
Hyla herpetological bulletin 2011_1
93
herpetological bulletin Vol. 2011., No 1., lipanj 2011. Hrvatsko herpetološko društvo
-
Upload
marija-kuljeric -
Category
Documents
-
view
246 -
download
0
description
Journal for publishing research on amphibians and reptiles of South-eastern Europe
Transcript of Hyla herpetological bulletin 2011_1

herpetological bulletin
Vol.
2011
., N
o 1.
, lip
anj 2
011.
Hrvatsko herpetološko društvo

Hyla herpetološki bilten
herpetological bulletin
Vol. 2011., No 1.
urednik/editor: Dušan Jelić
Hrvatsko herpetološko društvo - HYLA
Croatian Herpetological Society - HYLA
Zagreb, lipanj 2011.

Impressum
HYLA, HERPETOLOGICAL BULLETIN
Ključni naslov: Hyla (Zagreb)
Skraćeni ključni naslov: Hyla (Zagreb)
Izdavač/Publisher:
Hrvatsko herpetološko društvo - HYLA
Croatian Herpetological Society - HYLA
Radučka cesta 15,
10 000 Zagreb, Croatia
Urednik/Editor:
Dušan Jelić, prof. biol.
ISSN: 1848-2007

Sadržaj:
Contents:
O HHD HYLA ……………………………………………………………………… 1
About HHD HYLA ………………………………………………………………… 2
Kuljeri ć, M. - Lombardijska smeđa žaba, Rana latastei B o u l e n g e r , 1 8 7 9 (Amphibia, Anura) …………………………………………………………… 3
Italian agile frog, Rana latastei B o u l e n g e r , 1 8 7 9 (Amphibia, Anura)..
Treer, D. - Črno blago Bele Krajine – čovječja ribica nazvana krampus………… ... 21
Black treasure from Bela Krajina (White Region) - the olm named devil…….
Marchand, M. A. – Preliminary data on a Vipera berus bosniensis (B o e t t g e r , 1 8 8 9 ) population from Central Croatia …………………………………….. 29
Preliminarni podatci o istraživanju populacije Vipera berus bosniensis (B o e t t g e r , 1 8 8 9 ) u Hrvatskoj …………………………………………...
Vörös, J. & Jelić, D. - First steps to survey chytrid fungus in Croatia…………...... 31
Prvi koraci u istraživanju „chytrid“ gljivica u Hrvatskoj……………………..
Jeran, N., Đuri ć, P., & Žganec, K. – Finding of the Alpine salamander (Salamandra atra L a u r e n t i , 1 7 6 8 ; Salamandridae, Caudata) in the Nature Park Žumberak - Samoborsko gorje (NW Croatia)…………………………..……. 35
Nalaz crnog daždevnjaka (Salamandra atra L a u r e n t i , 1 7 6 8 ; Salamandridae, Caudata) u Parku prirode Žumberak - Samoborsko gorje (SZ Hrvatska)........
Barišić, F. & Bogdanović, T. - Gmazovi (Reptilia) Parka prirode Papuk ………. .. 47
Reptiles (Reptilia) of the Papuk Nature Park ………………….…………….
Jelić, D. & Bogdanović, T. – Preliminary data on existence of Zootoca vivipara ssp. pannonica (L a c & K l u c h , 1 9 6 8 ) in Croatia ……………………..... 77
Preliminarni podatci o postojanju Zootoca vivipara ssp. pannonica (L a c & K l u c h , 1 9 6 8 ) u Hrvatskoj ……………………………………………….
Upute autorima ……………………………………………………………………. 81
Author guidelines …………………………………………………………………. 85

1
O HHD – Hyla
Hrvatsko herpetološko društvo – Hyla osnovano je 1997. godine pod imenom "Društvo za
zaštitu i proučavanje vodozemaca i gmazova Hrvatske-Hyla". Osnovano je od strane biologa i
zaljubljenika u vodozemce i gmazove zbog potrebe zaštite ovih životinja, koje su često i
bezrazložno proganjane i ubijane, te ekosustava i mnogih staništa na kojima obitavaju ove, ali
i ostale skupine životinja.
Društvo je 2004. preimenovano u današnji naziv te sa razvila unutrašnja infrastruktura u vidu
web stranice (www.hyla.hr) i mailing liste koje održavaju povezanost članova i mreže
regionalnih i lokalnih udruga i organizacija partnera. Društvo je registrirano kao strukovna
organizacija te je većina članova biološke struke. Međutim, otvoreni smo za sve koje zanima
zaštita i proučavanje hrvatske herpetofaune (vodozemaca i gmazova) i staništa.
HHD-Hyla je punopravna članica IUCN (International Union for Conservation of Nature),
najstarije i najveće međunarodne mreže za zaštitu prirode koja pod svojim okriljem okuplja
više od 1000 članica - nevladinih i državnih organizacija - u više od 160 zemalja širom
svijeta.
Projekti i aktivnosti usmjereni su na istraživanja te zaštitu vrsta i staništa, edukaciju lokalnog
stanovništva i šire javnosti (u sklopu projekata ali i zasebna predavanja i radionice), edukaciju
studenata te izdavanje publikacija i ostalog edukativnog materijala. Društvo je aktivno na
nacionalnoj razini te provodimo projekte u raznim dijelovima Hrvatske uz suradnju s
državnim i lokalnim institucijama, udrugama, stručnjacima u zemlji i inozemstvu, školama te
lokalnim stanovništvom.
VODSTVO DRUŠTVA
Upravni odbor
Marija Kuljerić - predsjednica
Ivona Burić - dopredsjednica
Mila Lončar - tajnica
Boris Lauš
Petra Svoboda
Nadzorni odbor
Dragica Šalamon
Olga Jovanović
Ivan Budinski
KONTAKT
Poštanski pretinac: Radučka 15, 10 000 Zagreb e-mail: [email protected]
Telefon: 095 - 9050736 (Marija Kuljerić)
098 - 9310854 (Ivona Burić)

About HHD - Hyla
Croatian Herpetological Society - Hyla was founded in 1997 under the name "Society for the
protection and study of amphibians and reptiles in Croatia-Hyla". It was established by the
biologists and nature enthusiasts because of the need to protect amphibians and reptiles,
which are often unduly persecuted and killed, as well as ecosystems and many habitats, on
which this and other groups of animals reside.
In 2004 Society was renamed to its present name and we developed an infrastructure, web site
(www.hyla.hr) and mailing list, through which we maintain cohesion between members and a
network of regional and local NGOs and partner organizations. Society is registered as a
professional organization, and the majority of our members are biologists. However, we are
open to all people interested in research and conservation of Croatian herpetofauna
(amphibians and reptiles) and habitats.
HHD-Hyla is a full member of the IUCN (International Union for Conservation of Nature),
the oldest and largest international network for the protection of nature, with more than 1000
members - government and non-government organizations - in over 160 countries around the
world.
Projects and activities are focused on research and protection of species and habitats,
education of the local inhabitants and public (during the projects, as well as separate lectures
and workshops), training of students and publishing of the various scientific, professional and
educational materials. Society is active at the national level and implements projects in
different parts of the Croatia in cooperation with national and local institutions, NGOs,
national and international experts and scientists, schools and local inhabitants.

HYLA VOL. 2011. No. 1 Str. 3-20 SPECIES SUMMARY
3
Lombardijska smeđa žaba, Rana latastei Boulenger, 1879 (Amphibia, Anura)
Italian agile frog, Rana latastei Boulenger, 1879 (Amphibia, Anura)
MARIJA KULJERIĆ
Hrvatsko herpetološko društvo – Hyla, Radučka 15, 10000 Zagreb, Croatia, [email protected]
SAŽETAK
U ovom radu prikazana su dosadašnja saznanja o biologiji i ekologiji lombardijske smeđe
žabe (Rana latastei, Boulenger 1879) uz pregled znanstvene i stručne literature.
SUMMARY
This paper presents the current knowledge about the biology and ecology of Italian Agile frog
(Rana latastei, Boulenger 1879) throught review of scientific literature.
UVOD
Lombardijska smeđa žaba (Rana latastei) je mala do srednje velika smeđa žaba, izgledom
vrlo slična široko rasprostranjenoj šumskoj smeđoj žabi (Rana dalmatina), kojoj je, uz vrstu
Rana graeca, i najsrodnija među smeđim žabama Zapadnog Paleartika (VEITH I SUR, 2003).
Taksonomija vrste se nije mijenjala od godine opisa te ne postoje opisane podvrste. Dugo
vremena se smatralo da između pojedinih populacija ne postoje izraženije varijacije u
morfologiji (SCHMIDTLER, 1977, ARNOLD I OVERDEN, 2002), kao ni velikih genetskih razlika
(CAPULA I SUR, 1991). Međutim, posljednja genetska istraživanja pokazuju značajnu
varijabilnost između populacija. Genetska raznolikost izraženo opada od istoka prema zapadu
areala vrste, što ukazuje na vjerojatnu postglacijalnu kolonizaciju "padano-veneta" nizine iz
refugija smještenog na istočnom dijelu današnjih granica rasprostranjenosti (GARNER I SUR,
2004a). Nedavna istraživanja isto tako ukazuju na moguće postojanje varijabilnosti

morfoloških karakteristika i glasanja između i unutar pojedinih populacija ove vrste
(GIACOMA I SUR, 2004, MARZONA I SUR, 2004).
OPIS VRSTE
Karakteriziraju je slijedeće morfološke osobine: dugačke stražnje noge (kod noge istegnute
prema glavi peta doseže preko vrha njuške), bubnjić je relativno malen i smješten malo dalje
od oka, uglavnom ima kratku njušku sa zašiljenim vrhom iako postoje varijacije unutar i
između populacija, a na leđima ima razdvojene dorzolateralne nabore. Obojenost je tipična za
europske smeđe žabe. S dorzalne strane je relativno uniformna, sivkasto ili crvenkasto smeđe
boje sa vrlo malo tamnih pjega (osim pruga na nogama). Moguća je prisutnost tamne pruge
između očiju te obrnuti znak V između ramena (ARNOLD I OVERDEN, 2002). Na bočnim
stranama glave prisutna je tamna maska karakteristična za smeđe žabe.
Slika 1. Lombardijska smeđa žaba (Rana latastei), odrasla jedinka
Picture 1. Italian agile frog (Rana latastei), adult

HYLA VOL. 2011. No. 1 Str. 3-20 SPECIES SUMMARY
5
Osobine po kojima se najlakše raspoznaje od često simpatrične šumske smeđe žabe, R.
dalmatina, su bijela pruga iznad gornje usne koja završava oštro u ravnini oka te tamno
ispjegano grlo sa svjetlom prugom po sredini. Trbuh je bijeli, često sa sivkastim oznakama.
Iako nema jako izraženog seksualnog dimorfizma, mužjaci za vrijeme sezone parenja
razvijaju tamno smeđe palčane žuljeve na prednjim nogama. Ženke su često intenzivnije
obojane od mužjaka, te mogu imati i crvene ili narančaste pjege na rubovima trbušnog
predjela (POZZI, 1980). Seksualno aktivni mužjaci također mogu razviti tamno crveno-smeđe
pjege na grlu te narančasto do crvenkasto obojenje na unutrašnjem dijelu bokova (NOLLERT I
NOLLERT, 1992, ARNOLD I OVERDEN 2002).
Ukupna dužina jedinki (SVL) je varijabilna između populacija. Obično su ženke veće od
mužjaka, ali mužjaci imaju veće i duže prednje noge. Ženke mogu narasti do 75 mm u dužinu,
mužjaci do 58 mm. Najmanje spolno zrele jedinke su dugačke oko 35 mm, prosječna dužina
mužjaka je 40-45 mm, a ženki 45-55 mm (POZZI, 1980, MARZONA I SUR., 2004, FICETOLA,
2005) .
Slika 2. Lombardijska žaba - ventralna strana
Picture 2. Italian agile frog - ventral side

Mužjaci lombardijske žabe glasaju se za vrijeme parenja kako bi privukli ženke. Rijetki su
slučajevi glasanja van sezone parenja. Zabilježena su dva različita tipa glasanja odnosno pjeva
te takozvani "release call". Glasanje koje ljudsko uho može čuti na kratkoj udaljenosti je
dugački, nježni i slabo primjetni zvuk, a mužjaci ga proizvode u vodi i ponekad na površini
vode, tijekom dana i noći. Drugi tip glasanja, kratki i intenzivni, može se čuti samo ispod
vode, a takozvani "release call" je kratki ponavljajući "kek...kek..." (TRILAR, 2003, GIACOMA I
SUR, 2004, FICETOLA, 2005).
RASPROSTRANJENOST
Lombardijska smeđa žaba je endem šireg područja Padano-Veneta nizine (porječje rijeke Po)
te se većina populacija nalazi u Italiji. Izvan granica Italije zabilježeni su lokaliteti u
Švicarskoj (područje Mendrisiotto, južni Ticino), u Sloveniji na području Vipavske doline i
doline rijeke Branice te na području središnje i sjeverne Istre u Hrvatskoj (CAPULA I SUR,
1991, GROSSENBACHER I SUR, 2002, POBOLJŠAJ I LEŠNIK, 2003, KULJERIĆ, 2006, SINDACO I
SUR, 2008).

HYLA VOL. 2011. No. 1 Str. 3-20 SPECIES SUMMARY
7
Slika 3. Područje rasprostranjenosti lombardijske žabe. Izvor: Sindaco i sur, 2008. www.iucnredlist.org
Picture 3. Distribution area of the Italian agile frog. From: Sindaco et al., 2008. www.iucnredlist.org
U Hrvatskoj je prvi put zabilježena 1879. god. u okolici Pazina (CEI, 1944). Zatim je
Schmidtler (1977) pronalazi u dolini rijeke Mirne (Motovunska šuma), na čijem je području
zabilježena u nekoliko navrata do 2005. god. (KOBAŠLIĆ, 2002, SCHMIDTLER 1977, BURLIN I
DOLCE, 1986, BRESSI, 1995, KLETEČKI, 2003). Burlin i Dolce (1986) navode i nove nalaze za
Hrvatsku: 3 male doline prema granici sa Slovenijom. Recentna istraživanja ove vrste u
Hrvatskoj provodi Hrvatsko herpetološko društvo - Hyla s početkom u 2005. godini tijekom
kojih je zabilježeno mnogo novih lokaliteta. Danas poznati areal vrste u Hrvatskoj obuhvaća
oko 310 km2 - područje od granice sa Slovenijom sve do pazinskog sliva na jugu (Lipa,
Pazinčica i Beramski potok) i gornjeg toka rijeke Mirne na istoku (područje naselja Kotli).
Najviši zabilježeni lokaliteti nalaze se na 355 m (potoci Pregon, Lipa te Mlaka kod Čepića).

Najveća populacija nalazi se na području Motovunskih šuma odnosno dolina rijeka Mirne i
Butonige (KULJERIĆ, 2008, KULJERIĆ, 2009).
STANIŠTE
Rana latastei je vrsta koja preferira sjenovita mikrostaništa s konstantnim vlažnim uvjetima te
bogato razvijenim pridnenim slojem vegetacije. Primarno stanište su listopadne šume s
visokim nivoom podzemnih voda, odnosno staništa poplavnih nizina rijeka gdje prevladavaju
crna joha (Alnus glutinosa), vrbe (Salix spp.), hrast lužnjak (Quercus robur), grab (Carpinus
sp.) i jasen (Fraxinus sp.). Osim u šumama poplavnih nizina nalazimo je i u vlažnim šumama
okolnog brežuljkastog područja.
Slika 4. Tipično šumsko stanište lombardijske žabe u rano proljeće
Picture 4. Tipical forest habitat of the Italian agile frog in early spring

HYLA VOL. 2011. No. 1 Str. 3-20 SPECIES SUMMARY
9
Na povoljnim nizinskim staništima možemo je naći u većem broju po cijeloj šumi, dok se na
višim lokalitetima zadržava na vlažnom šumskom staništu u neposrednoj blizini vodotoka
(POZZI, 1980, DOLCE I SUR, 1985., FASOLA I SUR, 2000, VERCESI I SUR, 2000, FICETOLA, 2005,
KULJERIĆ, 2006). U vodu ulazi samo za vrijeme parenja, ostatak vremena provodi na kopnu.
Povremeno se može naći i na vlažnim livadama vezanim uz listopadne šume, plantažama
topola, kao i na krškom području (BRESSI, 2001, ARNOLD I OVENDEN, 2002, EDGAR I BIRD,
2006). Dolazi do 500 m nadmorske visine, ali su lokaliteti iznad 300 m rijetki
(GROSSENBACHER, 1997, FICETOLA, 2005, EDGAR I BIRD, 2006). Tako naprimjer BARBIERI I
BERNINI (2004) navode da od 75 istraživanih populacija na području Italije čak 85% dolazi na
nadmorskim visinama manjima od 300 m.
Mrijest polažu u mirnim, sporo protočnim vodenim tijelima unutar ili vrlo blizu šumskih
područja. Često su to mali bazeni u potocima sa čistom vodom, mrtvi rukavci i kanali te, kad
prije navedeno nije dostupno, lokve i ostala mirna i sporoprotočna vodena staništa. Za razliku
od drugih žaba područja izbjegava sunčana i otvorena staništa za polaganje jaja te preferira
zasjenjena mjesta (POZZI, 1980, BERNINI I SUR, 2004, FICETOLA I DE BERNARDI, 2004).
ŽIVOTNI CIKLUS I PONAŠANJE
R. latastei je vrsta koja se razmnožava u rano proljeće, većinom od sredine veljače do sredine
travnja (POZZI, 1980, GROSSENBACHER I SUR, 2002). Ženke se na lokalitetima razmnožavanja
zadržavaju nekoliko sati do nekoliko dana dok ne polože mrijest, a mužjaci ostaju na samim
mrjestilištima ili u neposrednoj blizini duže vrijeme, čak i do 4-5 tjedana (VERCESI I SUR,
2000). Jaja polažu u kuglastim nakupinama mrijesta, koje su kompaktnije i s manjim
promjerom želatinozne ovojnice jaja nego kod drugih smeđih žaba područja. Mrijest pričvrste
za otpalo i potopljeno suho granje, potopljene grane obalne vegetacije i drugu podvodnu
vegetaciju na prosječnoj dubini od 10-50 cm. U optimalnim uvjetima mnogo ženki polaže

mrijest jedan do drugoga na istoj mikro-lokaciji, te na jednom komadu potopljenog drveta
možemo naći čak 150 i više pojedinačnih mrijestova u grozdastoj formaciji (POZZI, 1980,
DOLCE I SUR, 1992). Prema starijim podacima pojedinačna nakupina sadržava 95-1200 jaja, u
prosjeku 300-400 (POZZI, 1980, ARNOLD I OVERDEN, 2002), međutim novija istraživanja
navode 676-2720 jaja (BERNINI I SUR, 2004). Moguće je da razlika u broju jaja predstavlja
metodološku pogrešku kod ranijih prebrojavanja budući je kod ove vrste vrlo teško prebrojati
jaja zbog vrlo kompaktnog mrijesta.
Slika 5. Mrijest lombardijske žabe
Picture 5. Spawn of the Italian agile frog
Brzina razvoja punoglavaca iz jaja ovisi o temperaturi vode te su za vrijeme razvoja jaja u
prirodi zabilježene vrijednosti od 1,5 do 15,5°C. U takvim uvjetima punoglavci se izliježu
nakon 10-25 dana. Daljnji razvoj odnosno metamorfoza traje oko 3 mjeseca i krajem lipnja -
početkom srpnja mlade žabe dužine oko 15 mm izlaze iz vode (POZZI, 1980, BARBIERI I
BERNINI, 2004). Granicom između juvenilnih i spolno zrelih žaba smatra se prosječna dužina
od 35 mm koju dosegnu na kraju prve sezone.

