Inhibitory Diffusible Factor 45 Bifunctional Activity - Journal of
6
THE JOURNAL OF BIOLOGICAL CHEMISTRY 0 1989 by The American Society for Biochemistry and Molecular Biology, Inc. Vol. 264. No. 21, Issue of July 25, pp. 12449-12454,1989 Printed in U. S. A. Inhibitory Diffusible Factor 45 Bifunctional Activity AS A CELL GROWTH INHIBITOR AND AS AN INSULIN-LIKE GROWTH FACTOR I-BINDING PROTEIN* (Received for publication, February 24, 1989) C. BlatS, J. DelbeS, J. Villaudyg, G. ChatelainS, A. Goldes, and L. HarelS From the Slnstitut de Recherches Scientifiques sur le Cancer, B.P. 8, 94802 Villejuif, Cedex, France and the SZnstitut Curie Pauillon Regaud, 75231 Paris, Cedex 05, France From medium conditioned by 3T3 cells, we had pre- viously purified to apparent homogeneity a novel in- hibitory diffusible factor of 45 kDa (IDFr6), and then determined the amino-terminal sequence. IDF46 pre- vented reversibly the growth of chick embryo fibro- blast (CEF). In these cells, DNA synthesis stimulated by 1% serum was 50% inhibited in the presenceof 45 ng/ml (1 nM) IDFIS. In the present article, we show that, in CEF, DNA synthesis stimulated by IGF-I was 100% inhibited in the presence of purified IDF46. Furthermore, the 45- kDa protein (IDFds) was, after Western blotting, able to bind IGF-I. The inhibitory effect of IDFrs upon serum stimula- tion did not seem to be the result of its inhibitory activity upon IGF-I stimulation, since stimulation by IGF-I and serum were additive. Moreover, it waspos- sibletodissociate thetwoinhibitoryeffects:when added to v-src transformed CEF, IDF4s was able to 100% inhibit stimulation induced by IGF-I and was unable to significantly decrease stimulation induced by serum, as was previously observed. Taken together, our results strongly suggest that IDFIS has two distinct functions, one of which was to bind IGF-I and the other to inhibit serum stimulation. Indeed, it was impossible to separate the two functions when IDFrswas purified by cation exchange fast protein liquid chromatogra- phy, a method very different from reverse-phase fast protein liquid chromatography previously used for pu- rification to apparent homogeneity of IDFr6. On the other hand, if the IGF binding activity and inhibitory activity effect upon serum stimulation were carried by two different proteins, the presence of IGF-I (in con- ditions where most of the 45-kDa proteins were bound to IGF-I) should not have affected the activity of the molecule inhibiting serum stimulation. However, we observed the contrary: when IDFr6 was bound to IGF- I, it lost its inhibitory effect upon stimulation induced by serum. This suggests that the two activities occurred on the same protein and that IDFIS is a bifunctional protein. Regulation of cell growth is probably under the control of positive and negative regulatory factors (1, 2). A number of proteins with growth inhibiting properties have been purified. Among these are the family of @-type transforming growth * This work was supported by I’Association pour la Recherche sur le Cancer andInstitutNational de la Sant6et de la Recherche MIdicale Contract 884011. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “aduertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. factors (3-6), interferons (7), tumor necrosis factors (8-ll), and interleukin (12). Furthermore, from medium conditioned by dense cultures of 3T3 cells, a 13-kDa inhibitory molecule (fibroblast growth regulator) was purified to apparent homo- geneity (13) and appears to be structurally related to a mam- mary-derived growth inhibitor (14-16). Some of us previously showed that 3T3 cells diffused inhib- itory molecules into medium (17). From medium conditioned by 3T3 cells we isolated an inhibitory diffusible factor (IDF45)’ of 45 kDa (18-20). More recently, we purified it to homoge- neity and the amino-terminal sequence was determined (21). In its chemical and biological properties, IDF45 differs from other inhibitory molecules known (21). It acts in the GI phase of the cell cycle and, contrary to transforming growth factor- @ (22), it inhibits early stimulation of RNA synthesis induced by serum (20). In chick embryo fibroblasts (CEF), DNA synthesis stimu- lated by 1% serum was 50% inhibited in the presence of45 ng/ml (1 nM) IDF45 (21). This inhibition was reversible. It is of interest that IDF45 was able to prevent CEF growth while little inhibitory activity was found when it was added to CEF transformed by the v-src oncogene (23). This agrees with the assumption that IDF45is involved in density-dependent in- hibition of growth observed in normal cells but not in trans- formed cells (24). The expression of oncogene v-SI%would allow the cells to escape this density-dependent growth inhi- bition. In order to understand the difference between the modes of action of IDF45 in normal and transformed CEF, we studied its effect upon stimulation of DNA synthesis induced by growth factors. We found that, in CEF, IGF-I was the only growth factor activity 100% inhibited by IDF45.2 In the present work, results suggest that the IDF45 has two distinct activities. The first would be to bindIGF-I and inhibitstimulation induced by this growth factor; the second would be to inhibit serum-induced DNA synthesis. MATERIALS AND METHODS Cek-Primary cultures of chick embryo fibroblasts were prepared from 10-day-old brown Leghorn chick embryos and were cultivated at 37 “C in modified Eagle’s medium supplemented with antibiotics and 5%calf serum as described (25). In some experiments CEF were infected as described (26) by a mutant of Rous sarcoma virus (NY68 strain) thermosensitive for the expression of the oncogene v-src. IDF4s Purification-1DFls was purified from medium conditioned by dense cultures of 3T3 cells as described (20, 21). Briefly, macromolecules from medium conditioned by dense cul- l The abbreviations used are: IDFd5, inhibitory diffusible factor 45; CEF, chick embryofibroblast;IGF-I,insulin-like growth factor-I; FPLC, fast protein liquid chromatography; SDS-PAGE, sodium do- decyl sulfate-polyacrylamide gel electrophoresis. J. Delbe, J. Villaudy, C. Blat, G. Desauty, A. GoldI, and L. Harel, manuscript in preparation. 12449
Transcript of Inhibitory Diffusible Factor 45 Bifunctional Activity - Journal of

THE JOURNAL OF BIOLOGICAL CHEMISTRY 0 1989 by The American Society for Biochemistry and Molecular Biology, Inc.
