
Human NLRP3 inflammasome senses multiple types of ... · PDF fileHuman NLRP3 inflammasome...
6
Human NLRP3 inflammasome senses multiple types of bacterial RNAs Wenwen Sha a,b , Hiroki Mitoma b,c , Shino Hanabuchi b,d , Musheng Bao b,d , Leiyun Weng b , Naoshi Sugimoto b,e , Ying Liu b,f , Zhiqiang Zhang a,b,g , Jin Zhong a , Bing Sun a,1 , and Yong-Jun Liu a,b,d,1 a Key Laboratory of Molecular Virology and Immunology, Institute Pasteur of Shanghai, Chinese Academy of Sciences, Shanghai 200031, China; b Baylor Institute for Immunology Research, Baylor Scott & White Health Service, Dallas, TX 75204; c Department of Clinical Immunology and Rheumatology/Infectious Disease, Kyushu University Hospital, 3-1-1 Maidashi, Higashi-ku, Fukuoka, 812-8582, Japan; d MedImmune, Gaithersburg, MD 20878; e Department of Hematology and Oncology, Graduate School of Medicine, Kyoto University, Kyoto 606-8507, Japan; f The Key Laboratory of Infection and Immunity, Institute of Biophysics, Chinese Academy of Sciences, Beijing 100101, China; and g Immunobiology and Transplant Research, Houston Methodist Hospital and Houston Methodist Research Institute, Texas Medical Center, Houston, TX 77030 Edited by Tak W. Mak, The Campbell Family Institute for Breast Cancer Research at Princess Margaret Cancer Centre, Ontario Cancer Institute, University Health Network, Toronto, ON, Canada, and approved October 10, 2014 (received for review July 3, 2014) Inflammasomes are multiprotein platforms that activate caspase-1, which leads to the processing and secretion of the proinflammatory cytokines IL-1β and IL-18. Previous studies demonstrated that bacte- rial RNAs activate the nucleotide-binding domain, leucine-rich-repeat- containing family, pyrin domain-containing 3 (NLRP3) inflammasome in both human and murine macrophages. Interestingly, only mRNA, but neither tRNA nor rRNAs, derived from bacteria could activate the murine Nlrp3 inflammasome. Here, we report that all three types of bacterially derived RNA (mRNA, tRNA, and rRNAs) were capable of activating the NLRP3 inflammasome in human macrophages. Bacte- rial RNA’s5′-end triphosphate moieties, secondary structure, and double-stranded structure were dispensable; small fragments of bac- terial RNA were sufficient to activate the inflammasome. In addition, we also found that 20-guanosine ssRNA can activate the NLRP3 inflammasome in human macrophages but not in murine macro- phages. Therefore, human and murine macrophages may have evolved to recognize bacterial cytosolic RNA differently during bacterial infections. bacterial RNA | single-stranded RNA | NLRP3 inflammasome | innate immunity | primary macrophages T he innate immune system is the first line of defense against microbial infections. Germ-line–encoded pattern-recognition receptors (PRRs) of the innate immune system recognize the presence of invariant evolutionarily conserved microbial com- ponents called “pathogen-associated molecular patterns” (1–3). In response to microbial infections, PRRs rapidly initiate signal- transduction pathways to induce type 1 IFN production, proin- flammatory cytokine production, and inflammasome activation. The inflammasome is a cytosolic large caspase-1–containing multi- protein complex that enables autocatalytic activation of caspase-1. Once caspase-1 is activated, it starts to cleave prointerleukin-1β (pro–IL-1β) and prointerleukin-18 (pro–IL-18) proteolytically into bioactive IL-1β and IL-18 (4–7). The mature forms of IL-1β and IL- 18 play roles in a variety of infectious and inflammatory processes. Cytosolic microbial nucleic acids are important activators of the innate immune system against both bacterial and viral infections, which induce type 1-IFN and proinflammatory cyto- kine responses as well as inflammasome activation. The role of microbial nucleic acids in inflammasome activation has been stud- ied mostly in murine bone marrow-derived dendritic cells (BMDCs) or bone marrow-derived macrophages (BMDMs). AIM2 has been identified as a specific cytosolic dsDNA sensor that directly binds ASC (apoptosis-associated speck-like protein containing a carboxyl-terminal CARD-like domain) and forms inflamma- some complexes in human and murine cells (8–11). Viral dsRNA was found to activate the nucleotide-binding domain, leucine-rich-repeat-containing family, pyrin domain- containing 3 (NLRP3) inflammasome in human and murine cells (12–15). Several groups have reported that cytosolic bacterial RNA activate the Nlrp3 inflammasome in murine macrophages (13, 16, 17). Our group also has reported that human THP-1– derived macrophages recognize cytosolic bacterial RNA and induce NLRP3 inflammasome activation (12). Bacterial RNA is composed of mRNA, tRNA, and three different sizes of rRNA (23s, 16s, and 5s). Sander et al. (18) reported that, of the different types of Escherichia coli RNA, only E. coli mRNA induced the secretion of IL-1β by murine BMDMs, but E. coli tRNA and E. coli rRNAs did not. We aimed to study (i ) whether a variety of cytosolic bacterial RNAs could activate the inflammasome in human myeloid cells and (ii ) what types of bacterial RNA activate the inflammasome in human and murine myeloid cells. Here, we demonstrate that a broad spectrum of cytosolic bacterial RNAs strongly induce the cleavage of caspase-1 and the secretion of IL-1β and IL-18 in human macrophages. Human macrophages can sense mRNA, tRNA, rRNAs, and small synthetic ssRNA through NLRP3, but murine macrophages can sense only the mRNA component. Bacterial RNA’s5′-end triphosphate moieties, secondary struc- ture, and double-stranded structure were dispensable, but small fragments of bacterial RNA were sufficient to activate the inflammasome. These findings suggest that upon bacterial infec- tions the human and murine NLRP3 inflammasomes sense cyto- solic bacterial RNAs differently. Significance The innate immune system has evolved to protect the host from potential pathogens. The nucleotide-binding domain, leucine-rich-repeat-containing family, pyrin domain-containing 3 (NLRP3) inflammasome is one of the platforms that can sense pathogenic bacteria, which can cause fatal bacterial infections. Here we show that all three types of bacteria-derived RNA— mRNA, tRNA, and rRNAs—as well as synthetic 20-guanosine ssRNA, are capable of activating the NLRP3 inflammasome and inducing human macrophages to secrete inflammatory cyto- kines. Interestingly, only bacterial mRNA is able to activate the murine Nlrp3 inflammasome. Therefore human macrophages may have evolved in a unique fashion, adapting to the bacte- rial environment. This study could provide important clues for developing efficient medicines for bacterial infections and various immunotherapies such as anticancer vaccines. Author contributions: W.S., H.M., J.Z., B.S., and Y.-J.L. designed research; W.S., H.M., M.B., and L.W. performed research; Y.L. contributed new reagents/analytic tools; W.S., H.M., and Y.-J.L. analyzed data; and W.S., H.M., S.H., N.S., Z.Z., and Y.-J.L. wrote the paper. The authors declare no conflict of interest. This article is a PNAS Direct Submission. 1 To whom correspondence may be addressed. Email: [email protected] or liuyo@ medimmune.com. This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10. 1073/pnas.1412487111/-/DCSupplemental. www.pnas.org/cgi/doi/10.1073/pnas.1412487111 PNAS | November 11, 2014 | vol. 111 | no. 45 | 16059–16064 IMMUNOLOGY AND INFLAMMATION
-
Upload
truongmien -
Category
Documents
-
view
218 -
download
1
Transcript of Human NLRP3 inflammasome senses multiple types of ... · PDF fileHuman NLRP3 inflammasome...

