
Influence of TAT-peptide polymerization on properties …chem1.eng.wayne.edu/~gzmao/Polyplexes Paper...
14
Influence of TAT-peptide polymerization on properties and transfection activity of TAT/DNA polyplexes Devika Soundara Manickam a , Harender S. Bisht a , Lei Wan b , Guangzhao Mao b , David Oupicky a, * a Department of Pharmaceutical Sciences, Wayne State University, Detroit, MI 48202, USA b Department of Chemical Engineering and Materials Science, Wayne State University, Detroit MI 48202, USA Received 25 August 2004; accepted 22 September 2004 Available online 21 November 2004 Abstract Use of bioactive cationic peptides as gene carriers is limited by instability of their DNA complexes in vivo and by the loss of their biological activity due to undesired interactions of their bioactive parts with the DNA. To overcome the two major limitations, biodegradable high-molecular-weight form of TAT peptide (POLYTAT) sensitive to cellular redox-potential gradients was synthesized in this study by oxidative polycondensation. Physicochemical and transfection properties of DNA polyplexes based on POLYTATwere investigated and compared with polyplexes based on TAT polymer prepared by in situ template-assisted polymerization. Physicochemical properties of TAT-based polyplexes were affected by the molecular weight and method of polymerization of the TAT peptide. All TAT-based DNA polyplexes exhibited reduced cytotoxicity when compared with control polyethylenimine (PEI) polyplexes. Polyplexes based on both high-molecular-weight TAT polypeptides exhibited increased transfection efficiency compared to control TAT peptide but lower than that of PEI polyplexes. The evidence shows that transfection activity of TAT-based polyplexes is strongly dependent on the presence of chloroquine and therefore suggests that TAT polyplexes are internalized by an endocytosis. Overall, high-molecular-weight reducible polycations based on bioactive peptides has the potential as versatile carriers of nucleic acids that display low cytotoxicity and can prove to be especially beneficial in cases that require surface presentation of membrane-active or cell-specific targeting peptides. D 2004 Elsevier B.V. All rights reserved. Keywords: Polyplex; TAT; Polypeptide; Transfection; Toxicity 1. Introduction Gene therapy has shown a potential to treat and prevent a wide variety of genetic and acquired diseases. In order to fully utilize this potential, safer and more efficient vectors for delivery of genes are 0168-3659/$ - see front matter D 2004 Elsevier B.V. All rights reserved. doi:10.1016/j.jconrel.2004.09.018 * Corresponding author. Tel.: +1 313 993 7669; fax: +1 313 577 2033. E-mail address: [email protected] (D. Oupicky). Journal of Controlled Release 102 (2005) 293 – 306 www.elsevier.com/locate/jconrel GENE DELIVERY
Transcript of Influence of TAT-peptide polymerization on properties …chem1.eng.wayne.edu/~gzmao/Polyplexes Paper...

www.elsevier.com/locate/jconrel NE
DELIV
ER
Y
Journal of Controlled Releas
GE
Influence of TAT-peptide polymerization on properties and
transfection activity of TAT/DNA polyplexes
Devika Soundara Manickama, Harender S. Bishta, Lei Wanb,
Guangzhao Maob, David Oupickya,*
aDepartment of Pharmaceutical Sciences, Wayne State University, Detroit, MI 48202, USAbDepartment of Chemical Engineering and Materials Science, Wayne State University, Detroit MI 48202, USA
Received 25 August 2004; accepted 22 September 2004
Available online 21 November 2004
Abstract
Use of bioactive cationic peptides as gene carriers is limited by instability of their DNA complexes in vivo and by the loss of
their biological activity due to undesired interactions of their bioactive parts with the DNA. To overcome the two major
limitations, biodegradable high-molecular-weight form of TAT peptide (POLYTAT) sensitive to cellular redox-potential
gradients was synthesized in this study by oxidative polycondensation. Physicochemical and transfection properties of DNA
polyplexes based on POLYTAT were investigated and compared with polyplexes based on TAT polymer prepared by in situ
template-assisted polymerization. Physicochemical properties of TAT-based polyplexes were affected by the molecular weight
and method of polymerization of the TAT peptide. All TAT-based DNA polyplexes exhibited reduced cytotoxicity when
compared with control polyethylenimine (PEI) polyplexes. Polyplexes based on both high-molecular-weight TAT polypeptides
exhibited increased transfection efficiency compared to control TAT peptide but lower than that of PEI polyplexes. The
evidence shows that transfection activity of TAT-based polyplexes is strongly dependent on the presence of chloroquine and
therefore suggests that TAT polyplexes are internalized by an endocytosis. Overall, high-molecular-weight reducible
polycations based on bioactive peptides has the potential as versatile carriers of nucleic acids that display low cytotoxicity and
can prove to be especially beneficial in cases that require surface presentation of membrane-active or cell-specific targeting
peptides.
D 2004 Elsevier B.V. All rights reserved.
Keywords: Polyplex; TAT; Polypeptide; Transfection; Toxicity
0168-3659/$ - see front matter D 2004 Elsevier B.V. All rights reserved.
doi:10.1016/j.jconrel.2004.09.018
* Corresponding author. Tel.: +1 313 993 7669; fax: +1 313
577 2033.
E-mail address: [email protected] (D. Oupicky).
1. Introduction
Gene therapy has shown a potential to treat and
prevent a wide variety of genetic and acquired
diseases. In order to fully utilize this potential, safer
and more efficient vectors for delivery of genes are
e 102 (2005) 293–306

D. Soundara Manickam et al. / Journal of Controlled Release 102 (2005) 293–306294
GEN
ED
ELIV
ER
Y
required. Vectors based on polyelectrolyte complexes
of nucleic acids and synthetic cationic polymers
(polyplexes) represent one of the major alternatives
to viral vectors that usually do not raise the safety
issues associated with viruses but unfortunately their
efficiency, despite enormous progress in recent years,
has yet to achieve competitive levels.
Bioactive cationic peptides capable of condensing
nucleic acids have attracted considerable attention as
parts of gene delivery vectors because of a highly
reproducible and scalable production and inherent
specific biological activity. Natural or artificial pep-
tides are often used to supplement other delivery
systems with various biological functions such as cell
specific targeting, membrane destabilizing activity, or
nuclear localization activity [1–3]. When used alone,
small cationic peptides usually exhibit lower cytotox-
icity and are also weaker activators of the complement
system than high-molecular-weight polycations [4,5].
However, the DNA complexes formed with short
cationic peptides are significantly less stable due to
less prominent cooperative effect [6]. Consequently,
such complexes may lack sufficient stability to
survive blood circulation and to protect DNA from
metabolism in vivo [7]. Additional obstacle hindering
efforts to use peptides as gene delivery vectors is that
some of the potentially useful cationic peptides
interact strongly with nucleic acids via their bioactive
parts, which leaves them unavailable for other
interactions and therefore inactive [8]. Published
evidence indicates that such peptides can constitute
effective components of synthetic gene delivery
complexes, as long as sufficient copies are displayed
on the outer surface of the complex [1,9].
A way of overcoming the two major limitations of
the small cationic bioactive peptides, i.e. low extrac-
ellular stability of their complexes with DNA and the
unwanted interactions with DNA resulting in reduc-
tion of their original biological activity, is to use
readily reversible polymeric forms of the peptides.
Such reversible polypeptides can be easily prepared
from peptides with terminal cysteine residues either
by oxidative polycondensation via the terminal
sulfhydryl groups using dimethylsulfoxide (DMSO)-
mediated oxidation [10] or by DNA template poly-
merization in situ [11,12]. In both cases, high-
molecular-weight polypeptides containing disulfide
bonds in the backbone are produced that can undergo
selective intracellular depolymerization mediated by
small redox molecules or redox enzymes [13,14]. An
additional attractive feature of using the disulfide
bonds in the polypeptide structure is their relative
extracellular stability, which reversibly increases the
affinity of the peptide to the nucleic acid during
extracellular delivery [15,16]. High-molecular-weight
reducible polypeptides can also offer better surface
presentation of the peptides on the surface of DNA
complexes. The intracellular depolymerization of
reducible polypeptides favorably affects cytotoxic
properties of their polyplexes and increases rates of
intracellular disassembly, which leads to enhanced
levels of transfection of both therapeutic DNA and
mRNA [13].
