
Inferring sockeye salmon (Oncorhynchus nerka...
12
Inferring sockeye salmon (Oncorhynchus nerka) population dynamics and water quality changes in a stained nursery lake over the past ~500 years Irene Gregory-Eaves, Bruce P. Finney, Marianne S.V. Douglas, and John P. Smol Abstract: Historical and paleolimnological studies have demonstrated that environmental changes in the North Pacific can strongly affect sockeye salmon (Oncorhynchus nerka) abundances. Whether these marine shifts would be influential on sockeye salmon from all lake types, however, has not yet been studied. This study represents the first paleolimnolo- gical analysis of past sockeye salmon population dynamics in a stained nursery lake (Packers Lake, Alaska). We adopted a multiproxy approach to determine whether salmon-derived nutrients (inferred from δ 15 N) would be available for algal uptake (inferred from the diatom species responses) in this stained lake, as high concentrations of humics and iron are known to sequester phosphorus. The strong degree of coherency between δ 15 N and diatoms, however, suggests that salmon-derived nutrients were bioavailable and enhanced productivity. Overall, our indicators responded to changes in sockeye salmon abundances and volcanic ashfalls over the past ~500 years. In a section of the core unaffected by tephras (AD ~1770–1882), our record suggests that the number of sockeye salmon spawners fluctuated widely. Com- parison of temporal shifts in inferred sockeye salmon abundances from Packers Lake with other clearwater nursery lakes reveals a broadly consistent pattern, likely influenced by past climatic changes. Résumé : Des études historiques et paléolimnologiques ont démontré que les changements environnementaux dans le Pacifique nord peuvent affecter l’abondance des saumons rouges (Oncorhynchus nerka). On n’a pas, toutefois, encore déterminé si ces changements marins affectent les saumons rouges provenant de tous les types de lacs. Notre étude est la première analyse paléolimnologique de la dynamique des populations de saumons rouges du passé dans un lac d’engraissement du saumon à eau fortement colorée, le lac Packers en Alaska. Une méthodologie impliquant de nom- breux indicateurs substituts nous permet de déterminer si les nutriments provenant des saumons (estimés d’après δ 15 N) sont disponibles pour l’absorption par les algues (estimée d’après les réactions des espèces de diatomées) dans ce lac à eau colorée, parce qu’on sait que les concentrations élevées de substances humiques et de fer emprisonnent le phos- phore. Le fort degré de cohérence entre δ 15 N et les diatomées, cependant, laisse croire que les nutriments provenant des saumons sont biodisponibles et qu’ils stimulent la productivité. Dans leur ensemble, nos indicateurs ont réagi aux chan- gements d’abondance des saumons rouges et des pluies de cendres volcaniques au cours d’environ les derniers 500 ans. Dans une section de carotte non affectée par les téphras (AD ~1770–1882), nos données indiquent que le nombre de saumons rouges reproducteurs a fluctué considérablement. La comparaison des changements temporels dans les abon- dances estimées de saumons rouges du lac Packers et d’autres lacs d’engraissement à eau claire montre des patterns en grande partie comparables, qui sont vraisemblablement causés par des changements climatiques au cours du passé. [Traduit par la Rédaction] Gregory-Eaves et al. 1246 Introduction A century’s worth of commercial catch data for Pacific salmon (Oncorhynchus spp.) in Alaska has revealed dra- matic variability, with the annual harvest fluctuating between ~25 million and ~200 million fish (Wertheimer 1997). Nu- merous analyses of Pacific salmon historical records have detected significant decadal-scale shifts and suggest that cli- matic fluctuations, manifested strongly in the marine envi- ronment (e.g., the Pacific Decadal Oscillation), explain these patterns (e.g., Hare and Mantua 2000 and papers cited therein). Paleolimnological studies extend sockeye salmon (Oncorhynchus nerka) records back hundreds to thousands of years (i.e., prior to extensive anthropogenic disturbances) Can. J. Fish. Aquat. Sci. 61: 1235–1246 (2004) doi: 10.1139/F04-071 © 2004 NRC Canada 1235 Received 10 December 2002. Accepted 16 March 2004. Published on the NRC Research Press Web site at http://cjfas.nrc.ca on 17 September 2004. J17235 I. Gregory-Eaves 1,2 and J.P. Smol. Paleoecological Environmental Assessment and Research Laboratory (P.E.A.R.L.), Department of Biology, Queen’s University, Kingston, ON K7L 3N6, Canada. B.P. Finney. Institute of Marine Science, University of Alaska Fairbanks, Fairbanks, AK 99775-7220, USA. M.S.V. Douglas. Paleoenvironmental Assessment Laboratory (P.A.L.), Department of Geology, University of Toronto, 22 Russell Street, Toronto, ON M5S 3B1, Canada. 1 Corresponding author (e-mail: [email protected]). 2 Present address: Department of Biology, McGill University, 1205 Docteur Penfield, Montreal, QC H3A 1B1, Canada.
Transcript of Inferring sockeye salmon (Oncorhynchus nerka...

Inferring sockeye salmon (Oncorhynchus nerka)population dynamics and water quality changes ina stained nursery lake over the past ~500 years
Irene Gregory-Eaves, Bruce P. Finney, Marianne S.V. Douglas, and John P. Smol
Abstract: Historical and paleolimnological studies have demonstrated that environmental changes in the North Pacificcan strongly affect sockeye salmon (Oncorhynchus nerka) abundances. Whether these marine shifts would be influentialon sockeye salmon from all lake types, however, has not yet been studied. This study represents the first paleolimnolo-gical analysis of past sockeye salmon population dynamics in a stained nursery lake (Packers Lake, Alaska). Weadopted a multiproxy approach to determine whether salmon-derived nutrients (inferred from δ15N) would be availablefor algal uptake (inferred from the diatom species responses) in this stained lake, as high concentrations of humics andiron are known to sequester phosphorus. The strong degree of coherency between δ15N and diatoms, however, suggeststhat salmon-derived nutrients were bioavailable and enhanced productivity. Overall, our indicators responded to changesin sockeye salmon abundances and volcanic ashfalls over the past ~500 years. In a section of the core unaffected bytephras (AD ~1770–1882), our record suggests that the number of sockeye salmon spawners fluctuated widely. Com-parison of temporal shifts in inferred sockeye salmon abundances from Packers Lake with other clearwater nurserylakes reveals a broadly consistent pattern, likely influenced by past climatic changes.
Résumé : Des études historiques et paléolimnologiques ont démontré que les changements environnementaux dans lePacifique nord peuvent affecter l’abondance des saumons rouges (Oncorhynchus nerka). On n’a pas, toutefois, encoredéterminé si ces changements marins affectent les saumons rouges provenant de tous les types de lacs. Notre étude estla première analyse paléolimnologique de la dynamique des populations de saumons rouges du passé dans un lacd’engraissement du saumon à eau fortement colorée, le lac Packers en Alaska. Une méthodologie impliquant de nom-breux indicateurs substituts nous permet de déterminer si les nutriments provenant des saumons (estimés d’après δ15N)sont disponibles pour l’absorption par les algues (estimée d’après les réactions des espèces de diatomées) dans ce lac àeau colorée, parce qu’on sait que les concentrations élevées de substances humiques et de fer emprisonnent le phos-phore. Le fort degré de cohérence entre δ15N et les diatomées, cependant, laisse croire que les nutriments provenant dessaumons sont biodisponibles et qu’ils stimulent la productivité. Dans leur ensemble, nos indicateurs ont réagi aux chan-gements d’abondance des saumons rouges et des pluies de cendres volcaniques au cours d’environ les derniers 500 ans.Dans une section de carotte non affectée par les téphras (AD ~1770–1882), nos données indiquent que le nombre desaumons rouges reproducteurs a fluctué considérablement. La comparaison des changements temporels dans les abon-dances estimées de saumons rouges du lac Packers et d’autres lacs d’engraissement à eau claire montre des patterns engrande partie comparables, qui sont vraisemblablement causés par des changements climatiques au cours du passé.