Vol. 264. No. 21, Issue of July 25, pp. 12449-12454,1989 Printed in U. S. A.
Inhibitory Diffusible Factor 45 Bifunctional Activity AS A CELL GROWTH INHIBITOR AND AS AN INSULIN-LIKE GROWTH FACTOR I-BINDING PROTEIN*
(Received for publication, February 24, 1989)
C. BlatS, J. DelbeS, J. Villaudyg, G. ChatelainS, A. Goldes, and L. HarelS From the Slnstitut de Recherches Scientifiques sur le Cancer, B.P. 8, 94802 Villejuif, Cedex, France and the SZnstitut Curie Pauillon Regaud, 75231 Paris, Cedex 05, France
From medium conditioned by 3T3 cells, we had pre- viously purified to apparent homogeneity a novel in- hibitory diffusible factor of 45 kDa (IDFr6), and then determined the amino-terminal sequence. IDF46 pre- vented reversibly the growth of chick embryo fibro- blast (CEF). In these cells, DNA synthesis stimulated by 1% serum was 50% inhibited in the presence of 45 ng/ml (1 nM) IDFIS.
In the present article, we show that, in CEF, DNA synthesis stimulated by IGF-I was 100% inhibited in the presence of purified IDF46. Furthermore, the 45- kDa protein (IDFds) was, after Western blotting, able to bind IGF-I.
The inhibitory effect of IDFrs upon serum stimula- tion did not seem to be the result of its inhibitory activity upon IGF-I stimulation, since stimulation by IGF-I and serum were additive. Moreover, it was pos- sible to dissociate the two inhibitory effects: when added to v-src transformed CEF, IDF4s was able to 100% inhibit stimulation induced by IGF-I and was unable to significantly decrease stimulation induced by serum, as was previously observed. Taken together, our results strongly suggest that IDFIS has two distinct functions, one of which was to bind IGF-I and the other to inhibit serum stimulation. Indeed, it was impossible to separate the two functions when IDFrs was purified by cation exchange fast protein liquid chromatogra- phy, a method very different from reverse-phase fast protein liquid chromatography previously used for pu- rification to apparent homogeneity of IDFr6. On the other hand, if the IGF binding activity and inhibitory activity effect upon serum stimulation were carried by two different proteins, the presence of IGF-I (in con- ditions where most of the 45-kDa proteins were bound to IGF-I) should not have affected the activity of the molecule inhibiting serum stimulation. However, we observed the contrary: when IDFr6 was bound to IGF- I, it lost its inhibitory effect upon stimulation induced by serum. This suggests that the two activities occurred on the same protein and that IDFIS is a bifunctional protein.
Regulation of cell growth is probably under the control of positive and negative regulatory factors (1, 2). A number of proteins with growth inhibiting properties have been purified. Among these are the family of @-type transforming growth
* This work was supported by I’Association pour la Recherche sur le Cancer and Institut National de la Sant6 et de la Recherche MIdicale Contract 884011. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “aduertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
factors (3-6), interferons (7), tumor necrosis factors (8-ll), and interleukin (12). Furthermore, from medium conditioned by dense cultures of 3T3 cells, a 13-kDa inhibitory molecule (fibroblast growth regulator) was purified to apparent homo- geneity (13) and appears to be structurally related to a mam- mary-derived growth inhibitor (14-16).
Some of us previously showed that 3T3 cells diffused inhib- itory molecules into medium (17). From medium conditioned by 3T3 cells we isolated an inhibitory diffusible factor (IDF45)’ of 45 kDa (18-20). More recently, we purified it to homoge- neity and the amino-terminal sequence was determined (21). In its chemical and biological properties, IDF45 differs from other inhibitory molecules known (21). It acts in the GI phase of the cell cycle and, contrary to transforming growth factor- @ (22), it inhibits early stimulation of RNA synthesis induced by serum (20).
In chick embryo fibroblasts (CEF), DNA synthesis stimu- lated by 1% serum was 50% inhibited in the presence of 45 ng/ml (1 nM) IDF45 (21). This inhibition was reversible. It is of interest that IDF45 was able to prevent CEF growth while little inhibitory activity was found when it was added to CEF transformed by the v-src oncogene (23). This agrees with the assumption that IDF45 is involved in density-dependent in- hibition of growth observed in normal cells but not in trans- formed cells (24). The expression of oncogene v-SI% would allow the cells to escape this density-dependent growth inhi- bition.
