
Metallothionein and occupational to cadmium · Metallothionein (MT) is a low molecular weight (
Upload
priyanka-sahaCategory
view
215download
2
In Vitro Radiation Induced Alterations in Heavy Metalsand Metallothionein Content in Plantago ovata Forsk
Priyanka Saha & Debadutta Mishra &
Anindita Chakraborty & Mathummal Sudarshan &
Sarmistha Sen Raychaudhuri
Received: 19 March 2008 /Accepted: 11 April 2008 /Published online: 21 May 2008# Humana Press Inc. 2008
Abstract Proton Induced X-ray emission (PIXE) and fluorescence-activated cell sorting(FACS) have been used to study the effects of gamma irradiation on heavy metalaccumulation in callus tissue of Plantago ovata—an important cash crop of India. PIXEanalysis revealed radiation-induced alteration in trace element profile during developmentalstages of the callus of P. ovata. Subsequent experiments showed antagonism between Feand Cu and also Cu and Zn and synergistic effect between Fe and Zn. FACS analysisshowed significant induction of the metallothionein (MT) protein following gamma-irradiation, and maximum induction was noted at the 50-Gy absorbed dose. This indicated aprogressive increment of MTs as a measure for protection against gamma-rays, to combatalteration in the homeostasis of heavy metals like Fe, Cu, Zn, and Mn.
Keywords Plantago ovata . Callus culture . Gamma irradiation . Trace elements .
Metallothionein
Introduction
Plant tissue culture, an important tool to study different aspects of genetic engineering, hasbeen widely used in fundamental and applied fields. The availability of efficient and long-term plant regeneration systems from callus culture is of prime importance in theapplication of tissue culture in crop improvements. In vitro morphological response inplants is influenced by various factors including initial explant type, culture conditions,genotype, and nutrient media composition [1–3]. These factors modulate cellulardifferentiation and morphological development. However, in vitro morphogenesis has also
Biol Trace Elem Res (2008) 124:251–261DOI 10.1007/s12011-008-8141-8
P. Saha : S. S. Raychaudhuri (*)Department of Biophysics, Molecular Biology and Genetics, University of Calcutta, 92, APC Road,Kolkata, 700009, Indiae-mail: [email protected]
D. Mishra :A. Chakraborty :M. SudarshanUGC-DAE Consortium for Scientific Research, Kolkata Centre, Radiation Biology Division,3/LB Bidhannagar, Salt Lake, Kolkata, 700098, India

been reported to be altered by various stress conditions [4]. The metabolic pathways orfactors involved in the alteration of morphogenesis under abiotic stress are an interestingarena to focus on.
Plantago ovata Forsk, the common Isabgul, is an economically important medicinalplant. The husks of the seeds of P. ovata are used as laxatives. Our group has been able toregenerate this species in vitro by callus culture [5] and somatic embryogenesis [6]. In thepresent work, the callus culture of P. ovata was used as the test system because of its lowchromosome number, amenable behavior in tissue culture, and rapid growth rate.
Earlier researchers established that trace elements of the culture media play a key role inthe induction of callus culture. These elements are needed for the structural and catalyticrole in association with proteins and other cell components to perform various metabolicactivity [7, 8]. Throughout the developmental sequence, calli are known to reveal changesin compositions of individual elements. In plants, ample variations in levels of thesemicronutrients have been recorded depending upon the environment in which the plants areexposed [9, 10]. In some cases, the chemical form of the concerned element is of paramountimportance in terms of their subsequent absorption or eventual incorporation to thefunctional tissues [11, 12]. As a consequence of these wide variability, plants have devisedpowerful physiological and biochemical systems to absorb, retain, and use the essentialmetals within a range for use in their specific metabolic functions [13, 14]. An alteration inmetabolic functions via heavy metal burden has been reported in many cases as aconsequence of exposure to radiation [15–17]. Under such stressed conditions, it has beenshown that a low-molecular-weight (6–7 kDa) cysteine-rich (25–33%) protein calledmetallothionein (MT) widely distributed in eukaryotic and prokaryotic organisms plays acritical role in the detoxification of heavy metals. In plants, a correlation has been observedbetween MT ribonucleic acid levels and tolerance to heavy metals in different Arabidopsisecotypes [18] suggesting a role in metal homeostasis of essential metal ions notably Zn andCu. In plants, earlier researchers have observed that MT is induced by heavy metal stress[19, 20]. Numerous studies over several years have implicated that MT synthesis are alsoinduced by the exposure of cells in vitro or tissues in vivo to either ionizing or UV radiation[21–23].
