
Introduction - Coweeta LTER | Coweeta LTERcoweeta.uga.edu/publications/771.pdf · cardis Ross...
15
Community structure of Trichoptera in a mountain stream: spatial patterns of production and functional organization % C ALEXANDER I). HURYN and .1. BRUCE WALLACE Department of Entomology. University of Georgia. Athens, Georgia SUMMARY. 1. Annual production was estimated' for Trichoptera occurring in each of three distinct habitats of a mountain stream: bedrock-outcrops, riffles and pools. Production was greatest on bedrock-outcrops (2608 mg ash-free dry weight m~ : ), followed by riffles (1038) and pools (950). 2. Annual production in bedrock-outcrops and pools was dominated by single functional groups, with collector-filterers and shredders contributing 73% and 15% of the annual production, respectively. Production in riffles was due primarily to shredders (46%), followed by collector-filterers (27%). 3. Taking account of the amount of stream area occupied by each habitat type, total annual production was estimated at 1336 mg AFDW m~ 2 . 53% of this production was attributable to four taxa: Parapsyche cardis Ross (25%), Pvcnopsyche gentilis (MacLachlan) (10%). Neophy- lax mitchelli Carpenter (9%) and Rhyacophila nigrita Banks (9%). 4. Habitat-weighted production was distributed among functional groups as follows: collector-filterers (41%), shredders (29%). engulfing- predators (\5%). scrapers (13%) and collector-gatherers (2%). 5. The distinct tuxonoinic and functional structures of irichopteran sub-communities were shaped by the distinct physical characteristics of their principal habitats. Bedrock-outcrops were characterized by low roughness and high current and were sites of low deposition or organic matter: thus the predominance of collector-filterers. In contrast, the other habitats of greater roughness (riffles) and/or lower current (pools) were sites of deposition of food (e.g. leaf litter) and greatest shredder production. 6. By distinguishing discrete mesoscale habitats, each with a function- ally distinct caddisfly sub-community, we speculate that small mountain streams provided the diverse physical templates essential for the evolution of the major feeding tactics (e.g. scraping, shredding, filter- feeding) of the. Trichoptera. • ( Jeoreiu rili
Transcript of Introduction - Coweeta LTER | Coweeta LTERcoweeta.uga.edu/publications/771.pdf · cardis Ross...

Community structure of Trichoptera in a mountain stream:
spatial patterns of production and functional organization
% C
A L E X A N D E R I). H U R Y N and .1. B R U C E WALLACEDepartment of Entomology. Univer s i ty of Georgia. Athens, Georgia
SUMMARY. 1. Annual production was estimated' for Trichopteraoccurring in each of three distinct habitats of a mountain stream:bedrock-outcrops, riffles and pools. Production was greatest onbedrock-outcrops (2608 mg ash-free dry weight m~ :), followed by riffles(1038) and pools (950).
2. Annual production in bedrock-outcrops and pools was dominatedby single functional groups, with collector-filterers and shredderscontributing 73% and 15% of the annual production, respectively.Production in riffles was due primarily to shredders (46%), followed bycollector-filterers (27%).
3. Taking account of the amount of stream area occupied by eachhabitat type, total annual production was estimated at 1336 mg AFDWm~2. 53% of this production was attributable to four taxa: Parapsychecardis Ross (25%), Pvcnopsyche gentilis (MacLachlan) (10%). Neophy-lax mitchelli Carpenter (9%) and Rhyacophila nigrita Banks (9%).
4. Habitat-weighted production was distributed among functionalgroups as follows: collector-filterers ( 4 1 % ) , shredders (29%). engulfing-predators ( \ 5 % ) . scrapers (13%) and collector-gatherers ( 2 % ) .
5. The d i s t i nc t tuxono in ic and f u n c t i o n a l s t ruc tures of i r ichopteransub-communi t ies were shaped by the dis t inct physical characterist ics ofthe i r pr incipal hab i ta t s . Bedrock-outcrops were characterized by lowroughness and high current and were sites of low deposition or organicmatter: thus the predominance of collector-filterers. In contrast, theother habi ta ts of greater roughness ( r i f f l e s ) and/or lower current (pools)were sites of deposition of food (e.g. leaf l i t t e r ) and greatest shredderproduction.
6. By distinguishing discrete mesoscale habitats, each with a function-ally distinct caddisfly sub-community, we speculate that small mountainstreams provided the diverse physical templates essential for theevolution of the m a j o r feeding tactics (e .g . scraping, shredding, f i l t e r -feeding) of the. Trichoptera .
•(Jeoreiu rili

Introduction
With the except ion of the Ch i ronomidae (C 'o t ' f -man & Ferrington. 1984; Piniler . 198h). streamdwell ing Trichoptera are genera l ly 'representedby more species t han are other croups of a n i m a l susing the same resources' (Mackay & Wiggins.1979). These authors a t t r ibute th is great ecolo-gical diversity to the secretion of silk which isu t i l i zed in various ways to en-nance explo i ta t ionof resources. Among trichopteran genera, allma jo r funct ional feeding groups are representedand are geographically widespread (Wiggins &Mackay. 1978: Wiggins, 1984): accordingly , thefunc t iona l composition of communi t ies occur-r ing in d i f f e r en t habi ta ts should serve as ageneral index to food a v a i l a b i l i t y and mode of itsacqu i s i t i on .
A l though c e r t a i n l y not a new deve lopment(iiynes. 1971); also see Statzner £ Higler.1985). a recent trend in stream ecology hasbeen to emphasize the influence of hydraulicsand geomorphology (Brussock, Brown & Dix-on, 1985; Statzner & Higler, 1985. 1986:Frissel et al., 1986; Huryn & Wallace, 1987).Channel form determines the spatial distribu-tion of physically distinct habitats, e.g. riffleversus pool, and the mode of food delivery,e.g. entrained versus deposited, to their resi-dent consumers (Brussock et al., 1985; Huryn& Wallace, 1987). The remarkable diversityamong the Trichoptera should enhance theirability to respond at the community level,taxonomically, and functionally to the physi-cally singularities of any given stream.
The objectives of this study were to examinethe autecologies of twenty-two species ofTrichoptera inhabiting upper Ball Creek, inthe southern Appalachian mountains(U.S.A.), and to evaluate the influence ofchannel form on spatial patterns of communitystructure, production, and functional organiza-tion.
Study area
General description
Upper Ball Creek dra ins watershed 27 of theCoweeta Hydrologic Labora to ry (MaconC o u n t y . Nor th C a r o l i n a . U . S . A . ) . Wa te r shed2~ is a 38.8 ha reference ca tchment wi th arugged topography ranc ine from ! !>35 to i J ' ~
m a . s . l . The vegetation of watershed 27 is .,mixed hardwood forest that has remainedgenera l ly und i s tu rbed . Mean a n n u a l precipita-tion approaches 250 cm and is evenly distributed throughout the year (Swank & Douo-lass. 1977). During the s tudy , streamflow \vas
continuous w i t h lowest and highest dischargesin the early autumn and late winter, re-spectively. Because of the perennially coolc l imate , the stream accumulates only 2800-3300 degree days \ ' . A more complete de-scr ipt ion of Ball Creek is given by Huryn &Wallace (1987).
The physical characteristics of the majorh a b i t a t s of Ball Creek were treated in de t a i l b\H u r y n & Wallace (19.87) . However, t he HIOMimportant features are summarized here. Thechanne l form of Ball Creek is heterogeneousand debris regulated, with about 19% bedrock-outcrop, 58% riffle and 23% pool (% totalstream bottom). Current velocities amongthese principal habitats were significantly diffe-rent as follows: bedrock-outcrop > riffle > pool(Fig. 1A).
Bedrock-outcrops were characterized by lowretention coupled with high entrainment ofmaterials as indicated by significantly lowerstanding crops of detritus compared to rifflesand pools. In contrast, pools were characterizedas habitats of relatively high retention and lowtransport. In terms of benthic organic matteraccumulation, riffles were consistently in-termediate to the bedrock-outcrops and pools(Fig. 2A).