Human NLRP3 inflammasome senses multiple types ofbacterial RNAsWenwen Shaa,b, Hiroki Mitomab,c, Shino Hanabuchib,d, Musheng Baob,d, Leiyun Wengb, Naoshi Sugimotob,e, Ying Liub,f,Zhiqiang Zhanga,b,g, Jin Zhonga, Bing Suna,1, and Yong-Jun Liua,b,d,1
aKey Laboratory of Molecular Virology and Immunology, Institute Pasteur of Shanghai, Chinese Academy of Sciences, Shanghai 200031, China; bBaylorInstitute for Immunology Research, Baylor Scott & White Health Service, Dallas, TX 75204; cDepartment of Clinical Immunology and Rheumatology/InfectiousDisease, Kyushu University Hospital, 3-1-1 Maidashi, Higashi-ku, Fukuoka, 812-8582, Japan; dMedImmune, Gaithersburg, MD 20878; eDepartment ofHematology and Oncology, Graduate School of Medicine, Kyoto University, Kyoto 606-8507, Japan; fThe Key Laboratory of Infection and Immunity, Instituteof Biophysics, Chinese Academy of Sciences, Beijing 100101, China; and gImmunobiology and Transplant Research, Houston Methodist Hospital and HoustonMethodist Research Institute, Texas Medical Center, Houston, TX 77030
Edited by Tak W. Mak, The Campbell Family Institute for Breast Cancer Research at Princess Margaret Cancer Centre, Ontario Cancer Institute, UniversityHealth Network, Toronto, ON, Canada, and approved October 10, 2014 (received for review July 3, 2014)
Inflammasomes are multiprotein platforms that activate caspase-1,which leads to the processing and secretion of the proinflammatorycytokines IL-1β and IL-18. Previous studies demonstrated that bacte-rial RNAs activate the nucleotide-binding domain, leucine-rich-repeat-containing family, pyrin domain-containing 3 (NLRP3) inflammasomein both human and murine macrophages. Interestingly, only mRNA,but neither tRNA nor rRNAs, derived from bacteria could activate themurine Nlrp3 inflammasome. Here, we report that all three types ofbacterially derived RNA (mRNA, tRNA, and rRNAs) were capable ofactivating the NLRP3 inflammasome in human macrophages. Bacte-rial RNA’s 5′-end triphosphate moieties, secondary structure, anddouble-stranded structure were dispensable; small fragments of bac-terial RNA were sufficient to activate the inflammasome. In addition,we also found that 20-guanosine ssRNA can activate the NLRP3inflammasome in human macrophages but not in murine macro-phages. Therefore, human and murine macrophages may haveevolved to recognize bacterial cytosolic RNA differently duringbacterial infections.
bacterial RNA | single-stranded RNA | NLRP3 inflammasome |innate immunity | primary macrophages
The innate immune system is the first line of defense againstmicrobial infections. Germ-line–encoded pattern-recognition
receptors (PRRs) of the innate immune system recognize thepresence of invariant evolutionarily conserved microbial com-ponents called “pathogen-associated molecular patterns” (1–3).In response to microbial infections, PRRs rapidly initiate signal-transduction pathways to induce type 1 IFN production, proin-flammatory cytokine production, and inflammasome activation.The inflammasome is a cytosolic large caspase-1–containing multi-protein complex that enables autocatalytic activation of caspase-1.Once caspase-1 is activated, it starts to cleave prointerleukin-1β(pro–IL-1β) and prointerleukin-18 (pro–IL-18) proteolytically intobioactive IL-1β and IL-18 (4–7). The mature forms of IL-1β and IL-18 play roles in a variety of infectious and inflammatory processes.Cytosolic microbial nucleic acids are important activators of
the innate immune system against both bacterial and viralinfections, which induce type 1-IFN and proinflammatory cyto-kine responses as well as inflammasome activation. The role ofmicrobial nucleic acids in inflammasome activation has been stud-ied mostly in murine bone marrow-derived dendritic cells (BMDCs)or bone marrow-derived macrophages (BMDMs). AIM2 has beenidentified as a specific cytosolic dsDNA sensor that directlybinds ASC (apoptosis-associated speck-like protein containinga carboxyl-terminal CARD-like domain) and forms inflamma-some complexes in human and murine cells (8–11).Viral dsRNA was found to activate the nucleotide-binding
domain, leucine-rich-repeat-containing family, pyrin domain-containing 3 (NLRP3) inflammasome in human and murine cells(12–15). Several groups have reported that cytosolic bacterialRNA activate the Nlrp3 inflammasome in murine macrophages
(13, 16, 17). Our group also has reported that human THP-1–derived macrophages recognize cytosolic bacterial RNA andinduce NLRP3 inflammasome activation (12). Bacterial RNA iscomposed of mRNA, tRNA, and three different sizes of rRNA(23s, 16s, and 5s). Sander et al. (18) reported that, of the differenttypes of Escherichia coli RNA, only E. coli mRNA induced thesecretion of IL-1β by murine BMDMs, but E. coli tRNA and E.coli rRNAs did not.We aimed to study (i) whether a variety of cytosolic bacterial
RNAs could activate the inflammasome in human myeloid cellsand (ii) what types of bacterial RNA activate the inflammasomein human and murine myeloid cells. Here, we demonstrate thata broad spectrum of cytosolic bacterial RNAs strongly induce thecleavage of caspase-1 and the secretion of IL-1β and IL-18 inhuman macrophages. Human macrophages can sense mRNA,tRNA, rRNAs, and small synthetic ssRNA through NLRP3, butmurine macrophages can sense only the mRNA component.Bacterial RNA’s 5′-end triphosphate moieties, secondary struc-ture, and double-stranded structure were dispensable, but smallfragments of bacterial RNA were sufficient to activate theinflammasome. These findings suggest that upon bacterial infec-tions the human and murine NLRP3 inflammasomes sense cyto-solic bacterial RNAs differently.
