
Intro popgen spring 2015
84
Hasan Alhaddad, Ph.D. Guest lecturer: Molecular Genetics (342) In Faculty of Allied Health Sciences (LT2-123) April 1 st 2015 (12:30 - 2:30) A Brief Introduction to Population Genetics
-
Upload
hhalhaddad -
Category
Science
-
view
97 -
download
3
Transcript of Intro popgen spring 2015
Hasan Alhaddad, Ph.D.
Guest lecturer: Molecular Genetics (342) In Faculty of Allied Health Sciences (LT2-123)
April 1st 2015 (12:30 - 2:30)
A Brief Introduction to Population Genetics

AIMS
• Review Mendelian genetics and its relation to population genetics.
• Review evolution and its relation to population genetics.
• Introduce the field of population genetics and its significance.
• Introduce the most basic model in population genetics (HW equation).
• Run some beanbag experiments.

Who am I?
• My name is Hasan Alhaddad.
• Ph.D. in Genetics from UC Davis.
• I studied insects and cats and I do population and evolutionary genetics for fun.
• I love science and I like to share this love with everybody especially you.
Who am I?• Check my website for more information or if you need to contact me.
hagenetics.org

Population genetics
Population Genetics
Evolutionary biologyMolecular Biology
Mendelianism
DNA
RNA
protein
Darwinism
dominant
recessive
co-dominant
mutation
mutation
species
neo-darwinism
genetic drift
mutation rate
substitution
deletion
natural selection
speciation
fitness
reproduction
survival
insertion transversion
transition
allele frequency
selection
gene
alleleMendeldiscrete
continuouslocus
loci
chromosome
different by state
identical by state
variation
polymorphism
homozygousheterozygous
segregation
monomorphic
morphology
haploid
diploid
biallelic
multi-allelic
Darwin
immigration
migration
isolationpopulation
phlyogenetics
phylogeny
taxonomy
genotype
phenotype
A
T C
G
U
replication
transcription
translation
population structure
genotype frequency
N
2N
pangenesis
hybridization
blending inheritance
visual traits
biochemical traits
Mendel’s first law
Mendel’s second law
Fst
Fis
Inbreeding
inbreeding depression
mRNA
tRNA
rRNA
gene expression
splicing
P
F1
F2 filial generation
parental generation adaptation
polyploidy
recombination rate
Fisher
Wright
sympatric speciation
allopatric speciation
genome
chromatin
histone
H3
H4
H2A
H2B
LD
Ne
generation
p
q
2pq
variancecontinous
epigenetics
motif
cis
trans
promoterexon
intron
transcriptome
proteome
double helix
bp
Kb
Mb
Gb
nucleotide
amino acid
alpha helix
Beta sheet

Population genetics is the field that results from combining Genetics and Evolution
We need to review both genetics and evolution to properly cover concepts
pertain to population genetics

Traditions of Genetics
Natural selection of small changes
Darwin Mendel
Transmission of large differences
Evolution and Population Genetics Analytical Genetics

Traditions of Genetics
Pearson
Darwin Mendel
Biometricians Mendelists
Yule Punnett Bateson
Hardy Weinberg
“one cannot help feeling that the speculations would have had more value had he kept his emotions under better control; the style and method of the religious revivalist are ill-suited to scientific controversy” (Edwards, 2008)
Yule to Bateson (1902)

Mendel and the general theory of inheritance and basic laws

The chief motives to understand heredity and the bases of it were:
1. Speciation 2. Hybridization 3. Similarities between
parents and offsprings
Scientific motives

What were the theories of inheritance at the time?

Theories of inheritance
Before Mendel the only proposed theory of inheritance was “blending inheritance”
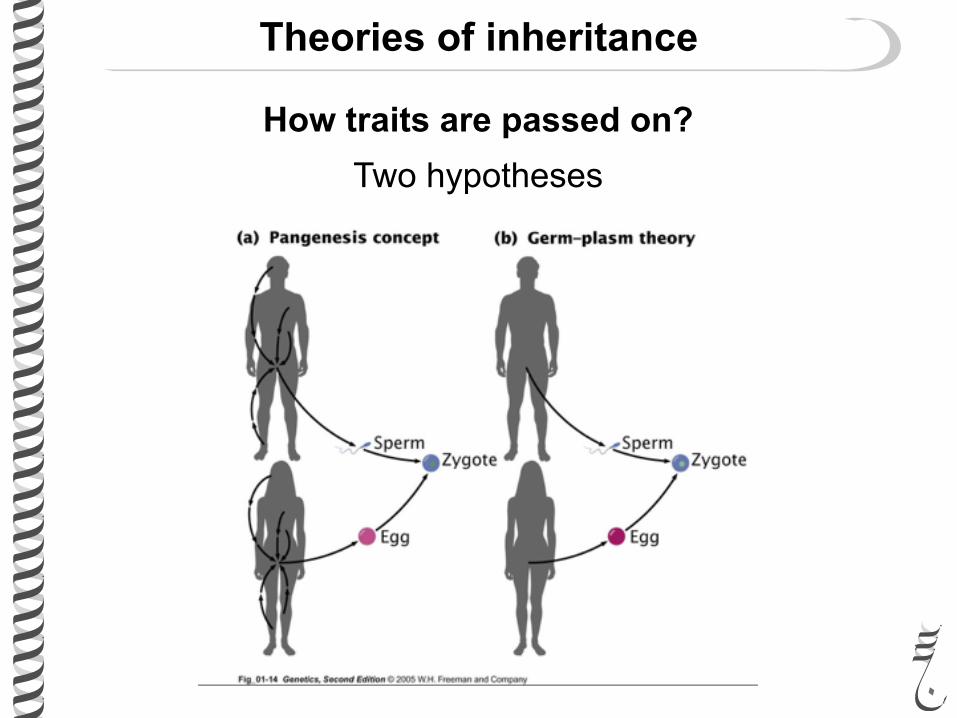
Theories of inheritance
How traits are passed on? Two hypotheses
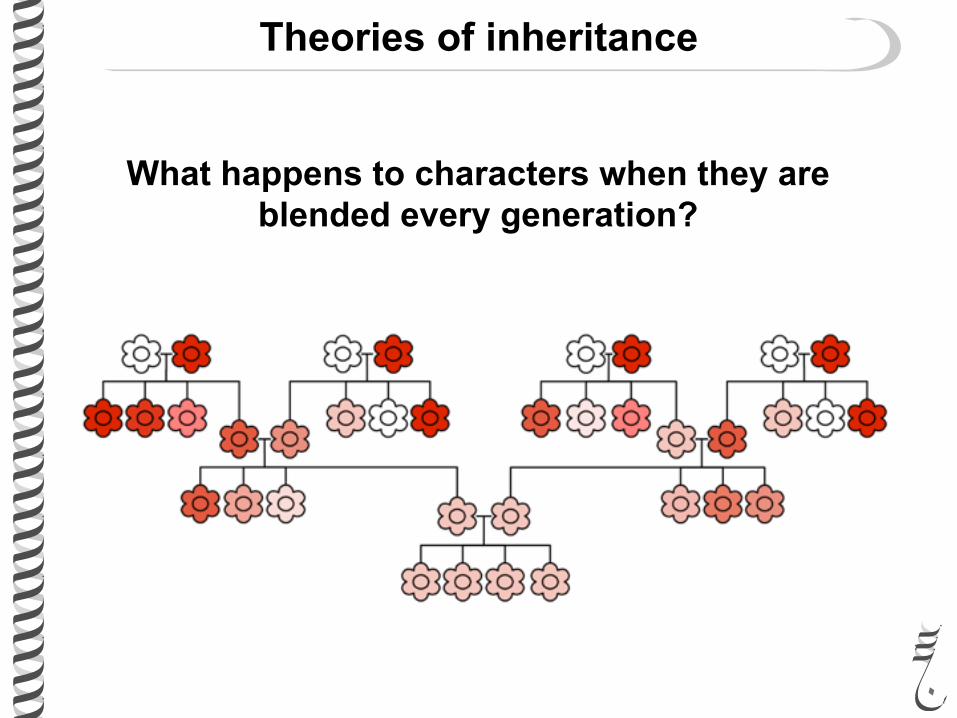
Theories of inheritance
What happens to characters when they are blended every generation?