[Traduit par la Rédaction] Gregory-Eaves et al. 1246
Introduction
A century’s worth of commercial catch data for Pacificsalmon (Oncorhynchus spp.) in Alaska has revealed dra-matic variability, with the annual harvest fluctuating between~25 million and ~200 million fish (Wertheimer 1997). Nu-merous analyses of Pacific salmon historical records have
detected significant decadal-scale shifts and suggest that cli-matic fluctuations, manifested strongly in the marine envi-ronment (e.g., the Pacific Decadal Oscillation), explain thesepatterns (e.g., Hare and Mantua 2000 and papers citedtherein). Paleolimnological studies extend sockeye salmon(Oncorhynchus nerka) records back hundreds to thousandsof years (i.e., prior to extensive anthropogenic disturbances)
Can. J. Fish. Aquat. Sci. 61: 1235–1246 (2004) doi: 10.1139/F04-071 © 2004 NRC Canada
1235
Received 10 December 2002. Accepted 16 March 2004. Published on the NRC Research Press Web site at http://cjfas.nrc.ca on17 September 2004.J17235
I. Gregory-Eaves1,2 and J.P. Smol. Paleoecological Environmental Assessment and Research Laboratory (P.E.A.R.L.), Departmentof Biology, Queen’s University, Kingston, ON K7L 3N6, Canada.B.P. Finney. Institute of Marine Science, University of Alaska Fairbanks, Fairbanks, AK 99775-7220, USA.M.S.V. Douglas. Paleoenvironmental Assessment Laboratory (P.A.L.), Department of Geology, University of Toronto,22 Russell Street, Toronto, ON M5S 3B1, Canada.
1Corresponding author (e-mail: [email protected]).2Present address: Department of Biology, McGill University, 1205 Docteur Penfield, Montreal, QC H3A 1B1, Canada.

and support the argument that climatic shifts are key factorsexplaining population dynamics of Pacific salmon (Finney etal. 2000; Gregory-Eaves et al. 2003; Sweetman and Finney2003). Paleolimnological studies from numerous lakes withinthe Kodiak Island and Bristol Bay regions also documentedsynchronous trends in sockeye salmon population dynamicsover the past ~2200 years (Finney et al. 2000, 2002).
Packers Lake is distinct from other sockeye salmon nurs-ery lakes whose sediment records have been studied to date.Packers Lake lies in a different geographic region and it is astained lake. Stained sockeye salmon nursery lakes are notuncommon in Alaska and have been defined as lakes withtrue colour >10 platinum cobalt units and turbidity is <3nephelometric turbidity units (Koenings and Edmundson1991). Stained lakes typically have a shallower euphoticzone than clearwater lakes (i.e., Secchi depths in stainedlakes are, on average, 2 m shallower than in clearwater lakes;Koenings and Edmundson 1991). In addition, elevated con-centrations of dissolved humic substances and iron instained lakes can render phosphorus less available for algaluptake, as these compounds are known to form complexeswith phosphorus (Jones 1998). In particular, Edmundson andCarlson (1998) found that, compared with clearwater lakes,stained Alaskan lakes had chlorophyll a concentrations thatwere 30% lower per unit total phosphorus. Lower productiv-ity, along with warmer temperatures and lower dissolvedoxygen conditions typical of stained lakes, can also limitsockeye salmon smolt production (Koenings et al. 1985).One of the objectives of this project was to evaluate whethersockeye salmon from a stained lake would show productiontrends coherent with those observed regionally in clearwaterlakes or whether local nursery lake conditions would bemore influential in determining sockeye salmon spawnerabundances.
We analysed a suite of paleolimnological proxy indicators(δ15N, diatoms, biogenic silica, and δ13C) to reconstruct pastsockeye salmon abundances and water quality conditions.Sockeye salmon accumulate >95% of their weight in the ma-rine environment and then return to their natal freshwaterenvironment to spawn and die (Burgner 1991). Thus, decom-position of their carcasses can represent a significant deliv-ery of nutrients to the freshwater environment, with up to60% of a lake’s annual phosphorus loading coming fromsalmon carcasses (Koenings and Burkett 1987a). Sedimentδ15N analysis can be used to track these salmon-derived nu-trients, as salmon are significantly enriched in δ15N relativeto terrestrial sources (Mathisen et al. 1988; Kline et al. 1993),and δ15N has been shown to be related to the density ofsockeye salmon spawners (number of spawners per squarekilometre; Finney et al. 2000). Diatoms, a group of algaethat are often well preserved in lake sediments, represent an-other proxy that has been used to infer past changes insalmon-derived nutrient loading, as diatom species assem-blages are very sensitive to shifts in nutrient concentrations(Finney et al. 2000, 2002; Gregory-Eaves et al. 2003). Nev-ertheless, it was not known whether diatoms would trackchanges in salmon-derived nutrients in a stained lake. Ifhumic substances and iron in a stained lake form strongcomplexes with phosphorus, then diatoms might be expectedto show little or no response to changes in salmon-derivednutrients inferred from δ15N.
Materials and methods
Site descriptionPackers Lake (60°28′30′′N, 151°55′30′′W) is located on
Kalgin Island, which is situated within Cook Inlet, Alaska(Fig. 1). This modestly sized sockeye salmon nursery lake(mean depth is 12.2 m and surface area is 2.1 km2) isslightly acidic, mesoeutrophic, and considerably coloured(Table 1) (Mazumder and Edmundson 2002). The dystrophicnature of this lake is likely due to seepage inputs that havecome from boggy vegetation such as sedges, meadow grasses,and Sphagnum spp. (Koenings et al. 1985). The catchmentalso contains some Sitka spruce (Picea sitchensis) and west-ern hemlock (Tsuga heterophylla).
Some nutrients in Packers Lake, however, probably comefrom the decay of sockeye salmon carcasses. Historical dataextending back to 1926 suggest that sockeye salmon escape-ment to Packers Lake has varied considerably, from <100 to100 000 fish (Koenings et al. 1985; Colvin 2001). However,it was not until 1973 that a consistent method for enumerat-ing adults returning to Packers Lake was established. There-fore, some of the variability in escapement predating 1973may be the result of differences in enumerating techniques.Since 1973, the number of returning spawners has beenmonitored by counting the adults that pass a weir at the out-let of Packers Lake. The annual return of sockeye salmonsince 1973 has varied between 168 and 58 010 spawners(Colvin 2001).
The year 1973 also marks the beginning of fisheries en-hancement programs at Packers Lake. In 1973, after 1.4 mil-lion sockeye salmon eggs were collected, the lake was treatedwith rotenone in an attempt to eliminate large populations ofthreespine (Gasterosteus aculeatus) and ninepine stickleback(Pungitius pungitius) (Koenings et al. 1985). Sockeye salmoneggs collected from Packers Lake in 1973 were incubated inthe Kasilof hatchery and reintroduced as fry into PackersLake in 1974 after the lake had detoxified (Koenings et al.1985).
