
Inhibition of Wnt/b-catenin signaling mediates ursolic ...€¦ · Inhibition of Wnt/b-catenin...
9
Inhibition of Wnt/b-catenin signaling mediates ursolic acid-induced apoptosis in PC-3 prostate cancer cells Ji-Hyuk Park 1+ , Hee-Young Kwon 1+ , Eun Jung Sohn 1+ , Kyung A Kim 2 , Bonglee Kim 1 , Soo-Jin Jeong 1 , Jun ho Song 1 , Jin Suk Koo 3 , Sung-Hoon Kim 1 College of Oriental Medicine, Kyung Hee University, Seoul 130-701, South Korea Department of Orthodontics, Kyung Hee University School of Dentistry, South Korea College of Natural Sciences, Andong National University, Andong, 760-749, South Korea Correspondence: Sung-Hoon Kim, e-mail: [email protected] Abstract: Background: Ursolic acid, a pentacyclic triterpenoid, is known to exert antitumor activity in breast, lung, liver and colon cancers. Nonetheless, the underlying mechanism of ursolic acid in prostate cancer cells still remains unclear. To investigate the antitumor mechanism, the apoptotic mechanism of ursolic acid via Wnt/b-catenin signaling was examined in PC-3 prostate cancer cells. Methods: Cytotoxicity assay, flow cytometry, immunofluorescence assay and western blotting were performed. Results: Ursolic acid showed cytotoxicity against PC-3, LNCaP and DU145 prostate cancer cells with IC of 35 μM, 47 μM and 80 μM, respectively. Also, ursolic acid significantly increased the number of ethidium homodimer stained cells and apoptotic bod- ies, and dose-dependently enhanced the sub-G1 apoptotic accumulation in PC-3 cells. Consistently, western blotting revealed that ursolic acid effectively cleaved poly (ADP-ribose) polymerase (PARP), activated caspase-9 and -3, suppressed the expression of sur- vival proteins such as Bcl-X , Bcl-2 and Mcl-1, and upregulated the expression of Bax in PC-3 cells. Interestingly, ursolic acid sup- pressed the expression of Wnt5a/b and b-catenin, and enhanced the phosphorylation of glycogen synthase kinase 3 b (GSK3b). Furthermore, the GSK3b inhibitor SB216763 or Wnt3a-conditioned medium (Wnt3a-CM) reversed the cleavages of caspase-3 and PARP induced by ursolic acid in PC-3 cells. Conclusions: Our findings suggest that ursolic acid induces apoptosis via inhibition of the Wnt5/b-catenin pathway and activation of caspase in PC-3 prostate cancer cells. These results support scientific evidence that medicinal plants containing ursolic acid can be applied to cancer prevention and treatment as a complement and alternative medicine (CAM) agent. Key words: ursolic acid, apoptosis, caspase, Wnt/GSK3b/b-catenin signaling 1366 Contributed equally to this work
Transcript of Inhibition of Wnt/b-catenin signaling mediates ursolic ...€¦ · Inhibition of Wnt/b-catenin...

Inhibition of Wnt/b-catenin signaling mediates
ursolic acid-induced apoptosis in PC-3 prostate
cancer cells
Ji-Hyuk Park1+, Hee-Young Kwon1+, Eun Jung Sohn1+, Kyung A Kim2,
Bonglee Kim1, Soo-Jin Jeong1, Jun ho Song1, Jin Suk Koo3, Sung-Hoon Kim1
1College of Oriental Medicine, Kyung Hee University, Seoul 130-701, South Korea
2Department of Orthodontics, Kyung Hee University School of Dentistry, South Korea
3College of Natural Sciences, Andong National University, Andong, 760-749, South Korea
Correspondence: Sung-Hoon Kim, e-mail: [email protected]
Abstract:
Background: Ursolic acid, a pentacyclic triterpenoid, is known to exert antitumor activity in breast, lung, liver and colon cancers.
Nonetheless, the underlying mechanism of ursolic acid in prostate cancer cells still remains unclear. To investigate the antitumor
mechanism, the apoptotic mechanism of ursolic acid via Wnt/b-catenin signaling was examined in PC-3 prostate cancer cells.
Methods: Cytotoxicity assay, flow cytometry, immunofluorescence assay and western blotting were performed.
Results: Ursolic acid showed cytotoxicity against PC-3, LNCaP and DU145 prostate cancer cells with IC50 of 35 µM, 47 µM and
80 µM, respectively. Also, ursolic acid significantly increased the number of ethidium homodimer stained cells and apoptotic bod-
ies, and dose-dependently enhanced the sub-G1 apoptotic accumulation in PC-3 cells. Consistently, western blotting revealed that
ursolic acid effectively cleaved poly (ADP-ribose) polymerase (PARP), activated caspase-9 and -3, suppressed the expression of sur-
vival proteins such as Bcl-XL, Bcl-2 and Mcl-1, and upregulated the expression of Bax in PC-3 cells. Interestingly, ursolic acid sup-
pressed the expression of Wnt5a/b and b-catenin, and enhanced the phosphorylation of glycogen synthase kinase 3 b (GSK3b).
