
Influence of some Trichoderma species on photosyntesis … M. Alexandru.pdf · 2013-08-27 ·...
12
Romanian Biotechnological Letters Vol. 18, No.4, 2013 Copyright © 2013 University of Bucharest Printed in Romania. All rights reserved ORIGINAL PAPER Romanian Biotechnological Letters, Vol. 18, No. 4, 2013 8499 Influence of some Trichoderma species on photosyntesis intensity and pigments in tomatoes Received for publication, February 1, 2013 Accepted, April 14, 2013 MIOARA ALEXANDRU 1* , DANIELA LAZĂR 2 , MIHAELA ENE 1 and TATIANA EUGENIA ŞESAN 2 1 National Institute for Physics and Nuclear Engineering “Horia Hulubei”, Department IRASM, Microbiological Laboratory, Romania 2 University of Bucharest, Faculty of Biology, Department of Botany and Microbiology, Romania *Corresponding author: Mioara Alexandru, National Institute for Physics and Nuclear Engineering “Horia Hulubei”, Department IRASM, Microbiological Laboratory, 30 Reactorului, Magurele, Ilfov, 077125, e-mail: [email protected] Abstract Trichoderma spp. are effective biocontrol agents against plant pathogens and they also enhance systemic resistance to plant diseases and overall plant growth. Root colonization with Trichoderma spp. induces changes in physiological processes like photosyntesis or other metabolism processes. The objective of this study was to analyze the influence of Trichoderma spp. on the intensity of photosynthesis and on the assimilatory pigment content in tomato plants treated with 6 species of Trichoderma: T. harzianum, T. koningii, T. longibrachiatum, T. tomentosum, T. virens (c.n. T. deliquescens), T. viride and plants treated with the pathogen Botrytis cinerea alone and the pathogen followed by treatment with Trichoderma spp. The results showed that the tested strains of Trichoderma were able to significantly increase the photosynthesis intensity in the leaves of tomato plantlets. Also, the photosynthesis intensity of plantlets infected with B. cinerea followed by inoculation with T. harzianum was higher than for the non-treated plantlets (control). Differences between assimilatory pigments content of tomato plants treated with various Trichoderma species were insignificant but a large variation of the chlorophyll a content was observed between the tomato plants treated with the two tested strains of the T. harzianum (MUCL 15569 and MUCL 29416) was recorded. Key words: Trichoderma spp., Botrytis cinerea, tomatoes, photosynthesis, assimilatory pigments. Abbreviations: CFU - colony-forming unit; KGy – kiloGray; MUCL - Mycothèque de l’Université Catholique de Louvain, Belgium; c.n. – current name of a fungus after Index Fungorum 42; A = Absorbance; SS - sum of squares; df – degrees of freedom; MS – mean squares; F – calculated F test; P-value – Probability; Fcrit – critical value of F test. Introduction Development of sustainable agriculture has been marked as one of the major strategies to reduce the harmful effects on the environment as a result of indiscriminate use of pesticides on agricultural crops [2]. The possible options include combinations of a gradual reduction in the use of pesticides and fertilizers and an increase in the application of biological control systems. One key aspect of sustainable agriculture is the reduction in agrochemical inputs with a gradual shift to alternative crop protection strategies, such as cultural practices, organic amendments, resistant or tolerant crop varieties and biocontrol systems [20].
Transcript of Influence of some Trichoderma species on photosyntesis … M. Alexandru.pdf · 2013-08-27 ·...

Romanian Biotechnological Letters Vol. 18, No.4, 2013 Copyright © 2013 University of Bucharest Printed in Romania. All rights reserved
ORIGINAL PAPER
Romanian Biotechnological Letters, Vol. 18, No. 4, 2013 8499
Influence of some Trichoderma species on photosyntesis intensity and pigments in tomatoes
Received for publication, February 1, 2013
Accepted, April 14, 2013
MIOARA ALEXANDRU1*, DANIELA LAZĂR2, MIHAELA ENE1 and TATIANA EUGENIA ŞESAN2 1National Institute for Physics and Nuclear Engineering “Horia Hulubei”, Department IRASM, Microbiological Laboratory, Romania 2University of Bucharest, Faculty of Biology, Department of Botany and Microbiology, Romania *Corresponding author: Mioara Alexandru, National Institute for Physics and Nuclear Engineering “Horia Hulubei”, Department IRASM, Microbiological Laboratory, 30 Reactorului, Magurele, Ilfov, 077125, e-mail: [email protected]
Abstract
Trichoderma spp. are effective biocontrol agents against plant pathogens and they also enhance
systemic resistance to plant diseases and overall plant growth. Root colonization with Trichoderma spp. induces changes in physiological processes like photosyntesis or other metabolism processes.
The objective of this study was to analyze the influence of Trichoderma spp. on the intensity of photosynthesis and on the assimilatory pigment content in tomato plants treated with 6 species of Trichoderma: T. harzianum, T. koningii, T. longibrachiatum, T. tomentosum, T. virens (c.n. T. deliquescens), T. viride and plants treated with the pathogen Botrytis cinerea alone and the pathogen followed by treatment with Trichoderma spp.
