
Hemicordados: graptolitos - DIGITAL.CSIC:...
31
Transcript of Hemicordados: graptolitos - DIGITAL.CSIC:...


1. INTRODUCCIÓN
El filo Hemichordata comprende deuterostoma-dos marinos, vermiformes a coloniales, caracteri-zados por tener hendiduras branquiales y un exiguocordón nervioso dorsal, que no es homólogo nicomparable con la notocorda de los cordados.
Los hemicordados fueron divididos tradicional-mente en tres clases: enteropneustos, pterobran-quios y graptolitos, siendo la tercera exclusiva-mente fósil. Algunos autores unificaron las dosúltimas en una sola clase Graptolithoidea, suma delas subclases Pterobranchia + Graptolithina. Otrasopiniones distintas, pero emergentes entre los pale-ontólogos, elevan por un lado los graptolitoideos alrango de subfilo (Graptolithina), integrado a su vezpor varias clases: los pterobranquios y cefalodísci-dos actuales, más los antiguos órdenes de graptoli-tos bentónicos y planctónicos. Una revisiónmoderna (Rickards & Durman, 2006) considera,sin embargo, la existencia de un subfiloPterobranchia con sólo tres clases: Graptolithina,Rhabdopleurina y Cephalodiscina, la primera delas cuales englobaría los seis órdenes de graptoli-tos clásicos.
Por su parte, los zoólogos discuten la existenciade otra categoría adicional viviente de hemicorda-dos (las larvas enigmáticas reunidas en el subfilo oclase Planctosphaeroidea), así como la propia enti-dad monofilética de los representantes actuales deeste grupo animal.
Los enteropneustos (subfilo Enteropneusta)son animales solitarios y relativamente grandes (2-180 cm de longitud), de hábitos excavadores y fre-cuentes en todos los mares y océanos actuales.Comúnmente reciben el sobrenombre de “gusanosbellota”, por poseer una neta probóscide anterior,individualizada por un tallo y un cuello cortos, delprominente tronco vermiforme (Fig. 8.1). Estosanimales carecen de lofóforos o brazos ciliados, yposeen un cordón nervioso suplementario (ven-tral). Su registro fósil se limita a un taxón delCámbrico medio conservado en circunstancias
8 Hemicordados: graptolitos
JUAN CARLOS GUTIÉRREZ-MARCO
Figura 8.1. Cordones fecales (en punteado) y madrigueras de algu-nos “gusanos bellota” actuales, con detalle de la organización delcuerpo de un enteropneusto (a la derecha, Saccoglossus).
Gutiérrez-Marco, J.C. 2009. Hemicordados: graptolitos. Capítulo 8, pp. 497-524 in: Paleontología de Invertebrados (Eds.M.L. Martínez Chacón & P. Rivas). Sociedad Española de Paleontología, Universidad de Oviedo, Universidad de Granada,Instituto Geológico y Minero de España (ISBN 978-84-613-4625-7), Oviedo, 524 pp.
1. Introducción ........................................................................497 2. Graptolitos: caractererísticas generales y conservación .....500 3. Estructura, composición y afinidades del esqueleto ...........502 4. Organización y elementos de la colonia .............................506 5. Clasificación .......................................................................512
5.1.Orden Crustoidea ..........................................................5135.2.Orden Camaroidea ........................................................5135.3.Orden Tuboidea ............................................................513
5.4.Orden Dithecoidea ........................................................5135.5.Orden Dendroidea .........................................................5135.6.Orden Graptoloidea ......................................................513
6. Evolución ............................................................................5147. Paleoecología ......................................................................516 8. Paleobiogeografía ...............................................................519 9. Interés bioestratigráfico ......................................................522 10. Bibliografía .........................................................................522
PC
Resaltado
excepcionales (yacimiento de Burgess Shale), deaspecto muy parecido a las formas actuales.Aunque los enteropneustos carecen de cutículafosilizable, sus hábitos sedimentívoros y excava-dores denotan que muchos icnofósiles de madri-gueras verticales en forma de U con varias abertu-ras (icnogénero Balanoglossites, obra también deciertos poliquetos), o incluso cordones fecalesmeandriformes en el plano horizontal, podríanimputarse a la actividad de estos hemicordados a lolargo del Fanerozoico (Fig. 8.1).
Los graptolitoideos (subfilo Pterobranchia) sonhemicordados primitivos de dimensiones milimé-tricas, que viven en tubos secretados por ellos mis-mos. Éstos se organizan bien en grupos (comopseudocolonias bentónicas) o bien forman auténti-cas colonias (bentónicas a planctónicas), con todoslos zooides interconectados, cuyo tamaño puedeascender a varios decímetros. Sus representantesactuales, algunos carentes de esqueleto, son anima-les raros y reúnen escasas especies agrupadas entres géneros y cuatro subgéneros. Dos de ellos,Cephalodiscus y Rhabdopleura, son claros “fósilesvivientes”, dado que la apariencia externa y lamicroestructura de sus tubos es prácticamente lamisma que la de las especies cámbricas (C. savits-kyi y R. obuti, respectivamente). A su vez, la seme-janza existente entre la organización de algunasformas actuales y la interpretación de sus homolo-gías frente a los fósiles de graptolitos, motivó undebate reciente sobre si las mismas son o no grap-tolitos vivos. En su transcurso se sucedieron títuloscomo Graptolites come to life, It’s alive and it’s agraptolite, Living non-graptolite, When is a grap-tolite not a graptolite?, When is a pterobranch agraptolite?, Lebende Graptolithen?-Leider nicht!,o Des graptolites vivants, actuels?, los cuales dancuenta de la difícil y sutil tarea de la discrimima-
2 GUTIERREZ-MARCO
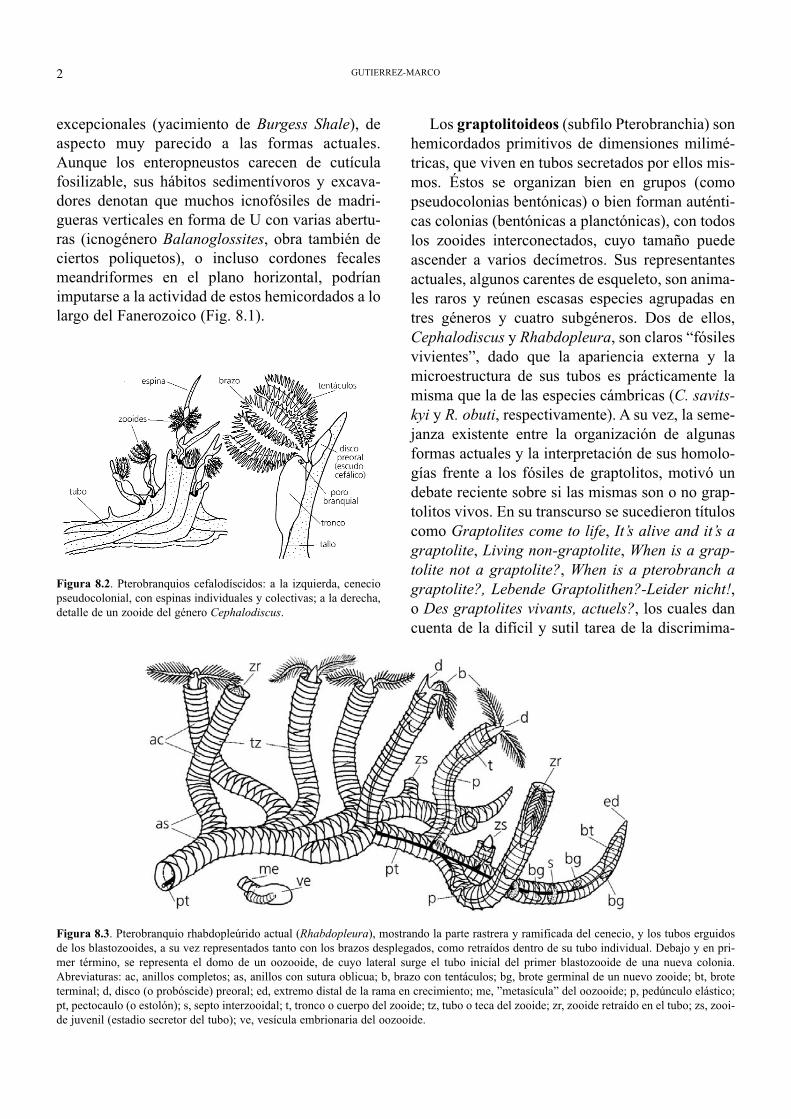
Figura 8.2. Pterobranquios cefalodíscidos: a la izquierda, ceneciopseudocolonial, con espinas individuales y colectivas; a la derecha,detalle de un zooide del género Cephalodiscus.
Figura 8.3. Pterobranquio rhabdopleúrido actual (Rhabdopleura), mostrando la parte rastrera y ramificada del cenecio, y los tubos erguidosde los blastozooides, a su vez representados tanto con los brazos desplegados, como retraídos dentro de su tubo individual. Debajo y en pri-mer término, se representa el domo de un oozooide, de cuyo lateral surge el tubo inicial del primer blastozooide de una nueva colonia.Abreviaturas: ac, anillos completos; as, anillos con sutura oblicua; b, brazo con tentáculos; bg, brote germinal de un nuevo zooide; bt, broteterminal; d, disco (o probóscide) preoral; ed, extremo distal de la rama en crecimiento; me, ”metasícula” del oozooide; p, pedúnculo elástico;pt, pectocaulo (o estolón); s, septo interzooidal; t, tronco o cuerpo del zooide; tz, tubo o teca del zooide; zr, zooide retraído en el tubo; zs, zooi-de juvenil (estadio secretor del tubo); ve, vesícula embrionaria del oozooide.

ción zoológica dentro de los graptolitoideos(Urbanek, 1986, 1994; Mierzejewski & Kulicki,2003; Maletz et al., 2005; Rickards & Durman,2006).
Tanto la mayoría de los pterobranquios actualescomo los graptolitos secretan un exoesqueletocolonial o tubario de naturaleza orgánica, llamadocenecio en el primer grupo y rabdosoma en elsegundo. El tubario se desarrolla a partir de un pri-mer zooide diferenciado, que en estado larvariocomienza con una vesícula, inicialmente cerradaen los hemicordados (domo) y abierta en los grap-tolitos (prosícula). La larva de este zooide inicial,fruto de reproducción sexual (oozooide), completasu crecimiento con la construcción de un tubo ometasícula formado por incrementos anulares, loscuales se corresponden con el peridermo fuselartan típico de todos los graptolitoideos (Figs. 8.3,8.4).
En los pterobranquios cefalodíscidos (claseCephalodiscina), el zooide inicial genera un largotubo que se asocia en forma de racimo con otrostubos adyacentes, creando pseudocolonias cuyosindividuos llegan a colaborar entre sí para cons-truir, mediante órganos especializados de su pro-bóscide, aberturas comunes y espinas aisladas omúltiples (Fig. 8.2). Cada zooide se fija al interiorde su tubo por un largo tallo retráctil, tal vez homó-logo de la cola post-anal con la que se anclan alsustrato las crías de los enteropneustos, y de sucuerpo parten de cinco a nueve pares de brazosprovistos de numerosos tentáculos ciliados.
Tanto en los pterobranquios rabdopléuridos(clase Rhabdopleurina) como en los graptolitos(la clase Graptolithina tradicional), el crecimien-to colonial se inicia a partir de un segundo indivi-duo que nace por gemación y se desarrolla, ya conperidermo fuselar, a partir de un poro abierto late-ralmente (por reabsorción) en la metasícula oincluso en la prosícula. Los restantes individuos dela colonia (blastozooides) brotan por gemación apartir del tallo del zooide que le precede o bien ger-minan del estolón (= pectocaulo) que relacionaentre sí los tallos de todos los zooides de unamisma colonia (Fig. 8.3). Cada blastozooide habi-ta la teca o tubo individual que él mismo secreta,aunque una vez edificada la colonia, si alguno
muere, puede ser reemplazado y el nuevo indivi-duo recién brotado, continuar con el crecimiento orefuerzo de la teca heredada. El cuerpo de los rab-dopléuridos tiene una constitución muy similar alde los cefalodíscidos, con un tronco en forma desaco acabado en una probóscide preoral y, por elotro extremo, en un largo tallo retráctil, que en estecaso se une al estolón común que interconectatodos los zooides, en vez de a la base de la teca(Fig. 8.3). Sin embargo, del cuerpo sólo parte unpar de brazos con tentáculos ciliados, cuya consti-tución (estructuras huecas en prolongación delmesoceloma) es muy parecida a la de un lofóforo.Además, carecen de hendiduras branquiales.
El registro geológico de rabdopléuridos y cefa-lodíscidos es bastante discontinuo en el Paleozoicoy Mesozoico (Chapman et al., 1995; Mierzejewski& Kulicki, 2002; Rickards & Durman, 2006), sinque existan apenas datos sobre fósiles cenozoicos,confundidos generalmente con restos de algas o de
3HEMICORDADOS: GRAPTOLITOS
Figura 8.4. Fósiles de graptolitos separados químicamente de lamatriz, en cuyo peridermo se distinguen los anillos fuselares, enimágenes de microscopía electrónica de barrido (A), óptica conven-cional (B-D) y videomicroscopía de infrarrojo (E), que vuelve trans-lúcida una colonia opaca. A, sícula inmadura de dicograptoideo (x6); B, sícula de diplográptido (x 3); C, sícula de monográptido concrecimiento incipiente de la primera teca (x 2,5); D, idem. con sícu-la y tres tecas ascendentes (Pseudomonoclimacis del Ludlow bálti-co, x 2,6); E, colonia biserial dipleural (Paraorthograptus delKatiense de Canadá, x 3), con la sícula en la parte izquierda, el nemaprolongado en vírgula y tecas sigmoides con espinas geniculares.Abreviaturas: c, cauda; ea, espina apertural; eg, espinas geniculares;m, metasícula; n, nema; p, prosícula; si, sícula; t, teca; vg, virguela.A, cortesía de Roberto Albani; E, tomada de un ejemplar de MikeMelchin.

cnidarios hidroideos. El desarrollo de las modernastecnologías de retrodispersión electrónica enmicroscopía de barrido, ha posibilitado el recono-cimiento del peridermo fuselar en muchos restosdudosos de conservación carbonosa, reasignablesen su mayoría a hemicordados paleozoicos. Porotra parte, los escasos fósiles de Stolonodendrum,en principio diferenciados como un orden aparte(“Stolonoidea”) de graptolitos ordovícicos, actual-mente se integran en el orden Rhabdopleurida.
En España, Maletz et al. (2005) reinterpretaroncomo posibles pterobranquios cuatro géneros decolonias bentónicas erguidas, procedentes delCámbrico medio de la Zona Cantábrica, los cualeshabían sido descritos por Sdzuy (1974) comograptolitos ancestrales (orden Dithecoidea: Fig.8.13C-D). Previamente, otros autores habíanapuntado una interpretación alternativa para estosfósiles, en el sentido de considerarlos como posi-bles restos de escifozoos coronados, que incluyenformas actuales de aspecto muy similar al de losditecoideos, por estar construidas a base de tubosorgánicos con incrementos transversos semejantesa los anillos fuselares. Sin embargo, la revisiónreciente de Rickards & Durman (2006) ha detec-tado evidencias claras del estolón en uno de losgéneros del Cámbrico ibérico (Sotograptus), loque, unido a la conservación del peridermo fuse-lar en muchas formas de ditecoideos, sitúa denuevo al grupo incuestionablemente entre losgraptolitos más primitivos.
2. GRAPTOLITOS: CARACTERÍSTICASGENERALES Y CONSERVACIÓN
Los graptolitos son hemicordados coloniales,exclusivamente paleozoicos, cuyo nombre alude alaspecto más habitual que presentan sus fósiles (delgr. graptós= escrito, y lithos= piedra). El sufijo -lithos obliga a que el término correcto en españolsea graptolito (pl. graptolitos), al igual que ocurrecon las palabras gastrolito, coprolito, monolito,aerolito, etc. Este proceder fue adoptado por lasReales Academias españolas y diversas academiascientíficas iberoamericanas, descartando la caste-llanización incorrecta más común (por ejemplo, latraducción de Bates, 1989) como “grapto-
lite”/“graptolites” (-ite es fem. clás.), en vana tras-literación vernácula de un anglicismo.
Los fósiles de graptolitos se conservan por logeneral como moldes mineralizados o como pelí-culas carbonosas aplastadas en pizarras y rocasarcillosas, asemejándose a trazos gráficos por suhabitual contraste de color frente a la matriz (Fig.8.5A-C). A grandes rasgos consisten en varillas deaspecto dentado que corresponden en realidad alesqueleto colonial o rabdosoma del graptolito,formado por una o más ramas o estipes (Figs.8.4D-E; 8.5D-M; 8.9A-H) cuyo número varíadependiendo del modo de vida de la colonia (ben-tónico o planctónico) y su tendencia evolutiva,encaminada en las colonias planctónicas hacia lasimplificación morfológica. Cada estipe compren-de una fila única o principal de tubos, llamadostecas, que crecen de forma consecutiva y se abrenen un mismo sentido, por lo que la rama adquiereun aspecto serrado por uno de sus bordes, en tantoque la parte opuesta se mantiene lisa, y cuyo perfilrecto o curvado va a depender de la morfología tri-dimensional del estipe (Fig. 8.5K-L).
Para entender un graptolito conviene tener pre-sente que cada fósil corresponde al esqueletoaplastado de una colonia completa o a parte de ella,que originalmente tuvo un cierto volumen y unacomposición exclusivamente orgánica, y en la quecada teca fue habitada por un individuo colonial.Esta naturaleza proteica del esqueleto de los grap-tolitos, carente de impregnaciones minerales, cons-tituyó sin duda un factor limitante para la conser-vación del mismo, lo que en principio sería contra-dictorio con la gran abundancia y notoriedad delregistro fósil del grupo en las rocas paleozoicas. Enefecto, los graptolitos proliferaron en todo tipo dehábitats planctónicos del Ordovícico y Silúrico, ysus esqueletos se acumularon por millones en ubi-cuos fondos anóxicos, fruto de estancamientos enla circulación profunda de las aguas marinas, vin-culados con ciclos eustáticos globales; o bien enáreas de alta productividad, relacionadas con aflo-ramientos (upwelling) de masas de agua oceánicarica en nutrientes (Fig. 8.16). Ello explica por quélos fósiles de graptolitos son tan abundantes enciertos sedimentos oscuros organógenos, en espe-cial las facies de pizarras negras carentes de biotur-
4 PALEONTOLOGÍA DE INVERTEBRADOS