Theories of inheritance
Darwin’s hypothesis of Pangenesis
“Gemmules” travel from every part of the body to the reproductive system to pass the traits to future generation.
Hypothesis NOT supported by scientific evidence.
O'Connor, C. & Miko, I. (2008) Developing the chromosome theory. Nature Education 1(1):44

Mendel and his peasGregor Mendel (Johann) studied heredity by the systematic breeding experiments of garden pea
(Pisum sativum)

Why Pea plants? Clear and distinct visual Traits/characters
Mendel’s Peas

What kind of traits/characters are these?
Why should we care?

Traits and characters
Traits can be continuous or discontinuous (also called discrete)
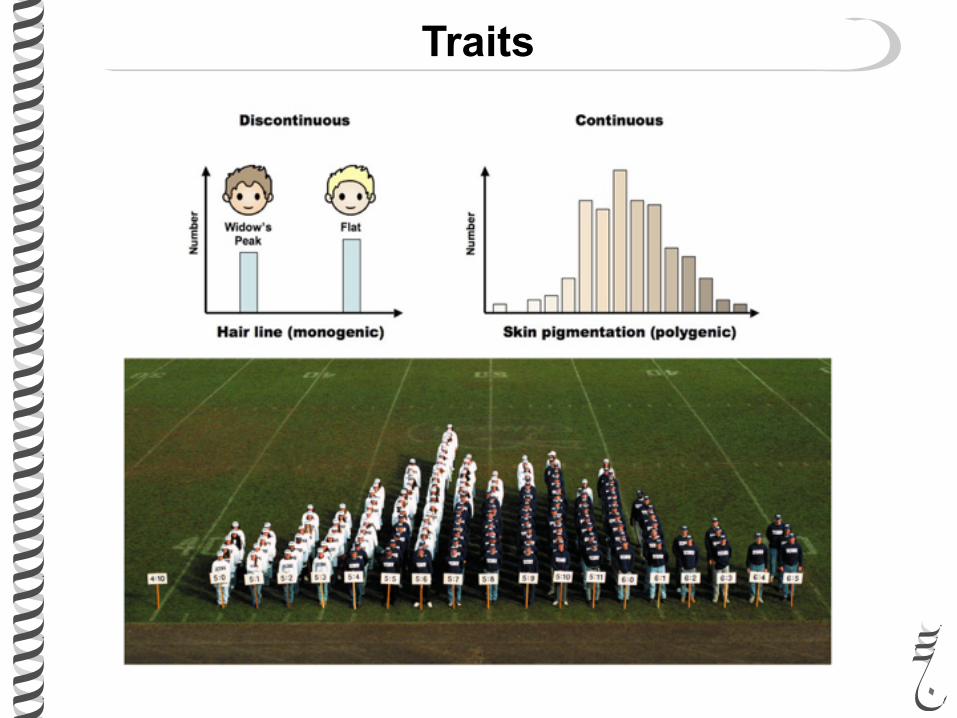
Traits

OK!?
What is the connection to Darwin and Mendel?

Mendel’s characters vs. Darwin’s
Mendel focused on variation of large effect while Darwin observed small variations that affect fitness

Lost?
Do not worry
We will get to Darwin in a bit

Mendel’s discrete characters
Mendel chose seven discrete characters that can be easily be visualized and identified.
Miko, I. (2008) Gregor Mendel and the principles of inheritance. Nature Education 1(1):134
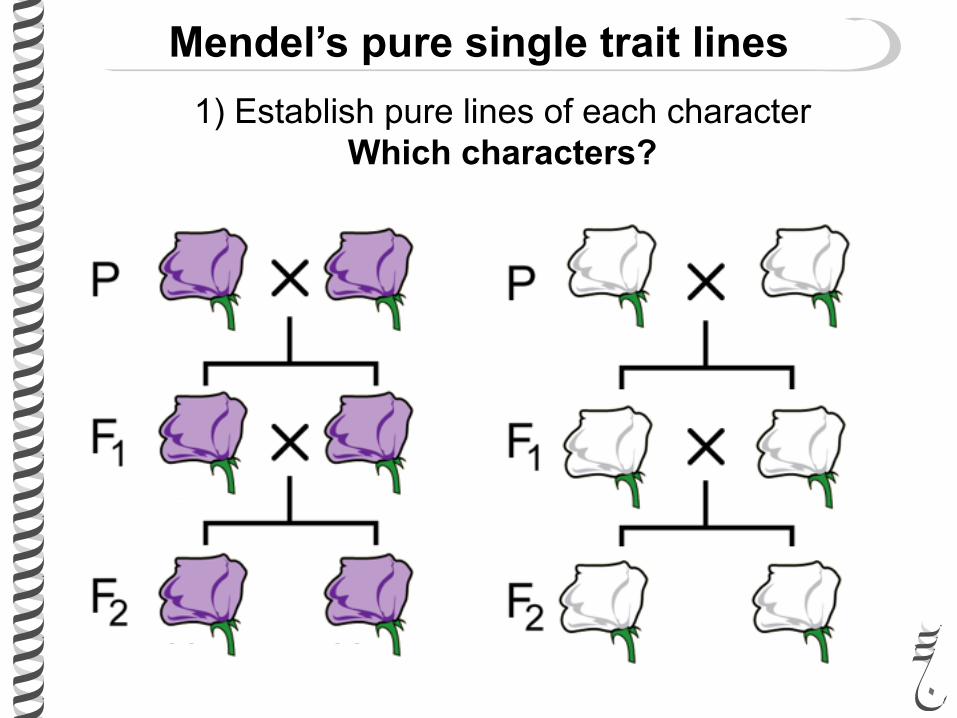
Mendel’s pure single trait lines1) Establish pure lines of each character
Which characters?

What are pure lines?
Homozygous?
Identical by state?
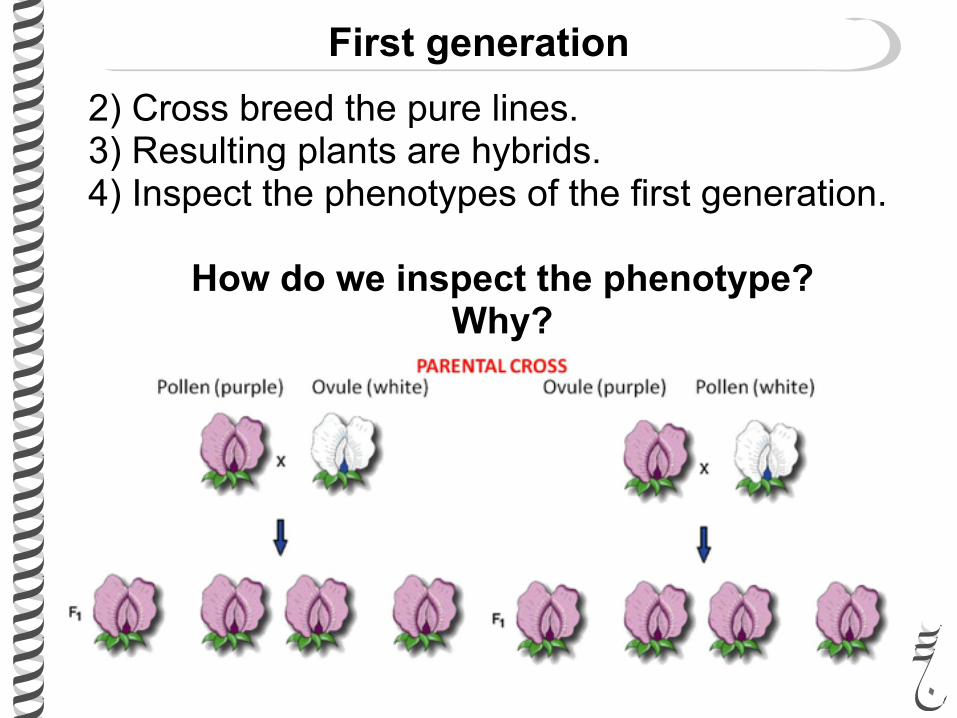
2) Cross breed the pure lines. 3) Resulting plants are hybrids. 4) Inspect the phenotypes of the first generation.
How do we inspect the phenotype? Why?
First generation

