
IMMUNOLOGY - National Science Digital Library at NISCAIR, India: Home
19
IMMUNOLOGY Immunoglobulin: Structure and Function Ashutosh Tiwari and Subrata Sinha Department of Biochemistry All India Institute of Medical Sciences New Delhi - 110029 CONTENTS Introduction Basic Antibody Structure Molecular structure of antibody Complementarity Determining Regions (CDRs) Antigen binding site Constant domains Classes subclasses and biological activities of antibodies Antigenic determinants of antibodies Monoclonal antibodies Functions of antibody Keywords Antibody, Immunoglobulin, Complementary Determining Regions(CDRs), Hinge Region, Constant Domains, Isotype, Allotype, Idiotype, Monoclonal antibody
Transcript of IMMUNOLOGY - National Science Digital Library at NISCAIR, India: Home

IMMUNOLOGY
Immunoglobulin: Structure and Function
Ashutosh Tiwari and Subrata Sinha Department of Biochemistry
All India Institute of Medical Sciences New Delhi - 110029
CONTENTS IntroductionBasic Antibody StructureMolecular structure of antibody Complementarity Determining Regions (CDRs) Antigen binding site Constant domainsClasses subclasses and biological activities of antibodiesAntigenic determinants of antibodiesMonoclonal antibodiesFunctions of antibody
Keywords Antibody, Immunoglobulin, Complementary Determining Regions(CDRs), Hinge Region, Constant Domains, Isotype, Allotype, Idiotype, Monoclonal antibody

Introduction
The antibody molecule is a Y-shaped protein present on the surface of B cells that is secreted into the blood or lymph in response to an antigenic stimulus, such as a bacterium, virus, parasite, or transplanted organ, and that neutralizes the antigen by binding specifically to it. Antibody molecule has evolved to perform two distinct functions- antigen recognition and antigen elimination. The antibody molecule has discrete globular domains to accomplish these two functions. One of these domains binds to the antigen, and the others mediate the neutralization functions. This chapter considers antibody structure and its relation to antibody functions. Basic Antibody Structure
The serum of a normal vertebrate contains a large variety of proteins. The globulin proteins of serum can be divided into three major fractions α, β and γ. The γ-globulin fraction of these proteins was identified as containing serum antibodies. To distinguish these antibodies from any other proteins present in γ-globulin fraction, these were called as immunoglobulins. With more information on the different class of antibodies it has been established that all antibodies need not necessarily present in the γ-globulin fraction. Significant amounts of immunoglobulin G (IgG), main class of antibody molecule and other important classes of antibody molecules are found in the α and the β fractions of serum. Antibodies consist of four polypeptide chains
Antibody molecules are made up two identical polypeptides of molecular weight of about 25,000 called light chains (L) and two identical larger polypeptides of molecular weight 50,000 or more called heavy chains (H) (Fig.1). Each light chain is linked to a heavy chain by noncovalent associations like hydrogen bonds, hydrophobic bonds and salt linkages and also by one covalent disulfide bridge, to form a heterodimer (H-L). The two identical light chain-heavy chain combinations (H-L) are linked together by similar noncovalent interactions and disulfide bridge to form the basic four chain (H-L)2 antibody structure, a dimmer of dimmers. The exact number and precise positions of these interchain disulfide bonds differ among antibody classes and subclasses. As shown in Fig.1, the molecule can be represented schematically in the form a Y, with the amino (N) termini of the four chains at the top and the carboxyl (C) termini of the two heavy chains at the bottom.
Variable and constant domains of light and heavy chain
When the amino acid sequences of various light chains and heavy chains were compared, a striking pattern emerged. In the amino terminal half of the polypeptide, the amino acid sequences were found to differ greatly among antibodies of different specificity. These regions of highly variable sequences were called V regions: VL in light chains and VH in heavy chains. By contrast, in the carboxy terminal half of the polypeptide, the sequences of all the antibody molecules were almost identical and these identical segments were called as constant (C) region. The variable region (V) begins at the amino terminus and is approximately 110 amino acid residues in length. The constant region (C) makes up the remainder of the chain, about 110 residues in length in case of light chain (CL) and about 330 residues long in case of heavy chain (CH) for the IgG molecule. Antibodies are glycoproteins; with few exceptions, constant region of heavy chains are involved in the attachment for carbohydrates. The glycosylation increases the
2