Significance
The innate immune system has evolved to protect the hostfrom potential pathogens. The nucleotide-binding domain,leucine-rich-repeat-containing family, pyrin domain-containing3 (NLRP3) inflammasome is one of the platforms that can sensepathogenic bacteria, which can cause fatal bacterial infections.Here we show that all three types of bacteria-derived RNA—mRNA, tRNA, and rRNAs—as well as synthetic 20-guanosinessRNA, are capable of activating the NLRP3 inflammasome andinducing human macrophages to secrete inflammatory cyto-kines. Interestingly, only bacterial mRNA is able to activate themurine Nlrp3 inflammasome. Therefore human macrophagesmay have evolved in a unique fashion, adapting to the bacte-rial environment. This study could provide important clues fordeveloping efficient medicines for bacterial infections andvarious immunotherapies such as anticancer vaccines.
Author contributions: W.S., H.M., J.Z., B.S., and Y.-J.L. designed research; W.S., H.M., M.B.,and L.W. performed research; Y.L. contributed new reagents/analytic tools; W.S., H.M., andY.-J.L. analyzed data; and W.S., H.M., S.H., N.S., Z.Z., and Y.-J.L. wrote the paper.
The authors declare no conflict of interest.
This article is a PNAS Direct Submission.1To whom correspondence may be addressed. Email: [email protected] or [email protected].
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1412487111/-/DCSupplemental.
www.pnas.org/cgi/doi/10.1073/pnas.1412487111 PNAS | November 11, 2014 | vol. 111 | no. 45 | 16059–16064
IMMUNOLO
GYAND
INFLAMMATION

ResultsCytosolic Gram-Negative and Gram-Positive Bacterial RNAs InduceInflammasome Activation in Human Macrophages. To investigatewhether bacterial RNAs induce inflammasome activation inhuman macrophages, we purified RNA from various kinds ofGram-negative and Gram-positive bacteria, including bacteriathat can cause human infectious diseases. We first stimulatedprimary human monocyte-derived macrophages (MDMs) withthese bacterial RNAs with or without Lipofectamine 2000 (L2K).The phenotype of primary human macrophages was tested byhuman macrophage markers CD14 and CD33 by flow cytometry(Fig. S1A), and the morphology of the human MDMs was ob-served under an electron microscope. We found that all types ofbacterial RNAs that were transfected with L2K led to the se-cretion of IL-1β and IL-18 (Fig. 1A) in human MDMs. On theother hand, bacterial RNAs without L2K did not induce secretionof IL-1β or IL-18 (Fig. 1A), indicating that bacterial RNAs arerecognized in the cytosol. As in human primary macrophages,all tested bacterial total RNA induced inflammasome activationin phorbol 12-myristate 13-acetate (PMA)-differentiated humanmonocytic THP-1 cells (THP-1–derived macrophages) (Fig.S2A). Secretion of IL-1β and IL-18 induced by L2K-transfectedbacterial RNAs was abolished almost completely in primary hu-man MDMs (Fig. 1B) and THP-1–derived macrophages (Fig.S2B) by treatment with an RNase mixture that cleaves the 3′ endof ssRNA, indicating that inflammasome activation is indeedmediated by RNA but not by potential contamination of otherbacterial components. Because E. coli total RNA is the best-studied bacterial RNA in murine macrophages, we decided tofocus on E. coli total RNA in human macrophages for furtheranalysis. E. coli total RNA with L2K induced IL-1β and IL-18secretion (Fig. 2A) and cleavage of IL-1β (p17) and caspase-1(Casp1) (p20 and p22) (Fig. 2B) in THP-1–derived macrophagesin a dose-dependent manner. To exclude the contamination ofDNA further, we treated E. coli total RNA with DNase I orRNase I. DNase I treatment did not affect the ability of E. colitotal RNA to induce inflammasome activation (Fig. 2 C and D).
These results clearly indicate that cytosolic bacterial RNAs in-duce inflammasome activation in human macrophages.
All Bacterial RNA Components (mRNA, tRNA, and rRNAs) InduceInflammasome Activation in Human Macrophages. A previous studyshowed that, of the various E. coli components, only mRNAcould activate the inflammasome in murine BMDMs (18). Todetermine which E. coli RNA components promote inflamma-some activation in human macrophages, we generated E. colimRNA, tRNA, 23s rRNA, 16s rRNA, and 5s rRNA by in vitrotranscription (Fig. 3A). The secondary structures for rRNAs andtRNA were predicted by RNAfold WebServer (Fig. S3), and thepurities were confirmed by Agilent Bioanalyzer (Fig. S4A). Westimulated murine BMDMs and human MDMs with these in vitro-transcribed RNAs and examined the secretion of IL-1β and IL-18and the cleavage of caspase-1. The phenotype of murine BMDMswas tested by F4/80 and CD11b (Fig. S1B). In murine BMDMs, weconfirmed that only mRNA induced secretion of IL-1β and IL-18(Fig. 3B) and cleavage of caspase-1 (p10) (Fig. 3C). Similar resultswere observed in the murine macrophage cell line Raw264.7 (Fig.S5). On the other hand, not only mRNA but also tRNA, 23s rRNA,16s rRNA, and 5s rRNA led to the secretion of IL-1β and IL-18 andthe cleavage of caspase-1 both in primary human MDMs (Fig. 3 Dand E) and in THP-1–derived macrophages (Fig. 3 F and G). Incontrast to in vitro-transcribed bacterial RNAs, a synthetic Toll-likereceptor 8 (TLR8) ligand, poly U, did not induce inflammasomeactivation (Fig. 3 F and G), a finding that is similar to a previousreport (19). RNase treatment of in vitro-transcribed bacterial RNAscompletely abolished IL-1β and IL-18 secretion (Fig. 3F), indicatingthat inflammasome activation is indeed mediated by these RNAs.To confirm bacterial RNA recognition in human cells further,
we isolated native tRNA, 23s rRNA, 16s rRNA, and 5s rRNAfrom E. coli total RNA via denaturing RNA gel (Fig. 4 A and B).We then stimulated THP-1–derived macrophages with individualnative E. coli RNA or poly U and examined the secretion ofIL-1β and IL-18 and the cleavage of IL-1β and caspase-1. All the
MockE. c P.
aC.v S.e M.c
S. aL. m E. f
0
1000
2000
3000
with L2K without L2KGram- Gram+
IL-1
8(p
g/m
l)
MockE. c P.
aC.v S.e M.c
S. aL. m E. f
0500
1000150020002500
_ RNaseGram- Gram+
IL-1
(pg/
ml)
MockE. c P.
aC.v S.e M.c
S. aL. m E. f
0500
1000150020002500
_ RNase
Gram- Gram+
IL-1
8(p
g/m
l)
MockE. c P.
aC.v S.e M.c
S. aL. m E. f
0
1000
2000
3000
with L2K without L2KGram- Gram+
IL-1
(pg/
m
A
B
Fig. 1. Cytosolic Gram-negative and Gram-positive bacterial RNAs induceinflammasome activation in primary human MDMs. (A) Human MDMs werestimulated with purified bacterial total RNAs isolated from Gram-negativebacteria including E. coli (E.c), Pseudomonas aeruginosa (P.a), Chromo-bacterium violaceum (C.v), Salmonella enterica (S.e), and Moraxella catar-rhalis (M.c) and from Gram-positive bacteria including Staphylococcus aureus(S.a), Listeria monocytogenes (L.m), and Enterococcus faecalis (E.f), with orwithout L2K. (B) Human MDMs were stimulated with bacterial total RNAspretreated with or without the RNase mixture. IL-1β and IL-18 production inthe culture supernatants was measured by ELISA. Values are means ± SD ofat least three independent experiments.