In this context, the present investigation is aimed at the determination of radiation-induced alteration in the profile of some essential metals such as iron (Fe), copper (Cu),zinc (Zn), and manganese (Mn) and its correlation with the MT content in the callus cultureof P. ovata. The role of trace elements such as Fe, Cu, Zn, and Mn in cell metabolism is ofparticular importance because many enzymes required these metals for their catalyticactivity [24, 25]. Deficiency and excess of some of these trace elements may lead to anumber of pathobiological conditions in different life forms [26–28]. The present studymainly focused on these four trace elements because MT has been suggested to regulate theintake of the essential metal ions like Fe, Zn, and Cu into the cell as well as participating asa metal donor and acceptor in different biosynthetic and catabolic processes, and thesemineral ions were also seen to be bound to the MT protein [29, 30]. The binding capacitiesof MTs are indicative of the role of these proteins in mineral homeostasis and enzymaticcatalytic activity. In the present study, PIXE technique has been utilized for the qualitativeand quantitative determination of trace elements of P. ovata. PIXE has been widely used fortrace element analysis in different fields of fundamental and applied sciences [31, 32]. Thistechnique offers advantages over other conventional techniques of element detection as itcan detect the multielemental profile of even a minute quantity of a sample and the preparedsample can be reused. MT content has been estimated by fluorescence-activated cell sorting(FACS), which permits the analysis of a great number of cells in a fraction of time and has
252 Saha et al.

been a powerful tool for diverse fields in biomedical research and clinical application [33,34]. The work has been designed with an aim to understand the precise role of somespecific elements during in vitro plant regeneration to counteract the effect of stress inducedby gamma-rays in the alteration of the MT content in a dose-dependent manner.
Materials and Methods
Reagents
Sucrose, agar, cellulase, pectinase, and D-mannitol were purchased from SRL (Mumbai,India). 2,4-dichlorophenoxyacetic acid (2,4-D), 6-furfurylaminopurine (kinetin), and bovineserum albumin (BSA) were obtained from Sigma (St Louis, MO, USA). The Murashige andSkoog (MS) medium was obtained from Himedia (Mumbai, India). Graphite powder andformaldehyde were purchased from E-Merck (Mumbai, India). Trypan blue was obtainedfrom Spectrochem. MT primary antibody and fluorescein isothiocyanate (FITC)-conjugatedsecondary antibody were obtained from Santa Cruz Biotechnology (California, USA).
Seed Germination and Callus Induction
Seeds of P. ovata were imbibed in 20 ml sterile distilled water about 24 h. The followingday, seeds were sterilized with 10% (v/v) liquid sodium hypochlorite (NaOCl) for 20 min andrinsed about five times with sterile distilled water to remove traces of sodium hypochlorite.Seeds were germinated aseptically in agar–sucrose media containing 3% (w/v) sucrose and0.9% (w/v) agar and germinated within 12 to 14 days. Callus cultures were established fromleaf explants (1 cm) of aseptically germinated seedlings of P. ovata following thestandardized method as described by Das Pal and Sen Raychaudhuri [6] in a MS medium[35] supplemented with 1 mg/l 2,4-D and 0.5 mg/l 6-furfurylaminopurine (kinetin). The pHof the medium was adjusted between 5.6 and 5.8 and autoclaved. The calli were grown at atemperature of 22–25°C and a relative humidity of 55–60% under Philips fluorescentdaylight tubes emitting 32×108 mol s−1m−2 for 16/8-h duration in light/dark periods. Calliwere maintained on the same fresh medium for two passages of 21-day duration.
Gamma Irradiation
Freshly subcultured callus weighing about 1 g each were taken in plastic packets separatelyand irradiated by a Cobalt-60 (60Co) gamma source (Gamma chamber 900, BRIT, NaviMumbai, India) at 25°C at the Chemical Science Division of Saha Institute of NuclearPhysics, Kolkata, India. Samples were irradiated at 10-, 20-, 50-, and 100-Gy absorbeddoses (Dose rate 1 Gy/min). Five sets of callus samples were exposed to each dose ofgamma-rays. Each of the five treatments was replicated thrice. The control set (not exposedto gamma rays) was maintained aseptically under laboratory conditions. All irradiated andcontrol samples were grouped for carrying out the PIXE and FACS analyses.
