
I. PENDAHULUAN Latar Belakang - repository.ipb.ac.id · Studi persebaran serbuk sari dapat...
119
I. PENDAHULUAN Latar Belakang Kelapa (Cocos nucifera L.) merupakan salah satu tanaman penghasil minyak yang paling penting di daerah tropis. Peranan kelapa sebagai komoditi perkebunan bagi masyarakat Indonesia dan negara sangat besar. Produksi kelapa selama bertahun-tahun menunjukkan penurunan bertahap karena berbagai alasan, tetapi masih bernilai ekonomi penting dengan adanya permintaan industri yang tinggi untuk minyak laurat. Agar produksi kelapa tidak menurun, maka pelaksanaan peremajaan dan rehabilitasi harus dilakukan pada sekitar 20-30% pertanaman kelapa (Prastowo 2007). Perbaikannya dilakukan melalui pemuliaan konvensional dan bioteknologi. Jenis kelapa di Indonesia sangat variatif diantaranya adalah kelapa kopyor yang merupakan kelapa mutan asli Indonesia dan berbeda fenotipenya dengan kelapa Makapuno yang berasal dari Filipina. Keberadaan kelapa kopyor yang unik dan asli Indonesia perlu terus dilestarikan dan dikembangkan lebih lanjut agar sumberdaya genetik asli Indonesia tersebut dapat dimanfaatkan sepenuhnya untuk kesejahteraan rakyat Indonesia. Berbeda dengan kelapa kopyor Indonesia, abnormalitas endosperma pada kelapa Makapuno menyebabkan jaringan ini menjadi lunak seperti jeli dan jika terlalu tua sebagian dari endospermanya akan terlarut dalam air kelapa, sehingga air kelapanya menjadi kental seperti minyak pelumas (Gambar 1.1). Perbedaan utama antara abnormalitas endosperma kelapa kopyor dan kelapa makapuno adalah pada kelapa kopyor endospermanya tetap mempunyai penampakan seperti endosperma kelapa tetapi terlepas dari cangkangnya (Gambar 1.1), rasa air kelapa dan endospermanya lebih manis dari kelapa normal dan tekstur endospermanya lembut seperti tekstur gabus (Maskromo 2005). Semakin tua buah kelapanya umumnya air kelapanya semakin berkurang dan volume endosperma yang terlepas serta mengumpul dalam rongga dalam cangkang biji kelapanya semakin banyak (Maskromo et al. 2007). Gambar 1.1 Variasi endosperma pada kelapa. Perbandingan endosperma abnormal pada (a) kelapa kopyor asal Indonesia, (b) kelapa Makapuno asal Filipina dan (c) endosperma normal pada kelapa normal (Sudarsono et al. 2014a) a b c
-
Upload
vuongquynh -
Category
Documents
-
view
236 -
download
0
Transcript of I. PENDAHULUAN Latar Belakang - repository.ipb.ac.id · Studi persebaran serbuk sari dapat...

I. PENDAHULUAN
Latar Belakang
Kelapa (Cocos nucifera L.) merupakan salah satu tanaman penghasil
minyak yang paling penting di daerah tropis. Peranan kelapa sebagai komoditi
perkebunan bagi masyarakat Indonesia dan negara sangat besar. Produksi kelapa
selama bertahun-tahun menunjukkan penurunan bertahap karena berbagai alasan,
tetapi masih bernilai ekonomi penting dengan adanya permintaan industri yang
tinggi untuk minyak laurat. Agar produksi kelapa tidak menurun, maka
pelaksanaan peremajaan dan rehabilitasi harus dilakukan pada sekitar 20-30%
pertanaman kelapa (Prastowo 2007). Perbaikannya dilakukan melalui pemuliaan
konvensional dan bioteknologi. Jenis kelapa di Indonesia sangat variatif
diantaranya adalah kelapa kopyor yang merupakan kelapa mutan asli Indonesia
dan berbeda fenotipenya dengan kelapa Makapuno yang berasal dari Filipina.
Keberadaan kelapa kopyor yang unik dan asli Indonesia perlu terus
dilestarikan dan dikembangkan lebih lanjut agar sumberdaya genetik asli
Indonesia tersebut dapat dimanfaatkan sepenuhnya untuk kesejahteraan rakyat
Indonesia. Berbeda dengan kelapa kopyor Indonesia, abnormalitas endosperma
pada kelapa Makapuno menyebabkan jaringan ini menjadi lunak seperti jeli dan
jika terlalu tua sebagian dari endospermanya akan terlarut dalam air kelapa,
sehingga air kelapanya menjadi kental seperti minyak pelumas (Gambar 1.1).
Perbedaan utama antara abnormalitas endosperma kelapa kopyor dan kelapa
makapuno adalah pada kelapa kopyor endospermanya tetap mempunyai
penampakan seperti endosperma kelapa tetapi terlepas dari cangkangnya (Gambar
1.1), rasa air kelapa dan endospermanya lebih manis dari kelapa normal dan
tekstur endospermanya lembut seperti tekstur gabus (Maskromo 2005). Semakin
tua buah kelapanya umumnya air kelapanya semakin berkurang dan volume
endosperma yang terlepas serta mengumpul dalam rongga dalam cangkang biji
kelapanya semakin banyak (Maskromo et al. 2007).
Gambar 1.1 Variasi endosperma pada kelapa. Perbandingan endosperma abnormal
pada (a) kelapa kopyor asal Indonesia, (b) kelapa Makapuno asal
Filipina dan (c) endosperma normal pada kelapa normal (Sudarsono
et al. 2014a)
a b c

2
Makapuno tidak memiliki aktivitas galaktosidase yang memungkinkan
mengubah struktur dinding sel dan adhesi sehingga menghasilkan endosperm
yang sangat kental (Luengwilai 2014). Besar kemungkinan bahwa abnormalitas
fenotipe endosperma kelapa kopyor juga diduga juga sebagai akibat dari defisiensi
enzim penting tertentu selama dalam proses perkembangan endospermanya.
Namun demikian, identitas enzim yang defisien dari endosperma kelapa kopyor
sampai saat ini masih belum diketahui. Karakteristik mutan pada kelapa kopyor
juga dapat diturunkan secara genetik dari tetua ke progeninya (Sukendah 2009).
Salah satu permasalahan yang dihadapi oleh petani kopyor adalah
rendahnya kuantitas hasil buah kopyor yang dipanen. Akibatnya produksi buah
kelapa kopyor masih belum dapat memenuhi kebutuhan konsumen. Adanya
kelapa normal di antara pertanaman kelapa kopyor diduga mempengaruhi
produktivitas buah kopyor yang didapat (Sudarsono et al. 2012). Keberadaan
pohon dewasa kelapa Dalam berbuah normal cenderung menyerbuk silang
(Pandin 2009a) diduga berpengaruh negatif terhadap produksi buah kopyor.
Pohon tersebut dapat menyebarkan serbuk sari pembawa sifat normal pada bunga
betina pohon kelapa kopyor, akibatnya buah yang terbentuk dari penyerbukan
akan menjadi buah normal (Sudarsono et al. 2012). Keberhasilan penyerbukan
pada tanaman juga memerlukan bantuan polinator untuk persebaran serbuk sari
tanaman kelapa maupun pada tanaman lainnya.
Studi evaluasi persebaran serbuk sari pada pertanaman kelapa kopyor
dianggap penting untuk dilakukan untuk membuktikan dan memberikan informasi
kepada petani mengenai jarak dan arah persebaran serbuk sari serta faktor yang
berpengaruh pada penyerbukan kelapa kopyor. Studi persebaran serbuk sari dapat
dipelajari dengan metode pewarnaan serbuk sari (Blair dan Williamson 2010) atau
menggunakan marka molekular (Austerlitz et al. 2004). Marka yang biasa
digunakan dalam analisis persebaran serbuk sari adalah marka RAPD pada Ilex
paraguariensis (Cansian et al. 2010) dan marka SSR pada tanaman Hymenaea
courbaril (Carneiro et al. 2011), tanaman pinus (Feng et al. 2010). Marka yang
sering digunakan adalah marka SSR (Single Sequence Repeat) yang mempunyai
keunggulan yaitu bersifat kodominan, polimorfismenya tinggi, lokus tersebar di
dalam genom dalam jumlah banyak (Lowe et al. 2004) dan sampel DNA yang
dibutuhkan sedikit karena dalam melakukan deteksi menggunakan PCR
(Polymerase chain reaction) yang dapat menggandakan DNA (Semagn et al.
2006). Penanda DNA berbasis SNAP adalah satu-satunya penanda DNA yang
memiliki sifat bi–alel dan kodominan, sehingga penanda SNAP mampu
membedakan alel homozigot dari heterozigot yang efisien (Morin et al. 2004).
Beberapa marker SSR dan marker SNAP sedang dikembangkan oleh PMB
Laboratorium dari kegiatan penelitian awal namun demikian penelitian lebih
lanjut masih sangat diperlukan.
Penelitian persebaran serbuk sari pada kelapa sampai saat ini belum ada
dilaporkan sehingga dianggap penting untuk mempelajari persebaran serbuk
sari pada kelapa kopyor. Hasil yang diharapkan adalah memberikan informasi
mengenai tipe persebaran serbuk sari dan besarnya persentase penyerbukan
silang ataupun penyerbukan sendiri pada pertanaman kelapa, efek keberadaan
pohon kelapa berbuah normal di dalam pertanaman kelapa berbuah kopyor,
efek polinator terhadap produksi buah kopyor sehingga diharapkan
produktivitas buah kelapa kopyor dapat ditingkatkan, selain itu juga

3
memberikan informasi dasar genomik dan molekuler yang akan sangat berguna
dalam mendukung program pemuliaan kelapa kopyor di masa yang akan
datang.
Tujuan Penelitian
1. Mengetahui besarnya persentase penyerbukan silang dan penyerbukan sendiri
yang terjadi pada populasi pertanaman kelapa kopyor tipe Genjah 2. Mengetahui besarnya persentase penyerbukan silang dan penyerbukan sendiri
yang terjadi pada populasi pertanaman kelapa kopyor tipe Dalam 3. Mengetahui peranan pendonor serbuk sari kelapa normal dalam pembentukan
progeni kelapa kopyor (efek xenia pada tanaman)
4. Mengetahui kemampuan dan efektifitas polinator lebah madu sebagai agen
penyerbuk dominan
Manfaat Penelitian
Dengan diperolehnya informasi mengenai pola persebaran dan agen
pernyerbuk yang dominan diharapkan akan memberikan informasi kepada petani
mengenai efek keberadaan pohon kelapa berbuah normal di dalam pertanaman
kelapa berbuah kopyor, efek polinator terhadap produksi buah kopyor sehingga
diharapkan produktivitas buah kelapa kopyor dapat ditingkatkan, selain itu juga
memberikan informasi dasar genomik dan molekuler yang akan sangat berguna
dalam mendukung program pemuliaan kelapa kopyor di masa yang akan datang.
Kerangka Penelitian
Pemanfaatan marka molekuler dalam mendeteksi dan membantu analisis
persebaran serbuk sari dalam populasi kelapa kopyor diharapkan mampu memberi
efek positif dalam peningkatan produktifitas hasil bagi petani. Dengan
pertimbangan tersebut maka dilakukan penelitian untuk melihat berbagai
pengaruh persebaran serbuk sari terhadap berbagai studi penelitian.
Kegiatan yang dilakukan dalam penelitian ini memiliki ruang lingkup
yang saling berkaitan untuk mencapai tujuan yang diharapkan yang terlihat pada
Gambar 1.3. Tahapan-tahapan yang dilakukan dalam penelitian ini adalah sebagai
berikut: Percobaan 1) Persebaran serbuk sari pada kelapa kopyor Genjah Pati
mengindikasikan pentingnya peranan polinator serangga dalam penyerbukan
Percobaan 2) Persebaran serbuk sari pada kelapa Dalam Kalianda membuktikan
adanya penyerbukan silang pada kelapa Dalam Percobaan 3) Persebaran serbuk
sari pada kelapa Dalam Dukuh Seti membuktikan pengaruh negatif kelapa normal
terhadap hasil buah kopyor Percobaan 4) Lebah sebagai polinator merubah pola
persebaran serbuk sari pada kelapa kopyor Pati. Informasi dari persebaran serbuk
sari pada pertanaman kelapa secara umum dapat dimanfaatkan untuk tujuan
pemuliaan tanaman kelapa di masa mendatang dan rekomendasi kebun bibit
kelapa yang efektif.

4
Bagan Alir Penelitian
Gambar 1.2. Bagan alir penelitian disertasi Analisis Persebaran Serbuk Sari Kelapa Kopyor (Cocos nucifera L.) Asal Pati dan
Kalianda Menggunakan Marka SSR dan SNAP Sebagai Penunjang Program Pemuliaan Tanaman
Plasma nutfah kelapa
kopyor Indonesia
( Pati dan Lampung)
Karakteristik
produktifitas kelapa
kopyor
Analisis persebaran
serbuk sari kelapa
kopyor
Percobaan 1
Persebaran serbuk sari pada kelapa kopyor
Genjah Pati mengindikasikan pentingnya
peranan polinator serangga dalam
penyerbukan
Percobaan 2
Persebaran serbuk sari pada kelapa Dalam
Kalianda membuktikan adanya
penyerbukan silang pada kelapa dalam
Percobaan 4
Lebah sebagai polinator merubah pola
persebaran serbuk sari pada kelapa
kopyor Pati
Data skoring tetua dan
progeni setiap populasi
Analisis parental
menggunakan CERVUS
Percobaan 3
Persebaran serbuk sari membuktikan
potensi pengaruh negatif kelapa normal
terhadap hasil buah kopyor
1. Mengetahui besarnya persentase penyerbukan
silang dan penyerbukan sendiri yang terjadi pada
populasi pertanaman kelapa kopyor tipe Genjah 2. Mengetahui besarnya persentase penyerbukan
sendiri dan penyerbukan silang yang terjadi pada
populasi pertanaman kelapa kopyor tipe Dalam 3. Mengetahui peranan pendonor serbuk sari
kelapa normal dalam pembentukan progeni
kelapa kopyor (efek xenia pada tanaman)
4. Mengetahui kemampuan dan efektifitas
polinator lebah madu sebagai agen penyerbuk
dominan

5
II. TINJAUAN PUSTAKA
Tanaman Kelapa
Tanaman kelapa (Cocos nucifera L.) adalah satu satunya spesies dari cocos
yang merupakan anggota dari subfamili Cocoideae dan famili Aracaceae
(Palmaceae). Kelapa merupakan tanaman diploid yang memiliki jumlah
kromosom 32 (2n=2×=32). Tanaman ini merupakan tanaman tahunan (perenial)
yang bersifat monoecious yaitu memiliki bunga jantan dan betina pada tandan atau
infloresensia yang sama (Chan dan Elevitch 2006).
Ciri-ciri pohon kelapa menurut Chan dan Elevitch (2006) adalah memiliki
batang tunggal dan beruas dengan tinggi mencapai 30 m dan diameter kanopi 8-9
m. Akar berbentuk serabut, tebal, berkayu dan adaptif pada lahan berpasir pantai.
Daun tersusun secara majemuk dan menyirip sejajar tunggal, pelepah terletak pada
ibu tangkai daun, duduk pada batang (roset batang). Warna pada tangkai daun
(petiole) mengindikasikan warna buah pada kelapa. Bunga kelapa merupakan bunga
majemuk yang dilindungi oleh spatha. Bunga jantan dan betina terdapat pada satu
tangkai utama yang disebut spadix, setiap spadix terdiri atas 40-60 cabang (spikelet)
dengan ribuan bunga jantan. Letak bunga bunga betina terletak di pangkal,
sedangkan bunga jantan di bagian atas bunga betina hingga ujung spikelet. Buah
kelapa memiliki tiga lapisan yaitu eksokarp (kulit tipis terluar yang memiliki
lapisan lilin) berwarna kuning, hijau, jingga atau coklat, mesokarp berupa lapisan
serat yang lebih tebal atau sering disebut sabut dan endokarp yang keras disebut
batok yang melindungi biji. Endokarp dan biji hanya dipisahkan oleh membran
yang melekat pada sisi dalam dari endokarp. Biji kelapa memiliki tiga mikrofil
(micropyle) dan hanya satu yang mengindikasikan keberadaan embrio. Embrio
kelapa berukuran kecil dan akan membesar ketika buah siap untuk berkecambah.
Endosperma biji kelapa terdiri atas endosperma cair yang mengandung banyak
enzim dan endosperma padat yang mengendap pada dinding endokarp ketika buah
menua (kernel).
Secara umum tanaman kelapa dibedakan atas dua tipe yaitu tipe Dalam
(typica) dan tipe Genjah (nana) (Kumar et al. 2011). Penggolongan kedua tipe ini
terutama didasarkan atas sifat munculnya pembungaan pertama, tinggi tanaman,
komponen buah dan tipe penyerbukan. Kelapa Dalam mempunyai tinggi sekitar
15-18 m, batang kekar dengan dasar membengkak atau disebut bole. Bunga
pertama muncul pada umur 6-10 tahun setelah tanam tetapi umur produktif dapat
mencapai 90 tahun. Mahkota pohon memiliki 25-40 daun yang terbuka penuh,
dengan panjang daun sekitar 5-7 m. Umumnya kelapa Dalam menyerbuk silang
dan dari penyerbukan sampai buah masak memerlukan waktu sekitar 12 bulan
dengan jumlah buah pertandan 6-12 butir. Ukuran buah besar sehingga produksi
kopra, minyak dan sabut umumnya berkualitas baik.
Pohon kelapa tipe Genjah berpenampilan pendek sekitar 8-10 m saat
berumur 20 tahun dengan batang agak kecil dan tanpa bole. Daunnya terbuka
penuh dengan panjang ≤ 4 m. Mulai berbunga umur 3-4 tahun setelah tanam tetapi
pembungaannya tidak teratur. Umumnya kelapa genjah menyerbuk sendiri,
dengan waktu yang diperlukan dari penyerbukan sampai buah masak 11-12 bulan.
Produksi buah sekitar 10-30 butir pertandan dengan ukuran buah kecil sehingga

6
kualitas buah dan kopranya kurang baik. Produksi akan mulai menurun setelah
tanaman berumur 25 tahun (Chan dan Elevitch 2006).
Penyerbukan atau polinasi adalah jatuhnya serbuk sari dari kotak sari
(antera) ke kepala putik (stigma) dalam satu bunga atau bunga yang berbeda.
Penyerbukan tumbuhan dapat terjadi secara biotik dan abiotik. Penyerbukan biotik
terjadi dengan bantuan hewan, sedangkan penyerbukan abiotik terjadi dengan
bantuan angin, air dan gravitasi (Liferdi 2008). Jarak persebaran serbuk sari pada
tanaman yang menyerbuk sendiri (autogamy) lebih rendah dibandingkan dengan
tanaman menyerbuk silang (Boer 2007).
Kelapa Dalam pada umumnya merupakan tanaman menyerbuk silang
sehingga tampilannya sangat beragam (Pandin 2009b). Kelapa Dalam memiliki
bunga jantan yang matang lebih dulu dibanding bunga betina. Bunga betina siap
diserbuki ketika bunga jantan umumnya sudah rontok sehinga terjadi penyerbukan
silang. Kelapa Genjah pada umumnya memiliki pola penyerbukan sendiri
meskipun memungkinkan terjadinya penyerbukan silang sehingga menyebabkan
tingginya tingkat kemiripan genetik pada kelapa Genjah. Bunga betina dan bunga
jantan pada kelapa Genjah masak secara bersamaan sehingga peluang untuk
menyerbuk sendiri sangat besar (Hannum et al. 2003).
Penelitian Ramirez et al. (2004) menyatakan sebanyak 59% penyerbukan
kelapa dibantu oleh serangga lebah madu. Lebah membantu proses penyerbukan
silang, sehingga meningkatkan produktivitas tanaman budidaya. Potensi ini
dimanfaatkan dengan cara meletakkan koloni lebah pada areal tanaman budidaya
yang daya serbuknya rendah. Perpindahan lebah dari satu bunga ke bunga yang
lain mempercepat proses polinasi karena serbuk sari banyak menempel pada kaki
dan perut dari lebah (Liferdi 2008).
Kelapa Kopyor
Kelapa berbuah kopyor dari segi morfologi sama dengan tanaman kelapa
lainnya. Maskromo et al. (2007) mengatakan buah kelapa kopyor hanya bisa
dipastikan setelah buah dipanen dengan cara mengguncang buah kelapanya. Pada
saat diguncang kelapa kopyor akan menghasilkan bunyi yang kurang nyaring
dibanding kelapa normal, karena sebagian atau seluruh endosperma fase padatnya
sudah lepas dari tempurungnya. Buah kopyor juga dapat diidentifikasi dengan
ketukan, tetapi memerlukan keterampilan khusus untuk dapat melakukannya.
Tukang ketuk kelapa yang sudah ahli dalam identifikasi buah kopyor disebut
“tukang totok”. Tingkat akurasi penentuan buah kopyornya dapat mencapai 99%
(Sudarsono et al. 2014a). Buah dengan sifat kopyor dihasilkan dari pohon kelapa
tertentu yang sebagian besar buahnya mempunyai endosperma normal dan
sebagian kecil abnormal (kopyor). Pohon kelapa kopyor hanya mempunyai buah
kelapa kopyor dengan frekuensi antara 3-4 buah kopyor per tandan.
Abnormalitas fenotipe endosperma kelapa Kopyor diduga juga sebagai
akibat dari defisiensi enzim penting tertentu selama dalam proses perkembangan
endospermanya. Namun demikian, identitas enzim yang mengalami defisien dari
endosperma kelapa Kopyor sampai saat ini masih belum diketahui. Karakteristik
mutan pada kelapa Kopyor juga dapat diturunkan secara genetik dari tetua ke
progeninya (Sukendah 2009). Berdasarkan hasil penelitian Maskromo (2005)
yang membedakan adalah bagian endospermanya seperti pada Gambar 2.1.

7
Gambar 2.1 Perbedaan fenotipe kelapa normal (kiri) dan kelapa kopyor (kanan)
Melalui serangkaian penelitian yang dilakukan sejak tahun 2005, pada tahun
2010 Balai Penelitian Tanaman Kelapa dan Palma Lain Manado telah berhasil
melepas tiga varietas unggul lokal kelapa Genjah kopyor asal Pati, Jawa Tengah
dengan potensi persentase rata-rata buah kopyor sekitar 40% pertandannya.
Potensi produksi buah kopyor tersebut masih berpeluang untuk ditingkatkan
dengan pemuliaan tanaman melalui penyerbukan silang buatan dengan tetua yang
memiliki persentase buah kopyor tinggi yaitu dengan serbuk sari dari tanaman
hasil kultur embryo yang menghasilkan buah kopyor mencapai 100% (Novarianto
dan Miftahorrachman 2000).
Kelapa kopyor ditemukan baik pada kelapa tipe Dalam maupun tipe Genjah,
dengan perbanyakan melalui kultur embrio dan menggunakan bibit alami. Pada
pengembangan menggunakan bibit alami, kelapa kopyor tipe Dalam hanya
menghasilkan buah kopyor antara 10 – 20 %, sedangkan tipe Genjah berpotensi
mencapai di atas 50 % pertandannya, dengan tingkat produksi yang beragam. Ini
terkait dengan pola pembungaan masing-masing tipe kelapa tersebut. Kelapa tipe
Dalam memiliki pola penyerbukan silang, sedangkan tipe Genjah menyerbuk
sendiri. Hal tersebut menyebabkan masih rendahnya jumlah buah kopyor yang
diperoleh petani, dan berdampak pada rendahnya produksi buah kopyor.
Penanda Genetik
Konservasi dan penggunaan sumber genetik tanaman sangat penting dalam
rangka produksi tanaman pertanian dan perkebunan serta pemeliharaan tanaman
secara berkesinambungan. Oleh karena itu plasma nutfah merupakan sumber
genetik tanaman yang perlu mendapat perhatian, tidak hanya pada tahap
pengumpulan dan pemeliharaan tetapi juga bagaimana mengkarakterisasi
keanekaragaman genetiknya, mengevaluasi sifat sifat yang dikehendaki dan
memanfaatkan untuk pemuliaan tanaman.
Penggunaan penanda sebagai alat karakterisasi sangat diperlukan untuk
pengkarakteran tanaman secara genetik. Penanda dapat dikategorikan sebagai
penanda morfologi, sitologi dan perkembangan terakhir adalah penenda
molekuler. Penanda yang banyak dilakukan adalah penanda morfologi, yaitu
dengan mengamati secara langsung karakter morfologi tanaman, namun penanda

8
tersebut mempunyai kelemahan karena karakter yang diamati kemungkinan
dipengaruhi oleh lingkungan. Suatu metode karakterisasi yang dikenal dengan
nama penanda molekuler telah dikembangkan untuk pengulangan keterbatasan
penanda morfologi.
Pemecahan kendala dalam pemuliaan konvensional mulai mendapat titik
terang dengan ditemukannya marka molekuler. Marka molekuler yang pertama
kali dikenal adalah marka protein yang secara genetik dikenal sebagai isozim
(Amar et al. 2011). Meskipun marka telah banyak digunakan dalam analisis
genetik tanaman namun dalam perkembangannya marka isozim masih sangat
terbatas jumlahnya. Beberapa sistem enzim tertentu dipengaruhi oleh regulasi
perkembangan jaringan, yaitu hanya mengekspresikan suatu sifat pada jaringan
tertentu dan pada stadia pertumbuhan tanaman. Kedua faktor tersebut merupakan
kendala utama pengunaan marka isozim dalam mengeksploitasi potensi genetik
tanaman (Mondini et al. 2009).
Semakin berkembangnya ilmu pengetahuan, maka pada awal tahun 1980-an
ditemukan teknologi molekuler yang berbasis pada DNA. Marka molekuler
tersebut dapat menutpi kekurangan dari marka isozim, karena jumlah yang tidak
terbatas dan dapat melingkupi seluruh genom tanaman, tidak dipengaruhi oleh
regulasi perkembangan jaringan, sehingga dapat dideteksi pada seluruh jaringan,
dan memiliki kemampuan yang sangat tinggi dalam menganalisis keragaman
karakter antar individu. Seiring dengan semakin berkembangnya teknologi yang
berbasis marka DNA, maka saat ini telah ditemukan tiga tipe marka DNA dengan
segala kelebihan dan kekurangan masing-masing. Semagn et al (2006)
menyatakan bahwa penanda molekuler secara garis besar dibagi menjadi tiga
kelompok berdasarkan metode deteksinya, yaitu (i) marka berbasis hibridisasi
seperti RFLP, (ii) marka berbasis PCR seperti RAPD, AFLP, ISSR, SSR, dan (iii)
marka berbasis sekuens DNA seperti SNP.
Teknologi marka DNA berdasarkan teknik PCR dapat bersifat spesifik atau
acak sesuai dengan tipe primer yang digunakan (Gupta et al. 2002). Marka DNA
hasil amplifikasi primer spesifik adalah marka yang bersifat kodominan.
Pengembangan marka-marka kodominan membutuhkan informasi sekuen dari
DNA target yang digunakan untuk merancang primer spesifik, umumnya
memiliki ukuran panjang 18-24 basa. Marka DNA hasil amplifikasi primer acak
(random primer) adalah marka DNA yang bersifat dominan, yaitu tidak dapat
membedakan antara genotipe tanaman homozigot dan heterozigot. RAPD
(Random Amplified Polymorphic DNA) adalah generasi pertama dari teknologi
marka DNA yang bersifat dominan. Generasi kedua dari marka-marka
berdasarkan teknik PCR adalah Amplified Fragment Length Polymorphisms
(AFLP) yang bersifat dominan dan Simple Sequence Repeats (SSR) yang bersifat
kodominan (Panaud et al. 1996).
Simple sequence repeats juga dikenal dengan mikrosatelit terdiri atas
pengulangan beberapa basa nukleotida, berupa dinukleotida, trinukleotida, atau
tetranukleotida, yang tersebar disepanjang genom kebanyakan spesies eukariotik
(Powell et al. 1996). Jumlah pengulangan nukleotida berkisar antara 5-40 kali
(Selkoe dan Toonen 2006) atau kurang dari 100 kali (Karp et al. 1997). Panjang
pengulangan ini bervariasi tergantung individu/varietas dan diwariskan kepada
generasi berikutnya. Motif pengulangan nukleotida yang paling banyak ditemukan
pada tanaman adalah AT, AG dan TC (Powell et al. 1996).

9
Primer spesifik dirancang pada runutan basa yang terkonservasi dan
selanjutnya digunakan untuk mengamplifikasi dan mengidentifikasi lokus yang
polimorfik dengan menggunakan metode standar elektroforesis pada gel
poliakrilamida (Jannati et al. 2009). Keunggulan analisis SSR adalah : 1)
mengidentifikasi polimorfisme secara akurat, 2) bersifat kodominan, dan 3) sangat
reproducible. Kemampuan teknik SSR membedakan individu-individu
berdasarkan kombinasi alel, menjadikan teknik ini sering digunakan untuk
mengidentifikasi dan menganalisis tetua (Zane et al. 2002) pada berbagai populasi
spesies tanaman (Holton et al. 2002). Kelemahan teknologi SSR adalah
memerlukan biaya dan curahan waktu yang tinggi untuk mengembangkannya,
sehingga penggunaan marka SSR terbatas pada tanaman-tanaman yang memiliki
nilai ekonomi tinggi (Ruan 2010).
Teknologi marka SSR telah mendominasi analisis genotyping tanaman
sebelum teknologi SNPs dikembangkan. Sejak sepuluh tahun terakhir, teknologi
marka SNPs mulai menggantikan teknologi marka SSR pada penelitian-penelitian
genetika. Marka SNPs adalah marka berdasarkan variasi perubahan satu basa (A,
T, G, atau C) pada situs-situs tertentu dari runutan basa DNA dalam genom
organisme (Ganal et al. 2009). Polimorfisme SNP tersedia paling melimpah dan
terdistribusi secara merata pada genom organism hidup (Aitken et al. 2009)
sehingga metode analisis marka DNA berdasarkan SNP mampu mengidentifikasi
variasi keragaman yang lebih tinggi dari metode analisis marka DNA berdasarkan
SSR (Li et al. 2009).
SNP (Single nucleotide polymorphism) adalah kelas mutasi yang disebabkan
oleh subsitusi atau insersi-delesi (indel) yang umum terjadi di genom organisme
hidup dan merupakan unit terkecil dari variasi genetik yang ditransmisikan dari
generasi ke generasi. Marka berdasarkan SNPs banyak digunakan pada studi-studi
tentang proses evolusi dari suatu genom atau gen, karena umumnya evolusi pada
sifat-sifat penting di tanaman adalah atribut dari keberadaan SNPs dan variasinya
(Shamay et al. 2004). SNP adalah perubahan posisi spesifik satu atau dua basa
nukleotida yang sifatnya melimpah dalam genom eukariot. Perbedaan basa
nukleotida diduga berpengaruh terhadap sifat fenotipik pada tiap-tiap individu
(McCouch et al. 2010). Jumlah SNP yang melimpah membuat marka SNAP lebih
menarik dibanding marka lainnya, termasuk dalam mengembangkan penanda bagi
gen target tertentu (Lestari dan Koh 2013). Deteksi marka SNAP yang bersifat ko-
dominan, berdasarkan pada amplifikasi PCR dengan primer yang berbasis pada
informasi sekuen untuk gen spesifik Marka SNAP saat ini telah digunakan
sebagai penanda genetik untuk berbagai fungsi pemuliaan tanaman, misalnya
analisis keragaman genetik, pembuatan linkage map dan Marker Assisted
Selection (Chen et al. 2011). Kelemahan dari teknik SNAP adalah memerlukan
informasi keragaman sekuen untuk suatu gen yang menjadi target analisis
(Mammadov et al. 2012).
Persebaran Serbuk Sari
Aliran gen atau gene flow adalah proses perpindahan informasi genetik
melalui persebaran serbuk sari (persebaran gamet jantan) dan melalui persebaran
benih (migrasi) (Mallet 2001). Aliran gen merupakan proses yang alami yang
terjadi pada tanaman yang menyebabkan gen-gen dalam tanaman berpindah.

