
Identification, Purification, and Partial Sequence ... · Identification, Purification, and Partial...
6
THE JOURNAL OF BIOLOCICAL CHEMISTRY Vol. 267, No. 4, Issue of February 5, pp. 2524-2529,1992 Printed in U.S.A. Identification, Purification, and Partial Sequence Analysis of Autotaxin, a Novel Motility-stimulating Protein* (Received for publication, August 28, 1991) Mary L.StrackeS, Henry C. Krutzsch, Edward J. Unsworth, Anders Arestad, Vittoria Cioce, Elliott Schiffmann, and Lance A. Liotta From the Laboratory of Pathology, National Cancer Institute, National Institutes of Health, Bethesda, Maryland 20892 Autotaxin (ATX) is a potent human motility-stimu- lating protein that has been identified in the condi- tioned medium from A2058 melanoma cells. This pro- tein has been purified to homogeneity utilizing a strat- egy involving five column steps. Homogeneity of ATX was verified by two-dimensional gel electrophoresis. The molecular size of ATX is 125 kDa, and it has an isoelectric point of 7.7 f 0.2. Purified ATX was di- gested with cyanogen bromide and trypsin, and the resulting ATX peptides were purified by reverse-phase high performance liquid chromatography. Eleven pep- tides were subjected to amino acid sequence analysis, and 114 residues were identified. The partial amino acid sequences and the amino acid composition ob- tained for ATX show that it does not exhibit any sig- nificant homology to known growth factors or previ- ously described motility factors. At picomolar concen- trations, ATX stimulates both random and directed migration of human A2058 melanoma cells. Pretreat- ment of the melanoma cells with pertussis toxin abol- ishes the response to purified ATX, indicating that ATX stimulates motility through a receptor acting via a pertussis toxin-sensitive G protein. Cell motility plays an important role in embryonic events, adult tissue remodeling, wound healing, angiogenesis, immune defense, and metastasis of tumor cells (1). In normal physio- logic processes, motility is tightly regulated, while tumor cell motility may be aberrantly regulated or autoregulated. Tumor cells can respond in a motile fashion to a variety of agents, including host-derived scatter factors (2, 3), growth factors (4-8), extracellular matrix components (9-11), and tumor- secreted or autocrine factors (12-17). Many types of tumor cells havebeenfound to produce “autocrine motility factors” which stimulate motility of the same tumor cells that make the factor (18). Our prior studies of human A2058 melanoma cells have demonstrated that these cells are a particularly rich source of autocrine motility factors. Wehavepreviouslyisolated an autocrine motility factor from the conditioned media of these cells which has a molecular mass of approximately 60 kDa (12). Similar tumor cell-derived or induced factors with the same molecular mass have subsequently been reported and purified by several other investigators (14-17, 19). Autocrine motility factors are not * The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “aduertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. $ T o whom correspondenceshould be addressed Laboratory of Pathology, National Cancer Institute, National Institutes of Health, Bldg. 10, Rm. 2A33, Bethesda, MD 20892. Tel.: 301-496-1843; Fax: 301-402-0043. specific for a given type of cancer cell and have a wide spectrum of activity on many types of cancer cells (20) but little effect onnormalfibroblasts or leukocytes. Theyact through cell-surface receptors (17, 21, 22) to stimulate both both random and directed migration (12, 14, 15). In the present report we describe the identification, purifi- cation to homogeneity, characterization, and partial amino acid sequence analysis of a novel 125-kDa motility-stimulat- ing protein. EXPERIMENTAL PROCEDURES’ Methods Cell Culture-The human melanoma cell line A2058, originally isolated by Todaro etal. (23), was maintained as previously described (12). Cell Motility Assays-The assay to determine motility was per- formed in triplicate using a 48-well microchemotaxis chamber as described previously in detail (24). The material to be tested was dialyzed into DPBS’ with calcium and magnesium supplemented with 0.1% (w/v) bovine serum albumin using Centricon 30 filtration units (Amicon). Chemotaxis was quantitated either by reading the stained membranes with a 2202 Ultroscan laser densitometer (Pharmacia- LKB