Sample Preparation for PIXE Studies
Nonirradiated and irradiated young calli weighing about 1 g each were freeze dried using aVertex Lyophilizer (Virtis, Gardiner, New York) at −80°C for 48 h and homogenized by abrittle fracture technique under liquid nitrogen. For PIXE analysis, these were then finally
In Vitro Radiation Induced Alterations in Plantago ovata Forsk 253

ground into powder and mixed thoroughly with extrapure graphite in a desired ratio(60:40). For each plant part, about 150 mg was palletized using a pelletizer and, using a 10-mm die, made into 1-mm-thick pellets according to the standardized protocol for thicktarget preparation of biological samples [36].
Blank targets, including the chemicals used and the standards from National Institute ofStandards and Technology (Trace elements in Apple Leaf Standard SRM 1515) were alsoprepared in the same technique. For the present work, we have considered Apple leafstandard SRM 1515 as the standard reference.
Experimental Setup of PIXE and Data Collection
A 2-mm collimated 2.5-MeV proton beam, generated from a 3-MV tandem pelletronaccelerator at the Institute of Physics, Bhubaneswar, India, was used to bombard thesamples. Each sample was first bombarded with a proton beam of a current of about 2–5 nA. Subsequently, the high-intensity, low-energy X-rays was suppressed using analuminum (Al) filter of 50 μm thickness using a beam current varying between 20 and40 nA, thus enhancing the X-rays due to higher Z elements. The characteristic X-raysproduced from the sample was extracted through a 25-μm Mylar window and detected by aCanberra-SL 30160 Si(Li) detector. The spectra were recorded on a personal computer-based multichannel analyzer using WINMCA software. The system was calibrated using a55Fe radioisotope standard X-ray source for the identification of the peaks. The spectralanalysis was carried out using GUPIX-2000 user-friendly software for PIXE analysis.
Single-cell Preparation for Fluorescence-activated Cell Sorting
Irradiated and nonirradiated calli weighing about 200 mg each were taken in a centrifugetube. Two hundred microliters of 1% cellulase and 80 μl of 1% pectinase and 520 μl of15% D-mannitol were added to the callus mass and incubated for 4 h for complete digestionof the cell wall of P. ovata. Stock solutions of 1% cellulase and 1% pectinase were preparedin 7.5% D-mannitol solution. After incubation, single cells were observed under afluorescence optical microscope (LEICA DC 300FX, Wetzler, Germany). Differenttreatments were carried our for ideal single-cell formation from the callus of P. ovata(Table 1).
Viability Analysis
Cellular viability was determined by microscope using the dye Trypan blue. About 500 μlof the single-cell suspension was taken in an Eppendorf tube, and 0.5 ml of 0.4% Trypanblue was added to it and allowed to stand for 15 min. This method is based on the principlethat live (viable) cells do not take up certain dyes, whereas dead (nonviable) cells do(Table 1). Percent cell viablility is computed with the equation:
% Cell viability ¼Total viable cells= total cells ðstained and unstainedÞ � 100
Fluorescence-activated Cell Sorting and Data Analysis
After the single cell was observed, the cell suspension was centrifuged at 2,000 rpm for5 min. The resulting supernatant was aspirated, and the cells were resuspended in 0.5–1 mlphosphate buffer saline (PBS; NaCl 0.136 M [w/v], KCl 0.004 M [w/v], Na2HPO4 0.012 M
254 Saha et al.

[w/v], KH2PO4 0.002 M [w/v]; pH 7.4). For fixation, formaldehyde was added to it (1–2%final concentration) and fixed for 10 min at 37°C. The cells were then chilled on ice for1 min. Then, the cells were permeabilized with 100% ice-cold methanol (v/v; 90% finalconcentration) and cooled for 30 min on ice. After permeabilization, 1×106 cells (cellscounts using heamocytometer) were aliquoted into each assay tube. For staining, 2–3 ml ofincubation buffer containing PBS (pH 7.4) and BSA (w/v) was added to each tube andrinsed by centrifugation. It was repeated twice. After centrifugation, the cells wereresuspended in 300 μl incubation buffer per assay tube and kept for 10 min at 25°C forblocking. The cells were then incubated with 2.5 μl MT-specific primary antibody [MT(FL-61)] for 1 h at 25°C with the necessary modification of the method used by themanufacturer. Following treatment with the primary antibody, the cells were washed
Table 1 Different Treatments Carried Out for Single Cell Formation from Callus of Plantago ovata forFACS Analysis
Numberoftreatments
Preparation ofcellulase andpectinase
Digestionperiod(h)
Treatments Inference (under fluorescenceoptical microscope)
Cellviability(%)
