
HeyL regulates the number of TrkC neurons in dorsal root ganglia
10
HeyL regulates the number of TrkC neurons in dorsal root ganglia Abhishek Mukhopadhyay ⁎, Jennifer Jarrett, Timothy Chlon, John A. Kessler Department of Neurology, Northwestern University's Feinberg School of Medicine, Chicago, IL 60611, USA abstract article info Article history: Received for publication 2 April 2009 Revised 6 July 2009 Accepted 10 July 2009 Available online 22 July 2009 Keywords: Dorsal root ganglion TrkC HeyL Hey1 Differentiation The basic-helix-loop-helix transcription factor HeyL is expressed at high levels by neural crest progenitor cells (NCPs) that give rise to neurons and glia in dorsal root ganglia (DRG). Since HeyL expression was observed in these NCPs during the period of neurogenesis, we generated HeyL null mutants to help examine the factor's role in ganglion neuronal specification. Homozygous null mutation of HeyL reduced the number of TrkC + neurons in DRG at birth including the subpopulation that expresses the ETS transcription factor ER81. Conversely, null mutation of the Hey paralog, Hey1, increased the number of TrkC + neurons. Null mutation of HeyL increased expression of the Hey paralogs Hey1 and Hey2, suggesting that HeyL normally inhibits their expression. Double null mutation of both Hey1 and HeyL rescued TrkC + neuron numbers to control levels. Thus, the balance between HeyL and Hey1 expression regulates the differentiation of a subpopulation of TrkC + neurons in the DRG. © 2009 Elsevier Inc. All rights reserved. Introduction The basic-helix-loop-helix (bHLH) transcription factors, hairy and Enhancer of split are transcriptional repressors in Drosophila that mediate some of the effects of the Notch signaling pathway on neural stem cell differentiation (Klambt et al., 1989; Lecourtois and Schweisguth, 1995). Related proteins in mammals include the Hes and Hey families of repressors (Akazawa et al., 1992; Leimeister et al., 1999; Nakagawa et al., 1999; Sasai et al., 1992; Thomas and Rathjen, 1992). The Hes family has seven members, Hes1–7, of which Hes1, Hes3, Hes5 and Hes6 are expressed in the developing nervous system (Akazawa et al., 1992; Sakagami et al., 1994; Sasai et al., 1992). In the mammalian central nervous system (CNS) Hes1 and Hes5 act as repressors of proneuronal genes like Mash1 thereby preventing differentiation (Akazawa et al., 1995; Castella et al., 1999; de la Pompa et al., 1997; Ishibashi et al., 1995, 1994; Jarriault et al., 1995; Nakamura et al., 2000). Hes3 has two isoforms, one of which has repressor activity similar to Hes1 while the other lacks repressor activity (Hirata et al., 2000, 2001). Hes6 represses Hes1 activity and conversely promotes neuronal differentiation (Bae et al., 2000; Gratton et al., 2003; Koyano-Nakagawa et al., 2000). Thus, different Hes proteins play contrasting roles in neural differentiation. Hey1, Hey2 and HeyL constitute the mammalian Hey (also known as CHF, Hesr, Herp, and Hrt) family that, like the Hes proteins, act downstream of the Notch signaling pathway (Iso et al., 2001; Leimeister et al., 1999; Nakagawa et al., 2000, 1999; Steidl et al., 2000). The Hey genes play an important role in the development of the cardiovascular system and loss of Hey function causes defects in vascular specification, septation and valve formation (Donovan et al., 2002; Fischer et al., 2004, 2007; Gessler et al., 2002; Kokubo et al., 2005, 2004, 2007; Sakata et al., 2002; Xin et al., 2007). In the developing brain, overexpression of Hey1 and Hey2 prevents neural precursors from differentiating during the neurogenic period and causes glial differentiation by antagonizing neuronal bHLH factors later in development (Sakamoto et al., 2003). HeyL overexpression in the developing retina increases rod differentiation whereas Hey2 increases gliogenesis (Satow et al., 2001). Hey proteins are expressed at high levels in the developing DRG and trigeminal ganglia (Leimeister et al., 1999, 2000; Nakagawa et al., 1999) but their roles in sensory ganglion development and neuronal specification are unknown. Sensory ganglion neurons are divided into three major subpopula- tions based on their expression of the neurotrophin receptors and the type of sensory information they convey—TrkA + which convey information from thermoreceptors and nociceptors, TrkB + that convey mechanosensory information and TrkC + that are propriocep- tive (Snider, 1994). Here, we report that mice carrying a targeted mutation in the HeyL gene have decreased numbers of large TrkC + neurons in the DRG. Moreover, HeyL in the DRG inhibits Hey1 expression and mice mutant for Hey1 alone have increased numbers of TrkC + neurons in the DRG suggesting that Hey1 and HeyL play antagonistic roles in TrkC + neuron differentiation. Materials and methods Generation and maintenance of mouse lines The targeting construct was generated from the BAC clone RP23- 25K8 (CHORI) containing the HeyL locus by recombineering as previously described (Liu et al., 2003). All vectors and bacterial strains used for creating the targeting vector were kind gifts from Dr. Developmental Biology 334 (2009) 142–151 ⁎ Corresponding author. Fax: +1 312 503 0872. E-mail address: [email protected] (A. Mukhopadhyay). 0012-1606/$ – see front matter © 2009 Elsevier Inc. All rights reserved. doi:10.1016/j.ydbio.2009.07.018 Contents lists available at ScienceDirect Developmental Biology journal homepage: www.elsevier.com/developmentalbiology
-
Upload
abhishek-mukhopadhyay -
Category
Documents
-
view
215 -
download
3
Transcript of HeyL regulates the number of TrkC neurons in dorsal root ganglia

Developmental Biology 334 (2009) 142–151
Contents lists available at ScienceDirect
Developmental Biology
j ourna l homepage: www.e lsev ie r.com/deve lopmenta lb io logy
HeyL regulates the number of TrkC neurons in dorsal root ganglia
Abhishek Mukhopadhyay ⁎, Jennifer Jarrett, Timothy Chlon, John A. KesslerDepartment of Neurology, Northwestern University's Feinberg School of Medicine, Chicago, IL 60611, USA
⁎ Corresponding author. Fax: +1 312 503 0872.E-mail address: [email protected] (A. M
0012-1606/$ – see front matter © 2009 Elsevier Inc. Aldoi:10.1016/j.ydbio.2009.07.018
a b s t r a c t
a r t i c l e i n f oArticle history:Received for publication 2 April 2009Revised 6 July 2009Accepted 10 July 2009Available online 22 July 2009
Keywords:Dorsal root ganglionTrkCHeyLHey1Differentiation
The basic-helix-loop-helix transcription factor HeyL is expressed at high levels by neural crest progenitorcells (NCPs) that give rise to neurons and glia in dorsal root ganglia (DRG). Since HeyL expression wasobserved in these NCPs during the period of neurogenesis, we generated HeyL null mutants to help examinethe factor's role in ganglion neuronal specification. Homozygous null mutation of HeyL reduced the numberof TrkC+ neurons in DRG at birth including the subpopulation that expresses the ETS transcription factorER81. Conversely, null mutation of the Hey paralog, Hey1, increased the number of TrkC+ neurons. Nullmutation of HeyL increased expression of the Hey paralogs Hey1 and Hey2, suggesting that HeyL normallyinhibits their expression. Double null mutation of both Hey1 and HeyL rescued TrkC+ neuron numbers tocontrol levels. Thus, the balance between HeyL and Hey1 expression regulates the differentiation of asubpopulation of TrkC+ neurons in the DRG.
© 2009 Elsevier Inc. All rights reserved.
Introduction
The basic-helix-loop-helix (bHLH) transcription factors, hairy andEnhancer of split are transcriptional repressors in Drosophila thatmediate some of the effects of the Notch signaling pathway on neuralstem cell differentiation (Klambt et al., 1989; Lecourtois andSchweisguth, 1995). Related proteins in mammals include the Hesand Hey families of repressors (Akazawa et al., 1992; Leimeister et al.,1999; Nakagawa et al., 1999; Sasai et al., 1992; Thomas and Rathjen,1992). The Hes family has seven members, Hes1–7, of which Hes1,Hes3, Hes5 and Hes6 are expressed in the developing nervous system(Akazawa et al., 1992; Sakagami et al., 1994; Sasai et al., 1992). In themammalian central nervous system (CNS) Hes1 and Hes5 act asrepressors of proneuronal genes like Mash1 thereby preventingdifferentiation (Akazawa et al., 1995; Castella et al., 1999; de laPompa et al., 1997; Ishibashi et al., 1995, 1994; Jarriault et al., 1995;Nakamura et al., 2000). Hes3 has two isoforms, one of which hasrepressor activity similar to Hes1 while the other lacks repressoractivity (Hirata et al., 2000, 2001). Hes6 represses Hes1 activity andconversely promotes neuronal differentiation (Bae et al., 2000;Gratton et al., 2003; Koyano-Nakagawa et al., 2000). Thus, differentHes proteins play contrasting roles in neural differentiation.
