
intracelular - sisne.org | Laboratório de Sistemas...
23
5915756 – Introdução à Neurociência Computacional – Antonio Roque – Aula 3 1 O Código Neural • A figura abaixo (retirada do artigo de Contreras citado na aula 2) dá exemplos de registros obtidos quando se insere um eletrodo de forma intracelular diretamente no soma de um neurônio que está sendo estimulado por um degrau de corrente constante. Os diferentes degraus de corrente são mostrados nas figuras por linhas horizontais abaixo dos gráficos. • Observe os trens de potenciais de ação registrados. Os spikes são vistos aparecendo no topo das flutuações subliminares de potencial. Como os spikes são eventos de duração muito mais curta que as flutuações de potencial, para a resolução temporal das medidas mostradas eles são vistos apenas como linhas verticais (os detalhes ao lado das figuras mostram as formas dos potenciais de ação). • A figura abaixo ilustra o que seria obtido para a mesma situação mostrada no topo da figura acima caso um registro extracelular fosse feito nas vizinhanças do soma do neurônio. Observe que os potenciais de ação aparecem como traços com partes positivas e negativas mais ou menos iguais com amplitudes muito menores do que as dos traços obtidos nos registros intracelulares,
Transcript of intracelular - sisne.org | Laboratório de Sistemas...

5915756 – Introdução à Neurociência Computacional – Antonio Roque – Aula 3
1
O Código Neural
• A figura abaixo (retirada do artigo de Contreras citado na aula 2) dá exemplos de
registros obtidos quando se insere um eletrodo de forma intracelular diretamente
no soma de um neurônio que está sendo estimulado por um degrau de corrente
constante. Os diferentes degraus de corrente são mostrados nas figuras por linhas
horizontais abaixo dos gráficos.
• Observe os trens de potenciais de ação registrados. Os spikes são vistos aparecendo
no topo das flutuações subliminares de potencial. Como os spikes são eventos de
duração muito mais curta que as flutuações de potencial, para a resolução temporal
das medidas mostradas eles são vistos apenas como linhas verticais (os detalhes ao
lado das figuras mostram as formas dos potenciais de ação).
• A figura abaixo ilustra o que seria obtido para a mesma situação mostrada no topo
da figura acima caso um registro extracelular fosse feito nas vizinhanças do soma
do neurônio. Observe que os potenciais de ação aparecem como traços com partes
positivas e negativas mais ou menos iguais com amplitudes muito menores do que
as dos traços obtidos nos registros intracelulares,

5915756 – Introdução à Neurociência Computacional – Antonio Roque – Aula 3
2
• É muito difícil fazer um registro intracelular do tipo dos mostrados acima no axônio
de um neurônio. Caso fosse possível, teríamos algo como o mostrado na figura
abaixo (apenas para o caso do primeiro neurônio da figura acima). Observe que os
mesmos spikes observados no registro feito no soma são observados no registro
feito no axônio, porém as flutuações subliminares do potencial estão ausentes
devido à forte atenuação.
• Este exemplo ilustra um fenômeno importante: apenas os spikes, e não as flutuações
subliminares, se propagam por grandes distâncias ao longo dos axônios.
• Isto sugere que os neurônios usam os spikes para se comunicar. Em particular,
neurônios localizados nos órgãos sensoriais na periferia do corpo (retina, cóclea,
epitélio olfativo, pele a língua), que precisam mandar informação sobre os estímulos
do mundo externo que bombardeiam o corpo para os neurônios do cérebro
(interneurônios), usariam os spikes para transmitir essa informação.
• Um argumento comumente usado em favor desta idéia é o caso dos neurônios da
retina (veja a figura abaixo). Com exceção das células ganglionares, que são os
neurônios de saída da retina e que enviam a informação visual para o cérebro, todos
os demais neurônios da retina não geram potenciais de ação. Como eles não
precisam mandar informação a grandes distâncias, eles podem se comunicar usando
apenas os potenciais graduados gerados pelos estímulos que chegam a eles.