10
Proses aliran gen dapat terjadi pada tanaman yang memiliki keserasian secara
seksual antara tanaman domestik maupun kerabat liarnya (Pandin 2009a). Analisis
aliran gen melalui serbuk sari dalam suatu populasi dapat digunakan untuk
menduga apakah terjadi perkawinan antara tanaman yang berbeda (outcrossing)
atau dengan tanaman yang sama (selfing) (Boer 2007). Hamrick dan Trapnell
(2011) mengatakan bahwa pola persebaran biji dapat dianalisis menggunakan dua
metode, yaitu :
a. Metode tak langsung meliputi analisis struktur genetik populasi menggunakan
marka genetik yang diwariskan secara maternal misalnya menggunakan
cpDNA (DNA kloroplas) dan mtDNA (DNA mitokondria) dalam satu
populasi.
b. Metode langsung menggambarkan pola persebaran biji menggunakan marka
molekuler untuk mengidentifikasi induk dari biji atau analisis parental.
Analisis metode langsung dibagi menjadi dua yaitu analisis induk jantan dan
betina dari biji dan analisis kecocokan antara induk jantan dengan induk
betina terhadap keturunannya.
Sistem perkawinan pada tanaman dapat diketahui melalui analisis pola
persebaran serbuk sari. Penelitian Carneiro et al. (2011) menyatakan bahwa
tanaman Hymenaea coubaril melakukan penyerbukan sendiri. Hal tersebut
bertentangan dengan penelitian sebelumnya oleh Dunphy et al. (2004) yang
menyatakan bahwa H. coubaril memiliki ketidaksesuaian secara seksual (self
incompability). Penebangan pohon H. coubaril secara bebas dalam areal
perhutanan dapat mengakibatkan berkurangnya pohon yang reproduktif. Kondisi
tersebut mengakibatkan tanaman terisolasi, sehingga persentase penyerbukan
sendiri dapat meningkat (Carneiro et al. 2011).
Informasi genetik dari suatu organisme tidak mengalami perubahan
sepanjang hayatnya namun tidak dapat dipertahankan karena masa hidup suatu
organisme tersebut sangat terbatas. Namun demikian setiap organisme
mempunyai potensi untuk menurunkan informasi genetik yang dimilikinya ke
keturunannya melalui pertukaran gamet dan hal ini akan menghasilkan
rekombinasi baru. Dengan demikian dinamika dari struktur genetik tidak dapat
diamati ditingkat organisme tunggal, tetapi diamati ditingkat populasi dimana
setiap anggota dari populasi tersebut saling bertukar gamet (Pandin 2009).

11
III. PERSEBARAN SERBUK SARI PADA KELAPA KOPYOR
GENJAH PATI MENGINDIKASIKAN PERANAN POLINATOR
SERANGGA DALAM PENYERBUKAN
Abstrak
Analisis parental dapat digunakan untuk mengevaluasi persebaran serbuk
sari pada kelapa kopyor. Tujuan penelitian untuk mengevaluasi (i) persebaran
serbuk sari, (ii) kisaran penyerbukan sendiri dan penyerbukan silang yang terjadi,
dan (iii) jarak serbuk sari ke tetua betina dalam populasi kelapa kopyor Pati. Hal
ini sangat penting dilakukan untuk petani dalam peningkatan panen buah kopyor
dan juga untuk mendukung pemuliaan kelapa mutan. Sebanyak 84 progeni
dipanen dari 15 tetua betina. Kandidat tetua sebanyak 95 pohon dianalisis untuk
melihat tetua jantan yang berptoensi untuk seluruh progeni. Provenan kelapa
dewasa dipetakan sesuai posisi GPS. Seluruh sampel di identifikasi menggunakan
6 lokus SSR dan 4 lokus SNAP. Analisis parental dilakukan menggunakan
CERVUS version 2.0 software. Hasil analisis mengindikasikan marka yang
digunakan efektif untuk mengidentifikasi kandidat tetua di seluruh progeni. Tidak
ada arah tertentu dari serbuk sari yang didonorkan tetua jantan ke tetua betina.
Serbuk sari yang didonorkan berasal tetua jantan dari arah relatif terhadap posisi
induk betina. Penyerbukan silang terjadi sebesar 82.1 % dari seluruh progeni
yang dianalisis. Penyerbukan silang antara kelapa dalam dengan kelapa dalam (D
x D), kelapa genjah dengan kelapa genjah (G x G), kelapa hibrida dengan kelapa
hibrida (H x H), D x G, G x D, D x H, G x H dan H x G seluruhnya diobservasi.
Penyerbukan sendiri (D x D dan G x G) terjadi sebesar 17,9% dari keseluruhan
progeni. Kelapa genjah tidak selamanya mengalami penyerbukan sendiri.
Keberadaan penyerbukan silang antara kelapa genjah dengan kelapa genjah,
kelapa dalam dengan kelapa genjah dan kelapa hibrida dengan kelapa genjah juga
diobservasi. Serbuk sari yang didonasikan dapat berasal dari serbuk sari donor
yang berada dalam kisaran jarak tempuh 0-58 meter dari tetua betina resipien
yang dievaluasi. Oleh karena itu, selain dengan adanya keberadaan angin,
polinator serangga juga dapat berperan penting dalam polinasi kelapa Kopyor.
Kata kunci : kelapa mutan, endosperma abnormal, analisis parental, persebaran
polen, penyerbukan silang, penyerbukan sendiri, marka SSR,
marka SNAP

12
III. Pollen Dispersal of Pati Kopyor Coconut Indicating Importance Roles Of
Insect Pollinator In Its Pollination *)
Abstract
Parentage analysis has been used to evaluate pollen dispersal in Kopyor
coconut (Cocos nucifera L.). The objectives of this research were to evaluate (i)
the dispersal of pollen, (ii) the rate of self and out-crossing pollination, and (iii)
the distance of pollen travel in Pati kopyor coconut population. The finding of this
activities should be beneficial to kopyor coconut farmers to increase their kopyor
fruit harvest and to support breeding of this unique coconut mutant. As many as
84 progeny arrays were harvested from 15 female parents. As many as 95 adults
coconut provenances surrounding the female parents were analyses as the
potential male parents for the progenies. The adult coconut provenances were
mapped according to their GPS position. All samples were genotyped using six
SSR and four SNAP marker loci. Parentage analysis was done using CERVUS
version 2.0 software. Results of the analysis indicated the evaluated markers were
effective for assigning candidate male parents to all evaluated seedlings. There is
no specific direction of donated pollen movement from assigned donor parents to
the female ones. The donated pollens could come from assigned male parents in
any directions relative to the female parent positions. Cross pollination occured in
as many as 82.1% of the progenies analyzed. Outcrossing among tall by tall
(TxT), dwarf by dwarf (DxD), hybrid by hybrid (HxH), TxD, DxT, TxH, DxH,
and HxD were observed. Self-pollination (TxT and DxD) occurred in as many as
17.9% of the progenies. The dwarf coconut is not always self pollinated. The
presence of DxD, TxD, and HxD outcrossing were also observed. The donated
pollens could come from pollen donor in a range of at least 0-58 m apart from the
evaluated female recipients. Therefore, in addition to the wind, insect pollinators
may play an important role in Kopyor coconut pollination.
KEY WORDS : Coconut mutant, abnormal endosperm, parentage analysis,
pollen movement, outcrossing rate, self pollination, SSR
marker, SNAP marker
--------------------
*) Sebagian dari penelitian Bab III. ini telah di-submit ke International Journal on
Coconut Research and Development (CORD)

13
Introduction
Kopyor coconuts are natural coconut mutants having abnormal endosperm
and only exist in Indonesia. The endosperm is soft, crumbly and detached from the
shell, forming flakes filling up the shell (Maskromo et al. 2007; Novarianto et al.
2014). The Makapuno coconut grown in the Philipines and other Asian countries is
another example of coconut mutant exhibiting endosperm abnormality (Samonthe
et al. 1989; Wattanayothin 2010). This mutant has been used as parent for
hybridizations in coconut breeding (Wattanayothin 2005). The Macapuno coconut
exhibits a soft and jelly-like endosperm (Santos 1999) that is phenotypically
different to Indonesian Kopyor coconut.
The kopyor coconut mutant phenotype is genetically inherited from parents to
their progenies (Sukendah 2009) and most probably is controlled by a single locus
(K locus) regulating the endosperm development of coconut (Sudarsono et al.
2014a). However, the identity of the regulatory locus has not yet been resolved.
The abnormal endosperm phenotype in kopyor coconut is controlled by the
recessive k allele; therefore, the genotype of kopyor fruit of coconut would be
homozygous kk for the zygotic embryos and homozygous kkk for the endosperm.
On the other hand, the genotype of the normal fruit of coconut would either be a
homozygous KK or a heterozygous Kk for the zygotic embryo and either a
homozygous KKK, heterozygous KKk, or heterozygous Kkk, respectively.
The origin of Kopyor coconut mutant is not well documented; however,
currently the kopyor provenances are found in a number of areas in Java and
southern part of Sumatera (Novarianto and Miftahorrachman 2000). The district
of Pati, Central Java Province is recognized as one of the Kopyor coconut
production centers. Kopyor coconuts have existed in this region for generations,
especially the dwarf type of Kopyor coconuts. Although only in a limited
numbers, Kopyor Tall and Kopyor Hybrid coconut types also exist along side of
the dwarf one.
The tall and dwarf coconut have different morphological characters and
pollination strategy. Tall coconuts are generally outcrossing since male flower
mature earlier than the female counterpart in the same inflorescence. Dwarf
coconut tends to self-pollinate because of an overlapping maturation period
between male and female flowers (Deb Mandal and Shyamapada 2011).
Pollination in coconut most probably is assisted by insect pollinators or by
the wind (Ramirez et. al. 2004). The family of Diptera, Coleoptera and
Hymenoptera are reported as effective pollinators of coconut (Ramirez et al.
2004). Distances of pollen transfer between male and female parents may be used
to predict the type of pollinator assisting pollination in coconut. Such question
may be answerred by studying pollen dispersal.
Evaluating pollen dispersal in various plant species usually use an approach
based on the parent – progeny genotype genotype (Austerlitz et al. 2004).
Evaluations have been done in pines (Schuster and Mitoon 2000), Dinizia excels -
Fabaceae (Dick et al. 2003), Quercus garryana - Fagaceae (Marsico et al. 2009)
and teak (Prabha et al. 2011). Availability of molecular markers capable of
identifying genotype of parents and their progenies should assist the pollen
dispersal studies. Using such markers, it should also be possible to estimate the

14
self-pollination and outcrossing rates in a certain population (Milleron et al.
2012).
To our understanding, pollen dispersal analysis has not been evaluated in
coconuts. With the development of kopyor coconut in Indonesia, availability of
information associated with pollen dispersal should be beneficial considering the
recessive nature of the kopyor character. Such coconut pollen dispersal evaluation
requires availability of some coconut progeny arrays and polymorphic loci for
molecular markers of coconut genome.
Co-dominant markers, such as SSR and SNAP markers for coconut have
been developed and routinely evaluated at PMB Lab, Department of Agronomy
and Horticulture, Faculty of Agriculture, Bogor Agricultural University (IPB),
Bogor, Indonesia for a number of plant species. These include coconut (Sudarsono
et al. 2014), cacao (Kurniasih 2012), and nut meg – Myristica sp. (Soenarsih
2012). Moreover, the gene specific SNAP markers have also been developed and
used successfully in coconut (Sudarsono et al. 2014).
The SSR markers have successfully been used in gene flow analysis of
pines (Lian et al. 2001; Burczyk and Koralewski. 2005). SNAP marker have also
been reported as an effective co-dominant marker for plant analysis (Morin et al.
2004, Sutanto et al. 2013) and proven to generate better data quality for the
majority of samples on plant genetic studies (Brumfield et al. 2003) and
population structure analysis (Herrera et al. 2007).
The objectives of this research were is to evaluate (i) the dispersal of pollen,
(ii) the rate of self and out-crossing pollination, and (iii) the distance of pollen
travel in Pati kopyor coconut population. The finding of these activities should be
beneficial to kopyor coconut farmers to increase their kopyor fruit yield and to
support breeding and cultivar development of this unique mutant.
Materials and Methods
Time and Location of Research
This research was conducted during the period of July 2012 up to January
2014. The field activities were at the Kopyor coconut plantation belonging to local
farmer‟s at Sambiroto, Pati District, Central Java, Indonesia. The research site was
at the following GPS location: S 6 32.182 E 11 03.354. The soil in the evaluated
Kopyor coconut plantation is sandy soil. The laboratory activities were done at
Plant Molecular Biology Laboratory (PMB Lab), Department of Agronomy and
Horticulture, Faculty of Agriculture, Bogor Agricultural University, Bogor,
Indonesia.
Selection of Parents and Progeny Arrays
There were 164 adult coconut trees in the field research site, consisted of a
mixture of both kopyor heterozygous Kk and normal homozygous KK coconut
trees. Only 95 out of 164 adult coconut trees in one block of 100x100 m2
were
sampled in this evaluation. Based on the coconut type, the sampled population
consisted of 68 dwarf, 14 tall and 13 hybrid coconuts. Moreover, based on their
phenotype, they were recognized as 22 normal homozygous KK and 73 kopyor
heterozygous Kk coconuts. Map of the existing coconuts in the research site was
generated using the GPS position of all individuals.

15
Six dwarf, seven tall, and two hybrid coconuts among the kopyor
heterozygous Kk trees were selected as female parents. They were selected using
purposive random sampling to represent different sites in the sampled population.
A single fruit bunch from each female parent containing 2-10 fruits/bunch was
harvested 10-11 months after pollination. The total harvested fruits were collected
and identified as either kopyor or normal fruits. The identified normal fruits were
germinated and DNA was isolated from leaf tissue of the germinated seedlings
(63 seedlings of normal fruits). The kopyor fruits are not able to naturally
germinate since this character is lethal. Zygotic embryos were isolated from the
identified kopyor fruits and DNA was isolated directly from the whole zygotic
embryo tissues (21 zygotic embryos). Among the 84 DNA samples, 26 samples
were from tall, 45 from dwarf, and 13 from hybrid female parents.
Genotyping of Parents and Progenies
DNA isolation was conducted using the CTAB method (Rohde et al. 1995).
Either young coconut leaf or zygotic embryo (0.3-0.4 g) was homogenized in 2 ml
of lysis buffer, containing 0.007 g PVP and 10 μl2-mercaptoetanol. The
homogenized tissues were then incubated in 65°C waterbath for 60 minutes and
the mixtures were centrifuged at 11000 rpm for 10 minutes using using the
Eppendorf 5416 centrifuge. Supernatant was then transferred to an Eppendorf tube
and an equal volume of chloroform:isoamyl-alcohol (24:1) was added. The
mixtures were mixed well; centrifuged at 11000 rpm for 10 minutes and the
supernatant was transferred into new microtube.
Cold isopropanol (0.8 volume of supernatant) and sodium acetate (0.1
volume of supernatant) were added into the supernatant. After overnight
incubation, the mixture was centrifuged at 11000 rpm for 10 minutes and DNA
pellet was retained. The DNA pellet was washed using 500 μl of cold 70%
ethanol, centrifuged and air dried before it was diluted into100 μl aquabidest.
RNA contaminants were remove using RNase treatment following standard
procedures (Sambrook and Russel 2001).
SSR marker at 37 loci (Lebrun et al. 2001) were evaluated for their
polymorphism 6 polimorphic loci were selected. In addition, four SNAP marker
loci developed based on nucleotide sequence variabilities of both SUS and
WRKY genes were also used to genotype all of the parents and progeny arrays.
To generate markers, PCR amplifications were conducted using the following
reaction mixtures: 2µl of DNA; 0.625 µl of primers, 6.25 µl PCR mix (KAPA
Biosystem), and 3 µl ddH20. Amplifications were conducted using the following
steps: one cycle of pre-amplification at 95°C for 3 minutes, 35 cycles of
amplification steps at 95 °C for 15 seconds (template denaturation), annealing
temperature for 15 seconds (primer annealing), and 72 °C for 5 seconds (primer
extension), and one cycle of final extention at 72 °C for 10 minutes as suggested
by KAPA Biosystem kit.
The generated SSR markers were separated using vertical 6%
polyacrilamide gel electrophoresis (PAGE) using SB 1x buffer (Brody and Kern
2004) and stained using silver staining. The silver staining was done following
methods developed by Creste et al. (2001). Electrophoregrams were visualized
over the light table and used to determine the genotype of the evaluated samples.

16
The generated SNAP markers were separated using 1% agarose gel
electrophoresis using TBE 1x buffer and stained using standard DNA staining
procedures (Sambrook and Russel 2001). The electrophoregrams were visualized
over the UV transluminescence table and recorded using digital camera. The
recorded pictures were used to determine the genotype of the evaluated samples.
Identification of the Candidate Male Parents
Each sample of the progeny arrays has a known female parent but unknown
pollen donor (the male parent). The candidate male parents could be any one of
the sampled adult population including the female parents. This steps were
conducted to determine the assigned male parent donating pollen to generate any
fruit in the progeny arrays.
Identification of the assigned male parent was done by analyzing genotype
of progeny and the respective female parent versus the genotype of all adult trees
in the selected samples. The ID of the potential male parent for any progeny was
determine based on the results of parentage analysis. Simulation was conducted to
determine the threshold level for confidence interval of 80% (relax) and 95%
(strict) levels before the final parentage analysis steps. Parentage analysis using
the genotype of progenies, female parents, and potential male parents was done
using CERVUS version 2.0 software (Marshall et al. 1998). Most likely approach
(potential male parent with the highest LOD score) based on the matching
genotype of progeny, female parent and potential male parent were used as the
basis for assigning certain adult individual as the potential male parent or pollen
donor of a progeny. The progeny and female parent genotype were compared with
those of other adult trees and the assigned male parent was selected based on the
output of CERVUS version 2.0 analysis results (Marshall et al. 1998).
Pattern of Pollen Dispersal
The location of the female and the asignmed male parents were plotted in
the map of adult individuals generated by Garmin MapSource GPS mapping
software version 76C5x. The distance between the known female parent and the
assigned male parent was calculated using the same software. The distances and
positions of both female and male parents in the generated map was then used to
ilustrate pattern of pollen dispersal in the location. Self pollination was defined if
the assigned male parent was the same as the female parent. Otherwise, they were
assigned as outcrossing. The outcrossing were further grouped as outcrossing
between either dwarf (dwarf parent pollinated other dwarf), tall (tall parent
pollinated other tall), or hybrid (hybrid parent pollinated other hybrid);
outcrossing between dwarf and either tall or hybrid (either tall or hybrid parent
pollinated by dwarf); outcrossing between tall by hybrid coconuts (tall parent
pollinated hybrid) or vice versa. The numbers of both self pollination and the
respective cross pollination were calculated.
Results and Discussions
The Parents and Progeny Arrays
Map of the existing coconut provenances in the research site are presented
in Figure 3.1. As indicated, the sample coconut population consists of a mixture of

17
normal homozygous KK and kopyor heterozygous Kk individuals and a mixture
of dwarf, tall and hybrid coconuts. All of these adult trees were used as potential
male parents capable of donating pollens to and pollinating the selected female
parents and generating the evaluated progeny arrays. The position of the selected
female parents (6 dwarf, 7 tall, and 2 hybrid kopyor heterozygous Kk coconuts)
are indicated in Figure 3.1. The harvested progenies from selected female parents
ranged from 2-10 progenies per female parent. Out of 84 selected progenies, 21
were kopyor nuts and 63 were normal ones. They were harvested from tall (26
progenies), dwarf (45 progenies), and hybrid (13 progenies) female parents,
respectively.
Genotyping of Parents and Progeny Arrays
The selected SSR and SNAP marker loci generated polymorphic markers in
the evaluated coconut population. An example of the polymorphic marker
generated by either the selected SSR (CnCir_56 locus) and SNAP (SUS 1_3
locus) primer pairs producing polimorphic markers is presented in Figure 2. and 3.
In Figure 2, the evaluated individuals are either homozygous cc (sample # 1), bb
(sample # 7-10), heterozygous bc (sample # 2-6, and 11), or heterozygous ab
(sample # 12) for the CnCir_56 SSR locus. On the other hand, the evaluated
individuals (sample # 1, 3, 4, 6) are heterozygous for reference and alternate
SNAP alleles and the other two (sample # 2 and 5) are homozygous for the
reference allele (Figure 3.3). All individuals were genotyped using the same
approaches. The summary of genotping results for a total of 179 individuals using
six SSR and four SNAP marker loci are presented in Table 3.1. The marker loci
generated a range of 2-4 alleles per locus (Table 3.1).
Mean number of alleles per locus is 3.4 and mean PIC for all marker loci
was 0.47. The polymorphic information content (PIC) for SSR marker loci ranges
from 0.31-0.68 while that of SNAP markers ranges from 0.28-0.37 (Table 3.1).
The PIC values represents measures of polimorphism between genotypes in a
locus using information of the allele numbers (Sajib et al. 2012). Total
exclusionary power using the ten marker loci is either 0.85 (first parent) or 0.97
(second parent), indicating the SSR and SNAP markers should be informative
enough for analyzing the evaluated coconut population.
Identification of the Candidate Male Parents
Results of simulation analysis using 10.000 iterations, 95 candidate male
parents, and the known female parent for each progeny, predicted the rate of
success in identifying male parents at 95% (strict) was 32% and at 80% (relax)
confidence interval was 62%. Parentage analysis was able to resolve the identity
of the male parent for every individual in the 84 progeny arrays using the most
likely parent approach. Moreover, the results of analysis also indicated that
assignment of the predicted male parents for the 20% (17 individuals) progenies
are at least in the minimum of 95% confidence and 43% (36 individuals) were at
least in the minimum of 80% confidence. The assignment for the male parents of
other 57% (48 individuals) progenies were at the level of less than 80 %
confidence. Although the confidence level was below 80 %, the male parent
assignment for these progenies shows LOD (likelihood of odds) value higher than
0. A positive LOD value indicates the suspected male parent might be the true

18
parents. According to Marshall et al. (1998), the higher the LOD value the the
higher the possibility the assigned male parent is the actual parent (Marshal et al.
1998).
Figure 3.1.Map of study site with the existed coconut provenances at Pati, Central
Java, Indonesia. The marks in this map indicated the positions of the
coconut provenances. The sampled provenances in an approximately
one hectare area are in the square box. The flags indicate the positions
of the selected female parents.
Note: The marks indicate the position of ( ) normal tall,( ) kopyor tall,
( ) normal dwarf, ( ) kopyor dwarf, ( ) normal hybrid, and ( ) kopyor
hybrid coconuts, respectively.
Figure 3.2. Polymorphism of SSR markers generated by PCR of the genomic
DNA sample # 1-12 with a pair of CnCir_56 SSR primers. M: 100 bp
DNA ladder markers. The a, b, and c are the three specific alleles of
the CnCir_56 locus.
1 2 3 4 5 6 7 8 9 10 11 12 M

19
Figure3.3. Polymorphism of SNAP markers generated by PCR of the genomic
DNA samples 1-6 with two pairs (the R: reference and the A: alternate
primer pairs) of the SUS 1_3 SNAP locus. M: 100 bp DNA ladder
markers. The R – PCR product of the R primer pairs and the A – PCR
product of the A primer pairs. The occurrences of both R and A PCR
products indicating the evaluated individuals are heterozygous, while
if either only A or B indicating they were homozygous.
Table 3.1. Numbers of alleles and individuals, numbers of heterozygous and
homozygous, observed (O) and expected (E) heterozygosity, and
polymorphic information content (PIC) at 10 molecular marker loci of
Kopyor coconut.
Locus Name No. of
alleles
No. of
individual
No. Of Heterozygosity
PIC Hetero-
zygous
Homozy-
gous O E
CnCir_87 2 179 27 152 0.15 0.39 0.31
CnCir_86 4 179 100 79 0.56 0.72 0.67
CnZ-18 4 179 74 105 0.41 0.61 0.57
CnZ_51 5 179 72 107 0.40 0.58 0.54
CnCir_B12 6 176 64 112 0.37 0.72 0.68
CnCir_56 5 179 78 101 0.44 0.64 0.57
CnSus1#14 2 179 143 36 0.80 0.49 0.37
CnSus1#3 2 174 147 27 0.85 0.50 0.37
WRKY19#1 2 176 111 65 0.63 0.47 0.36
WRKY 6#3 2 179 76 103 0.43 0.34 0.28
Cross pollination is pollination of female flower by male pollen from
different parents. Cross pollination produces half-sib progenies. The tall, dwarf
and hybrid coconuts could reciprocally donate their pollens. Based on the
assigned male parent of the 84 progeny arrays, cross pollination occured in as
many as 69 events (82.1 %). Among those identified as outcrossing, 4 events are
cross pollination between tall x tall (TxT), 16 tall by dwarf (TxD), and 4 tall by
hybrid (TxH) parents. Moreover, outcrossing among DxD (15 events), DxT (6
1 2 3 4 5 6 M R A R A R A R A R A R A

20
events), DxH (11 events), HxH (2 event) and HxD coconuts (11 events) were also
observed. Complete scheme and pollination types identified based on results of
pollen dispersal analysis are presented in Table 3.2.
The general understanding stated that because of the open flower
morphology and the differences in flower maturation, tall coconut is probably
always cross pollinated (Ramirez et al. 2004; Maskromo et al. 2011). However,
our data indicated there are at least 2.38% of self pollination among the tall
coconut (Table 3.2).
Self pollination is characterized by the pollination of female flower by male
pollen of the same parent. Self pollination produces full-sib progenies. Total
numbers of self pollination are observed in as many as 15 events (17.9 %) in the
evaluated progeny arrays (Table 3.2). They consist of two self pollination events
in the tall kopyor coconut (2.38 %) and 13 self pollination events in the dwarf
kopyor one (15.48%). Based on 13 progeny arrays harvested from the hybrid
parents, no self pollination in the hybrid coconut is recorded (Table 3.2).
The general understanding stated that because of the overlapping period
between male and female flower maturation, dwarf coconut is always self
pollinated (Maskromo et al. 2011). However, our data indicated the dwarf coconut
is not always self pollinated. Contrary to the basic understanding, our data
indicated the presence of more dwarf to dwarf (15 events, 17.86%), dwarf to tall
(6 events, 7.14%) and dwarf to hybrid (11 events, 13.1%) outcrossing (Table 3.2).
Finding by Rajesh et al. (2008) has previously indicated cross pollination
did occur in dwarf coconuts. Availability of new tools, such as molecular markers,
for analyzing outcrossing rate may change the previous understanding. Such
changes have been shown in Hymenaea coubaril which was previously reported
as more cross pollinated because of self incompatibility (Dunphy et al. 2004).
Table 3.2. Crossing schemes and pollination types identified based on results of
pollen dispersal analysis of the progeny arrays
Crossing
Scheme Pollination types Event numbers Percentage
TxT Self 2 2.4%
DxD Self 13 15.7%
HxH Self 0 0
Sub-total Self 15 17.9
TxT Outcross 4 4.8%
TxD Outcross 16 19.3%
TxH Outcross 4 4.8%
DxD Outcross 15 18.1%
DxT Outcross 6 7.2%
DxH Outcross 11 13.3%
HxH Outcross 2 2.4%
HxT Outcross 0 0
HxD Outcross 11 13.1%
Sub-total Outcross 69 82.1%
Total progenies 84 100.0%
Note: T – tall coconut, D – dwarf coconut, and H - hybrid coconut

21
However, more recent pollen dispersal studies indicated that H. coubaril is more
self pollinated (Carneiro et al. 2011).
Other alternative explanation for this findings is it is just a special case in
the evaluated site. In the study site, coconut provenances were planted in high
density planting. Moreover, population of honey bees exist in the coconut
plantation. Honey bees are known to roam around the male and female flowers
and function as effective pollinators for coconuts. The high density planting and
the availability of pollinators may have caused the unexpected outcrossing rate.
However, those are subjects of further investigations.
One assigned male parent may donate one or more pollens to the evaluated
female coconut parent, with a range of 1-5 pollens per assigned male one. Number
of assigned male parents donating certain numbers of pollen to the evaluated
female parents is presented in Figure 3.4. The data indicate that most of the
assigned male parents contribute only one pollen to the evaluated female parents.
Only three assigned male parents (two dwarf, and one hybrid coconuts) donated 4
or 5 pollens to the surrounding female parents.
The same female parents may receive donated pollens from different
numbers of assigned male parents, with a ranged of 1-7 assigned male parents
donated pollen to the same female one. The numbers female parents receiving
donated pollens from different number of assigned male parent iss presented in
Figure 3.5. The data indicated a single female parent most frequently received
pollens from 2, 4 or 5 different assigned male parents. Only three female parents
evaluated in this experiment (two dwarf and one hybrid coconuts) are found
receiving pollens from at least 6 assigned male parents (Figure 3.5).
Figure 3.4. Numbers of assigned male parent donating different numbers of
pollen to evaluated female parents.
Nu
mb
er
of m
ale
pare
nt
Number of donated pollen
27
11 7
2 1 0
10
20
30
40
1 2 3 4 5 6 7

22
Figure 3.5. Numbers of female parent receiving donated pollens from different
number of assigned male parents.
Pattern of Pollen Dispersal
The distances between female to the assigned male parents have been
determined based on their GPS positions. The distance of pollen travel between
assigned male to female parents as measured in this evaluation ranged from 0 - 58
m. Numbers of pollination events of each distance class from the assigned male to
the female coconut parents are presented in Figure 3.6. The assigned male parents
are distributed almost evenly in the different class distances from the female
parents. The 0 m distance between parents indicates self pollination events.
Figure 3.6. Numbers of pollination events for each distance class from the
assigned male to the female coconut parents.
Number of assigned male parents
Nu
mb
er
of fe
ma
le p
are
nt
1
3
1
4
3
1
2
0
2
4
6
1 2 3 4 5 6 7
Nu
mb
er
of eve
nts
Class distance between male to female (m)
15
6
13
18
12 12
7
0
10
20
0 < 10 < 20 < 30 < 40 < 50 < 60 > 60

23
To evaluate pattern of pollen dispersal among the assigned male parent to
the female, the positions of assigned male parents as pollen donors to one female
parent are plotted to a map using their GPS positions. Representative samples of
the assigned male parent positions to a single female recipient parent are
presented in Figures 3.7-3.11.
As the female parent, Hybrid kopyor # 059 (Figure 3.7) received 6 donated
pollens from six different assigned pollen donors. The pollen contributors to the
progeny array harvested from Hybrid kopyor # 059 female parent were all kopyor
heterozygous Kk coconuts. However, the seven progenies harvested from this
female parent were all phenotypically normal, i.e. genetically either a normal
heterozygous Kk or homozygous KK. The positions of the assigned male parents
relative to the female parent # 059 in the study site were presented in Figure 3.7.
The Dwarf kopyor # 067 (Figure 3.8) received 10 donated pollens from
eight different assigned male parents. The assigned pollen contributors to the
Dwarf kopyor # 067 female parent were all kopyor heterozygous Kk coconuts.
Only one out of the 10 progenies harvested from this female parent was
phenotypically kopyor. The assigned male parent for the harvested kopyor fruit
was the tall kopyor # 089. The positions of the assigned male parents relative to
the female parent # 067 were presented in Figure 3.8.
Figure 3.7. Pattern of pollen movement to female parent # 059 inferred from
parentage analysis. The mark indicates position of ( ) Dwaf kopyor,
( ) Hybrid kopyor as the assigned male (pollen donor) parents, and
( ) hybrid kopyor # 59 as the female recipient, respectively.