Hey1, Hey2 and HeyL constitute the mammalian Hey (also knownas CHF, Hesr, Herp, and Hrt) family that, like the Hes proteins, actdownstream of the Notch signaling pathway (Iso et al., 2001;Leimeister et al., 1999; Nakagawa et al., 2000, 1999; Steidl et al.,2000). TheHey genes play an important role in the development of thecardiovascular system and loss of Hey function causes defects in
ukhopadhyay).
l rights reserved.
vascular specification, septation and valve formation (Donovan et al.,2002; Fischer et al., 2004, 2007; Gessler et al., 2002; Kokubo et al.,2005, 2004, 2007; Sakata et al., 2002; Xin et al., 2007). In thedeveloping brain, overexpression of Hey1 and Hey2 prevents neuralprecursors from differentiating during the neurogenic period andcauses glial differentiationbyantagonizingneuronal bHLH factors laterin development (Sakamoto et al., 2003). HeyL overexpression in thedeveloping retina increases rod differentiation whereas Hey2increases gliogenesis (Satow et al., 2001). Hey proteins are expressedat high levels in the developingDRGand trigeminal ganglia (Leimeisteret al., 1999, 2000; Nakagawa et al., 1999) but their roles in sensoryganglion development and neuronal specification are unknown.
Sensory ganglion neurons are divided into three major subpopula-tions based on their expression of the neurotrophin receptors and thetype of sensory information they convey—TrkA+ which conveyinformation from thermoreceptors and nociceptors, TrkB+ thatconvey mechanosensory information and TrkC+ that are propriocep-tive (Snider, 1994). Here, we report that mice carrying a targetedmutation in the HeyL gene have decreased numbers of large TrkC+
neurons in the DRG. Moreover, HeyL in the DRG inhibits Hey1expression and mice mutant for Hey1 alone have increased numbersof TrkC+ neurons in the DRG suggesting that Hey1 and HeyL playantagonistic roles in TrkC+ neuron differentiation.
Materials and methods
Generation and maintenance of mouse lines
The targeting construct was generated from the BAC clone RP23-25K8 (CHORI) containing the HeyL locus by recombineering aspreviously described (Liu et al., 2003). All vectors and bacterialstrains used for creating the targeting vector were kind gifts from Dr.

143A. Mukhopadhyay et al. / Developmental Biology 334 (2009) 142–151
Neal Copeland. In the targeting construct loxP sequences wereintroduced such that they flanked the exons 2–5 of the HeyL geneand a neomycin resistance cassette (pGK-neo) flanked by FRT siteswas introduced downstream of the HeyL gene (Fig. 1A). The targetedregionwas flanked both upstream and downstream by 3 kb homologyregions for recombination. The targeting construct was electroporatedinto HM1 mouse ES cells and the neomycin resistant colonies werescreened by southern hybridization with a ∼700 bp probe down-stream of the 3′ homology region. Targeted ES cells were used by theNorthwestern University Transgenic and Targeted Mutagenesis Facil-ity to generate chimeras that transmitted the targeted (HeyLfn) alleleto their progeny. EIIa-cre mice (Jackson Labs) and ROSAFlp1/Flp1
(Jackson Labs) were bred to HeyLfn/+ and the progeny werebackcrossed to WT C57BL6 mice to generate HeyL+/− and HeyLfx/+
mice, respectively. Genotyping was done by polymerase chainreaction (PCR) using a combination of four primers a-5′-GAGCTCTC-TCATGCATGTTCTGCG-3′, b-5′-GGCTAGTTTCTCAGAGCTGAAGGATC-3′,c-5′-CCCCCATACACACACCCTGTTATTCTAG-3′ and d-5′-GAAGCTCTA-AGAGGAAATGCTGGGG-3′ as shown in Fig. 1C. The Hey1+/− micewere kind gifts from Dr. Manfred Gessler and have been describedpreviously (Fischer et al., 2004). Hey1−/−; HeyL+/− and Hey1+/−;HeyL−/− mice were bred to obtain the control (Hey1+/−; HeyL+/−),Hey1 mutant (Hey1−/−; HeyL+/−), HeyL mutant (Hey1+/−; HeyL−/−)and Hey1/L double mutant (Hey1−/−; HeyL−/−) mice.
Southern hybridization and quantitative real time PCR
Genomic DNA was extracted from ES cell colonies and 10 μg ofgenomic DNA samples were digested overnight at 37 °C with BssSI(New England Biolabs). The digested DNA was run on a 0.5% agarosegel, denatured using a solution containing 1.5 M NaCl and 0.5 N NaOHbefore being neutralized in 1M Tris (pH 7.4). The DNA fragments werethen transferred from the gel to a nitrocellulose membrane overnightby capillary transfer. The membrane was baked at 80 °C for 2 h to fixthe DNA on the filter. DIG labeled probe was generated from genomicDNA by PCR using DIG DNA Labeling and Detection Kit (Roche) and
Fig. 1. Targeting and genotyping strategies for HeyL knock out mice. (A) Exons 2–5 of the Heysequences in the targeted HeyLfn allele after homologous recombination. A neomycin resistan5. The targeted allele in the presence of germline flpe or cre recombinases generated the HeyLrestriction digestionwith the 3′ probe in (A) detected a smaller 6.66 kb fragment showing thbetween different alleles. Note the ∼2 kb band (arrow) for the HeyLfn allele when the genommRNA with GAPDH as internal control showed the lack of HeyL mRNA in HeyL−/−. B—BssS
hybridization and detection was done as per manufacturer's instruc-tions. RNAwas extracted from the tissue using RNA Aqueous 4 PCR Kit(Ambion) following manufacturer's instructions. cDNA was preparedfrom the RNA samples with OligodT primers using Thermoscript RT(Invitrogen) following manufacturer's instructions. Real time PCR wasdone on a Mastercycler ep realplex system (Eppendorf) with SYBRgreen (Applied Biosystems) PCR mix.
Immunohistochemistry
Mouse embryo, P0 mouse spinal columns with DRGs or P0 mousethoracic segment with DRGs and sympathetic ganglia were fixed in 4%paraformaldehyde for 2 h and cryoprotected in 30% sucrose solutionovernight. The dehydrated tissue samples were frozen in OCT (TissueTek) before being cryosectioned on a CM3050S cryostat (Leica) at10 μm thickness for immunohistochemistry. The tissue sections wereincubated with primary antibody overnight at 4 °C in a solutioncontaining 1% BSA, 0.25% Triton-X100 and 2% goat serum. For someantibodies antigen retrieval was done by boiling the sections in 10mMsodium citrate solution pH 6 for 10 min before incubation withprimary antibodies. After washes the tissue sections were incubatedwith Alexa Fluor conjugated goat secondary antibodies (1:500)(Invitrogen) in a solution containing 1% BSA, 0.25% Triton-X andDAPI nuclear stain (Invitrogen) for 1 h before a second set of washesand being mounted in ProLong Gold antifade reagent (Invitrogen).The primary antibodies used were Rabbit anti-ER81 (1:1000;Covance), Rabbit anti-p75 (1:500; Dr. Louis Reichardt), Rabbit anti-Parvalbumin (1:1000; Swant), Rabbit anti-Sox10 (1:1000; Abcam),Mouse anti-Substance P (1:500; R&D), Rabbit anti-TrkA (1:500; Dr.Louis Reichardt), Goat anti-TrkB (1:100; R&D), Goat anti-TrkC (1:50;R&D), and Mouse anti-Tyrosine hydroxylase (1:500; Sigma).
RNA in situ hybridization
Whole mount RNA in situ hybridization was done as describedpreviously (Avilion et al., 2000). For in situ hybridization on tissue
L locus that code for the bHLH and orange domains were flanked by loxP (red triangles)ce cassette flanked by FRT sequences (blue triangles) was also introduced 3′ of the exonfx and HeyL− alleles respectively. (B) Southern hybridization of genomic DNA after BssSIe presence of HeyLfn allele. (C) Genotyping primers a–d in (A) were used to distinguishic DNA is amplified with primers c and d. (D) Quantitative real time PCR from trigeminalI, H—HinfI, B’—BslI.
144 A. Mukhopadhyay et al. / Developmental Biology 334 (2009) 142–151
sections 20 μm sections were cut on the cryostat after 4% formalde-hyde fixation and 30% sucrose cryoprotection. Plasmid vectorscontaining cDNA for Hey1, Hey2 and HeyL were kind gifts from Dr.Gessler and have been used previously to generate antisense RNAprobes (Leimeister et al., 1999, 2000). DIG labeled probes wereprepared from the cDNA by PCR using DIG RNA Labeling Kit (Roche) asper manufacturer's instructions. The sections were hybridized inhybridization solution (50% Formamide, 5× SSC, 1% SDS, 500 μg/mltRNA, 200 μg/ml acetylated BSA, 50 μg/ml heparin) overnight at 68 °Cafter which three stringency washes were done for 45 min in a washsolution (50% Formamide, 2× SSC, 1% SDS) at 68 °C. The DIG labelingwas detected using a peroxidase conjugated anti DIG antibody(1:2500, Roche) and developed in NBT/BCIP solution (Roche).