Observations and findings:
• All resulting plants exhibits the phenotype of one of the parents.
• One of the parental phenotypes disappears in the first hybrid generation.
First generation

5) Self cross the F1 individuals. 6) Inspect the phenotypes of the resulting F2 generation.
Second generation

How did Mendel inspect the phenotypes of the F2 generation?
Second generation

Mendel’s monohybrid results

Observations and findings:
• The selfing of the first generation results in the reappearance of one of the parents’ characteristics.
• A factor/particle is within the plant that results in the appearance of the plant.
• Both male and female contribute equally to the phenotype.
• The absence or appearance of a specific character depends on the combination of factors.
Mendel’s Monohybrid Experiment
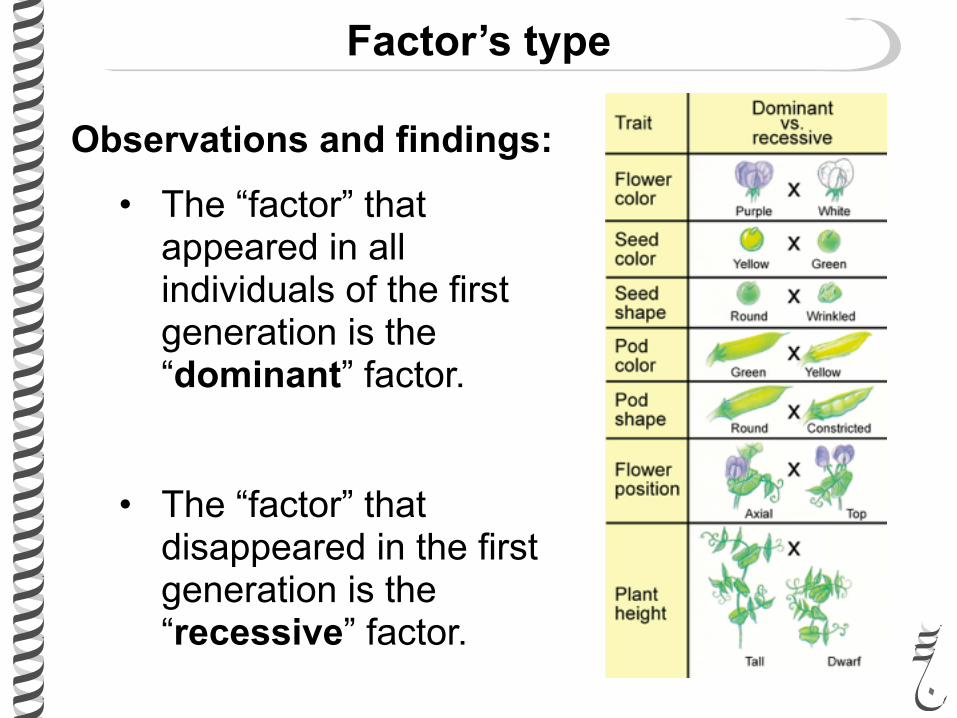
Observations and findings:
• The “factor” that appeared in all individuals of the first generation is the “dominant” factor.
• The “factor” that disappeared in the first generation is the “recessive” factor.
Factor’s type
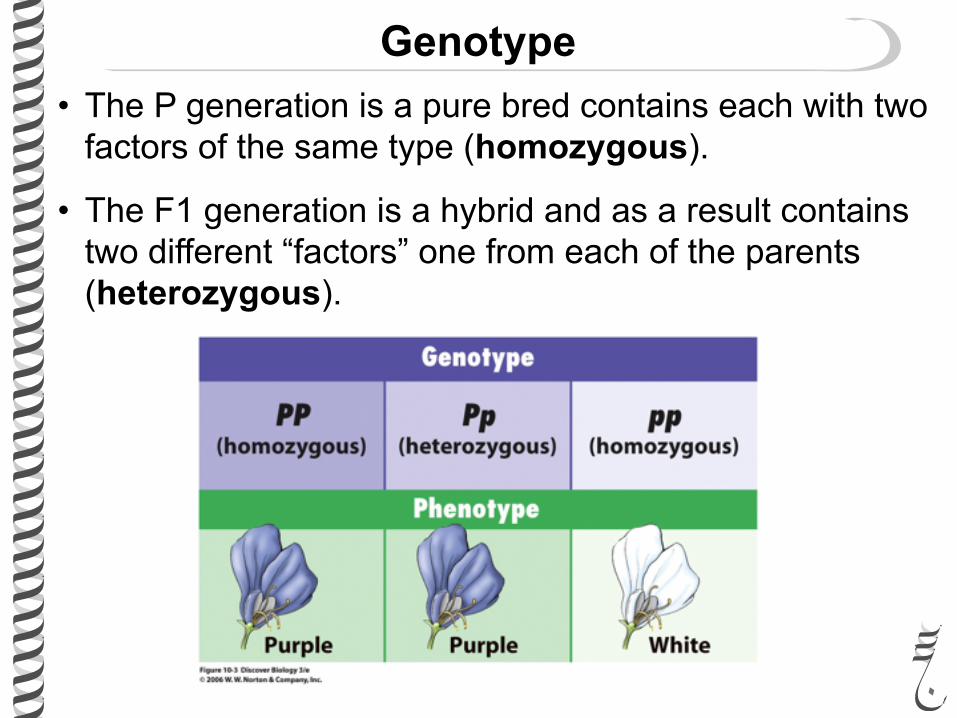
• The P generation is a pure bred contains each with two factors of the same type (homozygous).
• The F1 generation is a hybrid and as a result contains two different “factors” one from each of the parents (heterozygous).
Genotype

• “Factors” within a plant separate during the formation of gametes.
• “Factors” unite during fertilization randomly.
• The phenotype of resulting union is determined by the combination of factors.
Segregation of factors (alleles)
Mendel’s 1st law

• Yellow : Green 3:1
• Round : Wrinkled 3:1
What is the ratio of each phenotype independently?
Mendel’s 2nd law
• Each factor segregate independently.
Independent Assortment

• What is “dominant” and “recessive” a description of?
• What is a phenotype?
• What is a genotype?
• What is a homozygous?
• Identical by state?
• What is a heterozygous?
• Different by state?
• Did Mendel observe or infer genotype?
• Did Mendel observe or infer phenotype?
Review

Genes and Genotype

What is an allele?
Alleles are Mendel’s factors that he could not see but infer by crosses
Do not get it?
They are the (A) and (a) that are being passed into gametes and unite to give the genotype of
an individual.

What is an allele?

Alleles on chromosomes?
Where?
Same location?
Same chromosome?

Locus• A specific location in the genome is called
locus (plural loci). • Alleles at the same locus are inherited each
from one parent.

Alleles at a locus
• DNA at a specific locus may differ in one individual.
• How?
• What are the alleles in the figure?

Mendel’s work
Important contributions by Mendel to biology:
1.Genotypic notation.
2.Quantitative framework.

Now we need to review some evolution
What is evolution?
What is to evolve?

Evolution in general
Change through time
Non-biological Biological
Is evolution a fact or a theory?
Evolution (biological and non-biological) is a fact that is explained by theories
Not buying it?

Evolution in general
Can you think of a theory that explains the evolution of:
1) Food and cuisine (ex. Machboos)
2) Cars (ex. Ferrari)
3) Houses and cities (ex. skyscrapers)
4) Clothes and fashion (ex. Deshdasha)

So what about biological evolution?
Let’s start with the theory before Darwin

Lamarckian inheritance and evolution
Jean Baptiste Lamarck
Lamarck proposed the inheritance of acquired
characteristics

Testing Lamarck’s idea
August F. Weismann
• Weismann tested Lamarck’s idea using mice.
• Cutting mice tails and breeding them.
• 5 generations!
What happened?