Biotechnology Inc.) or by counting five randomly chosen high power fields (HPF) under light microscopy (400X) for each replicate. Densitometer units (wavelength = 633 nm) have been shown to be linearly related to the number of cells per H P F (10, 24). Two-dimensional Gel Electrophoresis-For two-dimensional elec- trophoresis, the protein, in 20% ethylene glycol, was dried in a Speedvac concentrator and redissolved in loading solution: 9 M urea, 1% (v/v) pH 3-10 Bio-Lyte, and 2.5% (v/v) Nonidet P-40. This sample was then subjected to isoelectric focusing (25) using a Bio- Rad tube cell in 120 X 3-mm polyacrylamide tube gels containing 9 M urea, 2% (v/v) pH 3-10 Bio-Lyte, 0.25% (v/v) pH 8-10 Bio-Lyte, and 2.5% (v/v) Nonidet P-40 (Bio-Rad). Reservoir solutions were 0.01 M phosphoric acid and 0.02 M NaOH. Nonequilibrium isoelectric focusing (26) was run initially with constant voltage (500 V) for 5 h. Since the protein was basic, we repeated the procedure under equilib- rium conditions (500 V for 17 h). Electrophoresis in the second dimension was performed on a 7.5% polyacrylamide gel using the conditions of Laemmli (27).The gel was stained with Coomassie Blue G-250 as described previously (28). Amino Acid Analysis-Acid hydrolysis of purified ATX and amino acid analysis (29) were performed at Yale University School of Medicine Protein and Nucleic Acid Facility, New Haven, CT. Preparation of Peptides for Internal Sequence of Autotaxin-Ho- mogeneous ATX was sequentially digested with cyanogen bromide Portions of this paper (including part of “Experimental Proce- dures,” Figs. 1-6, and Table I) are presented in miniprint at the end of this paper. Miniprint is easily read with the aid of a standard magnifying glass. Full size photocopies are included in the microfilm edition of the Journal that is available from Waverly Press. The abbreviations used are: DPBS, Dulbecco’s phosphate-buff- ered saline; ATX, autotaxin; HPLC, high pressure liquid chromato- graph; PT, pertussis toxin; SDS, sodium dodecyl sulfate; HPF, high power field. 2524
Transcript of Identification, Purification, and Partial Sequence ... · Identification, Purification, and Partial...

THE JOURNAL OF BIOLOCICAL CHEMISTRY Vol. 267, No. 4, Issue of February 5, pp. 2524-2529,1992 Printed in U.S.A.
Identification, Purification, and Partial Sequence Analysis of Autotaxin, a Novel Motility-stimulating Protein*
(Received for publication, August 28, 1991)
Mary L. StrackeS, Henry C. Krutzsch, Edward J. Unsworth, Anders Arestad, Vittoria Cioce, Elliott Schiffmann, and Lance A. Liotta From the Laboratory of Pathology, National Cancer Institute, National Institutes of Health, Bethesda, Maryland 20892
Autotaxin (ATX) is a potent human motility-stimu- lating protein that has been identified in the condi- tioned medium from A2058 melanoma cells. This pro- tein has been purified to homogeneity utilizing a strat- egy involving five column steps. Homogeneity of ATX was verified by two-dimensional gel electrophoresis. The molecular size of ATX is 125 kDa, and it has an isoelectric point of 7.7 f 0.2. Purified ATX was di- gested with cyanogen bromide and trypsin, and the resulting ATX peptides were purified by reverse-phase high performance liquid chromatography. Eleven pep- tides were subjected to amino acid sequence analysis, and 114 residues were identified. The partial amino acid sequences and the amino acid composition ob- tained for ATX show that it does not exhibit any sig- nificant homology to known growth factors or previ- ously described motility factors. At picomolar concen- trations, ATX stimulates both random and directed migration of human A2058 melanoma cells. Pretreat- ment of the melanoma cells with pertussis toxin abol- ishes the response to purified ATX, indicating that ATX stimulates motility through a receptor acting via a pertussis toxin-sensitive G protein.
Cell motility plays an important role in embryonic events, adult tissue remodeling, wound healing, angiogenesis, immune defense, and metastasis of tumor cells (1). In normal physio- logic processes, motility is tightly regulated, while tumor cell motility may be aberrantly regulated or autoregulated. Tumor cells can respond in a motile fashion to a variety of agents, including host-derived scatter factors (2, 3), growth factors (4-8), extracellular matrix components (9-11), and tumor- secreted or autocrine factors (12-17).