Cell counts and statistical analysis
Sections from six different thoracic DRGs (T1–T6) per animal orthe adjacent sympathetic ganglia were immunostained and imageswere taken using an Axiovert epifluorescence microscope (Zeiss).Cells were counted from four of the largest sections from each DRG,trigeminal and sympathetic ganglia manually using NIH ImageJsoftware. The area of each DRG, trigeminal and sympathetic gangliasection was also measured using ImageJ and the average number ofcells per square millimeter (sq. mm) was calculated for each animal.At least three animals of each genotype were used to get the averagenumber of cells per sq. mm. The examiner was not aware of thegenotypes of the specimens while acquiring images and counting thecells. In experiments with only two conditions Student's t-testassuming unequal variance was used while for multi conditionexperiments one-way ANOVA followed by Tukey's post hoc text wasused to determine statistical significance. All data is presented asmean±SEM.
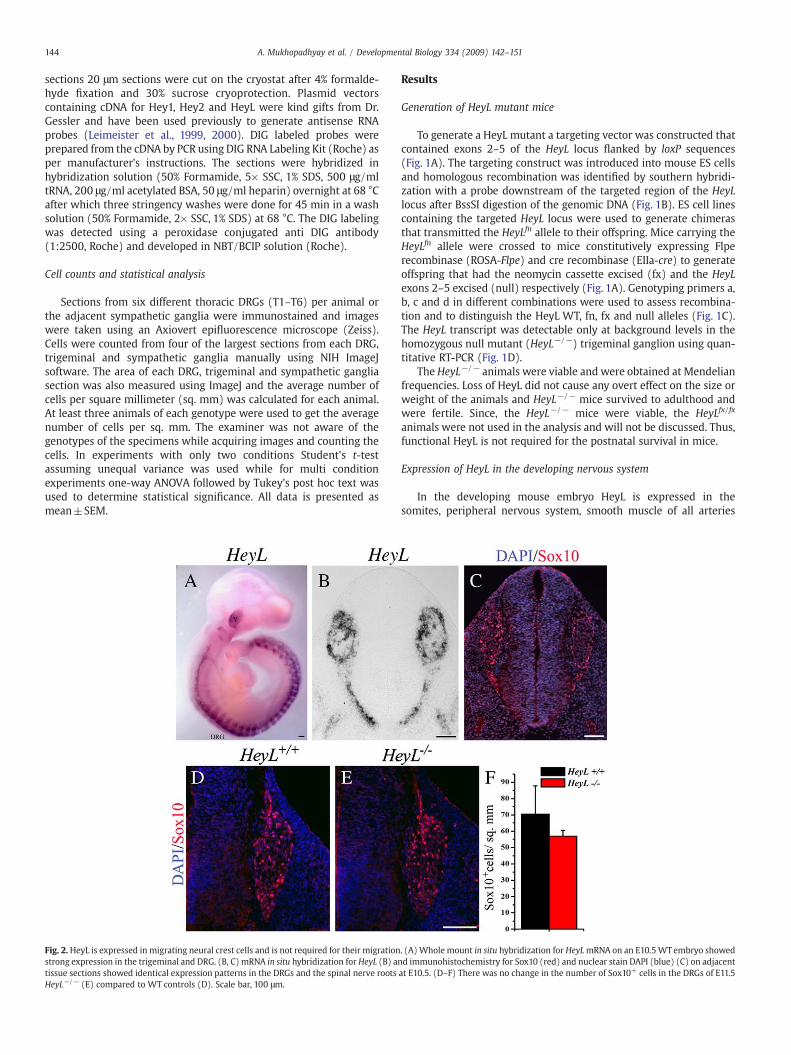
Fig. 2. HeyL is expressed in migrating neural crest cells and is not required for their migrationstrong expression in the trigeminal and DRG. (B, C) mRNA in situ hybridization for HeyL (B) antissue sections showed identical expression patterns in the DRGs and the spinal nerve rootsHeyL−/− (E) compared to WT controls (D). Scale bar, 100 μm.
Results
Generation of HeyL mutant mice
To generate a HeyL mutant a targeting vector was constructed thatcontained exons 2–5 of the HeyL locus flanked by loxP sequences(Fig. 1A). The targeting construct was introduced into mouse ES cellsand homologous recombination was identified by southern hybridi-zation with a probe downstream of the targeted region of the HeyLlocus after BssSI digestion of the genomic DNA (Fig. 1B). ES cell linescontaining the targeted HeyL locus were used to generate chimerasthat transmitted the HeyLfn allele to their offspring. Mice carrying theHeyLfn allele were crossed to mice constitutively expressing Flperecombinase (ROSA-Flpe) and cre recombinase (EIIa-cre) to generateoffspring that had the neomycin cassette excised (fx) and the HeyLexons 2–5 excised (null) respectively (Fig. 1A). Genotyping primers a,b, c and d in different combinations were used to assess recombina-tion and to distinguish the HeyL WT, fn, fx and null alleles (Fig. 1C).The HeyL transcript was detectable only at background levels in thehomozygous null mutant (HeyL−/−) trigeminal ganglion using quan-titative RT-PCR (Fig. 1D).
The HeyL−/− animals were viable and were obtained at Mendelianfrequencies. Loss of HeyL did not cause any overt effect on the size orweight of the animals and HeyL−/− mice survived to adulthood andwere fertile. Since, the HeyL−/− mice were viable, the HeyLfx/fx
animals were not used in the analysis and will not be discussed. Thus,functional HeyL is not required for the postnatal survival in mice.
Expression of HeyL in the developing nervous system
In the developing mouse embryo HeyL is expressed in thesomites, peripheral nervous system, smooth muscle of all arteries
. (A) Whole mount in situ hybridization for HeyLmRNA on an E10.5WT embryo showedd immunohistochemistry for Sox10 (red) and nuclear stain DAPI (blue) (C) on adjacentat E10.5. (D–F) There was no change in the number of Sox10+ cells in the DRGs of E11.5
145A. Mukhopadhyay et al. / Developmental Biology 334 (2009) 142–151
and the mesenchymal tissue surrounding the neural tube (Leime-ister et al., 2000; Nakagawa et al., 1999). To investigate which cells indeveloping peripheral ganglia express HeyL we performed RNA insitu hybridization. In E10.5 embryos HeyL mRNA was expressed indorsal root, trigeminal, and sympathetic ganglia and their nerveroots (Fig. 2A). This expression pattern was very similar to theexpression pattern of the SRY family transcription factor Sox10 thatis expressed in migrating neural crest progenitor cells (Britsch et al.,2001; Kuhlbrodt et al., 1998). For closer examination we probedDRGs from adjacent tissue sections for HeyL mRNA and Sox10protein and found that the HeyL expression (Fig. 2B) was identical tothat of Sox10 (Fig. 2C) in the DRG suggesting they are coexpressed.HeyL expression was also detected in the mesenchyme surroundingthe neural tube (Fig. 2B) as has been previously described(Nakagawa et al., 1999).
Since, HeyL was expressed in migrating neural crest cells weinvestigated if loss of HeyL function affected the neural crestprogenitor cell population in the embryonic DRG. Comparison ofE11.5 HeyL−/− DRGs with their wild type (WT) littermates did not
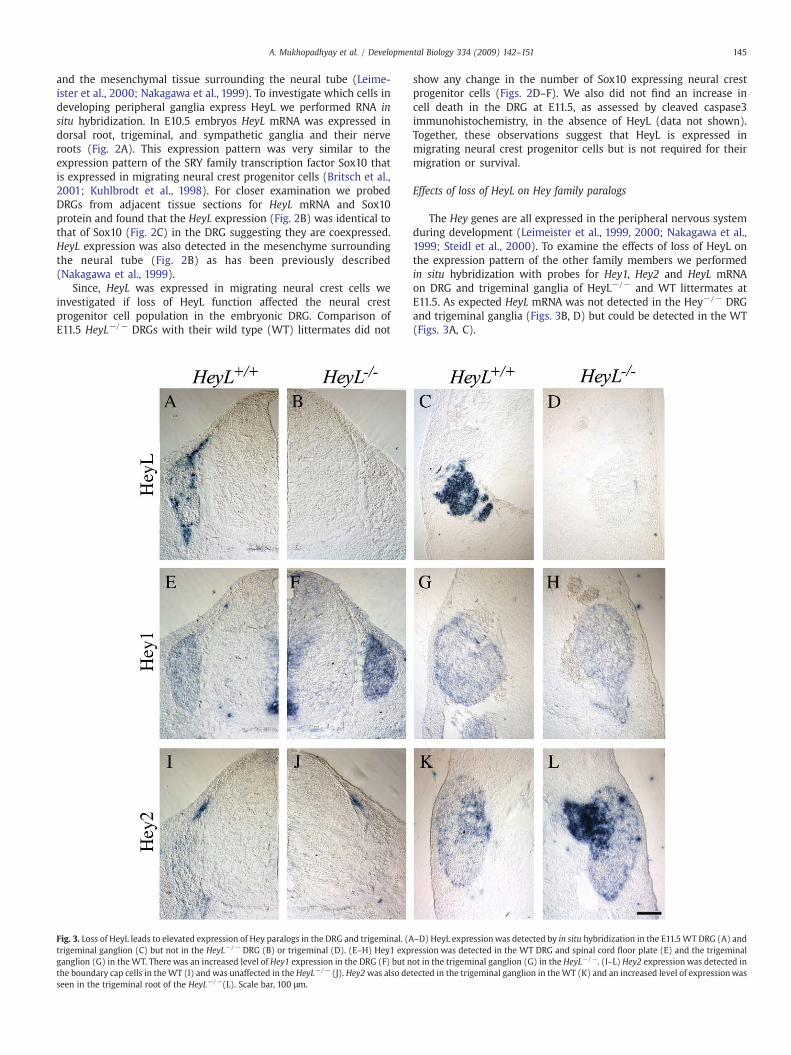
Fig. 3. Loss of HeyL leads to elevated expression of Hey paralogs in the DRG and trigeminal. (Atrigeminal ganglion (C) but not in the HeyL−/− DRG (B) or trigeminal (D). (E–H) Hey1 expganglion (G) in the WT. There was an increased level of Hey1 expression in the DRG (F) but nthe boundary cap cells in theWT (I) and was unaffected in the HeyL−/− (J). Hey2was also deseen in the trigeminal root of the HeyL−/−(L). Scale bar, 100 μm.