Natural selection
Darwin and Wallace, independently, proposed a theory of biological evolution and called it
“Natural selection”
Charles Darwin Alfred Russel Wallace

Variation!
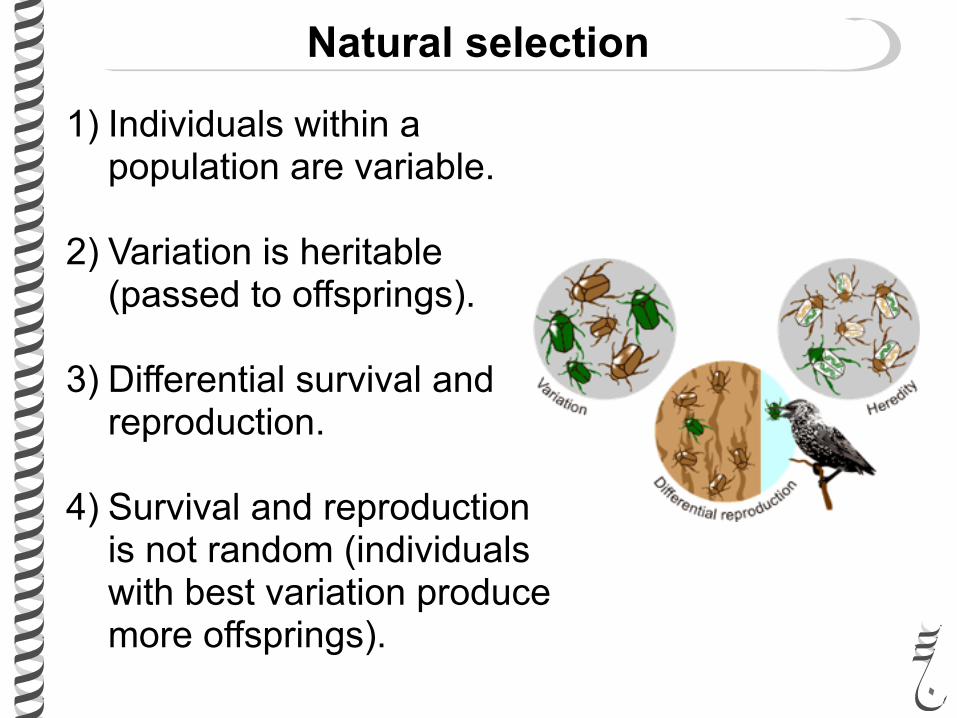
1) Individuals within a population are variable.
2) Variation is heritable (passed to offsprings).
3) Differential survival and reproduction.
4) Survival and reproduction is not random (individuals with best variation produce more offsprings).
Natural selection
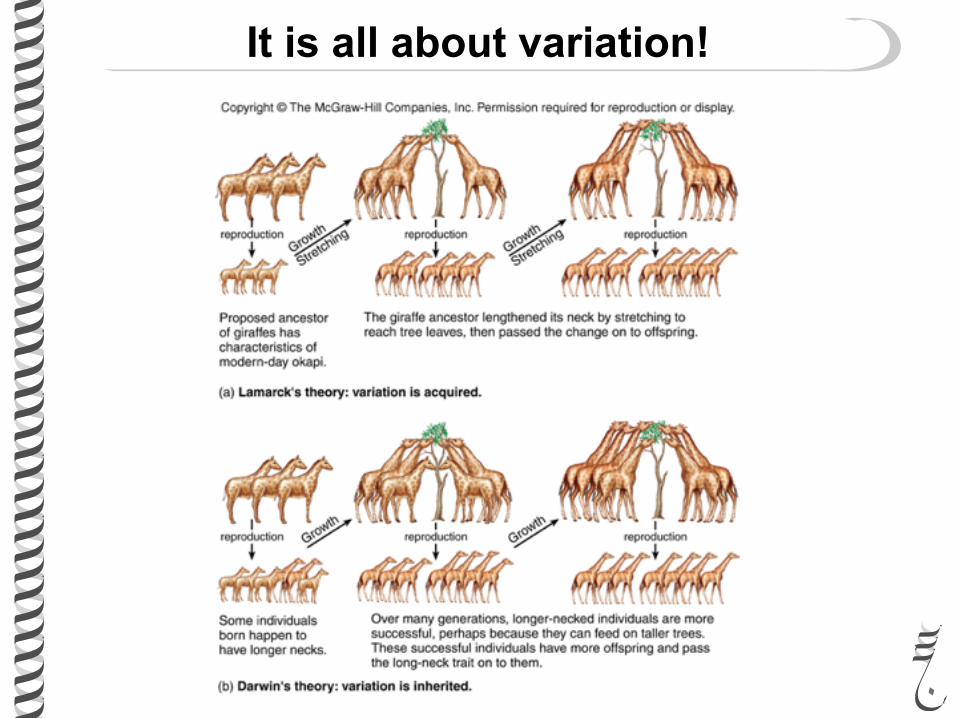
It is all about variation!

Did Darwin answer where variation come from?
Did he know about heredity?
Did he know about genes/factors?
Did evolutionary biology stop and end with Darwin’s theory?
Is natural selection the only force of evolution?
The field of evolutionary biology has itself evolved and evolutionary studies have gone way beyond
Darwin

Population genetics is the answer

Some significance
“Nothing in biology makes sense except in the light of evolution”
Theodosius Dobzhansky, 1973

Some significance
“Nothing in evolution makes sense except in the light of population
genetics”
Jeffrey Ross-Ibarra (2010 citing his mentor)

It is the study of the evolutionary historical record of a group of individuals documented
in the DNA of their descendants
What is population genetics?