5915756 – Introdução à Neurociência Computacional – Antonio Roque – Aula 3
3
• Assumindo que os spikes emitidos pelos neurônios fornecem uma representação (ou
codificação) dos estímulos recebidos por eles, passa a ser importante encontrar
maneiras de analisar os trens de spikes de neurônios para se tentar entender como se
dá essa codificação e que relações existem entre as características dos estímulos e as
seqüências de spikes emitidas pelos neurônios.
• Também é importante encontrar maneiras de se conseguir reconstruir o estímulo a
partir da leitura do trem de potenciais de ação que um neurônio emite. Isto é
chamado de decodificação.
• A figura abaixo ilustra os conceitos de codificação e de decodificação.
• Analisar o trem de disparos de um neurônio é uma tarefa bastante difícil de ser feita
experimentalmente, pois as respostas neuronais costumam variar de experimento
para experimento mesmo quando o mesmo estímulo é apresentado repetidas vezes.
• Quando se analisa os trens de disparos de neurônios, trata-se os spikes como
eventos estereotipados idênticos e sem estrutura temporal (como os traços verticais
da figura acima). Desta forma, o que é importante para se entender a codificação
neural são os instantes de ocorrência dos disparos.
• A maneira como os spikes são identificados e isolados experimentalmente a partir
de um registro de potencial (intra- ou extracelular) não será tratada aqui. Basta saber
que o sinal é filtrado para eliminar componentes de baixa frequência em relação aos
potenciais de ação e que algum tipo de detector de padrões (por exemplo, um
detector de limiar ou de forma de onda) é aplicado ao sinal resultante.
• A figura a seguir ilustra o procedimento de identificação de spikes (spike sorting) a
partir de um registro extracelular (a figura foi retirada do artigo de revisão: Quiroga,
R. Q. & Panzeri, S., Extracting information from neuronal populations: information
theory and decoding approaches. Nature Reviews Neuroscience, 10:173-185, 2009).
5915756 – Introdução à Neurociência Computacional – Antonio Roque – Aula 3
4
Nature Reviews | Neuroscience
500
0
0 1 2 3 4 5–500
Time (s)
Time (s)
500
0
0 1 2 3 4 5–500
Time (s)
Local field potential
0 1 2 3 4 5–50
50100
0
Spikes
Low-pass filter
High-pass filter
–500
50100150200
0 20 40 60
–500
50100150200
0 20 40 60
121 spikes
–500
50100150200
0 20 40 60
–500
50100150200
0 20 40 60
42 spikes16,273 spikes 269 spikes
Volta
ge (µ
V)
Volta
ge (µ
V)
Data point
Information theoryA mathematical theory that deals with measures of information and their application to the study of communication systems. In neuroscience it is used to establish the amount of information about a stimulus or behaviour that is contained in the neural responses.
Local field potential(LFP). A neurophysiological signal that is obtained by low-pass filtering extracellular recordings. It represents the mean field potential generated by the slow components of synaptic and neural events in the vicinity of the recording electrode.
clearly indicate whether an apple or an orange is being seen, but this uncertainty might be resolved when the activity of other neurons is considered. In fact, the popu-lation analysis based on decoding or information theory can reveal how the other neurons resolve this ambigu-ity — for example, by coordinating their firing to tag particularly salient events22,23 or by having each neuron represent a particular stimulus feature24–26. Second, postsynaptic neuronal systems must usually interpret neuronal responses obtained in only one trial. Both the information-theoretic and the decoding approaches quantify stimulus knowledge obtained with the observa-tion of single-trial population responses, thus providing a framework that is compatible with the strict timescales
of online brain processing. Third, the stimulus features encoded by the spike trains can be discovered by assess-ing whether the population response can discriminate different stimuli containing a particular feature. Fourth, it is possible to systematically evaluate how different features of the spike trains affect the performance of a decoding algorithm or the amount of extracted infor-mation. Fifth, the information given by different meas-ures of neuronal activity, such as spike trains and local field potentials (LFPs), can be analysed and combined. Although these two signals have very different charac-teristics and signal-to-noise ratios, information theory and decoding algorithms allow a direct comparison between LFPs and spikes because they project the two
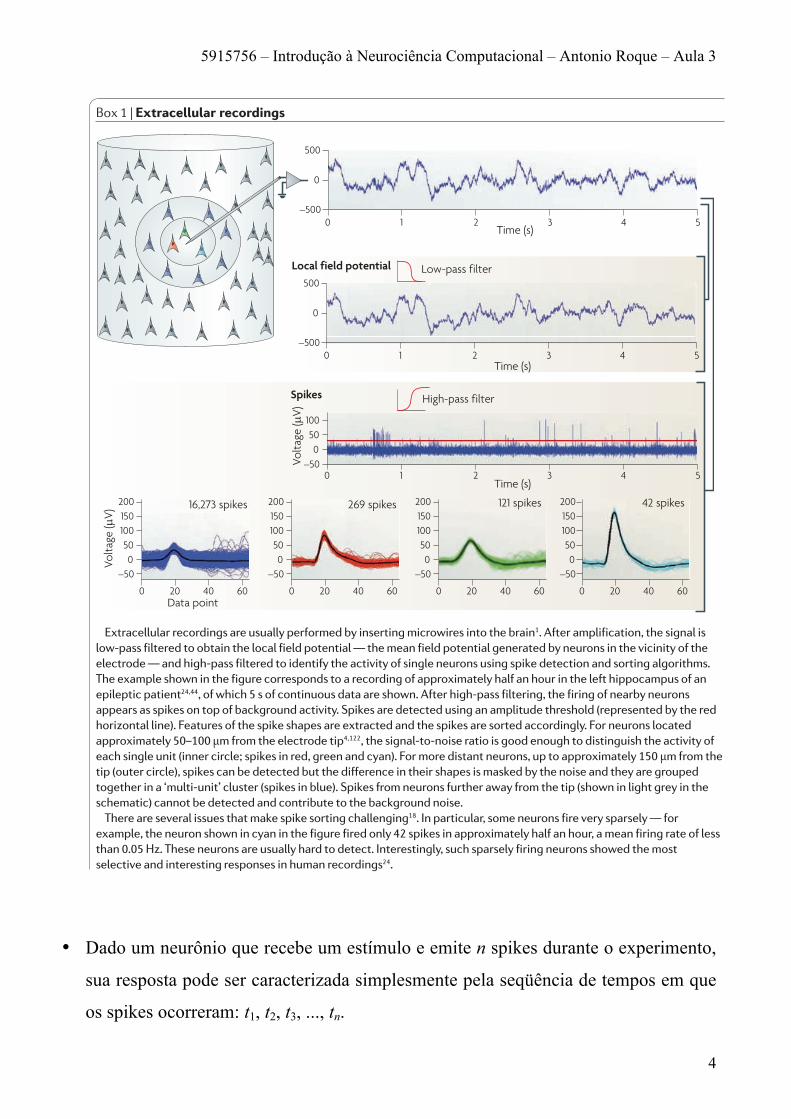
Box 1 | Extracellular recordings
Extracellular recordings are usually performed by inserting microwires into the brain1. After amplification, the signal is low-pass filtered to obtain the local field potential — the mean field potential generated by neurons in the vicinity of the electrode — and high-pass filtered to identify the activity of single neurons using spike detection and sorting algorithms. The example shown in the figure corresponds to a recording of approximately half an hour in the left hippocampus of an epileptic patient24,44, of which 5 s of continuous data are shown. After high-pass filtering, the firing of nearby neurons appears as spikes on top of background activity. Spikes are detected using an amplitude threshold (represented by the red horizontal line). Features of the spike shapes are extracted and the spikes are sorted accordingly. For neurons located approximately 50–100 μm from the electrode tip4,122, the signal-to-noise ratio is good enough to distinguish the activity of each single unit (inner circle; spikes in red, green and cyan). For more distant neurons, up to approximately 150 μm from the tip (outer circle), spikes can be detected but the difference in their shapes is masked by the noise and they are grouped together in a ‘multi-unit’ cluster (spikes in blue). Spikes from neurons further away from the tip (shown in light grey in the schematic) cannot be detected and contribute to the background noise.
There are several issues that make spike sorting challenging18. In particular, some neurons fire very sparsely — for example, the neuron shown in cyan in the figure fired only 42 spikes in approximately half an hour, a mean firing rate of less than 0.05 Hz. These neurons are usually hard to detect. Interestingly, such sparsely firing neurons showed the most selective and interesting responses in human recordings24.
REVIEWS
174 | MARCH 2009 | VOLUME 10 www.nature.com/reviews/neuro
• Dado um neurônio que recebe um estímulo e emite n spikes durante o experimento,
sua resposta pode ser caracterizada simplesmente pela seqüência de tempos em que
os spikes ocorreram: t1, t2, t3, ..., tn.
5915756 – Introdução à Neurociência Computacional – Antonio Roque – Aula 3
5
• Supondo que o experimento durante o qual os spikes foram registrados começou no
instante t = 0 e terminou em t = T, os tempos ti devem satisfazer 0 ≤ ti ≤ T para
qualquer i.
• Formalmente, pode-se representar a seqüência de spikes de um neurônio como:
( ),)(1∑=
−=n
iitttS δ
onde δ(x) é a função delta de Dirac, caracterizada pelas seguintes propriedades:
∫+∞
∞−
= ;1)( dttδ
).(´´)(´)( tfdttttf =−∫+∞
∞−
δ
Note que a função delta tem dimensão de t-1.
• Dado que existe variabilidade nas seqüências de spikes emitidas por um neurônio
quando se repete um mesmo estímulo várias vezes, costuma-se caracterizar as
respostas neuronais do ponto de vista estatístico. Uma das maneiras de se fazer isso
é através da determinação de taxas de disparos. Há várias definições possíveis de
taxa de disparo sendo usadas na literatura. É importante que se saiba claramente
qual delas está sendo usada em um dado momento e para o que ela serve.
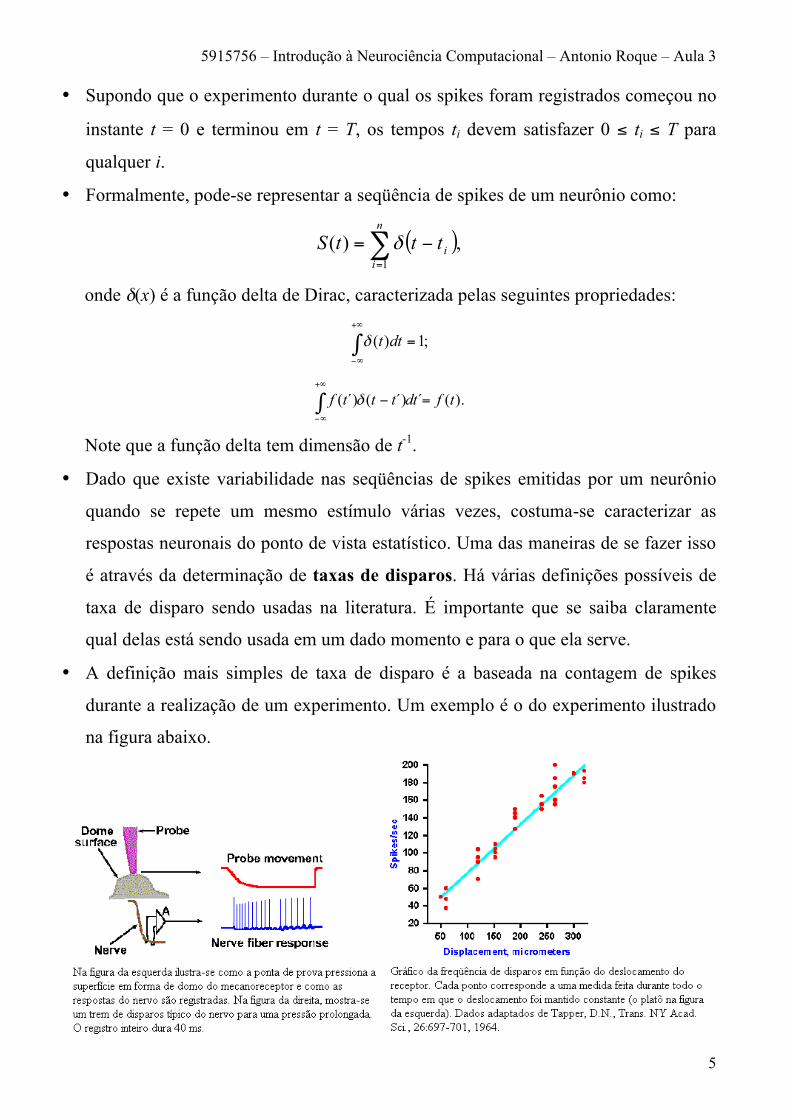
• A definição mais simples de taxa de disparo é a baseada na contagem de spikes
durante a realização de um experimento. Um exemplo é o do experimento ilustrado
na figura abaixo.