The results showed that the tested strains of Trichoderma were able to significantly increase the photosynthesis intensity in the leaves of tomato plantlets. Also, the photosynthesis intensity of plantlets infected with B. cinerea followed by inoculation with T. harzianum was higher than for the non-treated plantlets (control).
Differences between assimilatory pigments content of tomato plants treated with various Trichoderma species were insignificant but a large variation of the chlorophyll a content was observed between the tomato plants treated with the two tested strains of the T. harzianum (MUCL 15569 and MUCL 29416) was recorded.
Key words: Trichoderma spp., Botrytis cinerea, tomatoes, photosynthesis, assimilatory pigments.
Abbreviations: CFU - colony-forming unit; KGy – kiloGray; MUCL - Mycothèque de l’Université Catholique de Louvain, Belgium; c.n. – current name of a fungus after Index Fungorum 42; A = Absorbance; SS - sum of squares; df – degrees of freedom; MS – mean squares; F – calculated F test; P-value – Probability; Fcrit – critical value of F test.
Introduction Development of sustainable agriculture has been marked as one of the major strategies
to reduce the harmful effects on the environment as a result of indiscriminate use of pesticides on agricultural crops [2]. The possible options include combinations of a gradual reduction in the use of pesticides and fertilizers and an increase in the application of biological control systems. One key aspect of sustainable agriculture is the reduction in agrochemical inputs with a gradual shift to alternative crop protection strategies, such as cultural practices, organic amendments, resistant or tolerant crop varieties and biocontrol systems [20].

MIOARA ALEXANDRU, DANIELA LAZĂR, MIHAELA ENE and TATIANA EUGENIA ŞESAN
8500 Romanian Biotechnological Letters, Vol. 18, No. 4, 2013
The genus Trichoderma, commonly present in all types of soils 11, is being continuously studied for its role in stimulating plant development and healthiness status since its first report as a parasite of other soil fungi 33, 34. Trichoderma is involved in biocontrol of plant diseases and in plant growth promotion 1, 5, 9, 12, 16, 17, 33, 34, 37, 39 in increasing of plant nutrient uptake 14, it is used as a fertilizer 9 or for maintenance of plant greenness for a long time by inducing high photosynthetic rates 10. Recently, Trichoderma spp. have been reported as plant symbionts 8, inducing systemic resistance to plant pathogens by reducing disease symptoms and increasing the expression of defence-related genes 8, 15-17, 25. Some Trichoderma strains have the abilities to improve photosynthetic efficiency and respiratory activity through their ability to reprogram plant gene expression 24, 35.
Botrytis cinerea Pers. is a plant necrotrophic fungal pathogen that colonizes senescent or dead plant tissues and causes gray mold, the first important damaging plant disease in Europe 43. B. cinerea attacks a broad host range of food crops, including tomato (Lycopersicon esculentum Mill.), potato (Solanum tuberosum L.), grapevine (Vitis vinifera L.), strawberry (Fragaria spp), causing important economic losses, either at pre- or postharvest stages [19, 23]. The virulence factors for B. cinerea pathogenicity on different hosts have been described, such as: phytoalexin detoxification, cell wall–degrading enzymes, toxins, and several genes involved in fungus signalling [22, 31].
The present study was conducted to investigate the changes in physiological aspects (photosynthesis and pigments content in leaves) of plants treated with different Trichoderma species and B. cinerea.
Materials and methods
Tomato plants (L. esculentum), variety San Pedro 778, were used in this study. They were maintained for 35 days, at 25-28o C, under a natural photoperiod alternance, between February and April 2012.
Soil substrate was prepared by mixing 1 parts of sand with 3 part of soil, pH 5-7 (Universal peat substrate), followed by sterilization with gamma radiations. The gamma irradiation was performed in an industrial plant, with a 60Co source at the exposure of an absorbed dose of 25 kGy. The treatment aimed to eliminate the nematodes and competitive fungi. Around 100 ml of substrate was added in each pot.
As biocontrol agents were used 7 strains belonging to 6 species of Trichoderma, supplied by the Mycothèque de l’Université Catholique de Louvain (MUCL) - Belgium 41 as follows: Trichoderma harzianum Rifai - MUCL 15569; T. harzianum - MUCL 29416; T. koningii Oudem. - MUCL 42800; T. longibrachiatum Rifai - MUCL 20461; T. viride Pers. - MUCL 7769; T. tomentosum Bissett - MUCL 44900; T. virens (J.H. Mill, Giddens A.A. Foster) Arx - MUCL 34687, c.n. T. deliquescens (Sopp.) Jaklisch.
Each Trichoderma species and the pathogen-test B. cinerea (from the Faculty of Biology, Department of Botany and Microbiology) were plated onto Potato dextrose agar (PDA, Merck) plates and incubated at 26 ± 1oC for seven days until extensive sporulation had occurred. Spores from each isolate were harvested by flooding each Petri with 5-10 ml sterile water with 0.05% Tween 80 and carefully scrape of conidia with sterile glass beads. The chosen suspension had for each Trichoderma species a final spore concentration of 107 – 108 conidia per millilitre and 105-106 CFU / ml for B. cinerea.