HYLA VOL. 2011. No. 1 Str. 3-20 SPECIES SUMMARY
11
R. latastei veći dio godine proboravi na kopnu dok se u vodi zadržava u periodu
razmnožavanja. Žive u blizini mrjestilišta i nemaju izražen jak selidbeni nagon. Obično
hiberniraju na udaljenosti do nekoliko stotina metara od vode, a vrlo rijetko do 1 km (POZZI,
1980, DOLCE I SUR, 1985). U razdoblju od studenog do ranog proljeća hiberniraju na kopnu,
uz privremene prekide za vrijeme toplijih dana. Tijekom hibernacije, ali i drugih perioda
smanjene aktivnosti često se koriste nastambama malih sisavaca (POZZI 1980). Hibernacija
može biti privremeno prekinuta za vrijeme povišenih temperatura. U periodu od sredine
veljače do sredine ožujka mužjaci prvi dolaze na mrjestilišta, dok ženke dolaze nekoliko dana
kasnije. Tijekom razmnožavanja najveća aktivnost odraslih je oko mrjestilišta kada su aktivne
i danju i noću. Samo parenje najčešće se odvija u sumrak i zoru (POZZI, 1980, DOLCE I SUR,
1985, VERCESI I SUR, 2000, ARNOLD I OVERDEN, 2002). Tijekom proljeća i ranog ljeta, kada je
u šumi pridneni sloj vegetacije slabije razvijen te mu je smanjena relativna vlažnost, aktivnost
odraslih je također smanjena. Aktivnost se na površini (izvan podzemnih nastamba) povećava
za kišnih dana te tijekom ljeta i rane jeseni, kada vegetacija postaje sve gušća i pridneni sloj
sve vlažniji. Kako jesen odmiče možemo naći mnoge jedinke i van šume (POZZI, 1980, DOLCE
I SUR., 1985). Na nadzemnim staništima tijekom ožujka, travnja i veljače prevladavaju odrasle
jedinke, tijekom lipnja i srpnja tek preobražene jedinke, tijekom kolovoza i rujna subadultne
jedinke, a u listopadu opet odrasle (MAZZOTTI I SUR, 2004).
Često se nalazi u simpatriji sa šumskom smeđom žabom (Rana dalmatina), međutim veća
specijaliziranost pri odabiru staništa ograničava kompeticiju sa tom, ali i drugim vrstama
žaba. U prirodi nisu poznati hibridi između R. dalmatina i R. latastei (HETTYEY I PEARMAN,
2003).
Lombardijska žaba ima nešto kraći životni vijek u odnosu na srodne vrste. Spolnu zrelost
dostižu nakon jedne godine (SVL ≥ 35 mm), a mogu doživjeti 4 do 5 godina (GUARINO I
MAZZOTTI, 2004, BARBIERI I BERNINI, 2004). U prosjeku ipak žive samo oko dvije do tri

godine (EDGAR I BIRD, 2006). Zbog toga, te velike smrtnosti (posebice zimi), imaju vrlo velik
obrtaj populacije te im brojnost može znatno varirati između pojedinih godina (CORBETT,
1989). To znači da se pojedine populacije mogu značajno povećati nakon samo jedne dobre
sezone. Međutim zbog velike fragmentiranosti staništa (time i odvojenosti populacija) uslijed
stohastičkih događaja i loših uvjeta u pojedinim sezonama može doći i do izumiranja
pojedinih populacija. Treba napomenuti da je moguće da nakon blagih zima, zbog veće
aktivnosti i trošenja energetskih zaliha, samo 35-40% ženki polaže jaja (POZZI, 1980), što u
sinergiji s ostalim negativnim utjecajima može drastično smanjiti brojnost populacije u
kratkom roku.
PREHRANA I PREDATORI
Prehrana lombardijske smeđe žabe, kao i ostalih pripadnika istog roda (Rana sp.), usmjerena
je na male životinje prizemnog šumskog sloja i podzemlja. Većinu hrane nalazi u sloju lišća
na tlu šume te u nastambama malih sisavaca, vrlo rijetko i na obalama vodenih staništa. Slabo
je selektivna za određenu vrstu plijena te sastav prehrane dijelom odražava prisutnost
različitih vrsta na pojedinom području. Glavninu hrane čine paučnjaci (Aranea, Opiliones,
Acarina), kukci (Diptera, Hemiptera, Coleoptera, Orthoptera, Lepidoptera, Hymenoptera...),
gujavice, mekušci (Gastropoda), izopodni kopneni rakovi rakovi ali i drugi beskralježnjaci.
Budući da jednim dijelom živi u podzemlju hrani se i maločetinašima (Oligochaeta). Za
perioda razmnožavanja hrani se i vodenim kukcima. Nema značajne razlike u ishrani odnosno
tipu plijena između odraslih i juvenilnih jedinki (POZZI 1980, MANSI 1992).
Kao i većina drugih žaba, Rana latastei je plijen mnogim predatorima. Razni vodeni kukci,
poput vodenog škorpiona (Nepa cinerea), vodenih stjenica odnosno nauznačarki
(Notonectidae), ličinki vretenaca (Odonata) - posebno porodice Aeshnidae, hrane se
punoglavcima od najranijeg do najkasnijeg stadija. Zanimljiv je međusobni odnos

HYLA VOL. 2011. No. 1 Str. 3-20 SPECIES SUMMARY
13
lombardijske smeđe žabe i kornjaša obrubljenog kozaka (Dytiscus marginalis). Odrasle se
žabe hrane ličinačkim razvojnim stadijima kozaka, ličinke kozaka se hrane punoglavcima,
dok odrasli kozaci jedu jaja, punoglavce i tek preobražene jedinke. Iako zbog svoje
kompaktnosti mrijest R. latastei u pravilu ne predstavlja dobar plijen, njime se ipak u
određenoj mjeri hrane već navedeni kozaci, zatim pijavice (Haemopsis sp.), veliki vodenjak
(Triturus carnifex), pojedine ptice močvarice (Anas sp., Gallinula chlorops) te sisavci poput
lisice (Vulpes vulpes). Punoglavcima se hrani niz životinja poput raznih vodenih kukaca i
pijavica, vodenjaka, riba, barske kornjače (Emys orbicularis) i zmija roda Natrix te mnogih
ptica močvarica. Međutim, u malim zasjenjenim vodenim tokovima koje ova vrsta preferira,
broj predatora obično je manji nego u otvorenijim vodenim tijelima. Preobražene jedinke
(mlade i odrasle žabe) su isto hrana za mnogo različitih vrsta životinja poput zmija (Natrix
natrix, povremeno i druge vrste), mnogih ptica močvarica, vrane (Corvus cornix), sova (Strix
aluco, Asio otus, Tyto alba), ptica grabljivica (Buteo buteo, Milvus sp.) i mnogih sisavaca
(npr. Erinaceus europaeus, Vulpes vulpes, Meles meles, Mustela nivalis, Rattus norvegicus,
Arvicola terrestris) (POZZI, 1980, VERCESI I SUR, 2000). Ovdje treba spomenuti i alohtone
predatorske vrste – prvenstveno razne vrste riba te crvenouha kornjača (Trachemys scripta
elegans), vrste koje su primjećene na području rasprostranjenosti lombardijske žabe (EDGAR I
BIRD, 2006).
UGROŽENOST I STATUS VRSTE
Rana latastei je endemska vrsta većim dijelom ograničena na sliv rijeke Po u Italiji, područje
kojega se nalazi pod velikim ljudskim pritiskom. Iako ima relativno široko područje
rasprostranjenosti, ugrožena je zbog vrlo ograničene količine te fragmentiranosti povoljnog
staništa, posebno preostalih poplavnih šuma, što je dodatno naglašeno zbog ekoloških
karakteristika vrste (ANDREONE I LUISELLI, 2000, BARBIERI I BERNINI, 2004). Glavni razlozi

ugroženosti su uništavanje velikog dijela vlažnih šuma "padano-veneta" nizine i istarske
regije, prvenstveno zbog intenzivnog razvoja poljoprivrede ali i infrastrukture, što uključuje
vodne regulacije, krčenje šuma, fragmentaciju staništa, snižavanje razine podzemnih voda i
zagađenje močvarnih staništa (kemikalije i otpadne vode). Dodatni razlozi ugroženosti su
stradavanje na prometnicama, unos invazivnih predatorskih riba u vodena staništa i
potencijalni unos novih patogena na područja gdje lombardijska žaba obitava (SINDACO I SUR,
2008). Naime, posljednja genetska istraživanja su pokazala da redukcija genetske raznolikosti
(od istoka prema zapadu areala vrste), gubitak imunokompetentnih sposobnosti te međusobna
izoliranost populacija zbog ozbiljnog uništavanja staništa čine ovu žabu vrlo podložnom
raznim novim patogenima poput Ranavirusa i gljivice Batrachochytrium dendrobatidis,
pogotovo jer je već zabilježena prisutnost Bd u nekih punoglavaca lombardijske žabe. Zbog
navedenih razloga veliki dio zapadnog i središnjeg areala vrste je u opasnosti od eliminacije
(GARNER I SUR, 2004a, GARNER I SUR, 2004b, PEARMAN I GARNER, 2005).
Glavni razlozi ugroženosti R. latastei u Hrvatskoj su također nestanak, fragmentacija i
degradacija staništa. Redom su to krčenje šuma za poljoprivredne površine, urbanizaciju i
ostale namjene, zatim regulacija, kanaliziranje i betoniranje vodenih tokova gdje se mora
naglasiti regulacija srednjeg toka rijeka Mirne i Butonige te izgradnja akumulacije Butoniga
(70-tih godina 20. stoljeća) što je uzrokovalo propadanje staništa najveće populacije u
Hrvatskoj, čišćenje i siječa vegetacije na rubovima vodenih tokova i kanala, eksploatacija
vodenih resursa. Veliku prijetnju predstavljaju planirane daljnje regulacije sliva rijeke Mirne,
uz izgradnju nekoliko retencija na tokovima koji su potvrđeno stanište vrste. Areal
lombardijske žabe u Hrvatskoj je vrlo fragmentiran i postoji nekoliko zasebnih subpopulacija.
Najveći postotak jedinki nalazi se u Motovunskoj šumi, ali je i to područje podijeljeno
kanalom Mirne i brzom cestom, te dodatno ispresijecano manjim prometnicama. Osim što
uzrokuju fragmentaciju staništa, prometnice su razlog direktnog masovnog stradavanja

HYLA VOL. 2011. No. 1 Str. 3-20 SPECIES SUMMARY
15
jedinki. Kao uzroke ugroženosti treba navesti i pesticide, herbicide i umjetna gnojiva te
otpadne vode domaćinstava i industrije. Na jednom od lokaliteta u blizini Motovuna (Brkač i
Vižinadske vale) u tijeku je izgradnja golf kompleksa što vodi potpunom uništavanju staništa
i dodatnom pritisku na vodne resurse Motovunske šume i okolice (KLETEČKI, 2006, KULJERIĆ,
2008, KULJERIĆ, 2009).
Lombardijska žaba je zaštićena nacionalnom legislativom u svim državama na području
rasprostranjenosti, u Hrvatskoj se nalazi u kategoriji strogo zaštićenih svojti. Na nivou Europe
(time i globalne rasprostranjenosti) navedena je na Dodatku II Konvencije o zaštiti europskih
divljih vrsta i ptitodnih staništa (Bernska konvencija), kao i na Dodacima II i IVa Direktive o
zaštiti prirodnih staništa i divlje faune i flore (Direktiva o staništima) Europske komisije.
Provedbom Direktive o staništima zemlje članice EU su proglasile niz zaštićenih područja za
R. latastei u sklopu Natura 2000 ekološke mreže. Na IUCN-ovom Crvenom popisu ugroženih
vrsta navedena je kao osjetljiva (VU) vrsta, odnosno svojta kojoj prijeti visoki rizik od izumiranja
u prirodi (SINDACO I SUR, 2008). Nalazi se i na crvenim popisima zemalja gdje je rasprostranjena:
Italija (VU), Švicarska (EN), Slovenija (E) i Hrvatska (VU).

L ITERATURA
ANDREONE, F., LUISELLI, L. (2000): The Italian batrachofauna and its conservation status: a
statistical assessment. Biological Conservation 96: 197-208.
ARNOLD, N., OVERDEN, D. (2002): Collins Field Guide: Reptiles and Amphibians of Britain
and Europe. Collins Publishers, London.
BARBIERI, F., BERNINI, F. (2004): Distribution and status of Rana latastei in Italy (Amphibia,
Ranidae). Italian Journal of Zoology, 71(4): 91-94.
BERNINI F., A. GENTILLI, E. MERLI, E. RAZZETTI. 2004. Rana dalmatina and R. latastei:
Habitat selection, fluctuation in egg clutch deposition and response to exceptional floods in
northern Italy.
BRESSI, N. 1995. Catalogo della collezione erpetologica del Museo civico di Storia naturale di
Trieste. I - Amphibia. Cataloghi I, Museo civico di Storia Naturale di Trieste.
BRESSI, N., (2001): Prima segnalazione di Rana latastei Boulanger, 1879 in ambiente carsico.
Pianura - Scienze e storia dell'ambiente padano. N. 13/2001. pp. 233-235.
BURLIN, M., DOLCE, S. (1986): Osservazioni faunistiche sull´erpetofauna dell´Istria. I:
“Amphibia”. Atti Museo civico di Storia Naturale di Trieste, 39(1): 65-85.
CAPULA M., DOLCE S., LAPINI L., NASCETTI G. (1991): Electrophoretic analysis of Rana
latastei populations (Amphibia: Ranidae) from Italy and Istria (NW Yugoslavia). Gortania –
Atti Museo Friulano di Storia Naturale 13: 203-212.
CEI, G. (1944): Analisi biogeografica e ricerche biologiche e sperimentali sul ciclo sessuale
annuo delle rane rosse d’Europa. Monitore Zoologico Italiano, 54: 1-117.
CORBETT, K. (1989): Conservation of European Reptiles and Amphibians. The Conservation
Committee of the Societas Europaea Herpetologica (IUCN/SSC European Reptile and
Amphibian Specialist Group), London.

HYLA VOL. 2011. No. 1 Str. 3-20 SPECIES SUMMARY
17
DOLCE, S., LAPINI, L., STOCH, F., (1985): Indagini ecologiche su Rana latastei Boul.
(Ampibia, Anura) nei boschi della bassa pianura friulana (Italia nordorientale). Gortania 6,
227-238.
DOLCE, S., LAPINI, L., STERGULO, F., (1992): Contributo prleminare allo studio
dell'erpetofauna della bassa pianura friulana. Note eco-etologiche sugli anfibi e i rettili di
Bosco Baredi e Selva di Arverchi (Muzzara del Turgano, Udine). CNR, Roma.
EDGAR, P., BIRD, D. (2006): Action Plan for the Conservation of the Italian Agile Frog Rana
latastei in Europe. The Herpetological Conservation Trust, Bournemouth, Dorset.
FASOLA, M., ANNICHINI, I., VEROLI, A., (2000): Anfibi di un gradiente geografico padano. In:
Giacoma, C. (Ed.), Atti del I Congresso Nazionale della Societas Herpetologica Italica,
Museo Regionale di Scienze Naturali di Torino, Torino, 1996, pp. 493-497.
FICETOLA, G.F. (2005): Distribution and diversity of the semi-aquatic herpetofauna in the
river Po basin: from the microhabitat analysis to the regional perspective. Doctoral thesis,
Dipartimento di Biologia, Facolta di Scienze Matematiche, Fisiche e Naturali, Università
degli studi di Milano.
FICETOLA G. F., DE BERNARDI, F. (2004): Amphibians in a human-dominated landscape: the
community structure is related to habitat features and isolation. Biological Conservation 119:
219–230.
GARNER, T., PEARMAN, P., ANGELONE, S. (2004a): Genetic diversity across a vertebrate
species’ range: a test of the central–peripheral hypothesis. Molecular Ecology 13: 1047–1053.
GARNER, T.W.J., PEARMAN, P.B., CUNNINGHAM, A.A., FISHER, M.C., (2004b): Population
genetics and disease threats acoss the entire range of Rana latastei. pp. 62. U: V° Congresso
Nazionale della Societas Herpetologica Italica, 29. settembre - 3. ottobre 2004. Abstract book,
Calci (Pisa), Universita di Pisa, S.H.I., Pisa.

GIACOMA, C., SEGLIE, D., MARZONA, E. (2004): Analisi acustica in Rana latastei: effetto dei
constraints e variabilita intrapopolazionale, risultati preliminari. pp. 63. U: V° Congresso
Nazionale della Societas Herpetologica Italica, 29. settembre - 3. ottobre 2004. Abstract book,
Calci (Pisa), Universita di Pisa, S.H.I., Pisa.
GUARINO, F.M., MAZOTTI, S. (2004): Applicazioni scheletrocronologiche in Rana latastei. pp.
63. U: V° Congresso Nazionale della Societas Herpetologica Italica, 29. settembre - 3. ottobre
2004. Abstract book, Calci (Pisa), Universita di Pisa, S.H.I., Pisa.
GROSSENBACHER K. 1997. Rana latastei Boulenger, 1879. pp. 146-147. U: Gasc J.-P. i sur.
(ur.) Atlas of Amphibians and Reptiles in Europe. Reedition. Museum national d'Histoire
naturelle, Paris, 520 p. (Patrimoines naturels, 29). Paris.
GROSSENBACHER, K., LIPPUNER, M., ZUMBACH, S., BORGULA, A., LÜSCHER, B. (2002):
Phenology and reproduction of the 3 brown frog species Rana latastei, R. dalmatina, R.
temporaria; Development and status of the R. latastei populations in Mendrisiotto, southern
Ticino, Switzerland. Atti del terzo Convegno “Salvaguardia Anfibi”, Lugano, 23-24 giugno
2000 – Cogecstre Ediz., Penne, 2002: 91 – 100.
HETTYEY, A., PEARMAN, P.B. (2003): Social environment and reproductive interference affect
reproductive success in the frog Rana latastei. Behavioral Ecology 14: 294–300.
KLETEČKI, E. (2003): Talijanska žaba (Rana latastei) vrsta koja nestaje. Meridijan - časopis za
zemljopis, povijest, ekologiju i putovanja, 10(74): 46-48.
KLETEČKI, E. (2006): Rana latastei – lombardijska žaba. U: Crvena knjiga vodozemaca i
gmazova Hrvatske. Ministarstvo kulture, Državni zavod za zaštitu prirode, Zagreb.
KOBAŠLIĆ, A. (2002): Rasprostranjenost vodozemaca Hrvatske prema podacima Hrvatskog
prirodoslovnog muzeja u Zagrebu. Diplomski rad. Prirodoslovno – matematički fakultet,
Sveučilište u Zagrebu.

HYLA VOL. 2011. No. 1 Str. 3-20 SPECIES SUMMARY
19
KULJERIĆ, M. (2006): Rana latastei, lombardijska žaba. Izvještaj 2006., Nacionalni program
monitoringa biološke raznolikosti. Hrvatsko herpetološko društvo-Hyla, Zagreb.
KULJERIĆ, M. (2008): Rana latastei, lombardijska žaba. Izvještaj 2007.-2008., Nacionalni
program monitoringa biološke raznolikosti. Hrvatsko herpetološko društvo-Hyla, Zagreb.
KULJERIĆ, M. (2009): Rana latastei, lombardijska žaba. Izvještaj 2009., Nacionalni program
monitoringa biološke raznolikosti. Hrvatsko herpetološko društvo-Hyla, Zagreb.
MANSI, M. (1992): Remarks about the feeding habits of Rana latastei Boul. (Amphibia,
Anura) in two protected areas of the Po Valley (Northern Italy). pp. 305-308. U: Korsos, Z. i
Kiss, I. (ur). Proceedings of the Sixth Ordinary General Meeting of Societas Europaea
Herpetologica., Budapest 1991. Hungarian Natural History Museum, Budapest.
MARZONA, E., SCEGLIE, D., GIACOMA, C. (2004): Variabilità morfometrica, età e dimorfismo
sessuale in differenti popolazioni di Rana latastei. pp. 63-64. U: V° Congresso Nazionale
della Societas Herpetologica Italica. 29. settembre - 3. ottobre 2004. Calci (Pisa), Programma
e Riassunti. Universita di Pisa, S.H.I., Pisa.
MAZZOTTI, S., LUNARDI, S., GUARINO, F.M. (2004): Ecologia di popolazione e ritmi
riproduttivi di Rana latastei in biotopi relitti della Pianura Padana. pp. 64. U: V° Congresso
Nazionale della Societas Herpetologica Italica. 29. settembre - 3. ottobre 2004. Calci (Pisa),
Programma e Riassunti. Universita di Pisa, S.H.I., Pisa.
NOLLERT, A., NOLLERT, C. (1992): Die Amphibien Europas. Kosmos, Stuttgart.
PEARMAN, P.B., GARNER, T.W.J. (2005): Susceptibility of Italian agile frog populations to an
emerging strain of Ranavirus parallels population genetic diversity. Ecology letters, 8: 401-
408.
POBOLJŠAJ, K., LEŠNIK, A. (2003): Strokovna izhodišča za vzpostavljanje omrežja Natura
2000: Dvoživke (Amphibia). Končno poročilo. Center za kartografiju faune i flore, Miklaž na
Dravskem polju.

POZZI, A. (1980): Ecologia di Rana latastei Boul. (Amphibia, Anura). Atti della Società
Italiana di Scienze Naturali e del Museo Civico di Storia Naturale di Milano, 121(4): 221-274.
SCHMIDTLER, J.F. (1977): Amphibien aus Feuchwaldern Istriens. Salamandra 13: 114-116.
SINDACO, R., ROMANO, A., ANDREONE, F., GARNER, T., SCHMIDT, B., CORTI, C., VOGRIN, M.
(2008): Rana latastei. In: IUCN 2010. IUCN Red List of Threatened Species. Version 2010.4.
<www.iucnredlist.org>. Preuzeto 18. siječnja 2011.
TRILAR, T. (2003): Slovenske žabe. Stereo Compact Disc Audio, Prirodoslovni muzej
Slovenije, Ljubljana.
VERCESI, A., BERNINI, F., BARBIERI, F. (2000): La sintopia di Rana dalmatina e Rana latastei
nei boschi planiziali del fiume Ticino: aspetti della biologia riproduttiva. Atti del I Congresso
Nazionale della Societas Herpetologica Italica (Torino, 1996). Museo Regionale di Science
naturali Torino, 2000: 353-358.
VEITH, M., KOSUCH, J., VENCES, M., (2003): Climatic oscillations triggered post-Messinian
speciation of Western Paleartic brown frogs (Amphibia, Ranidae). Molecular Phylogenetics
and Evolution 26: 310-327.

HYLA VOL. 2011. No. 1 Str. 21-27 POPULAR ARTICLE
21
Črno blago Bele Krajine – čovječja ribica nazvana krampus
Black treasure from Bela Krajina (White Region) - the olm named devil
DAG TREER
Hrvatsko herpetološko društvo – Hyla, Radučka 15, 10000 Zagreb, Croatia, [email protected]
Crna čovječja ribica (Proteus anguinus parkelj Sket & Arntzen, 1994), ili „črni močeril“ na
slovenskom, podvrsta je čovječje ribice čiji su predstavnici nađeni samo na dva susjedna
izvora u području Bele Krajine, u jugoistočnoj Sloveniji, zapadno od Žumberka (Bela Krajina
dobila je ime po brezama kojih je mnogo u tom kraju). Ime podvrste – parkelj – na
slovenskom znači krampus. Takav je naziv ova čovječja ribica dobila jer ima pigmentiranu,
sivo, smeđe do crno obojenu kožu. Slične asocijacije izazvale su i jedinke njene srodne, bijele
podvrste za koje je lokalno stanovništvo u srednjem vijeku govorilo da su mladi zmajevi.
Osim pigmentiranom kožom, crna čovječja ribica razlikuje se od bijele i po tome što odrasle
jedinke imaju razvijene oči i drugačiju tjelesnu konstituciju – poput kraće glave i drugih
morfoloških karakteristika. Analizom mitohondrijske DNA utvrđeno je da je crna čovječja
ribica genetski srodnija bijeloj čovječjoj ribici iz susjednih izvora, nego što su populacije
bijele čovječje ribice međusobno srodne diljem svojeg područja rasprostranjenja. To znači da
se bijele čovječje ribice, na primjer iz Hrvatske i jugoistočne Slovenije, međusobno genetski
razlikuju više nego što se bijele čovječje ribice iz jugoistočne Slovenije razlikuju od crne
čovječje ribice. Kako bijele čovječje ribice izvana na prvi pogled izgledaju slično, ta razlika
se najlakše može uočiti uspoređujući njihovu genetsku srodnost. Unatoč tome, crna čovječja
ribica svoju posebnost dokazuje i nešto drugačijim načinom života. Noću jedinke dolaze u
površinske vode koje okružuju ujezereni izvor gdje se vjerojatno hrane jer tu mogu pronaći
više hrane nego u hranom siromašnijem, podzemnom staništu.