Materials and Methods
Benihic sampling
Twenty benthic samples were taken monthly(Ju ly 1983 to June 1984) from randomlyaligned ' location- along a 5 ( H I m section ofBail Creek tha t extended ups t ream from thewate r shed 27 weir . Two methods of samplingwere utilized depending upon local flow andsubstrate characteristics. In pools a 400 cm:
conn;: device was used. In n t f l e areas andmo--s-covered bedrock outcrops , a l>30 cnrSurber Sampler (mesh = 23(J / / m i w a s used. All<.;mp!e^ Here preserved in iS-SO formalin

143
—1
pin.
d SCR
PIG. \. Comparison of habitat characteristics withproduction of selected trichopteran funct ionalgroups in upper Ball Creek: (A) water velocity,biomass of aquatic mosses and median substrateparticle size measured in bedrock-outcrops, r i f f lesand pools. Uni t s of left y-axis are cm s ! for watervelocity and g AFDW m : for mosses. Water veloc-ity and moss hiomass are from H u r y n iV Wallace(19S~) . ( B ) A n n u a l production t m g AFDW m ~ )ca lcu la ted lo r c o l l e c t o r - f i l t e r i n g , -crapci and
Bedrock-outcrop
Riffle Poo!
FIG. 2. Comparison of habitat characteristics withproduction of selected trichopteran func t iona lgroups in upper Ball Creek: (A) biomass of finepaniculate organic mailer (FPOM>1 m m ) , coursepaniculate organic m a t t e r (CPOM> 1 mm exclusiveof wood) and woody mater ia l measured in bedrock-outcrops, r i f f les and pools. Uni t s are mg AFDWm :. Data from Huryn & Wallace (1987). (B)A n n u a l product ion i m g AFDW m : ) measured torcol lcc tor - ia thcrers and shredders .
con ta in ing a small amount of p h l o x i n e B dye.I n v e r t e b r a t e s wore removed f rom samples byhand p i c k i n g under 15x m a g n i f i c a t i o n . Amore d e t a i l e d account of the sampl ing proce-dure is provided by Huryn & Wallace (14X7) .
I't.h' historic* and fiaiciional Jn'clin^ ^rt>iif>assignment
Flight periods of adul t caddisflies were asses-sed by eol lect ins! wi th malaise or emergencetraps. Unless otherwise i n d i c a t e d . l a r \ a l ms tarswere d i s l i n g u i s h e d from h e a d - w i d t h f r e q u e n c yhistograms Seasonal changes i n ! a r \ a l mstarsw<is assessed hv examina t ion ot siiccessi\'esamples , ( n o w t h p a t t e r n s w e r e desc r ibed f romPlots 01 l a r v a l ash-free dry w e i g h t ( A F D W )
(see below) against t ime from i n i t i a l occurr-ence in the benthic samples. Funct ional feed-ing groups were assigned fo l lowing Wiggins(19S4) or personal observation.
Production
Most month ly estimates of larval AFDWwere based upon weights of at least fiveindividuals of the dominant instars present.For first i n s t a r larvae of several taxa. f ivelarvae were w e i g h e d toge ther and the meanweight ca lcula ted . Average mon th ly AFDWwas ca lcu la ted as the sum of the product ofeach in s t a r ' s mean AFDW and the i n s t a r sr e l a t i v e f r e q u e n c v w i t h i n i t s r e spec t ive cohor t .L a r v a e w e i « h e d were selected trom formal in

144
preserved samples , d r ied (>5'O for 24 h anddesiccated (t'a('(.)-,) tor an a d d i t i o n a l 14 h.Dried specimens were weighed to the nearestP.O. on a Calm 23 Electrobalance to obta in drymass, ashed (5()0°C) for 1 h and re weighed toobtain AFDVV.
Larval production was calculated by the sizefrequency method (Hami l ton , 1969) and cor-rected for cohort production in terval (CPI )(Benke . 1979). Cohort production in te rva l swere calculated as the interval in days from thepeak occurrence of f i r s t instar larvae un t i l theonset of pupa t ion . The method of Kreuger &Mar t in (1980) was used to ob ta in 95'V Cl forsize-frequency production est imates . For taxat h a t spent a disproport ionate amount of t imein a par t icular ins tar . the dura t ion of t ha t stagewas subdivided anil each d iv i s ion t reated as aseparate w e i g h t class ( B e n k e iV Wal lace .l^NO). Temporal patterns of production (e.g.Fig. 3A-C) were obtained from mon th ly stand-ing stocks and the instantaneous growth method(Benke, 1984). Growth was assumed to beexponential and growth rates were calculatedfrom changes in average monthly AFDW. Dueto low recovery of early instars of Palaegapetuscelsus (Ross) (Hydroptilidae), production wascalculated for the final instar only using theinstantaneous growth method.
Since all samples were taken at randomly-assigned locations, the resulting mean standingstocks of all samples combined were assumedto be weighted by the relative amount of eachhabitat in the stream. Habitat-weighted pro-duction was obtained by combining all samplesand util izing the grand mean annual density forthe various size classes.
Leaf litter biomass and substrate particle sizedistributions
Leaf litter (>\ cnr) was removed from sam-ples, air-dried, weighed to nearest 0.01 g andhomogenized in LI l abora tory b lende r . Subsam-ples were o \ e n d r i ed (55 : C) lor 24 h. t h e ndesiccated oxer CaCO-, for an additional 24 h.Subsamples were then weighed to obta in drymass, ashed (5()()°C) for about 8 h. thenreweished to obta in '"< AFDW. These datawere used to demons t r a t e m o n t h l y ehanue-- inleaf l i t t e r biomass.
Es t imates of per cent composit ion of sub-s t r a t e by panic le si/e w a s made w i t h i n the area
20 1
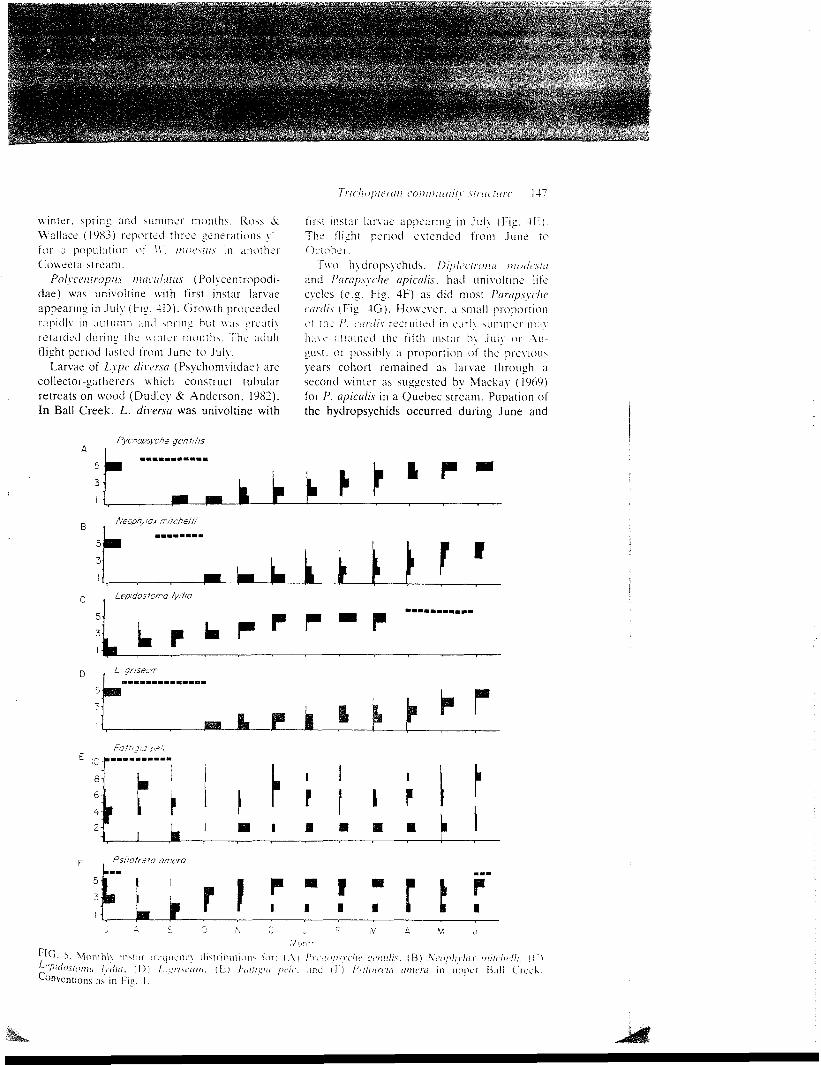
J A
FIG. 3. Temporal patterns of the production ofLepidostoma Ivdia (A). L. griseum (B) and Pycno-psyche gcniilis (C) in the r i f f le (squares) and pool (cir-cles) habitat of upper Ball Creek (July 1983 to June1984).
of each sample u n i t following Gurtz & Wallace(1984) and was used to estimate median Phivalues (Cummins. 1962) for each habitat .