solubility of the molecules. Inappropriate glycosylation, or its absence, affects the rate at which antibodies are cleared from the serum, and it also renders the efficiency of interaction between antibody and the complement system and between antibodies and Fc receptors.
Fig.1: Schematic diagram of an antibody molecule composed of two light chains and two heavy chains. The amino terminal domains, corresponding to the variable regions, form the antigen binding site. The carboxy terminal domains (constant regions) mediate the effector
functions of the antibody. Enzymatic and chemical method unfolds the basic antibody structure
When the γ-globulin fraction of serum was separated into high and low molecular weight fractions, antibodies of around 150,000 MW (Low mol.wt. fraction) were designated as immunoglobulin G (IgG). When the IgG was subjected to brief digestion with the enzyme papain, three fragments were produced, two of which were identical fragments (each with a MW of 45,000), had antigen binding activity and were called Fab fragments (‘Fragment- antigen binding‘). The other third fragment (MW of 50,000) has no antigen binding activity at all and has ability to crystallize during cold storage. It is called as Fc fragment (‘Fragment crystallizable’). Pepsin digestion gives a single fragment of 100,000 MW composed of two Fab-like fragments termed as F(ab’)2 fragment and a Fc fragment. Hetrodimer nature of IgG was deduced when the molecule was subjected to mercaptoethanol reduction and alkylation by chemicals (irreversible cleavage of disulfide bonds). When the treated IgG molecules were chromatographed on a column that separates molecules by size
3
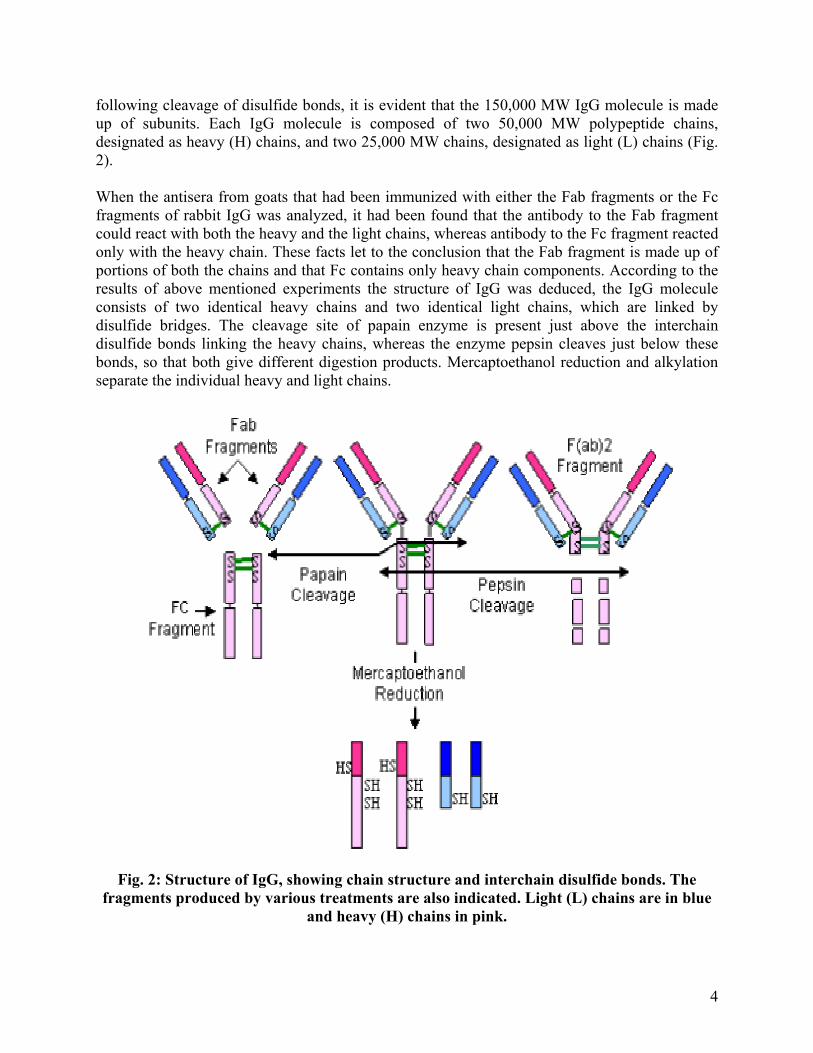
following cleavage of disulfide bonds, it is evident that the 150,000 MW IgG molecule is made up of subunits. Each IgG molecule is composed of two 50,000 MW polypeptide chains, designated as heavy (H) chains, and two 25,000 MW chains, designated as light (L) chains (Fig. 2). When the antisera from goats that had been immunized with either the Fab fragments or the Fc fragments of rabbit IgG was analyzed, it had been found that the antibody to the Fab fragment could react with both the heavy and the light chains, whereas antibody to the Fc fragment reacted only with the heavy chain. These facts let to the conclusion that the Fab fragment is made up of portions of both the chains and that Fc contains only heavy chain components. According to the results of above mentioned experiments the structure of IgG was deduced, the IgG molecule consists of two identical heavy chains and two identical light chains, which are linked by disulfide bridges. The cleavage site of papain enzyme is present just above the interchain disulfide bonds linking the heavy chains, whereas the enzyme pepsin cleaves just below these bonds, so that both give different digestion products. Mercaptoethanol reduction and alkylation separate the individual heavy and light chains.
Fig. 2: Structure of IgG, showing chain structure and interchain disulfide bonds. The fragments produced by various treatments are also indicated. Light (L) chains are in blue
and heavy (H) chains in pink.
4