24
Figure 3.8. Pattern of pollen movement to female parent # 067 inferred from
parentage analysis. The marks indicate position of ( ) Dwaf kopyor,
( ) Hybrid kopyor, ( ) Tall kopyor as the assigned male (pollen
donor), and ( ) Dwarf kopyor # 067 as the donor pollens and female
recipient, respectively.
Dwarf kopyor # 068 (Figure 3.9) received 9 donated pollens from four
assigned male parents. The four progenies were the result of outcross with either
hybrid (# 59) or dwarf (#87 or # 90) and from self pollination. The assigned
pollen contributors to the Dwarf kopyor # 068 were all kopyor heterozygous Kk
coconuts. Three out of the 9 progenies harvested from Dwarf kopyor # 069 were
phenotypically kopyor. These kopyor fruits received one donated pollen from
either the tall kopyor # 059, dwarf kopyor # 68 or # 87. The positions of the
assigned male parents relative to the female parent # 68 were presented in Figure
3.9.
Dwarf kopyor # 084 (Figure 3.10) received 8 donated pollens from
surrounding pollen donors. The pollen contributors to the Dwarf kopyor # 084
female parent were all kopyor coconuts. Only two out of the 8 progenies
harvested from Dwarf kopyor # 084 were phenotypically kopyor. These two
kopyor fruits received donated pollens from either The two assigned male parents,
either dwarf kopyor # 056 (one pollen) and hybrid kopyor # 057 (one pollen),
each contributed a one pollen to the evaluated progenies. Moreover, assigned male
parent # 32 is the most distance pollen contributor among the evaluated trees. The
positions of the assigned male parents (pollen contributors) relative to the female
parent # 084 were presented in Figure 3.10.

25
Figure 3.9. Pattern of pollen movement to female parent # 068 inferred from
parentage analysis. The marks indicate position of ( ) Dwaf kopyor
and ( ) Hybrid kopyor as the assigned male parents (pollen donors),
and ( ) Dwarf kopyor # 068 as the donor pollens and female
recipient, respectively.
Figure 3.10. Patterns of pollen movement to female parent # 084 inferred from
parentage analysis. The marks indicate positions of ( ) Dwaf
kopyor, ( ) Hybrid kopyor, ( ) the Tall kopyor as the assigned
male - pollen donors, and ( ) Dwarf kopyor # 84 as the female
recipient, respectively.

26
Dwarf kopyor # 089 (Figure 3.11) received 7 donated pollens from
surrounding pollen donors. The pollen contributors to the Dwarf kopyor # 089
female parent were all kopyor coconuts. None of the 7 progenies harvested from
Dwarf kopyor # 089 was phenotypically kopyor. The positions of the assigned
male parents relative to the female parent # 089 were presented in Figure 3.11.
Figures 3.7-3.11 indicated there is no specific direction of donated pollen
movement from assigned male parents to the female parents. The donated pollen
could come from assigned male parents in any directions relative to the female
parent positions. The other positions of the assigned male parents were presented
in Figure attachment 5-14.
In the reseach location, wind blows from left to right during the night and
from right to left during the day. If the wind is the major pollinators, there should
be a specific pattern of pollen movement. Moreover, the distance of pollen
dispersals should be close to the pollen donors. Our data did not support the wind
as the only major pollinator in Kopyor coconut since pollens disperse in random
directions and the assigned male parents are as far as 58 m apart from the
evaluated female recipients. Our data also indicated that insect pollinators may
play an important role in Kopyor coconut pollination. Numbers of insects are
associated with inflorescence of kopyor coconuts. Such insects may aid
pollination and promote cross pollination in kopyor coconuts, as it happens to
other plant species (Bown 1988). These findings, however, do not rule out the role
of wind in the Kopyor coconut pollination, especially from closely spacing male
pollen donors.
Figure 3.11. Pattern of pollen dispersal plotted based on the assigned male parent
as pollen donor to female parent # 089. The marks indicate position
of ( ) Dwaf kopyor, ( ) the Tall kopyor as the assigned pollen
donor, and ( ) Tall kopyor # 89 as the donor pollens and female
recipient, respectively.

27
This might have been the first report of using molecular marker to study
pollen dispersal in coconut. Results of this study point to new finding about pollen
dispersal and pollination, selfing and out-crossing rates among dwarf, hybrid, and
tall coconuts, respectively. However, to generalize the finding will require more
further research and evaluation since these findings may be only specific for the
current study site.
Conclusion
The evaluated markers were effective for assigning candidate male parents
to all evaluated seedlings. There is no specific direction of donated pollen
movement from assigned donor parents to the female ones. The donated pollens
could come from assigned male parents in any directions relative to the female
parent positions. Based on the assigned male parent of the 84 progeny arrays,
cross pollination occured in as many as 69 events (82.1 %) including one among
tall by tall (TxT), dwarf by dwarf (DxD) and hybrid by hybrid (HxH) cross
pollination events. Moreover, outcrossing among TxD, TxH, DxH and vice-versa
were also observed. This finding also indicated the dwarf coconut is not always
self pollinated. The presence of 17.86 DxD, 19.05% TxD and 13.10% HxD were
also observed. In Kopyor coconut, the pollens could travel from pollen donors as
far as 58 m apart from the evaluated female recipients. Therefore, insect
pollinators may play an important role in these distances of Kopyor coconut
pollination.

28

29
IV. PERSEBARAN SERBUK SARI PADA KELAPA DALAM KALIANDA
MEMBUKTIKAN ADANYA PENYERBUKAN SILANG PADA KELAPA
DALAM
Abstrak
Analisis paternitas digunakan untuk mengetahui pola persebaran serbuk
sari pada kelapa kopyor (Cocos nucifera L.) Dalam Kalianda. Tujuan spesifik
penelitian ini untuk mengevaluasi (i) persebaran serbuk sari serta (ii) kisaran
besarnya penyerbukan sendiri dan penyerbukan silang pada Kelapa Dalam
Kopyor Kalianda, asal Kalianda, Lampung. Populasi yang digunakan dalam
penelitian ini terdiri atas 60 pohon kelapa dewasa yang semuanya tipe kelapa
Dalam. Dari 60 pohon kelapa dewasa tersebut dipilih 14 pohon dewasa yang
dijadikan sebagai induk betina. Sebanyak 49 progeni dipanen dari 14 induk betina
terpilih dan dikecambahkan untuk sumber DNA dalam analisis paternitas. Calon
tetua jantan terdiri atas 47 pohon kelapa berbuah normal (homozigot KK) dan 13
pohon kelapa berbuah kopyor (heterozigot Kk) sebanyak 13 pohon. Enam lokus
marker SSR polimorfik yaitu CnCir_B12, CnCir_86, CnCir_87, CnCir_56,
CnZ_51, CnZ_18 dan empat lokus marker SNAP polimorfik yaitu CNSUS1#14,
CNSUS1#3, WRKY6#3 dan WRKY19#1 digunakan untuk menentukan genotipe
seluruh progeni, seluruh kandidat tetua jantan, dan semua tetua betina yang
digunakan. Hasil percobaan menunjukkan serbuk sari kelapa kopyor Dalam
Kalianda menyebar dengan jarak sejauh 63 m. Jarak penyebaran serbuk sari
terbanyak yang teramati adalah pada jarak 40-50 m, dengan frekuensi sebesar 13
kejadian polinasi. Diantara 47 progeni yang dievaluasi, hanya satu (2%) progeni
yang berasal dari penyerbukan sendiri (self pollination) dan 48 (98%) berasal dari
penyerbukan silang. Hasil penelitian juga menunjukkan 13 progeni (27%) berasal
dari penyerbukan silang antar tetua yang sama-sama kopyor heterosigot Kk dan
35 (71%) progeni berasal dari penyerbukan silang antara tetua kopyor heterosigot
Kk dengan donor serbuk sari yang berasal dari kelapa normal homosigot KK.
Kata kunci : Kelapa Dalam Kopyor, penyerbukan sendiri, penyerbukan silang

30
IV. POLLEN DISPERSAL IN KALIANDA TALL COCONUT PROVED
OUTCROSSING NATURE OF TALL COCONUT POLLINATION
Abstract
Paternity analysis is used to determine the pattern of pollen spread among
kopyor coconut (Cocos nucifera L.) in Kalianda, Lampung. The specific objective
of this study are to evaluate (i) the spread of pollen (ii) the magnitude of the range
of self-pollination and cross-pollination in Kalianda Tall Kopyor coconut at
Kalianda, Lampung. The population used in this study consisted of 60 adult palm
trees, mixtures of heterozygous kopyor (Kk) and homozygous normal (KK)
coconut trees. Progeny arrays (47 nuts) were harvested from 14 kopyor
heterozygous Kk female parents and the progeny were germinated. As many as 60
adult trees surrounding the identified female parents were selected as potential
male parents, consisted of 47 normal homozygous KK palm and 13 kopyor
heterozygous Kk parents. Six polymorphic SSR marker loci used were
CnCir_B12, CnCir_86, CnCir_87, CnCir_56, CnZ_51, CnZ_18 and the four
polymorphic SNAP markers used were CNSUS1 # 14, CNSUS1 # 3, WRKY6#1
and WRKY19 # 3. The markers were used to genotype all the progeny, the
potential male and the female parents. Results of the experiment indicated pollen
of Kalianda Tall Kopyor coconut disperse as far as 63 m. The most frequent
pollen dispersal distance are between 40-50 m with the frequency of 13
pollination events. Among the evaluated progenies, only one (2%) comes from
self pollination event and 48 (98%) comes from cross pollination ones. Results of
the progeny evaluation also indicated that 13 progenies (27%) are the results of
outcrossing among kopyor heterozygous Kk parents and 35 events (71%) are from
outcrossing among kopyor heterozygous Kk parents and the normal homozygous
KK assigned male parents.
Key words : Tall coconut kopyor, self polination, outcrossing pollination, pollen
dispersal

31
Pendahuluan
Kelapa kopyor merupakan komoditas andalan yang bernilai ekonomi tinggi
dan dicirikan oleh daging buah yang bertekstur gembur dan sebagian besar tidak
melekat di tempurungnya serta rasa yang gurih pada buah yang muda. Di Filipina,
jenis kelapa ini disebut makapuno,di Sri Lanka dan Thailand disebut dikiri. Buah
kopyor ini diduga berasal dari tanaman kelapa yang mengalami mutasi genetik
secara alamiah. Kelapa berbuah kopyor adalah mutan kelapa yang ditemukan di
antara populasi kelapa normal. Buah kelapa kopyor dapat dipasarkan dalam
bentuk segar dan siap saji maupun melalui pengolahan lebih dahulu. Di Indonesia,
pemanfaatan kelapa kopyor lebih ditujukan untuk kebutuhan konsumsi bahan
pangan berupa es kopyor, es krim kopyor, koktil, selei kopyor dan bahan
campuran kue (Sudarsono et al. 2012) .
Hasil survei yang dilaksanakan Balitka pada tahun 2006 menunjukkan
bahwa kelapa kopyor terdiri atas dua tipe, yaitu tipe Dalam dan tipe Genjah. Tipe
Dalam terdapat di Kalianda (Lampung Selatan), Ciomas (Bogor), Sumenep dan
Jombang (Jawa Timur) dan Pati (Jawa Tengah). Pertanaman kelapa kopyor yang
ditanam secara populasi dijumpai di Dukuhseti, Kabupaten Pati (Jawa Tengah),
Sumenep (Jawa Timur), Ciomas (Bogor, Jawa Barat), Kalianda (Lampung
Selatan), Riau dan Kalimantan Timur. Kelapa kopyor yang ditanam secara
individu terdapat di Kabupaten Pati, Jombang dan Sumenep (Sudarsono et al.
2014a). Secara alami, tanaman kelapa kopyor tipe Dalam hanya menghasilkan
buah kopyor 1-2 butir per tandan. Hal ini disebabkan kelapa tipe Dalam termasuk
tanaman menyerbuk silang sehingga peluang bertemunya gen resisif pada bunga
betina dan serbuk sari relatif kecil. Kelapa kopyor tipe Genjah menghasilkan buah
kopyor per tandan lebih banyak dari tipe Dalam, kadang-kadang dapat mencapai
50%.
Kabupaten Lampung merupakan salah satu sentra kelapa di provinsi
Lampung, dengan total areal pertanaman pada tahun 2010 seluas 34.730 ha dan
produksi 30 435.60 ton. Luas areal pertanaman kelapa tersebut terus menurun
seiring dengan alih fungsi lahan untuk pengembangan komoditi lainnya dan untuk
perluasan areal pemukiman. Pada pertanaman kelapa untuk produksi kopra
maupun pemanfaatan lainnya ternyata terdapat kelapa Dalam kopyor dengan
luasan dan jumlah tegakan yang bervariasi. Kondisi pertanamannya dalam bentuk
populasi dan individu tanaman yang menyebar di area kebun kelapa beberapa
petani setempat. Hingga saat ini asal usul tanamannya belum diketahui. Umumnya
tanaman kelapa Dalam kopyor di Lampung berasal dari warisan orang tua atau
pengembangan dari tanaman kelapa Dalam kopyor yang diperoleh dari relasi
petani setempat (Sudarsono et al. 2014b).
Berdasarkan pola penyerbukan, karakter morfologi seperti tinggi pohon,
warna buah dan perbedaan kuantitatif dan kualitatif dalam komponen buah, dan
kecepatan berbunga pertama kelapa dikelompokkan menjadi dua tipe. Tipe kelapa
Dalam (Typica) cenderung menyerbuk silang disebabkan oleh bunga jantan yang
lebih duluan matang dibandingkan bunga betina. Setelah penyerbukan, periode
untuk menjadi buah sadalah 12 bulan. Tipe kelapa Genjah (Nana) cenderung
menyerbuk sendiri karena adanya overlapping antara fase bunga jantan dan
betinanya (DebMandal dan Shyamapada 2011). Kelapa memiliki tipe bunga
berumah satu (monoecious), secara fisik bunga jantan dan betina terpisah dalam

32
individu pohon yang sama. Walaupun bunga jantan dan betina ada pada individu
pohon yang sama, tetapi bunga jantan dan betina tersebut biasanya mekar pada
waktu yang berbeda. Persebaran serbuk sari pada berbagai spesies tanaman kini telah banyak
diteliti contohnya pada tanaman pinus (Schuster dan Mitoon 2000) dan Quercus
garryana famili Fagaceae (Marsico et al. 2009). Dengan mengidentifikasi semua
sumber serbuk sari yang potensial dalam suatu wilayah studi, penanda dari tetua
secara langsung dapat digunakan untuk mengidentifikasi tetua jantan dari progeni
yang dianalisis, dan dengan demikian memberikan wawasan tentang bagaimana
sifat-sifat individu yang diberikan oleh tetua jantan dan menduga kisaran peluang
tanaman menyerbuk sendiri atau menyerbuk silang dalam suatu populasi
(Milleron et al. 2012). Untuk tanaman kelapa belum pernah dilaporkan mengenai
pola persebaran serbuk sarinya secara spesifik.
Untuk mengembangkan pertanaman kelapa Dalam kopyor di Kabupaten
Lampung Selatan, maka dianggap perlu untuk melakukan analisis persebaran
serbuk sari pada kelapa Dalam Kalianda sebagai sumber informasi berbagai tipe
penyerbukan tanaman kelapa Dalam. Adapun tujuan spesifik penelitian ini untuk
mengevaluasi (i) persebaran serbuk sari (ii) kisaran besarnya penyerbukan sendiri
dan penyerbukan silang pada Kelapa Dalam Kalianda Lampung.
BAHAN DAN METODE
Waktu dan Lokasi Penelitian
Penelitian dilaksanakan mulai bulan Juli 2013 hingga Agustus 2014.
Kegiatan lapang dilakukan di perkebunan kelapa kopyor di Agom Jaya, Kalianda
Kabupaten Lampung Selatan, Lampung dengan lokasi GPS S5 39.462 E105
34.962. Tipe tanah yang dievaluasi pada perkebunan kelapa kopyor merupakan
tipe tanah berpasir. Kegiatan laboratorium dilakukan di Laboratorium Biologi
Molekuler Tanaman (PMB Lab) Departemen Agronomi dan Hortikultura Fakultas
Pertanian Institut Pertanian Bogor.
Pemilihan Tetua dan Pemanenan Progeni
Populasi terdiri atas 282 pohon. Populasi yang dipilih berdasarkan jumlah
tanaman yang lebih dari 10 pohon kopyor. Populasi tersebut merupakan campuran
dari pohon kelapa kopyor heterozigot (Kk) dengan pohon kelapa normal (KK)
atau kelapa yang tidak pernah menghasilkan buah kopyor. Berdasarkan tipe
kelapa tersebut, populasi yang digunakan dalam penelitian ini terdiri dari 60
pohon kelapa dewasa yang semuanya tipe kelapa Dalam, dari 60 pohon tersebut
dipilih 14 pohon yang dijadikan sebagai kandidat induk betina. Lebih lanjut
berdasarkan fenotipenya, pohon kelapa berbuah normal (homozigot KK) sebanyak
47 pohon dan pohon kelapa berbuah kopyor (heterozigot Kk) sebanyak 13 pohon.
Keberadaan setiap pohon kelapa di perkebunan ditunjukkan dengan menggunakan
pemetaan GPS.
Semua pohon-pohon dewasa yang berada di sekitar pohon induk betina
dijadikan sebagai calon tetua jantan untuk dijadikan donor serbuk sari. Satu
tandan buah kelapa (progeni) dipanen dari setiap induk betina yang terpilih.

33
Genotyping tetua dan progeni
Isolasi DNA dilaksanakan menggunakan metode CTAB (Rohde et al. 1995)
dengan modifikasi. Daun kelapa muda maupun embrio kelapa (0.3-0.4 g) digerus
dengan buffer lisis 2 ml, yang mengandung PVP 0.007 g dan 2-mercaptoetanol
sebanyak 10 μl. Hasil gerusan daun atau embrio diinkubasi dalam waterbath pada
suhu 65 °C selama 60 menit dan dihomogenkan menggunakan sentrifugasi
Eppendorf Centrifuge 5416 dengan kecepatan 11000 rpm selama 10 menit.
Supernatan dipindahkan ke tube eppendorf kemudian ditambahkan
kloroform:isoamil-alkohol (24:1) sebanyak volume supernatant lalu disentrifugasi
dengan kecepatan 11000 rpm selama 10 menit dan dipindahkan ke mikrotube
baru.
Supernatan dipindah ke tube eppendorf baru kemudian ditambahkan
isopropanol dingin (0.8 volume dari supernatan) dan sodium asetat (0.1 volume
supernatant). Setelah diinkubasi dalam freezer semalaman,suspensi kemudian
disentrifugasi dengan kecepatan 11000 rpm selama 10 menit hingga diperoleh
pellet DNA. Pellet DNA dicuci dengan menggunakan ethanol 70% dingin 500 μl.
lalu disentrifugasi dan dikeringkan. Pellet DNA diencerkan menggunakan
aquabidest sebagai suspensi DNA. Kontaminan RNA dihilangkan dengan
menggunakan RNase sesuai standar prosedur (Sambrook dan Russel 2001).
Primer SSR yang diseleksi dari 36 primer (Lebrun et al. 2001) (Lampiran 1)
diperoleh 6 primer yang polimorfik. Sebagai tambahan, empat lokus marka SNAP
berdasarkan keragaman sekuens nukleotida (gen SUS dan WRKY) (Lampiran 2)
juga digunakan untuk menganalisis seluruh tetua dan progeni. Amplifikasi PCR
dilakukan dengan menggunakan 2µl DNA working solution, 0.625 µl primer, 6.25
pcr mix (KAPA Biosystem), dan 3 µl ddH2O untuk setiap reaksi. Amplifikasi
dilakukan sebanyak 35 siklus, yang dimulai pada suhu 95 °C selama 3 menit
sebagai denaturasi awal, diikuti 95 °C selama 15 detik sebagai denaturasi siklus
pertama, penempelan primer spesifik - suhu disesuaikan dengan masing-masing
pasangan primer - selama 15 detik, pemanjangan primer pada suhu 72 °C selama
5 detik. Final extention 72 °C selama 10 menit sesuai rekomendasi kit KAPA
Biosystem.
Produk amplifikasi PCR untuk primer SSR dipisahkan dengan elektroforesis
gel poliakrilamid 6% menggunakan Buffer SB 1x (Brody dan Kern 2004) dan
pewarnaan gel dengan pewarna silver. Pewarnaan silver mengikuti metode Creste
et al. (2001) dengan modifikasi. Elektroforegram di visualisasi diatas meja UV
transluminesen di foto menggunakan kamera digital. Pemberian skor setiap
genotipe dilakukan juga di atas light table.
Produk amplifikasi PCR untuk primer SNAP dipisahkan menggunakan gel
agarose 1% menggunakan buffer TBE 1x dan pewarnaan menggunakan gel untuk
melihat hasil skoring yang dihasilkan (Sambrook and Russel 2001).
Elektroforegram di visualisasi diatas meja UV transluminesen di foto
menggunakan kamera digital. Pemberian skor setiap genotipe dilakukan untuk
setiap sampel.
Identifikasi Kandidat Tetua Jantan
Setiap progeni yang disampling sudah diketahui induk betinanya tetapi tidak
diketahui pohon yang menjadi donor serbuk sarinya. Kandidat tetua jantan bisa
menjadi salah satu induk betina dalam populasi pohon dewasa tersebut. Tahapan

34
ini dilakukan untuk memprediksi calon pendonor serbuk sari diantara calon tetua
jantan yang dievaluasi di setiap progeni.
Identifikasi donor serbuk sari dilakukan dengan menganalisis genotipe
progeni versus seluruh induk jantan yang dievaluasi. Semua induk jantan atau
induk betinanya dievaluasi dan berpotensi sebagai donor serbuk sari. Analisis
molekuler menggunakan software CERVUS analysis parentage program
komputer Cervus 2.0 (Marshall 1998). Hasil Cervus diperoleh data alel frekuensi,
PIC, heterozigositas, homozigositas. Dilanjutkan dengan data simulasi dan data
analisis tetua. Data analisis tetua akan diperoleh kandidat tetua dengan lambang
(*): artinya tingkat kepercayaan 95%. (+): tingkat kepercayaan 80%. dan (-):
tingkat kepercayaan < 80% .
Analisis Pola Persebaran Serbuk Sari
Lokasi induk betina dan tetua jantan terpilih hasil analisis Cervus
diplotkan menggunakan perangkat lunak pemetaan Garmin MapSource GPS versi
76C5x. Jarak antara induk betina dan tetua jantan dihitung menggunakan software
yang sama. Jarak dan posisi dari kedua induk betina dan tetua jantan yang
dihasilkan kemudian digunakan untuk mengilustrasikan pola persebaran serbuk
sari di lokasi penelitian. Penyerbukan sendiri didefinisikan jika induk jantan yang
teridentifikasi sama dengan tetua betinanya. Selain itu semuanya didefinisikan
sebagai penyerbukan silang. Penyerbukan silang bisa dikelompokkan dengan
kategori penyerbukan silang genjah (induk betina genjah diserbuki oleh tetua
jantan genjah lainnya), penyerbukan silang dalam (induk betina dalam diserbuki
oleh tetua jantan dalam lainnya), penyerbukan silang hibrida ((induk betina
hibrida diserbuki oleh tetua jantan hibrida lainnya), penyerbukan silang antara
induk betina genjah dengan Dalam atau hibrida, atau sebaliknya. Kedua tipe
persilangan dan kuantitasnya semuanya dihitung.
Hasil dan Pembahasan
Pemetaan Populasi Tanaman
Peta lokasi penelitian dapat dilihat pada Gambar 4.1. Populasi tersebut
menggambarkan populasi yang digunakan dalam analisis pola persebaran serbuk
sari sebanyak 60 pohon kelapa dewasa yang semuanya tipe kelapa Dalam, dari 60
pohon tersebut dipilih 14 pohon yang dijadikan sebagai kandidat tetua betina.
Pohon kelapa yang ada di areal pertanaman terdiri atas pohon kelapa berbuah
normal sebanyak 68 pohon dan pohon kelapa berbuah kopyor sebanyak 13 pohon.
Semua pohon-pohon dewasa yang berada di sekitar pohon induk betina dijadikan
sebagai calon induk jantan untuk dijadikan donor serbuk sari. Satu tandan buah
kelapa (progeni) dipanen dari setiap induk betina yang terpilih dengan kisaran
jumlah progeni per induk betina antara 2 sampai 10 buah. Progeni dari setiap
pohon induk betina dipanen masing-masing satu tandan dengan jumlah total
progeni 51 buah.
Berdasarkan tipe kelapa tersebut, populasi yang digunakan dalam
penelitian ini terdiri dari 60 pohon kelapa dewasa yang semuanya tipe kelapa
Dalam, dari 60 pohon tersebut dipilih 14 pohon yang dijadikan sebagai kandidat
induk betina. Lebih lanjut berdasarkan fenotipenya, pohon kelapa berbuah normal

35
(homozigot KK) sebanyak 47 pohon dan pohon kelapa berbuah kopyor
(heterozigot Kk) sebanyak 13 pohon.
Genotyping tetua dan progeni
Berdasarkan analisis molekuler menunjukkan bahwa seluruh primer yang
digunakan menunjukkan pita polimorfik baik mikrosatelit dan SNAP. Salah satu
contoh primer SSR (CnCir_56) dan SNAP ( WRKY 6_3 ) yang menghasilkan alel
polimorfik pada populasi kelapa kopyor yang dianalisis dapat dilihat pada Gambar
4.2. dan 4.3. Pada Gambar 4.2. individu yang dievaluasi untuk lokus SSR
CnCir_56 menunjukkan alel homozigot cc (sampel nomor 1), homozigot bb
(sampel nomor 7-10), heterozigot bc (sampel 2-6 dan sampel 11) atau heterozigot
ab (sampel nomor 12). Sedangkan individu sampel nomor 1,2,3,4 dan 6
heterozigot untuk referens dan alternate untuk alel SNAP (WRKY 6_3) dan
sampel 5 homozigot untuk alel referens (Gambar 4.3). Setiap individu di genotipe
dengan pendekatan yang sama.
Gambar 4.1. Lokasi studi penelitian tegakan kelapa kopyor di Agom Jaya
Kalianda Lampung Selatan. Simbol mengidentifikasikan posisi ( )
induk betina Dalam normal, ( ) tetua jantan Dalam normal, ( )
induk betina Dalam kopyor, dan ( ) tetua jantan Dalam kopyor.