Why population genetics?
1) Understand and refine theory
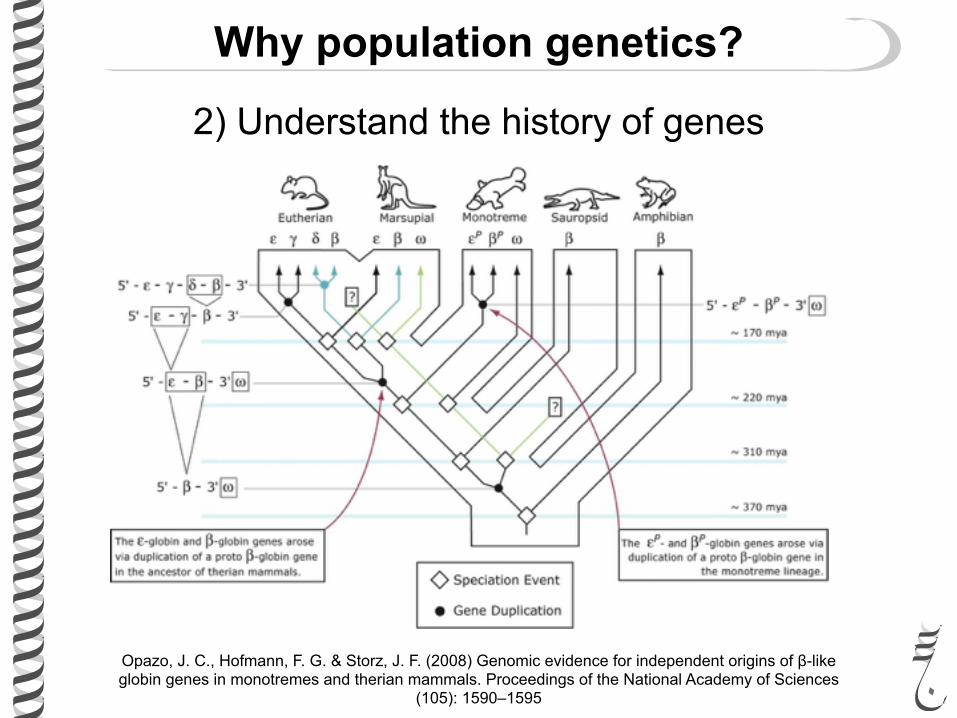
Why population genetics?
2) Understand the history of genes
Opazo, J. C., Hofmann, F. G. & Storz, J. F. (2008) Genomic evidence for independent origins of β-like globin genes in monotremes and therian mammals. Proceedings of the National Academy of Sciences
(105): 1590–1595
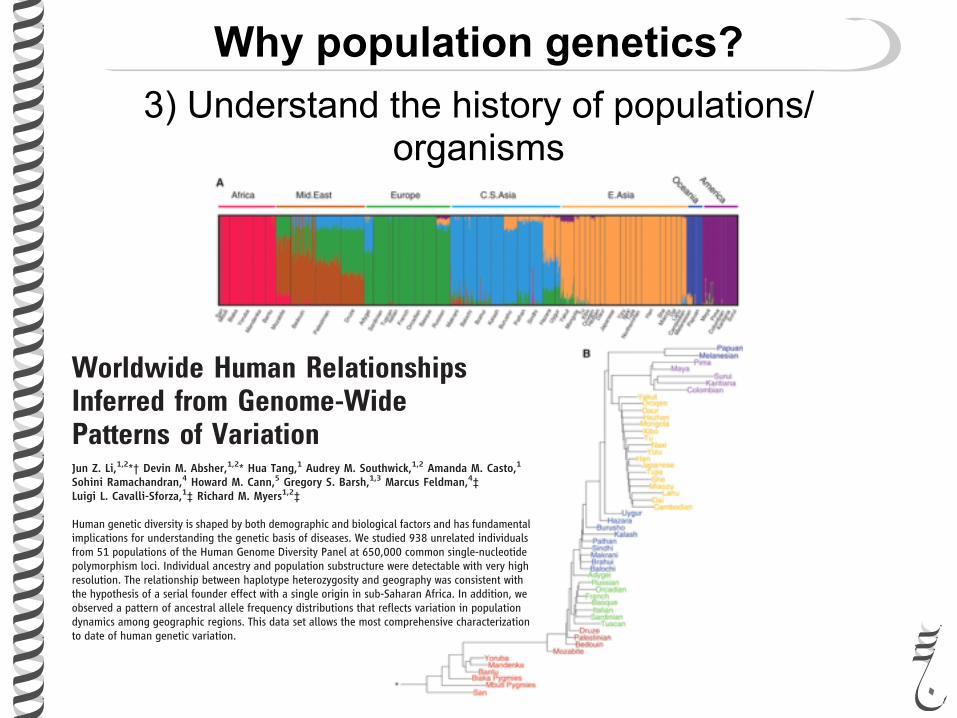
Why population genetics?3) Understand the history of populations/
organisms
21. J. Parsonnet, in Microbes and Malignancy, J. Parsonnet,Ed. (Oxford Univ. Press, New York, 1999), pp. 3–18.
22. Y. Xu et al., Genomics 81, 329 (2003).23. We thank the National Cancer Institute–supported
Cooperative Human Tissue Network for tissues usedin this study, M. Aquafondata for tissue staining,P. S. Schnable for sharing cDNA data sets used in DTSpilot testing, O. Gjoerup and R. D. Wood for helpfulcomments, and J. Zawinul for help with the manuscript.
Supported in part by funds from NIH R33CA120726and the Pennsylvania Department of Health. ThePennsylvania Department of Health specificallydisclaims responsibility for any analyses, interpretations,or conclusions.
Supporting Online Materialwww.sciencemag.org/cgi/content/full/1152586/DC1Materials and Methods
Figs. S1 to S3Tables S1 to S5References
5 November 2007; accepted 8 January 2008Published online 17 January 2008;10.1126/science.1152586Include this information when citing this paper.
Worldwide Human RelationshipsInferred from Genome-WidePatterns of VariationJun Z. Li,1,2*† Devin M. Absher,1,2* Hua Tang,1 Audrey M. Southwick,1,2 Amanda M. Casto,1Sohini Ramachandran,4 Howard M. Cann,5 Gregory S. Barsh,1,3 Marcus Feldman,4‡Luigi L. Cavalli-Sforza,1‡ Richard M. Myers1,2‡
Human genetic diversity is shaped by both demographic and biological factors and has fundamentalimplications for understanding the genetic basis of diseases. We studied 938 unrelated individualsfrom 51 populations of the Human Genome Diversity Panel at 650,000 common single-nucleotidepolymorphism loci. Individual ancestry and population substructure were detectable with very highresolution. The relationship between haplotype heterozygosity and geography was consistent withthe hypothesis of a serial founder effect with a single origin in sub-Saharan Africa. In addition, weobserved a pattern of ancestral allele frequency distributions that reflects variation in populationdynamics among geographic regions. This data set allows the most comprehensive characterizationto date of human genetic variation.
In the past 30 years, the ability to study DNAsequence variation has dramatically increasedour knowledge of the relationships among
and history of human populations. Analyses ofmitochondrial, Y chromosomal, and autosomalmarkers have revealed geographical structuringof human populations at the continental level (1–3)and suggest that a small group of individuals mi-grated out of eastern Africa and their descendantssubsequently expanded into most of today’s pop-ulations (3–6). Despite this progress, these studieswere limited to a small fraction of the genome, to
limited populations, or both, and yield an in-complete picture of the relative importance ofmutation, recombination, migration, demogra-phy, selection, and random drift (7–10). Tosubstantially increase the genomic and popula-tion coverage of past studies (e.g., the HapMapProject), we have examined more than 650,000single-nucleotide polymorphisms (SNPs) insamples from the Human Genome DiversityPanel (HGDP-CEPH), which represents 1064fully consenting individuals from 51 popula-tions from sub-Saharan Africa, North Africa,
Europe, the Middle East, South/Central Asia,East Asia, Oceania, and the Americas (11). Thisdata set is freely available (12) and allows adetailed characterization of worldwide geneticvariation.
We first studied genetic ancestry of eachindividual without using his/her populationidentity. This analysis considers each person’sgenome as having originated from K ancestralbut unobserved populations whose contributionsare described by K coefficients that sum to 1 foreach individual. To increase computational effi-ciency, we developed new software, frappe, thatimplements a maximum likelihood method (13)to analyze all 642,690 autosomal SNPs in 938unrelated and successfully genotyped HGDP-CEPH individuals (14). Figure 1A shows theresults for K = 7; those for K = 2 through 6 are infig. S1. At K = 5, the 938 individuals segregateinto five continental groups, similar to those re-
1Department of Genetics, Stanford University School ofMedicine, Stanford, CA 94305–5120, USA. 2StanfordHuman Genome Center, Stanford University School ofMedicine, Stanford, CA 94305–5120, USA. 3Department ofPediatrics, Stanford University School of Medicine, Stanford,CA 94305–5120, USA. 4Department of Biological Sciences,Stanford University, Stanford, CA 94305–5120, USA. 5Foun-dation Jean Dausset-Centre d’Etude du PolymorphismeHumain (CEPH), 75010 Paris, France.
*These authors contributed equally to this work.†Present address: Department of Human Genetics, Universityof Michigan, 5789A MS II, Ann Arbor, MI 48109–5618, USA.‡To whom correspondence should be addressed. E-mail:[email protected] (M.F.); [email protected](L.L.C.S.); [email protected] (R.M.M.)
Table 2. PCR for MCV DNA in comparison control tissues (n = 84). For detailed description of tissues and tissue sites, see table S2. MCV positivitiesmarked with plus and minus symbols together are as in Table 1. For the various body site tissues, there were 59 samples; for the skin and skin tumortissues, the sample size was 25 (table S2).
MCV positivityVarious body site tissues
Total MCV negative (%) 54/59 (92)Total MCV positive (%) 5/59 (8)
Appendix control 1 –/+Appendix control 2 –/+Gall bladder –/+Bowel –/+Hemorrhoid –/+
Skin and skin tumor tissuesTotal MCV negative (%) 21/25 (84)Total MCV positive (%) 4/25 (16)
Skin –/+KS skin tumor 1 –/+KS skin tumor 2 –/+KS skin tumor 3 –/+
22 FEBRUARY 2008 VOL 319 SCIENCE www.sciencemag.org1100
REPORTS