Four experimental variants, among them three fungal treatment experimental variants and one control (untreated) were performed in the rhizosphere, at different developmental stages: (1) Treatment with Trichoderma spp. (20 ml suspension from each

Influence of some Trichoderma species on photosyntesis intensity and pigments in tomatoes
Romanian Biotechnological Letters, Vol. 18, No. 4, 2013 8501
isolate/pot) applied simultaneously with seeding; (2) Infection of young plantlets of 14 days old with B.cinerea (20 ml suspension/pot), followed by a treatment with T. harzianum MUCL 15569 (20 ml suspension/pot), after a supplementary 7 days after infection; (3) Infection of young plantlets of 14 days old B. cinerea alone and (4) Control (untreated plants).
Four to five tomato seeds were planted into gamma sterilized substrate. All plants were grown in a protected space, under natural light conditions, at 25 to 28°C; all pots were light protected and were watered every 48 h with the same amount of sterile water.
The plants have been analyzed after 35 days for the intensity of photosynthesis and content of pigments in the leaves. After this analysis, fresh roots were harvested from the plants treated with Trichoderma spp. and B. cinerea, cleaned in tap running water, rinsed in distilled water and drained on sterile Watman paper. Half of the roots, randomly chosen, were put apart for microscopy and the second half of roots from plants treated with Trichoderma spp. were immediately deepen in liquid nitrogen and crushed in a mortar, in the presence of the extraction buffer, for DNA extraction.
A DNA extraction kit from Metabion was used, following the manufacturer instructions. DNA presence and quality was checked spectrophotometrically. Conventional PCR reaction was performed, using a Promega Go-Taq – Gren Master Mix. Primer sequences used for Trichoderma genus detection were as follows [38]: TGP4 (Fw): CTCCCAAACCCAATGTGAAC, TGP4 (Rw): GCGAGTGTGCAAACTACTG. Electrophoretic migration in 1,5% agarose gel reveled amplicon size comparing to a 100 bp ladder.
For microscopically analysis fresh roots were subjected to a clearing in KOH solution in 10% (w/v) KOH at 90o C for 10 min, in a water bath, followed by a whitening step in peroxide solution (5%), rinsing and acidifying with few drops of 2N HCl. A staining solution of classical trypan-blue staining protocols for fungi were applied on root [3]. Microscopic observations were conducted in a Zeiss Axio Imager D1.m Microscope, on squash preparations. Pictures were taken with a Canon MCD 20 camera and processed with AxioVision Rel. 4.6 software.
The photosynthesis rate was investigated with Warburg method [4, 18, 21, 26-30, 36] using pieces of known surface (expressed in dm2) cut from leaves of the tested plants. Each leaf fragment included a portion from mid rib which in the lower part was prolonged in an “artificial petiole”. The Warburg buffer solution number 6 (50 ml Na2CO3 0.1 M + 50 ml NaHCO3 0.1M) was used and the leaf fragment was put with the ”artificial petiole” in 0.3 ml of water in the central cylinder of reaction flask [26].
A spectrophotometric method with a spectrophotometer CECIL 1020 was used to determine the content of chlorophyll and carotenoid pigments in leaves. The absorbance of the solutions extracted with 100% acetone were measured at 661.6, 644.8 and 470 nm and the calculation of the content in a, b chlorophylls and carotenoids was performed according to Lichtenthaler [13]; the values were expressed in mg/g dry matter (DM).
The formulas used for the calculus of the content in chlorophyll and carotenoid pigments were:
Ca = 11.24*A661.6 – 2.04*A644.8 Cb = 20.13*A644.8 - 4,19*A661.6 Cx+c = (1000*A470 - 1.90*Ca - 63.14*Cb)/214 Carotenoid and xanthophylls pigments were evaluated quantitatively together. The differences between treatments were analysed with ANOVA method using the
Microsoft EXCEL add-on statistical facilities. For graphical comparison it was included the standard deviation.

MIOARA ALEXANDRU, DANIELA LAZĂR, MIHAELA ENE and TATIANA EUGENIA ŞESAN
8502 Romanian Biotechnological Letters, Vol. 18, No. 4, 2013
Results and discussion
Trichoderma isolates have important economical implications on plant development such as shortening of the plant growth period, as well as improving plant vigour to overcome biotic and/or abiotic stresses, resulting in increase plant productivity and yields. We showed here that Trichoderma-treated plants present longer roots and higher shoots and an improved plant vigour. Our results are in accordance with those of Yedidia et al. [14] and Harman et al. [8]. The aerial part of the plants presented significantly enhanced growth characteristics, mainly in plants treated with Trichoderma fungi, in comparison with non-treated (control) plants. Plants of 28 days old treated with Trichoderma strains, especially with T. longibrachiatum MUCL 20461, T. harzianum MUCL 15569, T. koningii MUCL 42800, showed improved plant vigour, they looked healthier and with more vigorous growth than the plant from the control (Fig. 1). Also, plants infected with pathogen B. cinerea alone, respectively, with pathogen followed by treatment with T. harzianum MUCL 15569 showed a good growth compared to control, but showed typical symptoms for infection with B. cinerea: white spots on leaf surfaces or whole plant wilted (Fig. 2 a, b).