Sequencing of Light and Heavy chains
Initially, to determine the amino acid sequence of heavy and light chain was held up because of the insufficient amounts of homogeneous protein were available. The heterogeneity of serum antibodies made them unsuitable for sequencing studies. The discovery of multiple myeloma, a cancer of antibody producing plasma cells made the sequencing of antibody chains feasible. In this case a cancerous plasma cell, called a myeloma cell, has been transformed and its protein synthesizing machinery and secretary functions are not altered; thus the cell continues to produce homogeneous antibody. This antibody is indistinguishable from normal antibody molecules but is called as myeloma protein. In most patients, the myeloma cells also secrete excessive amounts of light chains. These excessive light chains were discovered in the urine of myeloma patients and were named Bence-Jones proteins, on the name of their discoverer. Light Chain
When the amino acid sequences of several myeloma light chains were first compared, a striking pattern emerged. The N-terminal half of the polypeptide chains, consisting of 100-110 amino acids, were found to vary greatly among different Bence-Jones proteins. This region was called the variable (V) region. By contrast, in the C-terminal half of the polypeptide, called the constant (C) region, had two basic amino acid sequences. This information deduced the fact that there were two light chain types, kappa (κ) and lambda (λ). In humans, 60% of the light chains are kappa and the remaining 40% are lambda, whereas in mice, 95% of the light chains are kappa and only 5% are lambda. There are four subtypes of lambda chain present in humans, whereas in mice, there are three subtypes (λ1, λ2 and λ3). Amino acid substitutions at some positions are accountable for the subtype differences. A single antibody molecule contains only one light chain type, either kappa (κ) or lambda (λ), never both. Heavy Chain
For the heavy chain sequencing studies, first the myeloma proteins are separated by mercaptoethanol reduction followed by gel filtration chromatography in a denaturing solution. The amino-terminal part of the chain consisting of 100-110 amino acids, showed great sequence variation among myeloma heavy chains just like myeloma light chains and was therefore called variable (VH) region. The remaining carboxy-terminal portion of the myeloma heavy chains revealed five basic sequence patterns. On the basis of these basic sequence patterns, constant region of heavy chains characterized into five isotypes (µ, δ, γ, ε and α). The length of the constant regions is approximately 330 amino acids for δ, γ, and α, and 440 amino acids for µ and ε. The heavy chains of the given antibody molecule determines the class of that antibody: IgM (µ), IgG (γ), IgA (α), IgD (δ) or IgE (ε). Each class can have either kappa (κ) or lambda (λ) light chain. A single antibody molecule has two identical light chains and two identical heavy chains, H2-L2 or a multiple (H2-L2) n of this basic four chain structure (Table. A). Minor differences in the amino acid sequences of the α and γ heavy chains led to further classification of the heavy chains into their subtypes. In humans, there are two subisotypes of α heavy chains – α1 and α2 (thus two subclasses – IgA1 and IgA2), four subisotypes of γ heavy chains: γ1, γ2, γ3, γ4 (IgG1, IgG2, IgG3 and IgG4). In mice, there are four subisotypes, γ1, γ2a, γ2b, and γ3, and the corresponding subclasses.
5