Slika 1. Ujezereni izvor Jelševnik.
Figure 1. Limnocrene spring of Jelševnik.
Slika 2. Info ploča kod izvora.
Figure 2. Info table at the spring.
Crnu čovječju ribicu zapazilo je 1986. lokalno stanovništvo u izvoru Dobličica. Iako su
slovenski znanstvenici nakon toga organizirali više potraga za njom, uključivši i ronioce u

HYLA VOL. 2011. No. 1 Str. 21-27 POPULAR ARTICLE
23
potragu, kako bi je pronašli u tom izvoru i službeno dokazali njezino postojanje, crna čovječja
ribica tamo nije više pronađena. Srećom, kasnije je pronađena na susjednom izvoru Jelševnik
(slika 1, slika 2) koji sada predstavlja jedino mjesto gdje ona sigurno postoji. Zanimljivo je da
su na svim okolnim izvorima na kojima su nađene čovječje ribice, nađene samo jedinke bijele
podvrste.
Zbog postojanja deponija iznad izvora čak je i na izvoru Jelševnik opstanak ove jedinstvene
svojte bio upitan kada je došlo do podzemnog onečišćenja vodom iz deponija, nakon čega
ribice mjesecima nisu viđene u izvoru. Srećom, preživjele su nepovoljno razdoblje negdje
duboko u svom podzemnom staništu i ponovno se pojavile kada je prošlo onečišćenje. Nakon
toga je deponij saniran, pa ne predstavlja više opasnost za ovu svojtu.
Slika 3. Zaštitni kavez unutar šatora.
Figure 3. Protective cage inside the tent.

Slika 4. Zupančičeva hiša.
Figure 4. Zupančič house.
Činjenica koja će, vjerujem, mnoge oduševiti kada govorimo o crnoj čovječjoj ribici, je to što
ju možete bez mnogo muke vidjeti u njenom prirodnom staništu. Područje izvora Jelševnik je
u posjedu jedne obitelji koja ga je kupila u 80-ima. Slovenski biolozi su u međuvremenu
sagradili šator koji pokriva dio izvora. Šator omogućuje znanstvenicima i posjetiteljima da
vide crnu čovječju ribicu ponekad čak i tijekom dana, iako one obično izlaze noću iz
podzemlja.
Ribice su u šatoru zaštićene metalnim kavezom (slika 3) kroz kojeg ih lako možete vidjeti
kako izlaze ispod kamenja vjerojatno u potrazi za hranom. Kraj izvora obitelj posjeduje kuću
koja se zove, prema prezimenu obitelji, „Zupančičeva hiša“ (slika 4). Uz kuću se nalaze
objekti seoskog turizma te uzgajalište pastrva za prehranu. U kući je moguće pogledati video
o čovječjim ribicama i porazgovarati s ljubaznim i susretljivim domaćinima. Trenutno još
nema mnogo turista i ne plaća se ništa za posjetu i ulazak u šator, ali proizvodnja suvenira u
obliku crne čovječje ribice već je započela. Kada posjećujete ribice imajte na umu kako je to
svojta jedinstvena u cijelom svijetu koju je moguće naći jedino na tom lokalitetu pa ih

HYLA VOL. 2011. No. 1 Str. 21-27 POPULAR ARTICLE
25
nastojte ne previše uznemiravati. U planu su stalni videonadzor crne čovječje ribice u šatoru,
koji je možda već zaživio. Osobno sam imao priliku prvo uočiti planinskog velikog vodenjaka
(Triturus carnifex) u šatoru (slika 5), a tek nešto kasnije dvije crne čovječje ribice izašle su
kroz pukotine među kamenjem iz svojeg podzemnog staništa i učinile potragu uspješnom
(slika 6, slika 7).
Slika 5. Triturus carnifex, planinski veliki vodenjak.
Figure 5. Triturus carnifex, alpine crested newt.

Slika 6. Proteus anguinus parkelj kraj ulaza u svoje podzemno stanište.
Figure 6. Proteus anguinus parkelj by the entrance to its underground habitat.
Slika 7. Proteus anguinus parkelj.
Figure 7. Proteus anguinus parkelj.

HYLA VOL. 2011. No. 1 Str. 21-27 POPULAR ARTICLE
27
Lokalitet gdje se nalazi crna čovječja ribica relativno je blizu Zagrebu. Iz Zagreba treba
putovati do gradića koji se ponovno, «simbolično», zove Črnomelj. Tamo je informacije kako
doći do izvora Jelševnik moguće dobiti u turističkom uredu, a možete uvijek pitati i domaće
ljude da vas upute. Lako ga je pronaći, a osim toga do samog izvora moguće je doći autom po
asfaltiranoj cesti što je dobro za posjetitelje ali nije baš dobro i za zaštitu ovih endemskih
životinja. Koordinate su 45.5727 N i 15.1498 E. Preporuča se da najavite svoj dolazak
vlasnicima izvora, obitelji Zupančič, kako biste bili sigurni da će biti tamo kada dođete.
Njihovi brojevi telefona su +386 7 305 18 48, +386 31 632 636 i +386 41 632 636.
POVEZNICE ZA POSJETITELJE
http://www.zrsvn.si/sl/informacija.asp?id_meta_type=63&id_informacija=466
http://www.slovenia.info/?naravne_znamenitosti_jame=7224&lng=1
http://www.dnevnik.si/tiskane_izdaje/dnevnik/38764
http://www.slovenia.info/si/ponudniki-podezelje/Turistična-kmetija-
Zupančič.htm?ponudniki_podezelje=56&lng=1


HYLA VOL. 2011. No. 1 Str. 29-30 STUDENT PAPER SUMMARY
29
Preliminary data on a Vipera berus bosniensis (Boettger, 1889) population from Central
Croatia
Preliminarni podatci o istraživanju populacije Vipera berus bosniensis (Boettger, 1889) u
Hrvatskoj
Student: MARC-ANTOINE MARCHAND, Tour du Valat - Centre de recherche pour la conservation des
zones humides méditerranéennes - Le Sambuc- 13200 Arles, France, [email protected]
Supervisor: DUŠAN JELIĆ, State Institute for Nature Protection, Trg Mažuranića 5, 10000 Zagreb, Croatia,
During 2009 we have been studying population ecology of a low-land population of Vipera
berus bosniensis in surroundings of Okuje (Central Croatia). The main objective has been to
examine the population status by a Capture Mark Recapture method. We also noted
morphological (18 bosniensis subspecies characteristics) and physiological (body, air and
substrat temperature) data. We captured a total of 19 individuals within 14 capture sessions.
Our small data set make us use closed population estimators which are Schnabel,
Schumacher-Eschmeyer and Removal method. The effective estimation results give an
individuals number of 20-23 snakes on the 2,4 hectares of the study field, so a density of 8-9
individuals per hectare. This population can be considered having a high density compared to
ssp. berus populations from Alps, north France, Scandinavian island and Finland. Looking to
those same places for SVL length, our studied population has a higher length than ssp. berus.
For bosniensis subspecies characters, we found that only one of the three characters given by
Toth & Farkas (2004) is well represented in the population. Indeed, only 18% of 28
individuals sampling within the studied area and surrounding had two rows of sub-ocular
scales, only 25% had 23 rows of dorsal scales, but 93% had an interrupted pattern. About
temperature measurement, we took temperature of cloacae (CT), ground (GT), air at 5cm (5T)
and 60cm (60T) above the ground. We can conclude to this relation: CT > GT > 5T > 60T.
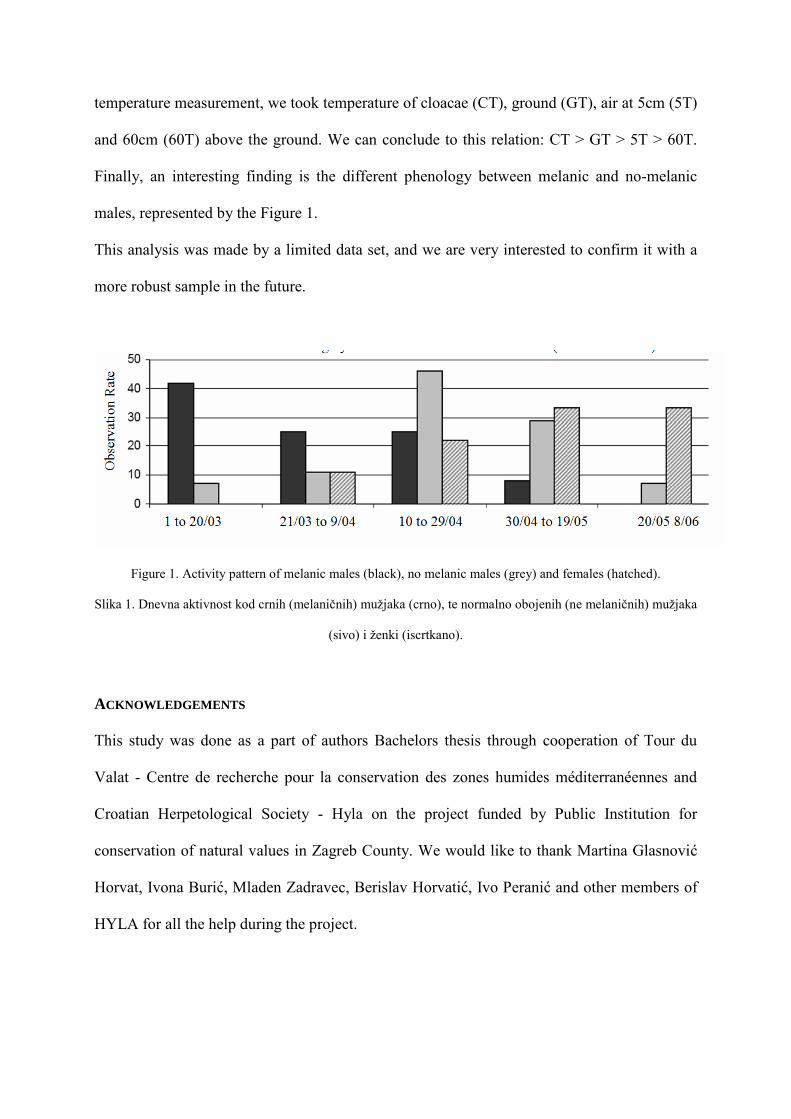
Finally, an interesting finding is the different phenology between melanic and no-melanic
males, represented by the Figure 1.
This analysis was made by a limited data set, and we are very interested to confirm it with a
more robust sample in the future.
Figure 1. Activity pattern of melanic males (black), no melanic males (grey) and females (hatched).
Slika 1. Dnevna aktivnost kod crnih (melaničnih) mužjaka (crno), te normalno obojenih (ne melaničnih) mužjaka
(sivo) i ženki (iscrtkano).
ACKNOWLEDGEMENTS
This study was done as a part of authors Bachelors thesis through cooperation of Tour du
Valat - Centre de recherche pour la conservation des zones humides méditerranéennes and
Croatian Herpetological Society - Hyla on the project funded by Public Institution for
conservation of natural values in Zagreb County. We would like to thank Martina Glasnović
Horvat, Ivona Burić, Mladen Zadravec, Berislav Horvatić, Ivo Peranić and other members of
HYLA for all the help during the project.

HYLA VOL. 2011. No. 1 Str. 31-34 SHORT NOTE
31
Fir st steps to survey chytrid fungus in Croatia
Prvi koraci u istraživanju chytrid gljivica u Hrvatskoj
JUDIT VÖRÖS*1, DUŠAN JELIĆ2
1Hungarian Natural History Museum, 1088 Budapest, Baross u. 13, Hungary, [email protected]
2State Institute for Nature Protection, Trg Mažuranića 5, 10000 Zagreb, Croatia
The Batrachochytrium dendrobatidis (Bd) (LONGCORE ET AL., 1999) is a recently emerged
amphibian pathogen, causing the disease chytridiomycosis, which has been identified as a
cause of mass amphibian mortalities and a factor influencing world-wide amphibian declines
(STUART ET AL., 2004, POUNDS ET AL., 2006). The disease has been considered responsible for
amphibian declines in the Americas, Caribbean, Africa, Australia, New Zealand, and the
range of the pathogen recently expanded into Europe (GARNER ET AL. 2005). Within Europe
the infection was detected already in about 1/3 of Europe’s amphibian fauna in 12 countries
(GARNER ET AL. 2005, VÖRÖS ET AL. 2009, SURA ET AL. 2010, SZTATECSNY & GLASER, 2011,
SPATIELEPIDEMIOLOGY, 2011).
The Easternmost occurrence of the pathogen in Europe is proved from the low elevation
mountains of Hungary, where no Bd-linked mortalities have been reported yet (VÖRÖS ET AL.,
2009). South-eastern Europe and the Balkan Peninsula seem to be mostly unexplored
regarding the distribution of the fungus. GARNER ET AL. (2005) sampled 8 individuals of Rana
latastei from low elevation area of coastal Croatia, which tested negative for Bd (GARNER,
pers. comm. 2005). Since Croatia has high elevation mountains and climatic conditions in

these regions could be suitable for Bd, a systematic survey is needed to infer the
presence/absence of the pathogen.
In 2010 we started sampling of Croatian amphibians on three species (Bombina variegata
(11), Ichthyosaura alpestris (1), Pelophylax esculentus (1)) and on two locations close to
Zagreb (Veliki Potok, Mikulići), in order to explore the possible Bd infection in the area. It is
well documented that introduced frog species can be vectors of chytridiomycosis and also
release of exotic species can be responsible for spread of the disease (GARNER ET AL., 2007,
SOTO-AZAT ET AL., 2010). For this reason we choose to start sampling mountainous habitats
near the capital.
The specimens were sampled on their back, groin and webbing area with a sterile cotton swab
(Biolab). We used the standard real-time PCR protocol (BOYLE ET AL., 2004) to detect Bd.
Tests were run in the Laboratory for Molecular Taxonomy of the Hungarian Natural History
Museum, Budapest, using an ABI Step One real-time PCR machine.
All the 13 individuals tested negative for chytrid fungus. This is a very low sampling size and
no significant consequences can be drawn from the results, but practice in Hungary shows that
Bombina variegata is a good indicator for presence of Bd (VÖRÖS ET AL., 2009). We plan
more studies focusing on high elevation areas and sampling Bombina variegata in order to
further assess the distribution of chytrid fungus within South-eastern Europe.
REFERENCES
BOYLE, D. G., BOYLE, D. B., OLSEN, V., MORGAN, J., HYATT A. (2004): Rapid quantitative
detection of chytridiomycosis (Batrachochytrium dendrobatidis) in amphibian samples using
real-time Taqman PCR assay. Disease of Aquatic Organisms 60:141-8.

HYLA VOL. 2011. No. 1 Str. 31-34 SHORT NOTE
33
GARNER T. W. J., PERKINS M. W., GOVINDARAJULU P., SEGLIE D., WALKER S., CUNNINGHAM
A. A., FISHER M. C. (2006): The emerging amphibian pathogen Batrachochytrium
dendrobatidis globally infects introduced populations of the North American bullfrog, Rana
catesbeiana. Biology Letters, doi:10.1098/rsbl.2006.0494
GARNER, T. W. J., WALKER, S., BOSCH, J., HYATT, A. D., CUNNINGHAM, A. A., FISHER, M. C.
(2005): Chytrid fungus in Europe. Emerging Infectious Diseases 11: 1639–1641.
LONGCORE, J., PESSIER, A., NICHOLS D. (1999): Batrachochytrium dendrobatidis gen. et sp.
nov., a chytrid pathogenic to amphibians. Mycologia 91: 219-27.
POUNDS, J. A., BUSTAMANTE, M. R., COLOMA, L. A., CONSUEGRA, J. A., FOGDEN, M. P. L.,
FOSTER, P. N., LA MARCA, E., MASTERS, K. L., MERINO-VITERI, A., PUSCHENDORF, R., RON,
S. R., SÁNCHEZ-AZOFEIFA, G. A., STILL, C. J., YOUNG, B. E. (2006): Widespread amphibian
extinctions from epidemic disease driven by global warming. Nature 439: 161-167.
SOTO-AZAT, C., CLARKE, B. T., POYNTON, J. C., CUNNINGHAM A. A. (2010): Widespread
historical presence of Batrachochytrium dendrobatidis in African pipid frogs, Diversity and
Distributions 16: 126–131.
SPATIALEPIDEMIOLOGY (2011): Spatialepidemiology.net <www.spatialepidemiology.net>.
Accessed 28.03.2011.
STUART, S. N., CHANSON, J. S., COX, N. A., YOUNG, B. E., RODRIGUES, A. S. L., FISCHMAN, D.
L., WALLER, R. W. (2004): Status and trends of amphibian declines and extinctions
worldwide. Science 306: 1783–1786.
SURA, P., JANULIS, E., PROFUS, P. (2010): Chytridiomycosis – a mortal danger for amphibians
- Chrońmy Przyrodę Ojczystą 66 (6): 406–42.
SZTATECSNY, M., GLASER, F. (2011): From the eastern lowlands to the western mountains:
first records of the chytrid fungus Batrachochytrium dendrobatidis in wild amphibian
populations from Austria. Herpetological Journal 21: 87-90.

VÖRÖS, J., HETTYEY, A., SÓS, E., DANKOVICS, R., GARNER T. (2009): Amphibian chytrid
fungus in Hungary. Second European Congress of Conservation Biology, abstract book: 105.

HYLA VOL. 2011. No. 1 Str. 35-46 SHORT NOTE
35
Finding of the Alpine salamander (Salamandra atra Laurenti, 1768; Salamandridae,
Caudata) in the Nature Park Žumberak - Samoborsko gorje (NW Croatia)
NINA JERAN*1, PETRA ĐURIĆ2, KREŠIMIR ŽGANEC
3
1 Požarinje 69, 10000 Zagreb, Croatia, [email protected]
2 State Institute for Nature Protection, Trg Mažuranića 5, 10000 Zagreb, Croatia
3 University of Zagreb, Faculty of Science, Rooseveltov trg 6, 10000 Zagreb, Croatia
ABSTRACT
This study confirms the presence of Alpine salamander (Salamandra atra) in the Nature Park
Žumberak - Samoborsko gorje, where previously only one specimen was recorded in 1989.
Species presence and distribution were investigated at ten different localities in stands of
montane beech forest, during the vegetation season 2004. In July 2004 five individuals (four
males and one female) of Alpine salamander were found in the virgin beech forest at site Kuta
(about 900 m a.s.l.), during weather conditions characterized by heavy rain. This is the
northernmost finding of the species in Croatia, as well as a confirmed disjunctive part of its
areal in the Dinarids. Conservation measures for the species are proposed but for more precise
conservation plan further research of species distribution and ecology is needed.
Keywords: Alpine salamander, Salamandra atra, Nature Park Žumberak - Samoborsko gorje,
distribution, amphibians

Alpine salamander (Salamandra atra) is a montane species occurring between 400 and 2,800
m a.s.l., but is usually found from 800 to 2,000 m a.s.l. (ARNOLD & BURTON, 1978). It
inhabits mainly forests (beech, mixed deciduous and mixed deciduous-coniferous), but may
also be found above the tree-line, in cool, damp alpine meadows, pastureland and other,
slightly modified habitats. The young are born completely metamorphosed, and therefore, its
life cycle is not associated with water. Alpine salamander is mainly nocturnal and during
daylight it hides in shady places, holes or beneath stones and logs, but can be encountered in
the open during the day, after rain or in overcast weather (ARNOLD & BURTON, 1978).
Distribution range of Alpine salamander (Figure 1a) is restricted into two areas: bigger area
on the north, encompassing Swiss, Austrian, Italian, German, French and Slovenian Alps, and
geographically isolated populations in the Dinaric Alps in Slovenia, Croatia, Bosnia and
Herzegovina, Serbia, Montenegro and northern Albania (ANDREONE ET AL., 2008). While in
most part of European Alps species still has abundant populations and is not considered
endangered, isolated populations of Dinaric Alps appear to be rarer and more fragmented
(KALEZIĆ & DŽUKIĆ, 2001, ANDREONE ET AL., 2008, LELO ET AL., 2008, ŠUNJE & LELO,
2010). Upon that, Dinaric populations are especially threatened by localised habitat alteration
through the cutting and removal of ground cover during forestry practices, intensification of
farming (in lower regions), tourism and infrastructure development. Negative effects of air
pollution, rain and soil acidification as well as climatic changes are also likely.
There are three currently recognized subspecies: S. atra atra, S. a. aurorae (restricted to an
extremely small area of northeastern Italy) and S. a. prenjensis. However, their status remains
questionable. While S. a. aurorae has sometimes been considered as a separate species, S. a.
prenjensis is often not considered to be valid subspecies (ANDREONE ET AL., 2008). Molecular
studies based on mtDNA cyt b gene (RIBERON ET AL., 2001) and AFLP markers (RIBERON ET
AL., 2004) failed to show clear genetic structure of the first two subspecies, however, the