TAT-peptide, amino acid residues 47–57 of the
transactivating transcriptional activator protein from
HIV-1, is an example of a so-called protein trans-
duction domain that recently attracted considerable
attention. TAT and similar peptides have been ascribed
with the unusual ability to translocate across cell
membranes in a receptor-independent and temper-
ature-independent manner [17,18]. The apparent lack
of a size limit and apparent ability of TAT-peptide to
transduce proteins and larger delivery vehicles
directly into cell cytoplasm seemed an attractive
feature that could have brought significant advantages
to cellular delivery of macromolecular therapeutics
[9,19,20]. The original reports suggested that TAT-
peptide transduction involves direct penetration of the
lipid bilayer caused by the localized positive charge of
the peptide. Recently, however, a number of studies
questioned the validity of the original reports and
suggested that (1) the internalization of TAT is a
temperature- and energy-dependent process, (2) endo-
somal transport is a key component of the mechanism,
and (3) TAT merely increases non-specific binding to
the cell surface [21–24].
The objective of this study is to design more
efficient and less toxic means of delivering nucleic
acids using TAT-derived polypeptides sensitive to
cellular redox-potential gradients. The central
hypothesis is that using high-molecular-weight form
of TAT peptide will permit more effective presenta-
tion of TAT residues on the surface of DNA
polyplexes, which will improve overall efficiency
of gene delivery process due to better interactions of
TAT polyplexes with cell membranes. To test the

D. Soundara Manickam et al. / Journal of Controlled Release 102 (2005) 293–306 295
EN
ED
ELIV
ER
Y
hypothesis, biophysical and transfection properties of
DNA polyplexes based on TAT polypeptides pre-
pared either in situ using template-assisted polymer-
ization or pre-formed by oxidative polycondensation
were investigated.
G
2. Materials and methods
2.1. Materials
Peptides, GTATG (GGRKKRRQRRRGG, Mr
1568) and CTATC (CGRKKRRQRRRGC, Mr 1660)
were synthesized using a standard Fmoc procedure by
Sigma-GENOSYS and purified to homogeneity by
HPLC using Discovery Bio-Wide-Pore C-18
(250�4.6 mm) column eluted with 0.1% trifluoro-
acetic acid and a gradient of acetonitrile (0�67.5%).
Polyethylenimine (PEI) with average molecular weigh
25,000 was from Aldrich. Plasmid DNA, gWizkHigh-Expression Luciferase (gWIZLuc), containing
luciferase reporter gene was purchased from Aldevron
(Fargo, ND) as 5 mg/mL aqueous solution and used
without further purification.
2.2. Synthesis of POLYTAT polypeptide
The POLYTAT polypeptide was synthesized by
oxidative polycondensation as previously described
[10]. Briefly, 20.8 mg of CTATC was dissolved in 145
AL phosphate buffered saline (PBS) and 70 ALDMSO. The reaction was carried out at room
temperature for 96 h. During the reaction, aliquots
were removed at 12-h time intervals and analyzed by
Size Exclusion Chromatography (SEC). The reaction
was terminated by dilution into 15 mL of 5 mM
HEPES buffer solution (pH 7) after no change in
molecular weight distribution was observed in two
consecutive SEC analyses. The SEC analysis was
performed using CATSEC-300 column eluted with
0.2 M NaCl and 0.1% TFA. Commercially available
poly(l-lysine) (PLL) (Sigma) samples were used to
estimate molecular weight of POLYTAT. Low molec-
ular weight impurities were removed by centrifugal
membrane filters with molecular weight cut-off
10,000. The concentration of POLYTAT was deter-
mined by a colorimetric 2,4,6-trinitrobenzenesulfonic
acid assay using GTATG calibration. Degradability of
POLYTAT was confirmed by SEC following reduc-
tion with 5 mM dithiothreitol.
2.3. Ethidium bromide exclusion assay
The ability of GTATG, CTATC, and POLYTAT to
condense DNAwas confirmed by a standard ethidium
bromide exclusion assay by measuring the changes in
ethidium bromide/DNA fluorescence. DNA (gWI-
ZLuc) solutions at a concentration of 20 Ag/mL were
mixed with ethidium bromide (1 Ag/mL) and fluo-
rescence measured using 360-nm excitation and 520-
nm emission and set to 100%. Background fluores-
cence was set to 0% using ethidium bromide (1 Ag/mL) solution alone. Fluorescence readings were taken
following a stepwise addition of a peptide solution,
and condensation curves for each peptide constructed.
2.4. Formulation of DNA polyplexes
Plasmid DNA (gWIZLuc) solution at a concen-
tration 20 Ag/mL in 10 mM HEPES buffer (pH 7) was
used to prepare all polyplexes in this study. Peptide/
DNA complexes were formed by adding a small
predetermined volume of a peptide (5 mg/mL) to
achieve the desired molar mixing ratio X (X=c (basic
amino acid residues of a peptide or amino groups of
PEI)/c (DNA phosphates)) and mixed by vigorous
vortexing for 10 s. Mass of 325 per one phosphate
group of DNA was assumed in the calculations.
Polyplexes were used in further experiments at least
90 min after formulation to permit efficient template-
assisted polymerization of CTATC in CTATC/DNA
polyplexes.
2.5. Molecular weight determination
Apparent weight-average molecular weights (Mw,a)
of DNA complexes were measured using a 7-angle
BiMwA Molecular Weight Analyzer equipped with
30 mW, vertically polarized solid state laser (660 nm)
as a light source (Brookhaven Instruments). The
instrument was calibrated with toluene and the
response of the CCD detectors was normalized with
a nominal 20-nm diameter polystyrene latex in water
(Duke Scientific). The static light scattering data were
analyzed in most cases by linear extrapolation to zero
scattering angle to obtain Mw,a of the DNA com-

D. Soundara Manickam et al. / Journal of Controlled Release 102 (2005) 293–306296
GEN
ED
ELIV
ER
Y
plexes. Calculation of refractive index increments of
the complexes as a function of the molar mixing ratios
was carried out using previously published model of
complex formation [25]. Previously published refrac-
tive index increment value of 0.185 mL/g was used
for DNA. Known refractive index increment of a
polypeptide poly(l-lysine) (0.188 mL/g) was used as
a good approximation for all three TAT formulations.
In all experiments, 1:1 charge stoichiometry and a full
release of counterions were assumed. Extrapolation to
zero concentration was not performed due to very low
concentrations of the complexes. GTATG, CTATC,
and POLYTAT polyplexes at various X were prepared
at DNA concentration of 20 Ag/mL in 10 mM HEPES
(pH 7) 90 min before analysis. Large dust particles
were removed by centrifugation and complexes were
diluted with 10 mM HEPES (pH 7) to the final DNA
concentration of 10 Ag/mL. Standard error of meas-
urement was calculated for each Mw,a determination.
To assess reproducibility of polyplex preparation,
three separate formulations of selected polyplexes
were also analyzed.
2.6. Hydrodynamic radius and zeta potential
determination
The determination of hydrodynamic diameters and
zeta potentials of peptide/DNA complexes was
performed using ZetaPlus Particle Size and Zeta
Potential analyzer (Brookhaven Instruments) equip-
ped with a 35-mW solid state laser (658 nm).
