
INTERAZIONI INTERMOLECOLARI NELLE PROTEINE -...
75
INTERAZIONI INTERMOLECOLARI INTERAZIONI INTERMOLECOLARI NELLE PROTEINE 1 1
Transcript of INTERAZIONI INTERMOLECOLARI NELLE PROTEINE -...

INTERAZIONI INTERMOLECOLARIINTERAZIONI INTERMOLECOLARINELLE PROTEINE
11

RICHIAMI DI TERMODINAMICAE t l i2 EntalpiaEntropiaEnergia Libera di Gibbs
2
g

ENERGIA INTERNA (U)L’energia interna riassume tutti i contributi cinetici e potenziali dell’energia di tutti gli atomi, ioni e molecole costituenti il sistema: è l’energia ioni e molecole costituenti il sistema: è l energia totale macroscopica del sistemaU non può essere misurata ma possono essere U non può essere misurata, ma possono essere misurate le sue variazioniΔU= w + qΔU w qDove w=energia fornita al sistema come lavoro e q= energia fornita al sistema come caloreq gPrimo principio: L’energia interna di un sistema isolato è costante 3g 3

ENTALPIA (H)Definizione formale di entalpiaDefinizione formale di entalpia
H=U + pV
dove U è l'energia interna del sistema, p la sua pressione e V il suo volume. L'entalpia risulta pertanto una grandezza termodinamica estensiva.
Nel caso, che capita molto spesso di studiare in chimica, di trasformazioni a pressione costante, la variazione di entalpia corrisponde esattamente alla variazione di calore:
ΔH= ΔQ
Endotermica aumento entalpiaEsotermica diminuzione entalpia
Entalpia di reazione: variazione di entalpia che accompagna una reazione quale è specificata dalla equazione chimica associata quantità di calore assorbito o ceduto nel corso della reazioneEntalpia standard di reazione (ΔHo)entalpia relativa alla trasformazione dei reagenti considerati nei rispettivi stati standard in prodotti considerati nei loro rispettivi stati standardΔH° = entalpia in condizioni standard biologiche p g
La legge di Hess stabilisce che in una reazione chimica, l'effetto termico a pressione costante è indipendente dagli stati intermedi attraverso i quali si evolve il sistema e dipende solo dal suo stato iniziale e finale.In altre parole ciò significa che la variazione di entalpia di una reazione che può essere scomposta 4In altre parole ciò significa che la variazione di entalpia di una reazione che può essere scomposta idealmente in più reazioni parziali è pari alla somma algebrica delle variazioni di entalpia dei singoli stadi.
4

ENTROPIA (S)ΔS= ΔQ /TΔS= ΔQrev/TLa variazione di entropia di una sostanza (ΔS) eguaglia il calore ad esso trasferito reversibilmente, diviso per la temperatura (inscala Kelvin) alla quale il trasferimento di calore ha avuto luogoIl t f i t è ibil d i t bi t i t h Il trasferimento è reversibile quando avviene tra un ambiente e un sistema che hanno una idfferenza di T infinitesima. Altrimenti la trasformazione diventa irreversibile (con formazioni di punti caldi che disperdono il calore e aumentano l’entropia del sistema) e l’equazione diventa:ΔS> ΔQ /TΔS> ΔQirrev/T
Maggiore è l’energia trasferita come calore, più viene stimolato il moto caotico. Maggiore risulta l’aumento di entropia
Secondo principio:Nel corso di una trasformazione spontanea l’entropia di un sistema isolato aumenta.
Entropia standard di reazione: il ΔSo è la differenza tra l’entropia standard dei prodotti e quella dei reagenti.
55

PROCESSI SPONTANEIL’O e L’H in stato gassoso reagiscono spontaneamente e violentemente L O2 e L H2 in stato gassoso reagiscono spontaneamente e violentemente a temperatura ambiente con la formazione di H2O, eppure si passa da uno stato più disordinato a uno più ordinato, con una variazione di entalpia molto negativa.
Questo non va contro il secondo principio?
Bi id l i i di t i di t tt il i t d t Bisogna considerare la variazione di entropia di tutto il sistema, dato che il processo passa del calore all’ambiente in cui viene disperso con un aumento di entropia molto maggiore della diminuzione di entalpia
L’aumento di entropia (a temperatura e pressioni costanti) è dato da:ΔSambiente= ΔQrev/T -ΔH/T essendo -ΔH=ΔQrev (calore trasferito all’ambiente)
La variazione totale di entropia che accompagna un qualunque processo è:ΔStotale= ΔSprocesso+ ΔSambiente ΔStotale= ΔSprocesso - ΔH/T
6TΔStotale= TΔSprocesso - ΔH 6

ENERGIA LIBERA DI GIBBS (G)D fi i l’ i lib di GibbDefiniamo l’energia libera di Gibbs comeG = H – TSE l i i t t t t àE la sua variazione a temperatura costante sarà:ΔG= ΔH – TΔSE di conseguenza a T e p costanti:E di conseguenza a T e p costanti:ΔG= -TΔStotale
Perché un processo sia spontaneo deve avvenire con ΔS>0 cioè con ΔG<0
L’energia libera esprime la variazione di entropia totale del processo e dell’ambiente 7entropia totale del processo e dell ambiente 7

PROCESSI SPONTANEI
Una reazione è spontanea se:Reazione esotermica: ΔH ti ΔS tiΔH negativa e ΔS negativa
Reazione endotermica: ΔH positiva ma TΔS è sufficientemente grande e positivo.ΔH positiva ma TΔS è sufficientemente grande e positivo.A determinate temperature anche le reazioni endotermiche possono diventare spontanee.La temperatura minima necessaria perché ciò avvenga è:La temperatura minima necessaria perché ciò avvenga è:ΔG= ΔH- TΔS=0 T= ΔH/ ΔS
88