Further manipulations of Packers Lake began in 1983. Itwas believed that more smolts could be produced if the lakewere limed to reduce the concentration of dissolved organicmatter and fertilized to stimulate primary and secondary pro-ductivity, which in turn could provide an enhanced foragebase for sockeye salmon fry (Koenings et al. 1985; Mazum-der and Edmundson 2002). In 1983, 21 tons of lime and3942 kg of nitrogen fertilizer were applied to the northernthird of the lake (Fandrei 1996). This liming treatment hadonly a slight effect on the water colour of Packers Lake, asthe lake is still coloured and experienced little to no changein water clarity (Table 1). From 1984 to 1998, nitrogen–phosphorus fertilizer was added to the northern third ofPackers Lake with the goal of increasing the phosphorusloading by 40% and nitrogen loading by 109% (Kyle et al.1997; Colvin 2001). A program to stock sockeye salmon fryinto Packers Lake was also implemented between 1988 and1997, and an average of 2 800 000 fry were introduced an-nually (i.e., mean between 1988 and 1994; Fandrei 1996).
Core and sediment analysisIn 1998, a 38-cm gravity core was collected from the
northern basin of Packers Lake (Fig. 1) and extruded contin-
© 2004 NRC Canada
1236 Can. J. Fish. Aquat. Sci. Vol. 61, 2004

uously in 0.5-cm intervals using a vertical Glew (1988) ex-truder. Chronostratigraphic techniques used in this studyincluded identifying tephra layers in the core and 210Pb,137Cs, and 14C analyses as described below.
Twenty-two samples were prepared for 210Pb and 137Cscounting following the methods described in Schelske et al.(1994) and analysed in a germanium gamma detector(EG&G Ortec, Oak Ridge, TN 37830, USA). For 14C analy-
© 2004 NRC Canada
Gregory-Eaves et al. 1237
Fig. 1. (a) Map of southern Alaska, USA, showing the location of Packers Lake on Kalgin Island and a nearby tephra reference lake, SkilakLake, on the Kenai Peninsula. Some of the Cook Inlet volcanoes are also highlighted (as triangles on the map): 1, Mt. Novarupta/Katmai; 2,Mt. Douglas; 3, Mt. St. Augustine; 4, Mt. Iliamna; 5, Mt. Redoubt; 6, Mt. Spurr – Crater Peak; 7, Mt. Hayes. (b) Bathymetric map ofPackers Lake showing the coring location and the portion of the lake that has been fertilized in recent years.
ParameterPrefertilization(1981–1982)
Fertilization withoutstocking (1983–1987)
Fertilization andstocking (1988–1995)
Euphotic zone depth (m) 4.0 (0.5) 4.9 (0.2)Secchi depth (m) 2.9 (0.2) 2.7 (0.1)Turbidity (nephelometric turbidity units) 1.3 (0.2) 1.0 (0.1)Color (platinum cobalt units) 27.0 (1.4)*pH 6.6 (0.4) 6.8 (0.1) 6.7 (0.0)Alkalinity (mg·L–1) 10.3 (1.3) 11.9 (1.0) 8.6 (0.3)Conductivity (µS·cm–1) 39.0 (1.0) 43.8 (0.9) 46.3 (1.3)Iron 720.0 (50.0) 479.8 (48.7) 516.1 (28.3)Total phosphorus (µg·L–1) 16.3 (2.3) 16.0 (1.5) 11.6 (0.7)Total Kjeldahl nitrogen (µg·L–1) 226.5 (0.5) 250.8 (16.2) 273.0 (15.7)Nitrate plus nitrite (µg·L–1) 68.5 (12.5) 82.8 (13.2) 215.0 (21.6)Total nitrogen to total phosphorus ratio (µg·L–1) 41.5 (7.5) 47 (2.8) 95.8 (7.8)Particulate organic carbon (µg·L–1) 271.3 (39.9)*Reactive silica (µg·L–1) 3237.0 (88.3) 3403.6 (74.6)*Chlorophyll a (µg·L–1) 1.4 (0.1) 2.0 (0.5) 1.7 (0.2)Macrozooplankton biomass (mg·m–2) 519 (86) 728 (88) 480 (75)
Note: Surface measurements (1 m) that have been seasonally averaged (May–October) are presented with the standard errors of seasonalmeans given in parentheses. Sources: Koenings et al. (1985), Fandrei (1996), and J. Edmundson (Limnology Lab, Alaska Department ofFish and Game, 43961 Kalifornsky Beach Road, Suite B, Soldotna, AK 99669-8367, USA, unpublished data).
*The 1990–1995 average from Alaska Department of Fish and Game (unpublished data).
Table 1. Physical, chemical, and biological water quality parameters from Packers Lake collected during theprefertilization, fertilization and no stocking, and fertilization and stocking periods.

sis, a spruce (Picea spp.) seed from near the bottom of thecore (36–36.5 cm) was analysed at the Lawrence LivermoreCenter for Accelerator Mass Spectrometry. The 14C date wascalibrated with the CALIB program version 4.3 using the at-mospheric decadal calibration curve (Stuiver et al. 1998).
Tephra layers were first identified using magnetic suscep-tibility analysis determined by employing a Bartington MS2magnetic susceptibility meter (Bartington Instruments Ltd.,Witney, Oxford, UK). Microscopic examination of the highmagnetic susceptibility intervals for the presence of volcanicglass confirmed these layers as tephras. Microprobe analysiswas conducted on volcanic glass from the 24.5-cm intervalusing a scanning electron microprobe quantometre (AppliedResearch Laboratories Ltd., Glendale, Calif.) to measure theoxide content of the glass and identify the source eruption ofthe tephra (Turney and Lowe 2001).
Physical, geochemical, and isotopic analyses conductedon the sediments included water content, biogenic silica,carbon, and nitrogen content, and δ15N and δ13C. Biogenicsilica analyses were performed on subsamples taken from asieved fraction (<420 µm) of the sediment and analysed ac-cording to the methods outlined in Mortlock and Froelich(1989) using a 0.1 mol·L–1 solution of Na2CO3 to dissolvethe biogenic silica. We have expressed our biogenic silica re-sults as percent opal (per unit dry weight), which accountsfor the presence of water in diatomaceous silica.
For all carbon and nitrogen analyses, bulk sediment sub-samples were first treated with 2 mol·L–1 HCl for approxi-mately 24 h to remove carbonates and to ensure that δ13Canalyses were performed only on carbon bound to organicmatter. Sediments were then washed with double-distilledwater four times, freeze-dried, homogenized, and analysedusing a Finnigan Delta Plus mass spectrometer (Bremen,Germany). Isotopic analyses are reported in standard δ nota-tion relative to atmospheric N2 for nitrogen and PeeDee Bel-emnite for carbon. Analytical precision was, on average,±0.2‰ for δ15N and ±0.3‰ for δ13C.
Diatoms and chrysophytes were also identified and enu-merated from 29 intervals in the core. Microfossil slideswere prepared according to standard methods (Battarbee etal. 2001). At least 400 diatom valves from each sample wereidentified and enumerated along parallel transects across slides(Gregory-Eaves et al. 2003). Chrysophyte cysts and scaleswere also enumerated and are expressed as a percentage rel-ative to the number of diatom frustules and chrysophytecysts or scales counted (Smol 1985).