In order to understand the difference between the modes of action of IDF45 in normal and transformed CEF, we studied its effect upon stimulation of DNA synthesis induced by growth factors. We found that, in CEF, IGF-I was the only growth factor activity 100% inhibited by IDF45.2 In the present work, results suggest that the IDF45 has two distinct activities. The first would be to bind IGF-I and inhibit stimulation induced by this growth factor; the second would be to inhibit serum-induced DNA synthesis.
MATERIALS AND METHODS
Cek-Primary cultures of chick embryo fibroblasts were prepared from 10-day-old brown Leghorn chick embryos and were cultivated at 37 “C in modified Eagle’s medium supplemented with antibiotics and 5% calf serum as described (25). In some experiments CEF were infected as described (26) by a mutant of Rous sarcoma virus (NY68 strain) thermosensitive for the expression of the oncogene v-src.
IDF4s Purification-1DFls was purified from medium conditioned by dense cultures of 3T3 cells as described (20, 21).
Briefly, macromolecules from medium conditioned by dense cul-
l The abbreviations used are: IDFd5, inhibitory diffusible factor 45; CEF, chick embryo fibroblast; IGF-I, insulin-like growth factor-I; FPLC, fast protein liquid chromatography; SDS-PAGE, sodium do- decyl sulfate-polyacrylamide gel electrophoresis.
J. Delbe, J. Villaudy, C. Blat, G. Desauty, A. GoldI, and L. Harel, manuscript in preparation.
12449

12450 IL)F45 Bifunctional Activity tures of 3T3 cells were concentrated on a YMlO Amicon filter. The molecules, soluble in acetic acid, were fractionated by Bio-Gel P150 chromatography. Proteins of the most inhibitory fractions were dis- solved in 1 ml of 0.05% trifluoroacetic acid adjusted to 17% acetoni- trile and fractionated by two successive fast protein liquid chroma- tographies (FPLC) on a reverse-phase C8 ProRPC HR5/2 column (Pharmacia LKB Biotechnology Inc.).
In some experiments, IDF15 was purified as described above except that the last step of purification by FPLC on a reverse-phase column was replaced by puri~cation on a cation exchange column (Mono-S HR5/5 Pharmacia) in conditions described in the legend to Fig. 7.
Assay of IDF4B Actiuity-Secondary cultures of CEF were seeded at 8 X 10' cells/weIl in 36-well culture plates. Medium was discarded 6 h later and cells were maintained for 60 h in serum-free medium. Fractions to be tested were solubilized either in culture medium or acetic acid (0.5 M) plus afbumin (200 pg/ml) and added to quiescent CEF culture in the presence or absence of 1% serum. DNA synthesis was determined by labeling the cells with ["Clthymidine (0.07 pCi/ well) between 5 and 24 h after addition of the fraction to be tested and of serum, as described (27). Protein was determined from optical density a t 280 nm and Lowry's method. One unit of IDFas is the quantity which inhibits 50% of DNA synthesis when added to 1 ml in quiescent CEF culture stimulated by 1% serum. Purified IDF45 at a concentration of 1 nM (45 ng/ml) corresponds to I unit (21).
IDF45 was also tested on CEF stimulated by IGF-I (from Amersham Carp.) in conditions described above except that serum was replaced by IGF-I a t the concentration indicated in the legend.
When IDF4s was tested on cells infected by NY68 virus, infected cells (5 X lo4 cells/well) were seeded at 37 "C in Eagle's medium plus 4% serum. They were transferred to 41 "C 24 h later and maintained at this temperature for 3 days. The medium was replaced by serum- free medium (100 pl) 18 h before the experiment and cells maintained at 41 "C. Then, one part of the culture was transferred to 37 "C, 4 h later, IDF,, with or without 1% serum was added to the cultures
I 10 x ) 4s ab NG/M. I GF
asx SERUM a2 0.4
FIG. 1. Stimulation by IGF-I of DNA synthesis. IGF-I (A) or serum (A) was added at different concentrations to quiescent CEF, and DNA synthesis was determined as described under "Materials and Methods."
either maintained at 41 "C or t,ransferred to 37 "C. DNA synthesis was determined by labeling the cells between 5 and 24 h after addition of IDF,,. Data in each experiment were the mean of three determi- nations (S.D.).
SDS-PAGE-Lyophilized proteins were solubilized in a SDS sam- ple buffer (10% glycerol, 2.3% SDS, 0.0625 M Tris-HC1, pH 6.8). SDS-PAGE and silver staining were performed with a PhastSystem (Pharmacia LKB Biotechnology Inc.) following the manufacturer's instructions. The staining method was able to reveal between 1 and 5 ng, depending on the proteins analyzed when decreasing concentra- tions of proteins were separated by SDS-PAGE.
