
Inducing Autophagy: Tat-D11, a highly - Novus BiologicalsInducing Autophagy: Tat-D11, a highly...
4
Inducing Autophagy: Tat-D11, a highly potent and specific autophagy-inducing peptide novusbio.com Abstract Many standard methods to induce autophagy lack desired specificity. Both starvation and rapamycin, an allosteric inhibitor of MTORC1, are known to regulate biological processes other than autophagy. The non-specific nature of these methods complicates data interpretation and has driven the preference for loss-of-function Atg mutants to analyze autophagy. In 2013, the discovery of engineered Tat-Beclin 1 provided the first method to induce autophagy without non-specifically regulating other pathways. Peptides comprised of the autophagy-inducing region of Beclin 1 fused to the HIV-Tat protein were demonstrated to increase autophagosome and autolysosome numbers, as well as protein degradation. Since then, Tat-Beclin 1 peptides have been widely used to successfully induce autophagy both in vitro and in vivo. Here we demonstrate the enhanced potency of a shorter Tat-Beclin 1 peptide to induce autophagy. This shorter peptide, Tat-Beclin 1 D11 (Tat-D11), increases autophagosome and autolysosome induction by over fivefold compared to the longer peptide, Tat-Beclin 1. These results demonstrate the superior potency of Tat-D11 over Tat-Beclin 1. Background Autophagy is an assembly of tightly regulated biological processes by which cellular components and bulk proteins are degraded, mostly during starvation and stress, and recycled. Components of the cytoplasm are sequestered and moved into the lysosomes or the vacuole lumen, where they are broken down into their basic components and returned to the cytosol for reuse. Along with the ubiquitin-proteasome system (UPS), autophagy is a critical mechanism for protein homeostasis whereby dysfunctional long-lived proteins and organelles are degraded as part of a quality control process [1, 2]. Three definitive types of autophagy exist in nature: macro-autophagy, micro-autophagy and chaperone-mediated autophagy [8]. Macro-autophagy involves a double-membrane vesicle, the autophagosome, to transport the cytoplasmic cargo to the lysosome. During this process, the autophagosome fuses with the lysosome resulting in the formation of the autolysosome. Micro-autophagy involves the direct transport of cytosolic cargo to the lysosome. Chaperone-mediated autophagy occurs when the cytosolic proteins are transported across the lysosome membrane involving the LAMP-2A (lysosomal-associated membrane protein 2A) receptor. Due to the vital role of autophagy in cellular and tissue homeostasis, disruption of autophagy can drive disease and pathophysiological conditions. Loss of autophagy has been shown to result in the accumulation of misfolded proteins in mice [3-5], and ubiquitylated proteins and inclusion bodies in neurons leading to neurodegeneration and cardiac hypertrophy [6, 7]. Common methods to induce autophagy including rapamycin, starvation, and carbamazepine elicit off-target effects and non-specifically regulate cellular processes instead of autophagy alone. Data produced using these methods must be interpreted carefully and with an understanding of how these compounds regulate other unrelated biological processes.
Transcript of Inducing Autophagy: Tat-D11, a highly - Novus BiologicalsInducing Autophagy: Tat-D11, a highly...

Inducing Autophagy: Tat-D11, a highly potent and specific autophagy-inducing peptide
novusbio.com
AbstractMany standard methods to induce autophagy lack desired specificity. Both starvation and rapamycin, an allosteric inhibitor of
MTORC1, are known to regulate biological processes other than autophagy. The non-specific nature of these methods complicates
data interpretation and has driven the preference for loss-of-function Atg mutants to analyze autophagy. In 2013, the discovery of
engineered Tat-Beclin 1 provided the first method to induce autophagy without non-specifically regulating other pathways. Peptides
comprised of the autophagy-inducing region of Beclin 1 fused to the HIV-Tat protein were demonstrated to increase autophagosome
and autolysosome numbers, as well as protein degradation. Since then, Tat-Beclin 1 peptides have been widely used to successfully
induce autophagy both in vitro and in vivo. Here we demonstrate the enhanced potency of a shorter Tat-Beclin 1 peptide to induce
autophagy. This shorter peptide, Tat-Beclin 1 D11 (Tat-D11), increases autophagosome and autolysosome induction by over fivefold
compared to the longer peptide, Tat-Beclin 1. These results demonstrate the superior potency of Tat-D11 over Tat-Beclin 1.
BackgroundAutophagy is an assembly of tightly regulated biological processes by which cellular components and bulk proteins are degraded, mostly during starvation and stress, and recycled. Components of the cytoplasm are sequestered and moved into the lysosomes or the vacuole lumen, where they are broken down into their basic components and returned to the cytosol for reuse. Along with the ubiquitin-proteasome system (UPS), autophagy is a critical mechanism for protein homeostasis whereby dysfunctional long-lived proteins and organelles are degraded as part of a quality control process [1, 2].