Results and discussion
Core chronology and tephrochronologyNumerous tephra layers were apparent in the Packers Lake
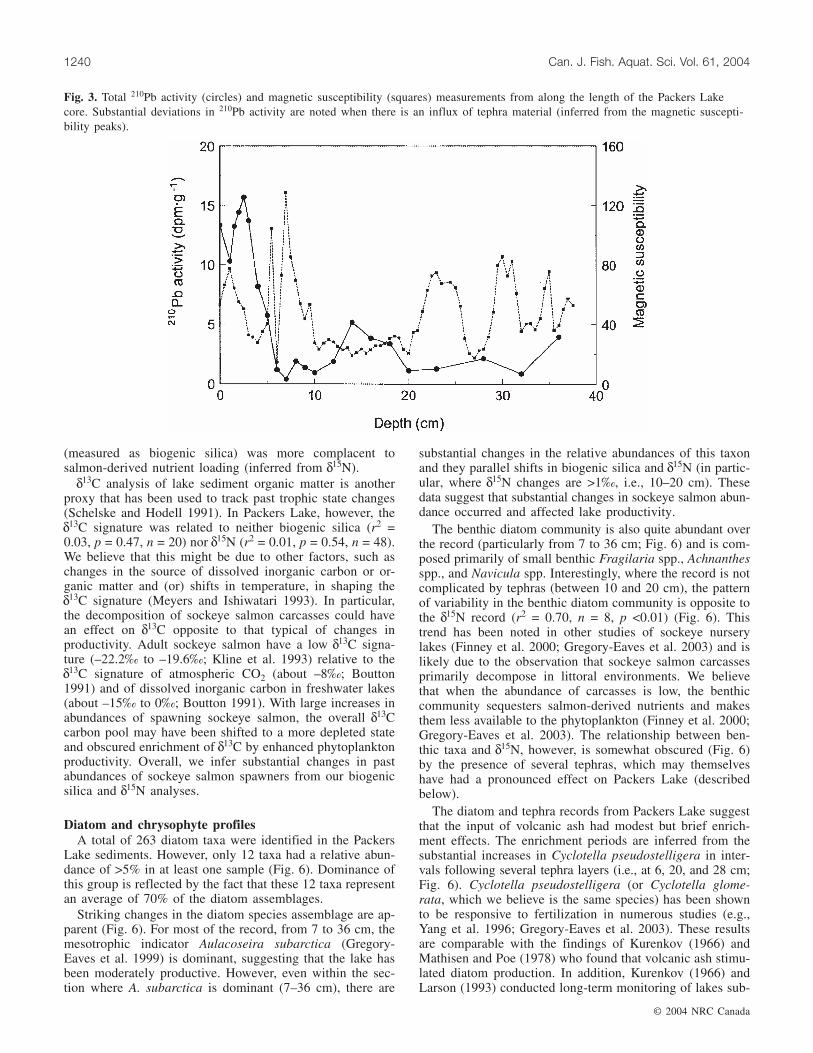
core (Fig. 2) and these layers provided both challenges andopportunities for constructing an age model. The total 210Pbprofile shows significant deviations where tephras are pres-ent, even at depths where we would expect there to be nounsupported 210Pb activity (i.e., only supported 210Pb)(Fig. 3). Using the historical tephras as age markers, wewould expect that sediments below ~10 cm would have littleto no unsupported 210Pb. Therefore, we suggest that the sud-den inputs of ash during a tephra sequence resulted in sub-stantial decreases in both unsupported (where present) and
supported 210Pb activity. The marked change in many of thegeochemical profiles (Fig. 4) supports the idea that tephralayers were deposited quickly and thus sources of 238U andits decay products would have been diluted. For example, asecond tracer in the 238U decay series, 214Pb, also shows de-creases with tephra inputs.
Based on the above interpretation, we believe that back-ground 210Pb levels are reached somewhere between 5 and14 cm at an activity of ~6 dpm·g–1, as all measurementswhere there were no tephras below 14 cm are close to thisnumber. Applying the constant rate of supply model (Appleby2001) to our 210Pb activities, the oldest age generatedthrough this technique was AD 1833 (at a depth of 4.5 cm)(Fig. 5). However, given that the 210Pb profile was compli-cated by tephras and reached background levels close to thetop of the core, we developed additional chronologies usingalternative methods (i.e., other radiometric isotopes andtephrochronology). We believe that most robust chronologyis based on using the following three markers: (i) our coringdate, (ii) the 137Cs peak, and (iii) a basal 14C date. Unlike the210Pb profile, the 137Cs peak, which indicates the height ofatmospheric nuclear weapons testing in 1963 (Appleby 2001),occurred in a section of the core that was not complicated bytephras (i.e., at 3 cm). Our 14C date of 370 ± 40 years BPwas calibrated to a calendar age of AD 1540 (Gregory-Eaves2002). This three-marker age model matches closely with anage model that we developed incorporating the age–depthrelationship of tephra layers (discussed below) and has goodoverall agreement with the 210Pb model (Fig. 5). The dis-crepancy between the older 210Pb ages and our other agemodels could be explained by our inability to clearly definebackground 210Pb activity as well as the larger errors associ-ated with older 210Pb dates.
A third ageing approach is to apply tephrachronology,which has been fairly well established in a paleolimnolo-gical study of a lake ~75 km east from our study site, SkilakLake (Figs. 1 and 2) (Beget et al. 1994). Through our micro-probe analyses, we identified the Mt. Spurr – Crater Peak(~AD 1650; Beget et al. 1994) tephra layer in the PackersLake core (Gregory-Eaves 2002). Knowing the position ofthis tephra layer and the sequence of historical eruptions al-lowed us to infer the sources of other tephras in the core bycomparing the magnetic susceptibility profile from PackersLake with that of Skilak Lake (Fig. 2). Unfortunately, for theSkilak Lake record, only the known historical eruptions (i.e.,Mt. Novarupta/Katmai in 1912, Mt. Redoubt in 1902, andMt. St. Augustine in 1883) and the 14C-dated Mt. St. Augus-tine tephra (~AD 1450) were dated reliably. Dates for allintervening tephras were interpolated. For this reason, wedid not include the position of tephra layers in the develop-ment of our final age model. However, a comparison of ourage model (using only the coring date, the 137Cs peak, andthe 14C date) with the alternative model that also includesthe tephra layers demonstrates a high degree of similarity,where the alternative model falls within the two-sigma rangeof our calibrated 14C date (Fig. 5).
Physical, geochemical, and isotopic dataAs mentioned above, tephra layers in the Packers Lake
core are represented as sharp changes in physical and chemi-cal measures of the sediment. Concurrent with the peaks in
© 2004 NRC Canada
1238 Can. J. Fish. Aquat. Sci. Vol. 61, 2004

magnetic susceptibility are dramatic changes in water, car-bon, and nitrogen content (Fig. 4). In contrast with thesemeasures, however, δ13C and δ15N show little relationship tothe input of tephras (this is especially true for the deviationsthat are >1‰) (Fig. 4). Similarly, the C:N ratio is relativelylow and stable (mean = 10.6 and SD = 1.0), which suggeststhat the organic matter throughout the sediment record isprimarily of aquatic origin.
Over the record, the δ15N profile varies between 4.9‰ and7.9‰. High values in δ15N are greatly enriched comparedwith δ15N values from non-salmon lakes in Alaska, whichdisplay mean δ15N of surface sediments of 1.5 ± 0.9‰ (n =33) (Finney et al. 2000). High sediment δ15N values arelikely due to the relatively high number of sockeye salmonthat return to Packers Lake (Table 2). The density of adultsockeye salmon, the number of spawners per lake surfacearea, at Packers Lake is comparable with that at Karluk andFrazer lakes (Table 2), where we have recorded a strong cor-respondence between paleolimnological indicators of sock-eye salmon, δ15N and diatoms, and historical escapement(Finney et al. 2000; Gregory-Eaves et al. 2003). In addition,
Packers Lake has a relatively slow flushing rate (Table 2),which is likely important in that salmon-derived nutrientscan be incorporated into the lentic food web and ultimatelyinto the lake sediments. Packers, Karluk, and Frazer lakeshave similar water residence times (Table 2).