Furthermore, the GSK3b inhibitor SB216763 or Wnt3a-conditioned medium (Wnt3a-CM) reversed the cleavages of caspase-3 and
PARP induced by ursolic acid in PC-3 cells.
Conclusions: Our findings suggest that ursolic acid induces apoptosis via inhibition of the Wnt5/b-catenin pathway and activation
of caspase in PC-3 prostate cancer cells. These results support scientific evidence that medicinal plants containing ursolic acid can be
applied to cancer prevention and treatment as a complement and alternative medicine (CAM) agent.
Key words:
ursolic acid, apoptosis, caspase, Wnt/GSK3b/b-catenin signaling
1366 Pharmacological Reports, 2013, 65, 1366�1374
Pharmacological Reports2013, 65, 1366�1374ISSN 1734-1140
Copyright © 2013by Institute of PharmacologyPolish Academy of Sciences
+Contributed equally to this work

Introduction
Prostate cancer is internationally the second most
common cancer diagnosed in men and the sixth most
common cause of cancer death among men. Accord-
ing to the NCI report, the incidence rate of prostate
cancer is approximately 16.22% in men born today [2,
6]. Recent clinical practice shows that medicinal
herbs with antitumor activities have chemopreventive
potential with few side effects [4, 35].
Ursolic acid as a pentacyclic triterpene acid is
mainly contained in several fruits such as hawthorn,
prunes, peppermint, apples, rosemary, lavender, and
also medicinal plants including Rosmarinus officina-
lis, Oldenlandia diffusa, Eriobotrya japonica, and
Glechoma hederaceae [31]. As a whole, ursolic acid
has been suggested as a non-toxic chemopreven-
tive/chemoprotective agent in clinical practice [20],
even though there are some evidences that ursolic
acid causes the DNA damages in human endothelial
cells [18] and reduces the sperm motility [3].
Previous evidence shows that ursolic acid induces
apoptosis in several cancers such as colon cancer [1],
breast cancer [11], leukemia [15], melanoma [12] and
prostate cancer [24]. Several papers studied the effect
of ursolic acid in vivo models. Ursolic acid signifi-
cantly inhibited tumor growth in transgenic mouse
prostate model [24] and prostate cancer xenograft in
nude mice [23]. Although ursolic acid exerts antitu-
mor activity via inhibition of CXCR4/CXCL12 [23],
matrix metalloproteinase 2 (MMP2) [12], P2Y2/Src/
p38/COX-2 [13] and the AKT pathway [37] as well as
autophagy [25], the antitumor mechanism of ursolic
acid still remains unclear in prostate cancers. Thus, in
the present study, the role of Wnt/b-catenin signaling
was elucidated in ursolic acid-induced apoptosis in
PC-3 prostate cancer cells using XTT assay, cell cycle
analysis, and western blotting.
Materials and Methods
Cell culture
PC-3, DU145, LNCaP prostate cancer cells, Raw
264.7 (leukemic monocyte macrophage cells) and
HEK293 (human embryonic kidney 293) cells were
obtained from American Type Culture Collection
(ATCC). HEK293, PC-3 and DU145 cells were main-
tained in Roswell Park Memorial Institute (RPMI)
1640 supplemented with 10% fetal bovine serum
(FBS) (Gibco, Carlsbad, CA, USA) and 1% antibiot-
ics at 37°C in a humidified atmosphere containing 5%
CO2. LNCaP cells were cultured in RPMI 1640 sup-
plemented with 10% FBS and 2 µmol/l L-glutamine,
10 µmol/l HEPES, 1 µmol/l sodium pyruvate, and
4.5% D-glucose without antibiotics at 37°C in a hu-
midified atmosphere containing 5% CO2. Wnt3a CM
was prepared as previously described [21].
Cytotoxicity assay
To investigate the cytotoxicity, 3-[4,5-dimethylthia-
zol-2-yl]-2,5-diphenyltetrazolium (MTT) bromide col-
orimetric assay was used. Prostate cancer cells were
plated at 104 cells in 96-well plates and incubated over-
night. Cells were treated with various concentrations of
ursolic acid. After 24-h incubation, 100 ml of MTT
(Sigma Chemical Co., St. Louis, MO, USA) was added
to each well and incubated for 4 h at 37°C. Formazan
crystals were dissolved by addition of 100 ml DMSO
solution. The microplate reader at 570 nM was used to
determine the absorbance of each well.
Ethidium homodimer assay
To measure cell death, we used the DAPI and ethid-
ium homodimer dye following the manufactures’ in-
structions (Molecular Probes). In brief, PC-3 cells
were treated with 30 µM ursolic acid for 24 h. After
incubation, cells were fixed in 4% methanol-free for-
maldehyde solution and stained with the 5 µM ethid-
ium homodimer and then incubated at 37°C for
30 min in the dark. Then, the cells were mounted with
mounting medium containing DAPI and visualized
under an Axio vision 4.0 fluorescence microscope
(Carl Zeiss Inc., Weimar, Germany).