Results
Life histories
Est imat ion of secondary production by thes i / e - f requency method r equ i r e s k n o w l e d g e otthe CPI i .Bcnke . 1984) w i n c h can be d e t e r m i -ned by detailed s tudy of l i f e h is tor ies . 'There-to re the f o l l o w i n g d iscuss ion o f the l i f e rhetor-

145
ies ot the B a l i Creek i n c h o p t e r a n t a u n a is ani n t e g r a l par! of the p roduc t ion da t a providedb e l o w . A O ' l n p i e i c l i s t k i t i n c h o p t c r a i i t a x aand l u n c t i o n a i a s s ignmen t s is p r o v i d e d i nTable 1. Figures of m o n t h l y ins ta r f requencydis t r ibut ions are provided only for those spe-cies for which Mich i n f o r m a t i o n is not read i lya v a i l a b l e i n i h e l i t e r a t u r e .
AV.'iVi i;/'/i:'/;.' inintini i R l u a c o p h i h d a e I t o l -lovved a 12 -mon th deve lopmen ta l period asreported by Mackay (1969) and Manuel cVFolsom ( 1 9 8 2 ) . whereas Rhvacopliilti glabcrri-ina had a short developmental period of about8 months starting in October. Rhyacophilatorva had an unusually long adult flight period(February-August) and a long period ofcontinuous rec ru i tment (Fig. 4A). OtherRhyacophila studied exhibited shorter flightperiods (<2 months) . The life cycle of Rhya-
co/>litlti tii^niti. the most a b u n d a n t Rlt\im>t'>h:luin the s t i idv area, was the most d i f f i c u l t toi n t e r p r e t , i i r s t u i s ta r l a r v a e w e r e p resen tt h r o u g h o u t t h e vea r Mimzestnis; c o n t i n u o u srec ru i tmen t from delayed ha tch ing of eggs (e .« .Thut. 1969). Rhyacophila nigriia and Rhva-cophihi carpenicri. which was rare , wereassumed to be u n i v o i t i n c w i t h ( P i s ot ?4() daw
Prepupae and pupae ot / ' , , / < • , ; : j i f / ' < 7 ! / \ cc!.\u.\occurred in May and J u n e , r e s p e c t i v e l y . Theflight period extended from early June to Ju ly .Development proceeded rapidly with larvaeattaining the penul t imate instar by late Ju ly .All P. celsus reached the final larval instar byAugust and grew steadily until prepupation inMay.
Four species of Philopotamidae occurred inupper Ball Creek: Dolophilodes distincnts, D.major, D. sisko and Wormaldia tnoesia. Lar-
TABLE 1. Habitat-weighted production measured tor Ball Creek Trichoptera. FG = functional group,cg=collector-gatherer, cf=collector-filterer. shr=shredder. scr-scraper, ep=engulfing-predator. CPI = cohortproduction in te rva l . Production (P) = mg AFDW m - y '. CL = 95Cf confidence l i m i t , hiomass (B) = mg AFDWm~~, abundance (A) = individuals m ' ~ .
TaxonRhyacophila carpenteri MilneR. m Carolina Banks 'R. glabherimu UlmerR. minora BanksR. nigrita BanksR. larva HagenPalcas'apciu'i n ' /v /<. \ (Ross)"Dfiliip/iiliuli^ i / /vmii '7 iH ( W a l k e r ) '
FGep—epepepepshref
CPI
365—209299365241_95
P6
—103
12146>1
4
±CL B
4 3— —
10 12 1
37 2325 9
— >12 > 1
A3
—95
10713
—d
P/B
2.3—
9.54.95.35.01.2
1 1 . 1
I). ni:iji>r (B.snk^i'I). v/.vAo t K o s x rWormnldia nme^nix ( B a n k s ) 'H'. mni'Mii.1*'I'(tl\:ti'iii)'t)i>it^ inticitluliis t h a n k sLypc iliversu I B a n k s )Parup.\\'chf Ciirili\ RossP- upicalis ( B a n k s )DipU'Clronu niddt'Shi BanksP\cnops\cht' wniilis MacLachlanP- sono ( M i l n e )Nfophyltu Miicht'l/i CarpenterCoerita scmaia Ross"Lepidostoma '^riscitm ( B a n k s )L. lydia RussMulannn hlcmii: S iMcv 'Psiloireiu iiincn: (Ross )Pseu(li>f>t>cra .^iiii>nliiri.\ C a r p e n t e rFauiflia pelt- I Ross)Sum
serscrshrshr
shrepshr
213655246241
4812
2940393d54232
132552416
141740
6.43 .74.54.45.08.44.312.72.78.16.4
3 . S2.33.3
'Ra re in uppci H a l l C i e e k . noi C k i n s u l e r e d i n t h e p r e s e n t s u u l v . p r o k l n e t i k U iinstantaneous growth: ' summer cohort: ' au tumn-sp r ing cohort: ' l l u r v n tV Wallace (

14(1 Alexander D. Hurvn and J. Bruce Wallace
I I p I ' '
L- w . r r '
i u
Doiophilodes aisftncfus
iL
Wormaldio moes'u'S
;
Porapsyche apicolis
P cardis
JMonth
FIG. 4. Monthly instar frequency distributions for: (A) Rhyacophila torva. (B) Dolophilodes distinaus, (C)\\'ormaldiu moestus, (D) Polycentropus maculatus. (E) Lype diversa, (F) Parapsyche apicalis, and (G) P. cardtsin upper Ball Creek (July 1983 to June 1984). Instars are designated 1-5. Width of each bar represents thepercentage of total individuals (combined from 20 samples m ' ) occurring in a given instar on each samplingdale. Dashed lines indicate fl ight periods.
vac of D. major occupy regions of under-ground flow (personal observation), and wereencountered rarely. Although adults of D.sisko were collected in emergence traps (lateJ u l y to A u g u s t s , larvae were n e \ e r d is t inguishedand may have been mis- ident i f ied as /).ilistincius. Dolophilodes ftisiinciiix was bivoltine!Fi». 4Bi. with a short summer cohort and a
au tu inn-spr ins ; cohort. Prepupae. pupae
and adults were colleclcd throughout the spring.summer and winter months. Benkc & Wallace(1980) and Ross & Wallace (1983) reportedmultivolt inism for D. distinctu.s. Wormaldianiocaui had at least two distinct cohorts (Fig.4C). ( i rowth of the autumn-spr ing cohort wasrapid dtirine October and November but sio\\er.yet relatively constant, from January to June.Pupae and a d u l t s were collected th roughou t the
147
win te r , spring and summer months . Ross &Wallace (1983) reported three generat ions y 'for a popula t ion of II ' . mocxiua in anotherCo\\:eeta stream.
Polvcemropus maciiUnus (Polycentropodi-dae) was un ivo l t ine with first instar larvaeappearing in J u l y (Fig. 4D). Growth proceededn'.pidh in a u t u m n and s p r i n g hu t was g r e a t l vretarded d u r i n g t h e w i n t e r m o n t h s . T h e a d u l tflight period lasted from June to Ju ly .