1 Enzymesprepared in7.5% mannitolsolution
4 Callus+1%cellulase+1%pectinase+15%mannitol
It gives best result. Ideal singlecells were found
83
2 Enzymesprepared in7.5% mannitol
4 Callus+1%cellulase+1%pectinase+7.5%mannitol
It gives poor result. Aftertreatment, only elongated cellswere found
80
3 Enzymesprepared in7.5% mannitolsolution
1 Callus+1%cellulase+1%pectinase+PBS
No single cells were found 79
4 Enzymesprepared in7.5% mannitolsolution.
4 Callus+1%cellulase+1%pectinase+PBS
Some of the cells were elongated,and some of the cells were round
82
5 Enzymesprepared in7.5% mannitolsolution
16 Callus+1%cellulase+1%pectinase+PBS
It gives an almost same result astreatment no. 1; single cells werefound, and all cells were round
80
6 Enzymesprepared inPBS
4 Callus+1%cellulase+1%pectinase+nomannitol or PBS
After treatment, most of the cellswere elongated
82
Table 2 Comparison of Calculated and Certified Values of Apple Leaf Standard (SRM1515)
Trace elements Calculated value (ppm) Certified value (ppm)
Fe 98±5.60 83±5.00Cu 5.7±0.53 5.64±0.24Zn 12.7±0.83 12.5±0.30Mn 65.5±2.29 54±3.00
In Vitro Radiation Induced Alterations in Plantago ovata Forsk 255

(2×3 ml) in incubation buffer and incubated for 1 h with a FITC-conjugated secondaryantibody (Goat anti-rabbit IgG–FITC) at 25°C. FITC is detected in a green channel or FL-1.A control (negative) sample was prepared in the same manner as described above withoutthe addition of primary and secondary antibodies for comparing this sample with the testsamples. The cells were then washed with 3 ml of incubation buffer and finally resuspendedin 500 μl PBS, and the MT content was analyzed using FACS Calliber Flow Cytometer(Becton Dickinson) equipped with an argon ion laser emitting at 488 nm. The experimentwas carried out at the user facility of Bose Institute, Kolkata, India. Finally, the data wereanalyzed using software Cell Quest. Cell Quest is the software developed by BD withcapabilities for acquiring and analyzing samples.
Results
The observed values of the selected elements were in good conformity with the certifiedvalues of the Apple leaf standard (Table 2). The variation in elemental content during invitro development of the callus is shown in Figs. 1, 2, 3, and 4. Radiation-induced alterationin the concentration of Fe and Zn was found to be synergistic in calli of P. ovata (Figs. 1
0
200
400
600
800
1000
1200
N 10 20 50 100
Gamma dose(Gy)
conc
(ppm
)
Fe Fig. 1 Variation in Fe content incalli of P. ovata under the normaland gamma-irradiated conditions.Elemental variation of differenttrace elements under the normaland gamma-irradiated conditionsby PIXE analysis. N stands forthe nonirradiated condition, 10stands for the 10-Gy gammadose, 20 stands for the 20-Gygamma dose, 50 stands for the50-Gy gamma dose, and 100stands for the 100-Gy gammadose
0
1
2
3
4
5
6
7
N 10 20 50 100
Gamma dose(Gy)
co
nc
(pp
m)
Cu Fig. 2 Variation in Cu content incalli of P. ovata under the normaland gamma-irradiated conditions.Elemental variation of differenttrace elements under the normaland gamma-irradiated conditionsby PIXE analysis. N stands forthe nonirradiated condition, 10stands for the 10-Gy gammadose, 20 stands for the 20-Gygamma dose, 50 stands for the50-Gy gamma dose, and 100stands for the 100-Gy gammadose