Fig. 1. Tomato plants (T) treatead with Trichoderma spp. (right) comparing with Control (non-treated; left)

Influence of some Trichoderma species on photosyntesis intensity and pigments in tomatoes
Romanian Biotechnological Letters, Vol. 18, No. 4, 2013 8503
Confirmation of colonization with Trichoderma spp. and B. cinerea was performed by
optical microscopic observations, which proved a successful infection for all Trichoderma strains and for B. cinerea. Infectious hyphae and intraradical spores were observed (Fig. 3-8).
Fig. 2. Tomato plants: (a) treatead with Botrytis cinerea and Trichoderma harzianum MUCL 15569 (right) comparing with Control (non-treated; left) and (b) treatead with Botrytis cinerea (right) comparing with Control (left)
a b
Fig. 3. T. harzianum spores in tomato roots cortex
Fig. 4. T. harzianum hypha in tomato roots
Fig. 5. T. viride hypha around tomato roots
Fig. 6. T. longibrachiatum hypha in tomato roots cortex

MIOARA ALEXANDRU, DANIELA LAZĂR, MIHAELA ENE and TATIANA EUGENIA ŞESAN
8504 Romanian Biotechnological Letters, Vol. 18, No. 4, 2013
PCR amplification for confirmation
Primers were designed to target a group of certain Trichoderma strains and an amplicon of 465 bp was expected for each species after migration in agarose gel. The presence of the DNA fragment of this size confirmed the colonization with Trichoderma fungus in each root replicate (Fig. 9).
Fig. 9. Electrophoregram of PCR amplified fragment from DNA of Trichoderma infected roots (lanes: 1 – T. harzianum MUCL 15569; 2 – T. harzianum MUCL 29416; 3 – T. tomentosum MUCL 44900; 4 – T. viride MUCL 7769; 5 – T. virens MUCL 34687; 6 – T. koningii MUCL 42800; 7 – T. longibrachiatum MUCL 20461; M – molecular weight marker (100 bp). Arrow: amplicon of expected size – 465 bp).
1 2 M 3 4 5 6 M 7
Fig. 7. Botrytis cinerea spores in tomato infected roots with B. cinerea alone (200x)
Fig. 8. Botrytis cinerea hypha in tomato roots infected with B.cinerea alone (200x)
Influence of some Trichoderma species on photosyntesis intensity and pigments in tomatoes
Romanian Biotechnological Letters, Vol. 18, No. 4, 2013 8505
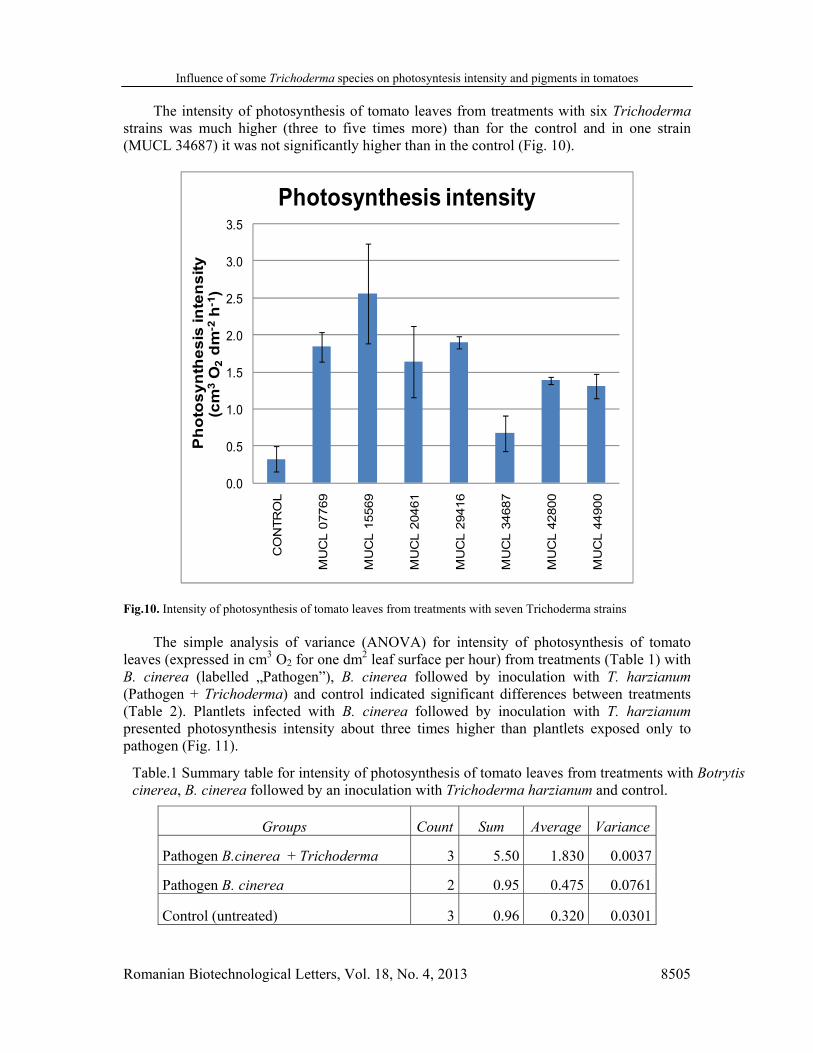
The intensity of photosynthesis of tomato leaves from treatments with six Trichoderma strains was much higher (three to five times more) than for the control and in one strain (MUCL 34687) it was not significantly higher than in the control (Fig. 10).