Larvae of Lype divcrsa (Psychomyiidae) arecollector-gatherers which construct t ubu la rretreats on wood (Dudley & Anderson, 1982).In Ball Creek, L. diversa was univoltine with
Pycnopsyche gen til is
f i r s t i n s t a r larvae appearing in J u l y (Fig. 4 E ) .The f l i gh t period extended from June toOctober.
Two hydropsychids. Diplecirviui in<iilt'auiand Parapsvche apicalis. had un ivo l t ine l i fecycles (e.g. Fig. 4F) as did most Parapsvcheciinlis (Fig . 4G). However, a small proportionol the /'. ciirdis r ec ru i ted in ea r l ) s u m m e r mayh a \ c a t t a i n e d the f i f t h i n s t a r b\ J u l y o r Au-gus t , or possibly a proportion of the previousyears cohort remained as larvae through asecond winter as suggested by Mackay (1969)for P. apicalis in a Quebec stream. Pupation ofthe hydropsychids occurred during June and
L gnseurr
11 • i f : l i : . i
Psi/otreto omero

14S Alexander 1). ilur\n and .1. Bruce Wallace
J u l y . The f l igh t period extended trom June 10September.
Pvcnopsvche geniilis ( L i m n e p h i h d a e ) andI'venopsvche son.so, were univoltine (Fi». 5A).First ins tar larvae of I', gcntilis appeared inSeptember and October and grew steadilythrough June. Pupation occurred in Augustand the flight period extended from August toOctober. Pvcnopsvche sonso followed a similarpat tern. Neophvlax initcliclli (Limnephi l idae)was univol t ine with an extended period ofaestivation through the summer months (Fig.5B). First instar larvae appeared in Octoberand were most abundan t by November.Growth was slow through January butincreased a b r u p t l y from Februan u n t i l theclosure of the larval case in June . The f l igh tperiod extended from August to October.
Lepidoslonni grisciun and Lipidoslo/iui l\diu(Lepidostomaiidae) exhibi ted discrete univol-tine cohorts (Figs. 5C, D). First instar larvaeof L. griseum appeared in October andpupated in late July. The flight period extendedfrom July to October. In contrast, first instarlarvae of L. lydia appeared in July and pupatedthe following March and April. The flight periodof L. lydia extended from April to June.Although adults Theliopsyche melas (Lepi-dostomatidae) were collected in an emer-gence trap in early June (1983 and 1984), larvaewere never found.
The head-width frequency-histogram for theBall Creek population of Fattigia pele (Serico-stomatidae) did not reveal discrete larval in-stars and, therefore, a range of size classes wasselected to follow development. The presenceof at least two cohorts on each collection dateindicated a 2-year life cycle (Fig. 5E), althoughsome individuals may be univoltine. Semivol-tinism is reported for the European S. persona-nun Spence (Iverson, 1973). Pupae of F. pelesvere found throughout the summer and theflight period extended from early July to lateAugus t .
Psilotreia aniera (Odon toce r idae j had anapparently univoltine life cycle. First instarlarvae appeared in August, and by December,two size groups consisting predominantly ofins ta r 11 and instar V larvae were present (Fig.5F) . A l t h o u g h the slowly developing groupapparent ly reached the f i n a l larval instar bylate summer , pupae were seen only early June.I t is possible that a port ion of the popu la t ion
may require 2 years to mature (see Para-ps\-che, above) . Females ca r ry ing ex t ruded eggmasses were observed in ear ly J u l y . Lowdensities of I'seudoyoera .lingnltiris (Odon-toceridae) made conclusions about its l i fe cycletenuous , a l though a 2-year cycle was indicatedAdults were reared from field collected pupaein late Ju ly .
Habitat-weighted production: toxonomicaspects
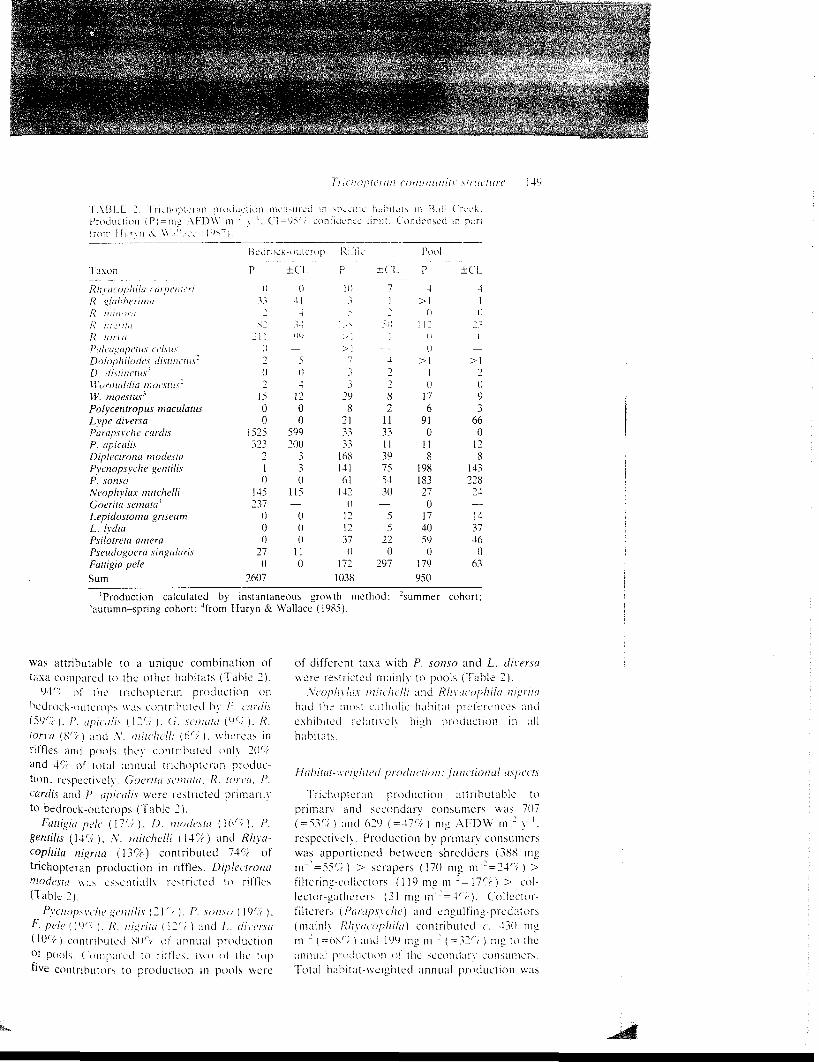
Total habi ta t -weighted trichopteran produc-tion was 1330 mg AFDW m : y ' wi th a meanbiomass of 275 mg AFDW m : (Table i)(P B = 5). These values do not include case andnet s i lk w h i c h may represent about 20rr ofeaddisfly production (Iverson. 1980: Huryn &Wallace. 1985). Four taxa c o n t r i b u t e d 5?'; Oftotal trichopteran production: /'. cardis ( 2 5 r i ) .P. geniilis (10 rf) . N. mitchelli (9 r r ) and R.nigrita (9%). Fattigia pele (8%), D. modesta(7%), P. apicalis (7%), P. sonso (6%). Goeri-la semata (4%), R. torva (3%) and P. amera(3%) contributed an additional 38%. Mostproduction was attributable to taxa with highfinal larval weights (i.e. P. cardis, F. pele), ora combination of short CPIs, high IGRs, andhigh densities (i.e. P. gentilis, N. mitchelli).
Neophylax mitchelli was the most abundanteaddisfly in the stream (134±17 ind. m~2)(X ±SE, 7J=240) followed by R. nigrita(119114 ind. rrf2). Other taxa had substan-tially lower abundances (Table 1). Totaltrichopteran density was 706 m"2.