Western Blotting and Detection of IGF-binding Proteins-SDS- PAGE was performed on a Phastsystem (Pharmacia) with radioactive standard proteins purchased from Du Pont-New England Nuclear: lactoglobulin A, carbonic anhydrase, ovalbumin, albumin, phospho- rylase b, [methyl-'*C]rnethylated. After electrophoresis, proteins were transferred by passive diffusion on a nitrocellulose sheet according to the following procedure; the gel plus one sheet of nitrocellulose (0.2 pm, Schleicher and Schuell BA83) and two sheets of paper (Whatman 3") were soaked for 5 min in phosphate-buffer saline. On a glass slab were set successively the gel, the wet nitrocellulose sheet, the two wet sheets of paper, two dried sheets of paper (What- man 3"); and a glass slab. The apparatus was compressed by pressure of 40 g/cm*. After 48 h at room temperature, about 80% of the total proteins applied to the gel was recovered on the nitrocellulose membrane. Following this, nitrocellulose was dried at 37 "C for 15 min and processed essentially as described by Hossenlopp et al. (28) to detect IGF binding activity. '"I-IGF-I was purchased from Amer- sham.
RESULTS
~ t i m u ~ ~ i o n by IGF-I of DNA Synthesis in CEF-Different concentrations of IGF-I were added to quiescent CEF, and DNA synthesis was determined by labeling the cells between 5 and 24 h after addition of IGF-I (Fig. 1). Stimulation of DNA synthesis reached a plateau a t a concentration of 20 ng/ ml IGF-I. At this concentration, DNA synthesis was 130% stimulated.
In the same exper~ment, stimulation by serum was deter- mined. Fig. 1 shows that 0.8% serum induced greater stimu- lation (320%) of DNA synthesis than that induced by IGF-I.
Inhibition by IDF4s of IGF-I Stimulation-DNA synthesis was determined in CEF stimulated by 30 ng/ml IGF-I in the presence of different concentrations of purified IDF4, added at time 0 with IGF-I. In this experiment DNA synthesis was 140% stimulated by IGF-I. The inhibition induced by IDF45 was proportional to the IDF4s concentration and reached 100% at a concentration of 2 units of IDF45 (Fig. 2). A t this concentration, IDF4:, inhibited the stimulation induced by 1% serum by only 66% (results not shown).
In the experiment shown in Fig. 3, 1 or 1.5 units of IDF~E, were added to cells in the presence of different concentrations of IGF-I. Stimulation of DNA synthesis induced by IGF-I was
FIGS. 2 (left) and 3 (right). Inhibition by IDFd6 of IGF-I stimulation. Fig. 2: IGF-I (30 ng/ml) was added to quiescent CEF in the absence or presence of dif- ferent concentrations of purified IDF45. In this experiment, DNA synthesis was 140% stimulated in the presence of 30 ng/ml IGF. The inhibitory activity was determined by the percent inhibition of the increase by IGF-I of ["Clthymidine inco~oration in the cells. DNA synthe- sis was 100% inhibited in the presence of IDFIs when incorporation of [14C]thy- midine in stimulated cells and control nonstimulated cells was the same. Fig. 3: in this experiment, no IDF45 (O), 1 unit (A), or 1.5 units (A) of IDF,, were added to quiescent CEF with different concen- trations of IGF-I.
CONCENTRATION OF I G F

12452 IDF45 Bifunctional Activity
F R A C T I O N NUMDER
B C
94- 67-
43- - 30-
20 -
0 - I D F m+IDi
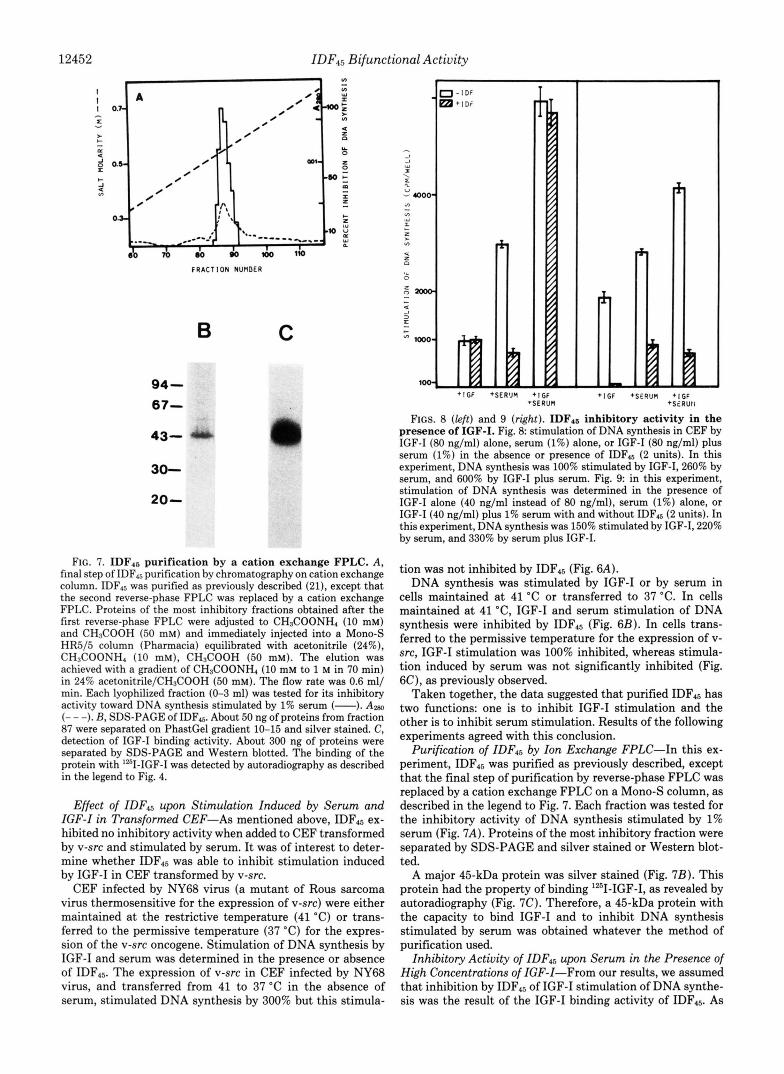
I, I FIGS. 8 (left) and 9 (right). IDFls inhibitory activity in the
presence of IGF-I. Fig. 8 stimulation of DNA synthesis in CEF by IGF-I (80 ng/ml) alone, serum (1%) alone, or IGF-I (80 ng/ml) plus serum (1%) in the absence or presence of IDF45 (2 units). In this experiment, DNA synthesis was 100% stimulated by IGF-I, 260% by serum, and 600% by IGF-I plus serum. Fig. 9: in this experiment, stimulation of DNA synthesis was determined in the presence of IGF-I alone (40 ng/ml instead of 80 ng/ml), serum (1%) alone, or IGF-I (40 ng/ml) plus 1% serum with and without IDF4s (2 units). In this experiment, DNA synthesis was 150% stimulated by IGF-I, 220% by serum, and 330% by serum plus IGF-I.