show any change in the number of Sox10 expressing neural crestprogenitor cells (Figs. 2D–F). We also did not find an increase incell death in the DRG at E11.5, as assessed by cleaved caspase3immunohistochemistry, in the absence of HeyL (data not shown).Together, these observations suggest that HeyL is expressed inmigrating neural crest progenitor cells but is not required for theirmigration or survival.
Effects of loss of HeyL on Hey family paralogs
The Hey genes are all expressed in the peripheral nervous systemduring development (Leimeister et al., 1999, 2000; Nakagawa et al.,1999; Steidl et al., 2000). To examine the effects of loss of HeyL onthe expression pattern of the other family members we performedin situ hybridization with probes for Hey1, Hey2 and HeyL mRNAon DRG and trigeminal ganglia of HeyL−/− and WT littermates atE11.5. As expected HeyL mRNA was not detected in the Hey−/− DRGand trigeminal ganglia (Figs. 3B, D) but could be detected in the WT(Figs. 3A, C).
–D) HeyL expressionwas detected by in situ hybridization in the E11.5WT DRG (A) andression was detected in the WT DRG and spinal cord floor plate (E) and the trigeminalot in the trigeminal ganglion (G) in the HeyL−/−. (I–L) Hey2 expressionwas detected intected in the trigeminal ganglion in theWT (K) and an increased level of expressionwas

146 A. Mukhopadhyay et al. / Developmental Biology 334 (2009) 142–151
In the ventricular zone of the spinal cord and the floor plate cells,regions where HeyL mRNAwas not detected (Figs. 3A, B), Hey1mRNAexpression was not affected in HeyL−/− compared to WT mice (Figs.3E, F). However, in the DRG where Hey1 and HeyL are both expressed,Hey1 mRNA was upregulated compared to the WT (Figs. 3E, F)suggesting that HeyLmay inhibit the expression of Hey1 in the DRG. Inthe trigeminal ganglion Hey1 transcript was detected only at lowlevels in the WT (Fig. 3G) and the expression was not different in theHeyL−/− mice (Fig. 3H). Interestingly, Hey2 mRNA was detected onlyin the boundary cap cells (Fig. 3I) that give rise to glia in the spinalnerve roots and some neurons and glia in the DRG (Maro et al., 2004).Loss of HeyL however, did not affect the expression of Hey2 in theboundary cap cells (Fig. 3J). In the trigeminal ganglia there was diffuseexpression of Hey2 both in the ganglion and the nerve root (Fig. 3K)and the expression was upregulated in the nerve root in the HeyL−/−
mice (Fig. 3L) suggesting that HeyLmight be involved in the inhibitionofHey2 in the trigeminal ganglion. Thus, loss of HeyL expression causesan upregulation of Hey paralogs in regions of overlapping expression.
Role of HeyL in differentiation of sensory neurons in the DRG
Soon after neurogenesis begins in the DRG (E11.5) almost all DRGneurons express TrkC and many co-express TrkA and TrkB. At thisstage neurotrophin receptor expression does not correspond to thefunctional subtypes of the DRG neurons (White et al., 1996). Later indevelopment neurotrophin receptor expression becomes specific tothe subtype of DRG neuron with the majority expressing TrkA bypostnatal day 0 (P0) (Carroll et al., 1992; Mu et al., 1993). Since HeyL isexpressed at high levels during the neurogenic period in the DRG weexamined if HeyL has a role in the differentiation of different subtypesof neurons.
We immunostained P0 thoracic DRGs with antibodies againstTrkA, TrkB, TrkC and the pan neuronal marker NeuN. The totalnumber of NeuN+ neurons in the HeyL−/− DRGs were comparableto the WT littermates (Supplementary Figs. 1A, B, Fig. 4G). Onanalyzing the specific subtypes, TrkA+ and TrkB+ neuronal popula-tions in the HeyL−/− compared to WT DRG were similar in numbersthough the TrkB+ neurons showed a statistically insignificantreduction (Figs. 4A–D and H, I). However, the large TrkC+ neuronsshowed a significant ∼19% reduction (260.6±7.5 cells/mm2 for WTvs 210.4±11.4 for HeyL−/−; p-valueb0.05) in the HeyL−/− DRGs.The TrkA+ neurons are distributed throughout the DRG, whilemajority of the TrkB+ and TrkC+ neurons are usually found at theperiphery (Mu et al., 1993). There was no overall change in thedistribution of the TrkA+, TrkB+ or TrkC+ neurons in the DRG of theHeyL−/− animal (Figs. 4A–E).
Another region of high HeyL expression is the developingtrigeminal ganglion (Fig. 2A). We also examined the neuronalsubtypes in trigeminal ganglia but did not find any significant changesin the numbers of different sensory neuron subtypes (SupplementaryFigs. 1C–N). Hence, loss of HeyL caused a reduction in the number ofproprioceptive large TrkC+ neurons specifically in the DRG withoutaffecting the nociceptive and thermoceptive TrkA+ neurons or themechanosensory TrkB+ neuron numbers or their distribution.
Effects of loss of HeyL on subtypes of TrkC expressing neurons
Since only a small subpopulation of the TrkC+ neurons was lost inthe HeyL mutant we investigated the expression of proteins thatcharacterize different subsets of TrkC+ DRG neurons. The calciumbinding protein parvalbumin (PV) is expressed in a large subpopula-tion of the proprioceptive neurons and is related to their functionalproperties (Antal et al., 1990; Carr et al., 1989a,b; Ringstedt et al.,1997). We immunostained thoracic DRGs from HeyL mutants and WTmice and found a small decrease in the number of PV immunoreactivecells in the mutant (Figs. 5A–C).
A subset of TrkC+ muscle sensory afferent neurons expresses theETS gene ER81 (Lin et al., 1998). Mice carrying mutations in ER81exhibit limb ataxia and abnormal flexor–extensor posturing of theirlimbs and proprioceptive Ia afferents are lost (Arber et al., 2000).Since ER81 is expressed in DRG neurons outside the TrkC+ popula-tion (Arber et al., 2000) we immunostained DRGs for both TrkCand ER81 to examine effects on the ER81+ subpopulation.Interestingly, the HeyL mutant DRGs exhibited a significant ∼20%reduction (90.4±6.4 cells/mm2 for WT vs 72.5±2.3 for HeyL−/−;p-valueb0.05) in the number of ER81+/TrkC+ neurons comparedto the WT (Figs. 5D–F). This is similar to the reduction in the totalnumber of TrkC+ neurons in the DRG of HeyL mutants (Fig. 4J) andsuggests that both ER81+ and ER81− subpopulations of TrkC+
neurons are affected in the HeyL mutant. Together, these observa-tions suggest a role for HeyL in the differentiation of multiplesubpopulations of TrkC+ neurons in the DRG.
Contrasting roles of Hey1 and HeyL in sensory neuron differentiation
Our previous results demonstrate an upregulation of Hey1expression and a reduction in the TrkC+ neurons in the DRGs of theHeyL−/− mice. This leads to two contrasting possibilities, either thatloss of HeyL expression causes a compensatory increase in theexpression of Hey1 that attenuated the reduction in TrkC+ neurons orthat HeyL inhibition of Hey1 is required for the differentiation of asubpopulation of TrkC+ neurons. To investigate these possibilities wegenerated mice carrying mutations in both Hey1 and HeyL andexamined neuronal differentiation in the thoracic DRGs. We hypothe-sized that if the upregulation of Hey1 is compensatory in nature, Hey1and HeyL should have similar roles in the differentiation process. Hey1mutants (Hey1−/−; HeyL+/−) would then be expected to have areduction in the number of TrkC+ neurons, and there might be a morepronounced effect on the differentiation of TrkC+ neurons in Hey1/Ldouble mutant (Hey1−/−; HeyL−/−) mice. Alternatively, if Hey1repression by HeyL plays a role in the differentiation of TrkC+ neuronsin the DRG, then the Hey1 null mice should have an increased numberof TrkC+ neurons compared to the controls.