Cell cycle analysis
PC3 cells treated with ursolic acid or GSK3b inhibitor
SB216763 (10 µM) for 24 h were fixed in 75% etha-
nol at –20°C, resuspended in PBS containing RNase
A (1 mg/ml), and incubated for 1 h at 37°C. The fixed
cells were stained with propidium iodide (50 µg/ml)
for 30 min at room temperature. The DNA contents
were analyzed using CellQuest Software with the
FACSCalibur flow cytometry (Becton Dickinson,
Franklin Lakes, NJ, USA).
Pharmacological Reports, 2013, 65, 1366�1374 1367
Wnt signaling mediates ursolic acid-induced apoptosisJi-Hyuk Park et al.

Western blotting
Whole cell lysates from the prostate cancer cells
exposed to ursolic acid or SB216763 (10 µM) or
Wnt3a-CM for 24 h were prepared using lysis buffer
(50 mM Tris-HCl, pH 7.4, 150 mM NaCl, 1% Triton
X-100, 0.1% SDS, 1 mM EDTA, 1 mM Na3VO4,1 mM NaF and protease inhibitor cocktail). The pro-
tein contents in the supernatants were measured by
using a Bio-Rad DC protein assay kit II (Bio-Rad,
Hercules, CA, USA), separated on 4–12% NuPAGE
Bis-Tris gels (Invitrogen, Carlsbad, CA, USA) and
electro-transferred onto a Hybond ECL transfer mem-
brane (GE Health Are Bio-Science, Piscataway, NJ,
USA). The membranes were blocked with 5% nonfat
dry milk and immunoblotted with anti-cleaved
caspase-3, caspase-3, cleaved caspase-9, caspase-9,
PARP, Bax, Bcl-2, Bcl-xL, Mcl-1L, Wnt5a/b, p-
GSK3b, GSK3b or b-catenin (Cell Signaling, Dan-
vers, MA, USA) antibodies.
Immunofluorescence assay
PC 3 cells in the absence or presence of ursolic acid
were fixed with 4% paraformaldehyde and permeabi-
lized in cold methanol for 15 min at –20°C. Fixed
cells were then washed twice with 1 × PBS, followed
by blocking with 10% normal goat serum blocking
solution (Zymed Laboratories, Carlsbad, CA, USA)
for 30 min. Cells were incubated with the primary an-
tibodies against b-catenin (Cell Signaling, Danvers,
MA, USA) for overnight at 4°C. The cells were
washed three times in 1 × PBS and incubated with Al-
exa Fluor 594 goat anti-rabbit IgG (Invitrogen, Carls-
bad CA, USA) for 45 min at room temperature in
a humidified chamber. Cells were washed in 1 × PBS,
mounted with Vectashield/DAPI (Vector Laborato-
ries, Burlingame, CA, USA) and visualized by a Carl
Zeiss LSM5 confocal microscope.
Statistical analysis
Statistical analysis of the data was conducted using
Sigmaplot version 12 software (Systat Software Inc.,
San Jose, CA, USA). All data were expressed as the
means ± standard deviation (SD). The statistically sig-
nificant differences between control and ursolic acid-
treated cells were calculated by the Student’s t-test.
Results
Cytotoxic effects of ursolic acid on PC-3, DU145and LNCaP prostate cancer cells
The cytotoxicity of ursolic acid (Fig. 1A) was evalu-
ated against PC-3, DU145 and LNCaP prostate cancer
cells using MTT assay. Cells were treated with vari-
ous concentrations of ursolic acid (0, 5, 10, 20, 40 or
80 µM) for 24 h. As shown in Figure 1B, ursolic acid
exerted cytotoxicity in PC-3, DU145 more than in
LNCaP cells. In contrast, ursolic acid did not show
significant cytotoxicity against Raw 264.7 leukemic
monocyte macrophage cells, while only high concen-
tration of ursolic acid (80 µM) affected the viability of
the cells.
1368 Pharmacological Reports, 2013, 65, 1366�1374
Fig. 1. Cytotoxic effect of ursolic acid in prostate cancer cells.(A) Chemical structure of ursolic acid (MW = 456). (B) Human pros-tate cancer cell lines PC-3, DU145, LNCaP or leukemic monocytemacrophage cell line Raw 264.7 were treated with various concentra-tions of ursolic acid (0, 5, 10, 20, 40 or 80 µM) for 24 h. Cell viabilitywas determined by XTT assay. The data represent the means ± SD.(C) Morphological feature of PC-3 cells were observed in inverted mi-croscope (100´). Cells were treated with ursolic acid (0, 7.5, 15 or30 µM) for 24 h. (D) PC-3 cells were treated with 30 µM of ursolic acidand then stained with ethidium bromide homodimer dye. Signalswere visualized under an Axio vision 4.0 fluorescence microscope

Apoptotic bodies were also observed in ursolic
acid-treated PC-3 cells under inverted microscope, in-
dicating apoptotic feature of ursolic acid, while intact
morphology was shown in untreated control (Fig. 1C).