256 Saha et al.

and 3). In comparison to the nonirradiated control groups, the concentrations of Fe and Znwere increased in all the gamma-exposed groups except at the 20-Gy absorbed dose, andthe maximum increment was noted at an absorbed dose of 50 Gy. Gamma irradiationinduced an antagonistic effect in Fe and Cu (Figs. 1 and 2) and also between Cu and Zn(Figs. 2 and 3). Callus samples of P. ovata expressed a linear dose-dependent increase inMn with respect to the control counterpart, though irradiation at the next higher dose of the100 Gy Mn level did not show a significant change (Fig. 4). Like Fe and Zn, the maximumincrement was noted at the 50-Gy radiation exposure in the concentration of Mn. The MTprotein level was determined by the flow cytometric technique. FACS analysis showedsignificant induction of the MT protein following gamma irradiation (Fig. 5a–e). Figure 5represents a single-parameter histogram, which shows the effect of gamma radiation on theMT content of P. ovata examined by flow cytometry. Figure 5a shows the negative (control)sample as described in the experimental part. In this type of graph, the control sample(Fig. 5a) was compared with the test samples (Fig. 5b–e). The observed graph reflects alinear dose-dependent increase in the MT protein with respect to the nonirradiated controlgroups, while at a higher dose of 100 Gy, the MT content was decreased in comparison tothe 50-Gy (17.9%) gamma dose and increased (72.1%) compared to the control counterpart.
0
20
40
60
80
100
120
140
160
N 10 50 100
Gamma dose(Gy)
conc
(ppm
)
Zn
20
Fig. 3 Variation in Zn content incalli of P. ovata under the normaland gamma-irradiated conditions.Elemental variation of differenttrace elements under the normaland gamma-irradiated conditionsby PIXE analysis. N stands forthe nonirradiated condition, 10stands for the 10-Gy gammadose, 20 stands for the 20-Gygamma dose, 50 stands for the50-Gy gamma dose, and 100stands for the 100-Gy gammadose
0
50
100
150
200
250
N 10 50 100
Gamma dose(Gy)
con
c(pp
m)
Mn
20
Fig. 4 Variation in Mn content incalli of P. ovata under the normaland gamma-irradiated conditions.Elemental variation of differenttrace elements under the normaland gamma-irradiated conditionsby PIXE analysis. N stands forthe nonirradiated condition, 10stands for the 10-Gy gammadose, 20 stands for the 20-Gygamma dose, 50 stands for the50-Gy gamma dose, and 100stands for the 100-Gy gammadose
In Vitro Radiation Induced Alterations in Plantago ovata Forsk 257
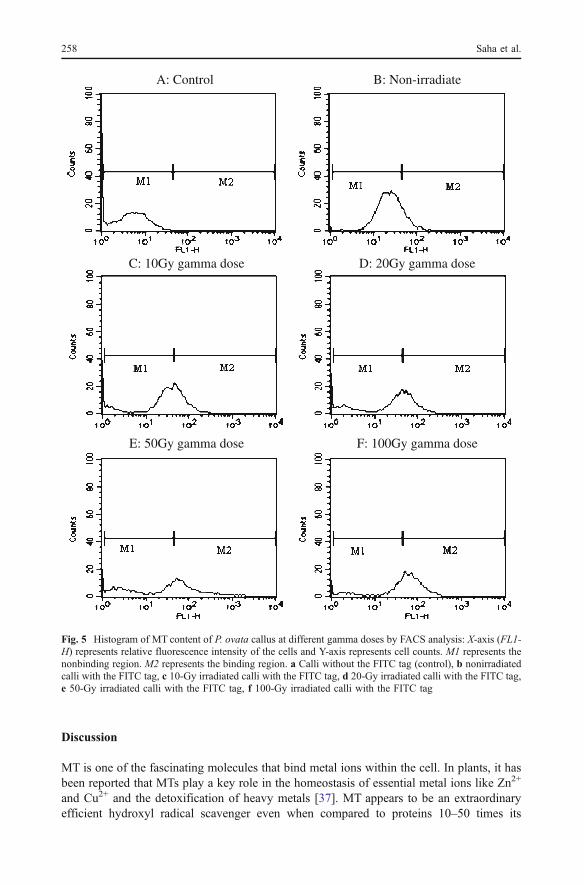
Discussion
MT is one of the fascinating molecules that bind metal ions within the cell. In plants, it hasbeen reported that MTs play a key role in the homeostasis of essential metal ions like Zn2+
and Cu2+ and the detoxification of heavy metals [37]. MT appears to be an extraordinaryefficient hydroxyl radical scavenger even when compared to proteins 10–50 times its
A: Control B: Non-irradiate
C: 10Gy gamma dose
D: 20Gy gamma dose
E: 50Gy gamma dose
F: 100Gy gamma dose
Fig. 5 Histogram of MT content of P. ovata callus at different gamma doses by FACS analysis: X-axis (FL1-H) represents relative fluorescence intensity of the cells and Y-axis represents cell counts. M1 represents thenonbinding region. M2 represents the binding region. a Calli without the FITC tag (control), b nonirradiatedcalli with the FITC tag, c 10-Gy irradiated calli with the FITC tag, d 20-Gy irradiated calli with the FITC tag,e 50-Gy irradiated calli with the FITC tag, f 100-Gy irradiated calli with the FITC tag