The total number of neurons in the DRGs of Hey1 mutant andHey1/2 double mutant mice assessed by NeuN immunostaining werecomparable to control DRGs (Figs. 6A–D). The TrkA+ and TrkB+
neuronal populations in the DRGs of Hey1 or Hey1/L double mutantmice also did not show any changes compared to that in control DRGs(Figs. 6E–L). Examination of other neuronal subtypes in the DRGbased on the expression of the peptide Substance P and the lowaffinity neurotrophin receptor p75 did not reveal any significantchanges in the Hey1, HeyL or Hey1/L double mutants compared to thecontrol animals (Supplementary Figs. 2A–J). Surprisingly, however,the number of TrkC+ neurons in the DRG of Hey1 mutants wassignificantly increased and there were no significant changes inthe Hey1/L double (325.3±12.6 for Hey−/− vs 253.41±29.2 forHey1−/−; HeyL−/− vs 236.5±11.4 for Hey1+/−; HeyL+/−; p-valueb0.05) mutants compared to the control DRGs (Figs. 6M–P).These results together with the observed increase in Hey1 expressionin the HeyL−/− ganglia suggest that Hey1 and HeyL have antagonisticroles in the differentiation of TrkC+ neurons in the DRG.
Discussion
We have created a mouse that carries mutation in the bHLHtranscription factor HeyL. The mutation in the HeyL locus deletessequences that code for all the conserved domains in the HeyL proteinincluding the bHLH, orange and the C-terminal TEIGAF and YHSWmotifs and presumably any function performed by the HeyL protein islost in the mutant animals. The growth, survival and fertility of themutant mice were not affected by the absence of HeyL function. Theonly other reported HeyL mutation in mice in which only the bHLH
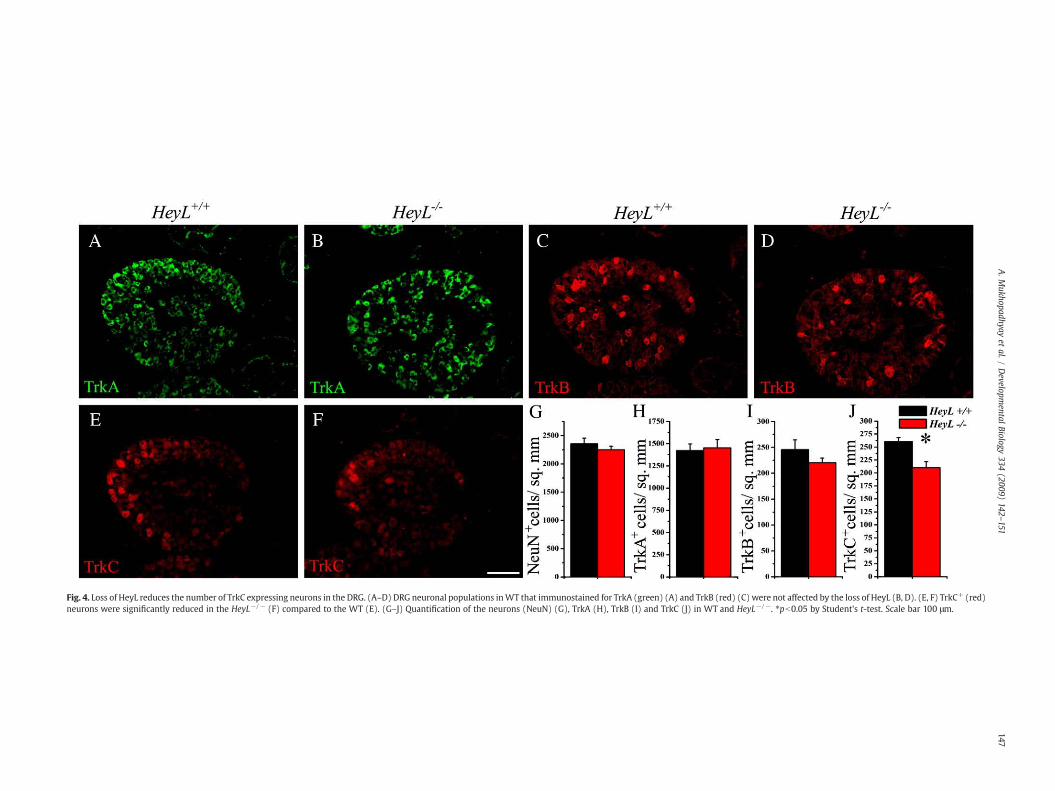
Fig. 4. Loss of HeyL reduces the number of TrkC expressing neurons in the DRG. (A–D) DRG neuronal populations inWT that immunostained for TrkA (green) (A) and TrkB (red) (C) were not affected by the loss of HeyL (B, D). (E, F) TrkC+ (red)neurons were significantly reduced in the HeyL−/− (F) compared to the WT (E). (G–J) Quantification of the neurons (NeuN) (G), TrkA (H), TrkB (I) and TrkC (J) in WT and HeyL−/−. ⁎pb0.05 by Student's t-test. Scale bar 100 μm.
147A.M
ukhopadhyayet
al./Developm
entalBiology
334(2009)
142–151

Fig. 5.Mutation in HeyL reduces the ER81 expressing subpopulation of TrkC+ neurons. (A–C) The PV+ (green) subpopulation of TrkC+ neurons showed a reduction in the HeyL−/−
that is not statistically significant (C). (D–F) The TrkC+ (red) neurons in the DRG that expressed ER81 (green) showed a significant reduction in the HeyL−/− (E, F) compared to theWT (D). ⁎pb0.05 by Student's t-test Scale bar, 100 μm.
148 A. Mukhopadhyay et al. / Developmental Biology 334 (2009) 142–151
and orange domains were affected (Fischer et al., 2007) also did notaffect survival. Thus, the additional loss of the conserved C-terminalmotifs in the HeyL mutant reported here did not cause any differentialeffect on survival.
Comparison of the expression of HeyL and Sox10 indicates thatHeyL is expressed by neural crest progenitor cells during the neu-rogenic period in dorsal root, sympathetic and trigeminal ganglia.Expression of HeyL in neural crest progenitors indicates twocontrasting possibilities for the role of HeyL. First, HeyL might berequired for the migration and maintenance of the neural crestprogenitor population. However the normal number of Sox10+ cells inthe DRG indicates that this is not the case. Alternatively, HeyL mightplay a role in the differentiation of neural crest progenitors intomature cell types. The reduction in the numbers of large TrkC+
neurons in the DRGs of HeyL null mice at P0 provides evidence for thesecond possibility. The TrkB+ neuronal population also showed amodest reduction in the in the HeyL mutants DRG that was notstatistically significant. The loss of HeyL did not change the totalnumber of neurons in the DRG of the mutants though there was atrend towards a reduction. However the TrkB+ and TrkC+ subpopula-tions of neurons in the DRG together constitute less than 20% of thetotal neurons (Mu et al., 1993). Thus the changes in the size of thesesmall populations in the HeyL mutant might not be sufficient todetectably change total neuron numbers given the threshold ofdetection using cell counting methods. The neurons of the sympa-thetic ganglia, which are derived from the neural crest cells and alsoexpressed HeyL, were not significantly affected in the HeyL mutantmice as assessed by tyrosine hydroxylase immunostaining (Supple-mentary Figs. 2K–O). Other mature cell types arising from Sox10+
neural crest cells include Schwann cells and satellite glia in the PNS(Britsch et al., 2001). We did not find any gross changes in Schwanncell differentiation after loss of HeyL, but we cannot exclude thepossibility of subtle molecular changes in Schwann cells that mightindirectly affect the differentiation of DRG neurons.