HYLA VOL. 2011. No. 1 Str. 35-46 SHORT NOTE
37
analyses of mtDNA D-loop sequence (STEINFARTZ ET AL., 2000) support the assignment of S.
a. atra and S. a. aurorae into separate subspecies, while S. a. prenjensis is shown to be more
closely related to S. a. atra.
Salamandra atra is strictly protected under the Bern Convention on the Conservation of
European Wildlife and Natural Habitats (Appendix II) and is listed on Annex IV of the
Council Directive 92/43/EEC on the conservation of natural habitats and of wild fauna and
flora (Habitats Directive). Subspecies S. atra aurorae is also listed on Annex II of the
Habitats Directive. According to the IUCN Red List of Threatened Species S. atra is
considered as Least Concern in view of its wide distribution, tolerance of a degree of habitat
modification, presumed large population, and because its decline is not considered fast, while
subspecies S. a. aurorae is considered as Critically Endangered. Salamandra atra is protected
by national legislation in most range countries (e.g. Switzerland, Austria, Slovenia, Croatia).
Findings of the Alpine salamander in Croatia include Gorski kotar area (Mala Kapela, Velika
Kapela, Samarske stijene, Bijele stijene, Risnjak), Učka and Žumberak Mountain, (KLETEČKI,
1990, ĐURIĆ ET AL., 2004, GRBAC ET AL., 2006). On the Žumberak Mountain, until this study,
Alpine salamander was recorded only once, in 1989, when one dead individual was found on
the bottom of jama Tomaševićka pit (KLETEČKI, 1990), located nearby bigger Pogana jama
cave. It is worth mentioning that the whole area of Žumberak Mountain is rich in karst
phenomena (dolines, blind valleys, caves and pits) (BUZJAK, 2002).
Aim of this study was to confirm the presence of Alpine salamander on the Žumberak
Mountain inside the Nature Park Žumberak - Samoborsko gorje, and to determine other
potential habitats in the scope of the Nature Park. Overall goal was to contribute to the
conservation of the Alpine salamander at Nature Park Žumberak - Samoborsko gorje but also
at national level where little is known about this species and its distribution.
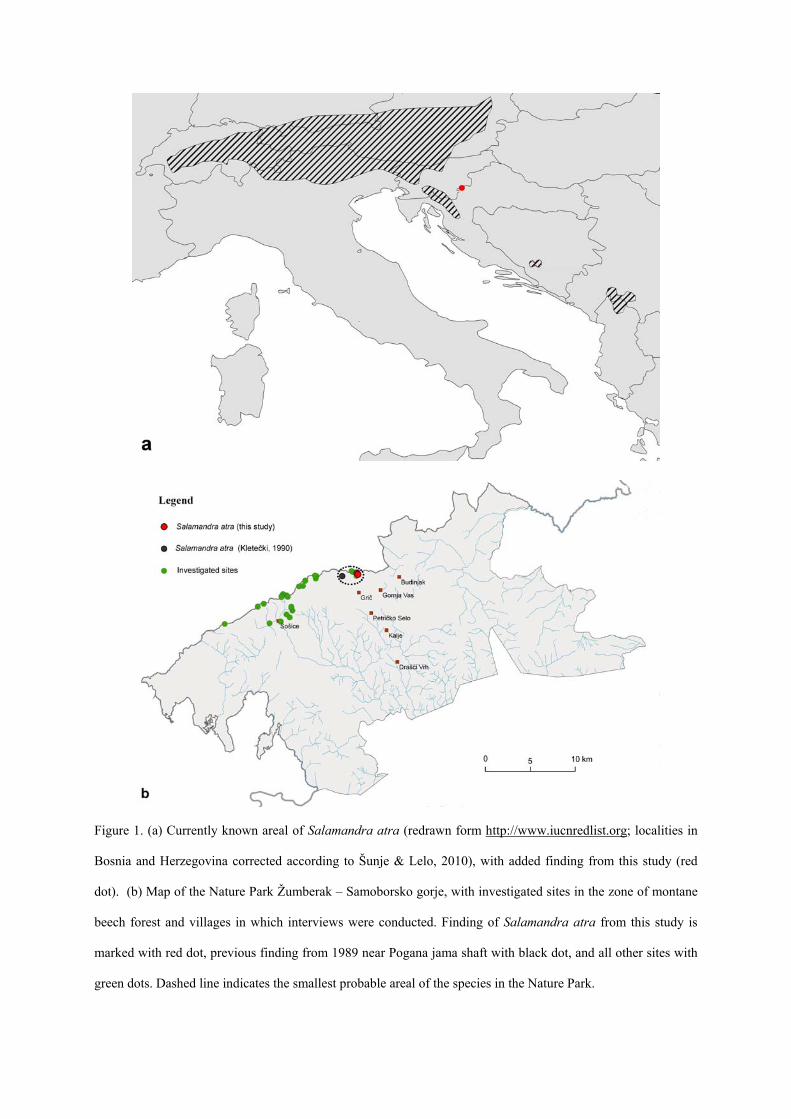
Figure 1. (a) Currently known areal of Salamandra atra (redrawn form http://www.iucnredlist.org; localities in
Bosnia and Herzegovina corrected according to Šunje & Lelo, 2010), with added finding from this study (red
dot). (b) Map of the Nature Park Žumberak – Samoborsko gorje, with investigated sites in the zone of montane
beech forest and villages in which interviews were conducted. Finding of Salamandra atra from this study is
marked with red dot, previous finding from 1989 near Pogana jama shaft with black dot, and all other sites with
green dots. Dashed line indicates the smallest probable areal of the species in the Nature Park.

HYLA VOL. 2011. No. 1 Str. 35-46 SHORT NOTE
39
Slika 1. (a) Trenutno poznati areal vrste Salamandra atra (nacrtano prema http://www.iucnredlist.org; lokaliteti
iz Bosne i Hercegovine izmijenjeni prema Šunje & Lelo, 2010), sa dodanim nalazom iz ovog istraživanja
(crvena točka). (b) Karta Parka prirode Žumberak – Samoborsko gorje, sa istraživanim lokalitetima planinske
bukove šume i lokacijama sela u kojima je provedena anketa. Nalaz vrste Salamandra atra iz ovog istraživanja
označen je crvenom točkom, prethodni nalaz iz 1989. g. kraj Pogane jame sa crnom točkom, a ostali istraživani
lokaliteti sa zelenim točkama. Iscrtana linija označava najmanji vjerojatni areal vrste u Parku prirode.
Field investigations of species presence and distribution were carried out during the
vegetation season 2004, in altogether nine days in May (3 days), July (5 days) and October (2
days). Field research was concentrated mostly on stands of montane beech forests scarce in
watercourses in upland parts of Žumberak Mountain, above 700 m a.s.l., which were regarded
as the most suitable habitats for Alpine salamander in this region. During the field research
we have searched ground vegetation layer in detail, and also turned logs, stones and layer of
deep leaves. Scanning of the terrain was conducted in conditions when, according to the
literature data, Alpine salamanders had the greatest activity (rainy and cloudy weather, mostly
early in the morning but also during other parts of the day, if weather conditions were
suitable). We used Garmin Summit GPS receiver for determination of coordinates and
ArcMap in ArcGIS 9.2 program package (1999-2006 ESRI Inc.) for production of maps.
In addition, we made personal interviews with local inhabitants (altogether 61 persons) from
Nature Park (villages Grič, Kalje, Petričko selo, Gornja Vas, Budinjak, Drašči Vrh and
Sošice) about their possible observations of Alpine salamander. Interviewees were shown the
picture of the Alpine salamander and they were warned about differences between this and
other possibly similar species (e.g. Fire salamander, newts).
We scanned ten different localities of beech forest stands in different stage of degradation and
in each we recorded coordinates for one or few points (Figure 1b). On 27th July 2004, between
about 1 and 2 p.m. in the old virgin montane beech forest Kuta, at about 900 m a.s.l., we

found five individuals of Alpine salamander, out of which four males and one female.
Weather conditions were characterised by heavy rain and high air humidity. First individual
was noticed two hours after the beginning of heavy rain, when deep layer of leaves was
impregnated with water (Figure 2). It was found at the bottom of shallow karst sinkhole;
female and another three males were found in a range of few hundred meters. After the rain
stopped at 3 p.m., we have continued the search for another hour but were unable to find any
new individual of Alpine salamander. One additional interesting finding was noted. Triggered
by putting them together into a plastic box, one male caught female in amplexus (Figure 2).
Thereafter, we have returned the pair on the forest soil and continued to observe and
document this reproductive behaviour for another 1.5 hour. This behaviour, together with
measured prominence of cloacae in male implies that in July this population is in reproductive
period.
Forest stand Kuta corresponds to an adequate habitat for Alpine salamander according to its
altitude (between 800 and 900 m a.s.l.) and its distance from the watercourses. The site is
positioned in the zone of montane beech forest (as. Lamio orvalae – Fagetum sylvaticae Ht.
1938/ Borhidi 1963). It is situated on a heavily-accessible terrain, with numerous rocks and
karst sinkholes of different depths and elevations (Figure 3). Old beech trees are developed,
since cutting is restricted. Rocks and trees are richly overgrown with mosses. Ground
vegetation layer is sparsely developed, somewhere with a dense bushes of ferns and young
trees of beech and sycamore maple. Many huge logs on the ground in different stage of
decomposition, as well as rocky terrain with numerous holes, provides abundant shelters for
salamanders. Forest at site Kuta is only about 1.5 km distant from the jama Tomaševićka pit,
the locality of the only previous finding of Alpine salamander at Nature Park Žumberak -
Samoborsko Gorje, also situated in a zone of old beech forest at around 900 m a.s.l
(KLETEČKI, 1990). Jama Tomaševićka is a pit located on the mountain slope nearby well-

HYLA VOL. 2011. No. 1 Str. 35-46 SHORT NOTE
41
known locality Pogana jama, which is actually a stratified cave that serves as a permanent
swallow hole for surface waters (BUZJAK, 2002). Both our and KLETEČKI (1990) findings
enabled us to identify the smallest probable area of distribution for Alpine salamander in the
Nature Park (Figure 1b). The area above 800 m a.s.l. where montane beech forests from
association Lamio orvalae – Fagetum sylvaticae are prevalent, represent potential habitat for
this species.
Figure 2. Alpine salamander (Salamandra atra) found in the virgin beech forest Kuta on 27th June 2004 (male -
left, amplexus - right).
Slika 2. Crni daždevnjak (Salamandra atra) nađen u bukovoj prašumi Kuta 27. srpnja 2004. godine. (mužjak -
lijevo, ampleksus - desno).
Figure 3. Stand of montane beech forest in Kuta, habitat of S. atra.
Slika 3. Sastojina planinske bukove prašume u području Kute (Žumberak), stanište vrste S. atra.

Regarding results from the interviews made among the local inhabitants, from altogether 61
examinees, 11 confirmed the observation of Alpine salamander in different sites, some of
which coincide with area close to our finding. However, possible confusion with similar
species should be taken into account.
Rare findings of Alpine salamander in Nature Park Žumberak - Samoborsko Gorje suggest
very small and reduced population. During our study we were searching the area in
appropriate weather conditions, for altogether 97 hours (per person) to find 5 individuals of
Alpine salamander (or 19.5 hours per person for finding one individual). Moreover, additional
fieldworks were undertaken (42 hours/person in 2009, and for 42 and 9 hours/person,
respectively, in 2010) in the key site Kuta, during appropriate weather conditions (JELIĆ, D.,
Zagreb, pers. comm. 2011), but without any finding. Furthermore, educational leaflet was
produced in hope that Nature Park visitors would report observations of the Alpine
salamander, but also without any success. For comparison, at Bijele stijene site in Mala
Kapela region (Gorski kotar), 18 hours of research yielded in finding 36 individuals of Alpine
salamander (or 30 mins per individual) implying to abundant and preserved population (JELIĆ,
D., pers. comm. 2011).
The conservation of Alpine salamander in the Nature Park Žumberak - Samoborsko Gorje
should be maintained through the preservation of its known and potential habitats, mainly
well developed beech forests. Since there are indications that this isolated population at
Žumberak Mountain is very small it is necessary to implement stricter protection measures at
its known distribution sites (site Kuta, jama Tomaševićka, with wider buffer area) where
forest cutting should be avoided as well as other activities that could have negative impact on
preservation of integrity of these beech forest habitats. If possible, we would suggest
protecting site Kuta as a special forest reserve. Also, further spreading of coniferous
plantations in Nature Park should be avoided at all known and potential habitats of the Alpine

HYLA VOL. 2011. No. 1 Str. 35-46 SHORT NOTE
43
salamander. Having in mind interesting disjunctive distribution area of Alpine salamander,
further research of its distribution and ecology is needed in order to establish its exact
distribution area and status of its populations in Croatia, as well as to propose and implement
appropriate management and protection measures for the species and its habitats.
REFERENCES
ANDREONE, F., DENOËL, M., MIAUD, C., SCHMIDT, B., EDGAR, P., VOGRIN, M., CRNOBRNJA
ISAILOVIC, J., AJTIC, R., CORTI, C. & HAXHIU, I. (2008): Salamandra atra. In: IUCN 2010.
IUCN Red List of Threatened Species. Version 2010.4., <www.iucnredlist.org>. Accessed
20th March 2010.
ARNOLD, E. N. & BURTON, J. A. (1978): Family Salamandridae – Typical Salamanders and
newts. pp. 32-34. In: ARNOLD, E. N. & BURTON, J. A. OVENDEN D. W. (eds.) A field guide of
the reptiles and amphibians of Britain and Europe. HarperCollinsPublishers, London.
BUZJAK, N. (2002): Speleological features of "Žumberak – Samoborsko gorje" Nature Park
(in Croatian). Geoadria 7(1): 31-49.
ĐURIĆ, P., JERAN, N. & ŽGANEC, K. (2004): Alpine salamander (Salamandra atra) in
Žumberak (in Croatian; expert report). Nature Park Žumberak - Samoborsko gorje.
GRBAC, I., TVRTKOVIĆ, N., KLETEČKI, E. & GRGUREV, M. (2006): New findings of the Alpine
salamander, Salamandra atra Laurenti, 1768 (Salamandridae, Amphibia) in Croatia. (in
Croatian). In: ARKO-PIJEVAC, M., KRUŽIĆ, B., KOVAČIĆ, M. (eds.) Prirodoslovna istraživanja
Riječkog područja. Prirodoslovni muzej Rijeka, Rijeka.
KALEZIĆ, M. & DŽUKIĆ, G. (2001): Amphibian status in Serbia and Montenegro (FR
Yugoslavia). FrogLog (Newsletter of the declining amphibian populations task force) 45: 2-3.
KLETEČKI, E. (1990): New finding of the alpine salamander (Salamandra atra Laurenti, 1768;
Salamandridae: Caudata) in Croatia. Arhiv bioloških nauka 42 (1-2): 5P.

LELO, S., KOTROŠAN, D., MUFTIĆ, K., TRAKIĆ, S., DURAKOVIĆ, E. & ŠUNJE, E. (2008): General
data regarding habitat and population of Salamandra atra Laurenti, 1768 (S. a. prenjensis
Mikšić, 1969) in Mt. Prenj (in Bosnian). Prilozi fauni Bosne i Hercegovine 4: 11-15.
RIBERON, A., MIAUD, C., GROSSENBACHER, K. & TABERLET, P. (2001): Phylogeography of the
Alpine salamander, Salamandra atra (Salamandridae), and the influence of the Pleistocene
climatic oscillations on population divergence. Molecular Ecology 10: 2555–2560.
RIBERON, A., MIAUD, C., GUYETANT, R. & TABERLET, P. (2004): Genetic variation in an
endemic salamander, Salamandra atra, using amplified fragment length polymorphism.
Molecular Phylogenetics and Evolution 31: 910–914.
STEINFARTZ, S., VEITH, M. & TAUTZ, D. (2000): Mitochondrial sequence analysis of
Salamandra taxa suggests old splits of major lineages and postglacial recolonisations of
Central Europe from distinct source populations of Salamandra salamandra. Molecular
Ecology 9: 397-410.
ŠUNJE, E. & LELO, S. (2010): Contribution to Alpine Salamander distribution, Salamandra
atra prenjensis Mikšić, 1969 (Amphibia, Salamandridae), in Bosnia and Herzegovina (in
Bosnian). Prilozi fauni Bosne i Hercegovine 6: 34-41.
ACKNOWLEDGEMENTS
This study was done by Croatian herpetological society – “Hyla” as a part of the project
“Alpine salamander (Salamandra atra) in Žumberak” that was supported by Public institution
“Žumberak - Samoborsko gorje Nature Park”. We are especially grateful to former
conservation manager of the Park - Biljana Janev Hutinec for support and advices, as well as
to Park ranger Slavko Struna for a huge help and engagement in a quest for Alpine
salamander. We also wish to thank to Eduard Kletečki for valuable advices. We are as well
grateful to Nina Popović on a help during the field work.

HYLA VOL. 2011. No. 1 Str. 35-46 SHORT NOTE
45
SAŽETAK
NALAZ CRNOG DAŽDEVNJAKA (SALAMANDRA ATRA LAURENTI, 1768; SALAMANDRIDAE,
CAUDATA) U PARKU PRIRODE ŽUMBERAK - SAMOBORSKO GORJE (SZ HRVATSKA)
NINA JERAN, PETRA ĐURIĆ, KREŠIMIR ŽGANEC
Ovim istraživanjem potvrđena je prisutnost crnog daždevnjaka (Salamandra atra) u Parku
prirode Žumberak - Samoborsko gorje, gdje je prethodno zabilježena samo jedna jedinka
1989. godine. Prisutnost i rasprostranjenost vrste istraživani su tijekom vegetacijske sezone
2004. godine na deset različitih lokaliteta u sastojinama planinske bukove šume. U srpnju
2004. g. pronađeno je pet jedinki (četiri mužjaka i jedna ženka) crnog daždevnjaka u bukovoj
prašumskoj sastojini Kuta (na oko 900 m n. v.), za vrijeme vremenskih prilika praćenih jakom
kišom. Ovo je najsjeverniji nalaz vrste u Hrvatskoj, koji ukazuje da Žumberak predstavlja
disjunktni dio njezinog areala u Dinaridima. U radu su predložene mjere zaštite vrste,
međutim, za donošenje preciznijeg plana zaštite potrebna su dodatna istraživanja
rasprostranjenosti i ekologije ove vrste.
Ključne riječi: crni daždevnjak, Salamandra atra, Park prirode Žumberak - Samoborsko
gorje, rasprostranjenost, vodozemci


HYLA VOL. 2011. No. 1 Str. 47-75 ORGINALNI ZNANSTVENI ČLANAK
47
Gmazovi (Reptilia) Parka prirode Papuk
Reptiles (Reptilia) of the Papuk Nature Park
FRANO BARIŠIĆ *, TOMISLAV BOGDANOVIĆ
Sveučilište J. J. Strossmayera, Odjel za biologiju, Trg Ljudevita Gaja 5, 31000 Osijek
SAŽETAK
Cilj istraživanja bio je načiniti sustavna faunistička i ekološka istraživanja te inventarizaciju i
valorizaciju faune gmazova na području Parka prirode Papuk. Rezultati dobiveni ovim radom
mogli bi doprinijeti daljnjem istraživanju cjelokupne faune gmazova Hrvatske. Sustavna
istraživanja faune i biologije pojedinih vrsta gmazova (Reptilia) na području Parka prirode
Papuk provedena su od veljače do kolovoza 2009. godine na petnaest istraživačkih postaja
koje su ciljano obuhvatile različite tipove staništa na kojima gmazovi obitavaju. Metode
prikupljanja gmazova podijeljene su dvojako prema vrstama gmazova koja obitavaju uz
vodena staništa i prema terestričnim vrstama. Korištene su metoda tzv. sit and wait, vizualna
metoda uzorkovanja, aktivno traženje i vizualna metoda uzorkovanja u pojasevima ili
kvadratnim plohama. Prikupljeni podaci su uneseni u bazu podataka i uz pomoć GIS
tehnologije načinjene su karte rasprostranjenosti pojedinih vrsta na istraživanom području.
Istražena je sezonska i dnevna dinamika sa utvrđenim eudominatnim i recentnim vrstama.
Ključne riječi: Inventarizacija, valorizacija, eudominatne vrste, recentne vrste, frekventnost

UVOD
Promatrajući istraživano područje, Park prirode Papuk možemo uvidjeti da postoji relativno
velika raznolikost ekosustava na relativno malom prostoru. Papučko-krndijsko gorje proteže
se smjerom istok–zapad, rezultati upućuju da je broj vrsta različit od sjevernijih postaja koje
su pod drugačijim klimatskim utjecajma od istraživanih postaja koje se nalaze na južnim
ekspozicijama Papuka. Na nešto višim nadmorskim visinama se ekološki uvjeti za život
gmazova lagano mijenjaju, ovdje treba imati u vidu činjenicu da je istraživano područje
gorski kraj i da spomenute razlike iznose nekoliko stotina metara nadmorske visine.
Najstarije nalaze herpetofaune sa ovog područja donosi KARAMAN (1921) koji je tijekom
svojih proputovanja posjetio područje Jankovca, Velike i Kutjeva te zabilježio 2 vrste
vodozemaca (Bombina variegata, Salamandra salamandra) i 4 vrste gmazova (Podarcis
muralis, Lacerta viridis, Anguis fragilis, Zamenis longissimus). Kasnije RADOVANOVIĆ (1951)
dodaje i Zootocu viviparu, a PAVLETIĆ (1964) pregledavajući zbirku Hrvatskog
prirodoslovnog muzeja samo potvrđuje isti sastav vrsta. Kasnijim usputnim istraživanjima
(KRIZMANIĆ I SUR., 1997, SOTIROPOULOS I SUR., 2001) zabilježene su na području Jankovca
još i dvije vrste vodenjaka, Lissotriton vulgaris i Ichthyosaura alpestris. Promatrajući povijest
pojedinih istraživanja iz ovog područja, vidimo da se od prvih do najnovijih nalaza fauna
gmazova relativno sporo obogaćuje novim taksonima, uspoređujući literaturne podatke o
zoogeografskoj rasprostranjenosti gmazova ovog područja (MIKUŠKA, 1979). Herpetološkim
istraživanjima tijekom 2008. godine po prvi put je za Papuk zabilježena vrsta ivanjski rovaš,
Ablepharus kitaibelii (SZÖVÉNYI I JELIĆ, 2011).
Herpetofauna Papuka je nedovoljno istražena, zastarjeli rezultati i moderne metode kartografije
uz primjerenu podršku informatike povod su dodatnom istraživanju herpetofaune papuka.
Rezultati ovih istraživanja doprinos su poznavanju faune gmazova sjeveroistočnog dijela

HYLA VOL. 2011. No. 1 Str. 47-75 ORGINALNI ZNANSTVENI ČLANAK
49
Hrvatske, te obzirom na sve veću ugroženost staništa na kojima obitavaju dovodi se u pitanje i
opstanak pojedinih vrsta.
MATERIJALI I METODE
Papuk je jedna od ključnih zemljopisnih odrednica Slavonije. Papuk, zajedno s Krndijom,
pripada slavonskome gorju, koje ima središnji zemljopisni položaj u panonskom, nizinskom
prostoru Slavonije, u istočnom dijelu Hrvatske (Slika 1.). Ove se dvije planine nalaze u
međuriječju Save i Drave, i oštro se izdižu iz područja Posavske i Podravske zaravni
(SAMARDŽIĆ, 2005).
N
1 : 17 000 000
Slika 1. Smještaj Parka prirode Papuk u Hrvatskoj (izvornik JU PP „Papuk“).
Picture 1. Locationof Papuk Nature Park in Croatia (original obtained from Papuk Nature Park Public
institution).