Three definitive types of autophagy exist in nature: macro-autophagy, micro-autophagy and chaperone-mediated autophagy [8]. Macro-autophagy involves a double-membrane vesicle, the autophagosome, to transport the cytoplasmic cargo to the lysosome. During this process, the autophagosome fuses with the lysosome resulting in the formation of the autolysosome. Micro-autophagy involves the direct transport of cytosolic cargo to the lysosome. Chaperone-mediated
autophagy occurs when the cytosolic proteins are transported across the lysosome membrane involving the LAMP-2A (lysosomal-associated membrane protein 2A) receptor. Due to the vital role of autophagy in cellular and tissue homeostasis, disruption of autophagy can drive disease and pathophysiological conditions. Loss of autophagy has been shown to result in the accumulation of misfolded proteins in mice [3-5], and ubiquitylated proteins and inclusion bodies in neurons leading to neurodegeneration and cardiac hypertrophy [6, 7]. Common methods to induce autophagy including rapamycin, starvation, and carbamazepine elicit off-target effects and non-specifically regulate cellular processes instead of autophagy alone. Data produced using these methods must be interpreted carefully and with an understanding of how these compounds regulate other unrelated biological processes.

novusbio.com
Peptide DescriptionsTat-Beclin 1 D11 (Tat-D11) [NBP2-49888]: Peptides comprising 11 amino acids derived from the autophagy inducer Beclin 1 linked to the HIV Tat protein to increase solubility. Tat-D11 is in the D-amino acid retro-inverso configuration.
Tat-Beclin 1 L11 (Tat-L11) [NBP2-49886]: Peptides comprising 11 amino acids derived from the autophagy inducer Beclin 1 linked to the HIV Tat protein to increase solubility. Tat-L11 is in the naturally occurring L-configuration.
Tat-Beclin 1 L11S (Tat-L11S) [NBP2-49887]: An inactive, scrambled control peptide derived from Tat-L11. Tat-L11S is recommended as a negative control for Tat-D11 and Tat-L11.
AUTOPHAGYINDUCTION
Tat-D11 Peptides
GAPR-1
Beclin 1
Illustration 1. GAPR-1/GLIPR2 is a negative regulator of autophagy and binds Beclin 1 to inhibit autophagy. In the presence of Tat-D11 peptides, Beclin 1 bound to GAPR-1 is released allowing Beclin 1 to mediate autophagosome formation and autophagy induction.
A potent and specific inducer of autophagy was engineered and detailed in a 2013 Nature publication [9]. Shoji-Kawata et al designed peptides that were mapped to a region of the autophagy-inducing protein, Beclin 1, and linked it to the HIV-1 Tat protein to facilitate transduction into the cell. The Tat-Beclin 1 peptides induced autophagy by interacting with the negative regulator of autophagy GAPR-1 (also known as GLIPR2). Tat-Beclin 1 peptides cleared small polyglutamine expansion protein aggregates, reduced titers of several positive-stranded
RNA viruses, and inhibited HIV replication in human macrophages [9].
Here, we demonstrate the vastly improved potency of a shorter Tat-Beclin 1 peptide, Tat-D11, to induce autophagy as evidenced by increased autophagosome and autolysosome numbers relative to Tat-Beclin 1. The shorter Tat-D11 peptides improved autophagy induction by over fivefold relative to the original Tat-Beclin 1 peptides.

novusbio.com
Tat-D11 10mMTat-L11S 10mM
Tat-L11 10mMTat-L11 10mM Tat-Beclin 1 10mM
Figure 1. Tat-D11 and Tat-L11 are more potent autophagy inducers than Tat-Beclin 1. HeLa GFP-LC3B were treated with Tat-D11, Tat-L11, Tat-L11S or Tat-Beclin for 1.5 hrs, and (A) the number of autophagosomes were assessed by fluorescent mi-croscopy and (B) the number of GFP+/ LC3B+ spots were quantified.
Figure 1. Tat-D11 is a more potent autophagy inducer than Tat-Beclin 1. HeLa GFP-LC3B were treated with Tat-D11, Tat-L11, or Tat-Beclin for 1.5 hr, and (A) the number of autophagosomes were assessed by fluorescent microscopy and (B) the number of GFP+/ LC3B+ spots were quantified.