bación (black shales), también llamadas sapropeli-tas o incluso “ampelitas” (de ampelos= vid, por suantiguo uso para abonar viñedos merced a sus con-centraciones en sulfatos y aluminosilicatos). A laacumulación de los graptolitos en estos sedimentostan particulares, seguiría el rápido aplastamientode los esqueletos coloniales durante la compacta-ción diagenética, y su conservación, bien comopelículas carbonosas (Fig. 8.5D, M) o preferente-mente como moldes, en los que el hueco dejado
por la degradación del rabdosoma se rellenó luegopor minerales de arcilla de alteración blanquecina(cloritas, pirofilitas: Figs. 8.5A, K, L; 8.6I), sustan-cias ferruginosas (Fig. 8.5B, G) o incluso cuarzo(variedades metamórficas fibrosas). Con una fre-cuencia menor, los rabdosomas de graptolitos con-servados en pizarras negras mantuvieron ciertogrado de su relieve original, hecho relacionado conla precipitación diagenética precoz de pirita porbacterias sulforreductoras, en el hueco interno del
5HEMICORDADOS: GRAPTOLITOS
Figura 8.5. Aspectos generales y morfología básica de la colonia de los graptoloideos planctónicos. A-B, moldes aplastados en pizarras, relle-nos por cloritas alteradas (A, x 0,3) u óxidos de hierro (B, x 1); C, rabdosomas tridimensionales de composición orgánica en caliza(Saetograptus del Ludlow báltico, x 1,8); D, colonia con dos estipes pendientes (Didymograptus del Darriwiliense de Cáceres, x 2); E, colo-nia con cuatro estipes ascendentes (Tetragraptus del Dapingiense de Noruega, x 2,4); F, rabdosoma de cuatro estipes ascendentes, unidos dor-salmente (Phyllograptus del Dapingiense de EEUU, x 2,1); G, colonia biserial dipleural, con los dos estipes unidos dorsalmente(Eoglyptograptus del Darriwiliense de Cáceres, x 2,7); H, rabdosoma biserial monopleural, con los estipes unidos por una de las caras latera-les (Glossograptus del Sandbiense de Perú, x 3,5); I, colonia biserial en conservación escalariforme (Normalograptus del Aeroniense checo,x 4); J, vista ventral de un rabdosoma ramificado en la horizontal, con estipes en conservación escalariforme (Clonograptus del Tremadociensede Huelva, x 1,4); K, rabdosomas uniseriales de estipes rectos (Colonograptus del Wenlock de León, x 5); L, colonia uniserial aplastada, conestipe arrollado en espiral turriculada (Spirograptus, Telychiense de Lugo, x 1,7); M, Didymograptus con membrana peridérmica que envuel-ve la sícula y las primeras tecas, Darriwiliense de Asturias (x 1,5). Abreviaturas: e, estipe; s, sícula; te, teca; ea, espina adapertural de la sícu-la; e(n) estipe secundario implicado en una dicotomía; et, espina tecal; mde, margen dorsal del estipe; mve, margen ventral del estipe; n, nema;r, rabdocórtex; vg, virguela; vi, vírgula.

esqueleto (Fig. 8.6E-F, J-K). A veces, la actividadde microorganismos en ambientes de oxigenaciónnormal, combinada con altas tasas de sedimenta-ción y ciertos enriquecimientos minerales, influyetambién en la piritización o fosfatización tempranade los graptolitos en el interior de nódulos y arenis-cas finas, que generan fenómenos de conservacióntridimensional secundariamente silicificados odesferruginizados (Fig. 8.6B, L).
Por otro, lado existen casos frecuentes, sobretodo en pizarras negras, de rabdosomas helicoida-les o de cierto volumen, que fueron aparentementeconservados bajo tapices bacterianos, quizá muyextendidos en los fondos marinos del Silúrico(Jones et al., 2002). Tales recubrimientos algales ymicrobianos dificultaron el enterramiento gradualde aquellos rabdosomas voluminosos que, tras lacompactación por la carga sedimentaria acumula-da, resultaron aplastados en un solo plano, sinmediación de sedimento intersticial (Fig. 8.5L).
La orientación de los restos de graptolitos en elsedimento, independientemente de la obrada por eloleaje y las corrientes, es importante también parainterpretar la morfología de muchos fósiles.Normalmente los estipes individuales tienen unasección ovalada, y cuando caen al fondo reposansobre uno de sus flancos laterales, por ser ésta laposición más estable (Fig. 8.5A, D, K). En estacomún circunstancia, los fósiles aparecen delimita-dos por el borde dorsal del estipe (recto o curvado),a un lado, y por el perfil ventral de las tecas, a otro(el borde denticulado opuesto). Cuando un mismograptolito presenta “tecas en ambos lados”, o bientiene un rabdosoma complejo, configurado entorno a dos o más estipes unidos dorsal o lateral-mente entre sí (Figs. 8.4E; 8.5F-H), o bien se tratade casos de superposición casual de estipes de dis-tintas colonias, en orientación opuesta, un casonada infrecuente en los yacimientos más ricos. Porúltimo, si un estipe no exhibe ninguno de sus dosperfiles denticulados, lo más probable es que sedisponga perpendicularmente al sedimento, por loque estamos ante sus contornos laterales, observa-bles bien en vista dorsal (“de canto”), o bien envista ventral (el sector al que se abren las tecas).Este segundo caso se denomina vista escalarifor-me cuando las aberturas tecales adoptan una dispo-
sición frontal, parecida a los huecos de una escale-ra de mano (Figs. 8.5I-J; 8.6B). Las orientacionesescalariformes o subescalariformes son frecuentesen rabdosomas o estipes de sección circular, en losque cualquier posición aleatoria resulta establesobre el fondo. Las vistas dorsales son comunesentre los rabdosomas cónicos o de aspecto dendroi-deo, donde las tecas se abren hacia el interior o unade las caras de la colonia (Figs. 8.9A-D, F-H;8.10A-D), y también en aquellos rabdosomas com-plejos ramificados en el plano horizontal, cuyoentramado de estipes hace que éstos fosilicen conlas aberturas tecales apoyadas sobre el fondo (Fig.8.10G-H, M). Pero a veces unos estipes que apa-rentan estar en vista dorsal, porque en ellos no seadvierten indicios de las denticulaciones tecales,en realidad corresponden a fosilizaciones lateralesde tecas muy elaboradas (por ejemplo, de tipogeniculado), subparalelas al eje del estipe y con lasaperturas en forma de ranura (Figs. 8.4E; 8.6B;8.10N; 8.11B-C, H, J).
Con todo, la conservación más espectacular yfidedigna del esqueleto de los graptolitos se da enciertas calizas pelágicas, sílex y nódulos, donde elperidermo mantiene su naturaleza orgánica e inclu-so cierto grado de la flexibilidad original, pudien-do extraerse sus restos de la roca mediante proce-dimientos químicos (Figs. 8.4; 8.6C-D). Este tipode material es el que ha permitido analizar la com-posición e histología del rabdosoma, extraordina-riamente significativa a la hora de interpretar lasafinidades biológicas de los graptolitos.
3. ESTRUCTURA, COMPOSICIÓN Y AFINI-DADES DEL ESQUELETO
El esqueleto o peridermo del graptolito está for-mado por una serie de tubos de naturaleza esclero-proteica que, a escala ultramicroscópica, se com-ponen de fibrillas con estriación espiral, posible-mente formadas por colágeno, delimitadas por unafábrica granular compleja de proteínas y polisacá-ridos (Towe & Urbanek, 1972; Crowther &Rickards, 1977; Rickards & Dumican, 1984;Armstrong et al., 1984; Briggs et al., 1995; Bateset al., 2009). El colágeno es la proteína fibrosa queconstituye la base de todos los tejidos conjuntivosanimales y forma, por ejemplo, los tendones de los
6 PALEONTOLOGÍA DE INVERTEBRADOS

vertebrados, así como el esqueleto de los rabdo-pléuridos actuales. Aunque los rabdosomas aparen-temente intactos de muchos graptolitos conservancierta elasticidad, los estudios moleculares revelanque el peridermo de estos fósiles tiene un compo-nente alifático predominante (similar a un queró-geno de tipo II), debido a su polimerización in situconsiguiente a la pérdida de ácidos grasos no satu-rados, que dio paso a la formación de unos agrega-dos lipídicos muy resistentes (Gupta et al., 2006).
Desde el punto de vista histológico, el esquele-
to de los graptolitos consta a grandes rasgos de untejido fuselar de textura esponjosa, con fibrillasdiminutas orientadas aleatoriamente, y de un teji-do cortical formado por haces superpuestos defibras de mayor grosor, tanto simples como entre-lazadas longitudinalmente (Fig. 8.7A). Las fibrascorticales se hallan interconectadas externamentepor trabéculas transversas, cuyo espaciamientoregular replica, de un modo indirecto, el bandeadointerno de las fibras de colágeno. Ambos tipos detejidos contactan entre sí y con el exterior a través
7HEMICORDADOS: GRAPTOLITOS
Figura 8.6. Fósiles y conservación de los graptolitos. A, fragmento de Monograptus que ostenta el récord de longitud para una colonia indi-vidual (1,45 m, fotografía por cortesía de David Loydell); B, rabdosomas de Normalograptus, conservados tridimensionalmente dentro denódulos ferruginizados en cuarcitas (Llandovery de Ciudad Real, x 6,2); C-D, fragmentos de graptolitos separados químicamente de la matriz(C, axil de un Dicranograptus del Ordovícico de EEUU, fotografiado con MEB, x 8; D, monográptidos y retiolítidos, sumergidos en gliceri-na, del Wenlock de Polonia, x 1,8); E-H y J-K, ejemplares piritizados de Baltograptus (E, Floiense de Perú, x 6,6), Eoglyptograptus (F y H,Darriwiliense de Asturias, x 10 y x 6 respectivamente: observar anillos fuselares en F), Undulograptus (G, Darriwiliense de Turquía, vaciadoen látex del molde externo, x 6), Parapetalolithus (J, Telychiense de Guadalajara, x 3) y Hustedograptus (K, Darriwiliense de Noruega, x3,6); I, molde de Metaclimacograptus en pirofilita (Telychiense de Guadalajara, x 6); L, vaciado en látex del interior de una colonia dePristiograptus, fosfatizada en un nódulo (Sheinwoodiense de Portugal, x 6).

de una serie de láminas, membranas y huecos(fábricas laminar, crasa, reticulada, verrucosa...;paracórtex, pseudocórtex, brastocripto...; membra-na granular, canalículos, vesículas, etc.), descritoso revisados en los trabajos de Crowther (1981),Urbanek & Mierzejewski (1984) o Rickards &Dumican (1985), entre otros.
El tejido fuselar es el responsable del creci-miento de la colonia y se desarrolla en anillosfuselares (Figs. 8.4, 8.6F, 8.7A) que, en la mayorparte de los graptolitos, están formados a su vezpor dos segmentos semicirculares o fúsulas, solda-dos oblicuamente en sus extremos. La sucesiónalterna de las suturas oblicuas, a lo largo de anillosconsecutivos, conforma las características líneasen zigzag visibles en las caras dorsal y ventral delas tecas. A su vez, cada anillo fuselar tiene unasección longitudinal cóncavo-convexa en el senti-do del crecimiento, donde el entrante central alar-gado del borde inferior de un determinado anillo,encaja en el saliente alargado del borde superiordel anillo precedente, y así sucesivamente (Fig.8.7A). La altura fuselar media es de 20-35 micras,y se va incrementando gradualmente dentro decada teca. Las partes protecales y las aberturas demorfología compleja, suelen implicar microfuse-los, que son incrementos más pequeños y disconti-nuos, de estructura interna mucho más densa quelos anillos normales.
El tejido cortical está normalmente superpuestoal fuselar y sirve de refuerzo al mismo (como ecto-córtex o endocórtex), disponiéndose en el exteriordel rabdosoma a modo de bandas corticales entre-cruzadas de distinto grosor (eucórtex: Fig. 8.7A).Con toda probabilidad, éstas fueron secretadas porla probóscide preoral de los zooides, sin duda dota-dos de un activo movimiento extratecal, únicamen-te restringido por la longitud y flexibilidad delestolón que los unía al resto de los individuos de lacolonia. Pero el tejido cortical no sólo se limita alreforzamiento estructural de las tecas y espinas,sino que a veces genera espesamientos localizadosen las porciones más delicadas de las colonias paraevitar su ruptura, envolviendo por ejemplo la sícu-la y las primeras tecas (rabdocórtex: Fig. 8.5M).En otras ocasiones crea o refuerza disepimentos,desactiva parásitos (encapsulándolos en vainas
protectoras llamadas tubotecas -Fig. 8.9B-, cica-triza agallas...) o bien contribuye a la flotabilidadde la colonia mediante la construcción de láminascorticales membranosas entre estipes adyacentes.
Con todo, el esqueleto de los graptolitos se dife-rencia del de los pterobranquios actuales por sucarácter heteromórfico, dado que su material fibro-so (fuselar y cortical) está organizado de distintamanera, mientras que en rabdopléuridos y cefalo-díscidos, de peridermo homeomórfico, el córtex yfuselos son estructuralmente similares, tras habersido secretados por el escudo cefálico de la probós-cide de los propios zooides. En virtud de estas yotras diferencias, algunos autores apuntaron que elrabdosoma de los graptolitos pudo haber permane-cido encerrado en un tejido blando externo, imagi-nariamente cubierto de cilios, cuyas únicas abertu-ras coincidirían con los orificios tecales de los queasomarían los apéndices nutritivo-respiratorios(semejantes a lofóforos), los cuales coadyuvarían ala propulsión de la colonia (Kirk, 1994). Esta dis-crepancia se debe a que el “modelo pterobranquio”de secreción no explica convincentemente el des-arrollo de estructuras complejas en los extremos dela colonia (bases de fijación de formas bentónicaserguidas, vírgulas, nematulario u órganos sicularesen planctónicos), algunas de ellas difícilmente ase-quibles por unos zooides sujetos por el estolón(Rickards, 1996), ni tampoco el armazón corticalde los retiolítidos, cuya construcción sigue siendoun enigma. Tanto para este último como para loselementos nemales, esencialmente formados portejido fuselar, se plantea un modo de secrecióninterna a partir de tejidos blandos que, o bien recu-brirían toda la colonia (Kirk, 1994), o bien queda-rían restringidos a ciertos bordes en crecimiento(Crowther & Rickards, 1977).
Otras diferencias entre los graptolitos y los pte-robranquios actuales, aparte de las adaptacionesrelacionadas con su modo de vida (predominante-mente planctónico en los primeros, bentónico sésilo incrustante en los segundos), tienen que ver conel polimorfismo tecal tan típico de los graptolitosbentónicos y con el estolón no esclerotizado de lasformas planctónicas. A la distinción relativa a lateca embrionaria del oozooide (sícula), se suma elhecho de que las formas vivientes, con tubos ergui-
8 PALEONTOLOGÍA DE INVERTEBRADOS

