
IDENTIFICATION OF ACTINOMYCES - Journal of …jb.asm.org/content/88/2/477.full.pdfces bovis (Harz,...
14
JOURNAL OF BACrERIOLOGY Vol. 88, No. 2, P. 477-490 August, 1964 Copyright © 1964 American Society for Microbiology Printed in U.S.A. IDENTIFICATION OF SPECIES OF ACTINOMYCES LUCILLE K. GEORG, GORDON W. ROBERTSTAD1, AND SHERRY A. BRINKMAN Comnmunicable Disease Center, Public Health Service, Atlanta, Georgia Received for publication 23 March 1964 ABSTRACT GEORG, LUCILLE K. (Communicable Disease Center, Atlanta, Ga.), GORDON W. ROBERTSTAD, AND SHERRY A. BRINKMAN. Identification of species of Actinomyces. J. Bacteriol. 88:477-490. 1964.-Four unusual isolates of Actinomyces species were compared with six control strains of well-identified Actinomyces species. Their oxygen requirements and their morphological and bio- chemical characteristics were determined. Final identifications were confirmed by cell-wall analy- ses for amino acid and carbohydrate composition and by agar-gel techniques. Two strains, A-13-R and A-13-S, isolated from the same cow head were shown to represent rough and smooth variants of A. bovis. The strains were not only morphologi- cally distinct, but had different biochemical reac- tions and antigenic makeup. The two other strains, X407 and X573, isolated from human pleural fluid and a lung abscess, respectively, were shown to represent a new Actinomyces species. For many years it was believed that Actinomy- ces bovis (Harz, 1877), isolated from lumpy jaw of cattle, and A. israelii (Kruse, 1896) Lachner- Sandoval (Breed, Murray, and Smith, 1957), isolated from human actinomycosis, were one species. However, studies by Erikson (1940, 1949), Thompson (1950), and Pine, Howell, and Watson (1960) provided morphological and bio- chemical evidence that there are two agents of actinomycosis: A. bovis, usually associated with bovine infection; and A. israelii, the common cause of human infections. Recently, Buchanan and Pine (1962) described a third agent of actino- mycosis, A. propionicus. This organism was isolated from human lacrimal canaliculitis. Division of the agents of actinomycosis into several species has received further support from cell-wall analyses by Cummins and Harris (1958), serological studies by King and Meyer (1963), and electron microscopy studies by Overman and Pine (1963). It has been reported also that A. I Supported by a National Institutes of Health postdoctoral fellowship (1962-1963). Present ad- dress: Division of Veterinary Science, University of Wyoming, Laramie. israelii is not host-specific as previously believed, as an organism isolated from "lumpy jaw" in a cow was identified as A. israelii by Pine et al. (1960). Although these organisms are referred to as the "anaerobic Actinomyces," their oxygen toler- ance varies from species to species, and even among strains of a single species. They are more correctly called "microaerophilic to anaerobic actinomycetes. " In the identification of these organisms, a fourth species, A. naeslundii, must also be taken into consideration. This organism, described by Thompson and Lovestedt (1951) and further delineated by Howell et al. (1959), has a greater oxygen tolerance than the other species, and has been called "facultative." However, under ana- erobiosis with increased C02 , isolation and main- tenance of the culture are more successful. A. naeslundii has not been proven to be a cause of human disease, but it is isolated frequently from human clinical materials and, because of its morphological and physiological similarity to the agents of actinomycosis, must be differentiated from them. The purpose of the present report is to describe four unusual isolates of Actinomyces, two of bovine (A-13-R and A-13-S) and two of human origin (X407 and X573), and to compare them with authenticated strains of A. bovis, A. israelii, A. naeslundii, and A. propionicus. MATERIALS AND METHODS Organisms. Control strains were well-identified isolates of A. israelii (ATCC 10048, 12102, and CDC A601), A. bovis (ATCC 13683 and CDC A-9), A. naeslundii (ATCC 12104 and Howell CS 1752), and A. propionicus (ATCC 14157). Strains ATCC 10048, 12102, and 13683 were studied recently by 13 members of an Interna- tional Committee for the Study of the Micro- aerophilic Actinomyces. All 13 members, who studied these strains as unknowns, were in agree- ment with the identifications as listed above (J. M. Slack, personal communication). 477 on June 2, 2018 by guest http://jb.asm.org/ Downloaded from
Transcript of IDENTIFICATION OF ACTINOMYCES - Journal of …jb.asm.org/content/88/2/477.full.pdfces bovis (Harz,...

JOURNAL OF BACrERIOLOGYVol. 88, No. 2, P. 477-490 August, 1964Copyright © 1964 American Society for Microbiology
Printed in U.S.A.
IDENTIFICATION OF SPECIES OF ACTINOMYCESLUCILLE K. GEORG, GORDON W. ROBERTSTAD1, AND SHERRY A. BRINKMAN
Comnmunicable Disease Center, Public Health Service, Atlanta, Georgia
Received for publication 23 March 1964
ABSTRACTGEORG, LUCILLE K. (Communicable Disease
Center, Atlanta, Ga.), GORDON W. ROBERTSTAD,AND SHERRY A. BRINKMAN. Identification ofspecies of Actinomyces. J. Bacteriol. 88:477-490.1964.-Four unusual isolates of Actinomycesspecies were compared with six control strains ofwell-identified Actinomyces species. Their oxygenrequirements and their morphological and bio-chemical characteristics were determined. Finalidentifications were confirmed by cell-wall analy-ses for amino acid and carbohydrate compositionand by agar-gel techniques. Two strains, A-13-Rand A-13-S, isolated from the same cow head wereshown to represent rough and smooth variants ofA. bovis. The strains were not only morphologi-cally distinct, but had different biochemical reac-tions and antigenic makeup. The two other strains,X407 and X573, isolated from human pleural fluidand a lung abscess, respectively, were shown torepresent a new Actinomyces species.
For many years it was believed that Actinomy-ces bovis (Harz, 1877), isolated from lumpy jawof cattle, and A. israelii (Kruse, 1896) Lachner-Sandoval (Breed, Murray, and Smith, 1957),isolated from human actinomycosis, were onespecies. However, studies by Erikson (1940,1949), Thompson (1950), and Pine, Howell, andWatson (1960) provided morphological and bio-chemical evidence that there are two agents ofactinomycosis: A. bovis, usually associated withbovine infection; and A. israelii, the commoncause of human infections. Recently, Buchananand Pine (1962) described a third agent of actino-mycosis, A. propionicus. This organism wasisolated from human lacrimal canaliculitis.