We find that a null mutation in HeyL increases levels of Hey1 inthe DRG and Hey2 in the trigeminal ganglion. Analogy to the Hesfamily of factors suggests that HeyL directly represses expression ofHey1 and Hey2 by binding to their promoters. In fact directrepression by HeyL of in vitro reporter expression from the Hey2promoter has been reported (Nakagawa et al., 2000) and our resultsprovide the first evidence of this mechanism in vivo. HeyL shows aweaker response to Notch signaling than Hey1 or Hey2 (Maier andGessler, 2000; Nakagawa et al., 2000). This suggests that HeyL mayalso be regulated by other signaling molecules and indeed HeyLexpression is upregulated in neural progenitor cells exposed tobone morphogenetic protein4 (BMP4) (Jalali et al., unpublished).Interestingly, expression of Hey1 can be synergistically induced inthe presence of Notch and BMP signaling in endothelial cells (Itohet al., 2004). BMP signaling regulates the expression of TrkC inneurons in the PNS (Chalazonitis, 2004; Chalazonitis et al., 2004;Zhang et al., 1998). The reduction in TrkC+ neurons in the DRGwhen HeyL function is lost suggests that HeyL might be aneffecter of BMP mediated TrkC expression in a subset of TrkC+
neurons.The Runt family transcription factor Runx3 is expressed in TrkC+
neurons and has been implicated in the cell fate specification in theDRG and projection of sensory afferents (Inoue et al., 2007, 2002;Kramer et al., 2006; Levanon et al., 2002). However Runx3 does notmediate the expression of TrkC in response to BMPs (Inoue et al.,2007). Recently, it has been shown that there is a late bornsubpopulation of DRG neurons that expresses TrkC independent ofRunx3 and that does not express PV (Nakamura et al., 2008). Howeverthe HeyL mutant does not exhibit a significant reduction in PV+
neurons indicating the reduction in TrkC+ neurons is predominantlyfrom the TrkC+/PV− subpopulation. In addition, the HeyL mutant hassimilar numbers of TrkC+ neurons in the DRG at an earlier time pointE11.5 (data not shown) and does not exhibit limb ataxia like the Runx3mutant (Inoue et al., 2002; Levanon et al., 2002). This suggests that
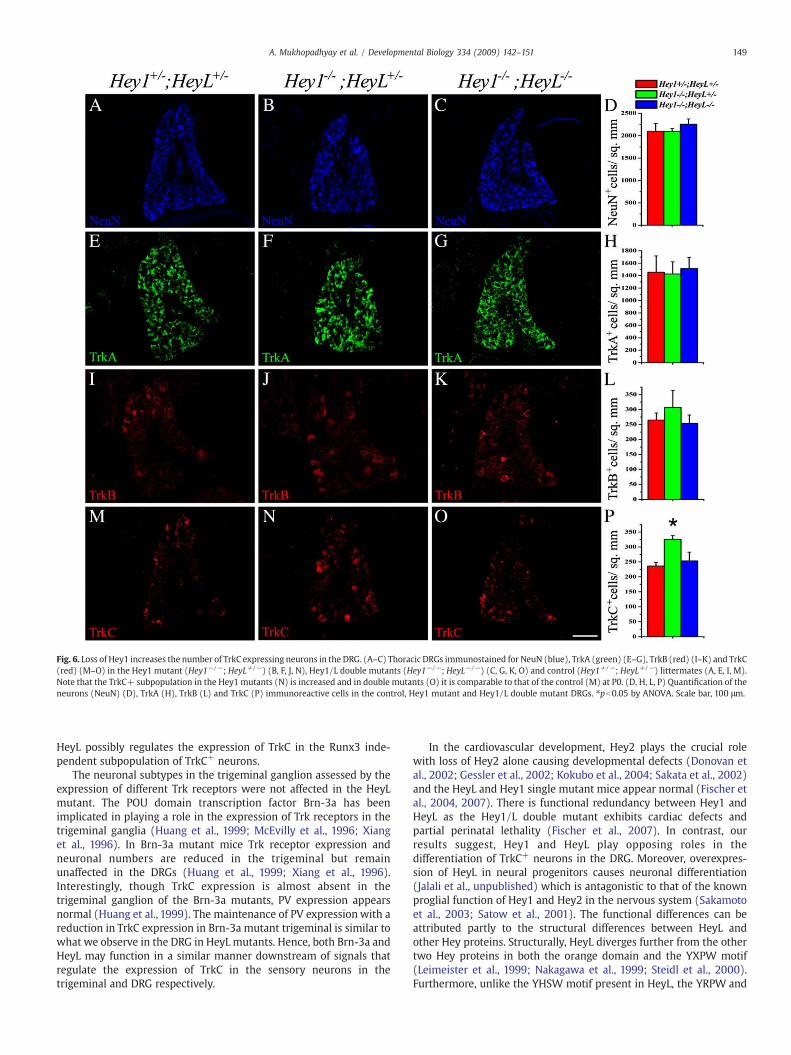
Fig. 6. Loss of Hey1 increases the number of TrkC expressing neurons in the DRG. (A–C) Thoracic DRGs immunostained for NeuN (blue), TrkA (green) (E–G), TrkB (red) (I–K) and TrkC(red) (M–O) in the Hey1 mutant (Hey1−/−; HeyL+/−) (B, F, J, N), Hey1/L double mutants (Hey1−/−; HeyL−/−) (C, G, K, O) and control (Hey1+/−; HeyL+/−) littermates (A, E, I, M).Note that the TrkC+ subpopulation in the Hey1 mutants (N) is increased and in double mutants (O) it is comparable to that of the control (M) at P0. (D, H, L, P) Quantification of theneurons (NeuN) (D), TrkA (H), TrkB (L) and TrkC (P) immunoreactive cells in the control, Hey1 mutant and Hey1/L double mutant DRGs. ⁎pb0.05 by ANOVA. Scale bar, 100 μm.
149A. Mukhopadhyay et al. / Developmental Biology 334 (2009) 142–151
HeyL possibly regulates the expression of TrkC in the Runx3 inde-pendent subpopulation of TrkC+ neurons.
The neuronal subtypes in the trigeminal ganglion assessed by theexpression of different Trk receptors were not affected in the HeyLmutant. The POU domain transcription factor Brn-3a has beenimplicated in playing a role in the expression of Trk receptors in thetrigeminal ganglia (Huang et al., 1999; McEvilly et al., 1996; Xianget al., 1996). In Brn-3a mutant mice Trk receptor expression andneuronal numbers are reduced in the trigeminal but remainunaffected in the DRGs (Huang et al., 1999; Xiang et al., 1996).Interestingly, though TrkC expression is almost absent in thetrigeminal ganglion of the Brn-3a mutants, PV expression appearsnormal (Huang et al., 1999). The maintenance of PV expressionwith areduction in TrkC expression in Brn-3a mutant trigeminal is similar towhat we observe in the DRG in HeyL mutants. Hence, both Brn-3a andHeyL may function in a similar manner downstream of signals thatregulate the expression of TrkC in the sensory neurons in thetrigeminal and DRG respectively.
In the cardiovascular development, Hey2 plays the crucial rolewith loss of Hey2 alone causing developmental defects (Donovan etal., 2002; Gessler et al., 2002; Kokubo et al., 2004; Sakata et al., 2002)and the HeyL and Hey1 single mutant mice appear normal (Fischer etal., 2004, 2007). There is functional redundancy between Hey1 andHeyL as the Hey1/L double mutant exhibits cardiac defects andpartial perinatal lethality (Fischer et al., 2007). In contrast, ourresults suggest, Hey1 and HeyL play opposing roles in thedifferentiation of TrkC+ neurons in the DRG. Moreover, overexpres-sion of HeyL in neural progenitors causes neuronal differentiation(Jalali et al., unpublished) which is antagonistic to that of the knownproglial function of Hey1 and Hey2 in the nervous system (Sakamotoet al., 2003; Satow et al., 2001). The functional differences can beattributed partly to the structural differences between HeyL andother Hey proteins. Structurally, HeyL diverges further from the othertwo Hey proteins in both the orange domain and the YXPW motif(Leimeister et al., 1999; Nakagawa et al., 1999; Steidl et al., 2000).Furthermore, unlike the YHSW motif present in HeyL, the YRPW and

150 A. Mukhopadhyay et al. / Developmental Biology 334 (2009) 142–151
YQPW motifs present in Hey1 and Hey2, respectively, are structurallycloser to the classical groucho binding WRPW domain present in Hesproteins. Hes6 which belongs to the closely related Hes family ofproteins also diverges in function from other Hes proteins like Hes1,Hes3 or Hes5 in the nervous system (Bae et al., 2000; Koyano-Nakagawa et al., 2000) whereas Hes6 functions similarly to other Hesproteins in non-neural tissues (Cossins et al., 2002). The difference inthe functions of Hes6 and other Hes proteins is partly due to ashorter loop in the bHLH sequence (Bae et al., 2000; Koyano-Nakagawa et al., 2000; Pissarra et al., 2000; Vasiliauskas and Stern,2000). Thus, the divergent structure and function of HeyL in theperipheral nervous system are similar to those of Hes6 in the centralnervous system.
Acknowledgments
We thank Neal Copeland for providing us the recombineeringvectors and bacterial strains, Louis Reichardt for the TrkA and p75antibodies and Manfred Gessler for providing us the Hey1, Hey2 andHeyL expression constructs and the Hey1 mutant mouse line. Wethank Lixin Kan and Vibhu Sahni for technical assistance. This projectwas supported by the Brinson Foundation andNIH grants NS 20778-24and NS 20013-25.
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at doi:10.1016/j.ydbio.2009.07.018.
References
Akazawa, C., Sasai, Y., Nakanishi, S., Kageyama, R., 1992. Molecular characterization of arat negative regulator with a basic helix-loop-helix structure predominantlyexpressed in the developing nervous system. J. Biol. Chem. 267, 21879–21885.