Why population genetics?
4) Understand the relationship between organisms
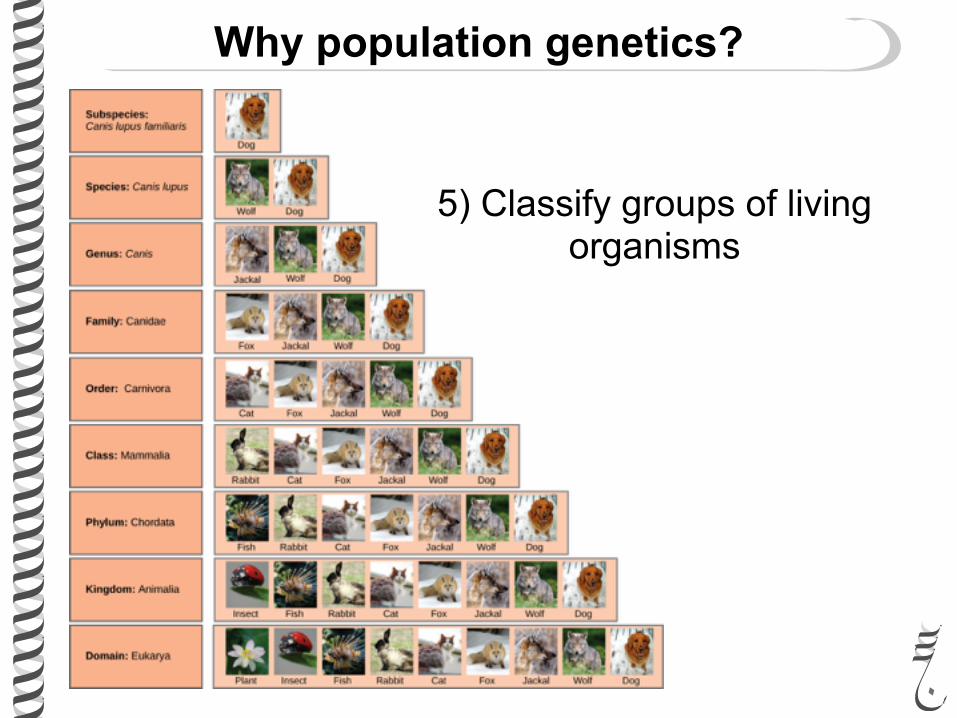
Why population genetics?
5) Classify groups of living organisms
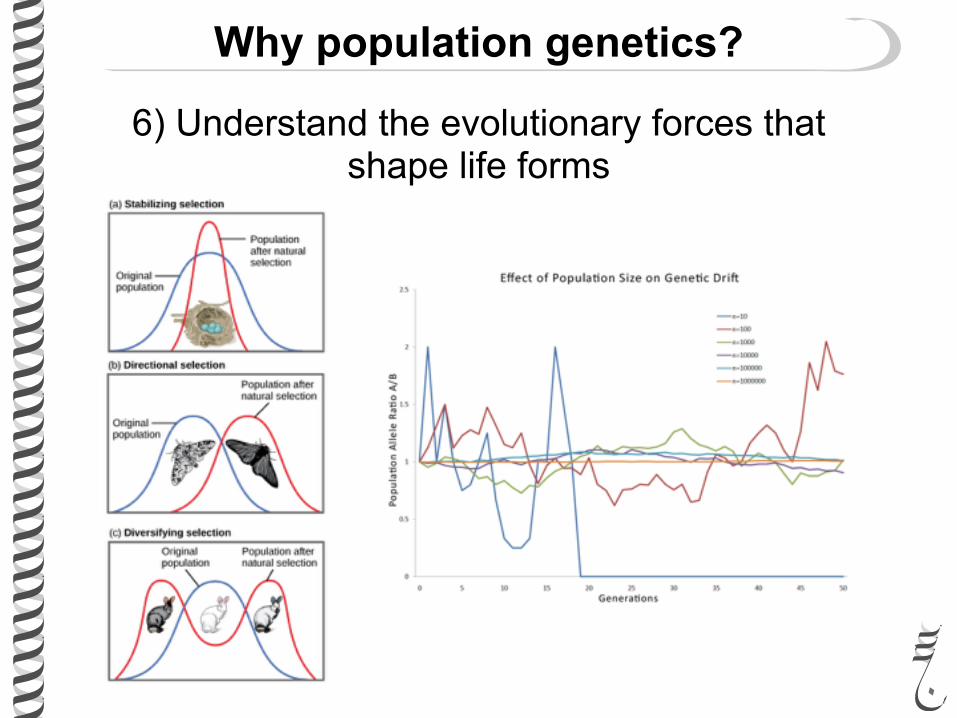
Why population genetics?
6) Understand the evolutionary forces that shape life forms
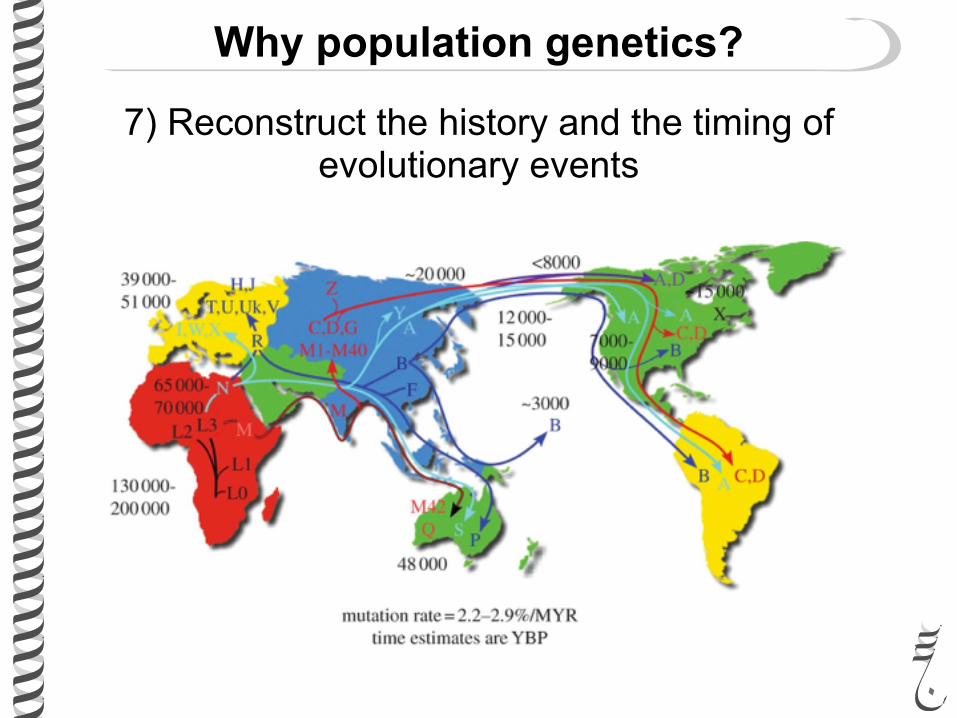
Why population genetics?
7) Reconstruct the history and the timing of evolutionary events