Division of the agents of actinomycosis intoseveral species has received further support fromcell-wall analyses by Cummins and Harris (1958),serological studies by King and Meyer (1963),and electron microscopy studies by Overman andPine (1963). It has been reported also that A.
I Supported by a National Institutes of Healthpostdoctoral fellowship (1962-1963). Present ad-dress: Division of Veterinary Science, Universityof Wyoming, Laramie.
israelii is not host-specific as previously believed,as an organism isolated from "lumpy jaw" in acow was identified as A. israelii by Pine et al.(1960).Although these organisms are referred to as
the "anaerobic Actinomyces," their oxygen toler-ance varies from species to species, and evenamong strains of a single species. They are morecorrectly called "microaerophilic to anaerobicactinomycetes. "
In the identification of these organisms, afourth species, A. naeslundii, must also be takeninto consideration. This organism, described byThompson and Lovestedt (1951) and furtherdelineated by Howell et al. (1959), has a greateroxygen tolerance than the other species, and hasbeen called "facultative." However, under ana-erobiosis with increased C02 , isolation and main-tenance of the culture are more successful. A.naeslundii has not been proven to be a cause ofhuman disease, but it is isolated frequently fromhuman clinical materials and, because of itsmorphological and physiological similarity to theagents of actinomycosis, must be differentiatedfrom them.The purpose of the present report is to describe
four unusual isolates of Actinomyces, two ofbovine (A-13-R and A-13-S) and two of humanorigin (X407 and X573), and to compare themwith authenticated strains of A. bovis, A. israelii,A. naeslundii, and A. propionicus.
MATERIALS AND METHODS
Organisms. Control strains were well-identifiedisolates of A. israelii (ATCC 10048, 12102, andCDC A601), A. bovis (ATCC 13683 and CDCA-9), A. naeslundii (ATCC 12104 and Howell CS1752), and A. propionicus (ATCC 14157).
Strains ATCC 10048, 12102, and 13683 werestudied recently by 13 members of an Interna-tional Committee for the Study of the Micro-aerophilic Actinomyces. All 13 members, whostudied these strains as unknowns, were in agree-ment with the identifications as listed above (J.M. Slack, personal communication).
477
on June 2, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

GEORG, ROBERTSTAD, AND BRINKMNIAN
Strains CDC A-9 and A-601 have been studiedby a number of workers. Their characteristics astypical A. bovis and A. israelii, respectively, havebeen delineated by Pine et al. (1960).The two A. naeslundii strains, ATCC 12104
and CS 1752, were studied by HowA-ell et al.(1959, 1962).
Strain ATCC 14157 is the tyl)e species A.propionicus described by Buchanan and Pine(1962), and is the only known strain of thisspecies.The unknown strains studied are identified as
A-13-R, A-13-S, X407, and X573. The sources ofthese strains are listed in Table 1.
TABLE 1. Actinontyces species studied
Strain no.* Identification Original source
ATCC 12102
ATCC 10048
CDC A601
ATCC 13683
CDC A-9
ATCC 12104
CS 1752
ATCC 14157
A. israelii
A. israelii
A. israelii
A. bovis
A. bovis
A. naesl'undii
A. naeslinr(lii
A. propionicois
A-13-R and A-13-S
X407
X573
Isolated by Luther Thompson from a human brain ab-scess, 1959 (Thompson 277).
Isolated by C. W. Emmons from human pleural fluid,1946 (Emmons '1829).
Isolated by the CDC Mycology Unit, from bovine''lumpy jaw,'" 1952.
Isolated by Leo Pine from bovine "lumpy jaw," 1959(Pine # 1).
Isolated by the CDC Mycology Unit, from bovine"lumpy jaw," 1956.
Isolated by Arden Howell, Jr., from a huiman sinus, 1951(Howell # 279).
Isolated by Arden Howell, Jr., from human tooth(plaque from root and crown surfaces), 1949.
Isolated from a human tear duct by Pine and Hardin(1959) and described by Buchanan and Pine (1962) as anew species. This is the only known strain of thisspecies.
Isolated from cow head # 13 at CDC Mycology Unit,6 March 19(61. The head, obtained from a local abat-toir, showed lesions suggestive of actinomycosis. Alarge quantity of granules was obtained from ab-scesses of the jaw. These were clubbed and composedof branched mycelium, 1 ,u or less in diameter.
Culture received by CDC Mycology Unit frorn FloridaState Board of Health, 11 May 1961 (CDC 45-332-61).Isolated from chest fluid of patient with empyema.
Culture received by CDC Mycology Unit from DexterH. Howard, University of California Medical Center,14 January 1963 (CDC 45-59-63). Isolated from humanlung abscess. Resected tissue showed masses of gram-positive filaments compatible to organisms of the genusA ctinomttyces.
* ATCC = American Type Culture Collection, Washington, D.C.; CD)C = Communicable DiseaseCenter, Atlanta, Ga.; CS = Dr. Howell's designation.
478 J. BACTERIOL.
on June 2, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

IDENTIFICATION OF ACTINOMYCES
Isolation methods. Primary isolation fromclinical material (A-13-R and A-13-S) was madeby streaking washed and crushed granules onBrain Heart Infusion (BHI) Agar plates. Thesewere incubated at 37 C under an atmosphere of5% C02 and 95% N2 (v/v). After 7 days, twocolony types were observed. These were smooth,slightly convex, soft colonies with an entire edge,designated "S," and raised, rough-surfaced,"molar tooth" colonies with irregular, sometimesfringed edges, designated "R." These wereselected and studied separately as A-13-S andA-13-R isolates.
These cultures, as well as the reference diagnos-tic cultures (X407 and X573) and the controlcultures, received from various investigators,were transferred to "reinforced thioglycolatebroth," i.e., thioglycolate broth with 10% horsemeat infusion (RT broth); after several days ofincubation, they were streaked on BHI and in-cubated anaerobically as above. Isolated colonieswere picked, and the process was repeated twiceto insure purity of strains.