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
CO
NTR
OL
MU
CL 0
7769
MU
CL 1
5569
MU
CL 2
0461
MU
CL 2
9416
MU
CL 3
4687
MU
CL 4
2800
MU
CL 4
4900
Ph
otos
ynth
esis
inte
nsi
ty(c
m3
O2
dm
-2h-
1 )Photosynthesis intensity
Fig.10. Intensity of photosynthesis of tomato leaves from treatments with seven Trichoderma strains The simple analysis of variance (ANOVA) for intensity of photosynthesis of tomato
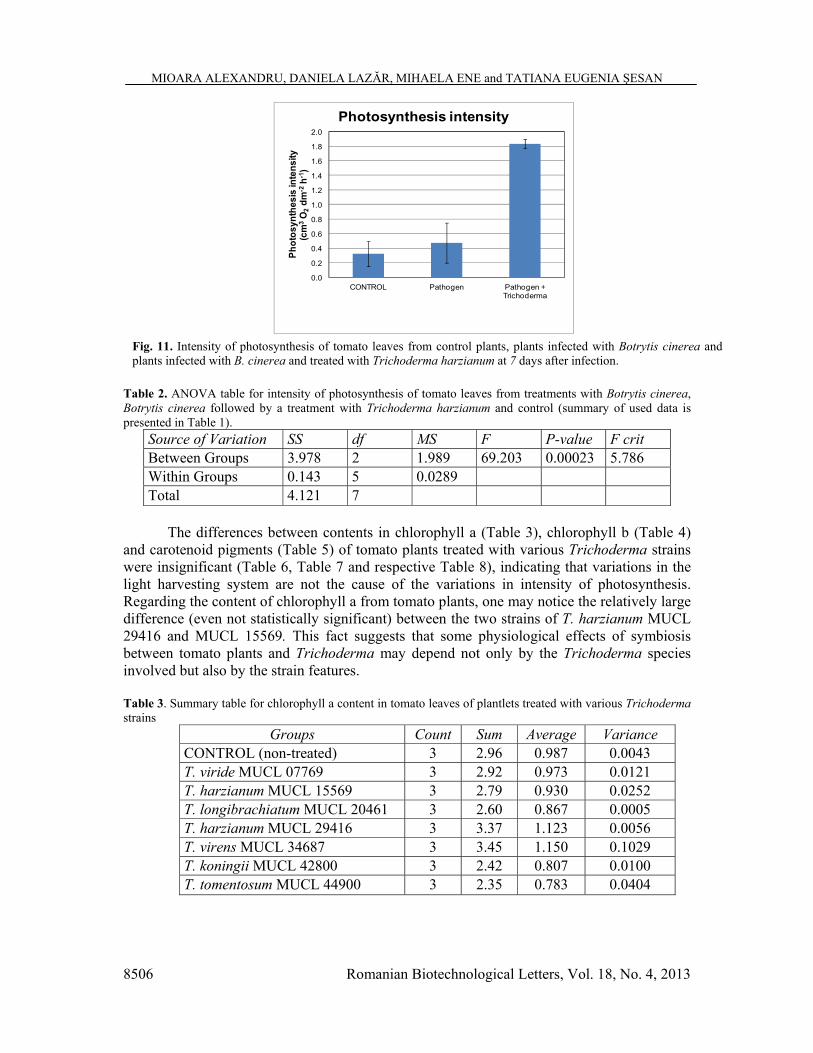
leaves (expressed in cm3 O2 for one dm2 leaf surface per hour) from treatments (Table 1) with B. cinerea (labelled „Pathogen”), B. cinerea followed by inoculation with T. harzianum (Pathogen + Trichoderma) and control indicated significant differences between treatments (Table 2). Plantlets infected with B. cinerea followed by inoculation with T. harzianum presented photosynthesis intensity about three times higher than plantlets exposed only to pathogen (Fig. 11).
Groups Count Sum Average Variance
Pathogen B.cinerea + Trichoderma 3 5.50 1.830 0.0037
Pathogen B. cinerea 2 0.95 0.475 0.0761
Control (untreated) 3 0.96 0.320 0.0301
Table.1 Summary table for intensity of photosynthesis of tomato leaves from treatments with Botrytis cinerea, B. cinerea followed by an inoculation with Trichoderma harzianum and control.
MIOARA ALEXANDRU, DANIELA LAZĂR, MIHAELA ENE and TATIANA EUGENIA ŞESAN
8506 Romanian Biotechnological Letters, Vol. 18, No. 4, 2013
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
1.6
1.8
2.0
CONTROL Pathogen Pathogen + Trichoderma
Phot
osyn
thes
is in
tens
ity(c
m3
O2
dm-2
h-1)
Photosynthesis intensity
Table 2. ANOVA table for intensity of photosynthesis of tomato leaves from treatments with Botrytis cinerea, Botrytis cinerea followed by a treatment with Trichoderma harzianum and control (summary of used data is presented in Table 1).