Production to biomass ratios (P/B) generallyreflected the CPIs assigned to the various taxaand, depending upon voltinism, fell within theexpected ranges cited by Waters (1977) forvarious aquatic invertebrates (Table 1). Un-usually low ratios were observed for R. carpen-teri, L. diversa, P. cetsus and P. aniera (Table2). and are attributed to accelerated growthearly in the l i fe cycle followed by a prolongedperiod in the f i n a l ins ta r which resu l ted in highbiomass (e.g. Figs. 4E. 5F).
'•• Habitat-specific production: taxonomic aspects
Trichopteran production (mg AFDW m~~y ') was greatest on bedrock-outcrops (2608)followed by riffles (1038) and pools (950)(Table 2). Product ion on bedrock-outcrops
149
TaxonRhvacophila carpcntcriR. "luhhi'rimaR. iniitortiR. i;!^n;,iR. uirvuPii/fugapetiis ff/.w/xDolophilodes distinciM'D. disiinciu.1''Wormuldia mocsnix'W. moestus*Polycentropus maculatustype diverseParapsvche cardi.sP. apicalisDiplecirona modestaPvcnopsvche gentilisP. sonsoNeophylax mitchelliGoerita semaia*Lepidostoma griseumL. lydiaPsilolreta ameraPseudogoera singularisFattigia peleSum
BainP
( I33-i,sj
2 1 !0-i0"i
1500
1525323
010
145237
000
270
2607
Kk-OUUTOp
±CL0
414
34W
—504
1200
599200
330
115—
000
110
Kin ' lcP
I D"̂5
^ N> i>1
733
298
213333
16814161
1420
1212370
1721038
±CL71-i
511I
—4•)-)82
11331139755430
—55
220
297
Pool
P4
>1( I
i 1 1( i(1
>110
176
910
118
198183270
1740590
179950
±CL41
( i^ ^
i i—
>1->093
660
128
143228
24—1437460
63
'Production calculated by instantaneous growth method: 2summer cohort;"'autumn-spring cohort; ''from Huryn & Wallace (19S5).
was attr ibutable to a unique combination oftaxa compared to the other hab i t a t s (Table 2).
94''; of the t r ichopteran product ion onbedrock-outcrops was c o n t r i b u t e d by /'. airdis(59%) . P. apicalis ( 1 2 % ) . C. scnuiia ( 9 % ) . R.iorni (8r; ) and .V. niiichclli (6rv ). whereas inriffles and pools they con t r ibu ted only 20'iand 4'/( of to ta l annual t r ichopteran produc-tion, respectively, (ioeriru scmutii. R. torva, P.canlix and P. npiculis were restricted p r imar i lyto bedrock-outcrops (Table 2).
Fattigia pele ( 1 7 r ; ) . D. modesta (Wt). P.gentilis ( 14Cr ) , A1, mitchelli ( \ 4 r ' t ) and Rhya-cophila nigrita ( 13%) contributed 749; oftrichopteran production in riffles. Diplectrnna'nodcsia was es sen t i a l ly restricted to r iff les(Table 2) .
Pycnopxychc iH'ntilis (21', ). P. \on.vo ( 19'V ),F. pele (}<•)'", ) . R. ni'^rita ( \ 2 ' " r ) and /,. dirersu(H) ' -v) con t r ibu ted SO'V of a n n u a l production°t pools. Compared to r i t f l e s . t w o oi the topfive contr ibutors to production in pools were
of different taxa with P. sonso and L. diversawere restr icted m a i n l y to pools (Table 2).
:\'coph\'lax niiichclli and Rlivucophila nigritahad the most c a t h o l i c h a b i t a t p re fe rences ande x h i b i t e d r e l a t i \ x ' l y high product ion in a l lhab i ta t s .
Habitat-u'cightetl production: functional aspects
Trichopteran production a t t r i b u t a b l e topr imary and secondary consumers was 707( = 53r; ) and 629 ( = 47% ) mg AFDW m : y ' .respectively. Production by pr imary consumerswas apportioned between shredders (388 mgm :=?5 rr) > scrapers (170 mg m' : = 241"^) >f i l te r ing-col lec tors ( 1 1 9 mg m":= 17 rt) > col-lector-gatherers (31 mg m - = 4'"r). Collector-fi l terers (Parapsvchc) and enaul f ing-preda tors( m a i n i \ Rhvacophilu) con t r ibu ted c. 430 mgm : (=nS 'V ) and 199 mg m ~ (=32 'V ) mg to thea n n u a l product ion of the secondary consumers.Total habi ta t -weighted annual production was

150 Alexander I.). Hurvn and J. Bruce Wallace
broken down among f u n c t i o n a l feeding groupsas follows: col lector-f i l terers (548 mg m ~ =4 1 ' r ) > shredders (388 mg m -- = 29 rr) > en-gulfing-predators (199 mg m ' ~ = l 5 f 7 r ) > scrapers(170 mg rrf:= 13rP ) > collector-gatherers (31
Habitat-specific production: functional aspects
Quali ta t ively , the funct ional structure of thetrichopteran communi ty occurring in pools,riffles and bedrock-outcrops could he brokendown in three groups. Production in pools andon bedrock-outcrops was dominated by singlefunct ional groups (Figs. I B . 2B). Shreddersand collector-fi l terers contr ibuted 75</v andiy/e of the a n n u a l production in pools andbedrockoutcrops. respect ively (Figs. IB . 2B).whereas production in riffles was more evenlyapportioned among the various funct ionalgroups (Figs. IB. 2B).
Leaf litter biomass and substrate particle sizedistributors
The annual pattern of leaf litter biomass inpools and riffles exhibited a distinct peak inautumn (Fig. 6). Apparently, litter accumu-lated in riffles (October) and later accumulatedin pools (November-December). Leaf litterbiomass on bedrock-outcrops was negligible.
The median Phi values for substrates of eachof the principal habitats exhibited the follow-ing trend: bedrock-outcrop > riffle > pool.Median Phi values are given in Fig. 1A.
Biology of dominant taxa
Collector-filterers
Both Parapsvche apicalis and Parapsvchecardis exhibited s ign i f ican t ly higher productionon bedrock-outcrops t h a n r iff les and pools (see95 f r C'ls. Table 2). w i t h /'. cardis e x h i b i t i n g asignif icant ly higher level of production than P.apicalis (Table 2). Production of Diplectroiwmodesta occurred almost exclusively in riffles,where it was s ign i f i can t ly greater than produc-tion measured for other h \dropsychids . In BallCreek, a ra ther obvious separation of habi ta tbetween the Arctopsychinae and Diplectroni-nae is apparent , wi th the former p r i m a r i l y
A s 0 N D J FMonth
FIG. 6. Temporal patterns of leaf l i t ter accumulationin the riffle (A) and pool (B) habitat of upper BallCreek (July 1983 to June 1984). Error bars = one95% CI. Samples of the pool habitat in October andJune were inadequate (n = \) and are not figured.Note the v-axis is scaled differently in A and B.
utilizing moss covered bedrock-outcrops andthe latter occurring upon unconsolidatedrubble. A similar habitat distribution wasobserved for the hydropsychid community ofHugh White Creek, a lower elevation streamat Coweeta (Gurtz & Wallace. 1986).
Other than the Hydropsychidae, collector-filtering caddisflies contributed little to totalproduction. The bivoltine philopotamids weredominated by Wormaldia moestiis. Dolophilo-des distinctus exhibited significantly lower pro-duction. This pat tern was s imi lar to thatobserved by Ross & Wallace (1983) with W.nujestus the d o m i n a n t p h i l o p o t a m i d at thehighest a l t i tude site which was replaced by D.distinctus throughout the remainder of theDryman Fork basin. Ross & Wallace (1983)observed a rather consistent level of philopota-mid production along Dryman Fork and a t t r i -buted this to the u n i f o r m supp ly of u l t ra f ineorganic seston th roughout the s t ream con-t i n u u m .