Similarly, cell death was confirmed in ursolic acid
treated PC-3 cells by ethidium homodimer assay. As
shown in Figure 1D, ethidium homodimer assay re-
vealed apoptotic features in ursolic acid treated PC-3
cells compared to untreated control.
Ursolic acid increased the sub-G1 apoptotic
portion in PC-3 cells
To confirm whether the cytotoxicity of ursolic acid
against PC-3 cells was due to apoptosis induction, cell
cycle analysis was carried out. As shown in Figure 2,
ursolic acid increased sub-G1 apoptotic portion by 1.30
± 0.02, 5.95 ± 1.43, 24.33 ± 0.16, and 49.93 ± 4.09% at
the concentrations of 7.5, 15, 30 and 60 µM, respec-
tively, compared to untreated control (1.45 ± 0.23%).
Pharmacological Reports, 2013, 65, 1366�1374 1369
Wnt signaling mediates ursolic acid-induced apoptosisJi-Hyuk Park et al.
Fig. 2. Effect of ursolic acid on thesub-G1 apoptotic portion in PC-3 cells.Cells were untreated or treated withvarious concentraions of ursolic acid(7.5, 15 or 30 µM) for 24 h. After fixingin 75% ethanol, cells were stained withpropidium iodide (PI) and cell cycle wasanalyzed by flow cytometry. (A) Histo-grams of flow cytometry analysis re-veal cell cycle distribution of eachsample. (B) Graphs represent per-centages of apoptotic portion. Datarepresent the means ± SD. * p < 0.05vs. untreated control

Ursolic acid activated caspase cascades andregulated the Bcl-2 family proteins in PC-3 cells
Generally, apoptosis is induced through two distinctive
pathways such as cell death extrinsic pathway and mi-
tochondrial dependent intrinsic pathway [8]. Western
blotting showed that ursolic acid activated caspase-9
and -3 and cleaved PARP in PC-3 cells as shown in
Figure 3A, implying that ursolic acid induces apoptosis
via mitochondrial dependent pathway in PC-3 cells.
Anti-apoptotic Bcl-2 family proteins such as Bcl-2
and Bcl-xL are frequently overexpressed in cancers [22].
In the present study, western blotting revealed that urso-
lic acid suppressed the expression of Bcl-XL, Bcl 2, and
Mcl-1L as anti-apoptotic genes and also up-regulated the
expression of Bax in PC-3 cells as shown in Figure 3B.
To further compare the apoptotic activity of ursolic
acid in other prostate cancer cells, the effect of ursolic
acid on apoptosis related proteins such as PARP, cas-
pase 3, Bcl-xL, Bcl-2 in Du145 and LNCaP cells was
examined by western blotting. Here, ursolic acid acti-
vated PARP and Bax as well as suppressed the expres-
sion of Bcl-xL and Bcl-2 in DU145 cells, while ursolic
acid cleaved PARP and suppressed the expression of
Bcl-xL in LNCaP cells (Figs. 3C and D).
Ursolic acid induced apoptosis through theWnt5/GSK3b/b-catenin signaling in PC-3 cells
Glycogen synthase kinase 3b (GSK3b), a regulator of
glycogen metabolism is involved in protein synthesis,
cell proliferation, cell differentiation, microtubule dy-
namics, cell motility and apoptosis [9]. GSK3b plays
a critical role in Wnt/b-catenin signaling pathway
[34]. Here, ursolic acid suppressed the expression of
Wnt5a/b and b-catenin, and enhanced the phosphory-
lation of GSK3b at Ser 9 in PC-3 cells (Fig. 4A).
GSK3b selective inhibitor SB216763 blocked the de-
creased expression of Wnt5a/b and b-catenin, and the
activated phosphorylation of GSK3b at Ser 9 induced
by ursolic acid (Fig. 4B). Likewise, GSK3 inhibitor
SB216763 prevented the cleavages of caspase-3 and
PARP induced by ursolic acid in PC-3 cells (Fig. 4C),
implying that ursolic acid-induced apoptosis is con-
trolled through the Wnt5/GSK3b/b-catenin signaling.
Wnt (Wnt1, Wnt3a, and Wnt8) signalings promote
the dissociation of b-catenin from Axin and inhibit
b-catenin phopsphorylation and subsequently b-ca-
tenin degradation [14, 16]. As shown in Figure 4D,
ursolic treatment induced the degradation of b-catenin
(Fig. 4D lower panel), while b-catenin exhibited sub-
1370 Pharmacological Reports, 2013, 65, 1366�1374
Fig. 3. Effect of ursolic acid on activa-tion and expression of apoptosis-related protein in PC-3 cells. Cellswere treated with various concentra-tions of ursolic acid (0, 3.75, 7.5, 15 or30 µM) for 24 h. Western blotting wasperformed to see apoptosis-relatedproteins on PC-3 (A, B), DU145 (C)and LNCaP (D) cells

cellular localization in PC-3 cells (Fig. 4D upper
panel). HEK 293 cells treated with 20 µM of ursolic
acid in the presence of Wnt3a-conditioned medium
blocked the apoptotic signaling such as PARP and
caspase 3 (Fig. 4E). Also, ursolic acid attenuated the
expression of Wnt5a and b-catenin in DU145 and
LNCaP cells (Figs. 4 F and G).