dos, tienen anillos fuselares esencialmente comple-tos (360º) y sin suturas (salvo en la parte reptante:Fig. 8.3), en vez de estar formados por dos semia-nillos de 180º como ocurre en la mayoría de losgraptolitos. Internamente, las fibrillas fuselares delos pterobranquios son finas y rectas, mientras queen los graptolitos son más irregulares y enmaraña-das. No obstante, uno de los grupos más raros degraptolitos bentónicos (orden Crustoidea) compar-te con los rabdopléuridos algunos caracteresestructurales, estolonales y el desarrollo de quistesresistentes (graptoblastos, presentes también en elorden Camaroidea), que se desconocen en los res-tantes graptolitos.
Las escasas evidencias directas de zooides grap-tolíticos proceden de un yacimiento delTremadociense de Australia (Rickards & Stait,1984). Los restos fosilizaron en el interior de tecas
adultas y consisten en cuerpos globosos piritizadosde menos de medio milímetro, conectados porestolones, que ocupan una longitud equivalente a20 anillos fuselares, y no conservan los apéndices(Fig. 8.8A). Posteriormente se han descrito otroscasos de conservación de tejidos blandos, princi-palmente estolones de graptoloideos, ditecoideos ysustancias de reserva en graptoblastos de crustoi-deos, pero sin fosilización de los propios zooides.
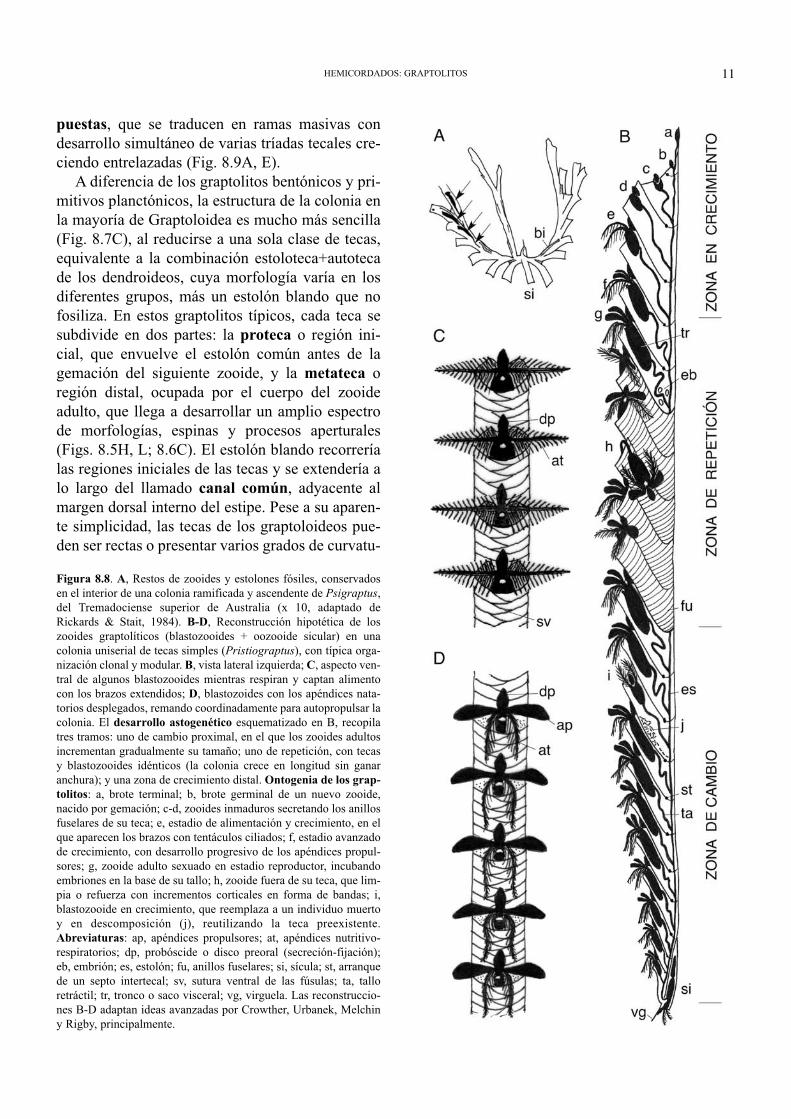
Un cálculo indirecto del tamaño de los zooidesgraptolíticos se basó en comparar las proporcionesdel órgano secretorio y el cuerpo de los rabdopléu-ridos actuales, con relación a la altura de los anillosfuselares y a la anchura de las bandas corticales delos graptolitos (Sudbury, 1991; Rigby & Sudbury,1995). Las dimensiones reconstruidas concuerdancon las de los exiguos fósiles y revelan que la onto-genia de los blastozooides atravesó probablementepor tres estadios: uno inicial de generación delesqueleto, en el que el animal construye una tecadesproporcionadamente grande para su tamaño, ydos posteriores, dedicados a la alimentación, elcrecimiento (doblando o triplicando las dimensio-nes del estadio esqueletal precedente) y, por últi-mo, la reproducción (Fig. 8.8B). En hemicordadosactuales, los zooides desarrollan el disco preoral yconstruyen su tubo antes de concluir el desarrollooperativo del “lofóforo”, dándose la maduraciónsexual en una minoría de blastozooides hermafro-ditas, capaces de incubar de uno a tres embrionesen sus tecas. En graptolitos, se ha sugerido que lafragmentación accidental de los rabdosomas y laregeneración de nuevas colonias a partir de estipesrotas, fue algo habitual dentro del ciclo vital de lasformas planctónicas, además de contar con unareproducción sexual comparable a la de los rabdo-pléuridos (Urbanek, 1990).
Antes de ser asignados a los hemicordados, losgraptolitos habían sido interpretados como fósilesde plantas marinas (siglo XVIII y principios delXIX), briozoos (mediados del s. XIX), cnidarioshidrozoos (desde mediados del s. XIX hastamediados del s. XX) e incluso animales incertaesedis (finales s. XIX a inicios del s. XX). Las pri-meras opiniones aisladas, que resaltaron sus afini-dades con Rhabdopleura, se remontan a la segun-da mitad del siglo XIX, cuando todavía muchos
9HEMICORDADOS: GRAPTOLITOS
Figura 8.7. A, esquema de una abertura tecal con los últimos seisanillos anillos fuselares (cada uno formado por dos fúsulas a-a’y b-b’), y sección parcial de los mismos (grosor exagerado 6 veces). Desu fino recubrimiento cortical interno y externo, se han representa-do algunas bandas corticales (eucórtex punteado). B-C, diferenciasentre los estipes de los graptoloideos primitivos (B, con tríadas teca-les) y las formas planctónicas evolucionadas (C, sin polimorfismotecal). En los esquemas se resaltan las homologías estoloteca-prote-ca (punteado) y autoteca-metateca (trama gris). D-F, esquema de lostipos básicos de desarrollo proximal de la colonia: isográptido (D),artus (E) y diplográptido (F, en este caso el llamado patrón A); lateca dicálica se destaca con puntos. Abreviaturas: a, autoteca; b,biteca; bc, banda cortical; e, estoloteca; es, estolón esclerotizado; p,proteca; t, teca (metateca); s, sícula; so, sutura oblicua entre dosfúsulas; tc, tejido cortical; tf, tejido fuselar.

zoólogos consideraban a dicho género como unbriozoo aberrante. Más tarde se vieron sustancia-das merced a los trabajos de Roman Kozlowskientre 1938 y 1966, y confirmadas sobre todo a par-tir del decenio de 1970, con la realización de estu-dios ultraestructurales mediante técnicas demicroscopía electrónica tanto en graptolitos comoen rabdopleúridos (Crowther, 1981; Rickards etal., 1982; Urbanek & Mierzejewski, 1984; Bates,1987; Mierzejewski & Kulicki, 2003, etc.)
4. ORGANIZACIÓN Y ELEMENTOS DE LACOLONIA
El rabdosoma de los graptolitos tiene una orga-nización clonal y modular que recuerda en ciertomodo a la del zoario de los briozoos (Urbanek,2004). El tamaño de la colonia varía normalmenteentre 2 y 15 cm, aunque el rango para las formasplanctónicas maduras se extiende desde los 0,5-0,8cm (rabdosomas con menos de cuatro zooidesagrupados en torno a la sícula) y los 145 cm de lon-gitud por 3 mm de anchura máxima, como es elcaso del ejemplar de monográptido de un solo esti-pe que ostenta el récord mundial (Fig. 8.6A,Loydell & Loveridge, 2001).
El aspecto del rabdosoma suele ser irregular enlas formas incrustantes y reptantes, aunque los grap-tolitos bentónicos más comunes desarrollan colo-nias erguidas de morfologías cónicas, arbustivas oen abanico (Fig. 8.9). Por su parte, los graptolitosplanctónicos primitivos tienen rabdosomas cónicos,campaniformes o radiados en el plano horizontal(Fig. 8.10A-D, G), en tanto que en las formas másmodernas, la colonia queda reducida a un estipe, detrazado recto a curvado (Fig. 8.12A-S).
La sícula o teca germinal del rabdosoma, habi-tada por el oozooide zigótico, es muy homogéneaestructuralmente y presenta pocos cambios en suforma y dimensiones a lo largo de toda la historiaevolutiva de los graptolitos. Las colonias bentóni-cas tienen una sícula tubular, de cuyo extremo ini-cial parten ramificaciones corticales que a vecescomponen un notorio disco basal de fijación. Enlas formas planctónicas, la sícula es uniforme yestá más elaborada, comprendiendo un extremocónico (prosícula) y otro apertural cilíndrico(metasícula). La prosícula presenta un alarga-
miento apical (cauda), que se halla separado delresto de la estructura (conus) por una membrana(diafragma), la cual desaparece al completarse elcrecimiento de la metasícula. La ornamentaciónprosicular consta de una línea espiral y varias cres-tas longitudinales de refuerzo, que crecen desde elápice hacia la metasícula (Fig. 8.4A-B). En lascolonias planctónicas, el vértice de la sícula seorienta convencionalmente hacia arriba y la caudapuede prolongarse más tarde en una estructurahueca y flexible llamada nema (Figs. 8.4C, 8.5B).El extremo de la colonia que engloba o se sitúacerca de la sícula es la región o parte proximal delrabdosoma; en tanto que el extremo opuesto, aleja-do de la sícula, forma la región distal del mismo.
A partir de la sícula se inicia el verdadero des-arrollo colonial del rabdosoma (astogenia),mediante la incorporación de nuevos individuos(blastozooides), interconectados por el estolón,que brotan por gemación asexual y construyentecas sucesivas. En los graptolitos bentónicos(orden Dendroidea) y primitivos planctónicos (ani-sográptidos: orden Graptoloidea), las tecas sonpolimorfas (Fig. 8.7B), diferenciándose en: estolo-tecas, o tecas portadoras de un estolón internoesclerotizado, que se ramifica en tríadas a medidaque crece la colonia; y autotecas y bitecas, deforma y tamaño algo distintos, interpretadas comocorrespondientes a zooides diferenciados sexual ofuncionalmente (los de las autotecas, relacionadostal vez con la alimentación, y los de las bitecas, conla reproducción o con la limpieza de la colonia).Este crecimiento se traduce generalmente en laconformación de rabdosomas cónicos o arbores-centes de ramificación compleja, en los que losestipes se presentan unidos por disepimentos(barras corticales transversas, a veces portandobitecas: Fig. 8.9F) o por procesos de anastomosis(coalescencia temporal entre estipes adyacentes,con habitual transferencia de tecas: Fig. 8.9A-B), yque llegan a albergar cientos o incluso miles deindividuos. En el caso más habitual, las coloniasbentónicas tienen estipes simples, apreciándoseuna fila única de autotecas rodeada por delgadasbitecas (Fig. 8.7B); sin embargo, algunos gruposde dendroideos (en especial los acantográptidos) yciertos Tuboidea llegan a presentar estipes com-
10 PALEONTOLOGÍA DE INVERTEBRADOS
puestas, que se traducen en ramas masivas condesarrollo simultáneo de varias tríadas tecales cre-ciendo entrelazadas (Fig. 8.9A, E).
A diferencia de los graptolitos bentónicos y pri-mitivos planctónicos, la estructura de la colonia enla mayoría de Graptoloidea es mucho más sencilla(Fig. 8.7C), al reducirse a una sola clase de tecas,equivalente a la combinación estoloteca+autotecade los dendroideos, cuya morfología varía en losdiferentes grupos, más un estolón blando que nofosiliza. En estos graptolitos típicos, cada teca sesubdivide en dos partes: la proteca o región ini-cial, que envuelve el estolón común antes de lagemación del siguiente zooide, y la metateca oregión distal, ocupada por el cuerpo del zooideadulto, que llega a desarrollar un amplio espectrode morfologías, espinas y procesos aperturales(Figs. 8.5H, L; 8.6C). El estolón blando recorreríalas regiones iniciales de las tecas y se extendería alo largo del llamado canal común, adyacente almargen dorsal interno del estipe. Pese a su aparen-te simplicidad, las tecas de los graptoloideos pue-den ser rectas o presentar varios grados de curvatu-
11HEMICORDADOS: GRAPTOLITOS
Figura 8.8. A, Restos de zooides y estolones fósiles, conservadosen el interior de una colonia ramificada y ascendente de Psigraptus,del Tremadociense superior de Australia (x 10, adaptado deRickards & Stait, 1984). B-D, Reconstrucción hipotética de loszooides graptolíticos (blastozooides + oozooide sicular) en unacolonia uniserial de tecas simples (Pristiograptus), con típica orga-nización clonal y modular. B, vista lateral izquierda; C, aspecto ven-tral de algunos blastozooides mientras respiran y captan alimentocon los brazos extendidos; D, blastozoides con los apéndices nata-torios desplegados, remando coordinadamente para autopropulsar lacolonia. El desarrollo astogenético esquematizado en B, recopilatres tramos: uno de cambio proximal, en el que los zooides adultosincrementan gradualmente su tamaño; uno de repetición, con tecasy blastozooides idénticos (la colonia crece en longitud sin ganaranchura); y una zona de crecimiento distal. Ontogenia de los grap-tolitos: a, brote terminal; b, brote germinal de un nuevo zooide,nacido por gemación; c-d, zooides inmaduros secretando los anillosfuselares de su teca; e, estadio de alimentación y crecimiento, en elque aparecen los brazos con tentáculos ciliados; f, estadio avanzadode crecimiento, con desarrollo progresivo de los apéndices propul-sores; g, zooide adulto sexuado en estadio reproductor, incubandoembriones en la base de su tallo; h, zooide fuera de su teca, que lim-pia o refuerza con incrementos corticales en forma de bandas; i,blastozooide en crecimiento, que reemplaza a un individuo muertoy en descomposición (j), reutilizando la teca preexistente.Abreviaturas: ap, apéndices propulsores; at, apéndices nutritivo-respiratorios; dp, probóscide o disco preoral (secreción-fijación);eb, embrión; es, estolón; fu, anillos fuselares; si, sícula; st, arranquede un septo intertecal; sv, sutura ventral de las fúsulas; ta, talloretráctil; tr, tronco o saco visceral; vg, virguela. Las reconstruccio-nes B-D adaptan ideas avanzadas por Crowther, Urbanek, Melchiny Rigby, principalmente.