36
2 3 4 5 6
Gambar 4.2. Visualisasi DNA hasil amplifikasi primer SSR CnCir_56 dengan
sampel DNA # 1-12. M: penanda 100 bp DNA . Posisi a, b, and c
menunjukkan posisi alel spesifik pada primer CnCir_56.
Gambar 4.3. Polimorfisme marker SNAP dua pasang dengan sampel DNA
genom 1-6 ( R: reference dan A: alternate pasang primer) pada
marka SNAP WRKY 6_3 SNAP. M: penanda 100 bp DNA . R -
produk PCR dari pasangan primer R dan A - produk PCR dari A
pasangan primer. Jika produk PCR menunjukkan R dan A
keduanya muncul maka individu yang dievaluasi heterozigot,
sedangkan jika hanya salah satunya A atau B menunjukkan
homozigot.
1 2 3 4 5 6
R A R A R A R A R A R A
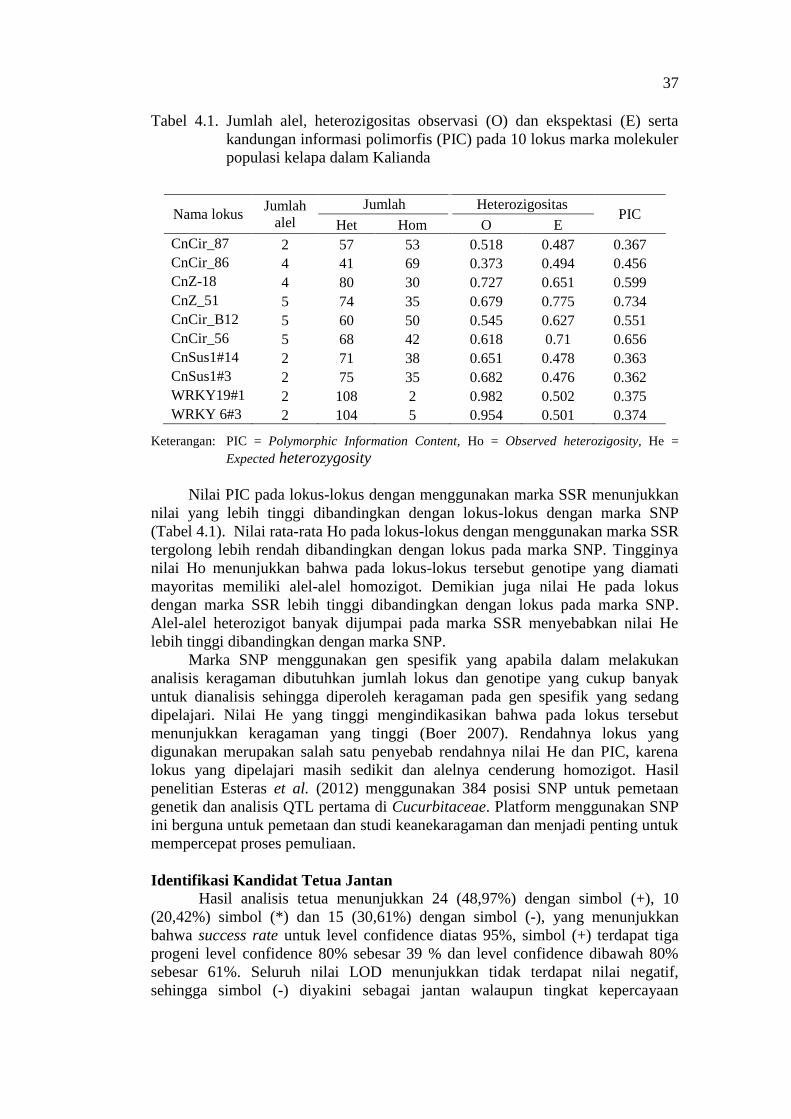
37
Tabel 4.1. Jumlah alel, heterozigositas observasi (O) dan ekspektasi (E) serta
kandungan informasi polimorfis (PIC) pada 10 lokus marka molekuler
populasi kelapa dalam Kalianda
Nama lokus Jumlah
alel
Jumlah Heterozigositas PIC
Het Hom O E
CnCir_87 2 57 53 0.518 0.487 0.367
CnCir_86 4 41 69 0.373 0.494 0.456
CnZ-18 4 80 30 0.727 0.651 0.599
CnZ_51 5 74 35 0.679 0.775 0.734
CnCir_B12 5 60 50 0.545 0.627 0.551
CnCir_56 5 68 42 0.618 0.71 0.656
CnSus1#14 2 71 38 0.651 0.478 0.363
CnSus1#3 2 75 35 0.682 0.476 0.362
WRKY19#1 2 108 2 0.982 0.502 0.375
WRKY 6#3 2 104 5 0.954 0.501 0.374 Keterangan: PIC = Polymorphic Information Content, Ho = Observed heterozigosity, He =
Expected heterozygosity
Nilai PIC pada lokus-lokus dengan menggunakan marka SSR menunjukkan
nilai yang lebih tinggi dibandingkan dengan lokus-lokus dengan marka SNP
(Tabel 4.1). Nilai rata-rata Ho pada lokus-lokus dengan menggunakan marka SSR
tergolong lebih rendah dibandingkan dengan lokus pada marka SNP. Tingginya
nilai Ho menunjukkan bahwa pada lokus-lokus tersebut genotipe yang diamati
mayoritas memiliki alel-alel homozigot. Demikian juga nilai He pada lokus
dengan marka SSR lebih tinggi dibandingkan dengan lokus pada marka SNP.
Alel-alel heterozigot banyak dijumpai pada marka SSR menyebabkan nilai He
lebih tinggi dibandingkan dengan marka SNP.
Marka SNP menggunakan gen spesifik yang apabila dalam melakukan
analisis keragaman dibutuhkan jumlah lokus dan genotipe yang cukup banyak
untuk dianalisis sehingga diperoleh keragaman pada gen spesifik yang sedang
dipelajari. Nilai He yang tinggi mengindikasikan bahwa pada lokus tersebut
menunjukkan keragaman yang tinggi (Boer 2007). Rendahnya lokus yang
digunakan merupakan salah satu penyebab rendahnya nilai He dan PIC, karena
lokus yang dipelajari masih sedikit dan alelnya cenderung homozigot. Hasil
penelitian Esteras et al. (2012) menggunakan 384 posisi SNP untuk pemetaan
genetik dan analisis QTL pertama di Cucurbitaceae. Platform menggunakan SNP
ini berguna untuk pemetaan dan studi keanekaragaman dan menjadi penting untuk
mempercepat proses pemuliaan.
Identifikasi Kandidat Tetua Jantan
Hasil analisis tetua menunjukkan 24 (48,97%) dengan simbol (+), 10
(20,42%) simbol (*) dan 15 (30,61%) dengan simbol (-), yang menunjukkan
bahwa success rate untuk level confidence diatas 95%, simbol (+) terdapat tiga
progeni level confidence 80% sebesar 39 % dan level confidence dibawah 80%
sebesar 61%. Seluruh nilai LOD menunjukkan tidak terdapat nilai negatif,
sehingga simbol (-) diyakini sebagai jantan walaupun tingkat kepercayaan

38
dibawah 80%. Nilai peluang kemungkinan atau LOD (Likelihood of odds)
menunjukkan nilai lebih besar dari 0 dan positif, nilai LOD positif lebih
mengindikasikan bahwa tetua jantan yang diduga adalah benar. Semakin besar
nilai LOD maka semakin besar peluang tetua tersebut merupakan tetua yang
sebenarnya (Marshal et al. 1998).
Dari hasil uji progeni dapat dibuat rangkuman skema persilangan dan tipe
penyerbukan pada populasi Kalianda yang menunjukkan bahwa terjadi selfing
genotipe kopyor sebanyak 1 kali (2%), persilangan antara genotipe kopyor
sebanyak 13 kali (27%) dan persilangan genotipe kopyor dan normal sebanyak 35
kali (71%) (Tabel 4.2). Jumlah progeni yang dihasilkan dari tetua jantan yang
teridentifikasi dapat dilihat pada Gambar 4.4. Penyerbukan polen dari tetua jantan
adalah sebanyak 1-3 serbuk sari, dengan frekuensi tertinggi yaitu 1 serbuk sari
sebanyak 15 kali penyerbukan oleh 1 tetua jantan (Gambar 4.5). Jarak persebaran
polen yang terdekat adalah 0 m dan terjauh > 60 m yaitu 63 m dengan frekuensi
penyerbukan tertinggi terjadi pada jarak >50 m yaitu sebanyak 13 kali (Gambar
4.6).
Tabel 4.2. Skema persilangan dan tipe penyerbukan yang diidentifikasi
berdasarkan hasil analisis persebaran dari uji progeni populasi
Kalianda.
Skema
Penyerbukan Tipe Penyerbukan
Jumlah peristiwa
penyerbukan Persentase
Kopyor x Kopyor Self 1 2%
Kopyor x Kopyor Outcross 13 27%
Kopyor x Normal Outcross 35 71%
Total progeni 49 100.00%
Gambar 4.4. Jumlah progeni yang dipanen per induk betina dan jumlah tetua
jantan yang mendonasikan serbuk sari untuk tetua betina tertentu
pada pertanaman kelapa kopyor populasi Agom Jaya, Lampung
Selatan
9
2 3
2
4 3
4
1
3
1
4
1
6 5
1
7
3 3
2 2
5 4
0
2
4
6
8
10
129
148
149
150
151
156
158
160
163
185
193
194
209
210
217
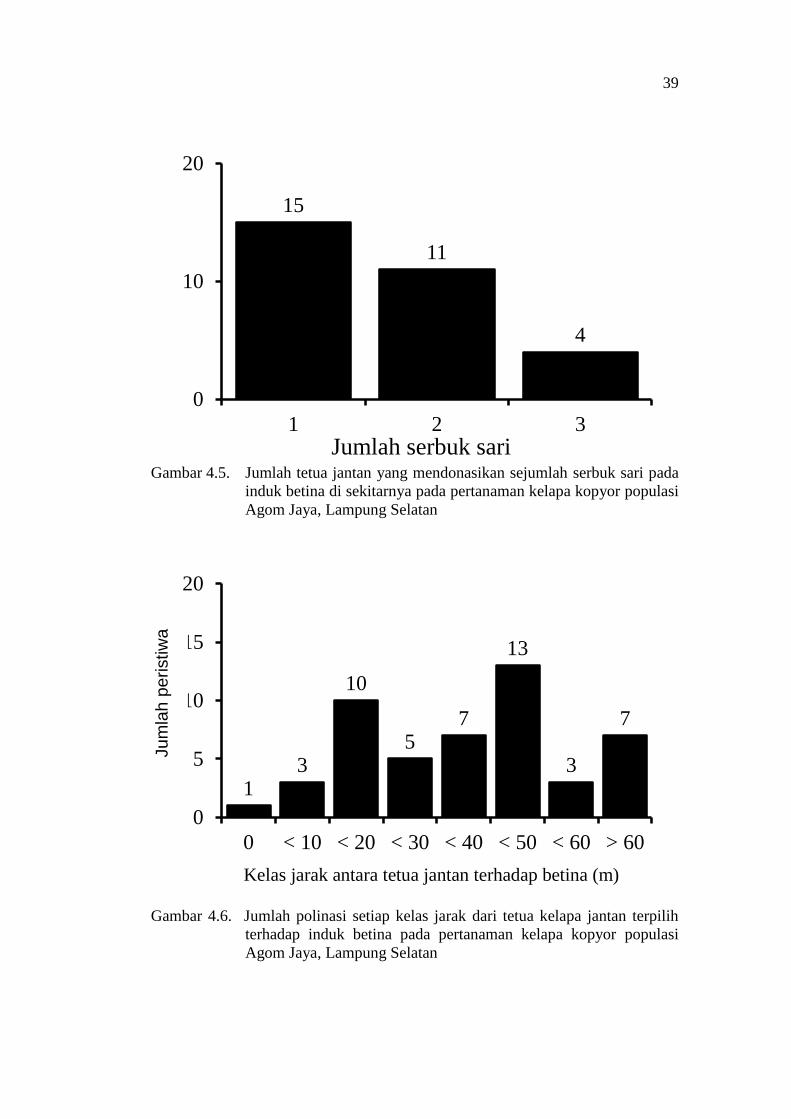
39
Gambar 4.5. Jumlah tetua jantan yang mendonasikan sejumlah serbuk sari pada
induk betina di sekitarnya pada pertanaman kelapa kopyor populasi
Agom Jaya, Lampung Selatan
Gambar 4.6. Jumlah polinasi setiap kelas jarak dari tetua kelapa jantan terpilih
terhadap induk betina pada pertanaman kelapa kopyor populasi
Agom Jaya, Lampung Selatan
1 3
10
5 7
13
3
7
0
5
10
15
20
0 < 10 < 20 < 30 < 40 < 50 < 60 > 60
Jumlah serbuk sari
15
11
4
0
10
20
1 2 3
Kelas jarak antara tetua jantan terhadap betina (m)
Jum
lah p
eristiw
a
pe
nye
rbu
kan

40
Analisis Pola Persebaran Serbuk Sari
Evaluasi pola persebaran polen dari tetua jantan yang sudah ditentukan
terhadap induk betina, posisi tetua jantan sebagai donor polen ke salah satu induk
betina di plotkan di peta menggunakan posisi GPS. Contoh gambaran dari posisi
tetua jantan yang sudah ditentukan terhadap induk betina resipien dipresentasikan
pada gambar 4.7-4.18. Beberapa pola penyerbukan pada pohon induk betina
kelapa Dalam kopyor yang memperlihatkan seluruh donor serbuk sari berasal dari
pohon Dalam kopyor terjadi pada induk betina nomor 148, 150, 163, 193, dan 194
(Gambar 4.7 - 4.11).
Gambar 4.7. Pola persebaran serbuk sari tanaman kelapa Dalam Kopyor Kalianda
induk betina nomor #148. Simbol mengidentifikasikan posisi ( )
induk betina Dalam kopyor dan ( ) tetua jantan Dalam kopyor.

41
Gambar 4.8. Pola persebaran serbuk sari tanaman kelapa Dalam Kopyor Kalianda
induk betina nomor 150. Simbol mengidentifikasikan posisi ( )
induk betina Dalam kopyor dan ( ) tetua jantan Dalam kopyor.
Gambar 4.9. Pola persebaran serbuk sari tanaman kelapa Dalam Kopyor Kalianda
induk betina nomor 194. Simbol mengidentifikasikan posisi ( )
induk betina Dalam kopyor dan ( ) tetua jantan Dalam Dalam
kopyor.

42
Gambar 4.10. Pola persebaran serbuk sari tanaman kelapa Dalam Kopyor
Kalianda induk betina nomor 163. Simbol mengidentifikasikan
posisi ( ) induk betina Dalam kopyor dan ( ) tetua jantan Dalam
kopyor.
Gambar 4.11. Pola persebaran serbuk sari tanaman kelapa Dalam Kopyor
Kalianda induk betina nomor 193. Simbol mengidentifikasikan
posisi ( ) induk betina Dalam kopyor dan ( ) tetua jantan Dalam
kopyor.
Beberapa pola penyerbukan pada pohon induk betina kelapa Dalam
kopyor yang memperlihatkan sebagian donor serbuk sari berasal dari pohon
Dalam normal terjadi pada induk betina nomor 149, 151, dan 156, (Gambar 4.12 -
4.14).

43
Gambar 4.12. Pola persebaran serbuk sari tanaman kelapa Dalam normal Kalianda
induk betina nomor 149. Simbol mengidentifikasikan posisi ( )
induk betina Dalam normal, ( ) tetua jantan Dalam normal dan ( )
tetua jantan Dalam kopyor.
Gambar 4.13. Pola persebaran serbuk sari tanaman kelapa Dalam normal Kalianda
induk betina no. 151. Simbol mengidentifikasikan posisi ( )
induk betina Dalam kopyor, ( ) tetua jantan Dalam kopyor, dan (
) tetua jantan Dalam normal.

44
Gambar 4.14. Pola persebaran serbuk sari tanaman kelapa Dalam Kopyor
Kalianda induk betina nomor 156. Simbol mengidentifikasikan
posisi ( ) induk betina Dalam kopyor dan ( ) tetua jantan Dalam
normal.
Gambar 4.15. Pola persebaran serbuk sari tanaman kelapa Dalam Kopyor
Kalianda induk betina nomor #129. Simbol mengidentifikasikan
posisi ( ) induk betina Dalam Normal, ( ) tetua jantan Dalam
kopyor, dan ( ) tetua jantan Dalam normal
Pohon induk betina kelapa Dalam normal yang ada di sekitar pohon kelapa
Dalam kopyor juga dapat menerima sumbangan serbuk sari yang berasal dari
kelapa Dalam kopyor. Gambar 4.15-4.17 menunjukkan pohon induk betina Dalam
normal nomor 129, 158, dan 209 menerima serbuk sari yang berasal dari donor
kelapa Dalam kopyor dan Dalam Normal. Sebaliknya, pohon induk betina Dalam

45
normal nomor 210 hanya menerima serbuk sari yang berasal hanya dari donor
kelapa Dalam kopyor (Gambar 4.18).
Gambar 4.16. Pola persebaran serbuk sari tanaman kelapa Dalam Kopyor
Kalianda induk betina nomor 158. Simbol mengidentifikasikan
posisi ( ) induk betina Dalam normal, ( ) tetua jantan Dalam
kopyor, dan ( ) tetua jantan Dalam normal.
Gambar 4.17. Pola persebaran serbuk sari tanaman kelapa Dalam Kopyor
Kalianda induk betina nomor 209. Simbol mengidentifikasikan
posisi ( ) induk betina Dalam, ( ) tetua jantan Dalam normal dan
( ) tetua jantan Dalam kopyor.

46
Gambar 4.18. Pola persebaran serbuk sari tanaman kelapa Dalam Kopyor
Kalianda induk betina nomor 210. Simbol mengidentifikasikan
posisi ( ) induk betina Dalam kopyor dan ( ) tetua jantan Dalam
kopyor.
Salah satu populasi kelapa berbuah kopyor dilaporkan di Kecamatan
Kalianda Lampung Selatan (Mahmud 2000). Daerah sentra kopyor seperti
Lampung dan Sumenep, petani memperbanyak dari pohon-pohon penghasil buah
kopyor tipe Dalam. Perkembangan tanaman kelapa berbuah kopyor ini mungkin
terjadi karena tumbuhnya buah kelapa normal yang memiliki gen kopyor dari
tanaman penghasil buah kopyor tersebut. Selain buah kopyor bergenotipe kk
(homozigot) yang letal, pada tanaman atau tandan yang sama juga dihasilkan buah
dengan endosperma normal yang memiliki embrio mengandung gen kopyor
dengan genotipe Kk (heterozigot) (Santos 1999).
Pola pertanaman kelapa Dalam berbuah kopyor di Kalianda yang bercampur
dengan kelapa Dalam normal. Pola tersebut memungkinkan terjadinya persilangan
alami antar tanaman kelapa Dalam berbuah kopyor dengan kelapa dalam berbuah
normal yang ada di sekitarnya. Hal tersebut disebabkan karena tipe kelapa Dalam
umumnya memiliki pola penyerbukan secara silang, karena periode kematangan
bunga betina (masa reseptif) tidak bersamaan dengan periode matangnya bunga
jantan (antesis) pada satu tandan yang sama (Maskromo et al. 2011).
Terbentuknya buah kopyor disebabkan oleh bertemunya gen kopyor pada bunga
betina kelapa yang membawa sifat kopyor dengan gen kopyor pada serbuk
sariyang menyerbuki bunga betina tersebut (Santos 1999).
Bunga betina kelapa Dalam kopyor heterozigot atau tanaman kopyor
homozigot yang dibuahi serbuk sari dari pohon kelapa normal akan membentuk
buah normal atau tidak kopyor. Pola pembungaan dan pembuahan pada tanaman
kelapa kopyor tipe Dalam tersebut menyebabkan rendahnya peluang terbentuknya
buah kopyor pada tanaman kelapa berbuah kopyor yang dikelilingi tanaman

47
kelapa normal. Produksi buah kopyor pada tanaman yang tumbuh berdekatan
dengan beberapa pohon kopyor lainnya lebih tinggi dibandingkan dengan tanaman
kopyor yang dikelilingi oleh banyak tanaman kelapa normal di sekitarnya.
Berdasarkan hasil pengamatan tersebut maka disarankan untuk menebang dan
mengganti tanaman kelapa normal secara bertahap dengan tanaman berbuah
kopyor (Sudarsono et.al. 2014a).
Simpulan
Pola persebaran polen pada tanaman kelapa kopyor tipe Dalam Kalianda
menunjukkan penyerbukan kelapa kopyor oleh kelapa normal dengan jarak
persebaran serbuk sari paling sering terjadi pada jarak <50 m dengan frekuensi 13
kali penyerbukan, dengan persebaran terjauh pada jarak 63 m dan persebaran
terdekat pada jarak 0 m. Hasil uji progeni menunjukkan persentase terjadinya
selfing sebanyak 1 kali (2%) dan persentase outcrossing terjadi sebanyak 48 kali
(98%) sehingga membuktikan besarnya penyerbukan silang pada tipe kelapa
Dalam.

48

49
V. PERSEBARAN SERBUK SARI MEMBUKTIKAN POTENSI
PENGARUH NEGATIF KELAPA NORMAL TERHADAP
HASIL BUAH KOPYOR
Abstrak
Salah satu tipe kelapa unik yang terdapat di Indonesia yaitu kelapa berbuah
kopyor. Buah kopyor ini diduga berasal dari tanaman kelapa yang mengalami
mutasi genetik secara alamiah. Upaya untuk mengatasi masalah produktifitas
buah kopyor yang rendah salah satunya adalah dengan mempelajari efek xenia
pada populasi kopyor. Xenia merupakan gejala genetik berupa pengaruh langsung
serbuk sari pada fenotipe biji dan buah yang dihasilkan tetua betina. Populasi yang
dipilih adalah area pertanaman di pekarangan rumah penduduk. Populasi tersebut
merupakan campuran dari pohon kelapa kopyor heterozigot (Kk) dengan pohon
kelapa normal (KK) atau kelapa yang tidak pernah menghasilkan buah kopyor.
Pohon kelapa yang ada di areal pertanaman terdiri atas pohon kelapa berbuah
normal sebanyak 33 pohon dan pohon kelapa berbuah kopyor sebanyak 9 pohon.
Semua pohon-pohon dewasa yang berada di sekitar pohon induk betina dijadikan
sebagai calon induk jantan untuk dijadikan donor serbuk sari. Hasil analisis
menunjukkan adanya pengaruh negatif xenia terhadap hasil panen buah kopyor
ketika pohon betina kopyor heterosigot Kk dikelilingi oleh kelapa normal
homosigot KK sebagai donor serbuk sari. Sebanyak 99 buah progeni yang
dipanen dari pohon induk betina kelapa kopyor heterosigot Kk, semuanya
mempunyai fenotipe normal. Hasil penelitian juga menunjukkan persebaran
serbuk sari dapat terjadi dengan kisaran jarak 0 m (self pollination) hingga 54 m
(outcrossing). Frekuensi persebaran serbuk sari terbesar terjadi pada jarak antar
tetua yang berkisar 20 m. Pada jarak tersebut, terjadi penyerbukan sebanyak 30
kali. Frekuensi terjadinya penyerbukan silang sebesar 95% dan penyerbukan
sendiri sebesar 5%.
Kata kunci : kelapa Dalam kopyor, xenia, persebaran serbuk sari

50
V. POLLEN DISPERSAL IN PATI TALL COCONUT INDICATED
NEGATIVE XENIA EFFECT FOR KOPYOR FRUIT YIELD*
Abstract
Kopyor coconut is one of the many unique coconut type existed in
Indonesia. Kopyor coconut is a naturally occuring coconut mutant. Efforts to
address the problem of low kopyor fruit yield is conducted by studying effect of
xenia on kopyor fruit yield. Xenia is a genetic phenomenon in the form of a direct
effect of pollen to the phenotype of fruits yielded by the female parents. The
mixture of kopyor and normal coconut population was selected in the farmers
coconut plantations. All adults trees surrounding the 9 kopyor heterozygous Kk
palms were evaluated as potential male candidate parents (pollen donors). Results
of the analysis indicated xenia reduce kopyor fruit yields. Kopyor heterozygous
Kk female parents produced low number of kopyor fruits when they are
surrounded by many normal homozygous KK pollen donors. Out of 99 harvested
progeny arrays from the kopyor heterozygous Kk female parents, none exhibited
kopyor phenotype. The results also indicated the pollen dispersal from normal
homozygous KK donor palms range from 0 m (self pollination) to 54 m
(outcrossing). The highest frequency of pollens are dispersed from a distance of
less than 20 m (30 pollination events). The occurence of outcrossing frequency is
at least 95% and the selfing frequency is 5%.
Key Words : Tall kopyor coconut, xenia, pollen dispersal
---------------------- *)
This chapter has been submitted to Buletin Palma : entitled ; Persebaran serbuk
sari pada kelapa dalam dukuh seti membuktikan potensi pengaruh negatif kelapa
normal terhadap hasil buah kopyor

51
Pendahuluan
Indonesia merupakan salah satu negara penghasil kelapa terbesar kedua di
dunia setelah Filipina. Selain memiliki areal pertanaman terluas, Indonesia juga
mempunyai keanekaragaman genetika kelapa yang besar. Hasil eksplorasi Balai
Penelitian Palma Manado diperoleh kurang lebih 105 aksesi kelapa Dalam dan
kelapa Genjah. Saat ini hasil eksplorasi tersebut dikoleksi secara ex situ di sam
beberapa kebun koleksi plasma nutfah kelapa (Novarianto et al. 2000). Diantara
berbagai jenis kelapa yang ada di Indonesia terdapat salah satu tipe kelapa yang
unik yaitu kelapa berbuah kopyor. Buah kopyor ini diduga berasal dari tanaman
kelapa yang mengalami mutasi genetik secara alamiah. Sebagai hasil mutasi
alamiah, jumlah tanaman kelapa berbuah kopyor sangat sedikit dibandingkan
dengan tanaman kelapa berbuah normal. Menurut Falconer (1985) peluang
terjadinya mutasi alamiah secara umum sangat rendah yaitu sebesar 10-5
sampai
10-6
untuk setiap generasi. Hal ini berarti bahwa hanya 1 diantara 100.000 sampai
1.000.000 peluang terjadinya mutasi alamiah di alam. Selain itu organisme hasil
mutasi cenderung letal, sehingga perkembangbiakannya terhambat dan akhirnya
punah.
Berdasarkan hal tersebut, kelapa mutan ini seharusnya tidak berkembang,
tetapi ternyata kelapa ini banyak ditemukan di beberapa sentra produksi kelapa di
Indonesia. Sifat kopyor dikendalikan oleh gen mutan pada lokus K. Buah kopyor
mempunyai embrio zigotik denga genotipe homozigot kk dan endosperma kkk.
Buah kelapa normal mempunyai embrio zigotik dengan genotipe homozigot KK
atau heterozigot Kk dan alternatif endosperma KKK, KKk atau Kkk. Buah kopyor
secara alami tidak dapat berkecambah sedangkan buah normal homozigot KK
atau heterozigot Kk dapat berkecambah dan menghasilkan bibit kelapa.
Keberadaan tanaman kelapa kopyor di Kab. Pati, Jawa Tengah sudah
diketahui oleh masyarakat luas terutama di daerah Jawa sejak tahun 1960an dan
pengembangannya terus dilakukan hingga saat ini (Dishutbun Pati 2004).
Tanaman ini tersebar di tujuh kecamatan yaitu Dukuhseti, Margoyoso, Tayu,
Wedarijaksa, Trangkil, Gunung Wungkal dan Cluwak. Luas pertanaman kelapa
kopyor di Kabupaten Pati 378.09 Ha dan areal terluas terdapat di tiga kecamatan,
yaitu Dukuhseti, Margoyoso dan Tayu, berturut-turut seluas 132.60 Ha, 131.55
Ha dan 69.50 Ha (Anonim 2004). Menteri Pertanian Republik Indonesia pada
tahun 2010 telah melepas secara resmi tiga varietas unggul lokal kelapa Genjah
Kopyor asal Pati. Tiga varietas yang dimaksud adalah Genjah Hijau Kopyor,
Genjah Coklat Kopyor dan Genjah Kuning Kopyor. Selain di tempat asalnya,
ketiga varietas kelapa kopyor tersebut telah ditanam sebagai koleksi di Kebun
Percobaan Kima Atas, Balitka Manado pada tahun 2006.
Salah satu permasalahan yang dihadapi petani adalah produksi buah kopyor
yang dipanen belum optimal. Tanaman kelapa kopyor di Pati dapat menghasilkan
jumlah buah kopyor sebanyak 1-4 butir per tandan dari jumlah total buah 7-15
butir. Secara teori, berdasarkan pola segregasi Hukum Mendel, maka peluang
terbentuk buah kopyor seharusnya mencapai 25% dari jumlah buah toal yang
dipanen (Maskromo 2005).
Upaya untuk mengatasi masalah produktifitas buah kopyor yang rendah
salah satunya adalah dengan mempelajari efek xenia pada populasi kopyor. Xenia
merupakan gejala genetik berupa pengaruh langsung serbuk sari pada fenotipe biji

52
dan buah yang dihasilkan tetua betina. Pada kajian pewarisan sifat, ekspresi dari
gen yang dibawa tetua jantan dan tetua betina diekspresikan pada generasi
berikutnya. Dengan adanya xenia, ekspresi gen yang dibawa tetua jantan dapat
diekspresikan pada tetua betina (buah) (Bulant et al. 2000). Fenomena xenia pada
tanaman kelapa telah diketahui berpengaruh terhadap ukuran buah, berat kopra
dan ketegaran hibrida (Satyabalan 1995).
Xenia berhubungan dengan fenotipe yang dikendalikan oleh gen resesif.
Terbentuknya buah kopyor pada kelapa tergantung pada genotipe serbuk sari yang
menyerbuki bunga betina tanaman kelapa kopyor, sehingga merupakan contoh
xenia. Adapun tujuan spesifik penelitian ini untuk mengevaluasi efek negatif
xenia pada pertanaman kelapa kopyor Dalam Pati dan mengetahui kisaran
besarnya penyerbukan sendiri dan penyerbukan silang yang terjadi pada populasi
Kelapa Dalam Pati.
BAHAN DAN METODE
Waktu dan Lokasi Penelitian
Penelitian dilaksanakan mulai bulan Maret 2013 hingga Desember 2013.
Kegiatan lapang dilakukan di perkebunan kelapa kopyor di Desa Dukuh Seti
Kabupaten Pati, Jawa Tengah dengan lokasi GPS S6 27.649 E111 02.747. Tipe
tanah yang dievaluasi pada perkebunan kelapa kopyor merupakan tipe tanah
berpasir. Kegiatan penelitian dilakukan di Laboratorium Biologi Molekuler
Tanaman (PMB Lab) Departemen Agronomi dan Hortikultura Fakultas Pertanian
Institut Pertanian Bogor.
Pemilihan Tetua dan Pemanenan Progeni
Populasi yang dipilih adalah area pertanaman di pekarangan rumah
penduduk. Populasi tersebut merupakan campuran dari pohon kelapa kopyor
heterozigot (Kk) dengan pohon kelapa normal (KK) atau kelapa yang tidak pernah
menghasilkan buah kopyor dengan total pohon dewasa 42 pohon. Pohon kelapa
yang ada di areal pertanaman terdiri atas pohon kelapa berbuah normal sebanyak
33 pohon dan pohon kelapa berbuah kopyor sebanyak 9 pohon. Semua pohon-
pohon dewasa yang berada di sekitar pohon induk betina dijadikan sebagai calon
induk jantan untuk dijadikan donor serbuk sari. Pemilihan pohon yang dijadikan
sebagai induk betina dilakukan dengan metode purpossive random sampling.
Induk betina yang digunakan sejumlah 7 pohon kelapa Dalam kopyor.
Masing-masing pohon induk betina dipanen tiga tandan buah kelapa (progeni)
dengan kisaran jumlah progeni per induk betina antara 2 sampai 12 buah. Progeni
buah kelapa normal langsung diisolasi DNA dari embrio zigotiknya. Semua
pohon-pohon dewasa yang berada di sekitar pohon induk betina dijadikan sebagai
calon tetua jantan untuk dijadikan donor serbuk sari. Satu tandan buah kelapa
(progeni) dipanen dari setiap induk betina yang terpilih dengan kisaran jumlah
progeni per induk betina antara 1 sampai 10 buah. Progeni dari setiap pohon induk
betina dipanen masing-masing satu tandan dengan jumlah total progeni 102 buah.
Progeni buah kelapa tadi langsung diisolasi dari embrio zigotiknya untuk
diperoleh DNA-nya.