FIG. 7. IDF4. purification by a cation exchange FPLC. A , final step of IDFt5 purification by chromatography on cation exchange column. IDF15 was purified as previously described (21), except that the second reverse-phase FPLC was replaced by a cation exchange FPLC. Proteins of the most inhibitory fractions obtained after the first reverse-phase FPLC were adjusted to CH3COONH4 (10 mM) and CH3COOH (50 mM) and immediately injected into a Mono-S HR5/5 column (Pharmacia) equilibrated with acetonitrile (24%), CH3COONH4 (10 mM), CH3COOH (50 mM). The elution was achieved with a gradient of CH3COONH4 (10 mM to 1 M in 70 min) in 24% acetonitrile/CH3COOH (50 mM). The flow rate was 0.6 ml/ min. Each lyophilized fraction (0-3 ml) was tested for its inhibitory activity toward DNA synthesis stimulated by 1% serum (-). A m (- - -). B, SDS-PAGE of IDF45. About 50 ng of proteins from fraction 87 were separated on PhastGel gradient 10-15 and silver stained. C, detection of IGF-I binding activity. About 300 ng of proteins were separated by SDS-PAGE and Western blotted. The binding of the protein with '*'I-IGF-I was detected by autoradiography as described in the legend to Fig. 4.
Effect of IDF45 upon Stimulation Induced by Serum and IGF-I in Transformed CEF-As mentioned above, IDF45 ex- hibited no inhibitory activity when added to CEF transformed by v-src and stimulated by serum. It was of interest to deter- mine whether IDF45 was able to inhibit stimulation induced by IGF-I in CEF transformed by v-src.
CEF infected by NY68 virus (a mutant of Rous sarcoma virus thermosensitive for the expression of v-src) were either maintained at the restrictive temperature (41 "C) or trans- ferred to the permissive temperature (37 "C) for the expres- sion of the v-src oncogene. Stimulation of DNA synthesis by IGF-I and serum was determined in the presence or absence of IDF45. The expression of v-src in CEF infected by NY68 virus, and transferred from 41 to 37 "C in the absence of serum, stimulated DNA synthesis by 300% but this stimula-
tion was not inhibited by IDF45 (Fig. 6A). DNA synthesis was stimulated by IGF-I or by serum in
cells maintained at 41 'C or transferred to 37 "C. In cells maintained a t 41 "C, IGF-I and serum stimulation of DNA synthesis were inhibited by IDR5 (Fig. 6B). In cells trans- ferred to the permissive temperature for the expression of v- src, IGF-I stimulation was 100% inhibited, whereas stimula- tion induced by serum was not significantly inhibited (Fig. 6C), as previously observed.
Taken together, the data suggested that purified IDF4s has two functions: one is to inhibit IGF-I stimulation and the other is to inhibit serum stimulation. Results of the following experiments agreed with this conclusion.
Purification of IDF45 by Ion Exchange FPLC-In this ex- periment, IDF45 was purified as previously described, except that the final step of purification by reverse-phase FPLC was replaced by a cation exchange FPLC on a Mono-S column, as described in the legend to Fig. 7. Each fraction was tested for the inhibitory activity of DNA synthesis stimulated by 1% serum (Fig. 7A). Proteins of the most inhibitory fraction were separated by SDS-PAGE and silver stained or Western blot- ted.
A major 45-kDa protein was silver stained (Fig. 7B). This protein had the property of binding lZ5I-IGF-I, as revealed by autoradiography (Fig. 7C). Therefore, a 45-kDa protein with the capacity to bind IGF-I and to inhibit DNA synthesis stimulated by serum was obtained whatever the method of purification used.