Preservation of stock cultures. Inoculation ofpurified strains was made to "Actinomyces main-tenance broth" (AM broth), in which the strainswere grown at 37 C under pyrogallol-carbonateseals as described by Howell and Pine (1956).After 5 to 7 days, the tubes were placed at -21 Cand maintained at that temperature for theduration of the experiment. AM broth, which is amodification of the synthetic medium of Howelland Pine (1956), contains (g per liter): KH2PO4,30.0; (NH4)2SO4, 2.0; MgSO4 7H20, 0.4; CaC12,0.04; Heart Infusion Broth, 25; dextrose, 5; cys-teine hydrochloride, 1; pancreatic digest of casein(Difco Casitone or BBL Trypticase), 4; yeastextract, 5; soluble starch, 1; distilled water, 1liter. Adjust pH to about 7.2. Dispense 8 ml pertube (18 X 150 ml), cotton-plugged. Autoclaveat 15 psi (120 C) for 10 min. The final pH shouldbe 6.8 to 7.0.
Mllorphological studies. Inoculum from RT brothwas streaked on BHI Agar plates for study ofcolony morphology. All plates were incubated at37 C under anaerobic conditions (5% C02 and95% N2) Young colonies (24 to 48 hr) wereexamined microscopically at 100 X. Maturecolonies were examined at 7 to 10 days. Gramstains of cells taken from the various media werealso made.
Oxygen-requirement determinations. Inoculumwas taken from 3- to 4-day-old cultures in AM
broth, and streaked on cotton-stoppered BHIAgar slants. To minimize variation in quantity ofinoculum, a sample of well-mixed culture suspen-sion was removed with a capillary pipette, andone drop was carefully streaked from the tip ofthe pipette in a single line from the base to thetop of an agar slant. Six tubes were inoculated inthis manner. All were incubated at 37 C: twoanaerobically (pyrogallol-carbonate seal), twounder microaerophilic conditions (candle jar), andtwo aerobically. The amount of growth was esti-mated visually as i to 4+ at 3 and 10 days. If asimilar pattern was not observed in replicatetubes, the tests were repeated.
Biochemical tests. Biochemical tests were doneaccording to the methods described by Howelland Pine (1956) and Howell et al. (1959), exceptthat the basal medium consisted of: Heart Infu-sion Broth, 25 g; pancreatic digest of casein, 4 g;and yeast extract, 5 g (per liter of distilled waterat pH 7.0).
Inoculum for all biochemical tests, except forthe carbohydrate fermentations, was taken from3-day-old cultures in AM broth. Inoculum forcarbohydrate fermentations was taken from cul-tures made in sugar-free Heart Infusion Broth.The following biochemical tests were per-
formed: catalase test, starch hydrolysis, nitratereduction, litmus milk reactions, H2S production,and acid from carbohydrates (dextrose, xylose,mannitol, raffinose, and mannose).
In the fermentation tests, the carbohydrateswere added to the basal medium at a concentra-tion of 0.5%. A drop in pH of less than 0.49 pHunit (as compared with control tubes) was con-sidered to be a negative reaction; a drop of 0.5 to0.9 units, a plus-minus reaction; and a drop of onepH unit or more was considered positive.
Starch hydrolysis was determined by growingthe strains on slants of basal medium to whichhad been added 5 g of soluble starch and 15 g ofagar per liter. Hydrolysis was determined byflooding the slants with Gram's iodine solution.
Litmus milk reactions were studied in litmusmilk reinforced with 0.5% yeast extract and 0.3%glucose.
Nitrate reduction was tested by growing thestrains on the basal medium containing 0.1%KNO3 .H2S production was demonstrated by growing
the organisms on BHI slants in cotton-pluggedtubes with strips of lead acetate paper suspendedabove the medium, and incubating at 37 C in an
479VOL. 88, 1964
on June 2, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

GEORG, ROBERTSTAD, AND BRINKMAN
anaerobic jar (95% N2 plus 5% CO2). (False reac-tions may be obtained with lead acetate paper, iftubes are sealed with pyrogallol-carbonate seals.)
Catalase tests were performed according tostandard procedures.
All cultures made for the biochemical tests(except for H2S production) were incubated at37 C under pyrogallol-carbonate seals and read at3 and 10 days.
Cell-wall analyses. Formalinized (0.5% forma-lin), washed cells were prepared from strainsA-13-R, A-13-S, X407, and X573 which had beengrown for 10 to 12 days at 37 C. The culturebottles contained 200 ml of AM broth with 200mg of sodium thioglycolate.
Cell-wall analyses by chromatographic tech-nique for amino acids and carbohydrates werekindly performed by C. S. Cummins, LondonHospital Medical College, University of London,London, England, according to methods ofCummins and Harris (1958).
Serological methods. Actinomyces antisera wereprepared in rabbits against the six control strainsand the four unidentified isolates. Methods forpreparing soluble antigens and cell suspensionsfor rabbit inoculation were based on the report byKing and Meyer (1963).
Formalinized, washed cells were prepared asabove for cell-wall analyses except that the super-natant broth was retained for preparation ofsoluble antigens. Suspensions were adjusted tobetween no. 4 and no. 5 McFarland nephelometerdensities, and were shaken in a paint shaker for1 hr with equal volumes of glass beads (diameterabout 420 ,u). (After shaking, microscopic examin-ation revealed disruption of all cell clumps andbreakage of some of the cells.)
After obtaining preimmunization serum speci-mens, 1-mi suspensions of each strain wereinoculated intravenously into each of three rab-bits three times per week for 4 weeks; 1 week aftercompletion of this schedule, serum specimenswere obtained for antigen analyses.
After removal of the organisms by centrifuga-tion, soluble antigens were precipitated by adding2 volumes of acetone in the cold (5 C) to 1 volumeof supernatant fluid; the mixture was thenallowed to stand overnight at 5 C. The acetonewas removed and the precipitate was redissolvedin sterile distilled water containing Merthiolate(1:10,000), in an amount equalling 3% of theoriginal supernatant. Soluble antigens were keptfrozen until needed for testing.
Agar-gel techniques for determining antigen-antibody reactions were carried out according toOuchterlony's (1949) methods, except that theagar-gel employed was prepared according to themethod of Schubert, Lynch, and Ajello (1961).Reactions were observed at 2, 3, 4, and 5 days.
RESULTSControl strains. Morphological studies of the
control strains are not presented here, as thesehave been described adequately in the literature.In general, our results coincided with those de-scriptions. Results of the tests for oxygen require-ments and the biochemical reactions are given inTable 2. Serological studies are reported inTable 5.
