
Immunogold localization of callose and other cell wall components in pea nodule transfer cells
9
Protoplasma (2000) 214:210-218 PROTOPLASMA Springer-Verlag 2000 Printed in Austria Immunogold localization of callose and other cell wall components in pea nodule transfer cells Preeti Dahiya and Nicholas J. Brewin* Department of Genetics,John Inues Centre, Norwich Received February 17, 2000 Accepted June 27, 2000 Summary. Transfer cells are located adjacent to xylem and phloem elements in pea nodule vascular tissues. The composition of the labyrinthine wall intrusions was investigatedby immunogold label- ing using specificantibody probes. Callose antigen was found at the base of newly formed cell wall intrusions and also in adjacent plas- modesmata. Sections through developed labyrinthine intrusions revealed that wall ingrowths had an internal structure with small domains of callose suggesting the presence of channels or vents. Xyloglucan and pectin antigens were uniformly distributed within the wall, but the distribution of extensin antigens was variable,with different antigens being detected in different regions of the wall ingrowth. A lectinlike glycoprotein, PsNLEC-1, was localized in intercellular spaces associated with nodule transfer cells.Previously, expression of this component was observed in other types of cells showing complex involution of the plasma membrane, namely root corticalcellsharboring arbuscularmycorrhizaeand nodule cellshar- boring nitrogen-fixing rhizobia. Keywords: Nodule lectin; Rhizobium; Legume; Symbiosis; Peri- cycle;Peribacteroid membrane; Symbiosome. Introduction Transfer cells are morphologically specialized cells with elaborate labyrinthine wall invaginations which amplify the symplast-apoplast interface (Pate and Gunning 1972, Ligrone et al. 1993). Many specialized plant organs involved in secretion or absorption have transfer cells as an integral part of their morphology. Examples include the gametophyte-sporophyte junc- tion (Ligrone et al. 1993), cotyledons (Offler et al. 1997), minor veins of the leaf (Pate and Gunning * Correspondenceand reprints: Department of Genetics,John Innes Centre, Norwich NR4 7UH, U.K. Email: [email protected] 1969), insectivorous plant glands (Heslop-Harrison and Knox 1971), and salt glands (Thomson et al. 1969). The situation is particularly interesting in nitrogen- fixing legume root nodules because these organs both receive and export substantial quantities of meta- bolites through their vascular connections. Photo- synthate is delivered as sucrose through the phloem and fixed nitrogen compounds, e.g., glutamine and asparagine, are exported by secretion into the xylem stream (Parsons and Baker 1996). In some (but not all) legumes, e.g., Pisum spp. and Phaseolus spp., the peri- cycle cells of the nodule vascular bundle are modified as transfer cells (Pate et al. 1968, Newcomb and Peterson 1979, Newcomb et al. 1979), and the xylem parenchyma of the root tissue adjacent to the nodule also develop wall ingrowths. Ineffective nodules of Pisurn spp. and uninvaded, spontaneous nodules in Medicago sativa have also been reported to possess transfer cells associated with their vascular tissues (Newcomb and Peterson 1979, Joshi et al. 1993). Newcomb and Peterson (1979), reported that, in an effective pea nodule, cell wall ingrowths begin to form in the pericycle cells 5 days after inoculation with Rhi- zobium spp. and in the xylem parenchyma cells of the adjacent roots after 7-8 days. In other parts of the root, not associated with nodules, wall ingrowths are very infrequent in root xylem parenchyma. At the ultrastructural level, transfer cells are char- acterized by labyrinthine invaginations consisting of branched and filiform protuberances of the cell wall. Typically, the plasma membrane follows the contours of the cell wall ingrowth. In most cases, cell wall invagi- nations are highly polarized and develop only on areas
-
Upload
preeti-dahiya -
Category
Documents
-
view
213 -
download
1
Transcript of Immunogold localization of callose and other cell wall components in pea nodule transfer cells

Protoplasma (2000) 214:210-218 PROTOPLASMA �9 Springer-Verlag 2000 Printed in Austria
Immunogold localization of callose and other cell wall components in pea nodule transfer cells
Preeti Dahiya and Nicholas J. Brewin*
Department of Genetics, John Inues Centre, Norwich
Received February 17, 2000 Accepted June 27, 2000
Summary. Transfer cells are located adjacent to xylem and phloem elements in pea nodule vascular tissues. The composition of the labyrinthine wall intrusions was investigated by immunogold label- ing using specific antibody probes. Callose antigen was found at the base of newly formed cell wall intrusions and also in adjacent plas- modesmata. Sections through developed labyrinthine intrusions revealed that wall ingrowths had an internal structure with small domains of callose suggesting the presence of channels or vents. Xyloglucan and pectin antigens were uniformly distributed within the wall, but the distribution of extensin antigens was variable, with different antigens being detected in different regions of the wall ingrowth. A lectinlike glycoprotein, PsNLEC-1, was localized in intercellular spaces associated with nodule transfer cells. Previously, expression of this component was observed in other types of cells showing complex involution of the plasma membrane, namely root cortical cells harboring arbuscular mycorrhizae and nodule cells har- boring nitrogen-fixing rhizobia.