53
Genotyping tetua dan progeni
Isolasi DNA dilaksanakan menggunakan metode CTAB (Rohde et al. 1995)
dengan modifikasi. Daun kelapa muda maupun embrio kelapa (0.3-0.4 g) digerus
dengan buffer lisis 2 ml, yang mengandung PVP 0.007 g dan 2-mercaptoetanol
sebanyak 10 μl. Hasil gerusan daun atau embrio diinkubasi dalam waterbath pada
suhu 65 °C selama 60 menit dan dihomogenkan menggunakan sentrifugasi
Eppendorf Centrifuge 5416 dengan kecepatan 11000 rpm selama 10 menit.
Supernatan dipindahkan ke tube eppendorf kemudian ditambahkan
kloroform:isoamil-alkohol (24:1) sebanyak volume supernatant lalu disentrifugasi
dengan kecepatan 11000 rpm selama 10 menit dan dipindahkan ke mikrotube
baru.
Supernatan dipindah ke tube eppendorf baru kemudian ditambahkan
isopropanol dingin (0.8 volume dari supernatan) dan sodium asetat (0.1 volume
supernatant). Setelah diinkubasi dalam freezer semalaman,suspensi kemudian
disentrifugasi dengan kecepatan 11000 rpm selama 10 menit hingga diperoleh
pellet DNA. Pellet DNA dicuci dengan menggunakan ethanol 70% dingin 500 μl.
lalu disentrifugasi dan dikeringkan. Pellet DNA diencerkan menggunakan
aquabidest sebagai suspensi DNA. Kontaminan RNA dihilangkan dengan
menggunakan RNase sesuai standar prosedur (Sambrook dan Russel 2001).
Primer SSR yang diseleksi dari 36 primer (Lebrun et al. 2001) (Lampiran 1)
diperoleh 6 primer yang polimorfik. Sebagai tambahan, empat lokus marka SNAP
berdasarkan keragaman sekuens nukleotida (gen SUS dan WRKY) (Lampiran 2)
juga digunakan untuk menganalisis seluruh tetua dan progeni. Amplifikasi PCR
dilakukan dengan menggunakan 2µl DNA working solution, 0.625 µl primer, 6.25
pcr mix (KAPA Biosystem), dan 3 µl ddH2O untuk setiap reaksi. Amplifikasi
dilakukan sebanyak 35 siklus, yang dimulai pada suhu 95 °C selama 3 menit
sebagai denaturasi awal, diikuti 95 °C selama 15 detik sebagai denaturasi siklus
pertama, penempelan primer spesifik - suhu disesuaikan dengan masing-masing
pasangan primer - selama 15 detik, pemanjangan primer pada suhu 72 °C selama
5 detik. Final extention 72 °C selama 10 menit sesuai rekomendasi kit KAPA
Biosystem.
Produk amplifikasi PCR untuk primer SSR dipisahkan dengan elektroforesis
gel poliakrilamid 6% menggunakan Buffer SB 1x (Brody dan Kern 2004) dan
pewarnaan gel dengan pewarna silver. Pewarnaan silver mengikuti metode Creste
et al. (2001) dengan modifikasi. Elektroforegram di visualisasi diatas meja UV
transluminesen, di foto menggunakan kamera digital. Pemberian skor setiap
genotype dilakukan juga di atas light table.
Produk amplifikasi PCR untuk primer SNAP dipisahkan menggunakan gel
agarose 1% menggunakan buffer TBE 1x dan pewarnaan menggunakan gel untuk
melihat hasil skoring yang dihasilkan (Sambrook and Russel 2001).
Elektroforegram di visualisasi diatas meja UV transluminesen di foto
menggunakan kamera digital. Pemberian skor setiap genotipe dilakukan untuk
setiap sampel.

54
Identifikasi Kandidat Tetua Jantan
Setiap progeni yang disampling sudah diketahui induk betinanya tetapi tidak
diketahui pohon yang menjadi donor serbuk sarinya. Kandidat tetua jantan bisa
menjadi salah satu induk betina dalam populasi pohon dewasa tersebut. Tahapan
ini dilakukan untuk memprediksi calon pendonor serbuk sari diantara calon tetua
jantan yang dievaluasi di setiap progeni.
Identifikasi donor serbuk sari dilakukan dengan menganalisis genotipe
progeni versus seluruh induk jantan yang dievaluasi. Semua induk jantan atau
induk betinanya dievaluasi dan berpotensi sebagai donor serbuk sari. Analisis
molekuler menggunakan software CERVUS analysis parentage program
komputer Cervus 2.0 (Marshall 1998). Hasil Cervus diperoleh data alel frekuensi,
PIC, heterozigositas, homozigositas. Dilanjutkan dengan data simulasi dan data
analisis tetua. Data analisis tetua akan diperoleh kandidat tetua dengan lambang
(*): artinya tingkat kepercayaan 95%. (+): tingkat kepercayaan 80%. dan (-):
tingkat kepercayaan < 80% .
Analisis Pola Persebaran Serbuk Sari
Lokasi induk betina dan tetua jantan terpilih hasil analisis Cervus
diplotkan menggunakan perangkat lunak pemetaan Garmin MapSource GPS versi
76C5x. Jarak antara induk betina dan tetua jantan dihitung menggunakan software
yang sama. Jarak dan posisi dari kedua induk betina dan tetua jantan yang
dihasilkan kemudian digunakan untuk mengilustrasikan pola persebaran serbuk
sari di lokasi penelitian.
Penyerbukan sendiri didefinisikan jika induk jantan yang teridentifikasi
sama dengan tetua betinanya. Selain itu semuanya didefinisikan sebagai
penyerbukan silang. Penyerbukan silang bisa dikelompokkan dengan kategori
penyerbukan silang genjah (induk betina genjah diserbuki oleh tetua jantan genjah
lainnya), penyerbukan silang dalam (induk betina dalam diserbuki oleh tetua
jantan dalam lainnya), penyerbukan silang hibrida ((induk betina hibrida diserbuki
oleh tetua jantan hibrida lainnya), penyerbukan silang antara induk betina genjah
dengan Dalam atau hibrida, atau sebaliknya. Kedua tipe persilangan dan
kuantitasnya semuanya dihitung.
Hasil dan Pembahasan
Pemetaan Populasi Tanaman
Peta lokasi penelitian dapat dilihat pada Gambar 5.1. Pohon kelapa
berbuah normal diberi warna merah, sedangkan untuk pohon kelapa berbuah
kopyor diberi warna hijau. jumlah total pohon Kopyor sebanyak 9 pohon (Dalam
kopyor 8 pohon dan Genjah kopyor 1 pohon) dan pohon normal sebanyak 33
pohon (Dalam normal 33 pohon dan Genjah normal 3). Yang dijadikan sebagai
pohon induk betina adalah semua pohon berbuah kopyor dengan pertimbangan
ingin melihat pengaruh pohon normal disekitar pohon berbuah kopyor tadi.

55
Gambar 5.1. Peta lokasi penelitian dengan tegakan kelapa kopyor pada desa
Dukuh Seti Kabupaten Pati Jawa Tengah. Simbol
mengidentifikasikan posisi ( ) induk betina Dalam kopyor, (….)
induk betina genjah kopyor, ( ) tetua jantan Dalam normal, ( )
tetua jantan Dalam kopyor, ( ) tetua jantan genjah normal, dan ( )
tetua jantan genjah kopyor
Tabel 5.1. Jumlah alel, heterozigositas observasi (O) dan ekspektasi (E) serta
kandungan informasi polimorfis (PIC) pada 10 lokus marka molekuler
hasil analisis Cervus pada populasi kelapa Dalam Pati
Nama lokus Jumlah Alel Jumlah Heterozigositas
PIC Het Hom O E
CnCir_87 2 64 80 0.444 0.496 0.372
CnCir_86 4 89 54 0.622 0.687 0.635
CnZ-18 4 116 28 0.806 0.741 0.691
CnZ_51 5 89 55 0.618 0.707 0.66
CnCir_B12 5 107 37 0.743 0.745 0.699
CnCir_56 5 81 63 0.563 0.695 0.647
CnSus1#14 2 39 100 0.281 0.252 0.22
CnSus1#3 2 96 43 0.691 0.483 0.365
WRKY19#1 2 116 24 0.829 0.5 0.374
WRKY 6#3 2 123 21 0.854 0.495 0.371 Keterangan: PIC= Polymorphic Information Content. Ho= Observed heterozigosity. He= Expected

56
Genotyping tetua dan progeni
Nilai PIC pada lokus-lokus dengan menggunakan marka SSR menunjukkan
nilai yang lebih tinggi dibandingkan dengan lokus-lokus dengan marka SNP
(Tabel 5.1). Semakin rendah tingkat polimorfisme, maka tingkat keragaman juga
semakin rendah. Hal ini disebabkan karena jumlah lokus pada marka SSR yang
digunakan lebih banyak dari pada jumlah lokus pada marka SNP. Nilai PIC
merupakan ukuran polimorfisme antar genotipe dalam suatu lokus yang
menggunakan informasi jumlah alel (Sajib et al. 2012).
Jumlah populasi homozigot dengan menggunakan lokus-lokus marka SSR
tergolong lebih rendah dibandingkan dengan lokus pada marka SNP. Demikian
juga nilai He pada lokus dengan marka SSR lebih tinggi dibandingkan dengan
lokus pada marka SNP. Alel-alel heterozigot banyak dijumpai pada marka SSR
menyebabkan nilai He lebih tinggi dibandingkan dengan marka SNP.Dari hasil
analisis cervus juga bisa diperoleh persentasi donor normal dan donor kopyor
yang menyerbuki pohon betina Dalam Kopyor yang ditunjukkan pada Tabel 5.2.
Pada penelitian ini juga memperlihatkan tidak adanya buah kopyor yang
bisa dipanen oleh petani. Hal ini mengindikasikan adanya dugaan bahwa kelapa
kopyor yang heterozigot Kk diserbuki oleh pohon normal KK. maka nilai harapan
produksi buah kopyornya menjadi nol dan semua buah yang dihasilkan
merupakan buah normal dengan genotipe embrio zigotik homozigot KK (50%)
atau heterozigot Kk (50%). Keberadaan pohon normal KK di sekitar pohon
kopyor heterozigot Kk dapat berpengaruh langsung terhadap fenotipe buah yang
dipanen sehingga dikategorikan sebagai efek xenia. Efek xenia ini bisa juga
dipelajari melalui analisis serbuk sari dengan mengidentifikasi tetua pendonor
polen yang terbaik untuk meningkatkan produksi buah. Hal ini juga dilakukan
oleh Olfati et al. (2010) dengan melihat efek xenia pada buah dan biji cucumber.
Sifat kopyor dikendalikan oleh satu lokus dengan alel resesif k. sedangkan
sifat normal dikendalikan oleh alel K. Pohon kelapa kopyor di lokasi penelitian
mempunyai genotipe heterozigot Kk. yang jika diserbuki oleh pohon kopyor
heterozigot Kk lainnya. menghasilkan nilai harapan 25% hasil buah kopyor
(genotipe embrio zigotik kk) dan 75% hasil buah normal (Sudarsono et al. 2014a)
Tabel 5.2. Skema persilangan dan tipe polinasi yang diidentifikasi berdasarkan
hasil analisis persebaran polen dari uji progeni pada populasi kelapa
Dalam Dukuh Seti, Pati
Skema polinasi Tipe polinasi Jumlah kejadian Persentase
Kopyor x Kopyor Self 3 3%
Kopyor x Kopyor Outcross 25 25%
Kopyor x Normal Outcross 74 73%
Total progeni 102 100.00%

57
Gambar 5.2. Jumlah progeni dipanen dan jumlah tetua jantan untuk tetua betina
tertentu populasi kelapa Dalam Dukuh Seti, Pati
Gambar 5.3. Jumlah serbuk sari yang didonasikan oleh tetua jantan terhadap
tetua betina tertentu populasi kelapa Dalam Dukuh Seti, Pati
Gambar 5.4. Jumlah polinasi setiap kelas jarak dari tetua kelapa jantan terpilih
terhadap tetua betina pada pertanaman kelapa kopyor populasi
Desa Dukuh Seti Kabupaten Pati Jawa Tengah
26
5
27
1
10
15 18 18
2
13
1
8 9 13
0
10
20
30
R1 R3 R6 R7 R10 R21 R32
4
11
4 3
1
4
1 2
0
5
10
15
1 2 3 4 5 6 7 8
5
21 30
15
22
5 1 0
0
10
20
30
0 < 10 < 20 < 30 < 40 < 50 < 60 > 60
Kelas Jarak antara tetua jantan terhadap betina (m)
Jum
lah p
eristiw
a
penyerb
ukan

58
Dari gambar 5.2. di atas terlihat bahwa jumlah progeni yang didapatkan
dari 7 tetua betina adalah sebanyak 102 progeni (batang hitam) dengan 64 pohon
tetua jantan (batang putih). Pada Gambar 5.3 menunjukkan terdapat 1-8 serbuk
sari yang dapat di donorkan oleh satu tetua jantan dengan frekuensi tertinggi
sebanyak 11 kali dengan 2 serbuk sari. Gambar 5.4 menunjukkan kelas jarak
polen dengan jarak terdekat 0 m (selfing) dan terjauh <60 m yaitu 54 m. Frekuensi
tertinggi terdapat pada jarak <20 m dengan 30 kali penyerbukan terjadi.
Analisis Pola Persebaran Serbuk Sari
Evaluasi pola persebaran polen dari tetua jantan yang sudah ditentukan
terhadap induk betina, posisi tetua jantan sebagai donor polen ke salah satu induk
betina di plotkan di peta menggunakan posisi GPS. Contoh gambaran dari posisi
tetua jantan yang sudah ditentukan terhadap induk betina resipien dipresentasikan
pada gambar 5.5 - 5.11.
Gambar 5.5. Pola persebaran serbuk sari kelapa kopyor Dukuh Seti dengan tetua
betina nomor R1. Simbol mengidentifikasikan posisi ( ) induk
betina Dalam kopyor, ( ) tetua jantan Dalam normal, ( ) tetua
jantan Dalam kopyor, dan ( ) tetua jantan genjah kopyor

59
Gambar 5.6. Pola persebaran serbuk sari kelapa kopyor Dukuh Seti dengan tetua
betina nomor R10. Simbol mengidentifikasikan posisi ( ) induk
betina Dalam kopyor tetapi sekaligus menjadi tetua jantan bagi
dirinya sendiri (selfing), ( ) tetua jantan Dalam normal dan ( )
tetua jantan Dalam kopyor.
Gambar 5.7. Pola persebaran serbuk sari kelapa kopyor Dukuh Seti dengan tetua
betina nomor R21. Simbol mengidentifikasikan posisi ( ) induk
betina Dalam kopyor, ( ) tetua jantan Dalam normal, ( ) tetua
jantan Dalam kopyor dan ( ) tetua jantan genjah normal.

60
Gambar 5.8. Pola persebaran serbuk sari kelapa kopyor Dukuh Seti dengan tetua
betina nomor R32. Simbol mengidentifikasikan posisi ( ) induk
betina Dalam kopyor tetapi sekaligus menjadi tetua jantan bagi
dirinya sendiri (selfing), ( ) tetua jantan Dalam normal, ( ) tetua
jantan Dalam kopyor dan ( ) tetua jantan genjah normal
Gambar 5.9. Pola persebaran serbuk sari kelapa kopyor Dukuh Seti dengan tetua
betina nomor R6. Simbol mengidentifikasikan posisi ( ) induk
betina Dalam kopyor tetapi sekaligus menjadi tetua jantan bagi
dirinya sendiri (selfing), ( ) tetua jantan Dalam normal, ( ) tetua
jantan Dalam kopyor, ( ) tetua jantan genjah normal, dan ( ) tetua
jantan genjah kopyor

61
Gambar 5.10. Pola persebaran serbuk sari kelapa kopyor Dukuh Seti dengan tetua
betina nomor R7. Simbol mengidentifikasikan posisi ( ) induk
betina genjah kopyor dan ( ) tetua jantan Dalam normal
Gambar 5.11. Pola persebaran serbuk sari kelapa kopyor Dukuh Seti dengan tetua
betina nomor R3. Simbol mengidentifikasikan posisi ( ) induk
betina Dalam kopyor tetapi sekaligus menjadi tetua jantan bagi
dirinya sendiri (selfing) dan ( ) tetua jantan Dalam normal.

62
Gambar 5.5-5.11 menunjukkan pola persebaran serbuk sari induk betina
kelapa Dalam kopyor memperlihatkan kecenderungan pohon pendonor serbuk sari
sebagian besar dari kelapa normal. Pengaruh negatif xenia ini terlihat pada
produktifitas buah kopyor dimana persentase buah yang dipanen semuanya
menjadi buah normal disebabkan populasi pohon kelapa normal (KK) lebih besar
di populasi tersebut. Sesuai dengan hasil penelitian Novarianto dan Lolong
(2012) bahwa dalam kondisi pertanaman campuran dengan sejumlah tanaman
kelapa normal rasio hanya menghasilkan rataan buah kopyor 15.8%.
Sehingga dalam rekomendasi penelitian Sudarsono et al (2014b) dalam
proyek Hi Link bahwa penebangan sebagian kelapa normal KK di antara
pertanaman kelapa Genjah Kopyor Pati Kk menyebabkan terjadinya peningkatan
persentase buah kopyor yang dipanen hingga 10%. Peningkatan persentase buah
kopyor yang dipanen diduga terkait dengan menurunnya pengaruh xenia pada
pertanaman akibat jumlah pohon normal KK yang semakin sedikit di lokasi
penelitian.
Populasi pertanaman yang cenderung kelapa Dalam juga terlihat 7 kali
terjadi persilangan sendiri (self pollination) berdasarkan hasil analisis cervus.
Kemungkinan ini juga bisa terjadi karena adanya tipe bunga kelapa dengan
morfologi terbuka. Xenia merupakan gejala genetik berupa pengaruh langsung
pollen pada fenotipe biji dan buah yang dihasilkan tetua betina.
Pada kajian pewarisan sifat. ekspresi dari gen yang dibawa tetua jantan dan
tetua betina diasumsikan baru diekspresikan pada generasi berikutnya. Dengan
adanya xenia, ekspresi gen yang dibawa tetua jantan secara dini sudah
diekspresikan pada organ tetua betina (buah) embrio dan/atau endosperm. Gejala
xenia tidak hanya mempengaruhi warna tetapi juga bentuk kadar gula, kadar
minyak, bentuk buah dan waktu pemasakan.
Xenia bukanlah penyimpangan dari Hukum Pewarisan Mendel melainkan
konsekuensi langsung dari pembuahan berganda (double fertilisation) yang terjadi
pada tumbuhan berbunga dan proses perkembangan embrio tumbuhan hingga biji
masak. Pada tahap perkembangan embrio sejumlah gen pada embrio dan
endosperm berekspresi dan mempengaruhi penampilan biji, bulir atau buah
(Denney 1992). Metaxenia tidak seperti xenia, tidak dapat dijelaskan dengan
elemen-elemen hereditas (kromosom) yang terbawa di dalam polen karena tidak
seperti kromosom yang terdapat pada jaringan yang menunjukkan pengaruh
langsung dari polen tetua (Bodor et al. 2008). Metaxenia menguraikan tentang pengaruh polen pada jaringan buah
maternal asal seperti pericarp dan komponen buah yang lain tidak dipengaruhi
oleh polen. Sedangkan xenia menguraikan tentang pengaruh polen pada jaringan
yang berisi sedikitnya satu satuan gen dari tetua jantan yakni embrio dan
endosperm (Ehlenfeldt 2003).
Simpulan
Pertanaman kelapa Dalam kopyor Pati di desa Dukuh Seti cenderung
mengalami pengaruh negatif xenia dengan keberadaan pohon-pohon kelapa
normal yang dalam jumlah lebih banyak di sekitar pohon kopyor. Selain itu,
pengaruh negatif xenia menyebabkan hasil buah kopyor yang dipanen hanya
sebesar 1% dan buah normal yang dipanen sebesar 99 % dari total buah yang

63
dipanen. Jarak penyebaran serbuk sari yang terdekat sejauh 0 m (self pollination)
dan terjauh sejauh 54 m. Frekuensi jarak penyebaran serbuk sari tertinggi terdapat
pada jarak < 20 m dengan sebanyak 30 kali penyerbukan. Persentase terjadinya
penyerbukan silang (outcrossing) sebesar 95% dan penyerbukan sendiri (selfing)
sebesar 5%.

64

65
VI. LEBAH MADU SEBAGAI POLINATOR MERUBAH POLA
PERSEBARAN SERBUK SARI KELAPA KOPYOR GENJAH PATI
Abstrak
Jarak dan arah persebaran serbuk sari dapat memberikan informasi tentang faktor
apa yang membantu penyerbukan kelapa kopyor di Pati. Analisis tetua jantan berdasarkan
enam lokus SSR dan empat lokus SNAP digunakan untuk melihat pola persebaran
serbuk sari kelapa kopyor (Cocos nucifera L.). Tujuan khusus dari penelitian ini adalah
untuk mengetahui (i) pola persebaran serbuk sari kelapa sebelum dan sesudah introduksi
lebah madu (ii) jarak distribusi serbuk sari kelapa sebelum dan sesudah introduksi lebah.
Enam lokus polimorfik marker SSR yang digunakan adalah CnCir_B12, CnCir_86,
CnCir_87, CnCir_56, CnZ 51, CnZ 18 dan empat lokus marker polimorfik SNAP yang
digunakan adalah CNSUS1Pos14. CNSUS1Pos3. WRKY6-Pos3 dan WRKY19-Pos1.
Sepuluh lokus marker tersebut digunakan untuk mengidentifikasi genotipe semua calon
tetua jantan, tetua betina dan semua progeninya. Hasil penelitian menunjukkan nilai PIC
dari marker yang dievaluasi setelah introduksi koloni lebah madu lebih tinggi
dibandingkan dengan sebelum introduksi. Jarak penyebaran serbuk sari terjauh setelah
introduksi koloni lebah madu sejauh 63 m dan sebelum introduksi sejauh 58 m.
Persentase terjadinya penyerbukan sendiri (selfing) sebelum introduksi lebah madu
sebesar 18% sedangkan setelah introduksi lebah madu sebesar 9.45%. Dengan demikian,
penyerbukan silang antar tanaman kelapa kopyor sesudah introduksi koloni lebah madu
lebih tinggi dibandingkan dengan sebelum introduksi.
Kata kunci : Lebah madu, polinator, persebaran polen, marka SSR, marka SNAP

66
VI. INTRODUCTION OF HONEY BEE POLLINATORS CHANGE THE
PATTERN OF POLLEN DISPERSAL AMONG PATI DWARF KOPYOR
COCONUTS
Abstract
Distance and direction of pollen spread can give information about the
factors assisting pollination in Pati kopyor coconut. Paternity analysis based on
six SSR markers and four SNAP marker loci were used to determine pattern of the
kopyor coconut (Cocos nucifera L.) pollen dispersal. The specific objectives of
this research are to evaluate: (i) the pattern of kopyor coconut pollen dispersals
before and after intoduction of honey bees colony in the coconut plantation and
(ii) the distribution of class distance among the female and the assigned male
coconut parents before and after the introduction of honey bee pollinators. The
evaluated SSR marker loci are CnCir_B12, CnCir_86, CnCir_87, CnCir_56, CnZ
51,CnZ 18 and the four SNAP marker loci are CNSUS1Pos14, CNSUS1Pos3,
WRKY6-Pos3 and WRKY19-Pos1. The markers were used to genotype all
candidate male parent, the female parent, and all the progeny arrays. Results of
the evaluation indicated PIC of the evaluated marker after honey bee colony
introduction is higher than before introduction. The longest pollen distance after
honey bee colony introduction is 63 m and before introduction is 58 m.
Percentage of self pollination before introduction of honey bee colony is 18%
while that after introduction is only 9.45%. Therefore, more outcrossing occurs
after introduction of honey bee colony into the kopyor coconut plantation.
.
Key Words: Honey bee, polinators, pollen dispersal, SSR marker, SNAP marker

67
Pendahuluan
Kelapa kopyor memiliki prospek yang cerah untuk dikembangkan sebagai
komoditas spesifik daerah karena mempunyai keunggulan kompetitif dibanding
kelapa normal. Selain itu. buah kopyor juga berpotensi sebagai komoditas ekspor.
Permintaan konsumen terhadap buah kopyor selalu tidak terpenuhi. karena
terbatasnya produksi buahnya. Berdasarkan hal tersebut di atas, agribisnis kelapa
kopyor saat ini menjadi sangat menjanjikan bagi petani. Pengembangan agribisnis
buah kelapa kopyor masih menghadapi beberapa kendala, di antaranya adalah
masih rendahnya produksi buah kopyor yang dihasilkan petani dari pohon kelapa
kopyornya. Hal ini terkait dengan jumlah buah kopyor yang dihasilkan setiap
pohon (Maskromo et al. 2007).
Data pertanaman kelapa kopyor di Kabupaten Pati diidentifikasi sebanyak
47 261 pohon yang dimiliki oleh 1 583 petani dengan produksi buah kopyor
296 279 butir pertahun (Anonim. 2004). Berarti produksi buah kopyor rata-rata
hanya 6 butir/pohon pertahun. Produksi buah kopyor tesebut masih sangat rendah
dibanding dengan potensi yang dimiliki oleh kelapa Genjah yang mampu
menghasilkan total buah perpohon80-120 butir pertahun. Jika produksi buah
kelapa Genjah rata-rata 50 butir/pohon/tahun. artinya produksi buah kopyor rata-
rata di Kabupaten Pati hanya 6/50×100%=12%. Walaupun pada beberapa pohon
terpilih ditemukan produksi buah kopyor antara 30-40%.Hal ini bisa terjadi karena
pohon kelapa kopyor alami (Genotipe heterozygot=Kk) tumbuh di antara pohon
kelapa bukan kopyor (Genotipe homozigot dominan=KK). Walaupun sifat kelapa
Genjah lebih dominan menyerbuk sendiri, tetapi peluang terjadi penyerbukan
silang masih cukup besar.
Salah satu faktor yang berpengaruh terhadap produktifitas pertanaman
kelapa kopyor adalah tingkat keberhasilan penyerbukan dan persentase bunga
betina yang berkembang menjadi buah dalam setiap tandan bunga. Vektor
persebaran serbuk sari dan biji terdiri atas vektor abiotik seperti olehangin
(anemochory), air (hydrochory), berat (barochory), dan vektor biotik yaitu
dibantu oleh hewan (zoochory) yang meliputi endozoochorous(setelah melalui
pencernaan) atau exozoochorous (tanpa melalui pencernaan). Keberhasilan
penyerbukan di kelapa sangat ditentukan oleh keberadaan serangga penyerbuk
(polinator) yang berkeliaran di tandan bunga tanaman kelapaterutama pada kelapa
tipe dalam. Dengan mengetahui faktor polinator juga dapat membantu dalam
mempelajari pola persebaran serbuk sari pada tanaman.
Produksi kelapa sawit di beberapa daerah di Indonesia masih belum
optimal meskipun kumbang penyerbuk untuk kelapa sawitElaeidobius
kamerunicusdidatangkan ke Indonesia sejak tahun 1982 (Sianturi 2001). Hal ini
disebabkan antara lain karena masih banyak bunga yang gagal diserbuki sehingga
buah kelapa sawit tidak berkembang. Agar jumlah buah kelapa sawit yang
berkembang semakin banyak, frekuensi penyerbukan perlu ditingkatkan dengan
cara meningkatkan jenis dan populasi serangga penyerbuknya.
Hasil eksplorasi Balai Penelitian Tanaman Kelapa dan Palma Lain Manado
(Balitka) di Kabupaten Pati, Jawa Tengah diperoleh adanya kelapa kopyor tipe
Genjah. Kelapa kopyor tipe Genjah memiliki beberapa keunggulan dibanding tipe
Dalam,yaitu berbuah lebih cepat pada usia 3-4 tahun setelah tanam,sedangkan
kelapa kopyor tipe Dalam berbuah pada umur 5-7 tahun setelah tanam. Kelapa

68
kopyor tipe Genjah mampu menghasilkan jumlah buah kopyor pertandan lebih
banyakbisa mencapai 50%, sedangkan tipe Dalam hanya 10% - 20% (Maskromo
dan Novarianto 2007). Peluang terjadinya sifat makapuno (kopyor) ini disebabkan
oleh penyerbukan antara bunga betina dan jantan yang membawa gen kopyor
(Santos 1999).
Penelitian persebaran serbuk sari pada kelapa sampai saat ini belum ada
dilaporkan sehingga dianggap penting untuk mempelajari persebaran serbuk sari
pada kelapa kopyor. Adapun tujuan spesifik penelitian ini untuk mengetahui (i)
pola persebaran serbuk sari pada kelapa sebelum dan sesudah introduksi pollinator
lebah madu (ii) jarak persebaran serbuk sari pada kelapa sebelum dan sesudah
introduksi polinator.
Bahan dan Metode
Waktu dan Lokasi Penelitian
Penelitian dilaksanakan mulai bulan Maret 2011 hingga Desember 2014.
Kegiatan lapang dilakukan di perkebunan kelapa kopyor di Desa Sambiroto
Kabupaten Pati, Jawa Tengah dengan lokasi GPS S6 27.649 E111 02.747. Tipe
tanah yang dievaluasi pada perkebunan kelapa kopyor merupakan tipe tanah
berpasir. Kegiatan penelitian dilakukan di Laboratorium Biologi Molekuler
Tanaman (PMB Lab) Departemen Agronomi dan Hortikultura Fakultas Pertanian
Institut Pertanian Bogor.
Introduksi Lebah Madu
Pertanaman lokasi penelitian seluas maksimum 0.5 ha diletakkan satu
kotak sarang lebah madu (Apis melifera) pada tahun 2012. Populasi sebelum
introduksi lebah dipanen pada tahun 2011 sementara untuk populasi setelah
introduksi lebah dipanen pada tahun 2014 .
Pemilihan Tetua dan Pemanenan Progeni
Populasi yang dipilih adalah area pertanaman di pekarangan rumah
penduduk dan merupakan campuran dari pohon kelapa kopyor heterozigot (Kk)
dengan pohon kelapa normal (KK) atau kelapa yang tidak pernah menghasilkan
buah kopyor. Populasi yang digunakan dalam analisis pola persebaran serbuk sari
sebanyak 95 pohon kelapa dewasa yang terdiri 68 pohon kelapa Genjah, 14 pohon
kelapa Dalam, dan 13 pohon kelapa Hibrida. Pohon kelapa yang ada di areal
pertanaman terdiri atas pohon kelapa berbuah normal sebanyak 22 pohon dan
pohon kelapa berbuah kopyor sebanyak 73 pohon. Semua pohon-pohon dewasa
yang berada di sekitar pohon induk betina dijadikan sebagai calon induk jantan
untuk dijadikan donor serbuk sari. Pemilihan pohon yang dijadikan sebagai induk
betina dilakukan dengan metode purpossive random sampling yang mewakili
masing-masing tipe kelapa Genjah, Dalam, Hibrida dan juga mewakili lokasi
didalam petak sampel (Gambar 2). Total progeni dari seluruh induk betina
sebanyak 127 buah. Satu tandan buah kelapa (progeni) dipanen dari setiap induk
betina yang terpilih dengan kisaran jumlah progeni per induk betina antara 2
sampai 10 buah.