Several studies reported that Wnt3a activates b-ca-
tenin dependent canonical Wnt signaling and inhibits
the proliferation [19, 26]. Thus, we investigated whether
or not Wnt3a-CM affected the apoptosis induced by ur-
solic acid. HEK 293 cells were treated with ursolic acid
in the absence and presence of Wnt3a-CM. As shown
in Figure 4E, Wnt3-CM treatment reversed the de-
creased b-catenin and increased cleavages of PARP
and caspase 3 induced by ursolic acid in HEK 293
cells, implying that Wnt signaling mediates the apopto-
sis induced by ursolic acid in HEK 293 cells.
Pharmacological Reports, 2013, 65, 1366�1374 1371
Wnt signaling mediates ursolic acid-induced apoptosisJi-Hyuk Park et al.
Fig. 4. Effect of ursolic acid on theWnt5/GSK3b/b-catenin signaling inPC-3 cells. (A) Cells were treated withvarious concentrations of ursolic acid(0, 3.75, 7.5, 15 or 30 µM) for 24 h.Western blotting was performed forWnt5a/b, GSK3b and b-catenin. (B, C)Cells were treated with ursolic acid (30µM) and/or GSK3b inhibitor SB216763(10 µM) for 8 h. Western blotting wascarried out with indicated antibodiessuch as Wnt5a/b, GSK3b and b-ca-tenin (B), cleaved PARP and cleavedcaspase-3 (C). (D) Cellular localizationof b-catenin was evaluated by indirectimmunofluorescence using b-cateninantibody. Immunofluorescence label-ing of b-catenin (red) in ursolic aciduntreated or treated PC-3 cells for 24 hare shown. Nuclei were shown withDAPI (blue). (E) HEK 293 cells weretreated with 20 µM of ursolic acid in theabsence or presence of Wnt3a-condi-tioned medium (Wnt3a-CM). Westernblotting was carried out with b-catenin,PARP and cleaved caspase 3. (F, G)Various concentrations of ursolic acid(0, 3.75, 7.5, 15 or 30 µM) were treatedin DU145 and LNCaP cells for 24 h.Western blotting was carried out to de-tect Wnt5a/b, GSK3b and b-catenin inDU145 (F) and LNCaP cells (G)
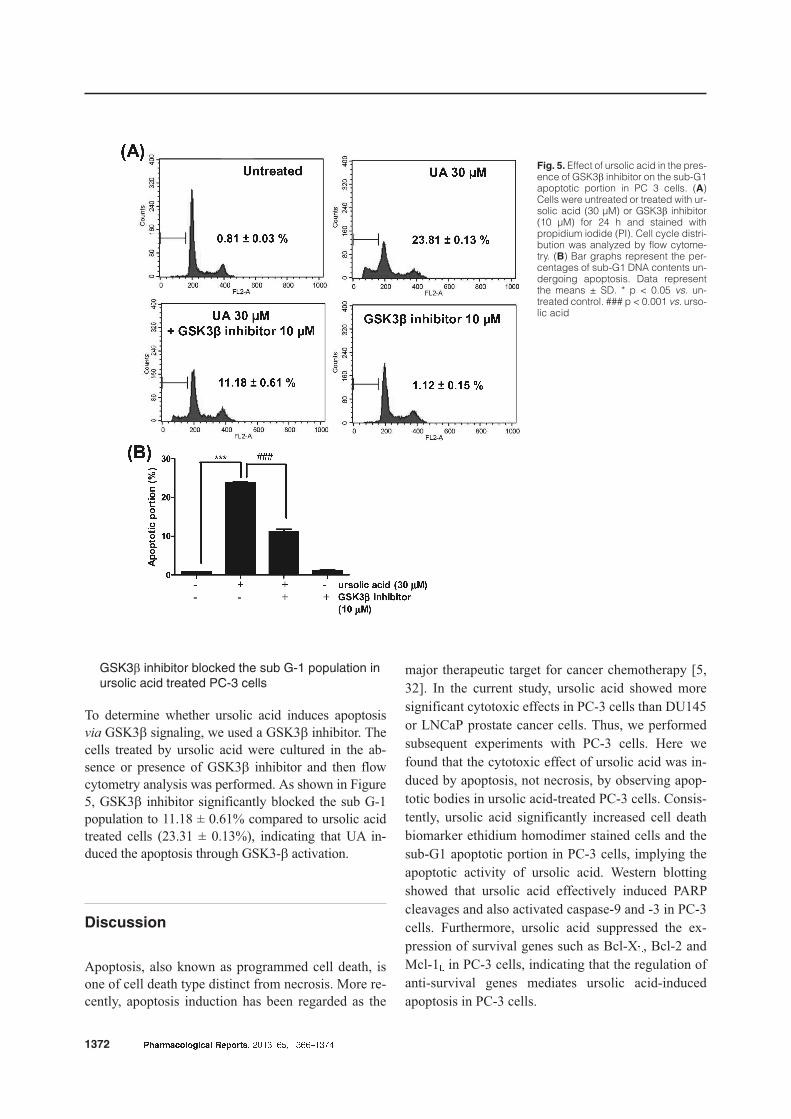
GSK3b inhibitor blocked the sub G-1 population inursolic acid treated PC-3 cells
To determine whether ursolic acid induces apoptosis
via GSK3b signaling, we used a GSK3b inhibitor. The
cells treated by ursolic acid were cultured in the ab-
sence or presence of GSK3b inhibitor and then flow
cytometry analysis was performed. As shown in Figure
5, GSK3b inhibitor significantly blocked the sub G-1
population to 11.18 ± 0.61% compared to ursolic acid
treated cells (23.31 ± 0.13%), indicating that UA in-
duced the apoptosis through GSK3-b activation.