Keywords: Nodule lectin; Rhizobium; Legume; Symbiosis; Peri- cycle; Peribacteroid membrane; Symbiosome.
Introduction
Transfer cells are morphologically specialized cells with elaborate labyrinthine wall invaginations which amplify the symplast-apoplast interface (Pate and Gunning 1972, Ligrone et al. 1993). Many specialized plant organs involved in secretion or absorption have transfer cells as an integral part of their morphology. Examples include the gametophyte-sporophyte junc- tion (Ligrone et al. 1993), cotyledons (Offler et al. 1997), minor veins of the leaf (Pate and Gunning
* Correspondence and reprints: Department of Genetics, John Innes Centre, Norwich NR4 7UH, U.K. Email: [email protected]
1969), insectivorous plant glands (Heslop-Harrison and Knox 1971), and salt glands (Thomson et al. 1969).
The situation is particularly interesting in nitrogen- fixing legume root nodules because these organs both receive and export substantial quantities of meta- bolites through their vascular connections. Photo- synthate is delivered as sucrose through the phloem and fixed nitrogen compounds, e.g., glutamine and asparagine, are exported by secretion into the xylem stream (Parsons and Baker 1996). In some (but not all) legumes, e.g., Pisum spp. and Phaseolus spp., the peri- cycle cells of the nodule vascular bundle are modified as transfer cells (Pate et al. 1968, Newcomb and Peterson 1979, Newcomb et al. 1979), and the xylem parenchyma of the root tissue adjacent to the nodule also develop wall ingrowths. Ineffective nodules of Pisurn spp. and uninvaded, spontaneous nodules in Medicago sativa have also been reported to possess transfer cells associated with their vascular tissues (Newcomb and Peterson 1979, Joshi et al. 1993). Newcomb and Peterson (1979), reported that, in an effective pea nodule, cell wall ingrowths begin to form in the pericycle cells 5 days after inoculation with Rhi- zobium spp. and in the xylem parenchyma cells of the adjacent roots after 7-8 days. In other parts of the root, not associated with nodules, wall ingrowths are very infrequent in root xylem parenchyma.
At the ultrastructural level, transfer cells are char- acterized by labyrinthine invaginations consisting of branched and filiform protuberances of the cell wall. Typically, the plasma membrane follows the contours of the cell wall ingrowth. In most cases, cell wall invagi- nations are highly polarized and develop only on areas

P. Dahiya and N. J. Brewin: Pea nodule transfer cells 211
involved in intensive plasma membrane transport activity, e.g., bordering xylem elements, or adjacent to sieve tubes, companion cells, and adjacent intercellu- lar spaces (Pate and Gunning 1972). Transfer cells possess dense cytoplasm, prominent nuclei, abundant rough endoplasmic reticulum (RER) and ribosomes (Pate et al. 1968). Almost all wall ingrowths are closely associated with individual mitochondria (Gunning 1977), suggesting the involvement of high ATPase activity during transport of solutes across the mem- brane (Maier and Maier 1972, Joshi et al. 1993). In addition RER-derived vesicles were reported to be in close association with the plasma membrane of the labyrinthine invaginations, which might facilitate more efficient solute transport across the membrane system (Joshi et al. 1993).
To investigate the development of labyrinthine walls in transfer cells, we examined their composition by immunogold labeling using a range of specific anti- body probes. This study was of particular interest in the context of pea and other legumes because these plants establish root symbioses with rhizobia and with arbuscular mycorrhizae (Glomales). In each case, host cell development progresses from an invasive phase (involving intrusive growth of the plant cell wall) to a phase of intense metabolic exchange (in which host cells develop an internal membrane system that enor- mously enlarges the surface area of the plant-microbe interface). These host cells contain specialized symbi- otic structures (Brewin 1998), which are somewhat similar to the labyrinthine intrusions of transfer cells (Pate et al, 1968). Therefore, the possible similarity between the morphological adaptations of host cells in symbiosis and of transfer cells was investigated by choosing a range of antibody probes which had been previously used to study host cells harboring endosymbionts (Dahiya et al. 1997, Rae et al. 1992, VandenBosch et al. 1989).
Material and methods
Biological material
Nodules of Pisum sativum variety Sprint2 were obtained as previ- ously described (Borisov et al. 1992, Sherrier et al. 1997). For pea nodulation, germinating seedlings were inoculated with Rhizobium leguminosarum bv. viciae wild-type strain 3841 (Wood et al. 1989). Plants were grown in gnotobiotic conditions (i.e., controlled micro- biological conditions) in conical flasks on agar medium containing mineraI salts but lacking a nitrogen source. Pea seeds were obtained from the pea germplasm collection at the John Innes Centre (cour- tesy of Mike Ambrose).