TIPI DI INTERAZIONITIPI DI INTERAZIONIINTERMOLECOLARI
99

TIPI DI INTERAZIONI
L i l tiLegami covalentiLegame ionicoL i idLegami a idrogenoInterazioni dipolo-dipoloInterazioni idrofobicheInterazioni idrofobicheInterazioni di Wan del Walls
1010

LEGAME COVALENTE
Energia: ΔG° da -40 a -110 Kcal/mole
Ponte disolfuroS-H + H-S S-S
1111

LEGAME IONICO
Energia: ΔG° da -5 a -10 Kcal/mole
Avviene tra due atomi aventi una carica netta
1212

INTERAZIONIIONE-DIPOLO E DIPOLO-DIPOLO
Energia: ΔG° da -1 a -7 Kcal/moleN, O e S sono più elettronegativi del C I legami C-X (dove X è un atomo fortemente elettronegativo) sono dei dipoliI di li i d l i di li d I dipoli possono essere attratti da altri dipoli o da ioni carichi
1313

LEGAME A IDROGENO
Energia: ΔG° da -3 a -5 Kcal/moleÈ un tipo particolare di interazione dipolo-dipoloAvviene tra un gruppo A-H e un atomo B (dove A e B sono due atomi fortemente elettronegativi) avente una coppia di elettroni di non legameavente una coppia di elettroni di non legame.
È l lt t di i l È un legame altamente direzionale
A H + B A H B A-H + :B A-----H-----B Se A e B hanno la stessa elettronegatività il protone è in comune tra i due 14protone è in comune tra i due 14

LEGAME A IDROGENO
Gli atomi elettronegativi che si trovano nelle catene proteiche sono N, O e S.
Tutti gli aminoacidi possono formare legami a idrogeno con gli atomi che formano il backboneidrogeno con gli atomi che formano il backbone
A i idi h f l i id l Aminoacidi che formano legami a idrogeno con la catena laterale:
Arg Lys Cys Asp Glu Met Thr Tyr Asn e GlnArg, Lys, Cys, Asp, Glu, Met, Thr, Tyr, Asn e Gln
1515

FORZE DI VAN DER WALLS
Energia: ΔG° circa -0,5 Kcal/mole
Deboli forze universali che diventano significative se c’è uno stretto contatto tra due molecolemolecole
S l di t è ffi i t t b i di li Se la distanza è sufficientemente bassa i dipoli temporanei di una molecola inducono dipoli contrari nell’altra molecola da cui deriva una contrari nell altra molecola da cui deriva una debole attrazione
1616

INTERAZIONI IDROFOBICHE(CONTRIBUTO ENTROPICO)
Energia: ΔG° da -0,7 a -1 Kcal/mole
Gruppi lipofili in soluzione acquosa tendono a d i l l lnascondersi al solvente polare.
Il guadagno di energia liberà è dato dall’aumento di entropia delle molecole di acqua circostanti.
1717
FOLDING1818

LEGAMI
Tre tipi di legami sono importanti per la formazione della struttura terziaria della proteina:proteina:
Legami a idrogenoLegami ioniciLegami ioniciPonti disolfuro
1919

LEGAMI A IDROGENO
Il 90% dei gruppi C=O e N-H del backbone forma legami a idrogenoI i N H f l i i li t il I gruppi N-H formano legami singoli, mentre il 35% dei gruppi C=O può essere coinvolto in due o tre legami a idrogenotre legami a idrogenoMolti di questi legami a idrogeno (50% dei CO e 30% dei NH) sono formati con l’acqua30% dei NH) sono formati con l acquaL’acqua è una parte fondamentale della struttura della proteina, potendo fungere sia da accettore p , p gche da donatore di legami a H, fa spesso da ponte.
2020

LEGAMI IONICI
Si formano tra coppie ioniche
A causa del momento di dipolo Asp e Glu si legano con l’N-ter delle alfa eliche, mentre Lys e Arg con il C terArg con il C-ter
Di t i 5 0 ÅDistanza: circa 5.0 ÅEnergia: 5-10 Kcal/mol
2121

LEGAME DISOLFURO
Avviene tra due cisteine.
La distanza del legame S-S è 2.07 Å contro gli 1.55 Å del C-C
2222

SPIEGAZIONE STRUTTURA COMPATTA
Nessuno di questi tre tipi di legame può spiegare la formazione della struttura compatta delle proteineproteine.
Legami disolfuro non sempre presentiLegami disolfuro non sempre presentiLegami ionici, non sono conservati evolutivamente e sono pochi: in media 5 ogni 150 residui Legami a Idrogeno: per formare dei legami idrogeno tra due residui della proteina si devono rompere quelli formati tra la proteina in struttura primaria e quelli formati tra la proteina in struttura primaria e l’acqua. Nella migliore delle ipotesi il numero totale di legami non cambia
2323

Il folding è guidato dall’effetto idrofobico che porta a nascondere i residui idrofobici all’interno della proteina Il peso da pagare in termini di della proteina. Il peso da pagare in termini di entalpia è la desolvatazione dei residui polari. Per questo motivo gran parte dei residui forma q g plegami a idrogeno nel backbone.
2424