Unknown strains. Results of studies with eachof the four unknown strains are discussed individ-ually. Table 2 lists the oxygen requirements andbiochemical reactions of the known and unknownstrains. Table 3 describes the morphological char-acteristics of the unknown strains. Cell-wall com-position of representative Actinomyces speciesand unknown strains is indicated in Table 4, andTable 5 displays the antigenic relationshipsamong these strains.Unknown strain A-13-R. Growth occurred only
under anaerobic conditions (Table 2). Cultures inRT broth consistently formed small discretegranular and large lobulated "bread-crumb"colonies distributed throughout the lower four-fifths of the medium. The broth itself remainedclear after shaking the tubes (Table 3). Coloniesgrown for 48 hr on BHI plates were microscopicin size. Under 100 X magnification they appearedto be composed of delicate branched filamentsthat radiated from a central point. They closelyadhered to the agar surface. The colonies weresimilar to the filamentous microcolonies of Actino-myces which have been described as "spider," or"A. israelii type," colonies by Pine et al. (1960).All of the A-13-R microcolonies were of this type,both on first isolation and after 2 years of re-peated subculture. Colonies (7 to 10 days old)were dull white and irregularly heaped. Some ofthe colonies had a fringe of mycelium. Many ofthe colonies had depressed centers and presentedthe "molar tooth" formation commonly seen inmature colonies of A. israelii. On first isolation,and in many subcultures, this was the only typeof mature colony observed. However, after 2 yearsof storage in frozen AM broth, it was found that,although all of the microcolonies were still of the
480 J. BACTERIOL.
on June 2, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

IDENTIFICATION OF ACTINOMYCES
O) O dtz
L ,
C.)
.;Q + +++
-0
P..
W B + ++ oo + t Q + +o + +
r + + + OCO+C) + + C) C)++
7+ +
|~~~~C ¢ O + 3) 0 +++°°
t2U' "3
±000+ + +000±o00nmu~~~~~~~~~~"
C~~~~~~~~~~~~~~~~~.
C> ++ + 0 + ~ +
1::4~00+~+
U00~~~~~~~~~"
++++~g
E-0 ++ "0 + CC)-
9' + + 0C)+.+)+c )c
0.
Cd
._1
OD~~~~~Ca C;. * *a °CC3*S*t o D;
4
4
. ,.) E E .C V° o 'o m
0 t )C .~0
481VOL. 88, 1964
C*
0C.)l
rsC)
-Z,t.)Q
,C.)0
q6)
CA)-It1
CsZ)l
CZ)
on June 2, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
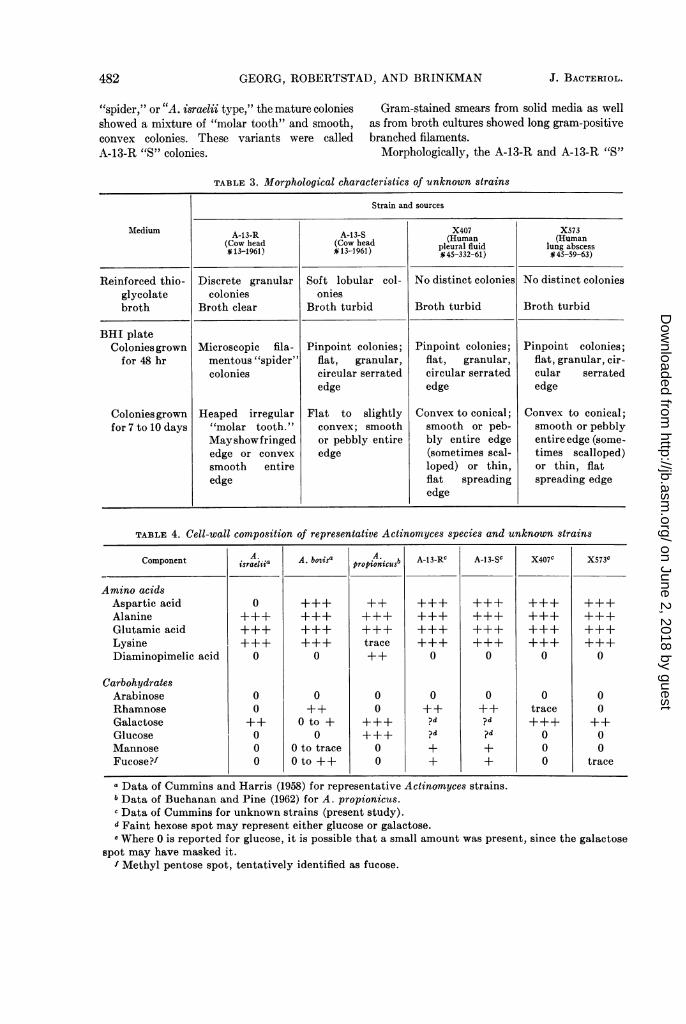
GEORG, ROBERTSTAD, AND BRINKMAN
"spider," or "A. israelii type," the mature coloniesshowed a mixture of "molar tooth" and smooth,convex colonies. These variants were calledA-13-R "S" colonies.
Gram-stained smears from solid media as wellas from broth cultures showed long gram-positivebranched filaments.