Many types of tumor cells have been found to produce “autocrine motility factors” which stimulate motility of the same tumor cells that make the factor (18). Our prior studies of human A2058 melanoma cells have demonstrated that these cells are a particularly rich source of autocrine motility factors. We have previously isolated an autocrine motility factor from the conditioned media of these cells which has a molecular mass of approximately 60 kDa (12). Similar tumor cell-derived or induced factors with the same molecular mass have subsequently been reported and purified by several other investigators (14-17, 19). Autocrine motility factors are not
* The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “aduertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
$ T o whom correspondence should be addressed Laboratory of Pathology, National Cancer Institute, National Institutes of Health, Bldg. 10, Rm. 2A33, Bethesda, MD 20892. Tel.: 301-496-1843; Fax: 301-402-0043.
specific for a given type of cancer cell and have a wide spectrum of activity on many types of cancer cells (20) but little effect on normal fibroblasts or leukocytes. They act through cell-surface receptors (17, 21, 22) to stimulate both both random and directed migration (12, 14, 15).
In the present report we describe the identification, purifi- cation to homogeneity, characterization, and partial amino acid sequence analysis of a novel 125-kDa motility-stimulat- ing protein.
EXPERIMENTAL PROCEDURES’
Methods
Cell Culture-The human melanoma cell line A2058, originally isolated by Todaro et al. (23), was maintained as previously described (12).
Cell Motility Assays-The assay to determine motility was per- formed in triplicate using a 48-well microchemotaxis chamber as described previously in detail (24). The material to be tested was dialyzed into DPBS’ with calcium and magnesium supplemented with 0.1% (w/v) bovine serum albumin using Centricon 30 filtration units (Amicon). Chemotaxis was quantitated either by reading the stained membranes with a 2202 Ultroscan laser densitometer (Pharmacia- LKB Biotechnology Inc.) or by counting five randomly chosen high power fields (HPF) under light microscopy (400X) for each replicate. Densitometer units (wavelength = 633 nm) have been shown to be linearly related to the number of cells per HPF (10, 24).
Two-dimensional Gel Electrophoresis-For two-dimensional elec- trophoresis, the protein, in 20% ethylene glycol, was dried in a Speedvac concentrator and redissolved in loading solution: 9 M urea, 1% (v/v) pH 3-10 Bio-Lyte, and 2.5% (v/v) Nonidet P-40. This sample was then subjected to isoelectric focusing (25) using a Bio- Rad tube cell in 120 X 3-mm polyacrylamide tube gels containing 9 M urea, 2% (v/v) pH 3-10 Bio-Lyte, 0.25% (v/v) pH 8-10 Bio-Lyte, and 2.5% (v/v) Nonidet P-40 (Bio-Rad). Reservoir solutions were 0.01 M phosphoric acid and 0.02 M NaOH. Nonequilibrium isoelectric focusing (26) was run initially with constant voltage (500 V) for 5 h. Since the protein was basic, we repeated the procedure under equilib- rium conditions (500 V for 17 h). Electrophoresis in the second dimension was performed on a 7.5% polyacrylamide gel using the conditions of Laemmli (27). The gel was stained with Coomassie Blue G-250 as described previously (28).
Amino Acid Analysis-Acid hydrolysis of purified ATX and amino acid analysis (29) were performed at Yale University School of Medicine Protein and Nucleic Acid Facility, New Haven, CT.
Preparation of Peptides for Internal Sequence of Autotaxin-Ho- mogeneous ATX was sequentially digested with cyanogen bromide
Portions of this paper (including part of “Experimental Proce- dures,” Figs. 1-6, and Table I) are presented in miniprint at the end of this paper. Miniprint is easily read with the aid of a standard magnifying glass. Full size photocopies are included in the microfilm edition of the Journal that is available from Waverly Press.
The abbreviations used are: DPBS, Dulbecco’s phosphate-buff- ered saline; ATX, autotaxin; HPLC, high pressure liquid chromato- graph; PT, pertussis toxin; SDS, sodium dodecyl sulfate; HPF, high power field.
2524
Purification of Autotaxin, a Tumor Autocrine Motility Factor 2525
and, following reduction and pyridylethylation, with trypsin, accord- ing to the protocol of Stone et al. (30). The resulting fragments were then separated by gradient elution on an Aquapore RP300 C-8 reverse phase column: 0.1% (v/v) trifluoroacetic acid and 0-70% acetonitrile utilizing a Dionex AI-450 BioLC system.
Sequence Analysis of Peptides-The amino acid sequences of pep- tides resulting from digestion and purification of ATX peptides 1-7 were determined on a Porton Instruments 2020 off-line sequenator using standard program 1. Phenylthiohydantoins from sequenator runs were analyzed on a Beckman System Gold HPLC using a modified sodium acetate gradient program and a Hewlett-Packard C- 18 column.
ATX peptides 9-12 (ATX-100 to ATX-103) were sequenced at the Yale University School of Medicine Protein and Nucleic Acid Facility from gel-purified ATX (29).