Table A: Chain composition of the five immunoglobulin classes in humans
Class Light Chain Heavy Chain Subclasses Molecular Formula
IgG
IgM
IgA
IgE
IgD
κ or λ
κ or λ
κ or λ
κ or λ
κ or λ
γ
µ
α
ε
δ
γ1, γ2, γ3, γ4
None
α1, α2
None
None
γ2κ2 or γ2λ2
(µ2κ2)n or (µ2λ2)n, n=1or5
(α2κ2)n or (α2λ2)n, n=1to4
ε2κ2 or ε2λ2
δ2κ2 or δ2λ2 Molecular structure of Antibody
It has been found that both heavy and light chains are made up of several homologous units of 110 amino acid residues termed as domain. Each domain forms a loop of 60 amino acids by an intrachain disulfide bond. Light chains contain one variable domain (VL) and one constant domain (CL), while heavy chains are made up of one variable domain (VH) and either three or four constant domains (CH1, CH2, CH3, and CH4), depending on the class of antibody. The fine structure of the immunoglobulin domains was solved by X-ray crystallography, which revealed that the domains are folded into a characteristic compact structure called ‘immunoglobulin fold’. This fold consists of two β-pleated sheets stabilized by hydrophobic interactions and a disulfide bond. Each β-pleated sheet is made up of antiparallel β strands of amino acids, which are connected by loop like structure of various lengths. The β strands within a sheet are stabilized by hydrogen bonds and the two β sheets within an immunoglobulin fold are stabilized by the hydrophobic interactions between them and by the conserved disulfide bond (Fig.3A). Although variable and constant domains have similar structure, but variable domain is slightly longer than the constant domain and have an extra pair of β strands and one extra loop within the β sheet structure (Fig.3B). The quaternary structure of the immunoglobulins is formed by facilitating noncovalent interactions between immunoglobulin domains across the faces of the β sheets. These interactions form link between nonidentical domains (VH/CL and CH1/CL) and between identical domains (CH2/CH2, CH3/CH3 and CH4/CH4). (Fig.4) Complementarity Determining Regions (CDRs)
By comparing the amino acid sequences of various VH and VL domains, it has been noted that variation in the amino acid sequences is highly concentrated in a few discrete regions of these domains. The maximum variability in the amino acid sequences is seen in those portions of the sequence that correspond to the loops that join the β strands (Fig.5). These regions were originally called hypervariable regions because of the high variability in sequence. These hypervariable regions form the antigen binding site of the antibody molecule. Since antigen binding site is complementary to the structure of the epitope, these regions are also called as
6
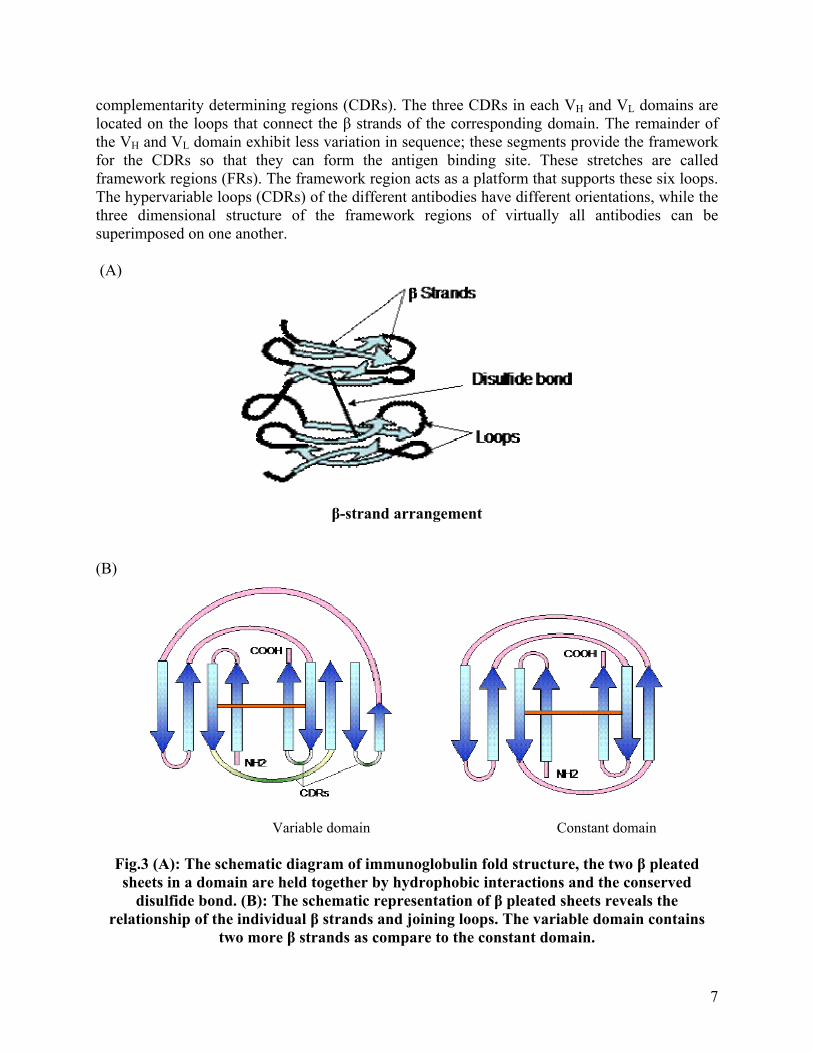
complementarity determining regions (CDRs). The three CDRs in each VH and VL domains are located on the loops that connect the β strands of the corresponding domain. The remainder of the VH and VL domain exhibit less variation in sequence; these segments provide the framework for the CDRs so that they can form the antigen binding site. These stretches are called framework regions (FRs). The framework region acts as a platform that supports these six loops. The hypervariable loops (CDRs) of the different antibodies have different orientations, while the three dimensional structure of the framework regions of virtually all antibodies can be superimposed on one another. (A)
β-strand arrangement (B)
Variable domain Constant domain
Fig.3 (A): The schematic diagram of immunoglobulin fold structure, the two β pleated sheets in a domain are held together by hydrophobic interactions and the conserved
disulfide bond. (B): The schematic representation of β pleated sheets reveals the relationship of the individual β strands and joining loops. The variable domain contains
two more β strands as compare to the constant domain.
7

Fig.4: Schematic representation of IgG molecule, showing associations between heavy and light chain domains. Carbohydrate moieties make the CH2/CH2 domains more accessible to
interact with complement components
Fig.5: Variability of amino acid residues in the variable region of heavy chain and light
chain of antibodies with different specificity. Three hypervariable regions (HV), also called complementarity-determining regions (CDRs) are present in both heavy and light chain
variable domains
8

Antigen Binding Site
Crystallographic analysis of many antigen antibody complex revealed that the complementarity determining regions (CDRs) are the antigen binding regions of the antibody molecule. The actual shape of the antigen-binding site is formed by the combination of CDRs of that particular antibody. X-ray diffraction analysis of many antibody-antigen complexes has shown that several CDRs may make contact with the antigen, and it has been observed in many of the complexes that all six CDRs contact the antigen. It has been also observed that the VH domain often contributes more to antigen binding than the VL domain. It is also evident in some cases; formation of antigen-antibody complex induces conformational changes in the antibody, antigen or both. This conformational change results in a closer fit between the epitope and the antibody’s binding site. In case of antibody these changes may occur in the orientation of the side chains of the antigen binding site and/or by overall conformational shift of the CDR loops. In addition to variability in the length and amino acid composition of the CDR loops, the ability of these loops to significantly change conformation upon antigen binding enables antibodies to attain a shape more effectively complementary to that of their epitope. Constant Domains
The constant domains of immunoglobulin participate in various biological functions that are determined by the amino acid sequence of each domain. CH1 and CL domains
The CH1 and CL domains help to hold the VH and VL domains together with the help of interchain disulfide bond between them (Fig.1). The random association between VH and VL domains and the stability of VH and VL interactions are mainly determined by the CH1 and CL domains, thus contributing to the overall diversity of antibody molecules that can be expressed by any individual. Hinge Region
This is the region at which the arms of the antibody molecule form a “Y” shape. It is called the hinge region because there is some flexibility in the antibody molecule at this point. This region is rich in proline and cystein residues. The proline residues confer an extended conformation and cystein residues provide sites for disulfide bonds which connect the two heavy chains in this region. The length and number of disulfide bonds is variable in hinge region in different classes and sub-classes of immunoglobulins (Fig.6). The hinge region is quite flexible and allows wide degree of angular movement of the Fab fragment, this helps in antigen binding. The IgE and IgM do not have a typical hinge region, CH2 domain of the heavy chains act as hinge region in case of these antibodies. Other Constant Region Domain
The heavy chains of IgG, IgA and IgD contain three constant region domains (CH1, CH2 and CH3) and a hinge region, whereas four constant region domains (CH1, CH2, CH3 and CH4) are present in case of IgE and IgM. There is no hinge region present in both these classes of immunoglobulins. The CH2/CH2 domains in IgE and IgM occupy the same position in the chains as the hinge region in the other classes of immunoglobulins. The function of this extra domain has not yet been determined. Structural analysis by X-ray crystallography revealed that
9