Morphologically, the A-13-R and A-13-R "S"
TABLE 3. Morphological characteristics of unknown strains
Strain and sources
Medium X407 X573A-13-R A-135S (Human (Human(Cow head (Cow head pleural fluid lung abscess# 13-1961) #13-1961) #45-332-61) #45-59-63)
Reinforced thio- Discrete granular Soft lobular col- No distinct colonies No distinct coloniesglycolate colonies oniesbroth Broth clear Broth turbid Broth turbid Broth turbid
BHI plateColonies grown Microscopic fila- Pinpoint colonies; Pinpoint colonies; Pinpoint colonies;
for 48 hr mentous "spider" flat, granular, flat, granular, flat, granular, cir-colonies circular serrated circular serrated cular serrated
edge edge edge
Coloniesgrown Heaped irregular Flat to slightly Convex to conical; Convex to conical;for 7 to 10 days "molar tooth." convex; smooth smooth or peb- smooth or pebbly
Mayshowfringed or pebbly entire bly entire edge entire edge (some-edge or convex edge (sometimes scal- times scalloped)smooth entire loped) or thin, or thin, flatedge flat spreading spreading edge
edge
TABLE 4. Cell-wall composition of representative Actinomyces species and unknown strains
Component A A. bovisa A b A-13-Rc A-13-Sc X407c X573cisraeliiapr io cu
Amino acidsAspartic acid 0 +++ ++ +++ +++ +++ +++Alanine +++ +++ +++ +++ +++ +++ +++Glutamic acid +++ +++ +++ +++ +++ +++ +++Lysine +++ +++ trace +++ +++ +++ +++Diaminopimelic acid 0 0 ++ 0 0 0 0
CarbohydratesArabinose 0 0 0 0 0 0 0Rhamnose 0 ++ 0 ++ ++ trace 0Galactose ++ 0 to + +++ ?d ?d +++ ++Glucose 0 0 +++ 0d0dO OMannose 0 O to trace 0 + + 0 0Fucose?f 0 0 to ++ 0 + + 0 trace
a Data of Cummins and Harris (1958) for representative Actinomyces strains.b Data of Buchanan and Pine (1962) for A. propionicus.c Data of Cummins for unknown strains (present study).d Faint hexose spot may represent either glucose or galactose.e Where 0 is reported for glucose, it is possible that a small amount was present, since the galactose
spot may have masked it.f Methyl pentose spot, tentatively identified as fucose.
482 J. BACTERIOL.
on June 2, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

IDENTIFICATION OF ACTINOMYCES
TABLE 5. Antigenic relationships*
Rabbit antisera
Soluble antigens CDC ATCC ATCC ATCC CDC ATCC Howell ATCCA-9 13683 10048 12102 A-601 12104 CS 1752 14157 CDC CDC CDC CDC(A~. (A. (A. (A. (A. (A. (A. (A. A-13-R A-13-S X407 X573
bovis) bovis) israelii) israelii) israelii) nlaes- faes- props-lundii) lundii) onicus)
CDC A-9 (A. bovis) + + 0 0 0 0 0 0 + + 0 0ATCC 13683 (A. bovis) + + 0 0 0 0 0 0 0 +. 0 0ATCC 10048 (A. is- 0 0 + + + 0 0 0 0 0 0 0
raelii)ATCC 12102 (A. is- 0 0 + + + 0 0 0 0 0 0 0
raelii)CDC A-601 (A. is- 0 0 + + + 0 0 0 0 0 0 0
raelii)ATCC 12104 (A. nae- 0 0 + 0 0 + + + 0 0 0 0
slundii)Howell CS 1752 (A. 0 0 + 0 0 + + + 0 0 0 0
naeslundii)ATCC 14157 (A. pro- 0 0 0 0 0 0 0 + 0 0 0 0
pionicus)CDC A-13-R 0 0 0 0 0 0 0 0 + 0 0 0CDC A-13-S + + 0 0 0 0 0 0 + + 0 0CDC X407 0 0 0 0 0 0 0 0 0 0 + +CDC X573 0 0 0 0 0 0 0 0 0 0 + +
* Determined from precipitin patterns in agar-gel; + = precipitin reaction; 0 = no reaction.
colonies resembled quite closely the three A.israelii control strains: 10048, 12102, and A601.The morphology of A-13-R is shown in Fig. 1 andis summarized in Table 3.The biochemical reactions of A-13-R are listed
in Table 2. They were the same whether theinoculum was taken from a typical "molar tooth"colony or from a convex, smooth-surfaced colony(A-13-R "S" colony). These reactions were nottypical of either the control A. bovis strains or thecontrol A. israelii strains. The inability to reducenitrate suggested A. bovis, while the inability tohydrolyze starch suggested A. israelii. A patternof acid production from carbohydrates appearedto be more consistent with that of A. bovis, as thisspecies has not been reported to ferment xylose orraffinose although about 80% of A. israelii strainsferment these two sugars.
Cell-wall analysis of A-13-R revealed a patternof amino acids and sugars that was compatiblewith that of the A. bovis strains studied by Cum-mins and Harris (1958). The principal aminoacids detected were aspartic acid, glutamic acid,alanine, and lysine. Sugars were detected corre-sponding in position to rhamnose, mannose,fucose, and glucose or galactose (Table 4).
Serological studies by the Ouchterlony tech-nique revealed an antigenic relationship withcontrol organism A. bovis (A-9). [Althoughno antigenic affinity was demonstrated againstthe second control A. bovis strain (ATCC 13683),A-13-R antiserum reacted with soluble antigensof two additional well-identified A. bovis strains,CDC A-8 and A-10, not included in this series.]A-9 antigen prxduced precipitin lines againstthe A-13-R antiserum. However, the reverserelationship was not shown i.e., A-13-R antigendid not react with A-9 antiserum, althoughit did react with its homologous antiserum(Table 5).The antigenic relationships between A-13-R
and A-13-S are discussed under A-13-S.Unknown strain A-13-S. Maximal growth was
obtained under anaerobic conditions; however,3+ growth was obtained under microaerophilicconditions (candle jar), and 1 + growth was ob-tained aerobically. Thus, A-13-S had a greater 02tolerance than did A-13-R.
Colonies grown for 48 hr on BHI plates werejust visible to the naked eye (pinpoint colonies).Under 100 X magnification, they appeared asflat, granular colonies with a central core of denser
VOL. 88, 1964 483
on June 2, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

GEORG, ROBERTSTAD, AND BRINKMAN
i0
.'_. ci
d
FIG. 1. Actinomyces bovis A-13-R rough "spider" variant. (a) Growth in thioglycolate broth; (b) Gram-stained smear, thioglycolate broth, 960 X; (c) 48-hr colony, BHI, 476 X; (d) 4-day-old colonies, BHI, 100X; (e, f, and g) mature colonies (7 to 10 days), BHI,'10 X.
