
Hypolipidemic Activity of HOE-402 Is Mediated by...
9
1005 Hypolipidemic Activity of HOE-402 Is Mediated by Stimulation of the LDL Receptor Pathway M. Huettinger, M. Hermann, H. Goldenberg, E. Granzer, and M. Leineweber HOE- 402 (4 -amlno-2 - [4,4 -dimethyl-2 -oxo-1 -imldazolldinyl] -pyrimldlne- 5 -A'- [trifluoromethylphenyl] - carboxamide-monohydrochloride) has been shown to exhibit hypolipidemic action in heterozygous Watanabe heritable hyperlipidemic rabbits. In all animals, elevated cholesterol levels were reduced to normal (from 3.0 to 1.5 mmol/L) after 3 weeks of HOE- 402 treatment This was due entirely to reduction of low density lipoprotein (LDL) cholesterol and was paralleled by accelerated removal of plasma '"I-LDL. This reduction of LDL levels was not found in homozygous LDL receptor-defective animals, emphasizing the necessity of a functional LDL receptor system for the hypolipidemic action. The effect of HOE-402 on LDL receptor activity in the cultured hepatoma cell line HepG2 was also determined. When cells were incubated with plasma from treated animals (containing cholesterol 1.5 mmol/L and HOE- 402 80 ng/mL), high-affinity cell-surface binding sites for LDL were induced more than threefold, as shown by Scatchard analysis of cell-surface binding data. Induction of the LDL receptor was detectable after 6 hours and was 300% after 18 hours. This induction was specific for LDL, as '"I-transferrin and ["Feltransferrin were internalized normally in HOE-402-treated cells. The increase of LDL receptor protein was related to induced LDL receptor mRNA levels (400%), as shown by quantification of Northern blotting experiments. These findings suggest that HOE-402 mediated its hypolipidemic action mainly via the LDL receptor pathway. It enhanced mRNA levels for LDL receptor, hence increasing its synthesis, which subsequently resulted in reduced plasma LDL levels. Apart from having pharmacological implications, this compound could be very useful in elucidating the components that regulate the gene expression of the LDL receptor. (Arteriosclerosis and Thrombosis 1993;13:1005-1012) KEY WORDS • hypolipidemia • WHHL rabbits • LDL receptor A nimal cells regulate their cholesterol supply /\ through a concerted regulation of two pathways J. A- that supply them with exogenous and endoge- nous cholesterol. When cells need cholesterol, they start the buildup of low density lipoprotein receptor (LDLr) protein to enhance the uptake of exogenous cholesterol. In addition, they increase the amount of enzymes responsible for de novo cholesterol synthesis. The bal- ance of these pathways is particularly delicate, as it controls not only the intracellular level of cholesterol but also the extracellular blood cholesterol level. As a therapeutic method of keeping blood cholesterol levels down, this balance can be shifted toward enhanced uptake of cholesterol into the cells. By blocking de novo synthesis of cholesterol with certain drugs, cells rely on exogenous uptake and mediate this demand by increas- ing the synthesis of mRNA coding for the LDLr pro- tein. 13 Besides blocking intracellular de novo synthesis, treatment with ethinyl estradiol also leads to dramati- cally enhanced LDLr activity. The enhanced LDL up- take from plasma into the liver leads to reduced plasma cholesterol levels. 4 Received October 12, 1992; revision accepted March 15, 1993. From the Department of Medical Chemistry, University of Vienna, Austria (Drs Huettinger, Hermann, and Goldenberg), and Hoechst AG, Frankfurt/M, FRG (Drs Granzer and Leineweber). Correspondence to University of Vienna, Department of Med- ical Chemistry, Wahringerstr 10, A-1090 Vienna, Austria (Dr M. Huettinger). In both systems, the series of events that are triggered follow the most powerful pathway to reduce plasma cho- lesterol: synthesis of large amounts of mRNA for LDLr leads to a proportional increase in cellular receptor num- bers. This rise in LDLr protein enhances the removal of LDL cholesterol (LDL-C) from the plasma, which results in the reduction of blood cholesterol levels. LDLr produc- tion operates within a variety of regulatory influences. The feedback regulation of LDLr synthesis via cholesterol and the influences of hormones are well accepted but are supplemented by new reports 14 describing additional ef- fects. In this study we investigated whether the induction of the LDLr pathway could be triggered by administration of HOE-402 (4-amino-2-[4,4-dimethyl-2-oxo-l-imidazo- lidinyl]-pyrimidine-5-/v'-[trifluoromethylphenyl]-carboxa- mide-monohydrochloride). This substance has been shown in preliminary experiments by Granzer 5 to possess hypolipidemic action in rats. Although detailed informa- tion is still lacking, we found no effect by HOE-402 on 3-hydroxy-3-methylglutaryl coenzyme A reductase in HepG2 cells (E. Granzer and M. Leinweber, unpublished observation), and in addition, no structural similarity to any known inhibitor of HMG CoA reductase was seen. For this first intensive investigation of the cellular mech- anism of the action of HOE-402, we used an animal model for decreased but inducible LDLr activity, the heterozy- gous Watanabe heritable hyperlipidemic (WHHL) rabbit, to show that the cellular effect of the substance is not overcome by regulatory influences in the intact organism. We followed plasma lipid parameters and measured 125 I- by guest on July 13, 2018 http://atvb.ahajournals.org/ Downloaded from
Transcript of Hypolipidemic Activity of HOE-402 Is Mediated by...