Inhibitory Activity of IDF45 upon Serum in the Presence of High Concentrations of IGF-I-From our results, we assumed that inhibition by IDF45 of IGF-I stimulation of DNA synthe- sis was the result of the IGF-I binding activity of IDF46. As

IDF45 Bifunctional Activity 12453
shown in Fig. 3 in the presence of 80 ng/ml of IGF-I, most of the IDF45 molecules (likely bound to IGF-I) were unable to inhibit stimulation induced by the free IGF-I molecules. Therefore, it was of interest to verify whether IDF45 bound to IGF-I was able to inhibit serum stimulation.
In the experiment in Fig. 8, cells were stimulated by serum (1%) alone, IGF-I (80 ng/ml) alone, or by the mixture of IGF- I and serum in the presence or absence of IDF45.
Stimulation of DNA synthesis by IGF-I was not inhibited. This result agrees with that shown in Fig. 3. DNA synthesis stimulated by serum alone was 73% inhibited; however, when serum and IGF-I were added simultaneously. DNA synthesis was not significantly inhibited (Fig. 8). The latter result clearly shows that IDF45 was unable to inhibit serum-stimu- lated DNA synthesis in the presence of a large concentration of IGF-I. In another experiment (Fig. 9), IGF-I was added at a concentration of 40 ng/ml instead of 80 ng/ml. Stimulation of DNA synthesis by IGF-I was inhibited by IDF45, like stimulation induced by serum. In this experiment, stimulation induced by IGF-I plus serum was also clearly inhibited in the presence of IDF45. Therefore, the inhibitory activity of IDF45 upon serum stimulation depended upon the IGF-I concentra- tion present in medium.
DISCUSSION
Our experiments demonstrate that, in CEF, IGF-I was able to stimulate DNA synthesis; this stimulation was 100% inhib- ited in the presence of IDF45. Furthermore, purified IDF45, which after SDS-PAGE, showed only one protein band of 45 kDa, was able to bind IGF-I. Inhibition by IDF45 of DNA synthesis stimulated by IGF-I is probably the result of the IDF45 ability to bind IGF-I.
The inhibitory effect of IDFd5 upon serum stimulation did not seem to result from its inhibitory effect upon IGF-I stimulation. Stimulation of DNA synthesis by IGF-I and by serum was additive; indeed in some experiments, we observed a synergy between the two effects. This suggests that the mode of action of IGF-I and serum are different, and that stimulation by serum is not due to cell stimulation by IGF-I present in the serum. Moreover, it was possible to dissociate the two inhibitory effects of IDF45 on IGF-I and serum stim- ulation. IDFIs inhibited the stimulation induced by serum or IGF-I when added to NY68-infected CEF, maintained at the restrictive temperature for the expression of v-src, whereas IDF45, when added to v-src-transformed CEF, was able to inhibit stimulation induced by IGF-I but was unable to sig- nificantly decrease serum-induced stimulation. These results agree with our previous ones (23) and also with the assump- tion that inhibition by IDF45 of IGF-I stimulation is due to binding of IGF-I to IDF45. Indeed, in this case, IDF45 would bind IGF-I in the culture medium and would prevent IGF-I stimulation whatever the cell type present in the culture.
Taken together, our results suggest that IDFh5 had two different functions, one of which was to inhibit serum stimu- lation and the other to bind IGF-I.
Two other experiments agree with this assumption. First, it was impossible to separate the two activities when they were purified by cation exchange FPLC, a method very dif- ferent from reverse-phase FPLC previously used for purifi- cation to apparent homogeneity of IDF15 (21). We also showed that IDF45 when bound to IGF-I (in the presence of a large concentration of growth factor), was unable to inhibit stimu- lation of DNA synthesis induced by serum. This result pro- vides an important argument in favor of our hypothesis. Indeed, as shown in Fig. 3, the inhibitory activity of IDF45 upon IGF-I stimulation depends upon the IGF concentration.
One unit of IDF45 (about 1 nM) mainly inhibited stimulation induced by 2 nM IGF-I. When the concentration of IGF-1 increased and was higher than 2 nM, inhibition decreased probably because IDF15 bound to IGF-I was unable to inhibit added free IGF-I. If the inhibitory effect upon serum stimu- lation and IGF binding activity was carried on two different proteins, the presence of IGF-I, under conditions in which most of the 45-kDa proteins bound to IGF-I, should have no effect upon the activity of the molecule which inhibits stim- ulation induced by serum. We observed the contrary: when IDF45 bound to IGF-I, it lost its inhibitory effect on stimula- tion induced by serum. These results thus strongly suggest that the two activities took place on the same protein.
It should be noted that other bifunctional proteins are known (29). One example is transforming growth factor-p, which has both inhibitory and stimulatory activities (3-6). Another is the IGF-I1 receptor, which has the property not only of binding IGF-I1 but also of binding and transporting mannose 6-phosphate (30,31).