'484 J. BAcTERIOL.
on June 2, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

IDENTIFICATION OF ACTINOMYCES
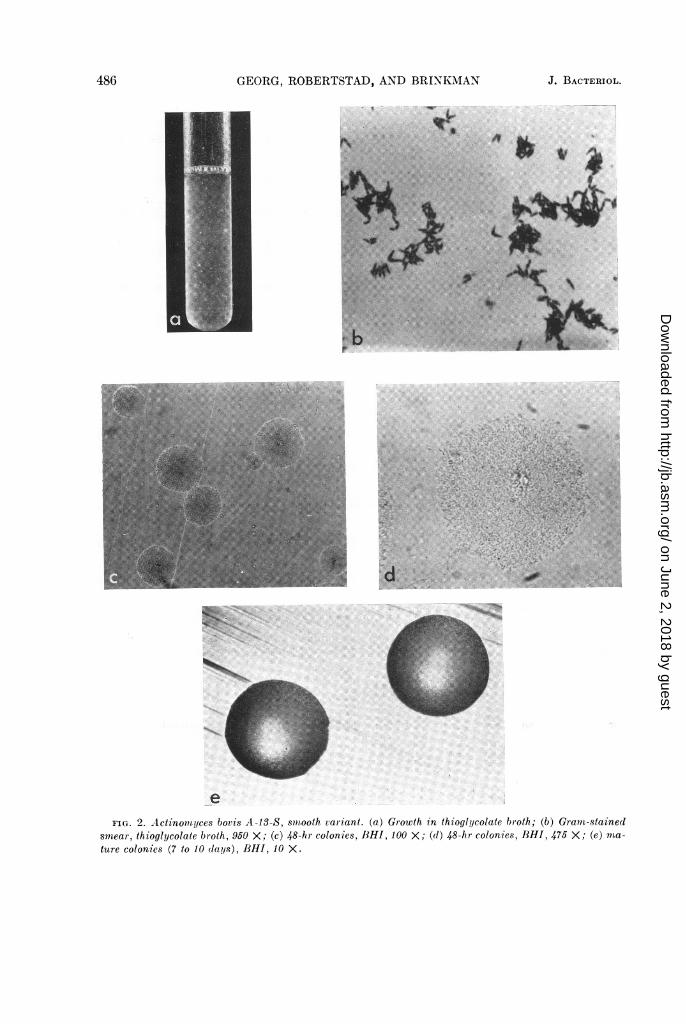
growth. The colony was circular, but the edge wasfinely serrated. No filamentous outgrowths wereobserved; 7- to 10-day-old colonies were dullwhite, slightly convex, with a smooth or slightlypebbly surface. The edges were entire and per-fectly smooth. Thus, both the microcolonies andthe mature colonies resembled the "S" or smoothA. bovis type described by Pine et al. (1960). Themicrocolonies and mature colonies remainedstable in morphology during 2 years of repeatedsubculture.
Cultures in RT broth consistently formed asoft cloudy growth throughout the lower four-fifths of the medium.
Gram-stained smears from solid media as wellas from broth cultures showed gram-positive shortdiphtheroid forms. These were usually thicker atone end, giving the organism a characteristic awlshape. These were frequently seen paired withthickened ends apposed. It was difficult to deter-mine true branching in young broth cultures;however, after 4 to 5 days in either RT or AMbroth, many long slender branched forms wereobserved among the short diphtheroid forms.
Morphologically, A-13-S resembled quiteclosely the two A. bovis control strains A-9 and13683. The morphology of A-13-S is illustrated inFig. 2 and summarized in Table 3.The biochemical reactions of A-13-S are listed
in Table 2. These reactions appeared similar tothose obtained for the two A. bovis control strains,and were typical for this species according to thestudies of Pine et al. (1960), except that fermenta-tion of xylose was not reported by these authors.However, in our experience, the fermentation ofthis sugar has been variable for several A. bovisstrains studied. The complete hydrolysis ofstarch by A-13-S was strikingly different fromresults with A-13-R. Hydrolysis of starch is acharacteristic that Pine et al. (1960) reported forall the strains of A. bovis which they had studied.
Cell-wall analysis of A-13-S (Table 4) wasidentical to that found for A-13-R. The pattern ofamino acids and carbohydrates was similar tothat usually encountered in A. bovis strains.
Serological studies by the Ouchterlony tech-nique revealed an antigenic affinity betweenA-13-S and A. bovis (Table 5). Both of the controlA. bovis strains, A-9 and 13683, reacted withA-13-S antiserum. The reverse reactions werepositive also. Common antigenic fractions couldbe demonstrated among these three strains; how-
ever, the antigens of A-9 and 13683 were notidentical to the A-13-S antigen. A-13-S antigenreacted with A-13-R antiserum, but did not reactwith A. israelii, A. naeslundii, or A. propionicusantisera.The antigenic relationships between A-13-R
and A-13-S antigens were interesting. A-13-Santigen reacted with both A-13-R and A-13-Santisera. A-13-R antigen, on the other hand, didnot react with the A-13-S antiserum. It reactedonly with its homologous antiserum, A-13-R(Table 5). It was demonstrated that A-13-Rshared an antigen related, but not identical, tothat found in A-13-S and A-9.Unknown strain X407. Growth occurred only
under anaerobic conditions (Table 2). Culturesin RT broth consistently showed diffuse growthcontaining soft lobular colonies (Table 3). Thegrowth was confined to the lower four-fifths of themedium. The broth became completely turbidupon gentle shaking. After 48 hr, growth on BHIplates was just visible to the naked eye as pin-point-sized glistening colonies. Under 100 X mag-nification, they appeared flat and granular withcentral cores of denser growth. The colonies werecircular, but their edges were finely serrated. Nofilamentous outgrowths were observed. After 7 to10 days of growth, colonies were dull white,slightly convex, with a smooth surface. The edgeswere entire and smooth. After 10 days, some ofthe larger colonies became irregularly conical andshowed irregular grooves from the edge of thecolony toward the center, giving a scalloped ap-pearance to the edge of the colony. Other coloniesdeveloped a pebbly surface and thin flat scallopedperipheral growth.Both the microcolonies and the mature colonies
resembled our strains of A. bovis.Gram-stained smears from solid media as well
as from broth cultures showed gram-positiveshort diphtheroid forms. Many showed clubbedor tapered ends. Few branched forms were ob-served, but long filaments were not formed. Theorganisms appeared very similar to A-13-S.Figure 3 illustrates the morphology of X407.The biochemical reactions of X407 (Table 2)
were not typical of any of the control cultures.The ability to hydrolyze starch completely andthe lack of ability to reduce nitrate suggested A.bovis; however, the sugar fermentation patternsuggested A. propionicus.