Protein databases (31) that were searched for homologies in amino acid sequence with the ATX peptides include: GenBankT" (68.0), EMBL (27.0), SWISS-PROT (18.0), and GenPept (64.3).
RESULTS
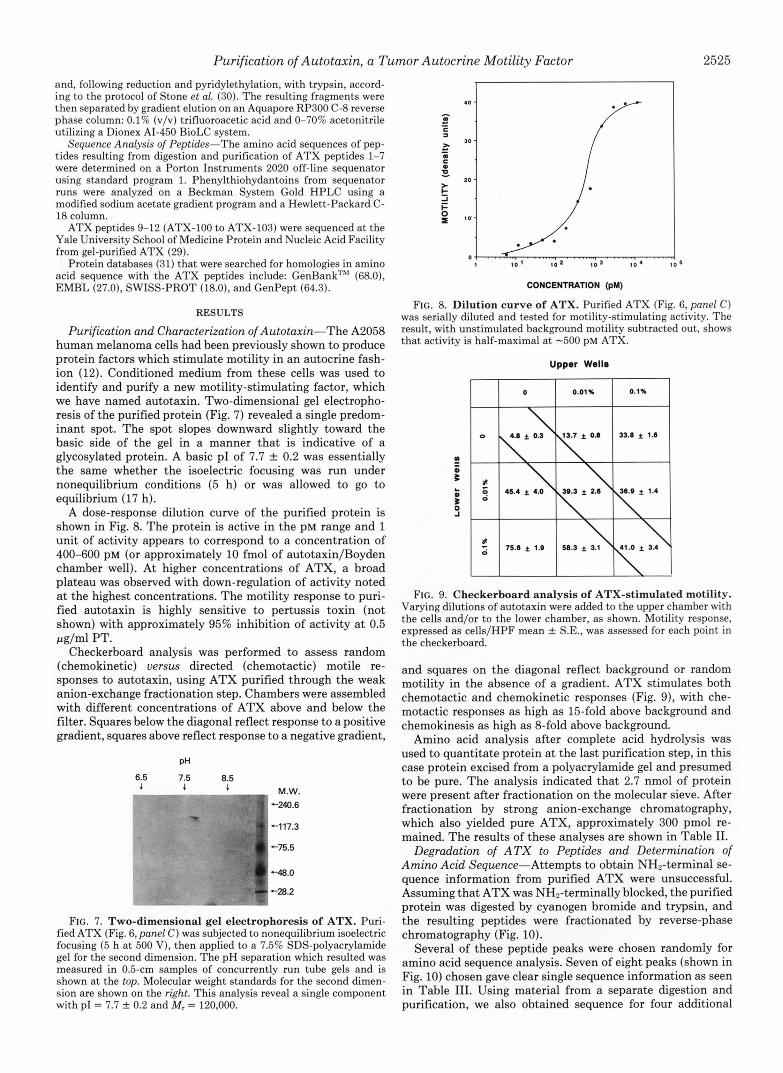
Purification and Characterization of Autotaxin-The A2058 human melanoma cells had been previously shown to produce protein factors which stimulate motility in an autocrine fash- ion (12). Conditioned medium from these cells was used to identify and purify a new motility-stimulating factor, which we have named autotaxin. Two-dimensional gel electropho- resis of the purified protein (Fig. 7) revealed a single predom- inant spot. The spot slopes downward slightly toward the basic side of the gel in a manner that is indicative of a glycosylated protein. A basic PI of 7.7 2 0.2 was essentially the same whether the isoelectric focusing was run under nonequilibrium conditions (5 h) or was allowed to go to equilibrium (17 h).
A dose-response dilution curve of the purified protein is shown in Fig. 8. The protein is active in the PM range and 1 unit of activity appears to correspond to a concentration of 400-600 PM (or approximately 10 fmol of autotaxin/Boyden chamber well). At higher concentrations of ATX, a broad plateau was observed with down-regulation of activity noted at the highest concentrations. The motility response to puri- fied autotaxin is highly sensitive to pertussis toxin (not shown) with approximately 95% inhibition of activity at 0.5 pg/ml PT.
Checkerboard analysis was performed to assess random (chemokinetic) uersus directed (chemotactic) motile re- sponses to autotaxin, using ATX purified through the weak anion-exchange fractionation step. Chambers were assembled with different concentrations of ATX above and below the filter. Squares below the diagonal reflect response to a positive gradient, squares above reflect response to a negative gradient,
pH
6.5 7.5 8.5 4 4 4 M.W.
