
Impact of microbiome on liver disease - ILSI...
53
Fondazione Edmund Mach Kieran Tuohy Fondazione Edmund Mach, San Michele all’Adige, Trento, Italy Impact of microbiome on liver disease
-
Upload
phungkhuong -
Category
Documents
-
view
217 -
download
0
Transcript of Impact of microbiome on liver disease - ILSI...

Fondazione Edmund Mach
Kieran Tuohy Fondazione Edmund Mach,
San Michele all’Adige, Trento, Italy
Impact of microbiome on liver disease

Ask not what your diet can do to your microbiome
but what your microbiome can do to your diet.

Liver fibrosis and repair: immune regulation of wound healing in a solid organ
Antonella Pellicoro, Prakash Ramachandran, John P. Iredale & Jonathan A. Fallowfield
Nature Reviews Immunology 14, 181–194 (2014)
Gut microbiome and liver disease
• Increased microbiota over-growth, dysbiosis and modified metabolic output
• Increased gut permeability – metabolic endotoxemia, hepatic inflammation

Cellular & Molecular Gastroenterology & Hepatology,
(2015) 1:275-284.

Cross talk between gut microbiota
and liver
• Metabolites – metabolic end products of ingested foods
(SCFA, small phenolic compounds, amino acids/amino acid
derivatives e.g. TRP metabolites, amines, phenols, ammonia)
• Enterohepatic circulation of bile acids – bile acid deconjugation,
dehydroxylation, 1° to 2° bile acids
• Bacterial translocation and inflammatory signals (e.g. LPS)
• Fats, lipids and choline metabolism
(e.g. TMA, Modified cholesterol species, microbial
biohydrogenation)

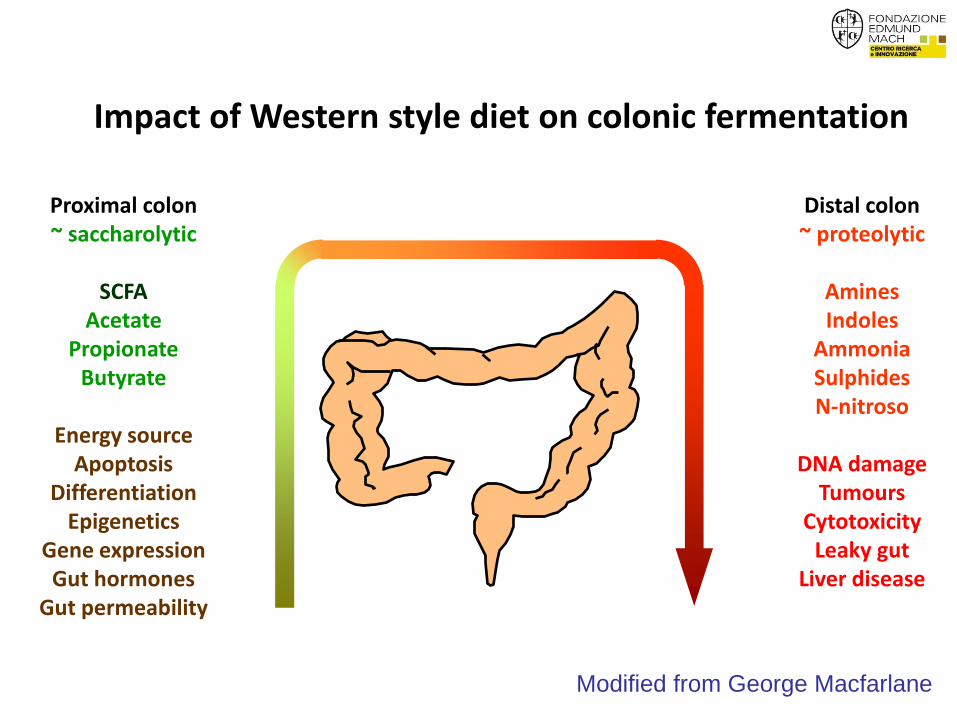
Impact of traditional diets rich in fiber on colonic fermentation
Proximal colon ~ saccharolytic
SCFA
Acetate Propionate
Butyrate
Energy source Apoptosis
Differentiation Epigenetics
Gene expression Gut hormones
Gut permeability
Distal colon ~ proteolytic
Amines Indoles
Ammonia Sulphides N-nitroso
DNA damage
Tumours Cytotoxicity
Leaky gut Liver disease
Modified from George Macfarlane
Impact of Western style diet on colonic fermentation
Proximal colon ~ saccharolytic
SCFA
Acetate Propionate
Butyrate
Energy source Apoptosis
Differentiation Epigenetics
Gene expression Gut hormones
Gut permeability
Distal colon ~ proteolytic
Amines Indoles
Ammonia Sulphides N-nitroso
DNA damage
Tumours Cytotoxicity
Leaky gut Liver disease
Modified from George Macfarlane