Park prirode Papuk površine je oko 335 km2. Granice Parka prirode definirane su Zakonom o
proglašenju PP Papuk (NN 61/99). Područje obuhvaća dio papučko-krndijskoga gorja, na
potezu Kutjevo – Velika –Kamenski Vučjak – Zvečevo – Voćin– Slatinski Drenovac –
Orahovica – Petrov vrh – Kutjevo (Slika 2). Većinom cjelovit šumski ekosustav je kako
reljefno tako i visinski odijeljen od slavonske ravnice, tako da je najniža točka ovog područja
na sjeveru 162 m.n.v., a najviša u središnjem dijelu – Vrh Papuka, 953 m.n.v. (SAMARDŽIĆ,
2000).
N
1 : 1 000 000
Slika 2. Granice PP Papuk prema Zakonu o proglašenju PP Papuk (NN 61/99) (izvornik JU PP Papuk).
Picture 2.Borders of Papuk Nature Park, according to the Papuk Public Institution Founding Act (NN 61/99)
(original obtained from Papuk Nature Park Public institution).
RELJEF I HIDROLOGIJA
Papuk se nalazi u kontinentalnom dijelu Republike Hrvatske, na prostoru dodira središnje i
istočne Hrvatske. Prema prirodno-zemljopisnoj regionalizaciji, prostor pripada panonskoj
megaregiji, odnosno području Slavonskoga gorja. Izvorne šume su dobro očuvane, a posebno

HYLA VOL. 2011. No. 1 Str. 47-75 ORGINALNI ZNANSTVENI ČLANAK
51
su vrijedne šume panonske bukve i jele u višim predjelima.U sklopu slavonskoga gorja Papuk
je najprostranija gora i zajedno s Krndijom oblikuje jednu reljefno-pejzažnu cjelinu.
Od glavnoga grebena razvedene su gorske kose i slivovi, u smjeru sjever–jug. Masiv
papučko-krndijskoga gorja u Parku prirode Papuk hidrološki je vrlo bogat nadzemnom i
podzemnom vodom. U navedenom području teren je razveden i obiluje brojnim stranama,
vrhovima, grebenima, jarcima i uvalama različitih nagiba. Površina Parka prirode Papuk od
33.600 ha obuhvaća 29.212,82 ha šumskih površina (87%).
POPIS ISTRAŽIVANIH NALAZIŠTA – LOKALITETA
Istraživanja su obavljena na 15 postaja (Tablica 1.)
Tablica 1. Popis nalazišta sa pripadajućim kraticama i zemljopisnim (GPS) koordinatama.
Table 1. List of sites with abbreviations and geographical (GPS) coordinates.
Nalazište OZNAKA X Y
Orahovačko jezero OJ 45°31′02.48″ 17°51′42.42″
Jankovac JA 45°31′24.45″ 17°41′24.91″
Zvečevo ZV 45°33′01.59″ 17°30′01.59″
Bistra BI 45°26′37.59″ 17°43′29.84″
Vetovo - ribnjaci VR 45°26′15.27″ 17°47′29.93″
Kutjevo - potok KP 45°26′31.01″ 17°53′03.96″
Dubočanka - potok DU 45°28′57.67″ 17°39′40.81″
Brzaja BR 45°27′41.01″ 17°29′57.58″
Duboka – potok DP 45°28′55.27″ 17°31′30.73″
Djedovica DJ 45°35′12.79″ 17°29′09.75″
Svinjarevac SV 45°30′39.79″ 17°30′10.16″
Fosilno nalazište FN 45°26′15.90″ 17°34′42.60″
Voćin – jezero VJ 45°36′53.53″ 17°33′11.67″
Sekulinci potok SP 45°31′09.01″ 17°29′13.77″
Slatinski drenovac SD 45°33′03.04″ 17°42′52.21″

Istraživano područje obuhvaćeno ovim istraživanjem, podjeljeno je, uvjetno rečeno, na tri
usporedna transekta, koji su obuhvaćeni slijedećim zemljopisnim pravcima:
pravac: Orahovica – Kutjevo,
pravac: Jankovac – Dubočanka – Velika,
pravac: Voćin – Zvečevo – Velika.
Promatranjem vegetacijskog pokrova, kao i hidroloških uvijeta, ustanovljeno je da istraživano
područje ima dosta različitih obilježja, pa su time i ekološki uvjeti života za vrste koje
obitavaju na tom prostoru raznoliki. Imajući to u vidu, istraživačke postaje odabrane su tako
da obuhvate različite tipove staništa na kojima gmazovi obitavaju, te je na taj način
zadovoljena prostorno-staništna pokrivenost (Tablica 1).
Mjesta uzorkovanja gmazova izabrana su tako da budu u što manjoj mjeri izložena
ometanjima te negativnim antropogenim utjecajima. Najvažnija staništa na kojima su
obavljana istraživanja gmazova na području Parka prirode Papuk su:
- mezofilne i kseromezofilne livade,
- prirodne, mješovite listopadne šume,
- potoci, brdski dijelovi riječnih tokova, privremene vodene površine,ribnjaci,
- iskopine kamena (kamenolomi), šljunka ili pijeska,
- kanali, sporiji dijelovi tekućica,
- antropogena staništa poput izletišta, voćnjaka, vinograda i sl.
Za uzorkovanje gmazova izabrana su takva mjesta koja posjeduju očuvane prirodne
vrijednosti, te postoji mogućnost dobivanja nekog od statusa zaštićenog područja. Važan
kriterij pri odabiru lokaliteta kojeg se mora posjećivati u više navrata jeste i njegova
pristupačnost.
U staništa značajnija za neke vrste gmazova u gorskom dijelu Hrvatske razvrstavaju se: rijeke,
potoci i poplavna polja, te lokve i jezera (prirodna i antropogena). Cretišta, bilo da su na nižoj

HYLA VOL. 2011. No. 1 Str. 47-75 ORGINALNI ZNANSTVENI ČLANAK
53
ili višoj nadmorskoj visini, s čistim kiselim vodama i vrlo bujnom vegetacijom, pogodna su
mjesta za razvoj većine vrsta. Cretovi su ugroženi, a zbog malog broja i malih površina
spadaju u kritično ugrožena staništa u Hrvatskoj (DZZP, 2010).
Osim podjele staništa koja se primijenjuje u istraživanjima većine europskih vrsta gmazova,
moguće je za konkretno istraživano područje načiniti i podjelu prema tipovima mikrostaništa.
Takvim pristupom može se detaljnije istražiti npr. vodeno stanište. Prvenstveno se utvrđuju
fizičko-kemijska obilježja vode, temperatura, kiselost/lužnatost (pH), stanje razvoja
makrofitske vegetacije, jedinstvo biljnog i životinjskog svijeta, ali i čitav niz drugih ekoloških
čimbenika, koji su bitni za razvoj pojedinih vrsta gmazova. Podrazumijeva se da takav tip
istraživanja zahtijeva značajna sredstva, kao i vremenski daleko opsežnija istraživanja od
inventarizacijskih.
TERENSKI RAD
Za potrebe ovoga rada obavljana su faunistička i dijelom ekološka istraživanja gmazova na
području Parka prirode Papuk, na petnaest lokaliteta. Istraživanja su vršena u razdoblju od
veljače do rujna 2009. godine.
Za prikupljanje podataka o vrstama i brojnosti gmazova na odabranim lokalitetima korištena
je metoda linijskog transekta ili presjeka. To je metoda u kojoj se područje, koje se želi
istražiti, podjeli na više manjih površina podjednake veličine (dužine oko 100 metara) na
kojima se uzorkovanje obavlja u određenom vremenskom razdoblju (nekoliko mjeseci,
godina).Tokom periodičnih posjeta lokalitetu – u približno istom trajanju (minuta, sati) –
prikupljaju se podaci koji podrazumijevaju bilježenje, promatranje, fotografiranje i dr., Podaci
koji su dobiveni na terenu bilježeni su u terenski dnevnik, koji je kasnije poslužio za izradu
tablica i grafičkih prikaza, te pri daljnjoj obradi podataka. Na terenu je provedeno ukupno 15

dana i posebna je pozornost posvećena da se uzorkovanja provode dosljedno, te da je na
svakoj postaji provedeno isto vremensko razdoblje.
Metode prikupljanja gmazova (NAGY I SUR., 2006) podijeljene su dvojako:
- VRSTE KOJE SU NAČINOM ŽIVOTA VEZANE ZA VODU
Broj pronađenih životinja trebalo bi izraziti po jedinici površine. Primjera radi, vrste iz
porodice Colubridae mogu se otkriti noćnim traganjem uz pomoć lampe, te je ova metoda bila
pogodna za njihovo praćenje. Za praćenje bjelouške i ribarice korištena je metoda „sit and
wait“, tj. „sjedi i čekaj“. Autori preporučuju čekanje od 30 minuta u tišini. Ova pasivna
metoda primjenjiva je na preglednim staništima. Uporaba dalekozora je također preporučljiva
i korisna.
- TERESTRIČNE VRSTE
Terestrični gmazovi pronalaženi su aktivnim traženjem, vizualnom metodom uzorkovanja u
pojasevima ili kvadratnim plohama. Uzorkovanje u pojasevima širine od 2–5 m i dužine 500–
1.000 m obavljano je u slučaju ako su to odlike terena dopuštale. Teren je tada bilo potrebno
podijeliti na uzdužne paralelne pojaseve,čija je širina određivana tako da ukupna površina
jednog pojasa bude 250 m2 Ukoliko to nije moguće zbog odlika terena, prikazana je stvarna
dužina površine uzorkovanja. Pri označavanju kvadratne plohe za uzorkovanje 50 × 50 m (ili
100 ×100 m) uzeto je u obzir da granice plohe budu u okviru istog tipa biljnog pokrova.
Uzorkovanje je obavljano u okviru plohe u pet paralelnih pojaseva širokih oko 2 m,
međusobno udaljenih oko 10 m (20 m). U slučaju uzorkovanja na manjim površinama dani su
podaci o dužini i širini pojasa po kojem je vršeno uzorkovanje. Ove metode su se pokazale
pogodnima pri utvrđivanju veličina populacija guštera. Na prostranim travnjacima lako je
označiti kvadratne plohe ili pojaseve, te su na taj način obavljana uzorkovanja, npr. zidnog
guštera ili zelembaća.

HYLA VOL. 2011. No. 1 Str. 47-75 ORGINALNI ZNANSTVENI ČLANAK
55
Prebrojavanje životinja duž linije transekta je metoda pogodna kod uzorkovanja npr. u uskom
pojasu pored puteva ili na mjestu posječene šume poput šumskih prosjeka. Gmazovi često
stradaju pri prelasku puteva i cesta, pa su i ovakvi nalazi redovito bilježeni. Tijekom
istraživanja prikupljeni su, između ostaloga, svlakovi gmazova (prvenstveno zmija), koji
također čine značajne podatke i trebaju biti pohranjeni u zbirci. U slučaju kada se razdoblja
razmnožavanja različitih vrsta poklapaju, kao i kod prisutnosti sličnih staništa na različitim
mjestima, smanjuje se broj neophodnih uzorkovanja. Nasuprot tome, broj neophodnih
uzorkovanja povećava se ako se istraživano područje prostire na velikoj dužini (NAGY I SUR.,
2006). Obilježavanje pojedinih istraživanih postaja bilo je izvršeno pomoću GPS sustava
(satelitski kompas Magellan), pri čemu su postaje paralelno zabilježene i na kartama.
Dobiveni podaci su zatim kompjuterski dorađivani satelitskim snimkama.
Prilikom kartiranja gmazova na području Parka prirode Papuk korišteni su najvećim dijelom
rezultati vlastitih istraživanja, osim u nekoliko slučajeva gdje su prikazani ranije prikupljeni
podaci (BOGDANOVIĆ, usmeno priopćenje). Podaci kartiranja prikazani su slikom za svaku
vrstu pojedinačno u obliku karte, te grafikonima. Tako, npr. za vrstu Natrix natrix na karti je
prikazana rasprostranjenost vrste (Slika 4.), a grafikonima su prikazani ukupan broj
zabilježenih jedinki po lokalitetu (Slika 5.) i razdoblje pojavljivanja, odnosno sezonska
dinamika (Slika 6.). Sve istraživane postaje obilježene su na kartama zelenim kružićima, a
postaje za koje postoji podatak o nalazu određene vrste obilježene su crvenim kružićima.
OBRADA I ANALIZA PODATAKA
Obrada materijala i analiza podataka dobivenih terenskim radom obavljeni su u kabinetu na
Sveučilištu J. J. Strossmayer u Osijeku. Za obradu i analizu podataka korišteno je osobno
računalo, te programski paket Microsoft Office (Excel, Access, Word) pomoću kojeg su
podaci iz terenskoga dnevnika prebačeni u tablice, grafikone i korišteni za izradu ovog rada.

Na osnovu podataka sa terena određena je frekventnost vrsta na lokalitetima –koliko je puta
svaka vrsta primijećena u određenom vremenskom intervalu. Osim toga, određen je stepen
dominantnosti za svaku vrstu na svakom lokalitetu. Dominantnost je ona vrijednost koja je u
nizu mjerenja najviša zabilježena vrijednost, dakle koja „dominira“ (PETZ, 1985).