5915756 – Introdução à Neurociência Computacional – Antonio Roque – Aula 3
6
• A figura ilustra a codificação em termos de taxa feita por um disco de Merkel, um
mecanoreceptor localizado abaixo da parte peluda da pele de gatos e do abdômen de
humanos. Observe que, quanto maior o valor do estímulo (o deslocamento do disco
mantido constante por um certo tempo), maior a taxa de disparos do receptor. Para a
faixa de valores de estímulos mostrada na figura, a relação entre taxa de disparos e
valor do estímulo é linear. A equação que aproxima essa relação é F = 0,55D + 18,
onde F é a taxa de disparos em spikes/seg e D é o deslocamento em micrômetros. A
partir desta equação, podemos calcular o deslocamento que o receptor sofre a partir
da medida da taxa de disparos do neurônio receptor. Isto sugere que o sistema
nervoso possa fazer o mesmo.
• A taxa de disparos definida pelo experimento ilustrado acima é a seguinte: Durante
um certo tempo T conta-se o número n de spikes que o neurônio emite em resposta a
um estímulo. Tomando-se a razão entre os dois tem-se a chamada taxa de disparos
baseada na contagem (em inglês, spike-count rate), que será denominada por r:
.)(1
0∫==T
dSTT
nr ττ
• Exercício: mostre a igualdade acima.
• Para que a taxa de disparos baseada na contagem tenha sentido, é necessário que o
intervalo de tempo T seja suficientemente grande para que vários spikes ocorram
dentro dele. A duração de T depende do tipo de neurônio e do estímulo. Valores
típicos são de algumas centenas de milisegundos (p. ex. 100 ou 500 ms).
• Medidas de taxa de disparos baseadas na contagem são muito comuns,
principalmente quando se estuda neurônios sensoriais e motores. Elas permitem que
se obtenha experimentalmente a função de transferência de um neurônio, conforme
definida na aula passada.
• Outro exemplo de uso da contagem de spikes como medida da resposta de um
neurônio ocorre na determinação da chamada curva de sintonia de um neurônio. Por
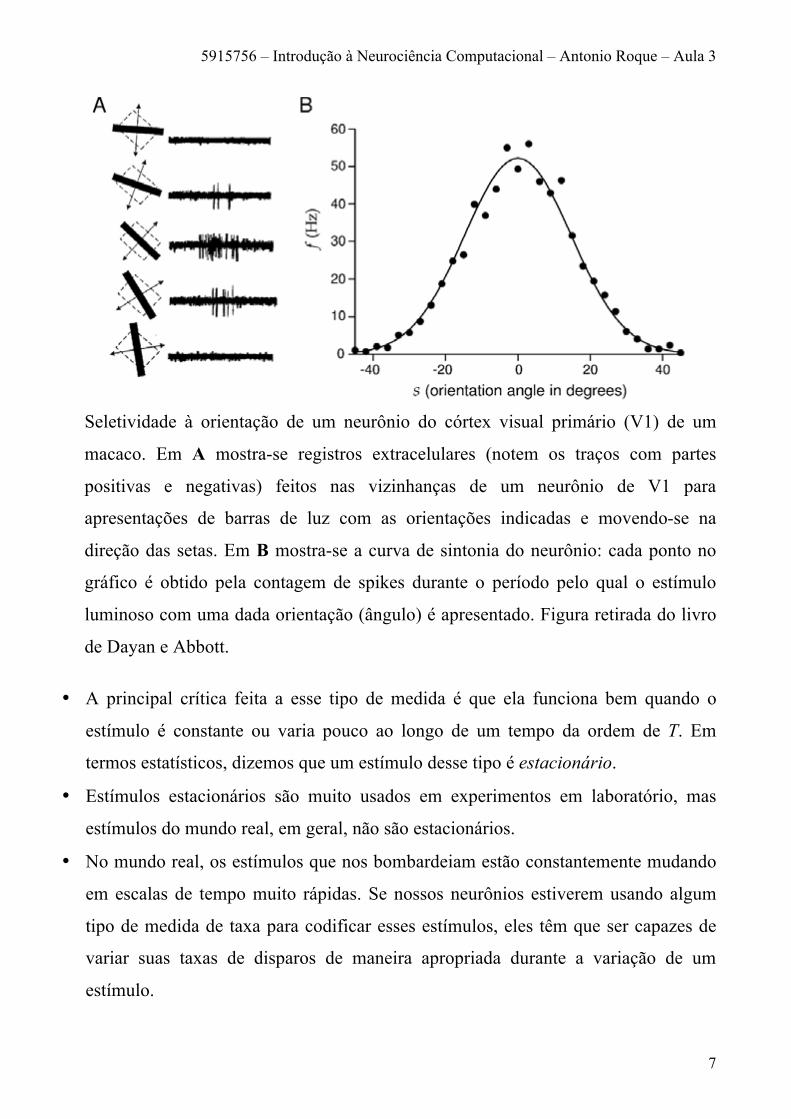
exemplo, a figura abaixo mostra a curva de sintonia de um neurônio do córtex visual
primário.
5915756 – Introdução à Neurociência Computacional – Antonio Roque – Aula 3
7
Seletividade à orientação de um neurônio do córtex visual primário (V1) de um
macaco. Em A mostra-se registros extracelulares (notem os traços com partes
positivas e negativas) feitos nas vizinhanças de um neurônio de V1 para
apresentações de barras de luz com as orientações indicadas e movendo-se na
direção das setas. Em B mostra-se a curva de sintonia do neurônio: cada ponto no
gráfico é obtido pela contagem de spikes durante o período pelo qual o estímulo
luminoso com uma dada orientação (ângulo) é apresentado. Figura retirada do livro
de Dayan e Abbott.
• A principal crítica feita a esse tipo de medida é que ela funciona bem quando o
estímulo é constante ou varia pouco ao longo de um tempo da ordem de T. Em
termos estatísticos, dizemos que um estímulo desse tipo é estacionário.
• Estímulos estacionários são muito usados em experimentos em laboratório, mas
estímulos do mundo real, em geral, não são estacionários.
• No mundo real, os estímulos que nos bombardeiam estão constantemente mudando
em escalas de tempo muito rápidas. Se nossos neurônios estiverem usando algum
tipo de medida de taxa para codificar esses estímulos, eles têm que ser capazes de
variar suas taxas de disparos de maneira apropriada durante a variação de um
estímulo.
5915756 – Introdução à Neurociência Computacional – Antonio Roque – Aula 3
8
• Uma maneira de se caracterizar a variação dos disparos de um neurônio ao longo de
um experimento é pela definição de uma taxa de disparos dependente do tempo: a
cada instante de tempo t durante um experimento, conta-se o número de spikes
emitidos entre t e t + Δt, para Δt sendo um intervalo pequeno, e divide-se por Δt:
( ) ,;)(1 ttttntr
ΔΔ+
=
onde o subíndice 1 foi usado para indicar que há outras maneiras de se definir uma
taxa de disparos dependente do tempo.
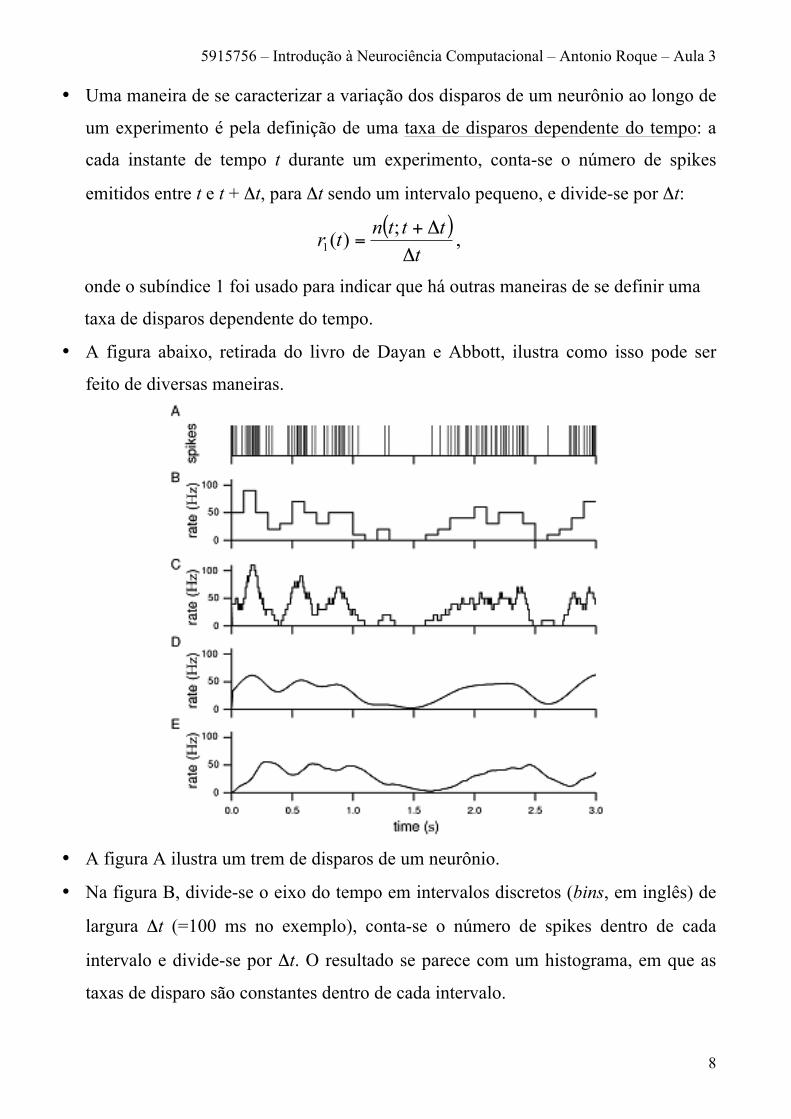
• A figura abaixo, retirada do livro de Dayan e Abbott, ilustra como isso pode ser
feito de diversas maneiras.
• A figura A ilustra um trem de disparos de um neurônio.
• Na figura B, divide-se o eixo do tempo em intervalos discretos (bins, em inglês) de
largura Δt (=100 ms no exemplo), conta-se o número de spikes dentro de cada
intervalo e divide-se por Δt. O resultado se parece com um histograma, em que as
taxas de disparo são constantes dentro de cada intervalo.
5915756 – Introdução à Neurociência Computacional – Antonio Roque – Aula 3
9
• Uma maneira (não mostrada na figura) de se evitar que a taxa de disparos tenha essa
aparência é trabalhar com um número constante de disparos n e um intervalo Δt de
tamanho variável. Escolhem-se intervalos de largura variável de maneira que dentro
de cada intervalo haja sempre o mesmo número n de disparos. Neste caso, a taxa
de disparos é aproximada por n dividido pelo tamanho variável do intervalo.
• A definição da taxa de disparos segundo a figura B depende não apenas do tamanho
do intervalo, mas da sua localização. Para evitar a arbitrariedade na localização dos
intervalos, pode-se definir um único intervalo ou janela de largura Δt e deslizá-la ao
longo do eixo do tempo, contando o número de disparos dentro da janela em cada
instante de tempo.
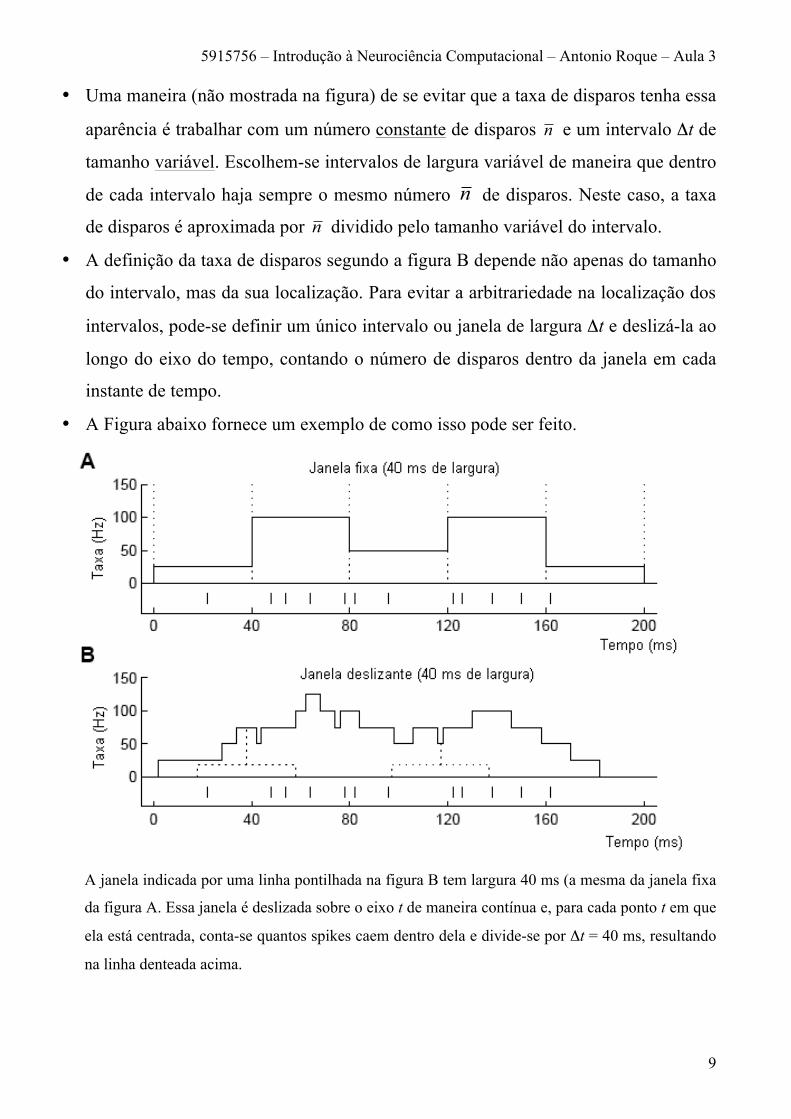
• A Figura abaixo fornece um exemplo de como isso pode ser feito.
A janela indicada por uma linha pontilhada na figura B tem largura 40 ms (a mesma da janela fixa
da figura A. Essa janela é deslizada sobre o eixo t de maneira contínua e, para cada ponto t em que
ela está centrada, conta-se quantos spikes caem dentro dela e divide-se por Δt = 40 ms, resultando
na linha denteada acima.