69
Genotyping tetua dan progeni
Isolasi DNA dilaksanakan menggunakan metode CTAB (Rohde et al. 1995)
dengan modifikasi. Daun kelapa muda maupun embrio kelapa (0.3-0.4 g) digerus
dengan buffer lisis 2 ml, yang mengandung PVP 0.007 g dan 2-mercaptoetanol
sebanyak 10 μl. Hasil gerusan daun atau embrio diinkubasi dalam waterbath pada
suhu 65 °C selama 60 menit dan dihomogenkan menggunakan sentrifugasi
Eppendorf Centrifuge 5416 dengan kecepatan 11000 rpm selama 10 menit.
Supernatan dipindahkan ke tube eppendorf kemudian ditambahkan
kloroform:isoamil-alkohol (24:1) sebanyak volume supernatant lalu disentrifugasi
dengan kecepatan 11000 rpm selama 10 menit dan dipindahkan ke mikrotube
baru.
Supernatan dipindah ke tube eppendorf baru kemudian ditambahkan
isopropanol dingin (0.8 volume dari supernatan) dan sodium asetat (0.1 volume
supernatant). Setelah diinkubasi dalam freezer semalaman,suspensi kemudian
disentrifugasi dengan kecepatan 11000 rpm selama 10 menit hingga diperoleh
pellet DNA. Pellet DNA dicuci dengan menggunakan ethanol 70% dingin 500 μl.
lalu disentrifugasi dan dikeringkan. Pellet DNA diencerkan menggunakan
aquabidest sebagai suspensi DNA. Kontaminan RNA dihilangkan dengan
menggunakan RNase sesuai standar prosedur (Sambrook dan Russel 2001).
Primer SSR yang diseleksi dari 36 primer (Lebrun et al. 2001) (Lampiran 1)
diperoleh 6 primer yang polimorfik. Sebagai tambahan, empat lokus marka SNAP
berdasarkan keragaman sekuens nukleotida (gen SUS dan WRKY) (Lampiran 2)
juga digunakan untuk menganalisis seluruh tetua dan progeni. Amplifikasi PCR
dilakukan dengan menggunakan 2µl DNA working solution, 0.625 µl primer, 6.25
pcr mix (KAPA Biosystem), dan 3 µl ddH2O untuk setiap reaksi. Amplifikasi
dilakukan sebanyak 35 siklus, yang dimulai pada suhu 95 °C selama 3 menit
sebagai denaturasi awal, diikuti 95 °C selama 15 detik sebagai denaturasi siklus
pertama, penempelan primer spesifik - suhu disesuaikan dengan masing-masing
pasangan primer - selama 15 detik, pemanjangan primer pada suhu 72 °C selama
5 detik. Final extention 72 °C selama 10 menit sesuai rekomendasi kit KAPA
Biosystem.
Produk amplifikasi PCR untuk primer SSR dipisahkan dengan elektroforesis
gel poliakrilamid 6% menggunakan Buffer SB 1x (Brody dan Kern 2004) dan
pewarnaan gel dengan pewarna silver. Pewarnaan silver mengikuti metode Creste
et al. (2001) dengan modifikasi. Elektroforegram di visualisasi di atas meja UV
transluminesen di foto menggunakan kamera digital. Pemberian skor setiap
genotype dilakukan juga di atas light table.
Produk amplifikasi PCR untuk primer SNAP dipisahkan menggunakan gel
agarose 1% menggunakan buffer TBE 1x dan pewarnaan menggunakan gel untuk
melihat hasil skoring yang dihasilkan (Sambrook and Russel 2001).
Elektroforegram di visualisasi di atas meja UV transluminesen di foto
menggunakan kamera digital. Pemberian skor setiap genotipe dilakukan untuk
setiap sampel.
Identifikasi Kandidat Tetua Jantan
Setiap progeni yang disampling sudah diketahui induk betinanya tetapi tidak
diketahui pohon yang menjadi donor serbuk sarinya. Tahapan ini dilakukan untuk

70
memprediksi calon pendonor serbuk sari diantara calon tetua jantan yang
dievaluasi dalam percobaan.
Identifikasi donor serbuk sari dilakukan dengan menganalisis genotipe
progeni versus seluruh induk jantan yang dievaluasi. Semua induk jantan atau
induk betinanya dievaluasi dan berpotensi sebagai donor serbuk sari. Analisis
molekuler menggunakan software CERVUS analysis parentage program
komputer Cervus 2.0 (Marshall 1998). Hasil Cervus diperoleh data alel frekuensi,
PIC, heterozigositas, homozigositas. Dilanjutkan dengan data simulasi dan data
analisis tetua. Data analisis tetua akan diperoleh kandidat tetua dengan lambang
(*): artinya tingkat kepercayaan 95%. (+): tingkat kepercayaan 80%. dan (-):
tingkat kepercayaan < 80% .
Analisis Pola Persebaran Serbuk Sari
Data genotipe progeni yang telah diketahui induk betinanya dibandingkan
dengan data pohon dewasa yang berpotensi menjadi tetua jantan (kandidat tetua
jantan). Identitas induk jantan diketahui berdasarkan metode all parent with plus
LOD. Hasil analisis selanjutnya digunakan untuk menentukan identitas induk
jantan berdasarkan nilai LOD tertinggi. Jarak antara induk betina dengan induk
jantan yang teridentifikasi dihitung menggunakan perangkat lunak GPS. Jarak
tersebut dihitung dengan memanfaatkan data koordinat tiap pohon. Induk betina
dan induk jantan kemudian digambarkan ke dalam peta sesuai dengan lokasi
penelitian sehingga membentuk pola persebaran serbuk sari. Induk jantan yang
teridentifikasi sama dengan induk betinanya, maka diasumsikan terjadi
penyerbukan sendiri, sedangkan jika induk jantan yang teridentifikasi berbeda
dengan induk jantan maka diasumsikan terjadi penyerbukan silang.
Gambar 6.1. Posisi pengambilan sampel di pertanaman kelapa kopyor Desa
Tayu Kabupaten Pati Jawa Tengah. Garis merupakan batasan
populasi yang dianalisis. Gambar bendera menunjukkan sebaran
induk betina yang disampling. Simbol menunjukkan posisi ( ) :
Dalam normal, ( ) : Dalam kopyor, ( ) : Genjah normal, ( ) :
Genjah kopyor, ( ) : hibrida normal, ( ) : hibrida kopyor dan
( ) sarang koloni lebah madu.

71
Hasil dan Pembahasan
Pemetaan Populasi Tanaman
Peta lokasi penelitian dapat dilihat pada Gambar 6.1. Populasi tersebut
menggambarkan bahwa pohon kelapa berbuah normal masih banyak tersebar di
dalam populasi yang dipilih. Jumlah total pohon normal sebanyak 22 pohon
dimana Genjah Normal 14 pohon. Dalam Normal 3 pohon dan Hibrida Normal 5
pohon. Jumlah total pohon Kopyor sebanyak 73 pohon dimana Genjah Kopyor
54 pohon. Dalam Kopyor 11 pohon dan Hibrida Kopyor 8 pohon. Induk betina
yang digunakan sejumlah 15 pohon yang terdiri dari 2 pohon kelapa Hibrida, 6
pohon kelapa Genjah dan 7 pohon kelapa Dalam. Satu tandan buah kelapa
(progeni) dipanen dari setiap induk betina yang terpilih dengan kisaran jumlah
progeni per induk betina antara 2 sampai 10 buah.
Genotyping tetua dan progeni
Berdasarkan analisis molekuler nilai rata-rata PIC dengan marka
SSRmenunjukkan bahwa pada populasi sebelum introduksi lebahmemiliki nilai
rata-rata PIC lebih tinggi dibandingkan dengan populasi sesudah introduksi lebah
(Tabel 6.1). Begitu pula dengan nilai rata-rata PIC dengan marka SNP baik pada
populasi sebelum dan sesudah introduksi lebah memiliki nilai yang hampir sama.
Nilai PIC merupakan ukuran polimorfisme antar genotipe dalam suatu lokus yang
menggunakan informasi jumlah alel (Sajib et al. 2012). Nilai PIC pada lokus-
lokus dengan menggunakan marka SSR menunjukkan nilai yang lebih tinggi
dibandingkan dengan lokus-lokus dengan marka SNP.
Tabel 6.1. Jumlah individu heterozigot dan homozigot, heterozigositas observasi
dan harapan, dan Polymorphic Information content (PIC) di 10 lokus
marka molekuler kelapa Kopyor sebelum (BF) dan setelah (AF)
introduksi koloni lebah madu pada pertanaman kelapa kopyor
populasi Desa Tayu Kabupaten Pati Jawa Tengah
Nama lokus
Jumlah
heterozigot Jumlah homozigot
Heterozigositas
teramati PIC
BF AF BF AF BF AF BF AF
CnCir_87 27 54 152 174 0.151 0.237 0.311 0.327
CnCir_86 100 102 79 126 0.559 0.447 0.666 0.663
CnZ-18 74 92 105 136 0.413 0.404 0.565 0.544
CnZ_51 72 92 107 136 0.402 0.404 0.541 0.375
CnCir_B12 64 101 112 126 0.364 0.445 0.677 0.682
CnCir_56 78 111 101 117 0.436 0.487 0.565 0.627
CnSus1#14 143 146 36 80 0.799 0.646 0.369 0.36
CnSus1#3 147 184 27 41 0.845 0.818 0.374 0.372
WRKY19#1 111 183 65 44 0.631 0.806 0.357 0.369
WRKY 6#3 76 185 103 43 0.425 0.811 0.283 0.368

72
Jumlah individu heterosigot yang teranalisis oleh kedua marka SSR dan
SNP menunjukkan kecenderungan yang lebih sedikit dibandingkan dengan jumlah
individu homozigot. Hal ini bisa dilihat dari nilai He observasi yang rata-rata
nilainya dibawah 0.5. Nilai He yang rendah menunjukkan bahwa kelapa-kelapa
yang dianalisis banyak terbentuk oleh hasil penyerbukan sendiri, karena gen-gen
terfiksasi/terkumpul dalam satu populasi hasil penyerbukan selfing atau dari polen
tanaman dengan sifat yang sama. Nilai PIC kedua marka juga menunjukkan nilai
yang rendah dengan rata-rata nilai dibawah 0.5 yang berarti bahwa tingkat
keragaman populasi kelapa-kelapa yang diamati rendah.
Jumlah individu yang homozigot pada lokus-lokus marka SSR tergolong
lebih tinggi dibandingkan dengan lokus pada marka SNP yang terlihat pada
rendahnya nilai heterosigositas (He). Rendahnya nilai He menunjukkan bahwa
pada lokus-lokus tersebut genotipe yang diamati mayoritas memiliki alel-alel
homozigot. Alel-alel heterozigot lebih banyak dijumpai pada marka SSR
menyebabkan nilai He SSR lebih tinggi dibandingkan dengan marka SNP,
sehingga menyebabkan rata-rata nilai PIC pada marka SSR lebih tinggi di
bandingkan dengan marka SNP yang membuktikan bahwa marka SSR lebih
efektif dalam menujukkan keragaman dalam suatu populasi.
Jumlah individu Heterozigot dan Homozigot pada populasi sebelum
introduksi lebah (BF) cenderung lebih rendah dari pada populasi setelah
introduksi lebah (AF). Dengan nilai heterosigositas obeservasi yang rata-rata
cenderung lebih tinggi pada setelah introduksi lebah. Hal ini menunjukkan bahwa
setelah introduksi lebah, populasi buah yang dihasilkan lebih banyak walaupun
buah yang dihasilkan tidak benyak berbuah kopyor seperti yang dikehendaki.
Nilai PIC pada kedua populasi menunjukkan nilai rata-rata yang lebih tinggi pada
sebelum introduksi lebah. Hal ini bisa dijelaskan karena pola hidup lebah yang
cenderung mencari makanan hanya disekitar pohon-pohon induk betina kelapa.
Perubahan parameter yang terjadi pada kedua populasi sebelum dan sesudah
introduksi lebah terjadi hanya pada jumlah rata-rata alel/lokus yang meningkat
2,85%, sedangkan pada parameter yang lain semuanya tetap (Tabel 6.2).
Identifikasi Kandidat Tetua Jantan
Hasil analisis tetua menunjukkan bahwapada populasi sebelum dan sesudah
introduksi lebah berturut turut terdapat 18dan 47simbol (+) yang menunjukkan
bahwa 80% yakin bahwa kandidat tetua tersebut yang menjadi tetua jantan.
Simbol (*) ditemukan sebanyak 3dan 13 pada kedua populasi yang menunjukkan
bahwa 95% yakin bahwa sebagai tetua jantan. dan simbol (-) sebanyak 84dan 123
pada kedua populasi yang menunjukkan dibawah 80% diduga sebagai tetua
jantan.
Seluruh nilai LOD menunjukkan tidak terdapat nilai negatif. sehingga
simbol (-) diyakini sebagai jantan walaupun tingkat kepercayaan dibawah 80%.
Nilai peluang kemungkinan atau LOD (Likelihood of odds) menunjukkan nilai
lebih besar dari 0 dan positif. nilai LOD positif lebih mengindikasikan bahwa
tetua jantan yang diduga adalah benar. Semakin besar nilai LOD maka semakin
besar peluang tetua tersebut merupakan tetua yang sebenarnya (Marshal et al.
1998). Pada populasi tanpa introduksi lebah menunjukkan kemampuan menduga
kandidat tetua jantan lebih rendah dibandingkan dengan populasi introduksi lebah.
Pada populasi setelah introduksi hampir 50% menunjukkan tingkat kepercayaan

73
lebih dari 80% untuk menduga kandidat tetua jantan dengan benar. dibandingkan
dengan populasi sebelum introduksi lebah. Hal ini menunjukkan bahwa dengan
introduksi lebah persebaran polen lebih terkontrol sehingga saat dilakukan analisis
dalam pendugaan kandidat tetua jantan lebih akurat dibandingkan tanpa adanya
introduksi lebah. Populasi sebelum introduksi memiliki tingkat kepercayaan lebih
rendah dalam menduga kandidat tetua jatan.Hal ini diduga karena penyerbukan
yang tidak terkontrol dan banyak kontaminasi dari polen pohon lain yang tidak
digunakan dalam kandidat tetua jantan.
Tabel 6.2. Perubahan parameter populasi sebelum dan setelah introduksi koloni
lebah madu pada pertanaman kelapa Kopyor di populasi Desa Tayu
Kabupaten Pati Jawa Tengah
Parameter Rata-rata
Perubahan parameter yang berhubungan dengan
introduksi lebah madu
Sebelum Sesudah
Jumlah rata-rata alel/lokus 3.4 3.6
Heterozigositas ekspektasi 0.55 0.55
PIC 0.47 0.47
Total exclusionary power:
- Tetua Pertama 0.85 0.85
- Tetua Kedua 0.97 0.97
Diagram batang jumlah tetua jantan yang berkontribusi terhadap betina
terpilih pada populasi kelapa yang diuji menunjukkan bahwa introduksi lebah
mampu meningkatkan jumlah polen tetua jantan yang menyerbuk yaitu dari 83
menjadi 127 peristiwa penyerbukan (Gambar 6.2). Hal ini dapat dijelaskan karena
sifat lebah yang lebih senang mengumpul pada populasi tanaman betina dan
membentuk koloninya disekitar tanaman tersebut, sehingga menyebabkan
tingginya peristiwa penyerbukan pada tanaman kelapa betina.
Jumlah serbuk sari yang didonorkan dari tetua jantan juga mengalami
peningkatan setelah dilakukan introduksi lebah. Frekuensi penyerbukan oleh tetua
jantan sebelum introduksi lebah maksimal hanya mampu mendonorkan 5 serbuk
sari, dengan adanya introduksi lebah dapat meningkatkan frekuensi penyerbukan
pollen dari tetua jantan yang sama sampai 7 serbuk sari. Terdapat 1 donor yang
berkontribusi 7 serbuk sari dan 4 donor yang berkontribusi 6 serbuk sari (Gambar
6.2).
Diagram batang jumlah tetua betina penerima donor dari tetua jantan
terpilih pada populasi kelapa yang diuji menunjukkan bahwa introduksi lebah
dapat meningkatkan jumlah polen yang menyerbuki betina. Dari diagram terlihat
bahwa pada sebelum introduksi lebah jumlah serbuk sari yang bisa di donorkan
oleh satu tetua jantan maksimal 7 serbuk sari, namun setelah dilakukan introduksi
lebah terdapat satu tetua jantan yang dapat menyerbuki 9 sampai 10 serbuk sari ke
satu betina(Gambar 6.3).

74
Gambar 6.2. Frekuensi tetua jantan yang mengkontribusikan serbuk sari terhadap
tetua betina yang diuji sebelum (atas) dan setelah (bawah)
introduksi koloni lebah madu pada pertanaman kelapa Kopyor
populasi Desa Tayu Kabupaten Pati Jawa Tengah
Jum
lah t
etua
janta
n y
ang
ber
kontr
ibusi
Jumlah polen donor ke tetua betina
27
11
7
2 1 0
10
20
30
40
1 2 3 4 5 6 7
Jum
lah t
etua
janta
n y
ang b
erkontr
ibusi
Jumlah polen donor ke tetua betina
20
14
7 3 3 4
1 0
10
20
30
40
1 2 3 4 5 6 7

75
Gambar 6.3. Jumlah tetua betina penerima donor serbuk sari dari tetua jantan
terpilih sebelum (atas) dan setelah (bawah) introduksi lebah madu
pada pertanaman kelapa Kopyor populasi Desa Tayu Kabupaten
Pati Jawa Tengah
Berdasarkan diagram kelas jarak dari tetua kelapa jantan terpilih terhadap
tetua betina sebelum dan sesudah introduksi lebah menunjukkan bahwa introduksi
lebah dapat meningkatkan jarak dan frekuensi persebaran polen. Jarak persebaran
polen yang mampu disebarkan oleh lebah hingga >60 m dengan 2 serbuk sari.
Sedangkan frekuensi penyerbukan pada introduksi lebah cenderung mengalami
peningkatan, dimana jarak dengan frekuensi tinggi yaitu berkisar 0-50 m, dengan
frekuensi tertinggi 36 serbuk sari pada jarak <20 m (Gambar 6.4).
1
3
1
4
3
1
2
0
2
4
6
1 2 3 4 5 6 7 8 9 10
Jum
lah
tet
ua
bet
ina
Jumlah tetua jantan donor
1
3 3
4
3
1 1 1
0
2
4
6
1 2 3 4 5 6 7 8 9 10
Jum
lah
tet
ua
bet
ina
Jumlah tetua jantan donor

76
Gambar 6.4. Jumlah penyerbukan setiap kelas jarak dari tetua kelapa jantan
terpilih terhadap tetua kelapa betina sebelum (atas) dan setelah
(bawah) introduksi koloni lebah madu pada pertanaman kelapa
kopyor populasi Desa Tayu Kabupaten Pati Jawa Tengah
Analisis Pola Persebaran Serbuk Sari
Beberapa pola penyerbukan pada pohon induk kelapa Genjah kopyor
sebelum dan sesudah introduksi lebah ditunjukkan pada gambar 6.5 – 6.10 dan
Gambar lampiran 17-27. Penyerbukan dengan jarak yang dekat diduga dibantu
oleh angin sedangkan jarak yang jauh dibantu oleh serangga (Boer 2007). Jarak
antar pohon memberikan pengaruh terhadap frekuensi penyerbukan. jarak yang
paling dekat (0-10 m) memiliki persentase kontribusi serbuk sari yang paling
tinggi. Hal tersebut disebabkan karena apabila penyerbukan dibantu oleh angin
maka perbedaan tinggi pohon pendonor serbuk sari yang lebih tinggi dibanding
pohon penerima serbuk sari mengakibatkan jarak edar serbuk sari bisa lebih jauh.
Selain itu. dengan data tersebut dapat diketahui bahwa yang berperan lebih
banyak dalam penyerbukan kelapa kopyor di Pati adalah faktor polinator biologis
yaitu lebah. Perlakuan setelah introduksi lebah memperlihatkan tipe penyerbukan
selfing yang terjadi sebanyak 15% dan jarak edar serbuk sari paling jauh adalah
83 km, kemungkinan disebabkan karena lebah mampu lebih jauh membawa
serbuk sari dibandingkan angin.
Jum
lah p
eristiw
a
penyerb
ukan
Kelas Jarak antara tetua jantan terhadap betina (m)
15
6 13
18 12 12
7 0
0
10
20
30
40
50
0 < 10 < 20 < 30 < 40 < 50 < 60 > 60
Kelas jarak antara tetua jantan terhadap betina (m)
Ju
mla
h p
eri
stiw
a
pe
nye
rbu
kan
12 14
36
25 20
16
2 2 0
10
20
30
40
50
0 < 10 < 20 < 30 < 40 < 50 < 60 > 60

77
Gambar 6.5. Pola pergerakan serbuk sari terhadap tetua betina #059 diduga
berdasarkan analisis parentage sebelum (atas) dan setelah (bawah)
introduksi koloni lebah madu pada pertanaman kelapa Kopyor
Tayu Kabupaten Pati. Simbol mengidentifikasikan posisi ( )
Genjah Kopyor, ( ) Hibrida Kopyor, ( ) Dalam Kopyor sebagai
tetua jantan pendonor serbuk sari, dan ( ) Hibrida Kopyor #059
sebagai tetua jantan terpilih (donor serbuk sari) dan resipien betina
(penyerbukan sendiri). Simbol ( ) menunjukkan posisi sarang
koloni lebah madu.

78
Gambar 6.6. Pola pergerakan serbuk sari terhadap tetua betina #067 diduga
berdasarkan analisis parentage sebelum (atas) dan setelah (bawah)
introduksi koloni lebah madu pada pertanaman kelapa Kopyor
Tayu Kabupaten Pati. Simbol mengidentifikasikan posisi ( )
Genjah Kopyor, ( ) Hibrida Kopyor, ( ) Dalam Kopyor sebagai
tetua jantan pendonor serbuk sari terpilih, dan ( ) Genjah Kopyor
#067 sebagai resipien betina. Simbol ( ) menunjukkan posisi
sarang koloni lebah madu

79
Gambar 6.7. Pola pergerakan serbuk sari terhadap tetua betina #069 diduga
berdasarkan analisis parentage sebelum (atas) dan setelah (bawah)
introduksi koloni lebah madu pada pertanaman kelapa Kopyor
Tayu Kabupaten Pati. Simbol mengidentifikasikan posisi ( )
Genjah Kopyor, ( ) Hibrida Kopyor, ( ) Dalam Kopyor sebagai
tetua jantan pendonor serbuk sari terpilih, dan ( ) Genjah Kopyor
#069 sebagai resipien betina. Simbol ( ) menunjukkan posisi
sarang koloni lebah madu

80
Gambar 6.8. Pola pergerakan serbuk sari terhadap tetua betina #084 diduga
berdasarkan analisis parentage sebelum (atas) dan setelah (bawah)
introduksi koloni lebah madu pada pertanaman kelapa Kopyor
Tayu Kabupaten Pati. Simbol mengidentifikasikan posisi ( )
Genjah Kopyor, ( ) Hibrida Kopyor, ( ) Dalam Kopyor sebagai
tetua jantan pendonor serbuk sari terpilih, dan ( ) Genjah Kopyor
#084 sebagai resipien betina. Simbol ( ) menunjukkan posisi
sarang koloni lebah madu

81
Gambar 6.9. Pola pergerakan serbuk sari terhadap tetua betina #089 diduga
berdasarkan analisis parentage sebelum (atas) dan setelah (bawah)
introduksi koloni lebah madu pada pertanaman kelapa Kopyor Tayu
Kabupaten Pati. Simbol mengidentifikasikan posisi ( ) Genjah
Kopyor, ( ) Dalam Kopyor sebagai tetua jantan pendonor serbuk
sari terpilih, dan ( ) Dalam Kopyor #089 sebagai resipien betina.
Simbol ( ) menunjukkan posisi sarang koloni lebah madu
Kondisi pertanaman di lapang juga memiliki jarak tanam yang berdekatan
sehingga memungkinkan bagi tanaman kelapa genjah untuk melakukan
outcrossing dengan tanaman genjah lainnya. Penelitian Perera et.al (2010)
menyatakan bahwa meskipun bunga jantan dan betina berada pada posisi
berdekatan pada satu bunga akan tetapi tetap berpeluang untuk outcrossing.

82
Gambar 6.10. Pola pergerakan serbuk sari terhadap tetua betina #092 diduga
berdasarkan analisis parentage sebelum (atas) dan setelah
(bawah) introduksi koloni lebah madu pada pertanaman kelapa
Kopyor Tayu Kabupaten Pati. Simbol mengidentifikasikan posisi
( ) Genjah Kopyor sebagai tetua jantan pendonor serbuk sari, dan
( ) Genjah Kopyor #092 sebagai resipien betina. Simbol ( )
menunjukkan posisi sarang koloni lebah madu
Varietas kelapa dalam Srilangka adalah menyerbuk silang (penyerbukan terbuka)
dimana serbuk sari matang dan terlepas dari bunga jantan sebelum bunga betina
siap untuk dikawini (reseptif).
Untuk populasi sebelum introduksi lebah jarak persebaran serbuk sari
terjauh adalah 58 m sedangkan populasi setelah introduksi lebah jarak persebaran
serbuk sari terjauh adalah 63 m. Untuk jarak yang terdekat adalah 0 m atau

83
penyerbukan sendiri pada kedua populasi. Penyerbukan paling banyak terjadi
dalam rentang jarak <30 m untuk populasi sebelum introduksi lebah yaitu 18 even
penyerbukan, untuk populasi setelah introduksi lebah penyerbukan paling sering
terjadi pada rentang jarak <20 m yaitu 36 even (Gambar 6.4)
Dalam mencari pakan, lebah mengunjungi berbagai kuntum bunga untuk
mencari nektar atau serbuk sari bunga. Ketika mengambil serbuk sari bunga,
secara tidak sengaja butir-butir serbuk sari bunga tersebut menempel pada bulu-
bulu yang ada di seluruh tubuh lebah dan terbawa pergi oleh lebah. Apabila lebah
madu tersebut hinggap pada bunga yang lain maka butir-butir serbuk sari bunga
yang menempel pada tubuh lebah itu dapat menempel pada kepala putik bunga
yang siap mengalami penyerbukan. Kepala putik yang siap mengalami
penyerbukan mempunyai substansi tertentu untuk mengikat serbuk sari bunga
yang jatuh diatasnya. Sehingga terjadilah penyerbukan yang akan dilanjutkan
dengan proses pembuahan. Apabila lebah madu tersebut terbang pada jenis bunga
yang berbeda maka akan terjadi penyerbukan silang. Kemampuan untuk
melakukan penyerbukan silang ini sangat tinggi dibandingkan dengan jenis
serangga lain (Suryanarayana et al. 1992). Lebah merupakan polinator terbaik
yang dapat meningkatkan produktivitas tanaman. Misalnya pada tanaman jarak
pagar, lebah dapat meningkatkan produksi sebesar 40% (Kasno et al. 2010).
Simpulan
Nilai PIC rata-rata yang terbentuk pada populasi setelah introduksi lebah
sedikit lebih tinggi dibandingkan sebelum introduksi lebah. Jarak persebaran
polen lebih jauh setelah introduksi lebah (63 m) dibandingkan sebelum introduksi
lebah (58 m). Persentase selfing lebih tinggi terjadi sebelum introduksi lebah
(18%) dibandingkan setelah introduksi lebah (9,45%). Persentase outcrossing
pada populasi sebelum introduksi lebah sebesar 82% dan setelah introduksi lebah
menjadi 90,55%.