both the CH2 domains of IgG, IgA and IgD (and the CH3 domains of IgE and IgM) are separated by oligosaccharide side chains; as a result these two globular domains are more accessible to the aqueous environment. The solvent accessibility of these domains is one of the key elements that contribute to the biological activity by activating complement components by IgG and IgM.
(A) γ, δ, α (B) µ, ε
Fig.6: (A) Schematic representation of domains of heavy and light chains, each domain is made up of 110 amino acid residues and an intrachain disulfide bond that forms a loop of
60 amino acids. Variable domains form the antigen binding site; while other domains mediate the effectors functions. (B) No hinge region in case of µ (IgM), ε (IgE) heavy
chains, and an additional domain replaces the hinge region. The last constant region domain of heavy chain is referred to as the carboxyl-terminal domain. In case of IgG, IgA and IgD, CH3/CH3 domains are designated as C-terminal domains whereas in case of IgE and IgM, one more domain is present at their C-terminal end designated as CH4/CH4. The five classes of antibody and their subclasses can be expressed either as secreted immunoglobulin or as membrane bound immunoglobulin. Secreted immunoglobulin has a hydrophilic amino acid sequence of various lengths at the carboxyl terminal end. In case of membrane bound immunoglobulin, the carboxyl-terminal domain contains a hydrophobic transmembrane sequence, a short cytoplasmic tail and an extracellular hydrophilic “spacer” sequence. The length of the transmembrane sequence is same among all immunoglobulin isotypes, whereas the lengths of spacer sequence and cytoplasmic tail vary.
10

Secreted IgM (sIgM) Membrane bound IgM (mIgM) Secreted IgM (sIgM) Membrane bound IgM (mIgM)
Fig.7: Structure of carboxyl-terminal domains of heavy chain in secreted and membrane bound immunoglobulin
Classes, Subclasses and Biological Activities of Antibodies
The classes and various isotypes of immunoglobulin have been discussed briefly earlier. Each class has unique amino acid sequences in the heavy chain constant region that determines class specific structural and functional properties. All classes of immunoglobulins are found in every human being. The subclasses within the class are found because of the subtle differences in the amino acid sequences of heavy chains of the same class. Immunoglobulin G (IgG)
It is the most abundant antibody in serum and constitutes about 75-80% of the total serum immunoglobulin. It has the longest half life of 23-25 days. The IgG molecule consists of two γ heavy chains and two κ or two λ light chains (Fig.8). There are four human IgG subclasses distinguished by differences in the γ heavy chain sequence and numbered according to their decreasing average serum concentration: IgG1, IgG2, IgG3 and IgG4.
• IgG1, IgG3 and IgG4 provide immunity to the developing fetus because they can readily cross the placenta and enter into neonatal circulation.
• IgG1 and IgG3 bind with high affinity to Fc receptors on the phagocytic cells and thus mediate phagocytosis. IgG4 has moderate affinity for Fc receptors while IgG2 has an extremely low affinity.
• IgG3 is the most effective complement activator, followed by IgG1; IgG2 is less efficient and IgG4 is not able to activate complement at all.
• IgG plays a major role in neutralization of toxins because it can easily diffuse into extravascular spaces.
11