5915756 – Introdução à Neurociência Computacional – Antonio Roque – Aula 3
10
• A taxa de disparos definida desta forma pode ser expressa matematicamente como a
soma de uma função janela ao longo dos tempos dos spikes, ti, i = 1, 2, ..., n:
,)()()(2 ∫+∞
∞−
−= τττ dtSwtr
onde a função janela é
⎩⎨⎧ Δ≤≤Δ−Δ
=contrário. caso0
22 se1)(
tttttw
• A figura C ilustra o resultado de se calcular a taxa de disparos dependente do tempo
segundo esta definição para uma janela de largura Δt = 100 ms.
• Observe que a medida de taxa parece ter uma resolução temporal melhor. Note,
porém, que por esta definição um mesmo spike pode cair dentro de mais de uma
janela e isto provoca uma correlação (dependente da largura da janela) entre taxas
medidas em tempos separados por menos que uma largura de janela.
• As descontinuidades observadas na figura C são causadas pelo fato de a função
janela usada não ser suave (faça um esboço dessa função janela para se convencer
disso).
• A operação matemática usada acima para se definir r2(t) é chamada de convolução
de S com w, e, nessa terminologia, a função w(t) é chamada de núcleo da
convolução. Também se pode chamar a integral acima de filtro linear e a função
w(t) de núcleo do filtro.
• Pode-se mostrar que a definição de r2(t) é coerente com a definição de taxa baseada
na contagem de spikes:
( )
( )
( )∑
∑ ∫
∫ ∑
∫
=
=
∞+
∞−
∞+
∞− =
+∞
∞−
−++−+−=−=
−−=
−−=
−=
n
ini
n
ii
n
ii
ttwttwttwttwtr
dttwtr
dttwtr
dtSwtr
1212
12
12
2
).()()()(
)()(
)()(
)()()(
…
ττδτ
ττδτ
τττ
5915756 – Introdução à Neurociência Computacional – Antonio Roque – Aula 3
11
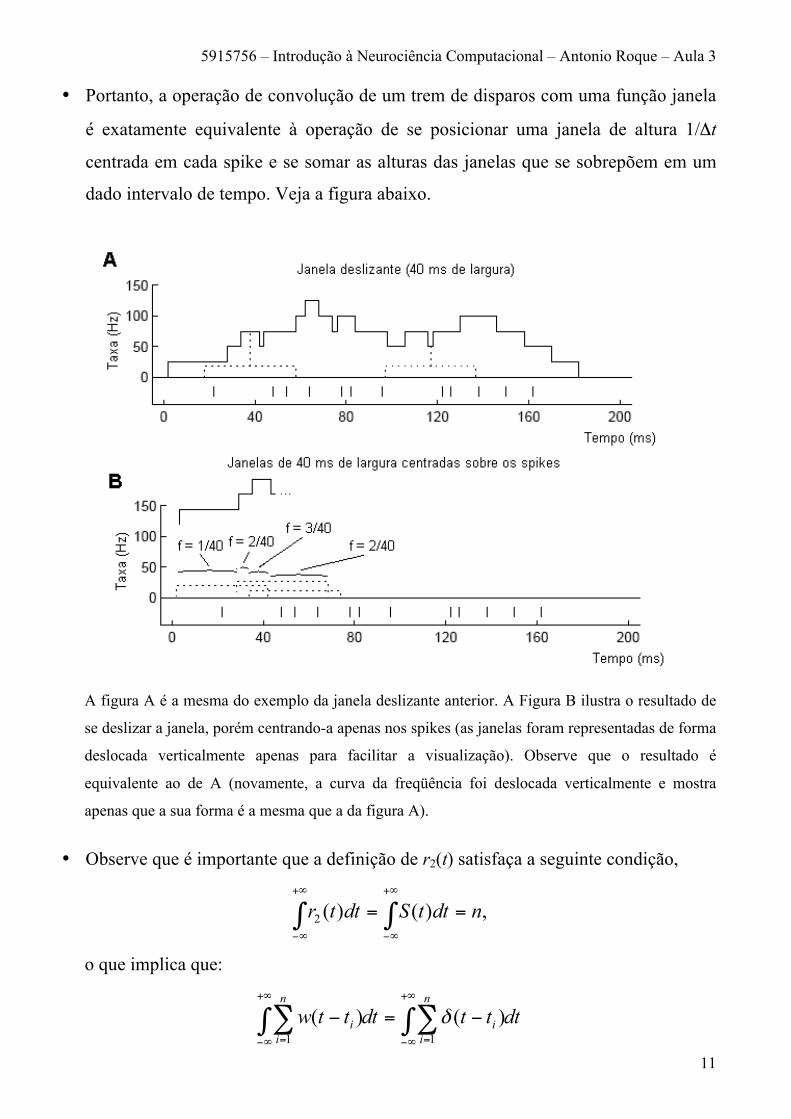
• Portanto, a operação de convolução de um trem de disparos com uma função janela
é exatamente equivalente à operação de se posicionar uma janela de altura 1/Δt
centrada em cada spike e se somar as alturas das janelas que se sobrepõem em um
dado intervalo de tempo. Veja a figura abaixo.
A figura A é a mesma do exemplo da janela deslizante anterior. A Figura B ilustra o resultado de
se deslizar a janela, porém centrando-a apenas nos spikes (as janelas foram representadas de forma
deslocada verticalmente apenas para facilitar a visualização). Observe que o resultado é
equivalente ao de A (novamente, a curva da freqüência foi deslocada verticalmente e mostra
apenas que a sua forma é a mesma que a da figura A).
• Observe que é importante que a definição de r2(t) satisfaça a seguinte condição,
∫∫+∞
∞−
+∞
∞−
== ,)()(2 ndttSdttr
o que implica que:
∫∑∫∑+∞
∞− =
+∞
∞− =
−=− dtttdtttwn
ii
n
ii
11)()( δ