151
Lt'vulouoina I\'dia exhib i ted highest abso-lu te g rowth from December to March , aperiod which coincided wi th the period ofgreatest litter biomass in Ball Creek (cf. Figs.3A and 6). Although, temporal and spatialpatterns of production of L. Ivdia indicates aclose l i n k w i t h changes in l i t t e r biomass o therl i t t e r sh redd ing taxa fo r w h i c h t e m p o r a l pat-terns of production are ava i l ab le , did notindicate clear relationships between litteravai labi l i ty and periods of maximum produc-tion (cf. Figs. 3B, C and 6).
Production of L. griseum also peaked inpools during the period of maximum litterbiomass; however, maximum monthly produc-tion occurred in riffles from March to June(Fig. 3B). Densities of L. griseum in rifflesduring this time were correlated significantlywith li t ter biomass (/•=().32, P>0.025. «=45),indicating that larval abundance was related tosize of litter patches. Grafius & Anderson(1980) reported similar biologies for Lepido-stoma cascadense (Milne) and Lepidostomaunicolor (Banks) which co-occur in riffle andpool areas of Oregon streams.
The temporal and spatial pattern of produc-tion of Pycnopsyche gentilis was remarkablysimilar to that of L. griseum and was highest inpools during periods of peak li t ter biomass (cf.Figs. 3C and 6). However, maximum monthlyproduction occurred in riffles during March-June, a period of low l i t t e r standing crop. I ncontrast to L. griseum, P. gentilis densities andl i t t e r biomass in r i f f l e samples t aken d u r i n gMarch. Apr i l . May and J u n e , were not corre-lated s ign i f i can t ly ( r=U.()2 . / '>(>.25. n = 45).indicating no relationship between larval densi-ty and size of l i t t e r patch. During th i s period,instar V larvae constructed m i n e r a l cases incontrast to leaf-disc cases of earl ierdevelopmental stages. Data for P. sonso werenot adequate to compare temporal productionpatterns with P. gentilis a l though growth pat-terns appeared similar. Unl ike P. gentilis. allinstars of P. sonso occurred mainly in pools(Table 2). The o ther shredding t r i c h o p t e r a nwith appreciable production, f-'iittigia pele.occur w i th in fine to relatively course sedimentsand are associated w i t h p a r t i a l l y buried l i t t e r(personal observa t ion) which appa ren t l y is notava i lab le to o ther t r i chop te ran t a \ a . w i t h thepossible exception of Psilotreia ameni.
Most shredders, t he re fo re , had spatial andtemporal patterns of production which w'ere:( i ) e i ther closeh l i n k e d to periods and loca-tions of l i t t e r accumula t ion , i .e . L. l\dia. or(i i) showed two phases: a period of relativelyhigh production which coincided with l i t teraccumulation in pools and an intense period ofproduction in r i f f l e areas which were low l i t t e rbiomass. Dur ing l a t e sp r ing , ava i lab le l i t t e rconsists of p r imar i ly robust and slowly con-ditioned leaves such as Rhododendron.Perhaps movement from pools to riffles by P.gentilis and L. griseum is related to util ization ofthese initially more refractory materials.
Scrapers
Despite heavy shading by the seasonallydense forest canopy and high ridges surround-ing the Ball Creek catchment, substantialproduction of scraping Trichoptera was found(13% of total). Goerita semata contributed toscraper production only on bedrock-outcrops(Huryn & Wallace, 1985). Most production ofN. mitchelli and G. semata occurred fromMarch until May and was correlated with risingstream temperatures as seasonal insolationincreased. As much as 849r of the productionof G. semata occurred during this period, and549r of the annual production was attributedto consumption of diatoms (Huryn & Wallace,1985).
Collector-gatherers
The only caddisfly designated a collector-gatherer was L\pe divcrsu which is well knownfor its association with submerged wood (e.g.Wiggins. 1977). A n n u a l production wasgreatest in pools where wood was mostabundant.
Engulfing-predators
Production of engulf ing-predators . R/iva-cophila spp., was low with R. nigrita and R.lorra being most productive. Rhvacophila nig-rita was remarkab ly widespread, w i t h similarlevels of production (c. K M ) mg AFDW m :) inal l h a b i t a t s sampled (Table 2) . F ina l i n s t a rlarvae ot R. nigrita are large (c. 3-10 x the

Alexander D. Hur\n utnl J. Bruce \\'ullace
AFDW of the other Rhvucophilu spp.) . acharacterist ic which may faci l i ta te r e l a t i \ c i \rapid movement through diverse h a b i t a t s ,Rhvacoplula lorva occurred almost exclus ivelyon bedrock-outcrops where its production wassigni f icant ly higher than that of R. nigriia. Theother Rhvacoplula species contributed l i t t l e tototal engulfing-predator production and wereconfined primarily to riffles (Table 2). Pseudo-goera singularis contributed little to engulfing-predator production and. l ike R. lorva. wasexclusively confined to bedrock-outcrops.
Discussion
The a b i l i t y of a stream channel to re ta in andcycle dissolved and paniculate materials inf lu-ences consumer production (Leopold. 1941).Retention may be mediated physically throughcharacteristics of channel form or biotically byactivities of consumers. Additionally, channelform and the resultant habitat structure servesas a physical template which is a major deter-minant of the structure and functional feedinggroup composition of the trichopteran com-munity. Production of the discrete trichopteransub-communities found in pools, riffles and onbedrock-outcrops is strongly subsidized by theprimary physically characteristics of these dis-tinctive meso-scale habitats.
Physical retention is a function of channelform (Likens & Bilby, 1982; Brussock et at..1985) and catchment history (Likens & Bilby,1982; Molles. 1982). Bilby & Likens (1980)and Bilby (1981) demonstrated that pooledareas upstream of debris dams are important inregulating release of particles. In Ball Creek,accumulation of leaf l i t ter in pools during lateautumn indicates that pools are primary sitesfor physical retention and storage of organicma t t e r . Accordingly, shredder production pre-domina ted in pools (12': . Fig. 2) andcollector-f i l terer production was min imal (4'<.Fig. 1). Trichopteran production in riffles wasdistributed more evenly among functionalgroups than in pools and on bedrock-outcrops,which reflects the greater d ivers i ty of physicalconditions w i t h i n this hab i t a t . The m a j o r pro-portion of production in r iff les (46% ) wascont r ibuted bv shredders (c t . Fiszs. I B and 2 B ) .
m a i n l y F. pele and P. !><'m;7/.v. Apparently thephysical r e t e n t i o n of lea! l i n e r predominatedtranspor t character is t ics in r i f f l e s , at least atbaseflow. Al though a s u b s t a n t i a l amount ofthe production by P. gentiUs occurred in poolsdur ing a u t u m n , the spatial pattern of produc-tion shifted from pools to riffles during early-spring and coincided with the decline of leaflitter in the former habitat. Ta.xa occupyingpools and, to a lesser degree, riffles, areexploiting areas of stream channel that aresites of physical re tent ion.
An obvious form of biotic retention is medi-ated by collector-f i l ter ing organisms which in-gest suspended mate r ia l s . 42% of the totaltrichopteran production in Ball Creek wasat t r ibutable to collector-t'ilterers (Table 1).Similar or h igher proportions of collector-f i l terer production appear to be general forstream reaches where there are suitable sitesfor retreat attachment [e.g. outcrop (Nelson &Scott, 1962; Flossner, 1976), cobble (Neves,1979; Kreuger & Waters, 1983), snags andemergent macrophytes (Cudney & Wallace,1980;"Smock Gilinsky & Stoneburner, 1985)].In Ball Creek, the dependence of the collector-filtering caddisfly community upon regions ofadequate current and stable attachment siteswas shown by the relatively high productionattained in bedrock-outcrop habitats whichwere characerized as shallow sluiceways ofwater over moss-covered bedrock-outcrops.Low physical retention and high potential forentrainment of materials over these outcropscan be inferred from their lower standing cropsof accumulated organic matter compared withriffles and pools (Fig. 2A: Huryn & Wallace,1987). Furthermore, the thick moss coverapparently provides retreat sites for Para-psyche, particularly those of P. cardis.Although const i tut ing only 19% of the streamarea, bedrock-outcrops contibuted 64% of theproduction of fi l ter-feeding Trichoptera.Collector-t'ilterers. th rough use of si lken nets,p r i m a r i l y exploi t e ros ional c h a r a c t e r i s t i c s ofthe bedrock-outcrop h a b i t a t .
