic therapy for
lymphoma. Clin Cancer Res 11, 1579-1587.
Summers BA, Appel MJ, 1994. Aspects of canine distemper virus and measles virus encephalomyelitis.
Neuropathol Appl Neurobiol 20, 525-534.
Svitek N, von Messling V, 2007. Early cytokine mRNA expression profiles predict Morbillivirus disease
outcome in ferrets. Virology 362, 404–410.
Tada Y, Nagasawa K, Ho A, Morito F, Ushiyama O, Suzuki N, Ohta H, Mak TW, 1999. CD28-deficient
mice are highly resistant to collagen-induced arthritis. J Immunol 162, 203–208.
Tarbell KV, Yamazaki S, Olson K, Toy P, Steinman RM, 2004. CD25+ CD4+ T cells, expanded with
dendritic cells presenting a single autoantigenic peptide, suppress autoimmune diabetes. J Exp
Med 199, 1467–1477.
Tatsuo H, Ono N, Tanaka K, Yanagi Y, 2000. SLAM ( CDw150 ) is a cellular receptor. Nature 406, 893–
897.
Tipold A , Moore P, Zurbriggen A, Burgener I, Barben G, Vandevelde M, 1999. Early T cell response in
the central nervous system in canine distemper virus infection. Acta Neuropathol 97, 45–56.
Tizard IR, 2009 Veterinary immunology: An introduction Sunders Elsevier,´ISBN 978-1-4160-4989-0, p
89-95.
Trifilo MJ, Hahm B, Zuniga EI, Edelmann KH, Oldstone MB, 2006. Dendritic cell inhibition: memoirs from
immunosuppressive viruses. J Infect Dis 194, 3–10.
Tsunoda I, Fujinami RS, 2002. Inside-out versus outside-in models for virus induced demyelination:
axonal damage triggering demyelination. Springer Semin Immunopathol 24, 105–125.
Valencia X, Stephens G, Goldbach-Mansky R, Wilson M, Shevach EM, Lipsky PE, 2006. TNF
downmodulates the function of human CD4+CD25hi T-regulatory cells. Blood 108, 253–261.
Vandevelde M, Kristensen B, Braund KG, Greene CE, Swango LJ, Hoerlein BF, 1980. Chronic canine
distemper virus encephalitis in mature dogs. Vet Pathol 17, 17–29.

132
Chapter 9: References
Vandevelde M, Kristensen F, Kristensen B, Steck AJ, Kihm U, 1982. Immunological and pathological
findings in demyelinating encephalitis associated with canine distemper virus infection. Acta
Neuropathol 56, 1–8.
Vandevelde M, Zurbriggen A, 2005. Demyelination in canine distemper virus infection: a review. Acta
Neuropathol 109, 56–68.
Vanloubbeeck Y, Hostetter J, Jones DE, 2003. The biology of dendritic cells and their potential use in
veterinary medicine. Health 4, 131–142.
Vidalain PO, Azocar O, Rabourdin-Combe C, Servet-Delprat C, 2001. Measle virus-infected dendritic
cells develop immuno-suppressive and cytotoxic activities. Immunobiol 204, 629–638.
Viglietta V, Baecher-Allan C, Weiner HL, Hafler DA, 2004. Loss of functional suppression by CD4+CD25+
regulatory T cells in patients with multiple sclerosis. J Exp Med 199, 971–979
Vignali DA, Collison WL, Workman JC, 2009. How regulatory T cells work. Nat Rev 8, 523–532.
von Messling V, Zimmer G, Herrler G, Haas L, Cattaneo R, 2001. The hemagglutinin of canine distemper
virus determines tropism and cytopathogenicity. J Virol 75, 6418-6427.
von Messling V, Svitek N, Cattaneo R, 2006. Receptor (SLAM [CD150]) recognition and the V protein
sustain swift lymphocyte-based invasion of mucosal tissue and lymphatic organs by a
morbillivirus. J Virol 80, 6084–6092.
von Rüden EL, Avemary J, Zellinger C, Algermissen D, Bock P, Beineke A, Baumgärtner W, Stein VM,
Tipold A, Potschka H, 2012. Distemper virus encephalitis exerts detrimental effects on
hippocampal neurogenesis. Neuropathol App Neurobiol 38, 426–442.
Vulcano M, Albanesi C, Stoppacciaro A, Bagnati R, D'Amico G, Struyf S, Transidico P, Bonecchi R, Del
Prete A, Allavena P, Ruco LP, Chiabrando C, Girolomoni G, Mantovani A, Sozzani S, 2001.
Dendritic cells as a major source of macrophage-derived chemokine/CCL22 in vitro and in vivo.
Eur J Immunol 31, 812–822.
Walker LS, Chodos A, Eggena M, Dooms H, Abbas AK, 2003. Antigen-dependent Proliferation of CD4+
CD25+ Regulatory T Cells in Vivo. J Exp Med 198, 249–258.
Wang YS, Chi KH, Liao KW, Liu CC, Cheng CL, Lin YC, Cheng CH, Chu RM, 2007a. Characterization of
canine monocyte-derived dendritic cells with phenotypic and functional differentiation. Can J Vet
Res 71, 165–174.
Wang YS, Chi KH, Chu RM, 2007b. Cytokine profiles of canine monocyte-derived dendritic cells as a
function of lipopolysaccharide- or tumor necrosis factor-alpha-induced maturation. Vet Immunol
Immunopathol 118, 186–198.
Weber M, Lange C, Günther W, Franz M, Kremmer E, Kolb HJ, 2003. Minor histocompatibility antigens
on canine hemopoietic progenitor cells. J Immunol 170, 5861–5868.
http://www.ncbi.nlm.nih.gov/pubmed?term=Transidico%20P%5BAuthor%5D&cauthor=true&cauthor_uid=11241286
http://www.ncbi.nlm.nih.gov/pubmed?term=Chiabrando%20C%5BAuthor%5D&cauthor=true&cauthor_uid=11241286