Cell-wall analysis of X407 revealed the follow-
VOL. 88, 1964 485
on June 2, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
GEORG, ROBERTSTAD, AND BRINKMAN
:: ^ i
:. w v s*: .:.: ,
X ... ....s: . ; .:^,, ::;:::
' i.' '.#*,^ ,t w. .__ ................ _r
:w ......... .: :. ^h._ j
s r... .... .'* .... ..... . .. ,,: JK ............ . . _ v.... :. ........... .... . ......... .. . , . 5E 5... ..
*, wWW b.... : ........... .. : : .: ...... . ..... . .. . . . .. . .. r ,,1 %
ss.
.. .. It, _.. .. ... .. .. . ... .. . ... . , . . .. .... . ^ .. i vc;-i .,wiE. .. , ... :.: - . i; ........... @.-_ * 4 + ;-&* * 4 ; Q ^t f_,. 5W. .< ; s45Lw. > Sz =,Ce*,w,wjis;t* :'' ' :' ,.,:#,',. , S'tw';,',8 ,- ., >
wR + ;. _s w.)*. .5ws fliF ..
x rs * *,, , t ., ri ffi ^ .....* , t * , , ' . +. , . . %s _ ** .:. .<; e,,,
t; a ^
.: . # sii '' ' .-1 'a' 'r.':. .. @
: ' + >..t
w a
.. I'.i-x. J! ,. u !. '..
ci
eFIG. 2. Actinomnyces boris A-13-S, smtooth variant. (a) Growth in thioglycolate broth; (b) Gramii-stained
smear, thioglycolate broth, 950 X; (c) 48-hr colonies, BHI, 100 X; (d) 48-hr colonies, BHI, 475 X; (e) ma-ture colonies (7 to 10 (lays), BHI, 10 X.
486 J. BACTERIOL.
on June 2, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

IDENTIFICATION OF ACTINOMYCES
d
51<N A
FIG. 3. Actinomyces (new species X407 and X573). (a) X407 and X573, growth in thioglycolate broth;(b) X407, Gram-stained smear, thioglycolate broth, 950 X; (c) X407, 48-hr colony, BHI, 100 X; (d) X407,mature colonies (7 to 10 days), BHI, 10 X; (e) X573, mature colonies (7 to 10 days), BHI, 10 X; (f)X407,mature colony (7 to 10 days), BHI, 10 X.
487VOL. 88, 1964
on June 2, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

GEORG(, ROBERTSTAD, AND) BRINKNIAN
TABLE 6. Suinmmary of characteristics of unknown strains
Strains and sources
Characteristics A-13-R A-13-S X407 X573(Cow head (Cow head (Human pleural fluid (Human lung abscess# 13-1961) # 13-1961) #45-332-61) § 45-59-63)
Morphological character-istics.................. Rough* Smooth Smooth Smooth
Biochemical character-istics....... Atypicalt Typical for A. Atypical Atypical
bovisCell-wall analysis........ Typical for A. Typical for A. Atypical Atypical
bovis bovisAntigenic affinity.... A. bovis A. bovis X573 X407Identification.... A. bovis A. bovis Actinomyces (new Actinomyces (new
species) species)
* "Rough" and "Smooth" are based on the characteristics of the microcolony.t Not typical of patterns of A. bovis, A. israelii, A. naeslhindii, or A. propionicus.
ing amnino acids: aspartic acid, glutamic acid,alanine, and lysine. The carbohydrates observedwere large amounts of galactose, and a trace ofrhamnose. Mannose and fucose were not detected.This l)attern appears to be distinctive from thatobserved by Cummiins and Harris for otherA ctinomtoyces species (Table 4).
Serological studies by the Ouchterlony tech-nique revealed no cross-reactions with any of thecontrol strains. Although X407 antigen did notreact with any of the control antisera, it didreact with its homologous antiserumln and alsowith X573 antiserum (Table 5).
Unknown strain X573. This strain apIeared tobe virtually identical to X407 in its morphology,oxygen requirements, and biochemical reactions.Cell-wall analysis revealed a lpattern of aminoacids and carbohydrates very similar, but notidentical, to that of X407 (Fig. 3; Tables 2, 3,and 4).
Serological studies revealed no antigenicaffinities with any of the control strains, but theX573 antigen reacted with the X407 antiserum aswell as with its homologous antiserum (Table 5).
'T'able 6 summarizes the findings on the four un-known strains. These are discusse(d below.
DiSCUSSIONA-13-R and A-13-S. On the primary isolation
plates from cow head no. 13, the different appear-ance of the two colony types wsas -ery strik-ing.Selection from the "R" and "S" colonies, followedby repeated streakings and picking of single col-onies, ga-ve evidence that these Nere distinct and
stable forms. The A-13-R strain regularly l)ro-duced filamentous "sl)ider" microcolonies (24 to48 hr) which developed into firm, heaped, roughcolonies (8 to 10 days). Both the microcolony andthe mature colony of A-13-R correslponded closelyto descriptions in the literature of A. israelii. TheA-13-S strain regularly produced flat, granularmicrocolonies that developed into convex, soft,smooth-surfaced colonies with entire edges. Boththe microcolonv and the mature colony corre-s)onded to descriptions of 4. botis. Therefore,the macroscopic morphology of A-13-R andA-13-S was suggestive of A. israelii and A. bovis,resp)ectively. This corresponds to the observationsof Pine et al. (1960) for P2R and P$.S.
Study of 02 requirements and biochemical re-actions further suggested identity with these twospecies. Both strains shared the important char-acteristics of the microaerophilic to anaerobicA4ctinornyces. The imlportant differences were thefinding that A-13-S had a greater 02 tolerancethan did A-13-R, and that it strongly hydrolyzedstarch although A-13-R utilized starch verylpoorly if at all. Neither strain reduced nitrate. Ingeneral, the 02 requirements and biochemical pat-tern of A-13-S w-ere characteristic of 'A. bovis asdescribed by Pine et al. (1960), and the findingson A-13-R were in general characteristic of A.israelii.However, the finding (Thompson, 1950) of
rough strains of A. bovis (1 of 11 strains of A.bovis studied), and the finding of a "spider" var-iant (P2R strain) of A. bovis by l'ine et al. (1960),coulpledl with the variability in the biochemical
488 J. BACTERIOL.
on June 2, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

IDENTIFICATION OF ACTINOMYCES
reactions of a number of human and bovine iso-lates we are currently studying, led us to searchfor further criteria before concluding that thesewere A. israelii and A. bovis.