1005
Hypolipidemic Activity of HOE-402 Is Mediatedby Stimulation of the LDL Receptor Pathway
M. Huettinger, M. Hermann, H. Goldenberg, E. Granzer, and M. Leineweber
HOE- 402 (4 -amlno-2 - [4,4 -dimethyl-2 -oxo-1 -imldazolldinyl] -pyrimldlne- 5 -A'- [trifluoromethylphenyl] -carboxamide-monohydrochloride) has been shown to exhibit hypolipidemic action in heterozygousWatanabe heritable hyperlipidemic rabbits. In all animals, elevated cholesterol levels were reduced tonormal (from 3.0 to 1.5 mmol/L) after 3 weeks of HOE- 402 treatment This was due entirely to reductionof low density lipoprotein (LDL) cholesterol and was paralleled by accelerated removal of plasma'"I-LDL. This reduction of LDL levels was not found in homozygous LDL receptor-defective animals,emphasizing the necessity of a functional LDL receptor system for the hypolipidemic action. The effect ofHOE-402 on LDL receptor activity in the cultured hepatoma cell line HepG2 was also determined. Whencells were incubated with plasma from treated animals (containing cholesterol 1.5 mmol/L and HOE- 40280 ng/mL), high-affinity cell-surface binding sites for LDL were induced more than threefold, as shown byScatchard analysis of cell-surface binding data. Induction of the LDL receptor was detectable after 6hours and was 300% after 18 hours. This induction was specific for LDL, as '"I-transferrin and["Feltransferrin were internalized normally in HOE-402-treated cells. The increase of LDL receptorprotein was related to induced LDL receptor mRNA levels (400%), as shown by quantification of Northernblotting experiments. These findings suggest that HOE-402 mediated its hypolipidemic action mainly viathe LDL receptor pathway. It enhanced mRNA levels for LDL receptor, hence increasing its synthesis,which subsequently resulted in reduced plasma LDL levels. Apart from having pharmacologicalimplications, this compound could be very useful in elucidating the components that regulate the geneexpression of the LDL receptor. (Arteriosclerosis and Thrombosis 1993;13:1005-1012)
KEY WORDS • hypolipidemia • WHHL rabbits • LDL receptor
A nimal cells regulate their cholesterol supply/ \ through a concerted regulation of two pathways
J. A- that supply them with exogenous and endoge-nous cholesterol. When cells need cholesterol, they startthe buildup of low density lipoprotein receptor (LDLr)protein to enhance the uptake of exogenous cholesterol.In addition, they increase the amount of enzymesresponsible for de novo cholesterol synthesis. The bal-ance of these pathways is particularly delicate, as itcontrols not only the intracellular level of cholesterolbut also the extracellular blood cholesterol level. As atherapeutic method of keeping blood cholesterol levelsdown, this balance can be shifted toward enhanceduptake of cholesterol into the cells. By blocking de novosynthesis of cholesterol with certain drugs, cells rely onexogenous uptake and mediate this demand by increas-ing the synthesis of mRNA coding for the LDLr pro-tein.13 Besides blocking intracellular de novo synthesis,treatment with ethinyl estradiol also leads to dramati-cally enhanced LDLr activity. The enhanced LDL up-take from plasma into the liver leads to reduced plasmacholesterol levels.4
Received October 12, 1992; revision accepted March 15, 1993.From the Department of Medical Chemistry, University of
Vienna, Austria (Drs Huettinger, Hermann, and Goldenberg),and Hoechst AG, Frankfurt/M, FRG (Drs Granzer andLeineweber).
Correspondence to University of Vienna, Department of Med-ical Chemistry, Wahringerstr 10, A-1090 Vienna, Austria (Dr M.Huettinger).
In both systems, the series of events that are triggeredfollow the most powerful pathway to reduce plasma cho-lesterol: synthesis of large amounts of mRNA for LDLrleads to a proportional increase in cellular receptor num-bers. This rise in LDLr protein enhances the removal ofLDL cholesterol (LDL-C) from the plasma, which resultsin the reduction of blood cholesterol levels. LDLr produc-tion operates within a variety of regulatory influences. Thefeedback regulation of LDLr synthesis via cholesterol andthe influences of hormones are well accepted but aresupplemented by new reports14 describing additional ef-fects. In this study we investigated whether the inductionof the LDLr pathway could be triggered by administrationof HOE-402 (4-amino-2-[4,4-dimethyl-2-oxo-l-imidazo-lidinyl]-pyrimidine-5-/v'-[trifluoromethylphenyl]-carboxa-mide-monohydrochloride). This substance has beenshown in preliminary experiments by Granzer5 to possesshypolipidemic action in rats. Although detailed informa-tion is still lacking, we found no effect by HOE-402 on3-hydroxy-3-methylglutaryl coenzyme A reductase inHepG2 cells (E. Granzer and M. Leinweber, unpublishedobservation), and in addition, no structural similarity toany known inhibitor of HMG CoA reductase was seen.For this first intensive investigation of the cellular mech-anism of the action of HOE-402, we used an animal modelfor decreased but inducible LDLr activity, the heterozy-gous Watanabe heritable hyperlipidemic (WHHL) rabbit,to show that the cellular effect of the substance is notovercome by regulatory influences in the intact organism.We followed plasma lipid parameters and measured 125I-
by guest on July 13, 2018http://atvb.ahajournals.org/
Dow
nloaded from

1006 Arteriosclerosis and Thrombosis Vol 13, No 7 July 1993
LDL disappearance from the plasma to find out whetherplasma LDL was reduced and, if so, to estimate whetherthis reduction occurred via reduction of synthesis orenhancement of removal. To add evidence for the neces-sity of a functional LDLr system for the hypolipidemicaction, we administered the substance to homozygousLDLr-defective animals. To pinpoint the cellular mecha-nism we studied the response of isolated cells to HOE-402. These experiments demonstrated an immediateresponse of LDLr activity that could overcome choles-terol repression. All results are compatible with theassumption that HOE-402 triggers a specific induction ofLDLr mRNA. The sequence of the other steps of thepathway, as described by Goldstein and Brown, lead tothe reduction of LDL levels in the plasma of an intactorganism.
MethodsAnalytical Methods and Chemicals
Total cholesterol, triglycerides, apoprotein B (apoB),and LDL-C were determined using commercially avail-able reagent kits (Boehringer Mannheim, Mannheim,FRG). Protein content of lipoproteins and cells wasdetermined by the method of Lowry et al.6 Chemicalswere obtained from Sigma Inc, and HOE-402 wassupplied by Hoechst AG, Frankfurt, FRG.
RabbitsMale New Zealand White rabbits (3.0 to 3.5 kg) were
obtained from Madoerin, Switzerland. WHHL rabbitswere raised in Vienna from a breeding pair obtainedfrom J.L. Goldstein, University of Texas SouthwesternMedical Center at Dallas. HOE-402 was administeredorally each morning in a 1-mL emulsion of liquefiedcorn starch and carrot juice.