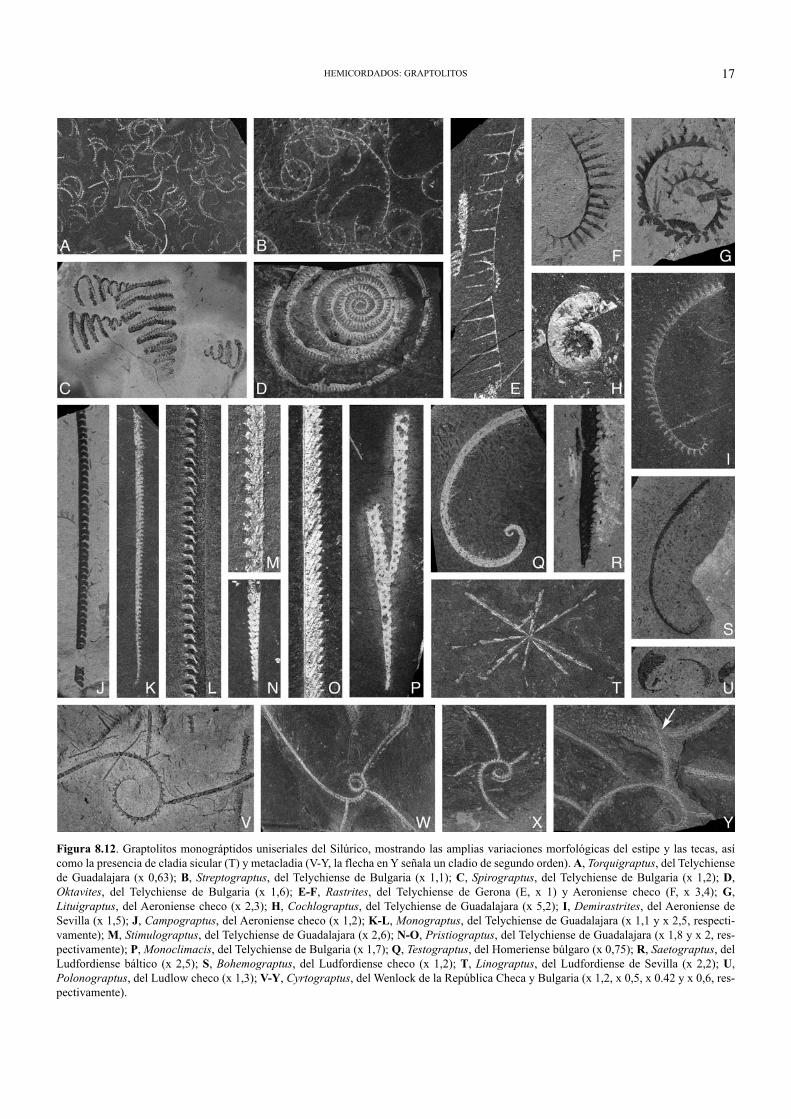
ra (sigmoidea, doble sigmoide con pliegues prote-cales, ganchudas, geniculadas, lobadas, globosas,etc.: Figs. 8.6 y 8.12); expandirse o estrecharseaperturalmente; solaparse con regularidad conotras tecas de su mismo estipe (la separación severifica mediante septos intertecales entre tecasconsecutivas) o bien mostrar metatecas aisladas(Fig. 8.12E-F); tener aberturas orientadas en elsentido de crecimiento (Fig. 8.6L) o en formaretroversa (al revés: Fig. 8.12J-L), introvertida(hacia el interior del estipe) o evertida (perpendi-cular al estipe); desarrollar aberturas digitadas, conpestañas y rebordes espesos, orejas asimétricas,etc.; y, finalmente, generar una gran variedad deespinas tecales (aperturales, geniculares, mesiales,protecales, etc.), orientadas dorsal, ventral o late-ralmente, las cuales forman a veces un entramadoespinoso muy notorio (la lacinia de ciertas formasbiseriales) y en otras ocasiones soportan procesosmembranosos con peridermo fuselar. También unamisma colonia puede desarrollar varios tipos mor-fológicos de tecas a lo largo de su astogenia, comoocurre entre los monográptidos biformes, en cier-tos casos de regeneración, o bien en rabdosomascon estipes secundarias especiales (cladia): Fig.8.12I, V.
Las formas más modernas de Graptoloidea secaracterizan por poseer una o dos series lineales detecas, que crecen y se abren en sentido opuesto a la
sícula (estipes ascendentes, Figs. 8.4D-E; 8.5E-H,K-L; 8.10I, Q-R), variando normalmente entreuniseriales (rabdosoma reducido a un solo estipe:Figs. 8.5K-L; 8.6L; 8.12A-S) y biseriales dipleu-rales (rabdosoma con dos estipes ascendentes, uni-dos dorsalmente y separados -o no- mediante unsepto central: Figs. 8.6F-K; 8.11A-K), aunqueexisten también raras formas cuadriseriales (concuatro estipes ascendentes, unidos completamentepor el dorso o con septos centrales perforados: Fig.8.5F) y biseriales monopleurales (rabdosoma condos estipes ascendentes, pero individualizados yunidos por las caras laterales: Figs. 8.5H; 8.10O-P). Al mismo tiempo, el nema hueco de la sículapuede llegar a prolongarse en una varilla fuselardenominada vírgula, que contribuye a reforzar yguiar el crecimiento de la colonia (Figs. 8.5K;8.11J), y que da lugar en algunas formas biserialesal llamado nematulario (o virgulario), relaciona-do con una de las adaptaciones al modo de vidaplanctónico (Fig. 8.14A). La vírgula se localiza enposición dorsal en las formas uniseriales evolucio-nadas, mientras que en las biseriales permanece enel centro de la colonia o rara vez se adosa a una desus caras laterales. En el extremo opuesto de lacolonia, el borde apertural de la sícula puede gene-rar un dentículo obtuso (rutelo), recrecer distal-mente en forma tubular (parasícula) o bien des-arrollar una virguela o espina ventral, además de
12 PALEONTOLOGÍA DE INVERTEBRADOS
Figura 8.9. Graptolitos bentónicos de los órdenes Tuboidea (A-B) y Dendroidea (C-H), del Ordovícico de España (A-E) y EEUU (F-H). A-B, Reticulograptus (x 1,2), con dos tubotecas de parásitos indicadas en B (flechas); C, Callograptus (x 1,3); D, H, Dendrograptus (x 1 y x1,3, respectivamente); E, Acanthograptus (x 1,2); F, Dictyonema (x 1,7); G, Desmograptus (x 1,3).
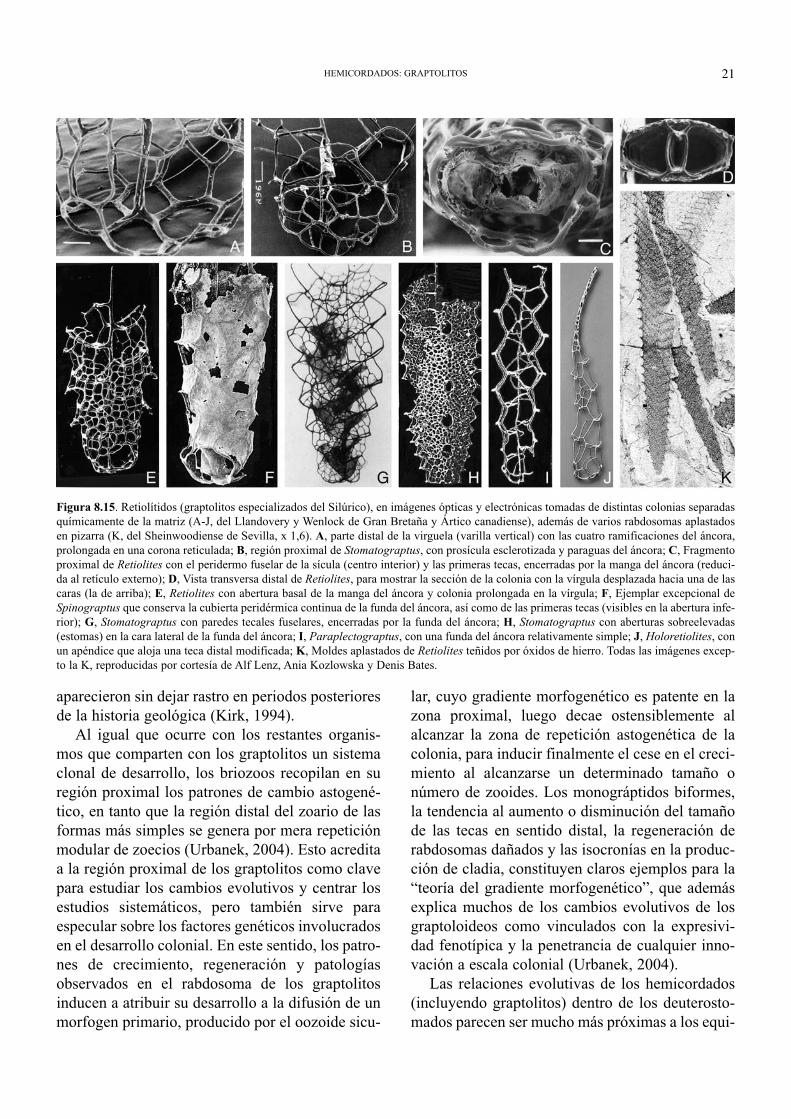
una o varias espinas dorsales (antivirguelares):Figs. 8.4 y 8.5. La virguela rara vez aparece rami-ficada, pero en algunos rabdosomas biseriales suápice distal se bifurca regularmente para formar eláncora, una estructura con cuatro varillas perpen-diculares al extremo de la virguela (Fig. 8.15A-B).
En un grupo especializado de graptolitos (losretiolítidos: Bates et al., 2005), el áncora de la vir-guela se ramifica distalmente para conformar unvarillaje en forma de paraguas invertido (corona oancora-umbrella), que se prolonga en sentidoascendente hasta envolver y rodear completamente
13HEMICORDADOS: GRAPTOLITOS
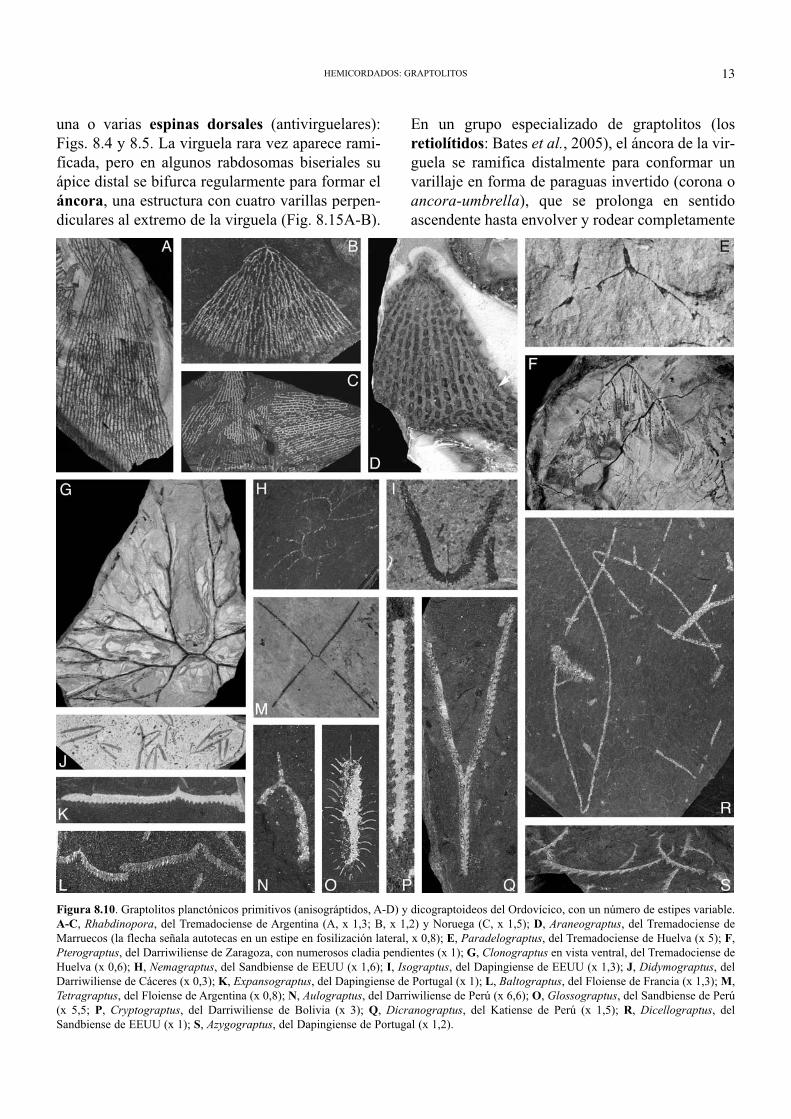
Figura 8.10. Graptolitos planctónicos primitivos (anisográptidos, A-D) y dicograptoideos del Ordovícico, con un número de estipes variable.A-C, Rhabdinopora, del Tremadociense de Argentina (A, x 1,3; B, x 1,2) y Noruega (C, x 1,5); D, Araneograptus, del Tremadociense deMarruecos (la flecha señala autotecas en un estipe en fosilización lateral, x 0,8); E, Paradelograptus, del Tremadociense de Huelva (x 5); F,Pterograptus, del Darriwiliense de Zaragoza, con numerosos cladia pendientes (x 1); G, Clonograptus en vista ventral, del Tremadociense deHuelva (x 0,6); H, Nemagraptus, del Sandbiense de EEUU (x 1,6); I, Isograptus, del Dapingiense de EEUU (x 1,3); J, Didymograptus, delDarriwiliense de Cáceres (x 0,3); K, Expansograptus, del Dapingiense de Portugal (x 1); L, Baltograptus, del Floiense de Francia (x 1,3); M,Tetragraptus, del Floiense de Argentina (x 0,8); N, Aulograptus, del Darriwiliense de Perú (x 6,6); O, Glossograptus, del Sandbiense de Perú(x 5,5; P, Cryptograptus, del Darriwiliense de Bolivia (x 3); Q, Dicranograptus, del Katiense de Perú (x 1,5); R, Dicellograptus, delSandbiense de EEUU (x 1); S, Azygograptus, del Dapingiense de Portugal (x 1,2).

todas las tecas de la colonia, con lo que el rabdoso-ma pasa a tener dos capas con un espacio huecointermedio (Fig. 8.15C-K). La capa externa sellama manga del áncora (ancora sleeve) y consis-te en un entramado complejo de varillas corticales(barras septales, transversas, conectivas, pleurales,geniculares, ventrales, laterales en zigzag, etc.) quellega a soportar un retículo y orificios sobreeleva-dos (estomas). Los fósiles de retiolítidos se limitannormalmente al citado armazón exterior, ya que elperidermo fuselar de las tecas propiamente dichas,y el que envolvía la manga del áncora, eran muytenues, y sólo se conservan en circunstanciasexcepcionales (Fig. 8.15F-G).
Las formas más primitivas de Graptoloideacarecen de vírgula, y de la sícula (generalmente sinvirguela) parten dos estipes primarios que se pue-den ramificar más adelante en varios órdenes dedicotomías, dando lugar a un número variable deestipes terminales (de dos a varias decenas). Lasprimeras tecas del rabdosoma se abren en el mismosentido que la sícula, o bien se orientan en direc-ción oblicua o perpendicular con relación al eje deésta, constituyendo el punto de partida de los lla-mados estipes pendientes (los que “cuelgan” de lasícula: Figs. 8.5B, D, M; 8.10A-D, J, N), declina-dos (divergen con ángulos inferiores a 180º: Figs.8.6E; 8.10E), deflexos (los inicialmente pendienteso declinados, que luego se horizontalizan: Fig.8.10L), horizontales (perpendiculares a la sícula:Fig. 8.10G, M, K), reflexos (cambiantes de recli-nados a horizontales) o reclinados (estipes queascienden desde la sícula sin llegar a tocarse entresí: Figs. 8.5E; 8.10I, R). Entre todas estas categorí-as existen formas compuestas, como rabdosomasde estipes biseriales que cambian a reclinados (Fig.8.10Q: a la parte temporalmente biserial se lallama axil) y ascendentes uni-biseriales (Fig.8.11L-M).
El proceso de ramificación de los estipes escomplejo y se produce por mediación de las llama-das tecas dicálicas, cada una de las cuales originados brotes del estolón, de los que germinan lastecas iniciales de un nuevo par de estipes, tanto enla primera dicotomía (contada a partir de la sícula)como en las restantes (Figs. 8.5J; 8.7D-F). En elcaso de un rabdosoma con varios órdenes de divi-
sión de estipes, las dicotomías pueden ser de tipoconsecutivo, cuando las tecas dicálicas se hallanseparadas por una sola teca normal, o retardado,cuando dos o más tecas unicálicas separan tecasdicálicas sucesivas (en la misma dirección de cre-cimiento). El sistema de notación de las tecas, en elrabdosoma ramificado de un graptoloideo, sueleexpresarse en términos binomiales, por ejemploteca 11 (primera teca formada en el primer estipederivado de la sícula), 21, 31, etc.; teca 12 (primerateca del segundo estipe), 22, 32, etc. A su vez losestipes siguen una notación polinómica a partir dela teca dicalical nn en las dicotomías posteriores ala primera, por ejemplo (nn+1)b, (nn+1)a, (nn+1)1,(nn+1)2..., 31 > 41a y 41b, 41b > 151b y 152b, etc., en laque se combina además si el crecimiento es dere-cho o izquierdo a partir de las tecas dicálicas impli-cadas en las divisiones. La ramificación en generalpuede responder a un estilo progresivo (aquél queda lugar a estipes que se dividen posteriormente),monoprogresivo (sólo se divide uno de los dossurgidos de una dicotomía) o terminal (ninguno delos dos estipes vuelve a dividirse). Finalmente ydependiendo de la divergencia a partir del punto dedivisión, los estipes pueden ramificarse en formadicótoma (con separación simétrica: Fig. 8.10G) olateral (cuando uno de los estipes mantiene ladirección de crecimiento y el otro se separa a unlado: Fig. 8.10F, H).
Un modelo alternativo de ramificación en rab-dosomas de graptoloideos, en el que no intervienenlas tecas dicálicas convencionales, es el que dalugar a las denominadas cladia. Éstos son estipes“secundarios” emanados, tanto de la apertura dealgunas tecas maduras de un estipe previo, comodesde el borde o el interior de la sícula, o inclusosurgidos a partir de fragmentos regenerados decolonias que perdieron la sícula. En algunos rabdo-somas, la emisión de cladia laterales ocurre deforma regular (Figs. 8.10F, H; 8.12V-Y), mientrasque otros presentan cladia sicular única o múltiple(Fig. 8.12T). Alternativamente, ciertas especiestienen la potestad de generar (o inhibir) un solocladio. Asimismo, la producción de cladia, en rab-dosomas uniseriales con vírgula, puede ir asociadaal desarrollo de pseudovírgula en cada cladio cre-ado por las metatecas del estipe principal, un pro-
14 PALEONTOLOGÍA DE INVERTEBRADOS