Biogenic silica analyses of sediment should be able to de-tect past changes in trophic state, as dissolved silica concen-trations in Packers Lake were not found to be limiting foralgal growth, and silica-requiring algae (i.e., diatoms andchrysophytes) dominated the modern phytoplankton assem-blages (Koenings et al. 1985; Kyle 1994). Similar to δ15N,biogenic silica exhibited substantial variation, ranging be-tween 6.2% and 22.7% opal (Fig. 4). Biogenic silica wasalso found to strongly covary with δ15N (r2 = 0.62, p < 0.01,n = 20). This observation supports our conclusion thatsalmon-derived nutrient loading to Packers Lake has beensubstantial. However, in the lower 20 cm of the core wherethere are large tephra layers, biogenic silica shows less vari-ation compared with that recorded in the δ15N profile. It ispossible that during this interval, other algae were moreabundant and thus the production by silica-requiring algae
© 2004 NRC Canada
Gregory-Eaves et al. 1239
Fig. 2. Magnetic susceptibility profiles from (a) Packers Lake and (b) Skilak Lake (data from Skilak Lake digitized from Beget et al.1994). Microprobe analyses confirmed the source volcanoes for tephras in Skilak Lake, whereas microprobe analysis only confirmedthe location of the Mt. Spurr – Crater Peak tephra in the Packers Lake core (in bold). The identification of tephras in the Packers Lakecore is based on matching magnetic susceptibility peaks between the Packers Lake and Skilak Lake profiles. The 1989–1990 Mt. Re-doubt tephra is not present in the Skilak Lake profile because the cores were collected in 1987 and 1988.
(measured as biogenic silica) was more complacent tosalmon-derived nutrient loading (inferred from δ15N).
δ13C analysis of lake sediment organic matter is anotherproxy that has been used to track past trophic state changes(Schelske and Hodell 1991). In Packers Lake, however, theδ13C signature was related to neither biogenic silica (r2 =0.03, p = 0.47, n = 20) nor δ15N (r2 = 0.01, p = 0.54, n = 48).We believe that this might be due to other factors, such aschanges in the source of dissolved inorganic carbon or or-ganic matter and (or) shifts in temperature, in shaping theδ13C signature (Meyers and Ishiwatari 1993). In particular,the decomposition of sockeye salmon carcasses could havean effect on δ13C opposite to that typical of changes inproductivity. Adult sockeye salmon have a low δ13C signa-ture (–22.2‰ to –19.6‰; Kline et al. 1993) relative to theδ13C signature of atmospheric CO2 (about –8‰; Boutton1991) and of dissolved inorganic carbon in freshwater lakes(about –15‰ to 0‰; Boutton 1991). With large increases inabundances of spawning sockeye salmon, the overall δ13Ccarbon pool may have been shifted to a more depleted stateand obscured enrichment of δ13C by enhanced phytoplanktonproductivity. Overall, we infer substantial changes in pastabundances of sockeye salmon spawners from our biogenicsilica and δ15N analyses.
Diatom and chrysophyte profilesA total of 263 diatom taxa were identified in the Packers
Lake sediments. However, only 12 taxa had a relative abun-dance of >5% in at least one sample (Fig. 6). Dominance ofthis group is reflected by the fact that these 12 taxa representan average of 70% of the diatom assemblages.
Striking changes in the diatom species assemblage are ap-parent (Fig. 6). For most of the record, from 7 to 36 cm, themesotrophic indicator Aulacoseira subarctica (Gregory-Eaves et al. 1999) is dominant, suggesting that the lake hasbeen moderately productive. However, even within the sec-tion where A. subarctica is dominant (7–36 cm), there are
substantial changes in the relative abundances of this taxonand they parallel shifts in biogenic silica and δ15N (in partic-ular, where δ15N changes are >1‰, i.e., 10–20 cm). Thesedata suggest that substantial changes in sockeye salmon abun-dance occurred and affected lake productivity.
The benthic diatom community is also quite abundant overthe record (particularly from 7 to 36 cm; Fig. 6) and is com-posed primarily of small benthic Fragilaria spp., Achnanthesspp., and Navicula spp. Interestingly, where the record is notcomplicated by tephras (between 10 and 20 cm), the patternof variability in the benthic diatom community is opposite tothe δ15N record (r2 = 0.70, n = 8, p <0.01) (Fig. 6). Thistrend has been noted in other studies of sockeye nurserylakes (Finney et al. 2000; Gregory-Eaves et al. 2003) and islikely due to the observation that sockeye salmon carcassesprimarily decompose in littoral environments. We believethat when the abundance of carcasses is low, the benthiccommunity sequesters salmon-derived nutrients and makesthem less available to the phytoplankton (Finney et al. 2000;Gregory-Eaves et al. 2003). The relationship between ben-thic taxa and δ15N, however, is somewhat obscured (Fig. 6)by the presence of several tephras, which may themselveshave had a pronounced effect on Packers Lake (describedbelow).
The diatom and tephra records from Packers Lake suggestthat the input of volcanic ash had modest but brief enrich-ment effects. The enrichment periods are inferred from thesubstantial increases in Cyclotella pseudostelligera in inter-vals following several tephra layers (i.e., at 6, 20, and 28 cm;Fig. 6). Cyclotella pseudostelligera (or Cyclotella glome-rata, which we believe is the same species) has been shownto be responsive to fertilization in numerous studies (e.g.,Yang et al. 1996; Gregory-Eaves et al. 2003). These resultsare comparable with the findings of Kurenkov (1966) andMathisen and Poe (1978) who found that volcanic ash stimu-lated diatom production. In addition, Kurenkov (1966) andLarson (1993) conducted long-term monitoring of lakes sub-
© 2004 NRC Canada
1240 Can. J. Fish. Aquat. Sci. Vol. 61, 2004
Fig. 3. Total 210Pb activity (circles) and magnetic susceptibility (squares) measurements from along the length of the Packers Lakecore. Substantial deviations in 210Pb activity are noted when there is an influx of tephra material (inferred from the magnetic suscepti-bility peaks).

jected to the deposition of volcanic ash and reported resultssimilar to ours, i.e., that lake enrichment appears to be ashort-term phenomenon.
Following the tephra layers between 5 and 10 cm, there isa noticeable rise in Aulacoseira distans var. tenella and a
continued decrease in A. subarctica. We infer from thesechanges a decrease in trophic state, as A. distans var. tenellais reported to have a lower total phosphorus optimum thanA. subarctica (Hall and Smol 1996). This inference is alsosupported by the decrease in δ15N and biogenic silica (Fig. 6).
© 2004 NRC Canada
Gregory-Eaves et al. 1241
Fig. 4. Physical, chemical, and isotopic properties of the Packers Lake core: (a) biogenic silica, (b) δ15N, (c) δ13C, (d) carbon content,(e) nitrogen content, (f) water content, and (g) magnetic susceptibility.