Preparation of Lipoproteins and SerumHuman LDL (d, 1.019 to 1.050 g/mL) was prepared as
described.7 Lipoprotein-deficient serum (LPDS) wasprepared by ultracentrifugation according to themethod of Goldstein et al7 with the thrombin treatmentomitted. Control serum was collected from the animalsin the pretreatment period, and conditioned serum wastaken in week 4 of the treatment. It was prepared bycentrifugation to remove the clots, followed by heatinactivation of the complement system and sterilizationthrough a 0.45-^m filter. Serum was then pooled andstored at -30°C.
CellsHepG2 cells were obtained from American Type
Culture Collection and grown in Eagle's minimumessential medium (MEM) (PAA, Linz, Austria) supple-mented as stated in the legends to the figures. Threedays before an experiment, cells were split into 5-cmdishes (NUNC, Roskilde, Denmark) and grown to 60%confluence in standard MEM. Additions of supple-mented media were made as stated, from 18 to 2 hoursbefore the uptake of LDL was determined.
Iodination of Lipoproteins and Assay ofInternalization and Surface Binding
In each preparation, 2 mg LDL at 10 mg/mL wasiodinated by the monochloride method to a specific
activity of 300 cpm/ng.8 For the in vivo determination ofLDL turnover, 40xl06 cpm labeled LDL was injectedinto the marginal ear vein, and the disappearance ofradioactivity was determined as described.8 Transferrinwas labeled with 125I by using IODO-GEN (Pierce,Europe BV) and with 59Fe according to Bates andSchlabach.9 Radioactivity was assayed in an LKBgamma counter (LKB, Wallach, Sweden). The uptakecapacity for radiolabeled transferrin and the 59Fe up-take was measured after incubation for 1 hour at 37°Cwith 125I-transferrin (105 cpm/nmol) and [59Fe]transfer-rin (3.7xlO4 cpm/nmol). Internalization was assessed asthe radioactivity found in cells after receptor-depen-dent input and degradation and output of radioactivityhad reached equilibrium. For determination of uptakeof labeled LDL, HepG2 cells were pregrown underconditions described and were assayed for internaliza-tion capacity or surface binding of labeled LDL accord-ing to Goldstein et al.7 Values given are for specificuptake, which was calculated from the difference oftotal uptake minus uptake measured when a 40-foldexcess of unlabeled LDL was present in the incubationmedium of duplicate or triplicate incubations.
Quantification of mRNA by Northern BlottingFor RNA preparations a procedure described by
Chomczynski and Sacchi10 was used with slight modifi-cations. Cells pretreated as described were washed andthen lysed with 2 mL of heated solution I (5 mol/Lguanidinium isothiocyanate and 10 mmol EDTA in 50mmol tris[hydroxymethyl]aminomethane [Tris]-HCl;pH 7.5) and 160 ^12-mercaptoethanol. Total RNA (5 to20 n%) was mixed with 2 volumes of FF-3-(N-morpholi-no)propanesulfonic acid (MOPS) (8% formaldehydeand 65% formamide in 350 mmol/L MOPS; pH 7) andseparated on a 1% horizontal agarose gel containing 2mmol/L EDTA, 5 mmol/L iodoacetamide, and 100ng/mL ethidium bromide in 40 mmol/L MOPS (pH 7.0).RNA was then transferred to a Hybond N membrane(Amersham) by using 20 x saline-sodium citrate (SSC)(3 mol/L NaCl and 0.3 mol/L sodium citrate; pH 7). AnLDLr cDNA probe (1) and a human /3-actin probe(kindly donated by D.W. Russell) were labeled by usinga random-primer kit to a specific activity of 5x10cpm/jig (Amersham). This labeled /3-actin probe wasdiluted sevenfold with unlabeled cDNA to give compa-rable intensities on exposure. After overnight prehy-bridization in a Hybaid hybridization oven (Biometra)at 42°C in 3x saline-sodium phosphate-EDTA buffer(50% formamide, 250 yuglmh denatured herring spermDNA, and 0.1% bovine serum albumin; pH 7), hybrid-ization of both probes was done overnight in the samebuffer. Filters were then washed with 2x SSC and l xSSC containing 0.1% sodium dodecyl sulfate (SDS) at42°C; 0.5 x SSC and 0.1 x SSC washes at 52°C; and afinal wash with O.lx SSC containing 0.1% SDS at 60°C.The dried filters were exposed to a Cronex4 film (notpreflashed; Du Pont) and quantified by densitometry.