5915756 – Introdução à Neurociência Computacional – Antonio Roque – Aula 3
12
⇒−=− ∑ ∫∑ ∫=
+∞
∞−=
+∞
∞−
n
ii
n
ii dtttdtttw
11)()( δ
.1)( =⇒ ∫+∞
∞−
dttw
• Desta forma, qualquer função w(t) que tenda para zero fora de uma pequena região
centrada em t = 0 e que satisfaça a condição acima pode ser usada como núcleo na
definição de r2(t).
• Um exemplo é a função gaussiana. A figura D ilustra o que acontece com r2(t)
quando se usa como kernel a função gaussiana:
.21)(
2
2
2 w
t
w
etw σ
σπ
−
=
• O parâmetro σw da função acima controla a largura da janela gaussiana, tendo papel
similar a Δt no caso anterior. Note que σw tem dimensão de t, de maneira que a
janela gaussiana tem dimensão de t−1 como deve ser para que a definição de r2(t)
tenha dimensão de freqüência (spikes/seg).
• Observe que o uso de uma função contínua como a gaussiana implica em uma
função r2(t) suave no tempo (veja a figura D da página 7).
• As duas funções janela vistas acima são simétricas em torno da origem, tendo
valores não nulos tanto para t negativo como para t positivo. Não há nada de errado
com isso quando se quer analisar um trem de spikes experimentalmente, após o
registro dos spikes, mas se se quer usar este modelo para entender como um
neurônio estaria decodificando a informação contida em um trem de disparos
emitido por um neurônio pré-sináptico a ele deve-se levar em consideração o fato de
que o neurônio pós-sináptico só tem acesso aos spikes que aconteceram antes do
tempo t.
• Chamamos a isso de condição de causalidade, ou seja, um fenômeno só pode ter
como explicação fenômenos que ocorreram antes dele no tempo.