Antibodies
Monoclonal antibodies and antisera used for immunogold labeling are listed in Table 1. Jan Peart and Dr. Paul Knox (University of Leeds) generously supplied monoclonal antibodies. The monoclonal antibody to (1-3)-[3-glucan was obtained from Biosupplies Australia Pty Ltd (Melbourne, Australia). Antiserum R76, reacting with the lectinlike glycoprotein PsNLEC-1, was made monospecific by immunoadsorption as previously described (Dahiya et al. 1997). For immunogold labelling, the secondary antibody was anti-rat antibody, conjugated to 10 nm diameter colloidal gold, or anti-mouse 10 nm, or anti-rabbit 20 nm diameter gold (as appropriate).
Microscopy
Pea nodules (three to four weeks old) were fixed in 1% (w/v) glu- taraldehyde, 4% formaldehyde in 100 mM sodium phosphate buffer, pH 7.2. Nodules were given cuts on both sides to allow better pen- etration of the fixative. After vacuum infiltration, floating nodules were removed and the fixative was replaced with fresh solution. After overnight incubation at 4 ~ nodules were dehydrated and embedded in LR White resin as described previously (VandenBosch et al. 1989). For transmission electron microscopy, gold sections, 90-150 nm thick, were collected on gold grids coated with 4% (w/v) parlodion and carbon. After blocking with 3% (w/v) bovine serum albumin (BSA) (Sigma, Poole, U.K.), 0.1% (v/v) Tween 20 (Sigma) in Tris-buffered saline (TBS) for 1 h, the sections were incubated overnight at 4 ~ with primary antibody (listed in Table 1). All of the monoclonal antibodies were diluted 1/5 and monospecific antiserum R76 was diluted 1/100 in 1% (w/v) BSA in TBS. After washing, by passing through 8-10 drops of TBS, sections were incubated with secondary anti-rat, -mouse, or -rabbit antibody con- jugated to 10-20 nm diameter colloidal gold (Amersham Interna- tional, Little Chalfont, U.K.), diluted 1/100 in 1% (w/v) BSA in TBS, for 1 h. The grids containing sections were then washed in water for 15-20 min and were counterstained in 2% (w/v) aqueous uranyl acetate for 15 min, followed by lead citrate for 4 s. All solutions used were filtered before use, and filter-sterilized deionised water was used throughout the experiment. Nodule tissues were viewed and photographed in a JEOL JEM-1200 EM transmission electron microscope at 80 kV.
Table 1. Antibodies and antisera
Antibody/antiserum a Component recognized Reference
Anti-(l-3)-~3-glucan JIM7 JIM5
M1
JIM13 LM2
MAC265
LM1, LM3
R76
(l-3)-[3-glucan Meikle et al. 1991 esterified pectin Knox 1992 unesterified pectin VandenBosch
et al. 1989 fucosylated xyloglycan Puhlmann et al.
1994 arabinogalactan protein Knox et al. 1991 arabinogalactan protein Smallwood et al.
1995 nodule extensin VandenBosch
et al. 1989 extensin Smallwood et al.
1995 lectinlike glycoprotein Dahiya et al. 1997
a Anti-(1-3)-[~-glucan and M1 are mouse derived, R76 is rabbit derived, all others are rat derived

212 R Dahiya and N. J. Brewin: Pea nodule transfer ceils
For each monoclonal antibody, 3-4 nodule replicates were routinely used for immunolocalization (but, in the experiments with antiserum R76, 12 nodules were used in order to confirm the variability of the result). In the case of R76, the serum was made monospecific by preadsorption to nitrocellulose sheets carrying PsNLEC-1 antigen that had been heterologously expressed in E. coli (Dahiya et al. 1997). In the case of the range of rat monoclonal anti- bodies, each served as a negative control for the others (because they showed different patterns of labeling) and therefore the alter- native negative control (omitting any first antibody) was not in- cluded in these experiments.
For light microscopy, median longitudinal sections (50 gm thick) were collected on gelatin (1%, w/v) coated slides. The sections were counterstained with basic fuchsin and photographed on a Zeiss Axiophot microscope.