ResultsIn Vivo Effect in the Intact Animal
We administered HOE-402 to a group of nine het-erozygous WHHL rabbits. In alternating 3- and 4-dayintervals, blood was drawn and assayed for total choles-
by guest on July 13, 2018http://atvb.ahajournals.org/
Dow
nloaded from
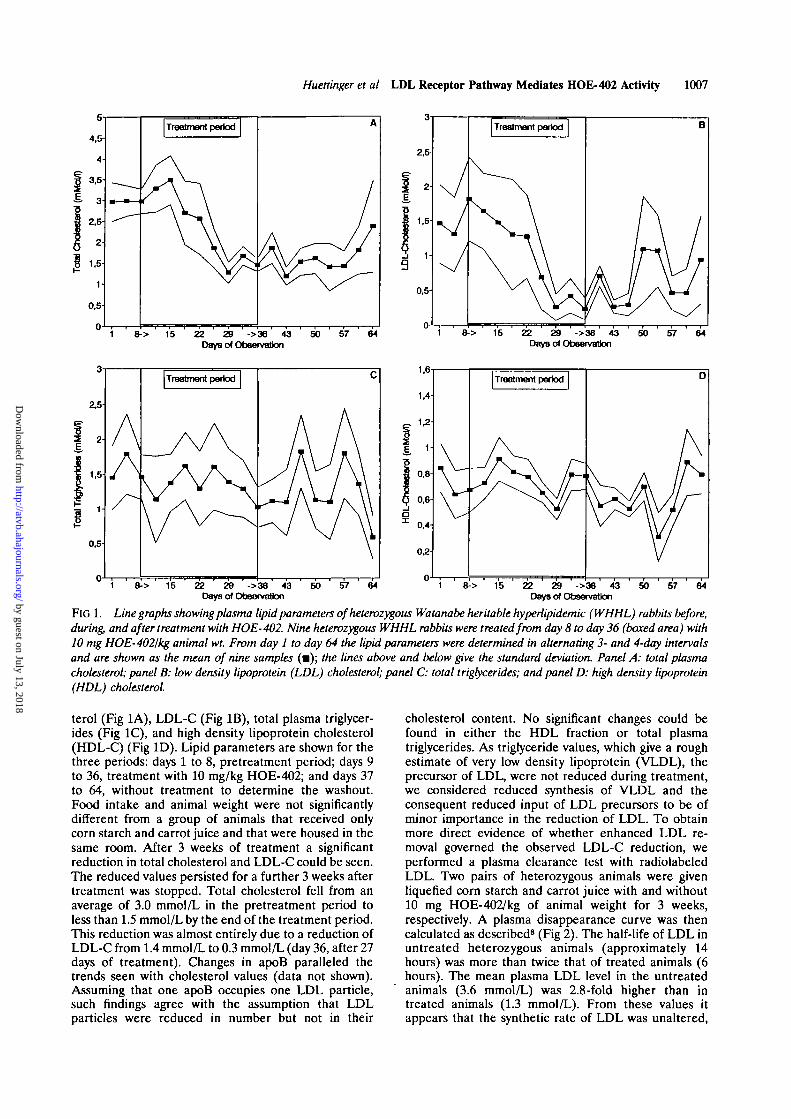
Huettinger et al LDL Receptor Pathway Mediates HOE- 402 Activity 1007
1 8-> 15 22 29 ->36 43 50 57 64
3
2,5
2
1,5-
1-
0,5-
0-
•7
Treatment period B
A /\J~\JT
1 8-> 15 22 29 ->36 43 50 57 64Days of Observation
50 57 6422 29 ->36 43 50 57 64 1 8-> 15 22 29 ->36 43Days of Observation Days of Observation
FlG 1. Line graphs showing plasma lipid parameters of heterozygous Watanabe heritable hyperlipidemic (WHHL) rabbits before,during, and after treatment with HOE-402. Nine heterozygous WHHL rabbits were treated from day 8 to day 36 (boxed area) with10 mg HOE-4021kg animal wt. From day 1 to day 64 the lipid parameters were determined in alternating 3- and 4-day intervalsand are shown as the mean of nine samples (•); the lines above and below give the standard deviation. Panel A: total plasmacholesterol; panel B: low density lipoprotein (LDL) cholesterol; panel C: total triglycerides; and panel D: high density lipoprotein(HDL) cholesterol
terol (Fig 1A), LDL-C (Fig IB), total plasma triglycer-ides (Fig 1C), and high density lipoprotein cholesterol(HDL-C) (Fig ID). Lipid parameters are shown for thethree periods: days 1 to 8, pretreatment period; days 9to 36, treatment with 10 mg/kg HOE-402; and days 37to 64, without treatment to determine the washout.Food intake and animal weight were not significantlydifferent from a group of animals that received onlycorn starch and carrot juice and that were housed in thesame room. After 3 weeks of treatment a significantreduction in total cholesterol and LDL-C could be seen.The reduced values persisted for a further 3 weeks aftertreatment was stopped. Total cholesterol fell from anaverage of 3.0 mmol/L in the pretreatment period toless than 1.5 mmol/L by the end of the treatment period.This reduction was almost entirely due to a reduction ofLDL-C from 1.4 mmol/L to 0.3 mmol/L (day 36, after 27days of treatment). Changes in apoB paralleled thetrends seen with cholesterol values (data not shown).Assuming that one apoB occupies one LDL particle,such findings agree with the assumption that LDLparticles were reduced in number but not in their
cholesterol content. No significant changes could befound in either the HDL fraction or total plasmatriglycerides. As triglyceride values, which give a roughestimate of very low density lipoprotein (VLDL), theprecursor of LDL, were not reduced during treatment,we considered reduced synthesis of VLDL and theconsequent reduced input of LDL precursors to be ofminor importance in the reduction of LDL. To obtainmore direct evidence of whether enhanced LDL re-moval governed the observed LDL-C reduction, weperformed a plasma clearance test with radiolabeledLDL. Two pairs of heterozygous animals were givenliquefied corn starch and carrot juice with and without10 mg HOE-402/kg of animal weight for 3 weeks,respectively. A plasma disappearance curve was thencalculated as described8 (Fig 2). The half-life of LDL inuntreated heterozygous animals (approximately 14hours) was more than twice that of treated animals (6hours). The mean plasma LDL level in the untreatedanimals (3.6 mmol/L) was 2.8-fold higher than intreated animals (1.3 mmol/L). From these values itappears that the synthetic rate of LDL was unaltered,
by guest on July 13, 2018http://atvb.ahajournals.org/
Dow
nloaded from

1008 Arteriosclerosis and Thrombosis Vol 13, No 7 July 1993
120 240 360 480 60060 180 300 420 540
TIME [MINUTES]
FIG 2. Line graph showing plasma decay curveof radiolabeled low density lipoprotein. Themean percentage of plasma radioactivity of twoanimals treated with HOE- 402 (A) and controls(•) was plotted against time. The radioactivitymeasured at 2 minutes after injection was set to100%. Numbers given are the mean plasmacholesterol levels at the time of the experiment.The dotted line was used to estimate the half-life.
and thus, the enhanced removal may be attributed toincreased production of functional LDLr's from thewild-type allele as a consequence of the treatment.Alternatively, a reduced synthesis of LDL could haveinduced the equilibrium we noted at the end of thetreatment. Such a situation would also result in en-hanced tracer clearance. To evaluate the involvement ofmechanisms not primarily dependent on the LDLr, wetreated homozygous WHHL rabbits with HOE-402.These animals have almost entirely lost functionalLDLr's due to a single gene defect. As a result, theyhave elevated production rates of LDL.11 Both condi-tions lead to markedly elevated plasma LDL levels. Themean total plasma cholesterol levels were estimated in apretreatment period and calculated for the high-choles-terol group (n=3) to be 15.3±4.7 and 10.2±1.7 mmol/Lfor the low-cholesterol group (n=2). The substance wasgiven for 28 days; the total plasma cholesterol levels areshown in Fig 3. The differences between mean choles-terol values in the third and fourth weeks of treatmentand the mean values of the control period were notstatistically significant (P>.02).