The lake fertilization program at Packers Lake (1983–1998) is marked by a large increase in C. pseudostelligera, adecrease in benthic diatoms and chrysophyte cysts, and anincrease in chrysophyte scales. As mentioned above,C. pseudostelligera has previously been shown to be respon-sive to fertilization. However, the dominance of this taxoninstead of the more nutrient-rich indicator A. subarctica (asinferred from its total phosphorus optimum; Gregory-Eaveset al. 1999) suggests that the enrichment caused by the fertil-ization program was not as strong as that evident earlier dur-ing the period of larger salmon returns (inferred from δ15N at~13 cm). The decrease in benthic diatoms is likely due to thefact that the fertilizer was applied to the pelagic zone andthus favored planktonic diatoms (which would cause them tomake up a greater proportion of the overall diatom assem-blage). Chrysophytes typically do well in nutrient-poor wa-ter (Smol 1985), and thus, the decrease in cysts (which are
formed by all chrysophyte taxa) with lake fertilization wasnot unexpected. The increase in scale-bearing chrysophytes(e.g., Mallomonas spp.), although opposite to the general-ized chrysophyte response, is similar to what we observed inFrazer Lake during periods of modest nutrient enrichment(Gregory-Eaves et al. 2003). The dominant chrysophytescales in both the Packers and Frazer lake records wereMallomonas crassisquama and Mallomonas pseudocoro-nata, taxa that are known to increase in abundance withslight nutrient enrichment (Fritz et al. 1993; Zeeb et al.1994). Interestingly, the pattern of increased abundances ofchrysophyte scales with nutrient enrichment at Packers Lakeis also apparent prior to fertilization. At 6 cm in the core,where we inferred a slight enrichment with the sharp rise inC. pseudostelligera, we also recorded a large increase in theabundance of chrysophyte scales, mostly M. pseudocoronataand M. crassisquama (Fig. 6).
In summary, the diatom and chrysophyte communities ofPackers Lake have undergone shifts that can be attributed tochanges in sockeye salmon spawner abundances, volcanicashfalls, and lake management practices. Many of the micro-fossil changes are synchronous with shifts in δ15N and bio-genic silica.
Relative importance of lake and ocean conditions inadult sockeye salmon production
Researchers over the past 10 years have clearly shownthat substantial quantities of salmon-derived nutrients may
© 2004 NRC Canada
1242 Can. J. Fish. Aquat. Sci. Vol. 61, 2004
Fig. 5. Age models developed for the Packers Lake core. The final age model (solid line) was generated by applying a linear fitthrough the following age markers (shaded diamonds): coring date (AD 1998 at 0 cm), the 137Cs peak (1963 at 3 cm), and the cali-brated 14C date (AD 1540 at 36 cm). The broken line represents an alternative age model, which is a linear fit through the coring date,the 137Cs peak, the calibrated 14C date, and the tephras (open diamonds). Estimated locations of regional tephras: Mt. Redoubt, AD1989–1990 at 1 cm; Mt. Novarupta/Katmai, AD 1912 at 5.5 cm; Mt. St. Augustine, AD 1883 at 7 cm; Mt. Spurr – Crater Peak, AD~1650 at 23 cm; Mt. Redoubt, AD ~1600 at 24.5 cm; Mt. Redoubt, AD ~ 1550 at 30 cm; Mt. St. Augustine, AD ~1450 at 35 cm.This alternative age model falls within the 2-sigma range of the calibrated 14C date. The ages generated from the 210Pb activities aremarked with × on the profile and were not incorporated into the two aforementioned models because of the difficulty in assigning abackground level of 210Pb.
Karluk Frazer Packers
Average escapement (1980–1998) 655 544 237 206 27 569Density of spawners (1000s·km–2) 16.6 14.3 13.4Water residence time (years) 4.8 2.1 2–4
Note: Sources: Koenings et al. (1987), Kyle et al. (1988), Schrof et al.(2000), and Colvin (2001).
Table 2. Comparison of water residence times and density ofspawning sockeye salmon among Karluk, Frazer, and Packerslakes from 1980 to 1998.

© 2004 NRC Canada
Gregory-Eaves et al. 1243
Fig
.6.
Mul
tipr
oxy
reco
rdfr
omPa
cker
sL
ake
high
ligh
ting
the
chan
ges
inm
agne
tic
susc
epti
bili
ty,
biog
enic
sili
ca, δ
15N
,do
min
ant
diat
omta
xa(>
5%re
lativ
eab
unda
nce)
,an
dre
l-at
ive
abun
danc
esof
chry
soph
yte
mic
rofo
ssil
s.S
hade
dar
eas
indi
cate
the
pres
ence
ofte
phra
laye
rs.

be incorporated into sockeye salmon smolts (e.g., Kline etal. 1993). It has also been demonstrated that larger smolts,up to 110 mm in size, have low ocean mortality and agreater chance of returning to their natal nursery grounds tospawn (Koenings and Burkett 1987b). Therefore, it seemsreasonable to suggest that there is a positive feedback loopamong generations of sockeye salmon in systems wheresalmon-derived nutrient loading is significant. Much of thisresearch, however, has been restricted to clearwater nurserylakes that have been monitored for short periods of time.
In our study of Packers Lake, we see a strong response inthe microfossils to changes in salmon-derived nutrients, asinferred from the δ15N profile, particularly between 10 and20 cm (Fig. 6), which suggests that these nutrients are avail-able for uptake by algae in this stained lake. The recentfertilization of Packers Lake also stimulated significant in-creases in zooplankton biomass and sockeye salmon smoltweights (Mazumder and Edmundson 2002) (Table 1).Mazumder and Edmundson (2002) suggested that the lack ofa significant response in chlorophyll a concentrations duringthe fertilization treatment could be explained by large in-creases in herbivorous zooplankton during this period. Over-all, these results suggest that, similar to clearwater lakes, itappears that nutrient dynamics in stained lakes can affectsockeye salmon smolt sizes, which, in turn, can be importantin determining the ocean survival of sockeye salmon.
Over longer time scales, however, it appears that environ-mental conditions in the North Pacific have a primary con-trol on adult sockeye production, as synchronous trends inpopulation dynamics are seen over large geographic areas.Comparison of sockeye salmon population dynamics inferredfrom the Packers Lake paleolimnological data from 10 to20 cm (i.e., ~1770–1880) in a section of the core free oftephra layers with records from Kodiak Island and BristolBay (Finney et al. 2000) reveals a coherent regional pattern(Fig. 7). Specifically, the early 1800s period of lower abun-dances of sockeye salmon at Packers Lake (inferred from thelow δ15N values (Figs. 6 and 7), lower abundances of A. sub-arctica, and higher abundances of benthic diatoms centeredaround 15 cm (Fig. 6) corresponds to a marked decrease insockeye salmon in all of the Kodiak Island and Bristol Baynursery lakes studied to date (Finney et al. 2000). The early1800s also correspond to a period marked by the coldest sea-surface temperatures in the last ~250 years, inferred fromtree-ring analysis (D’Arrigo et al. 1999). Over the twentiethcentury, a steady decline in inferred sockeye salmon escape-ment is apparent in all of the lakes studied to date (Finney etal. 2000; this study) and can be attributed, at least in part, tocommercial fishing.
The anadromous life cycle of sockeye salmon has ledmany investigators to look at both freshwater and marineenvironments to determine their roles in controlling adultsockeye salmon productivity. Generally, researchers have fo-cused on either the freshwater or marine environment, inpart because it would be logistically challenging to studyboth. However, the study of lake sediments from sockeyesalmon nursery lakes allows us to infer some of the dynam-ics from both environments. From our paleolimnological stud-ies of both clear and stained lakes, it appears that salmon-derived nutrients can be important in controlling freshwaterproductivity, which is important to the development of sock-
eye salmon smolts. The ocean environment, however, whichis the habitat shared by all sockeye salmon populations, hasa primary control on adult production over longer timesscales.