Results
Ultrastructure of pea nodule vascular elements
The general m o r p h o l o g y of the pea nodule is exhib-
ited in Fig. 1 A, which shows a per ipheral vascular
e lement and infected cells of the nodule. Nodu le vas-
cular e lements are in direct connect ion with those of
the roo t (Fig. 1B). A transverse sect ion of the nodule
vascular system shows t ransfer cells associated with
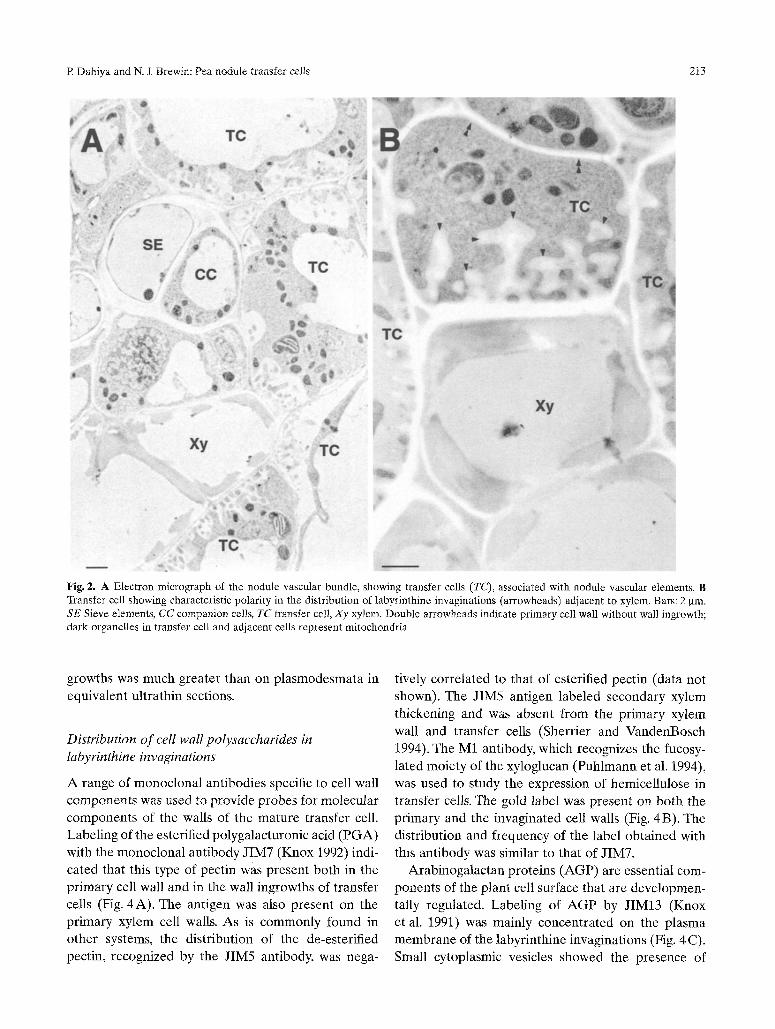
xylem and ph loem elements (Fig. 2A) . The wall
ingrowths of the transfer cells are usually polar ized
(Fig. 2B) and contain dense cytoplasm, enr iched in
mi tochondr ia and R E R .
Immunogold localization of callose antigen
Antical lose monoc lona l an t ibody (Meikle et al. 1991)
was used to p robe t ransfer cells in ul trathin sections
of the nodule vascular bundle (Fig. 3). In developing
transfer cells, there was significant a c c u m u l a t i o n of
colloidal gold label at the e lect ron-dense base of the
new wall ingrowths (Fig. 3A) . Associa ted with the
newly invaginated cell wall were channeMike struc-
tures, which also showed the presence of callose
antigen. I n mature t ransfer cells, there was no
detectable antigen at the base of the wall ingrowth and
the label was restr icted to small e lec t ron-dense
domains in the main b o d y of the labyrinthine invagi-
nat ions (Fig. 3 B). Channel- l ike structures with callose
antigen were also found within these walls, and thus it
is possible that the callose domains simply represent
channel- l ike structures in t ransverse section. These
structures connec ted the invaginated wall to the
pr imary wall, and at the distal end of the ingrowth they apparent ly open into the cytoplasm. Additionally,
some diffuse labeling of the pr imary cell wall and the
Fig. 1A, B, Micrographs illustrating the general morphology of the pea noduIe and connection of vascular elements between the root and the nodule. Nodule sections were stained with basic fuchsin. A Longitudinal section of a pea nodule showing infected cells (IC), nodule vascular bundles in transverse section (double arrowheads) and vascular connections (arrows) with root (Rt). Bar: 100 gin. B Enlargement from a different nodule, illustrating open vascular con- nection (arrows) between the nodule (Nd) and root (Rt). Transfer cells adjacent to xylem elements are also visible (arrowheads). Bar: 400 pm
cytoplasm was observed in young transfer cells, but
this was absent f rom the mature cells.