The bifunctional activity of IDF45 could have important repercussions in terms of precise regulation of cell prolifera- tion i n vitro and i n uiuo. Regulation would depend not only on the IGF-I or IDF45 concentration, but also, and more importantly, on the concentration ratio of the two molecules. For example, i n uiuo, the increase by growth hormone of IGF- I secretion might stimulate cell proliferation by preventing the inhibitory effect of an IDF15-like molecule. One might also question to what extent the increase in IGF-I secretion by transformed cells, as shown in mammary cancer cells (32,33) could stimulate cell proliferation by binding IDF45-like protein and preventing its inhibitory activity. However, the results described herein indicate that this type of mechanism does not enable v-src-transformed cells to escape inhibition by IDF45. Indeed, if the loss of IDF45 activity was the result of increased secretion of IGF-I, then stimulation induced by IGF-I should not be inhibited by IDF45 as was the case for stimulation induced by serum.
Different IGF-binding proteins are present in serum (34- 36). They form large complexes (of about 150 kDa) which are under growth-hormone control and small complexes (of about 40 kDa). After SDS-PAGE different molecular forms were observed from 41.5 to 24 kDa (37). IGF-binding proteins of 41.5 to 24 kDa are also secreted by cells in culture (38-42). While IGF-binding proteins secreted by rat liver cells were shown to inhibit the stimulatory effect of IGF on fibroblast DNA synthesis (43), an IGF-binding protein purified from amniotic fluid markedly potentiated replication of several cell types in response to human IGF-I (44).
Some of these IGF-binding proteins have been recently purified and characterized (38, 39, 45, 46). The amino-termi- nal sequences of 30,000-34,000 IGF-binding proteins of BRL3A rat cell line (39) and of human amniotic fluid (45-46) are different than the IDF45 amino-terminal sequence (21).
A recent article (which appeared after our experiments were concluded) revealed, in rat serum, a 45-kDa IGF-binding protein whose amino-terminal sequence resembles that of IDF45 (47). We do not yet know the extent of the homology. This protein is glycosylated (47), while IDF45 is not. Moreover, no inhibiting activity was shown for this molecule.
In conclusion, our results strongly suggest that IDF45 is bifunctional, and is able to inhibit serum stimulation and to bind IGF-I. This is the first time that this bifunctional activity of an IGF-I-binding protein has been pointed out.
Acknowledgments-Special thanks go to Drs. M. Binoux and P. Hossenlopp for helpful advice concerning the IGF binding rnethod-

12454 IDF,, Bifunctional Activity ology and Dr. J. Escribano for advice concerning the protein transfer on nitrocellulose.
REFERENCES
2. 3
4.
5.
6.
7.
8.
9.
10.
11.
12. 13. 14.
15.
16.
17.
18.
19.
20.
21.
22.
23.
1. Harel, L. (1981) Growth Factors Handbook of Experimental Phar- macology (Baserga, R., ed) Vol. 57, pp. 313-341, Springer- Verlag, Heidelberg
. Sporn, M. B., and Roberts, A. (1985) Nature 313, 745-747
. Holley, R. W., Bohlen, P., Fava, R., Baldwin, J. H., Kleeman, G., and Armour, R. (1980) Proc. Natl. Acad. Sci. U. S. A . 77,5989- C n n "
Tucker, R. F., Shipley, G. D., Moses, H. L., and Holley, R. W. (1984) Science 2 2 6 , 705-707
Roberts, A. B., Anzano, M. A., Wakefield, L. M., Roche, N. S., Stern, D. F., and Sporn, M. B. (1985) Proc. Natl. Acad. Sci. U. S. A. 82, 119-123
Hanks, S. K., Armour, R., Baldwin, J. H., Maldonado, F., Spiess, J., and Holley, R. W. (1988) Proc. Natl. Acad. Sci. U. S. A. 85, 79-82
Gresser, I., and Tovey, M. G. (1978) Biochem. Biophys. Acta 516,
Niitsu, Y., Watanabe, N., Sone, H. N., Neda, H., Yamauchi, N., and Urushizaki, I. (1985) Jpn. J. Cancer Res. 76, 1193-1197
Sugarman, B. J., Aggarwal, B. B., Hass, P. E., Figari, J. S., Palladino, M. A., Jr., and Shepard, H. M. (1985) Science 230, 943-945
Stolphen, A. H., Guinan, E. C., Fiers, W., and Pober, J. S. (1986) Am. J . Pathol. 123, 16-24
Sato, N., Goto, T., Haranaka, K., Satomi, N., Nariuchi, N., Mano- Hirano, Y., and Sawasaki, Y. (1986) J. Natl . Cancer Inst. 7 6 ,
Tsai, S. C., and Gaffney, E. V. (1986) Cancer Res. 4 6 , 1471-1477 Hsu, Y. M., and Wang, J. L. (1986) J. Cell Biol. 1 0 2 , 362-368 Bohmer, F. D., Kraft, R., Otto, A,, Wernstedt, C., Hellman, U.,
Kurtz, A., Muller, Th., Rhode, K., Etzold, G., Lehmann, W., Langen, P., Heldin, C.-H., and Grosse, R. (1987) J. Biol. Chem.
Hsu, Y. M., Barry, J. M., and Wang, J. L. (1984) Proc. Natl.
Bohmer, F. D., Sun, Q., Pepperle, M., Muller, Th., Erikson, U., Wang, J. L., and Grosse, R. (1987) Biochem. Biophys. Res. Commun. 148, 1425-1431
Harel, L., Jullien, M., and De Monti, M. (1978) J. Cell. Physiol.