Production of cngulf ing-predators followedthat of collector-filterers. being greatest onbedrock-outcrops > riff les > pools (Fig. 1).Engult ' ing-predator production on outcropswas p r imar i l y a t t r i b u t a b l e to the habi ta tspecialist, R. lorva. The specific adaptationsand t roph ic re la t ions of R. lorva are unc l ea r

Trichoplcran community structure 153
and. at present, we have no simple explanat ionfor the above trend of engulfing-predator pro-duction among h a b i t a t s .
Physical ly s table minera l surfaces are mostsuited for epilithon attachment and growth(Wallace & Gurtz . 19S6). Trichoptera whichconsume e p i l i t h o n f r e q u e n t l y have mouthpar ts(Cummins & Mer r i t t . h<>4) and case forms(Mol les . 19S4) t h a t are h i g h l y adapted toexploit physical characteristics of such localenvironments. Therefore, the contribution ofscrapers to total tr ichopteran production inBall Creek is probably related to the amountof stream area occupied by stable mineralsurfaces (Fig. 1). Scraper production in thebedrock-outcrop and riffle habitats was sig-nificantly higher than rates measured in pools.
Importance of habitat: evolutionaryconsiderations
It has often been proposed that the Tricho-ptera originated in cool lotic habitats (Ross,1967; Lepneva. 1969: Wiggins. 1977: Mackay& Wiggins, 1979); however, the implications ofstream channel form with respect to the evolu-tion of the functional diversity found amongthe recent fauna have never been developed(cf. Ross, 1967; Mackay & Wiggins, 1979;Weaver & Morse. 1986). The present study, bydistinguishing discrete mesoscale habi ta ts eachw i t h a f u n c t i o n a l l y d i s t inc t caddisfh sub-communi ty , demonstrates thai small streamss i tua ted in m o u n t a i n o u s t e r r a in may haveprovided the diversity of physical templatesessential for the in i t ia l adaptive radiation ofthe Trichoptera. H e a d w a t e r streams in moun-ta inous regions tend to be r emarkab ly diversein channel form and habitat structure, thuspresenting suites of u n i q u e selective pressurestha t , on an evo lu t ionary t ime scale, may con-strain and shape the functional adaptations oftheir fauna . These site-specific factors, whencombined with the potential for isolation ofindividual catchments in rugged and moun-tainous landscapes for long periods of t ime(Ross. 1950. 1972). as wel l as the appa ren t lylow vagility of many stream inhabi t ing insectsoccurring in such region-- (e .g. Wallace, V'ogel& Cuffney. 1986) ma) have promoted thel s<>lation of small popula t ions leading to theevolution of taxa representing the ancestors of
al l the present-day funct ional feeding groups.Subsequent periods of dispersal, perhaps re-lated to coalescence of catchments and streamsdur ing prolonged periods of downwast ing.would allow for development of the functional-ly dis t inct sub-communities presently occupy-ing physica l ly discrete habi ta ts of mountainstreams. These ecological!) diverse trichopter-an communi t i e s may have in turn providedfortui tously adapted forms which invaded themore spatially homogenous habitats such asponds, lakes and larger rivers and resulted intheir characteristically more derived fauna(Ross. 1956).
Weaver & Morse (1986) presented a revisedphylogeny based upon a more logical andwell-documented systematic analysis than thewidely cited phylogeny of Ross (1967). Theseauthors stated that the ancestral trichopteranprobably occurred along 'aquatic environmentssuch as littoral regions, slowly moving water-ways, or marshes'. In essence, it was impliedthat this hypothetical animal occurred withindetritus rich margins of depositional zones ofthe ancestral environment. Similar to those ofthe present, the mountain streams of Pangeamust have had substantial regions of organicmatter deposition within and along their mar-gins to provide just the type of habitat deducedto be ancestral by Weaver & Morse (1986). Itseems that the ecological aspects of the phy-logeny proposed by these authors are consis-tent w i t h the more t r ad i t iona l and welldocumented idea that the Trichoptera evolvedin association wi th low-order, cool, moderatelyhigh-gradient woodland streams (Ross. 1956.1967; Lepneva. 1969; Wiggins. 1977; Mackay& Wienins . 1979; l l enn i " . 1981).
Acknowledgments
This study was supported by grants from theNational Science Foundation (BSR-8516497),Georgia Power Company, and the Southeast-ern Forest Experiment Station. U.S.D.A.Forest Service. The authors express t h a n k s toB. C a n n a m e l l a . I. Goldman. E. Jakubowskiand A. Lingle for assistance in the laboratoryand Held . Drs D. A. Crossley. A. Hildrew..1. L. Meyer and an anonymous reviewerprovided const ruct ive cr i t ic ism of earl ier ver-sions of the manuscript .

154 Alexander D. Huryn and J. Bruce Wallace
References
Bcnkc A.C. (1979) A modif icat ion of the Hynesmethod for secondary production w i t h par t iculars ignif icance for mul t ivo l t ine populat ions. Limnol-ogy and Oceanography. 24, 168-174.
Bcnkc A.C. (1984) Secondary production of aquatic-insects. Ecology of Ac/italic Insects (Eds V. I I .Rcsh and D. M. Rosenberg), pp. 289-322. Prae-gcr Publishers, New York.
Bcnke A.C. & Wallace J.B. (1980) Trophic basis ofproduction among net-spinning caddisflies in asouthern Appalachian stream. Ecology. 61, 108-118.
Bilby R.E. (1981) Role of organic debris dams inregulating export of dissolved and paniculatemat te r from a forested watershed. Ecology, 62,1234-1243.
Bilby R.E. & Likens G.E. (1980) Importance oforganic debris dams in the structure and funct ionof stream ecosystems. Ecology. 61, 1107-1113.
Brussock P.P.. Brown A.V. & Dixon J.C. (1985)Channel form and stream ecosystem models.Water Resources Bulletin. 21, 859-866.
Coffman W.P & Ferrington L.C. (1984) Chironomi-dae. An Introduction to the Aquatic Insects ofNorth America, 2nd edn (Eds R. W. Merritt andK. W. Cummins), pp. 551-652. Kendall-Hunt,Dubuque, Iowa.
Cuclney M.D. & Wallace J .B . (1980) Life cyclesmicrodistribution and production dynamics of sixspecies of net-spinning caddisflies in a largesoutheastern (U.S.A.) river. Holarctic Ecologv,3, 169-182.
Cummins K.W. (1962) An evaluation of some tech-niques for the collection and analysis of benthicsamples with special emphasis on lotic waters.American Midland Naturalist, 67, 477-504.
Cummins K.W. & Merritt R.W. (1984) Ecology anddistribution of aquatic insects. An Introduction tothe Aquatic Insects of North America, 2nd cdn(Eds R. W. Merritt and K. W. Cummins), pp.59-65 Kendall-Hunt, Dubuque. Iowa.
Dudley T. & Anderson N.H. (1982) A survey ofinvertebrates associated with debris in aquatichabitats. Melanderia 39, 1-21.
Flossner D. (1976) Biomasse und Produktion desMakrobenthos der mittleren Saale. Limnologica,10. 123-153.
Frissell C.A.. Liss W.J . . Warren C.E. & HurleyM.D. (1986) A hierarchical framework forstream habitat classification: viewing streams in awatershed context. Environmental Management,10, 199-214.
Graf ius E .J . <k Anderson N . H . (19801 Popu la t iondynamics and role of two species of Lepiilosionm(Trichoptera: Lepidostomatidae) in an Oregonconiferous forest stream. Ecologv. 61, 808-816.