ceso restringido a taxones silúricos y devónicos. De todos modos, durante la astogenia de los
graptolitos se producen otros muchos procesos quedan lugar a estructuras clave para la taxonomía yclasificación de los graptoloideos, los más impor-tantes de los cuales tienen que ver con el modo dedesarrollo proximal del rabdosoma. Dicho térmi-no expresa la secuencia de crecimiento de las tecasiniciales de la colonia, que se halla relacionadafilogenéticamente en los distintos grupos taxonó-micos (Fortey & Cooper, 1986). Para poder reco-nocer la disposición de los brotes tecales en la pri-mera dicotomía, es necesario orientar el rabdoso-ma por su cara reversa, a fin de observar los “cana-les” de cruce o tubos protecales que se superponena la sícula y a las primeras tecas en cada estipe (lacara opuesta de la colonia -anverso- se distinguebien porque en ella la sícula permanece libredurante la mayor parte de su longitud mesio-dis-tal). Básicamente existen tres tipos de desarrolloproximal, llamados isográptido (con teca 12 dicá-lica y dos o más canales de cruce), artus (con teca11 dicálica y un canal de cruce), y diplográptido
(con teca dicálica 21, 22 o posteriores): Fig. 8.7D-F.El tipo isográptido es el más primitivo por estarrepresentado en las autotecas de los dendroideos,entre los primeros graptoloideos planctónicos (losanisográptidos) y en numerosos dicograptoideosordovícicos, cuyos representantes más profusa-mente ramificados muestran también el mismotipo de división en todas las dicotomías de la colo-nia. Su precedencia temporal revela que a partir deldesarrollo isográptido se originaron los otros dostipos, bien por anticipación (artus) o retardo (diplo-gráptido) en la aparición de la primera teca dicáli-ca, siguiendo procesos que se repitieron separada-mente en varias líneas filogenéticas.
Además de los tres tipos básicos de desarrolloinicial, los graptolitos registran numerosas varia-ciones en la estructura proximal de la colonia,siguiendo patrones de gran relevancia para abordaruna clasificación más natural de los graptoloideos.Dicha estructura se refiere a la orientación, sentidode crecimiento y disposición de las primeras tecasde la colonia: implica factores como el punto ymodo de origen de la teca 11 en la sícula (posición
15HEMICORDADOS: GRAPTOLITOS
Figura 8.11. Diplograptoideos (Diplograptina y Monograptina) de rabdosoma biserial (A-K) y uni-biserial (L-M). A, Gymnograptus, delDarriwiliense de Portugal (x 5); B-C, Climacograptus, del Sandbiense de EEUU y Katiense de China, respectivamente (B, con espesamien-tos corticales de las espinas antivirguelares, x 1,6; C, con parasícula descendente en la virguela, x 4,3); D, Anticostia, del Katiense de China(x 5); E, Pseudamplexograptus, del Darriwiliense de Perú (x 1,2); F, Geniculograptus, del Katiense de EEUU (x 1,9); G, Neodiplograptus,del Aeroniense de Sevilla (x 1,4); H y J, Metaclimacograptus, del Telychiense de Guadalajara (x 6,5) y Aeroniense checo (x 10), respectiva-mente; I, Parapetalolithus, del Telychiense de Guadalajara (x 0,7); K, Petalolithus, del Aeroniense checo (x 3,1); L, Dimorphograptus, delRhuddaniense de Barcelona (x 2,2); M, Parakidograptus, del Rhuddaniense de León (x 1,5).

prosicular o metasicular, brotada por reabsorción oinvaginación del borde sicular, por formación deun poro o lacuna en el peridermo, etc.), la orienta-ción primordial de las tecas (tipos estreptoblástico,prosoblástico, pericálica/monopleural, platicálica,etc.), la geometría inicial de la colonia (involucra,por ejemplo, términos como “simetría isográptida”-de la sícula con la teca 11-, “simetría meandro-gráptida” -de las tecas 11 y 12 en torno a la sícula-,“asimetría sigmagráptida”, manubrio, etc.), creci-mientos dextrales (sentido horario) y sinistrales(antihorario), etc. El caso paradigmático de aplica-ción de los conocimientos derivados de la estructu-ra proximal es la nueva clasificación de los diplo-graptoideos (= “Virgellina”; también llamadosaxonóforos), que reúnen formas extraordinaria-mente variadas con rabdosomas biseriales, unise-riales, biseriales-declinados, uni-biseriales, birra-mosos horizontales a reclinados, con cladia, conáncora, etc. Hasta hace poco, la clasificación de losdiplograptoideos atendía a patrones morfológicosgenerales del rabdosoma y las tecas (Bulman,1970), los cuales desembocaron en agrupamientostaxonómicos sin correspondencia filogenética. Losnuevos criterios aplicados al desarrollo proximaldiplográptido han conducido al establecimiento decatorce diseños básicos de estructura proximal, quecontribuyen decisivamente a la distinción cladísti-ca de varias superfamilias reunidas en los subórde-nes tradicionales Diplograptina y Monograptina,que aglutinan a buena parte de los graptolitosplanctónicos ordovícico-devónicos (Mitchell,1987; Melchin, 1998; Fortey et al., 2005; Mitchellet al., 2007; Maletz et al., 2009). Estas catorcepautas en la astogenia primordial se nombran porletras (A-M, R, U), diagnosticándose cada tipo enfunción del origen de la teca 12, la ontogenia inicialde las tecas 21 y 22, el nivel de aparición de la tecadicálica dentro de cada patrón de diseño y detallessiculares concretos.
Los nuevos avances en el conocimiento del des-arrollo y estructura proximal del rabdosoma de losgraptolitos exigen, para poder completar la clasifi-cación, disponer de un material muy bien conser-vado de los taxones que restan aún por conocerseen detalle. Es por ello por lo que se ha intensifica-do la búsqueda de material piritizado o susceptible
de preparación química, bien por remoción delperidermo cortical mediante soluciones ácidashasta hacerlo transparente, o bien por el empleo, enejemplares opacos, de técnicas de videomicrosco-pía por infrarrojos (Fig. 8.4E) y de microscopía debarrido con retrodispersión electrónica.
5. CLASIFICACIÓN
La clase Graptolithina reúne a más de 3000especies, que se agrupan en unos 300 géneros y enmedio centenar de familias o subfamilias. La clasi-ficación presentada en la última edición delTreatise (Bulman, 1970) ha quedado obsoleta, ymuchos géneros atribuidos en ella a graptolitos,además del antiguo “orden Stolonoidea”, se repar-ten ahora entre rabdopléuridos, algas e hidrozoos(Rickards & Durman, 2006). Los graptolitosauténticos se dividen en seis órdenes de los cualessólo dos, Dendroidea y Graptoloidea, alcanzannotoriedad en el registro fósil. Cabe la posibilidadde que en el futuro se opte por elevar cada uno delos órdenes al rango de clase o subclase, y queincluso la mayoría de los subórdenes aquí citadosalcance el rango de orden (Mitchell et al., 2007).El análisis cladístico moderno de los graptolitosplanctónicos (Maletz et al., 2009) los integra bási-camente en una Supercohorte Eugraptoloida, com-puesta por dos cohortes (Pan-Sinograpta y Pan-Reclinata), la segunda de las cuales reúne a lamayoría de las formas del Ordovícico Medio alDevónico Inferior. De todos modos y como la sis-temática de los graptolitos dista de estar resuelta, acontinuación se expone la morfología y caracteresdistintivos de una serie de grupos básicos, rete-niéndolos provisionalmente como órdenes tradi-cionales de la clase Graptolithina.
5.1. ORDEN CRUSTOIDEA (Ordovícico Inferior-Silúricosuperior). Colonias incrustantes con tríadas tecales; bitecas yestolotecas restringidas a la porción incrustante del rabdoso-ma; autotecas vesiculares, con suturas en zigzag en la partesuperior reptante, previa a unas aberturas erectas de morfolo-gía distintiva, con cuello y pliegues lobulares (Fig. 8.13B).Prosícula vesicular. Desarrollo de quistes resistentes (grapto-blastos). Peridermo fuselar heteromórfico y homeomórficocon respecto al córtex; estructuras fibrilares polimórficas.
16 PALEONTOLOGÍA DE INVERTEBRADOS
17HEMICORDADOS: GRAPTOLITOS
Figura 8.12. Graptolitos monográptidos uniseriales del Silúrico, mostrando las amplias variaciones morfológicas del estipe y las tecas, asícomo la presencia de cladia sicular (T) y metacladia (V-Y, la flecha en Y señala un cladio de segundo orden). A, Torquigraptus, del Telychiensede Guadalajara (x 0,63); B, Streptograptus, del Telychiense de Bulgaria (x 1,1); C, Spirograptus, del Telychiense de Bulgaria (x 1,2); D,Oktavites, del Telychiense de Bulgaria (x 1,6); E-F, Rastrites, del Telychiense de Gerona (E, x 1) y Aeroniense checo (F, x 3,4); G,Lituigraptus, del Aeroniense checo (x 2,3); H, Cochlograptus, del Telychiense de Guadalajara (x 5,2); I, Demirastrites, del Aeroniense deSevilla (x 1,5); J, Campograptus, del Aeroniense checo (x 1,2); K-L, Monograptus, del Telychiense de Guadalajara (x 1,1 y x 2,5, respecti-vamente); M, Stimulograptus, del Telychiense de Guadalajara (x 2,6); N-O, Pristiograptus, del Telychiense de Guadalajara (x 1,8 y x 2, res-pectivamente); P, Monoclimacis, del Telychiense de Bulgaria (x 1,7); Q, Testograptus, del Homeriense búlgaro (x 0,75); R, Saetograptus, delLudfordiense báltico (x 2,5); S, Bohemograptus, del Ludfordiense checo (x 1,2); T, Linograptus, del Ludfordiense de Sevilla (x 2,2); U,Polonograptus, del Ludlow checo (x 1,3); V-Y, Cyrtograptus, del Wenlock de la República Checa y Bulgaria (x 1,2, x 0,5, x 0.42 y x 0,6, res-pectivamente).

5.2. ORDEN CAMAROIDEA (Ordovícico Inferior-Medio). Rabdosoma incrustante con desarrollo en díadas;hay grandes autotecas vesiculares (cámaras) acabadas entubos erectos (fustes o collumae) de abertura simple, y esto-lotecas tubulares que forman una malla bifurcada por encimade las cámaras autotecales (Fig. 8.13A). Ocasionalmente pre-sentan bitecas pequeñas, restringidas a la parte incrustante yembebidas en tejido extra-autotecal. Estolón esclerotizadoincorporado a la pared externa. Presencia rara de graptoblas-tos u otras estructuras de enquistamiento. Peridermo fuselarheteromórfico.
5.3. ORDEN TUBOIDEA (Cámbrico superior-Silúricosuperior). Graptolitos sésiles, con colonias erguidas deaspecto dendroideo y carentes de tallo, o bien incrustantescon disco basal (tecorriza) y autotecas erectas.Ramificaciones tecales espaciadas irregularmente, con divi-sión del estolón en díadas y polimorfismo acusado, com-prendiendo autotecas helicoidales o modificadas (microte-cas, tecas umbeliformes), bitecas y, de forma esporádica,conotecas (tecas grandes en forma de embudo invertido,exclusivas de este orden). Estipes uniseriales irregulares ode tipo compuesto, con estolón interno esclerotizado.Prosícula vesicular con línea helicoidal; metasícula fuselar.Peridermo fuselar heteromórfico con relación al cortical. Elgénero Reticulograptus, de aspecto dendroideo y estipescompuestas anastomosadas, es común en el OrdovícicoSuperior de España (Fig. 8.9A-B).
5.4. ORDEN DITHECOIDEA (Cámbrico medio-Silúricoinferior). Colonias erectas, formadas por una hilera uniaxialo ramificada de tecas cónicas alargadas o tubulares, con pro-bable desarrollo en díadas (estoloteca axial y autoteca aisla-da). Estipes sin orientación dorsoventral definida, con tecasinsertas irregularmente. Disco basal y tallo reconocible enalgunas formas. Presentan estolón esclerotizado. Anillosfuselares irregulares, con sutura en zigzag poco patente. Elregistro español del Cámbrico medio comprende diversasespecies de los géneros Ovetograptus, Tarnagraptus,Sotograptus y Archaeolafoea (Fig. 8.13C-D). Las formasavanzadas de ditecoideos, diferenciadas por algunos autorescomo un orden aparte (“Mastigograptida”) desarrollan tría-das tecales (hay un dimorfismo incipiente, con bitecas peque-ñas y estolotecas con envueltas corticales), hábito dendroideocon ramificación irregular y sícula cilíndrica. El peridermofuselar y cortical es heteromórfico.
5.5. ORDEN DENDROIDEA (Cámbrico medio-Carbonífero superior). Comprende graptolitos bentónicossésiles, con sícula cilíndrica, estructuras de fijación, tallo yrabdosoma profusamente ramificado, con estipes que puedenpresentarse unidos mediante disepimentos transversos o pro-cesos de anastomosis. Tecas polimorfas de pequeño tamaño,con típico desarrollo en tríadas (autoteca, biteca, estoloteca)y alineadas en estipes simples o compuestos. Estolón internoesclerotizado.
El Orden Dendroidea se divide en cinco familias, desta-cando los Dendrograptidae (Cámbrico medio-Carbonífero),que agrupan las formas dendroideas más típicas, de rabdoso-ma cónico o flabeliforme aplanado (Dendrograptus,Callograptus, Aspidograptus, Dictyonema: Fig. 8.9C-D, F-H); y los Acanthograptidae (Cámbrico Superior-Devónicomedio), de estipes compuestos, ramificados o anastomosa-dos irregularmente, con tecas cilíndricas muy alargadas yaberturas evertidas en grupos aislados (Acanthograptus: Fig.8.9E).
5.6. ORDEN GRAPTOLOIDEA (Ordovícico Inferior-Devónico Inferior). A diferencia de los graptolitos bentóni-cos, el rabdosoma de los graptoloideos se compone de pocosestipes, que van disminuyendo progresivamente en númeroen el transcurso de la evolución, hasta quedar reducidos a unúnico estipe ascendente. La sícula es cónica y se prolonga enel nema. Sólo existe un tipo de tecas, que pueden presentargran diversidad morfológica. Todos los graptoloideos forma-ron colonias planctónicas y comprenden cinco subórdenes,cuya caracterización filogenética aún no ha sido aclarada aldetalle.
El suborden Anisograptina (Ordovícico Inferior) es elmás primitivo de todos los graptoloideos e incluye la impor-tante familia de los anisográptidos, en la que se verifica latransición evolutiva de los dendroideos a los graptoloideos.Sus representantes del Tremadociense inferior conservan eldesarrollo en tríadas tecales, pero gradualmente van perdien-do las bitecas y en las formas más evolucionadas éstas se res-tringen a las autotecas iniciales y, finalmente, a la sícula. Elgénero Rhabdinopora (Fig. 8.10A-C) tiene un rabdosoma
18 PALEONTOLOGÍA DE INVERTEBRADOS
Figura 8.13. Graptolitos bentónicos de los órdenes Camaroidea (A,Bithecocamara, x 48), Crustoidea (B, Bulmanicrusta, x 30) yDithecoidea (C, Tarnagraptus; D, Sotograptus; ambos x 3). A y B,del Tremadociense de Polonia; C y D, del Cámbrico medio deAsturias. Abreviaturas: ea, estolón de la autoteca; eb, estolón de labiteca.