Scattered light was detected at 908 angle and a
temperature of 25 8C. Complexes of gWIZLuc
plasmid with GTATG, CTATC, and POLYTAT were
prepared in 10 mM HEPES (pH 7) as described
above. Mean hydrodynamic diameters were calculated
for size distribution by weight, assuming a lognormal
distribution using the supplied algorithm and the
results are expressed as meanFS.D. of three runs.
Zeta potential values were calculated from measured
velocities using Smoluchowski equation and results
are expressed as meanFS.D. of 10 runs.
2.7. AFM characterization
Ten microliters of DNA or peptide/DNA complex
solution (20 Ag DNA/mL, X=3.0) was placed on
freshly cleaved mica. After 10 min, excess of solution
was removed and surface dried with a gentle stream of
nitrogen. AFM images were acquired using a Multi-
mode Nanoscope III AFM (Digital Instruments) in
tapping mode in air. Height, amplitude, and phase
images were captured, but only height images are
shown here. Silicon probes with a nominal radius of
curvature 10 nm (NSG10, NanoTechnology Instru-
ments, Europe) were used. Scan rate was 1 Hz.
Integral and proportional gains were approximately
0.4 and 0.7, respectively. In some cases, in situ
imaging was conducted by injecting the solution of a
DNA polyplexes into an AFM liquid cell, which was
sealed by an o-ring, and the adsorbed polyplex
structure was obtained in tapping mode in the
solution.
2.8. Cell lines
Murine melanoma cell line B16F10 (CRL-6475)
and human cervical carcinoma cell line HeLa (CCL-2)
were obtained from ATCC. Human endothelial cell
line EA.hy926, derived by a fusion of human
umbilical vein endothelial cells with a human lung
carcinoma A549, was a kind gift from Dr. Edgell
(University of North Carolina). All three cell lines
were maintained in Dulbecco’s Modified Eagle
Medium (DMEM) supplemented with 4 mM l-
alanyl-l-glutamine (GlutamaxR) and 10% fetal
bovine serum (FBS).
2.9. Cytotoxicity
Cytotoxicity of TAT-based DNA polyplexes was
determined by the CellTiter 96R Aqueous Cell
Proliferation Assay (Promega) and compared with
that of PEI/DNA polyplexes. Twenty thousand
EA.hy926 cells were seeded in a 96-well plate. Two
days after reaching confluence, the cells were
incubated in 150 AL of DMEM/FBS with 2.9 AgDNA/mL dose of GTATG/DNA, CTATC/DNA,
POLYTAT/DNA, and PEI/DNA complexes. The
medium was removed after 24 h and replaced with a
mixture of 100 AL fresh DMEM and 20 AL MTS
reagent solution. The cells were incubated for 1 h at
37 8C in CO2 incubator. The absorbance of each well
was then measured at 505 nm to determine cell
viability. The results are expressed as mean% cell
viability relative to untreated cellsFS.D.

D. Soundara Manickam et al. / Journal of Controlled Release 102 (2005) 293–306 297
GEN
ED
ELIV
ER
Y
2.10. Transfection efficiency in vitro
Transfection activity was analyzed using DNA
(gWIZluc) complexes of GTATG, CTATC, POLY-
TAT, and PEI prepared as described above (DNA
concentration 20 Ag/mL) at the indicated molar
mixing ratio X. All transfection studies were per-
formed in 48-well plates with cells plated 24 h before
transfection at the seeding density of 30,000 cells per
well using a previously described general protocol
[26]. On a day of transfection, the cells were
incubated with the complexes in 150 AL of FBS
supplemented DMEM. Unless stated otherwise, plas-
mid DNA concentration in the incubation media was
2.9 Ag/mL. After a 3-h incubation, cells were washed
with PBS and cultured for additional 24 h in a fresh
DMEM/FBS prior to analysis of reporter gene
expression. The culture medium was discarded and
cell lysates harvested after incubation of cells for 30
min at room temperature in 100 AL of cell lysis
reagent buffer (Promega). To measure the luciferase
content, 100 AL of luciferase assay buffer (20 mM
glycylglycine (pH 8), 1 mM MgCl2, 0.1 mM EDTA,
3.5 mM DTT, 0.5 mM ATP, 0.27 mM coenzyme A)
was automatically injected into 20 AL of cell lysate
and the luminescence was integrated over 10 s using
single tube Sirius luminometer (Zylux). Total cellular
protein in the cell lysate was determined by the BCA
protein assay using calibration curve constructed with
standard bovine serum albumin solutions. The trans-
fection results are expressed as Relative Light Units
(RLU) per mg of cellular protein. At least three
determinations on two separate occasions were
performed for each transfection experiment. Unless
stated otherwise, the results are expressed as mean
RLUFS.D. and where necessary, significant differ-
ences between two groups are determined by Stu-
dent’s t-test and between multiple groups by ANOVA
using Holm test for multiple comparisons. A value of
Pb0.05 was considered statistically significant.
3. Results and discussion
3.1. POLYTAT synthesis
Artificial or naturally derived cationic peptides
have been widely used as part of synthetic gene
delivery systems because of their DNA condensation
ability and possibility to exploit their intrinsic bio-
logical activity for enhancing the gene delivery
process. While DNA condensation is largely unaf-
fected by the amino acid sequence of a cationic
peptide, its biological activity is strongly dependent
on it. Current knowledge suggests that systemic
intravenous delivery of DNA complexes benefits
from using complexes with low disassembly rates.
This requires using high-molecular-weight polyca-
tions and severely limits the use of bioactive cationic
peptides for systemic gene delivery. Synthesis of high-
molecular-weight peptides with a defined sequence
using traditional methods of peptide synthesis or
methods of genetic engineering is complicated and
often not feasible. If a peptide can be engineered to
contain terminal cysteinyl residues (i.e. if the peptide
termini are not required for its biological activity) then
a simple method of oxidative polycondensation can be
used for the synthesis of high-molecular-weight
polypeptides [10]. In this study, a method was used,
which is based on a mild DMSO oxidation of
sulfhydryl groups into disulfides, to synthesize poly-
peptide consisting of the TAT repeating unit (Scheme
1). The reaction reached equilibrium after 4 days,
furnishing high-molecular-weight form of the TAT
peptide (POLYTAT) with molecular weight distribu-
tion shown in Fig. 1 and with an estimated average
molecular weight 9.4�104. The low molecular weight
residues seen in the chromatogram are likely to be
impurities not containing any sulfhydryl groups or
cyclic by-products. The presence of such impurities
was observed also in polypeptides of different amino
acid sequences used in other studies.
3.2. Formulation and physical properties of DNA
polyplexes
One of the objectives of this study was to
determine how a polymerization method influences
properties of polyplexes based on TAT polypeptides.
DNA polyplexes of POLYTAT (POLYTAT/DNA) and
polyplexes prepared by DNA-template-assisted poly-
merization of CTATC peptide (CTATC/DNA) per-
formed as described in Ref. [7] were studied.