Influence of HOE-402 on the Internalizationof LDL in HepG2 Cells
From the results in the intact animal we reasoned thatHOE-402 was taken up via intestinal absorption andtransported to the liver, where it induced LDL removal.To gain direct evidence of an effect on hepatocytes, weused a human hepatocyte-derived cell line (HepG2)and incubated it in heat-inactivated serum collectedfrom the rabbits in the pretreatment period (controlserum) and at the end of the treatment period (condi-tioned serum). By using serum from treated animals weavoided the problems of solubility and biological avail-ability of this extremely hydrophobic substance. Theconcentration of HOE-402 in conditioned serum was 80ng/mL (0.2 ^mol/L). Incubation of HepG2 cells for 18hours with MEM including from 1% to 50% of condi-tioned serum resulted in a proportional increase in LDLinternalization (Fig 4). From 20% to 50% of thisincrease of 125I-LDL internalization reached approxi-mately 400% of the value measured in cells grown in
control serum. Control serum in concentrations of 10%,25%, and 50% showed no changes in uptake. The timecourse of induction was measured in 25% conditionedserum and was found to occur very rapidly. The induc-tion was detectable within 3 to 6 hours of incubation andreached a maximal value after 18 hours (Table). So far,we could not rule out that association of HOE-402 withLDL might have altered the affinity of LDL to itsreceptor. To investigate changes of affinity induced bysuch association, saturation kinetics of cell-surfacebinding sites for LDL at 4°C were determined. Thebinding of 125I-labeled LDL to the surface of HepG2cells incubated in 25% conditioned serum or grown in25% control rabbit serum is shown in Fig 5. Serum fromthe pretreatment period gave a maximal total binding ofapproximately 9 ng (4.5 fmol) labeled LDL/mg cell
25-i
2O
15-
ID-
1410 17
Days erf Observation
21 2824
FIG 3. Line graph showing plasma cholesterol levels of fivehomozygous Watanabe heritable hyperlipidemic rabbits dur-ing administration of HOE-402. The rabbits were treated with10 mg HOE-402/kg of animal weight, and plasma cholesterolwas determined in alternating 3- and 4-day intervals. Themean values of the two high-cholesterol and three low-cholesterol animals before treatment were 15.3±4.7 and10.2±l.7 mmol/L, respectively.
by guest on July 13, 2018http://atvb.ahajournals.org/
Dow
nloaded from

Huettinger et al LDL Receptor Pathway Mediates HOE-402 Activity 1009
10 16 20% PLASMA M HBMUH
FIG 4. Line graph showing induction of internalization ofradiolabeled low density lipoprotein (LDL) into HepG2 cellsby conditioned serum supplemented in the culture medium.Serum prepared from treated animals (•) and from the sameanimals in the pretreatment period (A) was added from 1%to 50% to the medium used to grow HepG2 cells. Afterincubation for 18 hours, cells were washed and incubated inEagle's minimum essential medium plus 2% bovine serumalbumin and 8 fig labeled LDL for 3 hours. Cells were thenwashed and solubilized in sodium hydroxide, and the contentof each dish was counted for radioactivity. An aliquot wasused to determine protein content. Values are percentages ofmean specific uptake (total minus unspecific of duplicates) permilligram of cell protein. Uptake into cells incubated in 10%control serum was set to 100%.
protein. A considerable portion was unspecific, as itcould not be suppressed by a 40-fold excess of coldLDL. HepG2 cells grown in conditioned serum exhib-ited a fourfold elevation in the number of high-affinitybinding sites for LDL (18 frnol) with no alteration inbinding affinity (by Scatchard analysis; Fig 5, lowerpanel).
Effect of HOE-402 on Internalizationof Iron and Transfer r in
The enhanced uptake of radiolabeled LDL may beattributed to a general stimulation of cell-surface recep-tors or, theoretically at least, to acceleration of theendocytotic machinery.12 To clarify this, we investigatedtransferrin uptake into HepG2 cells under conditionsthat stimulated LDL uptake. An accelerated turnover ofthe endocytotic machinery could be detected by com-paring the steady-state level of 125I-labeled transferrin
Time Course of Induction of LDLr Activity in HcpG2 Cells AfterIncubation in Serum From Treated Animals
TimeSpecific internalization
(ng LDL/mg cell protein)
18 Hours control plasma
3 Hours HOE-402 plasma
6 Hours HOE-402 plasma
9 Hours HOE-402 plasma
18 Hours HOE-402 plasma
87/103
91/109
122/131
165/209
250/303
LDLr, low density lipoprotein receptor. The values given arespecific uptake (total cell-associated radioactivity minus unspe-cific binding) from duplicate dishes from one representativeexperiment.
Bound[ng]
'l-LDL
FIG 5. Lower panel; line graph showing cell-surface bindingofl25I-labeled low density lipoprotein (LDL) at 4°C. Standardmedium was removed 18 hours before the experiments, thecells were washed, and the medium was changed to 25%rabbit serum (conditioned serum and control serum) asindicated HepG2 cells were then incubated for 18 hours in25% rabbit serum from untreated animals (o, •&) and 18hours in 25% rabbit serum from animals treated with HOE-402 (•, *) . For determination of cell-surface binding, eachdish received, after washing in ice-cold Eagle's minimumessential medium, ice-cold medium containing 2% bovineserum albumin and I25I-LDL with (£, *) or without excessunlabeled LDL. After incubation for 2 hours and washing, thetotal radioactivity bound to the cells was determined andplotted against the concentration of labeled LDL. Each valueis the mean of duplicate incubations. Upper panel: Scatchardanalysis of these values.
undergoing retroendocytosis with the amount of irondelivered during several rounds of retroendocytosisunder stimulation by HOE-402. Under these conditionstransferrin was endocytosed via the transferrin receptor(370 fmol/mg cell protein, steady-state level), and ironcollected in the endocytotic compartment (9120fmol/mg per hour). Fig 6 shows the values of cell-associated 125I-LDL, 125I-transferrin, and [59Fe] transfer-rin. Incubation with conditioned serum over an 18-hourperiod followed by measurement of the uptake withlabeled LDL resulted in a 260% induction of LDLaccumulation. Despite this considerable enhancementof LDLr activity, the cells continued to internalizetransferrin at a rate equal to the control level. Theaccumulation of iron in cells was also unaltered over themeasured time interval.