STRUTTURA DELLE PROTEINE2525

AMINOACIDI E STRUTTURAAMINOACIDI E STRUTTURAPRIMARIA
2626
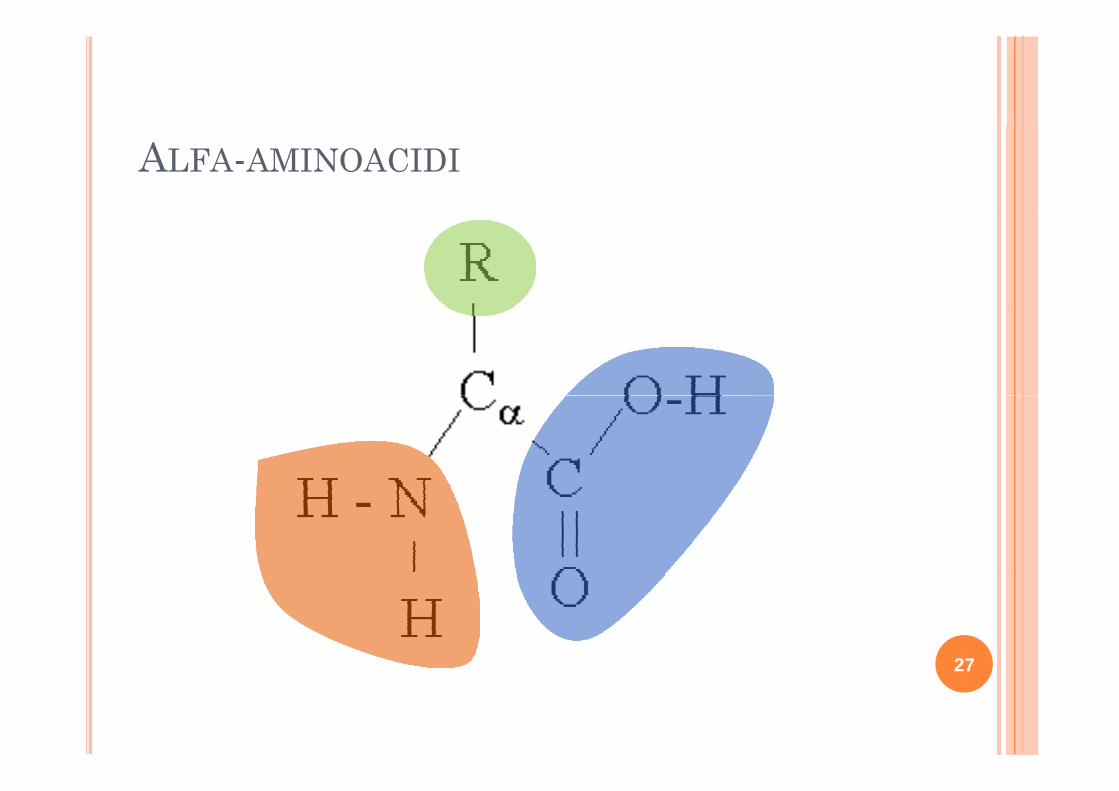
ALFA-AMINOACIDI
2727

TIPI DI AMINOACIDI
2828

LEGAME PEPTIDICO
Formazione Angoli di rotazione
2929

RAMACHANDRAN PLOT
Psi ( )
La glicina può assumere piùassumere più angoli di rotazione degli altri 30
Phi ( )
altri 30

LIVELLI DI STRUTTURA
Primaria
SecondariaSecondaria
Terziaria
Quaternaria3131
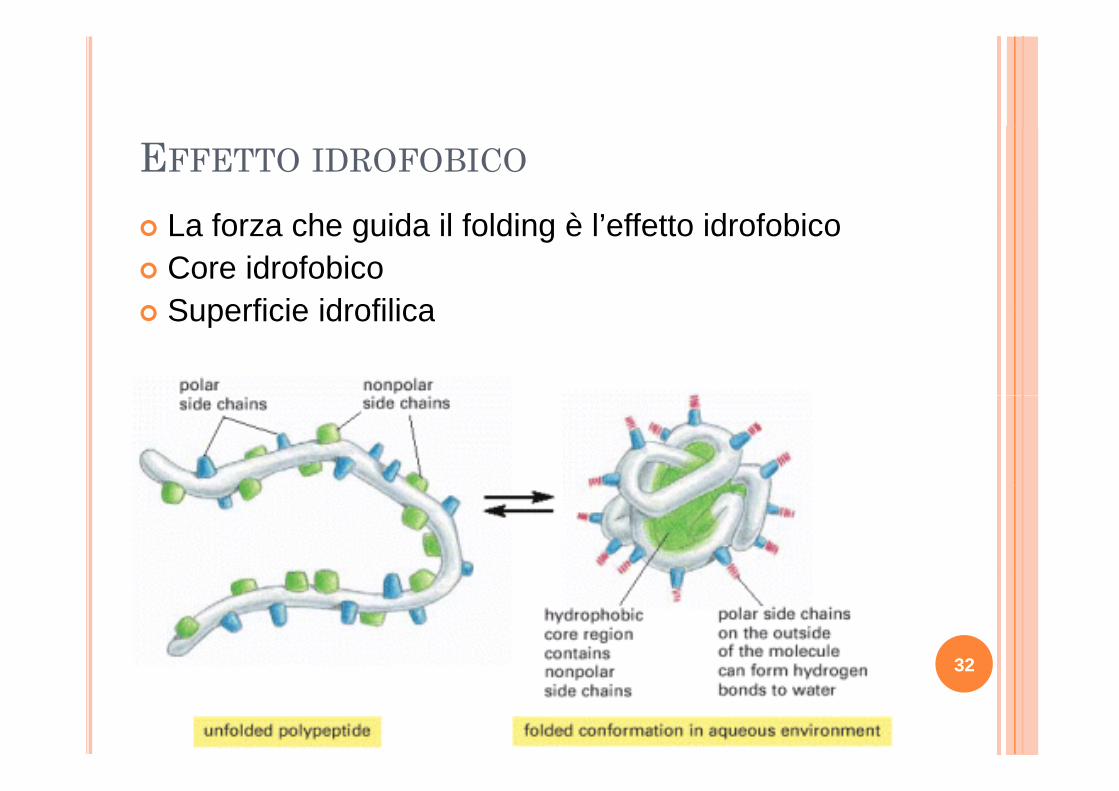
EFFETTO IDROFOBICO
L f h id il f ldi è l’ ff tt id f biLa forza che guida il folding è l’effetto idrofobicoCore idrofobicoSuperficie idrofilicaSuperficie idrofilica
3232