Discussion
Apoptosis, also known as programmed cell death, is
one of cell death type distinct from necrosis. More re-
cently, apoptosis induction has been regarded as the
major therapeutic target for cancer chemotherapy [5,
32]. In the current study, ursolic acid showed more
significant cytotoxic effects in PC-3 cells than DU145
or LNCaP prostate cancer cells. Thus, we performed
subsequent experiments with PC-3 cells. Here we
found that the cytotoxic effect of ursolic acid was in-
duced by apoptosis, not necrosis, by observing apop-
totic bodies in ursolic acid-treated PC-3 cells. Consis-
tently, ursolic acid significantly increased cell death
biomarker ethidium homodimer stained cells and the
sub-G1 apoptotic portion in PC-3 cells, implying the
apoptotic activity of ursolic acid. Western blotting
showed that ursolic acid effectively induced PARP
cleavages and also activated caspase-9 and -3 in PC-3
cells. Furthermore, ursolic acid suppressed the ex-
pression of survival genes such as Bcl-XL, Bcl-2 and
Mcl-1L in PC-3 cells, indicating that the regulation of
anti-survival genes mediates ursolic acid-induced
apoptosis in PC-3 cells.
1372 Pharmacological Reports, 2013, 65, 1366�1374
Fig. 5. Effect of ursolic acid in the pres-ence of GSK3b inhibitor on the sub-G1apoptotic portion in PC 3 cells. (A)Cells were untreated or treated with ur-solic acid (30 µM) or GSK3b inhibitor(10 µM) for 24 h and stained withpropidium iodide (PI). Cell cycle distri-bution was analyzed by flow cytome-try. (B) Bar graphs represent the per-centages of sub-G1 DNA contents un-dergoing apoptosis. Data representthe means ± SD. * p < 0.05 vs. un-treated control. ### p < 0.001 vs. urso-lic acid

The Wnt/b-catenin pathway is involved in regulat-
ing the cell proliferation and cell differentiation of
neural stem/progenitor cells [30]. Also, there are ac-
cumulating evidences that GSK3b is closely associ-
ated with b-catenin signaling [27, 36]. The axin/
GSK3/APC complex normally promotes the prote-
olytic degradation of the b-catenin intracellular sig-
naling molecule [10]. GSK3b is involved in a wide
range of cellular processes including differentiation,
growth, motility and apoptosis. Thus, the aberrant
regulation of GSK3b has been implicated in various
diseases including Alzheimer’s disease, non-insulin-
dependent diabetes mellitus and cancer [7, 17]. Since
the balance of inhibitory phospho-serine9 (pSer9; in-
active form) and stimulatory phospho-tyrosine216
(pY216; active form) of GSK3b is considered a hall-
mark of its transient kinase regulation in normal
CD34+ cells and several human cell types, previous
studies demonstrated that GSK3b phosphorylation at
serine enhances apoptosis in breast cancer [29], me-
dulloblastoma [33] and SK-OV-3 cells [28]. Here, ur-
solic acid suppressed the expression of Wnt5a/b and
b-catenin, and induced the phosphorylation of GSK3b
at Ser 9, which is an indicator of GSK3b deactivation
in PC-3 cells. These data imply that ursolic acid-
induced apoptosis can be mediated by Wnt5a/b sup-
pression, b-catenin degradation and GSK3b inactiva-
tion in PC-3 cells. Consistently, GSK3b inhibitor
SB216763 or Wnt3a-CM blocked the cleavages of
caspase- 3 and PARP in ursolic acid-treated PC-3
cells, further confirming the involvement of GSK3b
phosphorylation in ursolic acid-induced apoptosis in
PC-3 cells. These results support scientific evidence
that medicinal plants such as Oldenlandia diffusa,
Eriobotrya japonica, Rosmarinus offýcinalis and Gle-
choma hederaceae containing ursolic acid as a lead-
ing compound can be applied to cancer prevention
and treatment as a complementary and alternative
medicine (CAM) agent.
Conflict of interest:
The authors declare no conflict of interest.
Acknowledgment:
This work was supported by the National Research Foundation of
Korea (NRF) grant funded by the Korea government [MEST] (No.
2012-0005755).
References:
1. Andersson D, Liu JJ, Nilsson A, Duan RD: Ursolic acid
inhibits proliferation and stimulates apoptosis in HT29
cells following activation of alkaline sphingomyelinase.