258 Saha et al.

molecular weight [38]. Earlier researchers have shown that MTs are induced by thecombined effect of heavy metal and gamma irradiation in the animal system and act ashomeostatic regulators and radical scavengers against oxidative stress [39, 40]. Mikio et al.[21] have shown that MT is induced by a small dose (0.5 Gy) of irradiation. In the currentstudy, the MT protein level was induced by gamma irradiation. Callus cells of P. ovataexpressed a linear dose-dependent increase in the MT protein with respect to the controlcounterpart, and MT induction was maximum at an absorbed dose of 50 Gy, while at ahigher dose of 100 Gy, the MT level declined (with respect to 50 Gy) due to thedegeneration of cells by high gamma exposure. Characterization and expression of differenttypes of MT genes under stress conditions in the plant system has been investigated bysome researchers [41, 42]. Reports on Allium cepa (heavy metal tolerance and heat shock)[41] and rice (salt, drought, and metals CuCl2, ZnCl2, and CdCl2 stress) [42] prompted anincrease in the MT content in different organs of these species in vivo. Murphy and Taiz[18] found that Ag+, Cd+, Zn+, Ni+, and heat shock all induced significant levels of MT2gene expression in Arabidopsis. In the present investigation, flow cytometric data of theMT protein is in good agreement with the PIXE result. While, the maximum increase inelemental content induced by radiation was noted at the 50-Gy absorbed dose compared tothe nonirradiated control in all trace elements except in the concentration of Cu. Tocounteract such increment in elemental concentration, the MT content increased and wasmaximum at 50 Gy. Present investigation showing radiation-induced alteration in the traceelement profile during in vitro development of calli of P. ovata is similar to reports of Desaiet al. [8]. Earlier workers [43, 44] have proposed that these changes in elemental profilehint at the need of specific elements during different dynamic stages of differentiation.Significant changes in elemental concentration in callus tissue under stressed conditionsreflect the importance of trace elements in growth cycle processes of plants in general andin vitro callus development, in particular [8]. The present observation clearly indicates aprogressive increment of MTs as a measure for protection against radiation stress to combatalteration in the homeostasis of heavy metals like Fe, Cu, Zn, and Mn. This result is alsosupported from evidences from our earlier report [45] on stress-induced enzymes like Cu,Zn, Fe, and Mn superoxide dismutases that are also enhanced when exposed to gamma-rays. It may be due to the reason that these free radical-scavenging enzymes require theseheavy metals for their catalytic activity, and hence a progressive increase in these metalsappears as a function of natural defense in a plant system exposed to gamma irradiation.
References
1. Blinstrubiene A, Sliesaravicius A, Burbulis N (2004) Factors affecting morphogenesis in tissue culture oflinseed flax (Linum usitatissimum L.). Acta Univ Latviensis Biol 676:149–152
2. Kothari SL, Agarwal K, Kumar S (2004) Inorganic nutrient manipulation for highly improved in vitroplant regeneration in finger millet—Eleusine coracana (L.) Gaertin. In Vitro Cell Dev Biol Plant40:515–519
3. Niedz RP, Evens TJ (2007) Regulating plant growth by mineral nutrition. In Vitro Cell Dev Biol Plant43:370–381
4. Wi SG, Chung BY, Kim JS, Kim JH, Baek MH, Lee JW, Kim YS (2007) Effects of gamma irradiationon morphological changes and biological responses in plants. Micron 38:553–564
5. Pramanik S, Chakraborty S, Sen Raychaudhuri S (1996) Changes in isozymes during in vitromorphogenesis in Plantago ovata Forsk. Plant Cell Tissue Org Cult 44:123–127
6. Das Pal M, Sen Raychaudhuri S (2000) Enhanced development of somatic embryos of Plantago ovataForsk by additives. In Vitro Cell Dev Biol Plant 37:568–571
7. Sahasrabudhe NA, Nandi M, Bahulikar RA (1999) Influence of boric acid on somatic embryogenesis ofa cytosterile line of indica rice. Plant Cell Tissue Org Cult 58:73–75
In Vitro Radiation Induced Alterations in Plantago ovata Forsk 259

8. Desai NS, Joseph D, Suprasanna P, Bapat VA (2006) Study of elemental variations during somaticembryogenesis in sugarcane using photon induced X-ray probe. Nucl Instrum Methods B 252:299–302
9. Caselles J, Colliga C, Zornoza P (2002) Evaluation of trace element pollution from vehicle emission inpetunia plants. Water Air Soil Pollut 136:1–9