STRUTTURA SECONDARIAAlf Eli h F li tti B t33 Alfa-Eliche e Foglietti-Beta33

FORMAZIONE DELLE STRUTTURESECONDARIE
Il backbone è polare anche se i residui sono idrofobici
Formazione di legami a idrogeno tra i gruppi C=O e N H dei legami peptidiciC=O e N-H dei legami peptidici
F i di t tt d iFormazione di strutture secondarie.
3434

ALFA-ELICHE
3535
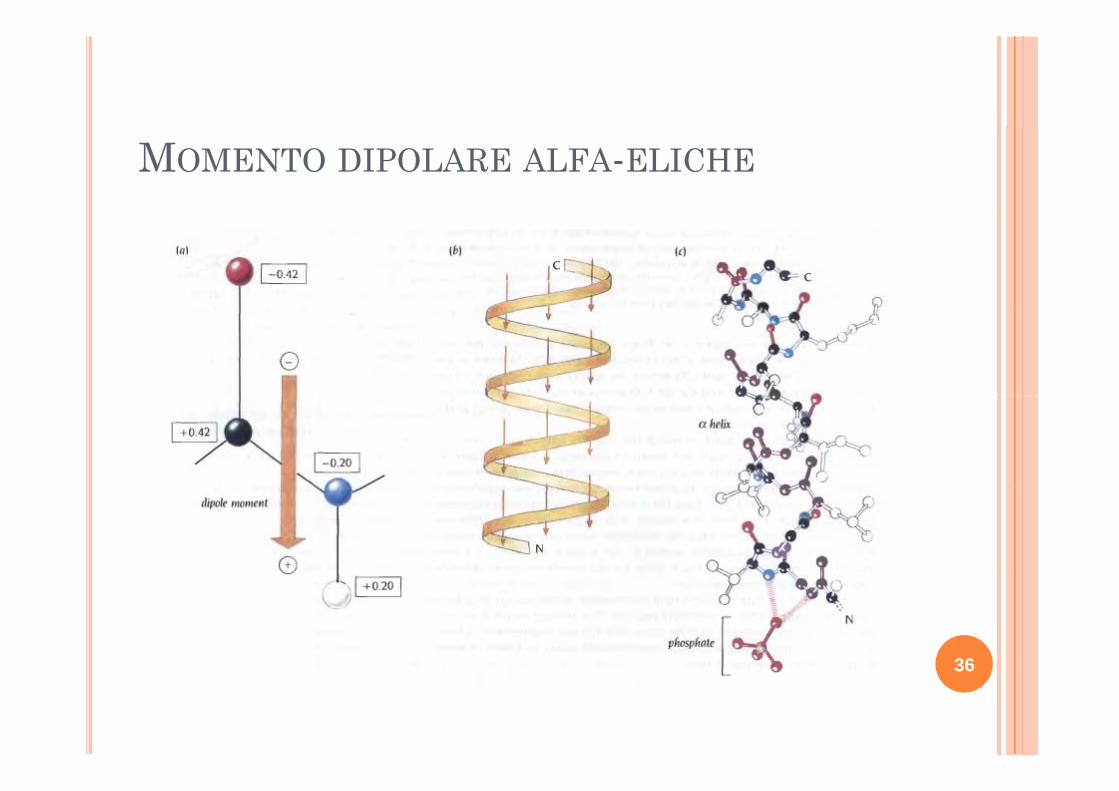
MOMENTO DIPOLARE ALFA-ELICHE
3636

AMINOACIDI PREFERITI
Alcuni aminoacidi hanno una leggera preferenza a trovarsi in alfa-eliche:
Al Gl L M tAla, Glu, Leu, Met
Altri vi si ritrovano meno spessoAltri vi si ritrovano meno spessoPro, Gly, Tyr, Ser
3737

FOGLIETTI BETA
Ogni strand è lungo da 5 a 10 residui in configurazione estesa
Gli angoli phi e psi sono nel quadrante in alto a sinistrasinistra
T ti iTre tipi:ParalleliAntiparalleliAntiparalleliMisti
3838

FOGLIETTI BETA PARALLELI
3939

FOGLIETTI BETA ANTIPARALLELI
4040

FOGLIETTI BETA MISTI
4141

REGIONI DI LOOP
I motivi sono formati dall’unione di più strutture secondarie, connesse tra loro da loopI l h t tt d fi it il l I loop non hanno una struttura definita e il loro backbone è esposto al solvente e libero di formare legami a idrogeno con il solventelegami a idrogeno con il solventeContengono maggiormente residui carichi o polaripolariSono evolutivamente meno stabili che le regioni organizzate in strutture secondariegIn natura assumono un numero definito di strutture
42Loop molto lunghi sono in genere molto flessibili
42

HAIRPIN LOOP
Le regioni di loop che connettono due filamenti beta adiacenti sono chiamati Hairpin loopS lt b i d d ll Sono spesso molto brevi ed assumono delle strutture caratteristiche
4343
STRUTTURA TERZIARIAM ti i D i i F ld44 Motivi, Domini e Fold44

DOMINI
L’unità fondamentale della struttura terziaria è il dominioD fi i i di d i i t li tidi Definizione di dominio: catena polipeptidica o parte di una catena polipeptidica che è in grado di assumere indipendentemente una struttura di assumere indipendentemente una struttura terziaria stabileLe proteine possono essere formate da un solo Le proteine possono essere formate da un solo dominio o da molti domini.La distinzione tra dominio e subunità è effimera:
In alcuni organismi una funzione è espressa da più catene polipeptidiche in altri è una catena unica f t d iù d i i 45formata da più domini 45

I QUATTRO DOMINI DI SRC
4646

I DOMINI SONO COMPOSTI DA MOTIVI DISTRUTTURE SECONDARIE
Motivi: combinazioni di strutture secondarie
I domini sono formati da diverse combinazioni di strutture secondarie e motivi
Il numero totale di fold esistenti in natura è ti t ll’ di di 1000stimato nell’ordine di 1000
P t i d i i h h Proteine, o domini che hanno una sequenza identica per almeno il 30% hanno lo stesso fold
4747