Polyplexes based on non-polymerizable GTATG
peptide (GTATG/DNA) served as controls in all
experiments. The ability of all three TAT peptides to
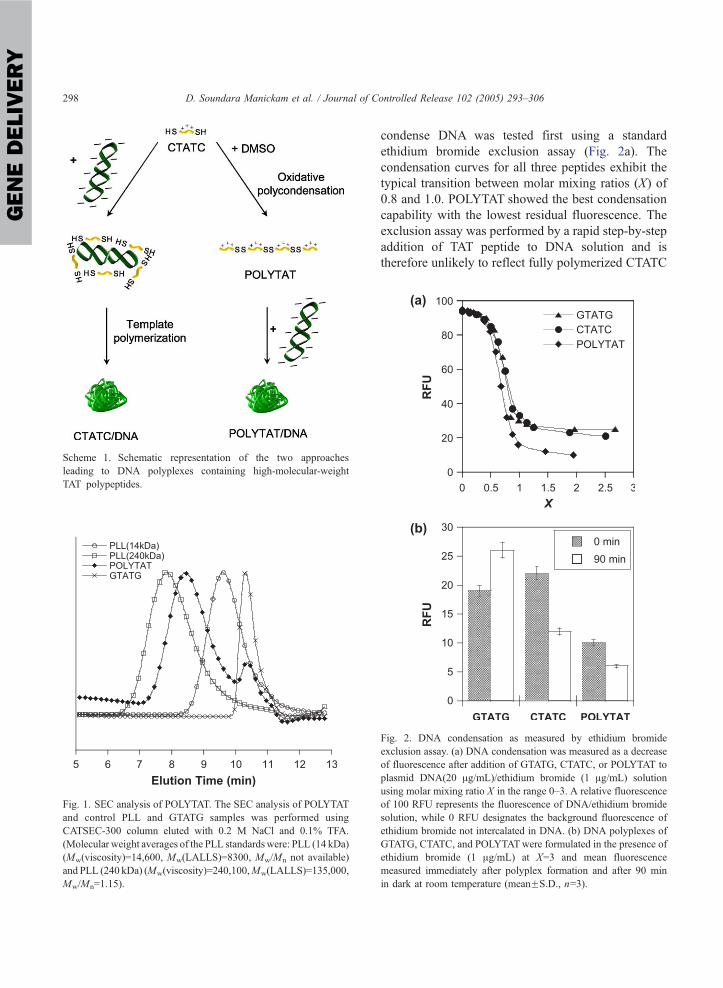
Fig. 1. SEC analysis of POLYTAT. The SEC analysis of POLYTAT
and control PLL and GTATG samples was performed using
CATSEC-300 column eluted with 0.2 M NaCl and 0.1% TFA.
(Molecular weight averages of the PLL standardswere: PLL (14 kDa)
(Mw(viscosity)=14,600, Mw(LALLS)=8300, Mw/Mn not available)
and PLL (240 kDa) (Mw(viscosity)=240,100,Mw(LALLS)=135,000,
Mw/Mn=1.15).
Scheme 1. Schematic representation of the two approaches
leading to DNA polyplexes containing high-molecular-weight
TAT polypeptides.
D. Soundara Manickam et al. / Journal of Controlled Release 102 (2005) 293–306298
GEN
ED
ELIV
ER
Y
condense DNA was tested first using a standard
ethidium bromide exclusion assay (Fig. 2a). The
condensation curves for all three peptides exhibit the
typical transition between molar mixing ratios (X) of
0.8 and 1.0. POLYTAT showed the best condensation
capability with the lowest residual fluorescence. The
exclusion assay was performed by a rapid step-by-step
addition of TAT peptide to DNA solution and is
therefore unlikely to reflect fully polymerized CTATC
Fig. 2. DNA condensation as measured by ethidium bromide
exclusion assay. (a) DNA condensation was measured as a decrease
of fluorescence after addition of GTATG, CTATC, or POLYTAT to
plasmid DNA(20 Ag/mL)/ethidium bromide (1 Ag/mL) solution
using molar mixing ratio X in the range 0–3. A relative fluorescence
of 100 RFU represents the fluorescence of DNA/ethidium bromide
solution, while 0 RFU designates the background fluorescence of
ethidium bromide not intercalated in DNA. (b) DNA polyplexes of
GTATG, CTATC, and POLYTATwere formulated in the presence of
ethidium bromide (1 Ag/mL) at X=3 and mean fluorescence
measured immediately after polyplex formation and after 90 min
in dark at room temperature (meanFS.D., n=3).

D. Soundara Manickam et al. / Journal of Controlled Release 102 (2005) 293–306 299
GEN
ED
ELIV
ER
Y
as suggested by similar condensation profiles for
GTATG and CTATC. Previous reports suggested that
the template-assisted polymerization of similar cati-
onic peptides proceeds rapidly and requires less than
30 min for completion [11]. Fig. 2b shows the residual
fluorescence intensity immediately after complex
formation and after a 90-min incubation. Significant
decrease of the fluorescence intensity observed for
CTATC/DNA suggests tightening of the polyplex
structure due to underwent polymerization. No such
decrease was observed for GTATG/DNA polyplexes.
Addition of a polycation to DNA solution results in
the DNA condensation and a formation of primary
complexes, which can then undergo a secondary
association until a colloidal equilibrium is reached.
Low molecular weight polycations usually form
polyplexes with a very low colloidal stability leading
to a formation of large particles. It can be assumed
that CTATC and GTATG peptides form primary
polyplexes of similar properties. In case of GTATG/
DNA, these polyplexes undergo rapid aggregation;
resulting in large hydrodynamic diameters (Fig. 3).
Size of the CTATC/DNA polyplexes, on the other
hand, will be determined by the balance between the
rate of aggregation and the rate of stabilizing template
polymerization. The stabilizing effect of the template
polymerization on the hydrodynamic diameter of
CTATC/DNA polyplexes is clearly demonstrated in
Fig. 3. Influence of template polymerization of CTATC on
aggregation of primary DNA polyplexes. Immediately after
preparation of CTATC and GTATG complexes with plasmid DNA
at X=3, hydrodynamic diameters were measured in 30-s intervals by
dynamic light scattering.
Fig. 3. Although the instrumentation available in the
authors’ laboratory does not permit capturing the early
phases of aggregation of primary complexes immedi-
ately after polycation addition, it nevertheless allows
to clearly demonstrate the stabilizing effect of
template polymerization of CTATC.
Biological activity of DNA polyplexes is affected
by their molecular parameters [27]. Parameters such
as size, surface charge, and molecular weight are
known to be controlled by the molecular weight of the
used polycations. Fig. 4 shows hydrodynamic diam-
eter, zeta potential, and apparent molecular weight for
all three TAT-based polyplexes prepared at various
molar mixing ratios, X. Fig. 4a confirms that increas-
ing X leads to a formation of smaller polyplexes. As
expected, POLYTAT forms polyplexes with the
smallest size of the three formulations due to its high
molecular weight that permits effective colloidal
stabilization via formation of stabilizing shell of
non-interacting parts of POLYTAT molecules on the
surface of the polyplexes [28]. This was further
confirmed by the observed highest positive zeta
potential of POLYTAT/DNA (Fig. 4b). Fig. 4c
confirms that GTATG forms a highly aggregated
polyplexes with the highest apparent molecular
weight of the three TAT-peptide forms tested. Appa-
rent molecular weight of the POLYTAT/DNA poly-
plexes decreases with increasing X, confirming the
effect of POLYTAT excess on stabilization of the
polyplexes against aggregation as observed for the
dependence of hydrodynamic diameters on X. In
contrast, apparent molecular weight of CTATC/DNA
increases with increasing X. The increase of molecular
weight, combined with decrease of the size of CTATC
polyplexes, would indicate increase of structural
density of CTATC polyplexes with increasing excess
of CTATC peptide. Typically, however, the structural
density of polyplexes based on high-molecular-weight
polycations increases at XN1 with increasing excess of
the polycation because of the intra-complex repulsive
action of excessive polycation [29–31]. Further
physical studies of CTATC/DNA polyplexes are
required to fully confirm and explain the molecular
weight behavior of CTATC/DNA polyplexes.
To determine possible influence of the polymer-
ization method on morphology of the polyplexes,
POLYTAT/DNA and CTATC/DNA were visualized
by AFM (Fig. 5). Both CTATC and POLYTAT

Fig. 5. Morphology of the polyplexes as determined by AFM. The
morphology was evaluated for (a) plasmid DNA (image size 1�1Am2 and z-range=1.5 nm), (b) CTATC/DNA (image size 4�4 Am2
and z-range=15 nm), and (c,d) POLYTAT/DNA formed in water
using X=3. POLYTAT/DNA polyplexes were visualized (c) in air
(image size 2�2 Am2 and z-range=15 nm) or (d) in a solution using
the liquid cell (image size 2�2 Am2 and z-range=50 nm).