As p la smodesmata are the mos t c o m m o n sites for
callose antigen in pr imary cell walls, these were exam- ined to indicate the relative level of ant igen abundance
(Fig. 3 C). It was repea ted ly found that the amoun t of
immunogo ld label at the base of the young wall in-
R Dahiya and N. J. Brewin: Pea nodule transfer cells 2t3
Fig. 2. A Electron micrograph of the nodule vascular bundle, showing transfer cells (TC), associated with nodule vascular elements. B Transfer cell showing characteristic polarity in the distribution of labyrinthine invaginations (arrowheads) adjacent to xylem. Bars: 2 gin. SE Sieve elements, CC companion cells, TC transfer cell, Xy xylem. Double arrowheads indicate primary cell wall without wall ingrowth; dark organelles in transfer cell and adjacent cells represent mitochondria
growths was much greater than on plasmodesmata in equivalent ultrathin sections,
Distribution of cell wall polysaccharides in labyrinthine invaginations
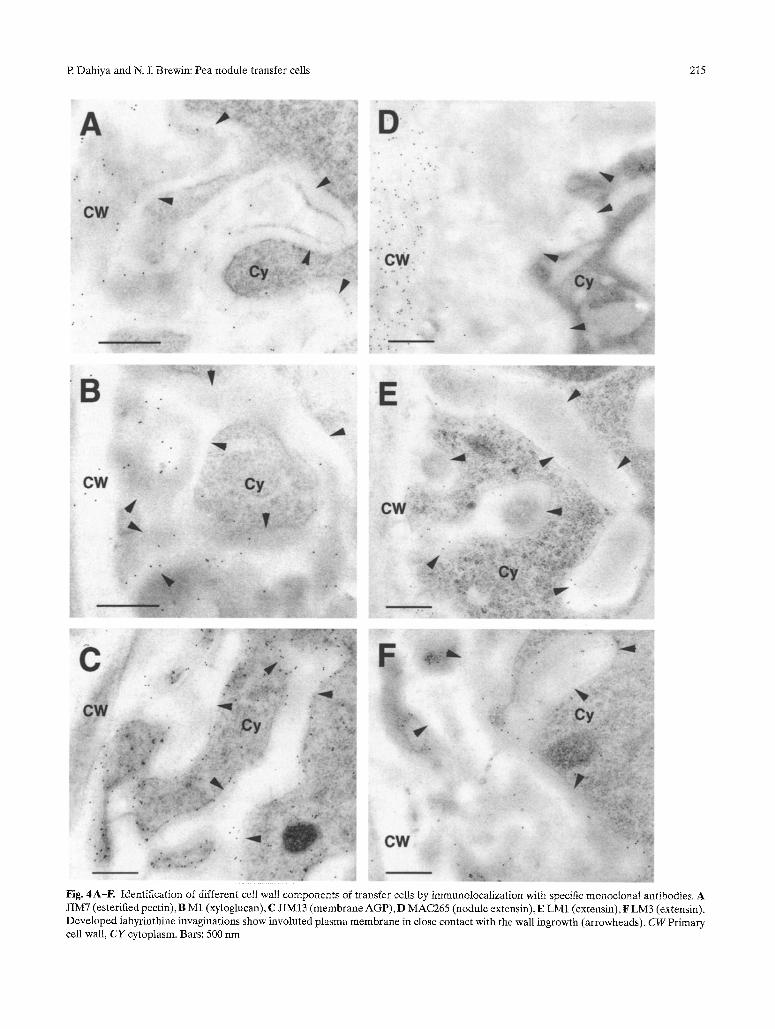
A range of monoclonal antibodies specific to cell wall components was used to provide probes for molecular components of the walls of the mature transfer cell. Labeling of the esterified polygalacturonic acid (PGA) with the monoclonal antibody JIM7 (Knox 1992) indi- cated that this type of pectin was present both in the primary cell wall and in the wall ingrowths of transfer cells (Fig, 4 A). The antigen was also present on the primary xylem cell walls. As is commonly found in other systems, the distribution of the de-esterified pectin, recognized by the JIM5 antibody, was nega-
tively correlated to that of esterified pectin (data not shown). The JIM5 antigen labeled secondary xylem thickening and was absent from the primary xylem wall and transfer cells (Sherrier and VandenBosch 1994). The M1 antibody, which recognizes the fucosy- lated moiety of the xyloglucan (Puhlmann et al. 1994), was used to study the expression of hemicellulose in transfer cells, The gold label was present on both the primary and the invaginated cell walls (Fig. 4B). The distribution and frequency of the label obtained with this antibody was similar to that of JIM7.
Arabinogalactan proteins (AGP) are essential com- ponents of the plant cell surface that are developmen- tally regulated, Labeling of AGP by JIM13 (Knox et al. 1991) was mainly concentrated on the plasma membrane of the labyrinthine invaginations (Fig. 4 C). Small cytoplasmic vesicles showed the presence of

214 R Dahiya and N. J. Brewin: Pea nodule transfer cells
Fig, 3A-C. Electron micrographs of transfer cells in a pea nodule section, showing immunogold localization of callose antigen. A Young transfer cell in nodule vascular bundle with developing, unbranched wall ingrowths (arrowheads) showing localized antigen (arrows) at the electron-dense base of the invaginated cell wall. Double arrowheads show channel-like structures associated with newly developed wall ingrowths. Cytoplasm (CY) and primary celt wall (CW) also show dispersed antigen. B Mature transfer cell showing fuliy developed labyrinthine invaginations (asterisks). Callose antigen is localized on small electron-dense aggregates in the main body of the labyrinth (arrows). Channel-like structures, similar to those of the young transfer cell, also showing callose antigen (double arrowheads). The label is absent from the base of the invaginated wall, primary ceil wail (CW) and cytoplasm (CY). 12 Plasmodesmata (double arrowheads) con- necting two cells in pea nodule, showing presence of callose antigen. Sy Symbiosome, M mitochondria. Bars: 500 nm
R Dahiya and N. J. Brewin: Pea nodule transfer cells 215
Fig. 4A-F. Identification of different cell wali components of transfer ceils by immunolocalization with specific monoclonal antibodies. A JIM7 (esterified pectin), B M1 (xyloglucan), C JIM13 (membrane AGP), D MAC265 (nodule extensin), E LM1 (extensin), F LM3 (extensin). Developed labyrinthine invaginations show involuted plasma membrane in close contact with the wall ingrowth (arrowheads). CW Primary cell wall, CY cytoplasm. Bars: 500 nm

216 R Dahiya and N. J. Brewin: Pea nodule transfer ceils
label but, as the cells became mature, the cytoplasmic label was progressively reduced, whereas the label on the plasma membrane remained unaffected. LM2, another AGP-recognizing antibody (Smallwood et al. 1996), also gave extensive cytoplasmic label, although the plasma membrane labeling was rare and dispersed (data not shown).