Table 1. Cellular actions described for TGR5 in different cell types. ∗Macrophages include alveolar macrophages,
Kupffer cells and THP-1 cells.
BA, fat & glucose
homeostasis
Inflammation (NF-κB)
Enterohepatic BA circulation
microbiota
(deconjugation, 1°→ 2°)
Journal of Hepatology (2011)
FXRα
VDR
PXR



• Lb. reuteri selected for Bile Salt Hydrolase activity (2 capsules/day at 2 x 109 CFU/capsule) for 9 weeks
• Randomized, double-blind, placebo-controlled, parallel-arm, multicenter study
• N=127 hypercholesterolemic patients
• Probiotic reduced plasma – TC by 9.14%
– LDL-C by 11.64%
– LDL-C/HDL-C ratio by 13.39%
– Non-cholesterol plant sterols
– Increased circulating deconjugated bile acids
• Proposed new cholesterol lowering activity of probiotics via modified absorption of lipids from the gut


Apple pectin and polyphenols increase faecal
excretion of bile acids and cholesterol derivatives

Apple pectin and polyphenols impact on lipid
metabolism and storage

TMA/TMAO confirmed strong link with CVD in patients •confirmed microbiota metabolism of L-carnitine/choline → TMA→TMAO •TMA not produced in vegans •confirmed inflammatory activity & linked to macrophage reverse cholesterol transport •TMAO reduced bile acid pool
Koeth et al., 2013 Nature Medicine
Gut flora metabolism of phosphatidylcholine
promotes cardiovascular disease
Wang et al., 2011 Nature

Established Therapies
• Unabsorbed antibiotics
(rifaxamin)
• Lactulose (prebiotic)
• Adjunt therapies: probiotics
Hepatic Encaphalopathy, a complication
of end stage liver failure


Cani et al., 2007 Diabetes
Plasma endotoxin (LPS) increased
with HF diet
• Upon high fat feeding or LPS injection inflammatory markers increased in liver and adipose tissue
– TNF-α, IL-1, IL-6
– insulin resistance and obesity
• In CD14 mutant mice this inflammatory response was blunted

Prebiotic relief of metabolic endotoxemia
through improved mucosal barrier function
LPS LPS LPS
High fat, low CHO diets induce microbiota dysbiosis
Low SCFA
•↑Inflammation •↑Insulin resistance •↑Hepatic fat deposition
Activation of WAT & liver inflammatory pathways
Prebiotic induced bifidogenesis & microbiota biosis
↑GLP-1, GLP-2, PYY ↑Tight junction proteins
High
SCFA
•↑ Satiety •↓Food intake •Maintenance of gut barrier function •Immune homeostasis
Mucosal
barrier

Metabolite Normal faecal concentration range*
Evidence for beneficial or harmful effects
Short chain fatty acid
•Acetate •Butyrate • Propionate
61-120 µmoles/g wet wt, 190-600 µmoles/g dry wt
Acetate: provides energy, substrate for lipogenesis and cholesterol Butyrate: provides energy for colonocytes, stimulates apoptosis and cell proliferation, inhibits histone deacetylase, regulates gut barrier and immune function Propionate: increases satiety, inhibits lipogenesis, increases absorption of water, regulates gut barrier and immune function
Branched chain fatty acids
2-3 µmoles/g wet wt Approx. 30 µmoles/g dry wt
Potentially harmful but also act against some less desirable bacteria
Hydrogen sulfide
Total sulfide 0.05-1.6 μmoles/g wet wt
Limited evidence in humans. Pro- and anti-inflammatory effects on colonic cells. Potentially toxic to colonic cells. Inhibits butyrate oxidation in vitro.
Ammonia 14-660 µmoles/g faeces No convincing evidence of harmful effects in healthy humans
Phenols
•p-Cresol
•Phenol
•Indole
Colonic generation rate is often
estimated from urinary
excretion/day
10-55 mg/d (urine), 5-8 mg/d
(faeces)
4-7.5 mg/d (urine), 0.25-0.66 mg/d
(faeces)
<50 mg/d (urine)
• Rapidly absorbed by colonic mucosa and excreted in urine
• Do not accumulate in healthy subjects
• Often considered as potentially toxic but no evidence for toxicity
in healthy humans
From polyphenols
•Simple phenols •Glycinated benzoic acids •Derivatives of
• Benzoic acid • Phenyl acetic acid • Phenylpropionic acid • Mandelic acids • Cinnamic acids
• Equol
varying concentration from
µmoles/l
(e.g. hydroxybenzoic acids)
to mmoles/l (e.g. phenyl propionic
acid)
Possible benefits in colon (anti-inflammatory, anti-oxidant)
but not well tested
*The normal concentration range is based on adults. This range may differ in stages of life as well as regions in the world. For detailed information on the references of the above-mentioned concentration values, please contact [email protected]