Anticancer Res, 2003, 23, 3317–3322.
2. Baade PD, Youlden DR, Krnjacki LJ: International epide-
miology of prostate cancer: geographical distribution and
secular trends. Mol Nutr Food Res, 2009, 53, 171–184
3. Chattopadhyay D, Dungdung SR, Mandal AB, Majum-
der GC: A potent sperm motility-inhibiting activity of
bioflavonoids from an ethnomedicine of Onge, Alstonia
macrophylla Wall ex A. DC, leaf extract. Contraception,
2005, 71, 372–378.
4. Chen XW, Sneed KB, Zhou SF: Pharmacokinetic profiles
of anticancer herbal medicines in humans and the clinical
implications. Curr Med Chem, 2011, 18, 3190–3210.
5. Cotter TG, Lennon SV, Glynn JG, Martin SJ: Cell death
via apoptosis and its relationship to growth, development
and differentiation of both tumour and normal cells.
Anticancer Res, 1990, 10, 1153–1159.
6. Crawford ED: Epidemiology of prostate cancer. Urology,
2003, 62, 3–12.
7. Forde JE, Dale TC: Glycogen synthase kinase 3: a key
regulator of cellular fate. Cell Mol Life Sci, 2007, 64,
1930–1944.
8. Fulda S: Evasion of apoptosis as a cellular stress re-
sponse in cancer. Int J Cell Biol, 2010, 370835.
9. Grimes CA, Jope RS: The multifaceted roles of glycogen
synthase kinase 3b in cellular signaling. Prog Neurobiol,
2001, 65, 391–426.
10. Gwak J, Oh J, Cho M, Bae SK, Song IS, Liu KH,
Jeong Y et al.: Galangin suppresses the proliferation
of b-catenin response transcription-positive cancer cells
Pharmacological Reports, 2013, 65, 1366�1374 1373
Wnt signaling mediates ursolic acid-induced apoptosisJi-Hyuk Park et al.
Fig. 6. A schematic diagram of ursolic acid induced apoptosis onWnt5/GSK3b/b-catenin signaling in prostate cancer cells

by promoting adenomatous polyposis coli/Axin/glyco-
gen synthase kinase-3b-independent b-catenin degrada-
tion. Mol Pharmacol, 2011, 79, 1014–1022.
11. Kassi E, Sourlingas TG, Spiliotaki M, Papoutsi Z,
Pratsinis H, Aligiannis N, Moutsatsou P: Ursolic acid
triggers apoptosis and Bcl-2 downregulation in MCF-7
breast cancer cells. Cancer Invest, 2009, 27, 723–733.
12. Kondo M, MacKinnon SL, Craft CC, Matchett MD,
Hurta RA, Neto CC: Ursolic acid and its esters: occur-
rence in cranberries and other Vaccinium fruit and effects
on matrix metalloproteinase activity in DU145 prostate
tumor cells. J Sci Food Agric, 2011, 91, 789–796.
13. Limami Y, Pinon A, Leger DY, Pinault E, Delage C,
Beneytout JL, Simon A, Liagre B: The
P2Y2/Src/p38/COX-2 pathway is involved in the resis-
tance to ursolic acid-induced apoptosis in colorectal and
prostate cancer cells. Biochimie, 2012, 94, 1754–1763.
14. Liu X, Rubin JS, Kimmel AR: Rapid, Wnt-induced
changes in GSK3b associations that regulate b-catenin
stabilization are mediated by Ga proteins. Curr Biol,
2005, 15, 1989–1997.
15. Liu XS, Jiang J: Induction of apoptosis and regulation of
the MAPK pathway by ursolic acid in human leukemia
K562 cells. Planta Med, 2007, 73, 1192–1194.
16. Logan CY, Nusse R: The Wnt signaling pathway in de-
velopment and disease. Annu Rev Cell Dev Biol, 2004,
20, 781–810.
17. Luo J: Glycogen synthase kinase 3b (GSK3b) in tumori-
genesis and cancer chemotherapy. Cancer Lett, 2009,
273, 194–200.
18. Messner B, Zeller I, Ploner C, Frotschnig S, Ringer T,
Steinacher-Nigisch A, Ritsch A et al.: Ursolic acid
causes DNA-damage, p53-mediated, mitochondria- and
caspase-dependent human endothelial cell apoptosis, and
accelerates atherosclerotic plaque formation in vivo.
Atherosclerosis, 2011, 219, 402–408.
19. Nygren MK, Dosen G, Hystad ME, Stubberud H,
Funderud S, Rian E: Wnt3a activates canonical Wnt sig-
nalling in acute lymphoblastic leukaemia (ALL) cells
and inhibits the proliferation of B-ALL cell lines. Br
J Haematol, 2007, 136, 400–413.
20. Ovesna Z, Vachalkova A, Horvathova K, Tothova D:
Pentacyclic triterpenoic acids: new chemoprotective
compounds. Minireview. Neoplasma, 2004, 51, 327–333.