Source of Variation SS df MS F P-value F crit Between Groups 3.978 2 1.989 69.203 0.00023 5.786 Within Groups 0.143 5 0.0289 Total 4.121 7
The differences between contents in chlorophyll a (Table 3), chlorophyll b (Table 4)
and carotenoid pigments (Table 5) of tomato plants treated with various Trichoderma strains were insignificant (Table 6, Table 7 and respective Table 8), indicating that variations in the light harvesting system are not the cause of the variations in intensity of photosynthesis. Regarding the content of chlorophyll a from tomato plants, one may notice the relatively large difference (even not statistically significant) between the two strains of T. harzianum MUCL 29416 and MUCL 15569. This fact suggests that some physiological effects of symbiosis between tomato plants and Trichoderma may depend not only by the Trichoderma species involved but also by the strain features. Table 3. Summary table for chlorophyll a content in tomato leaves of plantlets treated with various Trichoderma strains
Groups Count Sum Average Variance CONTROL (non-treated) 3 2.96 0.987 0.0043 T. viride MUCL 07769 3 2.92 0.973 0.0121 T. harzianum MUCL 15569 3 2.79 0.930 0.0252 T. longibrachiatum MUCL 20461 3 2.60 0.867 0.0005 T. harzianum MUCL 29416 3 3.37 1.123 0.0056 T. virens MUCL 34687 3 3.45 1.150 0.1029 T. koningii MUCL 42800 3 2.42 0.807 0.0100 T. tomentosum MUCL 44900 3 2.35 0.783 0.0404
Fig. 11. Intensity of photosynthesis of tomato leaves from control plants, plants infected with Botrytis cinerea and plants infected with B. cinerea and treated with Trichoderma harzianum at 7 days after infection.

Influence of some Trichoderma species on photosyntesis intensity and pigments in tomatoes
Romanian Biotechnological Letters, Vol. 18, No. 4, 2013 8507
Table 4. Summary table for chlorophyll b content in tomato leaves of plantlets treated with various Trichoderma strains
Table 5. Summary table for carotenoid pigments content in tomato leaves of plantlets treated with various Trichoderma strains
Groups Count Sum Average Variance CONTROL (non-treated) 3 0.59 0.197 0.000 T. viride MUCL 07769 3 0.55 0.183 0.002 T. harzianum MUCL 15569 3 2.1 0.700 0.843 T. longibrachiatum MUCL 20461 3 0.61 0.203 0.001 T. harzianum MUCL 29416 3 2.14 0.713 0.775 T. virens MUCL 34687 3 0.64 0.213 0.003 T. koningii MUCL 42800 3 0.47 0.157 0.001 T. tomentosum MUCL 44900 3 0.47 0.157 0.001
Table 6. ANOVA table for chlorophyll a content in tomato leaves of plantlets treated with various Trichoderma strains
Source of Variation SS df MS F P-value F crit Between Groups 0.383 7 0.055 2.176 0.094 2.657 Within Groups 0.402 16 0.025 Total 0.785 23
Table 7. ANOVA table for chlorophyll b content in tomato leaves of plantlets treated with various Trichoderma strains
Source of Variation SS Df MS F P-value F crit Between Groups 0.197 7 0.028 1.061 0.433 2.707 Within Groups 0.399 15 0.027 Total 0.596 22
Table 8. ANOVA table for carotenoid pigments content in tomato leaves of plantlets treated with various Trichoderma strains
Source of Variation SS Df MS F P-value F crit Between Groups 1.24 7 0.18 0.867 0.552 2.657 Within Groups 3.25 16 0.203 Total 4.49 23
Groups Count Sum Average Variance CONTROL (non-treated) 3 1.51 0.503 0.021 T. viride MUCL 07769 3 1.87 0.623 0.029 T. harzianum MUCL 15569 3 1.75 0.583 0.058 T. longibrachiatum MUCL 20461 3 1.96 0.653 0.039 T. harzianum MUCL 29416 3 2.44 0.813 0.003 T. virens MUCL 34687 2 1.04 0.520 0.039 T. koningii MUCL 42800 3 1.62 0.540 0.001 T. tomentosum MUCL 44900 3 1.85 0.617 0.028

MIOARA ALEXANDRU, DANIELA LAZĂR, MIHAELA ENE and TATIANA EUGENIA ŞESAN
8508 Romanian Biotechnological Letters, Vol. 18, No. 4, 2013
Increased photosynthesis intensity of plantlets infected with B. cinerea followed by inoculation with T. harzianum suggests a higher assimilate availability. That may support the recovery effort of tomato plants under beneficial influence of T. harzianum, but further studies are required for confirmation of the effect and understanding of involved mechanisms.
All tested strains of Trichoderma were able to significantly increase the photosynthesis intensity in the leaves of tomato plantlets. The highest effect on photosynthesis intensity was noticed in treatments with the strain T. harzianum MUCL 15569 and the lowest in the treatments with strain T. virens MUCL 34687.
The increased photosynthetic efficiency could be explained by the fungal improvement of the redox status of the plant. When plants are under stress, the content of reactive oxygen species may increase to toxic concentrations. Several pathways in plants convert oxidized glutathione and ascorbate to the reduced form [32]. The Trichoderma strains enhance the activity of these pathways, in part by enhancing the expression of genes encoding the component enzymes [6, 7]. Enhancement of these pathways in chloroplasts would logically be expected to increase photosynthetic efficiency by reducing damage by the superoxide anion and other reactive species involved in photosynthesis. Conclusions
Photosynthesis intensity of plantlets infected with B. cinerea followed by inoculation
with T. harzianum was higher than in the control and infected plantlets. All of tested strains of Trichoderma were able to significantly increase the photosynthesis intensity in the leaves of tomato plantlets.