IL-1β p170 0.10.2 0.5 1 2 5 10 (μg/ml)
BA4000
6000
(pg/
ml)
Cas-1
p22
p20p22
IL 1βSN
Cas 1IL-1β
p
pro0 1 2 5 1 2 5 10
0
2000
IL-1
β
p20p
Cas-1LY
IL-18
Cas-1
propro
400
600
g/m
l)
0 0.1 0.2 0.5 1 2 5 10μg/ml
Αctin
0
200
400
IL-1
8 (p
g
E.c - + + + DNase I - - - + RNase I
DC0 0.1 0.2 0.5 1 2 5 10
0
μg/ml
p17
p20
RNase I - - + -IL-1β
SN Cas-1 p221000
1500
2000
(pg/
ml)
1000
1500
2000
(pg/
ml)
p
Cas-1LY
IL-1β
p20p22Cas-1pro
pro
k I0
500
1000
IL-1
β
k I0
500
1000
IL-1
8
proCas 1
Αctin
IL-18pro
Mock —
RNase I
DNase I
Mock —
RNase I
DNase I
Fig. 2. Purified E. coli total RNA induces inflammasome activation in a dose-dependent manner. (A–D) Human THP-1–derived macrophages were stim-ulated with different concentrations of E. coli total RNA plus L2K (A and B)or stimulated with medium (Mock), E. coli total RNA pretreated with DNase Ior RNase I plus L2K or not pretreated (–) (C and D). Culture supernatants (SN)were analyzed for IL-1β and IL-18 by ELISA (A and C) and for cleavage of IL-1β(p17) and caspase-1 (Casp1) (p20 and p22) by immunoblotting (B and D). Celllysates (LY) were analyzed for pro–IL-1β, cleaved caspase-1 (p20 and p22),procaspase-1, pro–IL-18 and β-actin (Actin) by immunoblotting (B and D).Values are means ± SD of at least three independent experiments.
16060 | www.pnas.org/cgi/doi/10.1073/pnas.1412487111 Sha et al.

native bacterial RNAs tested induced IL-1β and IL-18 secretion(Fig. 4C) and cleavage of IL-1β and caspase-1 (Fig. 4D), but polyU did not. Therefore we conclude that human macrophagesrecognize all the individual bacterial RNA components.
Small Fragments of Bacterial RNA Are Sufficient for InflammasomeActivation in Human THP-1–Derived Macrophages. Virus-derivedRNA bearing a 5′-triphosphate group (5′-pppRNA) is reportedto induce retinoic acid-inducible gene-1 (RIG-1)–mediatedinflammasome activation in murine BMDCs and human PBMCs
(20). Because bacterial RNAs also have 5′-end triphosphatemoieties, we next tested whether 5′ triphosphate moieties playa role in inflammasome activation in human macrophages. Westimulated THP-1–derived macrophages with E. coli total RNApretreated with calf intestinal alkaline phosphatase (CIP), whichremoves the 5′-end triphosphate moieties. CIP treatment did notaffect E. coli RNA-induced IL-1β and IL-18 secretion (Fig. 5A),indicating that 5′-end triphosphate moieties are not required forbacterial RNA-induced inflammasome activation. We nexttested the length of RNA required for the inflammasome acti-vation. RNA fragmentation reagents were used to fragmentRNA into 60–200 nucleotides. E. coli total RNA-induced IL-1βand IL-18 secretion was not altered by treatment with RNAfragmentation reagents (Fig. 5A). RNase III digests dsRNA intosmall 12- to 15-base fragments. E. coli mRNA, tRNA, andrRNAs form short double-stranded stems and were digested intosmall fragments by treatment with RNase III (Fig. 5B). In con-trast to RNase A or RNase I treatment, the treatment withRNase III did not affect secretion of IL-1β and IL-18 or cleavageof IL-1β and caspase-1 (Fig. 5 C and D), suggesting that thedouble-stranded structure of bacterial RNA is not required.RNA poly I:C is well known to induce NLRP3 inflammasomeactivation in both human and murine macrophages. To in-vestigate further whether dsRNA structures are necessary for theinflammasome activation that is induced by bacterial RNA, weheat-denatured E. coli total RNA and the double-stranded RNAanalog poly I:C to disrupt the double-stranded structure. Westimulated THP-1–derived macrophages with poly I:C or E. colitotal RNA with or without heat denaturation. As previouslyreported, poly I:C-induced IL-1β and IL-18 secretion was abrogatedcompletely by heat denaturation of RNA in THP-1–derived mac-rophages (Fig. 5E). In contrast to poly I:C, heat-denatured E. colitotal RNA led to similar amounts of IL-1β and IL-18 secretioncompared with untreated RNA in THP-1–derived macrophages(Fig. 5E). In addition, 5s short rRNA (5s short) consisting of 1–47bases, which does not contain complementary sequences to forma double-stranded stem, also induces secretion of IL-1β and IL-18 inTHP-1–derived macrophages (Fig. 5F). These results suggest that5′-end triphosphate moieties, the full-length intact secondarystructure, and the double-stranded structure of bacterial RNA are
Fig. 3. In vitro-transcribed RNA components can induce inflammasomeactivation in human macrophages but not in murine macrophages. (A) E.colitotal RNA and in vitro-transcribed RNAs [23s rRNA (23s), 16s rRNA (16s), 5srRNA (5s), mRNA, and tRNA] were separated by agarose gel electrophoresis.(B and C) Murine BMDMs were primed with 100 ng/mL LPS for 3 h and thenwere stimulated with E. coli total RNA or in vitro-transcribed RNAs plus L2Kfor 16 h. Culture supernatants were analyzed for IL-1β and IL-18 by ELISA (B)and for cleavage of caspase-1 (p10) by immunoblotting (C). Cell lysates wereanalyzed for procaspase-1 and β-actin (C). NS, nonspecific band. (D and E)Human MDMs were stimulated with E. coli total RNA or in vitro-transcribedRNAs plus L2K. Cell supernatants were analyzed for IL-1β and IL-18 by ELISA(D) and for cleavage of IL-1β and caspase-1 (p20) by immunoblotting (E). Celllysates were analyzed for pro–IL-1β, procaspase-1, pro–IL-18, and β-actin (E).(F and G) THP-1–derived macrophages were stimulated with E. coli totalRNA, in vitro-transcribed RNAs, or poly U plus L2K, which were pretreatedwith RNase mixture or were not pretreated. IL-1β and IL-18 production in theculture supernatant was measured by ELISA (F). Cell supernatants stimulatedwith E. coli total RNA, or in vitro-transcribed RNAs without RNase mixture pre-treatment were analyzed for cleavage of IL-1β and caspase-1 by immunoblotting(G). Cell lysates were analyzed for pro–IL-1β, cleaved caspase-1, procaspase-1,pro–IL-18, and β-actin by immunoblotting (G). Values are means ± SD of at leastthree independent experiments.