Obese vs lean gut microbiota
Lean –open diet (LOD ■) Obese open diet (OOD ■) Obese on a saturated fat diet for 1 month (OHSFA ■) n=13
•Bacteroides uniformis and Prevotella copri more common in the microbiota of LOD than OOD – not present in the OHSFA
•Bacteroides vulgatus and Bacteroindes stercoris very frequently found in OHSFA, less frequent in OOD – not present in LOD
-5
0
5
-10 -9 -8 -7 -6 -5 -4 -3 -2 -1 0 1 2 3 4 5 6 7 8 9 10
PC
2
PC1
DGGE bands pattern (PCA)
[PC2]/[PC1]
%[1] = 0.092199 %[2] = 0.0642478
LOD
OHS
OOD
Fava et al in preparation

Fermentation end-products
• Faecal SCFA measured by GC
• Higher acetate in obese irrespective of diet
• Higher butyrate in obese (open diet)
0,0
5,0
10,0
15,0
20,0
25,0
30,0
35,0
40,0
ACETIC
PRO
PIO
NIC
I-BUTY
RIC
N-B
UTY
RIC
I-VALE
RIC
N-V
ALE
RIC
N-C
APRO
IC
mm
ol/g
faeces
LOD
OOD
OHSFA


SCFA as biomarker of healthy gut Increased faecal SCFA excretion due to decreased SCFA uptake?
MCT transporters
expression and
apical location is
promoted by
luminal SCFA
Increased faecal SCFA excretion could be due to decreased MCT active uptake in high fat/low CHO diets
Borthakur et al., 2012 Am J Physiol Gastrointest Liver Physiol
Goncalves et al., 2012 J Cell Biochemistry
Bile salt CDCA
and E. coli EPEC
inhibit butyrate
uptake

Biohydrogenation
• Recognised & studied in
rumenants
• Under studied & bearly
recognised in humans
Rosburia
Butryvibrio
Lactobacillus
Bifidobacterium

Lipids 2010

Chen et al. (2015)
Gastroenterology 148:203-214

Chen et al. (2015)
Gastroenterology 148:203-214

Chen et al. (2015)
Gastroenterology 148:203-214

•POLYPHENOLS….. 90% resistant to digestion and reach the colon, plant secondary metabolites, usually antioxidant, antimicrobial activities, enzyme/nutrient binding properties and possibly prebiotic type properties, e.g. red-wine polyphenols, apple tannins
Polyphenols

When to drink red wine, the French
paradox revisited?
Natella et al. Brit J Nutr (2011)

“the modality of drinking wine (during the
meal) could represent a decisive factor”
Natella et al. Brit J Nutr (2011)

American Journal of Clinical Nutrition, 2012
0
2
4
6
8
10
12
14
Ent
eroc
occu
s
Lactob
acillus
Clostrid
ium
Clostrid
ium
histo
lytic
um g
roup
Blaut
ia coc
coid
es-E
ubac
teriu
m re
ctal
e gr
oup
Bac
tero
ides
Bac
tero
ides
unifo
rmis
Pre
vote
lla
Bifido
bacter
ium
Egg
erth
ella
lent
a
16S
rR
NA
gen
e c
op
ies/g
faeces,
mean
+/-
SD
, n
=10
Basline
De-alcoholized red wine
Red wine
Gin
* * * *
* *
*
* *


Targeting Microbiota Polyphenol
metabolism
• Targeted MS based quantitative metabolite analysis
•139 fruit polyphenols
•UPLC/QqQ-MS/MS
•Modified to accurately quantitify about 150 common fruit polyphenols
and their catabolites

Microbiota modulation - in vitro
faecal batch cultures
•4 commercial apples
•Simulated gastric and small intestinal digestion
•Fermentation pH and temperature controlled anaerobic
faecal batch cultures
•FISH microbial enumeration
•Profile of microbial polyphenol catabolites

GASTRO-DUODENAL DIGESTION IN VITRO vs IN VIVO
pH 2.5 Add gastric pepsin & lipases 37°C/ 2h
pH 6 (6N NaOH) Bile salts & enzymes (amylase, trypsin,
chymotrypsin,
colipase ) 37°C/ 1h
•Strawberry (commercial var.) •Wild Strawberry
In collaboration with
Dr Chris Gill,
Ulster University

200 g Wild Strawberry “smoothy”
•Ileal fluid (n=5) collected from ileostomy patients before (T0) and after (T8) consuming 200 g Wild Strawberry smoothy (WS). •5 healthy controls fed in parallel
Ileostomy bag
GASTRO-DUODENAL DIGESTION IN VITRO vs IN VIVO
In collaboration with
Dr Chris Gill,
Ulster University

Increasing fruit and vegetable intake
in vivo – FLAVURS project
Flavonoid-poor F&V
Flavonoid-rich F&V +2 + 4
+2
+ 6
+ 4 + 6
Habitual diet
Wk 0
Visit 1 Wk 6
Visit 2
Wk 12
Visit 3 Wk 18
Visit 4
?