-717.3
-75.5
W . 0
..-
FIG. 7. Two-dimensional gel electrophoresis of ATX. Puri- fied ATX (Fig. 6, panel C ) was subjected to nonequilibrium isoelectric focusing (5 h a t 500 V), then applied to a 7.5% SDS-polyacrylamide gel for the second dimension. The pH separation which resulted was measured in 0.5-cm samples of concurrently run tube gels and is shown at the top. Molecular weight standards for the second dimen- sion are shown on the right. This analysis reveal a single component with PI = 7.7 f 0.2 and M, = 120,000.
CONCENTRATION (pM)
FIG. 8. Dilution curve of ATX. Purified ATX (Fig. 6, panel C) was serially diluted and tested for motility-stimulating activity. The result, with unstimulated background motility subtracted out, shows that activity is half-maximal a t -500 pM ATX.
Upper Wells
FIG. 9. Checkerboard analysis of ATX-stimulated motility. Varying dilutions of autotaxin were added to the upper chamber with the cells and/or to the lower chamber, as shown. Motility response, expressed as cells/HPF mean f S.E., was assessed for each point in the checkerboard.
and squares on the diagonal reflect background or random motility in the absence of a gradient. ATX stimulates both chemotactic and chemokinetic responses (Fig. 9), with che- motactic responses as high as 15-fold above background and chemokinesis as high as %fold above background.
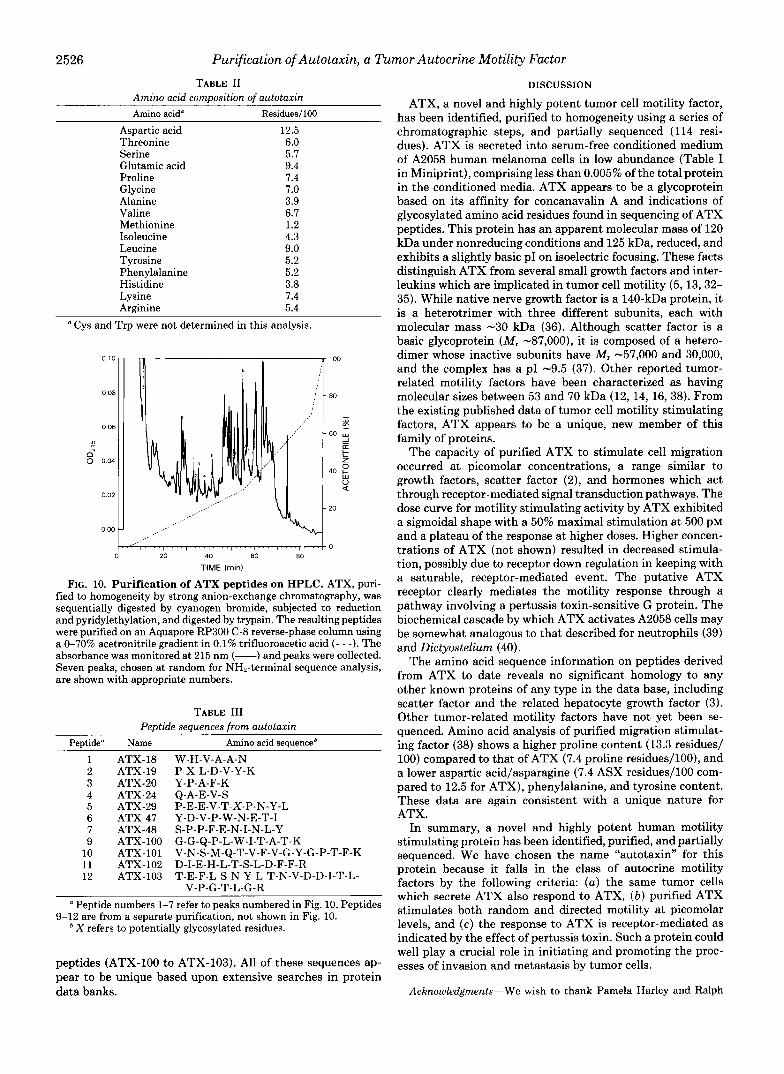
Amino acid analysis after complete acid hydrolysis was used to quantitate protein at the last purification step, in this case protein excised from a polyacrylamide gel and presumed to be pure. The analysis indicated that 2.7 nmol of protein were present after fractionation on the molecular sieve. After fractionation by strong anion-exchange chromatography, which also yielded pure ATX, approximately 300 pmol re- mained. The results of these analyses are shown in Table 11.