HYLA VOL. 2011. No. 1 Str. 47-75 ORGINALNI ZNANSTVENI ČLANAK
57
REZULTATI
Kvalitativni sustav ukazuje na bogatstvo faune gmazova na području Parka prirode Papuk,
koja iznosi jednu trećinu ukupne faune gmazova Hrvatske. Utvrđeno je ukupno 5 porodica
(Colubridae, Viperidae, Anguidae, Lacertidae, Scincidae), 7 rodova (Natrix, Zamenis,
Coronella, Vipera, Anguis, Lacerta, Ablepharus) i 11 vrsta gmazova (Tablica 2.).
Tablica 2. Popis utvrđenih porodica, rodova i vrsta gmazova na istraživanom području
Table 2. List of families, genera and species of reptiles recorded in the study area
Porodica Rod Vrsta
1. Colubridae Natrix Natrix natrix LINNAEUS, 1758
2. Colubridae Natrix Natrix tessellata LAURENTI, 1768
3. Colubridae Zamenis Zamenis longissimus LAURENTI, 1768
4. Colubridae Coronella Coronella austriaca LAURENTI, 1768
5. Viperidae Vipera Vipera berus LINNAEUS, 1758
6. Anguidae Anguis Anguis fragilis LINNAEUS, 1758
7. Lacertidae Lacerta Lacerta agilis LINNAEUS, 1758
8. Lacertidae Lacerta Lacerta viridis LAURENTI, 1768
9. Lacertidae Lacerta Lacerta vivipara JACQUIN, 1787
10. Lacertidae Lacerta Podarcis muralis LAURENTI, 1768
11. Scincidae Ablepharus Ablepharus kitaibelii BIBRON i BORY, 1833
UKUPNO 5 7 11
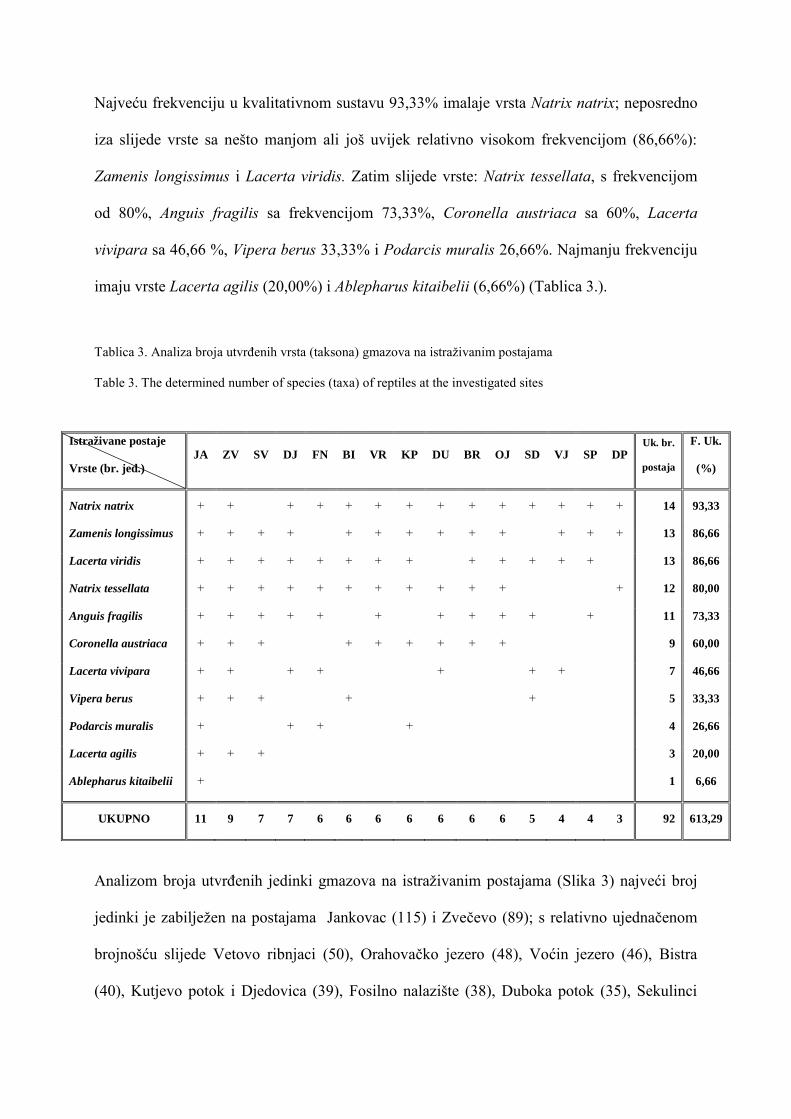
Najveću frekvenciju u kvalitativnom sustavu 93,33% imalaje vrsta Natrix natrix; neposredno
iza slijede vrste sa nešto manjom ali još uvijek relativno visokom frekvencijom (86,66%):
Zamenis longissimus i Lacerta viridis. Zatim slijede vrste: Natrix tessellata, s frekvencijom
od 80%, Anguis fragilis sa frekvencijom 73,33%, Coronella austriaca sa 60%, Lacerta
vivipara sa 46,66 %, Vipera berus 33,33% i Podarcis muralis 26,66%. Najmanju frekvenciju
imaju vrste Lacerta agilis (20,00%) i Ablepharus kitaibelii (6,66%) (Tablica 3.).
Tablica 3. Analiza broja utvrđenih vrsta (taksona) gmazova na istraživanim postajama
Table 3. The determined number of species (taxa) of reptiles at the investigated sites
Analizom broja utvrđenih jedinki gmazova na istraživanim postajama (Slika 3) najveći broj
jedinki je zabilježen na postajama Jankovac (115) i Zvečevo (89); s relativno ujednačenom
brojnošću slijede Vetovo ribnjaci (50), Orahovačko jezero (48), Voćin jezero (46), Bistra
(40), Kutjevo potok i Djedovica (39), Fosilno nalazište (38), Duboka potok (35), Sekulinci
Istraživane postaje
Vr ste (br. jed.) JA ZV SV DJ FN BI VR KP DU BR OJ SD VJ SP DP
Uk. br.
postaja
F. Uk.
(%)
Natrix natrix + + + + + + + + + + + + + + 14 93,33
Zamenis longissimus + + + + + + + + + + + + + 13 86,66
Lacerta viridis + + + + + + + + + + + + + 13 86,66
Natrix tessellata + + + + + + + + + + + + 12 80,00
Anguis fragilis + + + + + + + + + + + 11 73,33
Coronella austriaca + + + + + + + + + 9 60,00
Lacerta vivipara + + + + + + + 7 46,66
Vipera berus + + + + + 5 33,33
Podarcis muralis + + + + 4 26,66
Lacerta agilis + + + 3 20,00
Ablepharus kitaibelii + 1 6,66
UKUPNO 11 9 7 7 6 6 6 6 6 6 6 5 4 4 3 92 613,29
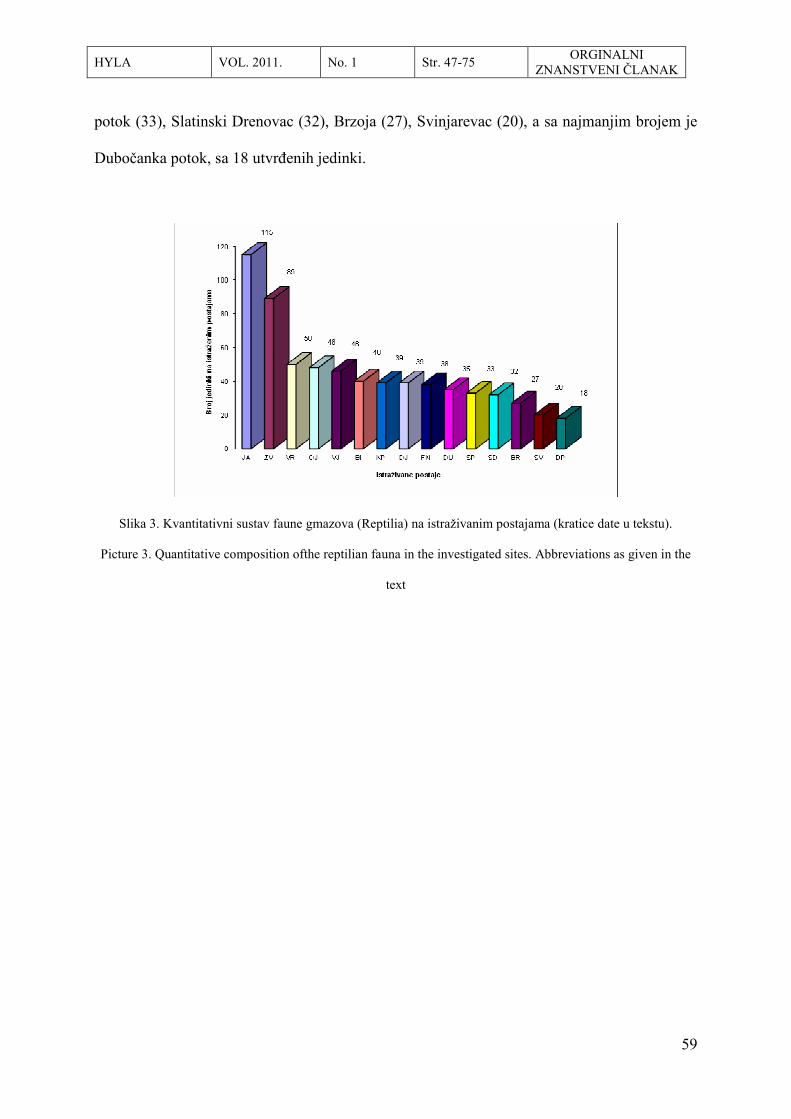
HYLA VOL. 2011. No. 1 Str. 47-75 ORGINALNI ZNANSTVENI ČLANAK
59
potok (33), Slatinski Drenovac (32), Brzoja (27), Svinjarevac (20), a sa najmanjim brojem je
Dubočanka potok, sa 18 utvrđenih jedinki.
Slika 3. Kvantitativni sustav faune gmazova (Reptilia) na istraživanim postajama (kratice date u tekstu).
Picture 3. Quantitative composition ofthe reptilian fauna in the investigated sites. Abbreviations as given in the
text
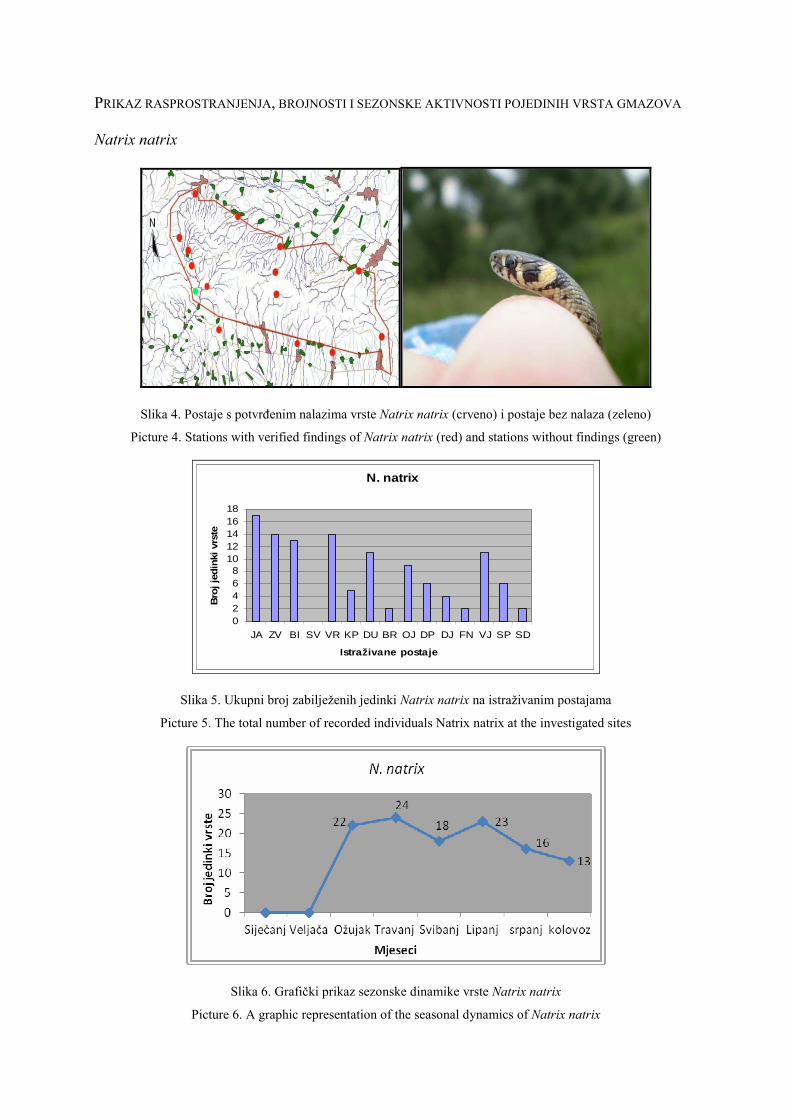
PRIKAZ RASPROSTRANJENJA, BROJNOSTI I SEZONSKE AKTIVNOSTI POJEDINIH VRSTA GMAZOVA
Natrix natrix
Slika 4. Postaje s potvrđenim nalazima vrste Natrix natrix (crveno) i postaje bez nalaza (zeleno)
Picture 4. Stations with verified findings of Natrix natrix (red) and stations without findings (green)
N. natrix
02468
1012141618
JA ZV BI SV VR KP DU BR OJ DP DJ FN VJ SP SD
Istraživane postaje
Bro
j jed
inki
vrs
te
Slika 5. Ukupni broj zabilježenih jedinki Natrix natrix na istraživanim postajama
Picture 5. The total number of recorded individuals Natrix natrix at the investigated sites
Slika 6. Grafički prikaz sezonske dinamike vrste Natrix natrix
Picture 6. A graphic representation of the seasonal dynamics of Natrix natrix
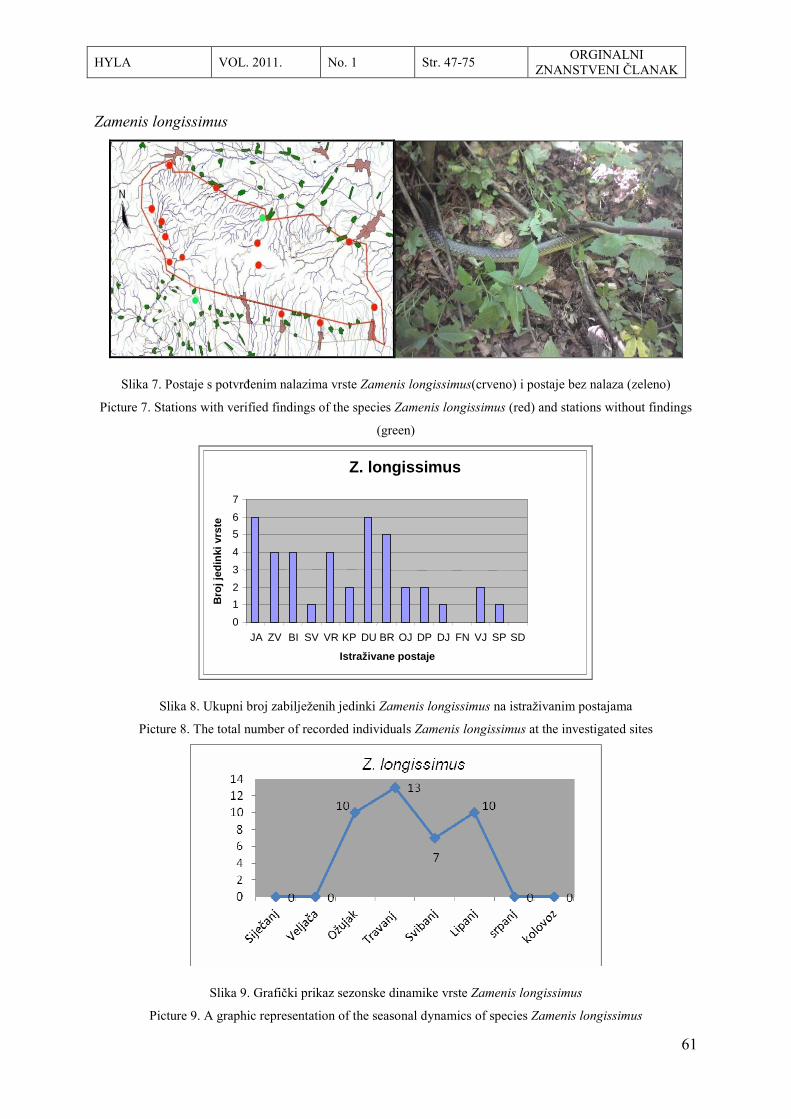
HYLA VOL. 2011. No. 1 Str. 47-75 ORGINALNI ZNANSTVENI ČLANAK
61
Zamenis longissimus
Slika 7. Postaje s potvrđenim nalazima vrste Zamenis longissimus(crveno) i postaje bez nalaza (zeleno)
Picture 7. Stations with verified findings of the species Zamenis longissimus (red) and stations without findings
(green)
Slika 8. Ukupni broj zabilježenih jedinki Zamenis longissimus na istraživanim postajama
Picture 8. The total number of recorded individuals Zamenis longissimus at the investigated sites
Slika 9. Grafički prikaz sezonske dinamike vrste Zamenis longissimus
Picture 9. A graphic representation of the seasonal dynamics of species Zamenis longissimus
Z. longissimus
0 1 2 3 4 5 6 7
JA ZV BI SV VR KP DU BR OJ DP DJ FN VJ SP SD Istraživane postaje
Bro
j jed
inki
vrs
te
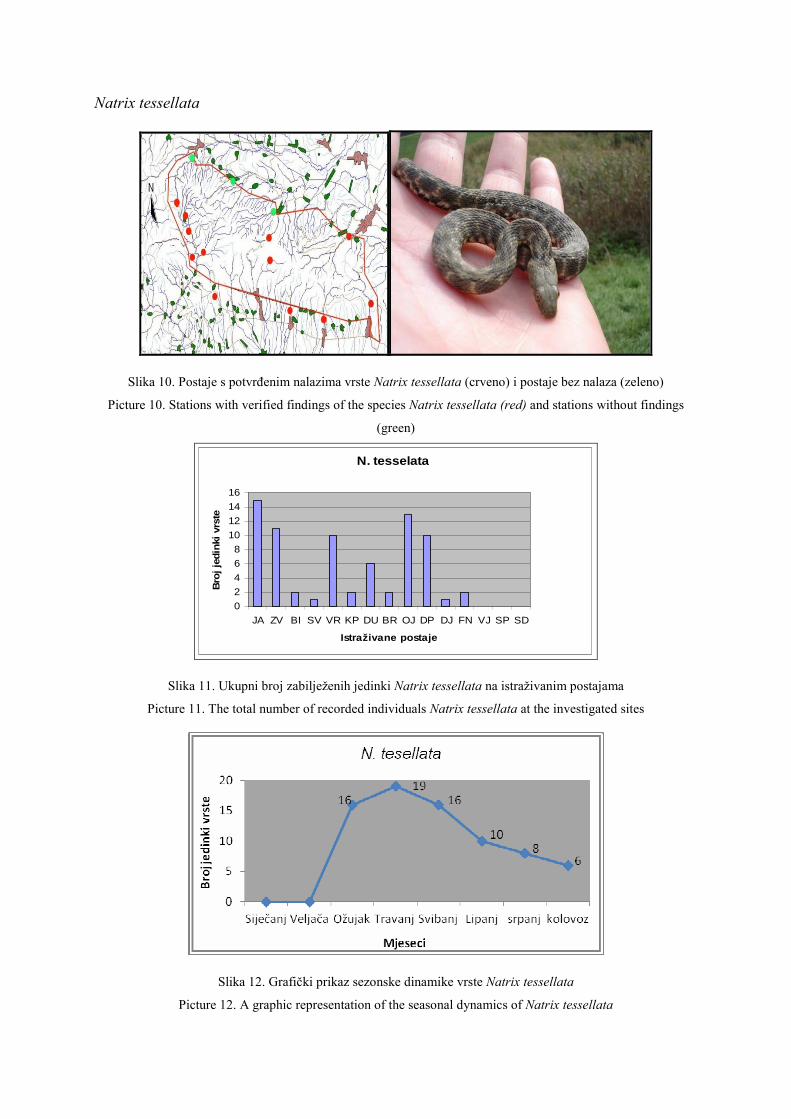
Natrix tessellata
Slika 10. Postaje s potvrđenim nalazima vrste Natrix tessellata (crveno) i postaje bez nalaza (zeleno)
Picture 10. Stations with verified findings of the species Natrix tessellata (red) and stations without findings
(green)
N. tesselata
0
2
4
6
8
10
12
14
16
JA ZV BI SV VR KP DU BR OJ DP DJ FN VJ SP SD
Istraživane postaje
Bro
j jed
inki
vrs
te
Slika 11. Ukupni broj zabilježenih jedinki Natrix tessellata na istraživanim postajama
Picture 11. The total number of recorded individuals Natrix tessellata at the investigated sites
Slika 12. Grafički prikaz sezonske dinamike vrste Natrix tessellata
Picture 12. A graphic representation of the seasonal dynamics of Natrix tessellata