Distribution of extensin antibodies
The distribution of MAC265 antigen has been studied in detail in pea nodules (Rae et al. 1991, 1992). It is abundantly present in the matrix glycoprotein of intra- cellular infection threads (VandenBosch et al. 1989) and has recently been characterized as an extensin type glycoprotein in the pea nodule (Brewin et al. 2000). Interestingly, it was found in the present study, that the LM1 and LM3 (Smallwood et al. 1995) anti- bodies, which also belong to the extensin family, recognize the infection thread matrix as strongly as MAC265 (data not shown).
In the present study, we localized the distribution of these antigens in transfer cells. MAC265 was found to label the cell wall in addition to intercellular spaces, of both infected and uninfected cells. The antigen, however, did not extend to the labyrinthine invagina- tions of the transfer cells (Fig. 4D). Unlike MAC265, the LM1 and LM3 antibodies also labeled the labyrinthine invaginations of the transfer cells. The antigen was present on the plasma membrane and in the space between the plasma membrane and the cell wall of the branched labyrinthine invaginations (Fig. 4 E, F).
Immunolocalization of PsNLEC-1 antigen
PsNlec-1 encodes a lectinlike glycoprotein that is strongly expressed in infected cells of the pea nodule (Kardailsky et al. 1996, Dahiya et al. 1997). Because a PsNlec-1 transcript had also been detected in cells associated with the nodule vascular tissue (Kardailsky et al. 1996), the distribution of protein antigen was also examined in these tissues. Interestingly, the PsNLEC- 1 antigen was localized in the nodule vascular bundle and the label was mainly present in the intercellular spaces adjacent to transfer cells. Some dispersed label was also localized in the cytoplasm of the transfer cells (Fig. 5). Other cells of the vascular bundle did not show any antigen except nodule xylem elements, where intense labeling was sometimes present in the
Fig. 5. Immunodetection of a lectinlike protein, PsNLEC- 1, in the intercellular space of the transfer cells (arrows). Dispersed label is also present in the cell cytoplasm. Arrowheads indicate invaginated cell wall. CW Primary cell wall, TC transfer cell. Bar: 2 gm
cell lumen. Although the label was sometimes very strong, the presence of it varied from one nodule to another: we could only localize the PsNLEC-1 antigen in 5 out of 12 of the nodules studied, but the basis for this variation is not understood.
Discussion
Labyrinthine invaginations of transfer cells are mor- phologically adapted intrusions of the cell wall. We used a range of antibodies in order to investigate the general composition of invaginated walls of the trans- fer cells associated with the pea nodule vascular bundle. Overall, the labeling given by all the anti- bodies on the xylem- and phloem-associated transfer cells showed no difference in composition, although, phloem-associated transfer cells deliver metabolites (secrete) from the plant to the nodule and those asso- ciated with xylem transfer fixed nitrogen from the nodule to the rest of the plant.
Antibodies that recognize esterified pectin and xyloglucan antigen were found to label the wall ingrowth uniformly, and the antibody recognizing the membrane arabinogalactan protein uniformly labeled plasma membrane surrounding the contours of the wall ingrowths. The intensity of the label given by all the above antibodies on labyrinthine invaginations was similar to that on the primary cell wall. These results suggest that in general, the major components of the invaginated cell wall are similar to those of primary cell wall.
Using a monoclonal antibody that recognizes callose, we localized channel-like structures or possi-

R Dahiya and N. J. Brewin: Pea nodule transfer cells 217
bly plasmodesmata next to the developing and dev- eloped invaginations (Fig. 3A), demonstrating that wall ingrowths develop in close proximity to plasmo- desmata. As hypothesized previously by Pate and Gunning (1972), the existence of open channels within the labyrinthine walls could serve to reduce the diffu- sion resistance in the apoplastic space. Interestingly, the intensity of the electron-dense domains and of channel-like structures in the developed labyrinth of transfer cells (Fig. 3A, B) resembled that of plas- modesmata shown in Fig. 3 C. This analogy and the fact that callose labels the collars of plasmodesmata (Radford et al. 1998) emphasizes the point that punc- tate antigen domains of the wall ingrowth probably represent callose-labeled openings similar to those of multiply branched plasmodesmata. These structures can be compared to the secondary plasmodesmata described by Lucas et al. (1993). Following this line of argument, it is possible that the accumulated antigen at the base of the young invaginations might there- fore represent developing secondary branching of the parent plasmodesma.