Why population genetics?
8) Find cool stuff!
Genome-wide Association studies
(GWAS)
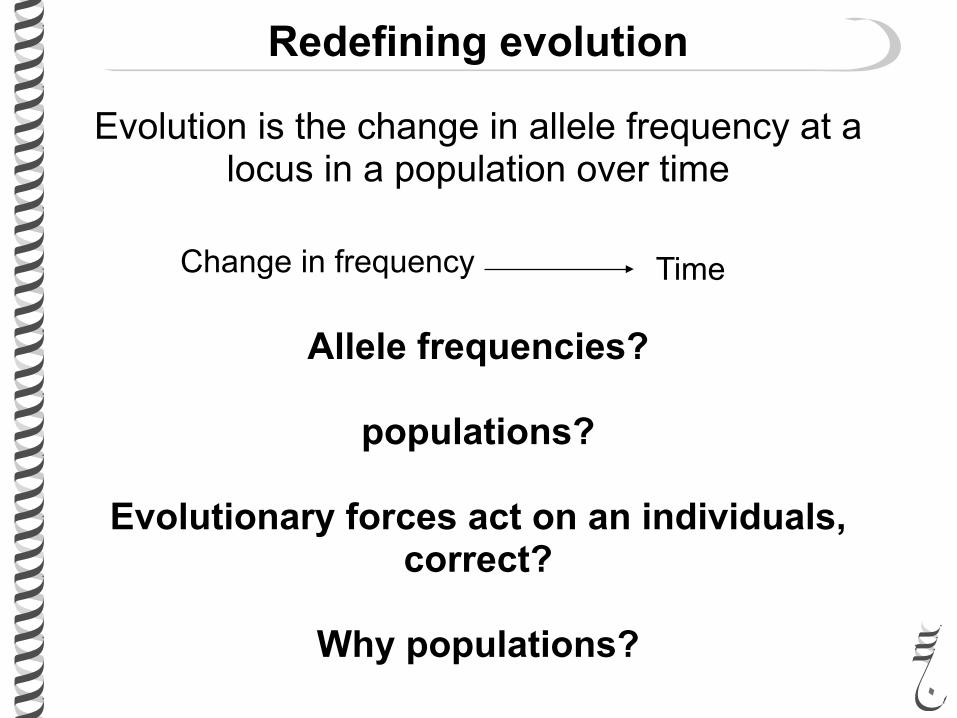
Redefining evolution
Evolution is the change in allele frequency at a locus in a population over time
Change in frequency Time
Allele frequencies?
populations?
Evolutionary forces act on an individuals, correct?
Why populations?

Redefining evolution
Evolutionary forces act on the individuals but the affects are seen in populations in the form
of changes in allele frequency
So what exactly do we do?
We study the effects of evolutionary forces on the frequency of a mutant allele
Ya Rabbi What are all these terms?
I am lost

Redefining evolution
One more thing about the changes
Molecular changes vs. Morphological changes
Small effects on fitness large effects on fitness
If they have small effects, then we will have to deal with chance
We need models to understand stochastic/random forces
vs.

(1) Edwards, 2008
Hardy-Weinberg Principle
“ The answer was in Mendel’s paper all the time”1
The first and most basic model in population genetics

Hardy-Weinberg Law
Weinberg Hardy
• Wilhelm Weinberg • (1862-1937) • German Physician
• Godfrey H. Hardy • (1877-1947 ) • English mathematician

(1) Edwards, 2008 (2) Jewett,1914
How did a mathematician, Hardy, get involved?
• February 28th 1908, Punnett gave a lecture on “Mendelism in relation to disease” 1.
•The lecture discusses brachydactylism.
• Brachydactylism means short-fingeredness 2.
• Shortness in fingers and toes relative to other body parts.
• The genetics disease is a dominant trait.

(1) Edwards, 2008
• Yule: if brachydactyl is a dominant trait (assuming random mating) ➔ 3:1 brachydactyl : normal. 1
• Yule misinterpreted Mendel’s theory. In order to get the 3:1 ratio, the gene frequency must be ½.
• Punnett: interpreted Yules remarks as “why the nation was not becoming ….. Brachydactylous”. 1
• Punnett was puzzled “why the dominant did not continually increase in frequency?” 1
• Hardy ….. Help!
How did a mathematician, Hardy, get involved?

(1) Hardy, 1908
Mendelian Proportions in a Mixed Population 1
• Assumptions: Aa is a Mendelian characters. The numbers of genotypes pure dominant (AA), heterozygotes (Aa), and pure recessives (aa) are 1:2:1 respectively.
• Conditions: “ … suppose that the numbers are fairly large, so that mating may be regarded as random, that the sexes are evenly distributed among the three varieties, and that all are equally fertile.”1
• Using “a little mathematics …” the allele frequencies in the next generation will be “ .. unchanged after the second generation.”
• “ I have … considered only the very simplest hypotheses possible.” 1

Assumptions• Single locus / Biallelic locus
• Diploid organism / Equal sexes.
• No natural selection: equal survival rates and reproductive success
• No mutation: no alleles created or converted
• No migration/gene flow: individuals do not move into or out of the population

Assumptions
• No genetic drift.
• Population is infinitely large: sampling errors and random effects insignificant.
• No population subdivision.
• Random mating (no inbreeding).
• Non overlapping generations.

The idea
Allele 2 (a)
Allele 1 (A)
Single locus and two alleles
q = frequency of Allele 2
q =
p = frequency of Allele 1
p =
What is the frequency of all alleles in a population?
p + q =
# of allele 1# allele 1+allele 2
# of allele 2# allele 1+allele 2
# of allele 1
# allele 1+allele 2
# of allele 2
# allele 1+allele 2+ =# allele 1+allele 2# allele 1+allele 2

Hardy-Weinberg Principle
(1) p + q = 1
A (p)
a (q)
a (q)
AA (p2) Aa (pq)
Aa (pq) aa (q2)
(2) p2+2pq+q2=1
Predictions(1) Allele frequency do not change over time
(2) Genotype frequencies can be calculated
A (p)
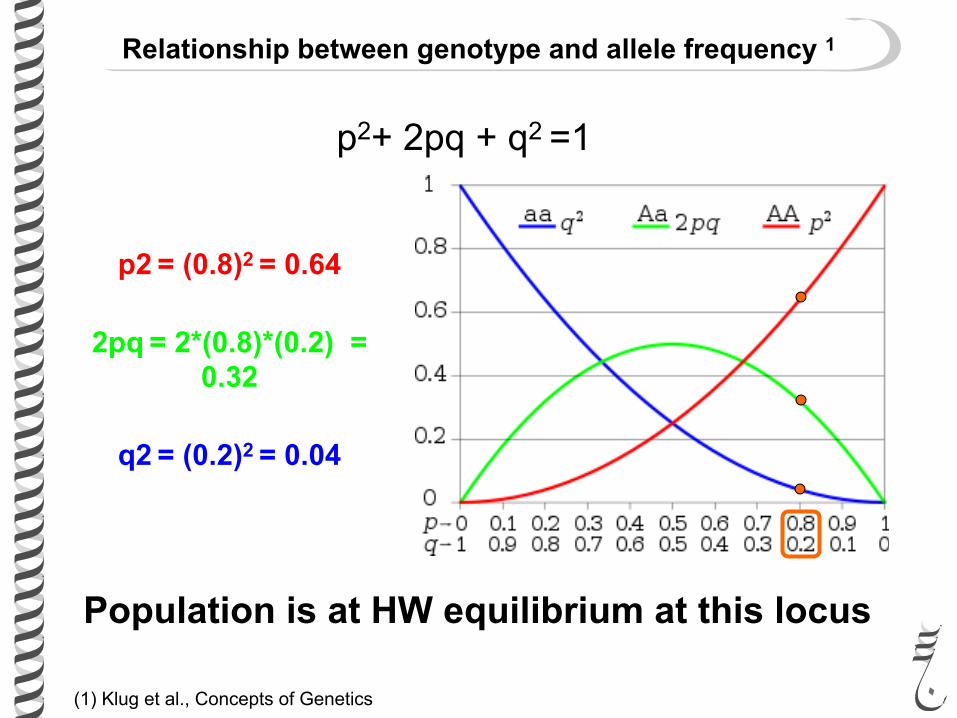
(1) Klug et al., Concepts of Genetics
Relationship between genotype and allele frequency 1
p2+ 2pq + q2 =1
p2 = (0.8)2 = 0.64
2pq = 2*(0.8)*(0.2) = 0.32
q2 = (0.2)2 = 0.04
Population is at HW equilibrium at this locus