Akazawa, C., Ishibashi, M., Shimizu, C., Nakanishi, S., Kageyama, R., 1995. A mammalianhelix-loop-helix factor structurally related to the product of Drosophila proneuralgene atonal is a positive transcriptional regulator expressed in the developingnervous system. J. Biol. Chem. 270, 8730–8738.
Antal, M., Freund, T.F., Polgar, E., 1990. Calcium-binding proteins, parvalbumin- andcalbindin-D 28k-immunoreactive neurons in the rat spinal cord and dorsal rootganglia: a light and electron microscopic study. J. Comp. Neurol. 295, 467–484.
Arber, S., Ladle, D.R., Lin, J.H., Frank, E., Jessell, T.M., 2000. ETS gene Er81 controls theformation of functional connections between group Ia sensory afferents and motorneurons. Cell 101, 485–498.
Avilion, A.A., Bell, D.M., Lovell-Badge, R., 2000. Micro-capillary tube in situ hybridisa-tion: a novel method for processing small individual samples. Genesis 27, 76–80.
Bae, S., Bessho, Y., Hojo, M., Kageyama, R., 2000. The bHLH gene Hes6, an inhibitor ofHes1, promotes neuronal differentiation. Development 127, 2933–2943.
Britsch, S., Goerich, D.E., Riethmacher, D., Peirano, R.I., Rossner, M., Nave, K.A.,Birchmeier, C., Wegner, M., 2001. The transcription factor Sox10 is a key regulatorof peripheral glial development. Genes Dev. 15, 66–78.
Carr, P.A., Yamamoto, T., Karmy, G., Baimbridge, K.G., Nagy, J.I., 1989a. Analysis ofparvalbumin and calbindin D28k-immunoreactive neurons in dorsal root ganglia ofrat in relation to their cytochrome oxidase and carbonic anhydrase content.Neuroscience 33, 363–371.
Carr, P.A., Yamamoto, T., Karmy, G., Baimbridge, K.G., Nagy, J.I., 1989b. Parvalbumin ishighly colocalized with calbindin D28k and rarely with calcitonin gene-relatedpeptide in dorsal root ganglia neurons of rat. Brain Res. 497, 163–170.
Carroll, S.L., Silos-Santiago, I., Frese, S.E., Ruit, K.G., Milbrandt, J., Snider, W.D., 1992.Dorsal root ganglion neurons expressing trk are selectively sensitive to NGFdeprivation in utero. Neuron 9, 779–788.
Castella, P., Wagner, J.A., Caudy, M., 1999. Regulation of hippocampal neuronaldifferentiation by the basic helix-loop-helix transcription factors HES-1 andMASH-1. J. Neurosci. Res. 56, 229–240.
Chalazonitis, A., 2004. Neurotrophin-3 in the development of the enteric nervoussystem. Prog. Brain Res. 146, 243–263.
Chalazonitis, A., D'Autreaux, F., Guha, U., Pham, T.D., Faure, C., Chen, J.J., Roman, D., Kan,L., Rothman, T.P., Kessler, J.A., Gershon, M.D., 2004. Bone morphogenetic protein-2and -4 limit the number of enteric neurons but promote development of a TrkC-expressing neurotrophin-3-dependent subset. J. Neurosci. 24, 4266–4282.
Cossins, J., Vernon, A.E., Zhang, Y., Philpott, A., Jones, P.H., 2002. Hes6 regulatesmyogenic differentiation. Development 129, 2195–2207.
de la Pompa, J.L., Wakeham, A., Correia, K.M., Samper, E., Brown, S., Aguilera, R.J.,Nakano, T., Honjo, T., Mak, T.W., Rossant, J., Conlon, R.A., 1997. Conservation of theNotch signalling pathway in mammalian neurogenesis. Development 124,1139–1148.
Donovan, J., Kordylewska, A., Jan, Y.N., Utset, M.F., 2002. Tetralogy of fallot and othercongenital heart defects in Hey2 mutant mice. Curr. Biol. 12, 1605–1610.
Fischer, A., Schumacher, N., Maier, M., Sendtner, M., Gessler, M., 2004. The Notch targetgenes Hey1 and Hey2 are required for embryonic vascular development. Genes Dev.18, 901–911.
Fischer, A., Steidl, C., Wagner, T.U., Lang, E., Jakob, P.M., Friedl, P., Knobeloch, K.P., Gessler,M., 2007. Combined loss of Hey1 and HeyL causes congenital heart defects becauseof impaired epithelial to mesenchymal transition. Circ. Res. 100, 856–863.
Gessler, M., Knobeloch, K.P., Helisch, A., Amann, K., Schumacher, N., Rohde, E., Fischer, A.,Leimeister, C., 2002. Mouse gridlock: no aortic coarctation or deficiency, but fatalcardiac defects in Hey2 −/− mice. Curr. Biol. 12, 1601–1604.
Gratton, M.O., Torban, E., Jasmin, S.B., Theriault, F.M., German, M.S., Stifani, S., 2003.Hes6 promotes cortical neurogenesis and inhibits Hes1 transcription repressionactivity by multiple mechanisms. Mol. Cell Biol. 23, 6922–6935.
Hirata, H., Ohtsuka, T., Bessho, Y., Kageyama, R., 2000. Generation of structurally andfunctionally distinct factors from the basic helix-loop-helix gene Hes3 byalternative first exons. J. Biol. Chem. 275, 19083–19089.
Hirata, H., Tomita, K., Bessho, Y., Kageyama, R., 2001. Hes1 and Hes3 regulatemaintenance of the isthmic organizer and development of the mid/hindbrain.EMBO J. 20, 4454–4466.
Huang, E.J., Zang, K., Schmidt, A., Saulys, A., Xiang, M., Reichardt, L.F., 1999. POU domainfactor Brn-3a controls the differentiation and survival of trigeminal neurons byregulating Trk receptor expression. Development 126, 2869–2882.
Inoue, K., Ozaki, S., Shiga, T., Ito, K., Masuda, T., Okado, N., Iseda, T., Kawaguchi, S., Ogawa,M., Bae, S.C., Yamashita, N., Itohara, S., Kudo, N., Ito, Y., 2002. Runx3 controls theaxonal projection of proprioceptive dorsal root ganglion neurons. Nat. Neurosci. 5,946–954.
Inoue, K., Ito, K., Osato, M., Lee, B., Bae, S.C., Ito, Y., 2007. The transcription factor Runx3represses the neurotrophin receptor TrkB during lineage commitment of dorsalroot ganglion neurons. J. Biol. Chem. 282, 24175–24184.
Ishibashi, M., Moriyoshi, K., Sasai, Y., Shiota, K., Nakanishi, S., Kageyama, R., 1994.Persistent expression of helix-loop-helix factor HES-1 prevents mammalian neuraldifferentiation in the central nervous system. EMBO J. 13, 1799–1805.
Ishibashi, M., Ang, S.L., Shiota, K., Nakanishi, S., Kageyama, R., Guillemot, F., 1995.Targeted disruption of mammalian hairy and Enhancer of split homolog-1 (HES-1)leads to up-regulation of neural helix-loop-helix factors, premature neurogenesis,and severe neural tube defects. Genes Dev. 9, 3136–3148.
Iso, T., Sartorelli, V., Chung, G., Shichinohe, T., Kedes, L., Hamamori, Y., 2001. HERP, anew primary target of Notch regulated by ligand binding. Mol. Cell Biol. 21,6071–6079.
Itoh, F., Itoh, S., Goumans, M.J., Valdimarsdottir, G., Iso, T., Dotto, G.P., Hamamori, Y.,Kedes, L., Kato, M., ten Dijke Pt, P., 2004. Synergy and antagonism between Notchand BMP receptor signaling pathways in endothelial cells. EMBO J. 23, 541–551.
Jalali, A., Bassuk, A.G., Kan, L., Israsena, N., Mukhopadhyay, A., Hu, M., Kessler, J.A.,unpublished. HeyL promotes neuronal differentiation of neural progenitor cells.unpublished.
Jarriault, S., Brou, C., Logeat, F., Schroeter, E.H., Kopan, R., Israel, A., 1995. Signallingdownstream of activated mammalian Notch. Nature 377, 355–358.
Klambt, C., Knust, E., Tietze, K., Campos-Ortega, J.A., 1989. Closely related transcriptsencoded by the neurogenic gene complex enhancer of split of Drosophilamelanogaster. EMBO J. 8, 203–210.
Kokubo, H., Miyagawa-Tomita, S., Tomimatsu, H., Nakashima, Y., Nakazawa, M., Saga, Y.,Johnson, R.L., 2004. Targeted disruption of hesr2 results in atrioventricular valveanomalies that lead to heart dysfunction. Circ. Res. 95, 540–547.
Kokubo, H., Miyagawa-Tomita, S., Nakazawa, M., Saga, Y., Johnson, R.L., 2005. Mousehesr1 and hesr2 genes are redundantly required to mediate Notch signaling in thedeveloping cardiovascular system. Dev. Biol. 278, 301–309.