10. Stenstrom A, Jonsdottir IS, Augner M (2002) Genetic and environmental effects on morphology inclonal sedges in the Eurasian Arctic. Am J Bot 89:1410–1421
11. Chen F, Vallyathan V, Castranova V, Shi X (2001) Cell apoptosis induced by carcinogenic metals. MolCell Biochem 222:183–188
12. Speranza A, Ferri P, Battistelli M, Falcieri E, Crinelli R, Scoccianti V (2007) Both trivalent andhexavalent chromium strongly alter in vitro germination and ultrastructure of kiwifruit pollen.Chemosphere 66:1165–1174
13. Poschenrieder C, Tolra R, Barcelo J (2006) Metals defend plants against biotic stress? Trends Plant Sci11:288–295
14. Qureshi MI, Abdin MZ, Qadir S, Iqbal M (2007) Lead induced oxidative stress and metabolic alterationsin Cassia angustifolia Vahl. Biol Plant 51:121–128
15. Babu TS, Akhtar TA, Lampi MA, Tripuranthakam S, Dixon DG, Greenberg BM (2003) Similar stressresponses are elicited by copper and ultraviolet radiation Aquatic plant Lemna gibba: implication ofreactive oxygen species as common signals. Plant Cell Physiol 44:1320–1329
16. Mishra S, Agarwal SB (2006) Interactive effects between supplemental ultraviolet-B radiation and heavymetals on the growth and biochemical characteristics of Spinacia oleracea L, Braz. J Plant Physiol18:307–314
17. Otero S, Olivera EN, Abaigar JM, Tomas R, Crespo MA, Beaucourt N (2006) Effects of cadmium andenhanced UV radiation on the physiology and the concentration of UV-absorbing compounds of theaquatic liverwort Jungermannia exsertifolia subsp. Cordifolia. Photochem Photobiol Sci 5:760–769
18. Murphy A, Taiz L (1995) Comparison of metallothionein gene expression and nonprotein thiols in tenArabidopsis ecotypes. Correlation with copper tolerance. Plant Physiol 109:945–954
19. Chongpraditnun P, Suzuki K, Kawaharada U, Nakajima K, Chino M (1991) Immunohistochemicallocalization of metallothionein in plant tissues. Water Air Soil Pollut 57–58:545–553
20. Hernandez MG, Murphy A, Taiz L (1998) Metallothioneins 1 and 2 have distinct but overlappingexpression patterns in Arabidopsis. Plant Physiol 118:387–397
21. Mikio S, Yanai T, Hasegawa H, Akata N, Kudo SK, Matsumoto T, Noda Y, Otsu H, Sato F (1998)Concentration of metallothionein in mice livers after a small dose of irradiation. J Radiat Res 39:239–242
22. Ono S, Cai L, Cherian MG (1998) Effect of gamma radiation on levels of brain metallothionein and lipidperoxidation in transgenic mice. Radiat Res 150:52–57
23. Ablett E, Whiteman DC, Boyle GM, Green AC, Parsons PG (2003) Induction of MT in human skin byroutine exposure to sunlight: evidence for a systemise response and enhanced induction at certain bodysites. J Invest Dermatol 120:318–324
24. Williams LE, Mills RF (2005) P1B-ATPases—an ancient family of transition metal pumps with diversefunctions in plants. Trends Plant Sci 10:491–502
25. Panda SK, Choudhury S (2005) Chromium stress in plants. Braz J Plant Physiol 17:95–10226. Krouma A, Drevon JJ, Abdelly C (2006) Genotypic variation of N2-fixing common bean (Phaseolus
vulgaris L.) in response to iron deficiency. J Plant Physiol 163:1094–110027. Bouazizi H, Jouili H, Ferjani EI (2007) Copper-induced oxidative stress in maize shoots (Zea mays L.):
H2O2 accumulation and peroxidases modulation. Acta Biol Hung 58:209–21828. Maksymiec W, Wojcik M, Krupa Z (2007) Variation in oxidative stress and photochemical activity in
Arabidopsis thaliana leaves subjected to cadmium and excess copper in the presence or absence ofjasmonate and ascorbate. Chemosphere 66:421–427
29. Winge DR, Nielson KB, Gray WR, Hamer DH (1985) Yeast metallothionein. Sequence and metalbinding properties. J Biol Chem 260:14464–14470
30. Duncan KER, Ngu TT, Chan J, Salgado MT, Merrifield ME, Stillman MJ (2006) Protein folding metal-binding mechanisms and binding site structure in metallothioneins. Exp Biol Med 231:1488–1499
31. Olabanji SO, Olubunmi P, Ceccato D, Buoso MC, Poli MD, Moschini G (2005) Evaluation of Nigeriananimal feeds by particle induced X-ray emission. Biol Trace Elem Res 107:181–197
32. Olabanji SO, Adesina SK, Ceccato D, Buoso MC, Moschini G (2007) PIXE analysis of some medicinalplants used in cleaning teeth in Southwestern Nigeria. Biol Trace Elem Res 116:171–184
33. Yurkow EJ, Makhijani PR (1998) Flow cytometric determination of metallothionein levels in humanperipheral blood lymphocytes: utility in environmental exposure assessment. J Toxicol Environ Health54:445–457
34. Lee HC, Lin TY (2005) Isolation of plant nuclei suitable for flow cytometry from recalcitrant tissue byuse of a filtration column. Plant Mol Biol Report 23:53–58