A. B.
Results and MethodsFigures 1 and 2 depict the vastly superior induction of autophagy by Tat-D11 in comparison to the currently used longer Tat-Beclin 1 peptide. The cells were plated in 96 black-well culture plates at 1.5 x 106 cells/mL and grown overnight at 37°C with 5% CO2 until ~80% confluent. Peptides were dissolved in sterile, ultrapure water to 1 mM and then added to Opti-MEM acidified with 0.15% 6N HCl. A 1:2 serial dilution, starting at 20 μM, was used to find the optimal induction concentration using the peptides: Tat-Beclin 1, Tat-D11, Tat-L11 and a scrambled version of the peptides, Tat-L11S. Cells incu-bated for 1.5 hrs at 37°C with 5% CO2. The peptides were removed and the cells fixed with 4% paraformaldeyhde for 20 min at room temperature. The cells were washed with sterile 1X PBS (pH 7.0) and then blocked with blocking buffer (1X PBS, 0.1% Triton-X, and 5% BSA) for 1 hr at room temperature. A 1:200 dilution of Neurotrace Red (Invitrogen), in addition to a 1:10,000 dilution of DAPI in 1X PBS were used to counterstain the cells. The plates were foil-sealed for analysis. Analysis was performed on the Operetta high content imaging platform and GFP-puncta from 11 spots within each well were enumerated at 40X (n=33).
Western blots showing the induction of LC3B and downregulation of p62 by Tat-D11 peptide is shown in Figure 3. 12-well plates were treated with peptides for 1.5 hrs as described above. Cell lysis was carried out using 150 μL of M-PER™ Mammalian Protein Extraction Reagent (Thermo 78501) with 1:100 Halt™ Protease and Phosphatase Inhibitor Single-Use Cocktail (Thermo 78442) per well. Plates were incubated at room temperature for 10 min and gently agitated. Cells were scraped from the plate and spun down at 13500 rpm for 10 min at 4oC. Cell debris was excluded and 30 μL of 6X Lamelli Reducing SDS loading buffer was added to the 150 μL supernatant. The solution was boiled for 5 min at 95°C and then cooled to room temperature before loading. A 5-20% gradient gel was loaded with 10 μL of reduced sample and run at 130 V for 1 hr. Gels were transferred to a nitrocellulose membrane at 100 V for 1 hr. Membranes were blocked for 1-2 hrs at room temperature in Pierce Protein-Free (PBS) Blocking Buffer (Thermo 37584). Cells were then stained overnight in Pierce Protein-Free Blocking Buffer with the respective antibodies: LC3B (Novus NB100-2220) at 2 μg/mL, SQSTM1/p62 (Novus Biologicals MAB8028) at 2 μg/mL, and
novusbio.com
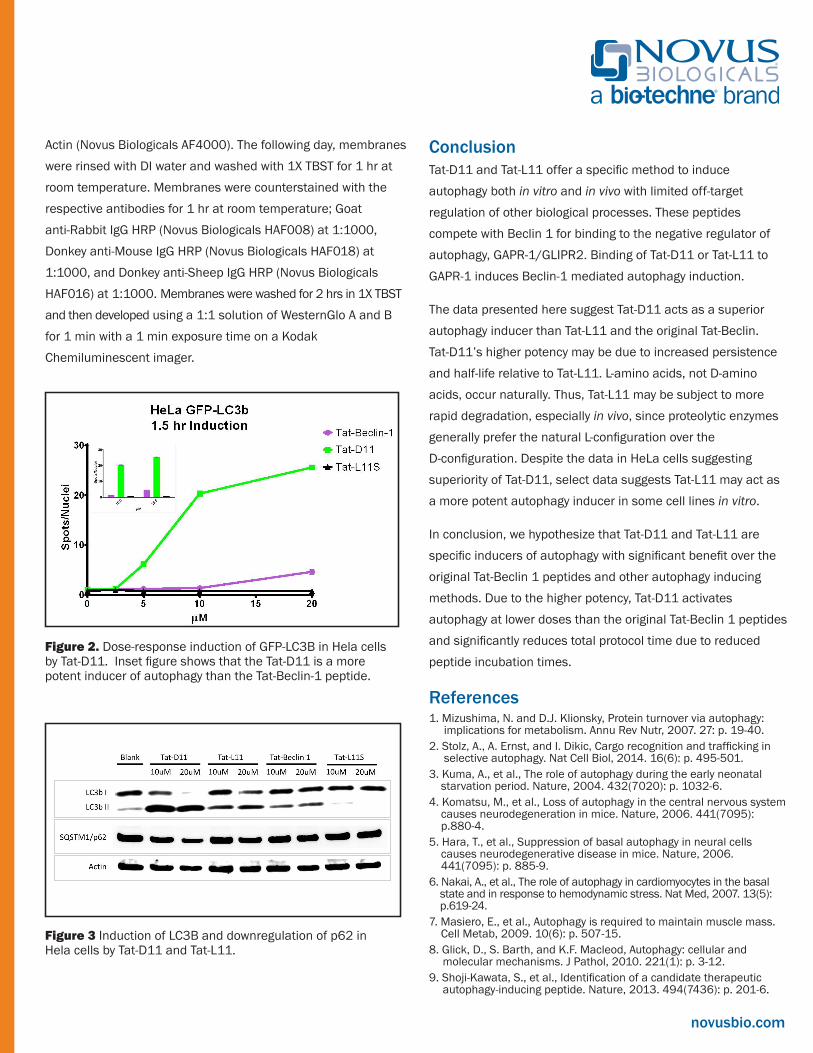
Figure 2. Dose-response induction of GFP-LC3B in Hela cells by Tat-D11. Inset figure shows that the Tat-D11 is a more potent inducer of autophagy than theTat-Beclin-1 peptide.