Kokubo, H., Tomita-Miyagawa, S., Hamada, Y., Saga, Y., 2007. Hesr1 and Hesr2 regulateatrioventricular boundary formation in the developing heart through the repres-sion of Tbx2. Development 134, 747–755.
Koyano-Nakagawa, N., Kim, J., Anderson, D., Kintner, C., 2000. Hes6 acts in a positivefeedback loop with the neurogenins to promote neuronal differentiation.Development 127, 4203–4216.
Kramer, I., Sigrist, M., de Nooij, J.C., Taniuchi, I., Jessell, T.M., Arber, S., 2006. A role forRunx transcription factor signaling in dorsal root ganglion sensory neurondiversification. Neuron 49, 379–393.
Kuhlbrodt, K., Herbarth, B., Sock, E., Hermans-Borgmeyer, I., Wegner, M., 1998. Sox10, anovel transcriptional modulator in glial cells. J. Neurosci. 18, 237–250.
Lecourtois, M., Schweisguth, F., 1995. The neurogenic suppressor of hairless DNA-binding protein mediates the transcriptional activation of the enhancer of splitcomplex genes triggered by Notch signaling. Genes Dev. 9, 2598–2608.
Leimeister, C., Externbrink, A., Klamt, B., Gessler, M., 1999. Hey genes: a novel subfamilyof hairy- and enhancer of split related genes specifically expressed during mouseembryogenesis. Mech. Dev. 85, 173–177.
Leimeister, C., Schumacher, N., Steidl, C., Gessler, M., 2000. Analysis of HeyLexpression in wild-type and Notch pathway mutant mouse embryos. Mech. Dev.98, 175–178.
Levanon, D., Bettoun, D., Harris-Cerruti, C., Woolf, E., Negreanu, V., Eilam, R., Bernstein,Y., Goldenberg, D., Xiao, C., Fliegauf, M., Kremer, E., Otto, F., Brenner, O., Lev-Tov, A.,Groner, Y., 2002. The Runx3 transcription factor regulates development andsurvival of TrkC dorsal root ganglia neurons. EMBO J. 21, 3454–3463.
Lin, J.H., Saito, T., Anderson, D.J., Lance-Jones, C., Jessell, T.M., Arber, S., 1998. Functionallyrelated motor neuron pool and muscle sensory afferent subtypes defined bycoordinate ETS gene expression. Cell 95, 393–407.
Liu, P., Jenkins, N.A., Copeland, N.G., 2003. A highly efficient recombineering-basedmethod for generating conditional knockout mutations. Genome Res. 13, 476–484.

151A. Mukhopadhyay et al. / Developmental Biology 334 (2009) 142–151
Maier, M.M., Gessler, M., 2000. Comparative analysis of the human and mouse Hey1promoter: Hey genes are new Notch target genes. Biochem. Biophys. Res. Commun.275, 652–660.
Maro, G.S., Vermeren, M., Voiculescu, O., Melton, L., Cohen, J., Charnay, P., Topilko, P.,2004. Neural crest boundary cap cells constitute a source of neuronal and glial cellsof the PNS. Nat. Neurosci. 7, 930–938.
McEvilly, R.J., Erkman, L., Luo, L., Sawchenko, P.E., Ryan, A.F., Rosenfeld, M.G., 1996.Requirement for Brn-3.0 in differentiation and survival of sensory and motorneurons. Nature 384, 574–577.
Mu, X., Silos-Santiago, I., Carroll, S.L., Snider, W.D., 1993. Neurotrophin receptor genesare expressed in distinct patterns in developing dorsal root ganglia. J. Neurosci. 13,4029–4041.
Nakagawa, O., Nakagawa, M., Richardson, J.A., Olson, E.N., Srivastava, D., 1999. HRT1,HRT2, and HRT3: a new subclass of bHLH transcription factors marking specificcardiac, somitic, and pharyngeal arch segments. Dev. Biol. 216, 72–84.
Nakagawa, O., McFadden, D.G., Nakagawa, M., Yanagisawa, H., Hu, T., Srivastava, D.,Olson, E.N., 2000. Members of the HRT family of basic helix-loop-helix proteinsact as transcriptional repressors downstream of Notch signaling. Proc. Natl. Acad.Sci. U. S. A. 97, 13655–13660.
Nakamura, Y., Sakakibara, S., Miyata, T., Ogawa, M., Shimazaki, T., Weiss, S., Kageyama,R., Okano, H., 2000. The bHLH gene hes1 as a repressor of the neuronal commitmentof CNS stem cells. J. Neurosci. 20, 283–293.
Nakamura, S., Senzaki, K., Yoshikawa, M., Nishimura, M., Inoue, K., Ito, Y., Ozaki, S., Shiga,T., 2008. Dynamic regulation of the expression of neurotrophin receptors by Runx3.Development 135, 1703–1711.
Pissarra, L., Henrique, D., Duarte, A., 2000. Expression of hes6, a new member ofthe Hairy/Enhancer-of-split family, in mouse development. Mech. Dev. 95,275–278.
Ringstedt, T., Kucera, J., Lendahl, U., Ernfors, P., Ibanez, C.F., 1997. Limb proprioceptivedeficits without neuronal loss in transgenic mice overexpressing neurotrophin-3 inthe developing nervous system. Development 124, 2603–2613.
Sakagami, T., Sakurada, K., Sakai, Y., Watanabe, T., Nakanishi, S., Kageyama, R., 1994.Structure and chromosomal locus of the mouse gene encoding a cerebellar Purkinjecell-specific helix-loop-helix factor Hes-3. Biochem. Biophys. Res. Commun. 203,594–601.
Sakamoto, M., Hirata, H., Ohtsuka, T., Bessho, Y., Kageyama, R., 2003. The basic helix-
loop-helix genes Hesr1/Hey1 and Hesr2/Hey2 regulate maintenance of neuralprecursor cells in the brain. J. Biol. Chem. 278, 44808–44815.
Sakata, Y., Kamei, C.N., Nakagami, H., Bronson, R., Liao, J.K., Chin, M.T., 2002. Ventricularseptal defect and cardiomyopathy in mice lacking the transcription factor CHF1/Hey2. Proc. Natl. Acad. Sci. U. S. A. 99, 16197–16202.
Sasai, Y., Kageyama, R., Tagawa, Y., Shigemoto, R., Nakanishi, S., 1992. Two mammalianhelix-loop-helix factors structurally related to Drosophila hairy and enhancer ofsplit. Genes Dev. 6, 2620–2634.
Satow, T., Bae, S.K., Inoue, T., Inoue, C., Miyoshi, G., Tomita, K., Bessho, Y., Hashimoto, N.,Kageyama, R., 2001. The basic helix-loop-helix gene hesr2 promotes gliogenesis inmouse retina. J. Neurosci. 21, 1265–1273.
Snider, W.D., 1994. Functions of the neurotrophins during nervous system develop-ment: what the knockouts are teaching us. Cell 77, 627–638.
Steidl, C., Leimeister, C., Klamt, B., Maier, M., Nanda, I., Dixon, M., Clarke, R., Schmid, M.,Gessler, M., 2000. Characterization of the human andmouse HEY1, HEY2, and HEYLgenes: cloning, mapping, and mutation screening of a new bHLH gene family.Genomics 66, 195–203.
Thomas, P.Q., Rathjen, P.D., 1992. HES-1, a novel homeobox gene expressed by murineembryonic stem cells, identifies a new class of homeobox genes. Nucleic Acids Res.20, 5840.
Vasiliauskas, D., Stern, C.D., 2000. Expression of mouse HES-6, a new member of theHairy/Enhancer of split family of bHLH transcription factors.Mech. Dev. 98,133–137.
White, F.A., Silos-Santiago, I., Molliver, D.C., Nishimura, M., Phillips, H., Barbacid, M.,Snider, W.D., 1996. Synchronous onset of NGF and TrkA survival dependence indeveloping dorsal root ganglia. J. Neurosci. 16, 4662–4672.
Xiang, M., Gan, L., Zhou, L., Klein, W.H., Nathans, J., 1996. Targeted deletion of the mousePOU domain gene Brn-3a causes selective loss of neurons in the brainstem andtrigeminal ganglion, uncoordinated limb movement, and impaired suckling. Proc.Natl. Acad. Sci. U. S. A. 93, 11950–11955.
Xin, M., Small, E.M., van Rooij, E., Qi, X., Richardson, J.A., Srivastava, D., Nakagawa, O.,Olson, E.N., 2007. Essential roles of the bHLH transcription factor Hrt2 in repressionof atrial gene expression and maintenance of postnatal cardiac function. Proc. Natl.Acad. Sci. U. S. A. 104, 7975–7980.
Zhang, D., Mehler, M.F., Song, Q., Kessler, J.A., 1998. Development of bone morphoge-netic protein receptors in the nervous system and possible roles in regulating trkCexpression. J. Neurosci. 18, 3314–3326.


