Labyrinthine walls and rhizobium-induced infection threads represent two different kinds of cell wall invagination. We used antiextensin antibodies, which label the infection thread matrix, as probes to compare the structure and development of labyrinthine walls with that of infection threads. The distribution of anti- bodies on wall ingrowths of transfer cells was found to be not as localized as in the case of infection threads. Although the LM1 and LM3 antibodies recog- nized the plasma membrane and the matrix between the cell wall and plasma membrane of the labyrinthine invaginations (Fig. 4E, F), the MAC265 antibody labeling was restricted to the primary cell wall at the base of the labyrinthine ingrowth (Fig. 4D). These results suggest that extensins are involved in both infection thread and labyrinthine wall development, but the composition of the in-growing cell wall is somewhat different in the two cell types.
Host cells harboring either rhizobial bacteroids or arbuscular mycorrhizae have previously been demon- strated to express a lectinlike glycoprotein, PsNLEC- 1 (Kardailsky et al. 1996, Balestrini et al. 1999). In the case of the nodule-infected cells, the protein was demonstrated to be mainly localized in the symbioso- mal space, a compartment which harbors the nitrogen- fixing bacteroid, and in the vacuole of the host cell. Interestingly, the PsNlec-1 gene transcript was also found to be expressed in the vascular bundle of the
nodule (Kardailsky et al. 1996). In the present study, this observation was confirmed by immunogold local- ization; the antigen was found to be present mainly in the intercellular" space adjacent to the transfer cells (Fig. 5) and some of the antigen was occasionally found in the lumen of xylem vessels. PsNLEC-1 is known to have an N-terminal targeting sequence (Kar- dailsky et al. 1996), but it is inlLeresting that from trans- fer cells the protein accumulates in the extracellular space, whereas in host cells containing bacteroids the protein is found in symbiosomes and vacuoles. The function of PsNLEC-1 is still unknown, but its expres- sion in symbiosomes, arbuscules, and transfer cells indicates an association with intensive membrane bio- genesis and secretory activity.
The localization of PsNLEC-1 in the lumen of xylem vessel was unexpected because tti~re are no previous reports of the transportation of a protein through the xylem, although, many proteins have been localized in the phloem sap (Thompson and Schulz 1999). In the case of nodulated legumes, xylem transport may be usual because the xylem serves not just as a conduit for soil-derived water and mineral salts but as the delivery route for the products of fixed nitrogen from root nodules. Thus, the localization of the PsNLEC-1 antigen seems to follow the traditional pathway pro- posed for the export of amides or ureides, and it is pos- sible to imagine that transfer cells are involved in the synthesis of this protein and its secretion into the tran- spiration stream of the root vasculature.
In conclusion, this study has thrown new light on the composition and development of the labyrinthine walls of transfer cells. The presence within labyrinthine invaginations of structures resembling secondary plas- modesmata might prove to be an important finding to establish functioning mechanisms for these elaborate structures, which play an important role at secretory interfaces. Furthermore, the localization of the PsNLEC-1 antigen in intercellular spaces adjacent to transfer cells is an intriguing observation, which might lead to further investigations of secretory processes, export of metabolites, and their pathways.
Acknowledgments
We thank Dr. M. McCann and N. L Stacey for helpful discussions and for their comments on the manuscript. This work was supported by GB-BBSRC.

218 R Dahiya and N. J. Brewin: Pea nodule transfer cells
R e f e r e n c e s
Balestrini R, Perotto S, Gasverde E, Dahiya P, Guldmann L, Brewin NJ, Bonfante P (1999) Transcription of a gene encoding a Iectin- like glycoprotein is induced in root cells harboring arbuscular mycorrhizal fungi in Pisum sativum. Mol Plant Microbe Interact 12:785-791
Borisov AY, Morzina EV, Kulikova OA, Tchetkova SA, Lebsky VK, Tikhonovich IA (1992) New symbiotic mutants of pea (Pisurn sativum L.) affecting either nodule initiation or symbiosome development. Symbiosis 14:297-313
Brewin NJ (1998) Tissue and cell invasion by Rhizobium: the struc- ture and development of infection threads and symbiosomes. In: Spaink HI', Kondorosi A, Hooykaas PJJ (eds) The Rhizobiaceae. Kluwer, Dordrecht, pp 417-429
- Rathbun EA, Wisniewski JP (2000) Structure and development of infection threads. In: Pedrosa FE, Hungria M, Yates MG, Newton WE (eds) Nitrogen fixation: from molecules to crop pro- ductivity. Kluwer, Dordrecht, pp 381-382