84

85
VII. PEMBAHASAN UMUM
Penggunaan analisis penyebaran polen pada sistem perkawinan baik pada
hewan maupun tumbuhan sangat efektif untuk melihat tipe persilangan,
inbreeding depression dan mempelajari evolusi biologi dalam suatu populasi
(Auld dan Rafael 2013). Populasi kelapa kopyor merupakan salah satu tanaman
dari family Arecaceae yang sangat menarik untuk dipelajari system
perkawinannya. Kelapa terdiri atas dua tipe pohon yaitu tipe Genjah dan tipe
Dalam yang berbeda pada performa (tinggi tanaman) dan umur berbuah tanaman.
Tanaman kelapa memiliki bunga jantan dan betina yang terpisah dan matang pada
waktu yang berbeda yang menyebabkan tanaman kelapa umumnya menyerbuk
silang.
Kelapa Dalam, kelapa Genjah dan kelapa Hibrida saling melakukan
penyerbukan silang satu sama lain dan tidak semua kelapa Dalam melakukan
penyerbukan silang. Begitupun dengan kelapa Genjah tidak semuanya melakukan
penyerbukan sendiri. Pada penelitian yang dilakukan pada percobaan 1 pada
populasi tanaman kelapa tipe Genjah terlihat terjadi selfing sebanyak 15 kali
(17,9%) dan jumlah outcrossing yang terjadi sebanyak 69 kali (82.1%). Pada
percobaan 2 yang dilakukan pada kelapa tipe Dalam terlihat terjadi selfing
sebanyak 1 kali (2%) dan jumlah outcrossing yang terjadi sebanyak 48 kali (98%).
Hal tersebut diduga disebabkan karena morfologi bunga kelapa yang terbuka,
sehingga kelapa Genjah juga memiliki peluang untuk menyerbuk silang
(Maskromo et al. 2011). Selain itu kondisi pertanaman di lapang juga memiliki
jarak tanam yang berdekatan sehingga memungkinkan bagi tanaman kelapa
Genjah untuk melakukan outcrossing dengan tanaman Genjah lainnya. Waktu
mekar bunga jantan (18-20 hari) dan betina (4-7 hari) yang berbeda juga dapat
meningkatkan besarnya penyerbukan silang (outcrossing) pada kedua tipe kelapa.
Adanya tanaman kelapa berbuah normal diantara tanaman kelapa berbuah
kopyor disebabkan oleh proses seleksi bibit yang belum dapat dipisahkan antara
bibit yang dapat berbuah kopyor dengan bibit kelapa normal pada saat penanaman
di lapang. Hal ini dibuktikan dengan masih banyaknya tanaman kelapa Dalam
normal yang ditemukan di lokasi pertanaman untuk tanaman berbuah kopyor.
Bibit kopyor alami yang ditanam petani berasal dari tanaman kelapa Dalam
kopyor heterozigot (Kk) yang secara teori membawa gen kopyor (k) namun juga
memiliki gen normal (K). Pada saat pembungaan, tanaman kopyor heterozigot
tersebut akan bersegregasi sehingga pada saat pembuahan berpeluang terbentuk
buah dengan genotipe (kk) dengan fenotipe endosperm kopyor yang embrionya
tidak dapat tumbuh seperti embrio pada buah kelapa normal. Pada tandan buah
yang sama berpeluang terbentuk buah normal dengan genotipe (Kk) dengan
fenotipe endosperm normal namun membawa gen kopyor (k). Dengan prinsip
memiliki peluang bergabung secara bebas saat proses meosis seperti pada Hukum
Mendel II, maka pada tandan buah yang sama akan terbentuk juga buah genotipe
(KK) dengan fenotipe endosperm dan embrionya tidak membawa gen kopyor (k).
Buah dengan genotipe KK akan menghasilkan tanaman yang tidak akan
menghasilkan tanaman berbuah kopyor atau disebut kelapa berbuah sayur
(Sudarsono et al. 2014b).

86
Fenomena pembentukan buah kelapa kopyor dan kelapa normal pada
pertanaman kelapa campuran disebabkan adanya efek xenia. Xenia merupakan
pengaruh langsung serbuk sari (pollen) pada fenotipe biji dan buah yang
dihasilkan tetua betina. Peristiwa xenia pada kelapa kopyor memberikan efek
negatif untuk pembentukan endosperma pertanaman kelapa kopyor pada
umumnya di populasi Dukuh Seti. Hal ini terlihat dari banyaknya donor serbuk
sari yang disumbangkan oleh tetua jantan kelapa normal terhadap induk betina
kopyor. Sehingga endosperma yang terbentuk semuanya menghasilkan
endosperma yang normal. Seperti yang dijelaskan pada laporan Hi Link 2014
(Sudarsono et al. 2014) bahwa buah kelapa dengan daging buah (endosperma)
yang keras dapat dihasilkan dari pohon kelapa normal KK (homosigot KK) atau
dari pohon kelapa kopyor Kk (heterosigot Kk). Buah kelapa ini mempunyai
embrio sigotik dengan konstitusi genetik KK dan Kk, daging buah (endosperma)
yang dihasilkan dengan konstitusi genetik KKK, Kkk atau KKk, jika diserbuki
oleh serbuk sari dari pohon kopyor Kk atau kk; atau dari pohon kelapa kopyor
Kk (heterosigot Kk). Buah kelapa dengan daging buah (endosperma) yang lembut
dan terlepas dari batok (cangkang)-nya, yang dalam kondisi tertentu dapat
dihasilkan dari pohon kelapa kopyor Kk (heterosigot Kk) – jika diserbuki oleh
serbuk sari dari pohon kelapa kopyor Kk (heterosigot Kk) atau kelapa kopyor kk
(homosigot kk); atau dari pohon kelapa kopyor kk (homosigot kk) - jika
diserbuki oleh serbuk sari dari pohon kelapa kopyor Kk; atau kelapa kopyor kk.
Buah kelapa kopyor kk (homosigot Kk) mempunyai embrio sigotik dengan
konstitusi genetik kk dan daging buah (endosperma) dengan konstitusi genetik
kkk.
Salah satu faktor yang berpengaruh terhadap produktifitas pohon kelapa
kopyor secara umum ditentukan tingkat keberhasilan penyerbukan dan persentase
bunga betina yang berkembang menjadi buah per tandan. Tingkat keberhasilan
penyerbukan pada kelapa salah satunya ditentukan oleh keberadaan serangga
polinator terutama pada kelapa tipe dalam. Berdasarkan pengamatan di lapangan,
tandan kelapa yang banyak dikunjungi oleh serangga penyerbuk umumnya
mempunyai tingkat keberhasilan penyerbukan yang tinggi. Hal ini ditunjukkan
dari jumlah buah total yang dipanen lebih banyak. Sebaliknya, tandan dengan
jumlah buah total yang dipanen rendah tetapi mempunyai jumlah betina yang
banyak, mengindikasikan rendahnya tingkat keberhasilan penyerbukan pada
tandan tersebut. Di lapangan, juga teramati banyaknya populasi lebah madu lokal
(Apis serana) yang terbang mengerumuni tandan yang sedang mekar.
Bantuan serangga sebagai polinator penyerbuk memanfaatkan infloresens
bunga untuk melengkapi aktivitas hidupnya seperti melakukan perkawinan,
meletakkan telur, dan mencari makanan sehingga aktivitas tersebut secara tidak
langsung menyebabkan terjadinya penyerbukan silang (Bown 1988). Lebih lanjut
Bown (1988) mengemukakan bahwa terdapat beberapa faktor yang merangsang
serangga untuk hinggap pada infloresens bunga antara lain bau atau aroma yang
dihasilkan oleh infloresens bunga yang terbuka seperti pada bunga tanaman
kelapa. Pada kebanyakan tanaman araceae, dihasilkan kombinasi senyawa berupa
amonia, protein, dan asam amino yang membuat serangga untuk mendatangi
organ reproduksi bunga. Berdasarkan hal tersebut, usaha untuk menjamin tingkat
keberhasilan penyerbukan yang tinggi diduga dapat dilakukan dengan
mengintroduksikan koloni lebah di pertanaman kelapa kopyor. Dengan

87
menerapkan budidaya lebah madu lokal di sekitar pertanaman kelapa kopyor,
diharapkan akan dapat mengatasi permasalahan rendahnya keberhasilan
penyerbukan sehingga meningkatkan jumlah buah kelapa total yang dipanen per
tandan dan meningkatkan produktifitas kelapa kopyor di lapangan.

88

89
VIII. SIMPULAN
1. Serbuk sari yang didonasikan pada populasi pertanaman kelapa kopyor
tipe Genjah Pati berasal dari serbuk sari donor yang berada dalam kisaran
jarak tempuh 0-58 meter dari tetua betina yang dievaluasi. Oleh karena itu,
selain dengan adanya keberadaan angin, polinator serangga juga dapat
berperan penting dalam polinasi kelapa Kopyor.
2. Populasi pertanaman kelapa kopyor tipe Dalam Kalianda hasil analisis
penyebaran serbuk sari menunjukkan persentase terjadinya outcrossing
sebesar 98% dan persentase selfing sebesar 2%. Hal ini mengindikasikan
bahwa kelapa kopyor tipe Dalam memiliki tipe penyerbukan yang
menyerbuk silang.
3. Kelapa Dalam tidak dianjurkan untuk dijadikan sebagai pendonor serbuk
sari dalam populasi pertanaman kelapa kopyor karena akan menyebabkan
menurunnya produksi buah kelapa kopyor yang disebabkan adanya efek
xenia.
4. Lebah madu efektif dijadikan sebagai polinator dalam pertanaman kelapa
kopyor karena dapat meningkatkan produksi buah dengan meningkatkan
jarak persebaran polen yang lebih jauh dibandingkan dengan populasi
sebelum introduksi lebah.

90

91
DAFTAR PUSTAKA
Amar Mohamed Hamdy, Manosh Kumar Biswas, Zongwen Zhang, Wen-Wu
Guo. 2011. Exploitation of SSR, SRAP and CAPS-SNP markers for
genetic diversity of Citrus germplasm collection. Scientia Horticulturae.
128 : 220-227.
Aitken N, Smith S, Schwarz C, Morin PA. 2009. Single nucleotide polymorphism
(SNP) discovery in mammals: a targeted-gene approach. Mol Ecol.
13:1423-1431.
Auld., Josh R, Rafael Rubio de Casas. 2013. The correlated evolution of dispersal
and mating-system traits. Evol Biol (2013) 40:185–193.
Austerlitz F, Dick CW, Klein EK, Muratorio SO, Smouse PE, Sork VL. 2004.
Using a genetic markers to estimate the pollen dispersal curve. Molecular
ecology. 13:937-954.
Blair AW, Williamson PS. 2009. Pollen dispersal in a star cactus (Astrophytum
asterias). Journal of Arid Environments. 74:525-527.
Boer D. 2007. Keragaman dan struktur genetik populasi jati sulawesi tenggara
berdasarkan marka mikrosatelit [disertasi]. Bogor (ID): Institut Pertanian
Bogor.
Brody Jonathan R, Scott E Kern. 2004. History and principles of conductive
media for standard DNA electrophoresis. Analytical Biochemistry 333 (1):
1–13.
Brumfield RT, Beerli P, Nickerson DA, Edwards SV. 2003. The utility of single
nucleotide polymorphisms in inferences of population history. Trends Ecol
Evol. 18:249-256.
Bown D. 1988. Aroid: Plants of the arum family. Oregon (US): Timber Press
Portland.
Bulant C, Gallais A., Matthys-Rochon E. Prioul J.L. 2000. Xenia effects in maize
with normal endosperm. Crop Science. Vol. 40 No. 1 : 182-189.
Burczyk J, Koralewski T. 2005. Parentage versus two-generation analyses for
estimating pollen-mediated gene flow in plant population. Mol.Ecol.
14:2525-2537.
Bodor P, Gaal M, Toth M. 2008. Metaxenia in apples cv. Rewena, Relinda,
Baujade as influenced by scab resistant pollinizers. Int J Hort Sci.14(3):11-
14.
Cansian RL, Mossi AJ, Luccio MD, Cechet ML, Mazutti M, Echeverrigaray S.
2010. Molecular identification of pollen donor plants on a progeny of
Cambona-4 female matrix of maté (Ilex paraguariensis St. Hil.
Aquifoliaceae). Acta Scientiarum Biological Sciences. 32(1):39-42.
Carneiro FS, Lacerda AEB, Lemes MR, Gribel R, Kanashiro M, Wadt LHO,
Sebbenn AM. 2011. Effects of selective logging on the mating system and
pollen dispersal of Hymenaea courbaril L. (Leguminosae) in the Eastern
Brazilian Amazon as revealed by microsatellite analysis. Forest Ecology
and Management. 262:1758-1765.
Chan E, Elevitch CR. 2006. Cocos nucifera (coconut). Species profile for pasific
island agroforestry. www.traditionaltree.org. Diakses 19 April 2013.

92
Chen H, He H, Zou Y, Chen W, Yu R, Liu X, Yang Y, Gao YM, Xu JL, Fan LM.
2011. Development and application of a set of breeder-friendly SNP
markers for genetic analyses and molecular breeding of rice (Oryza sativa
L.). Theor Appl Genet. 123:869-879.
Creste S, Tulmann AN, Figueira A. 2001. Detection of single sequence repeat
polymorphism in denaturing polyacrylamide sequencing gels by silver
staining. Plant Mol Biol Reporter 19:299-306.
DebMandal M, Shyamapada M. 2011. Coconut (Cocos nucifera L.:
Arecaceae): In health promotion and disease prevention. Asian Pacific
Journal of Tropical Medicine. 4(3):241-247.
Denney JO. 1992. Xenia includes metaxenia. HortScience 27:722–728.
Dick CW, Etchelecu G, Austerlitz F. 2003. Pollen dispersal of tropical tree
(Dinizia excels: Fabaceae) by native insects and African honeybees in
pristine and fragmented Amazonian rainforest. Mol Ecol. 12:753-764.
Dunphy BK, Hamrick JL, Schwagerl, J. 2004. A comparison of direct and indirect
measures of gene flow in the bat-pollinated tree Hymenaea courbaril in
the dry forest life zone of south-western Puerto Rico. Int J Plant Sci.
165:427-436.
Dishutbun. 2004. Selayang pandang komoditi kelapa kopyor di Kabupaten Pati.
Dinas Kehutanan dan Perkebunan Kabupaten Pati.
Ehlenfeldt M.K. 2003. Investigations of metaxenia in northern highbush blueberry
(Vaccinium corymbosum L.) cultivars. Journal of the American
Pomological Society. 57(1):156:162.
Esteras Cristina, Pedro Gómez, Antonio JM, José B, Nelly V D, Cristina R,
Fernando N and Belén P. 2012. High-throughput SNP genotyping in
Cucurbita pepo for map construction and quantitative trait loci mapping.
BioMedCentral. (13- 80) :1471-2164
Falconer DS, Mackay TFC. 1985. Introduction to quantitative genetic. New York
(US): Longman.
Feng FJ, Sui X, Chen MM, Zhao D, Han SJ, Li MH. 2010. Mode pollen spread in
clonal seed orchard of Pinus koraiensis. Journal of Biophysical Chemistry.
1(1):33-39.
Ganal MW, Altmann T, Roder MS. 2009. SNP identification in crop plants. [curr
opin]. Plant Biol. 12:211-217.
Gary NE. 1992. Activities and Behavior of Honey Bee. Di dalam: The Hive and
the honey bee. Illinois: Dadant & Hamilton, Ltd.
Gupta PK, Roy JK, Prasad M. 2002. Single nucleotide polymorphisms: a new
paradigm for molecular marker technology and DNA polymorphism
detection with emphasis on their use in plants. Curr Sci. 80:524-535.
Hamrick JL, Trapnell DW. 2011. Using population genetic analyses to understand
seed dispersal patterns. Acta oecologica. 37:641-649.
Hannum S, Hartana A, Suharsono. 2003. Kemiripan genetika empat populasi
kelapa genjah berdasarkan pada Random Amplified Polymorphic DNA.
Hayati 4:125-129.
Herrera MM, Alan WM, James WB, David NK, Raymond JS. 2007. Usefulness
of WRKY gene-derived markers for assessing genetic population
structure: An example with Florida coconut cultivars. Scientia
Horticulturae 115:19–26.

93
Holton TA, Christopher JT, McClure L, Harker N, Henri HJ. 2002. Identification
and mapping of polymorphic SSR markers from expressed gene sequences
of barley and wheat. Mol Breed. 9:63-71.
Jannati M, Fotouhi R, Pourjan A, Salehi A Z. 2009. Genetic diversity analysis of
Iranian citrus varieties using microsatellite (SSR) based markers. J Hortic
For. 1:120-125.
Karp AS, Kresnovich, Bhat KV, Ayad WG, Hodgkin T. 1997. Molecular Tools in
Plant Genetic Resources Conservation: A Guide to The Technologies.
IPGRI Technical Bulletin No.2. Italia (IT): International Plant Genetic
Resources Institute.
Kasno, Hasan HES, Efendi DS, Syaefuddin. 2010. Efektifitas 3 spesies lebah
madu sebagai agen penyerbukan untuk meningkatkan produktifitas
(>40%) biji jarak pagar (Jatropa curcas) pada ekosistem iklim basah.
Jurnal Ilmu Pertanian Indonesia. 15(1):25-33.
Kurniasih Surti. 2012. Pemanfaatan marka molekuler untuk mendukung perakitan
kultivar unggul kakao (Theobroma cacao L.) [disertasi]. Bogor (ID):
Institut Pertanian Bogor.
Kumar Pradeep, Manimekalai R, Ranjitha Kumari. 2011. Microsatellite Marker
based Characterization of South Pacific Coconut (Cocos nucifera L.)
Accessions. International Journal of Plant Breeding and Genetics. 5(1)
:34-43.
Lebrun P, Baudouin L, Bopurdeix T, Konan JL, Barker JHA, Aldam C, Herran A,
Ritter E. 2001. Construction of linkage map of the Rennell Island Tall
coconut type (Cocos nucifera L.) and QTL analysis for yield characters.
Genome. 44:962-970.
Lestari P, Koh HJ. 2013. Development of new CAPS/Dcaps and SNAP markers
for rice eating quality. Hayati Journal Biosciences. 20(1): 15-23.
Li S, Wan H, Ji H, Zou K, Yang G. 2009. SNP discovery based on CATS and
genotyping in the finless porpoise (Neophocaena phocaenoides). Conserv
Genet 10:2013-2019.
Liferdi L. 2008. Lebah polinator utama pada tanaman hortikultura. Iptek
Hortikultura. 4:1-5.
Lian C, Miwa M, Hogetsu T. 2001. Outcrossing and paternity analysis of Pinus
densiflora (Japanese red pine) by micro- satellite polymorphism. Heredity
87:88–98.
Luengwilai K, Diane M. B., Orrawan P., Jingtair S. 2014. Postharvest quality and
storage life of „Makapuno‟ coconut (Cocos nucifera L.). Scientia
Horticulturae. 175: 105–110 .
Lowe A, Harris S, Ashton P. 2004. Ecological Genetics. London (UK): Blackwell
Publishing.
Mahmud Z. 2000. Petunjuk teknis budidaya kelapa kopyor. Departemen
Kehutanan dan Perkebunan. Jakarta (ID): Dirjen Perkebunan.
Mallet J. 2001. Gene flow. Di dalam Woiwod IP, Reynolds DR, Thomas CD,
editor. Insect Movement: Mechanisms and Consequences. Wallingford
(GB): CAB International.
Mammadov JA, Aggarwal R, Buyyarapu R, Kumpatla SP. 2012. SNP Markers
and their impact on plant breeding. Int J Plant Genom. 2012:1-11.

94
Marsico TD, Jessica JH, Romero-Saverson J. 2009. Patterns of seed dispersal and
pollen flow Quercus garryana (Fagaceae) following post-glacial climatic
changes. J Biogeogr. 36(5):929-941.
Maskromo I dan H. Novarianto. 2007. Potensi genetik kelapa kopyor Genjah.
Warta Penelitian dan Pengembangan Pertanian. Vol. 29 No. 1.
Maskromo I, Novarianto H, Mashud N. 2007. Potensi pengembangan kelapa
kopyor di Indonesia. Warta Penelitian dan Pengembangan Tanaman
Industri Vol 13 No. 1.
Maskromo I, Novarianto H, Sudarsono. 2011. Fenologi pembungaan tiga varietas
kelapa genjah kopyor pati. Di dalam: Roedhy P, Slamet S, Anas D, Nurul
K, Dewi S, Sintho WA, editor. Prosiding Seminar PERHORTI
Kemandirian Produk Hortikultura untuk Memenuhi Pasar Domestik dan
Ekspor; 2011 Nov 23-24; Lembang, Indonesia. Bogor (ID): Perhimpunan
Hortikultura . Indonesia. Hlm 1002-1010.
Maskromo I. 2005. Kelapa Kopyor, dari normal jadi abnormal. Trubus No. 429.
XXXVI.
Marshal TC, Slate J, Krilek LEB, Pemberton JM. 1998. Statistical confidence for
likehood based paternity inference in nature populations. Mol Ecol. 7:639-
655.
McCouch SR, Zhao K, Wright M, Tung CW, Ebana K, Thomson M, Reynolds A,
Wang D, DeClerck G, Ali ML. 2010. Development of genome wide SNP
assays for rice. Breed Sci. 60:524-535.
Milleron Matias, Unai Lopez de Heredia, Zaida Lorenzo, Ramon Perea, Aikaterini
Dounavi, Jesus Alonso, Luis Gil, Nikos Nanos. 2012. Effect of canopy
closure on pollen dispersal in a wind-pollinated species (Fagus sylvatica
L.). Plant.Ecology. 213:1715-1728.
Mondini L, Arshiya Noorani, Mario A. Pagnotta. 2009. Assesing plant genetic
diversity by molecular tools. Diversity. 1:19-35.
Moran., Emily and James SC. 2011. Estimating seed and pollen movement in a
monoecious plant: a hierarchical Bayesian approach integrating genetic
and ecological data. Molecular Ecology 20 : 1248–1262.
Morin PA, Luikart G, Wayne RK, The SNP working group. 2004. SNPs in
ecology, evolution and conservation. Trends Ecol Evol. 19:208-216.
Muntamah L. 2009. Aktivitas Apis Cerana mencari polen dan indentifikasi polen
di perlebahan tradisional bali [tesis]. Bogor (ID): Institut Pertanian Bogor.
Novarianto H, Miftahorrachman. 2000. Koleksi dan konservasi jenis-jenis kelapa
unik. Makalah poster dalam Simposium Pengelolalan Plasma nutfah dan
pemuliaan Bandung 22-23 September. Perhimpunan Ilmu Pemuliaan
Indonesia.
Novarianto H, Kangiden DI, Tampake H, Rompas T. 2000. Penyerbukan buatan
pada kelapa. Pemberitaan Penelitian Tanaman Industri. 8(50):13-16.
Novarianto H, Lolong AA. 2012. Peningkatan persentase buah kopyor melalui
penyerbukan sendiri. Bul Palma. 13 (1): 7-16.
Novarianto H, Maskromo I, Dinarti D, and Sudarsono. 2014. Production
technology for Kopyor coconut seednuts and seedlings in Indonesia.
International Journal on Coconut R & D. 30(2):31-40.

95
Olfati JA, Sheykhtaher Z, Qamgosar R, Sabet AK, Peyvast GH, Samizadeh H,
Rabiee B. 2010. Xenia dan metaxenia on cucumber fruit and seed
characteristics. Int J Veg Sci. 16:243-252.
Panaud O, Chen X, McCouch SR. 1996. Development of microsatellite markers
and characterization of simple sequence length polymorphism (SSLP) in
rice (Oryza sativa). Mol.Gen.Genet. 252, 597-607.
Pandin DS. 2009a. Inbreeding depression analysis based on morphological
characters in four generations of selfed Mapanget Tall Coconut no 32
(Cocos nucifera L.). Indonesian Journal of Agriculture 2(2):110-114.
Pandin DS. 2009b. Depresi penangkaran dalam empat generasi penyerbukan
tertutup tanaman kelapa dalam mapanget No.32 berdasarkan sifat
morfologi dan penanda mikrosatelit. [disertasi]. Bogor (ID): Institut
Pertanian Bogor.
Piotto., FA, Katherine DBP, Marcílio de A, Giancarlo CXO. 2013. Interspecific
xenia and metaxenia in seeds and fruits of tomato. Sci. Agric. (70.2) :
p.102-107
Perera PIP, Hocher V, Weerakoon LK, Yakandawala DMD, Fernando SC,
Verdeil JL. 2010. Early inflorescence and floral development in Cocos
nucifera L. (Arecaceae: Arecoideae). South African Journal Botany
76 ; 482-492.
Powel W, Gordon CM, Jim P. 1996. Polimorphism revealed by simple sequence
repeat. Trend Plant Sci. 1:215-221
Prabha SS, Indira EP, Nair PN. 2011. Contemporary gene flow and matting
system analysis in natural teak forest using microsatellite markers.
Current Science 101 (9):1213-1219.
Prastowo B. 2007. Potensi sektor pertanian sebagai penghasil dan pengguna
energi terbarukan. Perspektif Vol. 6 No. 2/ Desember . Hal 84-92.
Rajesh MK, Arunachalam V, Nagarajan P, Lebrun P, Samsudeen K, Thamban C.
2008. Genetic survey of 10 Indian coconut landraces by Simple Sequence
Repeats (SSRs). Scientia Horticulturae 118:282–287.
Ramirez VM, Tablat VP, Kevan PG, Morillo IR, Harries H, Barrera MF, Villareal
DZ. 2004. Mixed mating strategies and pollination by insects and wind in
coconut palm (Cocos nucifera L. (Arecaceae)): importance in production
and selection. Agricultural and Forest Entomology. 6:155-163.
Rohde W, Kullaya A, Rodriguez J, Ritter E. 1995. Genome analysis of Cocos
nucifera L by PCR amplification of spacer sequences separating a subset
of copis-like EcoR1 repetitive elements. J Genet and Breed. 49:170 -186.
Ruan C. 2010. Germplasm regression combined marker trait association
identification on plants. Afr J Biotechnol. 9:573-580.
Sajib AM, M.M. Hossain, A.T.M.J. Mosnaz, H. Hossain, M.M. Islam, M.S. Ali,
SH. Prodhan. 2012. SSR marker-based molecular characterization and
genetic diversity analysis of aromatic landreces of rice (Oryza sativa L.). J.
BioSci. Biotech 1(2): 107-116.
Samonthe LJ, Mendoza EMT, Ilag LL, De La Cruz ND, Ramirez DA.1989.
Galactomannan degrading enzym in maturing normal and makapuno and
germinating normal coconut endosperm. Phytochemistry. 28(9):2269-
2273.

96
Sambrook J. and Russel DW. 2001. Molecular cloning: a Laboratory Manual.
Third Edition. Cold Spring Harbor Laboratory Press, Cold Spring Harbor,
New York.
Santos GA. 1999. Potensial use of clonal propagation in coconut improvement
program. Di dalam Oropeza C, Verdiel JL, Ashburner GR, Cardena R,
Samantha JM, editors. Current advances in coconut biotechnology.
Current Plant Science and biotechnology in Agriculture. London (GB):
Kluwer Academic Publisher. hlm 419-430.
Santoso U, Kubo K, Ota T, Tadokoro T, Maekawa A. 1996. Nutrient composition
of kopyor coconuts (Cocos nucifera L.). Food Chem. 57:299-304.
Satyabalan K. 1995. Xenia effect and hybrid vigor in coconuts. Int J Coconut
R&D. 11(1):41-47.
Schuster WSF, Mitoon JB. 2000. Paternity and gene dispersal in limberpine
(Pinus flexilis James). Heredity. 84:348-361.
Selkoe KA, Toonen RJ. 2006. Microsatellites for ecologists: a practical guide to
using and evaluating microsatellite markers. Ecol Lett. 9:615-629.
Semagn K, Bjornstad A, Ndjiondjop MN. 2006. An overview of molecular
marker methods for plants. African J Biotech. 5:2540-2568.
Shamay A, Fang J, Pollak N, Yonash N (2006). Discovery of c-SNPs in Anemone
coronaria and assessment of genetic variation. Genet. Resour. Crop Evol.
53: 821-829.
Sianturi, H.S.D., 2001. Budidaya Kelapa Sawit. Fakultas Pertanian Universitas
Sumatera Utara, Medan.
Soenarsih Sri. 2012. Pala (Myristica spp.) maluku utara berdasarkan keragaman
morfologi, kandungan atsiri, pendugaan seks tanaman dan analisis marka
SSR [disertasi]. Bogor (ID): Institut Pertanian Bogor.
Sudarsono, Sudrajad, Novarianto H, Hosang MLA, Dinarti D, Rahayu MR,
Maskromo I. 2012. Produksi bibit kopyor true to type dengan persilangan
terkontrol dan peningkatan produksi buah kopyor dengan polinator lebah
madu. Laporan Akhir Program Hi Link. Bogor (ID): Institut Pertanian
Bogor.
Sudarsono, Sudrajad, Novarianto H, Hosang MLA, Dinarti D, Rahayu MR,
Maskromo I. 2014a. Pendekatan genomik dan molekuler untuk
pengembangan kultivar unggul kelapa eksotik asal Indonesia, penyediaan
bibit dan pengendalian hamanya. Laporan Akhir KKP3N. IPB
bekerjasama dengan Badan Penelitian dan Pengembangan Pertanian.
Sudarsono, Sudrajad, Novarianto H, Hosang MLA, Dinarti D, Rahayu MR,
Maskromo I. 2014b. Produksi bibit kopyor true to type dengan persilangan
terkontrol dan peningkatan produksi buah kopyor dengan polinator lebah
madu. Laporan Akhir Program Hi Link. Bogor (ID): Institut Pertanian
Bogor.
Suryanarayana MC, Rao GM, Singh TSMS. 1992. Studies on pollen sources for
Apis cerana Fabr and Apis mellifera L bees at Muzaffarpur, Bihar, India.
Apidologie. 23: 33-46.
Sukendah 2009. Teknologi pembiakan kultur in vitro dan analisis molekuler pada
tanaman kelapa kopyor [disertasi]. Bogor (ID): Institut Pertanian Bogor.

97
Sutanto A, D. Sukma, C. Hermanto, and Sudarsono. 2014. Isolation and
characterization of Resistance Gene Analogue (RGA) from Fusarium
resistant banana cultivars. Emirates Journal of Food and Agriculture.
26(6):508-518.
Wattanayothin, S. 2005. The study on curd coconut hybrids. J. TNCEL 1(3):6-7.
Wattanayothin, S. 2010. Variety improvement of Makapuno. Proceedings of the
XLIV COCOTECH Meeting, 5-9 July 2010, Samui Island, Thailand. pp.
96-108.
Zane L, Bargelloni L, Patarnello T. 2002. Strategies for microsatellite isolation
[review]. Mol Ecol. 11:1-16.