Degradation of ATX to Peptides and Determination of Amino Acid Sequence-Attempts to obtain NH2-terminal se- quence information from purified ATX were unsuccessful. Assuming that ATX was NHP-terminally blocked, the purified protein was digested by cyanogen bromide and trypsin, and the resulting peptides were fractionated by reverse-phase chromatography (Fig. 10).
Several of these peptide peaks were chosen randomly for amino acid sequence analysis. Seven of eight peaks (shown in Fig. 10) chosen gave clear single sequence information as seen in Table 111. Using material from a separate digestion and purification, we also obtained sequence for four additional
2526 Purification of Autotaxin, a Tumor Autocrine Motility Factor
TABLE I1 Amino acid composition of autotaxin Amino acid" Residues/100
Aspartic acid Threonine Serine Glutamic acid Proline Glycine Alanine Valine Methionine Isoleucine Leucine Tyrosine Phenylalanine Histidine Lysine Arginine
12.5 6.0 5.7 9.4 7.4 7.0 3.9 6.7 1.2 4.3 9.0 5.2 5.2 3.8 7.4 5.4
Cys and Trp were not determined in this analysis.
__,' 1 " " 1 " " 1 " " 1 " " , ' " ' " ' ' ' , ' " , " ' ' 0
0 20 40 60 80
TIME (rnin)
FIG. 10. Purification of ATX peptides on HPLC. ATX, puri- fied to homogeneity by strong anion-exchange chromatography, was sequentially digested by cyanogen bromide, subjected to reduction and pyridylethylation, and digested by trypsin. The resulting peptides were purified on an Aquapore RP300 C-8 reverse-phase column using a 0-70% acetronitrile gradient in 0.1% trifluoroacetic acid ( - - -). The absorbance was monitored at 215 nm (-) and peaks were collected. Seven peaks, chosen at random for NH2-terminal sequence analysis, are shown with appropriate numbers.
TABLE I11 Peptide sequences from autotaxin
Peutide" Name Amino acid sequenceb
1 2 3 4 5 6 7 9
10 11 12
ATX-18 ATX-19 ATX-20 ATX-24 ATX-29 ATX-47 ATX-48 ATX-100 ATX-101 ATX-102 ATX-103
W-H-V-A-A-N P-X-L-D-V-Y-K Y-P-A-F-K Q-A-E-V-S P-E-E-V-T-X-P-N-Y-L Y-D-V-P-W-N-E-T-I S-P-P-F-E-N-I-N-L-Y G-G-Q-P-L-W-I-T-A-T-K V-N-S-M-Q-T-V-F-V-G-Y-G-P-T-F-K D-I-E-H-L-T-S-L-D-F-F-R T-E-F-L-S-N-Y-L-T-N-V-D-D-I-T-L-
V-P-G-T-L-G-R Peptide numbers 1-7 refer to peaks numbered in Fig. 10. Peptides
9-12 are from a separate purification, not shown in Fig. 10. * X refers to potentially glycosylated residues.
peptides (ATX-100 to ATX-103). All of these sequences ap- pear to be unique based upon extensive searches in protein data banks.
DISCUSSION
ATX, a novel and highly potent tumor cell motility factor, has been identified, purified to homogeneity using a series of chromatographic steps, and partially sequenced (114 resi- dues). ATX is secreted into serum-free conditioned medium of A2058 human melanoma cells in low abundance (Table I in Miniprint), comprising less than 0.005% of the total protein in the conditioned media. ATX appears to be a glycoprotein based on its affinity for concanavalin A and indications of glycosylated amino acid residues found in sequencing of ATX peptides. This protein has an apparent molecular mass of 120 kDa under nonreducing conditions and 125 kDa, reduced, and exhibits a slightly basic PI on isoelectric focusing. These facts distinguish ATX from several small growth factors and inter- leukins which are implicated in tumor cell motility (5, 13,32- 35). While native nerve growth factor is a 140-kDa protein, it is a heterotrimer with three different subunits, each with molecular mass -30 kDa (36). Although scatter factor is a basic glycoprotein (Mr -87,000), it is composed of a hetero- dimer whose inactive subunits have M, -57,000 and 30,000, and the complex has a PI -9.5 (37). Other reported tumor- related motility factors have been characterized as having molecular sizes between 53 and 70 kDa (12, 14, 16, 38). From the existing published data of tumor cell motility stimulating factors, ATX appears to be a unique, new member of this family of proteins.