Differences between assimilatory pigments (chlorophylls a and b and carotenoids) content of tomato plants treated with various Trichoderma species were insignificant. A large difference in chlorophyll a content has been observed between the tomato plants treated with the two tested strains of the T. harzianum (MUCL 15569 and MUCL 29416).
Trichoderma spp. have important implications on plant growth and some physiological aspects such shortening the plant growth period, as well as improving plant vigour to overcome biotic and/or abiotic stresses, resulting in increase plant productivity. Acknowledgements This work was supported by CNCSIS - UEFISCSU, project number PNII - IDEI 1931 /2008.
References
1. A. TRONSMO, L.G. HJELORD, Biological control with Trichoderma species. In: Plant Microbe Interactions and Biological Control. USA: Marcel Dekker Inc: 111-125 (1998).
2. B. SCHIPPERS, R.J. SCHEFFER, B.J.J. LUGTENBERG, P.J. WEISBEEK, Biocoating of seed with plant growth promoting rhizobacteria to improve plant establishment. Outlook Agr., 24: 179–185 (1995).
3. C. GRACE, D.P. STRIBLEY, A safer procedure for routine staining of vesicular-arbuscular mycorrhizal fungi. Mycological Research, 95: 1160-1162 (1991).
4. D. BURK, S. HENDRICKS, M. KORZENOVSKY, V. SCHOCKEN, O. WARBURG, The maximum efficiency of photosynthesis: A rediscovery. Science, New Series, 110(2853): 225-229 (1949).
5. D.L. LINDSEY, R. BAKER, Effect of certain fungi on dwarf tomatoes grown under gnotobiotic conditions. Phytopathology, 57: 1262–1263 (1967).
6. F. MASTOURI, T. BJORKMAN, G.E. HARMAN, Seed treatments with Trichoderma harzianum alleviate biotic, abiotic and physiological stresses in germinating seeds and seedlings. Phytopathology, 100: 1213–1221 (2010).

Influence of some Trichoderma species on photosyntesis intensity and pigments in tomatoes
Romanian Biotechnological Letters, Vol. 18, No. 4, 2013 8509
7. F. MASTOURI, Use of Trichoderma spp. to improve plant performance under abiotic stress. PhD thesis, Cornell University, Ithaca, NY, USA (2010).
8. G. E. HARMAN, C. R. HOWELL, A.VITERBO, I. CHET & M. LORITO, Trichoderma species – opportunistic, avirulent plant symbionts. Nature Rev. Microbiol., 2: 43-56 (2004).
9. G. E. HARMAN, Myths and dogmas of biocontrol. Changes in perceptions derived from research on Trichoderma harzianum T22. Plant Dis., 84, 377–393 (2000).
10. G. E. HARMAN, Overview of mechanisms and uses of Trichoderma spp. Phytopathology, 96: 190-4 (2006).
11. G. J. SAMUELS, Trichoderma: a review of biology and systematics of the genus. Mycological Research, 100: 923-935 (1996).
12. G.G. PAPAVIZAS, Trichoderma and Gliocladium: Biology, ecology and potential for biocontrol. Ann. Rev. Phytopathol., 23: 23 – 54 (1985).
13. H.K. LICHTENTHALER, Chlorophylls and carotenoids: pigments of photosynthetic biomembranes. Methods in enzymology, 148: 350-382 (1987).
14. I. YEDIDIA, A. K. SRIVASTVA, Y. KAPULNIK, I. CHET, Effect of Trichoderma harzianum on microelement concentrations and increased growth of cucumber plants. Plant Soil, 235:235-42 (2001).
15. I. YEDIDIA, M. SHORESH, Z. KEREM, N. BENHAMOU, Y. KAPULNIK, I. CHET, Concomitant induction of systemic resistance to Pseudomonas syringae pv. lachrymans in cucumber by Trichoderma asperellum (T-203) and accumulation of phytoalexins. Appl Environ Microbiol., 69: 7343–7353 (2003).
16. I. YEDIDIA, N. BENHAMOU, I. CHET, Induction of defense responses in cucumber plants (Cucumis sativus L.) by the biocontrol agent Trichoderma harzianum. Appl Environ Microbiol, 65: 1061–1070 (1999).
17. I. YEDIDIA, N. BENHAMOU, Y. KAPULNIK, I. CHET, Induction and accumulation of PR proteins activity during early stages of root colonization by the mycoparasite Trichoderma harzianum strain T-203. Plant Physiol Biochem., 38: 863–873 (2000).
18. J.H.C. SMITH, A. BENITEZ, Chlorophylls: Analysis in plant material. In Modern Methods of Plant Analysis (K. Paech and M.V. Tracey, eds.) Springer, Berlin: 142-196 (1955).