5915756 – Introdução à Neurociência Computacional – Antonio Roque – Aula 3
13
• Olhando para a definição de r2(t), ∫+∞
∞−
−= τττ dtSwtr )()()(2 , vemos que se
considerarmos valores negativos de τ estaremos usando spikes emitidos após o
tempo t. Para evitar isso, e, portanto, respeitarmos a condição de causalidade,
podemos definir que só vamos usar valores positivos de τ.
• Uma maneira de se fazer isso sem alterar a fórmula acima é definir o núcleo w(t) de
maneira que ele só tenha valores não-nulos para t > 0.
• Um kernel definido dessa forma é chamado de causal. Ele pode ser definido pela
notação:
[ ]⎩⎨⎧
<
≥== + .0 se0
0 se)()()(
tttw
twtw
• A figura E da página 7 mostra o efeito sobre r2(t) do uso de um núcleo causal dado
pela chamada função alfa,
( )[ ] .exp)( 2+−= tttw αα
• Exercício: faça o gráfico da função alfa para ver como ela é. Note que o parâmetro
α determina a extensão da janela (e tem dimensão de t−1).
• Compare as figuras D (que usa um núcleo não causal) e E (que usa um núcleo
causal). Observe que os picos da taxa de disparos calculados com o uso do núcleo
causal tendem a ocorrem depois dos picos calculados com o uso do núcleo simétrico
no tempo.
• A determinação da taxa de disparos dependente do tempo depende criticamente do
intervalo de tempo Δt usado.
• Valores pequenos de Δt implicam em poucos spikes sendo considerados (para um
intervalo muito pequeno é provável que o neurônio emita apenas um spike ou
nenhum), resultando em uma amostragem insuficiente para que se possa tomar uma
decisão. Por outro lado, intervalos muito longos reduzem a capacidade de se
acompanhar variações rápidas no estímulo.
• Do ponto de vista do cérebro, parece pouco provável que o esquema de codificação
apresentado acima seja usado de maneira geral.

5915756 – Introdução à Neurociência Computacional – Antonio Roque – Aula 3
14
• Para entender o problema, pense nos tempos de reação. Por exemplo, uma mosca
pode detectar um estímulo novo e mudar sua trajetória de vôo dentro de 30−40 ms.
Humanos podem detectar imagens apresentadas em uma sequência de imagens não
relacionadas mesmo que cada imagem seja apresentada por apenas 14-100 ms.
• Um neurônio pós-sináptico que usasse uma janela de tempo com uma largura Δt tão
pequena seria capaz de contar apenas uns dois ou três spikes e isso não seria
suficiente para uma estimativa confiável da freqüência (por causa da irregularidade
nos tempos dos spikes).
• A solução encontrada experimentalmente (em estudos offline) para este problema é
tomar médias por várias repetições de um mesmo estímulo, gerando o que se chama
de histograma de tempos pós-estímulo (ou peri-estímulo) PSTH (do inglês post-
stimulus time histogram ou peri-stimulus-time histogram).
• A idéia de um PSTH está ilustrada na figura abaixo (retirada do livro de Gerstner e
Kistler). K é o número de repetições.
• A apresentação gráfica das seqüências dos tempos de disparo do neurônio para cada
repetição do experimento de forma ordenada, uma seqüência em cima da outra, é
muito conveniente para se ter uma visualização da variabilidade dos disparos entre
as repetições.
5915756 – Introdução à Neurociência Computacional – Antonio Roque – Aula 3
15
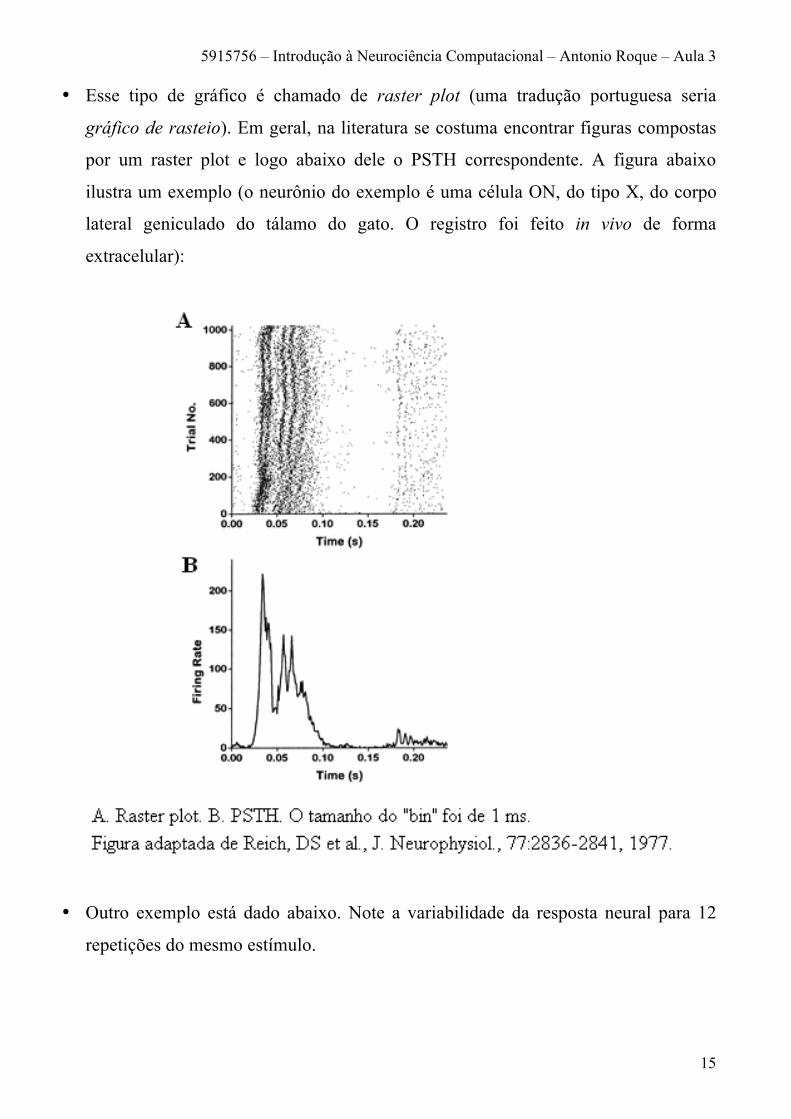
• Esse tipo de gráfico é chamado de raster plot (uma tradução portuguesa seria
gráfico de rasteio). Em geral, na literatura se costuma encontrar figuras compostas
por um raster plot e logo abaixo dele o PSTH correspondente. A figura abaixo
ilustra um exemplo (o neurônio do exemplo é uma célula ON, do tipo X, do corpo
lateral geniculado do tálamo do gato. O registro foi feito in vivo de forma
extracelular):
• Outro exemplo está dado abaixo. Note a variabilidade da resposta neural para 12
repetições do mesmo estímulo.