Figure 2. Dose-response induction of GFP-LC3B in Hela cells by Tat-D11. Inset figure shows that the Tat-D11 is a more potent inducer of autophagy than the Tat-Beclin-1 peptide.
Figure 3 Induction of LC3B and downregulation of p62 in Hela cells by Tat-D11 and Tat-L11.
Actin (Novus Biologicals AF4000). The following day, membranes were rinsed with DI water and washed with 1X TBST for 1 hr at room temperature. Membranes were counterstained with the respective antibodies for 1 hr at room temperature; Goat anti-Rabbit IgG HRP (Novus Biologicals HAF008) at 1:1000, Donkey anti-Mouse IgG HRP (Novus Biologicals HAF018) at 1:1000, and Donkey anti-Sheep IgG HRP (Novus Biologicals HAF016) at 1:1000. Membranes were washed for 2 hrs in 1X TBST and then developed using a 1:1 solution of WesternGlo A and B for 1 min with a 1 min exposure time on a Kodak Chemiluminescent imager.
ConclusionTat-D11 and Tat-L11 offer a specific method to induce autophagy both in vitro and in vivo with limited off-target regulation of other biological processes. These peptides compete with Beclin 1 for binding to the negative regulator of autophagy, GAPR-1/GLIPR2. Binding of Tat-D11 or Tat-L11 to GAPR-1 induces Beclin-1 mediated autophagy induction.
The data presented here suggest Tat-D11 acts as a superior autophagy inducer than Tat-L11 and the original Tat-Beclin. Tat-D11’s higher potency may be due to increased persistence and half-life relative to Tat-L11. L-amino acids, not D-amino acids, occur naturally. Thus, Tat-L11 may be subject to more rapid degradation, especially in vivo, since proteolytic enzymes generally prefer the natural L-configuration over the D-configuration. Despite the data in HeLa cells suggesting superiority of Tat-D11, select data suggests Tat-L11 may act as a more potent autophagy inducer in some cell lines in vitro.
In conclusion, we hypothesize that Tat-D11 and Tat-L11 are specific inducers of autophagy with significant benefit over the original Tat-Beclin 1 peptides and other autophagy inducing methods. Due to the higher potency, Tat-D11 activates autophagy at lower doses than the original Tat-Beclin 1 peptides and significantly reduces total protocol time due to reduced peptide incubation times.
References 1. Mizushima, N. and D.J. Klionsky, Protein turnover via autophagy: implications for metabolism. Annu Rev Nutr, 2007. 27: p. 19-40. 2. Stolz, A., A. Ernst, and I. Dikic, Cargo recognition and trafficking in selective autophagy. Nat Cell Biol, 2014. 16(6): p. 495-501. 3. Kuma, A., et al., The role of autophagy during the early neonatal starvation period. Nature, 2004. 432(7020): p. 1032-6. 4. Komatsu, M., et al., Loss of autophagy in the central nervous system causes neurodegeneration in mice. Nature, 2006. 441(7095): p.880-4. 5. Hara, T., et al., Suppression of basal autophagy in neural cells causes neurodegenerative disease in mice. Nature, 2006. 441(7095): p. 885-9. 6. Nakai, A., et al., The role of autophagy in cardiomyocytes in the basal state and in response to hemodynamic stress. Nat Med, 2007. 13(5): p.619-24. 7. Masiero, E., et al., Autophagy is required to maintain muscle mass. Cell Metab, 2009. 10(6): p. 507-15. 8. Glick, D., S. Barth, and K.F. Macleod, Autophagy: cellular and molecular mechanisms. J Pathol, 2010. 221(1): p. 3-12. 9. Shoji-Kawata, S., et al., Identification of a candidate therapeutic autophagy-inducing peptide. Nature, 2013. 494(7436): p. 201-6.


