3 CLASSI PRINCIPALI DI STRUTTUREPROTEICHE
AlfAlfaIl core della proteina è costituito esclusivamente da alfa-eliche
BetaIl d ll t i è tit it d f li tti b t ti ll li Il core della proteina è costituito da foglietti beta antiparalleli. Generalmente sono presenti due foglietti disposti l’uno contro l’altro.
Alfa/BetaAlfa/BetaSono formate dalla combinazione di motivi alfa-beta-alfa. In generale è presente un foglietto beta parallelo circondato da alfa-eliche
4848

CLASSI MINORI
Alfa + BetaCombinazioni discrete di motivi alfa e beta. Formano un foglietto antiparallelo da una parte e le alfa eliche un foglietto antiparallelo da una parte e le alfa eliche si raggruppano dall’altra
Metalloproteine proteine con molti ponti disolfurop p pSono versioni distorte di proteine a struttura più regolare, la struttura è fortemente influenzata dalla
di t i t lli i ti di lfpresenza di atomi metallici e ponti disolfuro.
4949

STRUTTURE A DOMINI ALFA5050

COILED COILS
ÈÈ un motivo strutturaleRipetizioni eptadiEffetto idrofobicoPonti salini
5151

COILED COILS
Posssono essere lunghe anche centinaia di residui in proteine filamentoseSi it h l h i Si ritrovano anche, con una lunghezza minore, come motivo di dimerizzazione nei fattori di trascrizione: leucine-zippertrascrizione: leucine zipper
5252

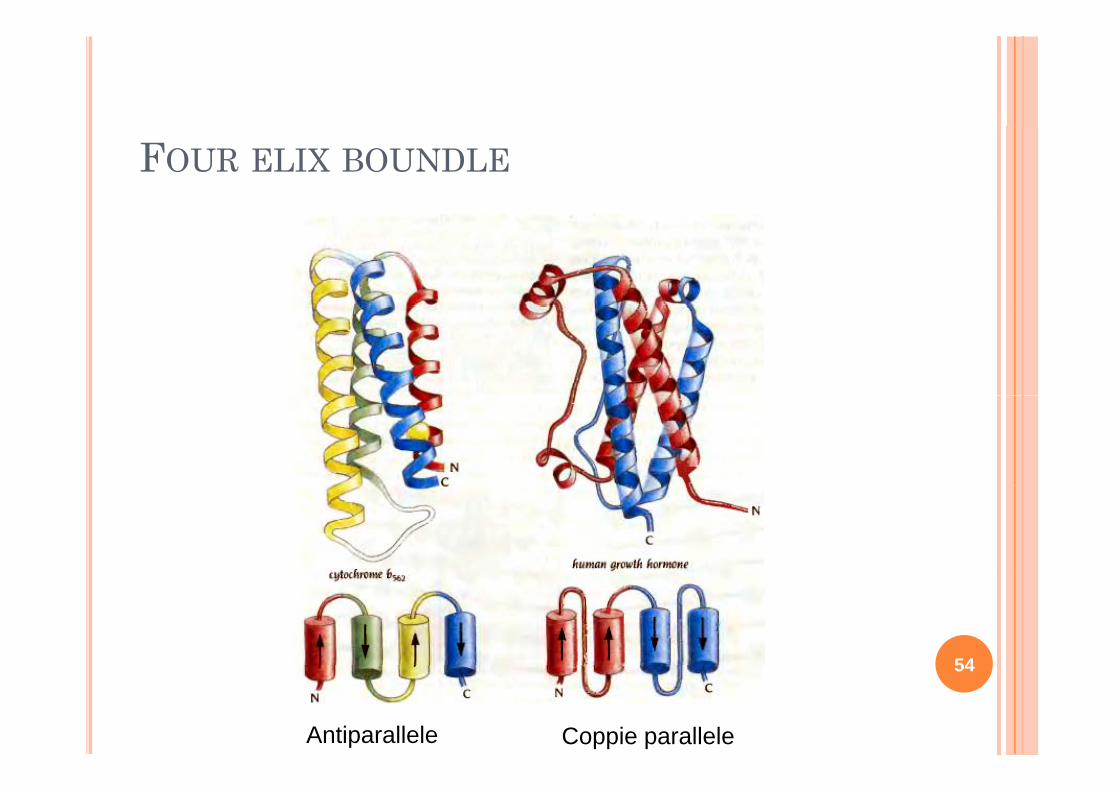
FOUR ELIX BOUNDLE
È il d i i d lfÈ il dominio ad alfa-eliche più semplice e più diffusop soLe catene laterali idrofobiche sono ll’i d ll all’interno delle
quattro eliche. L’impaccamento è L impaccamento è molto stretto e quindi questa regione non è accessibile all’acquaaccessibile all acquaLe catene idrofilichesono invece esposte 53sono invece esposte al solvente
53
FOUR ELIX BOUNDLE
54
Antiparallele Coppie parallele
54