Fig. 4. Molecular parameters of TAT-based DNA polyplexes. The
polyplexes were formulated at X=1.2, 2, and 3 using GTATG,
CTATC, and POLYTAT and (a) hydrodynamic diameter, (b) zeta
potential, and (c) apparent molecular weight of the polyplexes were
determined at 25 8C.
D. Soundara Manickam et al. / Journal of Controlled Release 102 (2005) 293–306300
GEN
ED
ELIV
ER
Y
changed the DNA morphology from wormlike chains
to compact particles. Although a distinct subpopula-
tion of rods among the typical spherical particles was
observed in CTATC/DNA polyplexes (Fig. 5b), no
clear difference in the morphology was observed
between CTATC and POLYTAT polyplexes (Fig. 5b
vs. c). The sizes of the rod-like particles were about 4
nm in height, 80 nm in length, and 40 nm in width.
The images in Fig. 5a–c were acquired in a dry state.
To determine the influence of drying on the polyplex
morphology, POLYTAT polyplexes were imaged
directly in the solution using a liquid cell (Fig. 5d).
The results show that sizes and shapes of the spherical
POLYTAT/DNA particles corresponded to those seen
in Fig. 5c (~45 nm in height, ~150 nm in diameter; the
actual diameter is smaller due to tip convolution in
AFM analysis).
3.3. Cytotoxicity of DNA polyplexes
A successful gene delivery carrier should be able
to deliver transcriptionally active gene to the cell
nucleus without negatively affecting normal func-
D. Soundara Manickam et al. / Journal of Controlled Release 102 (2005) 293–306 301
GEN
ED
ELIV
ER
Y
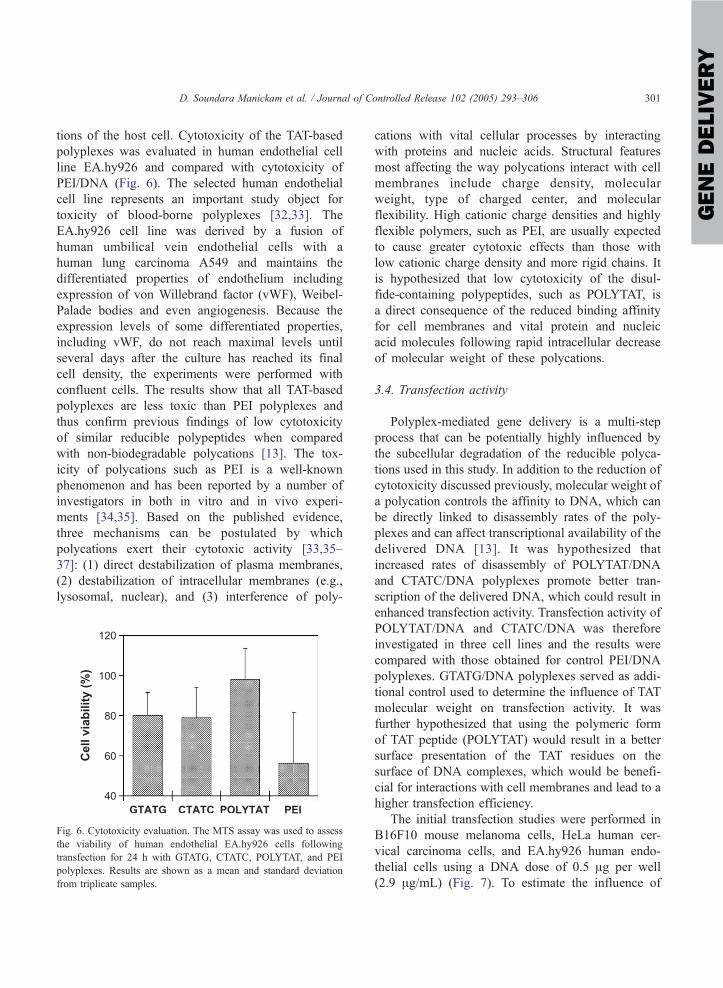
tions of the host cell. Cytotoxicity of the TAT-based
polyplexes was evaluated in human endothelial cell
line EA.hy926 and compared with cytotoxicity of
PEI/DNA (Fig. 6). The selected human endothelial
cell line represents an important study object for
toxicity of blood-borne polyplexes [32,33]. The
EA.hy926 cell line was derived by a fusion of
human umbilical vein endothelial cells with a
human lung carcinoma A549 and maintains the
differentiated properties of endothelium including
expression of von Willebrand factor (vWF), Weibel-
Palade bodies and even angiogenesis. Because the
expression levels of some differentiated properties,
including vWF, do not reach maximal levels until
several days after the culture has reached its final
cell density, the experiments were performed with
confluent cells. The results show that all TAT-based
polyplexes are less toxic than PEI polyplexes and
thus confirm previous findings of low cytotoxicity
of similar reducible polypeptides when compared
with non-biodegradable polycations [13]. The tox-
icity of polycations such as PEI is a well-known
phenomenon and has been reported by a number of
investigators in both in vitro and in vivo experi-
ments [34,35]. Based on the published evidence,
three mechanisms can be postulated by which
polycations exert their cytotoxic activity [33,35–
37]: (1) direct destabilization of plasma membranes,
(2) destabilization of intracellular membranes (e.g.,
lysosomal, nuclear), and (3) interference of poly-
Fig. 6. Cytotoxicity evaluation. The MTS assay was used to assess
the viability of human endothelial EA.hy926 cells following
transfection for 24 h with GTATG, CTATC, POLYTAT, and PEI
polyplexes. Results are shown as a mean and standard deviation
from triplicate samples.
cations with vital cellular processes by interacting
with proteins and nucleic acids. Structural features
most affecting the way polycations interact with cell
membranes include charge density, molecular
weight, type of charged center, and molecular
flexibility. High cationic charge densities and highly
flexible polymers, such as PEI, are usually expected
to cause greater cytotoxic effects than those with
low cationic charge density and more rigid chains. It
is hypothesized that low cytotoxicity of the disul-
fide-containing polypeptides, such as POLYTAT, is
a direct consequence of the reduced binding affinity
for cell membranes and vital protein and nucleic
acid molecules following rapid intracellular decrease
of molecular weight of these polycations.
3.4. Transfection activity
Polyplex-mediated gene delivery is a multi-step
process that can be potentially highly influenced by
the subcellular degradation of the reducible polyca-
tions used in this study. In addition to the reduction of
cytotoxicity discussed previously, molecular weight of
a polycation controls the affinity to DNA, which can
be directly linked to disassembly rates of the poly-
plexes and can affect transcriptional availability of the
delivered DNA [13]. It was hypothesized that
increased rates of disassembly of POLYTAT/DNA
and CTATC/DNA polyplexes promote better tran-
scription of the delivered DNA, which could result in
enhanced transfection activity. Transfection activity of
POLYTAT/DNA and CTATC/DNA was therefore
investigated in three cell lines and the results were
compared with those obtained for control PEI/DNA
polyplexes. GTATG/DNA polyplexes served as addi-
tional control used to determine the influence of TAT
molecular weight on transfection activity. It was
further hypothesized that using the polymeric form
of TAT peptide (POLYTAT) would result in a better
surface presentation of the TAT residues on the
surface of DNA complexes, which would be benefi-
cial for interactions with cell membranes and lead to a
higher transfection efficiency.
The initial transfection studies were performed in
B16F10 mouse melanoma cells, HeLa human cer-
vical carcinoma cells, and EA.hy926 human endo-
thelial cells using a DNA dose of 0.5 Ag per well
(2.9 Ag/mL) (Fig. 7). To estimate the influence of

Fig. 7. Transfection efficiency of TAT- and PEI-based polyplexes in
a panel of cell lines. GTATG, CTATC, and POLYTAT polyplexes
were formulated at X=3, while control PEI polyplexes at X=8. A
single DNA dose (0.5 Ag/well; 2.9 Ag/mL) was used in all three cell
lines. The luciferase reporter gene expression was measured 24 h
after 4-h incubation of the polyplexes with the cells either in the
absence or presence of 100 AM chloroquine. The luciferase
expression in relative light units (RLU) is normalized to mg of
cellular protein and reported as the mean and standard deviation
obtained from triplicate transfections.