260 Saha et al.

35. Murashige T, Skoog F (1962) A revised medium for rapid growth and bio-assays with tobacco tissuecultures. Physiol Plant 15:473–497
36. Chakraborty A, Selvaraj S, Sudarshan M, Dutta RK, Ghugre SS, Chintalapudi SN (2000) Modulatoryrole of vanadium on trace element profile in diethylnitrosamine-induced rat hepatocarcinogenesis. NuclInstrum Methods B 170:156–162
37. Zhigang A, Cuijie L, Yuangang Z, Yejie D, Wachter A, Gromes R, Raush T (2006) Expression ofBjMT2, a metallothionein 2 from Brassica juncea, increases copper and cadmium tolerance inEscherichia coli and Arabidopsis thaliana, but inhibits root elongation in Arabidopsis seedlings. J ExpBot 57:3575–3582
38. Thornally PJ, Vasak M (1985) Possible role for metallothionein in protection against radiation-inducedoxidative stress. Kinetics and mechanism of its reaction with superoxide and hydroxyl radicals. BiochimBiophys Acta 827:36–44
39. Matsubara J, Tajima Y, Karasawa M (1987) Metallothionein induction as a potent means of radiationprotection in mice. Radiat Res 111:267–275
40. Koterov AN, Trebenok ZA, Pushkareva NB, Nikol’skii AV (1997) The diverse effects of the combinedaction of cadmium chloride and ionizing radiation on the content of metallothioneins in mouse bonemarrow and liver. Radiat Biol Radioecol 37:196–201
41. Zhang H, Xu W, Guo J, He Z, Ma M (2005) Coordinated responses of phytochelations andmetallothioneins to heavy metals in garlic seedlings. Plant Sci 169:1059–1065
42. Jin S, Cheng Y, Guan Q, Liu D, Takano T, Liu S (2006) A metallothionein-like protein of rice (rgMT)functions in E. coli and its gene expression is induced by abiotic stresses. Biotechnol Lett 28:1749–1753
43. Ramage CM, Williams RR (2003) Mineral uptake in tobacco leaf discs during different developmentalstages of shoot organogenesis. Plant Cell Rep 21:1047–1053
44. Kintzios S, Drossopoulos JB, Lymperopoulos C (2001) Effect of vitamins and inorganic micronutrientson callus growth and somatic embryogenesis from leaves of chilli pepper. Plant Cell Tissue Organ Cult67:55–62
45. Roy S, Begum Y, Chakraborty A, Sen Raychaudhuri S (2006) Radiation-induced phenotypic alterationsin relation to isozymes and RAPD markers in Vigna radiata (L.) Wilczek. Int J Radiat Biol 82:823–832
In Vitro Radiation Induced Alterations in Plantago ovata Forsk 261







![Ostreopsis oVata meter [OVmeter] - onairweb.com · Ostreopsis oVata meter [OVmeter] PARTNERS OnAIR srl (capofila) CNR-Istituto di Biofisica Gruppo SIGLA srl SITEM srl Università](https://static.fdocuments.net/doc/165x107/5ecfe4ff5dc0cd06e4103cf4/ostreopsis-ovata-meter-ovmeter-ostreopsis-ovata-meter-ovmeter-partners-onair.jpg)