Immunoglobulin M (IgM)
IgM is the first immunoglobulin class produced in a primary response to an antigen, and it is also the first immunoglobulin to be synthesized by the neonate. IgM accounts for 5-10% of the total serum immunoglobulin. IgM is secreted by plasma cells as a pentamer in which five monomer units are held together by disulfide bonds. The five monomer subunits are arranged with their Fc regions in the center of the pentamer and the ten antigen binding sites on the periphery of the molecule. Each pentamer contains an additional polypeptide called the “J” (joining) chain, which is linked by two of the ten carboxyl terminal domains. The J chain is required for the polymerization of the monomers to form pentameric IgM. Monomeric IgM is also expressed as membrane bound antibody on B cells. Pentameric IgM has 10 antigen-binding sites thus it is more efficient than other isotypes in binding antigens with many repeating epitopes such as viral particles. Less IgM than IgG is required to neutralize viral infectivity. IgM is also more efficient than IgG at activating complement. Complement activation requires two Fc regions in close proximity, and the pentameric structure of single IgM molecules fulfills this requirement. IgM molecule does not diffuse well because of the large size, therefore is found in low concentrations in intercellular tissue fluids. J chain facilitates the binding of IgM to receptors on secretary cells, which transports it across epithelial linings to enter the external secretions that immerse mucosal surfaces. Immunoglobulin A (IgA)
It is the predominant immunoglobulin class in external secretion such as saliva, tears, breast milk, respiratory secretion, and genitourinary and digestive track secretions. IgA constitutes 10-15% of the total immunoglobulin in serum. IgA exists primarily as a monomer, but sometimes polymeric forms (dimers, trimers and some tetramers) containing J chain polypeptide is also seen (Fig.8). The secretory IgA consists of a dimer or tetramer, a J chain polypeptide and a secretory component. The J chain polypeptide in IgA facilitates polymerization function like in case of IgM. The secretory component is produced by epithelial cells of mucous membranes. The secretory component is made up of five immunoglobulin like domains that binds to the Fc region of IgA dimer. (Fig.8) The daily production of secretory IgA is greater than that of any other immunoglobulin class. IgA secreting plasma cells are concentrated along mucous membrane surfaces. The plasma cells that produce IgA preferentially migrate to subepithelial tissue, where secretory IgA binds to poly-Ig receptor present on the basolateral surface of most mucosal epithelia. After IgA binds to poly-Ig receptor, the receptor-IgA complex is then transported across the epithelial cell to luminal membrane by receptor mediated endocytosis. The poly-Ig receptor is then cleaved enzymatically from the membrane and becomes the secretory component, which is bound to and released together with polymeric IgA into mucosal secretion. The secretory component masks the sites susceptible to protease cleavage thus allowing the polymeric IgA to exist longer in the protease rich mucosal environment. Pentameric IgM is also transported into mucosal secretion by this mechanism. Polymeric secretory IgA helps in trapping the pathogen like bacteria and viruses by preventing their attachment to the epithelial cell wall. Secretory IgA can cross-link large antigens with multiple epitopes thus inhibiting viral infection and bacterial colonization. Complexes of secretory IgA and antigen are easily entrapped in mucous and then eliminated by ciliated
12
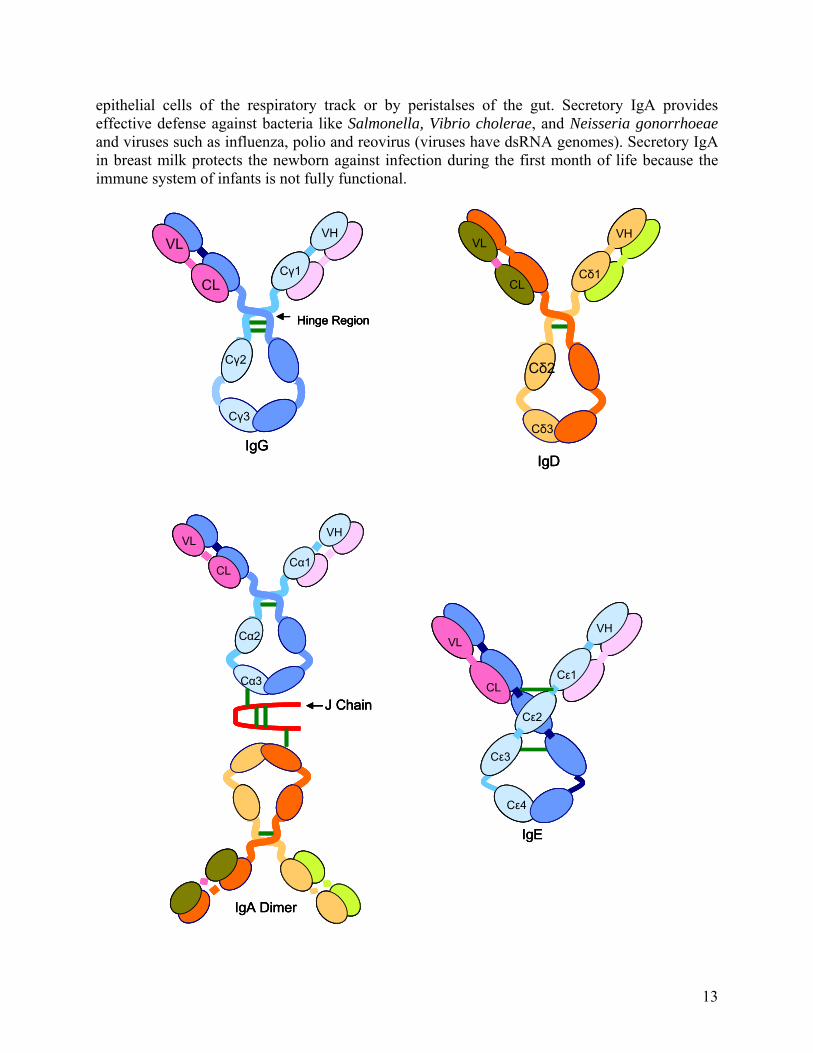
epithelial cells of the respiratory track or by peristalses of the gut. Secretory IgA provides effective defense against bacteria like Salmonella, Vibrio cholerae, and Neisseria gonorrhoeae and viruses such as influenza, polio and reovirus (viruses have dsRNA genomes). Secretory IgA in breast milk protects the newborn against infection during the first month of life because the immune system of infants is not fully functional.
VL
CL
VH
Cγ2
Cγ1
Cγ3
Hinge Region
IgG
CL
VL
Cδ2
VH
Cδ1
Cδ3
IgD
VL
CL
VH
Cγ2
Cγ1
Cγ3
Hinge Region
IgG
VL
CL
VH
Cγ2
Cγ1
Cγ3
Hinge Region
IgG
CL
VL
Cδ2
VH
Cδ1
Cδ3
IgD
CL
VL
Cδ2
VH
Cδ1
Cδ3
IgD
VHVL
CL
Cε2
Cε1
Cε3
Cε4
IgE
VL
CL
VH
Cα2
Cα1
Cα3
J Chain
IgA Dimer
VHVL
CL
Cε2
Cε1
Cε3
Cε4
IgE
VHVL
CL
Cε2
Cε1
Cε3
Cε4
IgE
VL
CL
VH
Cα2
Cα1
Cα3
J Chain
IgA Dimer
VL
CL
VH
Cα2
Cα1
Cα3
J Chain
IgA Dimer
13