by guest on July 13, 2018http://atvb.ahajournals.org/
Dow
nloaded from
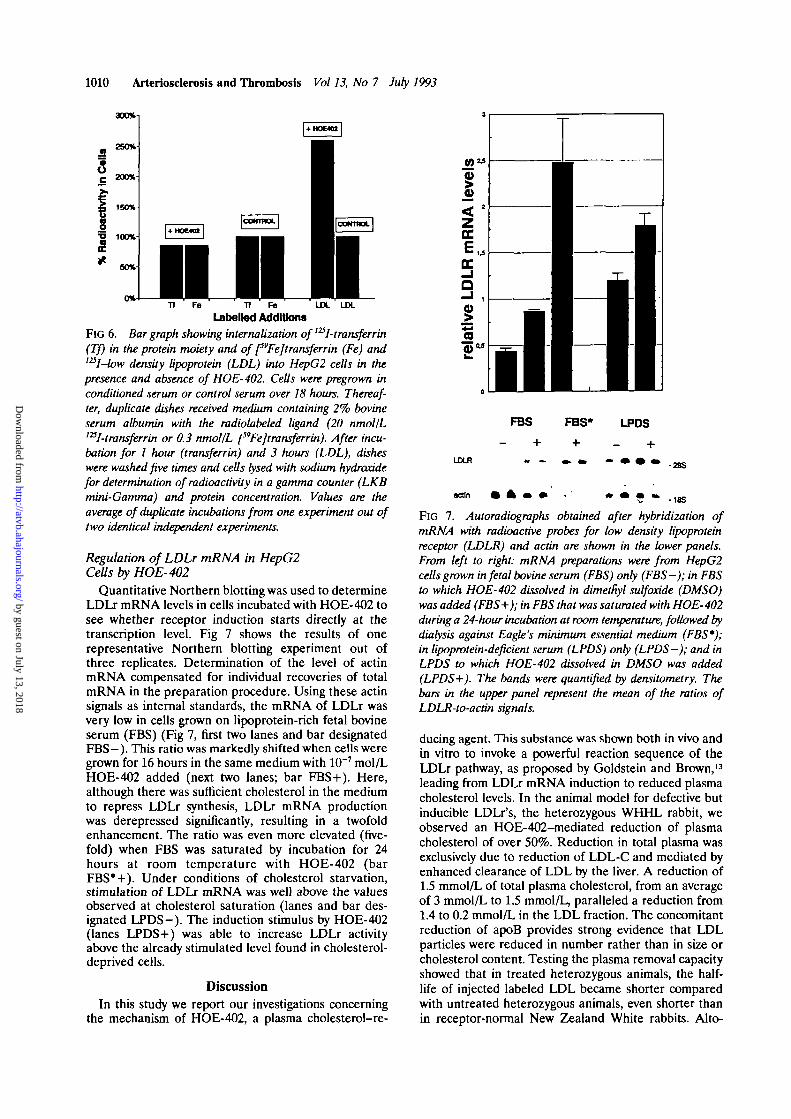
1010 Arteriosclerosis and Thrombosis Vol 13, No 7 July 1993
300%
Fe H Fi UX LDL
Labelled Additions
FlG 6. Bar graph showing internalization of l2Sl-transferrin(Tf) in the protein moiety and of [^Fejtransferrin (Fe) andl25I-low density lipoprotein (LDL) into HepG2 cells in thepresence and absence of HOE-402. Cells were pregrown inconditioned serum or control serum over 18 hours. Thereaf-ter, duplicate dishes received medium containing 2% bovineserum albumin with the radiolabeled ligand (20 nmol/LmI-transferrin or 0.3 nmol/L ^Fejtransferrin). After incu-bation for 1 hour (transferrin) and 3 hours (LDL), disheswere washed five times and cells lysed with sodium hydroxidefor determination of radioactivity in a gamma counter (LKBmini-Gamma) and protein concentration. Values are theaverage of duplicate incubations from one experiment out oftwo identical independent experiments.
Regulation of LDLr mRNA in HepG2Cells by HOE-402
Quantitative Northern blotting was used to determineLDLr mRNA levels in cells incubated with HOE-402 tosee whether receptor induction starts directly at thetranscription level. Fig 7 shows the results of onerepresentative Northern blotting experiment out ofthree replicates. Determination of the level of actinmRNA compensated for individual recoveries of totalmRNA in the preparation procedure. Using these actinsignals as internal standards, the mRNA of LDLr wasvery low in cells grown on lipoprotein-rich fetal bovineserum (FBS) (Fig 7, first two lanes and bar designatedFBS—). This ratio was markedly shifted when cells weregrown for 16 hours in the same medium with 10~7 mol/LHOE-402 added (next two lanes; bar FBS + ). Here,although there was sufficient cholesterol in the mediumto repress LDLr synthesis, LDLr mRNA productionwas derepressed significantly, resulting in a twofoldenhancement. The ratio was even more elevated (five-fold) when FBS was saturated by incubation for 24hours at room temperature with HOE-402 (barFBS*+). Under conditions of cholesterol starvation,stimulation of LDLr mRNA was well above the valuesobserved at cholesterol saturation (lanes and bar des-ignated LPDS-). The induction stimulus by HOE-402(lanes LPDS+) was able to increase LDLr activityabove the already stimulated level found in cholesterol-deprived cells.