The capacity of purified ATX to stimulate cell migration occurred at picomolar concentrations, a range similar to growth factors, scatter factor (Z), and hormones which act through receptor-mediated signal transduction pathways. The dose curve for motility stimulating activity by ATX exhibited a sigmoidal shape with a 50% maximal stimulation at 500 pM and a plateau of the response at higher doses. Higher concen- trations of ATX (not shown) resulted in decreased stimula- tion, possibly due to receptor down regulation in keeping with a saturable, receptor-mediated event. The putative ATX receptor clearly mediates the motility response through a pathway involving a pertussis toxin-sensitive G protein. The biochemical cascade by which ATX activates A2058 cells may be somewhat analogous to that described for neutrophils (39) and Dictyostelium (40).
The amino acid sequence information on peptides derived from ATX to date reveals no significant homology to any other known proteins of any type in the data base, including scatter factor and the related hepatocyte growth factor (3). Other tumor-related motility factors have not yet been se- quenced. Amino acid analysis of purified migration stimulat- ing factor (38) shows a higher proline content (13.3 residues/ 100) compared to that of ATX (7.4 proline residues/lOO), and a lower aspartic acid/asparagine (7.4 ASX residues/100 com- pared to 12.5 for ATX), phenylalanine, and tyrosine content. These data are again consistent with a unique nature for ATX.
In summary, a novel and highly potent human motility stimulating protein has been identified, purified, and partially sequenced. We have chosen the name "autotaxin" for this protein because it falls in the class of autocrine motility factors by the following criteria: (a) the same tumor cells which secrete ATX also respond to ATX, ( b ) purified ATX stimulates both random and directed motility at picomolar levels, and ( c ) the response to ATX is receptor-mediated as indicated by the effect of pertussis toxin. Such a protein could well play a crucial role in initiating and promoting the proc- esses of invasion and metastasis by tumor cells.
Acknowledgments-We wish to thank Pamela Harley and Ralph

Purification of Autotaxin, a Tumor Autocrine Motility Factor 2527
Hopkins 111 at the Fermentation Unit, Frederick Cancer Research and Development Facility, PRI/DyneCorp, Frederick, MD, for their patience and help in the large scale production of A2058-conditioned medium.
REFERENCES
1. Singer, S. J., and Kupfer, A. (1986) Annu. Reu. Cell Biol. 2, 337- 365
2. Rosen, E. M., Meromsky, L., Setter, E., Vinter, D. W., and Goldberg, I. D. (1990) Proc. SOC. Exp. Bid. Med. 195, 34-43
3. Weidner, K. M., Behrens, J., Vandekerckhove, J., and Birch- meier, W. (1990) J . Cell. Biol. 111, 2097-2108
4. Kahan, B. W., and Kramp, D. C. (1987) Cancer Res. 47, 6324- 6328
5. Stracke, M. L., Kohn, E. C., Aznavoorian, S. A., Wilson, L. L., Salomon, D., Kmtzsch, H. C., Liotta, L. A., and Schiffmann, E. (1988) Biochem. Biophys. Res. Commun. 153, 1076-1083
6. Tamm, I., Cardinale, I., Kmeger, J., Murphy, J. S., May, L. T., and Sehgal, P. B. (1989) J. Exp. Med. 170, 1649-1669
7. Wang, J. M., Taraboletti, G., Matsushima, K., Van Damme, J., and Montovani, A. (1990) Biochem. Biochem. Res. Commun.
8. Jouanneau, J., Gavrilovic, J., Caruelle, D., Jaye, M., Moens, G., Caruelle, J. P., and Thiery, J. P. (1991) Proc. Natl. Acad. Sci.
9. McCarthy, J. B., and Furcht, L. T. (1984) J. Cell B i d . 98, 1474-
169,165-170
U. S. A. 88, 2893-2897
* ." 10.
11.
12.
13.
14.
15.
16.
17.
148U Taraboletti, G., Roberts, D. D., and Liotta, L. A. (1987) J. Cell
Biol. 105, 2409-2415 Aznavoorian, S., Stracke, M. L., Krutzsch, H., Schiffmann, E.,
and Liotta, L. A. (1990) J. Cell Biol. 110, 1427-1438 Liotta, L. A,, Mandler, R., Murano, G., Katz, D. A., Gordon, R.