Fig. 4. Inflammasome activation induced by native E. coli tRNA and rRNAsis similar to that induced by in vitro-transcribed RNAs in human THP-1–derived macrophages. (A and B) Components of native E. coli total RNA, 16srRNA (n16s), and 23s rRNA (n23s) were identified by agarose gel electro-phoresis (A). Components of native E. coli total RNA, tRNA (ntRNA), and 5srRNA (n5s) were identified by PAGE (B). (C and D) THP-1–derived macro-phages were stimulated with E. coli total RNA or ntRNA, n23S, n16s, n5s, andpoly U, plus L2K. Culture supernatants were analyzed for IL-1β and IL-18 byELISA (C) and for cleavage of IL-1β and caspase-1 by immunoblotting (D). Celllysates were analyzed for pro–IL-1β, procaspase-1, pro–IL-18, and β-actin (D).Values are means ± SD of at least three independent experiments.
Sha et al. PNAS | November 11, 2014 | vol. 111 | no. 45 | 16061
IMMUNOLO
GYAND
INFLAMMATION

not required for inflammasome activation in human THP-1–derivedmacrophages.
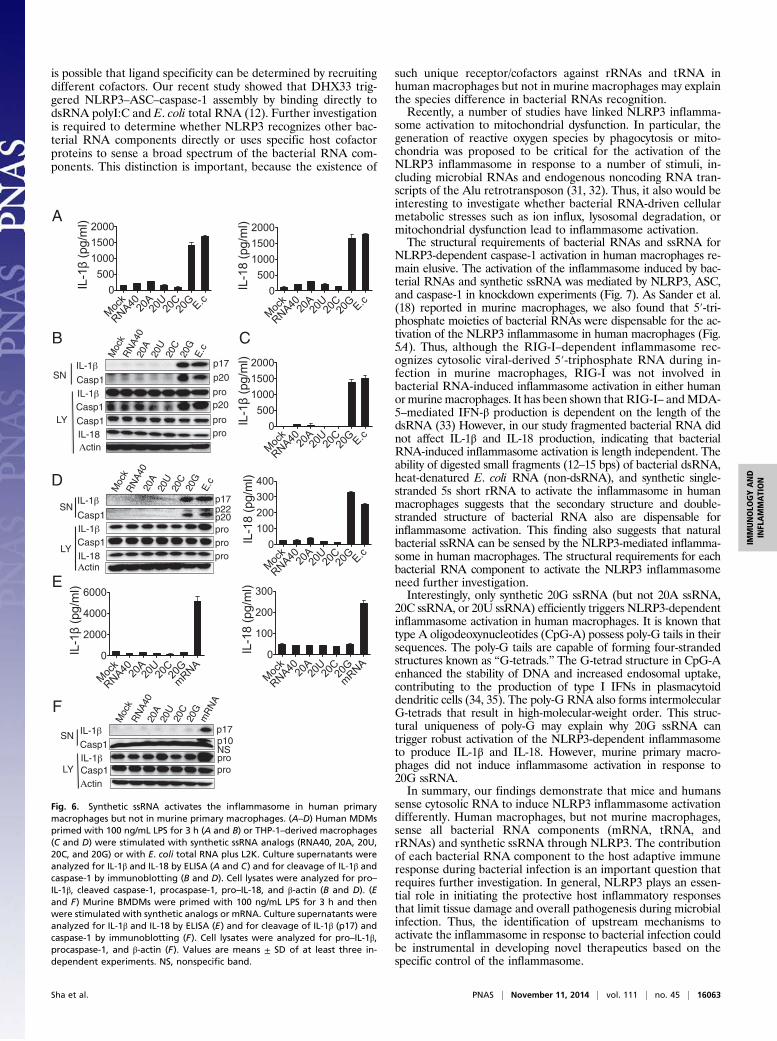
Synthetic ssRNA Activates the Inflammasome in Human PrimaryMacrophages but Not in Murine Primary Macrophages. Allen et al.(15) reported that ssRNA40, also known as “R-1075” (a U-richssRNA derived from the HIV-1 LTR), is sufficient to induce IL-1β secretion in human macrophages. We synthesized ssRNA40(RNA40), 20-adenosine-ssRNA (20A), 20-uridine ssRNA (20U),20-cytidine ssRNA (20C), and 20-guanosine ssRNA (20G) andthen stimulated human MDMs and THP-1–derived macrophageswith these synthetic ssRNAs. The purities of these ssRNAs wereshown in Fig. S4B. We found that 20G led to the highest amountof secretion of IL-1β and IL-18 (Fig. 6 A and C). It also was themost effective ssRNA at cleaving IL-1β and caspase-1 in bothhuman MDMs (Fig. 6B) and THP-1–derived macrophages (Fig.6D). In contrast, 20G failed to induce IL-1β and IL-18 secretion(Fig. 6E) and cleavage of IL-1β and caspase-1 (Fig. 6F) in murineBMDMs. In our system, ssRNA40 did not induce inflammasomeactivation in human MDMs or murine BMDMs (Fig. 6). Theability of 20G to activate the human but not the murine inflam-masome further demonstrates the differences between thesetwo systems.
NLRP3, ASC, and Caspase-1 Are Required for Cytosolic Bacterial RNA-Induced Inflammasome Activation in Human Macrophages. We in-vestigated which inflammasome was activated by individualbacterial RNA components or synthetic 20G in THP-1–derivedmacrophages. Our group and others (12, 13, 21) have reportedthat NLPR3 mediates dsRNA- or total bacterial RNA-inducedinflammasome activation. We developed specific human NLRP3-targeting, ASC-targeting, or caspase-1–targeting shRNA constructs(12). These shRNAs for NLRP3, ASC, or caspase-1 specificallydown-regulated the expression levels of the targeted proteinswithout affecting the expression of other proteins in THP-1–derivedmacrophages (Fig. 7A). Knockdown of NLRP3, ASC, or caspase-1in THP-1–derived macrophages predominantly abolished IL-1β inthe supernatant (Fig. 7B) and completely blocked the secretion ofIL-18 (Fig. S6), which are induced by E. coli total RNA, in vitro-transcribed individual E. coli RNA components and synthetic 20GssRNA. This targeting also blocked the cleavage of IL-1β and cas-pase-1 (Fig. 7C), which are induced by E. coli total RNA. These
observations suggest that the NLRP3/ASC/caspase-1 signalingpathway is involved in small ssRNA-induced inflammasome acti-vation. Therefore, the NLRP3/ASC/caspase-1 inflammasome iscritical for the activation of the cytosolic bacterial RNA-inducedinflammasome in human macrophages.