21. Park S, Gwak J, Cho M, Song T, Won J, Kim DE,
Shin JG, Oh S: Hexachlorophene inhibits Wnt/b-catenin
pathway by promoting Siah-mediated b-catenin degrada-
tion. Mol Pharmacol, 2006, 70, 960–966.
22. Sasi N, Hwang M, Jaboin J, Csiki I, Lu B: Regulated cell
death pathways: new twists in modulation of BCL2 fam-
ily function. Mol Cancer Ther, 2009, 8, 1421–1429.
23. Shanmugam MK, Manu KA, Ong TH, Ramachandran L,
Surana R, Bist P, Lim LH et al.: Inhibition of CXCR4/
CXCL12 signaling axis by ursolic acid leads to suppres-
sion of metastasis in transgenic adenocarcinoma of mouse
prostate model. Int J Cancer, 2011, 129, 1552–1563.
24. Shanmugam MK, Ong TH, Kumar AP, Lun CK, Ho PC,
Wong PT, Hui KM, Sethi G: Ursolic acid inhibits the ini-
tiation, progression of prostate cancer and prolongs the
survival of TRAMP mice by modulating pro-
inflammatory pathways. PLoS One 2012, 7, e32476.
25. Shin SW, Kim SY, Park JW: Autophagy inhibition en-
hances ursolic acid-induced apoptosis in PC3 cells.
Biochim Biophys Acta, 2012, 1823, 451–457.
26. Sonderegger S, Haslinger P, Sabri A, Leisser C, Otten
JV, Fiala C, Knofler M: Wingless (Wnt)-3a induces tro-
phoblast migration and matrix metalloproteinase-2 secre-
tion through canonical Wnt signaling and protein kinase
B/AKT activation. Endocrinology, 2010, 151, 211–220.
27. Song S, Mazurek N, Liu C, Sun Y, Ding QQ, Liu K,
Hung MC, Bresalier RS: Galectin-3 mediates nuclear
b-catenin accumulation and Wnt signaling in human co-
lon cancer cells by regulation of glycogen synthase
kinase-3b activity. Cancer Res, 2009, 69, 1343–1349.
28. Song YH, Jeong SJ, Kwon HY, Kim B, Kim SH,
Yoo DY: Ursolic acid from Oldenlandia diffusa induces
apoptosis via activation of caspases and phosphorylation
of glycogen synthase kinase 3 beta in SK-OV-3 ovarian
cancer cells. Biol Pharm Bull, 2012, 35, 1022–1028.
29. Soto-Cerrato V, Viñals F, Lambert JR, Kelly JA, Perez-
Tomas R: Prodigiosin induces the proapoptotic gene NAG-1
via glycogen synthase kinase-3 b activity in human breast
cancer cells. Mol Cancer Ther, 2007, 6, 362–369.
30. Syed Khaja AS, Helczynski L, Edsjo A, Ehrnstrom R,
Lindgren A, Ulmert D, Andersson T, Bjartell A: Ele-
vated level of Wnt5a protein in localized prostate cancer
tissue is associated with better outcome. PLoS One,
2011, 6, e26539.
31. Taniguchi S, Imayoshi Y, Kobayashi E, Takamatsu Y,
Ito H, Hatano T, Sakagami H et al.: Production of bioac-
tive triterpenes by Eriobotrya japonica calli. Phytochem-
istry, 2002, 59, 315–323.
32. Ulukaya E, Acilan C, Yilmaz Y: Apoptosis: why and
how does it occur in biology? Cell Biochem Funct, 2011,
29, 468–480.
33. Urbanska K, Trojanek J, Del Valle L, Eldeen MB,
Hofmann F, Garcia-Echeverria C, Khalili K, Reiss K:
Inhibition of IGF-I receptor in anchorage-independence
attenuates GSK-3b constitutive phosphorylation and
compromises growth and survival of medulloblastoma
cell lines. Oncogene, 2007, 26, 2308–2317.
34. Wu D, Pan W: GSK3: a multifaceted kinase in Wnt sig-
naling. Trends Biochem Sci, 2010, 35, 161–168.
35. Yang AK, He SM, Liu L, Liu JP, Wei MQ, Zhou SF:
Herbal interactions with anticancer drugs: mechanistic
and clinical considerations. Curr Med Chem, 2010, 17,
1635–1678.
36. Zhang X, Yin WK, Shi XD, Li Y: Curcumin activates
Wnt/b-catenin signaling pathway through inhibiting the
activity of GSK-3b in APPswe transfected SY5Y cells.
Eur J Pharm Sci, 2011, 42, 540–546.
37. Zhang Y, Kong C, Zeng Y, Wang L, Li Z, Wang H, Xu
C, Sun Y: Ursolic acid induces PC-3 cell apoptosis via
activation of JNK and inhibition of Akt pathways in vi-
tro. Mol Carcinog, 2010, 49, 374–385.
Received: October 8, 2012; in the revised form: May 10, 2013;
accepted: June 7, 2013.
1374 Pharmacological Reports, 2013, 65, 1366�1374


