133
Chapter 9: References
Wenzlow N, Plattet P, Wittek R, Zurbriggen A, Gröne A, 2007. Immunohistochemical demonstration of the
putative canine distemper virus receptor CD150 in dogs with and without distemper. Vet Pathol
44, 943–948.
Weiss DJ, Wardrop KJ, 2010. Schalm's Veterinary Hematology Wiley-Blackwell, Iowa State, pp: 958-967;
1039-1041.
Wijewardana V, Sugiura K, Oichi T, Fujimoto M, Akazawa T, Hatoya S, Inaba M, Ikehara S, Jayaweera
TS, Inaba T, 2006. Generation of canine dendritic cells from peripheral blood monocytes without
using purified cytokines. Vet Immunol Immunopathol 114, 37–48.
Wright NG, Cornwell HJ, Thompson H, Lauder IM, 1974. Canine distemper: current concepts in
laboratory and clinical diagnosis. Vet Rec 94, 86-92.
Wu L, Liu Y J, 2007. Development of dendritic-cell lineages. Immunity, 26, 741–750.
Wu GF, Laufer TM, 2007. The role of dendritic cells in multiple sclerosis. Curr Neurol Neurosci Rep 7,
245-252.
Wyss-Fluehmann G, Zurbriggen A, Vandevelde M, Plattet P, 2010. Canine distemper virus persistence in
demyelinating encephalitis by swift intracellular cell-to-cell spread in astrocytes is controlled by
the viral attachment protein. Acta Neuropathol 119, 617–630.
Wünschmann A, Alldinger S, Kremmer E, Baumgärtner W, 1999. Identification of CD4+ and CD8+ T cell
subsets and B cells in the brain of dogs with spontanious acute, subacute and chronic-
demyelinating distemper encephalitis. Vet Immunol Immunopathol 67, 101–116.
Wünschmann A, Kremmer E, Baumgärtner W, 2000. Phenotypical characterization of T and B cell areas
in lymphoid tissues of dogs with spontaneous distemper. Vet Immunol Immunopathol 73, 83–98.
Yamazaki S, Bonito AJ, Spisek R, Dhodapkar M, Inaba K, Steinman RM, 2007. Dendritic cells are
specialized accessory cells along with TGF- for the differentiation of Foxp3+ CD4+ regulatory T
cells from peripheral Foxp3 precursors. Blood 110, 4293–4302.
Yanagi Y, Takeda M, Ohno S, 2006. Measles virus: cellular receptors, tropism and pathogenesis. J Gen
Virol 87, 2767–2779.
Yang LJS, Schnaar RL, 2008. Axon regeneration inhibitors. Neurol Res 30, 1047–1052.
Yu X, Cheng YM, Shi BS, Qian FX, Wang FB, Liu XN, Yang HY, Xu QN, Qi TK, Zha LJ, Yuan ZH,
Ghildyal R, 2008. Measles virus infection in adults induces production of IL-10 and is associated
with increased CD4 + CD25 + regulatory T cells. J Immunol 181, 7356–7366.
Zaba LC, Krueger JG, Lowes MA, 2009. Resident and “inflammatory” dendritic cells in human skin. J
Invest Dermatol 129, 302–308.
Zhang X, Lu G, Qi J, Li Y, He Y, Xu X, Shi J, Zhang CW, Yan J, Gao GF, 2013. Structure
of measles virus hemagglutinin bound to its epithelial receptor nectin-4. Nat Struct Mol Biol 20,
67-72. Zheng Y, Rudensky AY, 2007. Foxp3 in control of the regulatory T cell lineage. Nat Immunol 8, 457–462.
http://www.ncbi.nlm.nih.gov/pubmed?term=Zurbriggen%20A%5BAuthor%5D&cauthor=true&cauthor_uid=18039911
http://www.ncbi.nlm.nih.gov/pubmed?term=Gr%C3%B6ne%20A%5BAuthor%5D&cauthor=true&cauthor_uid=18039911
http://www.ncbi.nlm.nih.gov/pubmed?term=Jayaweera%20TS%5BAuthor%5D&cauthor=true&cauthor_uid=16904755
http://www.ncbi.nlm.nih.gov/pubmed?term=Jayaweera%20TS%5BAuthor%5D&cauthor=true&cauthor_uid=16904755
http://www.ncbi.nlm.nih.gov/pubmed?term=canine%20distemper%3A%20current%20concepts%20in%20laboratory
http://www.ncbi.nlm.nih.gov/pubmed?term=Zurbriggen%20A%5BAuthor%5D&cauthor=true&cauthor_uid=20119836