DiscussionRecent studies have shown that human and murine innate im-mune cells respond differently to microbial components. MurineToll-like receptor 7 (TLR7) and Toll-like receptor 9 (TLR9)favor sequence-specific motifs that are distinct from those effi-ciently recognized by human TLR7 and TLR9 (22, 23). TLR8,which recognizes GU-rich ssRNA in humans, is nonfunctional inmice (24). Murine myeloid cells, but not human cells, expressTLR13 and recognize bacterial 23s rRNA in the endosome toactivate the NF-k B pathway (25–28) Moreover, cytosolic LPSactivates the caspase-11–mediated inflammasome in murinemacrophages (29). However, human cells do not have the cas-pase-11 gene and cannot respond to cytosolic LPS (30). Theseresults suggest that humans and mice have undergone majorevolutionary divergence in sensing microbes. In this study, weshow that human macrophages sense all bacterial RNA com-ponents (mRNA, tRNA, and rRNAs) and synthetic ssRNA toactivate the NLRP3 inflammasome, whereas murine cells pref-erentially recognize bacterial mRNA.It is not clear how murine cells selectively recognize mRNA
among the bacterial RNA components and why human macro-phages can sense such a broad spectrum of cytosolic RNAs. Al-though tRNA and rRNAs are composed of ssRNA, they internallyform a partial double-stranded secondary structure. Interestingly,Sander et al. (18) showed that the absence of 3′ polyadenylation inbacterial mRNA is the signal that activates the inflammasome.Thus, murine and human macrophages may have different sensingmachinery in the NLRP3 inflammasome to distinguish a differencein structure or the presence or the absence of unique nucleicmodifications among the individual bacterial RNA components.The actual PRR that recognizes a broad spectrum of bacterial
RNAs to activate NLRP3 remains poorly defined. NLRP3responds to numerous stimuli including viral/bacterial DNA andRNA, pore-forming toxins, silica, asbestos, uric acid, ion flux,and ATP (31). Given such a broad range of NLRP3 activators,direct binding of all these activators to NLRP3 seems unlikely. It
31.5
0.5
M III A Ikb
B
Mock
Heat-D
Heat-D
0100200300400
E.cpIC
IL-1
8(p
g/m
l)
E
Mock
Heat-D
Heat-D
0
1000
2000
3000 E.cpIC
IL-1
(pg/
ml)
C
Mock III A I
0
500
1000
1500
IL-1
8(p
g/m
l)
Mock III A I
0
1000
2000
3000
IL-1
(pg/
ml)
A
Mock
Frag0
500
1000
1500
IL-1
8(p
g/m
l)
Mock
CIP0
200400600800
IL-1
8(p
g/m
l)
Mock
CIP0
1000
2000
3000
IL-1
(pg/
ml)
Mock
Frag0
1000200030004000
IL-1
(pg/
ml)
p22p20
p22p20
p17
pro
IL-1
Casp1
IL-1
IL-18ctin
LY
Casp1
Casp1
Mock _ III IA
pro
pro
DSN
F
Mock
E.c 5s
5s sh
ort0
500
1000
1500
IL-1
(pg/
ml)
Mock
E.c 5s
5s sh
ort0
200400600800
IL-1
8(p
g/m
l)
Fig. 5. Small bacterial RNA fragments are sufficientfor inflammasome activation in human THP-1–derivedmacrophages. (A) THP-1–derived macrophageswere stimulated with E. coli total RNA pretreatedwith or without CIP or RNA fragmentation reagent(Frag). IL-1β and IL-18 production in the supernatantswas analyzed by ELISA. (B–D) THP-1–derived macro-phages were stimulated with E. coli total RNA pre-treated with RNase III (III), RNase A (A), RNase I (I), orwithout any RNase (–). RNase digestion was visualizedby agarose gel separation (B). Culture supernatantswere analyzed for IL-1β and IL-18 by ELISA (C) and forcleavage of IL-1β and caspase-1 by immunoblotting(D). Cell lysates were analyzed for pro–IL-1β, cleavedcaspase-1, procaspase-1, pro–IL-18, and β-actin (D).(E and F) THP-1–derived macrophages were stimu-lated with poly I:C (pIC) or with E. coli total RNA plusL2K with or without heat denaturation (Heart-D) (E)or with E. coli total RNA, 5s, or 5s short rRNA (5sshort) plus L2K (F). Culture supernatants were ana-lyzed for IL-1β and IL-18 by ELISA. Values are means ±SD of at least three independent experiments.
16062 | www.pnas.org/cgi/doi/10.1073/pnas.1412487111 Sha et al.
is possible that ligand specificity can be determined by recruitingdifferent cofactors. Our recent study showed that DHX33 trig-gered NLRP3–ASC–caspase-1 assembly by binding directly todsRNA polyI:C and E. coli total RNA (12). Further investigationis required to determine whether NLRP3 recognizes other bac-terial RNA components directly or uses specific host cofactorproteins to sense a broad spectrum of the bacterial RNA com-ponents. This distinction is important, because the existence of
such unique receptor/cofactors against rRNAs and tRNA inhuman macrophages but not in murine macrophages may explainthe species difference in bacterial RNAs recognition.Recently, a number of studies have linked NLRP3 inflamma-
some activation to mitochondrial dysfunction. In particular, thegeneration of reactive oxygen species by phagocytosis or mito-chondria was proposed to be critical for the activation of theNLRP3 inflammasome in response to a number of stimuli, in-cluding microbial RNAs and endogenous noncoding RNA tran-scripts of the Alu retrotransposon (31, 32). Thus, it also would beinteresting to investigate whether bacterial RNA-driven cellularmetabolic stresses such as ion influx, lysosomal degradation, ormitochondrial dysfunction lead to inflammasome activation.The structural requirements of bacterial RNAs and ssRNA for
NLRP3-dependent caspase-1 activation in human macrophages re-main elusive. The activation of the inflammasome induced by bac-terial RNAs and synthetic ssRNA was mediated by NLRP3, ASC,and caspase-1 in knockdown experiments (Fig. 7). As Sander et al.(18) reported in murine macrophages, we also found that 5′-tri-phosphate moieties of bacterial RNAs were dispensable for the ac-tivation of the NLRP3 inflammasome in human macrophages (Fig.5A). Thus, although the RIG-I–dependent inflammasome rec-ognizes cytosolic viral-derived 5′-triphosphate RNA during in-fection in murine macrophages, RIG-I was not involved inbacterial RNA-induced inflammasome activation in either humanormurinemacrophages. It has been shown that RIG-I– andMDA-5–mediated IFN-β production is dependent on the length of thedsRNA (33) However, in our study fragmented bacterial RNA didnot affect IL-1β and IL-18 production, indicating that bacterialRNA-induced inflammasome activation is length independent. Theability of digested small fragments (12–15 bps) of bacterial dsRNA,heat-denatured E. coli RNA (non-dsRNA), and synthetic single-stranded 5s short rRNA to activate the inflammasome in humanmacrophages suggests that the secondary structure and double-stranded structure of bacterial RNA also are dispensable forinflammasome activation. This finding also suggests that naturalbacterial ssRNA can be sensed by the NLRP3-mediated inflamma-some in human macrophages. The structural requirements for eachbacterial RNA component to activate the NLRP3 inflammasomeneed further investigation.Interestingly, only synthetic 20G ssRNA (but not 20A ssRNA,
20C ssRNA, or 20U ssRNA) efficiently triggers NLRP3-dependentinflammasome activation in human macrophages. It is known thattype A oligodeoxynucleotides (CpG-A) possess poly-G tails in theirsequences. The poly-G tails are capable of forming four-strandedstructures known as “G-tetrads.” The G-tetrad structure in CpG-Aenhanced the stability of DNA and increased endosomal uptake,contributing to the production of type I IFNs in plasmacytoiddendritic cells (34, 35). The poly-G RNA also forms intermolecularG-tetrads that result in high-molecular-weight order. This struc-tural uniqueness of poly-G may explain why 20G ssRNA cantrigger robust activation of the NLRP3-dependent inflammasometo produce IL-1β and IL-18. However, murine primary macro-phages did not induce inflammasome activation in response to20G ssRNA.In summary, our findings demonstrate that mice and humans
sense cytosolic RNA to induce NLRP3 inflammasome activationdifferently. Human macrophages, but not murine macrophages,sense all bacterial RNA components (mRNA, tRNA, andrRNAs) and synthetic ssRNA through NLRP3. The contributionof each bacterial RNA component to the host adaptive immuneresponse during bacterial infection is an important question thatrequires further investigation. In general, NLRP3 plays an essen-tial role in initiating the protective host inflammatory responsesthat limit tissue damage and overall pathogenesis during microbialinfection. Thus, the identification of upstream mechanisms toactivate the inflammasome in response to bacterial infection couldbe instrumental in developing novel therapeutics based on thespecific control of the inflammasome.