J Chain
Disulfide bond
IgM
J Chain
Disulfide bond
J Chain
Disulfide bond
IgM
Fig.8: Schematic representation of general structures of the five major classes of antibody. IgG, IgA and IgD heavy chain contain four domains and a hinge region, whereas the IgM
and IgE heavy chains contain five domains but no hinge region. The polymeric form of IgA and IgM contain a polypeptide, called J chain that is linked by two disulfide bonds. Serum
IgM is always present as a pentamer
Immunoglobulin E (IgE)
The serum concentration of IgE is very low (0.3µg/ml) but it is very potent in action. It mediates the allergic reaction or hypersensitivity reactions like asthma, hay fever and anaphylaxis reactions. IgE binds to the Fc receptors on the membranes of basophils and mast cells. Cross linkage of receptor bound IgE by antigen (allergen) induces basophils and mast cells to translocate their granules to the plasma membrane and release their contents to the extra cellular environment, process known as degranulation. Therefore a variety of pharmacologically active mediators like histamine, bradykinin and other vasoactive mediators are released and give rise to allergic and anaphylaxis manifestations. Immunoglobulin D (IgD)
It is found on B-cell surface along with IgM and act as a receptor for antigen binding. It is present in low concentration (30µg/ml) and constitutes about 0.2% of the total immunoglobulin in serum. It activates the B cells after binding to the antigen. No other function is known of IgD. Antigenic Determinants of Antibodies
Antibody molecules are complex glycoprotein so they themselves can act as immunogens and induce anti-Ig antibodies against themselves. It is seen that the whole antibody molecule is not
14

immunogenic to the host system which produce antibodies to them. Small regions or sites on Ig molecule can act as immunogens and called as antigenic determinants. Based on their location on the Ig molecule, they are divided into three types: isotypes, allotypes and idiotypes. Isotype
These antigenic determinants formed by unique sequences located in the constant region of heavy chain and light chain in an Ig molecule. The isotypes collectively define each heavy chain class and sub class and each light chain type and subtype with in a species (Fig.9a). All members of a species carry the same constant-region genes thus express the same isotypes, while different species inherit different constant-region genes therefore express different isotypes. Therefore, when an antibody from one species is injected into another species, the isotypic determinant will be recognized as foreign and anti-isotype antibody will be expressed. Allotype
With in a species all the members inherit the same set of isotype genes, but multiple alleles exist for some of the genes (Fig.9b). These alleles encode slight amino acid difference in the constant-region domains called allotypic determinants. Allotypic determinants occur in some, but not all, members of a species. Antibody to allotypic determinants can be produced by injecting antibodies from one member of a species into another member of the same species who carries different allotypic determinants. Antibodies to allotypic determinants can also arise from a blood transfusion. Idiotype
The amino acid sequence of the variable domain of heavy chain and light chain of a given antibody can not only function as antigen binding site but also as a set of antigenic determinants. Each individual antigenic determinants of the variable region is referred to as an idiotype (Fig.9c). In some cases an idiotope may be the actual antigen binding site, and in some cases an idiotope may comprise variable region sequence away from the antigen binding site. Each antibody will present multiple idiotopes and the sum of the individual idiotopes is called the idiotype of the antibody. Idiotopes arises from the mutations in the variable region genes during affinity maturation, mainly by somatic hypermutations. The antibodies produced against these idiotypes are called as anti-idiotypic antibodies, which are present in small quantities in normal individuals. They are increased in case of autoimmune disease. (a) Isotypic determinants
15