Cell-wall studies performed for us by C. S.Cummins indicated a similar pattern of aminoacids and sugars for both A-13-R and A-13-S. Thepattern found was typical of those found previ-ously by Cummins and Harris (1958) and Cum-mins (1962) for A. bovis. The presence of asparticacid and rhamnose and other methyl sugars inboth strains ruled out the possibility that eithercould be A. israelii. Essentially identical resultswere found previously by Cummins for strainsP2R and P2S as described by Pine et al. (1960).
Results of agar-gel precipitin tests indicatedthat A-13-S represented A. bovis. The A-13-Rantigen deviated from the expected pattern bynot reacting with any control A. bovis antisera;however, the A-13-R antisera reacted with A-13-Santigen and the antigens of three authenticatedA. bovis isolates (A-8, A-9, and A-10). It did notreact with 13683 antigen.The study of A-13-R and A-13-S emphasizes
the variability in morphology, oxygen require-ments, and biochemical reactions in A. bovts. Thefinding of these two distinct morphological typeswithin the species perhaps explains previous disa-greement of workers regarding the causal agentof bovine actinomycosis and their reluctance toaccept A. israelii as a separate species. It em-phasizes the potential danger of reliance uponmorphology for species identification within thetwo species, A. bovis and A. israelii. Finally, itpoints out a second morphological variance in thespecies A. bovis: (i) smooth to rough colonialvariation, (ii) smooth and "spider" colonialvariants reflecting diphtheroid and mycelial ele-ments, respectively.X407 and X573. There appeared to be little
doubt that X407 and X573 represented the samespecies. Morphologically they resembled A. bovisrather closely, and at first it was suspected thatwe might have evidence for the first authenticatedisolations of A. bovis from human materials. How-ever, both strains were less oxygen-tolerant thanwere the A. bovis strains we had studied previ-ously, and although both strains hydrolyzedstarch strongly, which is a characteristic of A.bovis, the biochemical pattern in general was dis-tinct from A. bovis.
Cell-wall analyses of these strains revealed adistinctive pattern of amino acids and sugars that
had not been observed in other Actinomycesspecies. Serological studies showed no antigenicrelationships between these strains and the con-trol strains studied. It was concluded that X407and X573 represented an actinomyces speciesthat was different from all the previously de-scribed species.These two strains and several other similar
strains being studied currently will be describedas a new Actinomyces species in a future publica-tion.
ACKNOWLEDGMENT
The writers acknowledge the photographicwork by John P. Gust, Jr., of the CommunicableDisease Center's Medical Audiovisual Branch.
LITERATURE CITED
BREED, R. S., E. G. D. MURRAY, AND N. R. SMITH.1957. Bergey's manual of determinative bac-teriology, 7th ed., p. 724-744. The Williams &Wilkins Co., Baltimore.
BUCHANAN, B. B., AND L. PINE. 1962. Characteri-zation of a propionic acid producing Actino-mycete, Actinonmyces propionicus, Sp. nov. J.Gen. Microbiol. 28:305-323.
CUMMINS, C. S. 1962. Chemical composition andantigenic structure of cell walls of Corynebac-terium, Mycobacterium, Nocardia, Actinomycesand Arthrobacter. J. Gen. Microbiol. 28:35-50.
CUMMINS, C. S., AND H. HARRIS. 1958. Studies onthe cell-wall composition and taxonomy ofActinomycetales and related groups. J. Gen.Microbiol. 18:173-189.
ERIKSON, D. 1940. Pathogenic anaerobic microor-ganisms of the Actinomyces group. Med. Res.Council, Spec. Rept. Ser. 240:1-63.
ERIKSON, D. 1949. The morphology, cytology, andtaxonomy of the actinomycetes. Ann. Rev.Microbiol. 3:23-54.
HARZ, C. 0. In 0. Bollinger. 1877. Uber eine neuePilzkrankheit beim Rinde. Zentr. Med. Wiss.15:481-485.
HOWELL, A., JR., W. C. MURPHY, III, F. Paul, ANDR. M. STEPHAN. 1959. Oral strains of Actino-myces. J. Bacteriol. 78:82-95.
HOWELL, A., JR., AND L. PINE. 1956. Studies on thegrowth of species of Actinomyces. I. Cultiva-tion in a synthetic medium with starch. J.Bacteriol. 71:47-53.
HOWELL, A., JR., R. M. STEPHAN, AND F. PAUL.1962. Prevalence of Actinomyces israelii, A.naeslundii, Bacterionema matruchotii and Can-dida albicans in selected areas of the oralcavity and saliva. J. Dental Res. 41:1050-1059.
VOL. 88, 1964 489
on June 2, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

GEORG, ROBERTSTAD, AND BRINKMAN
KING, S., AND E. MEYER. 1963. Gel diffusion tech-nique in antigen-antibody reactions of Actino-myces species and "anaerobic diphtheroids."J. Bacteriol. 85:186-190.
OUCHTERLONY, 0. 1949. Antigen-antibody reac-
tions in gels. Acta Pathol. Microbiol. Scand.26:507-515.
OVERMAN, J. R., AND L. PINE. 1963. Electron mi-croscopy of cytoplasnmic structures in faculta-tive and anaerobic Actinomyces. J. Bacteriol.86:656-665.
PINE, L., AND H. HARDIN. 1959. Actinomyces isra-elii, a cause of lacrimal canaliculitis in man.
J. Bacteriol. 78:164-170.PINE, L., A. HOWELL, JR., AND S. J. WATSON. 1960.
Studies on the morphological, physiological,and biochemical characteristics of Actinomy-ces bovis. J. Gen. Microbiol. 23:403-424.
SCHUBERT, J. H., H. J. LYNCH, ANI) L. AJELLO.1961. Evaluation of the agar plate precipitintest for histoplasmosis. Am. Rev. Respirat.Diseases 84:845-849.
THOMPSON, L. 1950. Isolation and comparison ofActinomyces from human and bovine infec-tions. Proc. Staff Meetings Mayo Clinic25:81-86.
THOMPSON, L., AND S. A. LOVESTEDT. 1951. AnActinomyces-like organism obtained from thehuman mouth. Proc. Staff Meetings MayoClinic 26:169-175.
490 J. BACTERIOL.
on June 2, 2018 by guesthttp://jb.asm
.org/D
ownloaded from


