19. J.R. COLEY-SMITH, K. VERHOEFF, W.R. JARVIS, The Biology of Botrytis. London: Academic Press, 181–218 (1980).
20. J.W. KLOEPPER, G.W. ZEHNDER, S. TUZUN, J. F. MURPHY, G. WEI, C. YAO, G. RAUPACH, Toward agricultural implementation of PGPR-mediated induced systemic resistance against crop pests. In: W. Tang, R.J. Cook, and A. Rovira, eds. Advances in Biological Control of Plant Diseases. China Agricultural University Press, Beijing: 165-174 (1996).
21. JESUS ROBERTO MILLAN-ALMARAZ, RAMON GERARDO GUEVARA-GONZALEZ, RENE DE JESUS ROMERO-TRONCOSO, ROQUE ALFREDO OSORNIO-RIOS, IRINEO TORRES-PACHECO, Advantages and disadvantages on photosynthesis measurement techniques: A review. African Journal of Biotechnology, 8 (25): 7340-7349 (2009).
22. M. CHOQUER, E. FOURNIER, C. KUNZ, C. LEVIS, J. PRADIER, A. SIMON, M. VIAUD, Botrytis cinerea virulence factors: New insights into a necrotrophic and polyphageous pathogen. FEMS Microbiol. Lett., 277: 1–10 (2007).
23. M. El OIRDI, K. BOUARAB, Plant signalling components EDS1 and SGT1 enhance disease caused by the necrotrophic pathogen Botrytis cinerea. New Phytol., 175: 131–139 (2007).
24. M. SHORESH, G. HARMAN, F. MASTOURI, Induced Systemic Resistance and Plant Responses to Fungal Biocontrol Agents. Annual Review of Phytopathology, 48: 21-43 (2010).
25. M. SHORESH, I. YEDIDIA, I.CHET, Involvement of jasmonic acid/ethylene signaling pathway in the systemic resistance induced in cucumber by Trichoderma asperellum T203. Phytopathology, 95: 76–84 (2005).
26. O. BOLDOR, O. RAIANU, M. TRIFU. Fiziologia plantelor, lucrări practice. Ed. Didactică şi Pedagogică, Bucureşti, 1983, 177-183.
27. O. WARBURG, G. KRIPPAHL, A. LEHMAN, Chlorophyll catalysis and Einstein’s law of photochemical equivalence in photosynthesis. Am. J. Bot., 56(9): 961-971 (1969).
28. O. WARBURG, About the speed of the photochemical decomposition of carbon acid in living cells. Biochem. Z., 100: 230-270 (1919).
29. O. WARBURG, Assimilatory quotient and photochemical yield. Am. J. Bot., 35(3): 194-204 (1948). 30. O. WARBURG, Photosynthesis. Science, New Series, 128(3315): 68-73 (1958). 31. R. GONZÁLEZ-LAMOTHE, G. MITCHELL, M. GATTUSO, M. DIARRA, F. MALOUIN, K.
BOUARAB, Plant antimicrobial agents and their effects on plant and human pathogens. Int. J. Mol. Sci., 10:3400–3419 (2009).

MIOARA ALEXANDRU, DANIELA LAZĂR, MIHAELA ENE and TATIANA EUGENIA ŞESAN
8510 Romanian Biotechnological Letters, Vol. 18, No. 4, 2013
32. R. MITTLER, Oxidative stress, antioxidants and stress tolerance. Trends in Plant Science, 7: 405–410 (2002).
33. R. WEINDLING, Experimental consideration of the mold toxins of Gliocladium and Trichoderma. Phytopathology, 31: 991–1003 (1941).
34. R. WEINDLING. Trichoderma lignorum as a parasite of other soil fungi. Phytopathology, 22: 837-45 (1932).
35. R.J. GEIDER, B.A. OSBORNE, Respiration and microalgal growth: a review of the quantitative relationship between dark respiration and growth. New Phytol., 112(3): 327-341 (1989).
36. S. HUNT, Measurements of photosynthesis and respiration in plants. Physiol. Plant., 117: 314-325 (2003). 37. T.E. ŞESAN, F. OANCEA. Trichoderma viride Pers. – Experimental Model for Biological and
Biotechnological Investigations of Mycromyceta with Importance in Obtaining Plant Protection Bioproducts. J. Plant Develop., 17: 49-62 (2010).
38. T.G. KIM, G.R. KNUDSEN, Quantitative Real-Time PCR effectively detects and quantifies colonization of sclerotia of Sclerotinia sclerotiorum by Trichoderma spp. Appl. Soil Ecol. Doi: 10.1016/j.apsoil.2008.03.013 (2008).
39. Y.-C. CHANG, Y.-C. CHANG, R. BAKER, O. KLEIFELD, I. CHET, Increased growth of plants in the presence of the biological control agent Trichoderma harzianum. Plant Dis. 70, 145–148 (1986).
40. Z. ŠESTÁK, Determination of chlorophylls a and b. In Plant Photosynthetic Production: Manual of Methods (Z. Šesták J., Catsky and P.G. Jarvis, eds.), Dr. W. Junk Publishers, The Hague: 672-701 (1971).
41. * * * http://bccm.belspo.be/mucl.php 42. * * * www.IndexFungorum.org 43. * * * news.Agrapages.com, cited in Sănătatea Plantelor (Plant's Health), 176(1): 29 (2013).


