D. Soundara Manickam et al. / Journal of Controlled Release 102 (2005) 293–306302
GEN
ED
ELIV
ER
Y
polyplex capability to avoid or to escape endo/
lysosomal pathway on the overall expression of
luciferase reporter gene, all experiments were con-
ducted in the presence and absence of chloroquine.
Polyplex-mediated transfection of fast growing
B16F10 cells resulted in the expected high levels
of luciferase gene expression (Fig. 7a). Almost a
100-fold difference between the transfection activity
of GTATG/DNA and high-molecular-weight forms of
TAT/DNA in the absence of chloroquine confirms
the importance of molecular weight of polycations
for high transgene expression levels. The difference
between low and high-molecular-weight forms of
TAT peptide further increased in transfection experi-
ments conducted in the presence of chloroquine.
Similar effect of molecular weight of TAT peptides
on transfection activity was observed in HeLa cells
(Fig. 7b). Polyplex-mediated transfection of the
endothelial cells EA.hy926 is very inefficient and
the levels of luciferase expression provided by all
three TAT formulations were near-background levels
in the absence of chloroquine (Fig. 7c). The presence
of chloroquine in the transfection media resulted in
more than 10-fold increase of luciferase expression
mediated by CTATC and POLYTAT polyplexes but
no increase was observed for GTATG/DNA. Control
PEI polyplexes proved to be superior to the TAT-
based polyplexes when transfections were conducted
in the absence of chloroquine in all three cell lines.
The data in Fig. 7 clearly demonstrate the benefits of
increasing the molecular weight of TAT peptide for
transfection activity of its DNA polyplexes. As
shown in the previous sections, the polymerization
method of the TAT peptides affects physicochemical
properties of their DNA polyplexes. The transfection
results obtained for CTATC/DNA and POLYTAT/
DNA were, therefore, compared to elucidate if the
polymerization method affects also the transfection
activity. In all three cell lines tested, no significant
differences in luciferase expression levels were
observed for CTATC and POLYTAT polyplexes.
The only statistically significant difference was
observed in transfections of HeLa cells conducted
in the presence of chloroquine, where CTATC/DNA
polyplexes unexpectedly displayed about 150-fold
increased activity compared to POLYTAT/DNA. In
repeated experiments, the increased activity of
CTATC/DNA was confirmed to exhibit an average
16-fold (n=4) higher luciferase expression than
POLYTAT/DNA. Finally, analysis of the influence
of the presence of chloroquine during transfection

Fig. 8. Influence of DNA dose on transfection efficiency. GTATG,
CTATC, and POLYTAT polyplexes were formulated at X=3 and
control PEI polyplexes at X=8. Transfections were performed in
B16F10 mouse melanoma cells (a) in the absence of chloroquine
and (b) in the presence of 100 AM chloroquine. Results are shown
as mean relative light units (RLU) expressed per mg of cellular
proteinFS.D. of three replicate assays.
D. Soundara Manickam et al. / Journal of Controlled Release 102 (2005) 293–306 303
GEN
ED
ELIV
ER
Y
showed that levels of luciferase expression mediated
by both CTATC and POLYTAT polyplexes are
significantly enhanced by chloroquine. On the other
hand, PEI/DNA polyplexes demonstrated only low
levels of potentiation by chloroquine due to their
inherent capability to escape endo/lysosomal traffick-
ing pathway. These results clearly suggest that the
TAT-based polyplexes are most likely internalized by
an endosomal cell uptake and that they do not have
any significant inherent membrane active properties
that would facilitate their direct translocation across
plasma membrane.
The efficiency of polyplex-mediated transfection
is greatly influenced by the DNA dose used. The
transfection studies showed in Fig. 7 were conducted
using a relatively high DNA dose (2.9 Ag/mL) that
could mask potential differences in the activity of
CTATC and POLYTAT polyplexes. Dependence of
luciferase reporter gene expression in B16F10 cells
on the DNA dose for CTATC, POLYTAT, and
control PEI polyplexes was therefore evaluated
(Fig. 8). In the absence of chloroquine (Fig. 8a),
the transfection activity of POLYTAT/DNA poly-
plexes decreases more slowly than that of CTATC/
DNA polyplexes in the DNA concentration range
above 0.5 Ag/mL, but is consistently lower than the
transfection activity of PEI/DNA polyplexes. At the
lowest DNA concentrations tested, the luciferase
expression of all three tested vectors drops signifi-
cantly to almost background levels. Transfecting the
cells in the presence of chloroquine results in high
levels of luciferase expression for all three tested
vectors. Unlike PEI polyplexes, the TAT-based
polyplexes appear to preserve their activity without
a significant decrease of luciferase expression to a
DNA dose of about 0.5 Ag/mL. As a result,
chloroquine potentiation of the TAT-mediated trans-
fection increases with decreasing DNA dose. In case
of CTATC/DNA polyplexes, for example, luciferase
expression increased 980-fold in the presence of
chloroquine at DNA dose 2.9 Ag/mL and 12,400-fold
at the lower DNA dose of 0.3 Ag/mL.
Molar mixing ratio used in polyplex formulation
not only affects the physicochemical properties as
demonstrated in Fig. 4, but more importantly for the
transfection activity, it determines the amount of free
polycation present in the formulations. For polyplexes
such as PEI/DNA that rely on buffering capability of
polycations for their activity, the presence of free
polycation is crucial for their transfection activity both
in vitro and in vivo [38,39]. To permit direct
comparison of the influence of CTATC and POLYTAT
polyplex composition on transfection activity with
that of PEI polyplexes, charge ratio Z was used
instead of molar mixing ratio X (Fig. 9). For both
TAT-based polypeptides tested, the Z and X ratios are
almost equal but they vary significantly for PEI
because of the variation in charge density in the PEI
chain. The compositions of the polyplexes are, there-
fore, expressed in terms of the charge ratio Z to
account for the variation in the content of charged

Fig. 9. Influence of charge ratio on transfection activity. GTATG,
CTATC, POLYTAT, and PEI polyplexes were formulated at various
charge ratios, Z, and transfections were performed in B16F10
mouse melanoma cells (a) in the absence of chloroquine and (b) in
the presence of 100 AM chloroquine using a single DNA dose (0.2
Ag/well; 1.25 Ag/mL). Results are shown as mean relative light units
(RLU) expressed per mg of cellular proteinFS.D. of three replicate
assays.
D. Soundara Manickam et al. / Journal of Controlled Release 102 (2005) 293–306304
GEN
ED
ELIV
ER
Y
groups in PEI. In the calculation of Z, it was assumed
that 45% of total amino groups of PEI is charged at
pH 7 [40]. In the absence of chloroquine, the
transfection activity of PEI/DNA decreased abruptly
at Zb3 while both CTATC/DNA and POLYTAT/DNA
did not show a significant decrease until Zb2 (Fig.
9a). POLYTAT/DNA polyplexes showed higher luci-
ferase expression than CTATC/DNA at all charge ratio
tested. The low DNA dose used in this transfection
experiment (0.2 Ag/well; 1.25 Ag/mL) disadvantaged
PEI polyplexes that require a critical mass of PEI for
efficient buffering function. Addition of chloroquine
to the transfection resulted in the expected increase of
luciferase expression for all three vectors tested (Fig.
9b). Similar to transfection conducted in the absence
of chloroquine, activity of PEI polyplexes exhibited a
strong dependence on the presence of free PEI and
decreased significantly as the charge ratio decreased.