In conclusion, changes in nutrients and productivity re-lated to adult sockeye salmon abundances and volcanic erup-tions are the main signals recorded in the Packers Lakesediments. Volcanic ashes appear to have triggered a short-term effect in the diatom community, reflecting slight nutri-ent enrichment. This enrichment response, which is shortterm in nature, is consistent with observations of waterchemistry and algal communities made in several lakes af-fected by volcanic ashfall (Kurenkov 1966; Mathisen andPoe 1978).
In contrast, sediments representing the period between~AD 1770 and 1882 were not influenced by volcanic erup-tions, and during this time, we infer dramatic changes insockeye salmon. The inferred pattern in sockeye salmonpopulation dynamics from Packers Lake tracks the sametrend as those apparent from analyses of other sockeyesalmon stocks from southern Alaska. The synchronicity ofsalmon trends among clearwater and stained lakes suggeststhat the marine phase of the sockeye salmon life cycle is acritical determinant of final adult spawner production, as thisenvironment is common to all salmon populations.
© 2004 NRC Canada
1244 Can. J. Fish. Aquat. Sci. Vol. 61, 2004
Fig. 7. Regional comparison of past sockeye salmon abundances(inferred from δ15N profiles) from (a) south-central Alaska and(b) Bristol Bay, Alaska. The δ15N profile from Packers Lake, thestained sockeye salmon nursery lake, reflects a trend that is co-herent with those from clearwater systems. Control lakes havesteep waterfalls at their outlets and thus have not been accessibleto sockeye salmon. However, a sockeye salmon population wasestablished in one of the control lakes (Frazer Lake) after thefish were introduced in the late 1950s and fish ladders were con-structed over the falls.

Acknowledgements
We thank Jon Sweetman for collecting the Packers Lakecore and providing helpful comments on the manuscript.Drs. Mark Brenner and Jules Blais and an anonymous re-viewer also had very helpful suggestions on the manuscript.The Alaska Department of Fish and Game, particularly JimEdmundson, provided fieldwork logistics. We also appreci-ate the help of Tom Brown, Fawn Ginn, Amy Hirons, NormaHaubenstock, Tim Howe, Andy Krumhardt, David Parker,and Peter Roeder in the laboratory. We acknowledge the as-sistance of Gary Fandrei (Cook Inlet Aquaculture Associa-tion), who provided historical and limnological data onPackers Lake, and John Glew, who drafted the map. Thisresearch was funded by the Natural Sciences and Engi-neering Research Council of Canada, the Ontario Scholar-ship Program, and the Northern Studies Training Program inCanada. The U.S. Global Ocean Ecosystems Dynamics pro-gram (contribution No. 406), jointly funded by the NationalScience Foundation and the National Oceanic and AtmosphericAdministration (NOAA), the Alaska Sea Grant College Pro-gram, and the NOAA Auke Bay Lab Ocean Carrying Capacityprogram supported this research in the United States.
References
Appleby, P.G. 2001. Chronostratigraphic techniques in recent sedi-ments. In Tracking environmental change using lake sediments.Vol. 1. Basin analysis, coring and chronological techniques.Edited by W.M. Last and J.P. Smol. Kluwer Academic Publishers,Dordrecht, Netherlands. pp. 171–203.
Battarbee, R.W., Carvalho, L., Jones, V.J., Flower, R.J., Cameron,N.G., Bennion, H., and Juggins, S. 2001. Diatoms. In Trackingenvironmental change using lake sediments. Vol. 3. Terrestrial,algal and siliceous indicators. Edited by J.P. Smol, H.J.B. Birks,and W.M. Last. Kluwer Academic Publishers, Dordrecht, Neth-erlands. pp. 155–202.
Beget, J.E., Stihler, S.D., and Stone, D.B. 1994. A 500-year-longrecord of tephras falls from Redoubt Volcanoes in upper CookInlet, Alaska. J. Volcanol. Geotherm. Res. 62: 55–67.
Boutton, T.W. 1991. Stable carbon isotope ratios of natural materi-als: II. Atmospheric, terrestrial, marine, and freshwater environ-ments. In Carbon isotope techniques. Edited by D.C. Colemanand B. Fry. Academic Press, Inc., San Diego, Calif. pp. 173–185.
Burgner, R.L. 1991. Life history of sockeye salmon (Oncorhynchusnerka). In Pacific salmon life histories. Edited by C. Groot and L.Margolis. University of British Columbia Press, Vancouver, B.C.pp. 1–117.
Colvin, R. 2001. Packers Lake sockeye salmon enhancement prog-ress report 2000. Cook Inlet Aquaculture Association. Availablefrom the Cook Inlet Aquaculture Association Corporate Office,40610 Kalifornsky Beach Road, Kenai, AK 99611, USA; e-mail: [email protected].
D’Arrigo, R.D, Wiles, G., Jacoby, G., and Villalba, R. 1999. NorthPacific sea surface temperatures: past variations inferred fromtree tings. Geophys. Res. Lett. 26: 2757–2760.
Edmundson, J.A., and Carlson, S.R. 1998. Lake typology influ-ences on the phosphorus–chlorophyll relationship in subarctic,Alaskan lakes. Lake Reservoir Manag. 14: 440–450.
Fandrei, G. 1996. Packers Lake sockeye salmon enhancement prog-ress report 1995. Cook Inlet Aquaculture Association. Availablefrom the Cook Inlet Aquaculture Association Corporate Office,
40610 Kalifornsky Beach Road, Kenai, AK 99611, USA; email:[email protected].
Finney, B.P., Gregory-Eaves, I., Sweetman, J., Douglas, M.S.V.,and Smol, J.P. 2000. Impacts of climatic change and fishing onPacific salmon abundances over the past 300 years. Science(Wash., DC), 290: 795–799.
Finney, B.P., Gregory-Eaves, I., Douglas, M.S.V., and Smol, J.P.2002. Fisheries productivity in the northeastern Pacific Oceanover the past 2200 years. Nature (Lond.), 416: 729–732.
Fritz, S.C., Kingston, J.C., and Engstrom, D.R. 1993. Quantitativetrophic reconstruction from sedimentary diatom assemblages: acautionary tale. Freshw. Biol. 30: 1–23.
Glew, J.R. 1988. A portable extruding device for close interval sec-tioning of unconsolidated core samples. J. Paleolimnol. 1: 235–239.
Gregory-Eaves, I. 2002. Tracking past sockeye salmon (Oncorhynchusnerka) population dynamics using paleolimnology. Ph.D. thesis,Queen’s University, Kingston, Ont.
Gregory-Eaves, I., Smol, J.P., Finney, B.P., and Edwards, M.E. 1999.Diatom-based transfer functions for inferring past climatic andenvironmental changes in Alaska, U.S.A. Arct. Antarct. Alp. Res.31: 353–365.
Gregory-Eaves, I., Smol, J.P., Douglas, M.S.V., and Finney, B.P.2003. Diatoms and sockeye salmon (Oncorhynchus nerka) popu-lation dynamics: reconstructions of salmon-derived nutrients overthe past 2000 years in two lakes from Kodiak Island, Alaska. J.Paleolimnol. 30: 35–53.
Hall, R.I., and Smol, J.P. 1996. Paleolimnological assessment oflong-term water-quality changes in south-central Ontario lakesaffected by cottage development and acidification. Can. J. Fish.Aquat. Sci. 53: 1–17.
Hare, S.R., and Mantua, N.J. 2000. Empirical evidence for NorthPacific regime shifts in 1977 and 1989. Prog. Oceanogr. 47:103–145.
Jones, R.I. 1998. Phytoplankton, primary production and nutrient cy-cling. In Aquatic humic substances: ecology and biogeochemistry.Edited by D.O. Hessen and L.J. Tranvik. Ecological Studies 133.Springer, Berlin. pp. 145–175.