DiscussionIn this study we report our investigations concerning
the mechanism of HOE-402, a plasma cholesterol-re-
LJXFt
actin
FBS
ft A •
FBS* LPDS
-2BS
-1SS
FIG 7. Autoradiographs obtained after hybridization ofmRNA with radioactive probes for low density lipoproteinreceptor (LDLR) and actin are shown in the lower panels.From left to right: mRNA preparations were from HepG2cells grown in fetal bovine serum (FBS) only (FBS—); in FBSto which HOE-402 dissolved in dimetfiyl sulf oxide (DMSO)was added (FBS+); in FBS that was saturated with HOE-402during a 24-hour incubation at room temperature, followed bydialysis against Eagle's minimum essential medium (FBS*);in lipoprotein-deficient serum (LPDS) only (LPDS-); and inLPDS to which HOE-402 dissolved in DMSO was added(LPDS+). The bands were quantified by densitometry. Thebars in the upper panel represent the mean of the ratios ofLDLR-to-actin signals.
ducing agent. This substance was shown both in vivo andin vitro to invoke a powerful reaction sequence of theLDLr pathway, as proposed by Goldstein and Brown,13
leading from LDLr mRNA induction to reduced plasmacholesterol levels. In the animal model for defective butinducible LDLr's, the heterozygous WHHL rabbit, weobserved an HOE-402-mediated reduction of plasmacholesterol of over 50%. Reduction in total plasma wasexclusively due to reduction of LDL-C and mediated byenhanced clearance of LDL by the liver. A reduction of1.5 mmol/L of total plasma cholesterol, from an averageof 3 mmol/L to 1.5 mmol/L, paralleled a reduction from1.4 to 0.2 mmol/L in the LDL fraction. The concomitantreduction of apoB provides strong evidence that LDLparticles were reduced in number rather than in size orcholesterol content. Testing the plasma removal capacityshowed that in treated heterozygous animals, the half-life of injected labeled LDL became shorter comparedwith untreated heterozygous animals, even shorter thanin receptor-normal New Zealand White rabbits. Alto-
by guest on July 13, 2018http://atvb.ahajournals.org/
Dow
nloaded from

Huettinger et al LDL Receptor Pathway Mediates HOE-402 Activity 1011
gether we observed a more-than-twofold acceleration ofcatabolism with a comparable reduction in pool size. Asthe synthetic rate of LDL at equilibrium is equal to thecatabolic rate (pool size x removal rate), our data suggestthat LDL synthesis was not affected by the treatment.
All reactions triggered by HOE-402 in heterozygousanimals, thereby leading to LDL reduction, are alsolikely to occur in homozygous animals.2 But if receptoractivity itself were the underlying principle of thecholesterol-lowering action, then the enhanced produc-tion of LDLr protein in homozygous WHHL rabbitswould not result in enhanced activity, as they producereceptor molecules that are functionally defective. Ourfinding that no plasma LDL reduction took place insuch a system emphasizes the role of LDLr-mediatedendocytosis.
Results from experiments with intact animalsprompted us to test the effect of HOE-402 on isolatedcells in tissue culture. A direct influence of HOE-402should be detected independent of any regulatorymechanism occurring in the liver cells of intact animals.HepG2 cells have been repeatedly shown to exhibitreceptor-mediated uptake of LDL sensitive to regula-tion.314 These cells were incubated in serum preparedfrom treated animals and, as a control, with serum fromthe same animals collected in the pretreatment period.After removal of HOE-402-containing medium bythorough washing, internalization capacity was deter-mined with labeled LDL. In these experiments wefound detectable stimulation of LDLr-mediated en-docytosis 3 to 6 hours after treatment with HOE-402serum, which was maximal after 18 hours in a dose-dependent manner. Increasing the concentration ofserum from 1% to 50% also raised the supply ofcholesterol, which normally leads to suppression ofLDL endocytosis. However, we observed no suppres-sion, indicating that HOE-402 can overcome downreg-ulation of receptor activity. Scatchard analysis of thebinding to cell-surface receptors at 4°C showed that thenumbers of high-affinity binding sites were enhanced. Amore than threefold increase in the number of bindingsites was found, which offered an explanation for theelevated capacity of internalization. There was no influ-ence on the affinity of the binding sites. Taken together,these findings extend our understanding of the mecha-nism underlying the reduction of plasma LDL-C medi-ated by HOE-402: plasma transports the active form ofHOE-402 to cells, where it leads to an induction of theLDLr system. As the liver is the dominating organ incholesterol removal, an extracellular to intrahepaticshift of LDL-C occurs. There is now general agreementthat induction of LDLr mRNA accounts for an increasein LDLr protein that leads to enhanced plasma LDLuptake. One well-documented example for such a se-quence in animals is the administration of pharmaco-logical doses of estradiol. In rabbits this leads to adose-dependent reduction of plasma LDL-C.4 It has notbeen determined whether this treatment leads to aspecific or composite regulation of mRNA or whether ittriggers metabolic events before these that result inLDLr stimulation. Here we show that HOE-402 did notinfluence endocytosis of transferrin and iron, whereas inthe same cells LDLr was induced by 250%. This em-phasizes a specific action: as transferrin uptake is alsoan indicator of a cell's growth status, this suggests that
the metabolic status of the cells was not otherwiseaffected.
In addition, we observed stimulation by HOE-402under conditions that suppress LDLr activity. When thecholesterol supply to cells was enhanced by elevatingserum concentration in the medium by severalfold, weobserved derepressed mRNA levels of LDLr. So far, wehave used in vivo prepared conditioned serum in ourexperiments. To rule out that a plasma factor inducedby HOE-402 was the active agent of receptor stimula-tion, we also dissolved HOE-402 directly in serum andtested for mRNA induction. The enhancement of LDLrmRNA was likewise found when HOE-402 was directly,without intestinal resorption, added to the tissue-cul-ture medium.
Considered together, the current data support theconcept that a dramatic stimulation by HOE-402 of theLDLr activity mediates the rapid clearance of LDL inintact animals as well as cultured cells. One reasonableassumption for the mechanism was the enhancement ofmRNA synthesis, which leads to an increased produc-tion of LDLr protein. The details of this enhancementare unclear. Our findings can be interpreted within theframework of the hypothesis that the compound itself ora complex with an intracellular protein controls LDLrgene expression. It is difficult, at this stage, to postulatea single and direct effect responsible for the hypolipid-emic action of HOE-402. Although the rapid responseof cells to the addition of HOE-402 suggests a directregulation of LDLr mRNA, it cannot be ruled out thatchanges in the regulatory cell cholesterol pool occurfirst. We have not detected any sign that would suggesta change in the intracellular cholesterol pool. Themolecular target of LDLr gene regulation by intracellu-lar cholesterol has been investigated in detail.15'16 Themolecules that couple LDLr gene expression to meta-bolic signals are under intensive investigation or are stillunknown.17 Our investigation might serve as an addi-tional tool to achieve that task.