In contrast, only a limited decrease in luciferase
expression was observed for POLYTAT and CTATC
polyplexes. Indeed, a 90-fold higher transfection
activity was measured for POLYTAT polyplexes at
the lowest Z tested compared with PEI polyplexes.
Large chloroquine potentiation effect was observed
for TAT polyplexes (7,400-fold increase in case of
CTATC polyplexes), again suggesting inefficient
cytoplasmic delivery of these vectors. The less
pronounced dependence of transfection activity of
TAT-based polyplexes on charge ratio used suggests
less significant reliance, if any at all, on the free
polycation than in case of PEI polyplexes. Higher
luciferase reporter gene expression observed for the
TAT-based polyplexes compared with PEI polyplexes
(Fig. 9b) is likely to be at least partly a result of
efficient intracellular degradation of TAT polypep-
tides and subsequent improved transcriptional avail-
ability of DNA. Better performance of POLYTAT
polyplexes is likely due to better surface presentation
of the TAT residues than in case of CTATC
polyplexes and therefore more efficient interactions
with cell membranes.
The central hypothesis of this study was that
using the polymeric form of the TAT peptide
(POLYTAT) will permit better surface presentation
of the TAT residues and that this results in a more
efficient cellular delivery of DNA due to TAT
translocating capability. In addition to the protein
and particle transduction properties of TAT peptide, a
nuclear localization function of the TAT peptides was
recently reported that can further enhance the trans-
fection activity of their DNA complexes [41].
Cellular uptake of TAT and other so-called protein
transduction domains have been ascribed in the
literature to an energy- and receptor-independent
mechanism that does not involve endocytosis and is
capable to deliver even large cargo [18,19]. These
studies were recently revisited and the mechanism of
cellular uptake reevaluated. The current evidence
therefore shows that the cell internalization of TAT
peptides and other protein transduction domains is an
energy-dependent process involving classical adsorp-

D. Soundara Manickam et al. / Journal of Controlled Release 102 (2005) 293–306 305
GEN
ED
ELIV
ER
Y
tive endocytosis [21–23]. The data presented in this
study fully confirm the current mechanism of TAT
internalization as documented by the strong depend-
ence of transfection activity of TAT-based polyplexes
on the presence of chloroquine, which suggests
endocytic cell uptake of these vectors. As suggested
by the very low luciferase expression in slowly
dividing endothelial cells, the data also do not seem
to confirm previous findings that the TAT nuclear
localization sequence is involved in enhancing gene
transfer [41].
4. Conclusion
Biodegradable, high-molecular-weight polycation
based on TAT peptide has been successfully synthe-
sized and its physicochemical properties, cytotoxicity,
and transfection activity evaluated. Physicochemical
properties of TAT-based polyplexes were affected by
the molecular weight and method of polymerization
of the TAT peptide. All TAT-based DNA polyplexes
exhibited reduced cytotoxicity when compared with
control PEI polyplexes. Evaluation of transfection
activity confirmed the importance of high molecular
weight of polycations for efficient transgene expres-
sion and suggested a contribution of the polymer-
ization method to the overall efficiency of the cellular
gene delivery process mediated by the TAT polypep-
tides. Higher luciferase reporter gene expression
observed for the TAT-based polyplexes compared
with the PEI polyplexes under selected experimental
conditions is likely to be, at least partly, a result of
efficient intracellular degradation of TAT polypep-
tides and subsequently improved transcriptional avail-
ability of DNA. TAT-based polyplexes appear to be
internalized by adsorptive endocytosis and their
transfection activity is strongly dependent on the
presence of chloroquine. Overall, it is expected that
reducible polycations based on bioactive peptides will
evolve into versatile carriers of nucleic acids that
display low cytotoxicity and will prove especially
beneficial in cases that require surface presentation of
membrane-active or cell-specific targeting peptides or
in development of non-toxic alternatives to PEI that
require a presence of large quantities of free high-
molecular-weight polycations to effectively buffer
endosomal pH.
Acknowledgment
This work was supported by AACP New Inves-
tigators Program from The American Foundation for
Pharmaceutical Education, by National Science Foun-
dation (CTS-0221586), and by Wayne State Univer-
sity Research Grant.
References
[1] M. Ogris, R.C. Carlisle, T. Bettinger, L.W. Seymour, Melittin
enables efficient vesicular escape and enhanced nuclear access
of nonviral gene delivery vectors, J. Biol. Chem. 276 (50)
(2001) 47550–47555.
[2] P. Erbacher, J.-S. Remy, J.-P. Behr, Gene transfer with
synthetic virus-like particles via the integrin-mediated endo-
cytosis pathway, Gene Ther. 6 (1) (1999) 138–145.
[3] M.A. Zanta, P. Belguise-Valladier, J.P. Behr, Gene delivery:
a single nuclear localization signal peptide is sufficient to carry
DNA to the cell nucleus, Proc. Natl. Acad. Sci. U. S. A. 96 (1)
(1999) 91–96.
[4] C. Plank, K. Mechtler, F.C. Szoka, E. Wagner, Activation of
the complement system by synthetic DNA complexes: a
potential barrier for intravenous gene delivery, Hum. Gene
Ther. 7 (12) (1996) 1437–1446.
[5] J.G. Elferink, Cytolytic effect of polylysine on rabbit
polymorphonuclear leukocytes, Inflammation 9 (3) (1985)
321–331.
[6] T. Reschel, C. Konak, D. Oupicky, L.W. Seymour, K. Ulbrich,
Physical properties and in vitro transfection efficiency of gene
delivery vectors based on complexes of DNA with synthetic
polycations, J. Control. Release 81 (1–2) (2002) 201–217.
[7] D.L. McKenzie, K.Y. Kwok, K.G. Rice, A potent new class of
reductively activated peptide gene delivery agents, J. Biol.
Chem. 275 (14) (2000) 9970–9977.
[8] K.H. Bremner, L.W. Seymour, A. Logan, M.L. Read, Factors
influencing the ability of nuclear localization sequence
peptides to enhance nonviral gene delivery, Bioconjug. Chem.
15 (1) (2004) 152–161.
[9] A. Eguchi, T. Akuta, H. Okuyama, T. Senda, H. Yokoi, H.
Inokuchi, S. Fujita, T. Hayakawa, K. Takeda, M. Hasegawa,
M. Nakanishi, Protein transduction domain of HIV-1 Tat
protein promotes efficient delivery of DNA into mammalian
cells, J. Biol. Chem. 276 (28) (2001) 26204–26210.
[10] D. Oupicky, A.L. Parker, L.W. Seymour, Laterally stabilized
complexes of DNA with linear reducible polycations: strategy
for triggered intracellular activation of DNA delivery vectors,
J. Am. Chem. Soc. 124 (1) (2002) 8–9.
[11] D.L. McKenzie, E. Smiley, K.Y. Kwok, K.G. Rice, Low
molecular weight disulfide cross-linking peptides as nonviral
gene delivery carriers, Bioconjug. Chem. 11 (6) (2000)
901–909.
[12] V.S. Trubetskoy, V.G. Budker, L.J. Hanson, P.M. Slattum, J.A.
Wolff, J.E. Hagstrom, Self-assembly of DNA–polymer com-

D. Soundara Manickam et al. / Journal of Controlled Release 102 (2005) 293–306306
GEN
ED
ELIV
ER
Y
plexes using template polymerization, Nucleic Acids Res. 26
(18) (1998) 4178–4185.
[13] M.L. Read, K.H. Bremner, D. Oupicky, N.K. Green, P.F.
Searle, L.W. Seymour, Vectors based on reducible polycations
facilitate intracellular release of nucleic acids, J. Gene Med. 5
(3) (2003) 232–245.
[14] M. Balakirev, G. Schoehn, J. Chroboczek, Lipoic acid-derived
amphiphiles for redox-controlled DNA delivery, Chem. Biol. 7
(10) (2000) 813–819.