98

99
LAMPIRAN
Lampiran 1. Pembuatan Larutan Stok
1. Buffer ekstraksi
Buffer ini merupakan campuran dari beberapa larutan, yaitu Tris, EDTA,
NaCl, CTAB, dan aquadest ,
Tris 1 M dibuat dengan cara Tris base 121,1 gr dilarutkan ke dalam 800
ml aquadest , kemudian pH diatur menjadi 7,5 menggunakan HCL pekat
(± 42 ml) untuk menurunkan pH, kemudian volume larutan dicukupkan
hingga 1000 ml,
EDTA 0,5 M dibuat dengan cara disodium ethylenediaminetetraacetate
dengan 186,1 gr dilarutkan dalam 800 ml aquadest , kemudian pH diatur
menjadi 8,0 menggunakan NaOH, setelah itu mencukupkan volume
hingga 1 liter,
NaCl 2,5 M dibuat dengan cara 292,2 gr NaCl dilarutkan dalam 1 liter
aquadest,
Buffer ekstraksi sebanyak 200 ml, dibuat dari Tris 20 ml, EDTA 8 ml,
NaCl 112 ml, CTAB (cetyltrimethylamoniumbromide) 4 gr kemudian
bahan-bahan tersebut dilarutkan dengan aquadest 50 ml kemudian
dicukupkan hingga volume mencapai 200 ml,
2. Chloroform-isoamylalkohol
Chloroform-isoamylalkohol merupakan campuran chloroform dengan
isoamyl alkohol dengan perbandingan 24:1, yaitu 480 ml chloroform dan 20
ml isoamyl alkohol,
3. SB Bufer 20X
Boric acid sebanyak 22,5 gr dan NaOH 4 gr dilarutkan dalam 300 ml
aquadest, kemudian volume dicukupkan hingga 500 ml, Stok 1 liter SB buffer
1x dibuat dengan melarutkan 50 ml stok SB buffer dengan 950 ml aquadest,
4. TBE Buffer 5 M
Tris base 54 gr dilarutkan dalam 800 ml aquadest , lalu ditambahkan 27,5 gr
boric acid dan 20 ml EDTA, kemudian volume dicukupkan hingga 1 liter,
Buffer TBE 0,5 M dibuat melalui pengenceran TBE buffer 5 M dengan
menggunakan rumus :
V1 x M1 = V2 x M2
V1 x 5 M = 1000 x 0,5 M
V1 = 10 ml TBE buffer 5 M + 990 ml aquadest
5. Etanol 70%
Etanol absolute sebanyak 350 ml dicampurkan dengan 150 ml aquadest ,
6. Ethidium bromide
Ethidium bromide sebanyak 1 gr dilarutkan ke dalam 100 ml aquadest, dan
penyimpanannya menggunakan botol gelap atau botol yang dibungkus
dengan aluminium foil,
7. Acrylamide Bis-acrilamide (19:1) 40%
Acrylamide 190 gr dan Bis-acrylamide 10 gr dimasukkan ke dalam beacker
glass 500 ml, kemudian ditambahkan aquadest sebanyak 400 ml, Setelah
larut sempurna, cukupkan volume menjadi 500 ml, Larutan disaring

100
menggunakan kertas saring dan menyimpannya ke dalam botol gelap di
lemari es,
8. Acrylamide solution 6%
Urea sebanyak 210 gr dilarutkan dalam 200 ml aquadest, Setelah larut,
kemudian ditambahkan 100 ml TBE buffer 5 M dan 75 ml stock akrilamid
40%, Volume dicukupkan sampai 500 ml, kemudian disaring dengan kertas
saring, Larutan disimpan dalam botol gelap di lemari es,
9. Ammonium persulfate (APS)
Ammonium persulfate sebanyak 1 gr dilarutkan dalam 10 ml aquadest,
kemudian larutan disimpan pada suhu 4ºC,
10. Sodium thiosulfate
Sodium thiosulfate 100 mg dilarutkan ke dalam 10 ml aquadest, larutan
disimpan pada suhu ruang,
11. Larutan dalam pewarnaan silver
Pewarnaan silver menggunakan lima larutan, yaitu larutan fiksasi, larutan
nitrit acid, larutan staining, larutan developer, larutan stop,
Larutan fiksasi dibuat dengan melarutkan 10 ml asam asetat glacial dan
100 ml ethanol 95% ke dalam 890 ml aquadest,
Larutan nitrit acid dibuat dengan melarutkan 15 ml nitrit acid ke dalam
985 ml aquadest,
Larutan staining dibuat dengan melarutkan 1,5 gr silver nitrat ke dalam
1 liter aquadest,
Larutan developer dibuat dengan melarutkan 30 gr sodium carbonat di
dalam 1000 ml aquadest, Larutan ini disimpan dalam freezer -20ºC
sampai pada saat akan digunakan, Pada saat akan digunakan, larutan
ditambahkan 1,5 ml formaldehid 37% dan 200 µl sodium thiosulfate,
Larutan stop dibuat dengan melarutkan 50 ml asam asetat glacial ke
dalam 950 ml aquadest

101
Lampiran 2 . Daftar 36 primer SSR
No Lokus Urutan basa (5’ – 3’)
Suhu
annealing
(°C)
Linkage
group
1 CnCir_74 F =GAG ATC CTC ACC TCC AC
R =CGG CAA CAA AGA GAA C 54 16
2 CnCir_J2 F =CCA TTG TCA TTG TTA TTT TG
R =GTC ACC ATC TTC TCA GTT TC 52 15
3 CnCir_87 F =ATA ACA TCC TCC AAC CTG
R =GAC TGA ATC CAA CCC TT 54 13
4 CnCir_G4 F =AGT ATA GTC ACG CCA GAA AA
R =AAA CCC ATA ACC AGC AAG 55 13
5 CnCir_K8 F=CCA GAC ATG AAA CAA ACA A
R =CAT GGC ACA TAG GAA GAA 53 13
6 CnCir_K1 F =TTA CCAGGC CAC AAA GAA
R =AGA GTG AAC AAAGAG GAA GAT T 55 12
7 CnCir_241 F =CCA CTC CAA CAA CAC C
R =AAT CAC CAA ACA CAT CTT C 53 11
8 CnCir_123 F =AAA GTG AAG TGG ATA ATG TG
R =AGA GAG GAT CTA GGG TTG T 54 11
9 CnCir_J10 F =GAG GGT ATG GTG CTG CTT G
R =ATC CTT CAT GTG GCT CTG C 60 10
10 CnCir_B11 F =TCT GCA TCC CTT CTT TAT TA
R =TTGTCT TTC TTT ATT CTA TTG G 53 10
11 CnCir_109 F =CCT ACC ACA CCT TCC A
R =ATC ATC TCA GTC CTT CTC A 55 10
12 CnCir_2 F =AGTCCT AAA AGT GTT GGC
R =GTA ATC CTA TGG CTG CTT 55 10
13 CnCir_86 F =ACT CAC GCA AAT ATA CTC A
R =ACT CACGCA AAT ATA CTC A 53 9
14 CnCir_89 F =GAG TTG GAG AAG AAG AGG
R =ACG ACA ATA GAT GGA ACA 53 9
15 CnCir_192 F =TTA GTT AGT GCT GTG GAT TG
R =TTG CTA TGA GTC CCT TGT 55 9
16 CnCir_D8 F =GCT CTT GAT GTG GCT GCT
R =AGG CGTGTT GAG ATT GTG 58 8
17 CnCir_121 F =GGA CAC TGG GTT CTG TT
R =CTC TGT AAT CTG CGG G 56 8
18 CnCir_56 F =AAC CAG AACTTA AAT GTC G
R =TTTGAA CTC TTC TAT TGG G 53 8
19 CnCir_H11 F =TCA TTC AGA GGA CAA AAG TT
R =TAA AAA TTC ATA AAG GTA AAA 46 5
20 CnCir_167 F =GGT GGG TAA GTG AAC ATC
R =GTG ATA CAA CGA ACC CTC 57 5
21 CnCir_C9 F = CAG AAA GGA GAA AGG AAA T
R =CTA CGA TAG AGG AAT GAG C 57 4
22 CnCir_C5 F =ACC AAC AAA GCC AGA GC
R =GCA GCC ACT ACC TAA AAA 57 4
23 CnCir_48 F =GTG AGG CTG CAA AGA AC
R =TCG TCA AAC CTG ACC A 51 4
24 CnCir_B12 F =GCT CTT CAG TCT TTC TCA A
R =CTG TAT GCC AAT TTT TCT A 51 3
25 CnCir_H4 F =TTA GAT CTC CTC CCA AAG
R =ATC GAAAGA ACA GTC ACG 55 3
26 CnCir_1 F =TTG GTC TAT TGC ATG TTC
R =TGG CAT TGA GAG GGT 53 3

102
27 CnCir_206 F =AAA GAG AAC GCA ACC A
R =CAA GTT CCA AAG AAC CA 51 2
28 CnCir_151 F =ACC ATG ATG TGC CTG T
R =GTT CAC AGT AGG TGG CTT 54 2
29 CnCir_147 F =TTT CTC ACC AAC AAA TAA AC
R =CTT GTG TGT TAG GGT CAT C 52 2
30 CnCir_51 F =TCT CGT GGA TCT CGT C
R =GCT CTT CCA GTT ACG TTT 55 2
31 CnCir_E4 F =GCA TGG TAT TCG GAT TTG
R =ATG GTT CAG ATT TGG ACA GT 55 1
32 CnCir_226 F =CTG AAG ATA TGT GTT TAT GC
R =TGT TCC AGA TTG AGG TT 52 1
33 CnCir_202 F =TTT AGA GGA AGA AGG ATG AG
R =GTG GTT GCT TGG TAT TGT 55 1
34 CNZ_21 F =ATGTTTTAGCTTCACCATGAA
R =TCAAGTTCAAGAAGACCTTTG 52 -
35 CNZ_51 F =CTTTAGGGAAAAAGGACTGAG
R =ATCCATGAGCTGAGCTTGAAC 52 -
36 CNZ_18 F =ATGGTTCAGCCCTTAATAAAC
R =GAACTTTGAAGCTCCCATCAT 52 -
Lampiran 3. Daftar primer SNAP
No Primer Id Urutan sekuens primer Panjang
Primer
(basa)
Suhu
Annealing
(°C)
1 CN_SUS1_P#3 Ref GAATGAGATGCTACAAAGAATAAATAAG 28 51,0
CN_SUS1_P#3 _REV TCTGTTCTAAATGGAACGCG 20 52,9
CN_SUS1_P#3 ALT GAATGAGATGCTACAAAGAATAAATAAA 28 50,7
2 CN_SUS1_P#14 REF GCTGAAAGTCACTGAAAAGGTAG 23 53,8
CN_SUS1_P#14 REV AAATTATTAAGAATGTCATGGTTTC 25 48,3
CN_SUS1_P#14 ALT GCTGAAAGTCACTGAAAAGGTAC 23 54,0
3 WRKY19-P#1Ref CGTCTTCTGCAAACTAAGCTCA 22 56,3
WRKY19-P#1Rev ATGATATATATACAAGACTACGCCGAT 27 55,0
WRKY19-P#1Alt GTCTTCTGCAAACTAAGCTCG 21 53,5
4 WRKY6-P#3Ref ATAAATATCACATATCCCTGAGCAA 25 55,0
WRKY6-P#3Rev CTACAGGTATTGTTAAATGTGCCA 24 55,0
WRKY6-P#3Alt TATAAATATCACATATCCCTGAGCAG 26 54,9

103
Lampiran 4. Glosari
1. Kelapa kopyor : kelapa mutan asli Indonesia dengan daging buah
(endosperma) yang terlepas dari cangkang (batok)-nya dan daging buahnya
lembut sehingga enak untuk campuran es atau es krim, Kelapa kopyor
umumnya dikonsumsi segar seperti kelapa muda,
2. Kelapa normal : kelapa dengan daging buah yang keras dan biasa digunakan
untuk menghasilkan kopra, santan, VCO atau minyak goreng, Di kalangan
masyarakat juga dikenal sebagai kelapa sayur,
3. Kelapa tipe Dalam : kelapa yang umur berbuah pertamanya antara 6-7 tahun
setelah tanam, Dicirikan dengan ukuran buah yang besar, jumlah buah per
tandan yang relatif sedikit, dan batang yang besar/kokoh,
4. Kelapa tipe Genjah : kelapa yang umur berbuah pertamanya antara 3-4
tahun setelah tanam, Dicirikan dengan ukuran buah yang relatif kecil, jumlah
buah per tandan yang relatif banyak, dan batang yang kecil,
5. Kelapa tipe Hibrida : kelapa hasil persilangan antara tipe Genjah (G) dengan
tipe Dalam (D), Karakteristiknya berbuah lebih cepat dari kelapa tipe Dalam ,
berbuah besar dan jumlah buah/tandan banyak, dengan batang yang lebih
besar/kokoh dibadingkan kelapa tipe Genjah,
6. Kelapa Makapuno : kelapa mutan asal Filipina dengan daging buah
(endosperma) yang lembek,
7. Gen K : gen yang menyebabkan daging buah (endosperma) kelapa menjadi
keras seperti kelapa normal,
8. Gen k : gen mutan yang menyebabkan daging buah(endosperma) kelapa
menjadi terlepas dari batoknya seperti pada kelapa kopyor,
9. Pohon kelapa normal KK (homosigot KK) : pohon kelapa dengan simbol
genetik (genotipe) KK merupakan pohon kelapa berbuah normal, Secara
teknis pohon kelapa ini juga disebut sebagai pohon kelapa normal homosigot
(KK) dan secara umum dikenal dengan istilah pohon kelapa sayur, Pohon
kelapa ini hanya akan menghasilkan buah kelapa normal (kelapa sayur) dan
tidak akan pernah menghasilkan buah kelapa kopyor,
10. Pohon kelapa kopyor Kk (heterosigot Kk) : pohon kelapa dengan simbol
genetik (genotipe) Kk merupakan pohon kelapa yang berbuah campuran
antara buah kelapa normal dan buah kelapa kopyor, Pohon kelapa ini di
daerah Pati, Jawa Tengah juga disebut kelapa kopyoran, Pohon kelapa kopyor
Kk, dalam setiap tandan buahnya berpotensi menghasilkan buah kelapa
campuran,yaitu: (a) buah kelapa normal KK, (b) buah kelapa normal Kk, dan
(c) buah kelapa kopyor kk,Dengan demikian, petani yang memanen buah dari
pohon kelapa kopyor Kk ini akan: (a) menjual buah kelapa kopyor kk yang
dipanen langsung ke konsumen dan(b) menjadikan buah kelapa normalyang
dipanen (campuran dari buah kelapa normal KK dan buah kelapa normal Kk)
sebagai bibit,
11. Pohon kelapa kopyor kk (homosigot kk) : pohon kelapa dengan simbol
genetik (genotipe) kk merupakan pohon kelapa yang berpotensi berbuah
100% buah kelapa kopyor dalam setiap tandan buahnya, Pohon kelapa kopyor
ini merupakan kelapa kopyor hasil kultur jaringan (embryo rescue), Pohon
kelapa kopyor kk, meskipun dalam setiap tandan buahnya berpotensi
menghasilkan buah kelapa 100%,tetapi: (a) jika terjadi penyerbukan dengan

104
serbuk sari dari kelapa normal KK,maka hasil buahnya menjadi 100% buah
kelapa normal, (b) jika terjadi penyerbukan dengan serbuk sari dari kelapa
kopyor Kk,maka hasil buahnya menjadi 50% buah kelapa normal Kk dan
50% buah kelapa kopyor kk, dan (c) jika terjadi penyerbukan dengan serbuk
sari dari kelapa kopyorkk,maka hasil buahnya menjadi 100% buah kelapa
kopyorkk,Dengan demikian, petani yang menanam pohon kelapa kopyor kk
inidisarankan : (a) mengisolasi pohon kelapa kopyor kk-nya daripohon kelapa
normal KK atau pohon kelapa kopyor Kk yang ada di sekitarnya, agar tidak
menurunkan produksi buah kopyor, dan(b) melakukan emaskulasi
(penghilangan bunga jantan penghasil serbuk sari) dari tanaman kelapa
normal KK atau kelapa kopyor Kk agar tidak menjadi sumber kontaminan
serbuk sari yang akan menurunkan produksi buah kopyor,
12. Buah kelapa normal KK (homosigot KK) : buah kelapa dengan daging
buah(endosperma) yang keras, yang dapat dihasilkan dari pohon kelapa
normal KK (homosigot KK) atau dari pohon kelapa kopyor Kk (heterosigot
Kk), Buah kelapa ini mempunyai embrio sigotik dengan konstitusi genetik
KK dan daging buah(endosperma) dengan konstitusi genetik KKK, Buah
kelapa normal KK (homosigot KK) ini jika dijadikan bibit akan menjadi bibit
kelapa normal, yang nantinya berkembang menjadi pohon kelapa normal KK
(homosigot KK) dan yang akan menghasilkan 100% buah kelapa normal,
13. Buah kelapa normal Kk (heterosigot Kk) : buah kelapa dengan daging
buah(endosperma) yang keras, yang dalam kondisi tertentu dapat dihasilkan
dari pohon kelapa normal KK (homosigot KK) – jika diserbuki oleh serbuk
sari dari pohon kopyor Kk atau kk; atau dari pohon kelapa kopyor Kk
(heterosigot Kk)– jika diserbuki oleh serbuk sari dari pohon kelapa normal
KK, kelapa kopyor Kk atau kelapa kopyor kk; atau dari kelapa kopyor kk
(homosigot kk) jika diserbuki oleh serbuk sari dari pohon kelapa normal KK
atau kelapa kopyor Kk, Buah kelapa normal Kk (heterosigot Kk) mempunyai
embrio sigotik dengan konstitusi genetik Kk dan daging buah(endosperma)
dengan konstitusi genetik Kkk atau KKk, Buah kelapa normal Kk
(heterosigot Kk) ini jika dijadikan bibit akan menjadi bibit kelapa kopyor
alami, yang nantinya berkembang menjadi pohon kelapa kopyorKk
(heterosigot Kk) dan yang akan menghasilkan 25-50% buah kelapa kopyor
per tandan,
14. Buah kelapa Kopyor homosigot kk : buah kelapa dengan daging
buah(endosperma) yang lembut dan terlepas dari batok (cangkang)-nya, yang
dalam kondisi tertentu dapat dihasilkan dari pohon kelapa kopyor Kk
(heterosigot Kk)– jika diserbuki oleh serbuk sari dari pohon kelapa kopyor
Kk (heterosigot Kk)atau kelapa kopyor kk (homosigot kk); atau dari pohon
kelapa kopyor kk (homosigot kk) - jika diserbuki oleh serbuk sari dari pohon
kelapa kopyor Kk; atau kelapa kopyor kk, Buah kelapa kopyor kk
(homosigot Kk) mempunyai embrio sigotik dengan konstitusi genetik kk dan
daging buah(endosperma) dengan konstitusi genetik kkk, Buah kelapa
kopyor kk (homosigot kk) ini tidak bisa dijadikan sebagai bibit, tetapi
langsung dijual kepada konsumen, Jika ingin dijadikan sebagai bibit maka
perlu dilakukan embryo rescue (penyelamatan embrio) dengan teknik kultur
jaringan, Hasil embrio rescue akan menjadi bibit kelapa kopyor homosigot
kk, yang nantinya berkembang menjadi pohon kelapa kopyor kk (homosigot

105
kk) dan yang akan berpotesi menghasilkan 100% buah kelapa kopyor per
tandan,
15. Bibit kelapa normal KK (homosigot KK): bibit kelapa yang ditumbuhkan
dari buah kelapa normal KK (homosigot KK), Bibit ini jika ditanam hanya
akan menghasilkan buah kelapa normal dan tidak akan menghasilkan buah
kelapa kopyor,
16. Bibit kelapa kopyor Kk (heterosigot Kk): bibit kelapa yang ditumbuhkan
dari buah kelapa normal heterosigot Kk, Bibit ini jika ditanam akan menjadi
pohon kelapa kopyor Kk dan berpotensi menghasilkan buah kelapa Kopyor
dengan persentase tertentu per tandannya,
17. Bibit kelapa kopyor alami: Bibit kelapa kopyor yang diperjualbelikan secara
komersial oleh petani penangkar kelapa kopyor, Pada umumnya, bibit kelapa
kopyor alami merupakan bibit campuran yang terdiri atas bibit kelapa kopyor
Kk (heterosigot Kk) dan bibit kelapa normal KK (homosigot KK), Secara
teoritis, proporsi campurannya berkisar antara 2:1 untuk bibit kelapa kopyor
Kk dan bibit kelapa normal KK,Bibit kelapa kopyor alamitelah dapat secara
rutin diproduksi dengan teknologi sederhana oleh petani,Karena merupakan
bibit campuran, konsumen pembeli bibit tidak mempunyai garansi akan
mendapatkan pohon kelapa yang menghasilkan kelapa kopyor atau yang
hanya menghasilkan kelapa normal, Jika bibityang dibeli adalah bibit kelapa
Kk (heterosigot Kk) maka akan menjadi pohon kelapa yang berbuah kopyor,
tetapi jika bibit yang dibeli adalah bibit kelapa KK (homosigot KK) maka
akan menjadi pohon yang tidak akan pernah berbuah kopyor, Adanya
ketidakpastian ini yang menjadi masalah bagi petani penangkar dan
konsumen yang membeli,
18. Bibit kelapa kopyor kk (homosigot kk): Bibit kelapa yang dihasilkan dari
kultur in vitro embrio sigotik kelapa kopyor, Karena kelapa kopyor kk tidak
bisa berkecambah secara alami, maka embrionya harus diisolasi dan ditanam
dalam media steril di laboratorium kultur jaringan, Hasilnya adalah bibit
kelapa kopyor kk (homosigot kk), yang hargaper bibitnyarelatif masih sangat
mahal, Produksi bibit kelapa ini memerlukan peralatan laboratorium yang
relatif canggih dan tidak efektif untuk dilakukanoleh petani,
19. Persilangan terbuka (open polination) : persilangan yang dilakukan secara
alami dengan bantuan serangga sehingga hanya dapat diidentifikasi induk
betinanya dan tidak dapat teridentifikasi (tidak jelas) identitas induk
jantannya, Dengan persilangan terbuka, identitas progeni hasil persilangan
hanya akan dapat dirunut ke satu tetua tertentu saja yang digunakan sebagai
induk betinanya, Bibit kelapa kopyor alami merupakan bibit kelapa yang
ditumbuhkan dari buah kelapa hasil persilangan terbuka,
20. Embryo sigotik : mata tunas yang ada di dalam buah kelapa, Ketika
berkecambah (secara alami atau dengan bantuan kultur jaringan),embryo
sigotik kelapanyaakan tumbuh dan berkembang menjadi bibit kelapa,
21. Daging buah (endosperma) : bagian dalam buah kelapa yang dapat dimakan
atau diperas untuk membuat santan dan minyak kelapa, Pada kelapa normal,
daging buahnya keras dan menempel pada batoknya, sebaliknya pada kelapa
kopyor, daging buahnya lembut dan terlepas dari batoknya,
22. Polinator: serangga yang membantu proses penyerbukan kelapa sehingga
bisa menghasilkan buah, Penyerbukan kelapa tipe Dalam memerlukan

106
bantuan serangga sedangkan tipe Genjah tidak harus dibantu serangga,
Tetapi, keberadaan serangga juga berpengaruh positif terhadap penyerbukan
kelapa tipe Genjah,
23. Polinator lebah madu : lebah madu juga dapat berfungsi sebagai serangga
yang membantu penyerbukan kelapa, Keuntungannya adalah serangga ini
menghasilkan produk tambahan berupa madu, selain sebagai polinator,
Serbuk sari dan nektar kelapa dapat dijadikan sebagai makanan bagi lebah
madu, sehingga budidaya lebah madu di bawah tegakan pohon kelapa dapat
berdampak positif karena menghasilkan madu dan membantu penyerbukan,
24. Produksi buah kelapa kopyor : budidaya kelapa kopyor untuk dipanen buah
kelapa kopyornya, Petani atau pekebun dapat menanam pohon kelapa kopyor
Kk (heterosigot Kk) yang akan menghasilkan buah kelapa kopyor 25-50%
atau pohon kelapa kelapa kopyor kk (homosigot kk) yang akan menghasilkan
buah kelapa kopyor 100%,
25. Produksi bibit kelapa kopyor : budidaya budidaya kelapa kopyor untuk
dipanen buah kelapa normalnya dan dijadikan sebagai benih untuk
menghasilkan bibit kelapa kopyor, Petani atau pekebun dapat menanam
pohon kelapa kopyor Kk (heterosigot Kk) yang akan menghasilkan buah
kelapa kopyor 25-50% dan buah normal yang akan dipanen umur 11 bulan
untuk dijadikan sebagai benih, Buah kelapa normal yang dipanen selanjutnya
dapat dikecambahkan menjadi bibit dan dijual sebagai bibit kelapa kopyor
alami (jika hasil persilangan terbuka) atau sebagai bibit kelapa kopyor T3
(jika hasil persilangan terkontrol dengan tetua jantan kelapa kopyor
homosigot kk)

107
Lampiran 5. Pola persebaran serbuk sari kelapa kopyor Genjah Pati desa Tayu
dengan tetua betina # 37
Lampiran 6. Pola persebaran serbuk sari kelapa kopyor Genjah Pati desa Tayu
dengan tetua betina # 39
108
Lampiran 7. Pola persebaran serbuk sari kelapa kopyor Genjah Pati desa Tayu
dengan tetua betina # 44
Lampiran 8. Pola persebaran serbuk sari kelapa kopyor Genjah Pati desa Tayu
dengan tetua betina # 51

109
Lampiran 9. Pola persebaran serbuk sari kelapa kopyor Genjah Pati desa Tayu
dengan tetua betina # 53
Lampiran 10. Pola persebaran serbuk sari kelapa kopyor Genjah Pati desa Tayu
dengan tetua betina # 58

110
Lampiran 11. Pola persebaran serbuk sari kelapa kopyor Genjah Pati desa Tayu
dengan tetua betina # 69
Lampiran 12. Pola persebaran serbuk sari kelapa kopyor Genjah Pati desa Tayu
dengan tetua betina # 85

111
Lampiran 13. Pola persebaran serbuk sari kelapa kopyor Genjah Pati desa Tayu
dengan tetua betina # 88
Lampiran 14. Pola persebaran serbuk sari kelapa kopyor Genjah Pati desa Tayu
dengan tetua betina # 92

112
Lampiran 15. Pola persebaran serbuk sari pada populasi Kelapa Dalam Kalianda,
Agom Jaya dengan pohon induk betina nomor 160
Lampiran 16. Pola persebaran serbuk sari pada populasi Kelapa Dalam Kalianda,
Agom Jaya dengan pohon induk betina nomor 185

113
Lampiran 17. Pola persebaran serbuk sari pada populasi Tayu sesudah introduksi
lebah dengan pohon induk betina nomor 31
Lampiran 18. Pola persebaran serbuk sari pada populasi Tayu sesudah introduksi
lebah dengan pohon induk betina nomor 38

114
Lampiran 19. Pola persebaran serbuk sari pada populasi Tayu sesudah introduksi
lebah dengan pohon induk betina nomor 46
Lampiran 20. Pola persebaran serbuk sari pada populasi Tayu sesudah introduksi
lebah dengan pohon induk betina nomor 48

115
Lampiran 21. Pola persebaran serbuk sari pada populasi Tayu sesudah introduksi
lebah dengan pohon induk betina nomor 51
Lampiran 22. Pola persebaran serbuk sari pada populasi Tayu sesudah introduksi
lebah dengan pohon induk betina nomor 57

116
Lampiran 23. Pola persebaran serbuk sari pada populasi Tayu sesudah introduksi
lebah dengan pohon induk betina nomor 58
Lampiran 24. Pola persebaran serbuk sari pada populasi Tayu sesudah introduksi
lebah dengan pohon induk betina nomor 68

117
Lampiran 25. Pola persebaran serbuk sari pada populasi Tayu sesudah introduksi
lebah dengan pohon induk betina nomor 85
Lampiran 26. Pola persebaran serbuk sari pada populasi Tayu sesudah introduksi
lebah dengan pohon induk betina nomor 88

118
Lampiran 27. Pola persebaran serbuk sari pada populasi Tayu sesudah introduksi
lebah dengan pohon induk betina nomor 167

119
RIWAYAT HIDUP
Penulis dilahirkan di Parepare pada tanggal 9 Februari 1982 sebagai anak
ke tiga dari tiga bersaudara dari pasangan Drs.Muh.Warkah Larekeng dan
Nadjmah Aminuddin,SPd. Pendidikan sarjana ditempuh di Jurusan Budidaya
Pertanian Universitas Hasanuddin Makassar pada tahun 2000 hingga 2004. Pada
tahun 2006 penulis menempuh studi Pascasarjana Sistem-Sistem Pertanian Unhas
dan lulus pada tahun 2008. Kesempatan untuk melanjutkan ke program Doktor
pada program studi Pemuliaan dan Bioteknologi Tanaman Departemen Agronomi
dan Hortikultura tahun 2009 dengan beasiswa BPPS dari Departemen Pendidikan
Tinggi Indonesia.
Penulis bekerja sebagai dosen yayasan di Universitas Muhammadiyah
Parepare, Kopertis IX dari tahun 2005 sampai 2014. Tahun 2015 hingga sekarang
menjadi dosen Fakultas Kehutanan Universitas Hasanuddin Makassar. Selama
mengikuti kuliah S3, penulis menjadi anggota Perhimpunan Pemulia Indonesia
(PERIPI) dan Perhimpunan Hortikultura Indonesia (PERHORTI). Penulis
mengikuti seminar Pakar Bioteknologi tahun 2012 sebagai penyaji dengan judul
artikel “Penggunaan 2,4 D dan Picloram untuk Induksi Kalus Embriogenik
Tanaman Kelapa Kopyor secara In Vitro”, dan pada acara Seminar Nasional
Hortikultura 2011 sebagai penyaji dengan judul artikel “Keefektifan Bahan
Pemadat Dan Pemotongan Haustorium Pada Kultur Embrio Zigotik Kelapa
Kopyor”. Karya ilmiah berjudul “Pollen Dispersal of Pati Kopyor Dwarf Coconut
Indicating Importance of Insect Pollinators role in its pollination” dikirimkan ke
Cord, International Journal on Coconut R & D, Asian and Pacific Coconut
Community dan akan diterbitkan pada Volume 31 Nomor 1 Tahun 2015. Artikel
yang lain dikirim ke Buletin Palma dengan judul “Penyebaran Serbuk Sari
Membuktikan Potensi Pengaruh Negatif Kelapa Normal Terhadap Hasil Buah
Kopyor”. Karya-karya ilmiah tersebut merupakan bagian dari Program S-3
penulis.


