134
Chapter 9: References
Zilliox MJ, Parmigiani G, Griffin DE, 2006. Gene expression patterns in dendritic cells infected with
measles virus compared with other pathogens. Proc Natl Acad Sci USA 103, 3363–3368.
Zou T, Caton AJ, Koretzky GA, Kambayashi T, 2010. Dendritic cells induce regulatory T cell proliferation
through antigen-dependent and -independent Interactions. J Immunol 185, 2790–2799.
Zozulya AL, Wiendl H, 2008. The role of regulatory T cells in multiple sclerosis. Nat Clin Pract Neurol 4,
384–398.
Zurbriggen A, Schmid I, Graber HU, Vandevelde M, 1998. Oligodendroglial pathology in canine
distemper. Acta Neuropathol 95, 71–77.
Örvell C, 1980. Structural polypeptides of canine distemper virus. Arch Virol 206, 193–206.

135
Chapter 10: Acknowledgements
10 Chapter 10: Acknowledgements
I would like to thank:
First of all, my main supervisors,
Prof. Dr. Wolfgang Baumgärtner for excellent mentorship, motivation and
continues support. I appreciate his guidance toward critical thinking, organizing
and giving me confidence for the future. Also, I am very grateful for a friendly and
cosy working environment.
Prof. Dr. Andreas Beineke, for continues advice and supervision, lively
discussions, ideas, continues motivation, and correcting the manuscripts.
Especially for his availability and openness to discuss different ideas and
encouragement for my work.
Special thanks to my co-supervisors, Prof. Dr. Andrea Tipold and Prof. Dr. Martin
Stangel, for their support, constructive comments, and thoughtful suggestions.
Yvonne Barthel and Max Iseringhausen for their collaboration and interesting
discussions that contributed to the realization of joint publication.
Ingo Spitzbarth, Vanessa Herder, Johannes Junginger, Florian Hansmann for
apprehensive reading and thoughtful corrections of the final manuscript.
Rich Roy, for his cordial help in reading and correcting my manuscript and
publications.
Unit for Reproductive Medicine, Small Animal Clinic, Prof. Dr. Anne-Rose
Günzel-Apel and Dr. Carola Urhausen, for providing the blood samples and the
entire staff for their positive and friendly spirit, making me feel welcome each
time I needed the samples.
Small Animal Clinic for providing us the possibility to use their laboratories. PD
Dr. Veronika Stein, for her support and advices in analysing the flow cytometry
data. Regina Carlson and Arianna Maiolini, PhD, for providing me a quick and
efficient help for all the technical problems in the lab. Dr. Marina Hoffmann for
collecting the blood samples.
Special thanks to coordination office of ZSN for continues support.

136
Chapter 10: Acknowledgements
Danuta Waschke, Claudia Herrmann, Kerstin Schöne, Kerstin Rohn, Petra
Grünig, and Bettina Buck for their excellent technical support during the
execution of this project and making the work at lab way more fun.
Dr. Karl Rohn at the Department of Biometry, Epidemiology and Information
Processing at the University of Veterinary Medicine Hannover, for helpful
discussions and statistical advice.
Office mates, Ingo Spitzbarth, for unreserved help and for the hips of answered
questions, ranging from German grammar to molecular analysis, Kristel Kegler
for always relaxed attitude and putting it up with my music, and Oscar for being
such a well behaved and friendly dog.
All the colleagues in the institute, Armend, Kerstin, Barbara, Florian, Vannessa,
Stephi, Christina, Yanyong , Andre, Wenhui, Anika, Johannes, and many others
whom I see on corridors and the others who left in meantime, for their advices,
help and hints for an easier life in the lab, and with whom I shared many smokes,
drinks, laughs and music.
To my small family, Lejla, for her unconditional love, and making me happy all
these years.
To my big family, my mom and dad, Olta, Vali, Laura, Greta, and Liam, for all the
hugs and support they gave me.
Young cell scheme, European Counsel and Kosovo government, Department of
Pathology, Veterinary University, Hannover, Germany, for providing me with a
financial support, and German Research Foundation (FOR 1103) for financing
the project.
All the people that unintentionally I have forgot to mention.


