ramificado y con disepimentos, muy parecido al del dendro-gráptido Dictyonema (Fig. 8.9F), por lo que en un principiose pensó que este último reunía especies bentónicas y planc-tónicas. Sin embargo, las primitivas formas planctónicas fue-ron transferidas al grupo de los anisográptidos, en virtud desu sícula cónica provista de nema (Fig. 8.10B), y a la tenden-cia de algunas subespecies de Rhabdinopora flabelliformis aformar sinrabdosomas. Otros géneros importantes sonAnisograptus, Adelograptus, Kiaerograptus y Araneograptus(Fig. 8.10D).
El suborden Didymograptina (= “Dichograptina”) reúnediversas familias de dicograptoideos ordovícicos, sin bitecasy con un número variable de estipes. La abertura de la sículasuele prolongarse en un rutelo y de modo ocasional generauna virguela. Las tecas suelen ser simples tubos de aberturanormal a denticulada, excepto en una familia (Sinograptidae)que desarrolla pliegues protecales y metatecales. Desarrolloproximal de los tipos isográptido y artus. El suborden com-prende formas inicialmente birradiadas (con dos estipes deprimer orden), susceptibles de ramificarse luego y componerrabdosomas con estipes horizontales, pendientes, declinados,ascendentes o reclinados, cuyo número varía de algunasdecenas de estipes a uno solo (Figs. 8.5B, D-F, M; 8.6E;8.10E-G, J-N, S). De los géneros representados en España,cabe mencionar Didymograptus, Aulograptus, Tetragraptus,Pterograptus, Acrograptus, Paradelograptus, Clonograptusy Azygograptus, entre otros.
El suborden Glossograptina (Ordovícico) incluye for-mas birradiadas reclinadas a ascendentes, e incluso rabdoso-mas biseriales monopleurales con vírgula y lacinia.Desarrollo proximal y simetría isográptidos. Tecas simples(Isograptus, Oncograptus), con pequeñas modificacionesaperturales y espinas notorias en géneros típicos (Glosso-graptus, Cryptograptus) (Figs. 8.5H; 8.6I, O, P).
El suborden Diplograptina (Ordovícico Medio-Superior)aglutina diplograptoideos de rabdosoma birramoso, biserial ouniserial, cuya sícula presenta virguela y el nema suele pro-longarse en una vírgula, a veces proyectada en el extremodistal de la colonia. Las tecas alcanzan gran variación morfo-lógica (simples, geniculadas, climacográptidas, pseudoclima-cográptidas, dicranográptidas, lasiográptidas, etc.). Incluyenumerosos géneros agrupados en tres superfamilias, diferen-ciadas con criterios fundados en la astogenia inicial:Climacograptoidea (que incluye formas de tecas muy elabo-radas como Pseudoclimacograptus, Haddingograptus,Climacograptus, etc.: Fig. 8.11B-C), Dicranograptoidea (conrabdosomas singulares como los de Dicranograptus,Dicellograptus y Nemagraptus: Figs. 8.6C; 8.10H, Q-R) yDiplograptoidea, que reúne formas de tecas relativamentesimples (Orthograptus, Rectograptus, Hustedograptus,Paraorthograptus, Eoglyptograptus) a espinosas (Gymno-graptus, Lasiograptus): (Figs. 8.4E; 8.5G; 8.6K; 8.11A, D-F), así como los “arquirretiolítidos” del Ordovícico, con peri-dermo atenuado y retículo, pero carentes de áncora y por lotanto del rabdosoma de doble capa propio de los verdaderosretiolítidos.
Finalmente, el suborden Monograptina (OrdovícicoMedio-Devónico Inferior) comprende diplograptoideos derabdosoma biserial dipleural, como los normalográptidosordovícico-silúricos (entre otros, Normalograptus,Undulograptus, Oelandograptus, Metaclimacograptus,Neodiplograptus, Cystograptus: Figs.8. 6B, G, I; 8.11G, H,J), los petalolítidos (Petalolithus, Parapetalolithus,Glyptograptus, etc.: Figs. 8.11I, K; 8.14A) y los auténticosretiolítidos (Retiolites, Stomatograptus, Gothograptus,Paraplectograptus, etc.: Figs. 8.6D; 8.15A-K). También losdimorfográptidos, con rabdosoma uni-biserial (génerosDimorphograptus, Parakidograptus, Akidograptus: Fig.8.11L-M), y los conocidos monográptidos, de rabdosomasuniseriales y gran variedad de formas (Monograptus,Rastrites, Monoclimacis, Pristiograptus, Oktavites,Spirograptus, Torquigraptus, etc.: Figs. 8.5A, C, K, L;8.6A, L; 8.12A-S, U), con cladia prominentes en ciertosgéneros (Cyrtograptus, Linograptus, Abiesgraptus, etc.: Fig.8.12T-Y).
Desde el punto de vista de la nueva clasifica-ción filogenética propuesta por Maletz et al.(2009), la inmensa mayoría de los graptoloideos seagrupa en el clado “reclinátidos” y en el nuevo“Orden Bireclinata”, que incluye los subórdenes“Pan-Glossograpta” (los glossograptinos clásicos)y “Pan-Axonophora” (diplograptinos + monograp-tinos).
19HEMICORDADOS: GRAPTOLITOS
Figura 8.14. A, colonia biserial de Parapetalolithus con nematula-rio de gran longitud, Telychiense de Checa, Guadalajara (x 1,3); B-D, Sinrabdosomas de Colonograptus, Homeriense de Salas de laRibera, León (x 2, x 1,5 y x 1,4, respectivamente).

6. EVOLUCIÓN
Existen numerosas evidencias de que los ptero-branquios rabdopléuridos fueron los antecesoresdirectos de los graptolitos, tal y como lo describenUrbanek (1986), Urbanek & Dilly (2000) yRickards & Durman (2006). Los graptolitos másantiguos aparecen ya diversificados en elCámbrico medio y son todos bentónicos. A partirdel Cámbrico superior, se inicia el desarrollo de losdendroideos que, sin alcanzar nunca importancianumérica, mantuvieron una gran uniformidadestructural hasta su desaparición a comienzos delCarbonífero superior. En el Tremadociense basalsurgen los graptoloideos planctónicos, cuyosrepresentantes más primitivos y difundidos sonmorfológicamente idénticos a los dendrográptidossésiles, excepto por la posesión de un nema quefacilitó su adaptación a la vida holoplanctónica. Lahistoria evolutiva posterior de los graptoloideos secaracteriza por una tendencia general hacia el per-feccionamiento en los mecanismos de flotabilidady autopropulsión de las colonias, lo cual involucrauna acusada simplificación estructural y la reduc-ción del número de individuos. El polimorfismotecal característico de los dendroideos desaparece,y el número de estipes disminuye progresivamentepara, al final, quedar reducido a uno solo (rabdoso-ma uniserial).
Estos cambios evolutivos se ven claramentereflejados por el registro estratigráfico de los grap-toloideos (las conocidas “faunas” anisográptida,dicográptida, diplográptida y monográptida deBulman, reexaminadas para el Ordovícico porChen et al., 2006). Así, los anisográptidos (o“graptodendroideos”) del Tremadociense inferiorpasan, de tener cuatro estipes primarios (condiciónproximal cuadrirradiada) a tres (idem. trirradiada),y finalmente a dos estipes de primer orden. De unmodo comparable, los rabdosomas birradiados delos anisográptidos más evolucionados, y de sussucesores los dicograptoideos cambian, a lo largodel Ordovícico Inferior y Medio, de tener variosórdenes de dicotomías y bastantes estipes (hori-zontales a pendientes), a un predominio de colo-nias con pocas ramas, reducidas finalmente a dosestipes terminales (o incluso a uno solo sin vírgu-la). En el Ordovícico Medio aparecen las formas
con rabdosoma biserial, que reemplazan a todas lasanteriores en el Ordovícico Superior y declinanpaulatinamente en el Silúrico (Llandovery), salvoun grupo, los retiolítidos, que pervive hasta elSilúrico superior. De los normalograptáceos ordo-vícicos con rabdosoma biserial, evolucionan, acomienzos del Silúrico, los monográptidos unise-riales, que alcanzaron un éxito adaptativo inusita-do durante ese periodo y perduran hasta finales delDevónico inferior, cuando se verifica la abruptadesaparición de todos los graptolitos planctónicos.
La historia evolutiva de los graptoloideos tam-bién estuvo jalonada de acontecimientos de extin-ción a escala global, relacionados probablementecon cambios climáticos u oceanográficos mundia-les (Melchin et al., 1998; Chen et al., 2006; Finneyet al., 2007). La glaciación de finales delOrdovícico trajo consigo una drástica extinciónentre los graptoloideos (sólo sobrevivieron doceespecies); el llamado Evento Lundgreni delWenlock superior dejó las formas uniseriales redu-cidas a dos especies (acompañadas por algunosretiolítidos que no acusaron la crisis y cuatro taxo-nes Lázaro), y al Evento Transgrediens (límiteSilúrico/Devónico) únicamente sobrevivieron tresespecies. Tan sólo en el Silúrico se sucedieron diezeventos de extinción a escala global entre los grap-tolitos planctónicos (Fig. 8.17). Su brusca desapa-rición del registro, en el Devónico Inferior, nocuenta con una explicación única, aunque proba-blemente la alteración en las corrientes oceánicas,vinculada con la colisión caledónica, pudo influirdecisivamente en la extinción de las escasas espe-cies devónicas. Éstas se encontrarían confinadas aunos biotopos concretos, en donde los graptolitosse habían refugiado tras las sucesivas extincionesdel Silúrico superior, pero ya en abierta competen-cia con nuevos y florecientes grupos pelágicos(dacrioconáridos, pterópodos...). En tales condi-ciones adversas, los graptolitos no lograron gene-rar una nueva radiación adaptativa que favorecierala recuperación evolutiva de todo el grupo. Hayautores que piensan que los graptolitos planctóni-cos no se extinguieron en el Devónico, sino queevolucionaron hacia formas carentes de esqueletoesclerotizado, desconocidas como fósiles (salvo undudoso ejemplar de Mazon Creek), las cuales des-
20 PALEONTOLOGÍA DE INVERTEBRADOS
aparecieron sin dejar rastro en periodos posterioresde la historia geológica (Kirk, 1994).
Al igual que ocurre con los restantes organis-mos que comparten con los graptolitos un sistemaclonal de desarrollo, los briozoos recopilan en suregión proximal los patrones de cambio astogené-tico, en tanto que la región distal del zoario de lasformas más simples se genera por mera repeticiónmodular de zoecios (Urbanek, 2004). Esto acreditaa la región proximal de los graptolitos como clavepara estudiar los cambios evolutivos y centrar losestudios sistemáticos, pero también sirve paraespecular sobre los factores genéticos involucradosen el desarrollo colonial. En este sentido, los patro-nes de crecimiento, regeneración y patologíasobservados en el rabdosoma de los graptolitosinducen a atribuir su desarrollo a la difusión de unmorfogen primario, producido por el oozoide sicu-
lar, cuyo gradiente morfogenético es patente en lazona proximal, luego decae ostensiblemente alalcanzar la zona de repetición astogenética de lacolonia, para inducir finalmente el cese en el creci-miento al alcanzarse un determinado tamaño onúmero de zooides. Los monográptidos biformes,la tendencia al aumento o disminución del tamañode las tecas en sentido distal, la regeneración derabdosomas dañados y las isocronías en la produc-ción de cladia, constituyen claros ejemplos para la“teoría del gradiente morfogenético”, que ademásexplica muchos de los cambios evolutivos de losgraptoloideos como vinculados con la expresivi-dad fenotípica y la penetrancia de cualquier inno-vación a escala colonial (Urbanek, 2004).
Las relaciones evolutivas de los hemicordados(incluyendo graptolitos) dentro de los deuterosto-mados parecen ser mucho más próximas a los equi-
21HEMICORDADOS: GRAPTOLITOS
Figura 8.15. Retiolítidos (graptolitos especializados del Silúrico), en imágenes ópticas y electrónicas tomadas de distintas colonias separadasquímicamente de la matriz (A-J, del Llandovery y Wenlock de Gran Bretaña y Ártico canadiense), además de varios rabdosomas aplastadosen pizarra (K, del Sheinwoodiense de Sevilla, x 1,6). A, parte distal de la virguela (varilla vertical) con las cuatro ramificaciones del áncora,prolongada en una corona reticulada; B, región proximal de Stomatograptus, con prosícula esclerotizada y paraguas del áncora; C, Fragmentoproximal de Retiolites con el peridermo fuselar de la sícula (centro interior) y las primeras tecas, encerradas por la manga del áncora (reduci-da al retículo externo); D, Vista transversa distal de Retiolites, para mostrar la sección de la colonia con la vírgula desplazada hacia una de lascaras (la de arriba); E, Retiolites con abertura basal de la manga del áncora y colonia prolongada en la vírgula; F, Ejemplar excepcional deSpinograptus que conserva la cubierta peridérmica continua de la funda del áncora, así como de las primeras tecas (visibles en la abertura infe-rior); G, Stomatograptus con paredes tecales fuselares, encerradas por la funda del áncora; H, Stomatograptus con aberturas sobreelevadas(estomas) en la cara lateral de la funda del áncora; I, Paraplectograptus, con una funda del áncora relativamente simple; J, Holoretiolites, conun apéndice que aloja una teca distal modificada; K, Moldes aplastados de Retiolites teñidos por óxidos de hierro. Todas las imágenes excep-to la K, reproducidas por cortesía de Alf Lenz, Ania Kozlowska y Denis Bates.

nodermos que a los cordados. En este sentido, losfilos Hemichordata y Echinodermata han sido reu-nidos cladísticamente en el término Ambulacraria,debido a las semejanzas en el desarrollo larvario(ambos poseen larvas tornaria), y a los datos apor-tados por la genética molecular (Sato et al., 2008).
7. PALEOECOLOGÍA
Todos los graptolitos dendroideos eran bentóni-cos y se fijaban al sustrato mediante el disco basalde la sícula. Éste era reforzado más tarde pornumerosas capas secundarias de tejido cortical,que llegaban a envolver toda la región proximal dela colonia, hasta generar un sólido anclaje del quepartía el tallo masivo, soporte de las restantes rami-ficaciones. Los modelos geométricos y los experi-mentos de flujo realizados sobre graptolitos den-droideos indican que el patrón morfológico delrabdosoma (cónico, arbustivo, en abanico, laminarirregular) representa adaptaciones a diferentesregímenes de paleocorrientes. En las colonias cóni-cas, por ejemplo, el flujo lateral generaría corrien-tes aferentes que saldrían por la parte superior delrabdosoma (Melchin & Doucet, 1996), introdu-ciendo nutrientes en el interior de una coloniacuyas fenéstrulas (huecos entre estipes) estaríanocupadas por los “lofóforos” extendidos de loszooides de las autotecas, los cuales se dispondríansiguiendo un patrón próximo-distal hasta generaruna superficie continua de filtrado (Starcher &McGhee, 2003).
Los graptolitos camaroideos, crustoideos ytuboideos son formas bentónicas incrustantes yreptantes, todo lo más con los extremos distales delas autotecas erguidos (Fig. 8.13A-B). Tan sóloalgunas especies del tuboideo Reticulograptus des-arrollan rabdosomas laminares erectos de ciertaentidad, con estipes profusamente anastomosados(Fig. 8.9A-B), y un modo de vida similar al de losdendroideos. Los ditecoideos constituyen coloniasbentónicas erguidas de ramas simples y poco onada divididas, de hábito semejante al de ciertasalgas e hidrozoos.
A diferencia de todos los grupos precedentes,los graptolitos típicos (orden Graptoloidea) esta-ban adaptados a una vida holoplanctónica, atesti-
guada por la distribución cosmopolita de muchasespecies y su relativa independencia de las faciessedimentarias. Hasta hace pocos años (y pervivien-do aún en dioramas y libros generales), se pensabaque todos los graptoloideos desarrollaban estructu-ras extratecales de flotación, rellenas de gas olíquido hidrostático, diseñadas bien como vejigasindividuales para cada colonia (nematulario, virgu-lario, virguelario), o bien como un flotador colecti-vo (“neumatóforo”) al que se adherían decenas decolonias por su vírgula o nema (sinrabdosomas).Sin embargo, tales interpretaciones han quedadodescartadas tras la revisión de los fósiles quesupuestamente apoyaban ambas hipótesis, asícomo por la realización de estudios ultraestructura-les. Los resultados confirmaron que los flotadoresno eran tales (Fig. 8.14): el nematulario no es unavesícula cerrada, sino láminas aplanadas de tejidofuselar, que irradian longitudinalmente en sustitu-ción de la vírgula, formando una triple o doblepaleta hidráulica (Rickards, 1996); en tanto que los“neumatóforos” de los sinrabdosomas (no los sin-rabdosomas en sí), más las supuestas cámarasincubadoras de sículas (“gonotecas”), resultaronser estructuras puramente imaginarias (Gutiérrez-Marco & Lenz, 1998).
La tendencia actual es considerar a los graptoli-tos como organismos clonales sencillos, cuyosmódulos (blastozooides) comparten el mismogenotipo que el zooide sicular (oozooide), e inte-graban “superindividuos” (los rabdosomas), capa-ces de realizar de forma coordinada y asumir aescala colonial diversas funciones, tales como laalimentación o la motricidad requerida para la auto-propulsión. La integración entre los zooides secanalizaría a través del tejido blando, homólogo delestolón esclerotizado de las formas sésiles (en estecaso sin envuelta peridérmica y flexible), que inter-conectaría entre sí todos los individuos de la colo-nia y contribuiría a la difusión del gradiente morfo-genético en la astogenia. Cada zooide dispondría deun “lofóforo” bipinado, formado por dos brazostentaculados y ciliados, dirigidos lateralmente, quecubriría un dominio elíptico perpendicular al eje delestipe (Fig. 8.8C). Con sus “lofóforos”, los indivi-duos coloniales batirían rítmicamente el agua, en unmovimiento de respiración y filtrado, para atrapar y
22 PALEONTOLOGÍA DE INVERTEBRADOS