5915756 – Introdução à Neurociência Computacional – Antonio Roque – Aula 3
16
• A taxa de disparos dependente do tempo obtida segundo este método pode ser
escrita como,
),;(11)(3 tttnKt
tr K Δ+Δ
=
onde K é o número de repetições do experimento e nk(t; t + Δt) é a soma do número
de spikes entre t e t + Δt ao longo das K repetições.
• Costuma-se representar a média de alguma variável u tomada por todas as K
repetições de um mesmo estímulo por <u>. Desta forma, pode-se escrever,
∫Δ+
Δ=
tt
t
dSt
tr ττ )(1)(3 .
• Pode-se definir r3(t) em termos de uma convolução com um núcleo w(t), como feito
no caso do registro do trem de disparos para uma única apresentação de um
estímulo, mas isto não será feito aqui.

5915756 – Introdução à Neurociência Computacional – Antonio Roque – Aula 3
17
• Apesar de sua utilidade prática na análise dos trens de disparo de neurônios, o
método de se fazer uma média sobre várias repetições de um estímulo não pode ser
o metido de decodificação usado pelos neurônios do cérebro. O motivo,
simplesmente, é que os animais têm que tomar suas decisões com base em uma
única repetição de um estímulo.
• Este problema com a definição de r3(t) nos leva a uma outra definição de taxa de
disparos que poderia ser utilizada por neurônios do cérebro. A idéia está em que ao
invés de se imaginar uma média por K repetições de um experimento pode-se
imaginar que um neurônio esteja fazendo uma média por K neurônios pré-sinápticos
a ele.
• Imagine que exista uma população composta por N neurônios com propriedades
idênticas (ou muito parecidas) e que recebam todos o mesmo estímulo. Como o
cérebro tem uma densidade muito grande de neurônios, uma tal situação não é
implausível.
• Imagine agora que exista um neurônio que receba sinapses de todos esses N
neurônios. Do seu ponto de vista, a variável importante é a quantidade de spikes que
ele recebe em seus dendritos a cada intervalo de tempo Δt dividida por N (a divisão
por N é feita para que se possa estimar o que cada neurônio da população pré-
sináptica está informando individualmente, em média). Dividindo-se esta
quantidade por Δt temos uma outra definição de taxa de disparos:
( )( )
,1;1)( 14 N
dttt
tNtttn
ttr
tt
t
N
j i
ji∫ ∑∑
Δ+
=
−
Δ=
Δ+
Δ=
δ
onde n(t; t + Δt) é o número de spikes emitidos por todos os N neurônios entre t e t +
Δt e, para levar em conta que agora temos mais de um neurônio, redefiniu-se a
expressão que representa a seqüência de disparos de um neurônio (digamos, o
neurônio j) por
( )∑=
−=n
i
ji
j tttS1
)( δ .

5915756 – Introdução à Neurociência Computacional – Antonio Roque – Aula 3
18
• O maior problema com a codificação por taxa baseada na média por uma população
de neurônios é que ela requer que todos os neurônios da população sejam idênticos.
Isso é pouco realista, mas evidências experimentais indicam que os neurônios
corticais das chamadas áreas sensoriais primárias (V1, A1 e S1, indicando os
córtices visual primário, auditivo primário e somestésico primário respectivamente)
estão arranjados em colunas cujas células têm propriedades similares.
• Há muito mais sobre o problema do código neural que poderia ser discutido nesta
aula. Para um aprofundamento neste tema, recomendam-se os seguintes livros:
- Rieke, F., Warland, D., de Ruyter van Steveninck, R. and Bialek, W., Spikes:
Exploring the Neural Code, MIT Press, Cambridge, MA, 1997. Capítulos 1 e
2.
- Dayan, P. and Abbott, L.F., Theoretical Neuroscience: Computational and
Mathematical Modeling of Neural Systems, MIT Press, Cambridge, MA,
2001. Capítulo 1.
- Gerstner, W. and Kistler, W., Spiking Neuron Models, Cambridge University
Press, Cambridge, UK, 2002. Capítulo 1.
- Quiroga, R. Q. & Panzeri, S., Extracting information from neuronal
populations: information theory and decoding approaches. Nature Reviews
Neuroscience, 10:173-185, 2009

5915756 – Introdução à Neurociência Computacional – Antonio Roque – Aula 3
19
Apêndice: Convolução Discreta
• O formalismo matemático usado acima, em que o tempo foi tratado como uma
variável contínua, é conveniente para um tratamento analítico (simbólico) dos trens
de disparos de neurônios. Porém, para uma análise numérica (usando o computador)
dos trens de disparos é necessário um formalismo em que o tempo é tratado como
uma variável discreta.
• Os registros intra- ou extra-celulares de células nervosas são sinais analógicos. Eles
podem ser discretizados a partir de um processo de amostragem a uma taxa
constante. Por exemplo, imagine um registro cuja duração total foi de 1 segundo. Se
amostrarmos o valor do sinal a cada 0,5 ms teremos, no fim do processo, gerado um
vetor com 1000/0,5 = 2000 componentes.
• Suponha que o sinal analógico já tenha sido pré-processado para isolar os spikes,
como na figura abaixo.
• Neste caso os valores amostrados serão, ou zero, ou um. O vetor que representa o
trem de disparos do neurônio será, portanto, um vetor binário S como o mostrado
abaixo.
S=[0000110001110000111110011001001000000010000].
• Supondo que temos um vetor de N componentes representando a atividade de um
neurônio registrada durante um certo tempo T. O número total de spikes emitidos
pelo neurônio durante esse período pode ser obtido simplesmente como,
.1
0∑−
=
=N
iiSn
• Dividindo-se n pelo tamanho do vetor, temos a fração dos componentes do vetor em
que houve um spike (o número de 1s dividido pelo número total de intervalos ou
bins). Em outras palavras, temos a probabilidade de que o neurônio emita um spike
em um intervalo:

5915756 – Introdução à Neurociência Computacional – Antonio Roque – Aula 3
20
.Nnp =
• Dividindo-se esta probabilidade pela duração Δt de cada intervalo (por exemplo, 0,5
ms), temos a taxa de disparos (spike-count rate) do neurônio baseada na contagem r,
definida na aula passada:
.1Nntt
prΔ
=Δ
=
• Como discutido nesta aula, essa medida de taxa só tem sentido em situações em que
o estímulo é estacionário durante o tempo T. Em situações mais realistas em que o
estímulo varia no tempo é mais conveniente calcularmos taxas para pequenos
segmentos do vetor S, gerando uma taxa de disparos dependente do tempo.
• Nesta aula, vimos que a taxa de disparos dependente do tempo pode ser expressa em
termos de uma convolução entre o sinal S (o trem de disparos) e um núcleo (kernel)
apropriado w (isto é, que satisfaça 1)( =∫+∞
∞−
dttw ),
.)()()(2 ∫+∞
∞−
−= τττ dtSwtr
• Um núcleo causal que pode ser usado é o seguinte:
⎩⎨⎧ Δ≤≤Δ
=contrário. caso0
0 se1)(
ttttw
• Isto corresponde a uma janela de largura Δt cujos valores no seu interior têm valor
1/Δt e os valores fora da janela têm valor 0. A versão de tempo discreto desse
núcleo (para uma janela de tamanho M < N) é um vetor w com as primeiras M
componentes diferentes de zero e iguais a 1/M e as demais componentes até N
iguais a zero:
w = [w0 w1 w2 … wM-1 wM wM+1 … wN-1] = [1/M 1/M 1/M ... 1/M 0 0 ... 0].
• A operação de convolução para tempo discreto pode ser escrita como (veja, por
exemplo, o livro Numerical Recipes cuja referência está dada no fim deste
apêndice):

5915756 – Introdução à Neurociência Computacional – Antonio Roque – Aula 3
21
( ) ,∑+∞
−∞=−=∗
jjjii wSwS
onde o símbolo f∗g é a notação usada para indicar a convolução das funções f e g.
• Para o núcleo causal definido acima, esta expressão torna-se,
( ) .1
0∑−
=−=∗
M
jjjii wSwS
• Podemos ter uma idéia do que uma convolução faz calculando seus termos:
( )
.00
11221100
110220110000
1
000
wSwSwSwSwS
wSwSwSwSwSwS
MM
MM
M
jjj
=
++++=
++++==∗
−+−−−
−+−−−−
−
=−∑
…
…
( )
.1001
111211001
111221111001
1
011
wSwSwSwSwSwS
wSwSwSwSwSwS
MM
MM
M
jjj
+=
++++=
++++==∗
−+−−
−+−−−−
−
=−∑
…
…
( )
.201102
112201102
112222112002
1
022
wSwSwSwSwSwSwS
wSwSwSwSwSwS
MM
MM
M
jjj
++=
++++=
++++==∗
−+−
−+−−−−
−
=−∑
…
…
...
( )
1122110
11221100
1
0
−+−−−
−+−−−−
−
=−
++++=
++++==∗ ∑MMiiii
MMiiii
M
jjjii
wSwSwSwS
wSwSwSwSwSwS
…
…
...
( )
.1231201
111221111001
1
011
−−−−−
−+−−−−−−−−
−
=−−−
++++=
++++==∗ ∑MMNNNN
MMNNNN
M
jjjNN
wSwSwSwS
wSwSwSwSwSwS
…
…
( )
.112211
1122110
11221100
1
0
−+−−−
−+−−−
−+−−−−
−
=−
+++=
++++=
++++==∗ ∑
MMNNN
MMNNNN
MMNNNN
M
jjjNN
wSwSwSwSwSwSwS
wSwSwSwSwSwS
……
…

5915756 – Introdução à Neurociência Computacional – Antonio Roque – Aula 3
22
( )
.1221
12211001
111221111001
1
011
−+−−
−+−−++
−+−+−+−+−+
−
=−++
++=
++++=
++++==∗ ∑
MMNN
MMNNNN
MMNNNN
M
jjjNN
wSwSwSwSwSwS
wSwSwSwSwSwS
……
…
...
( )
( )( ) .112
112413022
1111221111110011
1
01111
−−−+
−−−+−+−+−+
−+−−+−−−+−−−+−−−+−
−
=−−+−−+−
=∗
++++=∗
++++==∗ ∑
MNMN
MNMNMNMNMN
MMMNMNMNMN
M
jjjMNMN
wSwSwSwSwSwSwS
wSwSwSwSwSwS
…
…
• Portanto, S∗w tem N + M − 1 componentes. Por exemplo, se o vetor S tiver N =
2000 componentes e o núcleo w tiver M = 5 componentes não nulas a convolução
S∗w terá 2004 componentes.
• Observe que os termos das extremidades têm menos componentes não nulas. Por
causa disso, não se deve confiar muito neles e é costume desprezar os primeiros L e
os últimos L termos de uma convolução, para um tamanho L apropriado.
• A página anterior ficou muito carregada, mas para entender o significado da
convolução podemos olhar apenas para o termo genérico:
( )
1122110
11221100
1
0
−+−−−
−+−−−−
−
=−
++++=
++++==∗ ∑MMiiii
MMiiii
M
jjjii
wSwSwSwS
wSwSwSwSwSwS
…
…
• Para ajudar na visualização, podemos ordenar as séries temporais de S e de w de
modo que os termos sobrepostos correspondam aos termos multiplicados nesta
expressão:
00000 0121
1211210
………………
wwwwSSSSSSSSS
M
NiiiiiMiMi
−
−++−−+−−
• O valor da convolução de S com o núcleo w no tempo i corresponde a “olhar” para o
passado do sinal S com uma janela de comprimento M começando no instante i e se
estendendo para trás no tempo até i − M + 1. O i-ésimo valor da convolução é então
calculado pela soma ponderada dos valores de S dentro dessa janela com os pesos
dados por w.

5915756 – Introdução à Neurociência Computacional – Antonio Roque – Aula 3
23
• O núcleo causal apresentado acima satisfaz à condição 1=∑i iw , de maneira que a
convolução (S∗w)i dá a probabilidade pi de que o neurônio dispare no tempo i. Por
esta razão, podemos escrever:
( ) .1
0∑−
=−=∗=
M
jjjiii wSwSp
• Esta probabilidade é um número adimensional. Para transformá-la em uma taxa de
disparos (em spikes por segundo) temos que dividir por Δt:
∑−
=−Δ
=Δ
=1
0.1 M
jjji
ii wS
ttp
r
• Referências:
• Para saber mais sobre o livro Numerical Recipes: The Art of Scientific Computing,
Cambridge University Press, Cambridge, UK, visite a página do livro:
http://www.nr.com/


