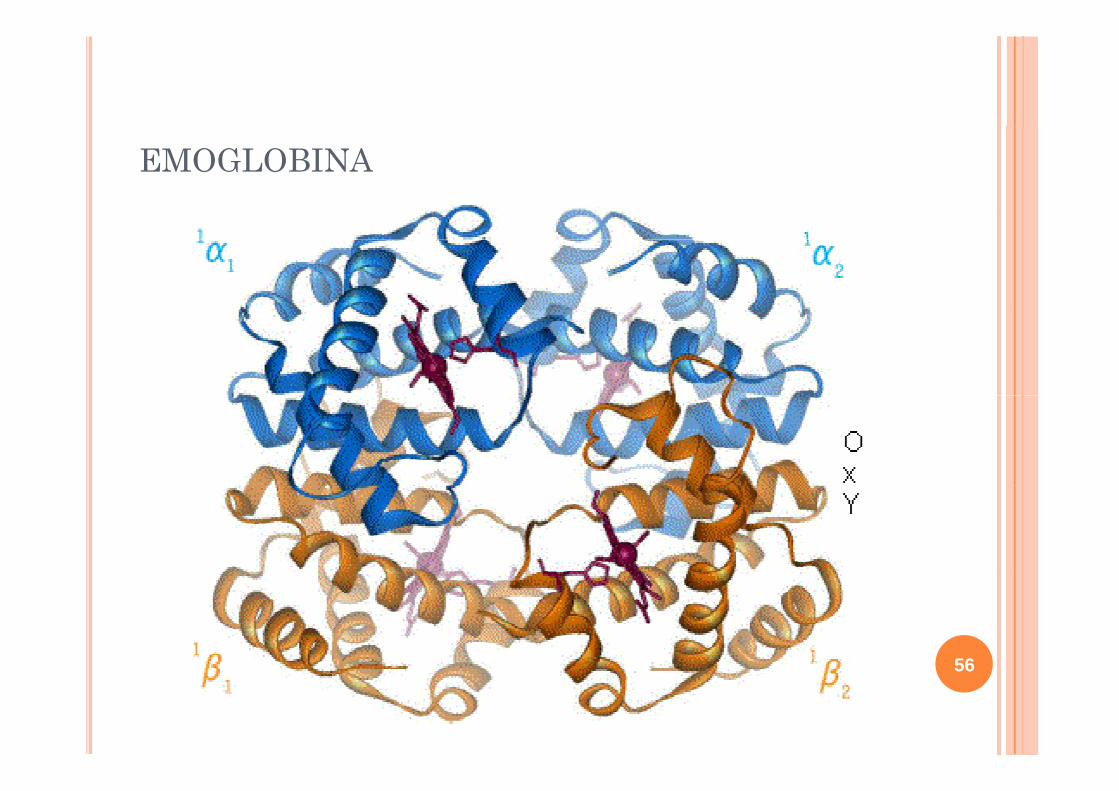
FOLD GLOBINICOMioglobina
Bundle di 8 alfa-eliche connesse tra loro da brevi looploro da brevi loopNon è costituito da unità più sempliciunità più semplici
5555
EMOGLOBINA
5656

STRUTTURE A DOMINI ALFA/ STRUTTURE A DOMINI ALFA/ BETA
5757

ALFA/BETAÈ l l t tt l iù t tÈ la classe strutturale più rappresentataConsiste di un foglietto-beta parallelo (o misto) interno circondato da alfa-elicheTutti gli enzimi della glicolisi appartengono a questa classeSono formati dal susseguirsi di motivi beta-alfa-betag
3 classi principali:TIM barrelRossman Fold 58Rossman FoldHorseshoe fold
58

TIM BARREL
f Core formato da un Beta-barrel di filamenti paralleli
Sono necessari almeno 5 Sono necessari almeno 5 filamenti per fare un barrel, in genere se ne osservano 8L’interno del barile è L interno del barile è completamente schermato dal solvete
Le eliche che connettono i filamenti sono all’esterno
Le interazioni eliche foglietti Le interazioni eliche-foglietti sono con residui idrofobiciI residui esterni delle eliche
id fili i 59sono idrofilici 59

ROSSMAN FOLD
Foglietto-beta parallelo apertoCi d d t d lfCirdondato da alfa-eliche da su entrambi i latii latiFormazione di una fessura nella porzione fessura nella porzione C-ter del foglietto-beta
In questa fessura spesso si ritrova il sito attivo dell’enzima
6060
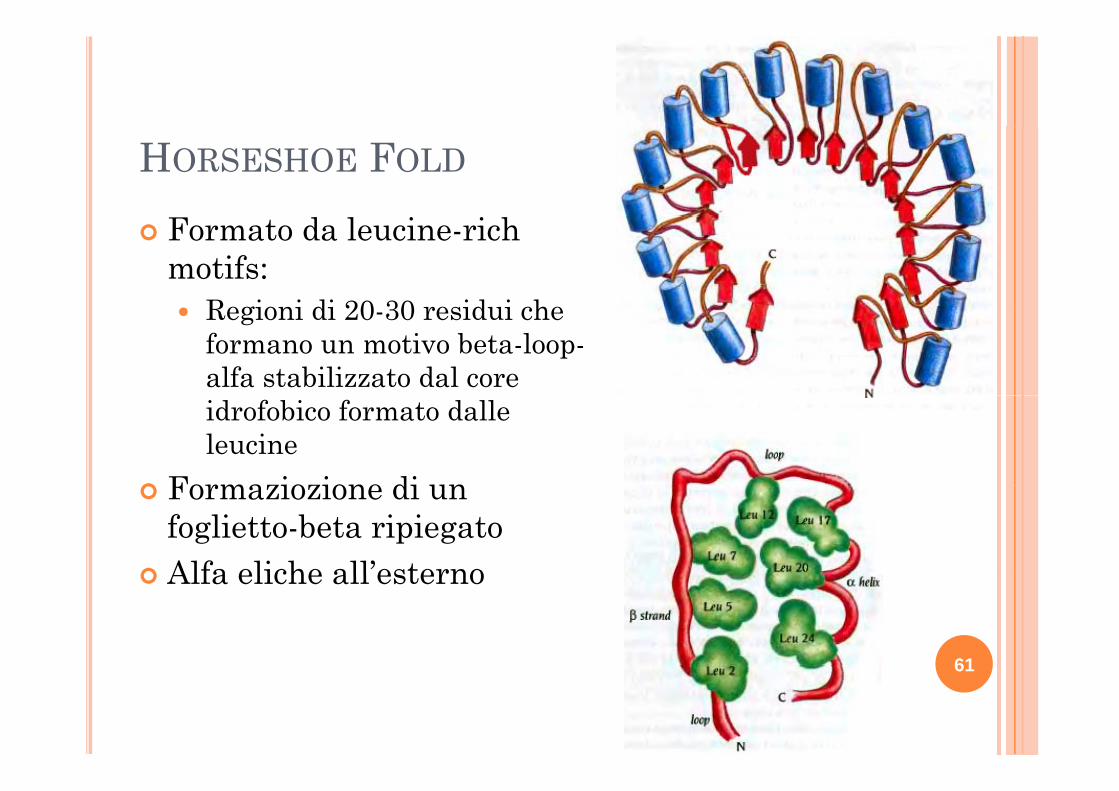
HORSESHOE FOLD
Formato da leucine-rich motifs:
R i i di 20 30 id i h Regioni di 20-30 residui che formano un motivo beta-loop-alfa stabilizzato dal core idrofobico formato dalle leucine
Formaziozione di un Formaziozione di un foglietto-beta ripiegatoAlfa eliche all’esternoAlfa eliche all esterno
6161