AcknowledgmentThis work was supported in part by Fonds zur FOrderung
der Wissenschaftlichen Forschung (project S4605).
References1. Ma PTS, GU G, Sudhoff T, Bilheimer D, Goldstein JL, Brown MS.
Mevinolin, an inhibitor of cholesterol synthesis, induces mRNA forlow density lipoprotein receptor in livers of hamsters and rabbits.Proc NatlAcad Sci USA. 1986;83:8370-8374.
2. Kume N, Kita T, Mikami A, Yokode M, Ishii K, Nagano Y, KawaiC. Induction of mRNA for low-density lipoprotein receptors inheterozygous Watanabe heritable hyperlipidemic rabbits treatedwith CS-514 (Pravastatin) and cholestyramine. Circulation. 1989;79:1084-1090.
3. Cohen LH, Griffionen M, Havekes L, Schouten D, VanhinsberghV, Kempen H. Effects of compactin, mevalonate and low densitylipoprotein on 3-hydroxy-3-methylglutaryl-coenzyme A reductaseactivity in the human hepatoma cell line HepG2. BiochemJ. 1984;222:35-39.
4. Ma P, Yamamoto T, Goldstein J, Brown MS. Increased mRNA forlow density lipoprotein receptor in livers of rabbits treated with17alpha-ethinyl estradiol. Proc NatlAcad Sci US A. 1986;83:792-796.
5. Granzer E. Antiatherosclerotic activity of the new compoundImanixil (HOE-402). Naunyn Schmiedcbergs Arch Pharmacol.1988;337(supp!):R70. Abstract.
6. Lowry OH, Rosenbrough NJ, Farr AH, Randall RJ. Protein deter-mination with Folin phenol reagent. J Bid Chan. 1951;193:265-275.
7. Goldstein JL, Basu SK, Brown MS. Receptor mediated endocyto-sis of LDL in cultured cells. Methods EnzymoL 1983;98:241-260.
by guest on July 13, 2018http://atvb.ahajournals.org/
Dow
nloaded from

1012 Arteriosclerosis and Thrombosis Vol 13, No 7 July 1993
8. Huettinger M, Schneider WJ, Ho YK, Goldstein JL, Brown M. Useof monoclonal anti-receptor antibodies to probe the expression of thelow density lipoprotein receptor in tissues of normal and Watanabeheritable hyperlipidemic rabbits. / Chn Invest 1984;74:1017-1026.
9. Bates GW, Schlabach MR. Reaction of ferric salts with transferrin.J Bioi Chem. 1973;248:3228-3232.
10. Chomczynski P, Sacchi N. Single step method of RNA isolation byacid guanidinium thiocyanate-phenol-chloroform extraction. AnalBiochem. 1987;162:156-159.
11. Spady DK, Huettinger M, Bilheimer DW, Dietschy JM. Role ofreceptor-independent low density lipoprotein transport in themaintenance of tissue cholesterol balance in the normal andWHHL rabbit. J Upid Res. 1987;28:32-41.
12. Nerup J, Mandrup-Poulsen T, Hokfelt B. Insulin may cause trans-location of proteins to the cell surface by stimulating membranetrafficking from the trans Golgi reticulum. In: Genes and GeneProducts in the Development of Diabetes MtUitus. New York, NY:Elsevier Science Publishing Co Inc; 1989013-328.
13. Goldstein JL, Brown MS. Progress in understanding the LDLreceptor and HMGCoA reductase, two membrane proteins thatregulate plasma cholesterol. J Upid Res. 1984;25:1450-1461.
14. Krempler F, Kostner GM, Friedl W, Pauhveber B, Bauer H, Sand-hofer F. Lipoprotein binding to cultured human hepatoma cells.J Clin Invest 1987;80:401-408.
15. Sudhoff T, Russel DW, Brown MS, Goldstein JL. A 42 bp elementfrom LDL receptor gene confers end-product repression by sterolswhen inserted into viral TK promoter. Cell. 1987;48:1061-1069.
16. Russell DW, Yamamoto T, Schneider WJ, Slaughter CJ, BrownMS, Goldstein JL. cDNA cloning of the bovine low density lipo-protein receptor: feedback regulation of a receptor mRNA. ProcNatlAcadSd USA. 1983;80:7501-7505.
17. Dawson PA, Ridgway ND, Slaughter CA, Brown MS, GoldsteinJL. cDNA cloning and expression of oxysterol-binding protein, anoligomer with a potential leucin zipper. J Biol Chem, 1989;264:16798-16803.
by guest on July 13, 2018http://atvb.ahajournals.org/
Dow
nloaded from

M Huettinger, M Hermann, H Goldenberg, E Granzer and M LeineweberHypolipidemic activity of HOE-402 is mediated by stimulation of the LDL receptor pathway.
Print ISSN: 1079-5642. Online ISSN: 1524-4636 Copyright © 1993 American Heart Association, Inc. All rights reserved.
Avenue, Dallas, TX 75231is published by the American Heart Association, 7272 GreenvilleArteriosclerosis, Thrombosis, and Vascular Biology
doi: 10.1161/01.ATV.13.7.10051993;13:1005-1012Arterioscler Thromb Vasc Biol.
http://atvb.ahajournals.org/content/13/7/1005World Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://atvb.ahajournals.org//subscriptions/
at: is onlineArteriosclerosis, Thrombosis, and Vascular Biology Information about subscribing to Subscriptions:
http://www.lww.com/reprints
Information about reprints can be found online at: Reprints:
document.Permissions and Rights Question and AnswerFurther information about this process is available in theis being requested is located, click Request Permissions in the middle column of the Web page under Services.Clearance Center, not the Editorial Office. Once the online version of the published article for which permission
can be obtained via RightsLink, a service of the CopyrightArteriosclerosis, Thrombosis, and Vascular Biology Requests for permissions to reproduce figures, tables, or portions of articles originally published inPermissions:
by guest on July 13, 2018http://atvb.ahajournals.org/
Dow
nloaded from


