Dahiya R Kardailsky IV, Brewin NJ (1997) Immunolocalization of PsNLEC-1, a lectin-like glycoprotein expressed in developing pea nodules. Plant Physiol 115:1431-1442
Fedorova E, Thomson R, Whitehead LF, Maudoux O, Udvardi MK, Day DA (1999) Localization of H+-ATPase in soybean root nodules. Planta 209:25-32
Gunning BES (1977) Transfer cells and their role in transport of solutes in plants. Sci Prog Oxford 64:539-568
Heslop-Harrison Y, Knox RB (1971) A cytochemical study of the leaf-gland enzymes of insectivorous plants of the genus Pingui- cula. Planta 96:183-211
Joshi PA, Caetano-Anolles G, Graham ET, Gresshoff PM (1993) Ultrastructure of transfer cells in spontaneous nodules of alfalfa (Medicago sativa). Protoplasma 172:6476
Kardailsky IV, Sherrier D J, Brewin NJ (1996) Identification of a new pea gene, PsNlecl, encoding a lectin-like glycoprotein isolated from the symbiosomes of root nodules. Plant Physiol 111:49-60
Knox JP (1992) Molecular probes for the plant cell surface. Proto- plasma 167:1-9
- Linstead PJ, Peart J, Cooper C, Roberts K (1991) Developmen- tally regulated epitopes of cell-surface arabinogalactan proteins and their relation to root-tissue pattern-formation. Plant J 1: 317-326
Ligrone R, Duckett JG, Renzaglia KS (1993) The gametophyte- sporophyte junction in land plants. Adv Bot Res 19:231-317
Lucas WJ, Ding B, Van Der Schoot C (1993) Plasmodesmata and the supracellular nature of plants. Plant Phytol 125:435-476
Maier K, Maier U (1972) Localization of 13-glycerophosphatase and Mg++-activated adenosine triphosphatase in a moss haustorium and the relation of these enzymes to the cell wall labyrinth. Pro- toplasma 75:91-112
Meikle J, Bonig I, Hoogenraad N J, Clarke AE, Stone BA (1991) The location of (l-3)q3-glucans in the walls of pollen tubes of Nico- tiana alata using (l-3)-[3-glucan-specific monoclonal antibody. Planta 185:1-8
Newcomb W, Peterson RL (1979) The occurrence and ontogeny of transfer cells associated with lateral roots and root nodules in Leguminosae. Can J Bot 57:2583-2602
- Sippell D, Peterson RL (1979) The early morphogenesis of Glycine max and Pisum sativum root nodules. Can J Bot 57: 2603-2616
Offer CE, Liet E, Sutton EG (1997) Transfer cell induction in cotyledons of Vicia faba L. Protoplasma 200:51-64
Parsons R, Baker A (1996) Cycling of amino compounds in symbi- otic lupin. J Exp Bot 47:421-429
Pate JS, Gunning BES (1969) Vascular transfer cells in angiosperm leaves: a taxonomic and morphological survey. Protoplasma 68: 135-156
- - (1972) Transfer cells. Annu Rev Plant Physiol 23:173-196 - - Briarty LG (1968) Ultrastructure and functioning of the
transport system of the leguminous root nodules. Planta 85: 11- 34
Puhlmann J, Bucheli E, Swain M J, Dunning N, Albersheim R Darvii1 AG, Hahn MG (1994) Generation of monoclonal antibodies against plant cell-wall polysaccharides 1: characterization of a monoclonal antibody to a terminal alpha-(l-2)-linked fucosyl- containing epitope. Plant Physiol 104:699-710
Radford JE, Vesk M, Overall RL (1998) Callose deposition at plas- modesmata. Protoplasma 201:30-37
Rac AL, Perotto S, Knox JR Kannenberg EL, Brewin NJ (1991) Expression of extracellular glycoproteins in the uninfected cells of developing pea nodule tissue. Mol Plant Microbe Interact 4: 563-570
- Bonfante-Fasolo R Brewin NJ (1992) Structure and growth of infection threads in the legume symbiosis with Rhizobium legu- minosarum. Plant J 2:385-395
Sherrier DJ, VandenBosch KA (1994) Localization of repetitive proline-rich proteins in the extracellular matrix of pea root nodules. Protoplasma 183:148-161
- Borisov AY, Tikhonovich IA, Brewin NJ (1997) Immunocytolo- gical evidence for abnormal symbiosome development in nodules of the pea mutant line Sprint2Fix- (sym31). Protoplasma 199: 57-68
Smallwood M, Martin H, Knox JP (1995) An epitope of rice threonine- and hydroxyproline-rich glycoprotein is cmnmon to cell wall and hydrophobic plasma-membrane glycoproteins. Planta 196:510-522
- Yates EA, Willats WGT, Martin H, Knox JP (1996) Immuno- chemical comparison of membrane-associated and secreted arabinogalactan-proteins in rice and carrot. Planta 198: 452- 459
Thomson WW, Berry WL, Liu LL (1969) Location and secretion of salt by the salt glands of Tamarix aphylla. Proc Natl Acad Sci USA 63:310-317
Thompson GA, Schulz A (1999) Macromolecular trafficking in the phloem. Trends Plant Sci 4:354-360
VandenBosch KA, Bradley D J, Knox JR Perotto S, Butcher GW, Brewin NJ (1989) Common components of the infection thread matrix and the intercellular space identified by immunocyto- chemical analysis of pea nodules and uninfected roots. EMBO J 8:335-342
Wood EA, Butcher GW, Brewin NJ, Kannenberg EL (1989) Genetic derepression of a developmentally regulated lipopolysaccharide antigen from Rhizobium leguminosarum 3841. J Bacteriol 171: 4549--4555


