Casp1
Casp1
IL-1
IL-1LY
SN
ctin
p17p10
proNS
pro
F
A
ctin
Casp1
p20 IL-1
LY
SN
IL-1
IL-18
Casp1
Casp1
p17
pro
p20
pro
pro
B
Mock
RNA4020A20
U20
C20
GmRNA
0
100
200
300
IL-1
8(p
g/m
l)
Mock
RNA4020A20
U20
C20
GmRNA
0
2000
4000
6000
IL-1
(pg/
ml)
C
Mock
RNA4020A20
U20
C20
G E.c0
500100015002000
IL-1
(pg/
ml)
Mock
RNA4020A20
U20
C20
G E.c0
500100015002000
IL-1
8(p
g/m
l)
ctin
Casp1p22
IL-1
LY
SN
IL-1
IL-18
p17
Casp1 pro
p20
pro
pro
Mock
RNA4020A20
U20
C20
G E.c0
500100015002000
IL-1
(pg/
ml)
Mock
RNA4020A20
U20
C20
G E.c0
100200300400
IL-1
8(p
g/m
l)
E
D
Fig. 6. Synthetic ssRNA activates the inflammasome in human primarymacrophages but not in murine primary macrophages. (A–D) Human MDMsprimed with 100 ng/mL LPS for 3 h (A and B) or THP-1–derived macrophages(C and D) were stimulated with synthetic ssRNA analogs (RNA40, 20A, 20U,20C, and 20G) or with E. coli total RNA plus L2K. Culture supernatants wereanalyzed for IL-1β and IL-18 by ELISA (A and C) and for cleavage of IL-1β andcaspase-1 by immunoblotting (B and D). Cell lysates were analyzed for pro–IL-1β, cleaved caspase-1, procaspase-1, pro–IL-18, and β-actin (B and D). (Eand F) Murine BMDMs were primed with 100 ng/mL LPS for 3 h and thenwere stimulated with synthetic analogs or mRNA. Culture supernatants wereanalyzed for IL-1β and IL-18 by ELISA (E) and for cleavage of IL-1β (p17) andcaspase-1 by immunoblotting (F). Cell lysates were analyzed for pro–IL-1β,procaspase-1, and β-actin (F). Values are means ± SD of at least three in-dependent experiments. NS, nonspecific band.
Sha et al. PNAS | November 11, 2014 | vol. 111 | no. 45 | 16063
IMMUNOLO
GYAND
INFLAMMATION

Materials and MethodsAll experiments with mice were approved by Baylor Institute for Immunol-ogy Research Animal Care and Use Committee and were consistent withlocal, state, and federal guidelines. All mice were bred in a sterile environ-ment. Details of summary of reagents, research methods and statisticalanalysis are available in SI Materials and Methods.
ACKNOWLEDGMENTS. We thank Dr. C. Harrod (Baylor Institute for Immu-nology Research) for critical reading of the manuscript and all the membersof the Y.-J.L. laboratory for helpful discussions. This study was supported bythe Chinese Academy of Sciences/State Administration of Foreign ExpertsAffairs International Partnership Program for Creative Research Teams, byNational 973 Key Project (2013CB530504), and by National Institutes of HealthGrant R37 AI091947.
1. Janeway CA, Jr, Medzhitov R (2002) Innate immune recognition. Annu Rev Immunol20(1):197–216.
2. Chen GY, Nuñez G (2010) Sterile inflammation: Sensing and reacting to damage. NatRev Immunol 10(12):826–837.
3. Kumar H, Kawai T, Akira S (2011) Pathogen recognition by the innate immune system.Int Rev Immunol 30(1):16–34.
4. Thornberry NA, et al. (1992) A novel heterodimeric cysteine protease is required forinterleukin-1 beta processing in monocytes. Nature 356(6372):768–774.
5. Ayala JM, et al. (1994) IL-lbeta-converting enzyme is present in monocytic cells as anInactive 45-kDa precursor. J Immunol 153(6):2592–2599.
6. Dinarello CA (1998) Interleukin-1 beta, interleukin-18, and the interleukin-1 betaconverting enzyme. Ann N Y Acad Sci 856:1–11.
7. Franchi L, Eigenbrod T, Muñoz-Planillo R, Nuñez G (2009) The inflammasome: Acaspase-1-activation platform that regulates immune responses and disease patho-genesis. Nat Immunol 10(3):241–247.
8. Fernandes-Alnemri T, Yu J-W, Datta P, Wu J, Alnemri ES (2009) AIM2 activates theinflammasome and cell death in response to cytoplasmic DNA. Nature 458(7237):509–513.
9. Hornung V, et al. (2009) AIM2 recognizes cytosolic dsDNA and forms a caspase-1-activating inflammasome with ASC. Nature 458(7237):514–518.
10. Bürckstümmer T, et al. (2009) An orthogonal proteomic-genomic screen identifiesAIM2 as a cytoplasmic DNA sensor for the inflammasome. Nat Immunol 10(3):266–272.
11. Roberts TL, et al. (2009) HIN-200 proteins regulate caspase activation in response toforeign cytoplasmic DNA. Science 323(5917):1057–1060.