Harel, L., Chatelain, G., and Golde, A. (1984) J . Cell. Physiol.
Harel, L., Blat, C., and Chatelain, G. (1985) J . Cell. Physiol. 123,
Blat, C., Chatelain, G., Desauty, G., and Harel, L. (1986) FEBS
Blat, C., Bohlen, P., Villaudy, J., Chatelain, G., GoldL, A., and
Chambard, J. C., and Pouyssigur, J. (1988) J . CeU. Physiol. 135,
Blat, C., Villaudy, J., Rouillard, D., Gold6, A., and Harel, L.
DYYL
231-247
1113-1121
262,15137-15143
A c d . Sci. U. S. A. 81, 2107-2111
96,327-332
119,101-106
139-143
Lett. 203, 175-180
Harel, L. (1989) J. Biol. Chem. 2 6 4 , 6021-6024
101-107
24.
25.
26.
27. 28.
29.
(1987) J. Cell. Physiol. 130, 416-419
(1984) Exp. Cell. Res. 152, 390-401
Res. 134, 121-128
Cell. Biol. Inter. Rep. 1 0 , 947-954
Jullien, M., Harel, L., Gold&, A., Villaudy, J., and Pugnet, P.
Blat, C., Harel, L., Villaudy, J., and Golde, A (1981) Exp. Cell.
Blat, C., Villaudy, J., Desauty, G., GoldL, A., and Harel, L. (1986)
Harel, L., Blat, C., and Chatelain, G. (1983) Biol. Cell. 4 8 , 11-16 Hossenlopp, P., Seurin, D., Segovia-Quinson, B., Hardouin, S.,
and Binoux, M. (1986) Anal. Biochem. 1 5 4 , 138-143 Sporn. M.. and Roberts. A. (1988) Nature 332. 217-219
30. MacDonald, R. G., Pfeffer,'S. R:, Coussens, L., Tepper, M. A., Brocklebank, C. M., Mole, J. E., Anderson, J. K., Chen, E., Czech, M. P., and Ullrich, A. (1988) Science 239, 1134-1137
31. Morgan, D. O., Edman, J . C., Standring, D. N., Fried, V. A., Smith, M. C., Roth, R. A., and Rutter, W. J. (1987) Nature
32. Huff, K. K., Kaufman, D., Gabbay, K. H., Spencer, E. M., Lippman, M. E., and Dikson, R. B. (1986) Cancer Res. 46,
33. Cariani, E., Lasserre, C. , Seurin, D., Hamelin, B., Kemeny, F., Franco, D., Czech, M. P., Ullrich, A., and Brechot, C. (1988) Cancer Res. 48,6844-6849
34. Zapf, J., Froesch, E. R., and Humbel, R. E. (1981) Curr. Top. Cell. Regul. 1 9 , 257-309
35. Moses, A. C., Nissley, S. P., Cohen, K. L., and Rechler, M. M. (1976) Nature 263, 137-140
36. Nissley, S. P., and Rechler, M. M. (1984) in Hormonal Proteins and Peptides (Li, C. H., ed) Vol. 12, pp. 127-203, Academic Press, New York
37. Hardouin, S., Hossenlopp, P., Segovia, B., Seurin, D., Portolan, G., Lassarre, C., and Binoux, M. (1987) Eur. J . Biochem. 1 7 0 ,
38. Lyons, R. M., and Smith, G. L. (1986) Mol. Cell. Endocr. 4 5 ,
39. Mottola, C., MacDonald, R. G., Brackett, J. L., Mole, J. E., Anderson, J. K., and Czech, M. (1986) J. Biol. Chem. 261, 11180-11188
40. Romanus, J. A., Yang, Y. W. H., Nissley, S. P., and Rechler, M. M. (1987) Endocrinology 1 2 1 , 1041-1050
41. Hossenlopp, P., Seurin, D., Segovia, B., Portolan, G., and Binoux, M. (1987) Eur. J. Biochem. 170, 133-142
42. Clemmons, D. R., Elgin, R. G., Han, V. K. M., Casella, S. J., D'Ercole, A. J., and Van Wyk, J. J. (1986) J. Clin. Invest. 77, 1548-1553
43. Knauer, D. J., and Smith, G. L. (1980) Proc. Natl. Acad. Sci.
44. Elgin, R. G., Busby, W. H., Jr., and Clemmons, D. R. (1987) Proc. Natl. Acad. Sci. U. S. A. 84, 3254-3258
45. Brinkman, A,, Groffen, C., Kortleve, D. J., Geurts van Kessel, A., and Drop, S. L. S. (1988) EMBO J. 7, 2417-2423
46. Brewer, M. T., Stetler, G. L., Squires, C. H., Thompson, R. C., Busby, W. H., and Clemmons, D. R. (1988) Biochern. Biophys. Res. Commun. 152,1289-1297
47. Zapf, J., Born, W., Chang, J. Y., James, P., Froesch, E. R., and Fischer, J. A,, (1988) Biochem. Biophys. Res. Commun. 156, 1187-1194
329,301-307
4613-4619
121-132
263-270
U. S. A. 77,7252-7254