encaminar a la boca las partículas alimenticias(seguramente nanoplancton, bacterias de aguassomeras a nitratorreductoras mesopelágicas, ydetritos orgánicos). Además de los brazos tentacu-lados, los zooides debían de poseer otros apéndicescarnosos a modo de remos (Fig. 8.8D), formadospor extensiones musculares en forma de pala que,movidos coordinadamente, lograrían propulsar lacolonia en la columna de agua (Melchin &DeMont, 1995). Por analogía con los hemicordadosactuales, la reconstrucción del zooide graptolíticose completa con la presencia de una probóscide pre-oral (Fig. 8.8B-D), con la que los individuos secre-tarían el esqueleto colonial y podrían reptar fuera desus tecas, realizando una serie de actividades queoptimizaran la alimentación (por ejemplo, treparpor las espinas hasta alcanzar zonas más energéti-cas donde extender los brazos tentaculados).
La capacidad de movimiento autónomo de lascolonias de graptolitos queda patente por el enro-llamiento espiral de muchos rabdosomas formadospor uno o más estipes, siempre con un leve a mar-cado componente helicoidal (Figs. 8.5L; 8.10H, R;8.12A-D, G). La flotabilidad de las colonias estaríaasegurada por el movimiento coordinado de loslóbulos carnosos de todos los zooides, en tanto quepara retardar el descenso o lograr una flotabilidadpasiva, los graptolitos desarrollaron una ampliagama de estructuras tendentes a incrementar lafricción en la masa de agua, tales como espinaslaterales y tecales (Figs. 8.5L; 8.6C; 8.10O;8.12M), láminas corticales desplegadas entre esti-pes proximales, etc. La experimentación hidrodi-námica y aerodinámica en modelos de rabdosomasbiseriales y uniseriales, ha mostrado que las colo-nias adultas se orientarían paralelas a la direccióndel flujo, con la abertura de la sícula enfrentada almismo (Rickards et al., 1998; Rantell & Rigby,2002). Las colonias juveniles resultarían establesverticalmente, con la abertura de la sícula haciaabajo, en tanto que la función del nema (vírgula) enlos rabdosomas adultos de estipes ascendentessería orientar y estabilizar la colonia frente a lascorrientes. Los nematularios de tres paletas y losespirales influirían además en la rotación de lacolonia, incrementando la capacidad de sus zooi-des para captar nutrientes.
A un nivel general, los patrones morfológicosde los graptolitos planctónicos responderían acinco arquetipos básicos (Underwood, 1993): “redde arrastre” (rabdosomas cónicos o turriculados:Figs. 8.5L; 8.10A-D; 8.12C); “red de deriva” (rab-dosomas con muchos estipes horizontales o enespiral rebajada: Figs. 8.10G-H; 8.12D, G, T-Y);colonias de uno o dos estipes curvos, contorsiona-dos y capaces de rotar sobre su eje (Figs. 8.10R;8.12A, F, I, Q, S); colonias simétricas bilateral-mente, con dos estipes divergentes, tal vez no dis-puestos en el mismo plano pero aún así con escasarotabilidad (Figs. 8.5A, M; 8.10I, Q); y, finalmen-te, colonias biseriales y uniseriales rectilíneas, queascenderían en la vertical con la sícula hacia arri-ba, para luego alimentarse en el descenso, con lacolonia oblicua u horizontal, pudiendo eventual-mente rotar sobre su eje (Figs. 8.6B, F-L; 8.11;8.12K-P, R). Del conjunto de estos arquetipos deri-van una serie de ecomorfotipos que, a lo largo dela evolución de las formas planctónicas, desapare-cen en unos grupos y vuelven a surgir en otros, yrepresentan ejemplos de convergencia morfológicaen relación con adaptaciones hidrodinámicas adeterminados nichos, en biotopos planctónicosdiversos (Underwood, 1998).
El modelo de automovilidad colonial requiereque las tecas permanezcan habitadas permanente-mente, con la brotación de nuevos individuos quereemplacen prontamente a los zooides muertos.Esta ocupación sucesiva de las tecas, por distintoszooides, ocurre en las colonias bentónicas de losrabdopléuridos y ha sido deducida también en losgraptolitos, a través de la identificación de discon-tinuidades y recrecimientos tecales imputables anuevos individuos (Rigby, 1994). La longevidadmedia de cada colonia no se conoce, pero, aplican-do el modelo pterobranquio, el monográptido máslargo del mundo podría haber vivido 25 años(Loydell & Loveridge, 2001).
Al igual que el resto del zooplancton, los grap-tolitos tendrían capacidad para migrar verticalmen-te en la columna de agua, bien siguiendo ciclosdiurno-nocturnos para eludir la actividad de preda-dores o los derivados fotosintéticos tóxicos delfitoplancton, o bien para mantenerse dentro de unrango de profundidad específica para cada especie
23HEMICORDADOS: GRAPTOLITOS

o grupo de especies, reduciendo de paso la compe-tición entre graptolitos de diferentes biotopos.
Un caso particular de adaptación la constituyenlos llamados sinrabdosomas, agrupaciones mono-específicas de graptolitos, unidos en forma radiadamediante el entrelazamiento de fibras emanadasdel extremo del nema o vírgula de cada colonia(rara vez por las virguellas): Fig. 8.14B-D. Elnúmero de colonias reunidas en un sinrabdosomaes de una veintena por término medio, y son siem-pre estructuras muy raras en el registro fósil de losgraptolitos, evidenciadas hasta el momento enmenos de 40 especies entre el Ordovícico basal yel Devónico inferior. La interpretación más proba-ble es considerar los sinrabdosomas como “estruc-turas de emergencia”, en virtud de las cuales lascolonias de ciertas especies de morfología simplecrecerían agregadas, con el fin de explotar losnutrientes presentes en masas de agua disaeróbi-cas, por coincidir su registro generalmente contiempos de crisis entre el plancton oceánico(Gutiérrez-Marco & Lenz, 1998). La sustentaciónpasiva por fricción, en las aguas profundas y den-sas, aumenta en proporción directa al tamaño, yésta es la razón por la que tampoco se necesitan los“flotadores” (neumatóforos) con los que se veníanrepresentando hasta ahora los sinrabdosomas, enuna reconstrucción vital de los graptolitos que,lejos de ser la regla, es claramente la excepción.
En la transición evolutiva de los graptolitosbentónicos sésiles a las primitivas formas planctó-nicas, algunos géneros, como Rhabdinopora, des-arrollaron nematulario, sinrabdosomas o bien unlargo nema terminado en un sencillo disco de fija-ción, con el que tal vez se sujetarían a algas flotan-tes del tipo sargazo, en un hábito pseudoplanctóni-co, restringido a los estadios astogenéticos inicia-les de pocas especies.
El varillaje estructural reticulado, característicode los retiolítidos (Fig. 8.15), supone la existenciade un espacio de separación entre el armazónexterno (manga del áncora) y las paredes tecales alas que envuelve, y ha sido interpretado por algu-nos autores como un órgano especializado, aunquede funciones todavía poco claras. En ejemplaresexcepcionales, se comprueba que tanto las paredestecales internas como el retículo externo estaban
cerrados por tejido fuselar, por lo que sus escasasaberturas funcionarían como orificios aferentes yeferentes, autopropulsando la colonia y compo-niendo un único aparato nutriente a escala colonial.Ello concuerda con la morfología hidrodinámicade muchos géneros (Fig. 8.15J). Antes de eso, sepensaba que el retículo constituía un refuerzoestructural del rabdosoma, como adaptación a lavida en aguas superficiales turbulentas, lo quepodría ser cierto en los falsos retiolítidos delOrdovícico. En cualquier caso, la manga del ánco-ra representó una estructura muy eficaz, dada suhomogeneidad y persistencia durante todo elSilúrico, sobreviviendo los retiolítidos a crisis evo-lutivas importantes que afectaron en forma dramá-tica a otros graptolitos más generalistas.
El hábito nectoplanctónico preponderante enalgunos otros graptoloideos podría haberse des-arrollado ya en formas primitivas de anisográpti-dos, como Araneograptus (Fig. 8.10D), cuyos rab-dosomas cónicos podrían medir 30 cm de longitudy haber generado corrientes aferentes-eferentesencauzadas por unos disepimentos oblicuos a losestipes. Su modo de vida se aproximaría al deltunicado actual Pyrosoma spinosum, cuyas colo-nias cónico-cilíndricas se propulsan lentamente yalcanzan decenas de metros de longitud.
8. PALEOBIOGEOGRAFÍA
Por tratarse de organismos mayoritariamenteplanctónicos, los graptolitos alcanzaron una enor-me dispersión en todos los mares ordovícico-devó-nicos, donde su distribución atravesó periodos deacusado provincialismo faunístico. Éste fue espe-cialmente patente en el Ordovícico Inferior yMedio, cuando se diferenciaron una ProvinciaPacífica (de aguas cálidas tropicales) y unaProvincia Atlántica (de aguas frías a templadas-frías), mediando zonas de transición en el inter-cambio de corrientes de uno u otro signo. Este con-trol primario aparente de la temperatura de lasaguas marinas sobre la distribución de los graptoli-tos, se combina, sin embargo, con otros muchosparámetros que regulan la abundancia y diversidadde los seres planctónicos actuales. Los más impor-tantes son la profundidad y la dinámica oceánica,
24 PALEONTOLOGÍA DE INVERTEBRADOS

las cuales condicionan el quimismo, propiedadesfísicas y contenido en nutrientes de ciertas masasde agua circundantes a la plataforma. Así, los grap-tolitos proliferaron en aquellas áreas con mayorproductividad para el fitoplancton, en especial, lasrelacionadas con áreas de afloramiento (upwelling)de aguas ricas en nutrientes, localizadas en ciertosmárgenes continentales de manera estable o esta-cional; en tanto que el centro de los océanos cons-tituiría, en líneas generales, un desierto para losgraptolitos (Finney & Berry, 1997).
La diferenciación lateral de masas de agua espe-cíficas, combinada con la profundidad, establecedos hábitats esenciales para los graptolitos (Fig.8.16): por un lado, el biotopo de aguas someras(hasta 100-150 m), en donde predominaron tantolas formas epipelágicas pandémicas (en océanos ymárgenes continentales), como las epipelágicasendémicas (restringidas, en este caso, a las áreas deplataforma interna); y, por otro, el biotopo deaguas profundas (150-1000 m), con claro predo-minio de las colonias mesopelágicas cónicas amultirramosas, de distribución cosmopolita(Cooper et al., 1991; Cooper, 1998; Egenhoff &Maletz, 2007). Así, en los depósitos correspon-dientes a las zonas neríticas, se registran graptoli-tos bentónicos a planctónicos epipelágicos, con
una neta proporción de formas endémicas en lasáreas de plataforma interna a media (la llamadabiofacies de didymográptidos del Ordovícico),cuya composición varía de un paleocontinente aotro dependiendo de la latitud y la temperaturasuperficial del agua. En los ambientes de platafor-ma distal y parte alta del talud continental (zonasnerítica externa a batial superior), los graptolitospredominantes son formas epipelágicas cosmopo-litas, con mezcla de endémicos en la primeraregión y de formas mesopelágicas en la segunda.El registro fósil corresponde entonces a la llamadabiofacies de isográptidos (Ordovícico), representa-da también en ambientes aún más profundos de laspartes medias y altas del talud continental, dondeconcurre una mezcla de graptolitos muy diversos,derivados de los diferentes biotopos de aguassomeras y profundas, ocasionalmente acompaña-dos por raros fragmentos de graptolitos bentónicos,transportados éstos por corrientes de turbidez agran distancia de sus ecosistemas neríticos origina-les. Por último, en los sedimentos batiales profun-dos a abisales, únicamente se acumulan restos degraptolitos mesopelágicos y raros epipelágicos,con baja diversidad, siendo improbable la existen-cia de graptolitos batipelágicos (de profundidadessuperiores a 1000 m).
25HEMICORDADOS: GRAPTOLITOS
Figura 8.16. Modelo esquemático de la distribución plataforma-océano de los graptolitos planctónicos en un margen continental con alta pro-ductividad orgánica (sin escala, modificado de Cooper, 1998, y Egenhoff & Maletz, 2007; con ideas de Finney & Berry, 1997). La prolifera-ción de formas pandémicas de graptolitos estuvo vinculada con aguas hipóxicas y desnitrificadas dentro de la “zona de oxígeno mínimo” (80-400 m), desarrollada en áreas de ascenso y afloramiento (upwelling) de masas oceánicas densas y ricas en nutrientes (flechas), lo cual sucedetanto en el margen continental, como en regiones costeras sometidas a fuertes vientos, coincidiendo especialmente con ciclos transgresivos.

El desarrollo del modelo plataforma-océano,con biotopos diferenciados por la profundidad,explica bastante bien las fluctuaciones en la com-posición de las asociaciones de graptolitos regis-tradas en ciertas localidades, sin duda vinculadascon ciclos eustáticos. De este modo, los máximostransgresivos estarían marcados por la penetraciónde las biofacies de isográptidos en las plataformasmarinas y su superposición a las biofacies previasde didymográptidos, y las regresiones, por el retor-no de estas últimas y el empobrecimiento en suregistro. Por otra parte, el modelo actualista deFinney & Berry (1997) interpreta satisfactoriamen-te la difícil supervivencia de los graptolitos en losocéanos y las áreas costeras, así como la naturale-za cambiante y efímera de sus hábitats más próspe-ros en términos de abundancia y diversidad, vincu-lados con las áreas de afloramiento de determina-das masas de agua en los márgenes continentales.La riqueza en nutrientes minerales y orgánicos deaquéllas, favorece la proliferación en su seno delplancton, además del florecimiento bacteriano enaguas fuertemente desnitrificadas en torno a lazona de oxígeno mínimo, que a menudo se estabi-liza durante largos periodos bajo las áreas de aflo-ramiento. Tanto aquellos graptolitos epipelágicosque prosperaron favorecidos por los blooms denanoplancton, como los que vivieron a expensas delas bacterias nitratorreductoras, en la zona de oxí-geno mínimo, contribuyeron decisivamente a laformación de “lutitas calientes” (hot-shales) en elSilúrico del norte de Gondwana, roca madre del4% de las reservas mundiales de hidrocarburos.Éstas prueban que el desarrollo de fondos anóxicosno es exclusivo de los márgenes continentales, sinoque ocurre ocasionalmente también en plataformasmarinas someras, por estratificación de masas deagua de diferente densidad, esencialmente durantelas etapas transgresivas (Finney et al., 2007). Enestas condiciones, las aguas superficiales bien oxi-genadas, calentadas por el sol y ricas en nutrientesminerales, fruto tanto del aporte fluvial como delas corrientes ascendentes de upwelling creadaspor vientos que soplan desde el continente, se com-binan con otras capas más densas, disaeróbicas,salinas y frías, que se sitúan por debajo de las ante-riores, y en las que se llega a formar una zona des-nitrificada. La elevada productividad de esta últi-
ma y de las aguas superficiales, hace que en elfondo se acumulen grandes cantidades de carbonoorgánico, hasta el punto de convertirlo en anóxico,lo que favorece la formación de sapropeles y laconservación de los graptolitos pelágicos.
9. INTERÉS BIOSTRATIGRÁFICO
Los graptolitos constituyen un grupo fósil deindudable interés geológico, especialmente en lossistemas Ordovícico y Silúrico, cuya subdivisión yestratotipos de límite coinciden a nivel mundialcon numerosas biocronozonas de graptoloideos.Ello se debe al hecho de que los graptolitos máscomunes reúnen todas las características exigiblesa los buenos fósiles-guía: amplia distribución geo-gráfica (relacionada con el modo de vida planctó-nico), abundancia en el registro (fosilizan en rocassedimentarias representativas de variados ambien-tes de plataforma marina, talud y fondo oceánico)y evolución rápida (duración media de las especiesrelativamente breve). Este último carácter se mani-fiesta ya a niveles supragenéricos, pudiéndose uti-lizar la sucesión clásica de “faunas” (anisográpti-da, dicográptida, diplográptida, monográptida)como guía primaria de edad. La pervivencia tem-poral de las especies de graptolitos es muy breve,con una media para el Silúrico inferior al millón deaños, alcanzando los doscientos cincuenta mil añosen muchas especies índices de biozona o subzona.Con ello, la resolución biocronológica alcanzadapor los graptoloideos ordovícico-devónicos supe-ra, en precisión y fiabilidad, a la derivada de losmétodos geocronométricos disponibles actualmen-te para estos mismos periodos, cuyo sólo margende error tiene una duración equivalente por lomenos a dos biozonas de graptolitos silúricas.
26 PALEONTOLOGÍA DE INVERTEBRADOS
Figura 8.17. Biocronozonas de graptolitos y cronoestratigrafía glo-bal del Ordovícico-Devónico Inferior, con indicación (a la derecha)de los principales acontecimientos de extinción ocurridos entre losgraptoloideos planctónicos. Las cifras expresan la antigüedad (enmillones de años) establecida por la IUGS para los estratotipos delímite respectivos. Las especies consignadas entre paréntesis denotanalternativas al taxón nominal de biocronozonas concretas, y no serefieren a equivalencias taxonómicas. Cuando una biocronozonacombina la extensión de dos taxones, el primero de los citados prece-de estratigráficamente al segundo. La revisión del límite Llandovery-Wenlock en su estratotipo británico, lo reubicará previsiblemente enla base de la Biocronozona de C. murchisoni (línea horizontal discon-tinua). Abreviaturas de la escala regional del Ordovícico mediterrá-neo: Ar, Arenigiense; Or, Oretaniense; Dob, Dobrotiviense; Ber,Berouniense; Kld, Kralodvoriense.
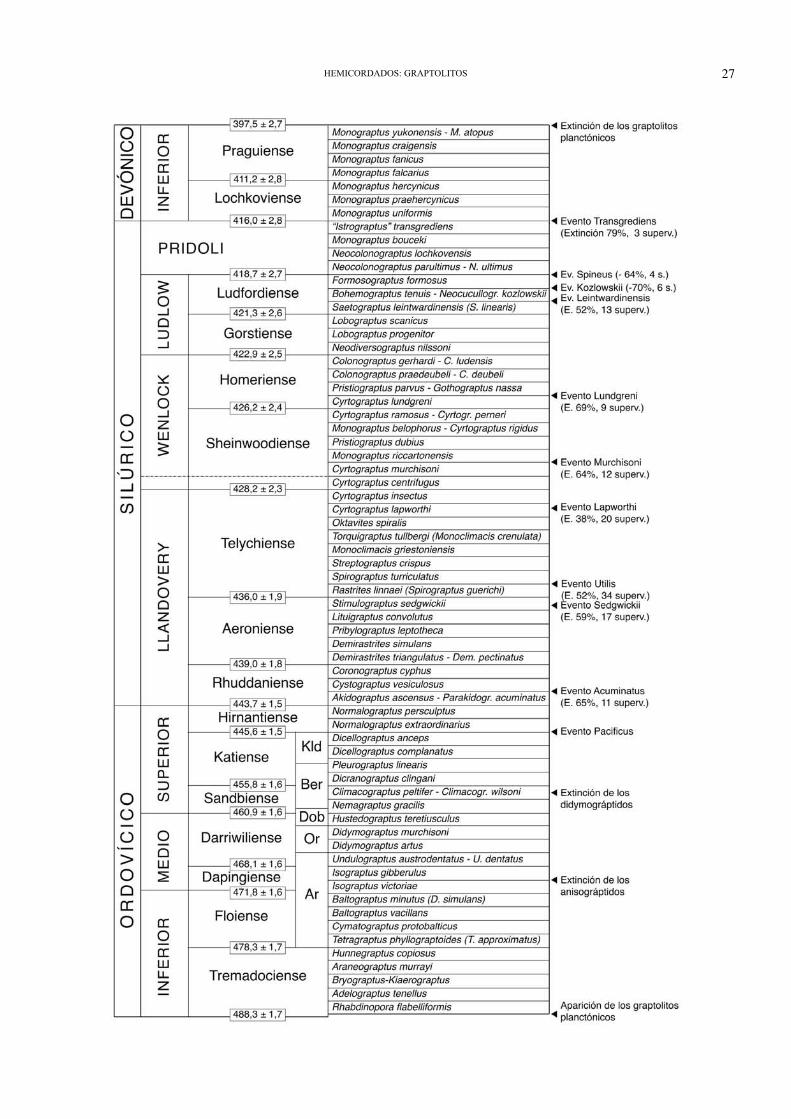
27HEMICORDADOS: GRAPTOLITOS