Kline, T.C., Jr., Goering, J.J., Mathisen, O.A., Poe, P.H., Parker,P.L., and Scalan, R.S. 1993. Recycling of elements transportedupstream by runs of Pacific salmon. II. δ15N and δ13C evidencein the Kvichak River Watershed, Bristol Bay, southern Alaska.Can. J. Fish. Aquat. Sci. 50: 2350–2365.
Koenings, J.P., and Burkett, R.D. 1987a. An aquatic rubic’s cube: res-toration of the Karluk Lake sockeye salmon (Oncorhynchus nerka).In Sockeye salmon (Oncorhynchus nerka) population biology andfuture management. Edited by H.D. Smith, L. Margolis, and C.C.Wood. Can. Spec. Pub. Fish. Aquat. Sci. No. 96. pp. 419–434.
Koenings, J.P., and Burkett, R.D. 1987b. Population characteristicsof sockeye salmon (Oncorhynchus nerka) smolts relative to tem-perature regimes, euphotic volume, fry density and forage basewithin Alaskan lakes. In Sockeye salmon (Oncorhynchus nerka)population biology and future management. Edited by H.D.Smith, L. Margolis, and C.C. Wood. Can. Spec. Publ. Fish.Aquat. Sci. No. 96. pp. 216–234.
Koenings, J.P., and Edmundson, J.A. 1991. Secchi disk and pho-tometer estimates of light regimes in Alaskan lakes: effects ofyellow color and turbidity. Limnol. Oceanogr. 36: 91–105.
Koenings, J.P., Kyle, G.B., and Marcuson, P. 1985. Limnologicaland fisheries evidence for rearing limitation of sockeye salmon,Oncorhynchus nerka, production from Packers Lake, Cook Inlet,Alaska (1973–1983). Alaska Department of Fish and Game, Di-vision of Fisheries Rehabilitation, Enhancement and Develop-ment. Available from Alaska Department of Fish and Game,
© 2004 NRC Canada
Gregory-Eaves et al. 1245

Division of Commercial Fisheries, 43961 Kailfornsky BeachRoad, Suite B, Soldotna, AK 99669, USA; telephone: (907)262–9368.
Kurenkov, I.I. 1966. The influence of volcanic ashfall on biologicalprocesses in a lake. Limnol. Oceanogr. 11: 426–429.
Kyle, G. 1994. Nutrient treatment of 3 coastal Alaskan lakes: trophiclevel responses and sockeye salmon production trends. AlaskaFish. Res. Bull. 1: 153–167.
Kyle, G., Koenings, J.P., and Barrett, B.M. 1988. Density-dependent,trophic level responses to an introduced run of sockeye salmon(Oncorhynchus nerka) at Frazer Lake, Kodiak Island, Alaska.Can. J. Fish. Aquat. Sci. 45: 856–867.
Kyle, G., Koenings, J.P., and Edmundson, J.A. 1997. An overviewof Alaska lake-rearing salmon enhancement strategy: nutrientenrichment and juvenile stocking. In Freshwaters of Alaska.Edited by A.M. Milner and M.W. Oswood. Ecological Studies119. Springer, New York. pp. 205–227.
Larson, D. 1993. The recovery of Spirit Lake. Am. Sci. 81: 166–179.Mathisen, O.A., and Poe, P.H. 1978. Effect of volcanic ash on
sockeye salmon lakes. Verh. Int. Ver. Limnol. 20: 165–172.Mathisen, O.A., Parker, P.L., Goering, J.J., Kline, T.C., Poe, P.H.,
and Scalan, R.S. 1988. Biogenic enrichment of sockeye salmonlakes and stock productivity. Verh. Int. Ver. Limnol 19: 487–495.
Mazumder, A., and Edmundson, J.A. 2002. Impact of fertilization andstocking on trophic interactions and growth of juvenile sockeyesalmon (Oncorhynchus nerka). Can. J. Fish. Aquat. Sci. 59: 1361–1373.
Meyers, P.A., and Ishiwatari, R. 1993. Lacustrine organic geo-chemistry — an overview of indicators organic matter sourcesand diagenesis in lake sediments. Org. Geochem. 20: 867–900.
Mortlock, R.A., and Froelich, P.N. 1989. A simple method for therapid determination of biogenic opal in pelagic marine sedi-ments. Deep-Sea Res. 36: 1415–1426.
Schelske, C.L., and Hodell, D.A. 1991. Recent changes in produc-tivity and climate in Lake Ontario detected by isotopic analysisof sediments. Limnol. Oceanogr. 36: 961–975.
Schelske, C.L., Peplow, A., Brenner, M., and Spencer, C.N. 1994.Low-background gamma counting: applications for 210Pb datingof sediments. J. Paleolimnol. 10: 115–128.
Schrof, S.T., Honnold, S.G., Hicks, C.J., and Wadle, J.A. 2000. Asummary of salmon enhancement, rehabilitation, evaluation andmonitoring efforts conducted in the Kodiak Management Area.Alaska Dep Fish Game Regional Inf. Rep. No. 4K00-57. Avail-able from Alaska Department of Fish and Game, Division ofCommercial Fisheries, 211 Mission Road, Kodiak, AK 99615,USA; telephone: (907) 486–1825.
Smol, J.P. 1985. The ratio of diatom frustules to chrysophyceanstatospores: a useful paleolimnological index. Hydrobiologia,123: 199–208.
Stuiver, M., Reimer, P.J., Bard, E., Beck, J.W., Burr, G.S., Hughen,K.A., Kromer, B., McCormac, G., Plicht, J.V.D., and Spurk, M.1998. INTCAL98 radiocarbon age calibration. Radiocarbon, 40:1041–1083.
Sweetman, J., and Finney, B.P. 2003. Differential responses of zoo-plankton populations (Bosmina longirostris) to fish predation andnutrient-loading in an introduced and a natural sockeye salmonnursery lake on Kodiak Island, Alaska, USA. J. Paleolimnol. 30:183–193.
Turney, C.S.M., and Lowe, J.J. 2001. Tephrachronology. In Trackingenvironmental change using lake sediments. Vol. 1. Basin analy-sis, coring and chronological techniques. Edited by W.M. Lastand J.P. Smol. Kluwer Academic Publishers, Dordrecht, Nether-lands. pp. 171–203.
Wertheimer, A.C. 1997. Status of Alaskan salmon. In Pacific salmonand their ecosystems: status and future options. Edited by D.J.Stouder, P.A. Bisson, and R.J. Naiman. Chapman and Hall, NewYork. pp. 179–197.
Yang, J.-R., Pick, F.R., and Hamilton, P.B. 1996. Changes in theplanktonic diatom flora of a large mountain lake in response tofertilization. J. Phycol. 32: 232–243.
Zeeb, B.A., Christie, C.E., Smol, J.P., Findlay, D.L., Kling, H.J.,and Birks, H.J.B. 1994. Responses of diatom and chrysophyteassemblages in Lake 227 sediments to experimental eutrophi-cation. Can. J. Fish. Aquat. Sci. 51: 2300–2311.
© 2004 NRC Canada
1246 Can. J. Fish. Aquat. Sci. Vol. 61, 2004






![Data Analysis Practice General [65 marks]...Data Analysis Practice General [65 marks]1a. [1 mark] Sockeye salmon (Oncorhynchus nerka) spend the first years of their lives in the freshwater](https://static.fdocuments.net/doc/165x107/608f09d6fe1f5a4d3b403934/data-analysis-practice-general-65-marks-data-analysis-practice-general-65.jpg)