STRUTTURE A DOMINI BETA6262

DOMINI BETA
Formati da foglietti beta con un numero di filamenti variabile tra 4 e più di 10G l t i ti i d Generalmente sono arrangiati in modo antiparalleloSono formati da due foglietti disposti uno di Sono formati da due foglietti disposti uno di fronte all’altro che interagiscono tra loro con catene idrofobiche creando un core non catene idrofobiche creando un core non accessibile al solventeI foglietti sono ripiegati e insieme i due foglietti g p g gformano una struttura simile a un barileI residui esterni dei foglietti e il loop formano la
63zona che interagisce con il solvente 63

MOTIVI
Data la quantità di filamenti, il numero di topologie teoricamente possibile per i foglietti-beta antiparalleli è enormebeta antiparalleli è enormeTuttavia in natura se ne ritrova un numero molto più esiguopiù esiguo
Tre motivi principali:Tre motivi principali:Up-and-down barrelGreek keyGreek keyJerry roll barrel
6464

UP-AND-DOWN BARREL
Topologia semplice:Serie di filamenti antiparalleli connessi antiparalleli connessi da hairpin
Residui interni idrofobiciResidui esterni idrofilici
6565

RETINOL BINDING PROTEIN - HUMAN
6666

GREEK KEY
ÈÈ il motivo più comune trovato nei beta barrelPrima si forma un lungo filamento antiparallelo, che
i i i ll t tt fi lpoi si arrangia nella struttura finaleUn loop per ogni motivo attraversa il barrel
6767

GAMMA-CRYSTALLIN
6868
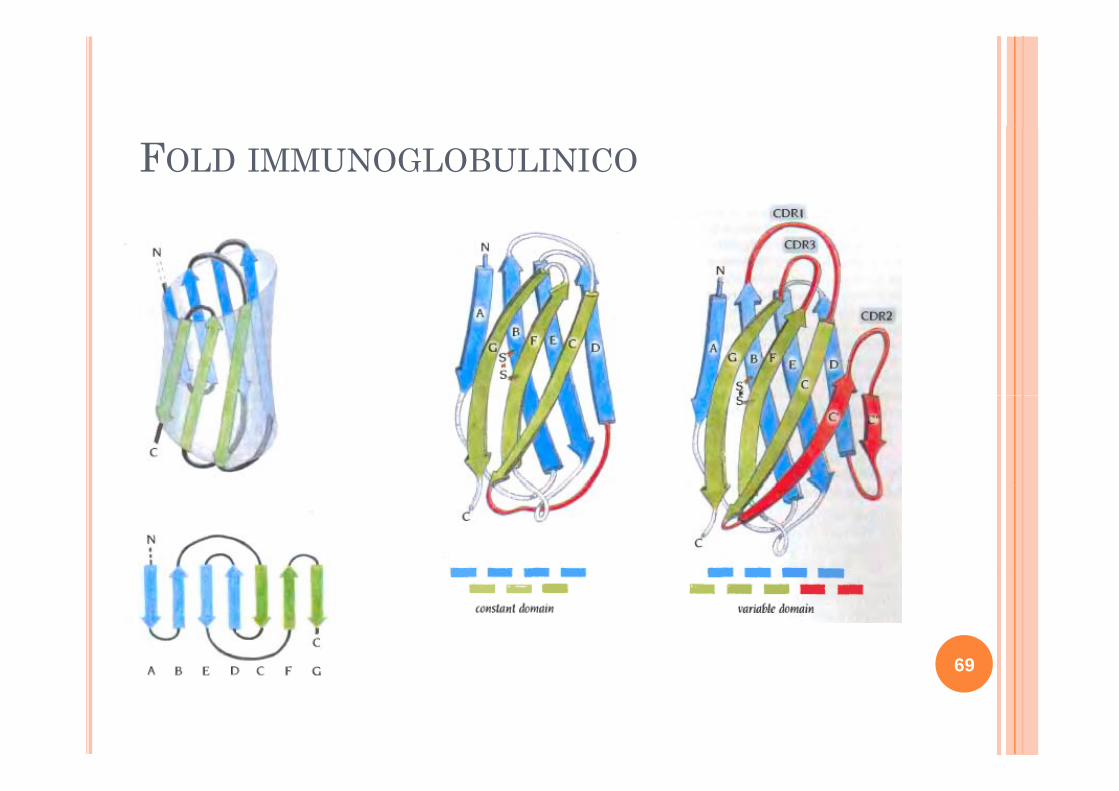
FOLD IMMUNOGLOBULINICO
6969