12. Mitoma H, et al. (2013) The DHX33 RNA helicase senses cytosolic RNA and activatesthe NLRP3 inflammasome. Immunity 39(1):123–135.
13. Kanneganti T-D, et al. (2006) Bacterial RNA and small antiviral compounds activatecaspase-1 through cryopyrin/Nalp3. Nature 440(7081):233–236.
14. Kanneganti TD, et al. (2006) Critical role for Cryopyrin/Nalp3 in activation of caspase-1in response to viral infection and double-stranded RNA. J Biol Chem 281(48):36560–36568.
15. Allen IC, et al. (2009) The NLRP3 inflammasome mediates in vivo innate immunity toinfluenza A virus through recognition of viral RNA. Immunity 30(4):556–565.
16. Brereton CF, et al. (2011) Escherichia coli heat-labile enterotoxin promotes protectiveTh17 responses against infection by driving innate IL-1 and IL-23 production.J Immunol 186(10):5896–5906.
17. Deshmukh SD, et al. (2011) Macrophages recognize streptococci through bacterialsingle-stranded RNA. EMBO Rep 12(1):71–76.
18. Sander LE, et al. (2011) Detection of prokaryotic mRNA signifies microbial viabilityand promotes immunity. Nature 474(7351):385–389.
19. Pétrilli V, et al. (2007) Activation of the NALP3 inflammasome is triggered by low
intracellular potassium concentration. Cell Death Differ 14(9):1583–1589.20. Poeck H, et al. (2010) Recognition of RNA virus by RIG-I results in activation of CARD9
and inflammasome signaling for interleukin 1 β production. Nat Immunol 11(1):
63–69.21. Rajan JV, Warren SE, Miao EA, Aderem A (2010) Activation of the NLRP3 in-
flammasome by intracellular poly I:C. FEBS Lett 584(22):4627–4632.22. Gantier MP, et al. (2008) TLR7 is involved in sequence-specific sensing of single-
stranded RNAs in human macrophages. J Immunol 180(4):2117–2124.23. Roberts TL, Sweet MJ, Hume DA, Stacey KJ (2005) Cutting edge: Species-specific TLR9-
mediated recognition of CpG and non-CpG phosphorothioate-modified oligonu-
cleotides. J Immunol 174(2):605–608.24. Heil F, et al. (2004) Species-specific recognition of single-stranded RNA via toll-like
receptor 7 and 8. Science 303(5663):1526–1529.25. Hochrein H, Kirschning CJ (2013) Bacteria evade immune recognition via TLR13 and
binding of their 23S rRNA by MLS antibiotics by the same mechanisms. OncoImmu-
nology 2(3):e23141.26. Hidmark A, von Saint Paul A, Dalpke AH (2012) Cutting edge: TLR13 is a receptor for
bacterial RNA. J Immunol 189(6):2717–2721.27. Oldenburg M, et al. (2012) TLR13 recognizes bacterial 23S rRNA devoid of erythro-
mycin resistance-forming modification. Science 337(6098):1111–1115.28. Li X-D, Chen ZJ (2012) Sequence specific detection of bacterial 23S ribosomal RNA by
TLR13. eLife 1(e00102):e00102.29. Hagar JA, Powell DA, Aachoui Y, Ernst RK, Miao EA (2013) Cytoplasmic LPS activates
caspase-11: Implications in TLR4-independent endotoxic shock. Science 341(6151):
1250–1253.30. Kayagaki N, et al. (2013) Noncanonical inflammasome activation by intracellular LPS
independent of TLR4. Science 341(6151):1246–1249.31. Bauernfeind F, Hornung V (2013) Of inflammasomes and pathogens—sensing of
microbes by the inflammasome. EMBO Mol Med 5(6):814–826.32. Tarallo V, et al. (2012) DICER1 loss and Alu RNA induce age-related macular de-
generation via the NLRP3 inflammasome and MyD88. Cell 149(4):847–859.33. Kato H, et al. (2008) Length-dependent recognition of double-stranded ribonucleic
acids by retinoic acid-inducible gene-I and melanoma differentiation-associated gene
5. J Exp Med 205(7):1601–1610.34. Guiducci C, et al. (2006) Properties regulating the nature of the plasmacytoid den-
dritic cell response to Toll-like receptor 9 activation. J Exp Med 203(8):1999–2008.35. Honda K, et al. (2005) Spatiotemporal regulation of MyD88-IRF-7 signalling for robust
type-I interferon induction. Nature 434(7036):1035–1040.
Casp1Actin
ASCNLRP3
IB
Casp1Casp1
ctin
Casp1
p20p22
LY
SN
IL-18
IL-1
p20p22
pro
Mock E.c
p17
IL-1
propro
A B
C
sh-S
cr
sh-N
LRP3
sh-A
SC
sh-C
asp1
sh-S
cr
sh-N
LRP3
sh-A
SC
sh-C
asp1
0200400600800
1000Mock E.c
IL-1
(pg/
ml)
sh-S
cr
sh-N
LRP3
sh-A
SC
sh-C
asp1
0100200300400500 20G
IL-1
(pg/
ml)
sh-S
cr
sh-N
LRP3
sh-A
SC
sh-C
asp1
0200400600800
100023s
IL-1
(pg/
ml)
sh-S
cr
sh-N
LRP3
sh-A
SC
sh-C
asp1
0200400600800
1000 16s
IL-1
(pg/
ml)
sh-S
cr
sh-N
LRP3
sh-A
SC
sh-C
asp1
0200400600800 5s
IL-1
(pg/
ml)
sh-S
cr
sh-N
LRP3
sh-A
SC
sh-C
asp1
0200400600800
1000mRNA
IL-1
(pg/
ml)
sh-S
cr
sh-N
LRP3
sh-A
SC
sh-C
asp1
0
500
1000
1500 tRNA
IL-1
(pg/
ml)
Fig. 7. NLRP3, ASC, and caspase-1 are required forcytosolic bacterial RNA-induced inflammasome ac-tivation in human macrophages. THP-1–derived macro-phages were targeted with scramble shRNA (sh-Scr)or shRNA specific for NLRP3 (sh-NLRP3), ASC (sh-ASC), or caspase-1 (sh-Casp1) and were stimulatedwith E. coli total RNA, 20G, or in vitro-transcribedbacterial RNAs plus L2K. (A) Knockdown efficienciesof NLRP3, ASC, or caspase-1 were tested by immu-noblotting. (B) Culture supernatants were analyzedfor IL-1β by ELISA. (C) Cell supernatants stimulatedwith E. coli total RNA or mock stimulated were an-alyzed by immunoblotting for cleavage of IL-1β andcaspase-1. Cell lysates stimulated with E. coli totalRNA or mock stimulated were analyzed for pro–IL-1β, cleaved caspase-1, procaspase-1, pro–IL-18, andβ-actin. Values are means ± SD of at least three in-dependent experiments.
16064 | www.pnas.org/cgi/doi/10.1073/pnas.1412487111 Sha et al.


