K., Chiang, P. K., and Schiffmann, E. (1986) Proc. Natl. Acad. Sci. U. S. A. 83, 3302-3306
Ruff, M., Schiffmann, E., Terranova, V., and Pert, C. B. (1985) Clin. Immunol. Immunopathol. 37, 387-396
Atnip, K. D., Carter, L. M., Nicolson, G. L., and Dabbous, M. K. (1987) Biochem. Biophys. Res. Commun. 146,996-1002
Ohnishi, T., Arita, N., Hayakawa, T., Izumoto, S., Taki, T., and Yamamoto, H. (1990) J. Neurosurg. 73,881-888
Silletti, S., Watanahe, H., Hogan, V., Nabi, I. R., and Raz, A. (1991) Cancer Res. 51, 3507-3511
Watanabe, H., Carmi, P., Hogan, V., Raz, T., Siletti, S., Nahi, I. R., and Raz, A. (1991) J . Biol. Chem. 266, 13442-13448
18. Liotta, L. A., and Schiffmann, E. (1988) Cancer Surveys 7, 631- 652
19. Schor, S. L., Schor, A. M., Grey, A. M., and Rushton, G. (1988) J. Cell Sci. 90, 391-399
20. Kohn, E. C., Francis, E. A., Liotta, L. A,, and Schiffmann, E. (1990) Int. J . Cancer 46, 287-292
21. Stracke, M. L., Guirguis, R., Liotta, L. A., and Schiffmann, E. (1987) Biochem. Biophys. Res. Commun. 146,339-345
22. Nabi, I. R., Watanahe, H., and Raz, A. (1990) Cancer Res. 50,
23. Todaro, G. J., Fryling, C., and DeLarco, J. E. (1980) Proc. Natl. Acad. Sci. U. S. A. 77, 5258-5262
24. Stracke, M. L., Engel, J. D., Wilson, L. W., Rechler, M. M., Liotta. L. A., and Schiffmann, E. (1989) J. Biol. Chem. 264,
409-414
21544121549' 25. O'Farrell. P. H. (1975) J. Biol. Chem. 250,4007-4021 26. O'Farrell; P. Z., Goodman, H. M., and O'Farrell, P. H. (1977) Cell
12.1133-1142 27. Laemmli, U. K. (1970) Nature 227, 680-685 28. Neuhoff, V., Arold, N., Taube, D, and Ehrhardt, W. (1988)
Electrophoresis 9, 255-262 29. Stone, K., McNulty, D., LoPresti, M., Crawford, J., DeAngelis,
R., and Williams, K. (1991) in Protein Chemistry, I I I (Angeletti, R., ed) Academio Press, NY, in press
30. Stone, K. L., LoPresti, M. B., Crawford, J. M., DeAngelis, R., and Williams, K. R. (1989) in A Practical Guide to Protein and Peptide Purification for Microsequencing (Matsudaira, P. T., ed) pp. 33-47, Academic Press, New York
31. Pearson, W. R., and Lipman, D. J. (1988) Proc. Natl. Acad. Sci.
32. Maciag, T., Mehlman, T., Friesel, R., and Schreiher, A. B. (1984) Science 225, 932-935
33. Gospodarowicz, D., Cheng, J., Lui, G. M., Baird, A., and Bohlent, P. (1984) Proc. Natl. Acad. Sci. U. S. A. 81,6963-6967
34. Van Snick, J. (1990) Annu. Reu. Immunol. 8, 253-278 35. Yoshimura, T., Matsushima, K., Tanaka, S., Robinson, E. A.,
Appella, E., Oppenheim, J. J., and Leonard, E. J . (1987) Proc. Natl. Acad. Sci. U. S. A . 84, 9233-9237
U. 5'. A. 85, 2444-2448
36. Varon, S., Nomura, J., and Shooter, E. M. (1967) Proc Natl. Acad. Sci. U. S. A. 57,1782-1789
37. Gherardi, E., Gray, J., Stoker, M., Perryman, M., and Furlong, R. (1989) Proc. Natl. Acad. Sci. U. S. A . 86, 5844-5848
38. Grey, A. M., Schor, A. M., Rushton, G., Ellis, I., and Schor, S. L. (1989) Proc. Natl. Acad. Sci. U. S. A. 86, 2438-2442
39. Verghese, M. W., Smith, C. D., and Snyderman, R. (1985) Biochem. Biophys. Res. Commun. 127, 450-457
40. Newell, P. C., Europe-Finner, G. N., Small, N. V., and Liu, G. (1988) J. Cell Sci. 89, 123-127
2528 Purification of Autotaxin, a Tumor Autocrine Motility Factor
TIME (min)
0
Flaura 2
J a 1
25, , 1.0
- 0.04
- 0.m - i 2 T r -0.02 2
- 0.01
- 0.00
A C
,173- cj - 480- (I! 75.5- 0
28.2- I, 19.4- 0
I 2
Purification of Autotaxin, a Tumor Autocrine Motility Factor 2529


