HYLA VOL. 2011. No. 1 Str. 47-75 ORGINALNI ZNANSTVENI ČLANAK
63
Coronella austriaca
Slika 13. Postaje s potvrđenim nalazima vrste Coronella austriaca (crveno) i postaje bez nalaza (zeleno)
Picture 13. Stations with verified findings of the species Coronella austriaca (red) and stations without findings
(green)
C. austriaca
0
1
2
3
4
5
6
JA ZV BI SV VR KP DU BR OJ DP DJ FN VJ SP SD
Istraživane postaje
Bro
j jed
inki
vrs
te
Slika 14. Ukupni broj zabilježenih jedinki Coronella austriaca na istraživanim postajama
Picture 14. The total number of recorded individuals Coronella austriaca at the investigated sites
Slika 15. Grafički prikaz sezonske dinamike vrste Coronella austriaca
Picture 15. A graphic representation of the seasonal dynamics of Coronella austriaca
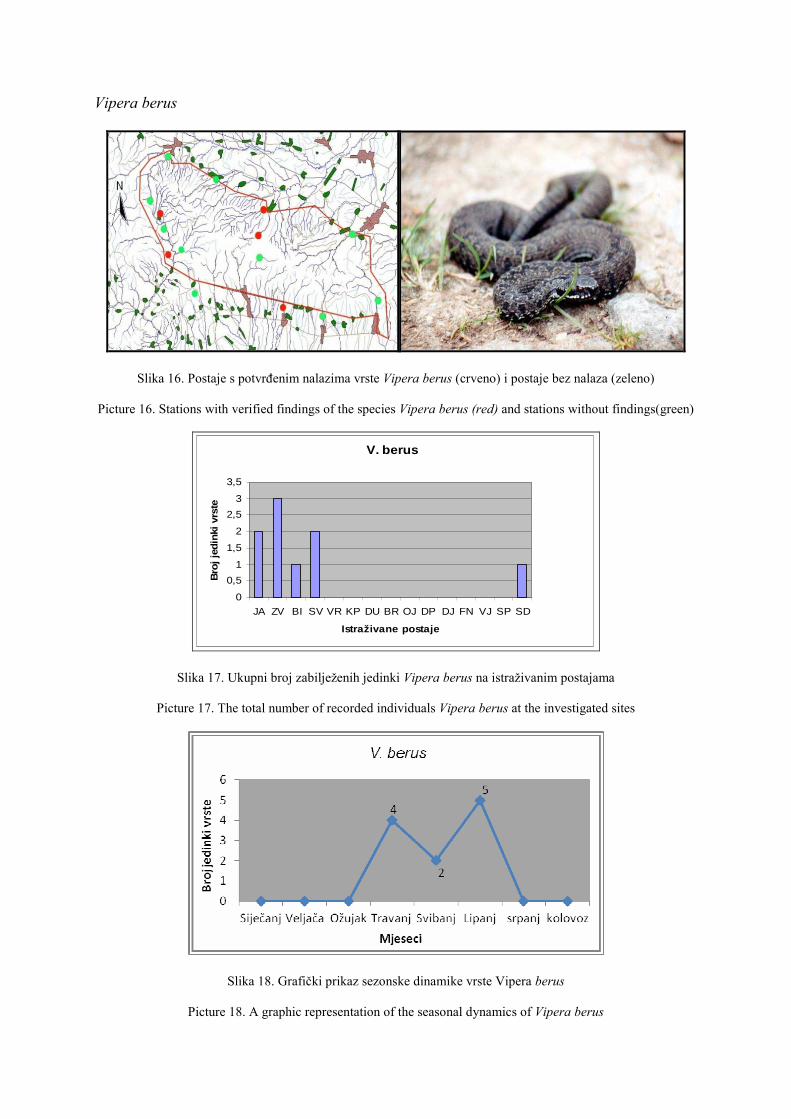
Vipera berus
Slika 16. Postaje s potvrđenim nalazima vrste Vipera berus (crveno) i postaje bez nalaza (zeleno)
Picture 16. Stations with verified findings of the species Vipera berus (red) and stations without findings(green)
V. berus
0
0,5
1
1,5
2
2,5
3
3,5
JA ZV BI SV VR KP DU BR OJ DP DJ FN VJ SP SD
Istraživane postaje
Bro
j jed
inki
vrs
te
Slika 17. Ukupni broj zabilježenih jedinki Vipera berus na istraživanim postajama
Picture 17. The total number of recorded individuals Vipera berus at the investigated sites
Slika 18. Grafički prikaz sezonske dinamike vrste Vipera berus
Picture 18. A graphic representation of the seasonal dynamics of Vipera berus
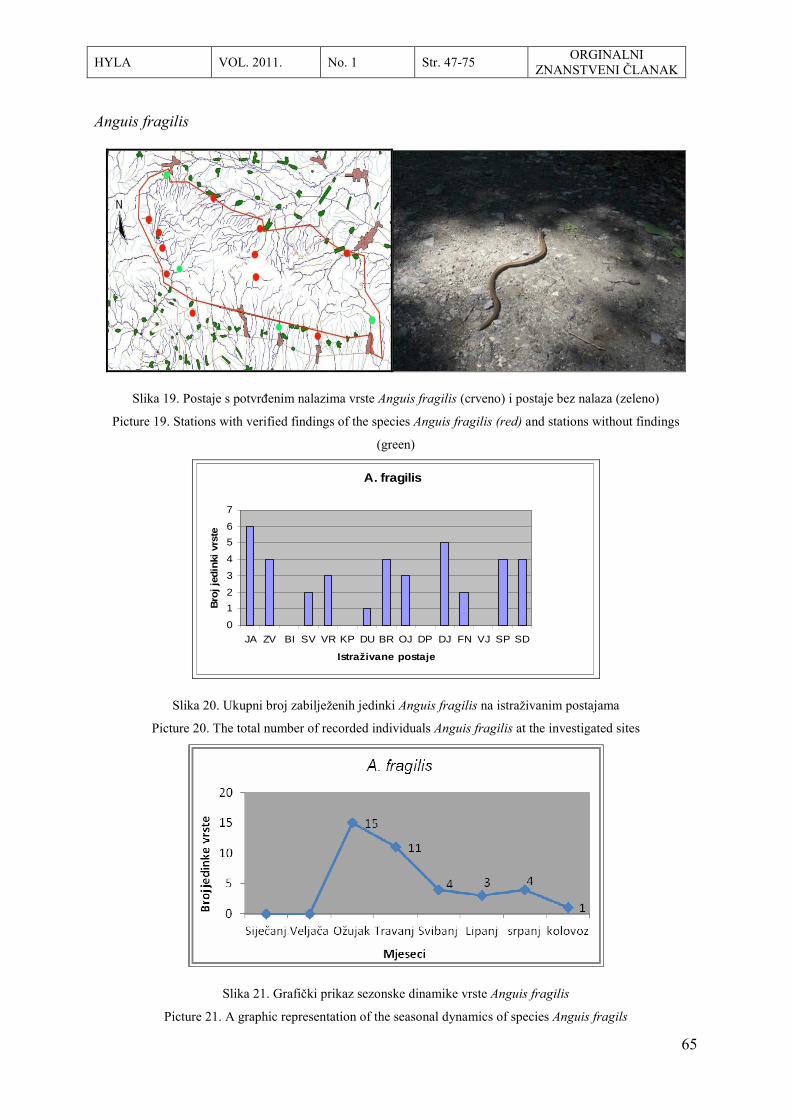
HYLA VOL. 2011. No. 1 Str. 47-75 ORGINALNI ZNANSTVENI ČLANAK
65
Anguis fragilis
Slika 19. Postaje s potvrđenim nalazima vrste Anguis fragilis (crveno) i postaje bez nalaza (zeleno)
Picture 19. Stations with verified findings of the species Anguis fragilis (red) and stations without findings
(green)
A. fragilis
0
1
2
3
4
5
6
7
JA ZV BI SV VR KP DU BR OJ DP DJ FN VJ SP SD
Istraživane postaje
Bro
j jed
inki
vrs
te
Slika 20. Ukupni broj zabilježenih jedinki Anguis fragilis na istraživanim postajama
Picture 20. The total number of recorded individuals Anguis fragilis at the investigated sites
Slika 21. Grafički prikaz sezonske dinamike vrste Anguis fragilis
Picture 21. A graphic representation of the seasonal dynamics of species Anguis fragils
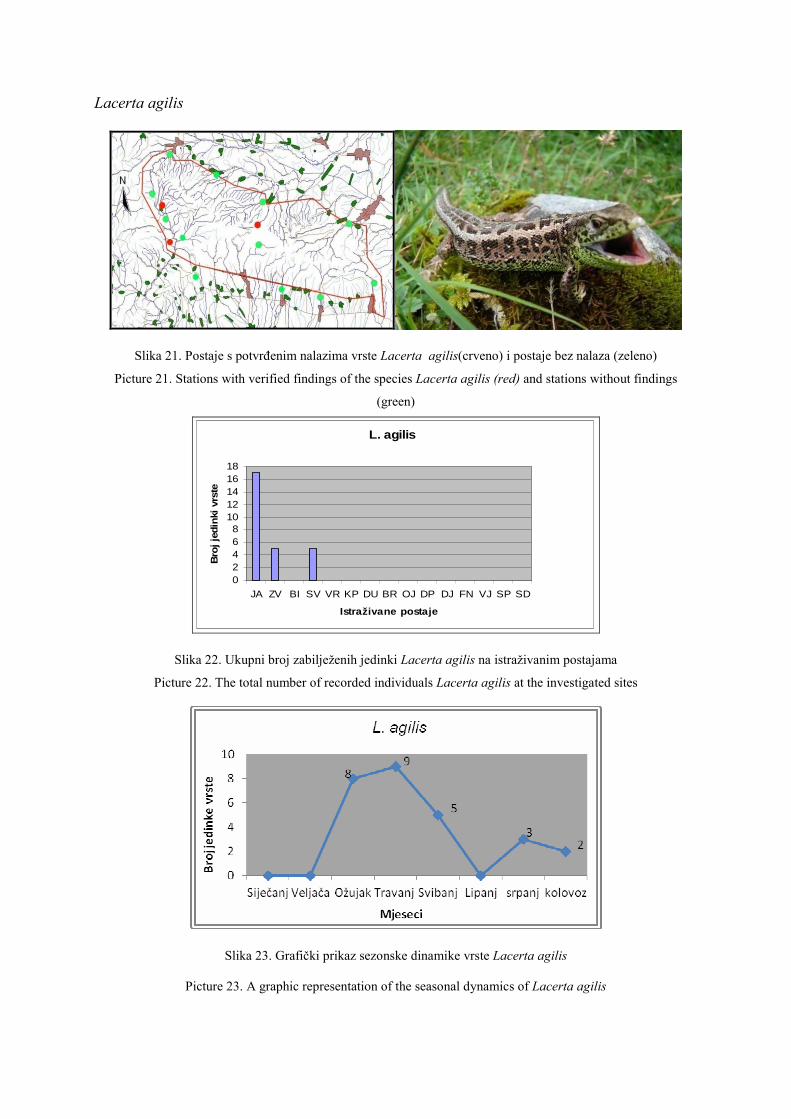
Lacerta agilis
Slika 21. Postaje s potvrđenim nalazima vrste Lacerta agilis(crveno) i postaje bez nalaza (zeleno)
Picture 21. Stations with verified findings of the species Lacerta agilis (red) and stations without findings
(green)
L. agilis
02468
1012141618
JA ZV BI SV VR KP DU BR OJ DP DJ FN VJ SP SD
Istraživane postaje
Bro
j jed
inki
vrs
te
Slika 22. Ukupni broj zabilježenih jedinki Lacerta agilis na istraživanim postajama
Picture 22. The total number of recorded individuals Lacerta agilis at the investigated sites
Slika 23. Grafički prikaz sezonske dinamike vrste Lacerta agilis
Picture 23. A graphic representation of the seasonal dynamics of Lacerta agilis
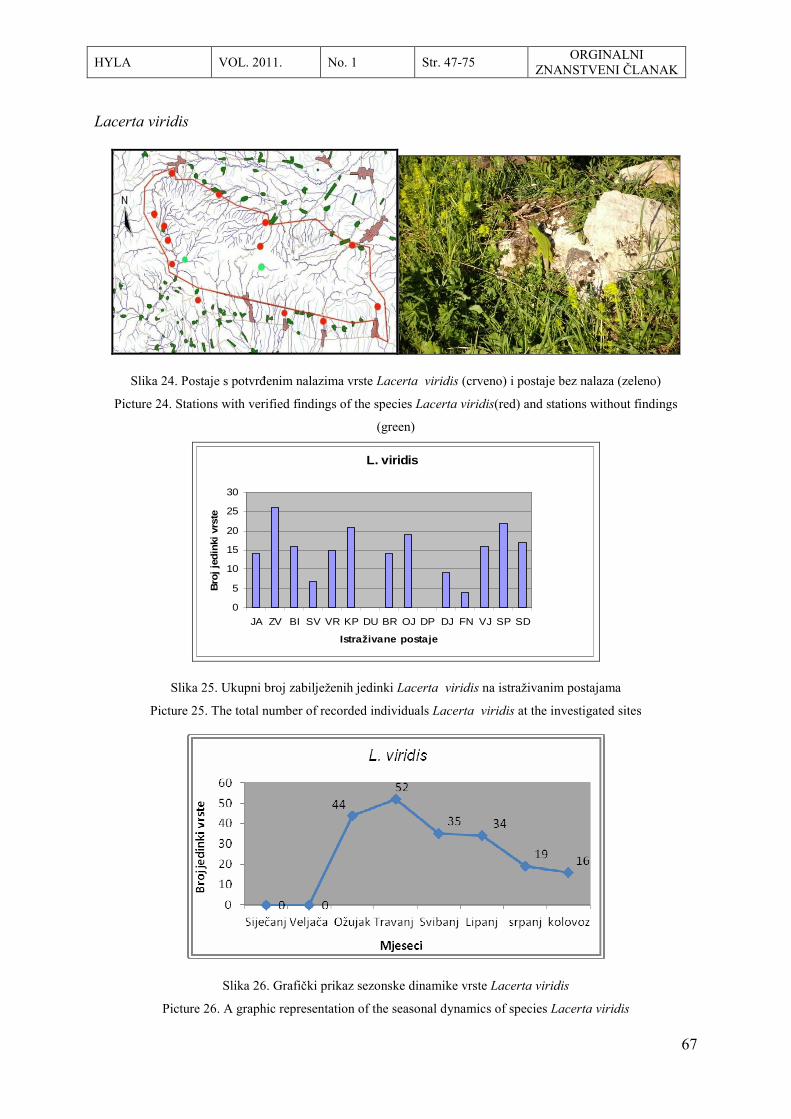
HYLA VOL. 2011. No. 1 Str. 47-75 ORGINALNI ZNANSTVENI ČLANAK
67
Lacerta viridis
Slika 24. Postaje s potvrđenim nalazima vrste Lacerta viridis (crveno) i postaje bez nalaza (zeleno)
Picture 24. Stations with verified findings of the species Lacerta viridis(red) and stations without findings
(green)
L. viridis
0
5
10
15
20
25
30
JA ZV BI SV VR KP DU BR OJ DP DJ FN VJ SP SD
Istraživane postaje
Bro
j jed
inki
vrs
te
Slika 25. Ukupni broj zabilježenih jedinki Lacerta viridis na istraživanim postajama
Picture 25. The total number of recorded individuals Lacerta viridis at the investigated sites
Slika 26. Grafički prikaz sezonske dinamike vrste Lacerta viridis
Picture 26. A graphic representation of the seasonal dynamics of species Lacerta viridis
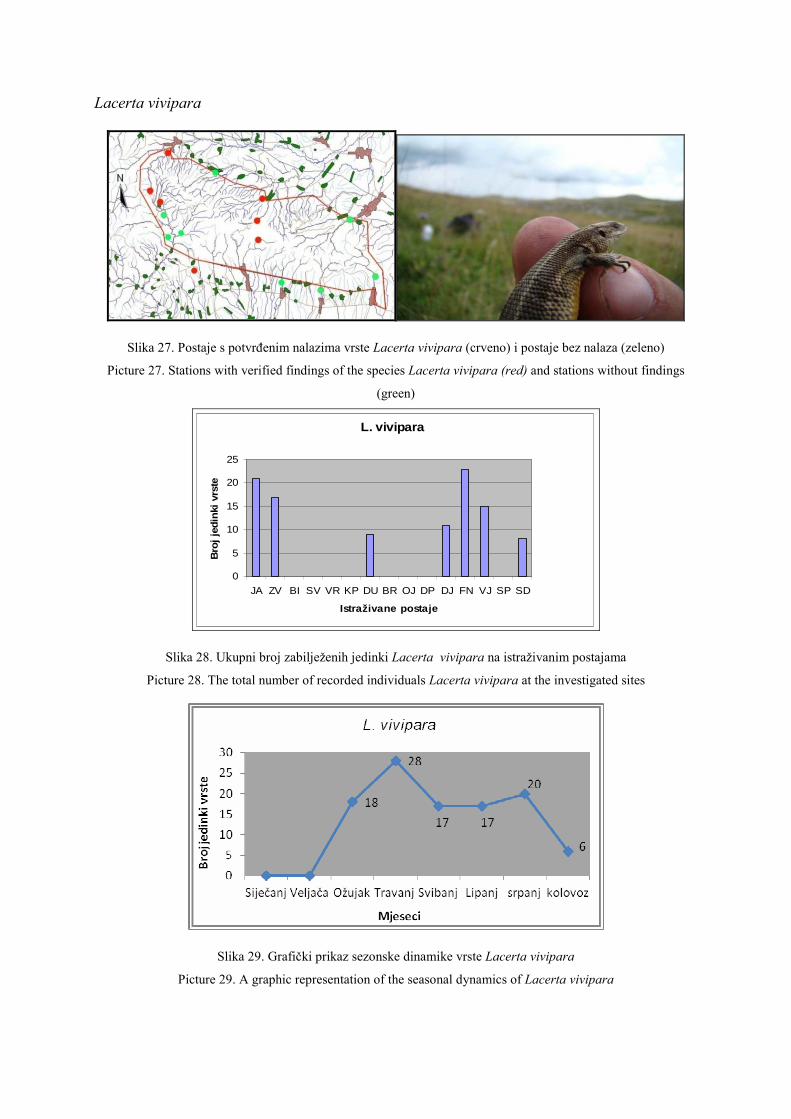
Lacerta vivipara
Slika 27. Postaje s potvrđenim nalazima vrste Lacerta vivipara (crveno) i postaje bez nalaza (zeleno)
Picture 27. Stations with verified findings of the species Lacerta vivipara (red) and stations without findings
(green)
L. vivipara
0
5
10
15
20
25
JA ZV BI SV VR KP DU BR OJ DP DJ FN VJ SP SD
Istraživane postaje
Bro
j jed
inki
vrs
te
Slika 28. Ukupni broj zabilježenih jedinki Lacerta vivipara na istraživanim postajama
Picture 28. The total number of recorded individuals Lacerta vivipara at the investigated sites
Slika 29. Grafički prikaz sezonske dinamike vrste Lacerta vivipara
Picture 29. A graphic representation of the seasonal dynamics of Lacerta vivipara
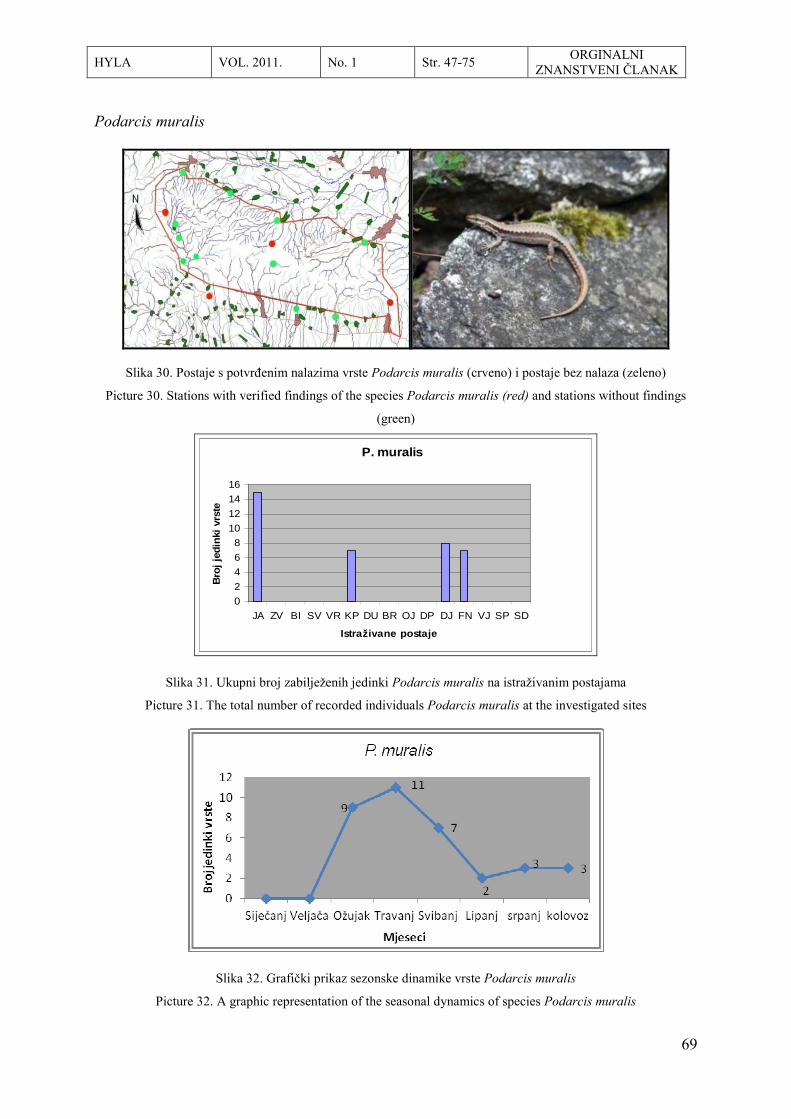
HYLA VOL. 2011. No. 1 Str. 47-75 ORGINALNI ZNANSTVENI ČLANAK
69
Podarcis muralis
Slika 30. Postaje s potvrđenim nalazima vrste Podarcis muralis (crveno) i postaje bez nalaza (zeleno)
Picture 30. Stations with verified findings of the species Podarcis muralis (red) and stations without findings
(green)
P. muralis
0
2
4
6
8
10
12
14
16
JA ZV BI SV VR KP DU BR OJ DP DJ FN VJ SP SD
Istraživane postaje
Bro
j jed
inki
vrs
te
Slika 31. Ukupni broj zabilježenih jedinki Podarcis muralis na istraživanim postajama
Picture 31. The total number of recorded individuals Podarcis muralis at the investigated sites
Slika 32. Grafički prikaz sezonske dinamike vrste Podarcis muralis
Picture 32. A graphic representation of the seasonal dynamics of species Podarcis muralis
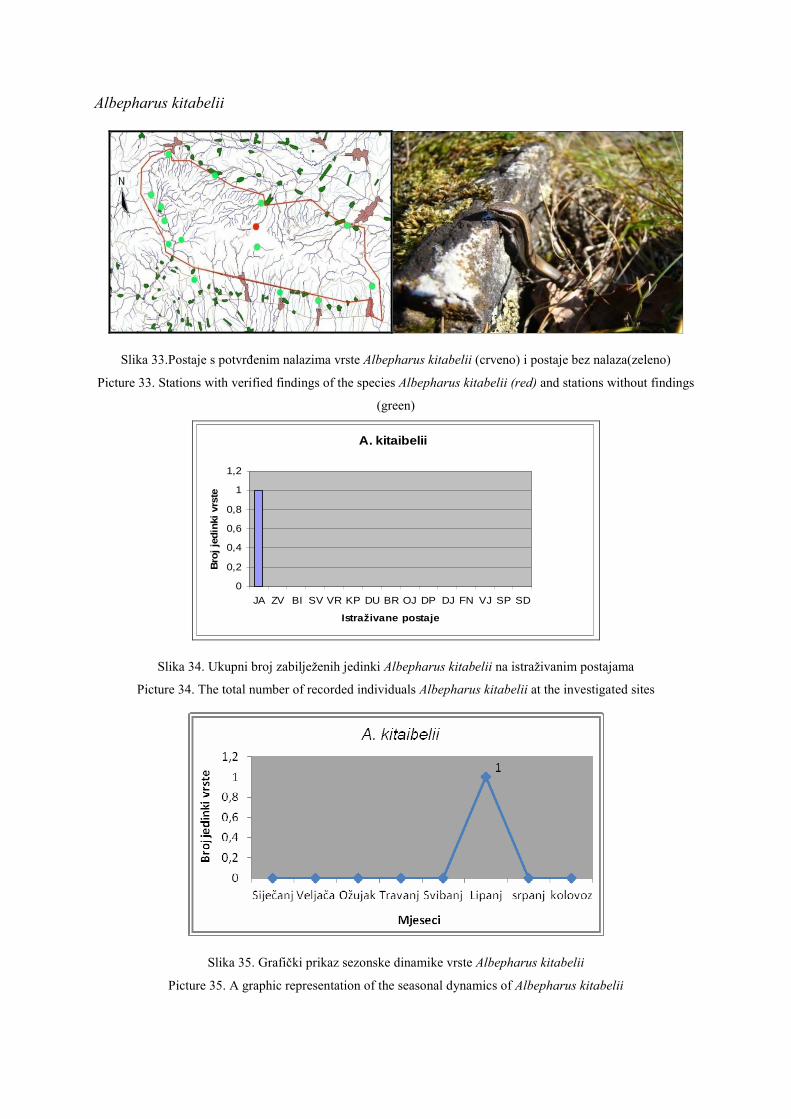
Albepharus kitabelii
Slika 33.Postaje s potvrđenim nalazima vrste Albepharus kitabelii (crveno) i postaje bez nalaza(zeleno)
Picture 33. Stations with verified findings of the species Albepharus kitabelii (red) and stations without findings
(green)
A. kitaibelii
0
0,2
0,4
0,6
0,8
1
1,2
JA ZV BI SV VR KP DU BR OJ DP DJ FN VJ SP SD
Istraživane postaje
Bro
j jed
inki
vrs
te
Slika 34. Ukupni broj zabilježenih jedinki Albepharus kitabelii na istraživanim postajama
Picture 34. The total number of recorded individuals Albepharus kitabelii at the investigated sites
Slika 35. Grafički prikaz sezonske dinamike vrste Albepharus kitabelii
Picture 35. A graphic representation of the seasonal dynamics of Albepharus kitabelii

HYLA VOL. 2011. No. 1 Str. 47-75 ORGINALNI ZNANSTVENI ČLANAK
71
RASPRAVA
Temeljem podataka dobivenih istraživanjem na odabranim nalazištima, načinjeni su grafički
prikazi rasprostranjenosti vrsta i brojnosti njihovih jedinki. U odnosu na ukupnu faunu u
Hrvastskoj koja broji 15 vrsta guštera i 17 vrsta zmija, broj utvrđenih vrsta u Parku Prirode
Papuk zabilježeno je 6 vrsta guštera i 5 vrsta zmija, drugim riječima, u Parku Prirode Papuk
zabilježena je gotovo trećina ukupne faune guštera i zmija u Hrvatskoj. Istočno hrvatska
ravnica (slavonsko gorje, gora Papuk), pruža optimalne ekološke uvjete za razvoj gmazova.
Na tom su području broj navodena i raznovrsna terestrična staništa, bogata različitim
mikrostaništima. Pregledom povijesnih istraživanja uočava se porast broja vrsta, u periodu od
1979 do 2009. godine što ukazuje na trend povećanja raznovrsnosti faune gmazova na tom
području (BOGDANOVIĆ, 2007).
Rezultati istraživanja nam pokazuju da možemo utvrditi tipologiju zajednica populacija
gmazova na istraživanim stanišima. Oni su vrlo važni biološki regulatori u populacijama
drugih vrsta organizama, prije svega kralježnjaka (Pisces, Amphibia, Reptilia, Aves,
Mammalia), a njihov položaj u hranidbenim lancima je od velike važnosti kako u vodenim
tako i kopnenim ekosustavima (KOVACS I BRANDON, 2005).
Analizirajući sezonsku aktivnost utvrđeno je da je cjelokupna sezonska aktivnost vrsta (u
razdoblju od veljače do kolovoza) imala odlike Gaussove krivulje. Dnevne aktivnosti, kako
vrsta tako i jedinki, na pojedinim staništima pokazuju oscilacije vrijednosti od jutarnjih i prije
podnevnih prema poslije podnevnim i noćnim satima. Ovim je potvrđeno pravilo da su
gmazovi nešto aktivniji za lijepa vremena (KOVACS I BRANDON, 2005). Sezonske aktivnosti
koje opisuje Gaussova krivulja poklapaju se s krivuljom srednje godišnje temperature okoliša,
pa je za gmazove optimalno vrijeme pojavljivanja od sredine ožujka do sredine rujna.
Opažanja dnevne dinamike ukazuju da bi optimalno vrijeme za gmazove bilo u
prijepodnevnim satima kada su najaktivniji vjerojatno zbog za njih same povoljnih ekoloških

uvjeta, ali i aktivnosti potencijalnog plijena. To u svakom slučaju ne isključuje mogućnosti
traženja, pronalaženja i uzimanja plijena i tijekom drugih dijelova dana, tako da su podaci koji
govore o oscilacijama aktivnosti tijekom dana brojni (KOVACS I BRANDON, 2005).
Rezultati ovih istraživanja doprinos su poznavanju faune gmazova sjevero istočnog dijela
Hrvatske, te s obzirom na sve veću ugroženost staništa na kojima obitavaju dovodi se u pitanje i
opstanak pojedinih vrsta. Pojedine postaje svojim se zemljopisnim smještajem nalaze
neposredno uz administrativne granice Parka prirode Papuk, što je za značaj istraživanja
svojevrstan „imperativ“ u smislu pridruženja tih prostora u budući sustav zaštite.
Tijekom istraživačkog rada na fauni gmazova Parka prirode Papuk prikupljena je ograničena
količina znanstvenog materijala, koja je pohranjena dijelom u „Zbirci Bogdanović“, a dijelom
u novo utemeljenoj „Zbirci gmazova Parka prirode Papuk“. Ova posljednja ima za svrhu
budućim istraživačima osigurati uvid u materijal prikupljen pri prvoj inventarizaciji gmazova,
a čuvati će se u prostorima Parka prirode Papuk. Nalazi navedenih 11 vrsta gmazova upućuju
da se faunistička istraživanja na ovom području trebaju nastaviti i aktivno uključiti u odgoj i
obrazovanje mladih u očuvanju flore i faune Parka prirode Papuk.
ZAKLJUČCI
• Istraživanja faune gmazova (Reptilia )i biologije pojedinih njihovih vrsta na području
Parka prirode Papuk od veljače do rujna 2009. Godine provedeno je, na petnaest istraživačkih
postaja koje su ciljano obuhvatile različite tipove staništa na kojima gmazovi obitavaju.
• Ukupno je utvrđeno 11 vrsta gmazova.
• Najveću frekvenciju susretanja na istraživanim postajama imale su vrste:Natrix natrix,
Lacerta viridis, Natrix tessellata i Lacerta vivipara, a najmanju vrste Vipera berus i
Ablepharus kitaibelii

HYLA VOL. 2011. No. 1 Str. 47-75 ORGINALNI ZNANSTVENI ČLANAK
73
• Utvrđene su kao eudominantne vrste Natrix natrix, Lacerta viridis, Zamenis
longissimus, Natrix tessellata, dok su recentne vrste Vipera berus i Ablepharus kitaibelii.
• Tijekom dana gmazovi su naj aktivniji u ranim jutarnjim i kasnim poslijepodnevnim
satima, a tijekom sezone – za vrijeme parenja, koje traje od ožujka do svibnja, (Sezona
parenja određena je temperaturom te ako je omogućena pravilna klima i prehrana do parenja
dolazi u proljeće).
• Broj vrsta raste od onih postaja (nalazište JA –Jankovac i ZV – Zvečevo) koje su manje
podložne utjecaju razlika u npr. vodostaju, temperaturi i drugim ekološkim faktorima, prema
istraživanim postajama koje su više izložene tim utjecajima (nalazište VR –Vetovo–ribnjaci i
nalazište KP – Kutjevo–potok).
• Sustavno i temeljito obrađeni su svi dosadašnji poznati podaci o nalazima gmazova, te je
načinjena dopuna cjelovite baze podataka o fauni za područje Parka prirode Papuk.
L ITERATURA
BOGDANOVIĆ, T. (2007): Inventarizacija i valorizacija faune vodozemaca (Amphibia) i
gmazova (Reptilia) Parka prirode „Papuk“. Izvještaj projekta, Osijek.
DZZP (2010): 2. veljače – Svjetski dan močvarnih staništa,
<http://www.dzzp.hr/kategorija/clanak/print.php?id=591> . Pristupljeno 22. kolovoza 2010.
KARAMAN, S. (1921): Beiträge zur Herpetologie von Jugoslavien. Glasn. Hrv. prir. dr.
XXXIII: 194-209.
KOVACS, T. I BRANDON, A. (2005): Herpetofauna of the Drava-valley (2002-2004). Natura
somogyiensis 7: 105-117.
KRIZMANIĆ, I., MESAROŠ, G., DŽUKIĆ, G. I KALEZIĆ, M.L. (1997): Morphology of the Smooth
Newt Triturus vulgaris) in formerYugoslavia: taxonomical implications and distrubution
patterns. Acta Zool. Acad. Sci. Hungaricae, 43 (4), 345-357.