What happens in frequencies ARE NOT in HW equilibrium?
We test which assumption of HW was violated and may, as a result, explain the
evolution of the population at this particular locus

Not so brief introduction after all
Sorry :-)

REPRINTS AND REFLECTIONS
A Defense of Beanbag Genetics*JBS Haldane
My friend Professor Ernst Mayr, of HarvardUniversity, in his recent book Animal Species andEvolution1, which I find admirable, though I disagreewith quite a lot of it, has the following sentences onpage 263.
The Mendelian was apt to compare the geneticcontents of a population to a bag full of coloredbeans. Mutation was the exchange of one kindof bean for another. This conceptualization hasbeen referred to as ‘‘beanbag genetics’’. Work inpopulation and developmental genetics has shown,however, that the thinking of beanbag geneticsis in many ways quite misleading. To considergenes as independent units is meaningless fromthe physiological as well as the evolutionaryviewpoint.
Any kind of thinking whatever is misleading outof its context. Thus ethical thinking involves theconcept of duty, or some equivalent, such as right-eousness or dharma. Without such a concept one islost in the present world, and, according to thereligions, in the next also. Joule, in his classicalpapers on the mechanical equivalent of heat, wroteof the duty of a steam engine. We now write ofits horsepower. It is of course possible thatethical conceptions will in future be applied toelectronic calculators, which may be given built-inconsciences!In another place2 Mayr made a more specific
challenge. He stated that Fisher, Wright, and I‘‘have worked out an impressive mathematicaltheory of genetical variaion and evolutionary change.But what, precisely, has been the contribution of thismathematical school to evolutionary theory, if I maybe permitted to ask such a provocative question?’’‘‘However,’’ he continued in the next paragraph,‘‘I should perhaps leave it to Fisher, Wright, andHaldane to point out what they consider their majorcontributions.’’ While Mayr may certainly ask thisquestion, I may not answer it at Cold Spring Harbor,
as I have been officially informed that I am ineligiblefor a visa for entering the United Statesy. Fisher isdead, but when alive preferred attack to defense.Wright is one of the gentlest men I have ever met,and if he defends himself, will not counterattack. Thisleaves me to hold the fort, and that by writing ratherthan speech.Now, in the first place I deny that the mathematical
theory of population genetics is at all impressive, atleast to a mathematician. On the contrary, Wright,Fisher, and I all made simplifying assumptions whichallowed us to pose problems soluble by the elemen-tary mathematics at our disposal, and even then didnot always fully solve the simple problems we setourselves. Our mathematics may impress zoologistsbut do not greatly impress mathematicians. Let megive a simple example. We want to know how thefrequency of a gene in a population changes undernatural selection. I made the following simplifyingassumptions3:
1. The population is infinite, so the frequency ineach generation is exactly that calculated, not justsomewhere near it.
2. Generations are separate. This is true for aminority only of animal and plant species. Thuseven in so-called annual plants a few seeds cansurvive for several years.
3. Mating is at random. In fact, it was not hard toallow for inbreeding once Wright had given aquantitative measure of it.
4. The gene is completely recessive as regardsfitness. Again it is not hard to allow forincomplete dominance. Only two alleles at onelocus are considered.
5. Mendelian segregation is perfect. There is nomutation, non-disjunction, gametic selection, orsimilar complications.
6. Selection acts so that the fraction of recessivesbreeding per dominant is constant from onegeneration to another. This fraction is the samein the two sexes.
*Haldane, J.B.S. A Defense of Beanbag Genetics. Perspectives inBiology and Medicine 7:3 (1964), 343-359. ! The JohnsHopkins University Press. Reproduced with permission of TheJohns Hopkins University Press.
yIn spite of this ineligibility I have, since writing this article,been granted an American visa, for which I must thank thefederal government. However, I am not permitted to lecture inNorth Carolina, and perhaps in other states, without answer-ing a question which I refuse to answer. Legislation to thiseffect does not, in my opinion, help American science.
Published by Oxford University Press on behalf of the International Epidemiological Association
! The Author 2008; all rights reserved.
International Journal of Epidemiology 2008;37:435–442
doi:10.1093/ije/dyn056
435
REPRINTS AND REFLECTIONS
A Defense of Beanbag Genetics*JBS Haldane
My friend Professor Ernst Mayr, of HarvardUniversity, in his recent book Animal Species andEvolution1, which I find admirable, though I disagreewith quite a lot of it, has the following sentences onpage 263.
The Mendelian was apt to compare the geneticcontents of a population to a bag full of coloredbeans. Mutation was the exchange of one kindof bean for another. This conceptualization hasbeen referred to as ‘‘beanbag genetics’’. Work inpopulation and developmental genetics has shown,however, that the thinking of beanbag geneticsis in many ways quite misleading. To considergenes as independent units is meaningless fromthe physiological as well as the evolutionaryviewpoint.
Any kind of thinking whatever is misleading outof its context. Thus ethical thinking involves theconcept of duty, or some equivalent, such as right-eousness or dharma. Without such a concept one islost in the present world, and, according to thereligions, in the next also. Joule, in his classicalpapers on the mechanical equivalent of heat, wroteof the duty of a steam engine. We now write ofits horsepower. It is of course possible thatethical conceptions will in future be applied toelectronic calculators, which may be given built-inconsciences!In another place2 Mayr made a more specific
challenge. He stated that Fisher, Wright, and I‘‘have worked out an impressive mathematicaltheory of genetical variaion and evolutionary change.But what, precisely, has been the contribution of thismathematical school to evolutionary theory, if I maybe permitted to ask such a provocative question?’’‘‘However,’’ he continued in the next paragraph,‘‘I should perhaps leave it to Fisher, Wright, andHaldane to point out what they consider their majorcontributions.’’ While Mayr may certainly ask thisquestion, I may not answer it at Cold Spring Harbor,
as I have been officially informed that I am ineligiblefor a visa for entering the United Statesy. Fisher isdead, but when alive preferred attack to defense.Wright is one of the gentlest men I have ever met,and if he defends himself, will not counterattack. Thisleaves me to hold the fort, and that by writing ratherthan speech.Now, in the first place I deny that the mathematical
theory of population genetics is at all impressive, atleast to a mathematician. On the contrary, Wright,Fisher, and I all made simplifying assumptions whichallowed us to pose problems soluble by the elemen-tary mathematics at our disposal, and even then didnot always fully solve the simple problems we setourselves. Our mathematics may impress zoologistsbut do not greatly impress mathematicians. Let megive a simple example. We want to know how thefrequency of a gene in a population changes undernatural selection. I made the following simplifyingassumptions3:
1. The population is infinite, so the frequency ineach generation is exactly that calculated, not justsomewhere near it.
2. Generations are separate. This is true for aminority only of animal and plant species. Thuseven in so-called annual plants a few seeds cansurvive for several years.
3. Mating is at random. In fact, it was not hard toallow for inbreeding once Wright had given aquantitative measure of it.
4. The gene is completely recessive as regardsfitness. Again it is not hard to allow forincomplete dominance. Only two alleles at onelocus are considered.
5. Mendelian segregation is perfect. There is nomutation, non-disjunction, gametic selection, orsimilar complications.
6. Selection acts so that the fraction of recessivesbreeding per dominant is constant from onegeneration to another. This fraction is the samein the two sexes.
*Haldane, J.B.S. A Defense of Beanbag Genetics. Perspectives inBiology and Medicine 7:3 (1964), 343-359. ! The JohnsHopkins University Press. Reproduced with permission of TheJohns Hopkins University Press.
yIn spite of this ineligibility I have, since writing this article,been granted an American visa, for which I must thank thefederal government. However, I am not permitted to lecture inNorth Carolina, and perhaps in other states, without answer-ing a question which I refuse to answer. Legislation to thiseffect does not, in my opinion, help American science.
Published by Oxford University Press on behalf of the International Epidemiological Association
! The Author 2008; all rights reserved.
International Journal of Epidemiology 2008;37:435–442
doi:10.1093/ije/dyn056
435
Let’s do beanbag genetics

Let’s do beanbag genetics


