JELLY ROLL
Motivo molto comuneI loop attraversano il b l tt ltbarrel quattro volte:
2 sopra 2 sotto2 sotto
7070
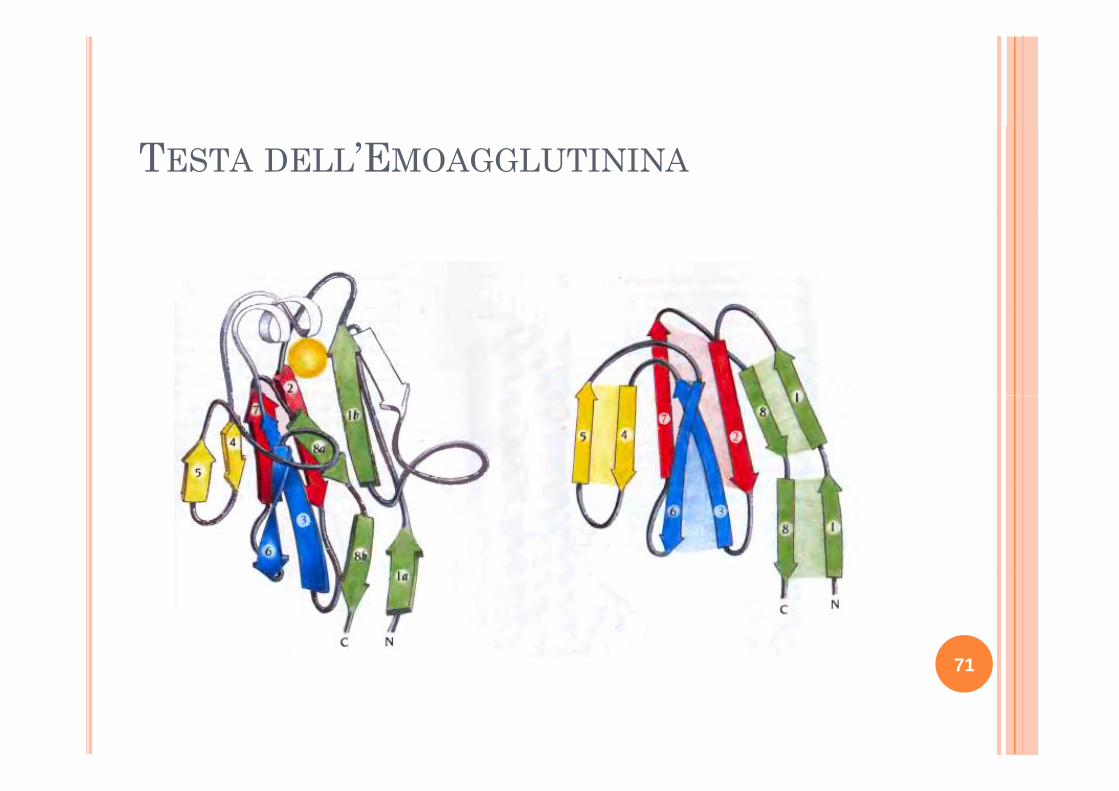
TESTA DELL’EMOAGGLUTININA
7171

PROTEINE TRANSMEMBRANA7272
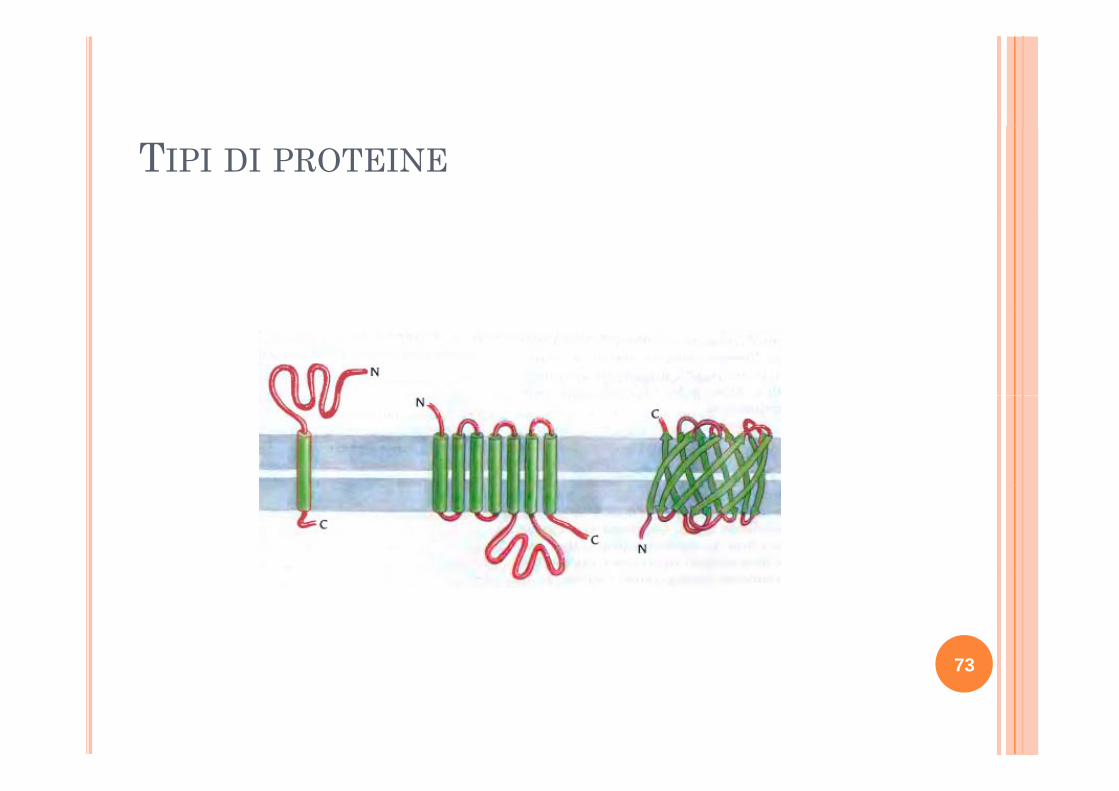
TIPI DI PROTEINE
7373

SEVEN MEMBRANE-SPANNING PROTEINS
Recettore associato a proteine G
7474

PORINE (ALL-BETA)
7575


