En Europa se han definido 23 biozonas de grap-tolitos para el Ordovícico, 36 para el Silúrico y 7para el Devónico inferior (Fig. 8.17). En las regio-nes pacíficas (Norteamérica, Australia), el númerode biozonas ordovícicas es aún mayor, y este hechoestá relacionado con el provincialismo faunísticodel Ordovícico Inferior y Medio, pues a comienzosdel Ordovícico Superior las biozonas toman uncarácter prácticamente cosmopolita. No obstante,en el Silúrico ibérico se registra también un ciertoprovincialismo a nivel específico durante elLlandovery y Wenlock, que comparte rasgos bio-geográficos con las plataformas perigondwánicasubicadas en el norte de África y el centro-sur deEuropa.
Los principales estudios bioestratigráficos sobregraptolitos españoles muestran la existencia desucesiones silúrico-devónicas especialmente rele-vantes en la Zona de Ossa Morena, Pirineos yCadenas Costeras Catalanas (Jaeger & Robardet,1979; Julivert et al., 1985; Degardin, 1988; Lenz etal., 1996; Gutiérrez-Marco et al., 1996; Piçarra etal., 1998), así como en el Silúrico de la CordilleraIbérica (Gutiérrez-Marco & Storch, 1998) y envariadas localidades del Ordovícico en el conjuntodel Macizo Hespérico (Gutiérrez-Marco, 1986).
10. BIBLIOGRAFÍA
Armstrong, W.G., Dilly, P.N. & Urbanek, A. 1984. Collagenin the pterobranch coenecium and the problem of grapto-lite affinities. Lethaia, 17, 145-152.
Bates, D.E.B. 1987. The construction of graptolite rhabdo-somes in the light of ultrastructural studies. Indian Journalof Geology, 9, 1-28.
Bates, D.E.B. 1989. Los graptolites. Mundo Científico, 9(88), 148-154 [traducción de La Recherche, 1988, 19(205), 1476-1482].
Bates, D.E.B., Kozlowska, A. & Lenz, A.C. 2005. Silurianretiolitid graptolites: Morphology and evolution. ActaPalaeontologica Polonica, 50, 705-720.
Bates, D.E.B., Kozlowska, A., Loydell, D., Urbanek, A. &Wade, S. 2009. Ultrastructural observations on some den-droid and graptoloid graptolites and on Mastigograptus.Bulletin of Geosciences, 84, 21-26.
Briggs, D.E.G., Kear, A.J., Baas, M., De Leeuw, J.W. &Rigby, S. 1995. Decay and composition of the hemichor-date Rhabdopleura: implications for the taphonom y ofgraptolites. Lethaia, 28, 15-23.
Bulman, O.M.B. 1970. Graptolithina with sections onEnteropneusta and Pterobranchia. In: Treatise on
Invertebrate Paleontology, part V (revised) (Ed. C.Teichert). Geological Society of America and TheUniversity of Kansas Press, Lawrence, 163 pp.
Chapman, A.J., Durman, P.N. & Rickards, R.B. 1995.Rhabdopleuran hemichordates: new fossil forms andreview. Proceedings of the Geologists’ Association, 106,293-303.
Chen, X., Zhang, Y.-d. & Fan, J.-x. 2006. Ordovician grapto-lite evolutionary radiation: a review. Geological Journal,41, 289-301.
Cooper, R.A. 1998. Towards a general model for the depthecology of graptolites. In: Proceedings 6th InternationalGraptolite Conference (Eds. J.C. Gutiérrez-Marco & I.Rábano). Madrid. Temas Geológico-Mineros ITGE, 23,161-163.
Cooper, R.A., Fortey, R.A. & Lindholm, K. 1991. Latitudinaland depth zonation of early Ordovician graptolites.Lethaia, 24, 199-218.
Crowther, P.R. 1981. The fine structure of graptolite perid-erm. Special Papers in Palaeontology, 26, 1-119.
Crowther, P.R. & Rickards, R.B. 1977. Cortical bandages andthe graptolite zooid. Geologica et Palaeontologica, 11, 9-46.
Degardin, J.-M. 1988. Le Silurien des Pyrénées.Biostratigraphie, Paléogéographie. Société Géologiquedu Nord, Publication 15, 525 pp.
Egenhoff, S. & Maletz, J. 2007. Graptolites as indicators ofmaximum flooding surfaces in monotonous deep-watershelf successions. Palaios, 22, 373-383.
Finney, S.C. & Berry, W.B.N. 1997. New perspectives ongraptolite distributions and their use as indicators of plat-form margin dynamics. Geology, 25, 919-922.
Finney, S.C., Berry, W.B.N. & Cooper, J.D. 2007. The influ-ence of denitrifying seawater on graptolite extinction anddiversification during the Hirnantian (latest Ordovician)mass extinction event. Lethaia, 40, 281-291.
Fortey, R.A. & Cooper, R.A. 1986. A phylogenetic classifica-tion of the graptoloids. Palaeontology, 29, 631-654.
Fortey, R.A., Zhang, Y.-d. & Mellish, C. 2005. The relation-ships of biserial graptolites. Palaeontology, 48, 1241-1272.
Gupta, N.S., Briggs, D.E.G. & Pancost, R.D. 2006.Molecular taphonomy of graptolites. Journal of theGeological Society, London, 163, 897-900.
Gutiérrez Marco, J.C. 1986. Graptolitos del Ordovícicoespañol. Tesis doctoral, Universidad Complutense deMadrid, 3 vols., 701 pp. (inéd.).
Gutiérrez-Marco, J.C. & Lenz, A.C. 1998. Graptolite syn-rhabdosomes: biological or taphonomic entities?.Paleobiology, 24, 37-48.
Gutiérrez-Marco, J.C. & Storch, P. 1998. Graptolite biostra-tigraphy of the Lower Silurian (Llandovery) shelf depositsof the Western Iberian Cordillera, Spain. GeologicalMagazine, 135 (1), 71-92.
28 PALEONTOLOGÍA DE INVERTEBRADOS

Gutiérrez-Marco, J.C., Lenz, A.C., Robardet, M. & Piçarra,J.M. 1996. Wenlock-Ludlow graptolite biostratigraphyand extinction: a reassessment from the southwesternIberian Peninsula (Spain and Portugal). Canadian Journalof Earth Sciences, 33, 656-663.
Jaeger, H. & Robardet, M. 1979. Le Silurien et le Dévonienbasal dans le nord de la province de Seville (Espagne).Geobios, 12, 687-714.
Jones, H., Zalasiewicz, J. & Rickards, B. 2002. Clingfilmpreservation of spiraliform graptolites: Evidence of organ-ically sealed Silurian seafloors. Geology, 30, 343-346.
Julivert, M., Durán, H., Rickards, R.B. & Chapman, A.J.1985. Siluro-Devonian graptolite stratigraphy of theCatalonian Coastal Ranges. Acta Geològica Hispànica,20, 199-207.
Kirk, N.H. 1994. Coordination of construction and function-ing of the graptolite colony. In: Graptolite Research Today(Eds. X. Chen, B.-D. Erdtmann &Y.-n. Ni). NanjingUniversity Press, 197-216.
Lenz, A.C., Robardet, M., Gutiérrez-Marco, J.C. & Piçarra,J.M. 1996. Devonian Graptolites from southwesternEurope: a review with new data. Geological Journal, 31,349-358.
Loydell, D.K. & Loveridge, R.F. 2001. The world’s longestgraptolite?. Geological Journal, 36, 55-57.
Maletz, J., Carlucci, J. & Mitchell, C.E. 2009. Graptoloidcladistics, taxonomy and phylogeny. Bulletin ofGeosciences, 84, 7-19.
Maletz, J., Steiner, M. & Fatka, O. 2005. Middle Cambrianpterobranch and the Question: What is a graptolite?.Lethaia, 38, 73-85.
Melchin, M.J. 1998. Morphology and phylogeny of someEarly Silurian ‘Diplograptid’ genera from CornwallisIsland, Artic Canada. Palaeontology, 41, 263-315.
Melchin, M.J. & DeMont, M.E. 1995. Possible propulsionmodes in Graptoloidea: a new model for graptoloid loco-motion. Paleobiology, 21, 110-120.
Melchin, M.J. & Doucet, K.M. 1996. Modelling flow pat-terns in conical dendroid graptolites. Lethaia, 29, 39-46.
Melchin, M.J., Koren, T.N. & Storch, P. 1998. Global diver-sity and survivorship patterns of Silurian graptoloids. NewYork State Museum Bulletin, 491, 165-182.
Mierzejewski, P. & Kulicki, C. 2002. Discovery ofPterobranchia (Graptolithoidea) in the Permian. ActaPalaeontologica Polonica, 47, 169-175.
Mierzejewski, P. & Kulicki, C. 2003. Cortical fibrils andsecondary deposits in periderm of the hemichordateRhabdopleura (Graptolithoidea). Acta PalaeontologicaPolonica, 48, 99-111.
Mitchell, C.E. 1987. Evolution and phylogenetic classifica-tion of the Diplograptacea. Palaeontology, 30, 353-405.
Mitchell, C.E., Goldman, D., Klosterman, S.L., Maletz, J.,Sheets, H.D. & Melchin, M.J. 2007. Phylogeny of the
Ordovician Diplograptoidea. Acta PalaeontologicaSinica, 46 (Suppl.), 332-339.
Piçarra, J.M., Gutiérrez-Marco, J.C., Lenz, A.C. & Robardet,M. 1998. Prídoli graptolites from the Iberian Peninsula: areview of previous data and new records. CanadianJournal of Earth Sciences, 35, 65-75.
Rantell, L. & Rigby, S. 2002. The hydrodynamic significanceof the graptoloid nema in scandent graptoloids.Senckenbergiana Lethaea, 82, 131-140.
Rickards, R.B. 1996. The graptolite nema: problems to allour solutions. Geological Magazine, 133, 343-346.
Rickards, R.B. & Dumican, L.W. 1984. The fibrillar compo-nent of the graptolite periderm. Irish Journal of EarthSciences, 6, 175-203.
Rickards, R.B. & Dumican, L.W. 1985. Graptolite ultrastruc-ture: evolution of descriptive and conceptual terminology.Geological Magazine, 122, 125-137.
Rickards, R.B. & Durman, P.N. 2006. Evolution of the earli-est graptolites and other hemichordates. In: Studies inPalaeozoic palaeontology (Eds. M.G. Basset & V.K.Deisler). National Museum of Wales Geological Series 25,Cardiff, 5-92.
Rickards, R.B. & Stait, B.A. 1984. Psigraptus, its classifica-tion, evolution and zooid. Alcheringa, 8, 101-111.
Rickards, R.B., Crowther, P.R. & Chapman, A.J. 1982.Ultrastructural studies of graptolites - a review.Geological Magazine, 119, 355-370.
Rickards, R.B., Rigby, S., Rickards, J. & Swales, C. 1998.Fluid dynamics of the graptolite rhabdosome recorded bylaser Doppler anemometry. Palaeontology, 41, 737-752.
Rigby, S. 1994. Hemichordate skeletal growth: shared pat-terns in Rhabdopleura and graptoloids. Lethaia, 27, 317-324.
Rigby, S. & Sudbury, M. 1995. Graptolite ontogeny and thesize of the graptolite zooid. Geological Magazine, 132,427-433.
Sato, A., Rickards, B. & Holland, P.W.H. 2008. The originsof graptolites and other pterobranchs: a journey from‘Polyzoa’. Lethaia, 41, 303-316.
Sdzuy, K. 1974. Mittelkambrische Graptolithen aus NW-Spanien. Paläontologische Zeitschrift, 48, 110-139.
Starcher, R.W. & McGhee, G.R., Jr. 2003. Fenestrate grapto-lite theoretical morphology: geometric constraints onlophophore shape and arrangement in extinct hemichor-dates. Journal of Paleontology, 77, 360-367.
Sudbury, M. 1991. The dimensions of the graptolite zooid.Geological Magazine, 128, 381-384.
Towe, K.M. & Urbanek, A. 1972. Collagen-like structures inOrdovician graptolite periderm. Nature, 237, 443-445.
Underwood, C.J. 1993. The position of graptolites within LowerPalaeozoic planktic ecosystems. Lethaia, 26, 189-202.
Underwood, C.J. 1998. Population structure of graptoliteassemblages. Lethaia, 31, 33-41.
29HEMICORDADOS: GRAPTOLITOS

Urbanek, A. 1986. The enigma of graptolite ancestry: a les-son for a phylogenetic debate. In: Problematic Fossil Taxa(Eds. A. Hoffmann & M.H. Nitecki). Oxford Monographson Geology and Geophysics 5, Clarendon Press, Oxford,184-226.
Urbanek, A. 1990. Reproductive strategy and the life cycle ingraptoloid colonies. Lethaia, 23, 333-340.
Urbanek, A. 1994. When is a pterobranch a graptolite?.Lethaia, 27, 324.
Urbanek, A. 2004. Morphogenetic gradients in graptolitesand bryozoans. Acta Palaeontologica Polonica, 49, 485-504.
Urbanek, A. & Dilly, P.N. 2000. The stolon system inRhabdopleura compacta (Hemichordata) and its phyloge-netic implications. Acta Palaeontologica Polonica, 45,201-226.
Urbanek, A. & Mierzejewski, P. 1984. The ultrastructure ofthe Crustoidea and the evolution of graptolite skeletal tis-sues. Lethaia, 17, 73-91.
30 PALEONTOLOGÍA DE INVERTEBRADOS
Nota: a la hora de componer esta separata, destinada a la difusión personal del capítulo del libro citadoen el encabezamiento (la paginación real es la que se indica al comienzo del capítulo), cabría añadir lasreferencias a otros tres trabajos aparecidos durante su publicación, y que ayudan a profundizar, respec-tivamente, en el conocimiento de las restricciones ecológicas del plancton graptolítico, nuevas especiesde graptolitos del Silúrico español, y en la actualización bioestratigráfica de 698 especies de graptolitosdel Ordovícico y Silúrico británico, cuna de la mayoría de los estudios tradicionales sobre este grupofósil. Boucot, A.J. & Chen, X. 2009. Fossil plankton depth zones. Palaeoworld, 18, 213-234.Loydell, D.K., Sarmiento, G.N., Storch, P. & Gutiérrez-Marco, J.C. 2009. Graptolite and conodont biostratigraphy of the
upper Telychian-lower Sheinwoodian (Llandovery-Wenlock) shales of the Jabalón River section, Corral de Calatrava,Spain. Geological Magazine, 146, 187-198.
Zalasiewicz, J.A., Taylor, L., Rushton, A.W.A., Loydell, D.K., Rickards, R.B. & Williams, M. 2009. Graptolites in Britishstratigraphy. Geological Magazine, 146, 785-850.
PC
Resaltado


![Hemicordados, Protocordados e Cordados[2]](https://static.fdocuments.net/doc/165x107/557203084979599169a47292/hemicordados-protocordados-e-cordados2-55b5160289be4.jpg)