[15] G. Saito, J.A. Swanson, K.D. Lee, Drug delivery strategy
utilizing conjugation via reversible disulfide linkages: role and
site of cellular reducing activities, Adv. Drug Deliv. Rev. 55
(2) (2003) 199–215.
[16] D. Oupicky, V. Diwadkar, Stimuli-responsive gene delivery
vectors, Curr. Opin. Mol. Ther. 5 (4) (2003) 345–350.
[17] S.R. Schwarze, K.A. Hruska, S.F. Dowdy, Protein trans-
duction: unrestricted delivery into all cells? Trends Cell Biol.
10 (7) (2000) 290–295.
[18] E. Vives, P. Brodin, B. Lebleu, A truncated HIV-1 Tat protein
basic domain rapidly translocates through the plasma mem-
brane and accumulates in the cell nucleus, J. Biol. Chem. 272
(25) (1997) 16010–16017.
[19] M. Lewin, N. Carlesso, C.H. Tung, X.W. Tang, D. Cory, D.T.
Scadden, R. Weissleder, Tat peptide-derivatized magnetic
nanoparticles allow in vivo tracking and recovery of progen-
itor cells, Nat. Biotechnol. 18 (4) (2000) 410–414.
[20] V.P. Torchilin, R. Rammohan, V. Weissig, T.S. Levchenko,
TAT peptide on the surface of liposomes affords their efficient
intracellular delivery even at low temperature and in the
presence of metabolic inhibitors, Proc. Natl. Acad. Sci. U. S. A.
98 (15) (2001) 8786–8791.
[21] J.A. Leifert, S. Harkins, J.L. Whitton, Full-length proteins
attached to the HIV tat protein transduction domain are neither
transduced between cells, nor exhibit enhanced immunoge-
nicity, Gene Ther. 9 (21) (2002) 1422–1428.
[22] J.P. Richard, K. Melikov, E. Vives, C. Ramos, B. Verbeure,
M.J. Gait, L.V. Chernomordik, B. Lebleu, Cell-penetrating
peptides. A reevaluation of the mechanism of cellular uptake,
J. Biol. Chem. 278 (1) (2003) 585–590.
[23] G. Drin, S. Cottin, E. Blanc, A.R. Rees, J. Temsamani, Studies
on the internalization mechanism of cationic cell-penetrating
peptides, J. Biol. Chem. 278 (33) (2003) 31192–31201.
[24] J.S. Wadia, R.V. Stan, S.F. Dowdy, Transducible TAT-HA
fusogenic peptide enhances escape of TAT-fusion proteins after
lipid raft macropinocytosis, Nat. Med. 10 (3) (2004) 310–315.
[25] H. Dautzenberg, A. Zintchenko, C. Konak, T. Reschel, V.
Subr, K. Ulbrich, Polycationic graft copolymers as carriers for
oligonucleotide delivery. Complexes of oligonucleotides with
polycationic graft copolymers, Langmuir 17 (10) (2001)
3096–3102.
[26] D. Oupicky, R.C. Carlisle, L.W. Seymour, Triggered intra-
cellular activation of disulfide crosslinked polyelectrolyte gene
delivery complexes with extended systemic circulation in vivo,
Gene Ther. 8 (9) (2001) 713–724.
[27] M. Ogris, P. Steinlein, M. Kursa, K. Mechtler, R. Kircheis, E.
Wagner, The size of DNA/transferrin-PEI complexes is an
important factor for gene expression in cultured cells, Gene
Ther. 5 (10) (1998) 1425–1433.
[28] H. Dautzenberg, J. Kriz, Response of polyelectrolyte com-
plexes to subsequent addition of salts with different cations,
Langmuir 19 (13) (2003) 5204–5211.
[29] H. Dautzenberg, Y.B. Gao, M. Hahn, Formation, structure, and
temperature behavior of polyelectrolyte complexes between
ionically modified thermosensitive polymers, Langmuir 16
(23) (2000) 9070–9081.
[30] D. Oupicky, C. Konak, K. Ulbrich, DNA complexes with
block and graft copolymers of N-(2-hydroxypropyl) meth-
acrylamide and 2-(trimethylammonio)ethyl methacrylate, J.
Biomat. Sci., Polym. Ed. 10 (5) (1999) 573–590.
[31] D. Oupicky, C. Konak, K. Ulbrich, M.A. Wolfert, L.W.
Seymour, DNA delivery systems based on complexes of DNA
with synthetic polycations and their copolymers, J. Control.
Release 65 (1–2) (2000) 149–171.
[32] D.M.L. Morgan, V.L. Larvin, J.D. Pearson, Biochemical-
characterization of polycation-induced cyto-toxicity to human
vascular endothelial-cells, J. Cell. Sci. 94 (3) (1989) 553–559.
[33] W.T. Godbey, K.K. Wu, A.G. Mikos, Poly(ethylenimine)-
mediated gene delivery affects endothelial cell function and
viability, Biomaterials 22 (5) (2001) 471–480.
[34] P. Chollet, M.C. Favrot, A. Hurbin, J.L. Coll, Side-effects of a
systemic injection of linear polyethylenimine–DNA com-
plexes, J. Gene Med. 4 (1) (2002) 84–91.
[35] D. Fischer, Y. Li, B. Ahlemeyer, J. Krieglstein, T. Kissel, In
vitro cytotoxicity testing of polycations: influence of polymer
structure on cell viability and hemolysis, Biomaterials 24 (7)
(2003) 1121–1131.
[36] D. Leroy, N. Schmid, J.P. Behr, O. Filhol, S. Pares, J. Garin,
J.J. Bourgarit, E.M. Chambaz, C. Cochet, Direct identification
of a polyamine binding domain on the regulatory subunit of
the protein-kinase casein kinase-2 by photoaffinity-labeling, J.
Biol. Chem. 270 (29) (1995) 17400–17406.
[37] S. Choksakulnimitr, S. Masuda, H. Tokuda, Y. Takakura,
M. Hashida, In-vitro cytotoxicity of macromolecules in
different cell-culture systems, J. Control. Release 34 (3)
(1995) 233–241.
[38] S. Boeckle, K. Gersdorff, S. Piepen, C. Culmsee, E. Wagner,
M. Ogris, Purification of polyethylenimine polyplexes high-
lights the role of free polycations in gene transfer, J. Gene
Med. 6 (10) (2004) 1102–1111.
[39] S.M. Zou, P. Erbacher, J.S. Remy, J.P. Behr, Systemic linear
polyethylenimine (L-PEI)-mediated gene delivery in the
mouse, J. Gene Med. 2 (2) (2000) 128–134.
[40] T.K. Bronich, H.K. Nguyen, A. Eisenberg, A.V. Kabanov,
Recognition of DNA topology in reactions between plasmid
DNA and cationic copolymers, J. Am. Chem. Soc. 122 (35)
(2000) 8339–8343.
[41] C. Rudolph, C. Plank, J. Lausier, U. Schillinger, R.H. Muller,
J. Rosenecker, Oligomers of the arginine-rich motif of the
HIV-1 TAT protein are capable of transferring plasmid DNA
into cells, J. Biol. Chem. 278 (13) (2003) 11411–11418.

















![TAT-902S [1 650] TAT- 1 600 102S F] TAT-312V TAT-322V ...TAT-902S [1 650] TAT- 1 600 102S F] TAT-312V TAT-322V TAT-332S 1/2 1/2 1/2 1/2 1/2 1/2 I OOOX420 IOOOX500 1200X500 TAT-1 52S](https://static.fdocuments.net/doc/165x107/6125a0cefb88a6479b4afa46/tat-902s-1-650-tat-1-600-102s-f-tat-312v-tat-322v-tat-902s-1-650-tat-.jpg)