MIKUSKA, J., MIKUSKA, T., MIKUSKA, A,. BOGDANOVIĆ, T. (2004): Gmazovi, Vodič kroz
biološku raznolikost Kopačkog rita, Filozofski fakultet, Osijek.
NAGY, K., GOROGH, Z., KOVACS, T., PECHY, T. & EREIFEJ, L. (2006): Keteltuek- es hullok
elterjedesenek orszagos terkepezese es monitorozasa (KHTM). Magyar Madartani es
Termeszetvedelmi Egyesulet, Keteltii- es Hiillovedelmi Szakosztaly, Budapest, Verzio: 1.1.
PAVLETIĆ J. (1964): Amphibia i Reptilia zbirke Hrvatskog narodnog zoološkog muzeja u
Zagrebu. Hrv. narod. zool. muz. 4: 1-37.
PETZ, B., (1985): Osnovne statističke metode za nematematičare, Nakladnik slap, Zagreb.
RADOVANOVIĆ, M. (1951): Vodozemci i gmizavci naše zemlje. Naučna knjiga, Beograd.
SAMARDŽIĆ, I., TOMAŠEVIĆ,M. (2000): Zaštićene, rijetke i ugrožene biljne vrste Slavonskoga
gorja. Spin Valis d. d. Požega.
SOTIROPOULOS, K., ELEFTHERAKOSA, K., DŽUKIĆ, G., KALEZIĆ, M.L., LEGAKISD, A. &
POLYMENI, R.M. (2007): Phylogeny and biogeography of the alpine newt Mesotriton alpestris
(Salamandridae, Caudata), inferred from mt DNA sequences. Molecular Phylogenetics and
Evolution Vol. 45, Issue 1: 211-226.
SZÖVÉNYI, G. I D. JELIĆ (2011): Distribution and conservation status of Snake eyed skink
(Ablepharus kitaibelii Bibron & Bory, 1833) in Croatia. NW Journal of Zoology, Volume 7,
Issue 1 (in press).

HYLA VOL. 2011. No. 1 Str. 47-75 ORGINALNI ZNANSTVENI ČLANAK
75
SUMMARY
INVENTARISATION AND VALORIZATION OF REPTILE (REPTILIA) FAUNA IN THE NATURE
PARK PAPUK
FRANO BARIŠIĆ *1, TOMISLAV BOGDANOVIĆ1
Research of fauna and biology of particular species of reptiles in the area of Nature park
Papuk were done in the period from February 2009 to September 2009, in 15 locations. The
transect method was used in data gathering on reptiles. The sampled units were measured for
length or photographed, and during sampling the environmental parametres were measured.
The collected data were entered in the database and with the help of GIS technology
distribution maps of particular species on the examined area were made. The species Natrix
natrix and Lacerta viridis had the largest quantitative structure. The eudominant species were
established to be Natrix natrix, Lacerta viridis, Natrix tessellata and Lacerta vivipara, while
the recent species are Vipera berus and Ablepharus kitaibelii. The numerousness of species in
the researched area as well as their distribution are the result of various conditions in
microhabitats, where species find the optimal conditions for survival. Furthermore, seasonal
and daily dynamics were analysed. Daily activities, of species and of individuals, show a
decrease of value from morning to afternoon, and as far as seasonal dynamics is concerned,
the most intensive period of appearance of reptile is in the period from July to September.
Keywords: species listing, evaluation, eudominat species, recent species, species frequencies


HYLA VOL. 2011. No. 1 Str. 77-79 PHOTO NOTE
77
Preliminary data on existence of Zootoca vivipara ssp. pannonica (Lac & Kluch, 1968) in
Croatia
Preliminarni podatci o postojanju Zootoca vivipara ssp. pannonica (Lac & Kluch, 1968)
u Hrvatskoj
DUŠAN JELIĆ1, TOMISLAV BOGDANOVIĆ2
1 State Institute for Nature Protection, Trg Mažuranića 5, 10 000 Zagreb, Croatia, [email protected]
2 Josip Juraj Strossmayer University Of Osijek, Department of Biology, Trg Ljudevita Gaja 5, 31000 Osijek,
Croatia, [email protected].
Figure 1. Zootoca vivipara ssp. pannonica from Spačva.
Slika 1. Zootoca vivipara ssp. pannonica iz Spačve.

Zootoca vivipara (VON JACQUIN, 1787) is known to appear in different mountain areas in
North - Western Croatia (RADOVANOVIĆ, 1951; DELY & BÖHME, 1984; AGASYAN et al.,
2009). It is usually found in colder and humid mountain habitats from 700 to 1700 m a.s.l.
These populations all belong to the subspecies Z. v. vivipara (VON JACQUIN, 1787) and Z. v.
carniolica MAYER, BÖHME, TIEDEMANN & BISCHOFF, 2000 (HEULIN et al., 2000; GUILLAUME
et al., 2006). In Europe, Z. v. pannonica is known to appear in lowland grasslands, marshes and
semi-steppe in Hungary and Austria (DELY & BÖHME, 1984; AGASYAN et al., 2009). Until
now mountain Papuk was the most Eastern locality in Croatia (RADOVANOVIĆ, 1951,
BOGDANOVIĆ, 2008, BARIŠIĆ & BOGDANOVIĆ, 2011) but existence of the lowland Z. v.
pannonica was never recorded. Authors bring new data that this taxa was recorded from 2009
till 2011 in several sites in Slavonia: Spačva, surroundings of Osijek, Vinkovci and Slavonski
Brod town. Primary habitats were lowland (80 – 120 m a.s.l.), partly flooded grasslands and
forests. Complete data will be published after further research in a separate publication. IUCN
category for Zootoca vivipara pannonica is Vulnerable B1+2bcd, C2a (ver 2.3) (AGASYAN et
al., 2009).
REFERENCES
AGASYAN, A., AVCI, A., TUNIYEV, B., CRNOBRNJA ISAILOVIC, J., LYMBERAKIS, P., ANDRÉN,
DAN COGALNICEANU, C., WILKINSON, J., ANANJEVA, N., ÜZÜM, N., ORLOV, N., PODLOUCKY,
R., TUNIYEV, S., KAYA, U., BÖHME, W., NETTMANN, H.K., CRNOBRNJA ISAILOVIC, J., JOGER,
U., CHEYLAN, M., PÉREZ-MELLADO, V., BORCZYK, B., STERIJOVSKI, B., WESTERSTRÖM, A. &
SCHMIDT, B. (2009): Zootoca vivipara. In: IUCN 2011. IUCN Red List of Threatened
Species. Version 2011.1. <www.iucnredlist.org>. Downloaded on 10th July 2011.
BARIŠIĆ, F. & BOGDANOVIĆ, T. (2011): Gmazovi (Reptilia) Parka Prirode Papuk. HYLA, Vol
2011., No 1., 42-74.

HYLA VOL. 2011. No. 1 Str. 77-79 PHOTO NOTE
79
BOGDANOVIĆ T. (2008): Inventarizacija i valorizacija faune vodozemaca (Amphibia) i
gmazova (Reptilia) Parka prirode Papuk, stručna studija, JU PP Papuk, Velika.
DELY, O.G. & BÖHME, W. (1984): Lacerta vivipara - Waldeidechse. In: Böhme,W. (ed.),
Handbuch der Reptilien und Amphibien Europas, Band 2/I., Echsen II (Lacerta). Aula-Verlag
Wiesbaden, pp. 362-393.
DŽUKIĆ, G. & KALEZIĆ, M. (2004); The biodiversity of amphibians and reptiles in the Balkan
Peninsula in: Balkan biodiversity. Papers from the ESF Exploratory Workshop on Balkan
biodiversity - Koper, September 2001. Griffiths, H.; Krystufek, B.; Griffiths, J. (eds.)
Guillaume, C.P., Heulin, B., Pavlinov, I.Y., Semenov, D.V., Bea, A., Vogrin, N. & Surget-
Groba, Y. (2006): Morphological variations in the common lizard, Lacerta (Zootoca)
vivipara. Russian Journal of Herpetology 13:1-10.
HEULIN, B., GUILLAUME, C.-P., VOGRIN, N., SURGET-GROBA, Y. & TADIĆ, Z. (2000): Further
evidence of the existence of oviparous populations of Lacerta (Zootoca) vivipara in the NW
of the Balkan Peninsula. Comptes Rendus de l'Académie des Sciences de Paris, Sciences de la
vie 323: 461-468.
RADOVANOVIĆ, M. (1951): Vodozemci i gmizavci naše zemlje. Naučna knjiga, Beograd.


81
Upute autorima
Rukopisi moraju biti dostavljeni isključivo u elektronskom obliku (na e-mail:
[email protected]), kao i originalne slike i tablice. Tekst rukopisa treba biti u formatu MS-
Word, font Times New Roman veličine slova 12, bez fusnota, s dvostrukim proredom na A4 (210
x 297 mm) papiru s marginama od najmanje 25 mm na svakoj strani. Sve stranice trebaju biti
numerirane u donjem desnom kutu.
Za sve vrste radova, naslov stranice mora sadržavati samo sljedeće: naslov rada (podebljan);
ime(na) autora (SMALL CAPS); adresu ustanove u kojoj je rad napravljen te adresu e-pošte
autora za korespondenciju (veličine slova 10). Naslov treba biti sažeta i informativna sinteza
istraživanja. Gdje je prikladno treba spomenuti porodicu ili višu svojtu - Lacerta agilis Linnaeus,
1758 (Squamata, Lacertidae).
- Izvorni znanstveni rad - treba sadržavati sažetak, uvod, materijale i metode, rezultate, raspravu
i literaturu (vidi primjer rukopisa dolje).
- Kratko priopćenje - općenito manje od četiri-pet stranica rukopisa koje su napisane kao
kontinuirani tekst sa sažetakom na početku s najviše 150 riječi.
- Slikovno priopćenje - izvješće o zanimljivim nalazima uhvaćenim na fotografiji i objašnjeno s
najviše 150 riječi teksta. Moguće je objaviti jednu ili više fotografija.
- Sažetak o vrsti - sažetak o trenutno poznatim podacima o određenoj vrsti (treba uključiti
distribuciju, ekologiju, ponašanje i pregled literature)
Literatura koja se citira treba biti napisana kako slijedi u tekstu, SMALL CAPS na sljedeći način:
(PIANKA, 1989, HUEY & PIANKA, 1981, HAYDON ET AL., 1997). Literatura koja se u tekstu navodi
zajedno treba biti napisana kronološki. Svi radovi navedeni u tekstu trebaju biti navedeni u
popisu literature. Literaturu "u tisku" treba navesti samo onda kada je prihvaćena za tisak. Imena
osoba koje su ustupile neobjavljene informacije trebaju biti citirana kao u primjeru:
“(ANDERSSON, Stockholm, pers. comm. 2005)”. U popisu literature pod naslovom Literatura
autore navodimo po abecednom redu te po datumu objave publikacije kod jednog autora.
Imena časopisa i serija moraju biti napisani u potpunosti.

Literaturu citirajte na sljedeći način:
HAYDON, D. T., CROTHER, B. I. & PIANKA. E. R. (1994): New directions in biogeography? Trends
in Ecology and Evolution 9: 403-406.
SOKAL, R. R. & ROHLF, F. J. (1995): Biometry. The principles and practice of statistics in
biological research. 3rd Edition. W. H. Freeman & co, New York.
HUEY, R. B. & PIANKA, E. R., (1983): Temporal separation of activity and interspecific dietary
overlap (with an Appendix by S. L. Pimm). pp. 281-296. In HUEY, R. B.,. PIANKA, E. R.,
SCHOENER T. W. (eds.) Lizard Ecology: Studies of a Model Organism. Harvard University Press.
IUCN (2010): Global Mammal Asessement. IUCN - International Union for Conservation of
Nature, <http://www.iucn.com> . Pristupljeno 21. siječnja 2010.
Svaki rad pregledati će 1-2 nezavisna recenzenta.

83
Naslov koji sažeto opisuje sadržaj rada
IME PREZIME1, IME PREZIME *2
1 Ime institucije, Radučka 15, 10000 Zagreb, Croatia, vaš[email protected] 2 Ime institucije, Heinzlova 55, 10000 Zagreb, Croatia
SAŽETAK : Sažetak treba biti jasan, pregledan i ne duži od 300 riječi, treba objediniti ključne
rezultate i zaključke. Također treba uključiti četiri do pet ključnih riječi.
Ključne riječi: četiri, pet, ključna riječ, uključen
UVOD
U uvodu treba staviti jezgrovit opis pozadine (problema), s logičkom podlogom, svrhom i
konkretnim ciljevima rada. Očekuje se dobra literaturna podloga, ako postoji.
MATERIJALI I METODE
Materijali i metode trebaju pružiti dostatne informacije da bi se dozvolilo ponavljanje
eksperimenta i/ili terenskog rada. Tehnički opis metoda istraživanja je poželjan samo ako se radi
o novim ili važnim metodama za razumijevanje rezultata.
REZULTATI
Ovaj dio mora sadržavati precizno predstavljanje rezultata istraživanja. Izbjegavajte
predstavljanje istih informacija kao teksta i/ili prikaza i/ili tablice. Ovdje ubacite tekst, tablice i
ilustracije. Za svaku ilustraciju potrebno je napisati referencu u tekstu. Također, spremite,
priložite i pošaljite vaše ilustracije zasebno, koristeći naziv datoteke TablicaBr.ekstenzija (npr.
Tab-1 korelacija SVL i mase kod ženki Lacerta vivipara.tiff).
Objašnjenje tablica, grafikona i slika treba staviti u legendu, koristeći Times New Roman font,
veličina 10. Objašnjenja za tablice pišu se iznad, lijevo poravnata, a objašnjenja slika i grafikona
pišu se ispod, centrirano poravnata.
Tablica 1. Kratak naslov razumljiv sam po sebi (Times New Roman 10)
no dostatno razumljiv NS 1 SPSV2 kratak,
NS2 NS 3 prvi drugi treći
kratak (cm) 16.69a 15.18b 15.95 15.94 15.92 0.31
kratak (kg) 14.83 14.83 14.97a 15.23a 14.29b 0.22
kratak (kg/min) 2.55 2.74 2.45 2.83 2.66 0.22 1 NS – Naslov stupca; 2 Standardna pogreška srednje vrijednosti; a,b= P<0.01.
RASPRAVA
Ovaj dio bi trebalo odvojiti od dijela s rezultatima kod izvornih znanstvenih radova, i trebao bi se
baviti značenjem rezultata i njihovom vezom s ciljevima istraživanja. Također bi trebalo uključiti
kako će rezultati istraživanja promijeniti odnosno utjecati na naše znanje o predstavljenom
organizmu ili staništu.
L ITERATURA
Slijedite upute na stranici 1.
ZAHVALE
Zahvale i bilo koje dodatne informacije koje se tiču financiranja istraživanja.

85
Author Guidelines
Manuscripts must be submitted in electronic version only (on [email protected]), as well as
the original figures and tables. The manuscript text should be MS-Word processed, typed
throughout in letter quality with font size 12, Times New Roman, without footnotes, double
spaced, on A4 (210 x 297 mm) paper, with margins of at least 25 mm on each side. All pages
should be numbered consecutively in the bottom, right-hand corner.
For all types of papers, the title page must contain and only contain the following: title of paper
(bold); name(s) of the author(s) (in Small caps); address of the Institution where the work
was done, and the email of the corresponding author (font size 10). Provide a title that is
concise but also an informative synthesis of the study.
- Original scientific paper – should include summary, introduction, materials and methods,
results, discussion and references (see example manuscript below).
- Short note - generally less than four-five manuscript pages; should be produced as continuous
text, preceded by an abstract of no more than 150 words.
- Photo note – reports of interesting findings captured on photographs, explained with maximum
150 words of text. It is possible to publish one or more photographs.
- Species summary – summary of currently known data about certain species (should include
distribution, ecology, behavior and literature overview)
Cited sources should written in Small caps and referred to as follows: (PIANKA, 1989, HUEY &
PIANKA, 1981, HAYDON ET AL., 1997). References cited together in the text should be arranged
chronologically. All publications cited in the text should be presented in a list of references.
References "in press" shall only be cited when they have been accepted for publication. Names
of persons who provided unpublished information should be cited as follows: “(ANDERSSON,
Stockholm, pers. comm. 2005)”. List references alphabetically by author under References and
date of publication at the same author.
Journal and series names have to be spelled out fully.

Use the following format for references section:
HAYDON, D. T., CROTHER, B. I. & PIANKA. E. R. (1994): New directions in biogeography? Trends
in Ecology and Evolution 9: 403-406.
SOKAL, R. R. & ROHLF, F. J. (1995): Biometry. The principles and practice of statistics in
biological research. 3rd Edition. W. H. Freeman & co, New York.
HUEY, R. B. & PIANKA, E. R. (1983): Temporal separation of activity and interspecific dietary
overlap (with an Appendix by S. L. Pimm). pp. 281-296. In Huey, R. B.,. Pianka, E. R., Schoener
T. W. (eds.) Lizard Ecology: Studies of a Model Organism. Harvard University Press.
IUCN (2010): Global Mammal Asessement. IUCN - International Union for Conservation of
Nature, <http://www.iucn.com> . Pristupljeno 21. sijecnja 2010.
Each paper will be reviewed by 1-2 independent reviewers.

87
Title that succinctly describes the contents of the paper
NAME SURNAME1, NAME SURNAME*2
1 AffiliationName, Radučka 15, 10000 Zagreb, Croatia, [email protected] 2 AffiliationName Heinzlova 55, 10000 Zagreb, Croatia
SUMMARY : The abstract should be clear, descriptive and not longer than 300 words, should
summarize the essential results and conclusions. Four or five keywords should also be included.
Key words: Four, Five, Keywords, Included
INTRODUCTION
Has to be a concise description of the background, rationale, aims and specific objectives of the
paper. Good literature background, if existing, is expected.
MATERIAL AND METHODS
Materials and methods have to provide sufficient information to permit repetition of the
experiment and/or fieldwork. The technical description of study methods should be given only if
such methods are new or important for the understanding the results.
RESULTS
Results section must be a concise presentation of the finding of the study. Avoid the presentation
of same information as text and/or figure and/or table. Insert text, tables and illustrations here.
References should be made in the text to each illustration. Also, save, attach and send your
illustrations separately using the file name TableNo.Explination.extension (ex. Tab-1Female
mass to SVL correlation in Lacerta vivipara.tiff).
Explanations of tables, graphs and pictures should be written in English and Croatian (note if one
is missing and it will be added by editor), given in the typewritten legend, use Times New
Roman font, size 10. Explanations of tables are written above, aligned left, and explanations of
pictures and graphs are written underneath, centre aligned.
Table 1. Brief and self-explanatory title (Times New Roman 10)
DISCUSSION
Discussion section should be separate from the results section at full-length papers and should
deal with the significance of the results and their relationship to the aims of the paper. Also
include how the findings of the paper will change, influence the state of our knowledge about
model organism or habitat.
REFERENCES
Follow the guidelines on page 1.
ACKNOWLEDGEMENTS
Acknowledgements and any additional information concerning research grants.



