Metabolomics workflow
Sample preparation:
extraction of all analytes
Statistical analysis Untargeted HR mass
spectrometry
Biomarker identification
Samples: urine,
plasma, fecal water
C+
NHO
F
CH3
NN
CH3
Separation on
LC column





Measuring the effect of apples (2 per day)
on the gut microbiome and heart health.
...from Trentino with love!

“Conslusions: Adherence to an MD pattern is associated with better HRQL. The
association is stronger with mental health than with physical health. Dietary total
antioxidant and fibre content independently explain this relationship”.

• Children with Prader-Willi syndrome (PWS), n=17 and children with diet associated “standard” obesity (n=21)
• Weight loss induced by reduced calorie intake and high fiber & polyphenol diet (whole grain/legume “grule”, plus 20g/prebiotic per day, fruit/veg, medicinal Chinese herbs)

Dietary intervention in obese children Before 30 day 60 day 90 day
Total Energy Intake 1559.56 (3577.4-916.2) 1232.7 (1992.4-858.3) 1167.9 (1648.8-74.2) 1169.2 (1919.3-940.7)
Protein (g) 46.9 (103.8-19.6) 46.2 (83.1-18.6) 38.2 (61.5-26.1) 36.9 (74.3-20.9)
Protein (%) 13. 6±1.1 13.7±0.5 13.0±0.5 12.7±0.5
Lipid (g) 64.6 (136.5-15.8) 26.6 (40.2-14.0) 25.2 (41.4-18.3) 24.8 (45.8-14.5)
Lipid (%) 34.0±4.0 19.7±0.4*** 20.3±0.7*** 19.7±0.6***
Carbohydrates (g) 218.3 (484.7-54.8) 191.9 (294.7-141.1) 189.7 (252.4-106.7) 187.1 (261.6-142.9)
Carbohydrates (%) 52.4±4.4 62.4±0.7* 62.6±0.7* 62.5±0.9*
Fiber (g) 6.2 (16.6-1.6)a 44.9 (58.7-30.7)b 48.5 (59.9-24.4)b 49.4 (66.9-37.3)b
Soluble fiber (g) 2.7 (7.2-0.7) 29.2 (40.9-10.7) 31.6 (39.3-11.1) 32.7 (45.6-23.7)
Zhang et al., 2015

Significantly reduced obesity and improved metabolic parameters in both genetically obese and diet associated obese children.
Zhang et al., 2015

Specific microbiota modulation
Zhang et al., 2015
•Dietary interventions modulate
gut microbiota (increased relative
abundance of Bifidobacterium
and other fiber degraders.
•Dietary interventions reduced
relative abundance of organisms
involved in TMAO, TRP and choline
metabolism

Summary • Metabolites derived from microbial activities in the gut play an
important role in liver disease risk (BA, lipid metabolism, inflammation)
• Probiotics (BSH), prebiotics/fiber and polyphenols all have potential to modulate both the flux of these metabolites and gut permeability/inflammatory output
• High resolution omics technologies have provided the necessary tools for tracking microbiome metabolic output and community structure/function
• Currently lack appropriately designed human studies to confirm theories generated from animal studies and human «case / control» observations


Fondazione Edmund Mach
• Thank you: ILSI Euroope, Prof Ian Rowland
• Fulvio Mattivi, Marynka Ulaszewska, Claudio Donati,
Fondazione Edmund Mach
• NN Group: Francesca Fava, Elena Franciosi, Athanasios Koutsos, Ilaria Caraffa,
Florencia Ceppa, Andrea Mancini
• University of Ulster: Dr Chris Gill
• University of Reading, Ian Rowland, Glenn Gibson, Julie Lovegrove, Parveen
Yaqoob, Christine Williams, John Swann














![IN THE SPOTLIGHT: The Microbiome and Cardiac Transplantation… Documents... · 2018. 4. 3. · after liver transplantation [20]. Tacrolimus and mammalian target of rapamycin inhibitor](https://static.fdocuments.net/doc/165x107/602d1d0a0ca80f72b650fe50/in-the-spotlight-the-microbiome-and-cardiac-transplantation-documents-2018.jpg)