Gurtz M.E. & Wallace J .B. (1984) Substrate-mediated response of stream invertebrates todis turbance . Ecology. 65. 1556-1559.
Gurtz M.E. & Wallace J .B . (1986) Subst ra tum-production relationships in ne t - sp inn ing cad-disflies (Tr ichoptera) in d is turbed and undis-
turbed hardwood catchments . Journal of theNorth American Hentliological Society. 5, 230-236.
Hami l ton A.L. (1969) On e s t i m a t i n g annua l produc-t ion . Limnology and Oceanography, 14, 771-78?
Hennig W.H. (1981) . Insect Phytogeny. Wiley NewYork.
Huryn A.D. & Wallace J .B . (1985) Life history andproduction of Gocrila semata (Trichoptera: Lirn-nephil idae) in the southern Appalachian Moun-tains. Canadian Journal of Zoology, 63, 2604-2611.
Huryn A.D. & Wallace J.B. (1987) Local geomor-phology as a determinant of macrofaunal produc-tion in a mountain stream. Ecology, 68, 1939-1942.
Hynes H.B.N. (1970) The Ecology of RunningWaters. Un ive r s i ty of Toronto Press, Toronto.
Iverson T.M. (1973) Life cycle and growth ofScricostoma personatinn Spence (TrichopteraSericostomatidae) in a Danish spring. Entomolo-gica Scandinavica, 4, 323-327.
Iverson T.M. (1980) Densi t ies and energetics of twostrcamliving larval populations of Sencosiomapersonatum (Trichoptera) . Holarctic Ecology, 3,65-73.
Kreuger C.C. & Martin F.B. (1980) Computation ofconfidence intervals for the size-frequencv(Hynes) methods of estimating secondary pro-duction. Limnology and Oceanography. 25, 773-777.
Krueger C.C. & Waters T.F. (1983) Annual pro-duction of macroinvertebrates in three streams ofdifferent water quality. Ecology, 64, 840-850.
Leopold L.B. (1941) Lakes in relation to terrestriallife patterns. Readings in Ecology (Ed. E. J.Kormondy), pp. 200-203. Prentice-Hall. NewJersev.
Lepneva S.G. (1969) Fauna SSSR, Rucheiniki.Vol. 1. Lichinki i kukolki podotryadakol'chatoshchupikovykh. Zoologicheskii InstitutAkademii Nauk SSSR. n.s. 88. [In Russian).Translated into English as: Fauna of the USSR;Trichoptera 1 (2) . Larvae and pupae of Annuli-palpia. Published by the Israel Program forScientific Translations, 1970.
Likens G.E. & Bilby R.E. (1982) Development,maintenance, and role or organic-debris dams inNew England streams. In: Swanson F.J., JandaR.J. . Dunne T.. Swanston D.N. (Eds). Work-shop on sediment budgets and rou t ing in foresteddrainage basins: proceedings. Gen. Tech. Rep.PNW-141. USDA. Forest Service. Pacific North-west Forest and Ranac Experiment Station. Port-land . Or.
Mackay R.J. (l%9) Aquatic insect communities of asmall stream on Mont St. Hilaire. Quebec. Jour-nal of the Fisheries Research Board of Canada,26, 1157-1183.
Mackay R.J . & Wiggins G.B. (1979) Ecologicald ivers i ty in Trichoptera. Annual Review of En-tomology, 24, 185-208.
Manuel ICL. & Folsom T.C. (1982) I n s t a r sizes, l i fecycles, and food habi t s of f ive Rhyacophila

Trichopteran communirc structure 155
(Trichoptcra: Rhyacophlidae) species from theAppalachian Mountains of South Carolina.U.S.A. Hydrobiologia. 97, 381-285.
Molles M.C.. Jr (1982) Trichopteran communi t ies ofstreams associated wi th aspen and coni fer forests:long term structural change. Ecology, 63, 1-6.
Molles M.C.. Jr (1984) Patterns of case b u i l d i n g bycaddisflies in the southern Rocky Mounta ins .Proceedings of /lie 4th International Symposiumon Trichoptera (F id . J. C. Morse), pp. 235-244.J u n k . The Hague.
Nelson D.J. & Scott D.C. (1962) Role of d e t r i t u s inthe product iv i ty of a rock-outcrop communi ty ina piedmont stream. Limnology and Oceanogra-phy, 7, 396-413.
Neves R.J. (1979) Secondary production of ep i l i th icfauna in a woodland stream. American MidlandNaturalist. 102, 209-224.
Finder L.C.V. (1986) Biology of freshwater Chir-onomidae. Annual Review of Entomology, 31,1-23.
Ross D.H. & Wallace J .B. (1983) Longitudinalpatterns of production, food consumption andseston u t i l i z a t i o n by net-spinning caddisflies(Trichoptera) in a southern Appalachian stream(U.S.A.). Holaraic Ecology, 6, 270-284.
Ross H.H. (1950) Synoptic notes on some nearcticlimnephilid caddisflies (Trichoptera: Limnephili-dae). American Midland Naturalist. 43, 410-429.
Ross H.H. (1956) Evolution and Classification of theMountain Caddis/lies. University of Illinois Press.Urbana.
Ross H.H. (1967) The evolution and past dispersalof the Trichoplera. Annual Review of Entomolo-gy, 12, 169-206.
Ross H.H. (1972) The origin of species diversity inecological communities. Taxon, 21, 253-259
Smock L.A.. Gi l insky E. & Stoneburner D.L.(1985) Macroinvertebrate production in a south-eastern Uni ted States b lackwater stream. Ecolo-gy. 66. 1491-1503.
Statzner B. <k Higler B. (1985) Questions andcomments on the River Cont inuum Concept.Canadian Journal of Fisheries and Aquatic Scien-ces. 42, 103.S-1044.
Statzner B. & Higler B. (1986) Stream hydraulics asa major de te rminant of hcnthic inver tebra te
zonation patterns. Freshwater Biology. 16, 12/-139.
Swank VV.T. & Douglass .I.E. (1977) N u t r i e n tbudgets for undisturbed and manipula ted hard-wood forest ecosystems in the m o u n t a i n s ofNorth Carolina. Watershed Research in EasternNorth America (Ed. D. L. Cornell) , pp. 343-364.Smithsonian. Washington. D.C.
Thut R.N. (1969) Feeding habi t s of la rvae of sevenRhyacophila (Trichoptera: R h y a c o - p h i l i d a e ) spe-cies wi th notes on other l i f e -h i s to ry fea tures .Annals of the Entomological Society of America.62, 894-898.
Wallace J .B. & Gurtz M.E. (1986) Response ofBaelis mayflies to catchment logging. AmericanMidland Naturalist, 115, 25-41.
Wallace J.B.. Vogal D.S. & Cuffney T.F. (1986)Recovery of a headwater stream from aninsecticide-induced community disturbance. Jour-nal of the North American Bentliological Society.5. 115-126.
Waters T.F. (1977) Secondary production in inlandwaters. Advances in Ecological Research. 10,91-164.
Weaver J.S. & Morse J.C. (1986) Evolution offeeding and case-making behaviour in Trichop-tera. Journal of the North American Benthologic-al Society, 5, 150-158.
Webster J .R. (1983) The role of benthic macro-invertebrates in detri tus dynamics of streams: acomputer simulation. Ecological Monographs,53, 383-404.
Wiggins G.B. (1977) Larvae of the North AmericanCaddisflv Genera (Trichoplera). Univers i ty ofToronto Press.
Wiggins G.B. (1984) Trichoptera. An Introduction tothe Aquatic Insects of North America. 2nd edn(Eds R. W. Merrit and K. W. Cummins) , pp.271-311. Kendal l -Hunt . Duhuque. Iowa.
Wiggins G.B. & Mackay R.J. (1978) Some relat ion-ships between systematic^ and t rophic ecology innearetic aquat ic insects. with specialreference to t r ichoptera. Ecology. 59, 1211-1220.
(Manuscript accepted 22 February /'AS'<Vy


