(b) Allotypic determinants
(c) Idiotypic determinants
Fig. 9: Types of Antigenic Determinants of Antibodies Monoclonal Antibodies
The most of the antigens comprise multiple epitopes and therefore induce proliferation and differentiation of a variety of B cells clones, each derived from a B cell that recognizes a particular epitope of that antigen. As a result serum antibodies are heterogeneous and mixture of antibodies, each specific for one epitope. Such a polyclonal antibody response facilitates the localization, phagocytosis and complement-mediated lysis of antigen in vivo. The polyclonal antibodies that increase immune protection in vivo often reduce the efficiency of an antiserum for various in vitro uses. So monoclonal antibodies, derived from single clone and specific for a single epitope are preferred for most research, diagnostic and therapeutic purposes. The biochemical purification of a monoclonal antibody from a polyclonal antibody preparation is not feasible. In 1975, George Kohler and Cesar Milstein devised a method ‘Hybridoma Technology’, for preparing monoclonal antibody, by fusing a normal activated antibody-producing B cells with a myeloma cell (a cancerous plasma cell). The fusion of these two cells leads to a hybrid cell, called as hybridoma that possessed the immortal growth properties of the myeloma cell and secreted the antibody produced by the B cell (Fig.10). The resulting clones of hybridoma cells, which secrete large quantities of monoclonal antibody, can be cultured indefinitely. Kohler and Milstein were awarded a Nobel Prize for this discovery.
16

Hybridoma screened for antibody production
Immortalized myeloma cells
Antibody producing cells fusedWith the myeloma cells
Antibody producing B cellsisolated from spleen
Immunization of mouse to stimulateantibody production
Culture of antibody producinghybridoma clones
Isolated monoclonal antibody
Hybridoma screened for antibody production
Immortalized myeloma cells
Antibody producing cells fusedWith the myeloma cells
Antibody producing B cellsisolated from spleen
Immunization of mouse to stimulateantibody production
Culture of antibody producinghybridoma clones
Isolated monoclonal antibody
Fig.10: Schematic representation of hybridoma technology used for monoclonal antibody production
Functions of Antibody
Antibodies recognize the antigen and participate in a broad range of other biological activities, which will result in removal of the antigen and death of the pathogen. The variable regions of the antibody bind to the antigen and the heavy chain constant region involves in a variety of collaborative interactions with other proteins, cells and tissues that result in the effector functions of the humoral response. These effector functions result from interactions between heavy chain constant regions and other serum proteins or cell membrane receptors, so not all the classes of immunoglobulin have the same functional properties. An overview of major effector functions mediated by antibodies is discussed here.
17

Opsonization
Opsonization is a process which promotes the phagocytosis of antigens by macrophages and neutrophiles. It is an important factor in antibacterial defenses. Fc receptors (FcR) are the protein molecules that present on the surfaces of macrophages and neutrophiles. The interaction between the Fc portions of the antibodies (complexed with the antigen or pathogen) and the FcR of phagocytic cells results in the binding of the pathogen to the phagocyte membrane. This cross linking of the FcR by binding to an array of antibody Fc regions initiates a signal-transduction pathway that results in the phagocytosis of the antigen-antibody complex. Complement Activation
Complement includes a collection of serum glycoproteins that can perforate cell membrane of the pathogens. IgM and most IgG subclasses can activate complement system. An important byproduct of the complement activation pathway is a protein fragment called C3b, which binds nonspecifically to cell and antigen-antibody complexes. Many cell types like macrophages and RBCs have receptors for C3b and binds to the C3b adhered complexes. The binding of adherent C3b by macrophages leads to phagocytosis of the molecular complex attached to C3b. The collaboration between antibody and the complement system is important for the inactivation and removal of antigens and killing of pathogens. Antibody Dependent Cell-Mediated Cytotoxicity (ADCC)
Coating of the target cells (virus infected cells of the host) by antibody and their subsequent linking with the Fc receptors of the number of cell types, particularly natural killer (NK) cells leads to their preferential killing. These cells secrete cytokines like interferon-γ (IFN- γ), tissue necrosis factor (TNF) and other granular enzymes to kill their target cells. Mucosal Immunity
IgA is the only class of antibody significantly present in body secretion and is transported across the mucosal lining by poly Ig mediated mechanism. This antibody protects the mucosal surfaces of the gastrointestinal tract, respiratory tract, genitourinary tract etc. by cross linking large antigens and preventing their attachment to mucosal cells. Neonatal Immunity
Significant amount of most subclasses of IgG transfer from mother to fetus. In human this transfer takes place during third trimester of gestation. Due to placental transfer, IgG gives protection to newborn immediately after birth. The transfer of IgG from mother to fetus is a form of passive immunization. The ability to transfer immunity from one individual to another by the transfer of antibodies is the basis of passive antibody therapy. Suggested Reading 1. Richard A Goldsby, Thomas J. Kindt, Barbara A. Osborne, Janis Kuby. Antibodies: structure and function.
Immunology.2003, 5th edition.W.H.Freeman and company, New York. 2. Kohlar.G and C.Milstein.1975.Continuous culture of fused cells secreting antibody of predefined
specificity.1975, Nature 256: 495. 3. Gerald Maurice Edelman. Antibody structure and molecular immunology. Scandinavian Journal of
Immunology 1991. Vol. 34 (1), 4-22.
18

4. Abul K. Abbas, Andrew H. Lichtman, Jordan S Pober. Cellular and Molecular Immunology.4th Edition. W.B. Saunders Company.
5. S. J. Singer and Russell F. Doolittle. Antibody Active Sites and Immunoglobulin Molecules. 1966 Science,Vol. 153(3731)13 – 25
6. Janeway CA, Jr et al Immunobiology. 2001,5th ed. Garland Publishing
19


















